Embed Size (px)
Citation preview

3ème partie : Neurosciences cognitives
Chapitre 2 –VIGILANCE, ATTENTION, MOTIVATION
La première étape du déroulement d'un comportement est permise s'il y a activation de l'organisme telle qu'elle résulte de la confrontation des conditions internes de la personne et des objets ou des situations présentes dans l'environnement. Cette confrontation est à l'origine de la motivation; elle-même naît de l'attention et de l'intention.
En d'autres termes, de l'activation de notre organisme dépend donc la motivation et les deux processus qui la sous-tendent, la perception, (l'attention) et l'orientation de notre action (l'intention). Sans la motivation, toute perception devient inutile. C'est le cas pour les individus en état de méditation profonde, dans le coma ou ayant atteint le seuil de la mort. D'un autre côté, sans la perception et la conscience que nous avons de la réalité extérieure (l'attention), nous serions dans l'incapacité d'orienter les comportements susceptibles de satisfaire nos besoins d'êtres vivant en interaction constante avec le monde (l'intention).
Dans ce chapitre, nous allons donc nous livrer à l'exploration de cette dimension de notre vie psychique, en tentant de comprendre les mécanismes à la base de nos motivations, de notre attention, ainsi que de la manière dont elles s'expriment en fonction des données du monde extérieur, c'est à dire notre intention.
I – LES ASPECTS DE LA CONSCIENCE
1) Activation de l'organisme Traditionnellement, la psychophysiologie occidentale reconnaît 2 états de conscience propres à tous les
individus : le sommeil d’une part, considéré comme une période repos, et l’état d’éveil (ou de veille, ou de vigilance active, ou de conscience ordinaire, ou encore de conscience de "surface") d’autre part. A noter que le sommeil (qu'il soit à ondes lentes –SOL- ou paradoxal –SP-) n'est pas déclenché comme l'a suggéré, au XIXème siècle, le psychologue William James, par une réduction de nos capacités perceptives. Bien au contraire, on admet aujourd'hui que le sommeil et la veille sont provoqués par l'activation de centres nerveux spécifiques (voir ci-dessous).
La vigilance active, la conscience ordinaire ou "de surface" correspond à une activation de tout l’organisme qui lui permet de capter, de sélectionner et de traiter les informations du monde extérieur, d’en garder certaines en mémoire ou de réagir à d’autres par des comportements appropriés ou non, selon les expériences ou les apprentissages antérieurs. L’état d’éveil est donc celui de l’adaptation à la réalité extérieure. Cet état de conscience normal et idéal se traduit, individuellement, essentiellement par la capacité de chacun de décoder et de répondre efficacement aux stimulations du milieu, d'une façon acceptée par la plupart des membres du groupe social d'appartenance. En effet, la manière dont nous prenons conscience à la fois de notre monde intérieur et du monde extérieur est largement "imprégnée" des habitudes culturelles du groupe social dans lequel nous nous identifions. Par ailleurs elle se transforme au cours du temps, à long terme, avec l'âge, et à court terme, le long d'une même journée. Le décodage des événements sera très différent selon par exemple que l'on est détendu ou anxieux, excité ou près de l'endormissement, jeune et inexpérimenté ou âgé.
Le terme de "conscience" est à l'évidence très vague et difficile à définir du point de vue neurobiologique, en raison de la polysémie de ce mot en langue française. On admet toutefois en général qu'il contient 2 concepts:
• celui de niveau de conscience ou d'éveil ("arousal"), apprécié par l'observation du comportement du sujet;
• celui de contenu de la conscience ("awareness"), beaucoup plus subjectif, et que l'on ne peut que présumer à partir d'indices indirects, comme le langage par exemple.
L'introduction du concept d'attention en neurologie s'est faite relativement récemment, à la lumière d'un certain nombre d'observations où des troubles comportementaux consécutifs à des lésions cérébrales suggéraient l'existence d'un système neuronal susceptible de sous-tendre les phénomènes dits attentionnels. Toutefois, il peut paraître hasardeux de suggérer qu'une fonction, jusqu'alors exclusivement explorée par les méthodes et les concepts de la psychologie, puisse être approchée d'un point de vue anatomique comme liée au fonctionnement de structures nerveuses propres et individualisables.
Vers le milieu des années 50, Hebb a introduit à l’usage des psychologues le terme d’activation aspécifique pour désigner le phénomène général qui fournit «l’énergie nécessaire au comportement sans diriger ce dernier». C’est le désir dépourvu de toute spécificité et considéré comme le fondement de la spontanéité. Mais pour que ce désir ou cette motivation s'exprime pleinement, il faut qu'il soit porté à un niveau optimal (au delà duquel il devient néfaste). Il a montré en effet que le niveau de performance augmente avec le niveau d'activation de l'organisme et que le meilleur ajustement de cette performance que permet la vigilance de l'organisme risque de se détériorer à partir d'un certain point si l'activation de l'individu se trouve augmentée de façon trop importante (fig. 1). Trop d’activation est néfaste à l’action. Hebb a défini un niveau optimal d’activation au delà duquel la performance décroît. Ce peut être le cas après une motivation trop forte ou une grave perturbation émotionnelle. L’étudiant qui doit à tout prix réussir un examen risque de perdre la concentration nécessaire à la compréhension des questions. Le sportif qui met fin à sa liaison amoureuse risque, pour sa part, de perdre tout intérêt pour le match qu’il va disputer .
204

Fig. 1 - La conscience de surface: les réactions aux stimuli dépendent du niveau de vigilance du
sujet. Celui-ci est capable de comportements d'autant plus efficaces que son état d'éveil n'est ni trop faible ni trop élevé. Quand le niveau d'éveil est trop bas, ou au contraire trop élevé, le niveau de performance devient médiocre (endormissement ou désorganisation complète des comportements). D'après Hebb PO, in "A Textbook of Psychology", Philadelphie, Baunders, 1966.
En définissant le concept d'activation, Hebb a également introduit la notion de "niveau optimal" de
performance. La courbe des performances comportementales d'un sujet en fonction de son niveau d'activation (représenté par exemple par la fréquence cardiaque) dessine une courbe en cloche. Ainsi s'explique l'effet paradoxal des tranquillisants qui, par exemple, chez un chauffeur très excité, améliorent les qualités de sa conduite alors que chez un sujet normal ils provoquent une diminution dangereuse des réflexes. On verra plus loin que le niveau d'activation défini par Hebb correspond à "l'état central fluctuant" défini par Vincent (Biologie des passions, éd. Odile Jacob, Paris, 1986).
Ainsi parmi les divers états de conscience qui constituent la toile de fond de notre vie mentale (cognitive et comportementale), l'état de vigilance active, propre à la conscience de surface, assure le lien de l'individu avec le monde extérieur en intégrant à tout moment les données qui en proviennent. Ce décodage de la réalité ne s'effectue cependant pas au hasard. L'attention que nous portons aux choses ou à certains événements peut, bien sûr, dépendre des caractéristiques mêmes de ceux-ci. Mais, le plus souvent, elle répond à un besoin de «faire» quelque chose. Ce besoin peut parfois être lié à la survie; nous sélectionnons alors les informations susceptibles de déboucher sur l'apaisement de la faim, de la soif ou sur l'évitement d'un danger. Quelquefois, il s'agit d'occuper le temps et notre attention se porte alors sur les stimulations de l'environnement les plus susceptibles de répondre à cette motivation. La plupart du temps néanmoins, nous orientons nos comportements en fonction des perceptions qui nous feront mener à bien un projet en cours, ou planifié à plus ou moins long terme.
Ici, comme souvent en neurologie et en neuro-psychologie, nos connaissances du substrat neurobiologique de ces fonctions résultent principalement de l'observation d'états pathologiques où elles ont été perturbées par des lésions cérébrales. On applique à ces états le terme générique de comas (voir ci-dessous).
2) Définition de l'attention Ici encore, les définitions et les concepts feront appel, plutôt qu'à la théorie psychologique classique, à
l'observation d'états mentaux particuliers survenant à la suite de lésions cérébrales et où la perturbation semble toucher électivement le domaine attentionnel. A cet égard, on peut dans un premier temps considérer l'attention comme une fonction cérébrale dont le but est de sélectionner parmi le flot des stimulations sensorielles qui parviennent simultanément et de manière incessante au cerveau, celles qui sont utiles, pertinentes, à la réalisation d'une activité motrice ou mentale. L'attention apparaît donc comme une propriété sensorielle supra-modale du cerveau. Ainsi considéré, le concept d'attention se situe à l'intersection de certains autres comme la vigilance, la concentration, la motivation, l'orientation, l'exploration, et ses frontières avec l'éveil et la conscience envisagés dans les paragraphes précédents ne sont certes pas très nettes. Toutefois, un certain nombre d'études expérimentales et surtout cliniques justifient une conception uniciste des phénomènes attentionnels, tout en conservant la distinction classiquement proposée entre une fonction tonique ou de base, responsable d'un éveil général (« arousal ») des fonctions mentales et habituellement rattachée au fonctionnement du système réticulaire ascendant, et une fonction phasique ou vectorielle, capable de contrôler l'efficacité d'un processus cérébral spécifique ("attention sélective"), et mise en relation avec des circuits neuronaux corticaux. Les comportements attentionnels quels qu'il soient (par exemple se concentrer sur l'écran d'un radar, ou sur une épreuve de calcul mental, ou sur une sensation viscérale interne) utilisent certes l'une des deux composantes de façon préférentielle mais nécessitent une activité optimale des deux, la perturbation d'un des 2 systèmes étant susceptible d'altérer à divers degrés la réalisation de la tâche.
Ainsi le niveau et la qualité de l'activation dépendent de 3 sources indissociables: • des niveaux de conscience et d'éveil dans lequel se trouve le cerveau. Une information offre peu
d'utilité lorsque l'organisme n'est pas dans un état d'éveil, un état de vigilance, un état de conscience suffisant pour être en état de l'intégrer. Cet éveil est notamment lié aux cycles naturels de veille-sommeil. Il peut être modifié par la méditation ou altéré par l'usage de certaines drogues;
• de la perception que nous avons du monde et qui nous est fournie par l'interprétation des informations captées par nos récepteurs. Il est à noter que nous sommes également sensibles à certaines informations de notre monde intérieur, aux images mentales et aux souvenirs conservés en mémoire;
• des besoins innés et des motivations acquises au cours de l'existence qui guident la recherche et le choix de notre "manière de vivre".
205

3) Troubles de la conscience autres que le coma
a) Mutisme akinétique On comprend sous ce terme divers états pathologiques où la vigilance elle-même paraît intacte mais où la
communication avec l'environnement est abolie ou sérieusement entravée. Ces états sont liés à des lésions intéressant de façon bilatérale les régions médianes du télencéphale lésions bilatérales des noyaux médians du thalamus, lésions des parois du 3ème ventricule, lésions de la région septale, lésions de la face interne des 2 lobes frontaux. Le tableau clinique, très difficile à appréhender, est dans tous les cas caractérisé par une absence de toute tentative de communication. Le sujet, les yeux ouverts, ne répond à aucune stimulation sensorielle. Seule la douleur peut donner une réaction, toutefois non dirigée vers sa source. Il peut parfois donner l'impression de suivre du regard un visage ou une lumière, mais le regard se perd rapidement dans des mouvements d'errance sans but apparent. Dans certains cas, toute motricité est abolie et le sujet est réellement mutique (c'est le cas, par exemple, de certaines lésions frontales bilatérales) dans d'autres, des mouvements spontanés dénués de propos existent, parfois accompagnés de gémissements incompréhensibles (les lésions sont alors situées plus bas, dans le thalamus ou l'hypothalamus médian). Ces états, qui ne sont ni paralysie, ni coma, semblent correspondre à une perturbation généralisée de l'activité corticale, comme en témoigne l'habituelle altération diffuse des tracés EEG, ou plus particulièrement des systèmes dopaminergiques corticaux.
b) Etats confusionnels Ils se rencontrent surtout lors de perturbations métaboliques ou toxiques, modifiant de façon globale
le fonctionnement cortical. Ici également, le trouble ne concerne pas le niveau de conscience mais son contenu. Les sujets sont éveillés,
bien que parfois en prise à des hallucinations quasi oniriques, mais la communication est encore très compromise; ils ne répondent pas à l'interlocuteur ou, à un moindre degré, font de multiples digressions dans leurs réponses. Lorsque l'on par vient toutefois à obtenir une "canalisation", même brève, du sujet, on peut noter qu'il est grossièrement désorienté dans le temps et dans l'espace, ne connaissant ni la date, ni le lieu où il se trouve. De même, il paraît atteint de troubles profonds de la mémoire. Bien que la littérature anglo-saxonne fasse référence à de tels états en tant que "trouble ou obscurcissement de la conscience", il semble plus adéquat de les inclure dans le cadre des troubles de l'attention.
c) Autres troubles: aphasie de Wernicke et syndrome de négligence contra-latérale Certains ont proposé de considérer comme troubles de la conscience tous les états neurologiques où, après
une lésion diffuse ou focale du cerveau, les sujets perdent la conscience de leur propre trouble. L'inclusion à ce titre, par exemple, de certains états aphasiques (aphasie de Wernicke, voir cours "langage"), de même que de nombreuses conséquences de lésions hémisphériques droites (voir cours "attention intention") relèvent d'un élargissement excessif du concept de conscience risquant, au moins du point de vue neurobiologique, de rendre la terminologie plus confuse plutôt que de l'expliciter.
En 1941, le neurologue britannique WR Brain publia les cas d'une série de patients porteurs de lésions
unilatérales du lobe pariétal s'accompagnant difficultés perceptives plus ou moins prononcées. Voici la description que fait Brain des cas les plus graves: "Trois patients présentaient une forme tout à fait différente de désorientation spatiale. Quoique leur mémoire des lieux fût intacte, de même que leur capacité de décrire des itinéraires familiers, ils se perdaient cependant chez eux, en allant d'une pièce à l'autre, commettant toujours la même erreur de tourner à droite au lieu de tourner à gauche ou de prendre la porte de droite au lieu de celle de gauche. Dans chaque cas, il y avait une lésion massive de la région pariéto~occipitale droite et l'on suggère que telle était la cause de l'inattention ou de la négligence à l'égard de la moitié gauche de l'espace externe". WR. Brain, 1941, (Brain, 64, p. 257)
À partir de ces observations et d'autopsies ultérieures, Brain résuma ainsi les caractéristiques de ce syndrome: "Le patient qui se trouve ainsi privé des sensations qui lui sont nécessaires pour construire un schéma de son corps, peut réagir à la situation de différentes façons. Il peut se souvenir que ses membres de gauche sont toujours là ou bien il peut les oublier régulièrement jusqu'à ce qu'on lui rappelle leur présence. Il peut avoir l'illusion qu'ils sont absents, c'est-à-dire avoir l'impression qu'ils sont absents tout en sachant qu'ils sont là; il peut croire qu'ils sont absents mais se rendre à l'évidence et se laisser convaincre du contraire; ou bien, enfin, sa croyance en leur absence peut être réfractaire à toute logique et à l'évidence du contraire et constituer ainsi une illusion Son état [est] ainsi comparable à une amnésie pour la moitié gauche du corps.."
On considère cette description comme le premier compte-rendu complet du lien entre les lésions du lobe pariétal et les déficits de la perception et de l'attention. Beaucoup d'autres patients ont été étudiés depuis les travaux d'avant-garde de Brain et l'on désigne aujourd'hui les troubles qu'ils présentent sous le nom de syndrome de négligence controlatérale.
La caractéristique de la négligence controlatérale est l'incapacité de percevoir son corps ou les objets de l'espace en relation avec le corps et d'y prêter attention en dépit d'une acuité visuelle, d'une sensibilité somatique et d'aptitudes motrices intactes. C'est ainsi que les personnes atteintes de ces troubles n'arrivent ni à signaler les stimulus présentés du côté du corps (ou de l'espace visuel) opposé à la lésion cérébrale, ni à y répondre ni à s'orienter vers eux. Ils peuvent également présenter des difficultés à exécuter des tâches motrices complexes du côté négligé, par exemple pour s'habiller, aller prendre des objets, écrire, dessiner et, à un degré moindre, pour s'orienter vers des sons. La négligence peut se manifester par des troubles aussi légers qu'un manque temporaire d'attention, se dissipant à mesure que le malade récupère, ou par une dénégation permanente et totale de tout le côté opposé à la lésion. Comme l'avait déjà fait remarquer Brain, les patients peuvent même refuser de reconnaître que les membres du côte négligé sont bien à eux.
Depuis la description originale de la négligence controlatérale et des ses relations avec le lobe pariétal, on a généralement admis que le cortex pariétal, et plus particulièrement le lobe pariétal inférieur, est la région corticale dont dépend essentiellement l'attention.
Ainsi, les patients présentant une lésion unilatérale du cerveau se comportent comme s'ils ignoraient les événements survenant dans la moitié opposée de l'espace extra-personnel. Ce syndrome, dit aussi d'héminégligence, peut se manifester de façon plus ou moins intense. Dans sa forme complète, généralement liée à une lésion
206

hémisphérique droite, le sujet semble «oublier» ou négliger tout ce qui concerne la moitié gauche de l'espace il ne mangera, par exemple, que la nourriture située sans la partie droite de son plateau ou son assiette, ne lira que la moitié droite d'un texte, dont la compréhension en sera par là-même compromise. Dans une épreuve consistant à décrire une scène représentée sur un dessin, il ne décrira que la partie droite de la scène. Si on lui demande d'écrire ou de copier un dessin, il n'utilisera que la moitié droite de la feuille et ne copiera que la partie droite du modèle. Bien entendu, le sujet ne présente aucun trouble perceptif, en particulier, il n'existe pas d'amputation du champ visuel (hémianopsie). Ce syndrome d'hémi-négligence est actuellement interprété comme une perturbation des processus attentionnels.
Les syndromes (ou états) confusionnels cités plus haut sont considérés comme des perturbations attentionnelles. Ici, hormis un état de vigilance fluctuant, le sujet a perdu une partie de ses capacités à se concentrer, à maintenir une suite cohérente à ses idées et donc à ses paroles; il apparaît hautement distractible(1) et semble avoir perdu toutes possibilités de se repérer dans le temps et dans l'espace. Les épreuves dites de contrôle mental (par exemple donner les mois de l'année à l'envers), lorsqu'elles peuvent être pratiquées, montrent que le sujet ne peut inhiber la tendance à donner la séquence dans l'ordre habituel, ce qui traduit une impossibilité de se concentrer sur un acte programmé. En bref, le sujet confus semble avoir perdu la capacité d'inhiber les stimuli parasites internes ou externes, ce qui rend impossible l'exécution d'une tâche suivie, même simple.
(1) Ce terme est l'un des nombreux néologismes qu'a créés la neurologie. Il s'oppose à l'adjectif distrait; est distractible un sujet dont l'attention pourra être attirée de façon excessive par des stimuli extérieurs même minimes. Le distrait, au contraire, est souvent à ce point absorbé par ses pensées qu'il ne « fera pas attention » à ce qui se passe autour de lui.
Le syndrome de négligence controlatérale est spécifiquement associé aux atteintes du lobe pariétal de l'hémisphère droit. On estime que l'inégalité du partage de cette fonction cognitive particulière entre les deux hémisphères est due au fait que le cortex pariétal droit intervient à la fois dans l'attention aux moitiés droite et gauche du corps et de l'espace péripersonnel, tandis que l'hémisphère gauche ne prend en charge que la moitié droite De la sorte, les lésions du lobe pariétal gauche tendent à être compensées par l'hémisphère droit intact. En revanche, quand c'est le lobe pariétal droit qui est atteint, l'hémisphère gauche est incapable de compenser le déficit d'attention au côté gauche. II – LES SYSTEMES ACTIVATEURS
1) Rôle de la substance réticulée dans le niveau d'activation L'activation psychologique constitue le prolongement de l'activation physiologique. Elle dépend du
décodage qui est fait de la réalité extérieure en fonction du niveau d'éveil ou de l'état de conscience de la personne ainsi que de ses besoins, de ses goûts, de ses intérêts et de ses projets. De même qu'un caillou brillant trouvé sur un chemin ne signifie pas la même chose pour le géologue que pour le promeneur, un plat de nourriture n'évoque pas la même attitude chez un individu qui a faim et chez celui qui est rassasié.
Le concept d’activation est inséparable de la mise en évidence à la même époque du rôle de la substance réticulée. Ce concept selon lequel l'activation ne résulterait pas -comme on pourrait le penser intuitivement- d'une réduction des capacités perceptives, mais au contraire de l'activation de centre nerveux spécifiques, a pour la première fois été suggéré par Magoun et Moruzzi, en 1949 après avoir stimulé électriquement la formation réticulaire du tronc cérébral.
Ainsi l'activation physiologique a son foyer dans les centres situés à la base du cerveau. Ces centres abritent les mécanismes de l'éveil. C'est également à cet endroit que les informations en provenance des mondes extérieur et intérieur de l'individu sont centralisées et triées avant d'être acheminées par le biais de la formation réticulée, vers le cortex, lorsqu'elles son jugées suffisamment importantes. L'activation des centres supérieurs qui en résulte permet ainsi à l'organisme de demeurer vigilant et attentif aux messages de l'environnement, assurant le maintien de son équilibre tant biologique que psychologique. Dans la mesure ou la stimulation électrique de la substance réticulée produit un état de vigilance et d'éveil, on l'a appelé le système réticulaire activateur (fig. 2 et 3).
La formation réticulée forme un manchon cellulaire autour du 4ème ventricule et de l'aqueduc de Sylvius, qui occupe, de la moelle cervicale au diencéphale, l'espace laissé libre situé entre les grandes voies ascendantes et descendantes et les noyaux des nerfs crâniens. Elle est parsemée de nombreux noyaux dont le locus niger, le noyau rouge, le corps de Luys sont les plus importants. A l'étage diencéphalique, elle pénètre dans l'hypothalamus et dans le thalamus (substance réticulaire thalamique).
Distribution diffuse des projections de la formation réticulaire sur l'écorce cérébrale
Une voie sensorielle spécifique (la voie somesthésique) est indiquée en flèches ponctuées
207
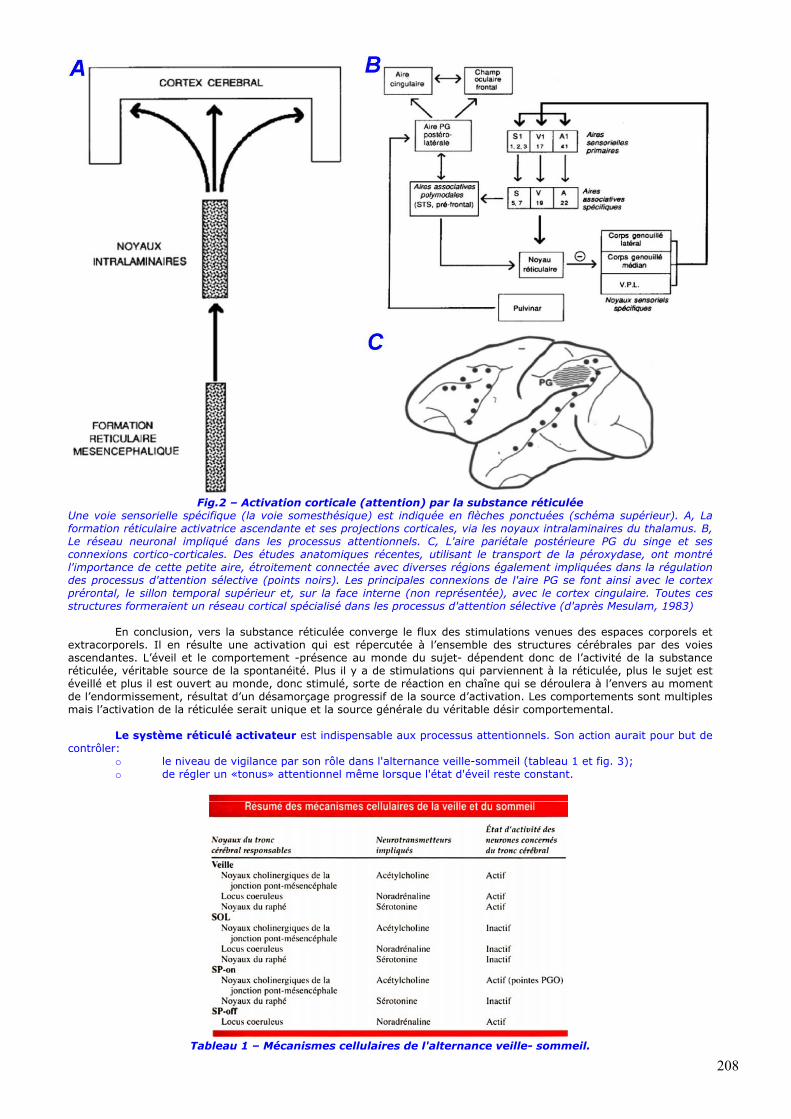
Fig.2 – Activation corticale (attention) par la substance réticulée
Une voie sensorielle spécifique (la voie somesthésique) est indiquée en flèches ponctuées (schéma supérieur). A, La formation réticulaire activatrice ascendante et ses projections corticales, via les noyaux intralaminaires du thalamus. B, Le réseau neuronal impliqué dans les processus attentionnels. C, L'aire pariétale postérieure PG du singe et ses connexions cortico-corticales. Des études anatomiques récentes, utilisant le transport de la péroxydase, ont montré l'importance de cette petite aire, étroitement connectée avec diverses régions également impliquées dans la régulation des processus d'attention sélective (points noirs). Les principales connexions de l'aire PG se font ainsi avec le cortex prérontal, le sillon temporal supérieur et, sur la face interne (non représentée), avec le cortex cingulaire. Toutes ces structures formeraient un réseau cortical spécialisé dans les processus d'attention sélective (d'après Mesulam, 1983)
En conclusion, vers la substance réticulée converge le flux des stimulations venues des espaces corporels et
extracorporels. Il en résulte une activation qui est répercutée à l’ensemble des structures cérébrales par des voies ascendantes. L’éveil et le comportement -présence au monde du sujet- dépendent donc de l’activité de la substance réticulée, véritable source de la spontanéité. Plus il y a de stimulations qui parviennent à la réticulée, plus le sujet est éveillé et plus il est ouvert au monde, donc stimulé, sorte de réaction en chaîne qui se déroulera à l’envers au moment de l’endormissement, résultat d’un désamorçage progressif de la source d’activation. Les comportements sont multiples mais l’activation de la réticulée serait unique et la source générale du véritable désir comportemental.
Le système réticulé activateur est indispensable aux processus attentionnels. Son action aurait pour but de
contrôler: o le niveau de vigilance par son rôle dans l'alternance veille-sommeil (tableau 1 et fig. 3); o de régler un «tonus» attentionnel même lorsque l'état d'éveil reste constant.
Tableau 1 – Mécanismes cellulaires de l'alternance veille- sommeil.
208
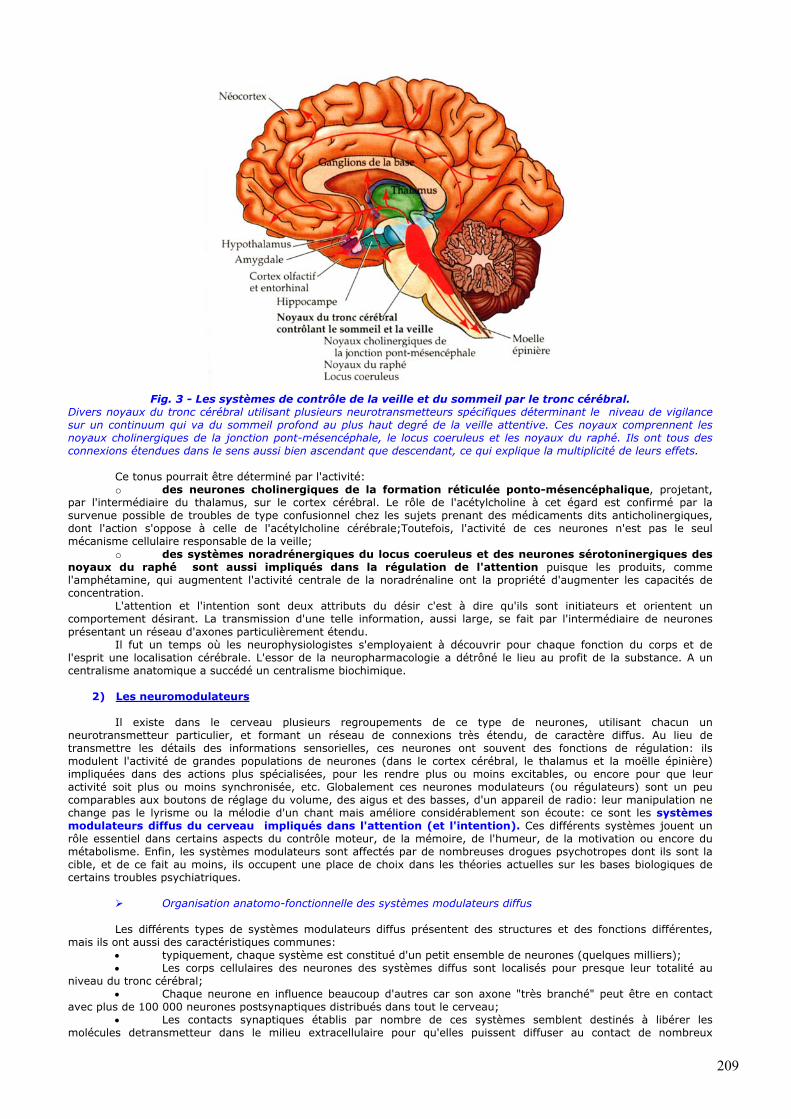
Fig. 3 - Les systèmes de contrôle de la veille et du sommeil par le tronc cérébral.
Divers noyaux du tronc cérébral utilisant plusieurs neurotransmetteurs spécifiques déterminant le niveau de vigilance sur un continuum qui va du sommeil profond au plus haut degré de la veille attentive. Ces noyaux comprennent les noyaux cholinergiques de la jonction pont-mésencéphale, le locus coeruleus et les noyaux du raphé. Ils ont tous des connexions étendues dans le sens aussi bien ascendant que descendant, ce qui explique la multiplicité de leurs effets.
Ce tonus pourrait être déterminé par l'activité: o des neurones cholinergiques de la formation réticulée ponto-mésencéphalique, projetant,
par l'intermédiaire du thalamus, sur le cortex cérébral. Le rôle de l'acétylcholine à cet égard est confirmé par la survenue possible de troubles de type confusionnel chez les sujets prenant des médicaments dits anticholinergiques, dont l'action s'oppose à celle de l'acétylcholine cérébrale;Toutefois, l'activité de ces neurones n'est pas le seul mécanisme cellulaire responsable de la veille;
o des systèmes noradrénergiques du locus coeruleus et des neurones sérotoninergiques des noyaux du raphé sont aussi impliqués dans la régulation de l'attention puisque les produits, comme l'amphétamine, qui augmentent l'activité centrale de la noradrénaline ont la propriété d'augmenter les capacités de concentration.
L'attention et l'intention sont deux attributs du désir c'est à dire qu'ils sont initiateurs et orientent un comportement désirant. La transmission d'une telle information, aussi large, se fait par l'intermédiaire de neurones présentant un réseau d'axones particulièrement étendu.
Il fut un temps où les neurophysiologistes s'employaient à découvrir pour chaque fonction du corps et de l'esprit une localisation cérébrale. L'essor de la neuropharmacologie a détrôné le lieu au profit de la substance. A un centralisme anatomique a succédé un centralisme biochimique.
2) Les neuromodulateurs
Il existe dans le cerveau plusieurs regroupements de ce type de neurones, utilisant chacun un
neurotransmetteur particulier, et formant un réseau de connexions très étendu, de caractère diffus. Au lieu de transmettre les détails des informations sensorielles, ces neurones ont souvent des fonctions de régulation: ils modulent l'activité de grandes populations de neurones (dans le cortex cérébral, le thalamus et la moëlle épinière) impliquées dans des actions plus spécialisées, pour les rendre plus ou moins excitables, ou encore pour que leur activité soit plus ou moins synchronisée, etc. Globalement ces neurones modulateurs (ou régulateurs) sont un peu comparables aux boutons de réglage du volume, des aigus et des basses, d'un appareil de radio: leur manipulation ne change pas le lyrisme ou la mélodie d'un chant mais améliore considérablement son écoute: ce sont les systèmes modulateurs diffus du cerveau impliqués dans l'attention (et l'intention). Ces différents systèmes jouent un rôle essentiel dans certains aspects du contrôle moteur, de la mémoire, de l'humeur, de la motivation ou encore du métabolisme. Enfin, les systèmes modulateurs sont affectés par de nombreuses drogues psychotropes dont ils sont la cible, et de ce fait au moins, ils occupent une place de choix dans les théories actuelles sur les bases biologiques de certains troubles psychiatriques.
Organisation anatomo-fonctionnelle des systèmes modulateurs diffus
Les différents types de systèmes modulateurs diffus présentent des structures et des fonctions différentes,
mais ils ont aussi des caractéristiques communes: • typiquement, chaque système est constitué d'un petit ensemble de neurones (quelques milliers); • Les corps cellulaires des neurones des systèmes diffus sont localisés pour presque leur totalité au
niveau du tronc cérébral; • Chaque neurone en influence beaucoup d'autres car son axone "très branché" peut être en contact
avec plus de 100 000 neurones postsynaptiques distribués dans tout le cerveau; • Les contacts synaptiques établis par nombre de ces systèmes semblent destinés à libérer les
molécules detransmetteur dans le milieu extracellulaire pour qu'elles puissent diffuser au contact de nombreux
209
neurones, plutôt que d'agir dans le voisinage de la fente de la synaptique. Ainsi ces contacts synaptiques ne présentent-ils pas, dans leur vaste majorité, des profils ultrastructuraux de synapses classiques.
Les principaux systèmes modulateurs du cerveau sont associés aux neurotransmetteurs suivants: la noradrénaline (NA), la sérotonine (5-HT), la dopamine (DA) ou l'acétylcholine (Ach). Tous ces neurotransmetteurs, au niveau cérébral, activent pour l'essentiel des récepteurs métabotropiques spécifiques (couplés aux protéines G).
Le rôle exact de ces systèmes sur le comportement n'est aujourd'hui encore pas connu avec précision. Seules quelques généralités peuvent être avancées les concernant.
• Voie cholinergiques du cerveau antérieur basal et du tronc cérébral (fig. 4).
L'acétylcholine est le neurotransmetteur reconnu de la jonction neuromusculaire, des synapses dans les
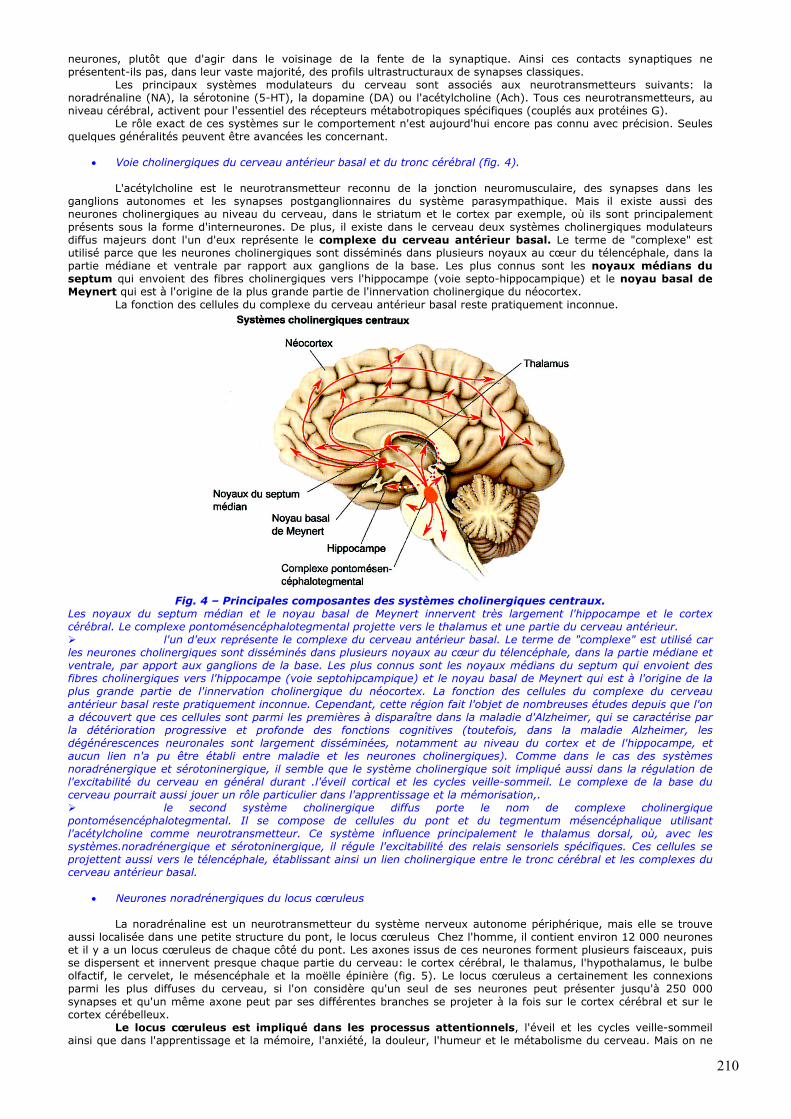
ganglions autonomes et les synapses postganglionnaires du système parasympathique. Mais il existe aussi des neurones cholinergiques au niveau du cerveau, dans le striatum et le cortex par exemple, où ils sont principalement présents sous la forme d'interneurones. De plus, il existe dans le cerveau deux systèmes cholinergiques modulateurs diffus majeurs dont l'un d'eux représente le complexe du cerveau antérieur basal. Le terme de "complexe" est utilisé parce que les neurones cholinergiques sont disséminés dans plusieurs noyaux au cœur du télencéphale, dans la partie médiane et ventrale par rapport aux ganglions de la base. Les plus connus sont les noyaux médians du septum qui envoient des fibres cholinergiques vers l'hippocampe (voie septo-hippocampique) et le noyau basal de Meynert qui est à l'origine de la plus grande partie de l'innervation cholinergique du néocortex.
La fonction des cellules du complexe du cerveau antérieur basal reste pratiquement inconnue.
Fig. 4 – Principales composantes des systèmes cholinergiques centraux.
Les noyaux du septum médian et le noyau basal de Meynert innervent très largement l'hippocampe et le cortex cérébral. Le complexe pontomésencéphalotegmental projette vers le thalamus et une partie du cerveau antérieur. l'un d'eux représente le complexe du cerveau antérieur basal. Le terme de "complexe" est utilisé car
les neurones cholinergiques sont disséminés dans plusieurs noyaux au cœur du télencéphale, dans la partie médiane et ventrale, par apport aux ganglions de la base. Les plus connus sont les noyaux médians du septum qui envoient des fibres cholinergiques vers l'hippocampe (voie septohipcampique) et le noyau basal de Meynert qui est à l'origine de la plus grande partie de l'innervation cholinergique du néocortex. La fonction des cellules du complexe du cerveau antérieur basal reste pratiquement inconnue. Cependant, cette région fait l'objet de nombreuses études depuis que l'on a découvert que ces cellules sont parmi les premières à disparaître dans la maladie d'Alzheimer, qui se caractérise par la détérioration progressive et profonde des fonctions cognitives (toutefois, dans la maladie Alzheimer, les dégénérescences neuronales sont largement disséminées, notamment au niveau du cortex et de l'hippocampe, et aucun lien n'a pu être établi entre maladie et les neurones cholinergiques). Comme dans le cas des systèmes noradrénergique et sérotoninergique, il semble que le système cholinergique soit impliqué aussi dans la régulation de l'excitabilité du cerveau en général durant .l'éveil cortical et les cycles veille-sommeil. Le complexe de la base du cerveau pourrait aussi jouer un rôle particulier dans l'apprentissage et la mémorisation,. le second système cholinergique diffus porte le nom de complexe cholinergique
pontomésencéphalotegmental. Il se compose de cellules du pont et du tegmentum mésencéphalique utilisant l'acétylcholine comme neurotransmetteur. Ce système influence principalement le thalamus dorsal, où, avec les systèmes.noradrénergique et sérotoninergique, il régule l'excitabilité des relais sensoriels spécifiques. Ces cellules se projettent aussi vers le télencéphale, établissant ainsi un lien cholinergique entre le tronc cérébral et les complexes du cerveau antérieur basal.
• Neurones noradrénergiques du locus cœruleus La noradrénaline est un neurotransmetteur du système nerveux autonome périphérique, mais elle se trouve
aussi localisée dans une petite structure du pont, le locus cœruleus Chez l'homme, il contient environ 12 000 neurones et il y a un locus cœruleus de chaque côté du pont. Les axones issus de ces neurones forment plusieurs faisceaux, puis se dispersent et innervent presque chaque partie du cerveau: le cortex cérébral, le thalamus, l'hypothalamus, le bulbe olfactif, le cervelet, le mésencéphale et la moëlle épinière (fig. 5). Le locus cœruleus a certainement les connexions parmi les plus diffuses du cerveau, si l'on considère qu'un seul de ses neurones peut présenter jusqu'à 250 000 synapses et qu'un même axone peut par ses différentes branches se projeter à la fois sur le cortex cérébral et sur le cortex cérébelleux.
Le locus cœruleus est impliqué dans les processus attentionnels, l'éveil et les cycles veille-sommeil ainsi que dans l'apprentissage et la mémoire, l'anxiété, la douleur, l'humeur et le métabolisme du cerveau. Mais on ne
210
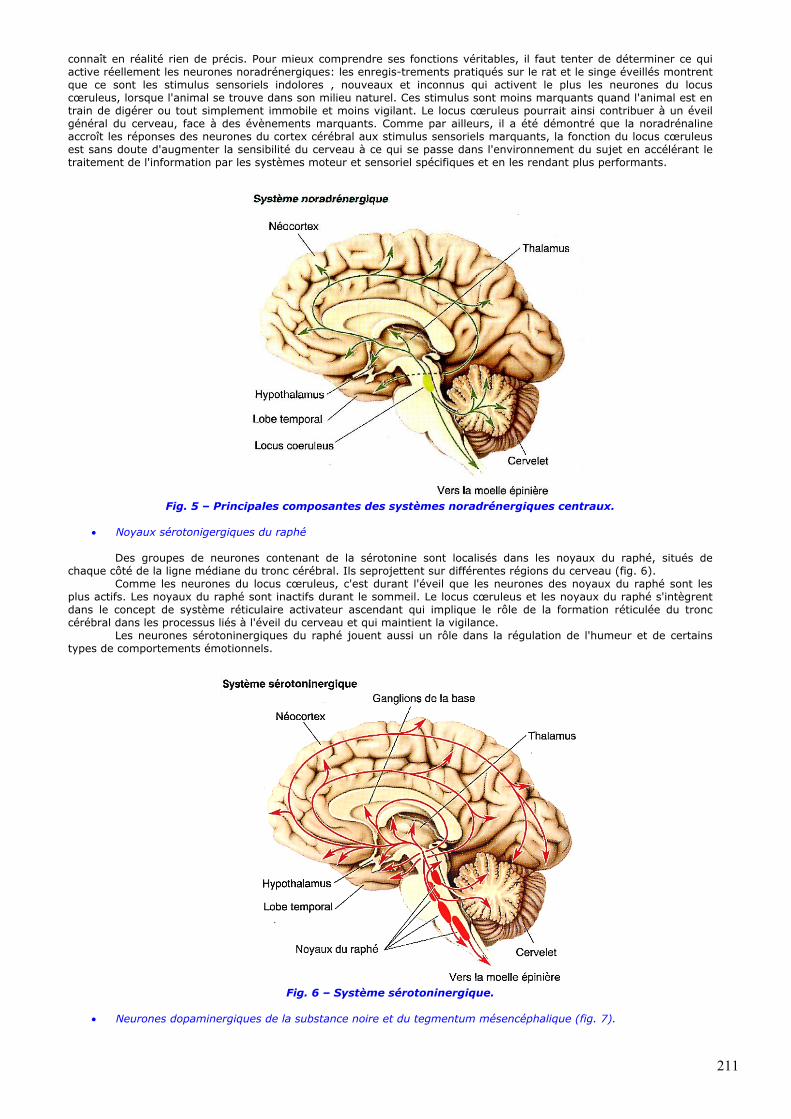
connaît en réalité rien de précis. Pour mieux comprendre ses fonctions véritables, il faut tenter de déterminer ce qui active réellement les neurones noradrénergiques: les enregis-trements pratiqués sur le rat et le singe éveillés montrent que ce sont les stimulus sensoriels indolores , nouveaux et inconnus qui activent le plus les neurones du locus cœruleus, lorsque l'animal se trouve dans son milieu naturel. Ces stimulus sont moins marquants quand l'animal est en train de digérer ou tout simplement immobile et moins vigilant. Le locus cœruleus pourrait ainsi contribuer à un éveil général du cerveau, face à des évènements marquants. Comme par ailleurs, il a été démontré que la noradrénaline accroît les réponses des neurones du cortex cérébral aux stimulus sensoriels marquants, la fonction du locus cœruleus est sans doute d'augmenter la sensibilité du cerveau à ce qui se passe dans l'environnement du sujet en accélérant le traitement de l'information par les systèmes moteur et sensoriel spécifiques et en les rendant plus performants.
Fig. 5 – Principales composantes des systèmes noradrénergiques centraux.
• Noyaux sérotonigergiques du raphé Des groupes de neurones contenant de la sérotonine sont localisés dans les noyaux du raphé, situés de
chaque côté de la ligne médiane du tronc cérébral. Ils seprojettent sur différentes régions du cerveau (fig. 6). Comme les neurones du locus cœruleus, c'est durant l'éveil que les neurones des noyaux du raphé sont les
plus actifs. Les noyaux du raphé sont inactifs durant le sommeil. Le locus cœruleus et les noyaux du raphé s'intègrent dans le concept de système réticulaire activateur ascendant qui implique le rôle de la formation réticulée du tronc cérébral dans les processus liés à l'éveil du cerveau et qui maintient la vigilance.
Les neurones sérotoninergiques du raphé jouent aussi un rôle dans la régulation de l'humeur et de certains types de comportements émotionnels.
Fig. 6 – Système sérotoninergique.
• Neurones dopaminergiques de la substance noire et du tegmentum mésencéphalique (fig. 7).
211
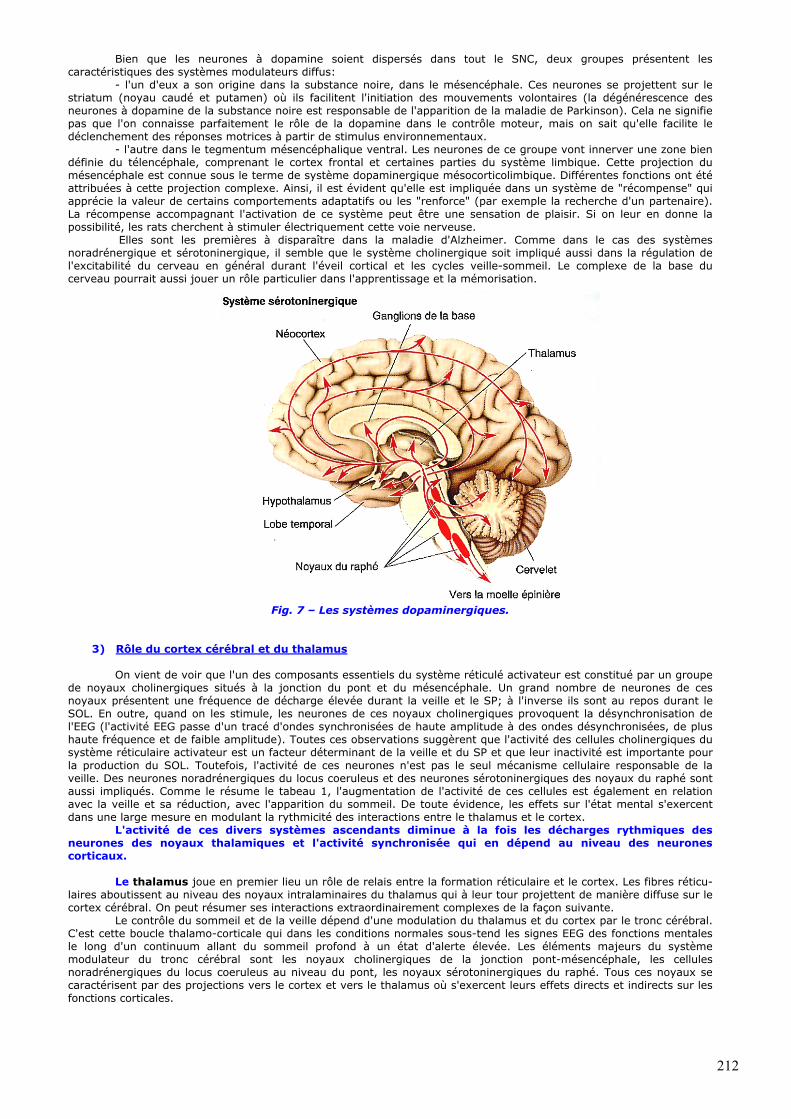
Bien que les neurones à dopamine soient dispersés dans tout le SNC, deux groupes présentent les caractéristiques des systèmes modulateurs diffus:
- l'un d'eux a son origine dans la substance noire, dans le mésencéphale. Ces neurones se projettent sur le striatum (noyau caudé et putamen) où ils facilitent l'initiation des mouvements volontaires (la dégénérescence des neurones à dopamine de la substance noire est responsable de l'apparition de la maladie de Parkinson). Cela ne signifie pas que l'on connaisse parfaitement le rôle de la dopamine dans le contrôle moteur, mais on sait qu'elle facilite le déclenchement des réponses motrices à partir de stimulus environnementaux.
- l'autre dans le tegmentum mésencéphalique ventral. Les neurones de ce groupe vont innerver une zone bien définie du télencéphale, comprenant le cortex frontal et certaines parties du système limbique. Cette projection du mésencéphale est connue sous le terme de système dopaminergique mésocorticolimbique. Différentes fonctions ont été attribuées à cette projection complexe. Ainsi, il est évident qu'elle est impliquée dans un système de "récompense" qui apprécie la valeur de certains comportements adaptatifs ou les "renforce" (par exemple la recherche d'un partenaire). La récompense accompagnant l'activation de ce système peut être une sensation de plaisir. Si on leur en donne la possibilité, les rats cherchent à stimuler électriquement cette voie nerveuse.
Elles sont les premières à disparaître dans la maladie d'Alzheimer. Comme dans le cas des systèmes noradrénergique et sérotoninergique, il semble que le système cholinergique soit impliqué aussi dans la régulation de l'excitabilité du cerveau en général durant l'éveil cortical et les cycles veille-sommeil. Le complexe de la base du cerveau pourrait aussi jouer un rôle particulier dans l'apprentissage et la mémorisation.
Fig. 7 – Les systèmes dopaminergiques.
3) Rôle du cortex cérébral et du thalamus
On vient de voir que l'un des composants essentiels du système réticulé activateur est constitué par un groupe
de noyaux cholinergiques situés à la jonction du pont et du mésencéphale. Un grand nombre de neurones de ces noyaux présentent une fréquence de décharge élevée durant la veille et le SP; à l'inverse ils sont au repos durant le SOL. En outre, quand on les stimule, les neurones de ces noyaux cholinergiques provoquent la désynchronisation de l'EEG (l'activité EEG passe d'un tracé d'ondes synchronisées de haute amplitude à des ondes désynchronisées, de plus haute fréquence et de faible amplitude). Toutes ces observations suggèrent que l'activité des cellules cholinergiques du système réticulaire activateur est un facteur déterminant de la veille et du SP et que leur inactivité est importante pour la production du SOL. Toutefois, l'activité de ces neurones n'est pas le seul mécanisme cellulaire responsable de la veille. Des neurones noradrénergiques du locus coeruleus et des neurones sérotoninergiques des noyaux du raphé sont aussi impliqués. Comme le résume le tabeau 1, l'augmentation de l'activité de ces cellules est également en relation avec la veille et sa réduction, avec l'apparition du sommeil. De toute évidence, les effets sur l'état mental s'exercent dans une large mesure en modulant la rythmicité des interactions entre le thalamus et le cortex.
L'activité de ces divers systèmes ascendants diminue à la fois les décharges rythmiques des neurones des noyaux thalamiques et l'activité synchronisée qui en dépend au niveau des neurones corticaux.
Le thalamus joue en premier lieu un rôle de relais entre la formation réticulaire et le cortex. Les fibres réticu-
laires aboutissent au niveau des noyaux intralaminaires du thalamus qui à leur tour projettent de manière diffuse sur le cortex cérébral. On peut résumer ses interactions extraordinairement complexes de la façon suivante.
Le contrôle du sommeil et de la veille dépend d'une modulation du thalamus et du cortex par le tronc cérébral. C'est cette boucle thalamo-corticale qui dans les conditions normales sous-tend les signes EEG des fonctions mentales le long d'un continuum allant du sommeil profond à un état d'alerte élevée. Les éléments majeurs du système modulateur du tronc cérébral sont les noyaux cholinergiques de la jonction pont-mésencéphale, les cellules noradrénergiques du locus coeruleus au niveau du pont, les noyaux sérotoninergiques du raphé. Tous ces noyaux se caractérisent par des projections vers le cortex et vers le thalamus où s'exercent leurs effets directs et indirects sur les fonctions corticales.
212

4) Cortex cérébral associatif (voir les chapitres suivants) La majeure partie du cerveau humain est dévolue à des tâches qui dépassent le codage des sensations
primaires ou la commande des actes moteurs. Les aires associatives des lobes pariétaux, temporaux et frontaux représentent à peu près 75% de la totalité du tissu cérébral. et prennent en charge les fonctions cognitives, c'est à dire le traitement des informations effectuées entre les entrées sensorielles et les sorties motrices. Littéralement ces fonctions cognitives désignent les processus par lesquels il nous est permis de connaître le monde.
⇒
⇒
L'apport de l'imagerie médicale Cette façon de voir est étayée par l'apport de techniques d'imagerie, telles que la tomographie par émission de
positons (TEP), utilisées au cours de tâches spécifiques d'attention chez des sujets normaux. Ainsi, le débit sanguin augmente dans le cortex pariétal droit durant des tâches exigeant une attention sélective à des stimulus visuels présentés dans le champ visuel gauche pour distinguer leur forme, leur couleur ou leur vitesse. Si la même tâche est à exécuter dans le champ visuel droit, il y a activation conjointe des cortex pariétaux droit et gauche. De plus, si l'on demande aux sujets de garder un haut niveau de vigilance, on observe, en plus de l'augmentation d'activité du cortex pariétal droit, une augmentation dans le cortex frontal droit. Ces recherches laissent envisager que d'autres régions que le lobe pariétal contribuent dans une certaine mesure aux comportements d'attention et peut-être aux syndromes de négligence. En tout état de cause, les cartes métaboliques confirment le fait que la négligence controlatérale provient typiquement d'une lésion pariétale droite et corroborent l'hypothèse d'une certaine spécialisation hémisphérique pour l'attention, au même titre que pour un certain nombre d'autres fonctions cognitives.
En conclusion, depuis une vingtaine d'année, l'apport des techniques d'imagerie cérébrale utilisée au cours de tâches spécifiques d'attention chez des sujets normaux a révélé que le syndrome d'héminégligence peut provenir de lésions siégeant dans une des trois régions corticales suivantes:
o Le gyrus angulaire du lobe pariétal; o Le gyrus cingulaire du lobe pariétal; o Le cortex frontal prémoteur. Ces 3 régions représentent donc des centres cruciaux d'un réseau cortical considéré comme le substrat
anatomique des processus attentionnels. Chez l'homme, ce réseau est organisé de manière asymétrique entre les hémisphères puisque, contrairement au singe, le syndrome d'héminégligence survient presque exclusivement après une lésion droite, ce qui traduit une dominance de l'hémisphère droit dans l'attention
Les apports de l'électrophysiologie: les "neurones attentionnels" du cortex pariétal
La complexité des fonctions cognitives et la difficulté de procéder, chez l'homme, à des études au niveau cellulaire a rendu difficile l'analyse des mécanismes mis en oeuvre par les aires corticales associatives de l'homme. Néanmoins, on a pu réaliser un certain nombre d'observations instructives sur des primates infra-humains et particulièrement sur le singe macaque (rhésus). Ces animaux ont des aptitudes cognitives qui font intervenir les cortex pariétaux, frontaux et temporaux et qui, à bien des égards, sont semblables à celles de l'homme. Il est possible de réaliser, chez le singe éveillé, des enregistrements électrophysiologiques de neurones isolés, au cours de divers comportements; on peut dès lors examiner les réponses physiologiques de cellules individuelles des cortex associatifs pendant l'exécution de tâches cognitives. Des études de ce genre portant sur le cortex pariétal ont tiré bénéfice du fait que les mouvement oculaires sont d'excellents indicateurs des comportements attentifs des primates. On peut ainsi utiliser la fixation des yeux sur une cible intéressante pour identifier, dans ce territoire cortical, les neurones sensibles à l'attention.
Ces travaux ont essentiellement été pratiqués chez le singe et se sont centrés sur une aire du lobule pariétal inférieur du singe dite aire PG (gyrus angulaire). Tout d'abord, des enregistrements cellulaires unitaires ont montré que les cellules de l'aire PG répondent à des événements sensoriels, mais sans spécificité pour une modalité donnée. En outre, cette réponse n'a lieu que si l'événement a une signification en termes de motivation (par exemple la vue d'un aliment chez un animal à jeun). Enfin, il existe une composante motrice au rôle de ces cellules dans la mesure où leur activité précède toujours un mouvement destiné à atteindre la cible («reaching»).
5) Les réseaux de l'attention
Ces données, de même que celles obtenues plus récemment à l'aide de la caméra à positons (TEP), ont amené
M. Posner à proposer une formulation originale des rapports entre attention et cerveau, et, plus hypothétiquement, entre attention et conscience. Pour cet auteur, les 3 régions cérébrales impliquées dans les processus attentionnels ont chacune un rôle distinct:
• le "réseau attentionnel postérieur", qui comprend les aires pariétales postérieures (gyrus angulaire), les noyaux réticulaires et pulvinarien du thalamus, et des parties du colliculus supérieur, est essentiellement un réseau d' «orientation», permet- tant de diriger l'attention vers les endroits pertinents, comme lors de la recherche visuelle. Il permettrait également, tel un zoom, de sélectionner l'échelle pertinente pour examiner l'influx visuel;
• le "réseau attentionnel antérieur" comprend essentiellement le gyrus cingulaire antérieur et l'aire prémotrice. Ces deux aires adjacentes sont activées lors de situations impliquant la détection d'événements. Une des situations, quel que peu artificielle, où ce système serait particulièrement sollicité est le test de Stroop, très largement utilisé en clinique neuropsychologique, qui consiste à imposer au sujet une consigne conflictuelle (en l'occurrence dénommer la couleur de mots imprimés dans des couleurs différentes, par exemple ('BLEU' écrit en vert, 'JAUNE' écrit en rouge);
• enfin, un troisième réseau dit "de vigilance" se situe dans la partie latérale du lobe frontal droit et son activité serait issue des voies noradrénergiques originaires du locus coeruleus. Le type de tâche impliquant l'utilisation de ce système est typiquement la situation où le sujet doit porter son attention vers une source de signaux en même temps qu'il attend l'apparition d'une cible peu fréquente. Il existerait une relation inverse entre l'activité dans le réseau antérieur et dans le réseau de vigilance. En effet, une tâche au cours de laquelle le sujet doit attendre des signaux de faible probabilité implique une suspension de toute autre activité attentionnelle, ce qui est possible grâce à l'action inhibitrice du système de vigilance sur le système attentionnel antérieur, action démontrée par les études en TEP.
213

Ces relations complexes entre les divers réseaux attentionnels pose la question passionnante, mais bien loin d'être résolue, des liens entre attention et conscience. La situation de la « cocktail party » est souvent prise comme exemple des caractéristiques de l'attention humaine. Dans cette situation, lors qu'on discute avec quelqu'un, il est plus difficile de suivre une conversation en raison d'un « parasitage » provenant des conversations voisines. Toutefois, les mécanismes de l'attention sélective permettent d'ignorer ce que disent les autres. Pour tant, on peut avoir conscience de bribes du contenu.
Des données anatomiques sont venues confirmer ces données fonctionnelles en montrant que l'aire PG reçoit des afférences provenant exclusivement d'aires associatives polymodales, elles mêmes en connexion avec les aires associatives unimodales, et de structures limbiques, en particulier le gyrus cingulaire. L'aire PG apparaît donc comme une région fonctionnellement et anatomiquement unique recevant des systèmes sensoriels une information déjà traitée et ne comportant plus la connotation de sa modalité d'origine, d'où le terme d'aire supramodale qui lui est attribué. Elle reçoit en outre une information motivationnelle par ses afférences limbiques et une activation réticulaire par le système formation réticulaire - noyaux intra-laminaires du thalamus. L'aire PG se projette ensuite elle-même sur un certain nombre de structures, en particulier la partie du cortex frontal (champ frontal oculaire) contrôlant la motricité oculaire et le colliculus supérieur, également impliqué dans l'activité oculo-motrice. Ces connexions efférentes de l'aire PG suggèrent qu'elle possède un rôle déterminant dans la modulation de l'exploration visuelle, des comportements d'orientation de la tête et des yeux et probablement aussi des comportements de « reaching » manuel.
6) Comas et pertes de connaissance
Hormis l'état de sommeil, la perte de la faculté d'interagir avec l'environnement est considérée comme un
trouble de conscience. L'expression «perdre conscience» ou «perdre connaissance» fait référence à une perturbation, en général brutale, du contact avec l'environnement. La circonstance la plus fréquente en est le "malaise vaso-vagal" ou état syncopal, banal évanouissement, où le sujet décrit une sensation de "tête vide" ou de vertige, parfois accompagnée d'une expérience corporelle de type "vide épigastrique" ou "sueurs". La cause en est habituellement une chute de la tension artérielle systémique, souvent due à une fragilité des systèmes neurovégétatifs régulateurs de la circulation sanguine qui provoque, à la suite d'une stimulation douloureuse ou émotionnelle, une baisse du débit sanguin cérébral diminuant temporairement l'irrigation de structures nécessaires au maintien de la conscience. Plus rarement, une syncope de ce type peut être due à un trouble cardiaque primitif. Très habituellement, le sujet reprend conscience dans les quelques minutes suivantes, ce qui traduit la normalisation de son débit sanguin.
Le coma est une circonstance beaucoup plus dramatique en raison de son caractère souvent persistant voire irréversible et de la gravité des lésions cérébrales qui en sont responsables. On distingue le coma de la "stupeur", qui en constitue une forme atténuée en individualisant 3 principales causes de comas:
o ceux dus à une lésion de structures du tronc cérébral critiques dans la fonction d'éveil; o ceux dus à une lésion des hémisphères cérébraux; o ceux dus à un trouble métabolique, comme l'hypoglycémie, qui déprime de façon globale la fonction
cérébrale. Plus généralement, on distingue plusieurs grades de coma depuis la simple obnubilation, où le sujet présente
une tendance à l'endormissement, proche de la somnolence des hypersomniaques, jusqu'au coma profond où les stimulations, même les plus intenses (comme celles qui provoquent normalement une douleur violente), ne modifient pas l'état de conscience (à en juger; du moins, par les réponses du sujet). L'étude de la topographie des lésions responsables de ces états est donc la source principale de nos connaissances de 1' «anatomie de la conscience».
⇒
⇒
Rôle de la formation réticulée, du thalamus et de l'hypothalamus Les lésions responsables de perte de conscience sont celles intéressant une aire mésencéphalo-
diencéphalique du cerveau allant depuis la partie supérieure du pont jusqu'à la partie basale du télencéphale. Une région critique, cependant, est la partie moyenne du mésencéphale, au niveau des tubercules quadrijumeaux, région où une lésion, même de petite taille, est susceptible de provoquer un coma. Depuis la fin des années 40, en particulier grâce aux travaux de Magoun et Moruzzi, l'on connaît le rôle déterminant de la formation réticulée paramédiane dans l'éveil cortical. La formation réticulée, qui s'étale de la partie rostrale du pont à la région paramédiane du mésencéphale, se prolonge au-dessus sous la forme de 2 voies, l'une pénétrant dans le thalamus, plus précisément dans sa substance centrale dite "noyaux intra-laminaires", l'autre parcourant l'hypothalamus latéral.
Le rôle des noyaux intra-laminaires du thalamus est suspecté de longue date, comme étant l'intermédiaire
indispensable aux projections issues de la formation réticulée et projetant de façon diffuse sur le cortex cérébral (projections dites non-spécifiques, par opposition à la projection d'autres noyaux thalamiques ou extra-thalamiques sur des régions spécifiques du cortex). La stimulation de cette région du thalamus est ainsi capable de provoquer une activité recrutant tout le cortex à l'électro-encéphalogramme, ce qui confirme le rôle de ces fibres de passage dans l'éveil cortical.
En fait, plus souvent que des lésions thalamiques, des lésions de l'hypothalamus postérieur, respectant le tronc cérébral et le thalamus, semblent, elles, susceptibles de provoquer le coma, ce qui suggère l'importance, encore mal connue, de cette zone dans l'éveil et la vigilance.
Rôle de régions plus caudales du tronc cérébral
Le rôle de régions plus caudales du tronc cérébral est moins clair. Le fait que ces régions contiennent des
structures vitales, indispensables à la régulation de fonctions telles que la respiration, le rythme cardiaque, la tension artérielle, explique que des lésions situées à ce niveau compromettent à court terme la survie du patient. Toutefois, l'observation de sujets survivant à de telles lésions montre que la conscience elle-même peut être totalement préservée. Tel est le cas de certains états consécutifs à des lésions gravissimes de la partie basse du pont (hémorragie ou ramollissement protubérantiels) où le sujet présente tous les signes extérieurs de coma (absence de mouvements spontanés des membres ou du visage, aucun signe de communication avec l'extérieur) mais l'EEG montre un tracé compatible avec un état de veille normale (désynchronisation), parfois partiellement réactif aux stimulations, parfois aréactif réalisant un rythme alpha permanent ou "alpha coma". En fait, tout porte à croire que, dans ces circonstances
214

particulièrement dramatiques, la conscience de ces sujets est strictement normale mais qu'ils sont seulement privés de tous moyens de l'exprimer (états dits de dé-efférentation ou "locked-in syndrome"). Fait intéressant, ces sujets semblent souvent totalement privés de sommeil (asomnie), ce qui corrobore les données de l'expérimentation animale suggérant un centre dans le tronc cérébral caudal (noyaux du raphé et noyau du faisceau solitaire) responsable de la production active du sommeil.
⇒ Troubles de la conscience autres que le coma Hormis lors de lésions, seules envisagées jusqu'ici, de parties basales du cerveau, des perturbations de
certains aspects de la conscience peuvent se rencontrer lors de lésions cérébrales plus diffuses ou touchant des structures plus proches du néocortex.
III – LES THEORIES DE LA MOTIVATION (VOIR CHAPITRE SUIVANT)
Il existe tout un vocabulaire de la motivation. "Bien avant que les psychologues ne tentent d'expliquer le pourquoi de nos actes de façon scientifique, le
langage courant avait développé un nombre important de termes désignant les divers aspects ainsi que les nuances intervenant dans l'expression de la motivation. À partir des définitions comme celles que fournit un dictionnaire, il est possible d'évaluer à quelle dimension les différents termes font référence et, partant, de les utiliser à bon escient.
Il existe tout d'abord des termes généraux. La motivation est l'ensemble des facteurs qui déterminent le comportement. Ce concept rend compte de la
relation entre un acte et les raisons qui l'expliquent ou le justifient. L'envie est associée au fait qu'un besoin est ressenti, qu'il soit d'ordre biologique et lié à la survie de
l'organisme, ou d'ordre purement intellectuel ou esthétique. Le motif est la raison qu'un organisme a d'agir. En général, le motif est d'ordre intellectuel. Le mobile constitue la raison ou le but pour lequel un acte a été effectué. Il ne peut donc être identifié
qu'après la manifestation du comportement. Certains termes font référence plus directement aux aspects physiologiques de la motivation. Le besoin constitue un état de manque, une insuffisance ou l'absence d'un élément nécessaire à l'équilibre de
l'organisme. Bien qu'il s'agisse le plus souvent d'une nécessité d'ordre biologique, ce terme peut également désigner une exigence. La tendance est ce qui amène l'individu à être ou à se comporter d'une façon déterminée (ce mot traduit, en français, le terme drive utilisé par les Anglo-Saxons).
La pulsion fait intervenir une dimension dynamique pour traduire certaines tendances instinctives ou innées, déclenchées après l'apparition du besoin, qui «poussent» l'individu à agir.
Quant à l'attirance, elle laisse sous-entendre l'existence d'un incitateur; En ce sens, elle rend compte d'une force qui attire vers quelqu'un ou vers quelque chose.
La dimension cognitive de la motivation n'est pas non plus oubliée. Les termes qui s'y rapportent mettent l'accent sur l'aspect raisonné ou réfléchi de certains comportements.
Le désir constitue la prise de conscience d'une tendance vers un objet connu ou imaginaire Dans un sens plus restrictif, il compose la base normale du comportement sexuel.
L'aspiration consiste en un élan, un mouvement vers un but donné, qui est le plus souvent d'un niveau élevé.
L'inclinaison fait intervenir la dimension affective. Il s'agit du goût pour un objet ou pour une fin donnée. L'intention concerne le fait de simplement se proposer ou de se fixer un certain but." (Les fondements de la
Psychologie, Godefroid, éd. Vigot, 1993). Il faut distinguer les motivations primaires, ou pulsions biologiques, nécessaires au bon
fonctionnement de l'organisme, des besoins n'ayant plus que de lointains rapports avec la survie. Trois tentatives d'explication les plus appropriées à la majorité des comportements humains.
1) La théorie des pulsions biologiques La faim, la soif, le besoin d'oxygène ou le contrôle de la température du corps sont des besoins primaires dont
la satisfaction est essentielle à la survie de tous les animaux. Tout déséquilibre en sucre, en eau, en oxygène ou en un quelconque élément de base nécessaire à l'organisme entraîne automatiquement un état de besoin et l'apparition d'une pulsion biologique qui "pousse" l'individu à combler ce besoin (fig. 3).
L’état pulsionnel primaire va alors engendrer une série d’activités coordonnées en vue du rétablissement de l’équilibre.
On appelle homéostasie cet état d’équilibre dans lequel se trouve l’organisme en l’absence de tout besoin à satisfaire. Un comportement homéostasique est donc un comportement visant à réduire un état pulsionnel en satisfaisant le besoin qui l’a provoqué.
La théorie des pulsions constitue donc une théorie de la motivation simple et directe permettant de justifier la façon dont les besoins biologiques sont satisfaits. Elle est cependant loin d’expliquer tous les types de motivations humaines. Comment comprendre par exemple qu’une personne même rassasiée se laisse encore tenter par un morceau de gâteau alléchant, ou qu’elle continue à boire, au cours d’une soirée, bien qu’elle ait depuis longtemps étanché sa soif. Il semble évident que, dans certains cas, la stimulation générée par certains objets du monde environnant tient un rôle d’incitateur qui peut être aussi important que l’état pulsionnel lui-même.
2) La théorie de l’activation optimale Cette théorie postule que l’organisme tenterait de maintenir constant un niveau optimal d’activation. Ce niveau ne correspondrait pas à zéro (comme c’est le cas dans la théorie pulsionnelle) mais serait relatif à
l’état physiologique d’une personne donnée à un moment donné. Certains individus auraient ainsi besoin d’une quantité plus importante de stimulations que d’autres qui n’en toléreraient qu’un nombre réduit. Ce besoin de stimulation varierait également avec l’état de conscience de la personne.
215

3) Les théories cognitives de la motivation ; le désir Les deux théories précédentes sont considérées comme trop grossières pour expliquer la diversité des
comportements humains. En effet la plupart du temps nous sommes toujours en train de faire quelque chose et la plupart du temps nous avons choisi de le faire. Même si, quand on étudie les comportements, on parle de motivation, ce terme est peut-être valable pour le rat de laboratoire qui passe sa vie "professionnelle" à appuyer sur des leviers et à franchir des labyrinthes, mais il est impropre à désigner les conduites de l’animal et celles de l’homme dans leur milieu naturel respectif. C’est pourquoi, concernant l’espèce humaine, on préfère parler de "désir" plutôt que de "motivation" pour désigner l'état sous-jacent aux passions ("comportement désirant est préféré à "comportement élémentaire" ou "passion" pour les comportements alimentaire, dipsique, et sexuel). Tandis que la motivation suppose l'acte, le désir désigne un état interne, une tendance vécue par le sujet sans le conduire nécessairement à l'action. Le désir se situe entre la jouissance et le besoin, le profit et la perte. La satisfaction d'un besoin conduit au renforcement, base des théories de l'apprentissage.
Le désir est d'abord un désir de récompense. Une forme de récompense est l'obtention d'un plaisir.
Le désir serait donc défini par le but à atteindre et justifié par la récompense, profit ou plaisir, obtenue. (voir plus loin la théorie hédoniste). Les fonctions cognitives de l’homme lui permettent d’anticiper sur le plaisir lié à la satisfaction d’un désir et de différencier le désir de l’instinct. Le désir serait donc défini par le but à atteindre et justifié par la récompense, profit ou plaisir, obtenue. Le physiologiste pourrait mesurer l’intensité du désir par celle de l’acte qui le sanctionne.
Pour le psychophysiologiste, le terme de motivation évoque deux notions complémentaires : • celle de motilité ou motricité, c’est à dire de production d’une énergie plus ou moins spécifique
nécessaire au déclenchement et à l’entretien d’une séquence de comportement ; • celle de motif d’action qui englobe à la fois -les processus par lesquels une signification motivante
ou démotivante (respectivement appétitive ou aversive) est conféré à tel stimulus- et ceux grâce auxquels cette signification sera ensuite évaluée par référence aux traces laissées par l’expérience passée (mémorisée).
• Une autre composante du désir est le besoin. Le besoin est ressenti comme une situation intolérable qu'il
faut faire cesser. Cet état interne que les psychologues appellent motivation provoque une tendance impérieuse (drive) à réaliser l'acte qui le soulagera. Mais plus que le besoin, c'est peut-être le manque, anticipation ou simulation du besoin, qui est à l'œuvre dans le désir et le place dans la durée. L'animal privé de nourriture pendant plusieurs heures se met en quête d'aliments susceptibles de soulager l'état désagréable (aversif) développé en lui par le jeûne; bien plus il apprend rapidement tout comportement capable de lui fournir l'aliment qui l'apaise. Lorsque l'animal mange, la tendance de faim se trouve réduite, ce qui réduit le besoin de nourriture. Les actes qui rendent la nourriture accessible sont renforcés, c'est à dire qu'ils sont plus susceptibles à se produire dans des situations semblables au cours de la privation de nourriture. L'école béhavioriste américaine a largement développé cette théorie (reduction drive theory ou réduction de tendance) qui lie l'apprentissage aux besoins. Un besoin est défini comme suit : "Quand un des produits ou une des conditions nécessaires à la survie de l'individu ou de l'espèce sont manquants, ou quand ils s'écartent naturellement d'un optimum, on dit qu'il existe un besoin primaire". Cette théorie de l'école béhavioriste -souvent dénigrée- a le mérite de relier les actions à l'état interne du sujet.
Pour l'école freudienne, le désir ne serait qu'une réactivation mnésique d'une perception anticipative d'un
plaisir déjà vécu. Ce serait le cas pour le désir sexuel. C'est le besoin qui serait imaginé et non son assouvissement qui serait à l'origine du désir amoureux: ce besoin n'est qu'une réactivation incessante du désir à travers la perte simulée de l'objet aimé. Mais pas plus qu'un jeûne sexuel prolongé ne peut être tenu pour responsable d'une passion amoureuse, une chute des matières énergétiques disponibles dans le sang ne suffit à expliquer que l'individu passe à table; un signal visuel ou olfactif ou simplement une habitude horaire peuvent réactualiser un déficit primaire virtuel ou déclencher le désir. Mais le désir n'est pas non plus que cela, une attente du plaisir. Il n'aurait sinon aucune raison de cesser, et le caractère événementiel d'un comportement ne pourrait s'expliquer.
En résumé, un état motivationnel représente une forme d’activation comportementale qui est fonction de
l’état présent et passé d’un individu : son action est ainsi tendue vers un objet-but signifiant qui le satisfait. Cependant en cas d’inadéquation entre le but recherché (la satisfaction d’un désir) et le résultat (le plaisir) ou encore si les exigences de la situation dépassent les potentialités du sujet, une émotion ressentie positivement ou négativement peut surgir comme si une énergie se diffusait dans l’organisme troublant ainsi aussi bien les régulations organiques que l’idéation (= enchaînement des idées) du sujet.
Le concept de motivation -que l’on parle de ressort interne, d’impulsion ou de raison d’agir en un mot du « pourquoi ? » du comportement- est une notion malaisée à cerner en raison du poids particulier que l’on accorde aux différents facteurs énergisants le comportement (le moteur), mais aussi en raison des significations fort diverses qui sont données (le motif) :
• pour les béhavioristes, tout comportement est conditionné exclusivement par ses conséquences renforçantes et il n’est pas nécessaire de connaître les mécanismes cérébraux pour prévoir un comportement donné. Pour eux, l’étude des motivations est plutôt celle des stimulations sensorielles motivantes sans aucun recours à la physiologie ;
• pour les existentialistes ou phénoménologues, les motivations sont chargées d’intentionnalité à ce point subtile qu’elles échappent presque par définition à l’analyse du biologiste ;
• quant aux neurophysiologistes, ils s’élèvent contre ces deux façons d’évacuer le cerveau de l’étude des motivations et le restituent dans un schéma stimulus-organisme-réponse. En effet, ce n’est pas parce que l’on peut prévoir le comportement d’un animal soumis à une stimulation sensorielle donnée que l’on a compris les mécanismes cérébraux qui permettent à l’individu de réagir de la sorte. Le neurophysiologiste s’efforce de définir l’état de motivation comme la probabilité de réponse à une stimulation en tenant compte de l’état physiologique du cerveau à ce moment là.
• Sur le plan de la psychologie, deux théories s’efforcent actuellement d’expliquer comment les motivations
déclenchent le comportement :
216

217
• la théorie homéostasique, héritée de la position des philosophes stoïciens, renvoie à la « drive reduction theory » des Anglo-Saxons. Elle soutient, comme par exemple dans la faim, que le besoin déclencherait un état de motivation qui fait apparaître un comportement consommatoire réduisant ce besoin et rétablissant un état neutre de non-besoin ou de satiété. Dans un tel contexte, le processus homéostasique est analysable en termes de systèmes de contrôle, dans un dispositif utilisant une valeur de référence pour enclencher ou débrancher le comportement motivé. Or cette théorie basée unique-ment sur la réduction des besoins ne suffit pas à expli-quer un comportement motivé impliquant aussi une dimension de plaisir/déplaisir;
• la théorie hédonistique, inspirée de la philosophie épicurienne, soutient que le comportement serait déterminé par la recherche du plaisir. Cette théorie s’appuie sur la mise en évidence récente dans le système nerveux central d’un système évaluatif de renforcement positif-négatif.
L’intérêt de ces deux positions théoriques -qui, loin de s’exclure, devraient contribuer à une explication globale de la motivation- sera apprécié par l’études de divers comportements motivés : les comportements d’autostimu-lation, alimentaire, dipsique et sexuel, dans lesquels les composantes homéostasiques et hédonistiques apparaîtront plus clairement, s’exerçant en outre dans une dialectique continue entre inné et acquis. D’ailleurs, la manière dont justement interagissent nature et culture chez l’être vivant constitue un des nœuds de la problématique contemporaine en biologie. Au stade actuel des connaissances, on peut admettre que ce qui est déterminé génétiquement dans un comportement répondant à un besoin inné, c’est avant tout une certaine probabilité d’apparition des apprentissages auxquels l’être vivant peut être soumis avec succès.
En fin sur le plan de la neurophysiologie, le comportement motivé résulterait de trois ensembles de
processus ayant des effets énergisants complémentaires : o une augmentation générale de la réactivité du système nerveux central, par le jeu
d’interactions entre l’hypothalamus latéral et la formation réticulée sous-tendant l’attention vigile ; o une facilitation sélective de la transmission sensorielle, entre autres olfactive, telles que
certaines stimulations «adéquates» (nourriture, partenaire sexuel) sont privilégiées selon le niveau de besoin ; o une modulation plus ou moins spécifique des effets renforçants (positifs ou négatifs) du
stimulus suivant l’état du milieu intérieur, ce qui induit chez le sujet humain des phénomènes d’alliesthésie (modifications ou altérations des sensations).





























