Embed Size (px)
Citation preview

Annts Limnol. 18 (3) 1982 : 217-262.
CONDIT IONS DE DEVELOPPEMENT DU PHYTOPLANCTON. ETUDE COMPARATIVE
DU COMPORTEMENT DE HUIT ESPECES EN CULTURE.
I. DETERMINATION DES PARAMETRES DE C R O I S S A N C E EN FONCTION DE LA LUMIERE ET DE LA TEMPERATURE
par A. D A U T A 1
La croissance de huit espèces d'algues du phytoplancton d'eau douce (6 Chlo-rophycées, 1 Diatomée, 1 Cyanophycée) a été étudiée en culture. Plusieurs méthodes d'évaluation de la biomasse ont été comparées : poids sec, comptages cellulaires, densité optique, chlorophylle. Selon les espèces, l'extraction de pigments par l'acétone donne des valeurs inférieures de 5 à 50 % par rapport à l'éthanol. S'il n'a a pas de carence en nutriments, une assez bonne approximation du taux de croissance peut être calculée à partir des paramètres acquis par des expériences de photosynthèse (Pmax). Les variations de taille des cellules au cours du cycle de division sont également décrites. Le taux de croissance des huit espèces est décrit en fonction de la température et de la lumière selon un système d'équations inspirées de la formulation de Steele (1965). Les paramètres obtenus permettent de bien modéliser l'évolution d'une culture en tenant compte du phénomène d'ombre propre.
Conditions for phytoplankton development, a comparative study of the behaviour of eight species in culture.
I. Determination of growth parameters in relation to light and temperature.
The growth of eight algal species from freshwater phytoplankton (6 Chloro-phyceae, 1 Diatom, 1 Cyanophyceae) has been studied in culture. Several methods of biomass evaluation have been compared: dry weight, cell counts, optical density, chlorophyll. Amongst the species, the extraction of pigments by acetone provides estimates that are lower 5 - 50 % compared with ethanol. If there is not a lack of nutrients, a close approximation of growth rate can foe calculated beginning with the parameters acquired from the processes of photosynthesis (Pmax)' Variations in the size of cells during their division cycle are likewise described. The growth rate of eight species is described as a function of temperature and light, and according to a basic equation based on Hhat of Steele (1965). The parameters obtained can be used to model the development of a culture, taking into account the phenomenon of self-shading.
1. Laboratoire d'Hydrobiologie, ERA 702 du CNRS, Université Paul-Sabatier, 118, route de Narbonne, 31062 Toulouse Cedex.
Ce travail a été accompli dans le cadre d'une action concertée DGRST (convention n° 78 7 2543) et de l'ASP PIREN CNRS « Ressources en Eau ». Nous remercions M"° Mur qui a effectué les analyses chimiques.
Article available at http://www.limnology-journal.org or http://dx.doi.org/10.1051/limn/1982005

218 A. DAUTA (2)
I. — INTRODUCTION
« L i m n o l o g i s t s h a v e adressée! t h e m s e l v e s for m o r e t h a n fifty y e a r s
t o t h e d i s t r i b u t i o n of p h y t o p l a n k t o n a n d t h e i r b i o m a s s in r e l a t i o n t o
the e n v i r o n m e n t f a c t o r s in l a k e s of di f ferent t rophy . A l t h o u g h th i s
ac t iv i ty h a s r e s u l t e d in a n e n o r m o u s a n d st i l l rap id ly g r o w i n g l i t téra
ture , p r o g r e s s in u n d e r s t a n d i n g a n d p r e d i c t i o n h a s b e i n g v e r y s l o w . »
Cet te c i t a t i o n e m p r u n t é e à Kalff e t K n o e c h e l (1978) p r e n d e n c o r e p l u s d e p o i d s si l 'on y j o i n t u n e ref lex ion de C h u (1942) :
« V a r i o u s f a c t o r s h a v e b e e n t a k e n i n t o c o n s i d e r a t i o n t o e x p l a i n t h e
p e r i o d i c i t y a n d d i s t r i b u t i o n of p h y t o p l a n k t o n d u r i n g t h e l a s t 50 y e a r s -
N u m e r o u s a t t e m p s h a v e b e e n m a d e t o c o r r e l a t e t h e o c c u r e n c e of
p l a n k t o n i c o r g a n i s m s w i t h o n e o r o t h e r f a c t o r , b u t al l h a v e m e t w i t h
s e r i o u s c r i t i c i s m in o n e r e s p e c t of a n o t h e r . »
Ce r a c c o u r c i d e p r e s q u e 100 a n s de r e c h e r c h e s d o n t d é c o u l e u n e
a b o n d a n t e b i b l i o g r a p h i e , i l l u s t r e :
— l ' intérêt m a n i f e s t é p a r les c h e r c h e u r s vis-à-vis d u p h y t o p l a n c t o n
— l e s p r o b l è m e s d ' in t erpré ta t ion q u e s o u l è v e n t l a d i s t r i b u t i o n , l'évo
l u t i o n e t la s u c c e s s i o n d e s a l g u e s d a n s l e s m i l i e u x n a t u r e l s
— l e s diff icultés d e m o d é l i s a t i o n e t d e p r é d i c t i o n re la t ive s a u x déve
l o p p e m e n t d e s a l g u e s .
E n s e m b l e h é t é r o g è n e d 'a lgues m i c r o s c o p i q u e s d o n t la ta i l le var ie
e n t r e 1 e t 250 ¡jura, l e p h y t o p l a n c t o n j o u e u n r o l e d a n s le m é t a b o l i s m e
d e s l a c s e t r iv i ère s r é g u l é e s . S o n d é v e l o p p e m e n t d é p e n d à c h a q u e ins
t a n t d e s f a c t e u r s c l i m a t i q u e s ( é o l a i r e m e n t , t e m p é r a t u r e ) qu i contrô
lent l 'act iv i té m é t a b o l i q u e d e s a l g u e s , d e la q u a n t i t é d ' é l é m e n t s nutri
t i f s d i s p o n i b l e s e t d e s f a c t e u r s b i o t i q u e s : n a t u r e d e s e s p è c e s , pré
dat ion , c o m p é t i t i o n ( C a p b l a n c q 1982).
L e s t ravaux re la t i f s a u p h y t o p l a n c t o n o n t c o n n u d e p u i s l 'ouvrage d e r é f é r e n c e d e H u t c h i n s o n (1967) u n d é v e l o p p e m e n t c o n s i d é r a b l e . C e p e n d a n t l e s p r e m i è r e s o b s e r v a t i o n s r e l a t i v e s à l ' é c a p h y s i o l o g i e d e s a l g u e s f u r e n t r é a l i s é e s p a r Zachar ias (1894) sur la v i t e s s e d e c h u t e d e s filaments d e Melosira. W h i p p l e (1895) p r o d u i t l e p r e m i e r travai l i m p o r t a n t s u r le ro l e d e s n u t r i m e n t s , d e la t e m p é r a t u r e e t d e la l u m i è r e sur le t aux d e c r o i s s a n c e d e s m i c r o o r g a n i s m e s , t a n d i s q u e l a p r e m i è r e i n t e r p r é t a t i o n d u c y c l e a n n u e l b a s é e s u r la t e m p é r a t u r e et la l u m i è r e e s t d é v e l o p p é e e n 1943 p a r F i n d e n e g g .
Avant 1950, la p l u p a r t d e s s u j e t s d ' intérêt a c t u e l s f u r e n t abor
d é s :
— a s s i m i l a t i o n de l 'azote p a r l e s C y a n o p h y c é e s : la p r e m i è r e publ i c a t i o n s u r c e p h é n o m è n e r e v i e n t à D r e w e s (1928), su iv i p a r A l l i s o n

(3) PARAMÈTRES DE CROISSANCE 219
e t M o r r i s (1930) ; d e n o m b r e u x c h e r c h e u r s d o n t F o g g (1942) e t T h a n
T u n (1949) p o u r s u i v r o n t c e s r e c h e r c h e s
—• a s s i m i l a t i o n d e n u t r i m e n t s t e l s a z o t e e t p h o s p h o r e , é t u d e s d e
c r o i s s a n c e s u r - d e s m i l i e u x d e c u l t u r e v a r i é s : K e t c h u m (1939) , H a r v e y
(1940) , C h u (1942, 1943), W u r t z (1949)
— p h y s i o l o g i e d e la n u t r i t i o n d e s C h l o r o p h y c é e s : A l g e u s (1946)
— a s s i m i l a t i o n d e d i v e r s e s s o u r c e s d e o a r b o n e : O s t e r l i n d (1947,
1948, 1949)
—• f lo t tabi l i té e t v i t e s s e d e s é d i m e n t a t i o n d e s c e l l u l e s : K l e b a h n (1895) , U t e r m o h l (1925) , R u t t n e r (1930) , Fr i tz (1935) , E i n s e l e e t G r i m m (1938), G r o s s e t Z e u t h e n (1948)
— q u o t a s ce l lu la i re s d e D i a t o m é e s et d e C y a n o p h y c é e s : E i n s e l e e t
G r i m m (1938) , G r i m m (1939)
— i n f l u e n c e d e la t e m p é r a t u r e s u r la c r o i s s a n c e d e s a l g u e s : R u t t n e r
( 1 9 3 7 ) ; L u n d (1949). R h o d e (1948) p r o p o s e u n e é t u d e e x p é r i m e n t a l e
d e la t e m p é r a t u r e s u r la b a s e d'un c o n c e p t p h y s i o l o g i q u e m o d e r n e .
L'analyse d e s t r a v a u x c o n c e r n a n t les 30 d e r n i è r e s m o n t r e n t ne t te
m e n t q u ' u n e ré -or i en ta t ion d e s a x e s d e r e c h e r c h e s s 'effectue d a n s
l e s a n n é e s s o i x a n t e .
—• Il s'agit d 'une p a r t d e l 'u t i l i sa t ion d e s m o d è l e s e n é c o l o g i e .
La p h o t o s y n t h è s e e n e s t l e p r e m i e r t h è m e ; l e b u t e s t d e m o d é l i s e r
l ' in tégrat ion d e la p r o d u c t i o n p r i m a i r e , so i t d a n s u n e t r a n c h e d'eau,
so i t s u r u n j o u r { T a l l i n g 1955, 1957, V o l l e n w e i d e r 1960 , S t e e l e 1965,
V o l l e n w e i d e r 1965).
Le d e u x i è m e t y p e d e m o d è l e e s t i n d u i t p a r l e s p r o b l è m e s d'eutro-p h i s a t i o n e t la n é c e s s i t é d e p r é d i r e l ' é v o l u t i o n à l o n g t e r m e d e l 'état t r o p h i q u e d'un lac , e n r é p o n s e à u n e m o d i f i c a t i o n d e la c h a r g e a n n u e l l e e n p h o s p h o r e . Les p r e m i e r s m o d è l e s s o n t p r o p o s é s p a r Bre-z o n i k (1968) e t V o l l e n w e i d e r (1968) , c e d e r n i e r u t i l i s a n t u n e formulat ion c o m p o r t a n t u n e s e u l e é q u a t i o n , s y s t è m e r e p r i s p a r D i l i o n e t Rig ler (1974) . D e p u i s , l 'u t i l i sa t ion d e s m o d è l e s d e s i m u l a t i o n s 'est c o n s i d é r a b l e m e n t é t e n d u e . C o m p t e t e n u d u c a r a c t è r e n o n exhaus t i f de m e s r e c h e r c h e s b i b l i o g r a p h i q u e s , l e s t ro i s ch i f fres s u i v a n t s s o n t t rè s s ignif icat i fs d e c e t t e p r o g r e s s i o n d a n s l 'u t i l i sa t ion d e s m o d è l e s :
p é r i o d e n o m b r e d e m o d è l e s p a r u s
( t o u s m o d è l e s c o n f o n d u s )
D u p r e m i e r m o d è l e , p r o p o s é
p a r F l e m i n g (1939) à 1970 1 9 7 0 — 1974 1975 — 1979
20
29
91
R e m a r q u e : d e 1939 à 1970, i l s 'agit e s s e n t i e l l e m e n t d e m o d è l e s
m a t h é m a t i q u e s , e t r e c e n s é s p a r P a t t e n (1968) .

220 A. DAUTA (4)
Cet te é v o l u t i o n e s t l i é e d e t o u t e é v i d e n c e , a u d é v e l o p p e m e n t d e s
o r d i n a t e u r s , e t à l eur u t i l i s a t i o n c r o i s s a n t e d a n s d e s d i s c i p l i n e s v a r i é e s
( C h a h u n e a u e t a l . 1980). D e p l u s , l e s s t r u c t u r e s d e s m o d è l e s d e v e n a n t
d e p l u s e n p l u s c o m p l e x e s , la f o r m u l a t i o n s t o c h a s t i q u e a fa i t p l a c e
à u n f o r m a l i s m e m a t h é m a t i q u e à b a s e d ' é q u a t i o n s d i f férent ie l les
i m p o s s i b l e à g é r e r a u t r e m e n t q u e p a r d e s m o d è l e s d e s i m u l a t i o n
(Meyer 1980).
— Le d e u x i è m e a x e d e r e c h e r c h e e s t d 'abord l i é à u n n o u v e a u
c o n c e p t d é v e l o p p é e n 1969 p a r D r o o p : la p r i s e e n c o m p t e d e s r é s e r v e s
in t race l lu la i re s e n n u t r i m e n t s . J u s q u ' a l o r s , l a c r o i s s a n c e d'une popu
la t ion e n f o n c t i o n d e s n u t r i m e n t s é ta i t d é c r i t e s e l o n u n e é q u a t i o n d e
t y p e M o n o d (1949), l iant la c r o i s s a n c e à la c o n c e n t r a t i o n d e l ' é l é m e n t
l imi tant p a r u n e f o n c t i o n h y p e r b o l i q u e . L 'ut i l i sa t ion d e s q u o t a s cel
lu la ires p e r m e t a lor s d e d é c o u p l e r d a n s l e s m o d è l e s les p h é n o m è n e s
d ' a s s i m i l a t i o n e t d e c r o i s s a n c e . Cet te n o u v e l l e a t t i t u d e vis-à-vis d e s
s t o c k s ce l lu la i re s e n n u t r i m e n t s g é n è r e u n e v a g u e d ' é tudes sur l'assi
m i l a t i o n d e s a l g u e s à l 'aide d e t u r b i d o s t a t s ( F u h s 1969, Azad e t Bor-
c h a r d t 1970) o u d e c h e m o s t a t s ( E p p l e y e t al. 1971). C a p e r o n et M e y e r
(1972) , D r o o p (1973) , R h e e (1973) s o n t l e s p r i n c i p a u x a r t i s a n s d e c e t t e
p e t i t e r é v o l u t i o n .
U n e d e s p r i n c i p a l e s p r é o c c u p a t i o n s d e s a n n é e s so ixante -d ix r e s t e
le p h o s p h o r e e t l ' e u t r o p h i s a t i o n ( V o l l e n w e i d e r 1968, B o w e n 1970,
K u e n t z e l 1970, J e n k i n s e t a l 1973, Griffith e t al. 1973). C e c o n s t a t
a m è n e d e n o m b r e u s e s é t u d e s s u r l e s p a r a m è t r e s re la t i f s à l 'ass imi la
t i o n d u p h o s p h o r e d o n t , e n t r e a u t r e s , c e l l e s d e R h e e (1973, 1974),
H e a l e y (1973) , D r o o p (1974) .
C e p e n d a n t l 'u t i l i sa t ion a c c r u e d e s m o d è l e s d e s i m u l a t i o n d e m a n d e d e p l u s e n p l u s d e d o n n é e s re la t ive s à l ' éco log ie d e s a l g u e s . Les s o u r c e s l e s p l u s u t i l i s é e s s o n t e n g r a n d e p a r t i e l e s m e s u r e s d e t erra in , e t quel q u e s v a l e u r s d é t e r m i n é e s e n c u l t u r e s ( S t r i c k l a n d 1958, C a p e r o n e t M e y e r 1972, R h e e 1973, 1974, D r o o p 1974, 1975, K i l h a m 1975) e t s u s c e p t i b l e s d'être m o d é l i s a b l e s . C e r t a i n s m o d è l e s o n t e u x - m ê m e s serv i d e s o u r c e s , à l ' e x e m p l e d e c e l u i p r o p o s é p a r L e h m a n et al. (1975). L'analyse d e s r é f é r e n c e s d e s m o d è l e s m o n t r e a l o r s q u e la p l u p a r t d'entre e u x u t i l i s e n t l e s m ê m e s p a r a m è t r e s , e n p a r t i c u l i e r p o u r la c r o i s s a n c e , l e s r a p p o r t s s t o e c h i o m é t r i q u e s C / N / P , l e s v i t e s s e s d'assim i l a t i o n e t l e s c o n s t a n t e s d e d e m i - s a t u r a t i o n ( K s ) , l e s c o n s t a n t e s de b i o d é g r a d a t i o n . Ces m o d è l e s b a s é s s u r d e s p a r a m è t r e s d 'or ig ines divers e s e t d e q u a l i t é s d i f férentes s o n t r e m i s e n q u e s t i o n par Ta l l ing (1979) qu i m e t l ' accent s u r l ' i n a d é q u a t i o n d e s f o r m u l e s m u l t i p l i c a t i v e s , e t p r é c o n i s e l ' e m p l o i d e p a r a m è t r e s i n t e r d é p e n d a n t s d a n s l e c a d r e de la m o d é l i s a t i o n .
E n o u t r e , l e s m o d é l i s a t e u r s s o n t g é n é r a l e m e n t c o n f r o n t é s a u x dif
ficultés s u i v a n t e s :

(5) PARAMÈTRES DE CROISSANCE 221
A. j , \-1-
7 ^ | Q
''M
,40JJ t V?^'/ $ *
1 —
[E * v r r J ; v ' f
Y g r. f f
i •>„.''- * r * ,
><* * * % m
.[i • » ^ V fr t :
. J*,50JJ f
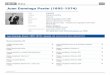
FIG. 1. — Cycle de division sur 2 4 heures, la à ld : Scenedesmus crassus, t = 0, t = 12, t = 19, t = 24 heures, le à lh : Coelastrum microsporum, t = 0, t = 14, t = 19, t = 2 4 heures.

222 A. DAUTA ( 6 )
FlG. 2. — Cycle de division sur 24 heures. 2a à 2 d : Dictyosphaerium pulchellum, t = 0, t = 14, t = 21, t = 24 heures. 2e à 2h : Scenedesmus quadricauda, t = 0, t = 13, t = 20, t = 24 heures.

FIG. 3. — Cycle de division sur 24 heures. 3a et 3b : Fragillaria bidens, t = 0 et t = 20 heures. 3c et 3d : Anabaena cylindrica, t = 0 et t = 21 heures. 3e à 3h : Monoraphidium minutum, t = 0, t = 12, t = 17, t = 23 heures.

FiG. 4. — 4a à 4f : Pediastrum boryanum, cycle de division sur 24 heures, t = 0. t = 13, t = 16, t = 20, t = 24. 4g et 4h : Cultures plurispéciûques.

( 9 ) PARAMÈTRES DE CROISSANCE 225
— réa l i s er u n m o d è l e d e s t i n é à ê t r e vérif ié sur d e s d o n n é e s n'ayant
p a s é t é c o l l e c t é e s d a n s l e b u t d 'une m o d é l i s a t i o n ,
— s o u s - i n f o r m a t i o n re la t ive a u x v a r i a b l e s d e forçage , u t i l i s a t i o n d e
v a l e u r s a p p r o x i m a t i v e s t i r é e s d e la b i b l i o g r a p h i e p o u r l e s p a r a m è t r e s
( t a u x d e c r o i s s a n c e , v i t e s s e s d 'a s s imi la t ion , q u o t a s , t a u x d e p h o t o s y n
t h è s e , e t c . ) .
C e r t a i n s p a r a m è t r e s p e u v e n t ê t r e é v a l u é s , d i r e c t e m e n t o u ind irec t e m e n t , p a r m e s u r e s s u r l e t erra in (ex . t aux d 'ex t inc t ion d e la l u m i è r e ) . D 'autres , é v a l u é s d 'après l e s e x p é r i e n c e s e n l a b o r a t o i r e ( taux d e c r o i s s a n c e , K s ) , n e s o n t p a s f o r c é m e n t r e p r é s e n t a t i f s d e s c o n d i t i o n s d e t erra in ( C h a h u n e a u e t al. 1980).
D a n s n o t r e c a s , u n e m a s s e d e d o n n é e s é ta i t d i s p o n i b l e sur u n
m i l i e u n a t u r e l , l e Lot : D a u t a (1975) , D a u t a e t al . (1975) , M a s s i o (1976) ,
D e n a t (1977) , C a p b l a n c q et T o u r e n q (1978), Ang e l i e r e t al. (1978) ,
D é c a m p s e t C a s a n o v a - B a t u t (1978) , C a p b l a n c q e t D a u t a (1978) .
L e s d e u x p r e m i e r s e s s a i s d e m o d é l i s a t i o n ( C a p b l a n c q e t D é c a m p s 1977, C a s a n o v a - B a t u t 1977) m o n t r a i e n t n e t t e m e n t la n é c e s s i t é d 'approf o n d i r l ' é tude e x p é r i m e n t a l e d e s t a u x d e c r o i s s a n c e e t d e s c i n é t i q u e s d ' a s s i m i l a t i o n .
Cet te réf lex ion n o u s a c o n d u i t s à é tud ier , e n l abora to i re , l e s param è t r e s d e c r o i s s a n c e d'une p o p u l a t i o n m o n o s p é c i f i q u e . L ' o r i en ta t i o n d e c e s p r e m i è r e s e x p é r i e n c e s ( G u e r r i e t al. 1981, D a u t a e t al. 1982, Brunei, e t al. 1982) c o n c e r n a i t la m i s e au p o i n t d e s t e c h n i q u e s et d e s m é t h o d e s . Le travai l p r é s e n t é ic i c o m p l è t e c e t t e p r e m i è r e é t u d e e t l ' é t end à 7 a u t r e s e s p è c e s d 'a lgues d u p h y t o p l a n c t o n d'eau d o u c e .
2 . — MATERIEL ET METHODES
2 . 1 . Mode de culture
N o u s a v o n s u t i l i s é le s y s t è m e d e c u l t u r e e n m i l i e u fini ( b a t c h cu l t u r e s ) . Ce m o d e d e c u l t u r e s i m p l e e t f iable d o n t Pr ib i l e t M a r v a n (1979) r a p p e l l e n t les a v a n t a g e s e t i n c o n v é n i e n t s , e s t v a l a b l e a u s s i b i e n p o u r l ' é tude d e la c r o i s s a n c e ( s u r p l u s i e u r s j o u r s ) q u e p o u r ce l l e d e l ' a s s imi la t i on ( q u e l q u e s h e u r e s ) ( K i l h a m 1978). Afin d e p r é v e n i r u n a p p a u v r i s s e m e n t d u m i l i e u e n d i o x i d e d e c a r b o n e , l e s c u l t u r e s s o n t f o r t e m e n t b u l l é e s a v e c d e l'air. Le m i l i e u d e c u l t u r e u t i l i s é ( T a b l e a u I ) e s t c e lu i é l a b o r é p a r B r u n e l - D e l c l a u x e t Guerr i (1980) : e s s e n t i e l l e m e n t m i n é r a l p o u r év i t er t o u t e p r o l i f é r a t i o n d e b a c t é r i e s , il s ' inspire d e s m i l i e u x d e c u l t u r e s p r o p o s é s p a r C h u 1942), R a y b u r n e t S tarr (1974), L e c o h u et G u e n é (1976) . L e s h u i t e s p è c e s y o n t p o u s s é d u r a n t t o u t e la d u r é e d e s é t u d e s ( s o i t 2 à 5 a n s s e l o n l e cas ) s a n s i n c o n v é n i e n t s e t s a n s a p p a r i t i o n d e s i g n e s d e d é g é n é r e s c e n c e . L ' é c l a i r e m e n t e s t d i s -

226 A. DAUTA ( 1 0 )
p e n s é s e l o n u n c y c l e d e 15 h e u r e s d e l u m i è r e p o u r 9 h e u r e s d ' o b s c u r i t é
p a r d e s t u b e s f l u o r e s c e n t s b l a n c s . P o u r d e s i n t e n s i t é s l u m i n e u s e s
s u p é r i e u r e s à 250 juE.m^.s ' 1 , u n e l a m p e d e 400 w a t t s ( t y p e P h y t o -
C l a u d e ) e s t u t i l i s ée . Les m e s u r e s d e l u m i è r e i n c i d e n t e o n t é t é r é a l i s é e s
à l 'aide d 'un q u a n t a m è t r e LI-COR 185 A.
TABLEAU I . — Composition du milieu de culture.
Composés chimiques Concentrations mg/litre
MgS04, 7H20 25 FeS04. 7H20 1
Milieu CaQ12. 2H20 25 de base Na2 E D T A 1
NaHCCB 50 Na2C03 5
Zn804, 7H20 0,020 CuC12, 2H20 0,020
Oligo MnC12, 4H20 0,400 éléments CoC12, 6H20 0,010
Bo3H3 0,001 Na2Mo04 0,035
KN03 200 Nutriments K2HP04 25
Si02 Na205, H20 212
2 .2 . Espèces étudiées
Les h u i t e s p è c e s é t u d i é e s a p p a r t i e n n e n t a u p h y t o p l a n c t o n d e s e a u x
d o u c e s .
Il s 'agit d e 6 C h l o r o p h y c é e s :
Scenedesmus crassus Chod . (fig. 1 a-1 d) Coelastrum microsporum N â g . (fig. 1 e-1 h ) Dictyosphaerium pulchellum W o o d . (fig. 2 a-2 d ) Scenedesmus quadricauda (Turp. ) B r e b . (fig. 2 e-2 h ) Pediastrum boryanum (Turp . ) M e n e g h . (fig. 4 a-4 f) Monoraphidium minutum (Les l i e ) K o m a r k o v a - L e g n e r o v a o u
Selenastrum minutum ( N a g . ) Co l l ins (fig. 3 e-3 h )
d ' u n e D i a t o m é e : Fragillaria bidens H e i b . (fig. 3 a-3 b )
•et d ' u n e ICyanophycée : Anabaena cylindrica L e m m . (fig. 3 c-3 d) .
L e s p h o t o g r a p h i e s 4 g e t 4 h r é a l i s é e s s u r u n e c u l t u r e p lur i spéc i -f ique i l l u s t r e n t l e s d i f f érences d e ta i l l e s e x i s t a n t e n t r e l e s e s p è c e s -é tudiées .
Les s o u c h e s d e Scenedesmus crassus e t d e Coelastrum microsporum
j n ' o n t é t é f o u r n i e s p a r l 'A lgo thèque d u L a b o r a t o i r e d e C r y p t o g a m i e

(11) PARAMÈTRES DE CROISSANCE 227
( M u s é u m d 'Hi s to i re N a t u r e l l e , Par i s ) . L a s o u c h e à'Anabaena cylin-
drica m'a é t é g r a c i e u s e m e n t p r ê t é e p a r C. B r a w e r s ( L a b o r a t o i r e d'Eco
l o g i e v é g é t a l e , U n i v e r s i t é c a t h o l i q u e d e L o u v a i n ) .
L e s a u t r e s s o u c h e s 1 o n t é t é i s o l é e s p a r imes s o i n s à p a r t i r d u m i l i e u
n a t u r e l ( L a c d e V i n ç a , P y r é n é e s - O r i e n t a l e s ) .
P a r m i c e s e s p è c e s , c e r t a i n e s o n t d é j à f a i t l 'obje t d ' é t u d e s é c o p h y -s i o l o g i q u e s a s s e z a p p r o f o n d i e s . Ains i , Scenedesmus quadricauda a é t é p a r t i c u l i è r e m e n t é t u d i é p a r l e s c h e r c h e u r s d e l 'Académie d e s S c i e n c e s T c h é c o s l o v a q u e :
— p h a s e s e t p a r t i c u l a r i t é s d e la d i v i s i o n ce l lu la i re : S e t l i k e t a l . 1972, B e r k o v a e t al. 1972),
— in f luence d e la t e m p é r a t u r e e t d e la l u m i è r e sur la m o r p h o l o g i e
d e s c e l l u l e s ( K o m a r e k e t R u z i c k a 1969, R u z i c k a 1971),
— r e l a t i o n e n t r e c r o i s s a n c e , t e m p é r a t u r e e t s y n c h r o n i s a t i o n ( K o m a
rek e t S i m m e r 1965, D o u c h a e t V e n d l o v a 1970, Z a c h l e d e r e t al. 1975,
D o u c h a 1979),
—• b e s o i n s n u t r i t i o n n e l s e n c u l t u r e s (Pribi l e t M a r v a n 1970 a, 1970 b , 1971 ; B u m b u et M o k r y a k 1973).
Anabaena cylindrica a é t é e t r e s t e e n c o r e le s u p p o r t d e s é t u d e s s u r
l e p h é n o m è n e d e f ixat ion d e l 'azote a t m o s p h é r i q u e , e n t r e a u t r e s :
— p r o c e s s u s d e f ixat ion d e l 'azote ( F o g g 1942, F o g g e t T h a n T u n
1949, C o x et Fay /1969),
— d i f f érenc ia t ion ce l lu la i re ( F a y 1969),
— p h o t o s y n t h è s e e t f ixat ion d'azote ( C o b b et M y e r s 1964),
— p r o d u c t i o n e x t r a c e l l u l a i r e (Fogg 1952, W a l s b y e t F o g g 1975),
— rô le d e s o l i g o - é l é m e n t s d a n s la c r o i s s a n c e (Arnon 1958, W a r d e t
W e t z e l 1975).
Les a u t r e s e s p è c e s o n t é t é p e u é t u d i é e s .
P o u r Scenedesmus crassus, o n p e u t c i t e r l e s r e c h e r c h e s e f f e c t u é e s s u r d e s e s p è c e s v o i s i n e s p a r T r a i n o r (1963), Tra inor e t B u r g (1965 b ) , S h u b e r t e t Tra inor (1974) , M a t h e w et C h o w d a r y (1977) , T e z u k a (1977) e t D o r (1978).
P o u r Coelastrum microporum, il f aut s i g n a l e r l e s é t u d e s d e T r a i n o r
e t B u r g (1965 a ) , G a t e s e t al. (1966) , Y a u e t C h a n (1973), C h a n (1976),
r e s p e c t i v e m e n t s u r la m o b i l i t é d e s c e l l u l e s , l 'u t i l i sa t i on d'un s u b s t r a t
o r g a n i q u e , la r é p o n s e à u n t r a i t e m e n t à l ' a u x i n e e t la f o r m a t i o n d e s
c o l o n i e s .
1. Je remercie MM. P. Bourrelly et R. (Lecohu qui ont bien voulu déterminer ces espèces.

228 A. DAUTA ( 1 2 )
Dictyosphaerium pulchellwn a serv i a u t e s t d'un m i l i e u d e cro i s
s a n c e ( T r a i n o r e t S h u b e r t 1973).
A n o t r e c o n n a i s s a n c e l e s a u t r e s e s p è c e s n 'ont p a s fait l 'obje t d'étu
d e s re la t ive s à l eur p h y s i o l o g i e . C e p e n d a n t , u n e m a s s e c o n s i d é r a b l e
d e m e s u r e s a é t é r é a l i s é e s u r u n e e s p è c e v o i s i n e d e Monoraphidium
minutum : Selenastrum capricornutum, s o u c h e m o n d i a l e m e n t u t i l i s é e
p o u r l e s b io - e s sa i s , d o n t la b i b l i o g r a p h i e a é t é e o l l a t i o n n é e p a r Le i sch-
m a n et a l . (1979) .
D a n s c e t t e b i b l i o g r a p h i e , il n 'est p a s t e n u c o m p t e d u n o m b r e t r è s
i m p o r t a n t d ' é tudes o u d e t e s t s d e t o x i c i t é ayant m i s e n j e u l 'une o u
l 'autre de c e s e s p è c e s c o n s i d é r é e s . %
2 . 3 . Estimation de la biomasse
La c r o i s s a n c e d'une c u l t u r e e s t g é n é r a l e m e n t e x p r i m é e c o m m e l'aug
m e n t a t i o n d'une b i o m a s s e , d'un n o m b r e de c e l l u l e s o u d 'un t a u x d e
c h l o r o p h y l l e p a r u n i t é d e t e m p s . L'éventai l d e s cr i t ère s u t i l i s é s p o u r
e x p r i m e r l a q u a n t i t é d 'a lgues p a r u n i t é d e v o l u m e e n g e n d r e d e s dif
f icul tés d e c o m p a r a i s o n d e d o n n é e s ( V o l l e n w e i d e r 1969). O n p e u t
c o n s i d é r e r ( L u k a v s k y e t al. 1979) q u e l e s m é t h o d e s d 'éva luat ion d e
c r i o s s a n c e p e u v e n t ê t r e r e g r o u p é e s :
—• e n m é t h o d e s d i r e c t e s ( n o m b r e d e c e l l u l e s , p o i d s s ec , b i o m a s s e ) ,
— e n m é t h o d e s i n d i r e c t e s ( d e n s i t é o p t i q u e , p i g m e n t s , p r o t é i n e s ,
A T P , C a r b o n e , Azote , e t c . ) .
Ic i , lie c h o i x d e la d e n s i t é o p t i q u e p o u r é v a l u e r la b i o m a s s e e s t l ié :
— à l ' ex i s t ence de t r è s b o n n e s c o r r é l a t i o n s e n t r e d e n s i t é o p t i q u e e t
d e n s i t é d e c e l l u l e s ,
— à la fac i l i t é d e c o n v e r s i o n d e l a d e n s i t é o p t i q u e , é t a l o n n é e p a r d e s c o m p t a g e s a u m i c r o s c o p e i n v e r s é ( s e l o n la m é t h o d e p r é c o n i s é e p a r U t h e r m o h l 1958).
Ce c h o i x d e la d e n s i t é o p t i q u e e n t a n t q u e p a r a m è t r e d ' e s t i m a t i o n d e b i o m a s s e e s t e n a c c o r d a v e c l ' é tude r é c e n t e d e B u t t e r w i c k e t al. (1982) qu i c l a s s e c e t t e m é t h o d e c o m m e é t a n t la p l u s r a p i d e e t la p l u s p r é c i s e , a p r è s l ' e s t i m a t i o n d e s p i g m e n t s ( e x t r a c t i o n p u i s m e s u r e e n fluorimetrie).
C e p e n d a n t , afin d e fac i l i t er u n e c o m p a r a i s o n a v e c d 'autres u n i t é s • c o u r a m m e n t u t i l i s é e s , l e s c o r r e s p o n d a n c e s a v e c la c h l o r o p h y l l e e t l e p o i d s s e c o n t é t é é t a b l i e s (Tab leau I I ) .
La c h l o r o p h y l l e a é t é é v a l u é e s e l o n la m é t h o d e . S C O R - U N E S C O , en
•comparant l'efficacité d ' e x t r a c t i o n d e d e u x s o l v a n t s : a c é t o n e (équa
t i o n s d 'après G o l t e r m a n 1969) o u é t h a n o l b o u i l l a n t ( é q u a t i o n s d 'après
M a r k e r e t al . 1980).

(13) PARAMÈTRES DE CROISSANCE 229
2
i
M
• a . v XI ^ Z
O N O N O N oq_ O N O N O N ON^
© o* © o" O o" o~ o"
ox ^ o' o- ô-
• ' + + + + + + +
• u-i m m r~ s S S! S 8 S! ¿ £ 2 a) ES »-i
r | IT) t* l * I T H O IH S O ° O f ~ 0 ° 2 S 5 N N p < N
V N . O N O N O N O N O N O N O N
O o O O O O O O
J + + + + + + + +
© © • * . f N CN ~ H
O N O N ' »-H
( S rO N ^ < N M N «
.g o" o" o" o o" o" o" o
^ + + + + + + + + § K 8 8 8 S 8 8 8
*3 t O» fN » f ) CO > «-H O" »-H O*
.5 a
2-5
g s ^
Wl R »» «1 a „ » s
s
§ S ? N „
C •« M K S t • « • s o i .a
y V O (U <J o - s O ,
5 5 •a «

230 A. DAUTA (14)
2 .4 . Estimation du taux de croissance
2.4.1. P R I N C I P E
La l u m i è r e , l a t e m p é r a t u r e e t l e s q u a n t i t é s d ' é l é m e n t s nutr i t i f s dis
p o n i b l e s s o n t l e s v a l e u r s s u s c e p t i b l e s d e d i m i n u e r l e t a u x spéc i f ique
d e c r o i s s a n c e l o r s q u ' e l l e s s ' é lo ignent d e s v a l e u r s o p t i m a l e s . P a r m i les
é l é m e n t s n u t r i t i f s , s o n t r e t e n u s l 'azote , l e p h o s p h o r e e t la s i l i c e , c o n s
t i t u a n t s l i m i t a n t g é n é r a l e m e n t la o r o i s s a n c e d e s c o m m u n a u t é s natu
re l l e s .
L ' équat ion p e r m e t t a n t d e décr i re l ' évo lu t ion d 'une p o p u l a t i o n A
s e l o n le t e m p s t s 'écrit : d A / d t = / iA. Le t a u x d e c r o i s s a n c e p. d e la
p o p u l a t i o n à c h a q u e i n s t a n t p e u t ê t r e c o n s i d é r é c o m m e u n t a u x maxi
m a l d e c r o i s s a n c e (;u,max), af fecté d e f a c t e u r s d e r é d u c t i o n l i é s a u x
v a r i a b l e s L u m i è r e ( I ) , t e m p é r a t u r e (T), e t a u x é l é m e n t s nutr i t i f s :
A z o t e ( N ) , P h o s p h o r e (P. ) . Ic i , la l u m i è r e i n t e r v i e n t d i r e c t e m e n t d a n s
l e c a l c u l d u t a u x d e c r o i s s a n c e ( i l n'y a p a s d e f a c t e u r d e r é d u c t i o n
l i é a u c a r b o n e , l ' e x p r e s s i o n i n c l u t la var ia t ion d u t a u x d e p h o t o s y n
t h è s e qu i a p p a r a î t à p a r t ) .
n = n m a x * R e d I * R e d T * R e d N * R e d P i t e r m e 1 1 1 t e r m e 2 '
o ù l e s s y m b o l e s « * » r e p r é s e n t e n t u n e c o m b i n a i s o n d e s d i f férents
f a c t e u r s .
+ D a n s c e p r e m i e r p a s v e r s la m o d é l i s a t i o n d'une p o p u l a t i o n d'al
g u e s , ¿i e s t c o n s i d é r é c o m m e c o n s t a n t p a r r a p p o r t a u t e m p s . C e c a s
c o r r e s p o n d à d e s c o n d i t i o n s o p t i m a l e s d e c r o i s s a n c e , o u l o r s q u e p.
d é p e n d d e v a r i a b l e s c o n s i d é r é e s e l l e s - m ê m e s c o m m e i n d é p e n d a n t e s du
t e m p s , à l ' e x e m p l e d e s v a r i a b l e s d e f o r ç a g e I e t T. Alors , o n p e u t é c r i r e
q u e p o u r d e u x é v a l u a t i o n s d 'une p o p u l a t i o n A (Al e t A2) r é a l i s é e s a u x
t e m p s t l e t t2 :
j* = L o g ( A 2 / A l ) * ( l / t 2 - t l ) .
S i l 'on a p p e l l e q l e r a p p o r t A 2 / A 1 , p o u r u n in terva l l e d e t e m p s ( t2- t l = 1 j o u r ) , a lor s ju. = L o g (q ) .
Cet te d é m a r c h e a é t é u t i l i s é e p o u r d é c r i r e /x ( I , T) e n c o n d i t i o n s n u t r i t i o n n e l l e s n o n l i m i t a n t e s .
+ D a n s l e c a s d e p o p u l a t i o n s e n m i l i e u fini ( c u l t u r e s e n b a t c h ) , p.
d e v i e n t u n e v a r i a b l e p a r r a p p o r t a u t e m p s , e t d é c r o î t a lors e n f o n c t i o n d e s v a r i a b l e s d é p e n d a n t e s d e l a d e n s i t é de la p o p u l a t i o n ( b a i s s e d e l u m i è r e d i s p o n i b l e p a r l ' o m b r e p r o p r e , é p u i s e m e n t d e s é l é m e n t s nutrit i f s e t v a r i a t i o n s d e s s t o c k s i n t e r n e s ) . L 'ex t inc t ion d e la l u m i è r e incid e n t e p a r l a d e n s i t é d e c e l l u l e s a é t é c a l c u l é e p o u r p o u v o i r p r e n d r e e n c o m p t e l'effet d ' o m b r e p r o p r e .

(15) PARAMÈTRES DE CROISSANCE 231
2.4.2. PROTOCOLE EXPÉRIMENTAL
U n e c u l t u r e m è r e e s t m a i n t e n u e n p h a s e e x p o n e n t i e l l e p a r d e s repi
q u a g e s f r é q u e n t s d a n s u n m i l i e u r i c h e e n n u t r i m e n t s (1 m m o l e K N 0 3 ,
70 jumoles K 2 H P 0 4 , 1 m m o l e S I 0 2 ) , d a n s d e s c o n d i t i o n s s u b - o p t i m a l e s
d e t e m p é r a t u r e e t d e l u m i è r e , e t s y n c h r o n i s é e s e l o n u n c y c l e d e
15 h e u r e s d e l u m i è r e p o u r 9 h e u r e s d 'obscur i t é . Avant l ' e x p é r i e n c e
d e s t i n é e à m e s u r e r /* (I , T ) , l a c u l t u r e e s t p l a c é e 24 h e u r e s à l 'obscu
r i té . A t = 0, so i t a u d é b u t d u c y c l e l u m i n e u x , l a c u l t u r e e s t a l o r s
t r a n s f é r é e e t d i l u é e d a n s u n m i l i e u neuf e n a b a i s s a n t la d e n s i t é opt i
q u e d e f a ç o n à a n n u l e r p r a t i q u e m e n t t o u t effet d ' o m b r e p r o p r e . Ains i
s o n t m i n i m i s é s l e s r i s q u e s é v e n t u e l s d e l i m i t a t i o n p a r l e s n u t r i m e n t s ,
le c a r b o n e o u la l u m i è r e ( M a r v a n e t Pribi l 1979).
P o u r c h a q u e t e m p é r a t u r e , la c u l t u r e d i l u é e , r épar t i e d a n s u n e q u i n
za ine d e f lacons , e s t s o u m i s e à d e s i n t e n s i t é s l u m i n e u s e s c o m p r i s e s
en tre 5 e t 750 / t E . m " 2 . s _ 1 .
La d e n s i t é o p t i q u e e s t e n s u i t e m e s u r é e le l e n d e m a i n ( t l ) e t le surl e n d e m a i n (t2) . E n d é b u t e t e n fin d ' expér i ence , o n t é t é m e s u r é e s la c h l o r o p h y l l e a, la b i o m a s s e s è c h e , l e s c o n c e n t r a t i o n s e n N e t P d u m i l i e u d e c u l t u r e .
2 .5 . Mesures d'assimilation du carbone
Il é ta i t i n t é r e s s a n t d e vérif ier l ' ex i s t ence d 'une c o r r é l a t i o n p e r m e t tant d ' e s t i m e r u n taux d e c r o i s s a n c e p a r u n e m e s u r e d u c a r b o n ass i m i l é , c e qui a u t o r i s e r a i t u n e m é t h o d e p l u s r a p i d e q u e ce l l e c o n s i s t a n t à s u i v r e la d e n s i t é d e c e l l u l e s . Le t a u x d ' a s s i m i l a t i o n d e c a r b o n e e s t e n effet u n e u n i t é u t i l i s é e p a r l e s o c é a n o g r a p h e s e t l e s l i m n o l o g i s t e s , m e s u r e q u i a l 'avantage d ' intégrer l ' inf luence d e s c o n d i t i o n s n u t r i t i v e s , de la l u m i è r e e t d e la t e m p é r a t u r e , m a i s a u s s i l ' i n c o n v é n i e n t d'être s u b o r d o n n é e à u n e lo i d e p r é c o n d i t i o n n e m e n t à la l u m i è r e e t d'être s o u v e n t e x p r i m é e p a r u n i t é d e c h l o r o p h y l l e a.
M a l g r é c e p r o b l è m e , n o u s a v o n s t e n t é d e re l i er la p h o t o s y n t h è s e
brute a u x t a u x d e c r o i s s a n c e m e s u r é s , d a n s l e c a s d e n u t r i m e n t s n o n
l i m i t a n t s .
L e s c u l t u r e s e n p h a s e e x p o n e n t i e l l e s o n t p r é c o n d i t i o n n é e s p e n d a n t 2 j o u r s à d e s l u m i è r e s fa ib l e s (20 Ju,E..m"2.s.). P o u r c e s m e s u r e s , le m i l i e u d e c u l t u r e e s t m o d i f i é p a r u n m é l a n g e a v e c u n e e a u n a t u r e l l e m i n é r a l e , t r è s c h a r g é e e n H C 0 3 — : c e t a j o u t p e r m e t d 'obten ir u n e a l ca l in i t é a s s e z é l e v é e (100 m g C 0 3 C a / I ) , é v i t a n t d e s v a r i a t i o n s t r o p i m p o r t a n t e s d e p H d u r a n t d'expérience e t u n e d i m i n u t i o n p o s s i b l e d e la v i t e s s e d ' a s s i m i l a t i o n p a r u n e b a i s s e t r o p r a p i d e d u s u b s t r a t e n c a r b o n e . I l e s t u t i l e d e s a v o i r q u e l 'eau m i n é r a l e e m p l o y é e (V i t t e l ) n e c o n t i e n t n i N 0 3 — N , n i P 0 4 — P , c e q u i a u t o r i s e s o n e m p l o i p o u r d e s c u l t u r e s c a r e n c é e s .

232 A. DAUTA (16)
A p r è s u n r a j o u t d e 14 C 0 3 N a H (lOjuCi.1" 1), l a c u l t u r e e s t r é p a r t i e
d a n s d e s flacons r o d é s d e 60 m l , e t p l a c é s d a n s u n e e n c e i n t e t h e r
m o s t a t é e c o n ç u e s p é c i a l e m e n t p o u r l e s e x p é r i e n c e s d ' a s s i m i l a t i o n de
c a r b o n e . Cet te e n c e i n t e (décr i t e p a r B r u n e l - D e l c l a u x e t Guerr i 1980,
p r o c h e d u s y s t è m e p r o p o s é p a r F e e 1973) e s t r é g l a b l e e n t e m p é r a t u r e
(10 à 35 °C) e t p e r m e t d ' incuber s i m u l t a n é m e n t u n e g r a n d e q u a n t i t é
d e flacons répar t i s d a n s d e s v a l e u r s d ' in tens i t é s l u m i n e u s e s c o m p r i s e s
e n t r e 0 e t 1 000 / i E . m " 2 . s n .
A p r è s u n e h e u r e d ' incubat ion , u n v o l u m e d e 10 m l d e c h a q u e flacon
e s t filtré s u r filtre e n n i t r a t e d e c e l l u l o s e . La r a d i o a c t i v i t é d u filtre e s t
m e s u r é e e n s c i n t i l l a t i o n l i q u i d e s u r c o m p t e u r à s c i n t i l l a t i o n B e c k m a n
L S 133. U n t é m o i n ( b r u i t d e f o n d ) pr i s à t = 0 e s t d é d u i t d e l a v a l e u r
o b t e n u e . Les r é s u l t a t s d e P m a x s o n t e x p r i m é s e n m g d e C a r b o n e ass i
m i l é / 1 0 9 c e l l u l e s / h e u r e , e t e n m g d e C a r b o n e a s s i m i l é p a r m g Chloro
p h y l l e a / h e u r e .
3. — RESULTATS
3 .1 . Résultats expérimentaux
3.1.1. E X P R E S S I O N DE LA BIOMASSE
L e s s p e c t r e s d ' a b s o r p t i o n e n f o n c t i o n d e la l o n g u e u r d 'onde o n t
é t é é t a b l i s p o u r l e s 8 e s p è c e s (fig. 5) . D a n s t o u s les c a s , l e s c u l t u r e s
o n t d e u x p i c s d ' a b s o r p t i o n d a n s l e s p l a g e s 400-440 n m et 660-700 n m .
P o u r t o u t e s l e s e s p è c e s , l e d e u x i è m e p i c (660-700 n m ) e s t p l u s s t a b l e
e t l e s m e s u r e s p l u s p r o d u c t i b l e s , à l ' e x c e p t i o n de Fragillaria qu i pré
s e n t e u n c o m p o r t e m e n t i n v e r s e . E n c o n s é q u e n c e , l e s d e n s i t é s o p t i q u e s
o n t é t é m e s u r é e s à 680 n m e t 440 n m , r e s p e c t i v e m e n t .
Ces l o n g u e u r s d ' o n d e c o r r e s p o n d e n t p a r a i l l eurs a u x c h o i x fa i t s p a r S o r o k i n ( 1 9 7 3 ) e t C l o e r n ( 1 9 7 6 ) : 678 n m , M a r g u l i e s ( 1 9 7 0 ) : 440 n m , F o y e t G i b s o n (1982) : 436 non, C o n d i t (1972) : 665 n m o u F e u i l l a d e e t al. (1976) : 650 n m .
Les r e l a t i o n s e n t r e d e n s i t é s o p t i q u e s e t n o m b r e d e c e l l u l e s (fig. 6) o n t é t é é t a b l i e s p a r c o m p t a g e s c e l l u l a i r e s (Tab leau I I ) . Le t r a i t e m e n t s t a t i s t i q u e d e s r é s u l t a t s m o n t r e q u e la r e l a t i o n n o m b r e d e ce l lul e s / d e n s i t é o p t i q u e e s t fiable ( p r é c i s i o n 80 %) t a n t q u e la d e n s i t é o p t i q u e n e d é p a s s e p a s 1,3. O n p e u t n o t e r q u e c e t t e i n c e r t i t u d e n'influe p a s s u r l e t aux de c r o i s s a n c e q u i p e u t ê t r e c a l c u l é d i r e c t e m e n t à partir d e s d e n s i t é s o p t i q u e s .
L e p l u s r e m a r q u a b l e (fig. 6) e s t l a var iab i l i t é d e la r e l a t i o n n o m b r e d e c e l l u l e s / d e n s i t é o p t i q u e e n f o n c t i o n d e l 'état n u t r i t i o n n e l ( c u l t u r e s n o n l i m i t é e s o u c a r e n c é e s e n N , P, S i ) e t s e l o n l e s e s p è c e s . Ce p h é n o -

(17) PARAMÈTRES DE CROISSANCE 2 3 3
.9-S .
.7.
densité optique
.6-1
.5
.6
.4.
. 5
.4
. 3
-i 1 1 r
Se .8 .7
6.
densité* optique
.1-
400 ' 600 ' 800 400 600 longueur d'onde en nm
-i r
Sq
-i 1 1 1 1 r
A c
Mm
800
FIG. 5. — Spectres d'absorption des différentes espèces étudiées. (Se) : Scenedesmus crassus; (Cm): Coelastrum microsporum; (Dp): Dictyosphaerium put-chellum ; (Sq) : Scenedesmus quadricauda ; (Ffo) : Fragillaria bidens ; (Ac) : Ana-baena cylindrica ; (Pb) : Pediastrum boryanum ; (Mm) : Monoraphidium minu-tum.
m è n e e s t s i m p l e m e n t l i é à la ta i l l e d e s c e l l u l e s o u a u n o m b r e d e cel
l u l e s c o n s t i t u a n t u n c é n o b e , c a r a c t è r e s v a r i a b l e s s e l o n q u e l ' e spèce e s t
c a r e n c é e o u n o n . C e p e n d a n t , p o u r c e r t a i n e s e s p è c e s (Dictyosphaerium,
Anàbaena, Monoraphidium) i l y a p e u d e v a r i a t i o n s .
P o u r l e s a u t r e s e s p è c e s , l e s l i m i t a t i o n s e n n u t r i m e n t s i n d u i s e n t les.
p h é n o m è n e s s u i v a n t s :
—• Coelastrum : la p r o p o r t i o n d e s c é n o b e s d i m i n u e , l e s c e l l u l e s i so
l é e s d e v i e n n e n t p r é p o n d é r a n t e s ,
— l e s d e u x e s p è c e s d e Scenedesmus : a u l i e u d e c é n o b e s de 4 c e l
lu l e s , i l appara î t d e s c é n o b e s d e 2, 4 c e l l u l e s e t d e s c e l l u l e s i s o l é e s .

234 A. DAUTA (18)
0 1 0 1 0 1 0 1 optique
FIG. 6. — Relations Nombre de Cellules = F (Densité optique) * F (Etat nutri-tionnel).
— Pediastrum : l a ta i l l e d e s c é n o b e s (fig. 4 a-4 f) e t s u r t o u t l e s pro
p o r t i o n s d e c é n o b e s d e 4, 8 e t 16 c e l l u l e s v a r i e n t (en m i l i e u c a r e n c e ,
l e s c u l t u r e s s o n t c o n s t i t u é e s d'un for t p o u r c e n t a g e de c é n o b e s d e 8
o u 16 c e l l u l e s d e f o r t e s d i m e n s i o n s ) .
— Fragillaria : s e u l e la c a r e n c e e n s i l i ce d o n n e d e s v a r i a t i o n s perc e p t i b l e s d e l a f r u s t u l e qu i p e u t a c c u s e r u n e d i m i n u t i o n de ta i l l e d e 20 % p a r r a p p o r t à la n o r m a l e .
Le t a b l e a u II d o n n e l e s r e l a t i o n s e t l e s r a p p o r t s p o i d s s e c e t ch lo r o p h y l l e a / 1 0 9 c e l l u l e s . Ces d e u x d e r n i e r s o n t é t é déf inis à p a r t i r de c u l t u r e s n o n c a r e n c é e s e t p o u r d e s s t a d e s d ivers : c r o i s s a n c e e x p o n e n t ie l le , p h a s e s t a t i o n n a i r e . L' interval le d e c o n f i a n c e d e s r a p p o r t s e s t g é n é r a l e m e n t l e p l u s é l e v é p o u r l e s e s p è c e s p r é s e n t a n t l e s p l u s f o r t e s var ia t i ons d e v o l u m e s a u c o u r s d e la c r o i s s a n c e (cf. § 3.1.2.) ; t a n d i s q u e l e s v a r i a t i o n s d e s t e n e u r s e n c h l o r o p h y l l e e n g l o b e n t d e s c u l t u r e s e x p o s é e s à d e s l u m i è r e s t rè s d i f férentes e n i n t e n s i t é s .
La figure 7 c o m p a r e l'efficacité d 'ex trac t ion d e d e u x s o l v a n t s (étha-n o l , a c é t o n e ) . Le r a p p o r t d 'ex trac t ion e s t t o u j o u r s f a v o r a b l e à l 'étha-n o l , e t var i e s e l o n l e s e s p è c e s . Les c a s l e s p l u s d é f a v o r a b l e s corres p o n d e n t a u x C h l o r o p h y c é e s . Ces r é s u l t a t s m o n t r e n t s a n s a m b i g u ï t é q u e l a m é t h o d e d 'ex trac t ion p a r l ' acé tone s o u s - e s t i m e g é n é r a l e m e n t la t e n e u r e n c h l o r o p h y l l e d 'une p o p u l a t i o n d 'a lgues s i ce l le-c i e s t à b a s e d e C h l o r o p h y c é e s .

(19) PARAMÈTRES DE CROISSANCE 235
Chlorophylle (j jg/l) Extraction-Alcool
Chlorophylle ( u g / l ) Extradion Acétone
FIG. 7. — Comparaison de d'efficacité d'extraction de l'éthanol et de l'acétone.
TABLEAU I I I . — Biovolumes et Quotas en Carbone. — Biovolume en micron 3 , Quotas en mg pour 1 0 9 Cellules.
Espèce Biovolume Quota en Carbone
mini moyen maxi mini moyen maxi Scenedesmus crassus 3 0 100 300 11 17 3 4 Coelastrum microsporum 5 0 80 700 1 0 14 100 Dictyosphaerium pulchellum 65 140 520 17 2 4 85 Scenedesmus quadricauda 80 180 680 2 4 3 0 72 Fragillaria bidens 200 280 560 3 4 41 84 Anabaena cylindrica 10 3 0 62 3 4,8 7,5 Pediastrum boryanum 60 350 900 12 60 144 Monoraphidium minutum 12 21 120 2 3,6 2 0
Le q u o t a ce l lu la i re e n c a r b o n e ( T a b l e a u I I I ) p e u t ê t re e s t i m é à par
tir d u b i o v o l u m e m o y e n ce l lu la i re e n u t i l i s a n t l e s f a c t e u r s de conver
s i o n d o n n é s p a r M u l l i n e t al. (1966) , e t S t r a t h m a n (1967) .
Ces r a p p o r t s C h l o r o p h y l l e / c a r b o n e ( d e 1,87 à 3,16 %) et Chlorop h y l l e / p o i d s s e c (0,38 à 1,44 %) c o r r e s p o n d e n t a u x v a l e u r s c i t é e s p a r P a r s o n s e t al. (1961), Ant ia e t al. (1963) , S l o a n et S t r i c k l a n d (1966) , P a r s o n s (1961) , M e e k s (1974) e t C a p b l a n c q (1982) qu i d o n n e n t u n pourc e n t a g e d e c h l o r o p h y l l e c o m p r i s e n t r e 0,3 e t 2 % d u p o i d s s e c . Le r a p p o r t e n t r e q u o t a m i n i m a l e t m a x i m a l d e c a r b o n e e x p r i m e à la fo i s l e s v a r i a t i o n s d e ta i l l e s d e s c e l l u l e s d 'une m ê m e e s p è c e , e t l e p o u v o i r de d i v i s i o n ( n o m b r e d e c e l l u l e s fi l les p r o d u i t e s à par t i r d 'une c e l l u l e a d u l t e ) . E n fait , c e s r a p p o r t s é v o l u e n t c o n s t a m m e n t d a n s le t e m p s a i n s i q u e l e m o n t r e n t D o u c h a e t V e n d l o v a (1970), S e t l i k e t al. (1972) e t D o u c h a (1979) . S e t l i k (1979) i n d i q u e q u e le t a u x é l evé d e c h l o r o p h y l l e

2 3 6 A. DAUTA ( 2 0 )
p a r u n i t é d e p o i d s s e c , d a n s l e c a s d 'une p o p u l a t i o n a p p r o c h a n t d e la p h a s e s tat iomnaire , e s t la c o n s é q u e n c e d'une par t d e l ' adapta t ion à u n e fa ib le i n t e n s i t é l u m i n e u s e , e t d 'autre p a r t d e la v a r i a t i o n d u profil d'âge d e la p o p u l a t i o n ( r a p p o r t c e l l u l e s j e u n e s / c e l l u l e s â g é e s ) .
3 . 1 . 2 . V A R I A T I O N S MORPHOLOGIQUES DES CELLULES AU COURS DE LA
CROISSANCE
Les d i f férentes p h a s e s d'un c y c l e d e d i v i s i o n ce l lu la i re o n t é t é étud i é e s p o u r c h a q u e e s p è c e . D e s c u l t u r e s s y n c h r o n i s é e s , e n p h a s e expon e n t i e l l e e t d a n s d e s c o n d i t i o n s c l i m a t i q u e s s u b - o p t i m a l e s , s o n t s u i v i e s ( m e s u r e d e d e n s i t é o p t i q u e e t p h o t o g r a p h i e ) t o u t e s les h e u r e s (fig. 8 ) d u r a n t u n j o u r . P o u r les C h l o r o p h y c é e s , la d i v i s i o n ce l lu la i re s e prod u i t d u r a n t la p h a s e o b s c u r e (fig. 8 ) . P o u r l e s e s p è c e s filamenteuses (Fragillaria e t Anabaena) la c r o i s s a n c e se m a n i f e s t e p a r u n e a u g m e n t a t i o n g r a d u e l l e d e la d e n s i t é d e la c l u t u r e . P a r m i l e s p h o t o g r a p h i e s p r i s e s à i n t e r v a l l e s régu l i er d u r a n t t o u t e la d u r é e d u cyc l e , j 'a i sé lect i o n n é l e s c l i c h é s l e s p l u s r e p r é s e n t a t i f s d e s d i f férentes é t a p e s d e la c r o i s s a n c e d'une a lgue (fig. 1 à 4 ) . I l s m o n t r e n t b i e n q u e l 'extraordina i re v a r i a t i o n d e ta i l le s u r v e n a n t a u s e i n d 'une e s p è c e l or s d e s a c r o i s s a n c e m o d i f i e s a n s c e s s e d e s v a l e u r s c o m m e le b i o v o l u m e ce l lula ire o u l e r a p p o r t c h l o r o p h y l l e / c e l l u l e . Le p o u v o i r d e m u l t i p l i c a t i o n d e s C h l o r o p h y c é e s y e s t m i s e n va leur .
Densité de cellules ( I 0 9 Cell. /litre)
0 15 24 0 15 2 4 0 15 2 4 0 15 2 4 Temps en heures
FIG. 8. — Suivi de l'évolution d'une culture sur 2 4 heurees.
Je m e s u i s u n p e u p l u s a t t a c h é à p h o t o g r a p h i e r l e s d i f férents s t a d e s d e g r o s s i s s e m e n t e t d e d i v i s i o n d e s c é n o b e s d e Pediastrum boryanum.
C e t t e e s p è c e q u i p o s s è d e u n t a u x d e c r o i s s a n c e l é g è r e m e n t m o i n s

( 2 1 ) PARAMÈTRES DE CROISSANCE 2 3 7
é l e v é q u e les a u t r e s C h l o r o p h y c é e s é t u d i é e s (cf. § 3 . 1 . 4 . ) p r é s e n t e u n e
v a r i a t i o n d e b i o v o l u m e p a s s a n t d e 6 0 à 9 0 0 p.m3 ( v a l e u r m o y e n n e
= 3 5 0 /Ш1 3. A par t i r d'une c e l l u l e a d u l t e p l u r i n u c l é é e , la f o r m a t i o n d e s
c é n o b e s fils d é b u t e p a r l ' e x p u l s i o n d'une v é s i c u l e (fig. 4 d ) o ù l e s zoo
s p o r e s l i b é r é e s à l ' intér ieur n a g e n t p e u d e t e m p s ( 5 à 1 5 m n ) , p u i s s e
r e g r o u p e n t a u t o u r d'une d 'entre e l l e s p o u r f o r m e r u n n o u v e a u c é n o b e
qu i s 'aplat i t (fig. 4 e e t 4 f) e t p r e n d p e u à p e u s e s c o n t o u r s déf in i t i f s
( 1 0 à 3 0 m n ) . Ce s c é n a r i o avai t é t é décr i t p a r B o u r r e l l y ( 1 9 6 6 ) . La
v é s i c u l e e n g l o b a n t le c é n o b e , b i e n v i s i b l e lorsqu' i l e s t j e u n e , s e m b l e
d i s p a r a î t r e lorsqu' i l a u g m e n t e d e ta i l le .
3 . 1 . 3 . IMPORTANCE DU FACTEUR LUMIÈRE E N CULTURE
Afin d'être c a p a b l e d e m o d é l i s e r u n e c u l t u r e d a n s u n m i l i e u fini, i l e s t i n d i s p e n s a b l e d 'évaluer le p h é n o m è n e d ' o m b r e p r o p r e . E n é c o s y s t è m e nature l , l a l u m i è r e s ' a t t énue r a p i d e m e n t a v e c la p r o f o n d e u r , c e qu i e s t g é n é r a l e m e n t e x p r i m é p a r la loi d e Beer^Lambert :
I = 1 0 * e x p . (— К * Z) , o ù Z e s t la p r o f o n d e u r , 1 0 l ' in tens i t é d i s p o n i b l e e t К u n coeff ic ient d 'ex t inc t ion . E n c u l t u r e , le coeff ic ient К d é p e n d à la fo i s d e la g é o m é t r i e ( p a r a m è t r e c o n s t a n t ) d u f lacon e m p l o y é ( p r i n c i p a l e m e n t sa h a u t e u r d'eau) e t d e la c o n c e n t ra t ion d e c e l l u l e s . Ains i , p o u r u n e c u l t u r e p l a c é e d a n s u n f lacon e x p o s é à u n e l u m i è r e i n c i d e n t e Ii , l ' in tens i t é l u m i n e u s e d i s p o n i b l e ( I d ) d e v i e n t u n e f o n c t i o n d e la d e n s i t é d 'a lgues ( e x p r i m é e e n d e n s i t é opt i q u e D O ) :
I d = I i * e x p . (— К * D O ) .
P o u r l e t y p e d e f lacon u t i l i s é d a n s l e c a s d e s m e s u r e s de t a u x d e c r o i s s a n c e ( e r l e n m e y e r d e 3 0 0 m l ) , l ' ex t inc t ion d e la l u m i è r e a été é v a l u é e p a r r a p p o r t à l a d e n s i t é de c e l l u l e s e t p o u r c h a q u e e s p è c e (fig. 9 ) . C o m p t e t e n u d e la l u m i è r e i n c i d e n t e , l ' a j u s t e m e n t d e la c o u r b e , e n t r e la d e n s i t é o p t i q u e d e la c u l t u r e ( v a l e u r u n i t a i r e qu i p e u t ê t r e c a l c u l é e à par t i r d u T a b l e a u I I ) e t la l u m i è r e d i s p o n i b l e ( v a l e u r m e s u rée a u q u a n t a m è t r e ) , d o n n e u n e v a l e u r К = — 0 , 8 (fig. 1 0 ) . C e t t e v a l e u r q u i do i t ê t r e r e d é f i n i e p o u r c h a q u e t y p e d e f lacon d e c u l t u r e u t i l i s é , p e r m e t e n p a r t i c u l i e r d'ef fectuer d e s c o r r e c t i o n s s u r l e s c o n d i t i o n s l u m i n e u s e s l i é e s à la m e s u r e d'un t a u x d e c r o i s s a n c e . ,K p e u t var i er d e — 0 , 5 à — 4 e t p l u s , s e l o n la g é o m é t r i e d u s y s t è m e d e c u l t u r e u t i l i s é e .
M y e r s ( 1 9 5 3 ) ava i t re l i é l e t a u x d e c r o i s s a n c e d'une c u l t u r e d e Chlo-
rel la à la b a i s s e d ' in tens i t é l u m i n e u s e p a r se l f - shading m a i s l ' impor
t a n c e d e c e t t e c o r r e c t i o n a p a r f a i t e m e n t é t é m i s e e n é v i d e n c e p a r
L i v a n s k y ( 1 9 7 9 ) q u i t i en t c o m p t e d e s r a d i a t i o n s n o n a b s o r b é e s p o u r
d é c r i r e u n t a u x d e c r o i s s a n c e . Cet te c o r r e c t i o n e n f o n c t i o n de la den
s i t é d e s c e l l u l e s e s t é g a l e m e n t i m p o r t a n t e p o u r l e s i n c u b a t i o n s des t i
n é e s a u x m e s u r e s d e p h o t o s y n t h è s e ( S e n g e r e t F l e i s c h h a c k e r 1 9 7 8 ) .

238 A. DAUTA (22)
Extinction de la Lumière
100%
50%
100%
50%
0 1.10 2.10 0 210 4.10 0 1.10 2.10 0 2.K) 4.10
Densité' de Cellules / litre
Fie 9 . — Extinction de la lumière (%) en fonction de la densité de cellules.
Intensité Lumineuse disponible ( u E / m 2 / s ) ( = Id )
-Intensité' lumineuse incidente ( = Ii )
I d = Il * e ^ 0 - 8 * 0 ^ 0
FIG. 10. ( K ) .
I 2 3 4 5 6 7 8 Densité Optique (=D.opt)
de la culture d'algues
Calcul de la lumière disponible. Ajustement du coefficient d'extinction

( 2 3 ) PARAMÈTRES DE CROISSANCE 2 3 9
3 . 1 . 4 . INTERACTION L U M I È R E - T E M P É R A T U R E
Les figures 1 1 à 1 8 r e p o r t e n t l e s t aux d e c r o i s s a n c e d e s h u i t e s p è c e s
c o n s i d é r é e s . Les c u l t u r e s o n t é t é s o u m i s e s à d e s t e m p é r a t u r e s c o m
p r i s e s e n t r e 1 5 °C e t 3 3 °C, e x c e p t é p o u r Fragillaria qu i fu t t e s t é e à
rée a u q u a n t a m è t r e ) , d o n n e u n e v a l e u r d e K = — 0 , 8 (fig. 1 0 ) . C e t t e
1 0 °C. P o u r c h a q u e t e m p é r a t u r e , p e u v e n t ê t r e déf inis ( a j u s t e m e n t gra
p h i q u e — trai t c o n t i n u ) u n t a u x d e c r o i s s a n c e o p t i m a l jim ( T ) , e t u n e
i n t e n s i t é l u m i n e u s e o p t i m a l e I o p t ( T ) . Les v a r i a t i o n s d e I o p t ( T ) e n
f o n c t i o n d e s e s p è c e s e t s e l o n la t e m p é r a t u r e e s t r e p o r t é e fig. 1 9 .
L ' e x a m e n d e s figures 1 1 à 1 8 a p p e l l e l e s r e m a r q u e s s u i v a n t e s :
— p o u r t o u t e s l e s e s p è c e s à l ' e x c e p t i o n d e Fragillaria, ¿u¿m ( T ) s'ac
cro î t a v e c la t e m p é r a t u r e , la v a l e u r a s y m p t o t i q u e c o r r e s p o n d a n t à
3 0 °C o u p l u s ;
— l e s C h l o r o p h y c é e s a t t e i g n e n t l e s t a u x d e c r o i s s a n c e l e s p l u s éle
v é s p o u r d e s t e m p é r a t u r e s e x c é d a n t 2 5 o u 3 0 °C, a l o r s q u e l ' o p t i m u m
es t d e 2 0 °C p o u r Fragillaria ;
— l e taux de c r o i s s a n c e l e p l u s é l e v é a é t é m e s u r é s u r u n e c u l t u r e d e Monoraphidium, p.m ( 3 5 °C) = 1 ,9 ;
— l a c r o i s s a n c e d e Anabaena r e q u i e r t d e s i n t e n s i t é s l u m i n e u s e s
n e t t e m e n t p l u s b a s s e s q u e l e s a u t r e s e s p è c e s ;
— I o p t ( T ) var i e d a n s l e m ê m e s e n s q u e la t e m p é r a t u r e . Fragillaria
s e m b l e n e p a s s u i v r e c e m ê m e s c h é m a .
L e s p a r a m è t r e s d e c r o i s s a n c e o n t é t é m e s u r é s a v e c u n e p h o t o p é
r i o d e d e 1 5 / 9 . Afin d ' e s t imer l e s c o n s é q u e n c e s de c e c h o i x arb i tra ire ,
u n e e x p é r i e n c e s u p p l é m e n t a i r e à 2 5 °C a é t é r é a l i s é e p o u r r e p l a c e r
c e t t e v a l e u r p a r m i u n e g a m m e d ' é c l a i r a m e n t s j o u r n a l i e r s a l lant d e 1
à 2 4 h e u r e s (fig. 2 0 ) .
Les r é s u l t a t s e x p r i m e n t l e s r e l a t i o n s c o m p l e x e s e x i s t a n t e s e n t r e
d 'une p a r t l e t a u x d e c r o i s s a n c e , l e s p h a s e s d e d iv i s ion , la m o r p h o l o g i e
d e s c é n o b e s , l ' adapta t ion à l ' é c la i rement , e t d'autre par t l ' in tens i t é
l u m i n e u s e , la p h o t o p é r i o d e , la t e m p é r a t u r e e t l e c a s é c h é a n t l e s cond i
t i o n s n u t r i t i o n n e l l e s . Le l i en e n t r e r y t h m e d e d i v i s i o n e t synchron i
s a t i o n a é t é é v o q u é p a r d e n o m b r e u x a u t e u r s ( K o m a r e k e t S i m m e r
1 9 6 5 ; J a m e s 1 9 6 6 ; Carol l e t al . 1 9 7 0 ; D a r l e y e t V o l c a n i 1 9 7 1 ) . L e s
m o d a l i t é s de m o r p h o g e n è s e d e s c e l l u l e s filles, r a p p e l é e s p o u r l e s pr in
c i p a l e s e s p è c e s p a r S e t l i k ( 1 9 7 9 ) , o n t é t é r e m a r q u a b l e m e n t d é c r i t e s
p o u r Scenedesmus quadricauda p a r D o u c h a e t V e n d l o v a ( 1 9 7 0 ) , S t e e n -
b e r g e n ( 1 9 7 5 ) , e t Z a c h l e d e r e t al. ( 1 9 7 5 ) . La f o r m a t i o n d e c h a q u e
c é n o b e o u s t a d e ce l lu la i re e s t d é t e r m i n é e p a r la d u r é e d e la p h o t o
p é r i o d e , la d i v i s i o n p r o p r e m e n t d i t e e t la l i b é r a t i o n d e s c e l l u l e s filles
s e p r o d u i s a n t d u r a n t l a p h a s e o b s c u r e , la d i s t r i b u t i o n e n ta i l l e d e s
c é n o b e s é t a n t l i é e a u t a u x d e c r o i s s a n c e (Se t l ik 1 9 7 9 ) . Les v a r i a t i o n s
d e la m o r p h o l o g i e d e s c é n o b e s o u d e s c e l l u l e s p e u t var ier ( S e t l i k

2 4 0 A. DAUTA ( 2 4 )
Toux de croissance ( j-1 )
2Ô 100 2 0 0 3 0 0 4 0 0 5 0 0 600(1.1) 2 0 ¡55 2 0 0 300 4 0 0 5 0 0 6 0 0 (1.1)
100 2 0 0 3 0 0 4 0 0 5 0 0 600(11) £ 100 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 (l.l)
Toux Oplîmol
de croitsance — * ,
*/ (33g 0 IÔÔ 2 0 0 3 0 0 4 0 0 5 0 0 « 0 0 ( U I 0 5 Q i5 2 0 2 5 3 0 3 5 «C
lntin»iM Lumineui* IpE/Cmî/s) Température
Fie. 11. — Scenedesmus crassus. Variations du taux de croissance pour 'différentes combinaisons Lumière-Température. Variation du uopt en fonction de la température.
Taux de croissance ( j-i )
o £ _ . . . . 2, KM 2 0 0 3 0 0 4 0 0 5 0 0 600(1.1) Z\
100 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 (IL)
KI9
100 200 3 0 0 4 0 0 5 0 0 600(11: 2 100 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 ( 1 . Il
TQUK Optimal .
0 ¡30"
rj« crol t u n e e
JJOpUT)
2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 (LU 0 5 10 15 20 2 5 3 0 3 5 "C Intensifs lumineu»e ( u £ / m 2 / i ) Temptroture
FIG. 12. — Coelastrum microsporum. Variations du taux de croissance pour différentes combinaisons Lumière-Température. Variation de uopt en fonction de la température.

(25) PARAMÈTRES DE CROISSANCE 241
Taux de croissance ( j-t )
100 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 01) ¿1 100 2 0 0 3 0 0 4 0 0 5 0 0 SCO (LU
100 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 U l ] 21 1 0 0 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 ( 1.1) Toux Optimal
AJOpttT)
O 100 2 0 0 3 0 0 4 0 0 . , 3 0 0 6 0 0 (U) O 5 K) 19 2 0 2 9 3 0 3 5 »c Intftntittf Lumlrwuu ( u E / m V i ) Tampéralura
FIG. 13. — Diciyosphaerium pulcheîlum. Variations du taux de croissance pour différentes combinaisons Lumière-Température. Variation de juppt en fonction de la température.
de croissance ( j _ 1 )
*••..:»•..,;„ MX)
U
200 3 0 0 4 0 0 5 0 0 600(11)
/ /
f (S) '
100
u
2 0 0 300~ 4 0 0 5 0 0 600(1.1)
/•t (S) II
K » 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 ( I I )
K » 2 0 0 3 0 0 4OO 5 0 0 6 0 0 (IL)
Taux Optimol
de cm)»*once « ' *
UOpt(T) /
hl ent i té lun*eu*e ( 1 1 E / m>A)
FIG. 14. — Scenedesmus quadricauda. Variations du «taux de croissance pour différentes combinaisons Lumière-Température. Variation de uopt en fonction de la température.

2 4 2 A. DAUTA ( 2 6 )
Taux de croissance { R
"~JOÔ 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 (.I.Lf 2* 2ÔÔ 3 0 0 4 0 0 5 0 0 6 0 0 ii.L] J
(22C)
100 2 0 0 3 0 0 4 0 0 5 0 0 ^ 6 0 0 ^ U L )
100 200 300 4 0 0 5 0 0 6 0 0 (LU
Taux OPTIMAL
D* CROISA
0 ¡00 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 (11) 0 « 10 15 * 2 0 25 3 0 35 *C Intensité" Lumineuse ( > £ / m^/s) Température
FIG. 15. — Fragïlîaria biâens. Variations du taux de croissance pour différentes combinaisons Lumière-Température. Variation de nopt en fonction de la température.
Toux de croissance ( r 1 )
@
100 4 0 0 3 0 0 4 0 0 5 0 0 600 (IL) ' 2
2-1 100 2 0 0 3 0 0 4 0 0 5 0 0 600(1.1) 2
u (S)
DO 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 ( I I ) D
(29t )
-100 2 0 0 3 0 0
TQUX Qptimo^
de croissance
UOpUT)
4 0 0 5 0 0 6 0 0 ( 1L)
•
0 ¡00 2 0 0 3 0 0 4 0 0 5 0 0 600(1.1} 5 ¡0 ¡5 2 0 2 5 3 0 3 5 * C ~ Intensité Lumineuse (JJE/ m z / s ) Température
FIG. 16. — Anabaena cylindrica. Variations du taux de croissance pour différentes combinaisons Lumière-Température. Variation de uopt en fonction de la température.

( 2 7 ) PARAMÈTRES DE CROISSANCE 243
Toux „ de croissance (r1)
""¡00 2 0 0 3 0 0 ' 4 0 0 SÔÔ 6 0 0 t u f 2
100 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 (IL) ' £ " ¡ 0 0 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 l l i j '
Taux Optimal
de croissance
U O P U T I
t ) ioÔ 2 0 0 3 Ù 0 4CO 5Ô0 « b O U l f 0 5 K> 15 2 0 2 5 3 9 3 5 *C Intensité 1 lumineuse (uE/mZ/t) Tempétoture
FIG. 17. — Pediastrum boryanum. Variations du taux de croissance pour différentes combinaisons Lumière-Température. Variation de uopt en fonction de la température.
de croissonce ( j - ' )
tOO 2 0 0 3 0 0 4 0 0 5 0 0 6 0 0 {Ut 2
100 2 0 0 3 0 0 4 0 0 5O0 60011.0 2l
/ (35 Ci
100 2 0 0 3 0 0 4 0 0 50O - 6 0 0 ( U )
/ (30CJ
100 2 0 0 3 0 0 4O0 5 0 0 6 0 0 ((IL)
Toux Optimol
de croissonce f
JJOpt.(T) /
O J ( 0 0 2 0 0 30Ô 4 0 0 5 0 0 6 0 0 ( U ) ^ 0 5 K> 6 ~20~ 25" 3 0 "35 "C Intensité Lgmneuse I t C / m ' / s ) Température
FIG. 18. — Monoraphidium minutum. Variations du taux de croissance pour différentes combinaisons Lumière-Température. Variation de uopt en fonction de la température.

244 A. DAUTA (28)
1979) : p o u r u n e p h a s e d e c r o i s s a n c e p l u s l o n g u e , la p r o p o r t i o n de
c e l l u l e s m è r e s d a n s la p o p u l a t i o n e s t s u p é r i e u r e à ce l l e q u i r é s u l t e
d'un taux d e c r o i s s a n c e é l evé . Les v a r i a t i o n s m o r p h o l o g i q u e s i n d u i t e s
par l e s l i m i t a t i o n s e n n u t r i m e n t s (§ 3.1.1.) o n t é t é r e m a r q u é e s par
d 'autres a u t e u r s :
— a p p a r i t i o n d e s t a d e s u n i o e l l u l a i r e s chez Scenedesmus obliquas,
c a s d 'une c a r e n c e e n p h o s p h o r e ( S h u b e r t e t T r a i n o r 1974),
— a p p a r i t i o n de c e l l u l e s i s o l é e s s e x u é e s c h e z Scenedesmus obliquus,
c a s d 'une c a r e n c e e n a z o t e ( T r a i n o r e t B u r g 1965 a ) ,
— c o n t r ô l e d u n o m b r e d e c e l l u l e s p a r c é n o b e (4 , 8, 16) c h e z Coelas-
trum microsporum p a r la c o n c e n t r a t i o n e n p h o s p h o r e d a n s le m i l i e u
( C h a n 1976).
eoo,
4 0 0
2 0 0
0
6 0 0
4 0 0
2 0 0
Intensité Lumineuse
S e
5 10 15 2 0 2 5 3 0 °C 5 10 № 2 0 2 5 30 °C 5 10 15 2 0 25 3 0 °C 5 10 15 2 0 2 5 3 0 °C
© © ©
0 5 10 15 2 0 25 3 0 °C 5 10 15 2 0 2 5 3 0 "C "5 10 15 2 0 25 3 0 ° C 5 10 15 2 0 2 5 3 0 °C T e m p é r a t u r e
FIG. 19. — Variation de Iopt en fonction de la température.
Taux de croissance
0 6 12 24 0 6 12 24 0 6 12 24 0 6 12 Nombre d'heures d'exposition
â la lumière FIG. 20. — Croissance d'une population : influence du nombre d'heures d'éclaire-
ment.

(29) PARAMÈTRES DE CROISSANCE 245
3.1.5. A S S I M I L A T I O N DE CARBONE
Les prof i l s d e p h o t o s y n t h è s e o b t e n u s n 'ont p a s é t é r e p r é s e n t é s . I l s
s o n t s e m b l a b l e s a u x prof i l s o b t e n u s p a r d e s i n c u b a t i o n s sur l e ter
ra in : i n h i b i t i o n p l u s o u m o i n s p r o n o n c é e p o u r l e s f o r t e s i n t e n s i t é s
l u m i n e u s e s et o p t i m u m v a r i a b l e s s e l o n la t e m p é r a t u r e e t l e s e s p è c e s .
C o n f o r m é m e n t a u m o d è l e p r o p o s é p a r Tal l ing (1955 e t 1957), d e u x
p a r a m è t r e s p e u v e n t ê t re re l i é s à la f o r m e d e s profi ls e t à l e u r s
v a r i a t i o n s :
— la c a p a c i t é p h o t o s y n t h é t i q u e o p t i m a l e ( P m a x ) ,
— l ' in tens i t é s eu i l d e s a t u r a t i o n d e la p h o t o s y n t h è s e p a r la l u m i è r e
( Ik) .
Ces d e u x p a r a m è t r e s s o n t d é t e r m i n é s g r a p h i q u e m e n t à part ir d e s
profi ls . La c a p a c i t é p h o t o s y n t h é t i q u e ( P m a x ) var ie d e 0,88 (Scenedes-
mus quadricauda, 15 °C) à 13,6 m g C a r b o n e a s s i m i l é / m g C h l o r o p h y l l e
a / h e u r e (Dictyosphaerium, 30 °C). P m a x e t Ik é v o l u e n t t o u s d e u x d a n s
le m ê m e s e n s q u e la t e m p é r a t u r e , Ik v a r i a n t d a n s u n e p l a g e a s s e z
é t r o i t e : e n t r e 90 (15 °C) e t 244 (30 °C) p o u r l ' e n s e m b l e d e s a u t r e s
e s p è c e s , à l ' e x c e p t i o n d e Anabaena p o u r l a q u e l l e l e s fa ib l e s v a l e u r s
d e I K (40 à 54 ju ,E .m" 2 . s _ 1 ) c o n f i r m e n t la s e n s i b i l i t é a u x f o r t s éc la ire -
m e n t s .
L e s v a l e u r s t r o u v é e s d e P m a x p a r a i s s e n t u n p e u p l u s é l e v é e s q u e
c e l l e s t r o u v é e s p a r Aruga (1965) e n c u l t u r e . O n r e m a r q u e c e p e n d a n t
q u e l ' a u g m e n t a t i o n d e P m a x a v e c la t e m p é r a t u r e (fig. 22 ) c o r r e s p o n d
à u n Q10 v o i s i n d e 2, c e qu i e s t e n a c c o r d a v e c la r e l a t i o n é t a b l i e p a r
( C a p b l a n c q 1982) à par t i r d e p l u s d e 1 000 d o n n é e s r e l e v é e s d a n s la
l i t t éra ture . D e m ê m e , l e s v a l e u r s p r i s e s p a r Ik a p p a r t i e n n e n t à l a
p lage d e v a l e u r s c i t é e s p a r R h o d e (1965) e t H a r r i s (1978) .
U n t r o i s i è m e p a r a m è t r e [ I o p t ( P S ) ] p e u t ê tre d é t e r m i n é sur l e s
c o u r b e s d e p h o t o s y n t h è s e e n f o n c t i o n d e l ' éc la i rement . M a l g r é l es
i m p r é c i s i o n s l i ée s à la d é t e r m i n a t i o n g r a p h i q u e e t l es effets t o u j o u r s
p o s s i b l e s d u p r é c o n d i t i o n n e m e n t d e s c u l t u r e s , c e p a r a m è t r e e s t a s s e z
v o i s i n d e I o p t (T) (cf. § 3.1.4. e t fig. 23) , l e s p l u s g r a n d s é c a r t s c o r r e s
p o n d e n t à Pediastrum e t Fragillaria.
3 .2 . Modélisation du taux de croissance
3.2.1. E C R I T U R E DU MODÈLE
Trois f a m i l l e s d ' é q u a t i o n s p e u v e n t r e n d r e c o m p t e d e la r e l a t i o n
c r o i s s a n c e - i n t e n s i t é l u m i n e u s e . C e s e x p r e s s i o n s o n t é t é e m p l o y é e s p a r
d i v e r s a u t e u r s p o u r décr i re u n t a u x d e c r o i s s a n c e o u u n t a u x d e p h o
t o s y n t h è s e e n f o n c t i o n d e la l u m i è r e . I l s'agit d e s é q u a t i o n s s u i v a n t e s :
— é q u a t i o n d e M i c h a e l i s - M e n t e n : p. = / tm * I / ( k I + 1), u t i l i s é e
Chen-Orlob (1968) ;

246 A. DAUTA (30)
— é q u a t i o n d e S t e e l e : JJ. = /un * a * I * e x p . ( 1 — a * I ) , d o n n é e p a r S t e e l e (1962) , u t i l i s é e p a r L e h m a n (1975) , C loern (1976) ;
— é q u a t i o n d e S m i t h : JJ. = /un * a * I / [ l + (a * I ) 2 ] * , d o n n é e
p a r S m i t h (1936) e t u t i l i s é e p a r Ta l l ing (1957), V o l l e n w e i d e r (1965) e t
F e e (1969), a v e c a = 1 / I o p t .
D a n s u n e p r é c é d e n t e é t u d e (Guerri e t al. 1981), l e s t e s t s d'ajuste
m e n t n o u s o n t c o n d u i t à r e t e n i r l ' é q u a t i o n d e S m i t h . C e p e n d a n t ,
c e t t e f o r m u l a t i o n n e r e s t i t u e p a s l ' inh ib i t ion d e la c r o i s s a n c e p a r l e s
for te s l u m i è r e s , p h é n o m è n e i m p o r t a n t p o u r c e r t a i n e s e s p è c e s t e l l e s
Anabaena ; d a n s c e c a s , l e t a u x d e c r o i s s a n c e e s t s u r e s t i m é . L ' é q u a t i o n
d e M i c h a e l i s - M e n t e n p r é s e n t a n t l e m ê m e dé faut , l ' équat ion d e S t e e l e
e s t c h o i s i e p o u r t radu ire l ' inf luence d e la l u m i è r e s u r l e t a u x d e
c r o i s s a n c e .
TABLEAU IV. — Paramètres de modélisation du taux de croissance. — Uopt (T) = A * T2 + B * T + C et Iopt (T) = D * T2 + E * T + F avec T en °C,
Iopt en jaE.m-2.s-l. et ju, en j-1. —> Domaine de définition : 15 °C < T < 35 °C pour toutes les espèces sauf 10 °C < T < 30 °C pour Fragillaria.
Uopt (T) Iopt (T) A B C D E F
Scenedesmus crassus —0,0062 0,3474 —3,2045 0,1157 1,2476 111,7 Coelastrum microsporum —0,0042 0,2805 —2,9170 0,30 — 6,50 160 Dictyosphaerium
pulchellum —0,0050 0,3180 —3,390 0,450 — 3,0 78,75 Scenedesmus quadricauda —0,0068 0,410 —4,4490 0,120 3,80 50 Fragillaria bidens —0,0062 0,252 —1,440 —1,110 47,30 —222 Anabaena cylindrica —0,0016 0,1031 —0,904 0,055 — 0,025 33,1 Pediastrum boryanum —0,0041 0,2597 —2,877 0,13 3,3 19 Monoraphidium minutum —0,0023 0,1995 —2,203 0,2875 0,425 9,25
3.2.2. CALIBRATION DES PARAMÈTRES
L e s e s t i m a t i o n s d e jxva. (T) e t I o p t ( T ) p e u v e n t ê t r e d é c r i t e s p a r u n e f o n c t i o n p o l y n o m i a l e d e la t e m p é r a t u r e ( a j u s t e m e n t p a r m é t h o d e s d e s m o i n d r e s c a r r é s ) .
E x e m p l e p o u r Coelastrum :
jum ( T ) = — 0 , 0 0 4 2 5 * T2 + 0,2805 * T — 2 , 9 1 7 (1 ) I o p t ( T ) = 0,3 * T2 — 6 , 5 * T + 1 6 0 (2 )
o ù / u n ( T ) : t a u x m a x i m a l spéc i f ique d e c r o i s s a n c e (par j o u r ) I o p t ( T ) : i n t e n s i t é l u m i n e u s e o p t i m a l e e n ju.E:m" 2 .s _ 1 .
T : t e m p é r a t u r e e n d e g r é C e l c i u s .
P o u r Coelastrum, c e s f o n c t i o n s s o n t déf in ies p o u r 15 < T < 35 "C.
L 'ut i l i sa t ion d'un s y s t è m e d ' é q u a t i o n f o r m é d e (1 ) e t (2 ) e t d e l 'équat ion d e S t e e l e p e r m e t d ' e s t imer s i m p l e m e n t u n t a u x d e c r o i s s a n c e m a x i m a l e n c o n d i t i o n s o p t i m a l e s e n f o n c t i o n d e la t e m p é r a t u r e , e t l ' in tens i t é l u m i n e u s e o p t i m a l e c o r r e s p o n d a n t e .


248 A. DAUTA ( 3 2 )
L e s v a l e u r s d e s p a r a m è t r e s p e r m e t t a n t d e m o d é l i s e r jum (T) e t I o p t
<T) e n f o n c t i o n d e la t e m p é r a t u r e (T) s o n t d o n n é s T a b l e a u I I I .
L ' incer t i tude re la t ive s u r le t a u x d e c r o i s s a n c e p r é d i t à l 'aide d e
c e s p a r a m è t r e s e s t c o m p r i s e e n t r e 3 e t 20 °/o ( B r u n e i e t Guerr i 1980).
3.2.3. V É R I F I C A T I O N ET VALIDATION DU MODÈLE
P o u r c h a q u e e s p è c e , e t p o u r c h a q u e c o m b i n a i s o n T e m p é r a t u r e -
L u m i è r e , l e t a u x d e c r o i s s a n c e a é t é m o d é l i s é p a r l ' équat ion d e S t e e l e
{ e n u t i l i s a n t l e s coeff ic ients d u t a b l e a u I V ) e t r e p o r t é s u r l e s f igures
11 à 18 ( tra i t d i s c o n t i n u ) .
O n e n r e g i s t r e c e r t a i n s é c a r t s a v e c l e s v a l e u r s e x p é r i m e n t a l e s ,
c o m m e p a r e x e m p l e p o u r Scenedesmus crassus (fig. 11 p o u r 15 °C) o u
Scenedesmus quadricauda (fig. 14 p o u r 20 °C), m a i s d a n s l ' e n s e m b l e
le t a u x d e c r o i s s a n c e e s t i m é p a r l e m o d è l e e s t p r o c h e d u t a u x m e s u r é
e x p é r i m e n t a l e m e n t e t l ' inh ib i t ion p a r l e s for te s i n t e n s i t é s l u m i n e u s e s
e s t b i e n r e t r a c é p o u r Anabaena.
Afin d e v a l i d e r c e s m o d è l e s , l es t a u x d e c r o i s s a n c e n 'ayant p a s s erv i
" " - - - a la d é t e r m i n a t i o n d e s p a r a m è t r e s e t m e s u r é s a u c o u r s d ' e x p é r i e n c e s
d i v e r s e s (ex. : su iv i 24 h e u r e s , c u l t u r e s m è r e s , c u l t u r e s u t i l i s é e s p o u r
e s t i m e r l e s r e l a t i o n s p o i d s s ec , c h l o r o p h y l l e , e t c . ) o n t é t é c o m p a r é s
a u x t a u x d e c r o i s s a n c e c a l c u l é s e n f o n c t i o n d e s c o n d i t i o n s e x p é r i m e n
t a l e s . D a n s d e n o m b r e u x c a s , l ' in tens i té l u m i n e u s e i n c i d e n t e d i sponi
b l e p o u r l e s a l g u e s a d û ê t r e c o r r i g é e (coeff ic ient K, cf. § 3.1.3.) p o u r c e
c a l c u l . La c o m p a r a i s o n e n t r e v a l e u r s c a l c u l é e s e t v a l e u r s m e s u r é e s
{fig. 21) m o n t r e q u e l e s m o d è l e s d o n n e n t u n e e s t i m a t i o n s a t i s f a i s a n t e
d u taux d e c r o i s s a n c e e n c o n d i t i o n s o p t i m a l e s e n n u t r i m e n t s .
3.3. Approximation du taux de croissance à l'aide des mesures d'assimilation photosynthétique
L e s d o n n é e s d e l ' a s s i m i l a t i o n d e c a r b o n e p a r p h o t o s y n t h è s e p e u v e n t ê t r e u t i l i s é e s p o u r c a l c u l e r u n t a u x d e c r o i s s a n c e p d e la p o p u l a t i o n . S i l 'on e s t i m e q u e le c a r b o n e ce l lu la i re r e p r é s e n t e u n e c o n s t a n t e p o u r u n e e s p è c e d o n n é e , t o u t d o u b l e m e n t d e c e s t o c k i m p l i q u e u n e divis i o n ce l lu la ire . La m u l t i p l i c a t i o n q d u c a r b o n e c e l l u l a i r e e s t t e l l e q u e :
q = e " , a v e c e" = 2 P (p = n o m b r e d e d i v i s i o n par j o u r ) . S i d C / d t
r e p r é s e n t e l e t a u x de p h o t o s y n t h è s e n e t t e d 'une p o p u l a t i o n d 'a lgues ,
i l e s t p o s s i b l e d e d é d u i r e u n taux d e c r o i s s a n c e ( E p p l e y 1968, P e t e r s o n
1978, C a p b l a n c q 1982) : = 1/dt) * L o g (1 + d C / C ) , o ù € r e p r é s e n t e
le c a r b o n e ce l lu la i re .
L 'appl i ca t ion a u x v a l e u r s m e s u r é e s a u § 3.1.5., p o u r u n c y c l e d'éclai-r e m e n t d e 15 h e u r e s e t e n s u p p o s a n t q u e le r a p p o r t r e s p i r a t i o n / P m a x

( 3 3 ) PARAMÈTRES DE CROISSANCE 2 4 9
50O
4 0 0
300
2x> 100
o
!K«(.uE/m2/s ]
( S e
500i
o 4 0 0
3 0 0
£ 0 0
* 100
0
500]
' 4 0 0
3 0 0
2 0 0
' 100-
o.
' 4 0 ° ! ( s q ) 3 0 0
2.00
ICO-0-
10 15 2 0 25 30 10 15 20 25 30 ß 15 2 0 25 3 0 10 15 20 2 5 30
IK«(j)E/m2/s ) 5 0 0
4 0 0
3 0 0
2 0 0
\oo ; *
o •
lOOl eo
o 6 0
» " 40-
2 0
0
5 0 0
4 0 0
' 300
200-
o 100
Ol
5 0 0
4 0 0
300
o î 200-
» ' 100-
0
15 ̂ max o
12 e n
Cossim.
m̂g ChLa/h
o
15 max o en
12
9 par 6 mgChl.a/h
3
o 10 15 20 25 30 10 15 20 25 30 10 15 2 0 25 3 0 " 1 0 15 2 0 25 30
Température
Fie. 22. — Paramètres de la photosynthèse. Evolution de Ik et Pmax en fonction de la température.
* 30°c o 25°C * 20°C * 15°C o I0°C
, 90O4COS0Û 0 100 200 300 400 500 0 100 200 300 400 600 0 100 200 3O0 400 5CO MotopUJ) 100,
»S/m*-/ s 0 100 200 300 400 5C0 0 20 40 60 80 100 0 100 200 300 4C0 503 0 ICO 200 300 400 503
Iopt (incubotion I4C )
FIG. 23. — Comparaison entre l'Intensité optimale estimée Iopt (incubation 14C) et l'Intensité optimale prédite par le modèle : Iopt (T).
2 JU, mod.
2 -Umod.
0 1 0 ° C o 15°C • 20"C * 2 5 ° C * 30°C
FIG. 24. — Comparaison entre le taux de croissance (uca) calculé à partir de Pmax et le taux de croissance (ujnod) calculé par modélisation.

250 A. DAUTA ( 3 4 )
= 0,1 [ v a l e u r m o y e n n e g é n é r a l e m e n t a d m i s e , b i e n q u e H a r r i s (1978)
i n d i q u e d e s v a r i a t i o n s e n t r e 0,01 e t 0 ,05] s 'écrit :
jt4 = I L o g { l + <15 * P m a x — 2 4 * P m a x * R ) / C ] (3) a v e c fi e x p r i m é p a r j o u r , e t P m a x e x p r i m é e n c a r b o n e a s s i m i l é par ce l lu l e .
La f igure 24 d o n n e la c o m p a r a i s o n e n t r e l e t a u x d e c r o i s s a n c e cal
c u l é p a r l e m o d è l e d e S t e e l e ( f tmod) e t c e lu i c a l c u l é d 'après l 'ass imi
l a t i o n d e c a r b o n e (juca). La c o r r é l a t i o n e s t r e l a t i v e m e n t b o n n e p o u r
la m a j o r i t é d e s e s p è c e s , sauf p o u r Fragillaria o ù l e ca l cu l p a r le car
b o n e a s s i m i l é p r é v o i t u n t a u x d e c r o i s s a n c e t r o p é l evé .
4 . — D I S C U S S I O N
E n c o n d i t i o n s n u t r i t i o n n e l l e s n o n l i m i t a n t e s , l e t a u x d e c r o i s s a n c e d 'une p o p u l a t i o n d 'a lgues e s t u n e f o n c t i o n d e l ' é c l a i r e m e n t e t d e la t e m p é r a t u r e . L ' i n t e r d é p e n d a n c e e n t r e c e s d e u x p a r a m è t r e s , e n c o r e m a l quant i f i ée m a l g r é le n o m b r e d ' é tudes c o n s a c r é e s à l ' éco log ie d u p h y t o -p l a n c t o n , a é t é é t u d i é e p o u r h u i t e s p è c e s d 'a lgues d 'eau d o u c e .
La d é t e r m i n a t i o n d e la c r o i s s a n c e d u n e p o p u l a t i o n d 'a lgues e n cult u r e é t a n t b a s é e s u r l ' a u g m e n t a t i o n d e la b i o m a s s e , p l u s i e u r s m o d e s d ' e x p r e s s i o n d e ce l le -c i o n t é t é c o m p a r é s . Le su iv i d 'une c u l t u r e p a r d e n s i t é o p t i q u e para î t ê t r e l e p a r a m è t r e l e p l u s f iable. La c o r r é l a t i o n d e c e t t e m e s u r e a v e c d 'autres u n i t é s d ' e x p r e s s i o n d e la b i o m a s s e e s t s a t i s f a i s a n t e m a i s d o i t ê t re c e p e n d a n t u t i l i s é e a v e c p r é c a u t i o n . Par e x e m p l e , si l 'on se ré fère à la d e n s i t é d e c e l l u l e s , l e s v a r i a t i o n s morp h o l o g i q u e s a u c o u r s d e la c r o i s s a n c e o u l i é e s a u x c o n d i t i o n s nutri t i o n n e l l e s p e u v e n t modi f i er la r e l a t i o n l inéa ire N o m b r e d e c e l l u l e s = A * D e n s i t é O p t i q u e . A u c o u r s d e l a c r o i s s a n c e , la v a r i a t i o n conti n u e l l e d e p a r a m è t r e s t e l s l e p o i d s s e c / c e l l u l e o u l e c a r b o n e / c e l l u l e n e p e r m e t p a s d 'é tab l i r a v e c p r é c i s i o n d e s r e l a t i o n s l inéa i re s a v e c l e n o m b r e d e c e l l u l e s . C e s v a r i a t i o n s , m i s e s e n é v i d e n c e p o u r d e s cult u r e s n o n l i m i t é e s e n n u t r i m e n t s p a r D o u c h a et V e n d l o v a ((1970), s o n t e n c o r e p l u s m a r q u é e s ( H e a l e y et H e n d z e l 1979) d a n s le c a s d e c u l t u r e s c a r e n c é e s : c h a n g e m e n t d e r a p p o r t s s t o e c h i o m é t r i q u e s , d e s ta i l l e s d e s c é n o b e s o u d e s c e l l u l e s — p a r e x e m p l e , l ' a c c r o i s s e m e n t d e ta i l le d e s c e l l u l e s o b s e r v é c h e z Pediastrum duplex p a r F u h s e t al. (1972) et L e h m a n (1976) d a n s l e c a s d e c a r e n c e e n a z o t e . D a n s c e travai l , le rô l e d e la p h o t o p é r i o d e a é t é é t u d i é d e f a ç o n t r è s f r a g m e n t a i r e . Les r e l a t i o n s c o m p l e x e s e x i s t a n t e n t r e p h o t o p é r i o d e , s y n c h r o n i s a t i o n et r y t h m e d e d i v i s i o n o n t é t é m i s e s e n v a l e u r p a r d e n o m b r e u x a u t e u r s d o n t P a a s c h e (1968) e t S e t l i k (1979). S i l e s c u l t u r e s p e r m e t t e n t d e c a l c u l e r l e s t a u x d e c r o i s s a n c e d e s p o p u l a t i o n s à par t i r d e p a r a m è t r e s d e b i o m a s s e m e s u r é s d a n s d e s c o n d i t i o n s d ' é c l a i r e m e n t t rè s p r é c i s e s .

(35) PARAMÈTRES DE CROISSANCE 251
il n 'en e s t p a s d e m ê m e d a n s l e s m i l i e u x n a t u r e l s o ù l e s m e s u r e s por
t e n t s u r u n e n s e m b l e p lur i spéc i f ique , e x t r ê m e m e n t d i l u é d a n s u n
m i l i e u m u l t i - d i r e c t i o n n e l n o n c o n t r ô l é . C'est p o u r q u o i il n o u s a paru
i n t é r e s s a n t d e t e s t e r la v a l e u r d e s p a r a m è t r e s g é n é r a l e m e n t u t i l i s é s
p a r l e s p h y t o p l a n c t o n o l o g i s t e s ( c h l o r o p h y l l e , a s s i m i l a t i o n p h o t o s y n
t h é t i q u e d e c a r b o n e ) .
N o s r é s u l t a t s c o n f i r m e n t d ' u n e p a r t l ' e x t r ê m e var iab i l i t é d e s r a p p o r t s
n o m b r e d e c e l l u l e s / p o i d s s e c / c h l o r o p h y l l e . I l s m o n t r e n t q u e s'il e x i s t e
u n e a s s e z b o n n e c o r r e s p o n d a n c e e n t r e l e s t a u x d e p h o t o s y n t h è s e e t d e
c r o i s s a n c e , l e s i n f o r m a t i o n s f o u r n i e s p a r l e s m e s u r e s d e p h o t o s y n
t h è s e s o n t p l u s a l é a t o i r e s . E n d e h o r s d e s p r o b l è m e s p o s é s p a r l e pré
c o n d i t i o n n e m e n t !à l a l u m i è r e ( J o r g e n s e n 1964), la c o n v e r s i o n d e s taux
d e p h o t o s y n t h è s e e n t a u x d e c r o i s s a n c e s u p p o s e la p r i s e e n c o m p t e
d e p a r a m è t r e s t e l s le q u o t a c e l l u l a i r e e n c a r b o n e e t la r e s p i r a t i o n . Or,
c e s p a r a m è t r e s s o n t s u s c e p t i b l e s d e var ier a u c o u r s d'un c y c l e d e
d i v i s i o n c e l l u l a i r e e t a v e c l e s c o n d i t i o n s n u t r i t i o n n e l l e s . S e n f t (1978)
m o n t r e p a r e x e m p l e q u ' e n c o n d i t i o n s n u t r i t i o n n e l l e s d é f a v o r a b l e s ,
P m a x d é p e n d d e s r é s e r v e s c e l l u l a i r e s e n p h o s p h o r e ; il é tab l i t a ins i
u n m o d è l e h y p e r b o l i q u e te l q u e : P o p t = P o p r 3 * (1 — K q / Q ) o ù K q
e t Q s o n t l e s s t o c k s i n t r a c e l l u l a i r e s e n p h o s p h o r e . P o u r d e s c u l t u r e s
c a r e n c é e s e n azote , R h e e (1978) , E p p l e y e t R e n g e r (1974) e t G l o v e r
(1980) t r o u v e n t u n e b o n n e c o r r é l a t i o n e n t r e P m a x et l e t aux d e c r o i s
s a n c e d a n s l e c a s o ù P m a x e s t e x p r i m é p a r u n i t é d e c h l o r o p h y l l e , e t
p a s d e c o r r e s p o n d a n c e s i P m a x e s t e x p r i m é p a r ce l lu l e , c e q u i r é s u l t e
à la fo i s d e s v a r i a t i o n s d u q u o t a ce l lu la i re e n c h o r o p h y l l e e t d e la
ta i l le d e s c e l l u l e s e n f o n c t i o n d u d e g r é d e c a r e n c e . Enf in , la v a l e u r
m o y e n n e d e r e s p i r a t i o n u t i l i s é e d a n s l ' é q u a t i o n (3) , d é d u i t e d e s v a l e u r s
de la l i t t éra ture , p e u t di f f ic i lement ê t r e c o n s i d é r é e c o m m e u n e cons
tante , m a i s p l u t ô t c o m m e u n e f o n c t i o n c o m p l e x e d e la t e m p é r a t u r e
(Jones 1977), de l a ta i l l e d e s c e l l u l e s ( B a n s e 1976), d e l 'âge d e la cu l
ture ( R y t h e r 1954, 1956 a, 1956 b ) e t v r a i s e m b l a b l e m e n t d e l 'état nutri-
t i o n n e l d e s a l g u e s ( C a p M a n c q 1982). iLaws e t W o n g (1978) d o n n e n t la
r e s p i r a t i o n c o m m e u n e f o n c t i o n c o m p l e x e — (A + Bp.)/{2fn + A + B / i )
o ù A e s t la r e s p i r a t i o n b a s a l e e t l ' excré t ion , e t Bj/i la p e r t e e n car
b o n e d é p e n d a n t d e la c r o i s s a n c e . M a l g r é c e n o m b r e i m p r e s s i o n n a n t
d ' incer t i tudes qu i d é m o n t r e n t la n é c e s s i t é d e p o u r s u i v r e l e s recher
c h e s s u r l e s p r o b l è m e s l i é s à la r e s p i r a t i o n , o n p e u t c o n s i d é r e r q u e l e s
m e s u r e s d e p r o d u c t i o n d o n n e n t d e s i n f o r m a t i o n s sur l e s p o t e n t i a l i t é s
d e d é v e l o p p e m e n t d 'une p o p u l a t i o n e n f o n c t i o n d e la l u m i è r e , à d é f a u t
d 'une m e s u r e d u t a u x d ' a c c r o i s s e m e n t rée l . T o u t e f o i s , d a n s l e c a s d e
c u l t u r e s n o n l i m i t é e s e n n u t r i m e n t s e t m o y e n n a n t q u e l q u e s précau
t i o n s re la t ive s a u p r o t o c o l e e x p é r i m e n t a l , o n p e u t u t i l i s e r c e t t e
m é t h o d e p o u r d é t e r m i n e r r a p i d e m e n t l e t a u x d ' a c c r o i s s e m e n t d e s
a l g u e s .

252 A. DAUTA
L e s t a u x d e c r o i s s a n c e l e s p l u s é l e v é s c o n c e r n e n t l e s C h l o r o p h y c é e s
e t c e p o u r d e s t e m p é r a t u r e s s u p é r i e u r e s à 20 °C. Les v a l e u r s d e l'opti
m u m t h e r m i q u e d é d u i t e s d e n o s e x p é r i e n c e s c o r r o b o r e n t l e s o bs erv a
t i o n s d e n o m b r e u x a u t e u r s ; l es t e m p é r a t u r e o p t i m a l e s d e s Chloro
p h y c é e s s o n t l e p l u s s o u v e n t s u p é r i e u r e s à 3 0 °C ( F e l f d l d y 1962, A r u g a
1965, Gargas 1978), c e l l e s d e s C y a n o p h y c é e s c o m p r i s e s e n t r e 27 et
35 °C { K r i ï g e r e t E lo f 1978). L ' o p t i m u m e s t p l u s b a s p o u r la D i a t o m é e
Fragillaria bidens (20 °C), e t c o m p a r a b l e à c e u x t r o u v é s p o u r différen
tes D i a t o m é e s m a r i n e s p a r E p p l e y (1977), l e s v a l e u r s p o u r l e s Diato
m é e s d'eau d o u c e r e s t a n t rares . Anabaena n e p o s s è d e p a s u n t a u x d e
c r o i s s a n c e m a x i m a l t r è s é l e v é ( f imax = 0,7) m a i s p o u s s e à d e fa ib l e s
i n t e n s i t é s l u m i n e u s e s ; V a n L i e r e e t al. (1978) o n t o b s e r v é u n c o m p o r
t e m e n t s i m i l a i r e e n c o m p a r a n t u n e C h l o r o p h y c é e e t Oscillatoria agar-
dhii, m a i s d o n t l ' in tens i t é l u m i n e u s e o p t i m a l e ( 2 0 à 40 w . m " a ) e s t
c e p e n d a n t p l u s é l e v é e q u e ce l l e d ' A n a b a e n a . L e s v a r i a t i o n s d e l ' inten
s i t é o p t i m a l e r e q u i s e , d é j à o b s e r v é e s p a r d ivers a u t e u r s t e l s S o r o k i n
e t K r a u s s (1962) , S m a y d a (1969) , C l o e r n (1976) o u E p p l e y (1977) , s o n t
ici c l a i r e m e n t quant i f i ées . U n e c o m p a r a i s o n d e s r é s u l t a t s o b t e n u s a v e c
l e s d o n n é e s b i b l i o g r a p h i q u e s m o n t r e q u e c e s v a l e u r s s o n t a s s e z pro
c h e s de c e l l e s r e c e n s é e s p a r F o g g (1965), S t a n k o v i c (1960) , H u t c h i n s o n
(1967) , M o s s (1973), E p p l e y (1977) , Patr i ck (1979) o u Gargas (1978) ,
v a l e u r s e s t i m é e s s u r l e t erra in o u m e s u r é e s e n l abora to i re .
A part ir d e s d e u x p a r a m è t r e s I o p t e t p-opt e t d e l eur r e l a t i o n a v e c la t e m p é r a t u r e , i l e s t p o s s i b l e d e p r é d i r e a v e c u n e a s s e z g r a n d e cert i t u d e le t a u x d e c r o i s s a n c e d'un p o p u l a t i o n . C e p e n d a n t , l e s v a l e u r s p r é d i t e s s u r e s t i m e n t g é n é r a l e m e n t l ' inh ib i t ion p a r l e s f o r t e s in tens i tés l u m i n e u s e s . D e u x e x p l i c a t i o n s p e u v e n t ê t re f o r m u l é e s :
— l ' équat ion d e S t e e l e n e d o n n e q u ' u n e a p p r o c h e e n c o r e impar
fa i te d u p h é n o m è n e d ' inh ib i t ion p a r la l u m i è r e ,
—• le t aux de c r o i s s a n c e m e s u r é a u x f o r t e s i n t e n s i t é s l u m i n e u s e s in tègre u n e c e r t a i n e c a p a c i t é d e s a l g u e s à s 'adapter r a p i d e m e n t (24 h e u r e s ) a u x c o n d i t i o n s l u m i n e u s e s ( S t e e m a n - N i e l s e n e t J o r g e n s e n 1968).
Le m o d è l e p r é s e n t é décr i t le t a u x d e c r o i s s a n c e e t l ' in tens i t é opti
m a l e c o m m e d e s f o n c t i o n s p a r a b o l i q u e s d e la t e m p é r a t u r e ( m é t h o d e
e m p i r i q u e u t i l i s é e p a r A h l g r e n 1978), c e qui p e r m e t e n p a r t i c u l i e r de
m o d é l i s e r u n e c h u t e a s s e z b r u t a l e d u taux d e c r o i s s a n c e l o r s q u e la
t e m p é r a t u r e o p t i m a l e e s t d é p a s s é e . Par c o n t r e , c e t y p e d e f o r m u l a t i o n
n e t e n d p a s t a n g e n t i e l l e m e n t v e r s z é r o p o u r l e s fa ib l e s t e m p é r a t u r e s ,
c e q u i c o r e s p o n d r a i t m i e u x a u x o b s e r v a t i o n s r é a l i s é e s d a n s l e m i l i e u
na ture l . E n fai t , d a n s la p l a g e de t e m p é r a t u r e qu i n o u s i n t é r e s s e et
qu i r e p r é s e n t e u n e réa l i t é é c o l o g i q u e (10 à 30 °C), o n p e u t a d m e t t r e
q u e le m o d è l e r e s t i t u e b i e n les p h é n o m è n e s m e s u r é s . La l i t t éra ture

(37) PARAMÈTRES DE CROISSANCE 253
fourn i t u n e v a r i é t é d ' é q u a t i o n s re l iant le taux d e c r o i s s a n c e o p t i m a l
à la t e m p é r a t u r e o u à la l u m i è r e . Ci tons p a r e x e m p l e :
/i = j u m * a * I * e x p ( l — b * I ) , V o l l e n w e i d e r (1965). Cet te
f o r m u l a t i o n p e r m e t d e m o d é l i s e r l e s effets s t i m u l a n t s e t i n h i b i t e u r s ,
m a i s p. r e s t e t o u j o u r s in f ér i eur à ,/u.opt.
/J. — / u n * T, Di T o r o et al. (1971), l e taux d e c r o i s s a n c e e s t cons i
d é r é d a n s u n e p r e m i è r e a p p r o c h e c o m m e l i n é a i r e m e n t d é p e n d a n t d e
la t e m p é r a t u r e .
= ju.ro. * exp (a * T) , E p p l e y (1972) . La d é p e n d a n c e e x p o n e n
t ie l le , é g a l e m e n t e m p l o y é e p a r C l o e r n (1976) , e s t la f o r m u l a t i o n la p l u s
u t i l i s é e e t s a n s d o u t e la p l u s e x a c t e tant q u e l 'on n e d é p a s s e p a s la
t e m p é r a t u r e o p t i m a l e .
p. = fum * e x p (—2,3 * (T — T o p t ) / ( T s — T o p t ) ) p o u r T > T o p t
o u p.m * exp (—2,3 * ( T — t o p t ) / ( T i — T o p t ) ) p o u r T < T o p t
Cet te a p p r o c h e b a s é e sur la loi N o r m a l e e s t d o n n é e par L e h m a n e t
al . (1975)
p. = / x m f f - T o p t ) , B i e r m a n (1976) .
E n réa l i t é , t o u t e s l e s f o r m u l a t i o n s c i t é e s , y c o m p r i s ce l l e a d o p t é e
d a n s c e t art ic le , p r é s e n t e n t u n d é f a u t , e t il a p p a r t i e n t a u m o d é l i s a t e u r
d e c h o i s i r la f o r m u l e qu i décr i t le m i e u x le t a u x d e c r o i s s a n c e d a n s
l ' interval le de t e m p é r a t u r e cho i s i . U n e f o r m u l a t i o n p a r u n s y s t è m e
d e d e u x é q u a t i o n s d é c r i v a n t i n d é p e n d a m m e n t le taux d e c r o i s s a n c e
r e s p e c t i v e m e n t a u - d e s s o u s e t a u - d e s s u s de la t e m p é r a t u r e sera i t peut -
ê t r e u n e s o l u t i o n à env i sager .
T R A V A U X C I T E S
AHLGREN (G.). 1978. — Growth of Oscillatoria agardhii in chemostat culture. 2 . Dépendance of growth constants on temperature. Mitt. Internat. Verein. Limnol., 21 : 88-102.
ALGEUS (S.). 1946. — Untersuchungen iiber die Ernâhrungsphysiologie des Chloro-phyceen. C. W. K. Gleerup, Forlag., Lund (Ed.) : 278 p.
ALLISON (F. E.) & MORRIS (H.J. ) . 1930. — Nitrogen fixation by blue-igreen algae.
Science, 71 : 221-223.
ANGELIER (E.), BORDES (J. M . ) , LUCCHETTA (JVC.) & ROCHARD (M . ) . 1978. — Analyse
statistique des paramètres physico-chimiques de la rivière Lot. Annls Limnol, 14 (1-2) : 39-57.
ANTIA (N. J . ) , MCALLISTER (CD.) , PARSONS (T. R.) & STEPHENS ( K . ) . 1963. — Further
measurements of primary production using a large volume plastic sphere. Limnol. Oceanogr., 8 : 166-183.
ARNON (D. I.). 1958. — The role of mieronutrients in plant nutrition with special reference to photosynthesis and nitrogen assimilation. In Trace elements, Ch. LAMB, O. G. BENTLEY and J. M . BEATTIE (Eds.), N. Y., Academic Press,
pp. : 1-32.
ARUGA (Y.). 1965. — Ecological studies of photosynthesis. II. Photosynthesis of algae. Bot. Mag. Tokio (78) : 360-365.

254 A. DAUTA ( 3 8 )
AZAD (H. S.) & BORCHARDT (J. A.)- 1970. — Variations in phosphorus uptake toy algae. Environ. Sci. Technol., 4 (9) : 737-743.
BANSE ( K . ) . 1976. — Rates of growth, respiration and photosynthesis of unicellular algae as related to cell size. A review. / . Phycol., 12 (2) : 135-140.
BERKOVA (E.), DOUCHA (J . ) , KUBIN ( S . ) , ZACHLEDER (V.) & SETLIK (I.). 1972. — Varia
tions in photosynthetic characteristics of Scenedesmus quadricauda during the cell cycle. In : G. FORTI, M. AVRON, A. MELANDRI (Eds.), Proc. Ilnd. Internat. Congr. Photosynth. Res. Stressa, 1971, W. Junk; The Hague: 2019-2032.
BIERMAM (V.J. Jr). 1976. — Multi-nutrient dynamic models of algal growth and species competition in eutrophic lakes. In : Modeling the Eutrophication processes in aquatic Ecosystems, Canale R. P. (Ed.), Ann Arbor Science Publishers, Inc. : 89-109.
BOURRELLY (P.). 1966. — Les algues d'eau douce. Tome 1 : Les algues vertes. Bou-bée & C e , Paris : 500 p.
BOWEN ( D . M. H.). 1970. — The Great Phosphorus controversy. Environ. Sci. Technol., 4 (9) : 725-726.
BREZONIK (P. L.). 1968. — Application of a mathematical model to the eutrophi-sation process. Proc. 11th Conf. Great Lakes Res. : 16-30.
BRUNEL-DELCLAUX (L.) & GUERRI (M. M.). 1980. — Cinétiques d'assimilation et de croissance de Scenedesmus crassus. Etude expérimentale et modélisation. Thèse 3° cycle, Toulouse : 226 p.
BRUNEI. (L.), DAUTA (A.) & GUERRI (M. M.). 1982. — Croissance algale : Validation d'un modèle à stock à l'aide de données expérimentales. Annls Limnol., 18 (2) : 91-100.
BUMBU (Y. U.) & MOKRYAK (A. S.). 1973. — Effect of some trace elements on the development of the algae Scenedesmus quadricauda (Turp.) Breb. Izv. Akad. Nauk. Mold. SSR., Ser. Biol. Kim. Nauk, USSR, 1 : 82-83.
BUTTERWICK (C), HEANEY (S.I.) & TAIXING (J. F . ) . 1982. — A comparison of eight methods for estimating the biomass and growth of planktonic algae. Br. Phycol. J., 17 : 69-79.
CAPBLANCQ (J.). 1982. — Phytoplancton et production primaire. In : Ecologie du plancton des eaux continentales. R. POURRIOT (Ed.), Masson, Paris : 1-48.
CAPBLANCQ (J.) & DAUTA (A.). 1978. — Phytoplancton et production primaire de la rivière Lot. Annls Limnol, 14 (1-2) : 85-112.
CAPBLANCQ (J.) & DÉCAMPS (H.). 1977. — Dynamics of the phytoplankton in the river Lot. Verh. Internat. Verein. Limnol, 20: 1479-1484.
CAPBLANCQ (J.) & TOURENQ (J. N.). 1978. — Hydrochimie de la rivière Lot. Annls Limnol, 14 (1-2) : 25-37.
CAPERON (J.) & MEYER (J.). 1972. — Nitrogen limited growth of marine phytoplankton. II. Uptake kinetics and their role in nutrient limited growth of phytoplankton. Deep Sea Res., 19 : 601-618.
CAROLL (J.W.), THOMAS (S.), DUNAWAY (C. H.) & O'KELLEY (J.C.). 1970. — Light induced synchronization of algal species that divide preferentially in darkness. Photo Chem. Photobiol, 12 : 91-98.
CASANOVA-BATUT (T.). 1977. — Etude préliminaire en vue de la modélisation de l'écosystème Lot. Thèse 3" cycle, Toulouse : 109 p.
CHAHUNEAU (F.), DES CLERS (S.) & MEYER (J. A . ) . 1980. — Les modèles de simula
tion en écologie lacustre. Présentation des différentes approches et analyse des modèles existants. Acta Oecologia, Oecol. Gêner., 1980, 1 (1) : 27-50.
CHAN (K. Y . ) . 1976. — Control of colony formation in Coelastrum microsporum (Chlorococcales, Chlorophyta). Phycologia, 15 (2) : 149-154.
CHEN (C. W.) & ORLOB (G. T.). 1975. — Ecologie simulations for aquatic environments in : Systems analysis and simulations in ecology. III. Patten (B. C.) Ed., Academic Press Inc. : 475-528.

( 3 9 ) PARAMÈTRES DE CROISSANCE 255
CHU (S. P.). 1942. — The influence of the mineral composition of the medium on the growth of plankton algae. Part I . Methods and culture media. / . Ecol., 30 : 284-325.
CHU (S. P.). 1943. — The influence of the mineral composition of the medium on the growth of plankton algae. Part I I . The influence of the concentration of inorganic nitrogen and phosphate phosphorus. J. Ecol., 31 (2) : 109-148.
CLOERN (J. E.). 1976. — Population dynamics of Cryptomonas ovata. A laboratory, field and computer study. These P.H.D. Washington State University. Department of Zoology : 102 p.
COBD (H. D.) & MYERS (J.). 1964. — Comparative studies of nitrogen fixation and photosynthesis in Anabaena cylindrica. Am. J. Bot., 51 : 753-762.
CONDIT (R. J.). 1972. — Phosphorus and algal growth in Spokane River. North Sei., 46 (3) : 177-189.
Cox (R. M.) & FAY (P.). 1969. — Special aspects of nitrogen fixation by blue green algae. Pre. R. Soc. B, 172 : 537-366.
DARLEY (W. M.) & VOCANI ( B E . ) . 1971. — Synchronised cultures: Diatoms, i n : Methods in Enzymology. Colowick (S. P.) & Kaplan (N. O.) Eds., (13) : 85-96.
DAUTA (A.). 1975. — Etude du phytoplancton du Lot. Annls. Limnol., 11 (3) : 219-23J.
DAUTA (A.), DÉCAMPS (H.), DENAT (D.), GALHARAGUE (J.), GIOT (D.), MASSIO (J.C.) &
TROLET (J. L.). 1975. — Etude de la turbidité et des matières en suspension dans les eaux du Lot et de leurs relations avec les écoulements. 3 . — Evolution des matières en suspension et de la turbidité dans une masse d'eau marquée. Rapport B.R.G.M. ; 75 S.G.N. 338 M.P.Y., 31 p.
DAUTA (A.), BRUNEL (L.) & GUERRI (M. M.). 1982. — Détermination expérimentale
des paramètres liés à l'assimilation de l'azote et du phosphore par Scene-desmus crassus. Annls Limnol., 18 (1) : 33-40.
DÉCAMPS (H.) & CASANOVA-BATUT (T. ) . 1978. — Les matières en suspension et la turbidité de l'eau dans la rivière Lot. Annls Limnol., 14 (1-2) : 59-84.
DENAT (D.). 1977. — Etude de la production primaire dans le réservoir de Cajarc (Lot). Thèse 3' cycle, Toulouse: 94 p.
DILLON (P. J.) & RIGLER (F. H.). 1974. — A test of a simple nutrient budget model predicting the phosphorus concentration in lake water. / . Fish. Res. Bd. Can., 31 (11) : 1771-1778.
D I TORO (D.M.), O'CONNOR (D.J.) & THOMANN (R. V . ) . 1971. — A dynamic model of phytoplankton populations in the Sacramento-San Joaquin Delta. In : Advances in Chemistry Series 106 : Non equilibrium Systems in natural water chemistry, R. F. Gould (Ed.), Washington, American Chemical Society : 131-180.
DOR (I.). 1978. — Effect of the green algae Scenedesmus obliquus and Chlorella vulgaris on heterotrophic bacteria in polluted waters. Verh. Internat. Verein. Limnol., 2 0 : 1930-1933.
DOUCHA (J.). 1979. — Continuous cultures of algae, in Algal Assays and Monitoring Eutrophication. Marvan P., Pribil S., Lhotsky O. (Eds.), Schweizer-bart'sche Verlagsbuchhandlung, Stuttgart ; pp. : 181-191.
DOUCHA (J.) & VENDLOVA (J.). 1970. — Growth and synthetic processes in continuous cultures of Scenedesmus quadricauda. Ann. Rep. Lab. Algol., 1969 : 129-140.
DREWES (K.). 128. — Uber die assimilation des Lufstikstoffs durch Blaualgen. Zentr. Bakteriol. Parasiteenk Abt., II. 7 6 : 88-101.
DROOP (M. R.). 1969. — New thoughts on nutrient limitation in mioroalgae. Br. Phycol. J., 4 : 212.

256 A. DAUTA ( 4 0 )
DROOP (M. R . ) . 1973. — Some thougths on nutrient limitation in algae. J. Phycol., 9 (3) : 264-272.
DROOP ( M . R . ) . 1974. — The nutrient status of algal cells in continuous cultures. J. Mar. Biol. Assoc. U.K., 54 : 825-855.
DROOP (M. R . ) . 1975. — The nutrient status of algal cells in batch culture. J. mar. Biol. Assoc. U.K., 55 : 541-555.
EINSELE (W.) & GRIMM (J.). 1938. — Uber den Kieselsauregehalt planktischer Diatomeen und dessen Bedeutung fur Einige Fragen ihrer Ökologie. Z. Bot., 32 : 545-590.
EPPLEY (R .W. ) . 1968. — An incubation method for estimating the carbon content of phytoplankton in natural samples. Limnol. Oceanogr. 13 : 574-582.
EPPLEY (R. W.). 1972. — Temperature and phytoplankton growth in the sea. Fishery Bulletin, 70 (4) : 1063-1085.
EPPLEY (R. W.). 1977. — The growth and culture of diatoms. I n : The Biology of Diatoms, Werner D. (Ed.), Botanical Monographs, 13 : 24-64.
EPPLEY (R. W.) & RENGER ( E . H . ) . 1974. — Nitrogen assimilation of an oceanic diatom in nitrogen-limited continuous culture. J. Phycol., 10 : 15-23.
EPPLEY (R.W.) , ROGERS (J.N.), MCCARTHY (J.J.) & SOURNIA (A.). 1971. — Light/ dark periodicity in nitrogen assimilation of the marine phytoplankton Skeletonema costatum and Coccolithus Huxleyi in N-lim:ted chemostat culture. I. Phycol, 7 : 150-154.
FAY (P.). 1969. — Cell différenciation in Anabaena cylindrica. Br. phycol ]., 4 : 212-213.
FEE (E. J.). 1969. — Numerical model for estimation of P .S . production, integrated over time and depth in natural waters. Limnol. Oceanogr., 14 (6) : 906-911.
FEE (E. J.). 1973. — A numerical model for determimining integral primary production and its application to Lake Michigan. J. Fish. Res. Board Can., 30 : 1447-1468.
FELFOLDY (L. J. M.). 1962. — On the role of pH and inorganic carbon sources in photosynthesis in unicellular algae. Acta Biol Acad. Sei. Hung., 13 : 207-214.
FEUILLADE (J.) & FEUILLADE (M.). 1976. — Importance des oligo-éléments pour la croissance d'Oscillatoria rubescens D. C. in vitro. Ann. HydrobioL, 7 (1) : 1-9.
FLEMING (R. H.). 1939. — The control of diatoms by grazing. J. Cons. Explor. Mer., 14 : 210-2277.
FINDENEGG (I.). 1943. — Untersuchungen über die Ökologie und die Produktionsve-rhältniss des Plankton in Kärntner Seegebiele. Int. Rev. Ges. Hydrobiol Hydrogr., 43 : 368-429.
FOGG (G. E.). 1942. — Studies on nitrogen fixation by blue green algae. I. Nitrogen fixation by Anabaena cylindrica. I. Expl. Biol, 19 : 78-87.
FOGG (G. E.) & THAN-TUN. 1949. — Photochemical reduction of elementary nitrogen in blue green algae Anabaena cylindrica. Biochem. Biophys. Acta, 30 : 29-219.
FOGG (G. E.). 1952. — The production of extracellular nitrogenous substrances by a blue green alga. Proc. R. Soc, B, 139 : 372-397.
FOGG (G. E.). 1965. — Algal cultures and phytoplankton ecology. University of Wisconsin Press. : 126 p.
FOY (R. H.) & GIBSON (C. E.). 1982. — Photosynthetic characteristics of planktonic blue green algae : the response of twenty strains grown under high and low light. Br. Phycol. I., 17 : 169-182.
FRITZ (F.). 1935. — Uber die Sinkgeschwindigkeit einiger Phytoplanktonorganis-men. Int. Rev. Hydrobiol. Hydrogr., 32 : 424-431.

(41) PARAMETRES DE CROISSANCE 257
FUHS (G. W.). 1969. — Phosphorus content and rate of growth in the Diatoms Cyclotella nana and Thalassiosim fluviatilis. J. Phycol. 5 : 312-321.
FUHS (G. W.), DEMMERLE (S.D.), CANELLI (E.) & CHEN (M.). 1972. — Characteristics or phosphorus limited algae. In : G. E. Likens (ed), Nutrients and Eutrophi-cation. Ann. Soc. Limnol. Oceanogr. Spec. Symp., 1 : 113-132.
GARGAS I E . ) . 1978. — Phytoplaneton production chlorophyll-a and nutrients in the Open Danish Waters 1975-77. The Belt Project. The National Agency of environmental Protection, Denmark. : 61 p.
GATES (J . ) , ANDERSON (J . ) , LYNCH (D. L.) & FENWICK (M. G. ) . 1966. — Some physiological studies on two strains of the green alga Coelastrum microsporum Naeg. Trans. III. State Acad. Sei., 59 : 399-400.
GLOVER (H. E.). 1980. — Assimilation numbers in cultures of marine phytoplankton. J. of Plankton Res.,:2 (1) : 69-79.
GOLTERMAN (H. L.). 1969. — Methods for chemical analysis of fresh waters. I.B.P. n° 8. Blackwel Scientific Publ., Oxford : 172 p.
GRIFFITH (E. J . ) , BEETON (A.), SPENCER (J. M.) & MITCHELL (D. T.) eds. 1973. — Environmental phosphorus hanbook. J. Wiley & sons. Inc., N.Y.: 718 p.
GRIMM (J.). 1939. — Beobachtungen Am Phytoplankton des Bodeersees (Obersee) sozie dren rechnerische Auswertung. Int. Rev. Ges. Hydrobiol., 3 9 : 193-315.
GROSS (F.) & ZEUTHEN (E.). 1948. — The buoyancy of plankton diatoms : a problem of cell biology. Proc. R. Soc, 135 B . : 382-389.
GUERRI (M. M,), BRUNEL (L.) & DAUTA (A.). 1981. — Interaction de la lumiere et de la temperature sur le taux de croissanice de Scenedesmus crassus. Annls Limnol, 17 (2) : 97-104.
HARRIS (G. P.). 1978. — Photosynthesis, production and growth: The physiological ecology of phytoplankton. Arch. Hydrobiol. beih., Ergebn. Limnol, 1 0 : 1-171.
HARVEY ( H . W.). 1940. — Nitrogen and phosphorus required for the growth of phytoplankton. / . Mar. Biol. Ass. U.K., 24 : 115-123.
HEALEY (F. P.). 1973. — Characteristics of phosphorus deficiency in Anabaena. J. Phycol, 9 : 383-394.
HEALEY (F. P.) & HENDZEL (L. L.). 1979. — Indicators of phosphorus and nitrogen deficiency in five algal cultures. J. Fish. Res. Board Can., 3 6 : 1364-1399.
HUTCHINSON (G. E.). 1967. — A treatise on Limnology, vol. 2 . J. Wiley & Sons, Inc., N.Y. : 1 100 p.
JAMES (J.W.). 1966. — Cell synchrony, a prologue to discovery. I n : Cell Prologue, Cameron ,1 . L.) & Padilla (G. M.) Eds., Academic Press, N.Y. : 1-13.
JENKINS (S. H. ) & IVES (K.I.) eds. 1973. — Progress in water technology (Vol. 2 ) . Pergamon Press.: 739 p.
JONES (R. I.). 1977. — The importance of temperature conditioning to the respiration of natural phytoplankton communities. Br. Phycol. J., 12 : 277-285.
JORGENSEN (S. E.). 1964. Adaptation to different light intensities in the diatom Cyclotella meneghiniana Ktitz. Physiol. Plant., 17 : 136-145.
JORGENSEN (S. E.). 1979. — Handbook of environmental Data and ecological parameters. Jorgensen (Ed.), Int. Soc. Ecol. Modell., 1162 p.
KALFF (J.) & KNOECHEL ( R . ) . 1978. — Phytoplankton and their dynamics in oligo-trophic lakes. Ann. Rev. Ecol. Syst., 9 : 4 7 5 4 9 5 .
KETCHUM (B. H . ) . 1939. — The absorption of phosphates and nitrates by iEumi-nated cultures of Nitzschia closterium. Am. J. Bot., 26 : 399-4 7.
KILHAM (S.S.). 1975. — Kinetics of silicon limited growth in freshwater diatom AsterioneUa formosa. J. phycol., »11 : 396-399.
KILHAM (S.S.). 1978. — Nutrient kinetics of freshwater planktonic algae using batch and semi-continuous methods. Mitt. Internal Verein. Limnol., 21 : 147-157.

258 A. DAUTA (42)
KLEBHAN (H.). 1895. — Gasvacuolen, ein Bestandtheil der Zellen der wasserblü-thebilden Phycochromacean. Flora, Jena, 80 : 241-242.
KOMAREK (J.) & SIMMER (J. ) . 1965. — Synchronisation of the cultures of Scenedes-mus quadricauda (Turp.) Breb. Biol. Plant. (Praha), 7 : 4 0 6 4 2 4 .
KOMAREK (J.) & RUZICKA (J. ) . 1969. — Effect of temperature on the growth and variability of Scenedesmus quadricauda (Turp.) Breb. Stud, in Phycology. (Praha) : 75-144.
KRUEGER (G.H.J.) & ELOFF (J. N.). 1978. — The effect of temperature on specific growth rate and activation energy of Microcystis and Synechococcus isolates relevant to the onset of natural blooms. / . Limnol. Soc. Sth. Afr., 4 (1) : 9-20.
KUENTZEL (L. E.). 1970. — Phosphorus Innocent pawn in pollution polities. In « Chem. Spec. Manufacturers Ass. Proc. 56 th Mid. Year Meetg. Chicago, 111., USA, 1970. ». New York, Chem. Spec. Manufacturers Ass. : 92-95.
LAWS (E. A.) & WONG (D. C. L.). 1978. — Studies of carbon and nitrogen metabolism by three marine phytoplankton species in nitrate-limited continuous culture. J. Phycol., 14 : 406-416.
LECOHU (R.) & GUENE (Y.). 1976. — Influence de différentes formes d'azote sur la croissance de Staurastrum polymorphum. C. R. hebd. Sc. Acad. Sei-, Paris, sei-. D , 283 (10) : 1165-1168.
LEHMAN (J. T.). 1976. — Photosynthetic capacity and luxury uptake of carbon during phosphate limitation in Pediastrum duplex (Chlorophyeeae). J. Phycol, 1 2 : 190-193.
LEHMAN (J. T.), BOTKIN (D. B.) & LIKENS (G. E.). 1975. — The assumptions and
rationales of a computer model of phytoplankton population dynamics. Limnol Oceanogr., 20, 3 : 343-364.
LEISCHMAN (A.), GREENE (J. C.) & MILLER (W. E.). 1979. — Bibliography of littéra
ture pertaining to genus Selenastrum. Report EPA-600/9-79. 21 : 192 p. LIYANSKY ( K . ) . 1979. — Productivity of algal suspensions in cultivation systems
with different geometry. Arch. Hydrobial. (Suppl), 53 (3) : 339-346. LUKAVSKY (J . ) , SIMMER (J.) & KUBIN (S.). 1979. — Methods for algal growth evalua
tion in algal assays. In : Algal Assays and monitoring eutrophication. Mar-van P., Pribil S . & IJhostsky O. (Eds.), Sdhweizerbart'sche Verlagbuchhandlung, Stuttgart : 77-85.
LUND ( J . W . G . ) . 1949. — Studies on Asterionella Hass. I. The origin and nature of cells producing seasonal maxima. J. EcoL, 37 : 389-419.
MARKER (A. F. H.), CROWTHER (CA.) & GUNN (R. J. M.). 1980. — Methanol and
phaeopigments by spectrophotometry. Ergebn. Limnol, 14 : 52-69. MARGULIES (M. M.). 1970. — Changes in absorbance spectrum of the diatom Phaso-
dactylum tricornutum upon modification of protein structure. 7. Phycol, 6 : 160-164.
MARVAN (P.) & PRIBIL (S.). 1979. — Factors limiting growth of algal cultures, in Algal Assays and monotoring Eutrophication, Marvan P. Pribil S. & Lhotsky O. (Eds.), Schweizerbart'che Verlagbuchandlung, Stuttgart : 47-56.
MASSIO (J. C) . 1976. — Facteurs d'évolution des matières en suspension minérales et organiques dans les eaux du Lot. Thèse 3° cycle, Toulouse : 122 p.
MATHEW (T.) & CHOUDARY (Y.B.K..). 1977. —(Colony development m Scenedesmus. Phykos, 16 (2) : 47-50.
MEEKS (J. C) . 1974. — Chlorophylls, i n : Algal Physiology and Biochemistry., Stewart W. D. P. (Ed.), Blackwel Scientific publications, Oxford: 161-175.
-MEYER (J. A.). 1980. — L'analyse des systèmes écologiques. Revue des principaux thèmes de recherche. In Textes des communications au Colloque « La contribution des disciplines scientifiques à la notion de système ». CNRS.

(43) PARAMETRES DE CROISSANCE 259
MONOD (J . ) . 1949. — The growth of bacterial cultures. Ann. Rev. Microbiol, 3 : 371-394.
Moss (В.). 1973. — The influence of environmental factors on the distribution of freshwater algae : an experimental study. 3. Effects of temperature, vitamin requirements and inorganic nitrogen compounds on growth. J. Ecol, 61 (1), 179-192.
MULLIN (M. M.), SLOAN (P. R.) & EPPLEY (R. W.). 1966. — Relationship between carbon content, cell volume and area in phytophlankton. Limnol. Oceanogr., 11 : 307-311.
MYERS (J . ) . 1953. — Growth characteristics of algae in relation to the problem of mass culture. In : Algal Culture from Laboratory to Pilot Plant., Burlew J. S. (Ed.), Carnegie Institution of Washington Publication, 600 : 37-54.
О STERLING (S.). 1947. — Growth of planktonic green alga at various carbonic and hydrogen-ion concentration. Nature, Land., 159 : 199-200.
OSTERLIND (S.). 1948. — Influence of low concentrations on the growth of a green alga. Nature, Land., 161: 319-320.
OSTERLIND (S.). 1949. — Growth conditions of Scenedesmus quadricauda with special reference to inorganic carbon sources. Symb. Bot. Upsal, 10 (2) : 1-141.
PAASCHE (Е.). 1968. — Marine plankton algae grown with light-dark cycles. II. Ditylum brightwellii and Nitzschia turgidula. Physiologia PI, 21 : 66-77.
PARSONS (T. R.). 1961. — On the pigment composition of eleven species of marine phytoplankton. Л Fish. Res. Board Can., 18 :1017-1025.
PARSONS (T. R.), STEPHENS (K.) & STRICKLAND (J. D. H.). 1961. — On the chemical composition of eleven species of marine phytoplankton. J. Fish. Res. Board Can., 18 : 1001-1016.
PATRICK (R.). 1977. Ecology of freshwater diatoms and diatoms communities. In : The Biology of Diatom, Werner D. (Ed.), Botanical Monographs, 13 : 284-332.
PATTEN (B.C.). 1968. — Mathematical models of phytoplankton production. Int. Rev. Ges. Hydrobiol, 53 (3) : 357-408.
PETERSON (B. J . ) . 1978. — Radiocarbon uptake, its relation to net particulate carbon production. Limnol. Oceanogr., 23 :179-184.
PRIBIL (P.) & MARVAN (P.). 1970a. — Der Veriauf des Mineralnahrstoffbedarfes in Kultur von Scenedesmus quadricauda (Turp.) Breb. 1. Kalium-aufnhame. Archiv. Hydrobiol. supipl 39, Algolog. stud., 1 : 41-46.
PRIBIL (S.) & MARVAN (P.). 1970b. — Der Veriauf des Mineralnahrstoffbedarfes in Kultur von Scenedesmus quadricauda (Turp.) Breb. 2. Phosphor-aufnhame. Arch. Hydrobiol. suppl. 39, Algolog. Stud., 2/3 : 17-25.
PRIBIL (S.) & MARVAN (P.). 1971. — Der Veriauf des Mineralnahrstoffbedarfes in Kultur von Scenedesmus quadricauda (Turp.) Breb. 3. Stickstoff-Aufnahme. Archiv. Hydrobiol. suppl. 39, Algolog. Stud., 5 : 334-348.
PRIBIL (S.) & MARVAN (P.). 1979. — Batch cultivations, in Algal assays and Monitoring Eutrophication. Marvan P., Pribil S., Lhotsky O. (Eds.), Schweizer-bart'sche Verlagbuchhandlung, Stuttgart: 175-180.
RAYBURN (W. R.) & STARR (R. C) . 1974. — Morphology and nutrition of Pandorina unicocca sp. nov. I. Phycol, 10 : 4249.
RHEE (G. Y.). 1973. — A continuous culture study of phosphate uptake, growth rate and polyphosphate in Scenedemus sp. I. Phycol, 9 : 495-506.
RHEE (G. Y.). 1974. — Phosphate under nitrate limitation by Scenedemus sp. and its ecological implications. / . Phycol, 10: 470475.
RHEE (G. Y.). 1978. — Effect of N : P atomic ratios nitrate uptake. Limnol Oceanogr., 23 (1) : 10-25.
RODHE (W.Y. 1948. — Environmental requirements of freshwater plankton algae.

260 A. DAUTA (44)
Experimental studies in the ecology of phytoplankton. Symb. Bot. Upsal., 10 (1) : 1-149.
RODHE (W.). 1965. — Standard correlations between pelagic photosynthesis and light. Mem. 1st. Ital Idrobiol., 18 (Suppl.) : 365-381.
RUTTNER (F . ) . 1930. — Das Plankton des Lunzer Untersees seine Verteilung in Raum und Zeit währed des Jahre 1908-1913. Int. Rev. Ges. Hydrobiol. Hydrogr., 23 : 1-138.
RUTTNER (F . ) . 1937). — Limnologische Studien au einigen Seen des Ostalpen, Arch. Hydrobiol, 32 : 167-319.
RUZICKA (J.). 1971. — Morphologische Variabilität der Algenhervorgerufen durch Kultivierungsbedingungen. Arch. Hydrobiol. suppl 39, Algol Stud., 4 : 146-177.
RYTHER (J. H.). 1954. — The ratio of photosynthesis to respiration in marine plankton algae and its effect on the measurement of productivity. Deep-Sea Res., 2 : 223-225.
RYTHER (J. H.). 1956 a. — Interrelation between photosynthesis and respiration in the marine flagellate, Dunaliella euchlora. Nature (Lond.), 178 : 861-862.
RYTHER (J. H.). 1956lb. — Photosynthesis in the ocean as a fonction of light intensity. Limnol Oceanogr., 1: 61-70.
SENFT (W. H.). 1978. — Dépendance of light saturated rates'of algal photosynthesis on intracellular concentrations of phosphorus. Limnol. Oceanogr., 23 (4) : 709-718.
SENGER (H.) & FLEISCHHACKER (Ph.). 1978. — Adaptation of the photosynthetic apparatus of Scenedesmus obliquus to strong and weak light conditions. I. Differences in pigments, photosynthetic capacity, quantum yield and dark reactions. Physiol. Plant., 43:35-42.
SETLIK (I.). 1979. — CeM growth and division of unicellular algae and Cyariophy-ceae. In : Algols assays and monitoring eutrophication. Marvan P., Pibril S. & Lhotsky O. (Eds.), Schweizerbarts'che Verlagbuchhandlung, Stuttgart: 23-40.
Sr.TLiK (I.), BERKOVA (E.), DOUCHA (J.), KUBIN (S.), VENDLOVA (J.) & ZACHLEDER (V.). 1972. — The coupling of synthetic, and reproduction processes in Scenedesmus quadricauda. Archiv. Hydrobiol suppl. 41, Algolog. Stud., 7 : 172-217.
SHUBERT (L. E.) & TRAINOR (F . R.). 1974. — Scenedesmus morphogenesis control of the unicellular stage with phosphorus. Br. Phycol. J., 9 : 1-17.
SLOAN (P. R.) & STRICKLAND (J. H. D.). 1966. — Heterotrophy of four marine phyto-plankters at low substrate concentrations. J. Phycol, 2 : 29-32.
SMAYDA (T. J.). 1969. — Experimental observations on the influence of temperature, light and salinity on cell division of the marine diatom Detonula confervacea (Cleve) Gran. / . Phycol, 5 : 150-157.
SMITH (E.L.). 1936. —• Photosynthesis in relation to light and carbon dioxide. Proc. Nat. Acad. Sc. Wash., 22 : 504-511.
•SOROKIN (C). 1973. — Dry weight, packed cell volume and optical density. In : Handbook on phycological methods, Stein J. R. (Ed.), Cambridge : 321-343.
SOROKIN (C.) & KRAUSS (R. W.). 1962. — Effects of temperature and illuminance on Chlorella growth uncoupled from cell division. PI. Physiol, 37 : 37-42.
STANKOVIC (S.). 1960. — The Balkan Lake Ohrid and its living world. Monographiae Biol, 9 : 357 p.
STEELE (J. H.). 1965. — Notes on some theoritioal problems in production ecology. Mem. 1st. Ital. Idrobiol, 18 suppl. : 383-398.
STEEMAN-NIELSEN (E.) & JORGENSEN (E.G.). 1968. — The adaptation of plankton algae. I. General part. Physiol. Plant., 21: 401413.
STEENBERGEN (C. L. M.). 1975. — Light dependant morphogenesis of unicellular ista-

(45) PARAMÈTRES DE CROISSANCE 261
ges in syneronized cultures of Scenedesmus quadricauda (Turp.) Breb., Chlorophyceae. Acta Bot. Neerl, 2 4 (5-6) : 391-396.
STRATHMAN (R. R . ) . 1967. — Estimating the organic carbon content of phytoplankton from cell volume or plasma volume. Limnol. Oceanogr., 12 : 411-418.
STRICKLAND (J. D.H.). 1958. — Solar radiation penetering the ocean. A review of requierements data and methods of measurements, with particular reference to photosynthetdc productivity. 7. Fish, Res. Board. Can., 15 :453493 .
TALLING (J. F.). 1955. — The light relations of phytoplankton populations. Verh. internat. Verein. Limnol., 12 : 141.
TALLING (J. F.). 1957. — The phytoplankton as a compound photosynthetic system. New Phyto, 56 : 133-149.
TALLING (J. F.). 1979. — Factors interactions and implications for the prediction of lake metabolism. Arch. HydrobioL, 13 : 96-109.
TEZUKA (Y.). 1977. — The effect of nutrient concentration on the standing crop Scenedesmus obliquus grown under continuous culture conditions. Jap. J. Limnol, 38 (3) : 90-93.
TRAINOR (F. R . ) . 1963. — Zoospores in Scenedesmus obliquus. Science, 1 4 2 : 1673. TRAINOR (F. R.) & BURG (C. A.). 1965 a). — Scenedesmus obliquus sexuality. Science,
148 : 1094-1095.
TRAINOR (F. R.) & BURG (C. A.). 1965 b. — Motility in Scenedesmus dimorphus, Scenedesmus obliquus, Coelastrum microsporum. J. Phycol, 1 : 15-18.
TRAINOR (F. R.) & SHUBERT (L. E.). 1973. — Growth of Dictyosiphaerium, Selenas-trum and Scenedesmus (Chlorophyceae) in a dilute algal medium. Phyco-logia, 12 ( 1 / 2 ) : 35-39.
UTERMOHL (H.). 1925. — Limnologische Plankton Studien beseidlung ostholsteini-scher Seen mit Schwespflanzen. Arch. Hydrobiol, SuppL, 5 : 1-524.
UTERMOHL (V. H.). 1958. — Zur Vervollkommung der quantitative phytoplankton metodik. Mitt. Intern. Verein. Theor. Angew. Limnol, 9 : 1-38.
VAN LIERE (L.), LOOGMAN (J. G.) & MUR (>L. R . ) . 1978. — Measuring light-irradiance in cultures of phototropic micro-organisms. FEMS Microbiology Letters, 3 : 161-164.
VOLLENWEIDER (R. A.). 1960. — Beiträge zur Kenntnis optischer Eignen chaf ten der Gewässer und Primärproduktion. Mem. Ist. Ital. Idrobiol, 13 : 87-113.
VOLLENWEIDER (R. A.). 1965. — Calculation models of photosynthesis depth curves and some implications regarding day rates estimates in primary production measurement. Mem. 1st. Ital Idrobiol. (suppl), 18 : 4 2 7 4 5 7 .
VOLLENWEIDER (R. A.). 1968. — Les bases scientifiques de l'euùrophisation des lacs et des eaux courantes sous l'aspect particulier du phosphore et de l'azote comme facteur d'eutrophisation. Rapport O.C.D.E. D.A.S./C.S.I., 68-27 : 128 p.
VOLLENWEIDER (R. A.) ed. 1969. — A manual on methods for measuring primary production in aquatic environments. I.B.P. n° 12. Blackwel Scientific Publ., Oxford, 213 p.
WALSBY (A. E.) & FOGG (G. E.). 1975. — The extracellular products of Anabaena cylindrica Lemm. III. Excretion and uptake of fixed nitrogen. Br. Phycol. }., 10 : 339-345.
WARD (A. K.) & WETZEL ( R . ) . 1975. — Some effects on blue green algal growth. / . Phycol, 11 (4) : 357-363.
WHIPPLE (G.C.). 1895. — Some observations of the temperature of surface waters and the effects of temperature on the growth of microorganisms. / . New Engl. Wat. Wks Ass., 9 : 202-222.
WURTZ (A.). 1949. — Croissance d'une Volvolcale : Pandorina morum (Müll.) Bory dans divers milieux minéraux et organiques. Comptes rendus des Séances de la Société de Biologie, tome CXLIII : pl. 483.

262 A. DATJTA (46 )
YAU (P.C.) & CHAN (K. Y.). 1973. — Some physiological responses of two strains of Coelastrum Naeg. after auxin treatment. New Asia College Acad. Chinese Univ. Hong-Kong, 15 : 17-27.
ZACHARIAS ( 0 . ) . 1894. — Beobachtungen auf Plankton des Grossen Plöner Sees. Forsch. Ber. Biol. Stn Plön., 2: 91-137.
ZACHLEDER ( V . ) , DOUCHA (J . ) , BERKOVA (E.) & SETLICK ( I . ) . 1975. — The effect of synchronizing dark period on populations of Scenedesmus quadricauda. Biol. Plant. (Praha), 1 7 : 4 1 6 4 3 3 .