Embed Size (px)
Citation preview

Couplage électronique et transferts de chargesdans l’ADN : étude du contrôle énergétique1
Mustapha Benrahmoune, Abdelali Filali-Mouhim et Jean-Paul Jay-Gerin
Résumé: Nous étudions, dans le cadre du mécanisme de superéchange, l'influence du gap énergétique sur le taux dedécroissance effectif du couplage électronique (βeff) dans l'ADN. L'ADN est décrit par un Hamiltonien électroniquemodèle, basé sur l'approximation des liaisons fortes, dans lequel les diverses orbitales ont toutes la même énergie etinteragissent entre elles selon une fonction exponentielle de la distance. Nos résultats numériques concernant lesvaleurs deβeff obtenues pour deux molécules différentes d'ADN sont analysés en s'appuyant sur la théorie de l'« ap-proximation du milieu continu », développée précédemment par Lopez-Castillo et al. (J.-M. Lopez-Castillo, A. Filali-Mouhim, I.L. Plante et J.-P. Jay-Gerin. J. Phys. Chem. 99 : 6864–6875, 1995). Nous mettons en évidence que le pontd'ADN intervenant entre le donneur et l'accepteur est défini par un paramètre de contrôle unique, sans dimension,Γ/E,où E est l'énergie séparant les orbitales de ce milieu de celles des sites rédox (gap énergétique) etΓ est la largeur debande électronique du pont considéré comme milieu continu. Dans la limite des larges gaps, notre modèle de couplage« à travers l'espace » reproduit les bons ordres de grandeur des valeurs deβeff trouvées tant expérimentalement que pard'autres approches théoriques. Nous trouvons de plus que, pour des conditions énergétiques équivalentes, les transfertsd'électron et de trou au sein de l'ADN diffèrent considérablement. Cette différence est attribuée au signe du paramètreΓ/E.
Mots clés: ADN, couplage électronique, coefficient de décroissance effectif, gap énergétique,mécanisme de superéchange, approximation du milieu continu, transferts d'électron et de trou à longue distance.
Abstract: The influence of the energetic gap on the effective distance–decay rate of electronic coupling (βeff) in DNAis investigated in the context of the superexchange mechanism. The DNA double helix is described by a tight-bindingelectronic Hamiltonian model, in which all orbitals have the same energy and interact with one another through an ex-ponentially decaying function of distance. Our numerical results concerning theβeff values obtained for two differentDNA molecules are analyzed within the theoretical framework of the “continuous-medium approximation,” previouslydeveloped by Lopez-Castillo et al. (J.-M. Lopez-Castillo, A. Filali-Mouhim, I.L. Plante, and J.-P. Jay-Gerin. J. Phys.Chem. 99 : 6864–6875, 1995). We find that the intervening DNA bridge between the donor and acceptor sites is de-fined by a unique dimensionless control parameterΓ/E, whereE is the energy of the orbitals of this medium with re-spect to those of the redox site orbitals (energetic gap) andΓ is the electronic band width of the bridge considered as acontinuous medium. In the narrow-band regime, our “through-space” coupling model predictsβeff values that are ingood order of magnitude agreement with those calculated by other theoretical approaches as well as with those ob-tained from experiment. Moreover, under equivalent energetic conditions, the DNA-mediated transfers of holes andelectrons differ considerably. This difference depends upon the sign of the parameterΓ/E.
Key words: DNA, electronic coupling, effective distance–decay rate, energetic gap, superexchange mechanism,continuous-medium approximation, long-distance electron and hole transfers.
129
Benrahmoune et al.Introduction
Les lésions subies par la molécule d’ADN dues à l’actiond’un rayonnement ionisant sont d’une importance capitale
dans les modifications de la létalité (risque de perte de lacapacité de prolifération) cellulaire, ainsi que dansl’induction des mutations, l’initiation des cancers et de façonplus générale le vieillissement des organismes (voir, parexemple : Tubiana et al. 1990; Frenkel 1992; Ames et al.1993; Cadet 1994; Loft et Poulsen 1996; Gros et al. 1999).Il a été montré que ces radiolésions peuvent donner lieu àdes réactions de transferts de charges (TC), le long de cettemolécule, sur des distances pouvant atteindre, en moyenne,une dizaine de paires de bases (~40 Å) (voir, par exemple :O’Neill et Fielden 1993; Faraggi et al. 1995; Barbara etOlson 1999). Ainsi, par le jeu de tels TC intramoléculaires, àlongue distance, les sites lésés de l’ADN par suite del’irradiation ne correspondent pas nécessairement aux sitesdes attaques initiales. Cette migration des charges crééesinitialement dans l’ADN irradié revêt donc une grande
Can. J. Physiol. Pharmacol.79: 122–129 (2001) © 2001 CNRC Canada
122
DOI: 10.1139/cjpp-79-2-122
Reçu le 2 mars 2000. Publié sur le site Web des Pressesscientifiques du CNRC janvier 26, 2001.
M. Benrahmoune, A. Filali-Mouhim et J.-P. Jay-Gerin.2
Département de Médecine Nucléaire et de Radiobiologie,Faculté de Médecine, Université de Sherbrooke, Sherbrooke,QC J1H5N4, Canada.
1L’article a été soumis au processus habituel de révision parles pairs.
2Auteur correspondant (courriel :[email protected]).

importance en radiobiologie fondamentale, puisqu’ellepermet de préciser la localisation et la nature chimique desdifférents sites radicalaires finalement formés, premièreétape d’une succession d’événements chimiques, bio-chimiques et biologiques pouvant conduire à des dégâtscellulaires. Par ailleurs, il faut souligner que les réactions deTC entre protéines et ADN sont tout aussi essentielles auniveau des interactions ADN–protéines impliquées dans lesprocessus biochimiques normaux d’une cellule, tels que laréplication, la transcription et la réparation de l’ADN (voir,par exemple : Demple et Harrison 1994; Sarasin 1997).
La structure de la molécule d’ADN, avec notamment sonempilement sous forme de plateaux presque parallèles debases complémentaires (adénine–thymine et guanine–cyto-sine) riches en électronsπ, a toujours été au coeur d’un vifdébat quant à la possibilité de TC à longue distance le longde cette molécule (Ratner 1999). En particulier, la classifica-tion de l’ADN en « bon » ou en « mauvais » conducteurdépendrait, selon certains auteurs (voir, par exemple :Warman et al. 1996; Pezeshk et al. 1996; Beratan et al.1997; Taubes 1997; Holmlin et al. 1997; Netzel 1998), del’importance des interactions de recouvrement mutuel desorbitales π entre paires de bases successives, les TCs’opérant alors le long de tels « cheminsπ » (Murphy et al.1993; Stemp et Barton 1996).
D’une manière générale, le couplage électronique àtravers un pont moléculaire (ou « milieu intermédiaire »),entre un site donneur (D) et un site accepteur (A), est l’undes facteurs déterminants du contrôle de la constante devitesse (k) de réaction de TC entre D et A. Dans le cas desTC à longue distance, ce couplage est faible et la théorie desperturbations permet alors le calcul dek, qui s’écrit (Ulstrup1979; Marcus et Sutin 1985) :
[1] k T= 2 2πh
DA FCWD( )
où h est la constante de Planck divisée par 2π, TDA est lecouplage électronique à travers le milieu intermédiaire entreD et A, et FCWD est le facteur de Franck–Condonnucléaire. L’équation 1, connue sous le nom de règle d’or deFermi, est fondamentale en théorie des TC intramoléculairesà longue distance (transferts dits « non adiabatiques »).
Tout comme pour les réactions de TC dans les protéines(voir, par exemple : Moser et al. 1992; Gray et Winkler1996; Lopez-Castillo et al. 1997), l’expérience nousenseigne que, pour l’ADN,k décroît généralement d’unemanière exponentielle en fonction de la distance (d) séparantD et A :
[2] k d∝ −e effβ
où βeff est le taux de décroissance effectif dont la valeur estune caractéristique du milieu intermédiaire. Les différentesvaleurs expérimentales deβeff rapportées à ce jour dans lalittérature pour l’ADN offrent une large dispersion (voir, parexemple : Barbara et Olson 1999). Certains travaux (Brun etHarriman 1992, 1994; Atherton et Beaumont 1995; Meadeet Kayyem 1995; Lewis et al. 1997; Fukui et Tanaka 1998;Meggers et al. 1998) montrent, en effet, quek décroîtrapidement avecd (0,6 Å–1 < βeff < 1,8 Å–1; 10 Å = 1 nm),rapprochant ainsi l’ADN des protéines (pour lesquelles,
typiquement,βeff ~ 0,8 à 1,5 Å–1). En revanche, d’autrestravaux (Murphy et al. 1993; Kelley et al. 1997; Kelley etBarton 1999; Núñez et al. 1999; Giese et al. 1999) mettenten évidence une faible dépendance dek en fonction de ladistance (βeff < 0,2 Å–1), conférant ainsi à l’ADN despropriétés conductrices semblables à celles des filsconducteurs moléculaires.
Parallèlement aux études expérimentales, un certainnombre d’approches théoriques ont été proposées récemmentafin d’interpréter les phénomènes de TC observés (Risser etal. 1993; Felts et al. 1995; Priyadarshy et al. 1996a, 1996b,1998; Olson et al. 1997; Jortner et al. 1998; Henderson et al.1999). De ces travaux, on retient particulièrement deuxmécanismes importants, dont la prédominance de l’un parrapport à l’autre dépend du gap énergétique entre lesénergies des sites donneur et accepteur et les énergies desétats du milieu intermédiaire (fig. 1) : (i) le mécanisme du« superéchange » qui, aux larges gaps, dominerait le TC en-tre les sites rédox (Priyadarshy et al. 1996a, 1996b, 1998),se traduisant par une forte décroissance dek en fonction ded, et (ii ) le mécanisme de transport « par sauts » qui, enparallèle avec le phénomène du superéchange, interviendraitaux faibles gaps et dans le cas où l’énergie des sites rédoxest située à l’intérieur de la bande de conduction du pont(régime dit d’« injection ») (Jortner et al. 1998; Meggers etal. 1998) et qui se traduirait alors par une faible dépendancede k avec la distance.
Si les paramètres énergétiques jouent donc un rôle impor-tant dans les TC le long de l’ADN, ils sont néanmoins maldéfinis et souvent inaccessibles à l’expérimentateur et, parconséquent, leur influence sur la valeur deβeff est mal connue(Netzel 1998). En particulier, une bonne connaissance du gapénergétique entre le système donneur–accepteur et les étatsintermédiaires oxydés et(ou) réduits des bases complémentairespermettrait d’évaluer la contribution des électronsπ aucouplage électronique, contribution qui, on le sait, demeure en-core controversée (voir, par exemple : Barbara et Olson1999).Afin de mieux préciser l’importance de ce paramètre clé,nous développons, dans le présent travail, une étudesystématique de l’influence du gap énergétique sur la valeurdeβeff pour deux structures moléculaires d’ADN. Ce travailse situe dans la continuité d’un projet initié il y a quelquesannées dans notre laboratoire concernant l’étude ducouplage électronique, via le mécanisme du superéchange,dans les grands systèmes tridimensionnels apériodiques ou« topologiquement » désordonnés (Lopez-Castillo et al.1995a, 1995b; Filali-Mouhim et al. 1996), incluant lessystèmes moléculaires complexes tels que les protéines(Lopez-Castillo et al. 1997).
Théorie du superéchange
Le mécanisme du superéchange (voir, par exemple : Kuki1991) implique le transfert, entre D et A, d’un électron viales orbitales inoccupées du pont moléculaire et(ou) d’un trouvia les orbitales occupées de ce dernier (fig. 1). Il traduit ledegré de recouvrement des orbitales des sites donneur etaccepteur avec celles du milieu intermédiaire. De cerecouvrement, il résulte une délocalisation accrue de lafonction d’onde de la particule permettant son passage, pareffet tunnel, de D vers A. Dans le cas où le couplage
© 2001 CNRC Canada
Benrahmoune et al. 123

électronique est dominé par un tel mécanisme,TDA estdonné par la somme :
[3] TDA = VDA + VS
où VDA est le couplage direct donneur–accepteur, le plussouvent négligeable pour les TC à longue distance, etVS estle couplage dit de superéchange. Dans la limite nonadiabatique,VS s’exprime par (Ulstrup 1979) :
[4] VS = VDI I FDA FA( )E I H V− −1
où VDI et VFA sont respectivement les couplages de D et Aavec les sites d’entrée ou « initial » (noté I) et de sortie ou« final » (noté F) du milieu intermédiaire,EDA est l’énergiedes sites rédox (énergie de l’effet tunnel),I est la matriceidentité et H est l’Hamiltonien électronique du pontmoléculaire. Diverses méthodes de chimie quantique, ab initioou semi-empiriques, sont utilisées pour le calcul des élémentsde matrice de H (voir, par exemple : Skourtis et Beratan1999).
Modèle
Les séquences des bases complémentaires des deuxmolécules d’ADN étudiées dans ce travail sont indiquées àla figure 2. Les coordonnées atomiques de ces molécules(incluant les atomes d’hydrogène) proviennent de la banquede données « Protein Data Bank » (fichiers : 1BBX.pdb et4GAT.pdb). Dans le présent modèle, nous associons àchaque atome une orbitale atomique de types.
Les deux systèmes de TC ici considérés sont représentéspar l’Hamiltonien électronique modèle suivant (Lopez-Castillo et al. 1995a, 1995b; Filali-Mouhim et al. 1996), basésur l’approximation des liaisons fortes :
[5] H E i i V i ji
N
iji j
= += ≠∑ ∑
1
où N est le nombre total d’orbitales (ou de sites atomiques),E correspond à l’énergie des orbitales (que nous supposonsidentique pour tous les sites) etVij est le couplage entre lesorbitalesi et j donné par une loi exponentielle de la forme :
[6] Vij = V0 e–β*ri
– rj*
V0 étant un préfacteur de couplage,β le coefficient dedécroissance des interactions électroniques directes inter-sites, et ri et rj les coordonnées respectives des sitesi et j.Soulignons que ce type de couplage a été testé avec succèspour le calcul deVS associé à différentes structures com-plexes moléculaires (Gruschus et Kuki 1993).
Pour la présente étude, nous avons adopté pourβ et V0 lesvaleurs respectivement de 1,7 Å–1 et –6 eV, valeursgénéralement utilisées dans la modélisation d’Hamiltoniensélectroniques mettant en jeu des couplages du type de ceuxemployés ici (Gruschus et Kuki 1993; Evenson et Karplus1993; Skourtis et Beratan 1999). Par ailleurs,VDI et VFA sontconsidérés comme des grandeurs arbitraires étant donnéqu’elles n’influencent pas le taux de décroissance deVS enfonction de d. Enfin, EDA est choisie comme référence
énergétique (EDA = 0), de sorte que le gap énergétique seréduit simplement àE.
Approximation du milieu continu
Pour tout degré de complexité et de désordre du pontmoléculaire, l’« approximation du milieu continu » (AMC)permet de traiter celui-ci comme un milieu effectifhomogène, possédant les mêmes propriétés moyennesconcernantVS, pour lequel un traitement analytique est alorspossible (Lopez-Castillo et al. 1995a, 1995b; Filali-Mouhimet al. 1996). Dans le cadre de cette approximation, Lopez-Castillo et al. (1995a) ont montré que, selon le signe deV0/E, βeff s’exprime de deux manières tout à fait distinctes, àsavoir :
[7] β βeff = +
+2
21 1
ΓE
dans le cas oùV0/E > 0, qui correspond,V0 ayant étéchoisi négatif dans ce travail, à un transfert de trou (E < 0) et
[8] β βeff = −
2 1
ΓE
dans le cas oùV0/E < 0, qui correspond à un transfertd’électron (E > 0).
Les équations 7 et 8 montrent que la valeur deβeff estentièrement définie par le rapportΓ/E, où le paramètreénergétiqueΓ est la largeur de bande électronique du pontconsidéré comme un milieu continu (fig. 3), donnée par :
© 2001 CNRC Canada
124 Can. J. Physiol. Pharmacol. Vol. 79, 2001
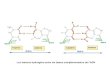
Fig. 1. Diagramme schématisant les énergies des orbitales dupont moléculaire (ou milieu intermédiaire). Les niveaux D et Areprésentent respectivement les énergies du donneur et del’accepteur. BV et BC : bande de valence (bande des orbitalesoccupées) et bande de conduction (bande des orbitalesinoccupées) du pont.∆EV et ∆EC représentent les gapsénergétiques entre l’énergie des sites donneur et accepteur et,respectivement, le haut de la BV et le bas de la BC.Vij est lecouplage électronique direct entre les orbitalesi et j du pont. Lestrois cas schématisés sur la figure correspondent aux régimessuivants (par rapport à la BC) : (a) large gap, (b) faible gap et(c) régime d’injection. D’après Priyadarshy et al. (1998).

[9] Γ = 8πρV0β–3
où ρ est la densité d’orbitales (nombre d’orbitales/Å3). Pourles deux molécules d’ADN ici étudiées,Γ = –2,5 eV, valeurcalculée à partir d’une densité de sites égale à 0,07 orbit-ales/Å3 que nous avons estimée à l’aide d’un modèle d’ADNproposé par Charlton et al. (1989).
Résultats et discussion
Dans toute notre étude, la position du site I est fixée(arbitrairement) sur celle du carbone en position 6 de lapremière base (ici, une cytosine) de l’extrêmité 5′ desmolécules d’ADN considérées, alors que celle du site F peutvarier, à volonté, le long des positions des autres atomes(fig. 2). À l’aide de l’équation 4, nous avons calculénumériquement le couplage électroniqueVS en fonction dela distance,dIF, séparant les sites I et F, pour différentesvaleurs du gap énergétique, à savoir :E = ±2, ±4 et ±10 eV.La figure 4 montre, à titre d’exemple, les résultats obtenuspour le système moléculaire 4GAT. Comme on peut le
remarquer sur cette figure, les valeurs deVS dans le cas dutransfert d’électron (V0/E < 0) sont de plusieurs ordres degrandeur supérieures à celles correspondant au transfert detrou (V0/E > 0). On peut noter aussi que la pente (βeff/2) descourbes représentant ln|VS/VDIVFA| en fonction dedIF, ainsique χ, le coefficient de corrélation linéaire, varientdifféremment avec le gap énergétique selon que l’on a af-faire à un transfert d’électron ou de trou. Par ailleurs, dans lecas du transfert de trou, des changements dans le signe deVSsont observés avec la distance : ce phénomène, quis’accompagne de réductions dramatiques de |VS|, trouve sonorigine dans les interférences destructives entre cheminsempruntés par la particule, comme il a été discuté en détaildans d’autres publications (Lopez-Castillo et al. 1993,1995a, 1995b; Filali-Mouhim et al. 1996; Wolfgang et al.1997).
La figure 5 compare, pour les deux molécules d’ADN iciétudiées, les valeurs deβeff/2β calculées numériquement enfonction de |Γ/E| avec celles obtenues à l’aide deséquations 7 et 8 dans le cadre de l’AMC. Comme on peut levoir, l’accord entre ces deux séries de valeurs est bon dans lalimite des larges gaps (ou bandes étroites) |Γ/E| << 1, où lesdeux systèmes moléculaires se comportent de la même façonque le milieu effectif « moyen », reflétant un phénomèned’auto-moyennage dans lequel seuls prédominent les effetsde densité (Lopez-Castillo et al. 1995a; Filali-Mouhim et al.
© 2001 CNRC Canada
Benrahmoune et al. 125
Fig. 2. Illustration de la séquence des bases complémentairespour les deux molécules d’ADN étudiées : (a) 1BBX (N = 758atomes) et (b) 4GAT (N = 823 atomes). D et A représententrespectivement les sites donneur et accepteur.
Fig. 3. Représentation des bandes de conduction et de valencedu pont, dans le cadre de l’approximation du milieu continu :(a) V0 < 0, les bandes électroniques s’élargissent vers le bas;(b) V0 > 0, les bandes électroniques s’élargissent vers le haut.Γreprésente la largeur de bande, donnée par l’équation 9, etEDA
est l’énergie des sites rédox (énergie de l’effet tunnel), prisecomme référence énergétique.

1996). En revanche, pour les grandes valeurs de |Γ/E| (~1),c’est-à-dire dans la limite des faibles gaps (ou bandes lar-ges), chacune des molécules se comporte de manière
« spécifique ». Dans cette limite, tous les détailsénergétiques et structuraux (séquence des bases comp-lémentaires, énergies des orbitales, couplages inter-sites, li-
© 2001 CNRC Canada
126 Can. J. Physiol. Pharmacol. Vol. 79, 2001
Fig. 4. Variation de ln|VS/VDIVFA| en fonction de la distancedIF séparant les sites initial (I) et final (F) du système moléculaire 4GAT,pour différentes valeurs du gap énergétique : |E| = 2 eV (a, d), |E| = 4 eV (b, e) et |E| = 10 eV (c, f). Les figures (a, b, c) correspon-dent à des transferts d’électron, alors que les figures (d, e, f) sont associées à des transferts de trou. Chaque point correspond à uneposition différente du site F le long de la molécule. (Õ) régression linéaire donnant les valeurs numériques deβeff/2 (VS est, d’aprèsles équations 1 à 3,proportionnel à exp[–(βeff/2)dIF]); χ est le coefficient de corrélation linéaire.

aisons hydrogène, couplages « à travers les liaisons », etc.)du pont intervenant devraient alors être pris en considérationafin d’obtenir une description adéquate deVS. On voit doncque le rapport Γ/E se présente comme un paramètreuniversel, contrôlant la valeur deβeff. L’existence d’un tel« paramètre de contrôle » a aussi été avancée dans le cas desTC dans les protéines (Lopez-Castillo et al. 1997) et certainssystèmes unidimensionnels (Evenson et Karplus 1992;Lopez-Castillo et al. 1993).
La distinction quantitative entre les valeurs deβeff misesen jeu lors d’un transfert de trou et d’un transfert d’électronpeut s’expliquer en termes de structure de bandesélectroniques (fig. 3). En effet, d’après l’équation 9, le sensde l’élargissement des bandes de valence et de conductiondu milieu intermédiaire se fait selon le signe deV0 : ainsi,selon les conventions de signe utilisées à la figure 3a, V0 < 0correspond à un élargissement des bandes vers le bas. Dansces conditions, le gap énergétique reste inchangé dans le casoù V0/E > 0 (transfert de trou), mais il est diminué lorsqueV0/E < 0 (transfert d’électron). L’effet tunnel étant d’autantplus probable que le gap énergétique est plus faible, le TCest ainsi facilité dans ce dernier cas : la valeur deβeff est, parconséquent, plus petite pour un transfert d’électron que pourcelui d’un trou. Par ailleurs, les effets de résonance se fontsentir plus précocement aux faibles gaps dans le cas oùV0/E < 0 que dans celui oùV0/E > 0, expliquant ainsi la dis-persion importante observée pourVS et βeff dans le cas dutransfert d’électron (figs. 4a et 5). Bien entendu, ces conclu-sions concernant la distinction entre les transferts de trou etd’électron devraient être inversées dans le cas où l’on auraitadoptéV0 > 0 : V0/E < 0 et V0/E > 0 correspondraient alorsrespectivement au transfert du trou et de l’électron.
Nos résultats montrent qu’il est possible d’utiliser leformalisme de l’AMC pour estimer la valeur deβeff à partirdu gap énergétique (ou bien l’inverse), dans la mesure
cependant où le couplage électronique est dominé par letransfert d’une seule particule : l’électron ou le trou. Lestravaux développés récemment par Priyadarshy et al. (1996a,1996b) nous offrent la possibilité d’obtenir une telle estima-tion. En effet, dans le cadre du mécanisme de superéchange età partir d’Hamiltoniens électroniques modèles, ces auteurs ontcalculé, pour trois systèmes expérimentaux d’ADN, desvaleurs deβeff égales à 1,2 et 1,6 Å–1. Si l’on se réfère à notrefigure 5, de telles valeurs deβeff correspondraient à untransfert d’électron. En adoptant alors les valeurs du gapénergétique (4 à 5 eV) données par ces mêmes auteurs,l’application des équations 8 et 9, avecβ = 1,7 Å–1 et Γ =–2,5 eV, conduisent àβeff = 1,6 à 1,8 Å–1, estimations dontl’ordre de grandeur est en bon accord, tant avec les valeurscalculées par Priyadarshy et al. (1996a, 1996b) qu’aveccelles trouvées expérimentalement.
Dans la région |Γ/E| << 1, notre modèle pourrait donc, enparticulier, décrire raisonnablement bienVS pour desmolécules d’ADN représentées par des Hamiltoniensélectroniques, tels que ceux employés par Priyadarshy et al.(1996a, 1996b), qui sont construits à partir d’unemodélisation plus réaliste que celle envisagée dans le présenttravail. La raison essentielle à cela réside dans le fait que no-tre modèle prend en compte correctement le caractèretridimensionnel du couplage électronique via les interactionsinter-sites « à travers l’espace » données par l’équation 6.Ainsi, même si chacune de ces interactions, par suite de leurnature à courte portée, contribue peu par son intensité aucouplage électronique total, il n’en reste pas moins qu’ellessont, dans leur ensemble, si nombreuses qu’il est nonseulement impossible de les négliger, mais égalementconcevable qu’elles puissent, dans certaines circonstances,prédominer (Lopez-Castillo et Jay-Gerin 1996).
Bien que les changements de signe deVS mentionnés plushaut dans le cas du transfert de trou (V0/E > 0) n’aient pasencore été mis en évidence expérimentalement, de tellesvariations ont néanmoins été observées théoriquement dansl’étude du TC, à longue distance, à travers des oligopeptides(Wolfgang et al. 1997), sous l’effet d’un changement de con-formation initié par simulation de dynamique moléculaire.Ce phénomène suscite actuellement un vif intérêt parmi leschercheurs qui s’intéressent aux transferts de chargescontrôlés, ces derniers constituant les processus élémentairesdes futurs dispositifs moléculaires (Jortner et Ratner 1997)qui, on le sait, incluent l’ADN (Fink et Schönenberger1999).
Conclusion
Nous avons étudié, dans le cadre du mécanisme desuperéchange, l’influence du gap énergétique surβeff dansl’ADN. Comme le couplage électroniqueVS est une fonctioncomplexe d’un grand nombre de paramètres d’originesénergétique et structurale, on s’est limité dans ce travail àl’étude systématique de ce paramètre clé. Les résultatsnumériques concernant les valeurs deβeff obtenues ici pourdeux molécules différentes d’ADN ont été analysés ens’appuyant sur la théorie de l’« approximation du milieucontinu ». Nous trouvons que, dans la région des larges gaps(|Γ/E| << 1), notre modèle de couplage « à travers l’espace »
© 2001 CNRC Canada
Benrahmoune et al. 127
Fig. 5. βef f /2β en fonction du paramètre de contrôle |Γ/E|.Résultats numériques pour les systèmes moléculaires 1BBX (d)et 4GAT (s); (- -) approximation du milieu continu pour letransfert de trou (éq. 7); (Õ) approximation du milieu continupour le transfert d’électron (éq. 8).

permet de reproduire les bons ordres de grandeur des valeursde βeff rapportées expérimentalement ou calculées pard’autres approches théoriques. Cependant, dans la région desfaibles gaps (|Γ/E| ~ 1), ce modèle apparaît insuffisant pourobtenir une bonne description deVS. Dans le but d’affinerquantitativement celui-ci, nous nous proposons donc, dansune étape subséquente, d’étudier en détail différents types destructures d’ADN et de prendre en considération untraitement plus réaliste des orbitalesπ (qui sont remplacéesici par des orbitaless), ainsi que la présence simultanée descouplages « à travers l’espace » et « à travers les liaisons »(développement en cours).
Remerciements
Ce travail a été réalisé grâce au soutien financier desInstituts de recherche en santé du Canada (subvention no
MT-14617).
Bibliographie
Ames, B.N., Shigenaga, M.K., et Hagen, T.M. 1993. Oxidants, an-tioxidants, and the degenerative diseases of aging. Proc. Natl.Acad. Sci. U.S.A.90 : 7915–7922.
Atherton, S.J., et Beaumont, P.C. 1995. Photoinduced electrontransfer reactions between copper ions and ethidium bromide inpolynucleotides. J. Phys. Chem.99 : 12 025 – 12 029.
Barbara, P.F., et Olson, E.J.C. 1999. Experimental electron transfer ki-netics in a DNA environment. Adv. Chem. Phys.107 : 647–676.
Beratan, D.N., et Skourtis, S.S. 1998. Protein-mediated electrontransfer: pathways, orbital interactions, and contact maps. Struc-ture–function relations for protein electron transfer.Dans Bio-logical electron transfer chains: genetics, composition and modeof operation.Éditeurs :G.W. Canters et E. Vijgenboom. KluwerAcademic Publishers, Dordrecht. NATO Science Series C,512 :9–27.
Beratan, D.N., Priyadarshy, S., et Risser, S.M. 1997. DNA: insula-tor or wire? Chem. Biol.4 : 3–8.
Brun, A.M., et Harriman, A. 1992. Dynamics of electron transferbetween intercalated polycyclic molecules: effect of interspersedbases. J. Am. Chem. Soc.114 : 3656–3660.
Brun, A.M., et Harriman, A. 1994. Energy- and electron-transferprocesses involving palladium porphyrins bound to DNA. J.Am. Chem. Soc.116 : 10 383 – 10 393.
Cadet, J. 1994. DNA damage caused by oxidation, deamination, ul-traviolet radiation and photoexcited psoralens.Dans DNA ad-ducts: identification and biological significance.Éditeurs : K.Hemminki, A. Dipple, D.E.G. Shuker, F.F. Kadlubar, D.Segerbäck et H. Bartsch. International Agency for Research onCancer, Lyon, France. IARC Scientific Publications no 125.pp. 245–276.
Charlton, D.E., Nikjoo, H., et Humm, J.L. 1989. Calculation of ini-tial yields of single- and double-strand breaks in cell nucleifrom electrons, protons and alpha particles. Int. J. Radiat. Biol.56 : 1–19.
Demple, B., et Harrison, L. 1994. Repair of oxidative damage toDNA: enzymology and biology. Annu. Rev. Biochem.63 :915–948.
Evenson, J.W., et Karplus, M. 1992. Effective coupling in bridgedelectron transfer molecules: computational formulation and ex-amples. J. Chem. Phys.96 : 5272–5278.
Evenson, J.W., et Karplus, M. 1993. Effective coupling in biologi-cal electron transfer: exponential or complex distance depend-ence? Science (Washington, D.C.),262 : 1247–1249.
Faraggi, M., Ferradini, C., et Jay-Gerin, J.-P. 1995. Migration etdevenir des charges formées initialement dans l’ADN irradié :une synthèse. New J. Chem.19 : 1203–1215.
Felts, A.K., Pollard, W.T., et Friesner, R.A. 1995. Multilevelredfield treatment of bridge-mediated long-range electron trans-fer: a mechanism for anomalous distance dependence. J. Phys.Chem.99 : 2929–2940.
Filali-Mouhim, A., Lopez-Castillo, J.-M., Plante, I.L., et Jay-Gerin, J.-P. 1996. Superexchange coupling and electron transferin homogeneous three-dimensional systems: site and bond disor-ders. J. Phys. Chem.100 : 12 311 – 12 320.
Fink, H.-W., et Schönenberger, C. 1999. Electrical conductionthrough DNA molecules. Nature (London),398 : 407–410.
Frenkel, K. 1992. Carcinogen-mediated oxidant formation and oxi-dative DNA damage. Pharmacol. Ther.53 : 127–166.
Fukui, K., et Tanaka, K. 1998. Distance dependence ofphotoinduced electron transfer in DNA. Angew. Chem. Int. Ed.Engl. 37 : 158–161.
Giese, B., Wessely, S., Spormann, M., Lindemann, U., Meggers,E., et Michel-Beyerle, M.E. 1999. On the mechanism of long-range electron transfer through DNA. Angew. Chem. Int. Ed.Engl. 38 : 996–998.
Gray, H.B., et Winkler, J.R. 1996. Electron transfer in proteins.Annu. Rev. Biochem.65 : 537–561.
Gros, F., Tubiana, M., Sarasin, A., Masse, R., Moustacchi, E.,Flüry-Hérard, A., et Rosa, J. (Éditeurs). 1999. Colloque« Risques cancérogènes dus aux rayonnements ionisants. » C.R.Acad. Sci. Paris, Sér. III,322 : 81–256.
Gruschus, J.M., et Kuki, A. 1993. New Hamiltonian model forlong-range electronic superexchange in complex molecularstructures. J. Phys. Chem.97 : 5581–5593.
Henderson, P.T., Jones, D., Hampikian, G., Kan, Y., et Schuster,G.B. 1999. Long-distance charge transport in duplex DNA: thephonon-assisted polaron-like hopping mechanism. Proc. Natl.Acad. Sci. U.S.A.96 : 8353–8358.
Holmlin, R.E., Dandliker, P.J., et Barton, J.K. 1997. Charge trans-fer through the DNA base stack. Angew. Chem. Int. Ed. Engl.36 : 2714–2730.
Jortner, J., et Ratner, M. (Éditeurs). 1997. Molecular electronics.Blackwell Scientific Publications Ltd., London.
Jortner, J., Bixon, M., Langenbacher, T., et Michel-Beyerle, M.E.1998. Charge transfer and transport in DNA. Proc. Natl. Acad.Sci. U.S.A.95 : 12 759 – 12 765.
Kelley, S.O., et Barton, J.K. 1999. Electron transfer between bases indouble helical DNA. Science (Washington, D.C.),283 : 375–381.
Kelley, S.O., Holmlin, R.E., Stemp, E.D.A., et Barton, J.K. 1997.Photoinduced electron transfer in ethidium-modified DNA du-plexes: dependence on distance and base stacking. J. Am. Chem.Soc.119 : 9861–9870.
Kuki, A. 1991. Electronic tunneling paths in proteins.DansLong-range electron transfer in biology.Éditeur : G.A. Palmer. Struct.Bonding,75 : 49–83.
Lewis, F.D., Wu, T., Zhang, Y., Letsinger, R.L., Greenfield, S.R., etWasielewski, M.R. 1997. Distance-dependent electron transferin DNA hairpins. Science (Washington, D.C.),277 : 673–676.
Loft, S., et Poulsen, H.E. 1996. Cancer risk and oxidative DNAdamage in man. J. Mol. Med.74 : 297–312.
Lopez-Castillo, J.-M., et Jay-Gerin, J.-P. 1996. Superexchange cou-pling and electron transfer in large molecules: through-spaceand through-bond interactions. J. Phys. Chem.100 : 14 289 –14 297.
© 2001 CNRC Canada
128 Can. J. Physiol. Pharmacol. Vol. 79, 2001

© 2001 CNRC Canada
Benrahmoune et al. 129
Lopez-Castillo, J.-M., Filali-Mouhim, A., et Jay-Gerin, J.-P. 1993.Superexchange coupling and non-nearest-neighbor interactionsin electron transfers. J. Phys. Chem.97 : 9266–9269.
Lopez-Castillo, J.-M., Filali-Mouhim, A., Plante, I.L., et Jay-Gerin, J.-P. 1995a. Superexchange coupling and electron trans-fer in homogeneous three-dimensional systems: the continuous-medium approximation. J. Phys. Chem.99 : 6864–6875.
Lopez-Castillo, J.-M., Filali-Mouhim, A., Plante, I.L., et Jay-Gerin, J.-P. 1995b. Superexchange coupling and electron trans-fers in large three-dimensional disordered media. Chem. Phys.Lett. 239 : 223–229.
Lopez-Castillo, J.-M., Filali-Mouhim, A., Nguyen Van Binh-Otten,É., et Jay-Gerin, J.-P. 1997. Electron transfer in proteins: struc-tural and energetic control of the electronic coupling. J. Am.Chem. Soc.119 : 1978–1980.
Marcus, R.A., et Sutin, N. 1985. Electron transfers in chemistryand biology. Biochim. Biophys. Acta,811 : 265–322.
Meade, T.J., et Kayyem, J.F. 1995. Electron transfer through DNA:site-specific modification of duplex DNA with ruthenium donorsand acceptors. Angew. Chem. Int. Ed. Engl.34 : 352–354.
Meggers, E., Michel-Beyerle, M.E., et Giese, B. 1998. Sequencedependent long range hole transport in DNA. J. Am. Chem. Soc.120 : 12 950 – 12 955.
Moser, C.C., Keske, J.M., Warncke, K., Farid, R.S., et Dutton, P.L.1992. Nature of biological electron transfer. Nature (London),355 : 796–802.
Murphy, C.J., Arkin, M.R., Jenkins, Y., Ghatlia, N.D., Bossmann,S.H., Turro, N.J., et Barton, J.K. 1993. Long-range photo-induced electron transfer through a DNA helix. Science (Wash-ington, D.C.),262 : 1025–1029.
Netzel, T.L. 1998. Present status and future directions of researchin electron-transfer mediated by DNA. J. Biol. Inorg. Chem.3 :210–214.
Núñez, M.E., Hall, D.B., et Barton, J.K. 1999. Long-range oxida-tive damage to DNA: effects of distance and sequence. Chem.Biol. 6 : 85–97.
Olson, E.J.C., Hu, D., Hörmann, A., et Barbara, P.F. 1997. Quanti-tative modeling of DNA-mediated electron transfer betweenmetallointercalators. J. Phys. Chem. B,101 : 299–303.
O’Neill, P., et Fielden, E.M. 1993. Primary free radical processesin DNA. Adv. Radiat. Biol.17 : 53–120.
Pezeshk, A., Symons, M.C.R., et McClymont, J.D. 1996. Electronmovement along DNA strands: use of intercalators and electronparamagnetic resonance spectroscopy. J. Phys. Chem.100 :18 562 – 18 566.
Priyadarshy, S., Beratan, D.N., et Risser, S.M. 1996a. DNA dou-ble-helix-mediated long-range electron transfer. Int. J. QuantumChem. Quantum Biol. Symp.23 : 65–71.
Priyadarshy, S., Risser, S.M., et Beratan, D.N. 1996b. DNA is not amolecular wire: protein-like electron-transfer predicted for an ex-tendedπ-electron system. J. Phys. Chem.100 : 17 678 – 17 682.
Priyadarshy, S., Risser, S.M., et Beratan, D.N. 1998. DNA-mediated electron transfer. J. Biol. Inorg. Chem.3 : 196–200.
Ratner, M. 1999. Electronic motion in DNA. Nature (Washington,D.C.), 397 : 480–481.
Risser, S.M., Beratan, D.N., et Meade, T.J. 1993. Electron transferin DNA: predictions of exponential growth and decay of cou-pling with donor–acceptor distance. J. Am. Chem. Soc.115 :2508–2510.
Sarasin, A. 1997. Lésions sur l’ADN : mécanismes de reconnais-sance et de réparation. Bull. Cancer,84 : 467–472.
Skourtis, S.S., et Beratan, D.N. 1999. Theories of structure–func-tion relationships for bridge-mediated electron transfer reac-tions. Adv. Chem. Phys.106 : 377–452.
Stemp, E.D.A., et Barton, J.K. 1996. Electron transfer betweenmetal complexes bound to DNA: is DNA a wire? Metal IonsBiol. Syst. 33 : 325–365.
Taubes, G. 1997. Double helix does chemistry at a distance — buthow? Science (Washington, D.C.),275 : 1420–1421.
Tubiana, M., Dutreix, J., et Wambersie, A. 1990. Introduction toradiobiology. Taylor & Francis, London.
Ulstrup, J. 1979. Charge transfer processes in condensed media.Lecture notes in chemistry, Vol. 10. Spinger-Verlag, Berlin.
Warman, J.M., de Haas, M.P., et Rupprecht, A. 1996. DNA: a mo-lecular wire? Chem. Phys. Lett.249 : 319–322.
Wolfgang, J., Risser, S.M., Priyadarshy, S., et Beratan, D.N. 1997.Secondary structure conformations and long range electronic in-teractions in oligopeptides. J. Phys. Chem. B,101 : 2986–2991.