Embed Size (px)
Citation preview

Adrian Lister
Critical reappraisal of the Middle Pleistocene deer species"Cervus" elaphoides Kaklke.In: Quaternaire - Volume 1 - Numéro 3-4 - 1990. pp. 175-192.
RésuméL'espèce "Cervus" elaphoides Kahlke 1960 fut fondée sur la base des bois provenant des Sables de Mosbach. Une étudenouvelle détaillée montre une continuité morphologique totale avec les restes, plus grands, du Cervus elaphus acoronatus dumême gisement. Cela, en plus des pédicules allongés du "C." elaphoides, suggère que les bois en question proviennent desjeunes individus de Ce. acoronatus, donc que "C." elaphoides soit un synonyme de ce dernier. Des ossements et des dents d'unpetit cervidé à Mosbach sont d'attribution incertaine, mais appartiennent peut-être au daim. Des fossiles provenant d'au moinsdouze autres gisements européens ont été attribués a "C." elaphoides, mais en chaque cas cela peut s'interpréter soit commedes jeunes C elaphus, soit comme une autre espèce de cervidé. Il est probable que parmi ce materiel il existe des taxa jusqu'àprésent indéfinis.
AbstractThe species "Cervus" elaphoides Kahlke 1960 was founded on antlers from the Mosbach Sands. A detailed restudy of thismaterial demonstrates complete morphological continuity with specimens of the larger Cervus elaphus acoronatus from the samesite. Together with the long pedicles of "C." elaphoides, this suggests that the antlers in question are young individuals of Ce.acoronatus, and that "C." elaphoides is therefore a junior synonym of the latter. Bones and teeth of a small cervid from Mosbachare of uncertain attribution, but may pertain to Dama dama. Fossils from at least twelve other European sites have been referredto "C." elaphoides, but in each case they can be interpreted as either young C. elaphus, or as pertaining to other cervid species.It is probable that among this material are some as yet undefined taxa.
Citer ce document / Cite this document :
Lister Adrian. Critical reappraisal of the Middle Pleistocene deer species "Cervus" elaphoides Kaklke. In: Quaternaire - Volume1 - Numéro 3-4 - 1990. pp. 175-192.
doi : 10.3406/quate.1990.1935
http://www.persee.fr/web/revues/home/prescript/article/quate_1142-2904_1990_num_1_3_1935

Quaternaire, 3-4, 1990, 175-192
CRITICAL REAPPRAISAL OF THE MIDDLE PLEISTOCENE DEER SPECIES ELAPHOIDES KAHLKE
'CERVUS
Adrian LISTER*
ABSTRACT
The species "Cervus" elaphoides Kahlke 1960 was founded on antlers from the Mosbach Sands. A detailed restudy of this material demonstrates complete morphological continuity with specimens of the larger Cervus elaphus acoronatus from the same site. Together with the long pedicles of "C." elaphoides, this suggests that the antlers in question are young individuals of Ce. acoronatus, and that "C." elaphoides is therefore a junior synonym of the latter. Bones and teeth of a small cervid from Mosbach are of uncertain attribution, but may pertain to Dama dama. Fossils from at least twelve other European sites have been referred to "C." elaphoides, but in each case they can be interpreted as either young C. elaphus, or as pertaining to other cervid species. It is probable that among this material are some as yet undefined taxa. Key-words : Antlers, Middle Pleistocene, Mosbach, Synonymy, "Cervus" elaphoides, Cervus elaphus acoronatus.
RÉSUMÉ
RÉÉVALUATION CRITIQUE DU CERVIDÉ PLEISTOCENE MOYEN "CERVUS" ELAPHOIDES KAHLKE L'espèce "Cervus" elaphoides Kahlke 1960 fut fondée sur la base des bois provenant des Sables de Mosbach. Une étude nouvelle
détaillée montre une continuité morphologique totale avec les restes, plus grands, du Cervus elaphus acoronatus du même gisement. Cela, en plus des pédicules allongés du "C." elaphoides, suggère que les bois en question proviennent des jeunes individus de Ce. acoronatus, donc que "C." elaphoides soit un synonyme de ce dernier. Des ossements et des dents d'un petit cervidé à Mosbach sont d'attribution incertaine, mais appartiennent peut-être au daim. Des fossiles provenant d'au moins douze autres gisements européens ont été attribués a "C." elaphoides, mais en chaque cas cela peut s'interpréter soit comme des jeunes C elaphus, soit comme une autre espèce de cervidé. Il est probable que parmi ce materiel il existe des taxa jusqu'à présent indéfinis. Mots-clés : Bois, Pleistocene moyen, Mosbach, Synonymie, "Cervus" elaphoides, Cervus elaphus acoronatus.
1. INTRODUCTION
The deer species "Cervus" elaphoides was founded by Kahlke (1960) on the basis of fossils from the Mosbach Sands. Since then, deer remains from several other Pleistocene localities have been referred to this species, and some authors (e.g. Heintz and Poplin, 1974; Crégut, 1979) have regarded it as a valuable stratigraphie marker for the early Middle Pleistocene. The fossils in question are believed to
correspond to a species approximately the size of a modern European fallow deer, Dama dama (L.). Some authors refer to such deer species as "medium sized", but in this paper they will be termed "small sized" because the chief comparison will be with the larger red deer (Cervus elaphus L.).
A consideration of the validity or otherwise of this species must begin with an analysis of the type sample, from Mosbach. The fluviatile sand deposits of the River Rhine at Mosbach, Germany, have yielded one of the most extensive collections of early Middle
Department of Zoology, Cambridge CB2 3EJ, England.
176
Pleistocene vertebrate fossils in Europe. The sequence of deposits covers an appreciable period of time, but the very large majority of fossils come from the unit known as Mosbach III, of Cromerian or early Elsterian age, c. 700,000-550,000 BP (Brii- ning, 1978; Igel, 1985; Berggren et a/., 1980). A much smaller amount of material comes from the late Lower Pleistocene unit Mosbach I. Enough fossils are provenanced to their precise horizon that separate faunal lists for these units have been established (Briining, 1978). All provenanced finds of the two species to be considered here, Cervus elaphus and "Cervus" elaphoides, are known to have come from Mosbach III. Thus, although many of the individual fossils of these species are not strictly stratified, it can safely be assumed that all or almost all of them pertain to the Mosbach III fauna.
Since Kahlke's (1960) study, a large amount of newly-discovered fossil material from Mosbach has been added to the collections. The present study is based on a detailed re-examination of all currently available remains from Mosbach, as well as from most of the other sites from which "Cervus"
phoides has been recorded. The following abbreviations for museum collections will be used : BM, Basle Museum; DM, Darmstadt Museum; IQW, Institut fur Quartârpalâontologie, Weimar; MNHN, Muséum National d'Histoire Naturelle, Paris; NHM, Natural History Museum, London; NMM, Naturhis- torisches Museum, Mainz; SMF, Senckenberg Museum, Frankfurt; WM, Wiesbaden Museum.
2. CERVUS ELAPHUS ACORONATUS AND "CERVUS" ELAPHOIDES FROM MOSBACH
a) Historical introduction
Red deer (Cervus elaphus) is one of the dominant large mammal species from the Mosbach Sands, represented by many hundreds of antlers, bones and teeth. The lower part of each antler bears the characteristic red deer feature of two basal tines, the
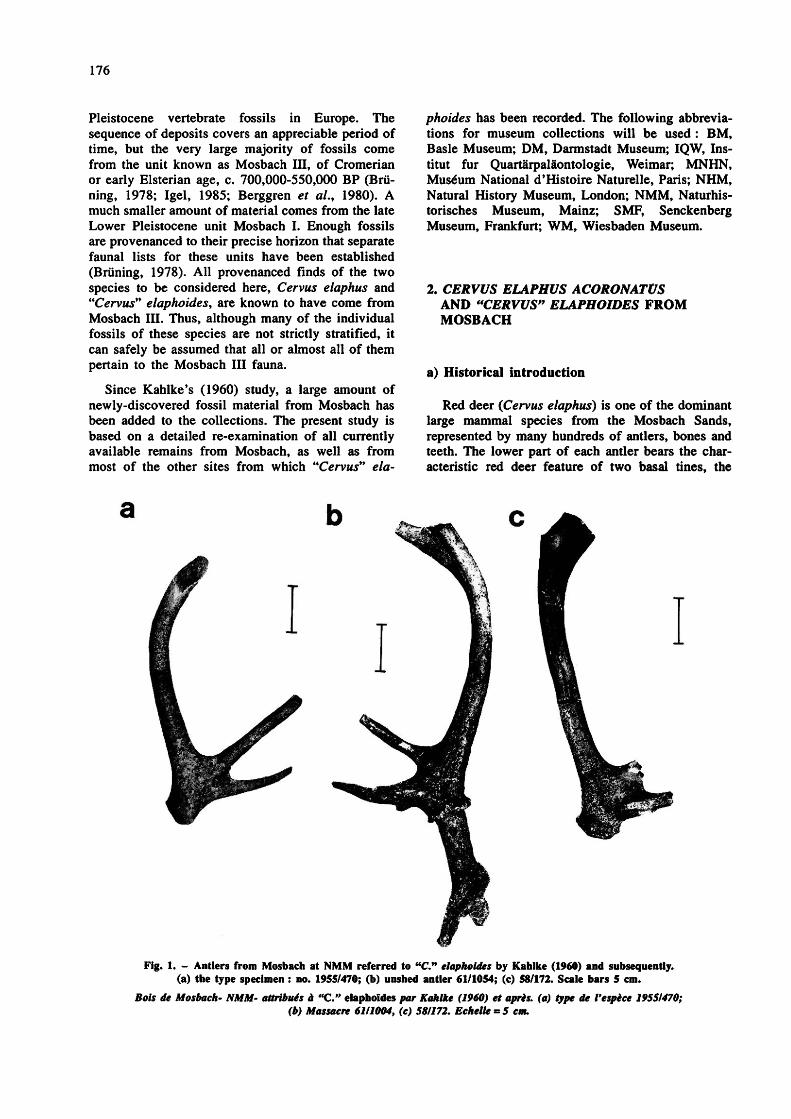
Fig. 1. - Antlers from Mosbach at NMM referred to "C." elaphoides by Kahlke (1960) and subsequently. (a) the type specimen : no. 1955/470; (b) unshed antler 61/1054; (c) 58/172. Scale bars 5 cm.
Bois de Mosbach- NMM- attribués à "C." elaphoïdes par Kahlke (1960) et après, (a) type de l'espèce 19551470; (b) Massacre 6111004, (c) 581172. Echelle = 5 cm.

177
brow and bez. The upper part of the antler is similar to that of red deer today inhabiting China (e.g. Ce. wallichi), in that it terminates, above the third (trez) tine, in a simple two-pointed fork oriented transverse to the long axis of the body. Unlike Recent European red deer, a multi-pointed "crown" does not form at the top of the antler, even in large, adult individuals. This lead Beninde (1937) to name the Mosbach deer Cervus acoronatus. Subsequent authors (e.g. Kurtén, 1968; Lister, 1986), recognising that the difference between the early Middle Pleistocene and modern red deer is no greater than that between the different modern subspecies, have relegated acoronatus to subspecific status within C. elaphus.
Bachofen-Echt (1929) was the first to suggest that the red-deer-like remains from Mosbach could be divided into two groups on the basis of size, and he believed that these corresponded to a small and a large "race" of red deer. The antlers of the two "races" were of similar structure, including the aco- ronate terminal fork, but differed in size, the larger (and more abundant) "race" having rose circumferences between 22 and 32 cm, the smaller "race" between 11 and 14 cm. In addition, cervine limb- bone remains from the site were found to fall into two distinct size categories, and these were referred to the two "races" of red deer.
Beninde (1937) accepted the existence of two size categories among these remains, but rejected Bacho- fen-Echt's suggestion that they represented "races" of a single species, since two races could not co-exist sympatrically without introgression. He therefore suggested that they must be systematically distinct on at least the species level. He mentioned the alternative possibility that the large and small antlers could be older and younger individuals of the same species, but asserted, without giving reasons, that this was unlikely.
In 1960, Kahlke created a formal taxonomic division between the large and small Mosbach antlers. The larger category was regarded as true red deer, retaining the name Cervus acoronatus Beninde. The smaller category was considered a separate species of uncertain generic attribution, and was named by Kahlke "Cervus" elaphoides. Kahlke (1960) described the "C." elaphoides antlers as differing from C. acoronatus not only in size, but also in morphology. While sharing with C. acoronatus the double lower tine and transverse terminal fork, "C" elaphoides was thought to lack the middle (trez) tine characteristic of C. elaphus and C. acoronatus. In addition, the surface of the antlers was smooth, lacking the "pearling" often found on red deer antlers. The brow and bez tines were said to be rather straight, the beam upright, and the pedicles unusually long. Kahlke (1960) chose as the type specimen of "C." elaphoides an incomplete antler at NMM, no. 1955/470 (fig. 1 a), and referred nine further
partial antlers from Mosbach to this species, as well as an almost complete antler figured by Bachofen- Echt (1929, Plate 11, no. 8; here fig. 2) but subsequently lost. Kahlke also tentatively referred some dental material from Mosbach to "Cervus" elaphoides.
Fig. 2. - Antler from Mosbach referred to "C." elaphoides by Kahlke (1960), traced from Bachofen-Echt (1929,
fig. 11.8). Scale not given, but scale bar approx. 10 cm. Mosbach, bois attribué à "C." elaphoides par Kahlke (1960), d'après Bachofen-Echt (1929, fig. 11.8). L'Echelle n'est pas
donnée mais la barre doit approcher 10 cm.
b) Biometrical and morphological analysis of the Mosbach antlers.
In what follows, I attempt to show that the "small" and "large" Mosbach antlers are part of a single, continuous range of variation. The analysis is based on all antlers and partial antlers of C. acoronatus and "C." elaphoides conserved at NMM, DM, SMF and WM, a total of nearly 300 specimens. All measurements in this and subsequent sections are in millimetres unless otherwise indicated.
Antler size
The most suitable general index of antler size is the circumference of the beam at the base of the antler. This is preserved in the large majority of specimens, and adequately reflects general beam stoutness. Total antler length could have been measured in only a small minority of the fossils.
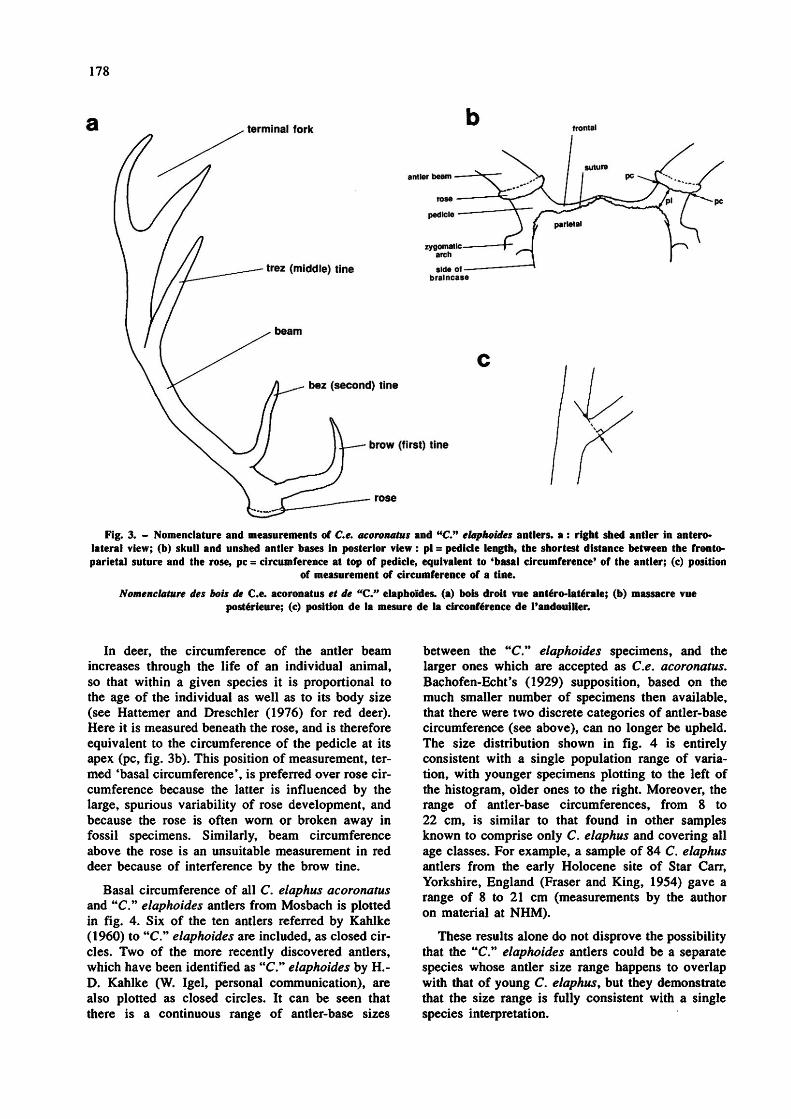
178
terminal fork
trez (middle) tine
frontal
antler beam pc
bez (second) tine
brow (first) tine
rose
Fig. 3. - Nomenclature and measurements of Ce. acoronatus and "C." elaphoides antlers, a : right shed antler in antero- lateral view; (b) skull and unshed antler bases in posterior view : pi = pedicle length, the shortest distance between the fronto- parietal suture and the rose, pc = circumference at top of pedicle, equivalent to 'basal circumference' of the antler; (c) position
of measurement of circumference of a tine. Nomenclature des bois de Ce. acoronatus et de "C." elaphoïdes. (a) bois droit vue antéro-latérale; (b) massacre vue
postérieure; (c) position de la mesure de la circonférence de l'andouiller.
In deer, the circumference of the antler beam increases through the life of an individual animal, so that within a given species it is proportional to the age of the individual as well as to its body size (see Hattemer and Dreschler (1976) for red deer). Here it is measured beneath the rose, and is therefore equivalent to the circumference of the pedicle at its apex (pc, fig. 3b). This position of measurement, termed 'basal circumference', is preferred over rose circumference because the latter is influenced by the large, spurious variability of rose development, and because the rose is often worn or broken away in fossil specimens. Similarly, beam circumference above the rose is an unsuitable measurement in red deer because of interference by the brow tine.
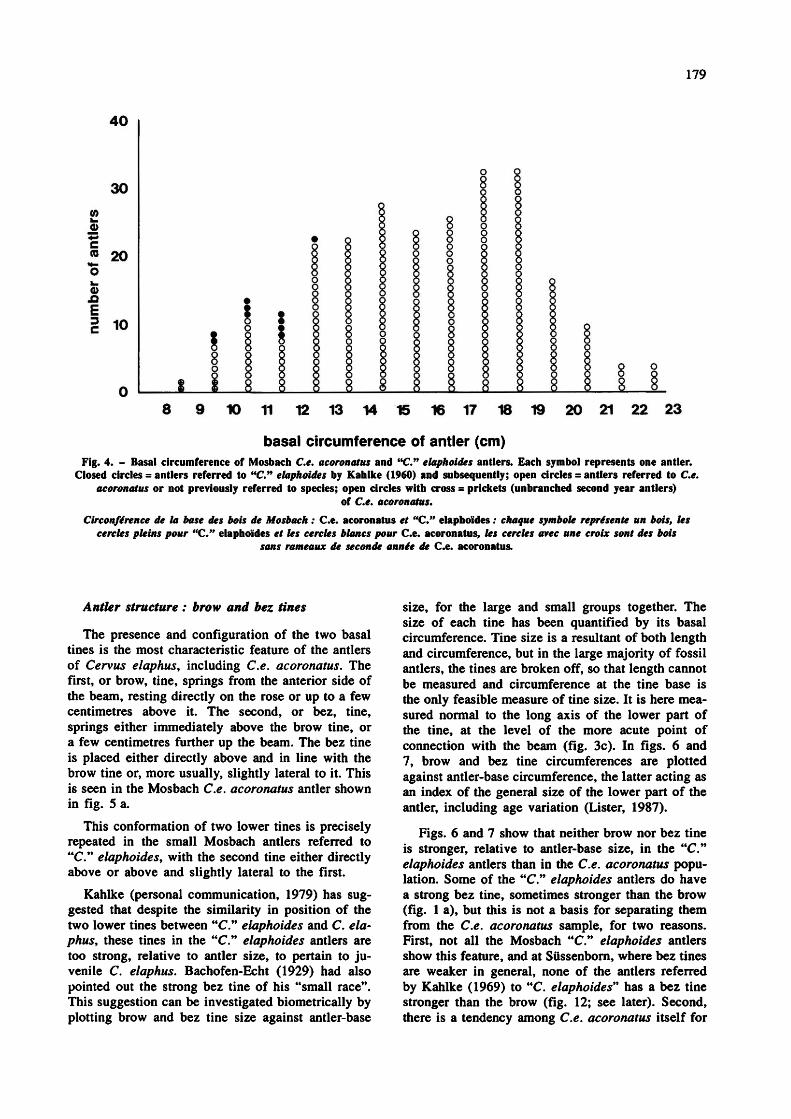
Basal circumference of all C. elaphus acoronatus and "C" elaphoides antlers from Mosbach is plotted in fig. 4. Six of the ten antlers referred by Kahlke (1960) to "C." elaphoides are included, as closed circles. Two of the more recently discovered antlers, which have been identified as C.n elaphoides by H.- D. Kahlke (W. Igel, personal communication), are also plotted as closed circles. It can be seen that there is a continuous range of antler-base sizes
between the "C." elaphoides specimens, and the larger ones which are accepted as Ce. acoronatus. Bachofen-Echt's (1929) supposition, based on the much smaller number of specimens then available, that there were two discrete categories of antler-base circumference (see above), can no longer be upheld. The size distribution shown in fig. 4 is entirely consistent with a single population range of variation, with younger specimens plotting to the left of the histogram, older ones to the right. Moreover, the range of antler-base circumferences, from 8 to 22 cm, is similar to that found in other samples known to comprise only C. elaphus and covering all age classes. For example, a sample of 84 C. elaphus antlers from the early Holocene site of Star Carr, Yorkshire, England (Fraser and King, 1954) gave a range of 8 to 21 cm (measurements by the author on material at NHM).
These results alone do not disprove the possibility that the "C." elaphoides antlers could be a separate species whose antler size range happens to overlap with that of young C. elaphus, but they demonstrate that the size range is fully consistent with a single species interpretation.
179
£
o
I C
40
30
20
10
8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
basal circumference of antler (cm) Fig. 4. - Basal circumference of Mosbach C.e. acoronatus and "C." elaphoides antlers. Each symbol represents one antler.
Closed circles = antlers referred to "C." elaphoides by Kahlke (1960) and subsequently; open circles = antlers referred to C.e. acoronatus or not previously referred to species; open circles with cross = prickets (unbranched second year antlers)
of C.e. acoronatus. Circonférence de la base des bois de Mosbach : Ce. acoronatus et "C" elaphoides ; chaque symbole représente un bois, les
cercles pleins pour "C." elaphoides et les cercles blancs pour Ce. acoronatus, les cercles avec une croix sont des bois sans rameaux de seconde année de Ce. acoronatus.
Antler structure : brow and bez tines
The presence and configuration of the two basal tines is the most characteristic feature of the antlers of Cervus elaphus, including C.e. acoronatus. The first, or brow, tine, springs from the anterior side of the beam, resting directly on the rose or up to a few centimetres above it. The second, or bez, tine, springs either immediately above the brow tine, or a few centimetres further up the beam. The bez tine is placed either directly above and in line with the brow tine or, more usually, slightly lateral to it. This is seen in the Mosbach C.e. acoronatus antler shown in fig. 5 a.
This conformation of two lower tines is precisely repeated in the small Mosbach antlers referred to "C." elaphoides, with the second tine either directly above or above and slightly lateral to the first.
Kahlke (personal communication, 1979) has suggested that despite the similarity in position of the two lower tines between "C." elaphoides and C. elaphus, these tines in the "C." elaphoides antlers are too strong, relative to antler size, to pertain to juvenile C. elaphus. Bachofen-Echt (1929) had also pointed out the strong bez tine of his "small race". This suggestion can be investigated biometrically by plotting brow and bez tine size against antler-base
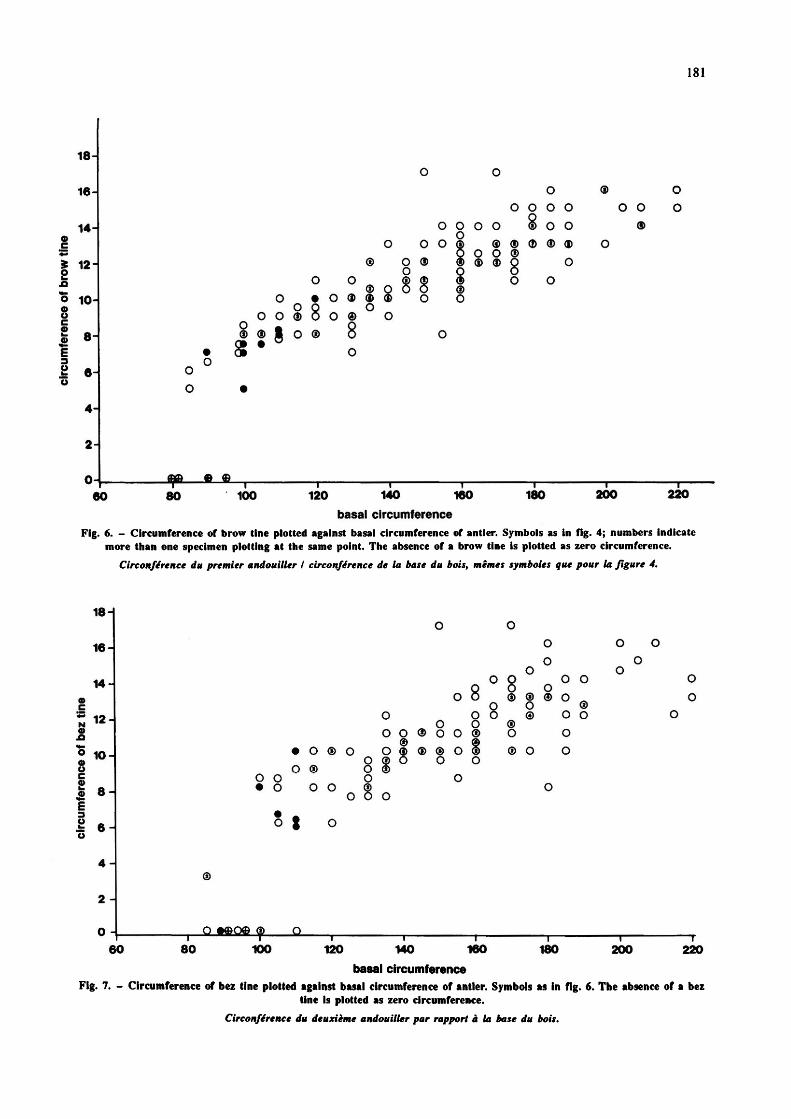
size, for the large and small groups together. The size of each tine has been quantified by its basal circumference. Tine size is a resultant of both length and circumference, but in the large majority of fossil antlers, the tines are broken off, so that length cannot be measured and circumference at the tine base is the only feasible measure of tine size. It is here measured normal to the long axis of the lower part of the tine, at the level of the more acute point of connection with the beam (fig. 3c). In figs. 6 and 7, brow and bez tine circumferences are plotted against antler-base circumference, the latter acting as an index of the general size of the lower part of the antler, including age variation (Lister, 1987).
Figs. 6 and 7 show that neither brow nor bez tine is stronger, relative to antler-base size, in the "C." elaphoides antlers than in the C.e. acoronatus population. Some of the "C." elaphoides antlers do have a strong bez tine, sometimes stronger than the brow (fig. 1 a), but this is not a basis for separating them from the C.e. acoronatus sample, for two reasons. First, not all the Mosbach "C." elaphoides antlers show this feature, and at Siissenborn, where bez tines are weaker in general, none of the antlers referred by Kahlke (1969) to "C. elaphoides" has a bez tine stronger than the brow (fig. 12; see later). Second, there is a tendency among C.e. acoronatus itself for
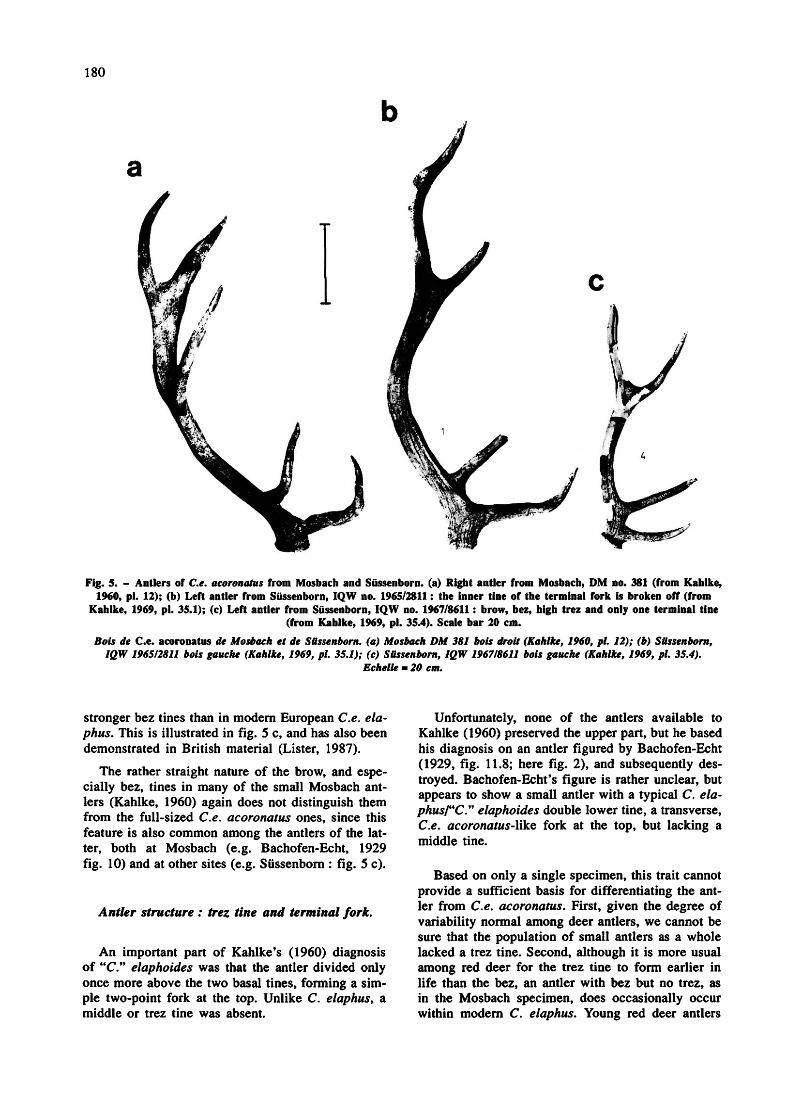
180
Fig. 5. - Antlers of Ce. acoronatus from Mosbach and Sussenborn. (a) Right antler from Mosbach, DM no. 381 (from Kahlke, 1960, pi. 12); (b) Left antler from Sùssenborn, IQW no. 1965/2811 : the inner tine of the terminal fork is broken off (from
Kahlke, 1969, pi. 35.1); (c) Left antler from Sussenborn, IQW no. 1967/8611 : brow, bez, high trez and only one terminal tine (from Kahlke, 1969, pi. 35.4). Scale bar 20 cm.
Bois de Ce. acoronatus de Mosbach et de Siissenborn. (a) Mosbach DM 381 bois droit (Kahlke, I960, pi. 12); (b) SUssenborn, IQW 196512811 bois gauche (Kahlke, 1969, pi 35.1); (c) Sussenborn, IQW 196718611 bois gauche (Kahlke, 1969, pi. 35.4).
Echelle = 20 cm.
stronger bez tines than in modem European Ce. ela- phus. This is illustrated in fig. 5 c, and has also been demonstrated in British material (Lister, 1987).
The rather straight nature of the brow, and especially bez, tines in many of the small Mosbach antlers (Kahlke, 1960) again does not distinguish them from the full-sized Ce. acoronatus ones, since this feature is also common among the antlers of the latter, both at Mosbach (e.g. Bachofen-Echt, 1929 fig. 10) and at other sites (e.g. Sussenborn : fig. 5 c).
Antler structure : trez tine and terminal fork.
An important part of Kahlke 's (1960) diagnosis of "C." elaphoides was that the antler divided only once more above the two basal tines, forming a simple two-point fork at the top. Unlike C. elaphus, a middle or trez tine was absent.
Unfortunately, none of the antlers available to Kahlke (1960) preserved the upper part, but he based his diagnosis on an antler figured by Bachofen-Echt (1929, fig. 11.8; here fig. 2), and subsequently destroyed. Bachofen-Echt's figure is rather unclear, but appears to show a small antler with a typical C. ela- phus/"C." elaphoides double lower tine, a transverse, Ce. acoronatus-like fork at the top, but lacking a middle tine.
Based on only a single specimen, this trait cannot provide a sufficient basis for differentiating the antler from Ce. acoronatus. First, given the degree of variability normal among deer antlers, we cannot be sure that the population of small antlers as a whole lacked a trez tine. Second, although it is more usual among red deer for the trez tine to form earlier in life than the bez, an antler with bez but no trez, as in the Mosbach specimen, does occasionally occur within modern C elaphus. Young red deer antlers
181
18
16
14
1 6 o
2
0-
®
O O O • O
® o
o o o o o
Q o o o o ® o o o 00® ®®®®® Q O O ® o ® ® ® ® o o
0 0 0 §® ® o o o ® o o
O o
s *#
o ® o 8
A A
o o o o
60 80 100 180 200 220 120 140 160 basal circumference
Fig. 6. - Circumference of brow tine plotted against basal circumference of antler. Symbols as in fig. 4; numbers indicate more than one specimen plotting at the same point. The absence of a brow tine is plotted as zero circumference.
Circonférence du premier andouiller I circonférence de la base du bois, mêmes symboles que pour la figure 4.
18
16
14
8H
S 6 o
0 -
o o • o o o
o o ® o o ® Q ® O o ® o
®
o
8
08 O O ® o o ® o
ô ô
O o
o o o ® ® ® o o
o o
© o o o o
o o o
o o
60 80 100 180 200 220 120 140 160 basal circumference
Fig. 7. - Circumference of bez tine plotted against basal circumference of antler. Symbols as in fig. 6. The absence of a bez tine is plotted as zero circumference.
Circonférence du deuxième andouiller par rapport à la base du bois.
182
with two lower tines and no further branching before a simple fork at the top have been figured by Ahlén (1965, figs. 29 and 78) from Sweden and Bachofen- Echt (1929, fig. 23) from the Carpathians.
The two antlers which have been referred to "C." elaphoides since Kahlke's (1960) study (see above) have an important bearing on this question. These specimens (fig. 1 b & c) preserve the major part of the beam, and although the distal tines are broken off, their bases can be seen. The two antlers share similar morphology towards their distal preserved end. In this region, the beam undergoes a slight outward twisting and flattening, above which a tine of circular cross-section originates in an anterior direction. The beam, antero-posteriorly flattened, is itself broken at this point, but had clearly continued a distal path. These details (outward twisting of the beam, flattening, anterior sprouting of a tine, continuation of the flattened beam) are repeated precisely, at a larger scale, in the numerous antlers of Ce. acoro- natus from Mosbach. In the better preserved specimens, the anterior tine is clearly seen to be the trez tine, and the flattened beam continues distally to form the final, transverse fork at the top. In the two small antlers, the rounded form and anterior position of the tine strongly suggest that this branching point represents the origin of a trez tine from the beam, rather than a transverse terminal fork. Further, the antero-posteriorly flattened cross-section of the broken end of the beam at this node implies that somewhat higher up, the beam had divided again, in a transverse plane, to form the terminal fork. This morphology agrees very closely with that of the full- sized Ce. acoronatus antlers. It also suggests that the lack of a trez tine on Bachofen-Echt's specimen, assuming the latter to be conspecific with the two more recently discovered antlers, represents intras- pecific variability rather than a species-diagnostic character.
Antler ornament
The type specimen of "C." elaphoides has a smooth surface, which does not appear to be the result of abrasion. However, several otherwise indistinguishable small antler bases (e.g. NMM 64/97 and 70/224) have clear longitudinal ridges and pearling. Among the larger, Ce. acoronatus antlers, there are many with the ridging and pearling typical of C. ela- phus, but some, which do not appear abraded, with smooth surfaces (e.g. NMM 64/1237). The presence of a smooth surface would therefore appear not to be a reliable feature for distinguishing "C." elaphoides.
Pedicle proportions
Pedicles are the bony outgrowths of the skull on which the antlers are borne. They are of particular
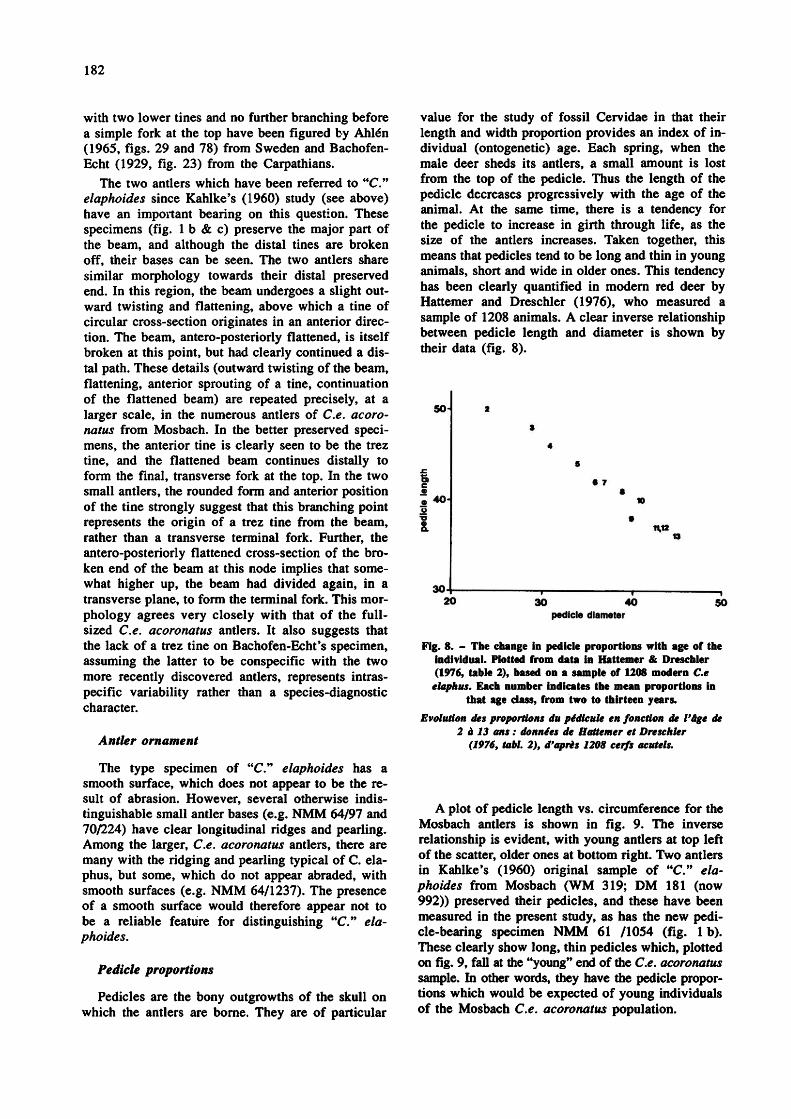
value for the study of fossil Cervidae in that their length and width proportion provides an index of individual (ontogenetic) age. Each spring, when the male deer sheds its antlers, a small amount is lost from the top of the pedicle. Thus the length of the pedicle decreases progressively with the age of the animal. At the same time, there is a tendency for the pedicle to increase in girth through life, as the size of the antlers increases. Taken together, this means that pedicles tend to be long and thin in young animals, short and wide in older ones. This tendency has been clearly quantified in modern red deer by Hattemer and Dreschler (1976), who measured a sample of 1208 animals. A clear inverse relationship between pedicle length and diameter is shown by their data (fig. 8).
50
40
30
• 7
20 30 40 pedicle diameter
50
Fig. 8. - The change in pedicle proportions with age of the individual. Plotted from data in Hattemer & Dreschler (1976, table 2), based on a sample of 1208 modern Ce
elaphus. Each number indicates the mean proportions in that age class, from two to thirteen years.
Evolution des proportions du pédicule en fonction de l'âge de 2 à 13 ans : données de Hattemer et Dreschler
(1976, tabl. 2), d'après 1208 cerfs acutels.
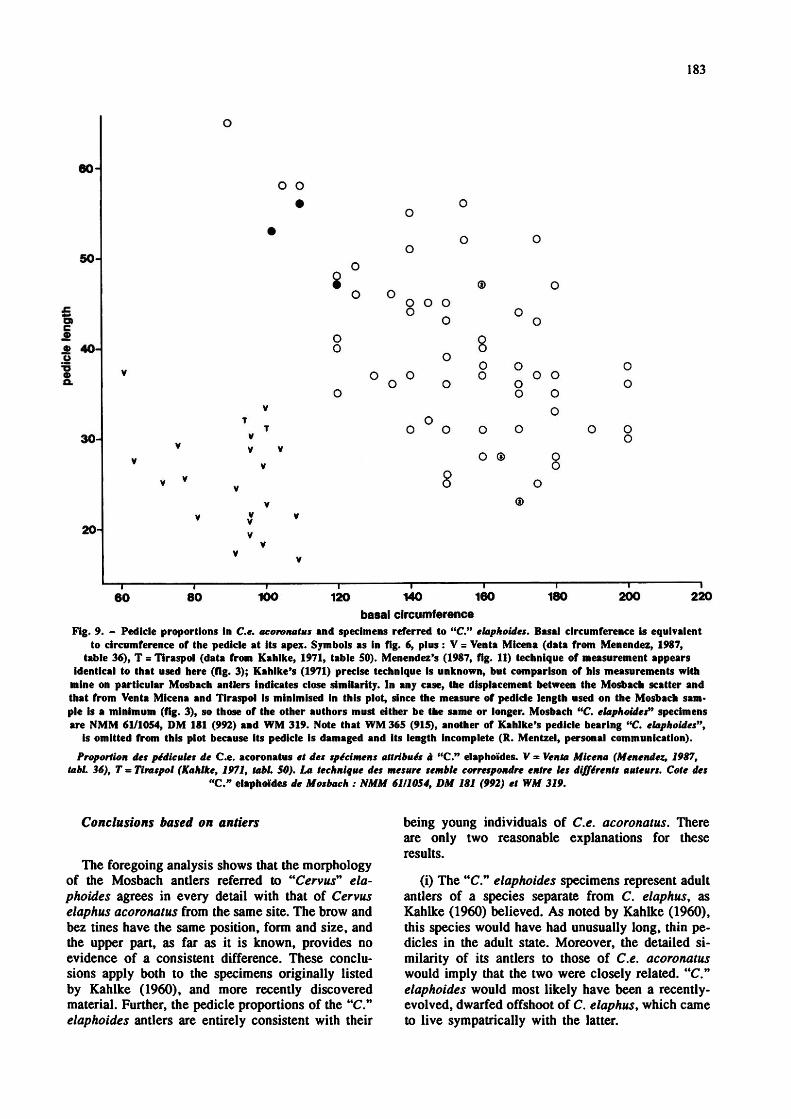
A plot of pedicle length vs. circumference for the Mosbach antlers is shown in fig. 9. The inverse relationship is evident, with young antlers at top left of the scatter, older ones at bottom right. Two antlers in Kahlke's (1960) original sample of "C." elaphoides from Mosbach (WM 319; DM 181 (now 992)) preserved their pedicles, and these have been measured in the present study, as has the new pedicle-bearing specimen NMM 61 /1 054 (fig. 1 b). These clearly show long, thin pedicles which, plotted on fig. 9, fall at the "young" end of the Ce. acoronatus sample. In other words, they have the pedicle proportions which would be expected of young individuals of the Mosbach Ce. acoronatus population.
183
60-
50-
« 40 u "g
30-
20-
o o
V v v
o
o
goo
o
o
o o o
o
o
8
O o o o o o o o
o
o o
o o o
8 ®
60 80 100 180 200 220 120 140 160 basal circumference
Fig. 9. - Pedicle proportions in C.e. acoronatus and specimens referred to "C." elaphoides. Basal circumference is equivalent to circumference of the pedicle at its apex. Symbols as in fig. 6, plus : V = Venta Micena (data from Menendez, 1987,
table 36), T = Tiraspol (data from Kahlke, 1971, table 50). Menendez's (1987, fig. 11) technique of measurement appears identical to that used here (fig. 3); Kahlke's (1971) precise technique is unknown, but comparison of his measurements with
mine on particular Mosbach antlers indicates close similarity. In any case, the displacement between the Mosbach scatter and that from Venta Micena and Tiraspol is minimised in this plot, since the measure of pedicle length used on the Mosbach sample is a minimum (fig. 3), so those of the other authors must either be the same or longer. Mosbach "C. elaphoides" specimens are NMM 61/1054, DM 181 (992) and WM 319. Note that WM 365 (915), another of Kahlke's pedicle bearing "C. elaphoides",
is omitted from this plot because its pedicle is damaged and its length incomplete (R. Mentzel, personal communication). Proportion des pédicules de C.e. acoronatus et des spécimens attribués à "C." elaphoïdes. V ■ Venta Micena (Menendez, 1987,
tabl. 36), T - Tiraspol (Kahlke, 1971, tabl. 50). La technique des mesure semble correspondre entre les différents auteurs. Cote des "C." elaphoïdes de Mosbach : NMM 6111054, DM 181 (992) et WM 319.
Conclusions based on antiers
The foregoing analysis shows that the morphology of the Mosbach antlers referred to "Cervus" elaphoides agrees in every detail with that of Cervus elaphus acoronatus from the same site. The brow and bez tines have the same position, form and size, and the upper part, as far as it is known, provides no evidence of a consistent difference. These conclusions apply both to the specimens originally listed by Kahlke (1960), and more recently discovered material. Further, the pedicle proportions of the "C." elaphoides antlers are entirely consistent with their
being young individuals of C.e. acoronatus. There are only two reasonable explanations for these results.
(i) The "C." elaphoides specimens represent adult antlers of a species separate from C. elaphus, as Kahlke (1960) believed. As noted by Kahlke (1960), this species would have had unusually long, thin pedicles in the adult state. Moreover, the detailed similarity of its antlers to those of C.e. acoronatus would imply that the two were closely related. "C." elaphoides would most likely have been a recently- evolved, dwarfed offshoot of C. elaphus, which came to live sympatrically with the latter.

184
(ii) The "C." elaphoides specimens are young antlers of C elaphus acoronatus. This interpretation would fit their pedicle proportions, their continuity of form with the larger antlers, and their size range. Given red deer of the body size of the Mosbach Ce. acoronatus population, one would expect the youngest antlers (second (pricket), third and fourth year heads) to have basal circumferences of 8-12 cm (Lister, unpublished measurements on British Ipswichian (Eemian) and Flandrian (Holocene) samples). This corresponds exactly to the lower part of the combined Mosbach range, including the antlers referred to "C." elaphoides. Moreover, the question must be asked : if these small antlers are not young Ce. acoronatus, where are the young antlers of the latter ?
It is very unlikely that among the large collection of this species from Mosbach, which includes several prickets (second year heads), no third or fourth year heads would occur. Thus, if "C." elaphoides were regarded as a separate species, it would have to be argued that the small antlers from Mosbach are a mixture of "C." elaphoides and young Ce. acoronatus. However, there is no reasonable morphological basis for dividing the available sample of small antlers into two groups. Among the very smallest specimens, some lack a bez tine while others possess one (fig. 7b). However, this is entirely normal variation within a single red deer population. In any case, specimens with and without a bez tine were included together in "C." elaphoides by Kahlke (1960)
Specimen mandible 1945/8921* Mi or M2 1956/215 Ml or M2 1962/423 M2 1961/892 M1 or M2 1956/569 scapula 1961/85 humérus 1955/152
proximal radius 1957/585
distal proximal
radius 1962/622 distal distal radius 1954/319 metacarpal 1954/617 metacarpal 1960/354 tibia 1958/604 tibia 1964/93 tibia 1949/382 tibia 1958/725 metatarsal 1954/615 phalanx I 1963/142
1
DCCO
DCD rxo
DCD CD
DDDDD DDDD
COD
Number of character for that particular tooth or bone, from Lister (in prep.) 2 3 4 5 6 7 8 D dama total C elaphus total
COCO COD COD COD
CODDDCOD COCO COCO
COD
CD
COD
OC D CD
CD
OC
OC CO
CCC
COCO
DCODDDDDD
DDDDDD
DDDDD DDDDD
CD
DDDDD DDDD
D D
COD
OC
CCCC
C
CCCC
* specimen said to be from the "lowest level" at Mosbach (Schmidtgen, 1930) Tabl. 1. - Mosbach dental and postcranial remains at NMM here referred to a small cervid species. Each specimen is scored
for characters known to distinguish C. elaphus from D. dama. Each character is scored on a scale from DDDDD (strongly supporting identification as D. dama) through zero (indeterminate or intermediate between D. dama and C. elaphus) to
CCCCC (strongly supporting identification as C. elaphus). The score awarded in each case is a product of the known degree of reliability of the character, and the degree to which the C. elaphus or D. dama character state is expressed in the specimen (see Lister, in prep., for details). Blanks indicate either that the character was not visible on the specimen due to damage, or
that it showed intermediate morphology. Dashes indicate that there is no character of that number for that element. Mosbach- NMM. Restes dentaires et post-crâniens attribués au petit cernai : Chaque os est noté d'après les caractères reconnus
distinguant le cerf du daim. SD : correspondance totale au daim, 5C : correspondance totale au cerf, blanc: le caractère n'est pas visible.
185
(fig. 7b). Similarly, Bachofen-Echt's (1929) lost specimen (fig. 2), apparently lacking a trez tine, could be regarded as a separate species from the antlers, small and large, which possess one, but a single antler must be regarded as insufficient evidence, given the known occasional ocurrences of trez-less young C elaphus individuals. Moreover, this would cause a taxonomic problem since the type specimen of "C." elaphoides (fig. 1) provides no evidence of the trez region, so it would be impossible to decide whether it grouped in the trez-less ("C." elaphoides) or trez- bearing (young C. elaphus) category.
It might be argued that the similarity of the "C." elaphoides antlers to those of adult Ce. acoronatus is misleading, and that young C elaphus antlers would not show such strong development of the brow, and especially bez, tines. However, modern red
deer stags, reared under good conditions, can show clear brow and bez tines in their second (third year) head, which may occasionally attain six-point antlers (de Nahlik, 1959, p. 73). Moreover, it has already been shown that Mosbach Ce. acoronatus, and other early Middle Pleistocene populations of this species, had unusually strongly developed bez tines. Thus, the preserved categories of antler form at Mosbach form a perfectly reasonable growth sequence for a single species, from the unbranched prickets, directly into four-point antlers (brow-trez-fork or brow-bez- fork), and thence into the full 5-point complement (brow-bez-trez-fork).
Finally, if the "C." elaphoides antlers were indeed adults of a small cervid species, one would expect also to find in the collection the very small young antlers (including prickets) of that species. No such
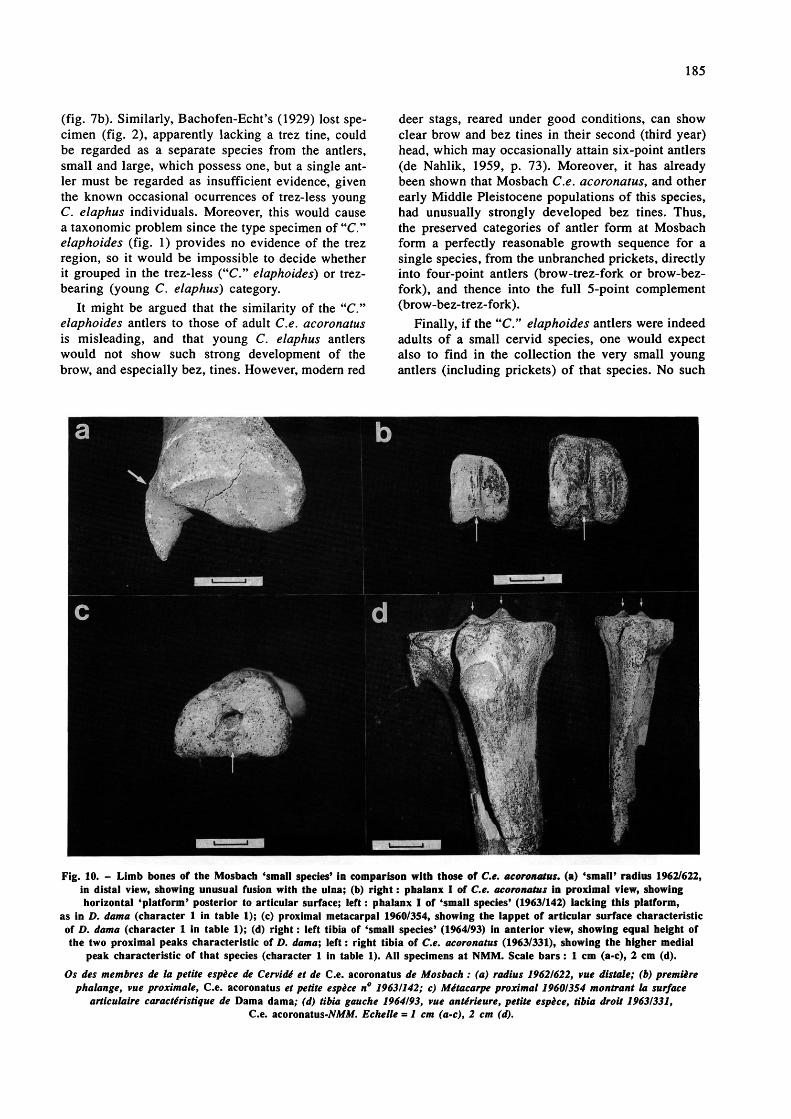
Fig. 10. - Limb bones of the Mosbach 'small species' in comparison with those of Ce. acoronatus. (a) 'small' radius 19(2/622, in distal view, showing unusual fusion with the ulna; (b) right : phalanx I of Ce. acoronatus in proximal view, showing horizontal 'platform' posterior to articular surface; left : phalanx I of 'small species' (1963/142) lacking this platform,
as in D. dama (character 1 in table 1); (c) proximal metacarpal 1960/354, showing the lappet of articular surface characteristic of D. dama (character 1 in table 1); (d) right : left tibia of 'small species' (1964/93) in anterior view, showing equal height of the two proximal peaks characteristic of D. dama; left : right tibia of Ce. acoronatus (1963/331), showing the higher medial
peak characteristic of that species (character 1 in table 1). AH specimens at NMM. Scale bars : 1 cm (a-c), 2 cm (d). Os des membres de la petite espèce de Cervidé et de Ce. acoronatus de Mosbach : (a) radius 19621622, vue dis taie; (b) première
phalange, vue proximate, Ce. acoronatus et petite espèce n° 19631142; c) Métacarpe proximal 1960I3S4 montrant la surface articulaire caractéristique de Dama dama; (d) tibia gauche 1964/93, vue antérieure, petite espèce, tibia droit 19631331,
Ce. acoronatus-M/Af. Echelle = 1 cm (a-c), 2 cm (d).

186
specimens are known, although this could conceivably be accounted for by preservation and collecting bias against them.
c) Bones and teeth
Kahlke (1960) mentioned a few dental remains from Mosbach which he tentatively referred to "C." elaphoides. Since Kahlke 's study, the Mosbach collections have expanded, and in the present investigation, one mandible, four isolated teeth, and thirteen partial or complete limb bones at NMM are referable to a cervid species smaller than Ce. aco- ronatus. Some of the specimens are clearly below the size range of the latter, and so are separable on that basis alone; others abut or slightly overlap in size the smallest Ce. acoronatus, but are separable on morphology (see below). All the 'small cervid' specimens are listed in table 1, and selected examples illustrated in fig. 10. Measurements of those which are sufficiently complete are presented in table 2, in comparison with Mosbach C.e. acoronatus.
The size range of the small skeletal elements is very conformable with that of Middle Pleistocene fallow deer, Dama dama (L.) (cf. Leonardi and Petronio, 1976). The other living European species in this size category is reindeer, Rangifer tarandus (L.). To investigate whether the Mosbach remains might pertain to one or other of these taxa, detailed morphological traits of the teeth and bones were examined. Reindeer teeth and bones have very distinctive and easily recognisable features; none of the Mosbach fossils falls into this category. The fallow
deer skeleton, however, is more similar to that of other Eurasian cervine deer (including Cervus), so identification requires more care. To this end, samples of known fallow and red deer skeletons (Recent, Holocene and Ipswichian (Eemian)) were compared, bone by bone and tooth by tooth, and an array of characters established by which the two species can be distinguished. Some of these characters are shown in fig. 10; full details are being published elsewhere (Lister, in preparation). In table 1, each of the Mosbach "small cervid" specimens has been scored according to these characters. The four teeth, and most of the limb bones, show overwhelmingly Dama dama-like character states (fig. 10, b-d). The only exception is distal tibia 1958/725, which is morphologically closer to Cervus elaphus, although radius 1962/622, while shows predominantly 'D. dama'- like character states (table 1), has the unusual feature of distal fusion to the ulna (fig. 10 a), which is unlike either C elaphus or D. dama.
The mandible (no. 1945/8921), now represented by a cast at NMM, is unfortunately too worn dentally to show the characters scored in the present study, and is in any case stated to have come from the "oldest level" (?Mosbach 1) (Schmidtgen, 1930). Interestingly, Schmidtgen (1930) referred this specimen to Dama dama.
The similarity of many of the Mosbach fossils to Dama dama does not make certain their identification to that species, since it is unknown to what extent some of the character states might be "primitive" among the Cervidae and be shared by a number of species. However, the identification of the Mosbach remains as Dama dama is quite plausible in view of the known occurrence of that species in the
Fig. 11. - Unshed antler base from Mosbach (NMM 1961/1002) here referred to D. dama. Scale bar 5 cm. Massacre, Mosbach, NMM 196111002, attribué à D. dama. Echelle = S cm.

187
Small species Specimen scapula 1961/85 humérus 1955/152 radius 1957/585 radius 1962/622 metacarpal 1954/617 tibia 1958/725 tibia 1964/93 metatarsal 1954/615 phalanx I 1963/142
dimension prox artic w dist artic w prox w prox w prox w distw proxd prox w prox w
measurement 35 6 42 5 46 2 47 5 32 8 37 3 51 0 29 0 19 0
Cervus elaphus n 36 46
48
54 72 3
64 82
range 38 0-50 0 47 6-63 0
50 0-69 0
35 5-49 0 46 0-63 0 78 0-92 0 32 0-48 0 18 7-28 2
acoronatus mean 45 39 54 24
60 03
43 12 53 27 86 00 39 18 22 39
st.dev. 3 06 3 13
4 51
3 03 3 73 7 21 5 58 1 56
Tabl. 2. - Measurements of Mosbach limb bones here referred to a small cervid species, in comparison with Mosbach C.e. acoronatus. Abbreviations : prox. = proximal, dist. = distal, artic. = articular, w. = width (medio-lateral dimension), d = depth
(antero-posterior dimension). Comparaison des mesures des membres attribuées à la petite espèce et à Ce. acoronatus. W = DMD et D = DAP.
early Middle Pleistocene of Europe; for example, at the Cromerian type site of West Runton, Norfolk (Azzaroli, 1953, p. 81-2; Lister, 1984). Moreover, since Kahlke's (1960) study, an antler base referable to D. dama has been discovered in the Mosbach Sands (NMM no. 61/1002). This specimen (fig. 11), still attached to the pedicle and part of the skull, shows all the features of fallow deer : a very inclined angle of departure of the beam (53° from the line of the pedicle), and a basal tine of circular section resting on the rose and making an obtuse angle (143°) with the beam. In addition, the large circumference of the basal tine (115 mm), approaching that of the beam (125 mm), is typical of early Middle Pleistocene Dama dama (Azzaroli, 1953, fig. 50).
3. OTHER SITES FROM WHICH "C." ELAPHOIDES HAS BEEN RECORDED
An indication of the currently suggested ages of the various sites here listed is given in table 3. Some of these sites are less well dated than others, and their dating may be subject to revision in the future.
a) Sites with small 'red-deer-like' antlers and small cervid skeletal remains
Sussenbom Germany
Kahlke (1969, p. 600) referred a total of five shed antlers and three isolated molars from the Sussen- born deposits to "C" elaphoides. Three of the antlers are complete enough to show the morphology of the lower region : of these, two possess a brow tine and
a rather weak bez tine, while the third bears a brow tine only, lacking the bez. All are broken above this region and so do not show the distal morphology. An example is illustrated in fig. 12.
Cervus elaphus acoronatus, of antler morphology very similar to that at Mosbach, occurs at Sussenbom (fig. 5 b & c), and it is difficult to see how the
myr BP
Arago
Mosbach III, Strénské Skàla Tiraspol Sussenbom
Solilhac, St Prest, Le Vallonnet
0.4
0.5
0.6
0.7
0.8
0.9
1.0
1.1
1.2
1.3
1.4
1.5 Tabl. 3. - Current estimates of the age of deposits from
which "C. elaphoides" has been recorded. Based on Agustf et al. (1987), Chaline (1974), Igel (1985) and Maglio (1973).
Estimation courante de l'âge des dépôts dans lesquels "C. elaphoides" a été déterminé, d'après Agusti et al. (1987),
Chaline (1974), Igel (1985), Maglio (1973).
Venta Micena
Cueva Victoria Sinzelles Durfort

188
Fig. 12. - Left shed antler base from Siissenborn (IQW 64/2107) referred to "C." elaphoides by Kahlke (1969). From Kahlke (1969, fig. 35); scale bar 5 cm.
Bois de chute, Siissenborn (IQW 6412107), attribué à "C." elaphoides par Kahlke (1969, fig. 35). Echelle = 5 cm.
Sussenborn antlers referred to "C." elaphoides can be excluded from being young antlers of Ce. aco- ronatus. Their basal circumferences range from 95- 135 mm (measurements by the author on material at IQW), comparable to young red deer (cf. fig. 4), and red-deer-like brow and bez tines are present. The relatively weak bez tine is a common feature of young C. elaphus antlers, and the difference from the Mos- bach antlers in this respect is typical of the type of variation found between red deer populations at the present day. Kahlke (1969) mentioned the lack of 'pearling' on the "C." elaphoides antlers, but they do possess distinct ridge-and-furrow ornament and so are not entirely smooth; this hardly seems a justifiable basis for taxonomic separation from Ce. aco- ronatus.
Nonetheless, it is conceivable, since none of the small Sussenborn antlers preserve a pedicle (Kahlke, 1969), that they could be adult antlers of a small species, perhaps the species found at Venta Micena (see later). Note that the Sussenborn antlers tabulated by Menendez (1987, table 38) as "C." elaphoides, including several with pedicles attached, are larger and are regarded even by Kahlke (1956, 1969) as Ce. acoronatus.
The three small cervid teeth from Sussenborn do betoken a small cervid species at the site. While the nature of this species cannot be known for certain, the teeth are very conformable morphologically to Dama dama, for example, in the presence of a weak buccal cingulum on the upper molars, and a lingual 'step' between the second and third lobes of the M3 (see Kahlke, 1969, fig. 34). These and other features are characteristic of D. dama rather than C. elaphus molars (Lister, in preparation).
Tiraspol, Moldavia At Tiraspol, Kahlke (1971) described several r
emains as "C." elaphoides : first, a small shed antler base of typical "C" elaphoides/young C elaphus form, with two well-developed basal tines; second, three small partial skulls with pedicles attached. The pedicles are short and stumpy (hence adult), but unfortunately their antlers have in each case been broken just above the base so that little or nothing of their morphology can be seen. Note that the pedicle proportions are quite different from those of putative "C" elaphoides from Mosbach (fig. 9). The skulls and limb bones demonstrate the presence of a small cervid species, but unfortunately the single sufficiently preserved antler is shed, so it is not certain whether it is conspecific with the small cervid remains (fitting a "C." elaphoides interpretation), or is merely a young antler of C. elaphus acoronatus, which also occurs at the site.
Solilhac, France The basal part of a single small shed antler at
MNHN (no. So 1.60), showing the typical double lower tine, was referred by Heintz and Poplin (1974) to "C" elaphoides, although they pointed out that the possibility of the specimen pertaining to C elaphus could not totally be ruled out. Menendez (1987) further referred to "C." elaphoides some small cervid dental remins from Solilhac, preserved at BM. He noted that they were very similar to teeth from Venta Micena, Spain (see later). ■
b) Sites with small cervid skeletal remains, but no sufficiently preserved antlers
Strânskâ Skâla, Czechoslovakia
Kahlke (1972) referred one partial molar of a small cervid to "C." elaphoides, but noted that on such limited material the identification was provisional.
Voigtstedt, Germany An upper second molar of a small cervid was ini
tially, and prudently, referred by Kahlke (1965) to "Cervus sp.", but has subsequently become regarded as "C." elaphoides (Kahlke, 1972; Heintz and Poplin, 1974; Menendez, 1987).
Sinzelles, France Cervid material of small size from this locality
was referred to "C." elaphoides by Heintz and Poplin (1974). In the present study, the following material at MNHN was examined : five partial or complete lower tooth rows, three partial or complete upper

189
tooth rows, and two isolated teeth. These remains are Dama-sized, but their morphology does not correspond well to either D. dama or C. elaphus. For example, the M3's lack the lingual 'step' between second and third lobes characteristic of D. dama, and unlike both species (Lister, in preparation), possess small lingual cuspules on the anterior part of the third lobe. Their specific attribution is therefore uncertain.
Arago, France
Abundant dental and some skeletal remains of a small cervid occur in the Lower Sequence at Arago. These fossils were referred by Crégut (1979, 1980) to "C." elaphoides, largely on the basis of size. Unfortunately, no antler remains corresponding to the small cervid have been found, so its specific identity remains uncertain. Morphological characters on many of the teeth and bones, however, are consistent with Dama dama (Lister, 1990-1991 [ce numéro du Quaternaire]).
Saint-Prest, France
Two antler bases showing the base of the first tine and with pedicles attached were referred to "C." elaphoides by Heintz and Poplim (1974). The attribution to this species (or to young C. elaphus) is uncertain, however, as the specimens are too damaged to show whether the characteristic second basal tine was present. Moreover, in at least one of the specimens, examined in the present study (MNHN, no. SPR-68), the pedicle is broken proximally so its full length, and hence the age of the individual, cannot be determined. The specific identity of the St. Prest material must therefore be regarded as unknown, although the rather high position and acute angle of the first tine on SPR-68 makes its attribution to Dama dama unlikely.
Cueva Victoria, Spain "C." elaphoides was recorded from this site by
Carbonell et al. (1981), but was omitted from a revised faunal list given by Gibert & Pèrez-Pèrez (1989). Menendez (1987) noted significant differences between the small cervid dental remains from Cueva Victoria and the putative "C." elaphoides material from Venta Micena (see later).
c) Sites with antlers differing from those of "C." elaphoides and C. elaphus
Le Vallonnet France Heintz and Poplin (1974), on the basis of the
limited small cervid material then available from Le Vallonnet, referred the remains to "C." elaphoides
on the basis of dental characters, noting their similarity to those from Sinzelles. Subsequently, much more small cervid material has become available from the site, including several shed antlers, two of which were figured by Lumley et al. (1988, fig. 14). These antlers, showing a broad, obtusely-angled, single lower tine, resting directly on the rose, are nothing like those of "C." elaphoides. This was recognised by Moullé (1986), who prudently demurred from identifying them to species, and by Lumley et al. (1988), who referred them to "Cervus" nestii (formerly Dama nestii Forsyth Major).
The new material from Le Vallonnet also includes a small frontal showing short, stumpy pedicles (Lumley et al. 1988, fig. 9), conforming to an adult individual of the small cervid species.
Durforty France The skull and antlers of a small cervid, referred
by Heintz and Poplin (1974) to "C." elaphoides, has been examined at MNHN in the present study. Each antler possesses a single lower tine based a few centimetres above the rose, above which the beam, long and sinuous, proceeds without further branching until 28 cm above the rose, at which point it is broken off. This morphology is quite unlike the Mosbach type material of "C." elaphoides, and also unlike C. elaphus or D. dama. Its greatest similarity is perhaps to Cervus perolensis, but the true specific attribution of this specimen remains to be established.
d) A genuine new species ?
Venta Micena, Spain
The large collection of fossils from this site includes the most important sample of "small cervid" material, apart from that of Mosbach, considered in this paper. There is no reason to doubt that the abundant antler, tooth and bone remains described in excellent detail by Menendez (1987) all belong to a single cervid species of small size. On the basis of the antler morphology, Menendez (1987) referred the Venta Micena material to "C." elaphoides. The antlers all show a basal tine placed close to the rose; in addition, the larger antlers of the sample possess a rudimentary second tine ("button") above the first, which is absent in smaller individuals. Menendez (1987) quite properly interpreted this as the normal, greater development of antler form in larger and older individuals of a species. He went on to homo- logise the two tines with the two basal tines of "C." elaphoides, particularly in the Siissenborn sample where the bez tine is somewhat weaker than at Mosbach (see above). Although the very reduced nature of the supposed bez tine at Venta Micena somewhat weakens the basis for equating it with

190
other "C." elaphoides samples, this attribution is still plausible if the Venta Micena sample represented a population of "C." elaphoides with particularly weak bez-tine development. Analogous inter-population variation occurs within C. elaphus at the present day (de Nahlik, 1959).
The identification of the Venta Micena species as "C" elaphoides is made difficult, however, by its pedicle proportions. The length and circumference of the 20 pedicles measured by Menendez (1987) have been plotted on fig. 9. A rough inverse correlation within the sample is evident, reflecting individual age variation as explained previously. Most significant, however, is the clear displacement of the scatter from that of Mosbach. The Venta Micena pedicles are much shorter at a given circumference than those from Mosbach, and indeed show exactly the proportions that would be expected in a genuine small- bodied cervid species. The long, thin pedicles of the small Mosbach antlers correspond, as explained above, either to their being young C. elaphus aco- ronatus, or if they do represent a separate species, it is one with unusually long pedicles in the adult stage. In either case, they cannot be equated with those from Venta Micena.
In his detailed description of the dental remains from Venta Micena, Menendez (1987) noted several traits which characterise this species, for example in the form of the entoconid of P4, and the third lobe of M3. A comparison of this material with the type sample of "C" elaphoides from Mosbach is clearly desirable. Unfortunately, the sample of small cervid dental material from Mosbach is very limited (see table 1). The four isolated first and second lower molars do not provide useful characters for comparison. There remains only the cast of the lower jaw (NMM no 1945/8921), which Schmidtgen (1930) referred to D. dama and Kahlke (1960) to "C." elaphoides. The teeth of this specimen are very worn, but the entoconid of P4 appears to show a free anterior end, as in the Venta Micena species. This similarity is intriguing, especially since the jaw is said to be from the lowest level at Mosbach, and is hence perhaps of comparable age to Venta Micena. By the same token, however, it may not relate to the "C." elaphoides remains from Mosbach III. Moreover, the variability of P4 morphology found in some species (Janis and Lister, 1985) means that a larger sample from Mosbach would be required for proper comparison.
Menendez (1987) noted a striking similarity between the dental remains of Venta Micena and those from Solilhac (see above), implying conspeci- ficity, while those from Sinzelles and Cueva Victoria showed different features, suggesting a separate species. This is a valid possibility which requires further investigation, but the referral of any of this material to "C." elaphoides as defined at Mosbach seems
bious. It is worth noting that these samples are up to c. 0.7 myr older than the type "C." elaphoides from Mosbach.
In summary, the Venta Micena material seems to represent a small cervid species showing normal short pedicles in the adult stage, and a basal antler morphology comprising first and vestigial second lower tines. This morphology suggests a genuinely new species, as yet unnamed.
4. CONCLUSIONS
According to the rules of zoological nomenclature (ICZN, 1985), the existence or otherwise of a species "Cervus" elaphoides depends on the validity of its type material. It is not possible, on the basis of specimens currently available, to totally disprove the existence of such a species at its type locality of Mosbach. If the species does exist, its great similarity in antler morphology to Cervus elaphus would suggest it was very closely related to the latter species, so the inverted commas should be dropped and the species styled Cervus elaphoides. Proof of its existence could be provided if a small skull bearing "C." elaphoides-type antlers with long pedicles but with the worn teeth of an adult animal, were found.
However, the data presented here shows that the currently available Mosbach small antlers are entirely consistent with their being young individuals of C. elaphus acoronatus, and this would appear to be the most parsimonious interpretation of the material. In this case, the name "Cervus" elaphoides is a junior synonym of C. elaphus, and should be dropped (cf. ICZN, 1985).
That a cervid species smaller than C. elaphus does also occur at Mosbach is quite clear from the dental and postcranial remains, and although the specific identity of this material is not certain, much of it is consistent with referral to Dama dama.
Since Kahlke 's (1960) study, fossils from at least 12 other European sites have been referred to "C" elaphoides by various authors. In some cases (Siis- senborn, Tiraspol, Solilhac), the situation is similar to Mosbach in that a small cervid species is evidenced by teeth and/or bones, but preserved antlers are not directly associated with them and can be referred to young Cervus elaphus. In other cases (Strânskâ Skâla, Voigtstedt, Sinzelles, Arago, St. Prest), dental and/or skeletal material indicates the presence of a small cervid, but in the absence of antler material, specific attribution is uncertain, although the detailed morphology of the teeth and bones may suggest groupings of the samples and/or referral of some samples to known species. In a third category of sites (Le Vallonnet, Durfort), antler remains are well-preserved, and patently indicate a species other than that

191
of the type material of "C." elaphoides. Finally, the very interesting material from Venta Micena comes closest to indicating the existence of a species with antlers in some respects similar to the type antlers of "C" elaphoides, but with pedicle proportions which preclude identity with the Mosbach material, suggesting rather a "new", and at present unnamed, species.
Heintz and Poplin (1974) and Crégut (1980) saw "C" elaphoides as a species restricted to the Cro- merian and early "Mindel", and thus a good biostra- tigraphic marker for the early Middle Pleistocene. However, the sites from which "C." elaphoides has now been recorded cover a much larger span of time : c. 1 myr (table 3). The referral to "C." elaphoides of material from all of these sites has probably inhibited recognition of a more diverse array of small cervid species in this time period, which should be the subject of future research. Antler characters still provide the most reliable basis for defining and identifying species, provided that individual (especially age) variation is properly taken into account. Dental and osteological characters have considerable potential, and require further careful research, using appropriate statistical samples to take account of variability. At our present limited state of knowledge, such characters must be used with particular care since apparently "species-specific" features might in fact be primitive characters which could characterise a number of different species. The • known array of small to medium-sized cervid species from the European late Lower and Middle Pleistocene, such as Dama dama, "Cervus" nestii, Cervus philisi Schaub and "Euctenoceros" médite rraneus Bonifay, may still be incomplete, with other species in this size category remaining to be properly defined.
ACKNOWLEDGEMENTS
I am particularly grateful to the curators of various museums who provided access to material in their care : Dr. W. Igel (NMM), Dr. G. Storch (SMF), Dr. W.v. Koenigswald (DM), Dr. E. Heintz (MNHN), Dr. J. Clutton-Brock (NHM) and Dr. H.- D. Kahlke (IQW). Dr. R. Mentzel (WM) kindly assisted with measurements. Drs. A.J. Stuart, H.-D. Kahlke, and E. Heintz provided valuable discussion. The research was supported by grants from Gonville and Cauis College and the DAAD, and a Royal Society/CNRS exchange fellowship.
RÉFÉRENCES
Agustf, J., Moyà-Solà, S. and Pons-Moyà, J. (1987) - La sucesiôn de Mamfferos en el Pleistoceno inferior de Europa : proposition de una nueva escala bioestratigrâ- fica. Paleont. i EvoL, Mem. Esp., 1, 287-295.
Ahlén, I. (1965) - Studies on the red deer, Cervus elaphus L., in Scandinavia. Viltrevy, 3, 1-247.
Azzaroli, A. (1948) - Revisione della fauna dei terreni fluvio-lacustri del Valdarno Superiore, 3. I cervi fossili della Toscana con particolare riguardo allé specie Vil- lafranchiane. Palaeontogr. ital., 43, 45-82.
Azzaroli, A. (1953) - The deer of the Wey bourne Crag and Forest Bed of Norfolk. Bull. Br. Mus. nat. Hist. (A. Geology), 2, 3-96.
Bachofen-Echt, A. (1929) - Die Stellung der Mosbacher Geweihe von Cervus elaphus in der Entwicklungs-Reihe dieses Hirsches. Notizbl. Ver. Erdk. Darmstadt, 1929, 5/12, 171-175.
Beninde, J. (1937) - Uber die Edelhirschformen von Mosbach, Mauer und Steinheim a.d. Murr. Palâont. Z., 19, 79-116.
Berggren, W.A., Burckle, L.H., Cita, M.B., Cooke, H.B.S., Funnell, B.M., Gartner, S., Hays, J.D., Ken- nett, J.P., Opdyke, N.D., Pastouret, L., Shackleton, N.J. and Takayanagi, Y. (1980) - Towards a Quaternary time scale. Quat. Res., 13, 277-302.
Briining, H. (1978) - Zur Untergliederung der Mosbacher Terrassenabfolge und zum klimatischen Stellenwert der Mosbacher Tierwelt im Rahmen des Cromer-Kom- plexes. Mz. Naturw. Arch., 16, 143-190.
Carbonell, A., Estevez, J., Moyà-Solà, S., Pons-Moyà, J., Agusti, J. and Villalta, J.F. (1981) - « Cueva Victoria » (Murcia, Espana) : Lugar de ocupacion humana mas antiguo de la Peninsula Iberica. Endins, 8, 47-57. Ciutat de Mallorca.
Chaline, J. (1974) - Essai de stratigraphie biologique et climatique du Pleistocene, du Pliocène et du Miocène supérieur continental eurasiatique, fondé sur l'étude des rongeurs. CNRS, Dijon.
Crégut, E. (1979) - La faune de mammifères du Pleistocene moyen de la Caune de l'Arago à Tautavel, Pyrénées Orientales. Unpublished thesis, Docteur de Spécialité, University of Provence, Marseille, 1-381.
Crégut, E. (1980) - La faune de mammifères du gisement pleistocene moyen antérissien de la Caune de l'Arago (Tautavel, Pyrénées Orientales, France). C.r. Acad, Se. Paris Ser. D, 290, 751-754.
Fraser, F.C. and King, J.E. (1954) - Faunal Remains in Clark J.G.D.C., Excavations at Star Carr. Cambridge University Press, Cambridge, ch. 3.
Gibert, J. and Pèrez-Pèrez, A. (1989) - A human phalanx from the Lower Palaeolithic site of Cueva Victoria (Murcia, Spain). Hum. EvoL, 4, 307-316.
Hattemer, H.H. and Dreschler, H. (1976) - Rosenstock- und Geweihmerkmale von Harz-Hirschen und ihr Zusammenhang mit dem Alter. Z. Jagdwiss., 22, 36-50.
Heintz, E., and Poplin, F. (1974) - Nouvelle interprétation des cervidés (Mammalia) de taille moyen des

192
faunes postvillafranchiens de France. C.r. Acad. Se. Paris Ser. D, 279, 397-400.
Igel, W. (1985) - Thesis, Univ. Mainz. International Commission for Zoological Nomenclature
(1985) - International Code of Zoological Nomenclature. International Trust for Zoological Nomenclature / British Museum (Natural History), London.
Janis, CM. and Lister, A.M. (1985) - The morphology of the lower fourth premolar as a taxonomic character in the Ruminantia (Mammalia : Artiodactyla), and the systematic position of Triceromeryx. J. Paleont., 59, 405-410.
Kahlke, H.-D. (1956) - Die Cervidenreste aus den alt- pleistozànen Ilmkiesen von Sussenborn bei Weimar. Akademie-Verlag, Berlin.
Kahlke, H.-D. (1960) - Die Cervidenreste aus den alt- pleistozànen Sanden von Mosbach (Biebrich-Wiesba- den). Abh. dt. Akad. Wiss., Kl. Chem. Geol. Biol., 7, 1-75.
Kahlke H.-D. (1965) - Die Cerviden-Reste aus den Tonen von Voigtstedt in Thiiringen. Palàont. Abh. A, 2, 379- 426.
Kahlke, H.-D. (1969) - Die Cerviden-Reste aus den Kie- sen von Sussenborn bei Weimar. Palàont. Abh. A, 3, 547-610.
Kahlke H.-D. (1971) - Family Cervidae in Nikiforova K.V. et al., The Pleistocene of Tiraspol. Acad. Sc. Mol- dav. SSR, Kishinev, 137-156.
Kahlke, H.-D. (1972) - Die Cerviden-Reste der Strânskâ Skâla bei Brno. Anthropos, Brno, 20, 193-197.
Kurtén, B. (1968) - Pleistocene Mammals of Europe. Weidenfeld and Nicholson, London.
Leonardi, G. and Petronlo, C. (1976) - The fallow deer of European Pleistocene. Geol. Rom., 15, 1-67.
Lister A.M. (1984) - Evolutionary and ecological origins of British deer. Proc. R. Soc. Edinb., 82B, 205-229.
Lister, A.M. (1986) - New results on deer from Swans- combe, and the stratigraphie al significance of deer in the Middle and Upper Pleistocene of Europe. J. Archaeol. Sc, 13, 319-338.
Lister, A. M. (1987) - Diversity and evolution of antler form in Quaternary deer in Wemmer CM., Biology and Management of the Cervidae. Smithsonian Institution, Washington.
Lister, A.M. (1990-1991) - Taxonomy and biostratigraphy of Middle Pleistocene deer remains from Arago, Pyrénées Orientales, France. Quaternaire, l (doit être publié dans le même numéro).
Lister, A.M. (in prep.) - Morphological characters for the separation of teeth and bones of red and fallow deer. J. Archaeol. Sc.
Lumley, H. de, Kahlke, H.-D., Moigne, A.-M. and Moul- lé, P.-E. (1988) - Les faunes de grands mammifères de la grotte du Vallonnet Roquebrune-Cap-Martin, Alpes- Maritimes. L'Anthropologie (Paris), 92, 465-496.
Maglio, V.J. (1973) - Origin and evolution of the Ele- phantidae. Trans. Am. Phil. Soc, 62, 1-149.
Menendez, E. (1987) - Cérvidos del yacimiento del Pleis- toceno inferior de Venta Micena-2, Orce (Granada, Espana). Paleont. i Evol., Mem. Esp., 1, 129-181.
Moullé, P.E. (1986) - Contribution à l'étude des faunes pleistocene inférieur de la grotte du Vallonnet (Roquebrune-Cap-Martin, Alpes-Maritimes) : les Cervidés. - Mémoire de DEA. MNHN, Paris, 1-83.
Nahlik, H. de (1959) - Wild Deer. Faber and Faber, London.
Schmidtgen, O. (1930) - Eine Unterkieferhâlfte vom Damhirsch aus dem Mosbacher Sand. Notizbl. Ver. Erdk. Darmstadt, 5/12, 114-118.