Embed Size (px)
Citation preview

Diversité de la végétation et du réservoir degraines du sol dans une pelouse calcicole dunord-ouest de la France
Olivier Chabrerie, Didier Alard et Blaise Touzard
Résumé : Dans une pelouse calcicole de la vallée de la Seine au nord-ouest de la France, la végétation et le réservoirde graines du sol ont été comparés selon leur composition, leur diversité et leur structure spatiale, grâce à un pland’échantillonnage spatialement explicite. Les résultats montrent (i) que l’on peut reconstituer un gradient de successiondes stades de pelouses rases aux stades pré-forestiers, à partir des données de la végétation; (ii) qu’il existe une faiblecorrespondance entre la végétation et le réservoir de graines (similarité moyenne de 41 %), dominé essentiellement pardes espèces annuelles ou bisannuelles (Hypericum perforatum, Blackstonia perfoliata); (iii) qu’il existe des différencesentre les patrons spatiaux de la végétation et du réservoir de graines. Ces patrons spatiaux sont souvent décalés et pré-sentent des niveaux d’hétérogénéité différents. L’hétérogénéité spatiale du réservoir de graines et son indépendance vis-à-vis de la structure de la végétation sont interprétées selon les modes de fonctionnement et les modalités des échanges(pluie de graines, recrutement des semis) entre les deux compartiments.
Mots clés : pelouse calcicole, biodiversité, patron spatial, succession, banque de graines, approche synchronique.
Abstract: In a chalk grassland of the Seine valley in northwestern France, vegetation and seed bank were comparedaccording to their composition, diversity, and spatial structure through use of a spatial sampling design. The resultsshow (i) that it is possible to establish a successional gradient from short grasslands to early stages of forests with veg-etation data; (ii) that there is a weak correspondence between vegetation and seed bank (mean similarity 41%), whichwas mainly dominated by opportunistic annual or biennial species (Hypericum perforatum, Blackstonia perfoliata);(iii) that there are differences between spatial patterns of vegetation and seed bank. These spatial patterns often exhibita time lag and represent different levels of heterogeneity. The spatial heterogeneity of the seed bank and its independ-ence from the vegetation structure were interpreted according to the functioning of the two compartments and theirtype of exchanges (through seed rain and seedling recruitment).
Key words: chalk grassland, biodiversity, spatial pattern, succession, seed bank, synchronic approach. Chabrerie et al.840
Introduction
Les pelouses sèches des coteaux calcaires de la vallée dela Seine au nord-ouest de la France sont des écosystèmesprairiaux très riches en espèces (Dutoit et Alard 1996a).Suite à l’abandon des pratiques agro-pastorales depuis lamoitié du 20e siècle, ces pelouses se boisent spontanément,et les espèces inféodées aux stades successionnels pionnierssont menacées de disparition (Dutoit et Alard 1996b). Leréservoir de graines du sol, représentant un potentield’espèces disponibles, a fait l’objet de beaucoup d’intérêtpour la restauration des pelouses calcicoles et le maintien deleur diversité (Hillier 1990; Bakker et al. 1996a; Willems etBik 1998).
Les relations entre la végétation et le réservoir de grainesont souvent été étudiées le long de gradients écologiques trèscontrastés. Ces gradients sont déterminés par des facteursabiotiques comme la salinité et l’aridité (Holzapfel et al.1993; Egan et Ungar 2000) ou correspondent à des succes-sions secondaires (Usher et Jefferson 1990; Milberg 1995;Hyatt et Casper 2000). L’étude du réservoir de graines dansles différents stades de succession permet d’aborder les ef-fets de la dynamique de la végétation sur la composition duréservoir de graines, puisque certaines espèces sont suscepti-bles de disparaître ou de persister dans les stades finaux dela succession (stades forestiers). De ce fait, la capacité de re-tour de l’écosystème vers un état initial (les pelouses rases)après la colonisation par les ligneux, dépend en partie du ré-servoir de graines du sol et des propriétés de persistance desespèces.
Selon les milieux étudiés, la similarité entre la végétationet le réservoir de graines présente des valeurs faibles dansles communautés végétales pérennes (Milberg 1995; Peco etal. 1998; Edwards et Crawley 1999) et des valeurs fortesdans les milieux fréquemment soumis à des perturbations(Holzapfel et al. 1993; Eriksson et Eriksson 1997; Egan etUngar 2000). Le réservoir de graines a été défini comme uncompartiment fonctionnel majeur des communautés végéta-
Can. J. Bot. 80: 827–840 (2002) DOI: 10.1139/B02-068 © 2002 CNRC Canada
827
Reçu le 15 novembre 2001. Publié sur le site Web desPresses scientifiques du CNRC, http://revcanbot.cnrc.ca, le2 août 2002.
O. Chabrerie, D. Alard1 et B. Touzard. Université deRouen, Laboratoire d’Écologie, Unité Propre de Recherche del’Enseignement Supérieur, Equipe d’Accueil 1293, 76821Mont Saint Aignan CEDEX, France.
1. Auteur correspondant (courriel : [email protected]).
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:23 AM
Color profile: Generic CMYK printer profileComposite Default screen

les (Templeton et Levin 1979). Sa fonction a été étudiéedans le cadre de mesures de restauration (Hutchings et Bo-oth 1996; Willems et Bik 1998) mais son rôle a été soulignéau sein des communautés végétales dans le cadre des nichesde régénération (Grubb 1977) suite aux micro-perturbations(Hillier 1990; Rusch 1992). Les espèces persistant dans lesréservoirs de graines sont essentiellement des espèces pion-nières (Thompson et al. 1996). Cette observation permettraitd’expliquer la faible similarité avec les formations végétalespérennes.
Le concept d’hétérogénéité a été largement utilisé en éco-logie (Kolasa et Pickett 1991; Pickett et Cadenasso 1995;Wilson 2000) et notamment dans le cadre des successionsvégétales (Armesto et al. 1991). Cependant les études despatrons spatiaux des réservoirs de graines sont moins abon-dantes (Henderson et al. 1988; Bertiller 1998).L’hétérogénéité spatiale est définie comme la variation pointpar point de la composition spécifique dans une commu-nauté végétale (Inouye et al. 1987). De nombreux facteurspeuvent être source d’hétérogénéité et ne sont pas nécessai-rement les mêmes pour la végétation et le réservoir de grai-nes. De ce fait, les deux compartiments sont susceptibles deprésenter des niveaux d’hétérogénéité différents. Cependant,la végétation et le réservoir de graines sont interdépendantespar leurs échanges constants d’individus à travers la pluie degraines et la germination. Il peut donc en résulter une corres-pondance dans la composition entre ces deux comparti-ments.
Les objectifs de conservation des écosystèmes prairiauxvisent généralement à maintenir une mosaïque diversifiéed’habitats (Pickett 1978). Le rôle de l’hétérogénéité spatialedu réservoir de graines sur le maintien de cette mosaïquereste cependant largement méconnu. Dans les milieux prai-riaux, les compositions en espèces de la végétation et du ré-servoir de graines sont très différentes, et à l’inverse lescompositions des réservoirs de graines sont similaires entreelles (Milberg 1995; Bakker et al. 1996a).
Dans le cadre de cette problématique, nous avons proposédeux hypothèses : (i) au cours de la succession secondaire, ily a indépendance à la fois de la composition et des patronsspatiaux entre la végétation et le réservoir de graines;(ii) l’hétérogénéité spatiale de la végétation est plus impor-tante que celle du réservoir de graines.
Nous avons étudié les relations entre la végétation et le ré-servoir de graines dans une mosaïque de végétation selon unéchantillonnage spatialement explicite. Un gradient de suc-cession a été reconstitué par modélisation statistique. Cetteapproche nous a permis de comparer la structure de la végé-tation et du réservoir de graines (richesse spécifique, indicede Shannon et équitabilité) à la fois dans le temps (approchesynchronique) et dans l’espace.
Matériel et méthodes
Site d’étudeCette étude a été réalisée dans une pelouse calcicole de la
réserve naturelle d’Hénouville (49°29′ N; 0°56′ E) au nord-ouest de la France. La réserve est située sur une pente de 20à 30°, comprise entre 15 et 75 m d’altitude, orientée vers lenord-ouest, le long de la berge convexe de la Seine. La végé-tation se développe sur des sols pauvres de type Rendolls
(United States Department of Agriculture 1998) reposant surun substrat sédimentaire calcaire du Crétacé. La végétationrelève de l’alliance du Mesobromion erecti (De Foucault etFrileux 1988). Cultivé jusqu’à la fin du 19e siècle, le site aconnu un pâturage généralisé (ovins, bovins) jusqu’en 1960,date à laquelle il a été abandonné. La physionomie de la vé-gétation actuelle est représentée par une mosaïqued’habitats : pelouses rases, pelouses hautes et fourrés.
Plan d’échantillonnageL’échantillonnage de la végétation et du réservoir de grai-
nes du sol a été réalisé le long d’un transect de 16 m ×108 m (fig. 1) disposé de haut en bas du coteau couvrantl’ensemble des stades de succession de la végétation. Letransect consiste en une grille de 140 points d’échantillonnagerégulièrement espacés de 4 m.
Échantillonnage de la végétationLa végétation a été échantillonnée en juillet 2000 autour
des 140 points de prélèvement du réservoir de graines(fig. 1). Pour chaque point, le recouvrement des plantes vas-culaires a été estimé sur des quadrats de 1 m2 selon uneéchelle à sept degrés (1, espèces représentées par un seul in-dividu à très faible recouvrement dans le quadrat (0,25 %);2, espèces avec un recouvrement <1 % (soit 0,5 %); 3, 1 à
© 2002 CNRC Canada
828 Can. J. Bot. Vol. 80, 2002
0 4 8 12 16
mètres
0
20
40
60
80
100
mèt
res
Haut de pente
Bas de pente
1 m
Transect d’échantillonnage(140 points)
(1)la végétation :
· estimation du recou-vrement des espècesdans un quadrat de 1 m2
N
(2) Echantillonnagede la banque degraines du sol
· prélèvementd’une carotte de sol
6 cm
25 cm
22
cm
1m
10
cm
· mise en germi-nation des carottes desol dans une serren o n c h a u f f é e e tcomptage hebdoma-daire des semis
Fig. 1. Plan d’échantillonnage. La végétation et le réservoir degraines ont été échantillonnés sur une grille-transect de 16 m ×108 m contenant 140 points. Ce transect est orienté du bas versle haut du coteau.
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:24 AM
Color profile: Generic CMYK printer profileComposite Default screen

5 %; 4, 5 à 25 %; 5, 25 à 50 %; 6, 50 à 75 %; 7, 75 à100 %). Pour chaque catégorie, la valeur moyenne du recou-vrement a été utilisée lors des analyses statistiques.
Échantillonnage du réservoir de grainesLe réservoir de graines du sol a été échantillonné en fé-
vrier 2000, période durant laquelle le nombre de graines via-bles est le plus élevé dans les réservoirs de grainespermanentes des pelouses calcicoles (Thompson et Grime1979; Graham et Hutchings 1988) parce que beaucoupd’entre elles proviennent de la pluie de graines de l’annéeprécédente. Cent quarante carottes de sol (diamètre de 6 cm,hauteur de 10 cm, soit un volume par carotte de 282,74 cm3)ont été collectées tous les 4 m sur la grille d’échantillonnage(fig. 1). Ces modalités d’échantillonnage permettentd’enregistrer la majorité des graines viables qui se trouventdans les tous premiers centimètres du sol (Roberts 1981). Levolume total de sol prélevé sur l’ensemble du transect(39 583,6 cm3) est supérieur au volume minimal de sol(1200 cm3) nécessaire pour échantillonner la majorité desespèces d’une prairie (Roberts 1981).
Les carottes de sol ont été déstructurées et étalées dansdes sacs plastiques puis séchées à l’air ambiant dansl’obscurité, au laboratoire. Les échantillons de sol ont étédébarrassés de leurs cailloux et des fragments de plantes parun tri manuel et disposés ensuite sous forme d’une couchede 0,5 cm d’épaisseur sur du sol stérile (stérilisation du solpar passage à l’autoclave à 100°C pendant trois périodes de2 h) dans des pots en plastique (diamètre de 25 cm, profon-deur de 22 cm). Les 140 pots ont été placés dans une serrenon chauffée et arrosés deux fois par semaine pour maintenirle sol humide. Afin de faciliter l’apparition de nouveaux se-mis, la terre des pots dans la portion de surface a été re-tournée tous les deux mois, et les échantillons ont été gardésau sec pendant 2 semaines en août (Roberts 1981; Willems1995). Quinze pots témoins contenant uniquement de la terrestérilisée ont été placés au hasard parmi les autres pots afinde contrôler une éventuelle contamination par des grainesdispersées dans la serre.
Les semis émergeant des pots ont été identifiés, comptéset éliminés une fois par semaine de mars à novembre 2000.Cette période de mise en germination relativement longuepermet de tenir compte des propriétés de persistance et desstratégies de germination saisonnière des espèces présentesdans le réservoir de graines (Thompson et Grime 1979; Aré-valo et Fernández-Palacios 2000). La détermination des es-pèces à été réalisée au stade de semis à partir des flores deMuller (Muller 1978) et Hanf (Hanf 1983) et de spécimen deréférence. Les semis non identifiables ont été transplantésdans de nouveaux pots jusqu’à ce que leur développementpermette de les identifier. La nomenclature des espèces suitcelle de Lambinon et al. (1992).
Analyse des données
Analyse de la composition de la végétation et duréservoir de graines
La composition de la végétation a été analysée par desanalyses statistiques multivariées effectuées sur le recouvre-ment des espèces (140 relevés × 104 espèces). Une analysefactorielle des correspondances (AFC) (Hill 1973) a permis
d’examiner les patrons de variation de composition en espè-ces entre les relevés. Cette analyse a été complétée par uneclassification hiérarchique (CH) réalisée à partir des coor-données factorielles des 140 points sur les deux premiersaxes de l’AFC (Roux 1985) selon la méthode de Ward(Ward 1963), afin d’identifier les principaux typesd’assemblages d’espèces sur le transect. Ces analyses ont étéréalisées à l’aide du programme informatique ADE-4(Thioulouse et al. 1997).
Analyse de la diversité de la végétation et du réservoir degraines
Afin d’appréhender la diversité de la végétation et du ré-servoir de graines, ont été calculés pour chaque relevé : la ri-chesse spécifique R (Whittaker 1972), l’indice de ShannonH′ (Shannon et Weaver 1949) et l’équitabilité J′ (Pielou1966). Les moyennes de R, H′ et J′ ont été calculées pourchaque assemblage d’espèces, déterminés précédemment parles analyses sur la végétation. Pour chaque typed’assemblage d’espèces identifié dans la végétation, le poold’espèces local, défini comme le réservoir d’espèces présen-tes dans chaque assemblage (Partel et al. 1996), du réservoirde graines et de la végétation a été évalué à travers la ri-chesse spécifique totale.
Similarité entre la végétation et le réservoir de grainesL’indice de similarité de Jaccard (Jaccard 1912) a permis
de comparer la composition de la végétation et celle du ré-servoir de graines pour chaque point du transect. La formulede cet indice est la suivante : % similarité = 100c/(a + b – c)où a représente le nombre d’espèces dans la végétation, b re-présente le nombre d’espèces dans le réservoir de graines etc représente le nombre d’espèces communes à la végétationet au réservoir de graines. Les moyennes de cet indice ontété calculées pour chaque assemblage d’espèces. La simila-rité entre les pools d’espèces des différents assemblages aégalement permis de mettre en évidence les décalages dyna-miques entre la végétation et le réservoir de graines.
Une série d’analyses de variance à un facteur (ANOVA) apermis de déterminer si ces indices (R, H′, J′, Jaccard)étaient significativement différents entre les assemblagesd’espèces. Des tests de normalité ont été préalablement ef-fectués pour vérifier la normalité de la distribution des don-nées. Une transformation logarithmique de base 10 a éténécessaire pour normaliser la distribution de l’indice de Jac-card. L’homogénéité des variances a été vérifiée. Quand uneffet significatif a été observé, des tests de comparaisons demoyennes Tukey–Kramer (Sokal et Rohlf 1981) ont permisde comparer les moyennes des indices entre les assemblagesd’espèces.
Patrons spatiaux de la végétation et du réservoir degraines
Les patrons spatiaux de la végétation et du réservoir degraines ont été étudiés au moyen de cartographies réalisées àl’aide du programme informatique S-PLUS 2000 (MathSoft1999). Plusieurs variables relatives à la végétation et au ré-servoir de graines ont été cartographiées (algorithme : inter-polation bivariée pour grille rectangulaire sous S-PLUS) :(i) les variations de la composition de la végétation, évaluéespar les coordonnées factorielles des points du transect le
© 2002 CNRC Canada
Chabrerie et al. 829
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:24 AM
Color profile: Generic CMYK printer profileComposite Default screen
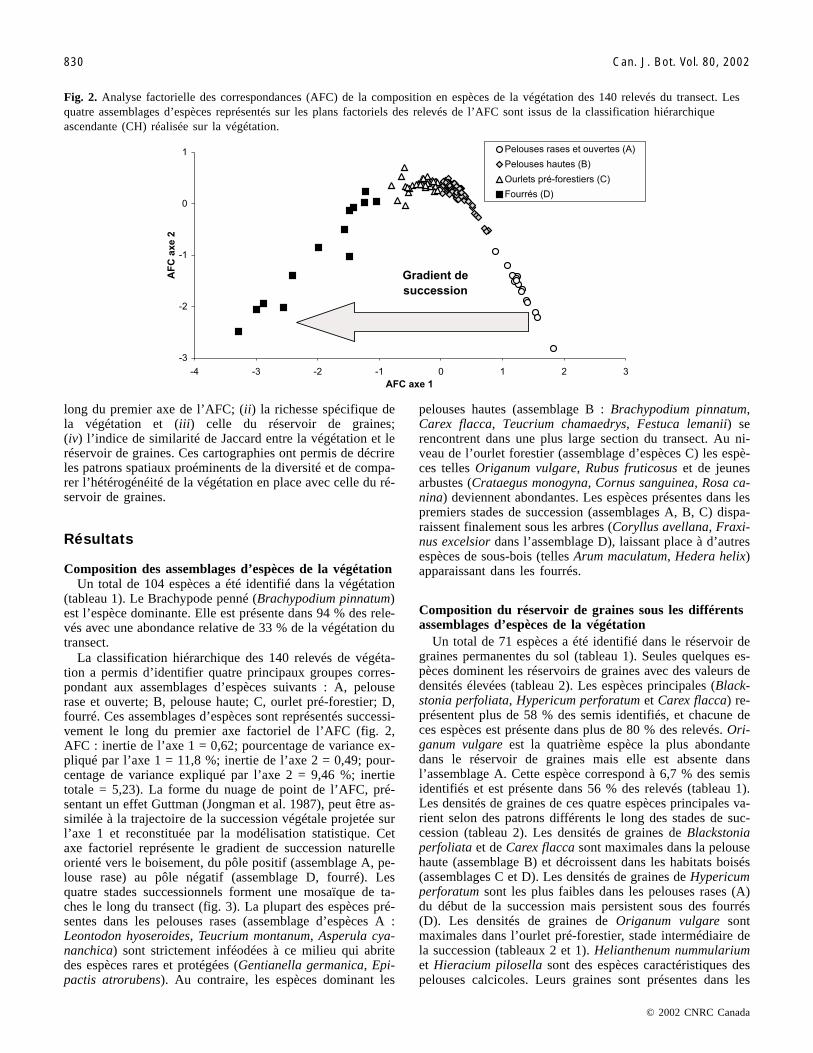
long du premier axe de l’AFC; (ii) la richesse spécifique dela végétation et (iii) celle du réservoir de graines;(iv) l’indice de similarité de Jaccard entre la végétation et leréservoir de graines. Ces cartographies ont permis de décrireles patrons spatiaux proéminents de la diversité et de compa-rer l’hétérogénéité de la végétation en place avec celle du ré-servoir de graines.
Résultats
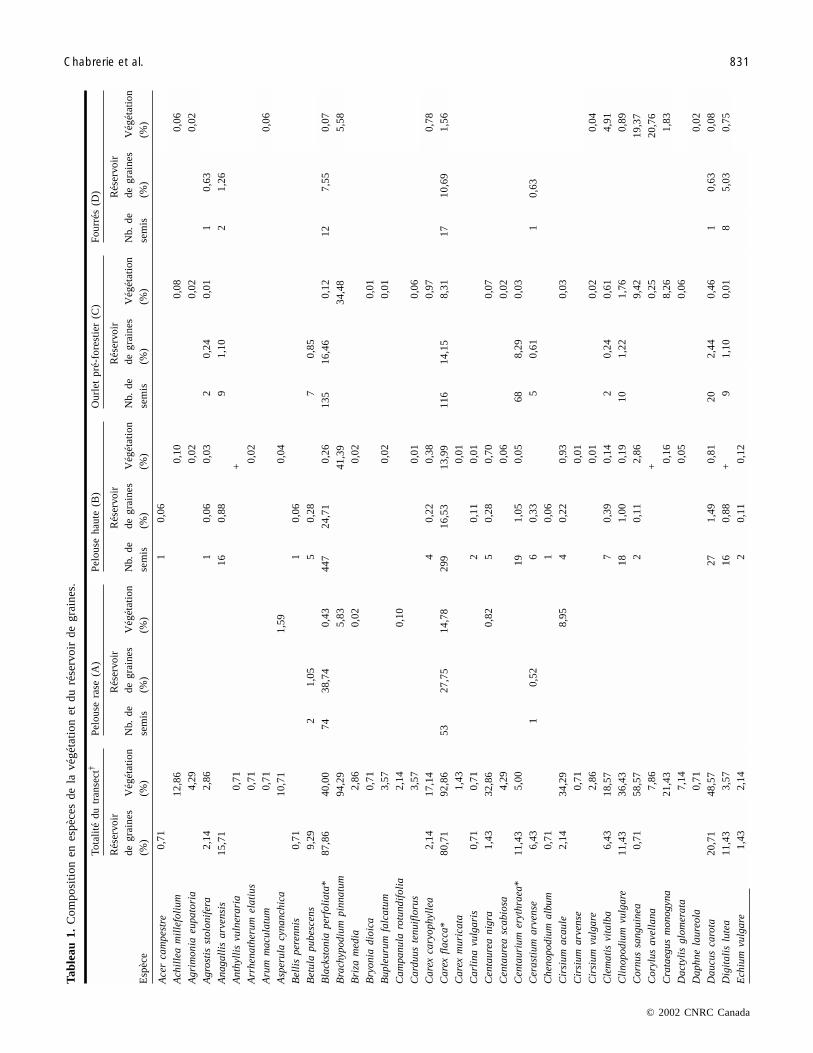
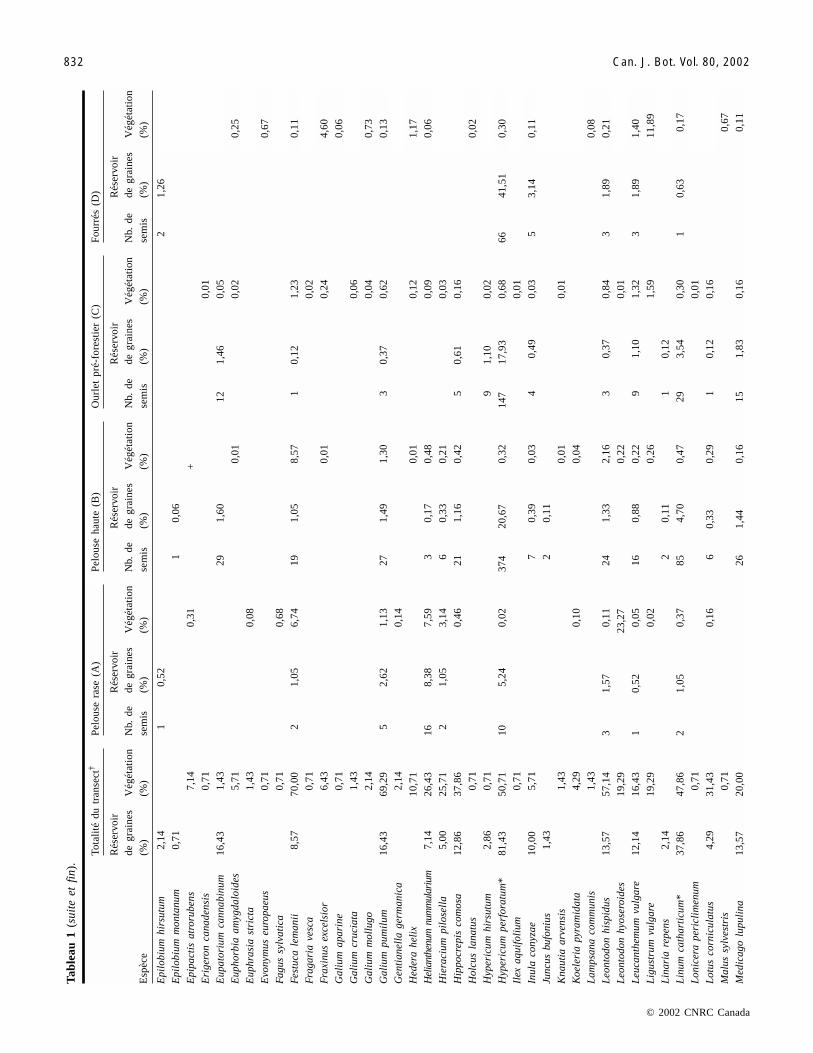
Composition des assemblages d’espèces de la végétationUn total de 104 espèces a été identifié dans la végétation
(tableau 1). Le Brachypode penné (Brachypodium pinnatum)est l’espèce dominante. Elle est présente dans 94 % des rele-vés avec une abondance relative de 33 % de la végétation dutransect.
La classification hiérarchique des 140 relevés de végéta-tion a permis d’identifier quatre principaux groupes corres-pondant aux assemblages d’espèces suivants : A, pelouserase et ouverte; B, pelouse haute; C, ourlet pré-forestier; D,fourré. Ces assemblages d’espèces sont représentés successi-vement le long du premier axe factoriel de l’AFC (fig. 2,AFC : inertie de l’axe 1 = 0,62; pourcentage de variance ex-pliqué par l’axe 1 = 11,8 %; inertie de l’axe 2 = 0,49; pour-centage de variance expliqué par l’axe 2 = 9,46 %; inertietotale = 5,23). La forme du nuage de point de l’AFC, pré-sentant un effet Guttman (Jongman et al. 1987), peut être as-similée à la trajectoire de la succession végétale projetée surl’axe 1 et reconstituée par la modélisation statistique. Cetaxe factoriel représente le gradient de succession naturelleorienté vers le boisement, du pôle positif (assemblage A, pe-louse rase) au pôle négatif (assemblage D, fourré). Lesquatre stades successionnels forment une mosaïque de ta-ches le long du transect (fig. 3). La plupart des espèces pré-sentes dans les pelouses rases (assemblage d’espèces A :Leontodon hyoseroides, Teucrium montanum, Asperula cya-nanchica) sont strictement inféodées à ce milieu qui abritedes espèces rares et protégées (Gentianella germanica, Epi-pactis atrorubens). Au contraire, les espèces dominant les
pelouses hautes (assemblage B : Brachypodium pinnatum,Carex flacca, Teucrium chamaedrys, Festuca lemanii) serencontrent dans une plus large section du transect. Au ni-veau de l’ourlet forestier (assemblage d’espèces C) les espè-ces telles Origanum vulgare, Rubus fruticosus et de jeunesarbustes (Crataegus monogyna, Cornus sanguinea, Rosa ca-nina) deviennent abondantes. Les espèces présentes dans lespremiers stades de succession (assemblages A, B, C) dispa-raissent finalement sous les arbres (Coryllus avellana, Fraxi-nus excelsior dans l’assemblage D), laissant place à d’autresespèces de sous-bois (telles Arum maculatum, Hedera helix)apparaissant dans les fourrés.
Composition du réservoir de graines sous les différentsassemblages d’espèces de la végétation
Un total de 71 espèces a été identifié dans le réservoir degraines permanentes du sol (tableau 1). Seules quelques es-pèces dominent les réservoirs de graines avec des valeurs dedensités élevées (tableau 2). Les espèces principales (Black-stonia perfoliata, Hypericum perforatum et Carex flacca) re-présentent plus de 58 % des semis identifiés, et chacune deces espèces est présente dans plus de 80 % des relevés. Ori-ganum vulgare est la quatrième espèce la plus abondantedans le réservoir de graines mais elle est absente dansl’assemblage A. Cette espèce correspond à 6,7 % des semisidentifiés et est présente dans 56 % des relevés (tableau 1).Les densités de graines de ces quatre espèces principales va-rient selon des patrons différents le long des stades de suc-cession (tableau 2). Les densités de graines de Blackstoniaperfoliata et de Carex flacca sont maximales dans la pelousehaute (assemblage B) et décroissent dans les habitats boisés(assemblages C et D). Les densités de graines de Hypericumperforatum sont les plus faibles dans les pelouses rases (A)du début de la succession mais persistent sous des fourrés(D). Les densités de graines de Origanum vulgare sontmaximales dans l’ourlet pré-forestier, stade intermédiaire dela succession (tableaux 2 et 1). Helianthenum nummulariumet Hieracium pilosella sont des espèces caractéristiques despelouses calcicoles. Leurs graines sont présentes dans les
© 2002 CNRC Canada
830 Can. J. Bot. Vol. 80, 2002
-3
-2
-1
0
1
-4 -3 -2 -1 0 1 2 3
AFC axe 1
AF
Caxe
2
Pelouses rases et ouvertes (A)
Pelouses hautes (B)
Ourlets pré-forestiers (C)
Fourrés (D)
Gradient de
succession
Fig. 2. Analyse factorielle des correspondances (AFC) de la composition en espèces de la végétation des 140 relevés du transect. Lesquatre assemblages d’espèces représentés sur les plans factoriels des relevés de l’AFC sont issus de la classification hiérarchiqueascendante (CH) réalisée sur la végétation.
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:24 AM
Color profile: Generic CMYK printer profileComposite Default screen
© 2002 CNRC Canada
Chabrerie et al. 831
Tota
lité
dutr
anse
ct†
Pel
ouse
rase
(A)
Pel
ouse
haut
e(B
)O
urle
tpr
é-fo
rest
ier
(C)
Four
rés
(D)
Esp
èce
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Nb.
dese
mis
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Nb.
dese
mis
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Nb.
dese
mis
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Nb.
dese
mis
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Ace
rca
mpe
stre
0,71
10,
06
Ach
ille
am
ille
foli
um12
,86
0,10
0,08
0,06
Agr
imon
iaeu
pato
ria
4,29
0,02
0,02
0,02
Agr
osti
sst
olon
ifer
a2,
142,
861
0,06
0,03
20,
240,
011
0,63
Ana
gall
isar
vens
is15
,71
160,
889
1,10
21,
26A
nthy
llis
vuln
erar
ia0,
71+
Arr
hena
ther
umel
atiu
s0,
710,
02A
rum
mac
ulat
um0,
710,
06A
sper
ula
cyna
nchi
ca10
,71
1,59
0,04
Bel
lis
pere
nnis
0,71
10,
06B
etul
apu
besc
ens
9,29
21,
055
0,28
70,
85B
lack
ston
iape
rfol
iata
*87
,86
40,0
074
38,7
40,
4344
724
,71
0,26
135
16,4
60,
1212
7,55
0,07
Bra
chyp
odiu
mpi
nnat
um94
,29
5,83
41,3
934
,48
5,58
Bri
zam
edia
2,86
0,02
0,02
Bry
onia
dioi
ca0,
710,
01B
uple
urum
falc
atum
3,57
0,02
0,01
Cam
panu
laro
tund
ifol
ia2,
140,
10C
ardu
uste
nuif
loru
s3,
570,
010,
06C
arex
cary
ophy
llea
2,14
17,1
44
0,22
0,38
0,97
0,78
Car
exfl
acca
*80
,71
92,8
653
27,7
514
,78
299
16,5
313
,99
116
14,1
58,
3117
10,6
91,
56C
arex
mur
icat
a1,
430,
01C
arli
navu
lgar
is0,
710,
712
0,11
0,01
Cen
taur
eani
gra
1,43
32,8
60,
825
0,28
0,70
0,07
Cen
taur
easc
abio
sa4,
290,
060,
02C
enta
uriu
mer
ythr
aea*
11,4
35,
0019
1,05
0,05
688,
290,
03C
eras
tium
arve
nse
6,43
10,
526
0,33
50,
611
0,63
Che
nopo
dium
albu
m0,
711
0,06
Cir
sium
acau
le2,
1434
,29
8,95
40,
220,
930,
03C
irsi
umar
vens
e0,
710,
01C
irsi
umvu
lgar
e2,
860,
010,
020,
04C
lem
atis
vita
lba
6,43
18,5
77
0,39
0,14
20,
240,
614,
91C
lino
podi
umvu
lgar
e11
,43
36,4
318
1,00
0,19
101,
221,
760,
89C
ornu
ssa
ngui
nea
0,71
58,5
72
0,11
2,86
9,42
19,3
7C
oryl
usav
ella
na7,
86+
0,25
20,7
6C
rata
egus
mon
ogyn
a21
,43
0,16
8,26
1,83
Dac
tyli
sgl
omer
ata
7,14
0,05
0,06
Dap
hne
laur
eola
0,71
0,02
Dau
cus
caro
ta20
,71
48,5
727
1,49
0,81
202,
440,
461
0,63
0,08
Dig
ital
islu
tea
11,4
33,
5716
0,88
+9
1,10
0,01
85,
030,
75E
chiu
mvu
lgar
e1,
432,
142
0,11
0,12
Tab
leau
1.C
ompo
siti
onen
espè
ces
dela
végé
tati
onet
duré
serv
oir
degr
aine
s.
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:25 AM
Color profile: Generic CMYK printer profileComposite Default screen
© 2002 CNRC Canada
832 Can. J. Bot. Vol. 80, 2002
Tota
lité
dutr
anse
ct†
Pel
ouse
rase
(A)
Pel
ouse
haut
e(B
)O
urle
tpr
é-fo
rest
ier
(C)
Four
rés
(D)
Esp
èce
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Nb.
dese
mis
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Nb.
dese
mis
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Nb.
dese
mis
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Nb.
dese
mis
Rés
ervo
irde
grai
nes
(%)
Vég
état
ion
(%)
Epi
lobi
umhi
rsut
um2,
141
0,52
21,
26E
pilo
bium
mon
tanu
m0,
711
0,06
Epi
pact
isat
roru
bens
7,14
0,31
+E
rige
ron
cana
dens
is0,
710,
01E
upat
oriu
mca
nnab
inum
16,4
31,
4329
1,60
121,
460,
05E
upho
rbia
amyg
dalo
ides
5,71
0,01
0,02
0,25
Eup
hras
iast
rict
a1,
430,
08E
vony
mus
euro
paeu
s0,
710,
67Fa
gus
sylv
atic
a0,
710,
68Fe
stuc
ale
man
ii8,
5770
,00
21,
056,
7419
1,05
8,57
10,
121,
230,
11F
raga
ria
vesc
a0,
710,
02F
raxi
nus
exce
lsio
r6,
430,
010,
244,
60G
aliu
map
arin
e0,
710,
06G
aliu
mcr
ucia
ta1,
430,
06G
aliu
mm
ollu
go2,
140,
040,
73G
aliu
mpu
mil
um16
,43
69,2
95
2,62
1,13
271,
491,
303
0,37
0,62
0,13
Gen
tian
ella
germ
anic
a2,
140,
14H
eder
ahe
lix
10,7
10,
010,
121,
17H
elia
nthe
num
num
mul
ariu
m7,
1426
,43
168,
387,
593
0,17
0,48
0,09
0,06
Hie
raci
umpi
lose
lla
5,00
25,7
12
1,05
3,14
60,
330,
210,
03H
ippo
crep
isco
mos
a12
,86
37,8
60,
4621
1,16
0,42
50,
610,
16H
olcu
sla
natu
s0,
710,
02H
yper
icum
hirs
utum
2,86
0,71
91,
100,
02H
yper
icum
perf
orat
um*
81,4
350
,71
105,
240,
0237
420
,67
0,32
147
17,9
30,
6866
41,5
10,
30Il
exaq
uifo
lium
0,71
0,01
Inul
aco
nyza
e10
,00
5,71
70,
390,
034
0,49
0,03
53,
140,
11Ju
ncus
bufo
nius
1,43
20,
11K
naut
iaar
vens
is1,
430,
010,
01K
oele
ria
pyra
mid
ata
4,29
0,10
0,04
Lam
psan
aco
mm
unis
1,43
0,08
Leo
ntod
onhi
spid
us13
,57
57,1
43
1,57
0,11
241,
332,
163
0,37
0,84
31,
890,
21L
eont
odon
hyos
eroi
des
19,2
923
,27
0,22
0,01
Leu
cant
hem
umvu
lgar
e12
,14
16,4
31
0,52
0,05
160,
880,
229
1,10
1,32
31,
891,
40L
igus
trum
vulg
are
19,2
90,
020,
261,
5911
,89
Lin
aria
repe
ns2,
142
0,11
10,
12L
inum
cath
arti
cum
*37
,86
47,8
62
1,05
0,37
854,
700,
4729
3,54
0,30
10,
630,
17L
onic
era
peri
clim
enum
0,71
0,01
Lot
usco
rnic
ulat
us4,
2931
,43
0,16
60,
330,
291
0,12
0,16
Mal
ussy
lves
tris
0,71
0,67
Med
icag
olu
puli
na13
,57
20,0
026
1,44
0,16
151,
830,
160,
11
Tab
leau
1(s
uite
etfi
n).
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:26 AM
Color profile: Generic CMYK printer profileComposite Default screen

© 2002 CNRC Canada
Chabrerie et al. 833O
noni
sre
pens
0,71
+O
riga
num
vulg
are*
55,7
162
,86
113
6,25
1,76
759,
154,
9211
6,92
1,00
Pim
pine
lla
saxi
frag
a1,
430,
020,
01P
lant
ago
lanc
eola
ta1,
4312
,86
10,
060,
191
0,12
0,02
Poa
prat
ensi
s15
,71
50,0
022
1,22
3,61
111,
342,
120,
02Po
lyga
laca
lcar
ea5,
0016
,43
10,
520,
065
0,28
0,09
10,
120,
12Po
tent
illa
rept
ans
2,14
1,43
10,
060,
012
0,24
0,02
Pote
ntil
last
eril
is1,
433,
571
0,06
0,01
10,
120,
040,
17P
rim
ula
veri
s2,
86+
0,05
Pru
nell
avu
lgar
is2,
8617
,14
0,02
20,
110,
185
0,61
1,20
0,78
Pru
nus
spin
osa
3,57
0,02
1,18
Que
rcus
petr
aea
2,86
0,01
Ran
uncu
lus
bulb
osus
5,71
40,0
01
0,52
70,
390,
594
0,49
0,49
0,08
Ran
uncu
lus
repe
ns1,
432,
140,
013
0,37
0,03
Res
eda
lute
a2,
862
1,05
20,
111
0,63
Ros
aca
nina
2,86
35,7
10,
051
0,06
0,59
40,
495,
4110
,83
Rub
usfr
utic
osus
17,8
643
,57
0,06
110,
610,
4416
1,95
4,25
106,
293,
57Sa
mbu
cus
nigr
a4,
294,
293
0,17
+2
0,24
10,
630,
88Sa
ngui
sorb
am
inor
11,4
370
,71
10,
520,
6616
0,88
2,29
20,
240,
410,
06Sc
abio
saco
lum
bari
a0,
7110
,00
0,25
10,
060,
030,
03Se
neci
oja
coba
ea0,
710,
01Se
seli
liba
noti
s0,
710,
02Se
sler
iaal
bica
ns20
,00
12,5
71,
03Si
lene
vulg
aris
0,71
10,
63So
lanu
mdu
lcam
ara
2,86
0,03
0,17
Sola
num
nigr
um2,
141
0,12
31,
89So
lida
govi
rgau
rea
1,43
0,01
Sonc
hus
aspe
r5,
717,
864
0,22
0,04
30,
370,
041
0,63
0,08
Sonc
hus
oler
aceu
s0,
711
0,06
Stel
lari
am
edia
2,14
10,
523
0,17
Succ
isa
prat
ensi
s0,
710,
01Ta
raxa
cum
offi
cina
le1,
4315
,71
10,
060,
101
0,12
0,02
0,06
Teuc
rium
cham
aedr
ys17
,14
80,0
02
1,05
6,29
191,
0510
,09
60,
731,
440,
02Te
ucri
umm
onta
num
0,71
9,29
10,
520,
820,
03Te
ucri
umsc
orod
onia
12,8
637
,86
70,
390,
2718
2,20
2,02
21,
261,
36T
hym
uspr
aeco
x1,
4337
,14
2,31
20,
110,
380,
14Tr
ifol
ium
repe
ns0,
711
0,52
Urt
ica
dioi
ca23
,57
0,71
84,
1947
2,60
161,
954
2,52
0,06
Verb
ascu
mth
apsu
s1,
432,
141
0,06
10,
630,
17Ve
rben
aof
fici
nali
s2,
142
0,11
10,
63Ve
roni
cach
amae
drys
0,71
10,
63Ve
roni
cam
onta
na0,
711
0,12
Viol
ahi
rta
12,1
455
,71
0,02
40,
220,
6025
3,05
3,16
1,19
Indé
term
inée
1,43
10,
521
0,06
*Les
espè
ces
poss
édan
tle
spl
usfo
rtes
dens
ités
degr
aine
sda
nsle
rése
rvoi
rde
grai
nes.
† Pour
cent
ages
depr
ésen
cede
ses
pèce
sda
nsle
sre
levé
sdu
rése
rvoi
rde
grai
nes
etde
lavé
géta
tion.
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:27 AM
Color profile: Generic CMYK printer profileComposite Default screen

premiers stades de la succession (assemblages A et B) et dis-paraissent sous une végétation boisée dans les assemblagesC et D (tableau 1). Les espèces arbustives (Cornus sangui-nea, Rosa canina, Sambucus nigra) sont peu représentéesdans le réservoir de graines, composant seulement 2,56 % dutotal des semis identifiés.
Les différences de densités de semis entre assemblagessont significatives (F[3,139] = 9,37; p < 0,001). Les tests decomparaisons de moyennes Tukey–Kramer indiquent que lespaires d’assemblage (B et C) et (A et D) présentent des dif-férences significatives. Les densités de graines sont élevéesdans les stades intermédiaires de la succession (assemblagesB et C) (tableau 2) et sont plus faibles au niveau des extré-mités du gradient (assemblages A et D). La présenced’arbres a un effet négatif sur la densité de graines (seule-ment 4326 graines en moyenne par m2 dans les fourrés)alors qu’au contraire l’ourlet, où se rencontrent à la fois lesespèces caractéristiques de la pelouse et celles de la forêt,possède la densité de graines la plus forte (8530 graines enmoyenne par m2).
Comparaison de la diversité de la végétation et duréservoir de graines
Les résultats de l’analyse de la diversité de la végétationet du réservoir de graines dans les quatre assemblagesd’espèces du transect sont donnés dans le tableau 3. La com-paraison de la végétation avec le réservoir de graines(ANOVA) montre que la richesse spécifique (F[1,279] =674,04; p < 0,001) et l’indice de Shannon (F[1,279] = 14,96;p < 0,001) sont plus faibles dans le réservoir de graines quedans la végétation (tableau 3). L’équitabilité J′ est dans tousles cas plus importante (F[1,279] = 273,37; p < 0,001) dans leréservoir de graines que dans la végétation, qui contient unplus grand nombre d’espèces à faible abondance coexistantavec quelques espèces dominantes.
La similarité entre la végétation et le réservoir de grainesest significativement plus élevée dans les assemblagesd’espèces B et C que dans les assemblages A et D (F[3,139] =8,32; p < 0,001) (tableau 4a). La similarité moyenne la plusfaible a été enregistrée au niveau des fourrés (26 %) où lenombre d’espèces communes à la végétation et au réservoirde graines est restreint (25 espèces présentes dans le réser-voir de graines contre 52 dans la végétation en place).L’indice de similarité a tendance à varier parallèlement à larichesse du réservoir de graines.
Comparaison de la diversité du réservoir de grainesentre les stades successionnels
Les ANOVA ont montré des différences significatives dela diversité du réservoir de graines entre les quatre assembla-ges d’espèces (A, B, C et D). La richesse spécifique R(F[3,139] = 14,38; p < 0,001) et l’indice de Shannon H′(F[3,139] = 20,58; p < 0,001) diffèrent de façon significativeentre les paires d’assemblages (B et C) et (A et D). Dans leréservoir de graines, R et H′ sont significativement plus éle-vés dans les assemblages B et C que dans les assemblages Aet D. L’équitabilité J′ ne varie pas de façon significativeentre les quatre assemblages d’espèces dans le réservoir degraines.
Les calculs de similarité des pools d’espèces de la végéta-tion et du réservoir de graines entre les différents assembla-
ges d’espèces (tableau 4b) montrent que : (i) les réservoirsde graines sont plus similaires entre eux qu’avec la végéta-tion quels que soient les stades de succession considérés;(ii) les réservoirs de graines des premiers stades de la suc-cession (assemblage A) sont plus similaires avec la végéta-tion de ces stades qu’avec la végétation des stades finaux;(iii) les réservoirs de graines des stades intermédiaires (as-semblages B et C) sont plus similaires avec la végétation desstades finaux (assemblage D) qu’avec la végétation dumême stade (pour l’assemblage B) ou des stades antérieurs(pour les assemblages B et C).
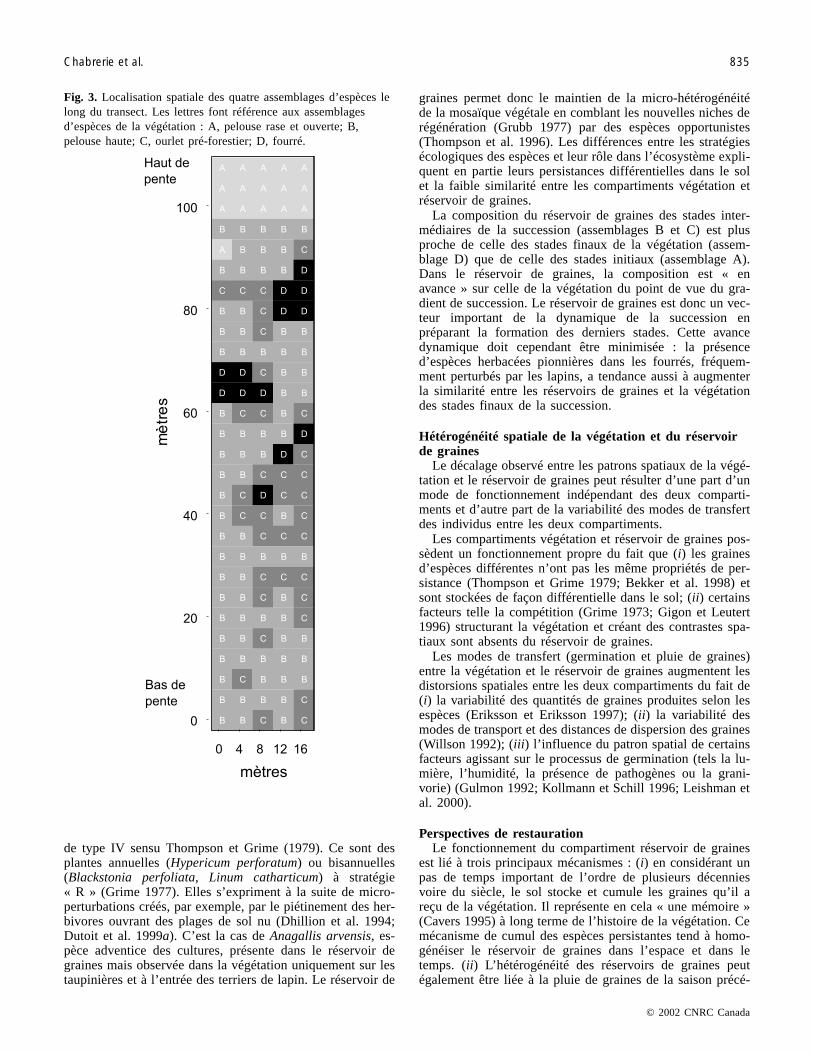
Analyse spatiale de la végétation et du réservoir degraines
Des courbes de richesse spécifique cumulée (Ferry etFrochot 1970), réalisées de façon préliminaire, présententdes pentes et des paliers différents selon les assemblagesd’espèces. Elles mettent en évidence des structures spatialesdifférentes entre la composition de la végétation et du réser-voir de graines. L’étude comparative des patrons spatiaux decomposition et de richesse spécifique met en évidence desdivergences entre la végétation et le réservoir de graines lelong du transect (fig. 4). La composition de la végétation(fig. 4a) présente de forts contrastes autour de trois taches defourré où les espèces de pelouse sont substituées par des es-pèces d’ourlet ou pré-forestières. Ces taches renferment unerichesse spécifique faible pour la végétation (fig. 4b) et pourle réservoir de graines (fig. 4c). Les points où la richessespécifique de la végétation et du réservoir de graines est laplus élevée sont situés dans les ourlets autour des taches ar-bustives. Cependant, ces zones de richesse élevée de la végé-tation (fig. 4b) ne sont pas toujours superposables à ceux duréservoir de graines (fig. 4c). Il existe donc un décalage spa-tial des patrons de richesse spécifique. Le patron spatial del’indice de similarité (fig. 4d) le long du transect est compa-rable à celui de la richesse spécifique du réservoir de graines(fig. 4c).
Discussion
Afin de répondre aux hypothèses, les différences entre lavégétation et le réservoir de graines sont abordées (i) le longdu gradient de succession secondaire et (ii) en comparantl’hétérogénéité des deux compartiments. Le rôle du réservoirde graines sera finalement discuté du point de vue de laconservation des pelouses calcicoles.
Gradient de successionDe nombreuses études confirment l’effet négatif à long
terme de la colonisation par les ligneux sur la richesse duréservoir de graines du sol (Donelan et Thompson 1980;Davies et Waite 1998; Kalamees et Zobel 1998). Les espècesqui disparaissent du réservoir de graines en fin du gradientde succession sont essentiellement des espèces de pelouse.De plus, les densités de graines de la majorité des espècesdiminuent le long du gradient de succession.
La faible similarité entre la végétation et le réservoir degraines montre que la composition des pelouses calcicolesne se retrouve pas dans le réservoir de graines du sol. Dansnotre site d’étude, les espèces dominant le réservoir de grai-nes sont essentiellement des espèces à graines persistantes
© 2002 CNRC Canada
834 Can. J. Bot. Vol. 80, 2002
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:28 AM
Color profile: Generic CMYK printer profileComposite Default screen
de type IV sensu Thompson et Grime (1979). Ce sont desplantes annuelles (Hypericum perforatum) ou bisannuelles(Blackstonia perfoliata, Linum catharticum) à stratégie« R » (Grime 1977). Elles s’expriment à la suite de micro-perturbations créés, par exemple, par le piétinement des her-bivores ouvrant des plages de sol nu (Dhillion et al. 1994;Dutoit et al. 1999a). C’est la cas de Anagallis arvensis, es-pèce adventice des cultures, présente dans le réservoir degraines mais observée dans la végétation uniquement sur lestaupinières et à l’entrée des terriers de lapin. Le réservoir de
graines permet donc le maintien de la micro-hétérogénéitéde la mosaïque végétale en comblant les nouvelles niches derégénération (Grubb 1977) par des espèces opportunistes(Thompson et al. 1996). Les différences entre les stratégiesécologiques des espèces et leur rôle dans l’écosystème expli-quent en partie leurs persistances différentielles dans le solet la faible similarité entre les compartiments végétation etréservoir de graines.
La composition du réservoir de graines des stades inter-médiaires de la succession (assemblages B et C) est plusproche de celle des stades finaux de la végétation (assem-blage D) que de celle des stades initiaux (assemblage A).Dans le réservoir de graines, la composition est « enavance » sur celle de la végétation du point de vue du gra-dient de succession. Le réservoir de graines est donc un vec-teur important de la dynamique de la succession enpréparant la formation des derniers stades. Cette avancedynamique doit cependant être minimisée : la présenced’espèces herbacées pionnières dans les fourrés, fréquem-ment perturbés par les lapins, a tendance aussi à augmenterla similarité entre les réservoirs de graines et la végétationdes stades finaux de la succession.
Hétérogénéité spatiale de la végétation et du réservoirde graines
Le décalage observé entre les patrons spatiaux de la végé-tation et le réservoir de graines peut résulter d’une part d’unmode de fonctionnement indépendant des deux comparti-ments et d’autre part de la variabilité des modes de transfertdes individus entre les deux compartiments.
Les compartiments végétation et réservoir de graines pos-sèdent un fonctionnement propre du fait que (i) les grainesd’espèces différentes n’ont pas les même propriétés de per-sistance (Thompson et Grime 1979; Bekker et al. 1998) etsont stockées de façon différentielle dans le sol; (ii) certainsfacteurs telle la compétition (Grime 1973; Gigon et Leutert1996) structurant la végétation et créant des contrastes spa-tiaux sont absents du réservoir de graines.
Les modes de transfert (germination et pluie de graines)entre la végétation et le réservoir de graines augmentent lesdistorsions spatiales entre les deux compartiments du fait de(i) la variabilité des quantités de graines produites selon lesespèces (Eriksson et Eriksson 1997); (ii) la variabilité desmodes de transport et des distances de dispersion des graines(Willson 1992); (iii) l’influence du patron spatial de certainsfacteurs agissant sur le processus de germination (tels la lu-mière, l’humidité, la présence de pathogènes ou la grani-vorie) (Gulmon 1992; Kollmann et Schill 1996; Leishman etal. 2000).
Perspectives de restaurationLe fonctionnement du compartiment réservoir de graines
est lié à trois principaux mécanismes : (i) en considérant unpas de temps important de l’ordre de plusieurs décenniesvoire du siècle, le sol stocke et cumule les graines qu’il areçu de la végétation. Il représente en cela « une mémoire »(Cavers 1995) à long terme de l’histoire de la végétation. Cemécanisme de cumul des espèces persistantes tend à homo-généiser le réservoir de graines dans l’espace et dans letemps. (ii) L’hétérogénéité des réservoirs de graines peutégalement être liée à la pluie de graines de la saison précé-
© 2002 CNRC Canada
Chabrerie et al. 835
A A A A A
A A A A A
A A A A A
B B B B B
A B B B C
B B B B D
C C C D D
B B C D D
B B C B B
B B B B B
D D C B B
D D D B B
B C C B C
B B B B D
B B B D C
B B C C C
B C D C C
B C C B C
B B C C C
B B B B B
B B C C C
B B C B C
B B B B C
B B C B B
B B B B B
B C B B B
B B B B C
B B C B C
Haut de
pente
Bas de
pente
0 4 8 12 16
mètres
0
20
40
60
80
100
mè
tre
s
Fig. 3. Localisation spatiale des quatre assemblages d’espèces lelong du transect. Les lettres font référence aux assemblagesd’espèces de la végétation : A, pelouse rase et ouverte; B,pelouse haute; C, ourlet pré-forestier; D, fourré.
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:28 AM
Color profile: Generic CMYK printer profileComposite Default screen
dente. Cette pluie de graines fournit des espèces à grainespersistantes et transitoires (Thompson et Grime 1979). Ellepeut être affectée entre autres par la variabilité saisonnière etspatiale du pâturage (Erkkila 1998; Woldu et Saleem 2000).(iii) Le dernier mécanisme concerne le recrutement des grai-nes du sol par la voie de la germination. Le sol représente unréservoir d’espèces disponibles pour la régénération futurede la végétation (Willems et Bik 1998).
Dans notre cas, la diversité enregistrée dans la végétation(104 espèces) n’est pas entièrement « gardée en mémoire »par le sol (71 espèces dont 51 communes avec la végéta-tion). De plus, les espèces dominant le réservoir de grainesn’ont pas les contributions les plus importantes dans la végé-tation; Blackstonia perfoliata représente 22 % des graines du
sol contre 0,2 % de la végétation. Les abondances desespèces dans le réservoir de graines ne sont donc pas propor-tionnelles à leur capacité d’expression dans la végétation.Cette observation confirme le rôle important de la multipli-cation végétative dans le développement et le maintiend’une pelouse calcicole (Schlapfer et Fischer 1998). Certai-nes graines transitoires ne sont pas échantillonnées dans leréservoir de graines permanente (Thompson et Grime 1979).Elles peuvent cependant participer à la composition descommunautés végétales sans se maintenir dans le réservoirde graines. Pour ces raisons, les perspectives de restaurationdu réservoir total des espèces d’une pelouse calcicole uni-quement par le réservoir de graines sont peu intéressantes.La plupart des études confirment ce constat (Dutoit et Alard
© 2002 CNRC Canada
836 Can. J. Bot. Vol. 80, 2002
Pelouse rase (A) Pelouse haute (B) Ourlet pré-forestier (C) Fourré (D)
Espèces principales Densité % Densité % Densité % Densité %
Blackstonia perfoliata 1636 39 2053 25 1404 16 326 8Carex flacca 1172 28 1373 17 1207 14 463 11Centaurium erythraea 707 8Daucus carota 208 2Digitalis lutea 218 5Galium pumilum 111 3Helianthenum nummularium 354 8Hypericum perforatum 221 5 1718 21 1529 18 1796 42Inula conyzae 136 3Linum catharticum 390 5 302 4Origanum vulgare 519 6 780 9 299 7Rubus fruticosusTeucrium scorodonia 187 2 272 6Urtica dioica 177 4 216 3 109 3Viola hirta 260 3Autres 553 13 2039 23 1945 24 707 15Densité totale 4222 8309 8530 4326
Tableau 2. Densités de graines (nb. de graines/m2) et fréquences relatives (%) des principales espèces (fréquence supérieure à 2 %)dans le réservoir de graines sous les quatre assemblages d’espèces (A, B, C et D) le long du transect.
Pelouse rase (A) Pelouse haute (B) Ourlet pré-forestier (C) Fourré (D) Ensemble des relevés
(a) Réservoir de grainesNb. de points 16 77 34 13 140Nb. total de semis 191 1809 820 159 2979Nb. moyen de semis par
point ± écart type11,94±1,94 23,49±2,58 24,12±2,80 12,23±3,30 21,28±3,30
(b) Diversité du réservoir de graines et de la végétation
(A) (B) (C) (D) Ensemble
R.G. Vég. R.G. Vég. R.G. Vég. R.G. Vég. R.G. Vég.
Richesse spécifique totale 23 37 62 80 45 72 25 52 71 104Richesse spécifique moyenne 4,19 17,75 7,91 18,32 8,68 17,94 5,23 14,62 7,42 17,82H ′ moyen 1,58 2,79 2,46 2,56 2,61 2,62 1,75 2,27 2,33 2,58J ′ moyen 0,80 0,67 0,85 0,61 0,87 0,64 0,78 0,60 0,84 0,62Total Rréservoir/total Rvégét. 0,62 0,78 0,63 0,48 0,68Rmoyenne réservoir/Rmoyenne végét. 0,24 0,43 0,48 0,36 0,42
Nota : Les moyennes de R, H ′ et J ′ ont été calculées pour chaque assemblage d’espèces. Richesse spécifique totale : stock d’espèces présentes danschaque assemblage d’espèces; R, richesse spécifique; H ′, indice de Shannon; J ′, équitabilité; R.G., réservoir de graines; Vég., végétation.
Tableau 3. Diversité de la végétation et du réservoir de graines parmi les quatre assemblages d’espèces (A, B, C et D).
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:29 AM
Color profile: Generic CMYK printer profileComposite Default screen
1995; Bakker et al. 1996b) excepté dans certains milieuxcomme les marais salés où la diversité de la végétation estdéjà très faible (Egan et Ungar 2000).
Les réservoirs de graines sous-jacents à une végétation ar-bustive sont appauvris en espèces. La modification de lacomposition du réservoir de graines le long du gradient desuccession ne permet pas de restaurer les stades antérieurs àla colonisation par les ligneux. Une gestion du milieu par undébroussaillage unique ne suffit pas à réorienter la dyna-mique de l’écosystème vers la pelouse initiale (Davies etWaite 1998; Stampfli et Zeiter 1999). D’autres techniquescomme l’apport de graines (Stevenson et al. 1997), notam-ment par les animaux d’élevages (Ghassali et al. 1998), ou latransplantation d’individus accompagnée d’un entretien ré-gulier (Davies et al. 1999; Van der Putten et al. 2000) se sontrévélées être plus efficaces.
La plupart des travaux de restauration des milieux à partirdu réservoir de graines se préoccupent du pool globald’espèces dormant dans le sol et ne tiennent pas compte del’aspect spatial. Or, le mode d’échantillonnage spatialementexplicite de notre étude suggère l’importance de l’hétéro-généité spatiale dans le fonctionnement des pelouses calcicoles.Chaque stade dynamique de la pelouse nécessite un pland’échantillonnage différent (nombre de prélèvements, vo-lume minimum de sol, distance entre les prélèvements) dansl’espace pour qu’il soit représentatif du pool d’espèces. Les
différents stades successionnels fonctionnent selon leurs pro-pres échelles de temps et d’espace.
L’hétérogénéité du réservoir de graines ne paraît pasessentielle pour maintenir l’hétérogénéité de la végétation.Le réservoir de graines contient aussi des espèces n’existantpas dans la végétation et qui peuvent s’exprimer suite àdes micro-perturbations (Rusch et van der Maarel 1992).D’autres espèces du réservoir de graines sont le témoin depratiques culturales anciennes comme la mise en labour.Parmi ces espèces qui sont des espèces généralistes (Cheno-podium album), il existe des espèces plus rares (Ajugachamaepitys) capables de s’exprimer à nouveau dans desopérations de restauration de vignoble ou de labour (Dutoitet al. 1999b). Les réservoirs de graines constituent donc unréservoir d’individus dormant, utile au maintien de certainespopulations (Menges 2000; Reinhold 2000) et à la survie desespèces rares (Sutcliffe et Kay 2000).
Remerciements
Les auteurs sont reconnaissants envers Jan Bakker (Uni-versité de Groningen, Pays-Bas) pour ses commentaires surle manuscrit. Les auteurs remercient également Mme IsabellePiquerez, M. Gilbert Mengy, M. Joël Hauville et M. LoicLecorgne du Jardin Botanique et le Service des EspacesVerts de la Ville de Rouen pour le soutien matériel de cette
© 2002 CNRC Canada
Chabrerie et al. 837
(a) Similarité moyenne entre la végétation et le réservoir de graines
A B C D TotalNb. de points 16 77 34 13 140Similarité moyenne (%) 30,43 46,39 46,25 26,23 41,13Nb. d’espèces communes 14 45 37 16 51
(b) Similarité des pools d’espèces entre les différents assemblages
A B C DVégétation/végétationA — 42,55 32,65 24,49B — — 54,72 44,44C — — — 55,10D — — — —Végétation/réservoir de graines
A 29,55 25,00 24,56 12,96B 23,44 37,31 32,86 15,28C 22,95 41,94 43,55 19,70D 21,67 40,98 38,10 26,23Réservoir de graines/réservoir de graines
A — 27,91 24,44 24,32B — — 53,33 40,48C — — — 33,33D — — — —
Nota : Assemblages d’espèces : A, pelouse rase et ouverte; B, pelouse haute; C, ourlet pré-forestier;D, fourré.
Tableau 4. Similarité moyenne (indice de Jaccard en pourcentage) comparant (a) la com-position floristique de la végétation et du réservoir de graines dans les quatre assemblagesd’espèces et (b) similarité des pools d’espèces de la végétation et du réservoir de grainesentre les différents stades de succession (calculée à partir de 13 points pris au hasard danschaque stade).
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:29 AM
Color profile: Generic CMYK printer profileComposite Default screen
© 2002 CNRC Canada
838 Can. J. Bot. Vol. 80, 2002
étude, M. Nicolas Courtier, M. Philippe Housset du Conser-vatoire des Sites Naturels de Haute-Normandie pourl’autorisation de travailler dans la Réserve Naturelled’Hénouville, M. Théophile Cousin, M. Ludovic Romain etl’équipe du chantier du Conservatoire des Sites Naturelspour l’aide logistique, ainsi que Mme Isabelle Poudevigne,M. Thibaud Decaëns et M. Ivan Mirkovic (Université deRouen, France) pour leur aide à la récolte des données. Cetteétude a été financée par le programme « Recréer la Nature2000 » du Ministère de l’Environnement (conventionDGAD/SRAE/95067/MNHN).
Bibliographie
Arévalo, J.R., et Fernández-Palacios, J.M. 2000. Seed bank analy-sis of tree species in two stands of the Tenerife laurel forest (Ca-nary Islands). For. Ecol. Manage. 130 : 177–185.
Armesto, J.J., Pickett, S.T.A., et McDonnell, M.J. 1991. Spatial he-terogeneity during succession: a cyclic model of invasion andexclusion. Dans Ecological heterogeneity. Éditeurs : J. Kolassaet S.T.A. Pickett. Springer-Verlag, New York. pp. 256–269.
Bakker, J.P., Poschlod, P., Strykstra, R.J., Bekker, R.M., etThompson, K. 1996a. Seed banks and seed dispersal: important
(a) (b) (c) (d)
0 5 10 150
20
40
60
80
100
2.9
4.8
4.8
4.8
6.6
6.6
6.6
6.6
6.6
6.6
8.5
10.4
12.3
14.1
0 5 10 150
20
40
60
80
100
10.3
12.5
12.5
14.8
14.8
17.017.0
17.0
17.0
17.0
19.3
19.3
19.3
19.3
19.3
19.3
21.5
21.5
21.5
0 5 10 150
20
40
60
80
100
0.1
0.1 0.2
0.2
0.20.2
0.2
0.2
0.2
0.2
0.2
0.2
0.3
0.3
0.3
0.3
0.3
0.3
0.4
0 5 10 15
0
20
40
60
80
100
mèt
res
-2.8
-2.3
-1.8
-1.8
-1.2
-1.2
-0.7
-0.7
-0.7
-0.7
-0.2-0.2
0.3
0.3
0.3
0.3
0.8
mètres mètres mètres mètres
Fig. 4. Patrons spatiaux de la végétation et du réservoir de graines. Les cartographies ont été réalisées avec le logiciel S-PLUS v.2000 : (a) coordonnées factorielles des relevés le long de l’axe 1 de l’AFC réalisée sur la végétation, (b) richesse spécifique de lavégétation, (c) richesse spécifique du réservoir de graines, (d) similarité (indice de Jaccard) entre la végétation et le réservoir degraines. Les flèches noires indiquent le centre des trois zones de fourré bordées des ourlets pré-forestiers.
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:29 AM
Color profile: Generic CMYK printer profileComposite Default screen

© 2002 CNRC Canada
Chabrerie et al. 839
topics in restoration ecology. Acta Bot. Neerl. 45 : 461–490.Bakker, J.P., Bakker, E.S., Rosen, E., Verweij, G.L., et Bekker, R.M.
1996b. Soil seed bank composition along a gradient from dry al-var grassland to Juniperus shrubland. J. Veg. Sci. 7 : 165–176.
Bekker, R.M., Bakker, J.P., Grandin, U., Kalamees, R., Milberg, P.,Poschlod, P., Thompson, K., et Willems, J.H. 1998. Seed size,shape and vertical distribution in the soil: indicators of seed lon-gevity. Funct. Ecol. 12 : 834–842.
Bertiller, M.B. 1998. Spatial patterns of the germinable soil seedbank in northern Patagonia. Seed Sci. Res. 8 : 39–45.
Cavers, P.B. 1995. Seed banks: memory in soil. Can. J. Soil Sci.75 : 11–13.
Davies, A., et Waite, S. 1998. The persistence of calcareous grass-land species in the soil seed bank under developing and establis-hed scrub. Plant Ecol. 136 : 27–39.
Davies, A., Dunnett, N.P., et Kendle, T. 1999. The importance oftransplant size and gap width in the botanical enrichment of spe-cies-poor grasslands in Britain. Restor. Ecol. 7 : 271–280.
De Foucault, B., et Frileux, P.N. 1988. Étude phytosociologiquedes corniches et côtes calcaires de la basse vallée de la Seine(des Andelys à Rouen). Doc. Phytosociol. 11 : 159–183.
Dhillion, S.S., McGinley, M.A., Friese, C.F., et Zak, J.C. 1994.Construction of sand shinnery oak communities of the LlanoEstacado: animal disturbances, plant community structure, andrestoration. Restor. Ecol. 2 : 51–60.
Donelan, M., et Thompson, K. 1980. Distribution of buried viableseeds along a successional series. Biol. Conserv. 17 : 297–311.
Dutoit, T., et Alard, D. 1995. Permanent seed banks in chalk grass-land under various management regimes: their role in the resto-ration of species-rich plant communities. Biodivers. Conserv. 4 :939–950.
Dutoit, T., et Alard, D. 1996a. Les pelouses calcicoles du Nord-Ouest de l’Europe (Brometalia erecti Br. Bl., 1936) : analyse bi-bliographique. Ecologie, 27 : 1–32.
Dutoit, T., et Alard, D. 1996b. Biodiversité actuelle et potentielledes écosystèmes prairiaux calcicoles : influence de la gestionsur les phytocénoses et les banques de graines. Acta Bot. Gal-lica, 143 : 431–440.
Dutoit, T., Roche, P., et Alard, D. 1999a. Influence de perturba-tions anthropiques sur la composition et la diversité botaniquedes pelouses calcicoles de la vallée de la Seine en Haute-Normandie (France). Can. J. Bot. 77 : 377–388.
Dutoit, T., Hill, B., Mahieu, P., et Gerbaud, E. 1999b. Restaurationet conservation de communautés d’espèces ségétales in situ : descoteaux de la basse vallée de la Seine aux cultures extensives duParc Naturel Régional du Lubéron. Bull. Soc. Bot. Centre-Ouest, 19(no spéc.) : 303–316.
Edwards, G.R., et Crawley, M.J. 1999. Herbivores, seed banks andseedling recruitment in mesic grassland. J. Ecol. 87 : 423–435.
Egan, T.P., et Ungar, I.A. 2000. Similarity between seed banks andabove-ground vegetation along a salinity gradient. J. Veg. Sci.11 : 189–194.
Eriksson, A., et Eriksson, O. 1997. Seedling recruitment in semi-natural pastures: the effects of disturbance, seed size, phenologyand seed bank. Nord. J. Bot. 17 : 469–482.
Erkkila, H.M.J.B. 1998. Seed banks of grazed and ungrazed Balticseashore meadows. J. Veg. Sci. 9 : 395–408.
Ferry, C., et Frochot, B. 1970. L’avifaune nidificatrice d’une forêtde Chêne pédonculés en Bourgogne : étude de deux successionsécologiques. Terre Vie, 24 : 153–250.
Ghassali, F., Osman, A.E., et Cocks, P.S. 1998. Rehabilitation ofdegraded grasslands in north Syria: The use of Awassi sheep todisperse the seeds of annual pasture legumes. Exp. Agric. 34 :
391–405.Gigon, A., et Leutert, A.G. 1996. The dynamic keyhole key model
of coexistence to explain diversity of plants in limestone andother grasslands. J. Veg. Sci. 7 : 29–40.
Graham, D.J., et Hutchings, M.J. 1988. Estimation of the seedbank of a chalk grassland ley established on former arable land.J. Appl. Ecol. 25 : 241–252.
Grime, J.P. 1973. Competitive exclusion in herbaceous vegetation.Nature (London), 242 : 344–347.
Grime, J.P. 1977. Evidence for the existence of three primary stra-tegies in plants and its relevance to ecological and evolutionarytheory. Am. Nat. 111 : 1169–1194.
Grubb, P.J. 1977. The maintenance of species-richness in plantcommunities: the importance of the regeneration niche. Biol.Rev. 52 : 107–145.
Gulmon, S.L. 1992. Patterns of seed germination in Californianserpentine grassland species. Oecologia, 89 : 27–31.
Hanf, M. 1983. The arable weeds of Europe with their seedlingsand seeds, BASF, R.U.
Henderson, C.B., Petersen, K.E., et Redak, R.A. 1988. Spatial andtemporal patterns in the seed bank and vegetation of a desertgrassland community. J. Ecol. 76 : 717–728.
Hill, M.O. 1973. Reciprocal averaging: an eigenvector method ofordination. J. Ecol. 61 : 237–249.
Hillier, S.H. 1990. Gaps, seed banks and plant species diversity incalcareous grasslands. Dans Calcareous grasslands — ecologyand management. Proceedings of a BES/NCC symposium atUniversity of Sheffield, 14–16 septembre 1987. Éditeurs :S.H. Hillier, D.W.A. Walton et D.A. Wells. Bluntisham books,Bluntisham. pp. 57–66.
Holzapfel, C., Schmidt, W., et Shmida, A. 1993. The role of seedbank and seed rain in the recolonization of disturbed sites alongan aridity gradient. Phytocoenologia, 23 : 561–580.
Hutchings, M.J., et Booth, K.D. 1996. Studies on the feasibility ofre-creating chalk grassland vegetation on ex-arable land. 1. thepotential roles of the seed bank and the seed rain. J. Appl. Ecol.33 : 1171–1181.
Hyatt, L.A., et Casper, B.B. 2000. Seed bank formation duringearly secondary succession in a temperate decidous forest. J.Ecol. 88 : 516–527.
Inouye, R.S., Huntly, N.J., Tilman, D., Tester, J.R., Stillwell, M., etZinnel, K.C. 1987. Old-field succession on a Minnesota sand-plain. Ecology, 68 : 12–26.
Jaccard, P. 1912. The distribution of the flora of the alpine zone.New Phytol. 11 : 37–50.
Jongman, R.H.G., ter Braak, C.F.J., et Tongeren, O.F.R. 1987.Dans Data analysis in community and landscape ecology. Cam-bridge University Press, Cambridge.
Kalamees, R., et Zobel, M. 1998. Soil seed bank composition in dif-ferent successional stages of a species rich wooded meadow inLaelatu, western Estonia. Acta Oecol.-Int. J. Ecol. 19 : 175–180.
Kolasa, J., et Pickett, S.T.A. 1991. Dans Ecological heterogeneity.Éditeurs : J. Kolasa et S.T.A. Pickett. Springer-Verlag, NewYork.
Kollmann, J., et Schill, H.-P. 1996. Spatial patterns of dispersal,seed predation and germination during colonization of abando-ned grassland by Quercus petraea and Corylus avellana. Vege-tatio, 125 : 193–205.
Lambinon, J., De Langhe, J.-E., Delvosalle, L., et Duvigneaud, J.1992. Nouvelle flore de Belgique, du Grand Duché du Luxem-bourg, du Nord de la France et des régions voisines. Édition duJardin botanique national de Belgique, Meise.
Leishman, M.R., Masters, G.J., Clarke, I.P., et Brown, V.K. 2000.
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:29 AM
Color profile: Generic CMYK printer profileComposite Default screen

© 2002 CNRC Canada
840 Can. J. Bot. Vol. 80, 2002
Seed bank dynamics: The role of fungal pathogens and climatechange. Funct. Ecol. 14 : 293–299.
MathSoft. 1999. S-PLUS 2000 Professional edition for Windows.Copyright Lucent Technologies, Inc., Kanata, Ont.
Menges, E.S. 2000. Applications of population viability analysesin plant conservation. Ecol. Bull. 48 : 73–84.
Milberg, P. 1995. Soil seed bank after eighteen years of successionfrom grassland to forest. Oikos, 72 : 3–13.
Muller, F.M. 1978. Seedlings of the north-western europeanlowland. Dans A flora of seedlings. Centre for AgriculturalPublishing and Documentation of Wageningen, Dr W. Junk B.V.Publishers, The Hague.
Partel, M., Zobel, M., Zobel, K., et van der Maarel, E. 1996. Thespecies pool and its relation to species richness: evidence fromEstonian plant communities. Oikos, 75 : 11–117.
Peco, B., Ortega, M., et Levassor, C. 1998. Similarity between seedbank and vegetation in Mediterranean grassland: a predictivemodel. J. Veg. Sci. 9 : 1–14.
Pickett, S.T.A. 1978. Patch dynamics and the design of nature re-serves. Biol. Conserv. 13 : 27–37.
Pickett, S.T.A., et Cadenasso, M.L. 1995. Landscape ecology: spa-tial heterogeneity in ecological systems. Science (Washington,D.C.), 269 : 331–335.
Pielou, E.C. 1966. The measures of diversity in different types ofbiological collections. J. Theor. Biol. 13 : 131–144.
Reinhold, K. 2000. Maintenance of a genetic polymorphism byfluctuating selection on sex-limited traits. J. Evol. Biol. 13 :1009–1014.
Roberts, H.A. 1981. Seed banks in soils. Adv. Appl. Biol. 6 : 1–55.Roux, M. 1985. Algorythmes de classification. Masson, Paris.Rusch, G. 1992. Spatial pattern of seedling recruitment at two dif-
ferent scales in a limestone grassland. Oikos, 65 : 433–442.Rusch, G., et van der Maarel, E. 1992. Species turnover and see-
dling recruitment in limestone grasslands. Oikos, 63 : 139–146.Schlapfer, F., et Fischer, M. 1998. An isozyme study of clone
diversity and relative importance of sexual and vegetative re-cruitment in the grass Brachypodium pinnatum. Ecography, 21 :351–360.
Shannon, C.E., et Weaver, W. 1949. The mathematical theory ofcommunication. University of Illinois Press, Urbana, Ill.
Sokal, R.R., et Rohlf, F.J. 1981. Biometry. Freeman, W.H. et Co.,San Francisco, Calif.
Stampfli, A., et Zeiter, M. 1999. Plant species decline due to aban-donment of meadows cannot easily be reversed by mowing. Acase study from the southern Alps. J. Veg. Sci. 10 : 151–164.
Stevenson, M.J., Ward, L.K., et Pywell, R.F. 1997. Re-creatingsemi-natural communities: vacuum harvesting and hand collec-tion of seed on calcareous grassland. Restor. Ecol. 5 : 66–76.
Sutcliffe, O.L., et Kay, Q.O.N. 2000. Changes in the arable flora ofcentral southern England since the 1960s. Biol. Conserv. 93 : 1–8.
Templeton, A.R., et Levin, D.A. 1979. Evolutionary consequencesof seed pools. Am. Nat. 114 : 232–249.
Thioulouse, J., Chessel, D., Doledec, S., et Olivier, J.M. 1997.ADE-4: a multivariate analysis and graphical display software.Stat. Comput. 7 : 75–83.
Thompson, K., et Grime, J.P. 1979. Seasonal variation in the seedbanks of herbaceous species in ten contrasting habitats. J. Ecol.67 : 893–921.
Thompson, K., Hillier, S.H., Grime, J.P., Bossard, C.C., etBand, S.R. 1996. A functional analysis of a limestone grasslandcommunity. J. Veg. Sci. 7 : 371–380.
United States Department of Agriculture. 1998. Keys to soil taxo-nomy. Soil survey staff, United States Department of Agricul-ture (USDA) – Natural Resources Conservation Service,Washington, D.C.
Usher, M.B., et Jefferson, J.B. 1990. The concepts of colonisationand succession: their role in nature reserve management. DansCalcareous grasslands — Ecology and management. Procee-dings of a BES/NCC symposium at University of Sheffield, 14–16 septembre 1987. Éditeurs : S.H. Hillier, D.W.A. Walton etD.A. Wells. Bluntisham books, Bluntisham. pp. 149–153.
Van der Putten, W.H., Mortimer, S.R., Hedlund, K., Van Dijk, C.,Browrn, V.K., Leps, J., Rodriguez-Barrueco, C., Roy, J.,Diaz Len, T.A., Gormsen, D., Korthals, G.W., Lavorel, S.,Santa Regina, I., et Smilauer, P. 2000. Plant species diversity asa driver of early succession in abandoned fields: A multi-siteapproach. Oecologia, 124 : 91–99.
Ward, J.H. 1963. Hierarchical grouping to optimize an objectivefunction. J. Am. Stat. 58 : 238–244.
Whittaker, R.H. 1972. Evolution and measurement of species di-versity. Taxon, 21 : 213–251.
Willems, J.H. 1995. Soil seed bank, seedling recruitment and ac-tual species composition in an old and isolated chalk grasslandsite. Folia Geobot. Phytotaxon. 30 : 141–156.
Willems, J.H., et Bik, L.P.M. 1998. Restoration of high speciesdensity in calcareous grassland: the role of seed rain and soilseed bank. Appl. Veg. Sci. 1 : 91–100.
Willson, M.F. 1992. The ecology of seed dispersal. Dans Seeds:The ecology of regeneration in plant communities. Éditeur :M. Fenner. CAB international, Southampton, R.U. pp. 61–85.
Wilson, S.D. 2000. Heterogeneity, diversity, and scale in plantcommunties. Dans Ecological consequences of habitat heteroge-neity. Éditeurs : M.J. Hutchings, E.A. John et A.J. Stewart.Blackwell Science, Oxford. pp. 53–69.
Woldu, Z., et Saleem, M.A.M. 2000. Grazing induced biodiversityin the highland ecozone of East Africa. Agric. Ecosyst. Environ.79 : 43–52.
J:\cjb\cjb8008\B02-068.vpTuesday, July 30, 2002 11:07:30 AM
Color profile: Generic CMYK printer profileComposite Default screen
![[LE CBS A BRUXELLES] · 2018. 6. 21. · Végétation sur dalle (ép. substrat 5 - 10 cm) 0.3 Végétation sur dalle (ép. substrat 10 - 20 cm) 0.4 Végétation sur dalle (ép. substrat](https://img.pdfslide.fr/doc/110x75/60da5a5c73a7c315177f61f5/le-cbs-a-bruxelles-2018-6-21-vgtation-sur-dalle-p-substrat-5-10.jpg)




























