Embed Size (px)
Citation preview

DOSSIER DEÀ LA QUALIFICATION
AUX FONCTIONS DE MAÎTRE DE CONFÉRENCES
« Sciences et Techniques des Activités Physiques et S
Matthieu CASTERAN
- Mme Isabelle SIEGLER - M Vincent NOUGIER
DOSSIER DE CANDIDATURE À LA QUALIFICATION
AUX FONCTIONS DE MAÎTRE DE CONFÉRENCES
SECTION 74 Sciences et Techniques
des Activités Physiques et Sportives
Par
Matthieu CASTERAN
(Campagne 2014) 1ère demande
Rapporteurs de ce dossier
Mme Isabelle SIEGLER M Vincent NOUGIER
CANDIDATURE
AUX FONCTIONS DE MAÎTRE
portives »

Table des matières
Dossier de candidature……………………………………………… page 1
Curriculum Vitae …………………………………………………… page 5
Déclaration de candidature internet ………………………………… page 7
Annexes
- N°1 Contrat d’ATER - N°2 Rapport de Thèse - N°3a Contrat de thèse - N°3b Avenant au contrat (Monitorat) - N°4 Attestation « Formation à la Vulgarisation Scientifique » - N°5a Article 1 (Neuroscience) - N°5b Article 2 (PloS One) - N°6 Résumés d’articles publiés - N°7 Communications - N°8 Conférence - N°9 Congrès organisés - N°10a Enseignements (résumé) - N°10b Fiches de service Dijon - N°10c Attestation cours Nancy - N°10d Fiche de service Marseille - N°11a Attestation des Conseils Scientifique et Documentaire - N°11b Attestation « Engagements au sein de l’Ecole Doctorale »

1
Exposé des activités de M. CASTERAN Matthieu I - Synthèse de la carrière
Je suis actuellement Attaché Temporaire d’Enseignement et de Recherche (ATER) à l’Institut des Sciences du Mouvement (UMR-CNRS 7287) de l’université d’Aix-Marseille et ceci jusqu’au 31 août 2014. J’ai réalisé mon doctorant au sein du laboratoire INSERM U-1093 « Cognition, Action et Plasticité Sensorimotrice » de Dijon rattaché à l’université de Bourgogne. J’ai obtenu pour la réalisation de cette thèse un financement INSERM-Région de 3 ans. De plus, lors de celle-ci j’ai eu la chance de pouvoir effectué des contrats de moniteur (sur 2 ans) me permettant ainsi de me familiariser avec l’enseignement et d’en comprendre les aboutissements.
Précédemment, j’ai réalisé une licence en entrainement sportif à la faculté des sciences du sport de l’université de Lorraine. Par la suite, je me suis dirigé vers un master de l’UFR « Sciences et Techniques, Mathématiques, Informatique et Automatisme » rattaché à la faculté des sciences de l’université de Lorraine me permettant d’acquérir des compétences en informatique, en traitement de l’image et en analyse de la performance. Désirant poursuivre en recherche, j’ai alors contacté le professeur Thierry Pozzo du laboratoire de Dijon qui m’a permis de réaliser un stage de 6 mois au cours de ma seconde année de master, me donnant ainsi une porte d’entrée sur le monde de la recherche. Suite à cela, j’ai obtenu la possibilité de réaliser ma thèse au sein de ce même laboratoire. II - Activités scientifiques et administration de l a recherche 1) Mes travaux de recherche ont eu pour but d’étudier le vieillissement selon une approche différente de celle rencontrée, faite à partir de données physiologiques ou cognitif. Nous avons initié une réflexion du vieillissement en analysant le contrôle « modulaire » des paramètres temporels et spatiaux de la réalisation de mouvements. Ceci par l’intermédiaire de deux paradigmes mettant en jeu une tâche focale et/ou une tâche de contrôle postural. - Le premier utilisant des déplacements du Centre de Masse (CoM) importants. Nous avons utilisés les mouvements de pointage de tout le corps chez des sujets jeunes et âgés sains permettant d’étudier le couple mouvement-équilibre.
- Le second avait pour but d’étudier des déplacements plus faibles du CoM. Nous avons analysé les déplacements de patient âgés sains et dépressifs afin de comprendre les mécanismes cognitifs (traitement de l’information) du contrôle postural lors d’une double tâche cognitive.
a) Mouvements de pointage de tout le corps Ces mouvements font intervenir une composante focale (l’atteinte de la cible) et une composante posturale (permettant l’atteinte de la cible par le doigt, mais aussi la conservation de l’équilibre). A l’aide de ce paradigme, nous avons montré que la vitesse de réalisation de ces mouvements n’a pas d’impact sur le contrôle global du corps (d’un point de vue géométrique), mais que des modifications temporelles sont observées (arrivée du pic de vitesse par exemple).
Dans un second temps, nous avons utilisé ce même paradigme dans le but de comprendre de quelle manière la variabilité de la géométrie du corps pouvait être contrôlée. L’étude du CoM et plus particulièrement de sa vitesse sur un axe antéropostérieur (codant l’atteinte de la cible) et un axe vertical (codant l’atteinte de la cible, mais aussi la conservation de l’équilibre) nous a permis de mettre en avant un mécanisme de contrôle en parallèle. En effet, l’axe vertical présente une variabilité extrêmement faible pouvant laisser penser à un contrôle en amont, à un programme préétabli pouvant être assimilé à une intégration de la composante gravitaire. A l’opposé, l’axe antéropostérieur présente une variabilité importante, montrant un contrôle soumis à des régulations.
Enfin, nous avons étudié l’impact du vieillissement sur ces mêmes paramètres afin de comprendre comment l’avancée en âge pouvait altérer le contrôle moteur de mouvements impliquant une tâche de précision et une tâche de conservation de l’équilibre. Nous avons alors montré que le vieillissement dit « normal », ne présente pas d’altération d’un point de vue de la mise en place de ce mécanisme de réduction de dimensionnalité. Cependant, des modifications d’ordre temporel ont été mises en lumière et interprétées comme des régulations mises en place afin de conserver l’équilibre lors de la réalisation de la tâche.

2
b) Dépression et contrôle posturale La dépression est un important problème de santé publique touchant différentes classes d’âge allant des plus jeunes aux plus âgés. D’un point de vue cortical, la dépression touche le cortex préfrontal et le cortex cingulaire antérieur. Le niveau sous-cortical est lui aussi atteint par des modifications constatées de l’hippocampe et des ganglions de la base. Cette dernière structure permet le contrôle des mouvements « non volontaires » tels que la posture par une intégration multi-sensorielle et plus précisément une intégration motrice-proprioceptive. De plus, la quantité de traitement de l’information étant limité et propre à chacun, nous nous sommes demandé si, lors d’une tâche cognitive en parallèle d’une tâche de contrôle postural, un sujet dépressif répondrait de manière similaire à un sujet sain.
Les études posturographiques nous ont permis de montrer que les sujets dépressifs présentaient une aire et une longueur de déplacement du Centre des Pressions (CoP) plus important que les sujets sains déjà en simple tâche de contrôle postural. De plus, les sujets dépressifs ne présentaient pas d’augmentation de leur surface lors de la double tâche cognitive. Ceci nous a permis de conclure que les patients dépressifs seraient déjà dans un processus de double tâche dus à la dépression et qu’elle agirait sur l'allocation des ressources du traitement de l’information. Ces résultats permettent de mettre en avant le fait qu’il est important, lors de la prise en charge de patients dépressifs, de prendre en compte l’aspect cognitif, mais aussi l’aspect physique de la personne qui peut devenir un risque supplémentaire. En effet, la dépression a déjà été mise en relation avec un risque de chute augmenté dans plusieurs études. Enfin, ceci montre que le contrôle postural, avec l’avancée en âge, n’est pas si « automatique » qu’il peut l’être présenté.
Ces deux paradigmes nous ont permis d’étudier les relations entre les paramètres spatiaux
(géométrie globale du corps et cinématique du mouvement) et temporels (temps de mouvement, arrivé des pics de vitesse et points de croisement), mais aussi de comprendre l’impact du traitement de l’information sur le contrôle postural. Nous pourrions alors donner une définition du vieillissement d’un point de vue du contrôle et de l’exécution du mouvement, et non plus seulement basé sur des paramètres physiologiques et/ou cognitif.
2) La dernière thématique abordée en parallèle de ma thèse concerne l’impact d’une maladie neurodégénérative liée au vieillissement lors de tâches de pointage de cible (la maladie d’Alzheimer). Lors de cette expérimentation nous avons pu tester des patients présentant une démence de type Alzheimer et des patients âgés sains lors de pointage de cibles en mouvement avec le membre supérieur. Nous avons voulu étudier les capacités d’imitation, mais aussi comprendre les relations entre la perception du mouvement et sa reproduction chez ces sujets. Ces deux groupes de sujets ont montré la capacité à reconnaitre des vitesses différentes de mouvements, mais aussi la faculté à la reproduire. Ceci montre une capacité interne motrice à faire le parallèle entre l’observation et la production du mouvement. Cependant, l’incapacité des patients Alzheimer lors de certain essaie à contrôler le départ prématuré de leur bras montre une insuffisance de ces patients lors du stade d’inhibition des commandes.
Ce dernier aspect de mes recherches nous a permis de mettre en avant une base sur laquelle les cliniciens peuvent s’appuyer lors d’interventions physiques et cognitives avec des patients âgés présentant une démence de type Alzheimer (phénomène d’imitation et difficultés d’inhibition).
Pour conclure, l’ensemble de ces recherches ont eu pour but d’étudier différents
caractéristiques auxquelles l’Homme est confronté (vitesse, vieillissement, dépression, traitement de l’information, démence par exemple) par l’analyse de l’exécution du mouvement et/ou du contrôle postural. C’est différentes thématiques font partie intégrante des connaissances dispensées et attribuées au champ des STAPS. L’étude du mouvement et du corps est un axe central de la formation donnée aux étudiants des différentes filières en faculté des sciences du sport. La connaissance du mouvement est essentielle pour la performance et l’entrainement, pour l’activité physique adaptée et la création d’instruments adaptés, pour l’enseignement, et enfin pour de nombreux domaines de recherche liés à l’Homme.

3
Publications - Casteran M , Pftizenmeyer F, Thomas E, Manckoundia P (2013) Postural Control in
Depressive Elderly Subjects. Journal of the American Geriatrics Society, en soumission. - Casteran M , Manckoundia P, Pozzo T, Thomas E (2013) Alterations with Movement Duration
in the Kinematics of a Whole Body Pointing Movement. PLoS ONE 8(1): e52477. doi:10.1371/journal.pone.0052477.
- Bisio A, Casteran M , Ballay Y, Manckoundia P, Mourey F, Pozzo T. (2012). Motor resonance mechanisms are preserved in Alzheimer’s disease patients. Neuroscience.
Direction de travaux - 3 Etudiants de L3 APAs (stage professionnel dans le domaine de l’activité physique adaptée). - Master 1 (Recherche - Postural Control in Depressive Elderly Subjects, étudiant en second auteur). Responsabilité d’animation de recherche Gestion administrative : PHRC – Impact de la dépression sur le contrôle postural de la personne âgée. Approuvé et soutenue par le CHU de Dijon, approuvé par le CPP Est 1 et enregistré à l’Agence Française de Sécurité Sanitaire des Produits de Santé (N° 2007-A01054-49). Organisation de colloques et conférences Comité d’organisation - 17ème Forum des Jeunes Chercheurs - Dijon 2011. - 14ème Journées Nationales des BIOTechno - Dijon 2011 (démarchage de partenaires industriels). - 1er congrès Européen de Stimulation Cognitive - Dijon 2012 (Stimco). Président du comité d’organisation - 19ème Forum des Jeunes Chercheurs - Dijon 2013. - 16ème Journées Nationales de BIOTechno - Dijon 2013 (Organisation de tables rondes, de conférences et démarchage de partenaires industriels). - 1er et 2nd concours photographique « Art et Sciences » - Dijon 2011 et 2013. - Nombreuses conférences dans le cadre de mon implication associative « EDIFICE » Participation en tant que conférencier invité Conférence décrivant et analysant les différents outils utilisés en neuroscience pour l’expérimentation et plus particulièrement les techniques d’imagerie. Université pour Tous de Bourgogne (2013). Collaboration scientifique L’étude des mécanismes d’imitation de la vitesse chez des patients présentant une dégénérescence de type Alzheimer est une collaboration avec le Dr. Ambra Bisio de l’Institut Italien des Technologies. Expertise scientifique - En tant que membre du conseil scientifique de l’université de Bourgogne, j’ai été nommé expert pour le PRES Bourgogne - Franche-Comté lors de l’appel à projet du Bonus Qualité Recherche 2013. - Enfin, en tant qu’élu de l’école doctorale, j’ai fait parti des 3 dernières campagnes de jury pour les bourses doctorales (INSERM, INRA, MRT, Région et CIFRE). III - Activités pédagogiques - Mon activité d’enseignement s’est fait durant 2 ans sous forme de contrat de monitorat avec 141,5h d’enseignement en UFR STAPS. Ceci sur les sites de Dijon, Le Creusot et Nancy sous des formats de TP, TD et CM. Enfin, j’ai encadré des étudiants de Licence et Master en stage. - En parallèle, j’ai participé durant deux ans au programme de vulgarisation scientifique de l’université de Bourgogne « l’Expérimentarium » permettant aux jeunes chercheurs de présenter leur recherche au grand public (primaires, collèges, lycées, et tout public lors de journées spéciales). - Pour finir, je suis actuellement ATER au sein de l’UFR STAPS de l’université d’Aix-Marseille.
4
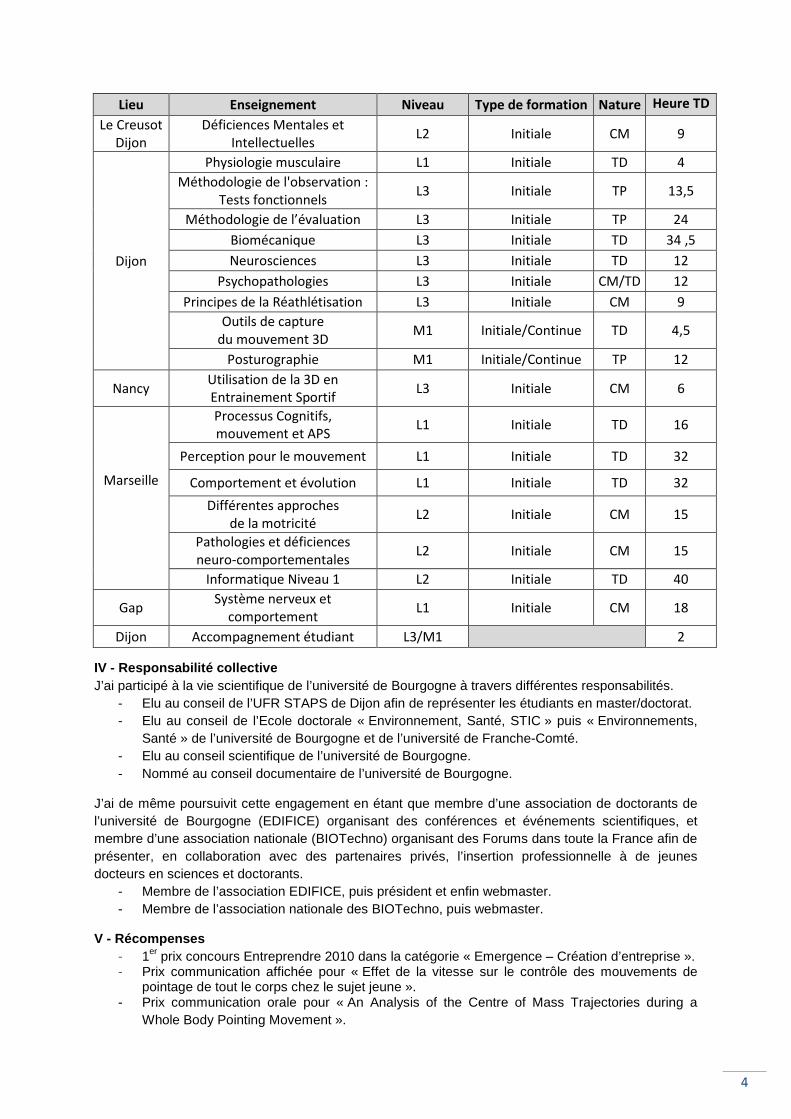
Lieu Enseignement Niveau Type de formation Nature Heure TD
Le Creusot
Dijon
Déficiences Mentales et
Intellectuelles L2 Initiale CM 9
Dijon
Physiologie musculaire L1 Initiale TD 4
Méthodologie de l'observation :
Tests fonctionnels L3 Initiale TP 13,5
Méthodologie de l’évaluation L3 Initiale TP 24
Biomécanique L3 Initiale TD 34 ,5
Neurosciences L3 Initiale TD 12
Psychopathologies L3 Initiale CM/TD 12
Principes de la Réathlétisation L3 Initiale CM 9
Outils de capture
du mouvement 3D M1 Initiale/Continue TD 4,5
Posturographie M1 Initiale/Continue TP 12
Nancy Utilisation de la 3D en
Entrainement Sportif L3 Initiale CM 6
Marseille
Processus Cognitifs,
mouvement et APS L1 Initiale TD 16
Perception pour le mouvement L1 Initiale TD 32
Comportement et évolution L1 Initiale TD 32
Différentes approches
de la motricité L2 Initiale CM 15
Pathologies et déficiences
neuro-comportementales L2 Initiale CM 15
Informatique Niveau 1 L2 Initiale TD 40
Gap Système nerveux et
comportement L1 Initiale CM 18
Dijon Accompagnement étudiant L3/M1
2
IV - Responsabilité collective J’ai participé à la vie scientifique de l’université de Bourgogne à travers différentes responsabilités.
- Elu au conseil de l’UFR STAPS de Dijon afin de représenter les étudiants en master/doctorat. - Elu au conseil de l’Ecole doctorale « Environnement, Santé, STIC » puis « Environnements,
Santé » de l’université de Bourgogne et de l’université de Franche-Comté. - Elu au conseil scientifique de l’université de Bourgogne. - Nommé au conseil documentaire de l’université de Bourgogne.
J’ai de même poursuivit cette engagement en étant que membre d’une association de doctorants de l’université de Bourgogne (EDIFICE) organisant des conférences et événements scientifiques, et membre d’une association nationale (BIOTechno) organisant des Forums dans toute la France afin de présenter, en collaboration avec des partenaires privés, l’insertion professionnelle à de jeunes docteurs en sciences et doctorants.
- Membre de l’association EDIFICE, puis président et enfin webmaster. - Membre de l’association nationale des BIOTechno, puis webmaster.
V - Récompenses - 1er prix concours Entreprendre 2010 dans la catégorie « Emergence – Création d’entreprise ». - Prix communication affichée pour « Effet de la vitesse sur le contrôle des mouvements de
pointage de tout le corps chez le sujet jeune ». - Prix communication orale pour « An Analysis of the Centre of Mass Trajectories during a
Whole Body Pointing Movement ».

5
Curriculum Vitae
CASTERAN Matthieu Né le 7 Avril 1986 à Epinal (Vosges) – 27 ans Avenue Jules Ferry Tél : 06.13.72.85.68 Résidence le Solémar Bat. B E-mail : [email protected] 13260 CASSIS
Situation actuelle ATER à l'UFR STAPS de l'Université d’Aix-Marseille, rattaché à « l’Institut des Sciences du Mouvement – Etienne-Jules Marey » membre de l'équipe « Comportement Perceptivo-Moteur » (Contrat en annexe 1) Diplômes universitaires 2005-2008 : Licence « Entrainement Sportif »; UFR STAPS - Université de Lorraine, Nancy 2008-2009 : Maîtrise « Information Numérique et Entreprise, spécialité Modélisation 3D, Ingénierie Sportive et Technologies »; UFR Sciences et Techniques, Mathématiques, Informatique et Automatismes (STMIA) – Université de Lorraine, Nancy Mention « Assez bien » 2009-2010 : Master « Ingénierie de la Mesure et de l’Image, spécialité Mesure, Performance et Certification », UFR STMIA de l'université de Lorraine, Nancy Titre du mémoire: « Intégration de la vitesse dans la planification motrice et le contrôle des mouvements orientés » - Direction du mémoire : Dr. Elizabeth Thomas
Mention « Assez-Bien » (2nd de la promotion) 2010-2013 : Doctorat de l'Université de Bourgogne ; STAPS - Neurosciences
« Vieillissement, Contrôle Modulaire de l’équilibre et des Mouvements Orientés » - Directeur: Pr. Patrick Manckoundia ; Co-encadrant: Dr. Elizabeth Thomas - Jury : Présidente du Jury : Dr. Agnès Roby-Brami Rapporteur : Pr. Vincent Nougier Rapporteur : Pr. Guy Chéron Invité : Pr. Thierry Pozzo Mention « Très Honorable » (Rapport en annexe 2) Allocation INSERM-Région / Contrat de moniteur sur 2 ans (Contrats en annexe 3) Autres diplômes ou compétences - Brevet d’Aptitude aux Fonctions d’Animateur – Surveillant de Baignade (BAFA-SB). - Brevet National de Sécurité et de Sauvetage Aquatique (BNSSA). - Premier Secours en Equipe Niveau 1 (anciennement AFCPSAM). - Entraineur Handball « Contrex Handball club » (-14 et -16ans durant deux ans). - 1er prix - concours Entreprendre 2010 Promotech CEI, catégorie "EMERGENCE" création d’entreprise. - Formation de vulgarisation Scientifique « Expérimentarium » de l’université de Bourgogne (Cf. annexe 4). Activité de recherche (mots clefs) Mots clefs : Contrôle moteur ; Relations mouvement-équilibre ; Relations temporelles-spatiales ; Mouvements de tout le corps ; Vieillissement ; Dépression ; Démence de type Alzheimer. Articles de Recherche Originaux (Annexe 5)
1) Casteran Matthieu , Pfitzenmeyer François, Thomas Elizabeth & Manckoundia Patrick (2013) “Postural control in depressive elderly subjects” En cours de soumission dans le JAGS.
2) Casteran Matthieu , Manckoundia Patrick, Pozzo Thierry & Thomas Elizabeth (2013). “Alterations with Movement Duration in the Kinematics of a Whole Body Pointing Movement” PLoS ONE 8(1): e52477. Doi: 10.1371/journal.pone.0052477.
3) Bisio Ambra, Casteran Matthieu , Ballay Yves, Manckoundia Patrick, Mourey France & Pozzo Thierry. (2012) “Motor resonance mechanisms are preserved in Alzheimer's disease patients” Neuroscience 222C: 58-68.Doi: 10.1016/j.neuroscience.2012.07.017.
Résumés de Congrès Publiés (Annexe 6) 1) Casteran Matthieu , Thomas Elizabeth, & Manckoundia Patrick (2012). Impact d’une tâche cognitive
sur la posture du sujet âgé dépressif comparé au sujet âgé non dépressif. Neurophysiologie Clinique/Clinical Neurophysiology, 42(6), 401-402. Doi: 10.1016/j.neucli.2012.09.049.
2) Casteran Matthieu , Pozzo Thierry, & Thomas Elizabeth (2012) "Contrôle du centre de masse lors de pointage de tout le corps chez le sujet jeune et âgé sain." Neurophysiologie Clinique/Clinical Neurophysiology 42.6: 401. Doi: 10.1016/j.neucli.2012.09.048.

6
Nombres de publications, ouvrages, travaux, brevets , etc.
1. Journal of the American Geriatrics Society, 2013, 1er/4 auteurs (En cours de soumission) 2. Plos One, 2013, 1er/4 auteurs 3. Neuroscience, 2012, 2ème/6 auteurs
Nombre de Conférences et congrès (Informations détaillées fournies en annexe 7) 1. Conférence invitée : 1 (Attestation annexe 8) 2. Communications affichées : 5
a. résumés publiés dans une revue indexée : 1/5 (Clinical Neurophysiology) b. résumés publiés dans des actes : 2/5
4. Communications orales : 4 a. résumés publiés dans une revue indexée : 1/4 (Clinical Neurophysiology) b. résumés publiés dans des actes : 2/4
5. Conférence organisé dans le cadre de l’association de doctorants : 5 Encadrement d'étudiants (nombres et % d’encadrement ) Étudiant de Master : François Pfizenmeyer (Kinésithérapeute) étudiant de Master 1 (50%), l’étude réalisée a fait l’objet d’une présentation orale et d’un article en collaboration en soumission. Evènement Scientifique Participation à l'organisation de congrès :
- 5 au total, 3 comme organisateurs, 2 comme président du comité d’organisation. (Compléments en annexe 9)
- Organisation du 1er et 2nd concours photographique « Art et Sciences ». Activité d'expertise :
- Expert pour l’AAP BQR du PRES Bourgogne-Franche Comté 2013 (Conseil Scientifique) - Jury concours des bourses de thèse de l’école doctorale Environnements-Santé
Participation à la vie de l'équipe de recherche :
- Représentant au conseil de laboratoire - Participation à l’organisation d’un congrès sur le site de l’UFR STAPS (StimCo)
Activité d'enseignement (volumes horaires, année, d iscipline) (Détail en annexe 10) Cours magistraux : 75h TD, L1-L2-L3, Etude du mouvement /Neuroscience / Déficiences / Syst. Nerveux Travaux dirigés : 162h TD, L1-L3, Bioméca./Neuroscience/Psychopathologies/Physiologie/Informatique Travaux pratiques : 49,5h TD, L3, Méthodologie Obs.-Eval./Posturographie/Capture 3D du Mouvement. Activité administrative Responsabilités électives :
- Représentant au conseil de laboratoire - Elu au conseil de l’UFR STAPS - Elu au conseil Scientifique de l’Université de Bourgogne - Nommé au conseil documentaire de l’Université de Bourgogne - Elu au conseil de l’école doctorale « Environnements-Santé » Bourgogne-Franche Comté
(Cf. annexe 11) Responsabilité associatives :
- Président, puis Webmaster de l’association de doctorants EDIFICE www.edifice-dijon.com
- Webmaster de l’association nationale des BIOTechno www.biotechno.eu
Pour plus d’informations : www.matthieu.casteran.com

1
Il est conseillé de joindre ce document au dossier transmis
aux rapporteurs désignés par le Conseil National des Universités
DÉCLARATION DE CANDIDATURE À LA QUALIFICATION
AUX FONCTIONS DE MAÎTRE DE CONFÉRENCES,
POUR LA SECTION 74-Sciences et techniques des activités physiques et sportives
(Campagne 2014)
1ère demande
Je soussigné(e) M.
Nom de famille : CASTERAN
Nom d'usage : CASTERAN
Prénom : MATTHIEU
Date et lieu de naissance : 07/04/1986 - EPINAL
Nationalité : Française
.
Date de création de la candidature
17/10/2013 à 16:10
Date de dernière modification de la candidature
17/10/2013 à 16:10
Titres universitaires français :
Doctorat
Diplôme au titre duquel la qualification est demandée : Doctorat
Titre : Vieillissement, contrôle modulaire de l'équilibre et des mouvements orientés
Date de soutenance : 25/11/2013
Lieu de la soutenance : UNIVERSITE DE BOURGOGNE
Mention :
Directeur : PR. PATRICK MANCKOUNDIA
Composition du jury : DR. ELIZABETH THOMAS
PR. GUY CHERON
DR. AGNES ROBY-BRAMI
PR. VINCENT NOUGIER
Adresse postale et électronique à laquelle seront acheminées toutes les correspondances
RESIDENCE LE SOLEMAR BATIMENT B
Code postal : 13260 Ville :CASSIS Pays : FRANCE Téléphone : 0613728568 Télécopie : Adresse électronique : [email protected]

2
Liste des étabs et labos d'exercice :
INSERM U-1093 Cognition, Action et Plasticité Sensorimotrice - Dijon
UMR CNRS 7287 Institut des Sciences du Mouvements - Étienne Jules Marey - Marseille
Activités en matière d'enseignement :
Dijon L2
CM Déficiences mentales et intellectuelles
L3
TD Neurosciences/Biomécanique
TP Méthodologie de l'Observation (Tests Fonctionnels) /Tests Cognitifs/Plateforme de force
CM Réathlétisation/Psychopathologies
Suivie de Stage
M1
TD+TP Cinématique 3D/Plateforme de force
Référent de Stage
Nancy L3
CM Utilisation de la 3D en Entrainement sportif
Marseille L1/L2
ATER en cours
Thème de recherche et mots clés :
1 - Contrôle moteur
Vieillissement
Redondance et Variabilité
Mouvement de pointages
Cinématique/EMG
2 - Impact de la dépression sur le sujet âgé
Vieillissement normal et pathologique
Contrôle postural
Double tâche
3 - Mécanisme de résonance chez le patient atteint de la maladie d'Alzheimer
Contrôle moteur
Imitation
Vieillissement normal et pathologique
Action-Perception
Activités en matière d'administration et autres responsabilités collectives :
Élu aux Conseils :
-Scientifique de l'Université de Bourgogne
-De l'Ecole Doctorale «Environnements-Santé»
-De l'UFR STAPS Master/Doctorat Dijon

3
Président / Webmaster de l'association de Doctorants «Edifice»
Webmaster du Réseau National des BIOTechno
Président des comités d'organisation :
-du Forum des Jeunes Chercheurs 2013
-des Journées Nationales des BIOTechnologies 2013
déclare faire acte de candidature à la qualification.
Fait à le
Signature
Dijon 17 Octobre 2013

Annexes

Annexe 1
Contrat ATER



Annexe 2
PV-Avis-Rapport
de soutenance




Annexe 3
Contrat de thèse
Avenant (monitorat)







Annexe 4
Vulgarisation Scientifique

Elise Cellier-Holzem Responsable de l'Experimentarium Université de Bourgogne [email protected] Tel : 03 80 39 35 91 Bureau R21 6, Bd Gabriel 21000 Dijon Objet : Attestation de participation au programme de culture scientifique
L’Experimentarium est un programme de culture scientifique de l’Université de Bourgogne. Il forme de jeunes chercheurs à la vulgarisation derecherche. Autour d’expériences ou d’objets insolites, les chercheurs racontent leur quotidien, invitent au questionnement et entraînent les visiteurs (petits et grands) au cœur de leur recherche. J'atteste, Elise CellierCasteran a participé activement au programme Experimentariumdoctorat.
En effet, depuis septembre 2011manifesté par la construction et la production d’unrecherche. Au totalses travaux devant un public d’information le temps de préparation des ateliers de l’des rencontres avec le public, pour chaque doctorant est estimé Matthieu a effectué Pour plus d’informations sur l’Experimentarium Fait à Dijon, le 5 décembre
Dijon, le
ttestation de participation au programme de culture scientifique « Experimentarium
L’Experimentarium est un programme de culture scientifique de l’Université de Bourgogne. Il forme de jeunes chercheurs à la vulgarisation derecherche. Autour d’expériences ou d’objets insolites, les chercheurs racontent leur quotidien, invitent au questionnement et entraînent les visiteurs (petits et grands) au cœur de leur recherche.
Elise Cellier-Holzem, responsable de l'Experimentarium, que a participé activement au programme Experimentarium
septembre 2011 et jusqu’à avril 2013, son investissement s’est a construction et la production d’un atelier de prés
total, Matthieu a effectué 46 heures durant lesquelles ses travaux devant un public allant des élèves de CM1 au grand public.d’information le temps de préparation des ateliers de l’Experimentarium, en amont des rencontres avec le public, pour chaque doctorant est estimé
a effectué 55 heures dans le cadre de ce programme.
Pour plus d’informations sur l’Experimentarium : http://experimentarium.u
5 décembre 2013
Elise Cellier-Holzem
Dijon, le 5 décembre 2013
Experimentarium »
L’Experimentarium est un programme de culture scientifique de l’Université de Bourgogne. Il forme de jeunes chercheurs à la vulgarisation de leur sujet de recherche. Autour d’expériences ou d’objets insolites, les chercheurs racontent leur quotidien, invitent au questionnement et entraînent les visiteurs (petits et grands) au
e de l'Experimentarium, que Matthieu a participé activement au programme Experimentarium au cours de son
son investissement s’est atelier de présentation de sa
durant lesquelles il a pu présenter au grand public. A titre
rimentarium, en amont des rencontres avec le public, pour chaque doctorant est estimé neuf heures. Ainsi,
http://experimentarium.u-bourgogne.fr/
Holzem

Annexe 5
Article Neuroscience
Article PloS One

Neuroscience 222 (2012) 58–68
MOTOR RESONANCE MECHANISMS ARE PRESERVEDIN ALZHEIMER’S DISEASE PATIENTS
A. BISIO, a* M. CASTERAN, b Y. BALLAY, b
P. MANCKOUNDIA, b,c F. MOUREY b AND T. POZZO a,b,d
aDepartment of Robotics, Brain and Cognitive Sciences, Istituto
Italiano di Tecnologia, via Morego 30, 16163 Genoa, Italy
b INSERM U1093 Cognition, Action et Plasticite Sensorimotrice,
Universite de Bourgogne, France
cService de Medecine Interne Geriatrique, Hopital de Champmaillot,
Centre Hospitalier Universitaire, rue Jules Violle, BP 87909, 21079
Dijon, Franced Institut Universitaire de France, Universite de Bourgogne, UFR
STAPS, Dijon, France
Abstract—This study aimed to better characterize the
sensorimotor mechanisms underlying motor resonance,
namely the relationship between motion perception and
movement production in patients suffering from Alzheimer’s
disease (AD). This work first gives a kinematic description of
AD patients’ upper limb movements, then it presents a sim-
ple paradigm in which a dot with different velocities is
moved in front of the participant who is instructed to point
to its final position when it stopped. AD patients’ actions,
as well as healthy elderly participants, were similarly influ-
enced by the dot velocity, suggesting that motor resonance
mechanisms are not prevented by pathology. In contrast,
only patients had anticipatory motor response: i.e. they
started moving before the end of the stimulus motion, unlike
what was requested by the experimenter. While the
automatic imitation of the stimulus suggests an intact ability
to match the internal motor representations with that of the
visual model, the uncontrolled motion initiation would
indicate AD patients’ deficiency to voluntarily inhibit
response production. These findings might open new
clinical perspectives suggesting innovative techniques in
training programs for people with dementia. In particular,
the preservation of the motor resonance mechanisms, not
dependent on conscious awareness, constitutes an intact
basis upon which clinicians could model both physical
and cognitive interventions for healthy elderly and AD
patients. Furthermore, the evaluation of the inhibitory
functions, less sensitive to the level of education than other
methods, might be useful for screening test combined with
the traditional AD techniques. However, further investiga-
0306-4522/12 $36.00 � 2012 IBRO. Published by Elsevier Ltd. All rights reservehttp://dx.doi.org/10.1016/j.neuroscience.2012.07.017
*Corresponding author. Tel: +39-010-71781406, +39-340-2435784;fax: +39-010-7170817.
E-mail addresses: [email protected], [email protected] (A.Bisio).Abbreviations: AD, Alzheimer’s disease; CE, control experiment; CG,control group; MMSE, mini-mental state examination; MNS, mirrorneuron system; MO, movement observation experiment; PM, pointingmovement experiment; rt, reaction time; SD, standard deviation; SE,standard error; TOM, theory of mind; vD, dot mean velocity; vp,participants’ mean velocity.
58
tions to understand if this feature is specific to AD or is pres-
ent also in other neurodegenerative diseases are needed.
� 2012 IBRO. Published by Elsevier Ltd. All rights reserved.
Key words: movement observation–execution, automatic imi-
tation, action–perception matching, dementia, ageing.
INTRODUCTION
Alzheimer’s disease (AD) is the most common form of
dementia that causes a decline of intellectual functioning
that interferes with daily life activities (Borson and
Raskind, 1997). Indeed, cognitive deterioration is the
first sign of the illness and the most documented aspect
of AD. Both imaging (Scahill et al., 2002) and
neuropathology (Double et al., 1996) studies have
described the brain in AD as characterized by
progressive cerebral atrophy, which increases as the
disease progresses. Despite large inter-individual
variability, the earlier change in AD patients are found in
the medial temporal structures, and the earlier clinical
sign is memory loss (Braak and Braak, 1991). At this
stage, a significant range of atrophy is present also in
the neocortical areas, but it does not differ from healthy
aged-matched individuals (Double et al., 1996). In mild
and moderate stages, a significant loss of volume is
observed not only in frontal and temporal areas but also
in parietal lobes (Scahill et al., 2002). Recent findings in
healthy people showed that parietal, temporal and frontal
lobes play a fundamental role in linking action to
perception (Grezes and Decety, 2001; Jeannerod, 2001;
Iacoboni, 2009b). Indeed, action–perception coupling is
crucial to allow humans to efficiently communicate with
other people and to interact with the environment, a
function that is markedly deficient in AD. The mechanism
associated with the link between perception and action is
known as motor resonance and is considered to stem
from the activity of the mirror neuron system (Rizzolatti
et al., 1999). As such, it was proposed to play a crucial
role in empathy and imitation (Iacoboni, 2009a), action
understanding (Rizzolatti et al., 2001), intention reading
(Iacoboni, 2005) and more generally all phenomena
underlying social interactions. Therefore, it appears
evident that the preservation of this mechanism is
fundamental for communicative purposes. Imitation, and
its inhibition during daily life activities (Bien et al., 2009),
is a special case of perception–action matching
(Wohlschlager et al., 2003) that supposes an intact
ability to perceive the external motion and to map it into
d.

A. Bisio et al. / Neuroscience 222 (2012) 58–68 59
internal motor representation either voluntarily or
automatically. Imitation mechanisms are associated with
learning (Meltzoff and Moore, 1977), empathizing
(Iacoboni, 2009a), and also considered at the basis of
social cognition (Meltzoff and Prinz, 2002).
Several neuroimaging studies have described the
neural activations during voluntary and automatic
imitation tasks, and found that both frontal and parietal
regions (i.e. fronto-parietal network of the human
mirroring system) were active when imitating an
observed motion (Iacoboni et al., 1999; Koski et al.,
2002; Iacoboni and Dapretto, 2006; Heyes, 2011).
Despite the fact that lesions to the frontal and parietal
lobes are well documented in AD, to the best of our
knowledge no study has characterized either the
automatic or the voluntary imitation capabilities of AD
patients. Likewise, little research has illustrated the
behavioural consequences of these cerebral damages at
perceptual and motor levels. Within this small literature
there are reports of alterations of motor abilities (Kluger
et al., 1997; Ghilardi et al., 1999; Manckoundia et al.,
2006), deterioration of objects’ motion and shape
perception (Gilmore et al., 1994; Rizzo and Nawrot,
1998), and impairments in transforming the visual input
into a motor output (Tippett and Sergio, 2006; Tippett
et al., 2007; Yan et al., 2008) starting from the mild
stage of the illness. Furthermore, while some have found
evidence for the involvement of both frontal and parietal
regions in developing and maintaining a social cognition
(Adolphs, 1999), a small number of works on this topic in
AD have actually found contrasting evidences. Indeed,
some studies (Cuerva et al., 2001; Verdon et al., 2007)
described AD patients’ impairment in Theory of Mind
tasks – TOM (for reviews see Meltzoff, 1999; Frith and
Frith, 2006). In contrast, Gregory et al. (2002) compared
AD patients with a population of frontal variant
frontotemporal dementia (fvFDT), and found that AD
patients’ difficulties in TOM were only for tasks requiring
heavy demands on working memory suggesting that AD
patients are not generally impaired in TOM tests, but
rather other underlying processes, like memory, are
responsible for the observed TOM deficits.
Given this, investigation of motor resonance
mechanisms in AD would cast light on patients’ ability to
relate with other people through sharing behavioural
states (e.g. imitating others’ movement). Moreover, in
patients whose brain’s lesions are difficult to
circumscribe, as in the case of AD, verifying the
preservation of perception–action coupling could be
informative about the remaining functionality of the
underling perceptual, motor and cognitive mechanisms.
In particular, by assessing whether both AD patients’
and healthy aged-matched people’s motor responses
are influenced by the observed movements, we would
gain insight about the preservation of motor resonance
mechanisms during this pathology.
If a ‘weak’ influence of the perceived movement
features in action production is synonymous with the
normal activation of motor resonance mechanisms,
exaggerated imitative response would be a sign of
abnormal functioning of the inhibitory circuitry (Bien
et al., 2009). In other words, despite the occurrence of
perception–action coupling at neural level, healthy adults
typically avoid exaggerated imitation behaviours
because these actions are not adaptive for most
everyday situations. Interestingly, previous clinical
studies performed on a group of patients with frontal
lobe damage, including AD individuals, report their
incapacity to inhibit the production of motor responses to
external stimuli. This is the case of the well known
environmental dependency syndrome that includes in its
symptoms the ‘‘imitation behaviour’’ (Lhermitte, 1986;
Lhermitte et al., 1986). Even if imitation behaviour is
marginally present in AD, although several experiments
have provided substantial evidence that AD patients
have a significant impairment in tasks requiring voluntary
inhibition (see for reviews (Amieva et al., 2004; Fournet
et al., 2007).
The present study has a threefold purpose. Firstly, it
provides a kinematic description of patients’ motor
deficiencies when performing a simple arm upwards
movement. Secondly, it characterizes the relationship
between motion observation and movement production
in AD patients by testing if and how their actions are
influenced by a previously observed motion. If motor
resonance mechanisms are still preserved, AD patients’
movements would be influenced by the observed
biological motion in so far as healthy aged-matched
participants are. Finally, this work investigates whether
AD patients’ deficiency in tasks requiring voluntary
inhibition causes inappropriate motor responses
irrespective of task demands. To these aims, the
imitation paradigm we proposed in Bisio et al. (2010)
was applied to simultaneously verify the occurrence of
normal automatic imitation phenomena and the voluntary
inhibition capabilities by providing a simple visual
stimulation and asking participants to produce a
movement in response to it.
EXPERIMENTAL PROCEDURES
Participants
The experimental group was composed of 25 elderly participants
(21 women and 4 men), ranging in age from 75 to 91 years of age
(mean age ± SD, 84.2 ± 4.5), with probable mild and moderate
AD (Perneczky et al., 2006) diagnosed according to the French
National Institute of Neurology and Communication Disorders
and Strokes – The Alzheimer’s Disease and Related Disorders
Association (NINCDS-ADRDA) and the Diagnostic and
Statistical manual-IV-Text Revised (DSM IV-TR) criteria. All the
patients lived at home or in a nursing home specializing in AD
and there was no reported difference in the severity of the
pathology based on the residence type. They underwent
comprehensive diagnostic evaluation, including clinical
assessment, brain Magnetic Resonance Imaging (MRI) and
examination of motor competencies. All of them presented with
progressive cognitive impairment. Their Mini-Mental State
Examination (MMSE) scores were between 12 and 24
(mean ± SD, 19 ± 4). Patients were excluded from the
present study if (A) their dementia was not considered due to
AD, (B) if severe vascular lesions were present, and (C) if they
were unable to perform simple arm pointing movement. The
Control Group (CG) was composed of 14 healthy participants
(10 women and 4 men), ranging in age from 74 to 89 years of

60 A. Bisio et al. / Neuroscience 222 (2012) 58–68
age (mean age ± SD, 82.4 ± 5), living at home. Their MMSE
scores were between 25 and 30 (mean ± SD, 28.6 ± 1.4).
They underwent a detailed medical and physical examination
before the study, and they were screened for cognitive deficits
using the MMSE. Participants with self-reported problems of
head injury, drug or alcohol abuse, psychiatric or neurological
disease were excluded. All participants were right-handed, and
had normal or corrected-to-normal vision. They were able to
hear adequately, to pay attention to the examiner’s behaviour
and to understand simple questions. The two groups did not
statistically differ in terms of age. In contrast, MMSE values of
the two groups were significantly different (F(1, 37) = 95.94,
p< 0.00001). Written informed consent was obtained from
each participant or their guardians, and the protocol was
approved by the Local Ethics Committee.
Materials and procedure
The experiment was performed in a darkened room. Participants
sat on a chair, in front of a large rear projection screen
(170 � 230 cm) placed 10 cm beyond the end of participants’
extended arm. A video-projector, with a refresh rate of 60 Hz
and placed behind the screen and connected to a PC, back-
projected the visual stimuli onto the display screen. The
projected visual stimulation was generated using MatLab
Psychtoolbox 3 (Brainard, 1997). An optoelectronic system
(SMART) with five cameras was used to record movements at
a sampling frequency of 120 Hz. One passive infrared reflective
marker (diameter = 20 mm) was applied onto a fingertip of the
participant’s right hand. Experiments lasted about 20 min.
Pointing movement experiment (PM). This experiment was
aimed at measuring participants’ natural pointing movements.
The kinematic data served as a baseline to be compared with
arm kinematics after motion observation (described in the
section Movement observation experiment (MO)). A green
cross appeared on the screen to indicate the starting position.
After 3 s, the cross disappeared and two vertically aligned light
blue dots (3.2 cm in diameter with a 51 cm gap between them)
were displayed for 3 s. One of the two dots replaced the green
cross and the other one was the target for the movement
(Fig. 1a). The participants’ shoulder level was roughly at the
middle of the distance between the two dots. Participants used
their right arm with an extended position to perform upwards
movements from the given starting position to the target dot
using a spontaneous natural velocity. Movement accuracy was
not emphasized. The pointing arm movement was repeated five
times. The beginning of the experiment was preceded by a
Fig. 1. Sequence of visual stimuli. (a) In pointing movement experiment (PM)
After 3 s, the cross disappeared and two vertically aligned light blue dots (3.2 c
(b) In movement observation experiment (MO) a green cross was displayed t
was replaced by a light blue dot (3.2 cm in diameter). The dot kept this pos
51 cm of space with 3 different velocities. (For interpretation of the references
of this article.)
training phase, which ended when the participant understood
the task and correctly accomplished all the experimental tasks
at least twice.
Movement observation experiment (MO). A moving stimulus
was used as a template to test the effect of motion perception
on subsequent pointing movement execution. A green cross
was displayed to indicate the movement’s starting position.
After 3 s, the green cross was replaced by a light blue dot
(3.2 cm in diameter). The dot kept this position for 1.5 s, and
then started to move vertically upwards, covering 51 cm of
space. Dot motions differed in mean velocity: slow (.39 m/s),
medium (.51 m/s), and fast (.64 m/s) (Fig. 1b). Participants did
not know if the stimulus motion was computer- or human-
generated. Stimulus velocities were randomized. Participants
were asked to point the green cross, then to watch the dot’s
movement, wait until the dot reach its final, visible position, and
finally point towards this position. Thus, the executed
movements were congruent with the observed ones in terms of
direction. Movement accuracy was not emphasized. Each dot
motion velocity was repeated four times. As in the PM, the
beginning of the experiment was preceded by a training phase,
which ended when the participant understood the task and
accomplished all the experimental tasks at least twice.
In both experiments, participants received verbal feedback
from the experimenter during the testing procedure in order to
eliminate any confusion about their aim.
Data treatment
Data processing. Data was low-pass filtered at 5 Hz using a
2nd order Butterworth filter. To define the onset and offset of
the movement, we chose a threshold corresponding to 10% of
the maximum value of the movement velocity profile.
Data analysis. Participants’ movement reaction time (rt) and
mean velocity (vp, mean value of participants’ arm velocity
module, the latter obtained as the square root of the three
spatial components of the velocity) were considered as the
main outcome variables. In PM, rt was calculated as the time
elapsed between the appearance of the two dots and arm’s
movement onset; for the MO, it was computed as the
difference in time between the end of dot motion and the onset
of participant’s pointing movement. Shapiro–Wilk normality test
was used to assess the normality of data before performing
statistical tests. The data were normally distributed and t-testsand ANOVA were conducted to do the statistic evaluations. In
a green cross appeared on the screen to indicate the starting position.
m in diameter with a 51 cm gap between them) were displayed for 3 s.
o indicate the movement’s starting position. After 3 s, the green cross
ition for 1.5 s, and then started to move vertically upwards, covering
to colour in this figure legend, the reader is referred to the web version
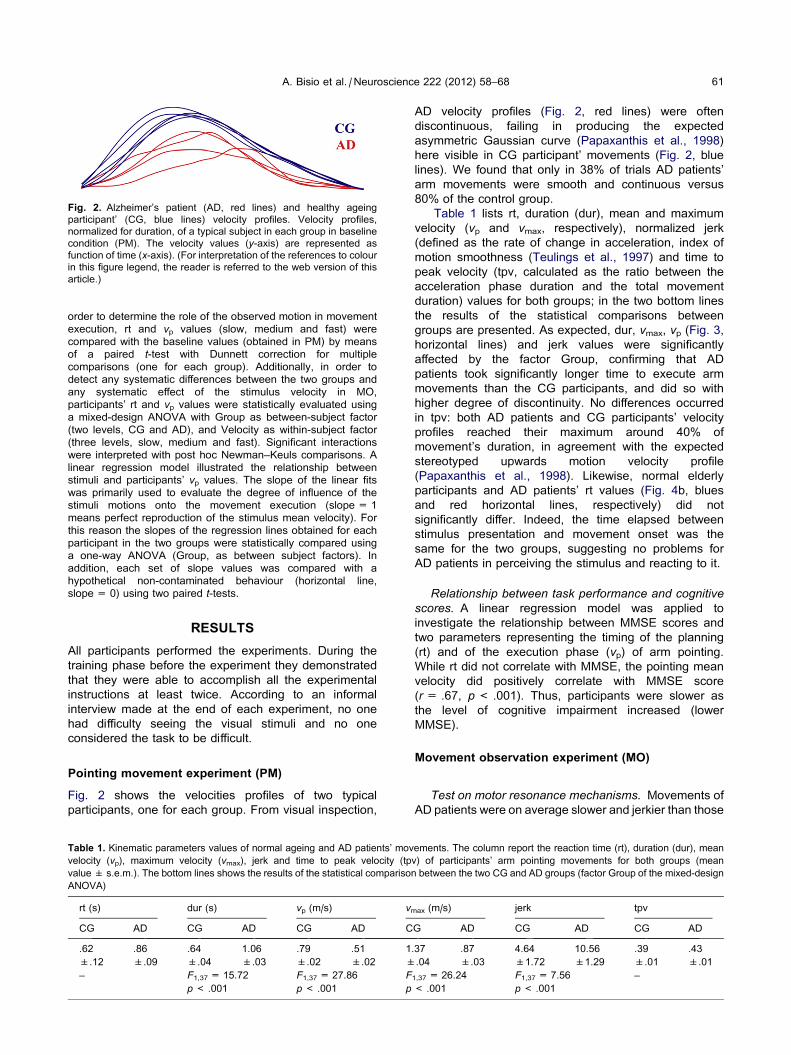
Fig. 2. Alzheimer’s patient (AD, red lines) and healthy ageing
participant’ (CG, blue lines) velocity profiles. Velocity profiles,
normalized for duration, of a typical subject in each group in baseline
condition (PM). The velocity values (y-axis) are represented as
function of time (x-axis). (For interpretation of the references to colour
in this figure legend, the reader is referred to the web version of this
article.)
A. Bisio et al. / Neuroscience 222 (2012) 58–68 61
order to determine the role of the observed motion in movement
execution, rt and vp values (slow, medium and fast) were
compared with the baseline values (obtained in PM) by means
of a paired t-test with Dunnett correction for multiple
comparisons (one for each group). Additionally, in order to
detect any systematic differences between the two groups and
any systematic effect of the stimulus velocity in MO,
participants’ rt and vp values were statistically evaluated using
a mixed-design ANOVA with Group as between-subject factor
(two levels, CG and AD), and Velocity as within-subject factor
(three levels, slow, medium and fast). Significant interactions
were interpreted with post hoc Newman–Keuls comparisons. A
linear regression model illustrated the relationship between
stimuli and participants’ vp values. The slope of the linear fits
was primarily used to evaluate the degree of influence of the
stimuli motions onto the movement execution (slope = 1
means perfect reproduction of the stimulus mean velocity). For
this reason the slopes of the regression lines obtained for each
participant in the two groups were statistically compared using
a one-way ANOVA (Group, as between subject factors). In
addition, each set of slope values was compared with a
hypothetical non-contaminated behaviour (horizontal line,
slope = 0) using two paired t-tests.
RESULTS
All participants performed the experiments. During the
training phase before the experiment they demonstrated
that they were able to accomplish all the experimental
instructions at least twice. According to an informal
interview made at the end of each experiment, no one
had difficulty seeing the visual stimuli and no one
considered the task to be difficult.
Pointing movement experiment (PM)
Fig. 2 shows the velocities profiles of two typical
participants, one for each group. From visual inspection,
Table 1. Kinematic parameters values of normal ageing and AD patients’ mov
velocity (vp), maximum velocity (vmax), jerk and time to peak velocity (tp
value ± s.e.m.). The bottom lines shows the results of the statistical comparison
ANOVA)
rt (s) dur (s) vp (m/s) vm
CG AD CG AD CG AD C
.62 .86 .64 1.06 .79 .51 1
±.12 ±.09 ±.04 ±.03 ±.02 ±.02 ±
– F1,37 = 15.72 F1,37 = 27.86 F
p< .001 p< .001 p
AD velocity profiles (Fig. 2, red lines) were often
discontinuous, failing in producing the expected
asymmetric Gaussian curve (Papaxanthis et al., 1998)
here visible in CG participant’ movements (Fig. 2, blue
lines). We found that only in 38% of trials AD patients’
arm movements were smooth and continuous versus
80% of the control group.
Table 1 lists rt, duration (dur), mean and maximum
velocity (vp and vmax, respectively), normalized jerk
(defined as the rate of change in acceleration, index of
motion smoothness (Teulings et al., 1997) and time to
peak velocity (tpv, calculated as the ratio between the
acceleration phase duration and the total movement
duration) values for both groups; in the two bottom lines
the results of the statistical comparisons between
groups are presented. As expected, dur, vmax, vp (Fig. 3,
horizontal lines) and jerk values were significantly
affected by the factor Group, confirming that AD
patients took significantly longer time to execute arm
movements than the CG participants, and did so with
higher degree of discontinuity. No differences occurred
in tpv: both AD patients and CG participants’ velocity
profiles reached their maximum around 40% of
movement’s duration, in agreement with the expected
stereotyped upwards motion velocity profile
(Papaxanthis et al., 1998). Likewise, normal elderly
participants and AD patients’ rt values (Fig. 4b, blues
and red horizontal lines, respectively) did not
significantly differ. Indeed, the time elapsed between
stimulus presentation and movement onset was the
same for the two groups, suggesting no problems for
AD patients in perceiving the stimulus and reacting to it.
Relationship between task performance and cognitive
scores. A linear regression model was applied to
investigate the relationship between MMSE scores and
two parameters representing the timing of the planning
(rt) and of the execution phase (vp) of arm pointing.
While rt did not correlate with MMSE, the pointing mean
velocity did positively correlate with MMSE score
(r= .67, p< .001). Thus, participants were slower as
the level of cognitive impairment increased (lower
MMSE).
Movement observation experiment (MO)
Test on motor resonance mechanisms. Movements of
AD patients were on average slower and jerkier than those
ements. The column report the reaction time (rt), duration (dur), mean
v) of participants’ arm pointing movements for both groups (mean
between the two CG and AD groups (factor Group of the mixed-design
ax (m/s) jerk tpv
G AD CG AD CG AD
.37 .87 4.64 10.56 .39 .43
.04 ±.03 ±1.72 ±1.29 ±.01 ±.01
1,37 = 26.24 F1,37 = 7.56 –
< .001 p< .001
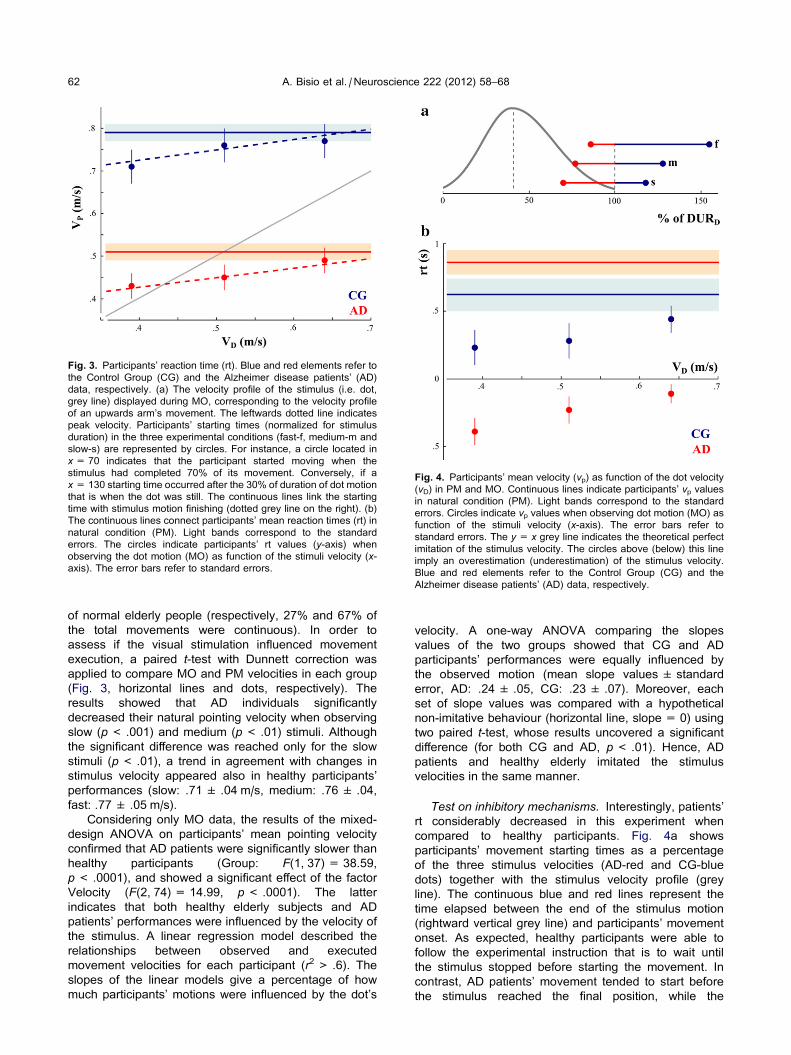
Fig. 3. Participants’ reaction time (rt). Blue and red elements refer to
the Control Group (CG) and the Alzheimer disease patients’ (AD)
data, respectively. (a) The velocity profile of the stimulus (i.e. dot,
grey line) displayed during MO, corresponding to the velocity profile
of an upwards arm’s movement. The leftwards dotted line indicates
peak velocity. Participants’ starting times (normalized for stimulus
duration) in the three experimental conditions (fast-f, medium-m and
slow-s) are represented by circles. For instance, a circle located in
x= 70 indicates that the participant started moving when the
stimulus had completed 70% of its movement. Conversely, if a
x= 130 starting time occurred after the 30% of duration of dot motion
that is when the dot was still. The continuous lines link the starting
time with stimulus motion finishing (dotted grey line on the right). (b)
The continuous lines connect participants’ mean reaction times (rt) in
natural condition (PM). Light bands correspond to the standard
errors. The circles indicate participants’ rt values (y-axis) when
observing the dot motion (MO) as function of the stimuli velocity (x-axis). The error bars refer to standard errors.
Fig. 4. Participants’ mean velocity (vp) as function of the dot velocity
(vD) in PM and MO. Continuous lines indicate participants’ vp values
in natural condition (PM). Light bands correspond to the standard
errors. Circles indicate vp values when observing dot motion (MO) as
function of the stimuli velocity (x-axis). The error bars refer to
standard errors. The y= x grey line indicates the theoretical perfect
imitation of the stimulus velocity. The circles above (below) this line
imply an overestimation (underestimation) of the stimulus velocity.
Blue and red elements refer to the Control Group (CG) and the
Alzheimer disease patients’ (AD) data, respectively.
62 A. Bisio et al. / Neuroscience 222 (2012) 58–68
of normal elderly people (respectively, 27% and 67% of
the total movements were continuous). In order to
assess if the visual stimulation influenced movement
execution, a paired t-test with Dunnett correction was
applied to compare MO and PM velocities in each group
(Fig. 3, horizontal lines and dots, respectively). The
results showed that AD individuals significantly
decreased their natural pointing velocity when observing
slow (p< .001) and medium (p< .01) stimuli. Although
the significant difference was reached only for the slow
stimuli (p< .01), a trend in agreement with changes in
stimulus velocity appeared also in healthy participants’
performances (slow: .71 ± .04 m/s, medium: .76 ± .04,
fast: .77 ± .05 m/s).
Considering only MO data, the results of the mixed-
design ANOVA on participants’ mean pointing velocity
confirmed that AD patients were significantly slower than
healthy participants (Group: F(1, 37) = 38.59,
p< .0001), and showed a significant effect of the factor
Velocity (F(2, 74) = 14.99, p< .0001). The latter
indicates that both healthy elderly subjects and AD
patients’ performances were influenced by the velocity of
the stimulus. A linear regression model described the
relationships between observed and executed
movement velocities for each participant (r2 > .6). The
slopes of the linear models give a percentage of how
much participants’ motions were influenced by the dot’s
velocity. A one-way ANOVA comparing the slopes
values of the two groups showed that CG and AD
participants’ performances were equally influenced by
the observed motion (mean slope values ± standard
error, AD: .24 ± .05, CG: .23 ± .07). Moreover, each
set of slope values was compared with a hypothetical
non-imitative behaviour (horizontal line, slope = 0) using
two paired t-test, whose results uncovered a significant
difference (for both CG and AD, p< .01). Hence, AD
patients and healthy elderly imitated the stimulus
velocities in the same manner.
Test on inhibitory mechanisms. Interestingly, patients’
rt considerably decreased in this experiment when
compared to healthy participants. Fig. 4a shows
participants’ movement starting times as a percentage
of the three stimulus velocities (AD-red and CG-blue
dots) together with the stimulus velocity profile (grey
line). The continuous blue and red lines represent the
time elapsed between the end of the stimulus motion
(rightward vertical grey line) and participants’ movement
onset. As expected, healthy participants were able to
follow the experimental instruction that is to wait until
the stimulus stopped before starting the movement. In
contrast, AD patients’ movement tended to start before
the stimulus reached the final position, while the
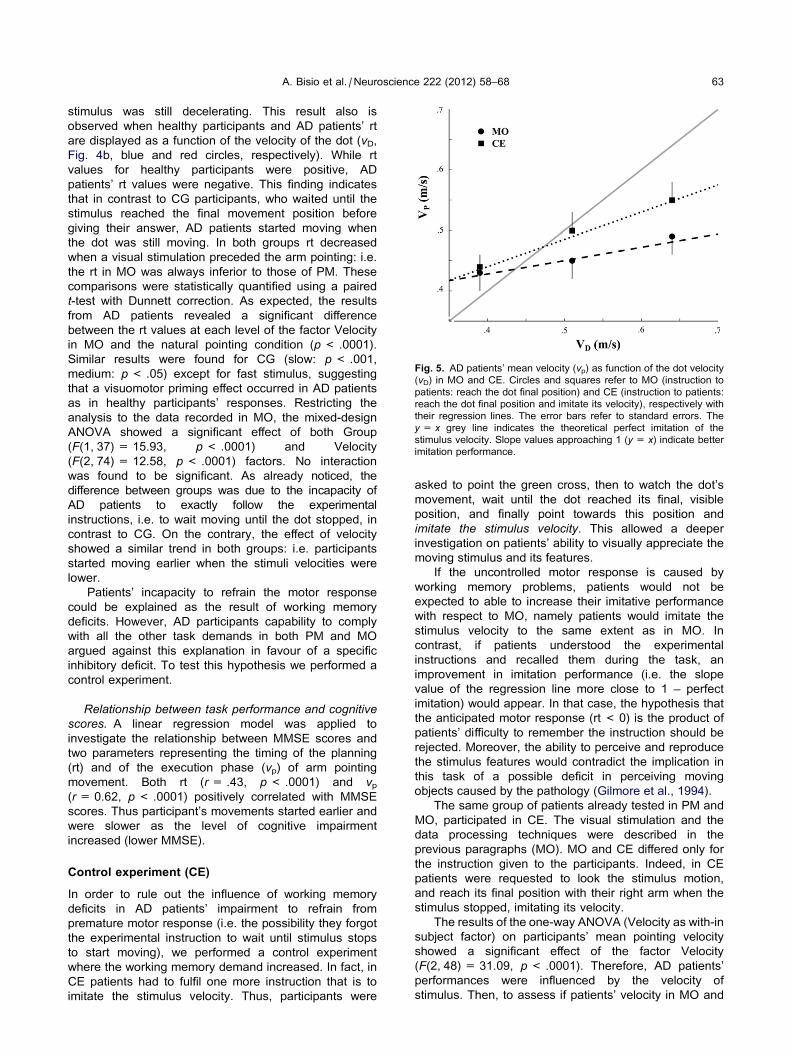
Fig. 5. AD patients’ mean velocity (vp) as function of the dot velocity
(vD) in MO and CE. Circles and squares refer to MO (instruction to
patients: reach the dot final position) and CE (instruction to patients:
reach the dot final position and imitate its velocity), respectively with
their regression lines. The error bars refer to standard errors. The
y= x grey line indicates the theoretical perfect imitation of the
stimulus velocity. Slope values approaching 1 (y= x) indicate better
imitation performance.
A. Bisio et al. / Neuroscience 222 (2012) 58–68 63
stimulus was still decelerating. This result also is
observed when healthy participants and AD patients’ rt
are displayed as a function of the velocity of the dot (vD,Fig. 4b, blue and red circles, respectively). While rt
values for healthy participants were positive, AD
patients’ rt values were negative. This finding indicates
that in contrast to CG participants, who waited until the
stimulus reached the final movement position before
giving their answer, AD patients started moving when
the dot was still moving. In both groups rt decreased
when a visual stimulation preceded the arm pointing: i.e.
the rt in MO was always inferior to those of PM. These
comparisons were statistically quantified using a paired
t-test with Dunnett correction. As expected, the results
from AD patients revealed a significant difference
between the rt values at each level of the factor Velocity
in MO and the natural pointing condition (p< .0001).
Similar results were found for CG (slow: p< .001,
medium: p< .05) except for fast stimulus, suggesting
that a visuomotor priming effect occurred in AD patients
as in healthy participants’ responses. Restricting the
analysis to the data recorded in MO, the mixed-design
ANOVA showed a significant effect of both Group
(F(1, 37) = 15.93, p< .0001) and Velocity
(F(2, 74) = 12.58, p< .0001) factors. No interaction
was found to be significant. As already noticed, the
difference between groups was due to the incapacity of
AD patients to exactly follow the experimental
instructions, i.e. to wait moving until the dot stopped, in
contrast to CG. On the contrary, the effect of velocity
showed a similar trend in both groups: i.e. participants
started moving earlier when the stimuli velocities were
lower.
Patients’ incapacity to refrain the motor response
could be explained as the result of working memory
deficits. However, AD participants capability to comply
with all the other task demands in both PM and MO
argued against this explanation in favour of a specific
inhibitory deficit. To test this hypothesis we performed a
control experiment.
Relationship between task performance and cognitivescores. A linear regression model was applied to
investigate the relationship between MMSE scores and
two parameters representing the timing of the planning
(rt) and of the execution phase (vp) of arm pointing
movement. Both rt (r= .43, p< .0001) and vp(r= 0.62, p< .0001) positively correlated with MMSE
scores. Thus participant’s movements started earlier and
were slower as the level of cognitive impairment
increased (lower MMSE).
Control experiment (CE)
In order to rule out the influence of working memory
deficits in AD patients’ impairment to refrain from
premature motor response (i.e. the possibility they forgot
the experimental instruction to wait until stimulus stops
to start moving), we performed a control experiment
where the working memory demand increased. In fact, in
CE patients had to fulfil one more instruction that is to
imitate the stimulus velocity. Thus, participants were
asked to point the green cross, then to watch the dot’s
movement, wait until the dot reached its final, visible
position, and finally point towards this position and
imitate the stimulus velocity. This allowed a deeper
investigation on patients’ ability to visually appreciate the
moving stimulus and its features.
If the uncontrolled motor response is caused by
working memory problems, patients would not be
expected to able to increase their imitative performance
with respect to MO, namely patients would imitate the
stimulus velocity to the same extent as in MO. In
contrast, if patients understood the experimental
instructions and recalled them during the task, an
improvement in imitation performance (i.e. the slope
value of the regression line more close to 1 – perfect
imitation) would appear. In that case, the hypothesis that
the anticipated motor response (rt < 0) is the product of
patients’ difficulty to remember the instruction should be
rejected. Moreover, the ability to perceive and reproduce
the stimulus features would contradict the implication in
this task of a possible deficit in perceiving moving
objects caused by the pathology (Gilmore et al., 1994).
The same group of patients already tested in PM and
MO, participated in CE. The visual stimulation and the
data processing techniques were described in the
previous paragraphs (MO). MO and CE differed only for
the instruction given to the participants. Indeed, in CE
patients were requested to look the stimulus motion,
and reach its final position with their right arm when the
stimulus stopped, imitating its velocity.
The results of the one-way ANOVA (Velocity as with-in
subject factor) on participants’ mean pointing velocity
showed a significant effect of the factor Velocity
(F(2, 48) = 31.09, p< .0001). Therefore, AD patients’
performances were influenced by the velocity of
stimulus. Then, to assess if patients’ velocity in MO and

64 A. Bisio et al. / Neuroscience 222 (2012) 58–68
CE were different, a repeated measure ANOVA was
applied (two factors: Experiment with two levels, MO and
CE, and Velocity with three levels, slow, medium, and
fast). The results showed a significant interaction
between Experiment and Velocity (F(2, 48) = 5.28,
p< 0.01). The Newman–Keuls post hoc comparison
revealed that a significant difference occurred among
each of the three levels of the factor velocity in MO and
CE (p< 0.01). Moreover, patients’ responses in MO and
CE differed in the medium and the fast conditions
(p< 0.001). At last, in order to evaluate if the imitation
performance increased in CE, the slopes values of the
linear regression models applied to each patients’ data
(see Data treatments – Data analysis) in MO and CE
were statistically compared. The results of the one-way
ANOVA showed that CE mean slope values were
significantly higher than MO mean slope values
(mean ± SE, MO: .24 ± .05, CE: .45 ± .06,
F(1, 24) = 8.62, p< 0.01). Namely, when AD patients
were explicitly asked to imitate the stimulus velocity,
imitation performance was better than when they were
only asked to reach the final stimulus position (Fig. 5).
Together with this confirmation of the patients’ ability
to visually perceive the modifications of the moving
stimulus, these results support the hypothesis that AD
patients understood the experimental instructions and
were able to use that during movement execution.
Therefore, these findings argue against the explanation
that AD patients’ incapacity to refrain the motor
response is caused by problems in memorizing the
experimental instruction.
DISCUSSION
This study had a threefold purpose: (1) measuring the
characteristics of natural pointing movements in
normal elderly participants and AD patients; (2) testing if
and how AD patients’ actions were influenced by a
previously observed motion (test on motor resonance
mechanisms); (3) assessing AD patients’ voluntary
capabilities to control motor response production (test on
inhibitory mechanisms). To these aims patients’
behaviour was measured using a simple arm’s upwards
pointing movement, which was recorded in natural
condition and after the observation of a dot displacing
vertically.
Kinematic features of the participants’ pointingmovement in natural condition (PM)
The mean reaction time (rt) of AD participants was similar
to that of healthy elderly subjects, in contrast to rt
recorded during more challenging tasks requiring
complex decision-making components (Pirozzolo et al.,
1981; Storandt and Beaudreau, 2004; van Deursen
et al., 2009) or complicated sensorimotor transformation
(Tippett and Sergio, 2006; Tippett et al., 2007) in which
the reaction time to the stimulus was greater in AD
patients. This result suggests that the planning of simple
movements has not been deteriorated by AD, probably
because it involves low level cognitive processes that
are not affected by the pathology. At the same time our
results indicate that rt associated with simple movement
is not a sensitive measure for discriminating healthy
people from AD patients as proposed by (Storandt and
Beaudreau, 2004).
Even though patients prepared the response in the
same amount of time as did the CG subjects, they
were not able to maintain the initial motor plan
throughout its course, as indicated by the increased
movements duration and jerk with respect to healthy
aged-matched participants (see also Ghilardi et al.,
1999, 2000; Tippett and Sergio, 2006; Yan et al.,
2008). The presence of these altered kinematic
parameters was correlated with the results of MMSE,
the most commonly used instrument for screening
cognitive functions. Indeed, when the cognitive
impairment increased (and MMSE score decreased) the
motion’s duration increased and the velocity profiles
became more fragmented (higher jerk value), which
indicates several online adjustments to the initial
planned trajectory. Unfortunately, we are not able to
provide a complete description of the results of the
neuropsychological assessment the patients underwent.
Therefore, we cannot speak about any possible
correlations between the kinematic performance and
the neuropsychological evaluation. Having stated this
major limitation to our study, we maintain that our data
suggest that AD patients need to continuously monitor
the ongoing action (in line with the findings of Bellgrove
et al., 1997; Ghilardi et al., 1999) and are dependent
on sensory feedback during the execution of
movements. This is consistent with the computational
theory on motor control that proposes that when
checking current motion with the desired one, sensory
feedbacks are compared to the instantaneous efferent
copy. This mechanism would allow the prediction of the
next state of the system (Wolpert and Kawato, 1998)
as well as providing updates to the internal model that
is the memory of the action. The altered corticocortical
connectivity however is specific to AD (Braak and
Braak, 1991) and might introduce a delay in the
forward and feedback mechanisms. This delay would
consequently compromise the online updating of the
motor response (Scott, 2004). Thus, a speculative
interpretation of the present findings is that slow and
jerky pointing movements represent an AD patients’
effort to compensate for this type of short term memory
deficiency created by the mismatch between the actual
and the predicted state of the body.
Conversely, the observed asymmetry of patients’
velocity profiles (Pozzo et al., 1998) suggests that the
intact representation and integration of the gravitational
force field in AD patients’ motor plan is preserved: i.e.
the time to peak velocity (tpv) occurred around the 40%
of the trajectory duration. Indeed, tpv values did not
significantly differ between the two groups.
Hence, the present results show that natural
movement pace (dur and vp) and smoothness (jerk),
rather than reaction time (rt) and movement timing (tpv),
are appropriate clinical markers to discriminate
pathological (AD) from normal ageing in simple motor
task.

A. Bisio et al. / Neuroscience 222 (2012) 58–68 65
Movement velocity of AD patients and healthycoetaneous is influenced by the stimulus velocity
The movements of healthy elderly participants were
implicitly influenced by the observed motion velocity,
behaviour previously noticed in healthy young adults
(Bisio et al., 2010). In that study (Bisio et al., 2010), we
showed that participants’ movements automatically
imitated the stimulus velocity only when the displayed
kinematics respected the biological law of motion. In
agreement with this result, the present findings show
that healthy elderly movements’ were influenced by the
stimulus velocities, suggesting ability to automatically
match the perceived kinematics with brain action
representation when getting older. Hence, we propose
that motor resonance mechanisms are not altered
across the lifespan.
Similarly, the AD patients’ behaviour was influenced
by the display velocity. This supports the hypothesis
that the resonance mechanisms (i.e. perception–action
matching) underlying automatic imitation are preserved
during AD. A possible objection to this interpretation is
that patients’ movement onset occurred before the end
of the stimulus motion, thus restricting the possibility to
appreciate its biological signature. Nevertheless, it was
demonstrated that the first 60% of the trajectory of a
similar moving target alone is sufficient for recognizing
biological kinematics (Pozzo et al., 2006). Thus,
because patients started moving when the target had
covered at least the 70% of its total displacements, this
alternative hypothesis seems unlikely.
The transformation of the visual input into a motor
command is commonly attributed to the activity of the
posterior parietal cortex (Decety et al., 2002). According
to the description of the neuropathological staging of AD
proposed by Braak and Braak (1991), the parietal cortex
is one of the primary anatomical area affected by early
stage Alzheimer’s disease.
This was recently confirmed and extended by a 3-
years long longitudinal neuroimaging study on the
evolution of brain atrophy in a population of aMCI
(amnestic Mild Cognitive Impairment) patients later
diagnosed with AD (Whitwell et al., 2007). Indeed,
1 year prior to the diagnosis, the parietal lobe
involvement was noticed as well as widespread cerebral
atrophy in the medial temporal lobe. Nevertheless, since
a behavioural influence of the visual model was present
in patients’ performance, one might suppose that intact
brain regions allow this translation to occur. Previous
imaging studies on healthy individuals (Grezes and
Decety, 2001; Jeannerod, 2001; Iacoboni, 2009a)
showed that motion observation and imitation induce
simultaneous activation of both the parietal and
premotor areas in the regions where the human mirror
neurons system are thought to be located. Rizzolatti
et al. (1999) proposed that these brain areas would give
rise to a resonance mechanism that directly (and
implicitly) maps a pictorial or kinematic description of the
observed action onto an internal motor representation of
the same action (i.e. the direct matching hypothesis). By
showing an automatic imitation of the observed motion
into movement production, the present paradigm gives
indirect cues about the current activity of this
mechanism in AD. Thus, a speculative interpretation of
the present findings is that the areas considered to be
part of the fronto-parietal mirror neuron system (MNS)
might be preserved from the alterations induced by AD.
However, although the frontal lobes are relatively spared
until the moderate stage of the illness (Double et al.,
1996; Salat et al., 1999, 2001; Whitwell et al., 2007),
this is not the case for the parietal regions. In this
regard, a very recent review (Jacobs et al., 2012)
describes the structural, functional and metabolic
changes observed in the parietal cortex (including the
inferior parietal lobule – the parietal region of the MNS)
in preclinical and early AD. An alternative explanation to
the preservation of the MNS areas is that protective-
compensatory strategies (where undamaged areas take
over the function of the injured ones (Hill and
Kolanowski, 2011)) maybe intervening to ensure these
functions that are crucial for everyday life. Therefore,
the present behavioural observations raise a question
regarding the way these mirror mechanisms appear in a
damaged brain like that of AD patients. Hence, this
study would like to promote the application of specific
neuroimaging and neurophysiological methodologies to
specifically tackle this issue and to describe the
evolution of MNS functioning in dementia. Moreover, the
results obtained by this work might have considerable
impact on therapeutic applications. Indeed, testing
motor resonance mechanisms might be a valid tool for
indicating the presence of intact social cognition abilities
in this kind of patients that often show difficulty to
interact with relatives or healthcare staff. Since this
methodology tests automatic and unconscious
responses, it could be more appropriate than other
conventional techniques where explicit choices and
explanations are required (e.g. example of TOM test
(Gregory et al., 2002)). Moreover, while explicit re-
learning seems to be inappropriate for AD patients,
implicit methods based on the influence exerted by the
observed stimuli on action production would represent
adequate tools for rehabilitation programs in addition to
conventional techniques. For instance, specific
treatments aiming at stimulating motor resonance
mechanisms, as in the case of imitation paradigms,
might be useful to maintain and/or improve AD patients’
communication skills.
Uncontrolled initiation of AD patients’ motorresponse while observing a moving stimulus
In MO, the rt values were velocity dependent for both
normal participants and AD patients: that is the rt
increased when the stimulus velocity increased. A
perceptual difficulty in perceiving and interacting with
fast moving objects could explain this effect (Rizzo and
Nawrot, 1998), and consequently would affect the first
step of the perception–action-matching system in the
two groups (see above).
Most interesting was the AD patients’ incapacity to
refrain from the pointing motion while observing the
moving stimulus. In fact, although healthy participants
were able to comply with the instructions given by the

66 A. Bisio et al. / Neuroscience 222 (2012) 58–68
experimenter, i.e. to wait starting moving until the stimulus
stopped, AD participants’ movement onsets always
occurred before the end of the dot’s motion (Fig. 4).
Thus, the mere presence of the stimulus was sufficient
to trigger the action. Since one of the most common
signs of AD is the memory loss, memory difficulties had
to be ruled out to ensure that difficulties remembering
the motion features or the task directions were not the
cause of the different pattern of compliance with the task
instructions. The fact that AD participants were able to
comply with all task demands in both the PM (to point
the initial green cross with the right arm, to wait until two
circles appear and to move towards the upwards one)
and the MO (e.g. to make a straight vertical arm
movement towards the stimulus final position)
experiments suggest memory deficits were not causing
the observed difference. Moreover, in order to further
rule out this ‘‘working memory deficit hypothesis’’ we
increased the difficulty of the task by adding an
additional instruction, that is to imitate the stimulus
velocity (control experiment – CE), and found that the
imitative performance actually increased with respect to
MO. Therefore, patients remembered the additional
request and were able to accomplish it. This finding
further argues against the explanation that AD patients’
impulsivity was due to problems with recalling the
experimental instruction. Most likely, their responses
may indicate that they are dependent on the visual
stimulus due to inadequate functioning of the inhibitory
mechanisms on the automatic perception–action
coupling. A similar behaviour was previously reported in
AD patients by (Lhermitte, 1986), who described
patients’ tendency to imitate the gesture of people
around them (i.e. imitation behaviour) and to
compulsively act on nearby objects (i.e. utilization
behaviour). The incapacity to refrain from movement
initiation while observing the moving stimulus would
confirm these clinical observations, uncovering a
dependence on environmental cues which is known as
environmental dependency syndrome, and commonly
seen in people with frontal lobes damages (Lhermitte,
1986). Indeed, topographical (Double et al., 1996),
imaging (Scahill et al., 2002) and behavioural (Rossit
and Harvey, 2008) studies reported morphology and
metabolism alterations of the frontal lobes in AD patients
that may explain this unrestrained behaviour. However,
alterations in frontal areas, similar to what is seen in AD
patients, have also been described in normal elderly
people (Double et al., 1996; Salat et al., 1999, 2001)
suggesting that the behavioural differences between AD
and healthy age-matched participants could not be
attributed only to frontal lobes damages. Thus, one could
speculate that lesions to other brain structures are
responsible of these findings (Perry and Hodges, 1999).
Actually, in a review on inhibitory functioning in AD
(Amieva et al., 2004), it was postulated that tasks
involving controlled/voluntary inhibition rely on the
activity of several structures distributed in the brain,
including the frontal regions and their cortical
connections. Interestingly, AD involves a breakdown of
the connections between anterior and posterior cortical
association areas (Braak and Braak, 1991) which could
prevent efficient communication between brain regions
and could provoke the immediate execution of motor
output in response to motion observation (i.e. action–
perception matching). The present results are supported
by and extend to the sensorimotor domain the findings of
previous experiments in which the inhibitory capabilities
were tested by asking participants to suppress an
overlearned response while executing a less overlearned
response (Logan and Cowan, 1984), as in the Stroop
test (Koss et al., 1984; Fisher et al., 1990) and in the
Stop Signal task (Amieva et al., 2002).
In conclusion, the present results designate motor
inhibitory deficiency as a feature that allows
discriminating AD patients from healthy cohort which is
in agreement with previous studies.
CONCLUSION
This work is the first to identify and quantify automatic
imitation phenomena occurring in healthy elderly people
and AD patients when observing a moving stimulus. The
results suggest that the resonance mechanisms
underlying social cognition are preserved during normal
and pathological ageing. The preservation of this
mechanism, independent of conscious awareness, is a
significant finding relevant for physical and cognitive
interventions. Moreover, the premature motor initiation
described here extends previous observations obtained
during more demanding cognitive tasks and generalizes
AD inhibition deficiencies to the sensorimotor domain.
This evidence, in addition to the specific kinematic
features of AD patients’ movements (high jerk and low
velocity), could represent an innovative new tool for
screening tests for AD. However, first this paradigm
needs to be assessed in patients affected by other
neurodegenerative diseases to shed light on the
specificity of this behaviour to AD. In conclusion, we
believe that this work characterizes the sensorimotor
abilities of AD patients and offers an innovative
approach to design new clinical interventions for healthy
elderly and patients with dementia.
Acknowledgement—The authors wish to thank Shannon Hennig
who kindly provided language editing.
REFERENCES
Adolphs R (1999) Social cognition and the human brain. Trends Cogn
Sci 3:469–479.
Amieva H, Lafont S, Auriacombe S, Carret NL, Dartigues J-F,
Orgogozo J-M, Colette F (2002) Inhibitory breakdown and
dementia of the Alzheimer type: a general phenomenon? J Clin
Exp Neuropsychol 24:503–516.
Amieva H, Phillips LH, Sala SD, Henry JD (2004) Inhibitory
functioning in Alzheimer’s disease. Brain 127:949–964.
Bellgrove MA, Phillips JG, Bradshaw JL, Hall KA, Presnell I, Hecht H
(1997) Response programming in dementia of the Alzheimer type:
a kinematic analysis. Neuropsychologia 35:229–240.
Bien N, Roebroeck A, Goebel R, Sack AT (2009) The brain’s intention
to imitate: the neurobiology of intentional versus automatic
imitation. Cereb Cortex 19:2338–2351.

A. Bisio et al. / Neuroscience 222 (2012) 58–68 67
Bisio A, Stucchi N, Jacono M, Fadiga L, Pozzo T (2010) Automatic
versus voluntary motor imitation: effect of visual context and
stimulus velocity. PLoS ONE 5(10):e13506. http://dx.doi.org/
10.1371/journal.pone.0013506.
Borson S, Raskind MA (1997) Clinical features and pharmacologic
treatment of behavioral symptoms of Alzheimer’s disease.
Neurology 48:S17–S24.
Braak H, Braak E (1991) Neuropathological staging of Alzheimer-
related changes. Acta Neuropathol 82:239–259.
Brainard DH (1997) The psychophysics toolbox. Spat Vis
10:433–436.
Cuerva AG, Sabe L, Kuzis G, Tiberti C, Dorrego F, Starkstein SE
(2001) Theory of mind and pragmatic abilities in dementia.
Neuropsychiatry Neuropsychol Behav Neurol 14:153–158.
Decety J, Chaminade T, Grezes J, Meltzoff AN (2002) A PET
exploration of the neural mechanisms involved in reciprocal
imitation. Neuroimage 15:265–272.
Double KL, Halliday GM, Kril JJ, Harasty JA, Cullen K, Brooks WS,
Creasey H, Broe GA (1996) Topography of brain atrophy during
normal aging and Alzheimer’s disease. Neurobiol Aging
17:513–521.
Fisher LM, Freed DM, Corkin S (1990) Stroop color-word test
performance in patients with Alzheimer’s disease. J Clin Exp
Neuropsychol 12:745–758.
Fournet N, Mosca C, Moreaud O (2007) [Deficits in inhibitory
processes in normal aging and patients with Alzheimer’s
disease: a review] Deficits des processus inhibiteurs dans le
vieillissement normal et la maladie d’Alzheimer. Psychol
Neuropsychiatr Vieil 5:281–294.
Frith CD, Frith U (2006) The neural basis of mentalizing. Neuron
50:531–534.
Ghilardi MF, Alberoni M, Marelli S, Rossi M, Franceschi M, Ghez C,
Fazio F (1999) Impaired movement control in Alzheimer’s
disease. Neurosci Lett 260:45–48.
Ghilardi MF, Alberoni M, Rossi M, Franceschi M, Mariani C, Fazio F
(2000) Visual feedback has differential effects on reaching
movements in Parkinson’s and Alzheimer’s disease. Brain Res
876:112–123.
Gilmore GC, Wenk HE, Naylor LA, Koss E (1994) Motion perception
and Alzheimer’s disease. J Gerontol 49:P52–P57.
Gregory C, Lough S, Stone V, Erzinclioglu S, Martin L, Baron-Cohen
S, Hodges JR (2002) Theory of mind in patients with frontal
variant frontotemporal dementia and Alzheimer’s disease:
theoretical and practical implications. Brain 125:752–764.
Grezes J, Decety J (2001) Functional anatomy of execution, mental
simulation, observation, and verb generation of actions: a meta-
analysis. Hum Brain Mapp 12:1–19.
Heyes C (2011) Automatic imitation. Psychol Bull 137:463–483.
Hill NL, Kolanowski AM (2011) Cognitive rehabilitation in people with
dementia. In: Aging well: gerontological education for nurses and
other health professionals, (Jones & Bartlett Learning, eds), pp
263–284.
Iacoboni M (2005) Neural mechanisms of imitation. Curr Opin
Neurobiol 15:632–637.
Iacoboni M (2009a) Imitation, empathy, and mirror neurons. Annu
Rev Psychol 60:653–670.
Iacoboni M (2009b) Neurobiology of imitation. Curr Opin Neurobiol
19:661–665.
Iacoboni M, Dapretto M (2006) The mirror neuron system and the
consequences of its dysfunction. Nat Rev Neurosci 7:942–951.
Iacoboni M, Woods RP, Brass M, Bekkering H, Mazziotta JC,
Rizzolatti G (1999) Cortical mechanisms of human imitation.
Science 286:2526–2528.
Jacobs HI, Van Boxtel MP, Jolles J, Verhey FR, Uylings HB (2012)
Parietal cortex matters in Alzheimer’s disease: an overview of
structural, functional and metabolic findings. Neurosci Biobehav
Rev 36:297–309.
Jeannerod M (2001) Neural simulation of action: a unifying
mechanism for motor cognition. Neuroimage 14:S103–S109.
Kluger A, Gianutsos JG, Golomb J, Ferris SH, George AE, Franssen
E, Reisberg B (1997) Patterns of motor impairment in normal
aging, mild cognitive decline, and early Alzheimer’s disease. J
Gerontol B Psychol Sci Soc Sci 52:P28–P39.
Koski L, Wohlschlager A, Bekkering H, Woods RP, Dubeau M-C,
Mazziotta JC, Iacoboni M (2002) Modulation of motor and
premotor activity during imitation of target-directed actions.
Cereb Cortex 12:847–855.
Koss E, Ober BA, Delis DC, Friedland RP (1984) The Stroop color-
word test: indicator of dementia severity. Int J Neurosci 24:
53–61.
Lhermitte F (1986) Human autonomy and the frontal lobes. Part II:
Patient behavior in complex and social situations: the
‘‘environmental dependency syndrome’’. Ann Neurol 19:335–343.
Lhermitte F, Pillon B, Serdaru M (1986) Human autonomy and the
frontal lobes. Part I: Imitation and utilization behavior: a
neuropsychological study of 75 patients. Ann Neurol 19:
326–334.
Logan GD, Cowan W (1984) On the ability to inhibit thought and
action: a theory on an act of control. Psychol Rev 91:295–327.
Manckoundia P, Mourey F, Pfitzenmeyer P, Papaxanthis C (2006)
Comparison of motor strategies in sit-to-stand and back-to-sit
motions between healthy and Alzheimer’s disease elderly
subjects. Neuroscience 137:385–392.
Meltzoff AN (1999) Origins of theory of mind, cognition and
communication. J Commun Disord 32:251–269.
Meltzoff AN, Moore MK (1977) Imitation of facial and manual gestures
by human neonates. Science 198:75–78.
Meltzoff AN, Prinz W (2002) The imitative mind: development,
evolution, and brain bases. Cambridge, England: Cambridge
University Press.
Papaxanthis C, Pozzo T, Stapley P (1998) Effects of movement
direction upon kinematic characteristics of vertical arm pointing
movements in man. Neurosci Lett 253:103–106.
Perneczky R, Wagenpfeil S, Komossa K, Grimmer T, Diehl J, Kurz A
(2006) Mapping scores onto stages: mini-mental state
examination and clinical dementia rating. Am J Geriatr
Psychiatry 14:139–144.
Perry RJ, Hodges JR (1999) Attention and executive deficits in
Alzheimer’s disease. Brain 122:383–404.
Pirozzolo FJ, Christensen KJ, Ogle KM, Hansch EC, Thompson WG
(1981) Simple and choice reaction time in dementia: clinical
implications. Neurobiol Aging 2:113–117.
Pozzo T, Papaxanthis C, Petit JL, Schweighofer N, Stucchi N (2006)
Kinematic features of movement tunes perception and action
coupling. Behav Brain Res 169:75–82.
Pozzo T, Papaxanthis C, Stapley P, Berthoz A (1998) The
sensorimotor and cognitive integration of gravity. Brain Res
Brain Res Rev 28:92–101.
Rizzo M, Nawrot M (1998) Perception of movement and shape in
Alzheimer’s disease. Brain 121(Pt. 12):2259–2270.
Rizzolatti G, Fadiga L, Fogassi L, Gallese V (1999) Resonance
behaviors and mirror neurons. Arch Ital Biol 137:85–100.
Rizzolatti G, Fogassi L, Gallese V (2001) Neurophysiological
mechanisms underlying the understanding and imitation of
action. Nat Rev Neurosci 2:661–670.
Rossit S, Harvey M (2008) Age-related differences in corrected and
inhibited pointing movements. Exp Brain Res 185:1–10.
Salat DH, Kaye JA, Janowsky JS (1999) Prefrontal gray and white
matter volumes in healthy aging and Alzheimer disease. Arch
Neurol 56:338–344.
Salat DH, Kaye JA, Janowsky JS (2001) Selective preservation and
degeneration within the prefrontal cortex in aging and Alzheimer
disease. Arch Neurol 58:1403–1408.
Scahill RI, Schott JM, Stevens JM, Rossor MN, Fox NC (2002)
Mapping the evolution of regional atrophy in Alzheimer’s disease:
unbiased analysis of fluid-registered serial MRI. Proc Natl Acad
Sci USA 99:4703–4707.
Scott SH (2004) Optimal feedback control and the neural basis of
volitional motor control. Nat Rev Neurosci 5:532–546.
Storandt M, Beaudreau S (2004) Do reaction time measures enhance
diagnosis of early-stage dementia of the Alzheimer type. Arch Clin
Neuropsychol 19:119–124.

68 A. Bisio et al. / Neuroscience 222 (2012) 58–68
Teulings HL, Contreras-Vidal JL, Stelmach GE, Adler CH (1997)
Parkinsonism reduces coordination of fingers, wrist, and arm in
fine motor control. Experimental neurology 146:159–170.
Tippett WJ, Krajewski A, Sergio LE (2007) Visuomotor integration is
compromised in Alzheimer’s disease patients reaching for
remembered targets. Eur Neurol 58:1–11.
Tippett WJ, Sergio LE (2006) Visuomotor integration is impaired in
early stage Alzheimer’s disease. Brain Res 1102:92–102.
van Deursen JA, Vuurman EFPM, Smits LL, Verhey FRJ, Riedel WJ
(2009) Response speed, contingent negative variation and P300
in Alzheimer’s disease and MCI. Brain Cogn 69:592–599.
Verdon CM, Fossati P, Verny M, Dieudonne B, Teillet L, Nadel J
(2007) Social cognition: an early impairment in dementia of the
Alzheimer type. Alzheimer Dis Assoc Disord 21:25–30.
Whitwell JL, Przybelski SA, Weigand SD, Knopman DS, Boeve BF,
Petersen RC, Jack Jr CR (2007) 3D maps from multiple MRI
illustrate changing atrophy patterns as subjects progress from
mild cognitive impairment to Alzheimer’s disease. Brain
130:1777–1786.
Wohlschlager A, Gattis M, Bekkering H (2003) Action generation and
action perception in imitation: an instance of the ideomotor
principle. Philos Trans R Soc Lond B Biol Sci 358(1431):501–515.
Wolpert DM, Kawato M (1998) Multiple paired forward and inverse
models for motor control. Neural Netw 11:1317–1329.
Yan JH, Rountree S, Massman P, Doody RS, Li H (2008) Alzheimer’s
disease and mild cognitive impairment deteriorate fine movement
control. J Psychiatr Res 42:1203–1212.
(Accepted 10 July 2012)(Available online 17 July 2012)

Alterations with Movement Duration in the Kinematicsof a Whole Body Pointing MovementMatthieu Casteran1, Patrick Manckoundia1,2, Thierry Pozzo1,3,4, Elizabeth Thomas1*
1 UFR-STAPS, INSERM U-1093, Cognition, Action and Sensorimotor Plasticity Universite de Bourgogne, Campus Universitaire, Dijon, France, 2 Service de Medecine Interne
Geriatrie, Hopital de Champmaillot, Centre Hospitalier Universitaire, Dijon, France, 3 Italian Institute of Technology, Genoa, Italy, 4 Institut Universitaire de France, Paris,
France
Abstract
Our aim was to investigate how the organization of a whole body movement is altered when movement duration (MD) isvaried. Subjects performed the same whole body pointing movement over long, normal and short MDs. The kinematictrajectories were then analyzed on a normalized time base. A principal components analysis (PCA) revealed that the degreeof coordination between the elevation angles of the body did not change with MD. This lack of significant differences in thecoordination was interesting given that small spatial and temporal differences were observed in the individual kinematictrajectories. They were revealed by studying the trajectories of the elevation angles, joint markers and center of mass. Theelevation angle excursions displayed modifications primarily in their spatial characteristics. These alterations were moremarked for the short rather than long duration movements. The temporal characteristics of the elevation angles asmeasured by the time to peak of angular velocity were not modified in the same fashion hence displaying a dissociation inthe tuning of the spatial and temporal aspects of the elevation angles. Modifications in the temporal characteristics of themovement were also studied by examining the velocity profiles of the joint markers. Interestingly, unlike the disorderednature of this variable for the elevation angles, the time to peak velocity was neatly ordered as a function of MD for the jointmarkers – It arrived first for the short duration movements, followed by those of the normal and finally long durationmovements. Despite the modifications observed in the kinematic trajectories, a PCA with the elevation angle excursions atdifferent MDs revealed that two principal components were sufficient to account for nearly all the variance in the data. Ourresults suggest that although similar, the kinematic trajectories at different MDs are not achieved by a simple time scaling.
Citation: Casteran M, Manckoundia P, Pozzo T, Thomas E (2013) Alterations with Movement Duration in the Kinematics of a Whole Body PointingMovement. PLoS ONE 8(1): e52477. doi:10.1371/journal.pone.0052477
Editor: Paul L. Gribble, The University of Western Ontario, Canada
Received August 3, 2012; Accepted November 19, 2012; Published January 14, 2013
Copyright: � 2013 Casteran et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: Matthieu Casteran was supported by a doctoral grant from the Institut National de la Sante et de la Recherche Medicale de France and the ConseilRegional de Bourgogne. Elizabeth Thomas was supported by the ANR-10-ORAR-006-03. The funders had no role in the study design, data collection and analysis,decision to publish or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected]
Introduction
One of the fundamental features that can be adjusted in any
movement is its duration. In this study we examine how the
kinematics of a whole body movement is adjusted for different
movement durations (MD). Several studies exist on the effects of
movement duration on the kinetics and kinematics of arm
movements [1,2,3,4,5,6,7,8,9,10]. Theoretical work has been
done to show the independance from MD of their velocity profiles
and their movement path during reaching [2,10]. Experimental
work has confirmed this invariance [1,3]. Further work has shown
that the kinematic and muscular activation features reflect a
strategy for arm movements that that is more in keeping with
optimizing dynamic forces rather than minimizing antigravity
torques [8,6]. While the study of arm movements has provided us
with much insight into the organization of movement, many of our
daily activities associate focal displacements with simultaneous
postural demands. Whole body pointing movements are therefore
interesting to study. The trunk is a much heavier segment than the
arm. Reaching over could bring into play a greater role for the
gravitational component, changing the forces that are optimized
and hence could reorganize the kinematics of the whole body
pointing.
There are several more immediate reasons for carrying out the
current study. Previous research has been done on the effects of
MD on whole body pointing. This research however was restricted
to examining the effects of decreasing movement duration (MD),
i.e. at higher movement speeds [11,12,13,14]. The previously cited
studies showed that the adjustments of the whole body pointing
movements for short MDs, was not achieved by a simple time
scaling. In this study, we extend this research to look into the
results of the opposite process, i.e. increasing MD. There are many
reasons for doing this. Firstly slow movements are more subject to
various types of modifying processes that have the potential for
changing their trajectories. One of these is sensorial and especially
proprioceptive feedback. An examination of the EMGs of slow
arm movements therefore reveals trajectories that are not as
smooth as those from normal or fast movements [4,15]. This
feedback is also probably the source of a greater variability that is
frequently observed in slow movements [16]. Another potential
source of modifications in slow movements is their relationship to
gravity. Especially for moving downwards, slowing down must
involve the use of force to counter the normal gravity dictated
PLOS ONE | www.plosone.org 1 January 2013 | Volume 8 | Issue 1 | e52477

speed of the body. Nishikawa et al. [8] in studies of arm
movements over long durations did not observe differences in the
way the movement was organized. But would this observation
extend to movements involving heavier segments of the body? An
answer to this question would allow us to contribute to the picture
on ‘speed sensitive strategies’ [4,17]. It would also permit us to
have a control for studies in which long MD could be a
confounding factor. One important example of this is studies on
ageing where it is always necessary to ask if the observed effects are
due to anything more than the lower speeds with which elderly
subjects perform most movements [18,19,20,21].
The current investigation was carried out by examining the
kinematics of a whole body pointing movement executed over
long, normal and short durations. An examination of the
individual kinematic trajectories as well as their degree of
coordination with each other was carried out. The kinematic
trajectories compared at the different MDs were the elevation
angles, the joint marker positions and the center of mass (CoM)
trajectory. A visual examination of the eight elevation angle
trajectories revealed that the movements were carried out using a
similar strategy in each case. This similarity was further quantified
with the use of correlation coefficients once the dimensionality of
the space had been reduced using the principal components
analysis technique. This procedure revealed elevation angle
excursions at different MDs that were similar albeit with small
differences. The application of the PCA also allowed us to
compare the degree of covariation among the elevation angles at
each MD. It was not found to be significantly altered by MD. As
the previously described analyses had revealed small differences in
the kinematic trajectories executed over different durations, we
carried out a detailed comparison of them. For the elevation angles
we compared the amplitudes and the time to peak of their velocity
profiles. At levels that involved more integrated effects, we
examined the temporal characteristics of the joint marker velocity
profiles and the spatial characteristics of the CoM trajectory. It
should be noted that the comparisons of all trajectories were done
using a normalized time base. Despite the differences observed in
the kinematic trajectories, PCA analyses, this time using trajecto-
ries from different MDs revealed that two common waveforms
could be combined to produce the movement trajectories at
different MDs. This therefore provides a means for reducing the
number of degrees of freedom for whole body pointing [22].
Materials and Methods
ParticipantsEleven healthy adults, 3 women and 8 men (mean age: 2666
years; mean height: 1.73 m60.08 m; mean weight: 66611 kg)
took part in the experiments. None of the subjects had any
previous neurological diseases and they had normal or corrected to
normal vision. The experiments conformed to the Declaration of
Helsinki and written consent was obtained from all the partic-
ipants. The study was approved by the Ethics Committee of the
University of Burgundy.
Motor TaskAll the participants performed a Whole Body Pointing (WBP)
movement. The experimental procedures have been used and
validated in previous studies [12,23,24,25]. We asked participants
to perform a WBP movement simultaneously with their two index
fingers in order to touch two targets. The targets (462 cm) were
separated by a distance of 0.5 m from their centres and positioned
on a piece of wood. They were placed at a distance corresponding
to 15% of each participant’s height in the anteroposterior (AP)
plane and in the vertical plane. Distances were measured from the
distal end of each participant’s big toe. Participants started from an
upright position. Their hands were positioned so that the
hypothenar eminence was in contact with the thighs. Only, the
index finger remained extended while the rest of the fingers were
bent. MD and target accuracy were the primary constraints
imposed on the participants.
Movement duration constraintsThe WBP movement was carried out over three different
durations. These were a self-selected duration (N), long duration
(Lo) and shorter than normal (Sh) (without asking participants to
go as fast as possible). For each movement time, all the subjects
carried out about 5 preliminary unrecorded WBP movements in
order to familiarize themselves with the movement and the
necessary durations. This was then followed by a block of ten
movements for each movement time. There was a two minutes
pause between movement blocks of each duration. Each subject
therefore performed a total of 30 trials.
Data collection and processingWe used an optoelectronic device (VICON, sampling frequency
200 Hz) with three cameras in order to capture movement
kinematics in 3 dimensions (3D). Twelve retro-reflective markers
(0.2 m in diameter) were placed at various anatomical locations on
the right side of the body (External cantus of eye, auditory meatus,
acromial process, humeral condyle, ulnar styloid process, apex of
the index finger, L5 vertebra, greater trochanter, knee interstitial
joint space, external malleolus, fifth metatarsal head of the foot,
and the middle of arm in order to have 3D with the VICON
system). We used a 9 segment model similar to our previous studies
of the same movement [24,25].
All processing of the 3D marker positions was performed with
custom software written in Matlab (Mathworks). Before the
computation of the angular trajectories, the recorded marker
position signals were low-pass filtered using a fourth-order
Butterworth filter at a cut-off frequency of 10 Hz (Matlab filtfilt
function). The filtering was followed by the use of interpolation
routines (Matlab spline function) so that all trajectories irrespective
of execution duration lay along a 200 point time base.
Kinematic computationsMovement onset was defined as the time when the velocity of
the finger exceeded 5% of its peak and movement cessation was
noted likewise when this velocity dropped below the 5% threshold
[26]. Kinematic parameters including angular displacements were
computed using previously reported techniques [27,24]. The
following eight elevation angles (angle between the vertical and the
segment) were calculated: Shank (Sh); Thigh (Th); Pelvis (Pe);
Trunk (Tr); Humerus (Hu); Forearm (Fo); Hand (Ha) and Head
(He) (figure 1b and 2). The amplitude of each angular
displacement was defined as the absolute value of the difference
between the initial and final angle.
Centre of Mass analysisWe calculated the CoM displacements in 3D in order to
characterize the manner in which equilibrium was managed
during the WBP movement. This estimation was made from an
eight-segment mathematical model using rigid segments (Head,
Trunk, Thigh, Shank, Foot, Upper arm, Forearm and Hand). For
this, we used the anthropometric parameters described by Winter
[29] and validated by Stapley [11] and Berret [24] in previous
studies of WBP movement. Stapley [11] had compared the
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 2 January 2013 | Volume 8 | Issue 1 | e52477

modeled CoM and measured Centre of Pressure (CoP) position
using a force platform, during quiet stance as well as the times
series of measured and estimated (modeled) ground reaction
forces. These studies showed that such a model provided a realistic
representation of the WBP CoM position.
Principal Component AnalysisA PCA [28] was applied to the angular displacements of the
eight elevation angles. As the z scores of the elevation angles and
their correlation matrices were used for this computation, the
results obtained provided us with information concerning the
linear correlation of these angles. In all cases, the PCA was
performed separately for each individual. For the results described
in section 3.2, the trials came from whole body pointing of only
one duration. For the results described in section 3.6, however, the
trials were either from the normal and short MDs or from the
normal and long MD trials.
In all cases two principal components were found sufficient to
account for more than 95% of the variance in the data. This is
referred to as the variance accounted for (VAF).
Statistical analysisAll statistical analyses were performed primarily using a
repeated measures analysis of variances (ANOVA) test. The test
was applied after ensuring a normal distribution of the data using
the Kolgomorov-Smirnov and Lillefors test. The MDs were in all
cases a repeated measures factor. The kinematic trajectory
amplitudes and peak times as well as the eigen values from the
PCA were also used as repeated measure factors depending on the
question at hand. The post hoc tests were done using a Tukey
HSD. Results were taken as statistically significant if p,0.05.
Results
In this section we will report on the analysis of the whole body
pointing (WBP) movements accomplished over different durations.
We will first report on the general characteristics of the movements
for the three durations - long, normal and short. The similarities
between the kinematic trajectories of the movements were
quantified using correlation coefficients after having reduced the
dimensionality of this space with the PCA technique. Since this
process revealed slight differences, we undertook a comparison of
the individual kinematic trajectories at different MDs. The
trajectories examined were those of the elevation angles, the joint
markers and the CoM. Both the spatial and temporal character-
istics of these kinematic trajectories were examined. Finally a
principal components analysis using kinematic trajectories from
different movement types was used to probe if common waveforms
can be combined to generate the kinematic trajectories generated
over different durations.
3.1 Movement at three speeds: General characteristicsOur first step was to verify that the subjects did indeed carry out
the movements at the instructed pace. The mean MD was
1.2360.11 s for normal MD, 1.9460.23 s for long MD
0.7760.06 s for short MD. The three were found to be
significantly different from each other (p,0.05, repeated measures
ANOVA). The mean difference between the long duration and
normal movements was found to be on average slightly higher
than those between the normal and short duration movements. As
the subjects were asked to touch a target that was sufficiently wide
and long (see Methods) all attempts to touch the target at the three
different speeds were successful. The peak velocities for the
normal, long duration and short duration movements were
0.8860.08, 0.5660.06 and 1.4460.11 m.s21 respectively.
Figure 1. Stick diagrams. a) Stick diagrams of a whole body pointing movement to a target that is placed at 15% on the anteroposterior axis andon the vertical axis. b) The computed elevation angles for the movements were the Shank (Sk), Thigh (Th), Pelvis (Pe), Truck (Tr), Head (He), Humerus(Hu), Forearm (Fo) and Hand (Ha).doi:10.1371/journal.pone.0052477.g001
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 3 January 2013 | Volume 8 | Issue 1 | e52477
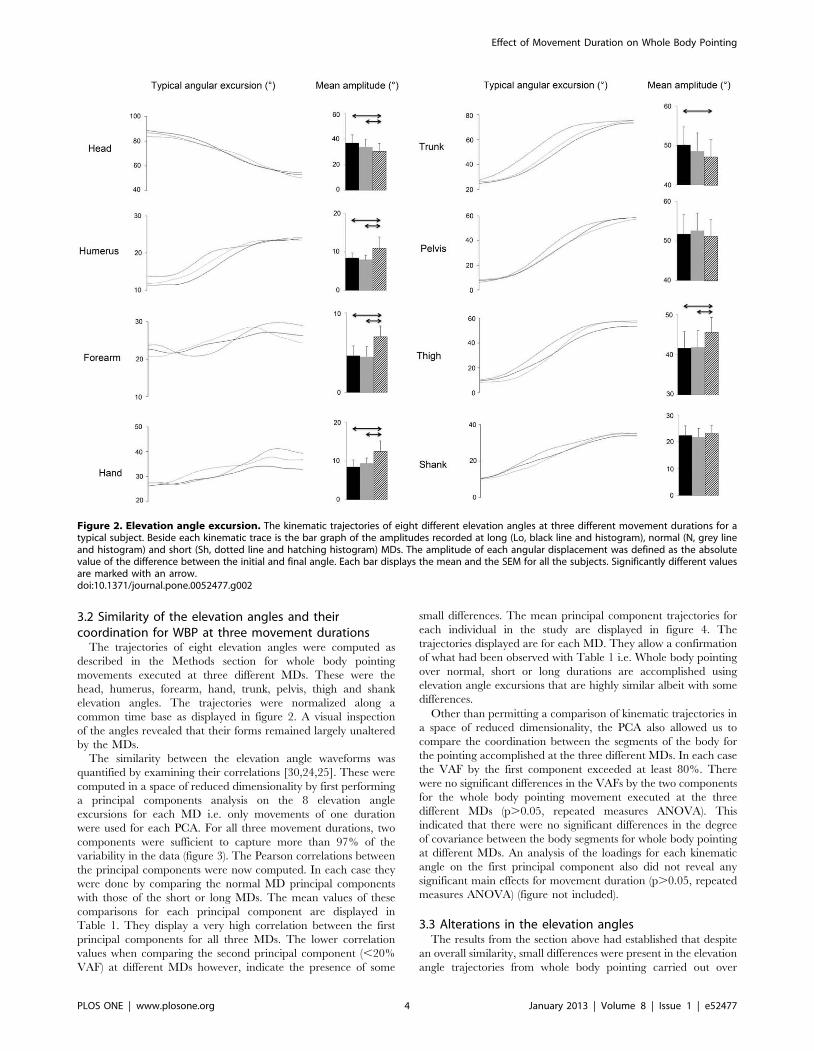
3.2 Similarity of the elevation angles and theircoordination for WBP at three movement durations
The trajectories of eight elevation angles were computed as
described in the Methods section for whole body pointing
movements executed at three different MDs. These were the
head, humerus, forearm, hand, trunk, pelvis, thigh and shank
elevation angles. The trajectories were normalized along a
common time base as displayed in figure 2. A visual inspection
of the angles revealed that their forms remained largely unaltered
by the MDs.
The similarity between the elevation angle waveforms was
quantified by examining their correlations [30,24,25]. These were
computed in a space of reduced dimensionality by first performing
a principal components analysis on the 8 elevation angle
excursions for each MD i.e. only movements of one duration
were used for each PCA. For all three movement durations, two
components were sufficient to capture more than 97% of the
variability in the data (figure 3). The Pearson correlations between
the principal components were now computed. In each case they
were done by comparing the normal MD principal components
with those of the short or long MDs. The mean values of these
comparisons for each principal component are displayed in
Table 1. They display a very high correlation between the first
principal components for all three MDs. The lower correlation
values when comparing the second principal component (,20%
VAF) at different MDs however, indicate the presence of some
small differences. The mean principal component trajectories for
each individual in the study are displayed in figure 4. The
trajectories displayed are for each MD. They allow a confirmation
of what had been observed with Table 1 i.e. Whole body pointing
over normal, short or long durations are accomplished using
elevation angle excursions that are highly similar albeit with some
differences.
Other than permitting a comparison of kinematic trajectories in
a space of reduced dimensionality, the PCA also allowed us to
compare the coordination between the segments of the body for
the pointing accomplished at the three different MDs. In each case
the VAF by the first component exceeded at least 80%. There
were no significant differences in the VAFs by the two components
for the whole body pointing movement executed at the three
different MDs (p.0.05, repeated measures ANOVA). This
indicated that there were no significant differences in the degree
of covariance between the body segments for whole body pointing
at different MDs. An analysis of the loadings for each kinematic
angle on the first principal component also did not reveal any
significant main effects for movement duration (p.0.05, repeated
measures ANOVA) (figure not included).
3.3 Alterations in the elevation anglesThe results from the section above had established that despite
an overall similarity, small differences were present in the elevation
angle trajectories from whole body pointing carried out over
Figure 2. Elevation angle excursion. The kinematic trajectories of eight different elevation angles at three different movement durations for atypical subject. Beside each kinematic trace is the bar graph of the amplitudes recorded at long (Lo, black line and histogram), normal (N, grey lineand histogram) and short (Sh, dotted line and hatching histogram) MDs. The amplitude of each angular displacement was defined as the absolutevalue of the difference between the initial and final angle. Each bar displays the mean and the SEM for all the subjects. Significantly different valuesare marked with an arrow.doi:10.1371/journal.pone.0052477.g002
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 4 January 2013 | Volume 8 | Issue 1 | e52477
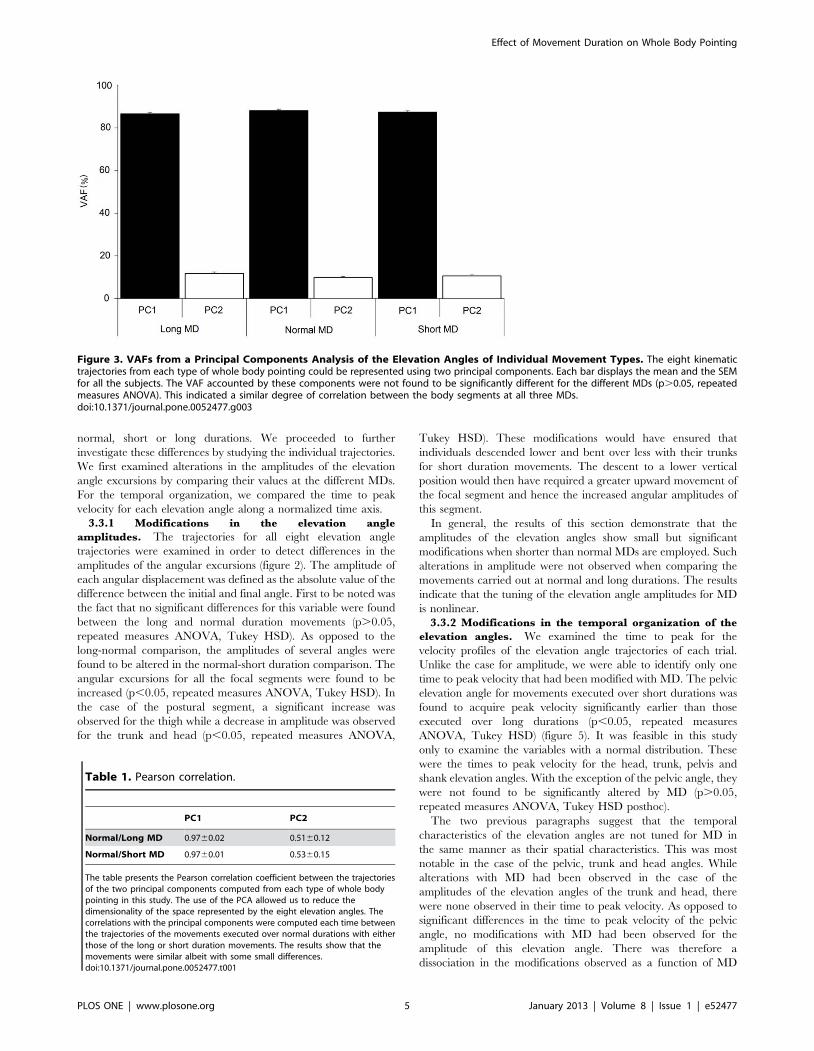
normal, short or long durations. We proceeded to further
investigate these differences by studying the individual trajectories.
We first examined alterations in the amplitudes of the elevation
angle excursions by comparing their values at the different MDs.
For the temporal organization, we compared the time to peak
velocity for each elevation angle along a normalized time axis.
3.3.1 Modifications in the elevation angle
amplitudes. The trajectories for all eight elevation angle
trajectories were examined in order to detect differences in the
amplitudes of the angular excursions (figure 2). The amplitude of
each angular displacement was defined as the absolute value of the
difference between the initial and final angle. First to be noted was
the fact that no significant differences for this variable were found
between the long and normal duration movements (p.0.05,
repeated measures ANOVA, Tukey HSD). As opposed to the
long-normal comparison, the amplitudes of several angles were
found to be altered in the normal-short duration comparison. The
angular excursions for all the focal segments were found to be
increased (p,0.05, repeated measures ANOVA, Tukey HSD). In
the case of the postural segment, a significant increase was
observed for the thigh while a decrease in amplitude was observed
for the trunk and head (p,0.05, repeated measures ANOVA,
Tukey HSD). These modifications would have ensured that
individuals descended lower and bent over less with their trunks
for short duration movements. The descent to a lower vertical
position would then have required a greater upward movement of
the focal segment and hence the increased angular amplitudes of
this segment.
In general, the results of this section demonstrate that the
amplitudes of the elevation angles show small but significant
modifications when shorter than normal MDs are employed. Such
alterations in amplitude were not observed when comparing the
movements carried out at normal and long durations. The results
indicate that the tuning of the elevation angle amplitudes for MD
is nonlinear.
3.3.2 Modifications in the temporal organization of the
elevation angles. We examined the time to peak for the
velocity profiles of the elevation angle trajectories of each trial.
Unlike the case for amplitude, we were able to identify only one
time to peak velocity that had been modified with MD. The pelvic
elevation angle for movements executed over short durations was
found to acquire peak velocity significantly earlier than those
executed over long durations (p,0.05, repeated measures
ANOVA, Tukey HSD) (figure 5). It was feasible in this study
only to examine the variables with a normal distribution. These
were the times to peak velocity for the head, trunk, pelvis and
shank elevation angles. With the exception of the pelvic angle, they
were not found to be significantly altered by MD (p.0.05,
repeated measures ANOVA, Tukey HSD posthoc).
The two previous paragraphs suggest that the temporal
characteristics of the elevation angles are not tuned for MD in
the same manner as their spatial characteristics. This was most
notable in the case of the pelvic, trunk and head angles. While
alterations with MD had been observed in the case of the
amplitudes of the elevation angles of the trunk and head, there
were none observed in their time to peak velocity. As opposed to
significant differences in the time to peak velocity of the pelvic
angle, no modifications with MD had been observed for the
amplitude of this elevation angle. There was therefore a
dissociation in the modifications observed as a function of MD
Figure 3. VAFs from a Principal Components Analysis of the Elevation Angles of Individual Movement Types. The eight kinematictrajectories from each type of whole body pointing could be represented using two principal components. Each bar displays the mean and the SEMfor all the subjects. The VAF accounted by these components were not found to be significantly different for the different MDs (p.0.05, repeatedmeasures ANOVA). This indicated a similar degree of correlation between the body segments at all three MDs.doi:10.1371/journal.pone.0052477.g003
Table 1. Pearson correlation.
PC1 PC2
Normal/Long MD 0.9760.02 0.5160.12
Normal/Short MD 0.9760.01 0.5360.15
The table presents the Pearson correlation coefficient between the trajectoriesof the two principal components computed from each type of whole bodypointing in this study. The use of the PCA allowed us to reduce thedimensionality of the space represented by the eight elevation angles. Thecorrelations with the principal components were computed each time betweenthe trajectories of the movements executed over normal durations with eitherthose of the long or short duration movements. The results show that themovements were similar albeit with some small differences.doi:10.1371/journal.pone.0052477.t001
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 5 January 2013 | Volume 8 | Issue 1 | e52477
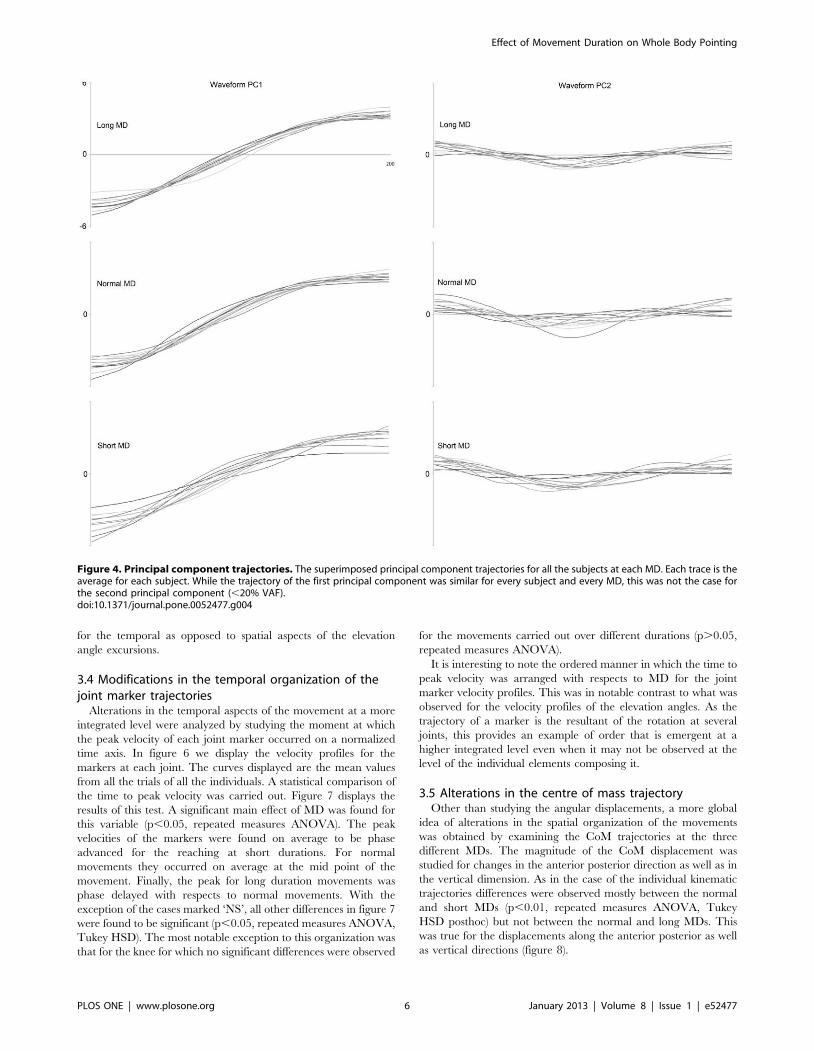
for the temporal as opposed to spatial aspects of the elevation
angle excursions.
3.4 Modifications in the temporal organization of thejoint marker trajectories
Alterations in the temporal aspects of the movement at a more
integrated level were analyzed by studying the moment at which
the peak velocity of each joint marker occurred on a normalized
time axis. In figure 6 we display the velocity profiles for the
markers at each joint. The curves displayed are the mean values
from all the trials of all the individuals. A statistical comparison of
the time to peak velocity was carried out. Figure 7 displays the
results of this test. A significant main effect of MD was found for
this variable (p,0.05, repeated measures ANOVA). The peak
velocities of the markers were found on average to be phase
advanced for the reaching at short durations. For normal
movements they occurred on average at the mid point of the
movement. Finally, the peak for long duration movements was
phase delayed with respects to normal movements. With the
exception of the cases marked ‘NS’, all other differences in figure 7
were found to be significant (p,0.05, repeated measures ANOVA,
Tukey HSD). The most notable exception to this organization was
that for the knee for which no significant differences were observed
for the movements carried out over different durations (p.0.05,
repeated measures ANOVA).
It is interesting to note the ordered manner in which the time to
peak velocity was arranged with respects to MD for the joint
marker velocity profiles. This was in notable contrast to what was
observed for the velocity profiles of the elevation angles. As the
trajectory of a marker is the resultant of the rotation at several
joints, this provides an example of order that is emergent at a
higher integrated level even when it may not be observed at the
level of the individual elements composing it.
3.5 Alterations in the centre of mass trajectoryOther than studying the angular displacements, a more global
idea of alterations in the spatial organization of the movements
was obtained by examining the CoM trajectories at the three
different MDs. The magnitude of the CoM displacement was
studied for changes in the anterior posterior direction as well as in
the vertical dimension. As in the case of the individual kinematic
trajectories differences were observed mostly between the normal
and short MDs (p,0.01, repeated measures ANOVA, Tukey
HSD posthoc) but not between the normal and long MDs. This
was true for the displacements along the anterior posterior as well
as vertical directions (figure 8).
Figure 4. Principal component trajectories. The superimposed principal component trajectories for all the subjects at each MD. Each trace is theaverage for each subject. While the trajectory of the first principal component was similar for every subject and every MD, this was not the case forthe second principal component (,20% VAF).doi:10.1371/journal.pone.0052477.g004
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 6 January 2013 | Volume 8 | Issue 1 | e52477
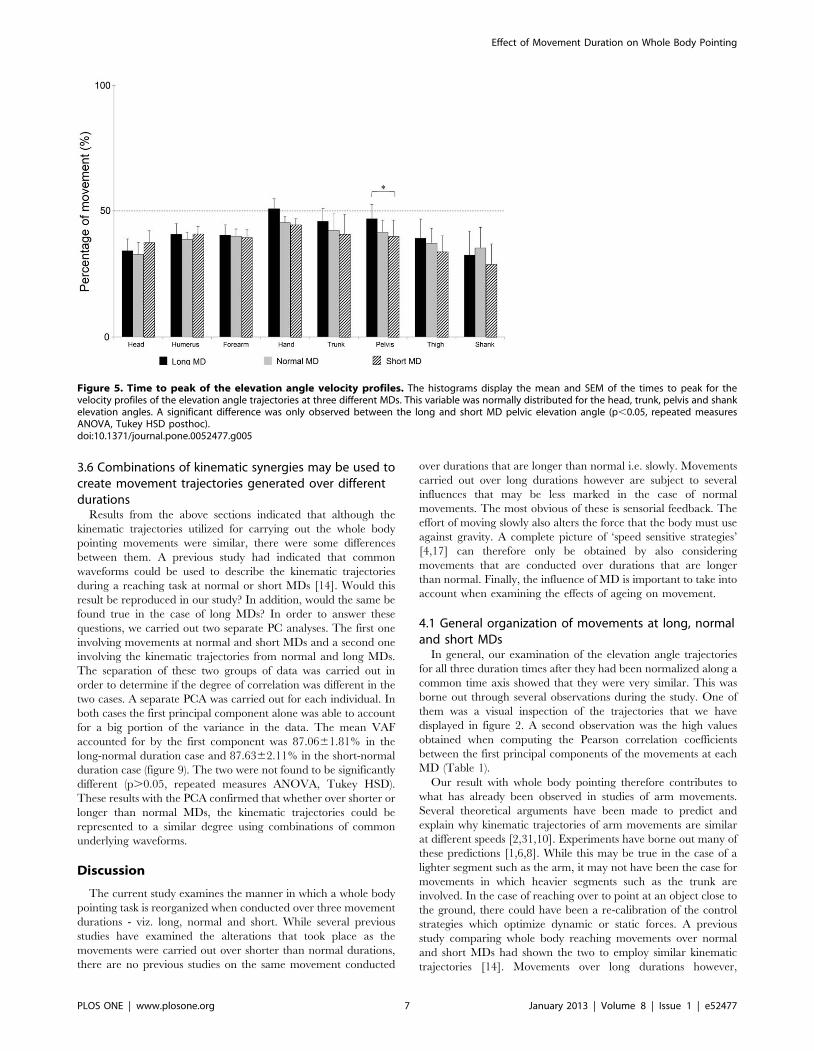
3.6 Combinations of kinematic synergies may be used tocreate movement trajectories generated over differentdurations
Results from the above sections indicated that although the
kinematic trajectories utilized for carrying out the whole body
pointing movements were similar, there were some differences
between them. A previous study had indicated that common
waveforms could be used to describe the kinematic trajectories
during a reaching task at normal or short MDs [14]. Would this
result be reproduced in our study? In addition, would the same be
found true in the case of long MDs? In order to answer these
questions, we carried out two separate PC analyses. The first one
involving movements at normal and short MDs and a second one
involving the kinematic trajectories from normal and long MDs.
The separation of these two groups of data was carried out in
order to determine if the degree of correlation was different in the
two cases. A separate PCA was carried out for each individual. In
both cases the first principal component alone was able to account
for a big portion of the variance in the data. The mean VAF
accounted for by the first component was 87.0661.81% in the
long-normal duration case and 87.6362.11% in the short-normal
duration case (figure 9). The two were not found to be significantly
different (p.0.05, repeated measures ANOVA, Tukey HSD).
These results with the PCA confirmed that whether over shorter or
longer than normal MDs, the kinematic trajectories could be
represented to a similar degree using combinations of common
underlying waveforms.
Discussion
The current study examines the manner in which a whole body
pointing task is reorganized when conducted over three movement
durations - viz. long, normal and short. While several previous
studies have examined the alterations that took place as the
movements were carried out over shorter than normal durations,
there are no previous studies on the same movement conducted
over durations that are longer than normal i.e. slowly. Movements
carried out over long durations however are subject to several
influences that may be less marked in the case of normal
movements. The most obvious of these is sensorial feedback. The
effort of moving slowly also alters the force that the body must use
against gravity. A complete picture of ‘speed sensitive strategies’
[4,17] can therefore only be obtained by also considering
movements that are conducted over durations that are longer
than normal. Finally, the influence of MD is important to take into
account when examining the effects of ageing on movement.
4.1 General organization of movements at long, normaland short MDs
In general, our examination of the elevation angle trajectories
for all three duration times after they had been normalized along a
common time axis showed that they were very similar. This was
borne out through several observations during the study. One of
them was a visual inspection of the trajectories that we have
displayed in figure 2. A second observation was the high values
obtained when computing the Pearson correlation coefficients
between the first principal components of the movements at each
MD (Table 1).
Our result with whole body pointing therefore contributes to
what has already been observed in studies of arm movements.
Several theoretical arguments have been made to predict and
explain why kinematic trajectories of arm movements are similar
at different speeds [2,31,10]. Experiments have borne out many of
these predictions [1,6,8]. While this may be true in the case of a
lighter segment such as the arm, it may not have been the case for
movements in which heavier segments such as the trunk are
involved. In the case of reaching over to point at an object close to
the ground, there could have been a re-calibration of the control
strategies which optimize dynamic or static forces. A previous
study comparing whole body reaching movements over normal
and short MDs had shown the two to employ similar kinematic
trajectories [14]. Movements over long durations however,
Figure 5. Time to peak of the elevation angle velocity profiles. The histograms display the mean and SEM of the times to peak for thevelocity profiles of the elevation angle trajectories at three different MDs. This variable was normally distributed for the head, trunk, pelvis and shankelevation angles. A significant difference was only observed between the long and short MD pelvic elevation angle (p,0.05, repeated measuresANOVA, Tukey HSD posthoc).doi:10.1371/journal.pone.0052477.g005
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 7 January 2013 | Volume 8 | Issue 1 | e52477
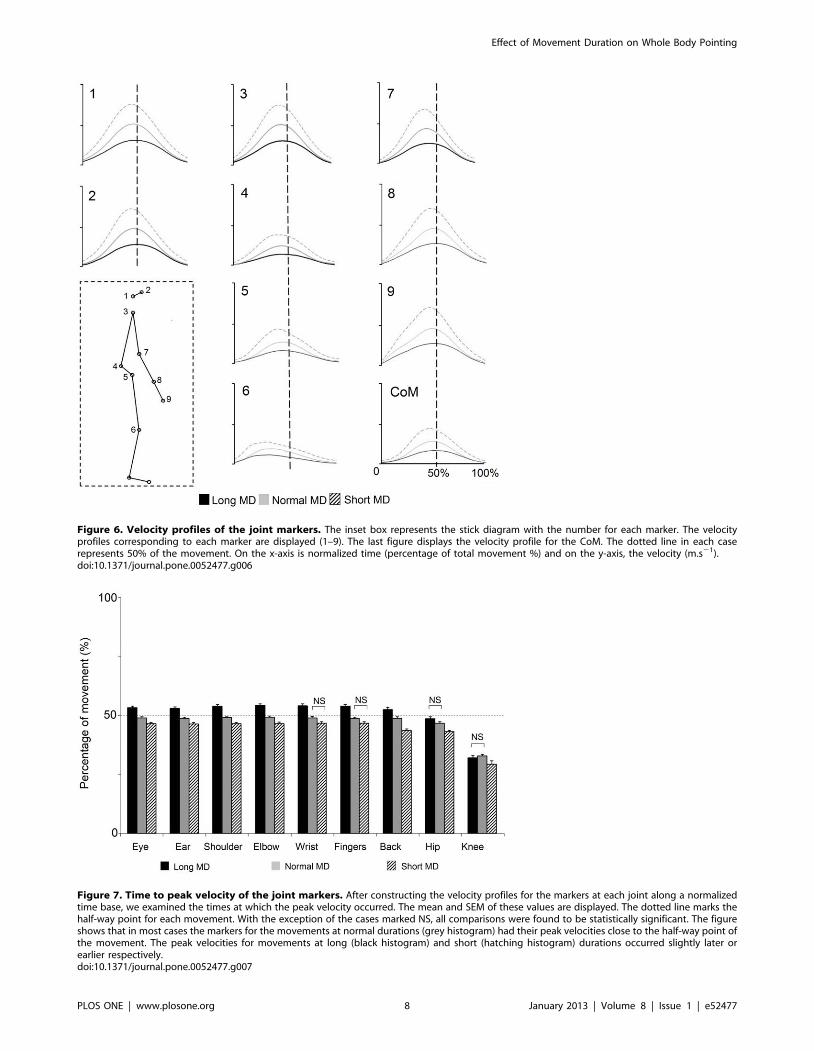
Figure 6. Velocity profiles of the joint markers. The inset box represents the stick diagram with the number for each marker. The velocityprofiles corresponding to each marker are displayed (1–9). The last figure displays the velocity profile for the CoM. The dotted line in each caserepresents 50% of the movement. On the x-axis is normalized time (percentage of total movement %) and on the y-axis, the velocity (m.s21).doi:10.1371/journal.pone.0052477.g006
Figure 7. Time to peak velocity of the joint markers. After constructing the velocity profiles for the markers at each joint along a normalizedtime base, we examined the times at which the peak velocity occurred. The mean and SEM of these values are displayed. The dotted line marks thehalf-way point for each movement. With the exception of the cases marked NS, all comparisons were found to be statistically significant. The figureshows that in most cases the markers for the movements at normal durations (grey histogram) had their peak velocities close to the half-way point ofthe movement. The peak velocities for movements at long (black histogram) and short (hatching histogram) durations occurred slightly later orearlier respectively.doi:10.1371/journal.pone.0052477.g007
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 8 January 2013 | Volume 8 | Issue 1 | e52477
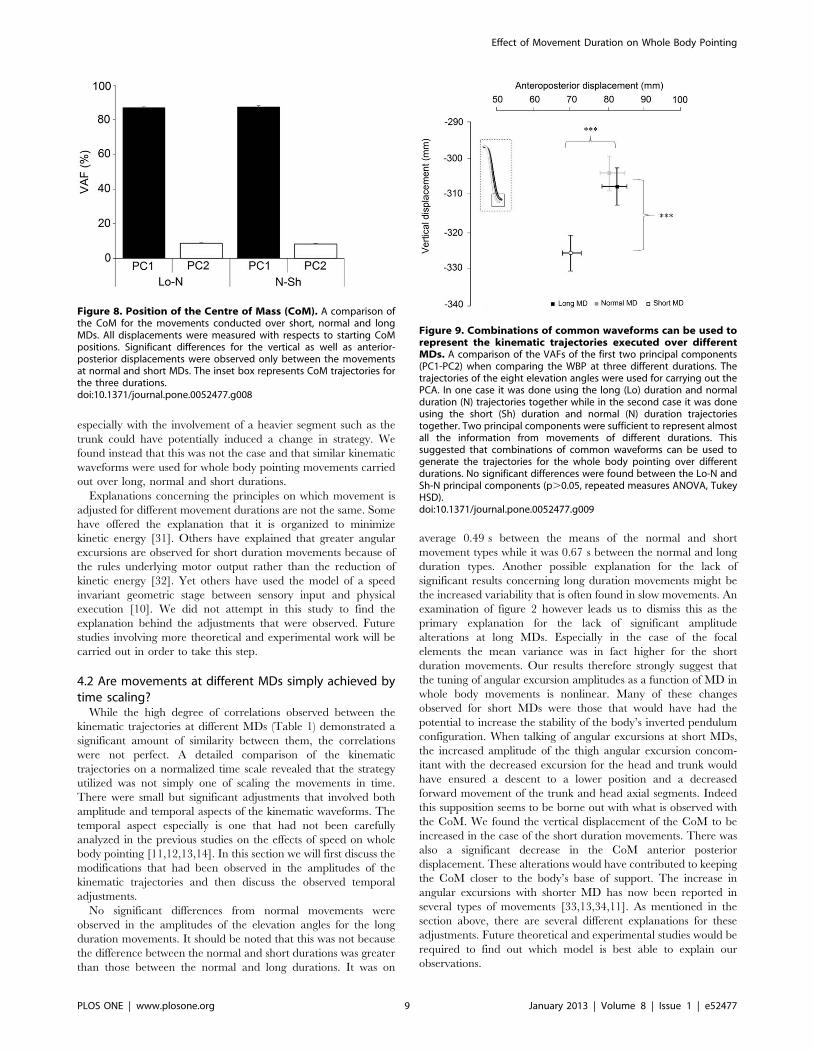
especially with the involvement of a heavier segment such as the
trunk could have potentially induced a change in strategy. We
found instead that this was not the case and that similar kinematic
waveforms were used for whole body pointing movements carried
out over long, normal and short durations.
Explanations concerning the principles on which movement is
adjusted for different movement durations are not the same. Some
have offered the explanation that it is organized to minimize
kinetic energy [31]. Others have explained that greater angular
excursions are observed for short duration movements because of
the rules underlying motor output rather than the reduction of
kinetic energy [32]. Yet others have used the model of a speed
invariant geometric stage between sensory input and physical
execution [10]. We did not attempt in this study to find the
explanation behind the adjustments that were observed. Future
studies involving more theoretical and experimental work will be
carried out in order to take this step.
4.2 Are movements at different MDs simply achieved bytime scaling?
While the high degree of correlations observed between the
kinematic trajectories at different MDs (Table 1) demonstrated a
significant amount of similarity between them, the correlations
were not perfect. A detailed comparison of the kinematic
trajectories on a normalized time scale revealed that the strategy
utilized was not simply one of scaling the movements in time.
There were small but significant adjustments that involved both
amplitude and temporal aspects of the kinematic waveforms. The
temporal aspect especially is one that had not been carefully
analyzed in the previous studies on the effects of speed on whole
body pointing [11,12,13,14]. In this section we will first discuss the
modifications that had been observed in the amplitudes of the
kinematic trajectories and then discuss the observed temporal
adjustments.
No significant differences from normal movements were
observed in the amplitudes of the elevation angles for the long
duration movements. It should be noted that this was not because
the difference between the normal and short durations was greater
than those between the normal and long durations. It was on
average 0.49 s between the means of the normal and short
movement types while it was 0.67 s between the normal and long
duration types. Another possible explanation for the lack of
significant results concerning long duration movements might be
the increased variability that is often found in slow movements. An
examination of figure 2 however leads us to dismiss this as the
primary explanation for the lack of significant amplitude
alterations at long MDs. Especially in the case of the focal
elements the mean variance was in fact higher for the short
duration movements. Our results therefore strongly suggest that
the tuning of angular excursion amplitudes as a function of MD in
whole body movements is nonlinear. Many of these changes
observed for short MDs were those that would have had the
potential to increase the stability of the body’s inverted pendulum
configuration. When talking of angular excursions at short MDs,
the increased amplitude of the thigh angular excursion concom-
itant with the decreased excursion for the head and trunk would
have ensured a descent to a lower position and a decreased
forward movement of the trunk and head axial segments. Indeed
this supposition seems to be borne out with what is observed with
the CoM. We found the vertical displacement of the CoM to be
increased in the case of the short duration movements. There was
also a significant decrease in the CoM anterior posterior
displacement. These alterations would have contributed to keeping
the CoM closer to the body’s base of support. The increase in
angular excursions with shorter MD has now been reported in
several types of movements [33,13,34,11]. As mentioned in the
section above, there are several different explanations for these
adjustments. Future theoretical and experimental studies would be
required to find out which model is best able to explain our
observations.
Figure 8. Position of the Centre of Mass (CoM). A comparison ofthe CoM for the movements conducted over short, normal and longMDs. All displacements were measured with respects to starting CoMpositions. Significant differences for the vertical as well as anterior-posterior displacements were observed only between the movementsat normal and short MDs. The inset box represents CoM trajectories forthe three durations.doi:10.1371/journal.pone.0052477.g008
Figure 9. Combinations of common waveforms can be used torepresent the kinematic trajectories executed over differentMDs. A comparison of the VAFs of the first two principal components(PC1-PC2) when comparing the WBP at three different durations. Thetrajectories of the eight elevation angles were used for carrying out thePCA. In one case it was done using the long (Lo) duration and normalduration (N) trajectories together while in the second case it was doneusing the short (Sh) duration and normal (N) duration trajectoriestogether. Two principal components were sufficient to represent almostall the information from movements of different durations. Thissuggested that combinations of common waveforms can be used togenerate the trajectories for the whole body pointing over differentdurations. No significant differences were found between the Lo-N andSh-N principal components (p.0.05, repeated measures ANOVA, TukeyHSD).doi:10.1371/journal.pone.0052477.g009
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 9 January 2013 | Volume 8 | Issue 1 | e52477

Amplitude was not the only feature that was altered as MD was
increased or decreased with respects to normal pointing. We also
noted the moment at which peak velocity occurred for each joint
marker and for each elevation angle. In the case of the elevation
angles, the time to peak velocity was only found to be significantly
different for the pelvic elevation angle. For this variable, we only
analyzed the elevation angles for which the time to peak velocity
was normally distributed. Because of the number of variables
involved in whole body pointing, posthoc pairwise comparisons in
the nonparametric cases would have led to a very low value of p
after the necessary Bonferroni corrections. Other features that
distinguished the time to peak velocities of the elevation angle
excursions (figure 5) from those of the joint marker trajectories
(figure 7), were their non parametric distributions in some
instances and finally the multidirectional nature of their modifi-
cations with MD. By this we mean that for some elevation angles,
a decreased MD led to the peak velocity time occurring earlier
than normal, while in other cases, the opposite was true.
The modifications observed in the spatial aspects of the
elevation angles were quite different from those observed in the
temporal domain. The specifics of this difference have already
been provided in the results section. This therefore provides an
example of dissociation between the temporal and amplitude
regulation for whole body pointing. A demonstration of such a
dissociation has already been made for arm pointing movements
[35,36]. Our study shows that coupling the arm movements to
postural control does not alter this aspect of motor control.
As opposed to changes observed with the elevation angles,
alterations in MD gave rise to very ordered shifts in the time to
peak velocity of the joint marker trajectories. We observed that
they had undergone phase shifts with respects to the trajectory at
normal duration. The word ‘phase’ is used here as all comparisons
are made using a normalised time axis. With the exception of the
knee, the peak of every marker had undergone a phase advance
for the short duration movements and a phase delay for the long
duration ones i.e. they occurred earlier than normal for the short
duration movements and later than normal for the long duration
ones. The utility of such phase shifts may best be understood by
noting that this means earlier transitions into the deceleration
phase as the MD is decreased. This would then mean a longer
period over which to break the movement when it is at a high
speed. Although they was speaking of electromyographic activity,
Gottlieb [4] had mentioned the use of such a ‘speed sensitive
strategy’ when they reported the earlier onset of antagonist muscle
activities for creating an earlier decelerating force in the case of
short duration or fast movements. These observations had been
made for arm movements. Our results suggest that a similar
strategy is also employed when the focal element is coupled to
postural displacements.
Although it was not the goal of this paper, one of the interesting
results in this study was the difference in the temporal re-
organization of the joint marker trajectories as opposed to those of
the angular excursions. As opposed to elevation angle excursions
which are created by a segment, the movements of joint markers
are the result of the angular excursions of several segments, not all
of which are displaced in the same direction. This sort of difference
between elevation angle excursions and joint marker trajectories is
likely to be more marked in the case of whole body movements
than for isolated arm movements. These individual changes at the
level of the joints then gave rise to the sort of changes described in
the paragraph above i.e. those that lead to earlier deceleration for
shorter MD movements.
4.3 High covariation between the body segments for allMDs
The degree of covariation between the segments was quantified
using a PCA. In all cases two principal components were sufficient
to capture practically all the variance in the data, hence indicating
a high degree of correlation between the angular displacements of
all the body segments. Despite significant differences in the
kinematic trajectories with MD, the coordination between the
segments remained similarly high for all the movements. This
suggested that modifications in any body segment were coordi-
nated with similar modifications in the remaining segments.
The VAF by the first principal component in our study is
slightly lower than what had been observed in the study of
Thomas et al [14] when analyzing fast reaching movements or by
Alexandrov et al [34] when looking at trunk bending movements
of various durations. The most likely reason for this might have
been the lower number of variables involved in the two cited
studies.
4.4 Combinations of a few kinematic synergies cangenerate movement trajectories at different MDs
A principal components analysis using kinematic trajectories of
different MDs showed that common waveforms could be used to
represent all the movements. We did not perform an analysis with
all three movement types at once. It was done instead with the
long-normal and short-normal kinematic trajectories separately.
This was done in order to probe if the inter-speed segmental
covariation might be different for these two groups. Two principal
components were sufficient to account for more than 95% of the
variance in the data in both cases. No significant differences were
found between the VAF by the first principal component in either
group. This was also the case for the second principal component
component. The ability to represent with two components, the
trajectories from different movement types, indicates that they
could be generated using combinations of a few common
waveforms.
4.5 Unintended modifications in movement durationsIt should be noted that movement duration is not always
something that is intentionally controlled. It has been found to
vary for the same target location despite the lack of any explicit
instructions or incentives to do so [36]. It can also change
unintentionally when the characteristics of the movement trajec-
tory change [37]. Fitt’s law [38] concerning the change of
movement speed with alteration in stimulus position or dimensions
is one of the most studied examples in motor control [33]. We
were unable in this study to make any claims concerning
unintentional alterations in movement duration as all subjects
had been given explicit instructions concerning this variable. It will
be interesting in the future to examine how movement duration
may be implicitly altered as different aspects of the whole body
pointing such as distance or directions are modified.
Author Contributions
Conceived and designed the experiments: MC TP ET. Performed the
experiments: MC. Analyzed the data: MC ET. Contributed reagents/
materials/analysis tools: MC ET. Wrote the paper: MC PM TP ET.
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 10 January 2013 | Volume 8 | Issue 1 | e52477

References
1. Soechting JF, Lacquaniti F (1981) Invariant characteristics of pointing
movement in man. J Neuroscience 7: 710–720.2. Hollerbach JM, Flash T (1982) Dynamic interaction between limb segments
during planar arm movements. Biol Cybern 44: 67–77.3. Atkeson CG, Hollerbach JM (1985) Kinematic features of unrestrained vertical
arm movements. J Neuroscience 5: 2318–2330.
4. Gottlieb GL, Corcos DM, Agarwal G (1989) Strategies for the control ofvoluntary movements with one mechanical degree of freedom. Behavioral and
Brain Sciences 12: 189–250.5. Gottlieb GL, Song Q, Hong DA, Almeida GL, Corcos D (1996) Coordinating
movement at two joints: a principal of linear covariance. J Neurophysiol 75:
1760–1764.6. Flanders M, Herrmann U (1992) Two components of muscle activation: scaling
with speed of arm movement. J Neurophysiol 67: 931–943.7. Buneo C, Soechting J, Flanders F (1994) Muscle Activation Patterns for
Reaching: The Representation of Distance and Time. Neurophysiology 71:1546–1158.
8. Nishikawa KC, Murray ST, Flanders M (1999) Journal of Neurophysiology 81:
2582–2586.9. Gaveau J, Papaxanthis(2011) The temporal structure of vertical arm movement.
Plos One 6 : 1–8.10. Torres EB, Zipser D (2002) Reaching to grasp with a multi-joint arm I
Computational Model. J Neurophysiol 88:2355–2367.
11. Stapley PJ, Pozzo T, Cheron G, Grishing A (1999) Does the coordinationbetween posture and movement during whole-body reaching ensure centre of
mass stabilization? Exp Brain Res 129: 134–146.12. Pozzo T, Stapley PJ, Papaxanthis C (2002) Coordination between equilibrium
and hand trajectories during whole body pointing movements. Exp Brain Res144: 343–350.
13. Thomas JS, Corcos DM, Hasan Z (2003) Effect of movement speed on limb
segment motions for reaching from a standing position. Exp Brain Res 148: 377–387.
14. Thomas JS, Corcos D, Hasan Z (2005) Kinematic and kinetic constraints onarm, trunk and leg segments in target-reaching movements. J Neurophysiol 93:
352–364.
15. Brooks VB (1986) The neural basis of motor control. Oxford University Press.16. Schmidt RA, Lee TD (2005) Motor Control and Learning. Human Kinetics,
Champaign, IL.17. Torres EB (2011) Two classes of movements in motor control. Exp Brain Res
215: 269–83.18. Tinetti ME, Speechley M, Ginter SF (1988) Risk factors falls among elderly
persons living in the community. N Engl J Med 319: 1701–1707.
19. Boulgarides LK, McGinty SM, Willett JA, Barnes CW (2003). Use of clinicaland impairment-based tests to predict falls by community-dwelling older adults.
Phys Ther 83: 328–339.
20. Houles M, Abellan van Kan G, Rolland Y, Andrieu S, Anthony P, et al. (2010)
La vitesse de marche comme critere de fragilite chez la personne agee vivant au
domicile. Cah Annee Gerontol 2: 13–23.
21. Paizis C, Papaxanthis C, Berret B, Pozzo T (2008) Reaching beyond arm length
in normal ageing: Adaptation of hand trajectory and dynamic equilibrium.
Behavioral Neuroscience 122: 1361–1370.
22. Bernstein N (1967) The coordination and regulation of movements. Oxford:
Pergamon.
23. Schmid M, Schieppati M, Pozzo T (2006) Effect of fatigue on the precision of a
whole-body. Neuroscience 139: 909–920.
24. Berret B, Bonnetblanc F, Papaxanthis C, Pozzo T (2009). Modular control of
pointing beyond arm’s length. J Neurosci 29: 191–205.
25. Chiovetto E, Berret B, Pozzo T (2010) Tri-dimensional and triphasic muscle
organization of whole-body pointing movements. Neuroscience 170: 1223–1238.
26. Sergio LE, Ostry DJ (1994) Coordination of mono- and bi-articular muscles in
multi-degree of freedom elbow movements. Experimental Brain Research 9:
551–555.
27. Papaxanthis C, Pozzo T, Intyre JMC (2005) Kinematic and dynamic processes
for the control of pointing movements in humans revealed by short-term
exposure to microgravity. Neuroscience 135: 371–383.
28. Jolliffe I (1986) Principal components analysis. New York: Springer.
29. Winter D (1990) Biomechanics and motor control of human movement. New
York: John Wiley and Sons.
30. Salton G (1989) Automatic text processing. Addison Welsley, New York
31. Soechting JF, Buneo CA, Herrmann U, Flanders M (1995) Moving effortlessly in
three dimensions: Does Donder’s law apply to arm movement? J Neurosci.
15:6271–80.
32. Corcos DM, Gottlieb GL, Agarwal GL (1989) Organizing principles for single
joint movements II A speed sensitive strategy. J Neurophysiol 62: 358–368.
33. Schillings JJ, Meulenbroek RGJ, Thomassen AJWM (1996) Limb segment
recruitment as a function of movement direction, amplitude and speed. J Mot
Behav 28: 241–254.
34. Alexandrov A, Frolov A, Massion J (1998) Axial synergies during human upper
trunk bending. Experimental Brain Research 118(2):210–20.
35. Biess A, Liebermann DG, Flash T (2007) A computational model for redundant
human three-dimensional pointing movements: integration of independent
spatial and temporal motor plans simplifies movement dynamics. J Neuroscience
27: 13045–64.
36. Torres EB (2010) New symmetry of intended curved reaches. Behavioral and
Brain Functions 6:21.
37. Vivian P, Terzuolo C (1982) Trajectory determines movement dynamics.
Neuroscience 7:431–7.
38. Fitts P (1954) The information capacity of the human motor system in
controlling the amplitude of movement. J Exp Psychol 47: 381–31.
Effect of Movement Duration on Whole Body Pointing
PLOS ONE | www.plosone.org 11 January 2013 | Volume 8 | Issue 1 | e52477

Annexe 6
Résumés publiés



Annexe 7
Communications

Communications
Affichées - 9th Congress Progress in Motor Control (PMC IX), « An Analysis of the Centre of Mass trajectories during a Whole Body Pointing Movement», 13-16 Juillet 2013, Montréal-Canada. - 19ème Congrès de l’Association Posture-Equilibre (APE) et 1er Congrès de la Société Francophone Posture, Equilibre, Locomotion (SOFPEL), « Contrôle du centre de masse lors de pointage de tout le corps chez le sujet jeune et âgé sain », 30 Novembre et 1Décembre 2012, Marseille-France. - Association des Chercheurs en Activité Physique et Sportive - ACAPS, « Indépendance spatiale et temporelle de l'organisation synergique d'un mouvement de tout le corps chez le sujet jeune », 24-26 Octobre 2011 Rennes-France. - 17ème Forum des Jeunes Chercheurs, « Effet de la vitesse sur le contrôle des mouvements de pointage de tout le corps chez le sujet jeune », 16-17juin 2011 Dijon-France. Prix Communication Poster - Institut Fédératif de Recherche, Santé - STIC (I.F.R. 100), « Effet de la vitesse sur le contrôle des mouvements de pointage de tout le corps chez le sujet jeune », 13 Avril 2011 Dijon-France Orales - 19ème Forum des Jeunes Chercheurs, « An Analysis of the Centre of Mass Trajectories during a Whole Body Pointing Movement », 13-14 Juin 2013 Dijon-France. Prix Communication Orale - 19ème Congrès de l’Association Posture-Equilibre (APE) et 1er Congrès de la Société Francophone Posture, Equilibre, Locomotion (SOFPEL), « Impact d'une tâche cognitive sur la posture du sujet âgé dépressif comparé au sujet âgé non-dépressif », 30 Novembre et 1Décembre 2012, Marseille-France. - 18ème Forum des Jeunes Chercheurs, « Modifications cinématiques d'un mouvement de pointage de tout le corps lors de différents temps de mouvement », 6-7 Septembre 2012 Besançon-France. - 28ème Colloque du Club Locomotion et Motricité Rythmique, « Effet de la vitesse sur le contrôle des mouvements de pointage de tout le corps chez le sujet jeune », 22-23Septembre 2011, Marseille-France.
Conférence « Université Pour Tous », conférence à caractère général sur les neurosciences, et plus en particulier sur les outils récents d’expérimentation (IRMf, EEG, TEP, TITD, etc.), qui sont utilisés dans les analyses du comportement humain. 4 Avril 2013, Chalon-sur-Saône, France. http://www.utb-chalon.org/

Annexe 8
Conférence

5 bis, Avenue Nicéphore Niepce – 71100 Chalon-sur-S aône – Tel/fax : 03.85.93.41.70. E-mail : [email protected] Secrétariat : mardi et jeudi 9 h 12 h – 14 h 17 h – vendredi 9 h 12 h
U N I V E R S I T É D E B O U R G O G N E
UNIVERSITÉ POUR TOUS Centre de Chalon-sur-Saône
ATTESTATION D’INTERVENTION A L’UNIVERSITE POUR TOUS DE BOURGOGNE
Antenne de Chalon-sur-Saône
Je soussigné Yves FOURNIER, président de l’Université pour Tous de Bourgogne à Chalon-sur-Saône, certifie que Monsieur Matthieu CASTERAN a présenté devant les adhérents de l’UTB une conférence intitulée : “l’Imagerie médicale fonctionnelle” le jeudi 4 avril 2013 à Chalon-sur-Saône.
Fait à Chalon-sur-Saône, le 5 décembre 2013
Le Président, Y. FOURNIER

Annexe 9
Congrès organisés

Organisation de Congrès
1) Forum des Jeunes Chercheurs - Membre du comité d’organisation du 17ème Forum des Jeunes chercheurs 2011 https://sites.google.com/site/fjc2011dijon/home - Président du comité d’organisation du 19ème Forum des Jeunes Chercheurs 2013 https://sites.google.com/site/fjc2013dijon/home
« Le Forum des Jeunes Chercheurs est une manifestation scientifique annuelle qui se déroule sur deux jours et permet aux doctorants de l’Ecole Doctorale "Environnements - Santé" de Dijon-Besançon de présenter leurs travaux de thèse lors de sessions de communications scientifiques orales ou affichées. Se tenant à tour de rôle à Dijon et Besançon, cet évènement est organisé par les doctorants de l'ED ES de Dijon en partenariat avec l'association EDIFICE. Au cours de ce forum, des conférences scientifiques, des ateliers, ainsi que des interventions de professionnels sont organisés afin de permettre aux doctorants d’obtenir des éléments leur permettant de mieux appréhender « l’après-doctorat » et le marché du travail en les aidant à élaborer leur projet professionnel. » www.edifice-dijon.com 2) Forum des BIOTechno (Association Nationale des BIOTechno) - Membre du comité d’organisation des 14ème Journées Nationales des BIOTechnologies 2011 - Président du comité d’organisation des 16ème Journées Nationales des BIOTechnologies 2013 www.biotechno.eu
« Cette association de loi 1901 a pour volonté de valoriser la formation des docteurs et d'œuvrer à l'amélioration de l'insertion des jeunes diplômés qui peuvent devenir créateurs de savoir (recherche), diffuseurs de savoir (enseignement) ou fournisseurs d'innovations (création d'entreprise). Aujourd'hui, le Réseau BIOTechno est le premier interlocuteur des ministères, entreprises et organismes publics de recherche et joue un rôle essentiel dans le développement du secteur des biotechnologies en France. Depuis maintenant 17 ans, nous organisons les Journées BIOTechno dans plusieurs grandes villes de France, et proposons aux jeunes chercheurs de découvrir les métiers des biotechnologies et les acteurs professionnels de ce secteur. Le nombre croissant de participants (étudiants, sociétés de biotechnologie, les structures publiques de recherche et structure de valorisation) montre l'intérêt de l'organisation de telles journées et leur reconduction. Des conférences, des tables rondes et des ateliers sont organisé autours de partenaire industriel. »
3) CEN StimCo Membre du comité d’organisation du 1er Congrès Européen de Stimulation Cognitive – StimCo 2012 www.censtimco.org
« Organisé sous l’égide du Centre d’Expertise National en Stimulation Cognitive (CEN STIMCO), ce premier colloque Européen aura pour objectifs d’étudier les enjeux, les difficultés et les apports possibles de la stimulation cognitive. A visée pluridisciplinaire, il est organisé par le laboratoire Inserm Cognition, Action, et Plasticité Sensorimotrice U1093, au sein de la Faculté des Sciences du Sport de Dijon (Université de Bourgogne). Durant deux jours et demi, des associations et collectivités publiques, des prescripteurs, des usagers, des chercheurs, des industriels associeront leurs efforts pour mieux comprendre les fondements, les méthodes et les pratiques de la stimulation cognitive. Les débats, sous formes de communications orales et affichées, de tables rondes et d’ateliers de démonstration, porteront également sur les besoins des populations. Des perspectives de développement éclairées par les aspects éthiques et règlementaires seront finalement proposées. »

Annexe 10
Enseignements
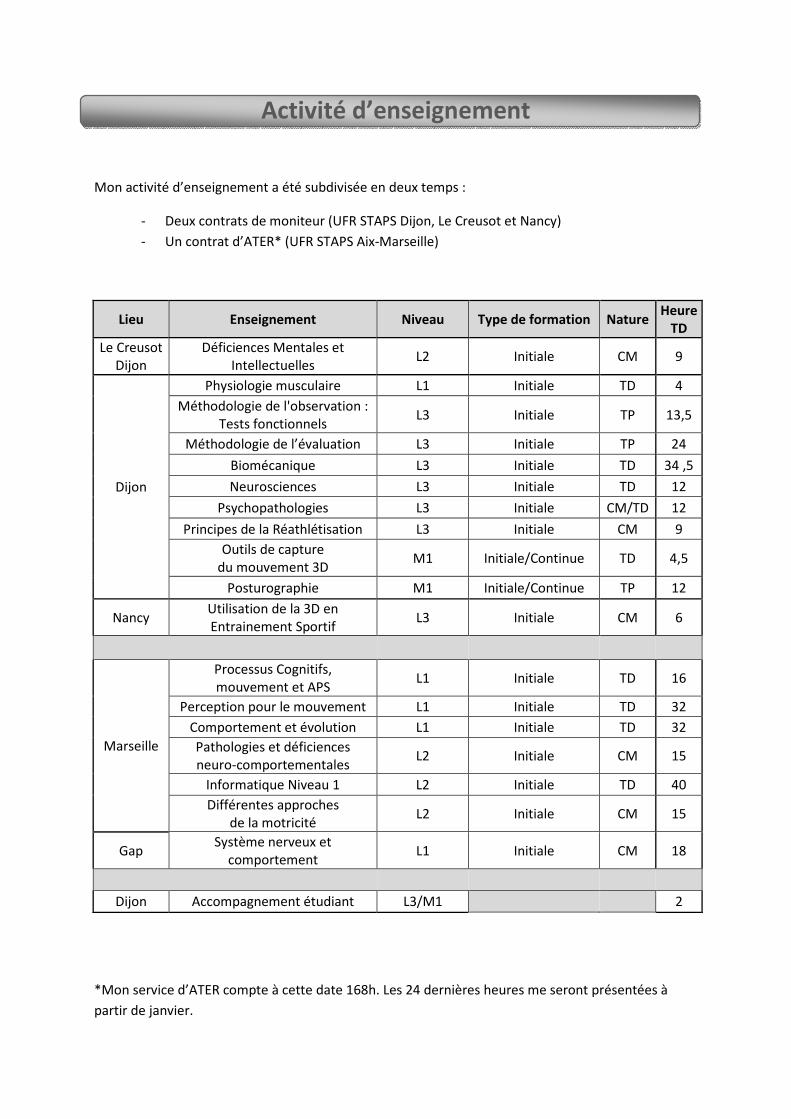
Activité d’enseignement
Mon activité d’enseignement a été subdivisée en deux temps :
- Deux contrats de moniteur (UFR STAPS Dijon, Le Creusot et Nancy)
- Un contrat d’ATER* (UFR STAPS Aix-Marseille)
Lieu Enseignement Niveau Type de formation Nature Heure
TD
Le Creusot
Dijon
Déficiences Mentales et
Intellectuelles L2 Initiale CM 9
Dijon
Physiologie musculaire L1 Initiale TD 4
Méthodologie de l'observation :
Tests fonctionnels L3 Initiale TP 13,5
Méthodologie de l’évaluation L3 Initiale TP 24
Biomécanique L3 Initiale TD 34 ,5
Neurosciences L3 Initiale TD 12
Psychopathologies L3 Initiale CM/TD 12
Principes de la Réathlétisation L3 Initiale CM 9
Outils de capture
du mouvement 3D M1 Initiale/Continue TD 4,5
Posturographie M1 Initiale/Continue TP 12
Nancy Utilisation de la 3D en
Entrainement Sportif L3 Initiale CM 6
Marseille
Processus Cognitifs,
mouvement et APS L1 Initiale TD 16
Perception pour le mouvement L1 Initiale TD 32
Comportement et évolution L1 Initiale TD 32
Pathologies et déficiences
neuro-comportementales L2 Initiale CM 15
Informatique Niveau 1 L2 Initiale TD 40
Différentes approches
de la motricité L2 Initiale CM 15
Gap Système nerveux et
comportement L1 Initiale CM 18
Dijon Accompagnement étudiant L3/M1
2
*Mon service d’ATER compte à cette date 168h. Les 24 dernières heures me seront présentées à
partir de janvier.
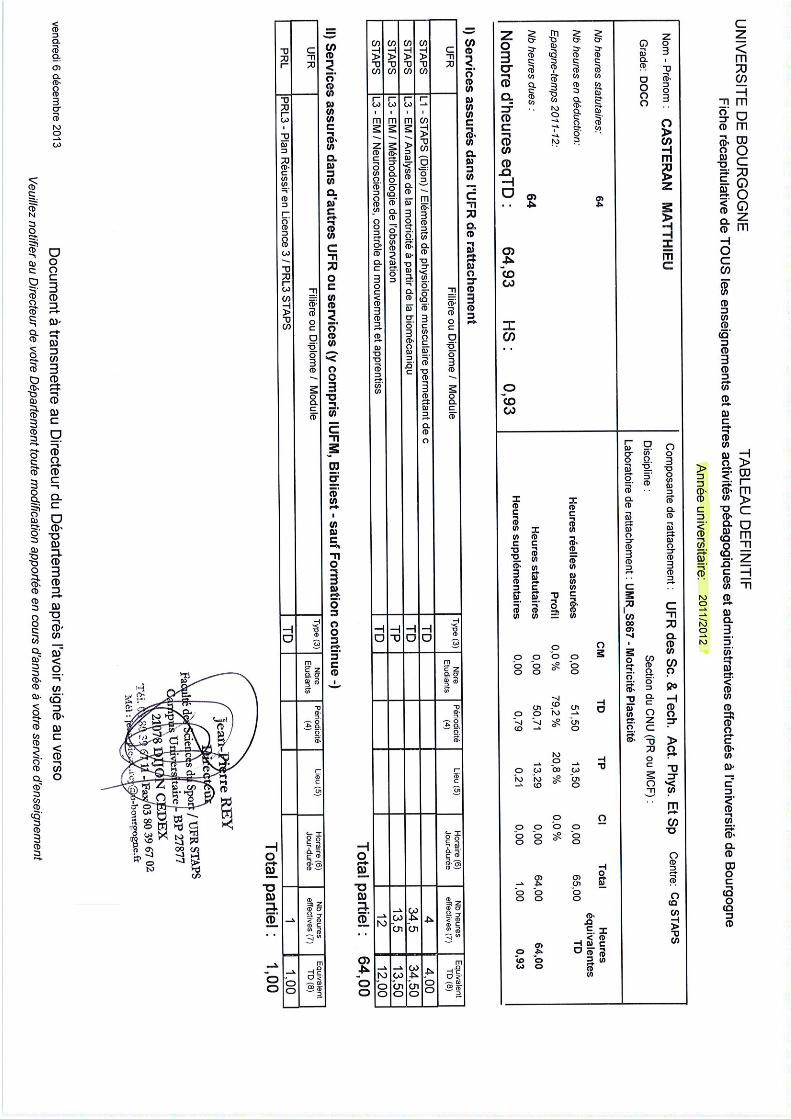


Nom :
Prénom :
Statut :
Si APSA, préciser l'APSA
concernéeh TP par
groupe
Nbre de
groupes
h TD par
groupe
Nbre
groupes
TD
h CM
Nbre
groupes
CM
h TP par
groupe
Nbre
groupes
h TD par
groupe
Nbre
groupes
TD
h CM Eq. TD
S1 8 2 ð 16
S1 1 12 ð 18
S2 8 4 ð 32
S2 8 4 ð 32
S4 10 1 ð 15
S4 10 1 ð 15
L2MSS4U7 - FLOUT108 S4 20 2 ð 40
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
Si APSA, préciser l'APSA
concernéeh TP par
groupe
Nbre de
groupes
h TD par
groupe
Nbre
groupes
TD
h CM Eq. TD
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
DIPLÔME ET LIEU DE
FORMATIONh TP par
groupe
Nbre
groupes
h TD par
groupe
Nbre
groupes
TD
h CM Eq. TD
ð 0
ð 0
ð 0
ð 0
ð 0
ð 0
TOTAL 168
à Marseille, le
192
-24
Code APOGEE - CODE ETAPE - TITRE DU COURS
Charge de service théorique (hTD)
Différence
ECOLE DOCTORALE - DU - AUTRES ENSEIGNEMENTS HORAIRES
Code APOGEE - CODE ETAPE - TITRE DU COURS
MASTER STAPS
FLCOM108 - FPS2T0 - Différentes approches de la motricité
FLCOM109 - FPS2T0 - Pathologies et déficiences neuro-comportementales
FLOUT108 - FPS2T0 - Informatique, Bureautique niveau 1
FICHE 1
CASTERAN
Matthieu
FLCOM102 - FPS1T0 - Perception pour le mouvement
ATER
MARSEILLELICENCE STAPS
Les cellules vertes doivent êtres renseignées par l'enseignant
Les cellules bleues sont des cellules calculées automatiquement et protégées (donc non modifiables)
FLCOM101 - FPS1T0 - Processus cognitifs, mouvement et APS
FLCOM100 - FPS1T0 - Système nerveux et comportement dans les APS
FLCOM103 - FPS1T0 - Comportement et évolution
CALCUL DES CHARGES D'ENSEIGNEMENT (2013/2014)
Code APOGEE - CODE ETAPE - TITRE DU COURS
GAP
MARSEILLE
Signature de l'enseignant
17 Décembre 2013

Annexe 11
Implications Scientifiques


Secrétariats : Université de Bourgogne, Bâtiment Gabriel, 6 Bd Gabriel, 21000 Dijon.
Tél. : 03 80 39 38 68 (corinne.aquilina@u‐bourgogne.fr) ou 03 80 39 38 60 (josette.thery@u‐bourgogne.fr).
Université de Franche‐Comté, UFR Sciences Médicales et Pharmaceutiques, 19 Rue Ambroise Paré, 25030 Besançon cedex. Tél. : 03 63 08 22 13 (martine.gautheron@univ‐fcomte.fr).
Dijon, le 05/12/2013
Pour faire valoir à qui de droit Madame, Monsieur, Je soussigné, Thierry Rigaud, Directeur de Recherche au CNRS et Directeur de l’Ecole Doctorale Environnements – Santé des Universités de Bourgogne et de Franche‐Comté, certifie que Matthieu Casteran a été représentant élu des doctorants dans le conseil de l’Ecole Doctorale pendant la totalité de sa durée de thèse. A ce titre il a participé aux diverses activités listées ci‐dessous : ‐ Participant à l’organisation du Forum des Jeunes Chercheurs de l’ED, en 2011 ‐ Président du comité d’organisation du Forum des Jeunes Chercheurs de l’ED, en 2013 ‐ Participant, en tant qu’assesseur, à trois jurys de concours de l’école doctorale (jurys de sélection des doctorants) ‐ Co‐responsable, avec un membre Enseignant‐Chercheur du conseil de l’ED, de la sélection des dossiers de « bourses de mobilité » (bourses attribuées par l’ED apportant aux doctorants un soutien financier pour participer à des manifestations scientifiques) Matthieu Casteran s’est acquitté de ces diverses responsabilités avec un sérieux, une efficacité et une assiduité remarquable, faisant de lui un des référents principaux de la représentation doctorale au sein de l’ED. Cordialement Thierry Rigaud





























