Embed Size (px)
Citation preview

Développement d’une méthode analytique pour la
spéciation du sélénium dans le plasma des Inuits du
Nunavik
Mémoire
Adel Achouba
Maîtrise en médecine expérimentale
Maître ès sciences (M.Sc.)
Québec, Canada
© Adel Achouba, 2015


iii
Résumé
Le sélénium (Se) est très abondant dans l’alimentation marine traditionnelle des Inuits et
par conséquent leur apport en Se est parmi les plus élevés au monde. Cette alimentation
marine est une source importante de méthylmercure (MeHg) et un apport élevé en Se
pourrait contrer certains de ses effets néfastes. Dans cette étude, une méthode de spéciation
et de quantification du Se plasmatique et des fractions du Hg associées a été développée
puis appliquée à des échantillons de plasma d’adultes inuits ayant participé à l’enquête de
santé Qanuippitaa-2004. La glutathion peroxydase 3 (GPx3), la sélénoprotéine P (SelP) et
la sélénoalbumine (SeAlb) représentaient en moyenne 25%, 52% et 23% du Se plasmatique
total respectivement. En outre, l’Hg coéluait avec chacune de ces protéines dont 50% était
associé à la SelP. La relation non linéaire observée entre les concentrations plasmatiques et
sanguines de Se suggère qu’un sélénocomposé, possiblement la sélénonéine, s’accumule
dans les cellules sanguines.


v
Abstract
Selenium (Se) is highly abundant in traditional marine foods consumed by Inuit and
accordingly their Se intake is one of the highest in the world. These marine foods are also a
replete of methylmercury (MeHg) and an elevated Se intake may offset some of its
deleterious effects. In this study, a method for the speciation and quantification of Se and
associated Hg fractions in human plasma was developed and then applied to plasma
samples of Inuit adults who participated to the 2004 Qanuippitaa health survey.
Glutathione peroxidase 3 (GPx3), selenoprotein P (SelP) and selenoalbumin (SeAlb)
represented on average 25%, 52% and 23% of the total plasma Se concentration,
respectively. In addition, Hg co-eluted with each Se-containing protein and 50% of plasma
Hg was associated with SelP. A non-linear relationship was observed between plasma and
blood Se, suggesting that a selenocompound, possibly selenoneine, accumulates in blood
cells of our participants.


vii
Table des matières
Résumé ............................................................................................................................................................... iii
Abstract ................................................................................................................................................................ v
Table des matières ......................................................................................................................................... vii
Liste des tableaux ............................................................................................................................................ xi
Listes des figures .......................................................................................................................................... xiii
Abréviations ..................................................................................................................................................... xv
Remerciements ............................................................................................................................................. xvii
Avant-propos .................................................................................................................................................. xix
Chapitre I : Introduction ................................................................................................................................. 1
1 Historique .................................................................................................................................................. 1
2 Propriétés physico-chimiques ............................................................................................................. 2
3 Présence dans l’environnement ......................................................................................................... 3
3.1 L’atmosphère ................................................................................................................................... 3
3.2 L’eau ................................................................................................................................................... 4
3.3 Le sol .................................................................................................................................................. 4
4 Sélénium dans l’organisme humain ................................................................................................. 6
4.1 Les composés organiques séléniés............................................................................................ 6
4.1.1 Les acides aminés séléniés ................................................................................................. 6
4.1.2 La sélénonéine ........................................................................................................................ 9
4.2 Sélénite et séléniate ........................................................................................................................ 9
4.3 Les sélénoprotéines ........................................................................................................................ 9
4.3.1 La famille des glutathion peroxydases (GPx) .............................................................. 9
4.3.2 La sélénoprotéine P (SelP) .............................................................................................. 12
4.3.3 La famille des iodothyronines désiodases (DIOs) ................................................... 13
4.3.4 La famille des thiorédoxines réductases (TrxR) ...................................................... 14
4.3.5 La méthionine R-sulfoxyde réductase 1 (MsrB1) ................................................... 16
4.3.6 La sélénoprotéine N (SelN) ............................................................................................. 16
4.3.7 La sélénoprotéine O (SelO) ............................................................................................. 17
4.3.8 La sélénoprotéine I (SelI) ................................................................................................ 17

viii
4.3.9 La sélénoprotéine W (SelW) ........................................................................................... 18
4.3.10 La sélénoprotéine H (SelH) ............................................................................................. 18
4.3.11 La sélénoprotéine M (SelM) ........................................................................................... 18
4.3.12 La sélénoprotéine de 15KDa (Sep15) .......................................................................... 19
4.3.13 Les sélénoprotéines K et S (SelK, SelS) ..................................................................... 19
4.3.14 La sélénoprotéine T (SelT) .............................................................................................. 20
4.3.15 La sélénoprotéine V (SelV) ............................................................................................. 20
4.3.16 La sélénophosphate synthétase 2 (SPS2) .................................................................... 20
5 Toxicocinétique du sélénium ........................................................................................................... 20
5.1 Absorption ...................................................................................................................................... 20
5.2 Distribution .................................................................................................................................... 21
5.3 Biotransformation ........................................................................................................................ 21
5.4 Élimination ..................................................................................................................................... 23
6 Sélénium et santé.................................................................................................................................. 23
6.1 Les maladies cardiovasculaires ............................................................................................... 23
6.2 Le diabète de type 2 .................................................................................................................... 24
6.3 Le système immunitaire ............................................................................................................. 25
6.4 Les maladies infectieuses virales ............................................................................................ 25
6.5 Le cancer ......................................................................................................................................... 25
7 Interaction entre le sélénium et le méthylmercure (Se-MeHg) ............................................. 26
7.1 Diminution de la toxicité par formation de composés MeHg-Se(H) .......................... 27
7.2 Le sélénium joue un rôle dans la biotransformation du MeHg .................................... 28
7.3 Le sélénium inhibe le produit de la dissociation du MeHg............................................ 29
8 Population d’étude ............................................................................................................................... 29
8.1 Les Inuits du Nunavik ................................................................................................................ 29
8.2 Alimentation traditionnelle des Inuits du Nunavik ........................................................... 30
8.3 Recrutement et échantillonnage .............................................................................................. 31
9 Objectifs .................................................................................................................................................. 32
10 Démarche expérimentale ................................................................................................................... 32
10.1 Spéciation du sélénium plasmatique ................................................................................. 33
10.2 Quantification du sélénium et du mercure ...................................................................... 33

ix
Chapitre 2: Article scientifique ............................................................................................................ 37
Circulating levels of selenium species in Inuit adults from Nunavik ..................................... 37
Chapitre 3 : Conclusion générale ........................................................................................................ 55
Références: ................................................................................................................................................. 57


xi
Liste des tableaux
Tableau 1 : L’abondance relative naturelle des isotopes du sélénium. ................................. 2
Tableau 2 : Répartition selon le groupe d'âge et âge médian des Inuits du Nunavik (année
2011) ..................................................................................................................................... 30
Table 3 : Operating conditions for the liquid chromatography (LC) and ICP-MS
instruments ............................................................................................................................ 52
Table 4 : Analytical performance of the ID-AF-ICP-MS method ....................................... 53
Table 5 : Concentrations of total blood Se, total plasma Se and plasma Se proteins in Inuit
of Nunavik (n=852) .............................................................................................................. 54
Table 6 : Concentrations of total blood Hg and plasma Hg associated to selenoproteins in
Inuit of Nunavik (n=845) ...................................................................................................... 54


xiii
Listes des figures
Figure 1 : Cycle biogéochimique du sélénium. ..................................................................... 5
Figure 2 : Structures chimiques de la sélénocystéine et la cystéine. ..................................... 6
Figure 3 : Les étapes de biosynthèse de la Sec-ARNt(Ser)Sec (A) et le mécanisme d’insertion
de la sélénocystéine dans les sélénoprotéines (B) chez les eucaryotes ................................... 7
Figure 4 : Structures chimiques de la sélénométhionine et la méthionine. ........................... 8
Figure 5 : Structure chimique de la sélénonéine. ................................................................... 9
Figure 6 : Mécanisme d’action des GPx selon Papp et al .................................................... 12
Figure 7 : Mécanisme d’action du système TRx selon Lu et al .......................................... 16
Figure 8 : La biotransformation du sélénium selon Alexander et al .................................... 22
Figure 9 : Carte géographique du Nunavik .......................................................................... 29
Figure 10 : Schematic diagram of the ID-AF-ICP-MS method for Se-containing proteins
separation and quantification. ............................................................................................... 50
Figure 11 : Typical mass flow chromatogram obtained by AF-ICP-MS analysis of Inuit
plasma for both (78Se/77Se) and (200Hg/204Hg) ratios. ........................................................... 50
Figure 12 : Relationship between blood and plasma Se concentrations in Inuit of Nunavik
(n=852).................................................................................................................................. 51


xv
Abréviations
ApoER2 récepteur 2 de l’apolipoprotéine E
ARNm acide ribonucléique messager
ARNt acide ribonucléique de transfert
C4H10O2 hydroperoxyde de tert-butyle
C9H12O2 hydroperoxyde de cumène
Ca++ calcium
CEPT choline/éthanolamine phosphotransférase
CHPT choline phosphotransférase
CNX calnexin
CTQ Centre de Toxicologie du Québec
DIOs iodothyronines désiodases
DMDSe diméthyldiséléniure
DMSe diméthylséléniure
EFSec facteur d’élongation spécifique
eIF4a3 facteur d’initiation eucaryotique
ERAD Endoplasmic-Reticulum-Associated protein Degradation
ERO espèces réactives de l’oxygène
Fe Fer
Fe2SeO3 sélénite ferrique
Gal-1 galectine 1
GPx glutathion peroxydase
Grx glutarédoxine
Grxr glutarédoxine réductase
Grx2 glutarédoxine 2
GSH glutathion
GSSeH glutathio-sélénol
H2O2 peroxyde d’hydrogène
H2Se séléniure d'hydrogène
Hg mercure
Hgin mercure inorganique
HgSe tiemannite
ICP-MS spectrométrie de masse à plasma induit
INSPQ Institut National de Santé Publique de Québec
LaOOH acide linoléique
LDLs lipoprotéines de basse densité
MeHg Méthylmercure
MeHgCl chlorure de méthyl-mercure
MeSeH méthane sélénol
Met méthionine
MsrB1 méthionine R-sulfoxyde réductase 1
Na2SeO3 sélénite de sodium
NO monoxyde d’azote
O2Se dioxyde de sélénium

xvi
OMS organisation mondiale de la santé
RyR récepteur ryanodine
SBP2 protéine de liaison
Se sélénium
Se+IV sélénite
Se+VI séléniate
Se0 sélénium élémentaire
SeAlb sélénoalbumine
Sec sélénocystéine
SECIS Selenocysteine insertion sequence
SeGalNAc méthyl 2-acetamido-2-deoxy-1-seleno-β-D-galactose
SeGluNAc méthyl 2-acetamido-2-deoxy-1-seleno-β-D--glucosamine
Se-II sélénure
SelH sélénoprotéine H
SelI sélénoprotéine I
SelK sélénoprotéine K
SelM sélénoprotéine M
SelN sélénoprotéine N
SelO sélénoprotéine O
SeLP sélénoprotéine P
SelR sélénoprotéine R
SelS sélénoprotéine S
SelT sélénoprotéine T
SelV sélénoprotéine V
SelW sélénoprotéine W
SelX sélénoprotéine X
SeMet sélénométhionine
Sep15 sélénoprotéine de 15KDa
Ser sérine
SeRS séryl-ARNt synthétase
SPS2 sélénophosphate synthétase 2
T3 triiodothyroxine
T4 thyroxine
TMSE triméthylsélénium
Trx thiorédoxine
TrxR thiorédoxine réductase
UGGT UDP-glucoseglycoprotéine glucosyltransférase
UPLC chromatographie en phase liquide à haute performance

xvii
Remerciements
Tout d’abord je tiens à remercier mon directeur de recherche Dr Pierre Ayotte pour m’avoir
donné la chance de réaliser ce projet et de m’avoir guidé avec patience et bienveillance tout
au long de mon parcours à la maîtrise.
Mes remerciements vont aussi à Pierre Dumas qui a été plus qu’un collègue, toujours
présent pour répondre à mes questions et m’aider dans la réalisation de mon projet.
À Nathalie Ouellette, merci pour tes conseils, ta gentillesse et ta présence à chaque fois que
j’avais besoin d’aide.
À tous les membres de l’Institut national de santé publique du Québec (INSPQ) que j’ai
côtoyés de près ou de loin et en particulier à Normand Fleury le directeur scientifique du
Centre de toxicologie du Québec (CTQ) qui m’a ouvert les portes de l’INSPQ pour la
première fois pour un stage qui s’est terminé par une maîtrise.
Enfin, je remercie ma chère épouse Meriem qui m’a soutenu tout au long de cette aventure,
mes adorables filles AYA et NADA qui me rendent la joie quand je la perds, mes chers
parents qui ne cessent de penser à moi, mes amis ainsi que tous les membres de ma famille.


xix
Avant-propos
Ce mémoire est présenté sous forme d’article scientifique. L’ébauche de cet article a été
rédigée par le candidat avec la participation des coauteurs. Pierre Ayotte a révisé l’article
d’une manière critique à plusieurs reprises jusqu’à la version finale, laquelle a été
approuvée par tous les coauteurs. Pierre Dumas et Nathalie Ouellet ont participé
grandement au développement de la méthode analytique.


1
Chapitre I : Introduction
1 Historique
Le sélénium fut découvert accidentellement par le suédois Jacob Berzelius en 1817 [1], lors
de ses travaux de recherche sur une maladie qui a touché des individus œuvrant dans la
fabrication de l’acide sulfurique. Le chercheur pensait à l’époque que c’était l’arsenic qui
était la cause de cette intoxication alors qu’il fit la découverte d’un nouveau composé qu’il
nomma Sélénium du nom grec de la déesse de la lune Selênê.
Le caractère toxique que lui a conféré cette découverte a été à l’origine de la mauvaise
réputation qui a précédé le sélénium pendant plusieurs années, laquelle fut aggravée par
certaines études comme celle de Nelson en 1943 évoquant le risque cancérigène de cet
élément [2]. Cependant, en 1957 le premier effet sur la santé d’une carence en sélénium a
été mis en évidence par les chercheurs Schwarz et Foltz au cours de leurs travaux sur la
nécrose hépatique chez les rats. En effet, leurs résultats montraient clairement qu’un apport
insuffisant en sélénium induit une nécrose fatale du tissu hépatique chez cette espèce [3].
Cette découverte fut à l’origine du statut d’oligo-élément essentiel à l’organisme attribué au
sélénium. Depuis, plusieurs études ont été réalisées pour essayer de bien cerner les effets
reliés au statut en sélénium.
En 1973, le sélénium a été identifié comme un composant essentiel d’un enzyme
antioxydant : la glutathion peroxydase (GPx) [4, 5]. Son importance comme élément trace
essentiel à l’humain a été confirmée par la première étude de supplémentation visant la
prévention de la cardiomyopathie ayant engendré la mort de plusieurs individus dans le
comté de Keshan, province chinoise d’Heilongjihang et dont la cause était une carence en
sélénium [6]. À partir de 1978, une supplémentation systématique en sélénite de sodium
(Na2SeO3) a permis l’éradication presque totale de la maladie dans cette région de la Chine
[7].

2
Durant ces dernières années, plusieurs études de supplémentation en sélénium ont été
réalisées afin de connaitre ses effets sur la santé humaine [8].
2 Propriétés physico-chimiques
Le Sélénium (Se) est un métalloïde appartenant à la famille des chalcogènes regroupant
l’oxygène, soufre, polonium et tellure. De masse atomique égale à 78.96 g.mol-1 il constitue
le 69ème élément sur terre. À l’état naturel, il se présente sous quatre formes d’oxydation: -II
(séléniure), 0 (sélénium élémentaire), +IV (sélénite) et +VI (séléniate) [1].
Le sélénium comporte six isotopes naturels stables à savoir le 74Se, 76Se, 77Se, 78Se, 80Se et
82Se. Les plus abondants sont le 78Se et 80Se représentant chacun 23 et 50% respectivement
du Se total (voir Tableau 1).
Le sélénium élémentaire est allotropique. Ces formes cristallines et amorphes sont
insolubles dans l’eau :
- Le sélénium métallique constitue la forme la plus courante, il est de couleur grise et
sa structure cristalline est hexagonale.
- Le sélénium amorphe rouge foncé monocyclique.
- Le sélénium vitreux de couleur noir.
La forme la plus réduite du sélénium (Se–II) est présente dans les séléniures inorganiques
ainsi que dans l’ensemble des composés organiques séléniés. Le séléniure précipite avec les
éléments métalliques, formant ainsi des séléniures métalliques à faible solubilité.
Tableau 1 : L’abondance relative naturelle des isotopes du sélénium.
Isotope 74Se 76Se 77Se 78Se 80Se 82Se
Abondance relative 0,87% 9,02% 7,58% 23,52% 49,82% 9,19%

3
3 Présence dans l’environnement
3.1 L’atmosphère
L’atmosphère constitue un réservoir transitoire important de sélénium où a lieu son
émission, transport et dépôt dans l’environnement. En effet, des estimations récentes
indiquent que 13 000 à 19 000 tonnes de sélénium par année sont recyclées dans la
troposphère [9].
Il existe deux catégories de sources d’émission de sélénium dans l’atmosphère: naturelles
(60%) et anthropiques (40%). Les sources naturelles, comprennent l’érosion des sols, les
volcans, les sels des océans et les processus de biométhylation des organismes vivants
(bactérie, algues et plantes). En ce qui concerne les sources anthropiques, la combustion des
énergies fossiles constitue la principale source d’émission de sélénium [9].
On distingue trois espèces de sélénium présentent dans l’atmosphère :
- Les composés organiques volatils: le diméthylséléniure (DMSe), le
diméthyldiséléniure (DMDSe) et le méthane sélénol (MeSeH).
- Les composés inorganiques volatils: le sélénium élémentaire Se(0), dioxyde de
sélénium (O2Se) et le séléniure d'hydrogène (H2Se).
- Les composés particulaires: Se(0) surtout lié aux cendres volantes et particules en
suspensions.
Les niveaux atmosphériques de sélénium dans les régions urbaines varient entre 0.1 et 10
ng/m3 provenant essentiellement de la combustion des sources d’énergie fossile et du
charbon [10]. Le sélénium dans l’air ambiant se trouve lier aux cendres volantes et
particules en suspension qui peuvent en contenir entre 1.4 à 11 µg/g et 1 à 110 µg/g
respectivement [10]. Ces valeurs augmentent au fur et à mesure qu’on se rapproche de la
source d’émission [11].

4
3.2 L’eau
Le sélénium dans les différentes nappes d’eau provient des dépôts atmosphériques ou par
drainage des sols et des sous-sols. Les niveaux varient de 0.06 à 0.12 µg/L dans les eaux de
mer [12] et de 0.06 à 400 µg/L dans les eaux de surface et souterraines [1].
Le pH influence grandement la teneur du sélénium dans les eaux de surface. En effet, à pH
alcalin (pH >7.5) les niveaux de sélénium sont élevés résultant de l’oxydation des sélénites
en séléniates solubles dans l’eau. Cependant, en présence du Fer (Fe), le sélénium se
précipite sous forme de sélénite ferrique (Fe2SeO3) ce qui fait diminuer sa concentration
dans ces eaux [1, 13].
Le sélénium dans l’eau potable dépasse rarement les 10 µg/L dans la plupart des pays
hormis quelques régions sélénifères [14]. Le seuil de potabilité fixé par l’Organisation
mondiale de la santé (OMS) pendant plusieurs années de 10 µg/L a été récemment révisé
pour atteindre 40 µg/L [15].
3.3 Le sol
Le sélénium est réparti d’une façon hétérogène dans la croûte terrestre. Son abondance est
faible et varie en moyenne entre de 0.05 à 0.09 µg/g [16]. La teneur des sols en sélénium
dépend en grande partie de la nature des roches mères qui constituent la source primaire par
lessivage [1]. Le sélénium dans ces roches est surtout associé à des minéraux sulfurés sous
forme de sélénites et séléniures. Ainsi, dans les roches volcaniques, le sélénium peut
atteindre des concentrations de 120 µg/g alors que dans le charbon les valeurs sont de
l’ordre 0.47 à 8.1 µg/g [13].
Le sélénium peut être présent dans les sols sous différentes formes : sélénites, séléniates,
sélénium élémentaire ou en association avec des minéraux. Le pH du sol joue un rôle
important en favorisant l’oxydation ou la réduction du sélénium. Ainsi dans les sols à pH
5
alcalin, le sélénium se présente sous forme de séléniate soluble dans l’eau le rendant ainsi
disponible aux plantes [13].
La concentration du sélénium dans les sols varie énormément d’une région à une autre.
Dans les zones séléniprives, comme le Danemark, Nouvelle Zélande et la Chine, la teneur
en sélénium varie entre 0.005 et 2 µg/g. Ces zones sélénifères sont localisées
essentiellement en Irlande, Australie, Canada, Colombie et États-Unis; la plus forte teneur
(1200 µg/g) a été enregistrée en Irlande [1, 13, 17].
Après avoir exposé la présence du sélénium dans l’environnement, on peut à présent
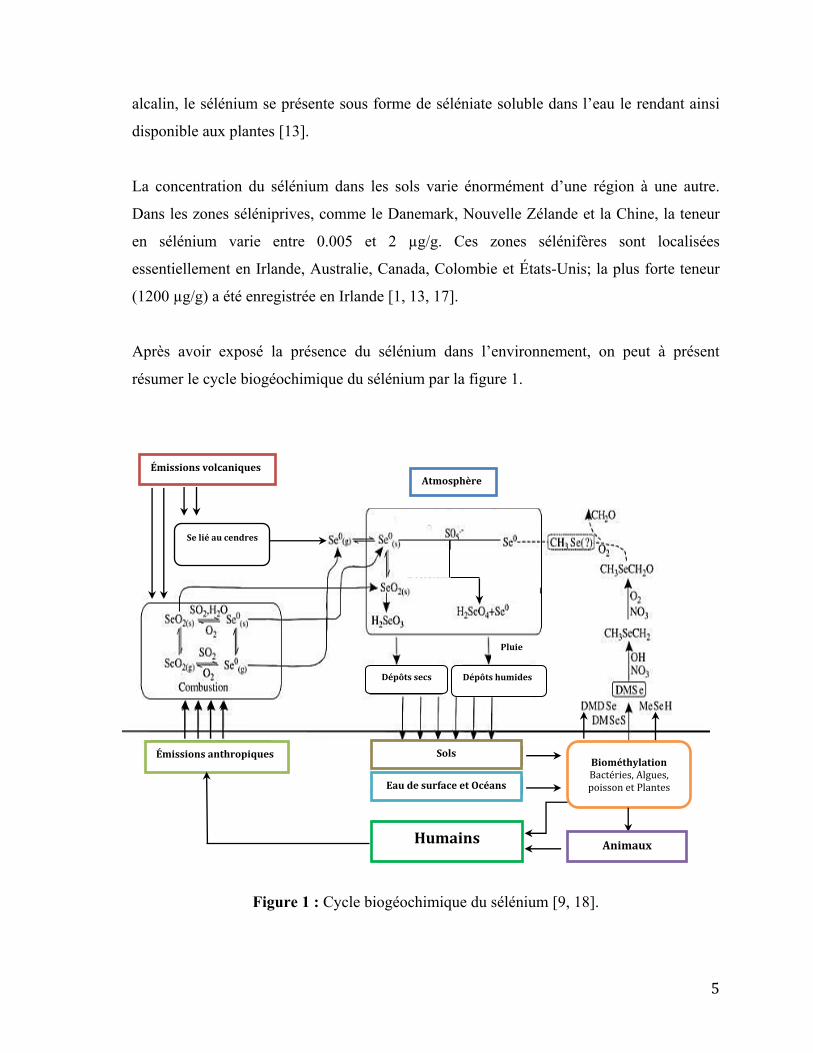
résumer le cycle biogéochimique du sélénium par la figure 1.
Figure 1 : Cycle biogéochimique du sélénium [9, 18].
Émissions volcaniques
Sols
Eau de surface et Océans
Se lié au cendres
Dépôts secs Dépôts humides
Pluie
Atmosphère
Émissions anthropiques Biométhylation
Bactéries, Algues, poisson et Plantes
Humains Animaux

6
4 Sélénium dans l’organisme humain
Le sélénium dans le corps humain est présent principalement sous forme de sélénites,
séléniates, composés organiques séléniés libres ou incorporés dans des protéines appelées
sélénoprotéines. Environ 90% du sélénium total est contenu dans des protéines qui exercent
leurs activités biologiques grâce aux groupements sélénol (R-Se-H) des résidus
sélénocystéine [19, 20].
4.1 Les composés organiques séléniés
4.1.1 Les acides aminés séléniés
4.1.1.1 La sélénocystéine
Analogue sélénié de la cystéine (figure 2), la sélénocystéine (Sec) constitue le 21ème acide
aminé du code génétique dans l’organisme humain [21, 22]. Sa découverte fut suite à la
mise en évidence de la première liaison covalente du sélénium dans le site actif d’une
sélénoenzyme appelé glutathion peroxydase (GPx) [5, 23].
Figure 2 : Structures chimiques de la sélénocystéine et la cystéine.
L’insertion de la sélénocystéine dans les sélénoprotéines est dite spécifique et originale car
en plus du fait qu’elle est codée génétiquement, le codon en question est un codon UGA
(Stop) impliquant une machinerie complexe faisant interagir plusieurs acteurs [24].
CystéineSélénocystéine

7
En effet, la sélénocystéine se différencie des autres acides aminés par le fait qu’elle ne se
présente pas comme un acide aminé libre lié à son acide ribonucléique de transfert (ARNt)
et qu’une une étape de biosynthèse d’un ARNt spécifique à elle est nécessaire. Ceci
commence par le chargement d’une serine (ser) sur un ARNtsec pour former l’ARNt(Ser)Sec
[25]. Ce dernier, subit une aminocylation par l’action de la séryl-ARNt synthétase (SeRS)
pour former la séryl-ARNt(Ser)Sec (ser-ARNt(Ser)Sec) [26, 27]. Par la suite une phosphoséryl-
ARNt(Ser)Sec kinase phosphoryle le résidu séryl en phosphoséryl-ARNt(Ser)Sec (Pser-
ARNt(Ser)Sec) [28, 29] avant sa conversion en sélénocystéyl-ARNt(Ser)Sec (Sec-ARNt(Ser)Sec)
par la sélénophosphate synthétase 2 (SPS2) [21, 30] (Voir figure 3 (A)).
Une séquence sur l’ARN messager (ARNm) appelée élément SECIS (pour SElenoCysteine
Insertion Sequence) est nécessaire à l’insertion de la sélénocystéine et joue un rôle
important dans le recodage du codon UGA en Sec [31, 32]. L’élément SECIS en « tige-
boucle » va être au centre d’un complexe formé par le facteur d’élongation spécifique
(EFSec) et une protéine de liaison (SBP2) pour permettre la présentation de Sec-
ARNt(Ser)Sec au ribosome [32]. Au moins 3 autres facteurs sont aussi impliqués à savoir la
protéine ribosomale L30 [33], la nucléoline [34] et le facteur d’initiation eucaryotique
eIF4a3 [35] (Voir figure 3 (B)).
Figure 3 : Les étapes de biosynthèse de la Sec-ARNt(Ser)Sec (A) et le mécanisme d’insertion
de la sélénocystéine dans les sélénoprotéines (B) chez les eucaryotes [24].
ARNt(Ser)Sec (ser) ser-ARNt(Ser)Sec
Sec-ARNt(Ser)Sec
Pser-ARNt(Ser)Sec
A
B

8
4.1.1.2 La sélénométhionine
Bien que la sélénométhionine (SeMet) ne soit pas codée génétiquement, elle est insérée
dans les protéines d’une façon non spécifique. En effet, elle constitue l’analogue sélénié de
la méthionine (met), et s’insère à sa place par substitution pour former des protéines
séléniées [36].
L’insertion de sélénométhionine dans les protéines n’altère pas leur structure tertiaire.
Cependant, lorsque la substitution de la sélénométhionine à la place de la méthionine a lieu
au voisinage d’un site actif enzymatique, cela peut perturber son activité car le groupement
CH3-Se de la sélénométhionine est plus hydrophobe que le groupement CH3-S de la
méthionine. Il en résulte aussi des perturbations au niveau de l’accès du substrat au site
actif [37].
L’incorporation de la sélénométhionine se fait essentiellement au niveau des globules
rouges et plus précisément dans l’hémoglobine [38]. Dans le plasma elle est incorporée à
l’albumine formant ainsi la sélénoalbumine [39] [36].
Les sélénoprotéines formées par cette insertion non spécifique constituent un réservoir
important de sélénium qui sera utilisé au besoin par l’organisme [37].
Figure 4 : Structures chimiques de la sélénométhionine et la méthionine.
Sélénométhionine Méthionine

9
4.1.2 La sélénonéine
Récemment découvert par le chercheur Japonais Michiaki Yamashita dans le sang du thon
[40], ce sélénocomposé semble être la forme organique prédominante chez les populations
dont l’alimentation repose en partie sur les produits de la mer [41]. On ne connait pas
beaucoup de choses sur la sélénonéine mais sa présence en tant que composé organique
sélénié majeur dans le sang est en faveur d’une activité antioxydante dans l’organisme [41].
Figure 5 : Structure chimique de la sélénonéine.
4.2 Sélénite et séléniate
Ce sont les deux formes oxydées du sélénium dans l’organisme qui partagent les différents
systèmes d’oxydoréduction dans le métabolisme du sélénium [13].
4.3 Les sélénoprotéines
Chez l’humain, il existe environ 25 sélénoprotéines comportant un résidu Sec dans leur site
actif [20]. Ces dernières sont impliquées dans plusieurs fonctions biologiques dont
plusieurs ne sont pas encore identifiées, pour certaines d’entre elles.
4.3.1 La famille des glutathion peroxydases (GPx)
La glutathion peroxydase fut la première sélénoprotéine identifiée chez les mammifères [4,
5]. À présent, il existe au moins 8 isoenzymes dont 5 seulement comportent un résidu Sec,
constituant ainsi des sélénoprotéines [13, 20]. Ces 5 isoformes sont réparties dans différents

10
compartiments cellulaires où elles exercent leur rôle de détoxication des espèces réactives
de l’oxygène (ERO) en utilisant le pouvoir réducteur du glutathion (GSH) (figure 6).
4.3.1.1 La glutathion peroxydase 1(GPx1)
La GPx1 est la sélénoprotéine la plus abondante dans l’organisme. C’est une protéine
ubiquitaire tétramérique localisée dans le cytosol. Elle réduit essentiellement le peroxyde
d’hydrogène (H2O2) et agit comme un antioxydant intracellulaire important [42, 43]. La
GPx1 est la sélénoprotéine la plus sensible au changement du statut en sélénium. En effet,
l’expression de son ARNm ainsi que sa synthèse diminuent lorsque le niveau de sélénium
est faible [44]. C’est aussi le cas, contrairement à ce que l’on peut imaginer, lors du stress
oxydatif [45]. Cette exclusion est en fait dans le but de préserver les ressources cellulaires
en sacrifiant la sélénoprotéine qui récupère rapidement après le stress oxydatif.
Les expériences sur des modèles animaux suggèrent un effet mineur de cette sélénoprotéine
dans des conditions physiologiques normales [43, 46]. Toutefois, on sait maintenant que le
peroxyde d’hydrogène est impliqué dans les voies de signalisation de certaines molécules
en relation avec des processus biologiques tels que la prolifération cellulaire, l’apoptose et
la réponse au stress [47]. Étant donné le grand pouvoir réducteur de la GPx1 sur ce
composé, on ne peut que présumer que cette sélénoenzyme est un acteur principal modulant
plusieurs de ces activités biologiques [48].
4.3.1.2 La glutathion peroxydase 2 (GPx2)
La GPx2 est un homotétramère exprimé dans tout le tractus gastro-intestinal y compris
l’épithélium squameux de l’œsophage [49]. Par sa localisation, elle constitue la première
barrière contre le stress oxydatif induit par les aliments pro-oxydants. Son rôle essentiel est
de lutter contre le stress oxydatif touchant l’épithélium intestinal en ayant la spécificité à
plusieurs substrats incluant le peroxyde d’hydrogène, l’hydroperoxyde de tert-butyle
(C4H10O2), l’hydroperoxyde de cumène (C9H12O2) et l’hydroperoxyde de l’acide linoléique
(LaOOH) [50]. De plus, son expression non uniforme dans l’épithélium, élevée dans les

11
cryptes et diminuant graduellement en allant vers la surface luminale, lui suggère un rôle
dans la prolifération cellulaire [51].
4.3.1.3 La glutathion peroxydase 3 (GPx3)
La GPx3 est la seule sélénoprotéine secrétée dans l’organisme. C’est un homotétramère
synthétisé principalement dans les reins par les cellules de l’épithélium tubulaire proximal
et les cellules pariétales de la capsule de Bowman puis relâché dans la circulation sanguine
[52]. Des études ont aussi révélé qu’elle est exprimée dans d’autres tissus y compris le
cœur, où elle représente la 3ème sélénoprotéine en termes d’abondance de son ARNm [53].
La GPx3 est la deuxième plus abondante parmi les sélénoprotéines plasmatiques et la seule
de sa famille à exercer une fonction antioxydante extracellulaire. Elle réduit le peroxyde
d’hydrogène, les hydroperoxydes d'acides gras et de phospholipides et permet aussi le
maintien de la biodisponibilité du monoxyde d’azote (NO) dans les vaisseaux [54, 55].
4.3.1.4 La glutathion peroxydase 4 (GPx4)
Contrairement aux autres membres de la famille des glutathion peroxydases, la GPx4
possède une structure monomérique et se distingue par son large éventail de substrats
qu’elle peut réduire [56]. En effet, c’est la seule à avoir une spécificité aux hydroperoxydes
de phospholipides et de cholestérol intégrés dans les membranes et les lipoprotéines et peut
agir directement en utilisant l’électron des groupements thiols d’autres co-substrats ou par
le biais du GSH [57, 58].
La GPx4 est la sélénoprotéine la plus abondante dans les testicules avec différentes
localisations : cytosolique, mitochondriale et nucléaire [59]. Elle joue un rôle important
dans la maturation des spermatozoïdes en subissant une transformation, au cours des
dernières étapes de la spermatogénèse, en une structure protéique entrant dans la
composition de la gaine mitochondriale des spermatozoïdes [60]. Elle joue aussi un rôle
dans la condensation de la chromatine et assure sa stabilité structurale dans le sperme [61].

12
Elle est aussi impliquée dans la régulation de l’apoptose [58] [62] et constitue un
antioxydant essentiel à l’embryogénèse [63].
4.3.1.5 La glutathion peroxydase 6 (GPx6)
La GPx6 est la dernière sélénoprotéine découverte dans la famille des GPx et son
expression semble être confinée aux embryons et à l’épithélium olfactif [20]. Il existe peu
de données sur le rôle et la fonction de cette sélénoprotéine mais on peut lui attribuer une
fonction antioxydante dans les tissus où elle est fortement exprimée.
Figure 6 : Mécanisme d’action des GPx selon Papp et al [54].
4.3.2 La sélénoprotéine P (SelP)
La SelP représente la sélénoprotéine la plus abondante dans le plasma incorporant plus de
50% du sélénium total plasmatique [64]. Son expression est confirmée dans tous les tissus
mais elle est synthétisée majoritairement par le foie ou elle est secrétée sous une forme
glycosylée [54]. La SelP possède la particularité de pouvoir contenir jusqu’à 10 résidus Sec
dans sa séquence. Cette caractéristique lui vaut son rôle important dans le transport du
sélénium du foie vers les différents compartiments du corps, essentiellement vers le cerveau
et les testicules [64-66]. Deux récepteurs endocytiques ont été récemment identifiés,
impliqués dans l’utilisation de la SelP par les tissus cibles. Il s’agit en premier du récepteur

13
2 de l’apolipoprotéine E (ApoER2) au niveau du cerveau et testicules [67] et en second de
la mégaline retrouvée au niveau des reins [68].
En plus de son rôle de transporteur de sélénium vers les tissus périphériques, la SelP peut se
lier à l’héparine ou encore aux métaux lourds pouvant aussi jouer le rôle d’agent chélateur
[69]. Des fonctions antioxydantes peuvent aussi lui être attribuées grâce à son pouvoir
protecteur, démontré in vitro, de cellules endothéliales et astrocytes contre le dommage
oxydatif et son inhibition de l’oxydation des lipoprotéines de basse densité (LDLs) [54].
4.3.3 La famille des iodothyronines désiodases (DIOs)
Cette famille regroupe 3 types de sélénoprotéines (DIO1, DIO2 et DIO3) impliquées dans
la régulation de l’activité de l’hormone thyroïdienne. Ces sélénoprotéines ont des structures
et localisations différentes mais elles partagent la caractéristique d’être des protéines
homodimères membranaires et d’avoir toutes un site actif contenant un résidu Sec et deux
histidines au sein d’un repliement de type thiorédoxine (Trx) [24].
4.3.3.1 La désiodase type 1 (DIO1)
La DIO1 est retrouvée au niveau des surfaces membranaires plasmiques des cellules et
exprimée essentiellement au niveau du foie, des reins et des glandes thyroïde et hypophyse
[70]. Elle catalyse la réaction de conversion de la prohormone thyroxine (T4) en une forme
active, la triiodothyroxine (T3) et se trouve être responsable de la régulation des niveaux
circulants de l’hormone thyroïdienne dans le sérum [71].
4.3.3.2 La désiodase type 2 (DIO2)
La DIO2 est localisée à la surface membranaire du réticulum endoplasmique et retrouvée
essentiellement dans le système nerveux central, la glande thyroïde ainsi que les muscles
cardiaques et squelettiques [70]. Tout comme la DIO1, elle assure la conversion de la T4 en
T3 et joue un rôle important dans la régulation locale intracellulaire des niveaux de T3,

14
nécessaire à plusieurs processus biologiques comme la thermogenèse dans les tissus
adipeux bruns [72] et la différenciation et régénération des muscles squelettiques [73].
4.3.3.3 La désiodase type 3 (DIO3)
Cette dernière sélénoprotéine est associée aux surfaces membranaires plasmiques et
retrouvée majoritairement au niveau de l’utérus, du placenta, du cerveau, de la peau et des
testicules [70]. Elle catalyse la réaction de conversion de la T3 en une forme inactive, la T2;
et impliquée aussi dans la régulation locale des niveaux de T3 dans les tissus [71].
4.3.4 La famille des thiorédoxines réductases (TrxR)
Les thiorédoxines réductases (TrxR) sont des enzymes oxydoréductases utilisant le
NADPH pour réduire leur substrat, la thiorédoxine (Trx). Une fois réduite, la Trx est
responsable de la réduction de protéines par le bris des ponts disulfures entre les résidus
cystéines (figure 7). Ce grand pouvoir antioxydant du système thiorédoxine est la raison
pour laquelle on retrouve la TrxR et son substrat dans pratiquement toutes les cellules [74].
Il existe 3 types de TrxR ayant la même structure homodimérique et agissant dans
différents compartiments cellulaires :
4.3.4.1 La thiorédoxine réductase 1 (TrxR1)
LA TrxR1 est essentiellement localisée dans le cytosol et le noyau. La forme cytosolique
est responsable de la réduction de la Trx qui joue un rôle important en tant qu’antioxydant,
régulateur de facteurs de transcription et de l’apoptose [74]. Ce sélénoenzyme réduit aussi
une protéine apparentée à la thiorédoxine, connu sous le nom de TRP14, qui possède le
même rôle que la Trx et se trouve être impliquée dans la stimulation du TNF-α [75, 76]. La
TrxR1 peut aussi agir sur plusieurs autres substrats tels que les sélénites, la vitamine C et
les hydroperoxydes [24]. Elle peut faciliter l’action antioxydante des peroxyrédoxines et la
méthionine sulfoxyde réductase en agissant comme un donneur d’électron [77, 78].

15
La TrxR1 est essentielle à l’embryogenèse [79] et impliquée dans la réparation de l’ADN,
le maintien et la régulation du statut redox intracellulaire [24].
La fonction du résidu Sec a été toujours associée à des activités antioxydantes
hypothétiques. Toutefois, une récente étude chez les souris, menée par le chercheur Peng et
ses collaborateurs, a démontré que le résidu Sec dans la TrxR1 était le seul responsable de
la détoxification du peroxyde d’hydrogène issu du métabolisme du glucose dans les
fibroblastes embryonnaires et que la forme équivalente avec un résidu Cys à la place de Sec
n’a pas pu empêcher la mort cellulaire massive de celles-ci [80].
4.3.4.2 La thiorédoxine réductase 2 (TrxR2)
De localisation mitochondriale, la TrxR2 réduit, en plus de la Trx, la glutarédoxine 2
(Grx2) [81]. Cette sélénoprotéine semble être nécessaire à l’hématopoïèse, le
développement du cœur et la fonction cardiaque [82]. Elle est impliquée aussi dans la
régulation du statut redox des mitochondries au cours du développement par la réduction du
cytochrome c [83]. Enfin, la TrxR2 joue un rôle aussi dans la prolifération cellulaire [84] et
vient d’être récemment associée à la régulation de l’apoptose [85] [86].
4.3.4.3 La thiorédoxine réductase 3 (TrxR3 ou TGR)
Retrouvée essentiellement dans les testicules [87], la TrxR3 réduit à la fois la Trx et le
glutathion (GSH) du fait qu’elle possède un domaine glutarédoxine (Grx) localisé dans sa
partie NH2 terminale [88] . Elle intervient en synergie avec la GPx4 au niveau de la
maturation des spermatozoïdes en catalysant l’isomérisation des ponts disulfures des
différentes protéines impliquées dans ce processus [89].

16
Figure 7 : Mécanisme d’action du système TRx selon Lu et al [90].
4.3.5 La méthionine R-sulfoxyde réductase 1 (MsrB1)
Il s’agit d’une sélénoprotéine cytosolique et nucléaire fortement exprimée au niveau du foie
et des reins connue aussi sous le nom de sélénoprotéine R ou X (SelR et SelX) [91]. C’est
la seule de sa famille, qui comprend deux autres homologues (MsrB2 et MrsB3), possédant
un résidu Sec dans son site actif. Elle est responsable de la réduction de l’énantiomère R de
la méthionine sulfoxyde assurant ainsi la réparation des dommages causés aux protéines par
le stress oxydatif due à l’oxydation de leurs méthionines [92]. La MsrB1 n’agit pas toute
seule; elle fait intervenir la thiorédoxine (Trx) pour sa réduction qui est à son tour réduite
par la TrxR, une sélénoprotéine [93].
Une étude récente a révélé l’action antagoniste de cette sélénoprotéine contre la mono-
oxygénase Mical sur l’assemblage de l’actine. En effet, la Mical oxyde sélectivement deux
méthionines parmi les 16 présentes dans l’actine qui sont réduites réversiblement par la
MsrB1 [94]. Plusieurs autres rôles biologiques ont été proposés incluant sa nécessité dans
la viabilité des cellules du cristallin [95], son effet protecteur contre la
neurodégénérescence [91] et son effet contre le vieillissement [96].
4.3.6 La sélénoprotéine N (SelN)
Il s’agit d’une protéine transmembranaire localisée dans la membrane du réticulum
endoplasmique [97] et codée par le gène SEPN1, dont la mutation a été associée à plusieurs
maladies musculaires [98]. Son expression est majoritaire dans les cellules fœtales et

17
diminue graduellement au fur et à mesure que les tissus deviennent matures incluant
essentiellement le muscle squelettique [97]. La SelN est essentielle au développement et la
différenciation du muscle squelettique [99]. Elle joue aussi un rôle important dans la
régénération des fibres musculaires, suite à une blessure ou un stress, par le maintien d’un
équilibre entre l’état de quiescence et la prolifération des cellules satellites responsables de
l’action régénératrice [100].
Récemment, la SelN a été impliquée dans la régulation de la mobilisation intracellulaire du
calcium (Ca++) par sa liaison au récepteur ryanodine (RyR), responsable de la libération du
Ca++ du réticulum sarcoplasmique lors de la contraction musculaire [101]. De plus, cette
sélénoprotéine semble jouer un rôle important dans le maintien du statut redox et la
protection des cellules contre le stress oxydatif [102].
4.3.7 La sélénoprotéine O (SelO)
La SelO est l’une des sélénoprotéines les moins caractérisées, malgré le fait qu’elle ait été
identifiée plusieurs années auparavant [20] . Cependant une récente étude a levé le voile sur
cette protéine en démontrant qu’il s’agissait d’une sélénoprotéine de localisation
mitochondriale impliquée dans l’activité redox qui s’y produit. Cette étude a révélé aussi
que la SelO est essentielle et que sa présence est affectée par le déficit en sélénium [103].
4.3.8 La sélénoprotéine I (SelI)
C’est une sélénoprotéine transmembranaire, retrouvée dans plusieurs tissus, renfermant un
domaine CDP-alcool phosphotransférase similaire à celui retrouvé dans la choline
phosphotransférase (CHPT) et la choline/éthanolamine phosphotransférase (CEPT) [20]
[104]. La SelI diffère des CHPT et CEPT par l’existence d’un résidu Sec dans sa
terminaison COOH. La fonction de son résidu Sec demeure encore inconnue, cependant
une étude sur l’E. coli a démontré que ce dernier possède une activité éthalonamine
transférase [104].

18
4.3.9 La sélénoprotéine W (SelW)
La SelW est une petite sélénoprotéine de 9 kDa localisée dans le cytosol et fortement
exprimée au niveau des muscles et du cerveau [105, 106]. Elle peut se lier au glutathion
(GSH) [107] et possède une activité antioxydante démontrée in vitro [108] et in vivo [109]
suggérant un rôle important dans la lutte contre le stress oxydatif dans les cellules
musculaires et neuronales. Cette sélénoprotéine a été récemment impliquée dans la
régulation de la protéine 14-3-3, une protéine très abondante dans les voies de signalisation
cellulaire, en favorisant sa dissociation de ses partenaires, à savoir les protéines CDC25B et
Rictor, d’une manière réductive [110, 111].
4.3.10 La sélénoprotéine H (SelH)
La SelH est une sélénoprotéine nucléaire fortement exprimée au cours du développement
embryonnaire [112]. Cette sélénoprotéine se lie à l’ADN et se trouve être impliquée dans la
détoxification cellulaire des espèces réactives de l’oxygène en régulant l’expression des
gènes responsables de la synthèse du glutathion [113]. Récemment, la SelH a été identifiée
comme une sélénoprotéine protectrice de la senescence cellulaire en réponse au stress
oxydatif en maintenant l’intégrité du génome à travers la voie de signalisation impliquant
les protéines ATM et p35 [114].
4.3.11 La sélénoprotéine M (SelM)
La SelM est majoritairement exprimée dans le cerveau et localisée spécifiquement dans le
réticulum endoplasmique [115]. Sa structure comportant un motif de type CysXXSec au
sein d’un repliement de type thiorédoxine lui suggère une activité d’oxydoréduction des
ponts disulfures des protéines [116]. Les études réalisées pour démystifier le rôle de cette
sélénoprotéine ont démontré que la SelM est impliquée dans la protection des neurones
contre les stress oxydatif et qu’elle participe à la mobilisation du Ca++ cytoplasmique [117].
De plus, la SelM interagit avec la galectine1 (Gal-1) laquelle est impliquée dans la
prévention de la neurodégénérescence et le la protection du cerveau [118]. La surexpression

19
de cette sélénoprotéine a induit l’inhibition de l’agrégation du peptide β-amyloïde lui
suggérant un rôle préventif contre la maladie d’Alzheimer [119]. Il demeure clair que la
SelM joue un rôle important dans le cerveau, cependant les mécanismes par lesquels elle
exerce ses fonctions sont loin d’être identifiés.
4.3.12 La sélénoprotéine de 15KDa (Sep15)
Cette sélénoprotéine appartient à la même famille que la SelM partageant toutes les deux
31% d’homologie de séquence [120]. La Sep15 est localisée au niveau du réticulum
endoplasmique avec des niveaux d’expression élevés dans les reins, le foie, la prostate et
les testicules [121]. Similaire à la SelM, sa structure lui suggère des fonctions
d’oxydoréduction des protéines [116]. Dans le réticulum endoplasmique, la Sep15 forme un
complexe avec l’UDP-glucoseglycoprotéine glucosyltransférase (UGGT) dont l’action
consiste à empêcher la sortie des glycoprotéines mal repliées du réticulum endoplasmique
et les rediriger vers des voies de dégradation [122]. De plus, la Sep15 est impliquée dans le
contrôle de qualité du repliement des protéines en régulant le cycle de la calnexin (CNX),
une protéine nécessaire à ce processus [123]. Enfin, une étude récente sur des souris
Knock-out a associé le déficit de la Sep15 avec la formation de cataractes par défaut de
repliement des protéines du cristallin [124].
4.3.13 Les sélénoprotéines K et S (SelK, SelS)
Ces deux sélénoprotéines ne partagent aucune homologie de séquence mais, par leur
topologie et leurs fonctions, elles peuvent être regroupées dans une seule famille. En effet
les deux sont des sélénoprotéines transmembranaires présentes dans le réticulum
endoplasmique [20] et sont impliquées dans la voie de dégradation des protéines mal
repliées par la machinerie ERAD (pour Endoplasmic-Reticulum-Associated protein
Degradation) [125].

20
4.3.14 La sélénoprotéine T (SelT)
La SelT est exprimée dans différents tissus embryonnaires et adultes avec une localisation
au niveau du réticulum endoplasmique et l’appareil de Golgi [126]. La SelT est impliquée
dans la mobilisation du Ca++ au niveau des cellules neuronales stimulée par le neuropeptide
PACAP [127]. Récemment, la SelT a été impliquée dans la régulation de la fonction des
cellules β et le maintien de l’homéostasie glucidique.
4.3.15 La sélénoprotéine V (SelV)
Exprimé essentiellement dans les testicules, la SelV possède la même structure que la SelW
et la SelH [20, 126]. On ne connait pas de rôles biologiques associés à cette sélénoprotéine
mais sa présence dans l’organe où a lieu la spermatogenèse souligne davantage le rôle du
sélénium dans ce processus.
4.3.16 La sélénophosphate synthétase 2 (SPS2)
Son rôle dans la synthèse des sélénoprotéines a été évoqué dans la section 4 [30].
5 Toxicocinétique du sélénium
5.1 Absorption
L’absorption du sélénium dans l’organisme humain se produit au niveau de l’intestin avec
une efficacité variable, dépendante de la forme ingérée et de la présence ou non des
aliments. En effet, l’absorption du sélénium augmente en présence des protéines, de la
vitamine E et A et diminue en présence des sulfates, de l’arsénique, du mercure et de la
vitamine C [128].
L’efficacité de l’absorption du sélénium, toutes formes confondues, est supérieure à 80%.
Toutefois, les formes organiques sont mieux absorbées que celles inorganiques. La

21
sélénométhionine est la forme la mieux absorbée par l’intestin avec une efficacité
d’absorption de 90%. Cette dernière l’est par un mécanisme actif similaire à celui de la
méthionine. L’absorption de la sélénocystéine quant à elle n’est toujours pas claire avec des
évidences que le mécanisme n’est pas actif [17].
En ce qui concerne les formes inorganiques, les séléniates sont mieux absorbés que les
sélénites avec une efficacité d’absorption moyenne de 60% [129]. En effet, le séléniate est
absorbé par un mécanisme de transport actif similaire à celui des sulfates alors que le
sélénite est absorbé par simple diffusion passive [17].
5.2 Distribution
Après absorption intestinale, le sélénium rejoint la circulation sanguine pour être distribué
vers différents organes. Chez les individus dont l’apport en sélénium est adéquat, le
sélénium est réparti comme suit : 30% dans le foie, 15% dans les reins, 10% dans le plasma
et 30% dans les muscles avec une prédominance au niveau du muscle cardiaque. Dans les
conditions où l’apport en sélénium est faible, l’accumulation du sélénium diminue dans
tous les organes à l’exception des reins ou a lieu la synthèse de la GPx3. Cette diminution a
pour but de favoriser sa distribution vers les organes nobles comme le cerveau [13, 17].
Dans le sang, la majeure partie du sélénium est retenue dans les globules rouges alors que
dans le plasma il est essentiellement transporté par la SelP (à plus de 50%). Le reste du
sélénium plasmatique est partagé entre la GPx3 et la SeAlb [17] [64].
5.3 Biotransformation
Le séléniure d’hydrogène (H2Se) constitue le métabolite intermédiaire central autour duquel
différentes voies de transformation de sélénium ont lieu (figure 8). Le sélénite et le
séléniate sont transformés en H2Se par le système glutarédoxine (Grx, Grxr et GSH) et/ou
le système thiorédoxine (TrxR et Trx) [130]. Le séléniate est aussi réduit en sélénite par la
voie de réduction des sulfates mais le mécanisme sous-jacent n’est toujours pas clair [131].

22
Les formes organiques issues de l’alimentation subissent aussi plusieurs
biotransformations. La sélénométhionine est soit incorporée non spécifiquement dans des
protéines [36] ou soit transformée en sélénocystéine par la cystathionine β-synthase et γ-
lyase [132]. La sélénocystéine provenant de l’alimentation ou de la conversion de la
sélénométhionine est transformée en H2Se par l’action de la sélénocystéine β-lyase [133].
La méthyl-sélénocystéine est convertie en méthyl-sélénol par la γ-lyase qui sera par la suite
transformée en H2Se [134].
Le H2Se ainsi formé est converti en sélénophosphate pour assurer la synthèse des 25
sélénoprotéines codées par le génome [135]. Il peut aussi être métabolisé en séléno-sucres
via la glutathio-sélénol (GSSeH) [136] ou subir une méthylation avant d’être excrété [137].
Figure 8 : La biotransformation du sélénium selon Alexander et al [13].

23
5.4 Élimination
L’élimination du sélénium est essentiellement rénale chez les individus dont l’apport en
sélénium est normal. Le triméthylsélénium (TMSE) et les séléno-sucres à savoir le méthyl
2-acétamido-2-déoxy-1-séléno-β-D-galactose/-glucosamine (SeGalNAc et SeGluNAc
respectivement) représentent les composés majeurs excrétés. À des doses toxiques, le
sélénium est aussi éliminé sous forme de TMSE par exhalation. Des formes non
métabolisées de séléniate et sélénite sont aussi retrouvées dans les urines [13].
6 Sélénium et santé
6.1 Les maladies cardiovasculaires
L’association de certaines maladies cardiovasculaires aux polymorphismes génétiques des
gènes codants pour les sélénoprotéines GPx1, GPx3 et SelS suppose que le sélénium
pourrait avoir un effet bénéfique dans la prévention des maladies cardiovasculaires [138-
141]. Cependant, les essais cliniques de supplémentation en sélénium n’ont permis de
déceler aucun effet protecteur [142, 143]. De plus, l’étude sur les éléments traces CARDIA,
réalisée auprès de 3112 jeunes adultes Américains, portant sur l’association de la teneur en
sélénium des ongles et l’athérosclérose, était aussi non concluante [144].
Flores-Mateo et ses collaborateurs ont réalisé une méta-analyse comportant 25 études
observationnelles et 6 essais randomisés de supplémentation en sélénium, provenant de
plusieurs pays de l’Europe, la Chine, les États-Unis et l’Australie. Ils ont pu mettre en
évidence une association significative inverse entre le statut en sélénium, mesuré dans le
sang et/ou dans les ongles, et le risque de maladies coronariennes. Cependant, l’auteur
conclut que ces résultats doivent être validés du fait de la nature observationnelle des études
utilisées [145].
L’association entre le sélénium et les maladies lipidiques est aussi complexe. Une
supplémentation de sélénium entre 100 et 200 µg par jour chez une population ayant un

24
statut en sélénium bas s’est avérée bénéfique, en diminuant considérablement le taux du
cholestérol total. Toutefois, dans cette même étude le statut élevé en sélénium et/ou la
supplémentation de plus de 300 µg par jour étaient associés à une augmentation du taux de
cholestérol total [146].
Les différences observées dans ces études sont probablement liées au statut en sélénium des
populations étudiées. Les effets bénéfiques du sélénium ont été observés dans les études où
le statut en sélénium des participants était bas. En outre, l’optimisation du sélénium au
départ des études est aussi un facteur pouvant annuler l’effet bénéfique de la
supplémentation en sélénium sur les maladies cardiovasculaires et lipidiques [8].
6.2 Le diabète de type 2
Les résultats des différentes études et essais cliniques visant à déterminer les effets du
sélénium sur le diabète divergent. Dans l’étude prospective EVA réalisée en France, le
statut élevé de sélénium chez les hommes (entre 94 et 155 µg/L) était associé à une
diminution du risque d’hyperglycémie [147]. De même, aux États-Unis, dans une récente
étude prospective sur deux cohortes de sexe différent incluant 3630 femmes et 3535
hommes, le taux de sélénium dans les ongles était inversement associé au risque de diabète
de type 2 [148]. À l’opposé, deux autres grandes études transversales réalisées aux États-
Unis ont montré que le sélénium était associé à une augmentation de la prévalence du
diabète [149, 150].
Les résultats de supplémentation en sélénium étaient aussi controversés. Certaines d’entre
elles n’ont trouvé aucune association [151, 152] alors que d’autres révèlent une
augmentation du risque de diabète du type 2 en relation avec l’apport en sélénium [153,
154].
L’explication de la non-concordance de tous ces résultats peut être attribuée à des
différences du statut en sélénium entre les populations et par conséquent à l’activité de
certaines espèces de sélénoprotéines, incluant la GPx1, la DIO2 et la SelP [155].

25
6.3 Le système immunitaire
Le sélénium se révèle bénéfique au système immunitaire. En effet, la supplémentation en
sélénium à raison de 400 µg par jour chez des personnes âgées a fait augmenter le nombre
de lymphocytes T de 27% par rapport au placebo [156]. Chez d’autres personnes
supplémentées, le sélénium était responsable de l’augmentation de l’immunité à médiation
cellulaire [157] et de la réponse immunitaire à une stimulation antigénique [158]. Le
sélénium semble jouer un rôle important dans la régulation de l’expression des cytokines et
la prolifération des lymphocytes T activés [159].
6.4 Les maladies infectieuses virales
Le sélénium joue un rôle important dans la lutte contre les maladies virales [159]. Le déficit
en sélénium a été associé à une diminution de la survie des enfants infectés par le VIH
[160] et une augmentation de la virulence et de la propagation d’autres infections virales
[161, 162]. Les essais cliniques randomisés ont démontré aussi des effets bénéfiques de la
supplémentation en sélénium chez des sujets VIH-positifs [163, 164].
6.5 Le cancer
De nombreuses études prospectives ont démontré que le sélénium diminue le risque de
plusieurs cancers. Cet effet bénéfique est d’autant plus observé chez les populations dont le
statut en sélénium est faible [8].
Le sélénium semble diminuer le risque de cancer de prostate dans plusieurs études et essais
cliniques [165-167]. De plus, l’essai clinique «The Nutritional Prevention of Cancer»
(NPC), regroupant 1312 personnes ayant reçu une supplémentation quotidienne de 200 µg
de sélénium a révélé une diminution de l’incidence du cancer de la prostate de 67%. Cette
observation était confinée au groupe de participants dont le statut en sélénium au début de
l’étude était inférieur à 106 µg/L [168]. Cependant, l’essai clinique SELECT regroupant
35 533 Américains n’a pas pu reproduire les résultats de l’étude NPC avec la même dose de

26
sélénium par jour, pour une même durée de traitement [154]. Cette différence peut être
expliquée par le fait que la forme de sélénium était différente entre les deux études et
surtout le statut en sélénium des participants au début de l’étude SELECT qui était
supérieur à 106 µg/L [8].
Le sélénium pourrait aussi avoir un effet protecteur contre le cancer du poumon. En effet,
une méta-analyse regroupant 16 études cliniques [169] a révélé que le risque relatif (RR) de
développer un cancer de poumon chez les individus ayant un statut en sélénium
plasmatique élevé était de 0.74 (intervalle de confiance à 95% [0.57-0.97]). De plus, le RR
était nettement plus bas, RR=0.46 (intervalle de confiance à 95% [0.24-0.87]), lorsque le
sélénium était mesuré dans les ongles.
L’effet du sélénium sur le cancer de la vessie a fait l’objet d’une méta-analyse en 2010 qui
a conclu qu’un apport élevé en sélénium est associé à une diminution du risque de cancer
de vessie [170].
Toutefois, ces évidences sont à l’opposé de ce qui été observé dans plusieurs études où le
statut ou l’apport en Se élevé était souvent non protecteur voire même augmente le risque
de certains cancers [154, 168].
7 Interaction entre le sélénium et le méthylmercure (Se-MeHg)
L’effet protecteur du sélénium contre la toxicité du mercure fut rapporté pour la première
fois en 1967 par Parizek et Ostadalova qui ont observé une diminution considérable de la
toxicité liée au mercure inorganique (Hgin) suite à l’administration simultanée du Hg2+ et du
sélénium sous forme de sélénite chez les rats [171]. Suite à cette découverte, les études
visant à déterminer les effets du sélénium sur le mercure et les mécanismes d’interaction
entre eux se sont accentuées.
Le méthylmercure (MeHg) avec sa capacité à traverser la barrière hémato-encéphalique
[172, 173] et placentaire [174] et son absorption gastro-intestinale voisine de 100% [175]

27
constitue la forme la plus toxique pour l’humain et suscite le plus grand intérêt des
chercheurs. En 1972, Ganther et al ont publié le premier rapport mettant en évidence une
interaction entre le Se et le MeHg. En effet, les animaux nourris avec une diète riche en
MeHg et en Se ont présenté une diminution des signes de toxicité et ont vécu plus
longtemps que ceux nourris avec une diète riche en MeHg seulement [176]. Suite à ces
observations, plusieurs autres études se sont succédées chez différentes espèces animales
confirmant l’effet antagoniste que peut avoir le Se sur le MeHg.
L’interaction entre le MeHg et le Se a fait l’objet de plusieurs revues de littérature, dont les
plus récentes publications sont celles de Khan et al (2009) [177] et Yang et al (2008) [178].
Le sélénium peut exercer un effet antagoniste sur le mercure par plusieurs mécanismes qui
sont présentés ci-dessous.
7.1 Diminution de la toxicité par formation de composés MeHg-Se(H)
Le MeHg présente une grande affinité pour les groupements thiols (SH) qui sont très
abondants dans les biomolécules. Cette affinité conduit à la formation de complexes MeHg-
S-R avec une constante de formation allant de 1015 à 1017 [179]. La liaison entre le MeHg
et le groupement SH, malgré sa force, est instable dans les fluides biologiques et le MeHg
peut être déplacé par un autre ligand. Cette instabilité conduit à la formation de complexes
méthylmercure sélénol (MeHg-SeH), par un échange de ligand entre les deux groupements
sélénol et thiol, dont la constante de formation est beaucoup plus grande que celle du
complexe MeHg-SH [180].
La formation de composés MeHg-Se-R entraîne une diminution de la biodisponibilité du
mercure et joue un rôle primordial dans la détoxification de celui-ci. En 1979, Magos et
coll. ont démontré qu’en incubant des érythrocytes de rat avec du chlorure de méthyl-
mercure (MeHgCl) et du sélénite de sodium (Na2SeO3), un composé MeHg-Se extractible
dans le benzène apparaissait [181]. Ce composé a été ultérieurement identifié comme étant
le séléniure bis(méthyl-mercure) [182]. Plusieurs autres composés MeHg-Se ont été

28
proposés ou synthétisés par la suite mais aucun d’entre eux n’a été retrouvé dans les
organismes vivants [177].
7.2 Le sélénium joue un rôle dans la biotransformation du MeHg
Les mammifères marins présentent de faibles concentrations de MeHg dans le foie et les
reins, malgré le fait que l’espèce dominante de mercure dans les muscles des poissons
qu’ils consomment soit le MeHg [183, 184]. En effet, le MeHg subit une biotransformation
par déméthylation produisant ainsi du mercure inorganique qui va s’accumuler dans ces
organes [185-187].
Le ratio molaire Hg:Se voisin de 1 observé dans le foie de plusieurs espèces de poissons
[183, 184, 188] a soulevé depuis longtemps le questionnement des chercheurs quant à
l’effet protecteur du sélénium contre la toxicité du mercure. L’hypothèse que le Se puisse
être directement impliqué dans la déméthylation du MeHg a été proposée par Martoja et
Berry en 1980 suite à la découverte de granules de tiemannite (HgSe) dans le foie des
dauphins de Cuvier [189, 190]. Ces chercheurs ont supposé que le Se est directement lié à
la déméthylation du MeHg et que les granules de HgSe seraient le produit final de cette
dégradation. Ils ont aussi suggéré que ce processus est prévu dans les espèces ou le Se et
l’Hg coexistent d’une façon équimolaire. Cette hypothèse a été soutenue par Nigro et
Leonzio qui ont identifié le même type de granules dans différentes espèces de mammifères
marins. Ils ont démontré aussi que ces granules sont principalement localisées dans le
cytoplasme des macrophages [190]. Une récente étude publiée en juillet 2012 a révélé, chez
les dauphins de Guyane, l’existence de cristaux accumulés dans les cellules de Kupffer qui
seraient probablement des granules HgSe [191]. Palmisano et ces collaborateurs ont
rapporté la présence de Hg lié à des protéines dans le foie des dauphins [192]. En effet, les
granules de HgSe peuvent former des polymères et se lier à une protéine qui semble être la
sélénoprotéine P [193]. Le mécanisme responsable de la déméthylation du mercure est
encore non élucidé. Il semblerait qu’un seuil de MeHg doive être atteint pour que le
processus de formation de granules HgSe se déclenche [192].

29
En résumé, le rôle exact du Se dans la biotransformation du MeHg est loin d’être claire
mais son implication, dans la formation de HgSe, est confirmée chez plusieurs espèces de
mammifères marins. Il reste à savoir si un tel mécanisme de détoxication pourrait exister
chez l’humain.
7.3 Le sélénium inhibe le produit de la dissociation du MeHg
Ganther et ces collaborateurs ont émis l’hypothèse que la toxicité du MeHg n’est pas
induite seulement par la molécule intacte et que les radicaux méthyl libres formés par une
rupture homolytique de la molécule pourraient être incriminés [194]. Si cette hypothèse
s’avère vraie, la glutathion peroxydase (GPx) pourrait contrecarrer l’effet que peuvent
induire ces radicaux.
8 Population d’étude
8.1 Les Inuits du Nunavik
Figure 9 : Carte géographique du Nunavik [195].

30
Le Nunavik représente la partie arctique du Québec, situé au nord de la 55ème parallèle
(Figure 9). Environ 11 000 Inuits y résident dans les 14 communautés qui longent les 2000
km de côtes de l’ouest de la Baie d’Hudson, du détroit d’Hudson et de la Baie d’Ungava.
La population inuit du Nunavik est la plus jeune de toutes les populations inuites du Canada
avec 37% d’enfants âgés de 0-14 ans et 58% pour le groupe d’âge de 0-24 ans (voir tableau
2) [196].
Groupes d'âge et médiane Nunavik
(%)
Total - Groupes d'âge 100,0
0 à 24 ans 58,4
0 à 14 ans 37,0
0 à 4 ans 13,1
5 à 9 ans 12,4
10 à 14 ans 11,5
15 à 24 ans 21,4
25 à 64 ans 39,0
65 ans et plus 2,7
Âge médian (années) 21
Tableau 2 : Répartition selon le groupe d'âge et âge médian des Inuits du Nunavik (année
2011) [197].
8.2 Alimentation traditionnelle des Inuits du Nunavik
Les Inuits perçoivent la nourriture traditionnelle comme une composante essentielle pour la
santé et le bien-être. Elle représente une partie de l’identité et des valeurs de ce peuple
autochtone. Le terme de «nourriture traditionnelle » fait référence à tous les aliments
récoltés dans l’environnement local. Il englobe plusieurs variétés et espèces incluant les
poissons, le caribou, les oiseaux, le phoque, les baleines, les plantes et les petites baies
[198].
Cette alimentation traditionnelle dont le savoir est transmis d’une génération à l’autre est
très bénéfique pour la santé par sa richesse en protéines, vitamines et minéraux. De plus, les
mammifères marins, consommés en grande quantité, sont riches en sélénium et en acides
gras polyinsaturés (oméga-3), lesquels sont essentiels à l’activité métabolique de

31
l’organisme [199]. Toutefois, par cette même alimentation, les Inuits sont exposés à
certains contaminants environnementaux qui ne cessent d’augmenter dans l’Arctique [200].
Parmi ceux ci, le méthylmercure (MeHg) est l’élément le plus préoccupant car il est
fortement bioaccumulé dans les mammifères marins. Ce dernier a été associé aux
accouchements prématurés et retards de croissance fœtale lorsque les parents Inuits sont
exposés [201]. De plus, l’exposition des parents Inuits au MeHg est associée au trouble de
déficit de l'attention avec hyperactivité (TDAH) observé chez les enfants d’âge scolaire
[202, 203].
La présence d’éléments comme le sélénium et les oméga-3 peuvent contrer les effets
néfastes du MeHg. Plusieurs études ont démontré que ces deux éléments sont bénéfiques
pour la santé. Chez les Inuits du Nunavik, les oméga-3 ont été associés à une augmentation
de poids à la naissance, une amélioration de la fonction de mémoire chez les enfants d’âge
scolaire et à une diminution de la prévalence des facteurs de risque cardiovasculaire chez
les adultes [204-207].
En ce qui concerne le sélénium, les évidences sont de plus en plus fortes en faveur d’une
action antagoniste contre le MeHg chez cette même population [208, 209]. Toutefois, des
doutes subsistent en ce qui concerne l’innocuité des apports élevés en sélénium observés
chez cette population.
8.3 Recrutement et échantillonnage
Notre étude se situe en marge de l’Enquête de santé auprès des Inuits du Nunavik 2004 -
Qanuippitaa? How are we? visant à vérifier l’évolution de l’état de santé de la population
Inuit du Nunavik. La méthode de recrutement aléatoire stratifiée en deux étapes était
utilisée pour maximiser le nombre de participants et avoir un échantillon le plus
représentatif de la population. Au terme de ce recrutement, 889 participants âgés de 18 à 74
ans ont été accueillis au bord du bateau de recherche Amundsen où l’équipe de recherche a
procédé aux prélèvements sanguins et à l’enregistrement des différents paramètres
anthropométriques et cliniques (pression, pouls, température et indice de masse corporelle).

32
Des questionnaires ont été aussi administrés visant à documenter les données
sociodémographiques et habitudes alimentaires détaillées de chacun des participants [210].
Notre étude a été réalisée à l’aide des échantillons plasmatiques issus de cette enquête et
qui ont été entreposés à -80 °C au niveau du Centre de Toxicologie du Québec (CTQ) de
l’Institut National de Santé Publique de Québec (INSPQ).
9 Objectifs
L’objectif principal est d’améliorer notre connaissance du statut en sélénium de la
population inuite du Nunavik, plus précisément les niveaux circulants des différentes
espèces de sélénoprotéines dans le plasma d’adultes inuits ayant participés à l’enquête de
santé Qanuippitaa-2004.
Les objectifs spécifiques sont :
1. Mettre au point une méthode analytique pour identifier et quantifier les
sélénoprotéines plasmatiques.
2. Appliquer la méthode à l’ensemble des échantillons plasmatiques des participants à
l’enquête de santé Qanuippitaa-2004.
L’objectif secondaire était de monitorer les fractions de mercure associées aux
sélénoprotéines lors de leur analyse.
10 Démarche expérimentale
La méthode développée est une adaptation de la méthode déjà publiée par Li et al en 2011
[211]. Elle implique la séparation des sélénoprotéines plasmatiques par un système de
chromatographie d’affinité en phase liquide à haute performance (UPLC) couplée à la
spectrométrie de masse à plasma induit (ICP-MS) avec quantification par dilution
isotopique.

33
10.1 Spéciation du sélénium plasmatique
En résumé, 100 µl de plasma est injecté dans le système UPLC à un débit de 1 ml/min et
véhiculé par une phase mobile à base d’acétate d’ammonium. Dans un premier temps
l’échantillon rejoint une première colonne contenant de l’héparine sépharose qui peut
retenir sélectivement la SelP. Ensuite, le plasma dépourvu de sa SelP atteint une deuxième
colonne contenant la blue sepharose comme phase solide, laquelle retient la SelAlb. Ainsi,
la GPx3 et les petites molécules séléniées non retenues seront donc les premières à
rejoindre l’ICP-MS. Après 5 minutes, un système de valve externe connectant les deux
colonnes va permettre l’exclusion de la deuxième colonne renfermant la SeAlb et l’élution
de la SelP. Par la suite, le système de valve est ramené à sa position initiale pour inclure la
deuxième colonne à nouveau et éluer la SelAlb.
10.2 Quantification du sélénium et du mercure
À la sortie de l’UPLC une solution enrichie en 77Se et 204Hg est infusée de façon continue et
se mélange au flux sortant avant de parvenir à l’ICP-MS. Une fois arrivé, le flux est
introduit dans la chambre de nébulisation puis arrive dans la torche à plasma où circule du
gaz d’argon. Le débit du gaz élevé dans un espace restreint favorise la friction de ses
atomes et génère ainsi une chaleur qui peut atteindre les 8000k. L’échantillon soumis à
cette chaleur est atomisé et ionisé. Un système de vide différentiel accélère les ions du
plasma, à travers les orifices des différents cônes situés après le plasma d’argon, vers un
ensemble de lentilles électrostatiques permettant d’extraire les ions chargés positivement et
les envoyer vers un analyseur de masse quadripolaire. L’analyseur de masse transmet
seulement les ions ayant un rapport masse sur charge particulier, déterminé en fonction des
voltages et de la fréquence appliquée au quadripôle. Les ions positifs parviennent ensuite au
détecteur qui comptabilise les ions à la masse considérée. Les masses détectées dans cette
méthode sont : le 76Se, 77Se, 78Se, 82Se, 200Hg, 203Hg et 204Hg.
Au terme de la séparation par UPLC et la détection par ICP-MS, des comptes par seconde
sont enregistrés pour les différentes masses surveillées. Ainsi, une élévation du signal de

34
sélénium de la ligne de base (pic) correspond à une espèce de sélénium séparée. Suivant
l’ordre de séparation sus-cité, l’ordre des sélénoprotéines détectées est : GPx3, SelP puis la
SeAlb. L’élévation du signal du mercure correspond au mercure associé aux
sélénoprotéines.
La quantification du sélénium et du mercure est basée sur les principes de la dilution
isotopique [212] . Les éléments chimiques en général présentent plusieurs isotopes stables.
Un isotope est un nucléide qui possède le même nombre de protons mais diffère par son
nombre de neutrons. L’abondance de ces isotopes naturels pour un élément donné est
conservée ce qui implique que les ratios isotopiques pour une solution donnée le sont aussi.
Supposant qu’on dispose d’une solution naturelle renfermant un analyte, qu’on veut doser,
composée de plusieurs isotopes dont (A) et (B) (les masses et les ratios isotopiques naturels
sont aussi connus). Par l’ajout d’une solution enrichie en isotope (B) qu’on mélange en
proportions connues avec l'échantillon; après homogénéisation, on peut mesurer le nouveau
rapport isotopique par ICP-MS. Avec la mesure de ce rapport altéré par l’ajout de l’isotope
B on peut déterminer la concentration de l’élément à doser par l’équation suivante:
C𝑥 =m𝑠𝑝 × C𝑠𝑝
m𝑥[R (
NANB)
𝑚𝑖𝑥× Ab (B)𝑠𝑝 − Ab (BA)𝑠𝑝
Ab (A)𝑥 − R (NANB
)𝑚𝑖𝑥
× Ab (B)𝑥
]
Cx = concentration inconnue à déterminer
msp = masse de l’isotope ajouté
Csp = concentration de l’élément dans la solution enrichie
mx = masse de l’échantillon prélevé
R (NA
NB)
𝑚𝑖𝑥 = ratio isotopique dans le mélange
AB = abondance en % pondéral
Ab (A)sp = abondance de l’isotope A dans la solution enrichie
Ab (B)sp = abondance de l’isotope B dans la solution enrichie
Ab (A)x = abondance de l’isotope A dans l’échantillon

35
Ab (B)x = abondance de l’isotope B dans l’échantillon
La solution d’infusion enrichie en 77Se et 204Hg mélangée au flux sortant de l’UPLC est
donc utilisée pour quantifier le sélénium et le mercure associé aux différentes espèces de
sélénoprotéines séparées. Après détection par ICP-MS le ratio isotopiques 78 Se/77Se et
200Hg/ 204Hg sont utilisés pour la quantification du Se et Hg respectivement.


37
Chapitre 2: Article scientifique
Circulating levels of selenium species in Inuit adults from Nunavik
A. Achoubaa, P. Dumasb, N. Ouelleta, M. Lemirea b c, P. Ayottea b c*
a Axe santé publique et pratiques optimales en santé, Centre de recherche du CHU de
Québec, Québec, Canada;
b Centre de toxicologie du Québec, Institut national de santé publique du Québec, Québec,
Canada.
c Département de médecine sociale et préventive, Faculté de médecine, Université Laval,
Québec, Québec, Canada
*Corresponding author: e-mail: [email protected], phone: 418-650-5115 ext.4654,
fax: 418-654-2148
KEYWORDS: selenium, selenoproteins, isotope dilution analysis, Inuit, plasma, blood,
mercury.
Abstract
Selenium (Se) is highly abundant in traditional marine foods consumed by Inuit and
accordingly their Se intake is among the highest in the world. Some of these foods are also
high in methylmercury (MeHg), which is bioaccumulated in fish and marine mammals. Se
is an important element in the antioxidant defense system and therefore high Se intake may
counteract some of the deleterious effects of MeHg exposure. However, elevated plasma Se
concentrations have recently been associated to type 2 diabetes, hypercholesterolemia
and/or hypertension. In this study, a method combining affinity chromatography and
inductively coupled plasma-mass spectrometry with quantification by post-column isotope
dilution was adapted in our laboratory and used to determine plasma levels of
selenoproteins in archived plasma samples of Inuit adults who participated to the Nunavik
Inuit Health Survey 2004. Mercury associated with selenoproteins was also quantified.
Results show that glutathione peroxidase 3 (GPx3), selenoprotein P (SelP) and

38
selenoalbumin (SeAlb) represented respectively 25%, 52% and 23% of total plasma Se
concentration (N = 852). In addition, small amount of Hg co-eluted with each Se-
containing protein and 50% of plasma Hg was associated to SelP. A non-linear relationship
was observed between plasma and blood Se levels in Inuit. Plasma Se concentration
(median = 139 µg L-1; interquartile range (IQR) = 22.7 µg L-1) were markedly lower and
less variable than these blood Se concentration (median = 261 µg L-1, IQR = 166 µg L-1).
The non-linear relationship between plasma and blood Se suggests that a selenocompound,
possibly selenoneine, accumulates in the blood cellular fraction of several Inuit adults.
1. Introduction
Selenium (Se) is an essential trace element for humans. It is mainly incorporated in
selenoproteins in the form of selenocyteine (SeCys) which is encoded by the UGA codon in
the human genome [20]. A total of 25 selenoproteins have been identified until now, which
contain a SeCys residue in their active site and play several biological functions, notably
protection from oxidative stress [20, 24]. In human plasma, three Se containing proteins,
namely selenoprotein P (SelP), glutathione peroxidase 3 (GPx3) and selenoalbumin
(SeAlb), are found so far [213]. Only SelP and GPx3 possess SeCys residues and belong to
the family of selenoproteins while Se in SeAlb occurs unspecifically as selenomethionine
(SeMet) by substitution of a methionine [214].
SelP is the most abundant plasma selenoprotein and contains more than 50% of the total
plasma Se [24]. Although its expression is observed in many tissues, SelP is mainly
secreted by the liver and can contain up to 10 SeCys residues [69]. It is mainly responsible
for the transport of Se from the liver to peripheral tissues [64, 66] especially the brain and
testis where specific receptors were recently identified [67, 68]. Further, SelP may also
exhibit antioxidant functions, thereby protecting neuronal and astrocyte cells [215] and
inhibiting the oxidation of low density lipoproteins [216]. GPx3 is secreted by kidneys
[217] and plays a major role in the detoxification of extracellular hydrogen peroxides and
fatty acid hydroperoxides [54]. It is also expressed in other tissues including the heart,
where it is the third selenoprotein in terms of mRNA abundance [53]. SeAlb has no known

39
Se dependent biological function; it likely represents a form of Se storage which can be
used by the liver to synthetize SelP [218].
Se is exceptionally abundant in certain traditional marine foods of Inuit and accordingly, Se
intake in this population is among the highest in the world [198]. However, some marine
foods also contain high concentrations of methylmercury (MeHg) which is bioaccumulated
by marine mammals and predatory fish species in the Arctic [198, 200]. MeHg with its
capacity to cross the blood-brain and placental barriers and its gastrointestinal absorption
near to 100% is a neurotoxic threat for Arctic residents [219]. This toxicity can be
influenced by certain elements present in foods which may offset some of the deleterious
effects of MeHg [220]. Data from several epidemiological studies suggest that an adequate
Se intake could play a protective role against the toxic effects of MeHg [208, 221], to
which the Inuit population is highly exposed. Several researchers have hypothesized that Se
is directly linked to the demethylation of MeHg and that granules of HgSe would be the
final product of this degradation [177]. These granules have already been identified in
marine mammals and are located in the cytoplasm of macrophages in the liver [190, 191].
However in other populations, an elevated Se status has been recently associated to type 2
diabetes, hypercholesterolemia and/or hypertension [8, 155]. Additionally, in some
seleniferous areas in China (County of Enshi, Hubei Province) where selenosis symptoms
were observed in the population, SeMet was found to be the main Se species involved
[222].
Typically, the Se status is assessed by measuring the Se content of the plasma/serum or
blood. To prevent human toxicity, the U.S. Environmental Protection Agency set Se blood
levels corresponding to the no observable adverse effect level (NOAEL) and the lowest
observable adverse effect level (LOAEL) at 1000 µg L-1 and 1350 μg L-1 respectively. At
the opposite, deficiency occurs when plasma Se status is below 70-90 μg L-1 [223].
All these findings suggest that, to better understand the impacts of high Se status on human
health of Inuit, new biomarkers other than the commonly measured total plasma Se must be
studied. The circulating levels of plasma selenoproteins may help to better characterize the

40
Se status and largely improve our capacity to identify the risks and benefit linked to the
exceptionally high Se status of the Inuit population.
Here, we adapted in our laboratory the method of Li et al (2011) combining affinity column
(AF) chromatography and inductively coupled plasma mass spectrometry (ICP-MS) with
post-column isotope dilution (ID-AF-ICP-MS) to determine plasma levels of GPx3, SelP
and SelAlb in a representative sample of the Inuit population of Nunavik, in order to better
assess their Se status.
2. Experimental
2.1. Study population
In the fall of 2004, a health survey was conducted among the 14 communities of Nunavik
living along the coasts of the East Hudson Bay, Hudson Straight and Ungava Bay. The
Nunavik territory covers a third of the total surface area of Quebec (Canada) and is home to
approximately 11000 Inuit [196]. The aim of this study was to gather social and health
information for this population and verify the evolution of its health status since the last
survey conducted in 1992 [224]. The targeted individuals were the permanent Inuit
residents of Nunavik aged between 18 and 74 years. The study design and the sampling
procedure were published elsewhere [210]. Briefly, a two-stage stratified random sampling
method was used. In the first stage, a proportional random sample selection of private Inuit
households was carried out, taking into account the size of each village. In the second
stage, eligible members of each household were asked to participate. Participants were
invited to fill out several questionnaires and attend a clinical session during which blood
samples and different clinical and anthropometric measurements were collected. The
overall participation rate was 50% and a total of 889 participants completed the clinical
session.
Our study sample consisted of 852 participants from whom plasma samples were available.
After blood collection and centrifugation, plasma was isolated then aliquoted in Sarstedt

41
(Nümbrecht, Germany) 1.5 mL tubes and stored at -80 °C at the Centre de Toxicologie du
Québec (CTQ) of the Institut National de Santé Publique du Québec, Canada (INSPQ).
2.2. Reagents and chemicals
Ammonium acetate (ACS grade) was purchased from Anachemia (Montreal, QC, Canada),
while methanol and ethanol (Omnisolv grade) were obtained from EMD (Omaha, NB) and
Commercial Alcohols (Brampton, ON, Canada) respectively. The enriched solution of 77Se
(98%) was obtained from Cambridge Isotope Laboratories (Andover, MA) and 204 Hg
(98%) from Isoflex (San Francisco, CA). Milli-Q water was purified by the advantage A10
ultrapure water system (Merck Millipore, Billerica, MA).
Two Se certified human serum reference materials (CRM) (BCR-637 and SRM-1950) were
used throughout. The SRM-1950 was purchased from the National Institute of Standards
and Technology (NIST, Gaithersburg, MD, USA) and the BCR-637 from the Institute for
Reference Materials and Measurements (Geel, Belgium). Proficiency testing material
(PTM) (QMEQAS 11-S-04) with a consensus value for Hg and Se were also used; they
were obtained from the quality assessment scheme for trace elements operated by the
INSPQ.
2.3. Instrumentation
The complete chromatography separation of the three selenoproteins was carried out using
an Acquity ultra performance liquid chromatography system (UPLC) (Waters, Milford,
MA) equipped with two 1-mL affinity columns heparin-sepharose and blue-sepharose, both
purchased from GE Healthcare (Uppsala, Sweden). The columns were connected to a six-
way Rheodyne automated switching valve (Model MXP9900-000; Oak Harbor, WA).
A NexION 300s ICP-MS (Perkin-Elmer, Waltham, MA) was coupled to the UPLC system
for Se and Hg detection and quantification by post-column isotope dilution. A daily
performance solution containing 1 µg L-1 of Be, Ce, Co, Fe, In, Li, Mg, Mn, Pb and U in
1% (v/v) HNO3 was used for instrument optimization. The kinetic energy discrimination

42
mode (KED) in the collision cell using H2 gas was employed to eliminate the polyatomic
interferences upon detection of both Se and Hg. The isotope solution containing 77Se and
204Hg was continuously infused using the main ICP-MS peristaltic pump and mixed to the
UPLC column outflow through a T connector. UPLC and ICP-MS operating conditions are
presented in Table 3.
2.4. Analytical procedures
2.4.1. Speciation analysis
The method was adaptated from the previous published work of Li et al [211].Plasma
samples were injected through the UPLC system following the conditions described in
table 3 and schematic representation of the procedure is given by Figure 10. While the
switching valve was in position 1, the flow passed through both heparin and blue sepharose
columns using the mobile phase A. The Heparin-sepharose column has the ability to retain
selectively the SelP while the blue-sepharose can retain both SelP and SeAlb. At this stage,
the SelP and SeAlb were retained by heparin and blue sepharose columns respectively
while the GPx and small Se compounds were the first eluted. Switching the valve to
position 2 excluded the blue-sepharose column and permitted the elution of the SelP using
mobile phase B. Finally the system was switched back to position 1 to elute SeAlb using
the mobile phase B.
Post-column isotope dilution was performed using two stock solutions of 77Se and 204Hg
prepared as follows: 4.92 mg of 77Se was weighed and dissolved in 10 mL of 5% (v/v)
HNO3; and 1 mg of 204Hg was dissolved in 5 mL of 10% (v/v) HNO3 then completed to
100 mL with 10% (v/v) HNO3 containing 1 mL of Au at 1000 mg L-1. Using the previous
stock solutions, a daily isotope solution was prepared for speciation analysis in 0.5 % (v/v)
HNO3 containing 2.95 µg L-1 of 77Se, 5 µg L-1 of 204Hg and 8% (v/v) MeOH.
ICP-MS data were treated with the IDSoft post column ID software [225]. After the input
of all ID parameters including the isotopic abundance, detector dead time and mass bias,

43
(78Se/77Se) and (200Hg/204Hg) chromatographic ratios were transformed to mass flow charts
and the integration of peaks yielded concentrations.
2.4.2. Blood Se and Hg concentrations analysis
The measurement of whole blood Se and Hg concentrations was performed at the
toxicology laboratory of INSPQ and the complete analytic method is detailed elsewhere
[226].
2.5. Statistical analysis
Descriptive statistics were used to present the distributions of blood Hg, Se and plasma Hg,
Se biomarkers. Normality was tested for both Se and Hg: Se in plasma follows a normal
distribution and mean concentrations were used for comparison. In contrast, blood Hg and
blood Se follow a log-normal distribution and accordingly medians were used. The
relationship between plasma and blood Se was tested using the Pearson correlation
coefficient.
3. Results
Figure 11 shows a mass flow chromatogram obtained after the complete separation of
selenoproteins by ID-AF-ICP-MS. The speciation time analysis was 20 min and the three
plasma Se-containing proteins (GPx3, SelP and SeAlb) were well separated. As shown in
Figure 11, there was a Hg signal associated with each Se-containing protein. To quantify
both Se and Hg, (78Se/77Se) and (200Hg/204Hg) ratios were used for isotope dilution analysis
respectively. Data on the analytic performance of the method are presented in table 4. For
Se-containing protein quantification, the limits of detection (LODs) were 0.70, 1.10 and
2.20 µg Se L-1 for GPx3, SelP and SeAlb respectively. The repeatability in terms of relative
standard deviation (RSD %) was 5.84% for GPX3, 3.36% for SelP and 2.65% for SeAlb.
Both SRM-1950 and BCR-637 had a certified value for total plasma Se and the method

44
shows a LOD of 3.71 µg L-1 and between-day precisions of 2.30% and 3.22% for SRM-
1950 and BCR-637 respectively. The quantification of Hg concentration associated with
Se-containing proteins was validated through a PTM with a consensus value for total
plasma Hg. The LODs were 0.02 µg L-1 for all Hg associated to selenoproteins (Hg-GPx3,
Hg-SelP and Hg-SeAlb) and 0.04 µg L-1 for the total plasma Hg concentration. The
repeatability was 3.49% for total plasma Hg and 19.14%, 7.15% and 19.37% for Hg
associated to GPx3, SelP and SeAlb respectively.
Results from the blood Se analysis and application of the adapted method for Se speciation
on 852 samples from Inuit adults are presented in table 5. High concentrations of blood Se
are observed in this population (mean = 350 µg L-1) and 50% of participants had a blood Se
level ≥ 261 µg L-1 (interquartile range (IQR) = 166). The maximum and minimum
concentrations registered for blood Se were 3355 µg L-1 and 119 µg L-1 respectively. Total
plasma Se concentrations were lower than the blood Se levels; concentrations ranged
between 84.5 to 229 µg L-1 (mean = 140 µg L-1) with a median of 139 µg L-1 (IQR =22.7).
Speciation analysis shows that 52% of total plasma Se was in the form of SelP (mean =
72.4 µg Se L-1) while GPx3 (mean = 35.6 µg Se L-1) and SeAlb (mean = 32.6 µg Se L-1)
represented 25% and 23% of total plasma Se, respectively.
Results of Hg analysis in both blood and plasma are shown in table 6. As expected, blood
Hg levels were higher than corresponding plasma levels in our participants (median in
blood = 11.8 µg L-1, range = 0.08 to 241 µg L-1 vs median in plasma = 1.78µg L-1, range =
0.02 to 24.5 µg L-1). Similarly to Se, more than half of total plasma Hg was associated to
SelP (mean = 1.46 µg L-1) and the rest was shared equally between GPx3 (mean = 0.70 µg
L-1) and SeAlb (mean =0.55 µg L-1).
The relationship between total blood and plasma Se concentrations in this population was
found to be non-linear (r = 0.33, p < 0.001) (Figure12). Plasma Se levels reach a plateau at
approximately 175 µg L-1 while blood Se continues to increase.

45
4. Discussion
4.1. Method development and validation
In this study, we developed a method for the simultaneous quantification of Se proteins in
human plasma and associated Hg content by ID-AF-ICP-MS. Although several methods
were published using the same approach for Se proteins [211, 227, 228], none of them
allowed to simultaneously quantify Se proteins and Hg. The use of H2 gas with KED mode
in the collision cell, permitted to eliminate argon (Ar) induced polyatomic interferences
(Ar38Ar40) on isotope 78Se. We selected post-column isotope dilution for quantification
because standards for selenoproteins were unavailable. Moreover, this method provides the
most accurate results for Se quantification when performed correctly [229]. Indeed, the
accuracy and repeatability of our method for both CRMs and PTM were excellent (less
than 5% in overall for total Se and Hg). We also investigated the possibility of using
standards which were provided in some ELISA kits to elaborate the standard curves, but no
Se was detected when analyzed by ICP-MS in those standards as they were fragments of
recombinant selenoproteins. Therefore, the validation process was also affected by the lack
of selenoprotein standards so that the evaluation of matrix effects and recovery could not be
performed.
4.2. Biomarkers of Se and Hg status among the Inuit of Nunavik
Our study is the first to conduct plasma Se speciation in a population with a very high
selenium status and also the largest one in terms of participant number (n = 852). To our
knowledge, this is the second population study focusing on the Se speciation in an
apparently healthy population, following that conducted in 2010 by Letsiou et al. in the
Greek population [230].
Mean plasma Se concentration, which represents the biomarker of Se status most often
used, was 140 µg L-1, with values ranging from 84.5 to 229 µg L-1. The mean concentration
in Inuit was similar to those reported in general populations of North America (USA mean

46
= 136.4 µg L-1)[149], and higher than those found in UK (mean = 85.3 µg L-1) [231],
Russia (mean = 93.6 µg L-1)[232], Korea (mean = 112 µg L-1) [233], Australia (mean = 103
µg L-1) [234], Japan (mean = 119.8 µg L-1) [235], France (mean = 88.0 µg L-1) [236] and
even in Finland (mean = 110.5 µg L-1) where soils are fertilized with selenium since 1984
[237]. The 25th and 75th percentiles of plasma Se concentrations were 128 and 151 µg L-1
respectively, indicating a very narrow distribution of Se plasma levels (IQR =22.7). With
regard to plasma Se distribution in selenoproteins, SelP contained 52% of total plasma Se
while GPx3 and SeAlb represented 25 and 23% respectively. These results are in
agreement with those noted in the Greek population, except for SeAlb which represented
only 11% of total plasma Se (25% in GPx3, 53% in SelP, mean plasma Se = 91 µg L-1, n =
399). The variation of results for SeAlb can be explained by the difference in SeMet status
between the populations, since Se in SeAlb is mainly incorporated unspecifically in the
form of SeMet substituting for Met [238]. Moreover, our results were also in complete
agreement with those reported in previous published studies involving a limited number of
healthy participants, using the same analytical method, in which Se status was above 100
µg L-1. Jitaru et al (2008) found Se incorporated at 23%, 56% and 20% in GPX3, SelP and
SeAlb respectively (plasma Se mean =110 µg L-1, n=3) [239] while Reyes et Al (2003)
found 19%, 55% and 15% (plasma Se mean =100 µg L-1, n=5) [240]. However, when
comparing our results to those in which Se status was relatively low (control groups in
Roman et al studies (2010) mean Se = 85 µg L-1, n = 15) results were inconsistent and the
percentage of Se-GPx3 tends to decrease with respect to SelP which remains the dominant
Se-containing protein [241]. Furthermore, more than 75% of our participants exhibited
plasma Se levels above 125 µg L-1, a Se status high enough to maximize activities and
concentrations of selenoproteins [242]. These findings taken together support the existence
of a regulation mechanism for plasma Se status. At high Se levels where selenoproteins
activity and concentration are maximized, little variation of Se distribution among Se-
containing proteins can be observed between populations. In contrast, variation may occur
at low Se status as GPx3 is known to be more sensitive to Se status, in contrast to SelP
which ensure the delivery of Se to organs and therefore it is less sensitive to the Se status
[243].

47
Blood Hg levels in Nunavik (median = 11.2 µg L-1 IQR=17.9) is the highest when
compared to other populations in Canada (median = 0.81 µg L-1, IQR =1.28) [244], USA
(median = 0.64 µg L-1, Q1< LOD Q3 = 2.87 µg L-1) [245], Korea (median = 3.37 µg L-1,
IQR = 3) [246], Hong Kong (median = 4.93 µg L-1, IQR = 3.23) [247] and Norway
(median = 4.0 µg L-1) [248]. However, it remains lower than levels founds in population of
both Brazilian Amazon (median= 57.2 µg L-1, IQR = 55.9) [249] and Inuit of Greenland
(median=18 µg L-1, IQR = 25.3) [250].This elevated blood Hg comes mostly from the
consumption of marine mammals which bioaccumulate MeHg in their tissues [200].
Lemire et al (2012) previously reported that beluga meat constitutes the most important
source of MeHg, contributing up to two thirds of MeHg intake in this population [198].
Blood Se concentrations are exceptionally high in this population (median =261 µg L-1,
IQR=166 µg L-1), with values comparable to those of the Inuit of Greenland (GM = 285.11
range = 68 to 5600 µg L-1) [250] and populations in the Brazilian Amazon (median = 284.3
μg L-1, IQR=161.9 µg L-1) [249]. However, when compared to general populations
including Canadians (median = 199 µg L-1, IQR=36.7 µg L-1) [251], Americans (median =
189 μg L-1, IQR= 31.3 µg L-1) [252] and other populations in Europe [253, 254], blood Se
levels in Nunavik are much higher. These high blood Se levels are also related to the
consumption of traditional marine foods in Nunavik. Beluga mattaaq, seal liver and walrus
meat were found to be the most important sources of Se in this population [198].
In this study, Hg was monitored and quantified simultaneously along with Se speciation.
Indeed, our results show that plasma Hg accounted for 15% of blood Hg (mean = 2.59 µg
L-1). In addition, this plasma Hg was found to be associated with the three peaks of Se
proteins (Figure 12). Having said that, the first peak includes all unretained molecules and
therefore Hg may not be associated to GPx3. However, Hg appears to bind to both SelP and
SeAlb since it is retained and eluted with these Se-containing proteins selectively.
Furthermore, our findings show that more than 50% of plasma Hg coeluted with SelP.
Likewise, Chen et al (2006) have shown that SelP binds Hg much more at higher Hg
exposure [255]. The binding capacity of Hg to these proteins is explained by its strong
affinity for selenol (Se-H) groups present in these Se-containing proteins [177, 178]. Thus,

48
given that SelP represents the major selenoprotein in plasma which may contain up to 10
residues of SeCys [69], it is likely the major Hg binding of all Se-containing proteins in
plasma.
Despite the high blood Se levels found in Nunavik, mean plasma Se concentration was not
exceptionally elevated and did not exceed 229 µg L-1. We observed a nonlinear relationship
blood and plasma Se levels in this population (r = 0.33, p < 0.001), with plasma Se reaching
a plateau at approximately 175 µg L-1 (Firgue 13). A similar relationship was previously
reported in Inuit of Greenland by Hansen et al in 2004 [256]. In contrast, Lemire et al
(2012) found a linear relationship between plasma and blood Se levels in populations living
in the Brazilian Amazon [223]. This may be explained by the fact that the main dietary
source of Se in the latter populations is Brazil nuts (Bertholletia excelsa), in which Se is
mainly present in the form of SeMet [257]. As mentioned previously, SeMet is
incorporated in albumin in lieu of methionine to form SeAlb [238]. Therefore in this
population, the increase of Se in whole blood is accompanied by a proportional increase in
plasma due to the fact that it is incorporated in Alb. Moreover, Li et al (2011) in their
supplemental study, using Se-enriched yeast (SelenoPrecise, Pharma Nord, Denmark) in
which SeMet is the main form of Se [258], have shown that plasma SeAlb was the most
increased of all Se-containing proteins after one year follow-up [211]. In Inuit populations
of Greenland and Nunavik, the main dietary sources of Se are likely marine mammal
tissues, but little information is available on Se species present in marine foods. Lemes et al
(2011) reported that organs from beluga whales contained methylselenocysteine (MeSecys)
as a major selenocompound [259]; however unknown Se species were detected and
remained to be identified.
Despite the lack of data on Se speciation in marine foods consumed by the Inuit, several
lines of evidence suggest that selenoneine might be a major selenocompound in these
foods. Yamashita et al have recently isolated from blue fin tuna blood an organic form of
Se identified as selenoneine, an analog of ergothioneine [40]. Selenoneine was shown to be
present in various fish species and a major form of selenium in blood cells obtained from a
fish-eating population in Japan [41]. Selenoneine is actively transported in red blood cells

49
via the OCTN1 membrane transporter, similarly to ergothioneine [260]. Such an active
transport of selenoneine in red blood cells of Inuit would explain the much higher Se
concentrations in whole blood compared to plasma in the present study. Confirmation of
selenoneine presence awaits the synthesis of the native compound and the isotope labeled
analog for definitive identification and quantification in the blood of Inuit.
5. Conclusion
In this study an analytic method for plasma Se speciation and quantification was developed
in order to document the levels of Se proteins in the Inuit population of Nunavik that
exhibit one of the highest Se status in the world. Our results indicate that SelP was the
predominant selenoprotein incorporating more than 50% of total plasma Se of the Inuit of
Nunavik. This selenoprotein appears to bind most Hg present in plasma. The non-linear
relationship between plasma and whole blood Se concentrations suggests the accumulation
of a selenocompound, possibly selenoneine, in red blood cells of Inuit. Future studies will
investigate Se species in red blood cells of Inuit and in marine foods rich in the element.
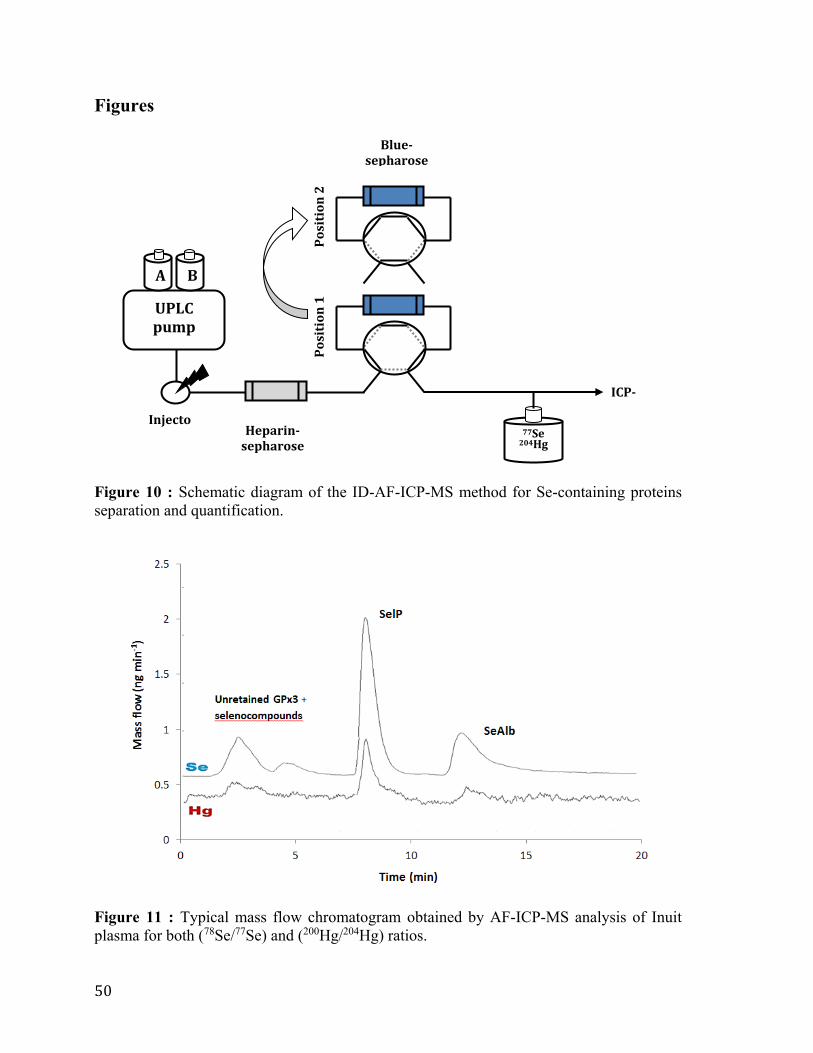
50
Figures
Figure 10 : Schematic diagram of the ID-AF-ICP-MS method for Se-containing proteins
separation and quantification.
Figure 11 : Typical mass flow chromatogram obtained by AF-ICP-MS analysis of Inuit
plasma for both (78Se/77Se) and (200Hg/204Hg) ratios.
ICP-MS
77Se 204Hg
UPLC pump
Heparin-sepharose
column
Blue-sepharose
column
Po
siti
on
1
Po
siti
on
2
B A
Injector
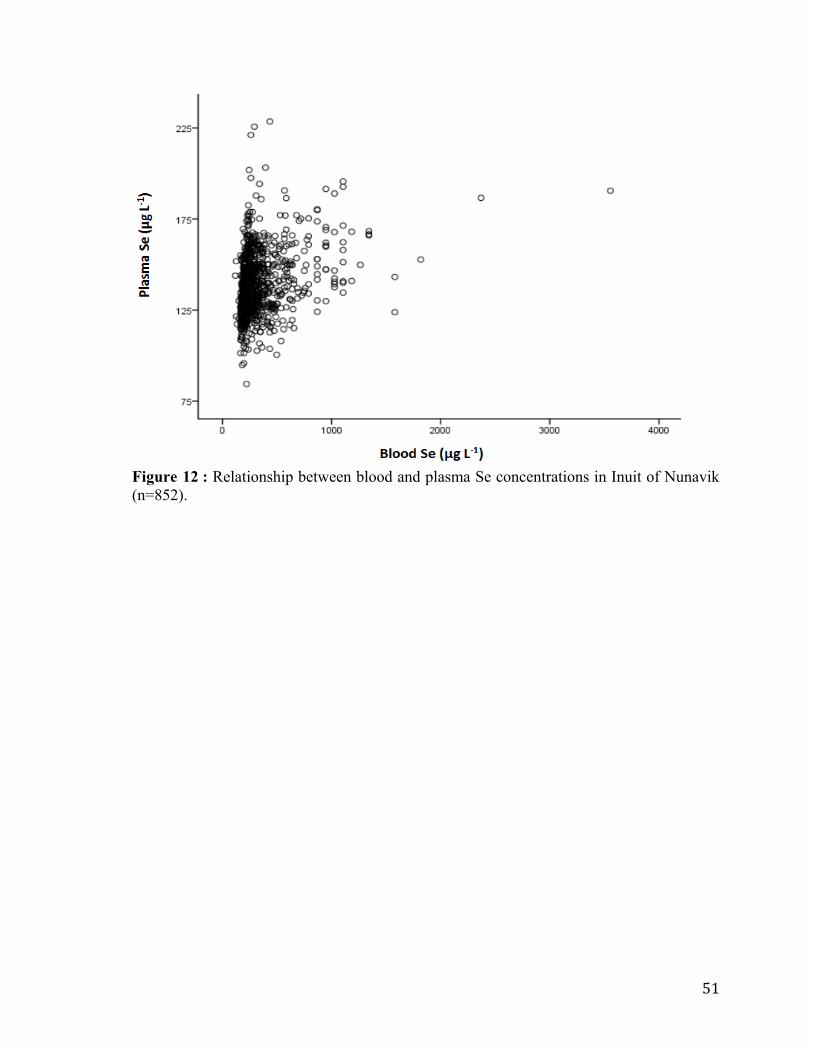
51
Figure 12 : Relationship between blood and plasma Se concentrations in Inuit of Nunavik
(n=852).

52
Tables
Table 3 : Operating conditions for the liquid chromatography (LC) and ICP-MS
instruments.
ICP-MS
Rf Power 1600w
Plasma Ar flow rate 18 L min-1
Auxiliary Ar flow rate 1.2 L min-1
Nebulizer Ar flow rate 1.06 L min-1
H2 flow rate in the collision cell 3 L min-1
Scan mode Peak Hopping
Dwell time (speciation analysis) 113 ms
Isotopes monitored 76 Se, 77 Se, 78 Se, 82 Se, 200Hg, 203 Hg and 204Hg
Isotope solution flow rate 0.207 g min-1
LC
Columns Heparin Sepharose and Blue Sepharose (1 mL
each)
Flow rate 0.8 ml min-1
Injection volume 100 µL
Mobile phase A (binding buffer) 0.05 mol L-1 ammonium acetate, , pH 7
Mobile phase B (elution buffer) 1.5 mol L-1 ammonium acetate, , pH 7
Gradient and valve position 1-6 min 100% A + position 1
6-10 min 100% B + position 2
10-20 min 100% B + position 1
20-25 min 100% A + position 1
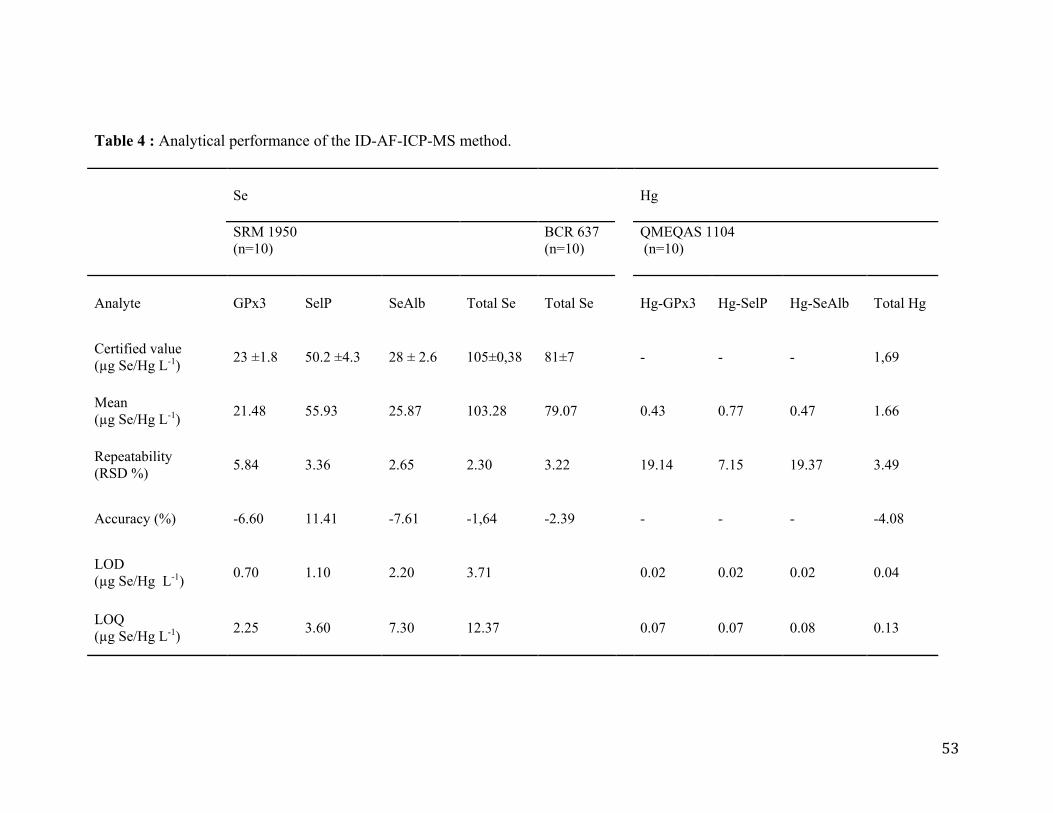
53
Table 4 : Analytical performance of the ID-AF-ICP-MS method.
Se
Hg
SRM 1950
(n=10)
BCR 637
(n=10)
QMEQAS 1104
(n=10)
Analyte GPx3 SelP SeAlb Total Se Total Se Hg-GPx3 Hg-SelP Hg-SeAlb Total Hg
Certified value
(µg Se/Hg L-1) 23 ±1.8 50.2 ±4.3 28 ± 2.6 105±0,38 81±7 - - - 1,69
Mean
(µg Se/Hg L-1) 21.48 55.93 25.87 103.28 79.07 0.43 0.77 0.47 1.66
Repeatability
(RSD %) 5.84 3.36 2.65 2.30 3.22 19.14 7.15 19.37 3.49
Accuracy (%) -6.60 11.41 -7.61 -1,64 -2.39 - - - -4.08
LOD
(µg Se/Hg L-1) 0.70 1.10 2.20 3.71
0.02 0.02 0.02 0.04
LOQ
(µg Se/Hg L-1) 2.25 3.60 7.30 12.37
0.07 0.07 0.08 0.13
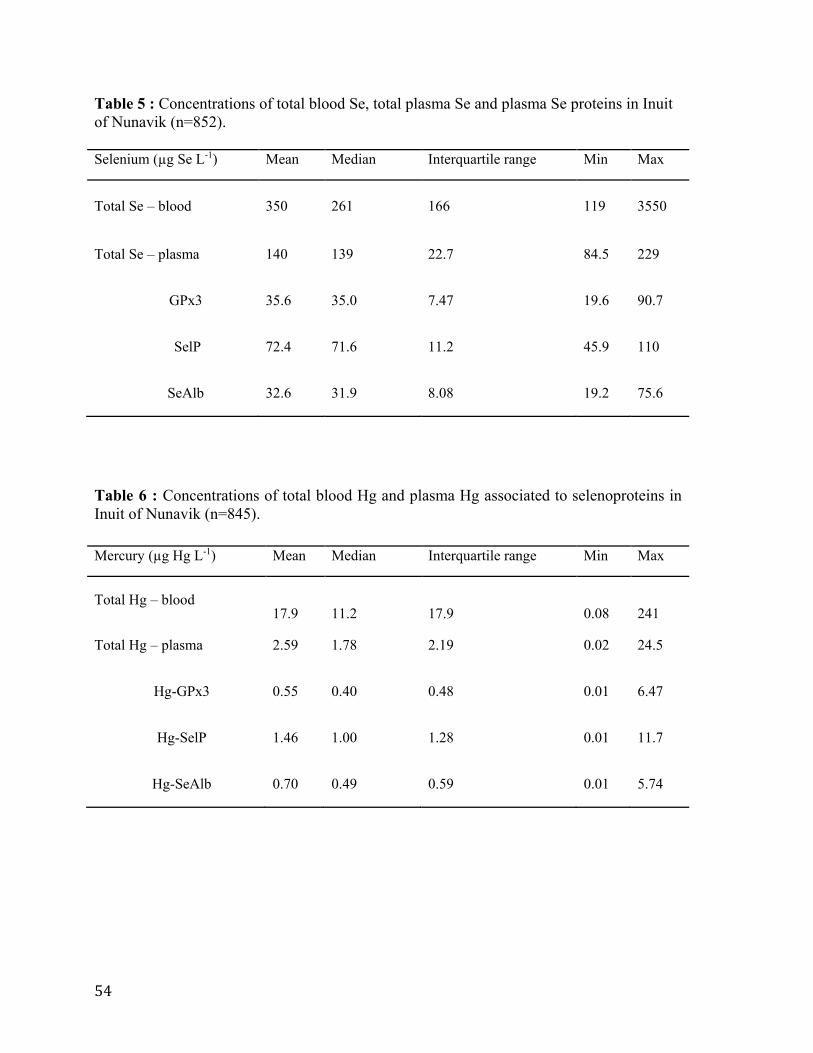
54
Table 5 : Concentrations of total blood Se, total plasma Se and plasma Se proteins in Inuit
of Nunavik (n=852).
Selenium (µg Se L-1) Mean Median Interquartile range Min Max
Total Se – blood 350 261 166 119 3550
Total Se – plasma 140 139 22.7 84.5 229
GPx3 35.6 35.0 7.47 19.6 90.7
SelP 72.4 71.6 11.2 45.9 110
SeAlb 32.6 31.9 8.08 19.2 75.6
Table 6 : Concentrations of total blood Hg and plasma Hg associated to selenoproteins in
Inuit of Nunavik (n=845).
Mercury (µg Hg L-1) Mean Median Interquartile range Min Max
Total Hg – blood 17.9 11.2 17.9 0.08 241
Total Hg – plasma 2.59 1.78 2.19 0.02 24.5
Hg-GPx3 0.55 0.40 0.48 0.01 6.47
Hg-SelP 1.46 1.00 1.28 0.01 11.7
Hg-SeAlb 0.70 0.49 0.59 0.01 5.74

55
Chapitre 3 : Conclusion générale
L’objectif principal de cette étude était de vérifier les niveaux circulants des
sélénoprotéines et du mercure associé dans le plasma d’adultes inuits du Nunavik, chez qui
le statut en sélénium et les concentrations du mercure sont élevés. Pour ce faire, une
méthode analytique combinant une séparation par chromatographie d’affinité couplée à la
spectrométrie de masse à plasma induit avec une quantification par dilution isotopique a été
développée. L’application de cette méthode à l’ensemble des échantillons plasmatiques
nous a permis de quantifier avec précision les concentrations des sélénoprotéines et d’avoir
une meilleure évaluation du statut en sélénium chez les Inuits du Nunavik. De plus, la
quantification du signal du mercure associé aux sélénoprotéines nous a permis de vérifier la
possibilité d’une interaction entre ces deux entités.
Ainsi, les espèces plasmatiques de sélénoprotéines (GPx3, SelP et SeAlb) ont été séparées
et quantifiées dans 852 échantillons. Nos résultats indiquent que le sélénium plasmatique,
en moyenne chez les Inuits du Nunavik, est incorporé à plus de 50% dans la SelP alors que
le reste est partagé entre les deux autres sélénoprotéines. Cette distribution du sélénium
n’était pas différente de celles retrouvées dans d’autres populations dont le statut en
sélénium était comparable. Ceci indique que les sélénoprotéines sont maintenues à un
équilibre lorsque le statut en sélénium plasmatique est élevé, à l’exception de la SeAlb qui
varie selon le statut de la SeMet.
Nous avons pu aussi quantifier le mercure plasmatique qui représentait 15% du mercure
total. Ce mercure n’était pas totalement libre dans le plasma et était associé aux pics de
sélénoprotéines lors de la séparation chromatographique. Ces résultats suggèrent que le
mercure se lie aux sélénoprotéines plasmatiques, probablement par affinité aux
groupements sélénols. Toutefois, cette liaison diminue la biodisponibilité du mercure mais
ne permet pas de prouver un quelconque antagonisme entre le mercure et les
sélénoprotéines.

56
Les analyses sanguines effectuées nous ont permis de comparer les niveaux sanguins du
sélénium avec ceux du plasma. Nous avons été surpris de l’écart existant entre les valeurs
sanguines et plasmatiques dans cette population. En effet, nos résultats démontrent que la
relation entre le sélénium plasmatique et sanguin était non-linéaire et que le sélénium dans
le sang continuait à augmenter alors qu’il atteignait un plateau dans le plasma. Cette
observation était semblable à ce qui a été rapporté par Hansen et al en 2004 chez les Inuits
du Groenland alors que chez les Brésiliens d’Amazonie, une relation linéaire était observée.
Nous pensons que cette différence entre les Inuits et les Brésiliens réside dans l’espèce de
sélénium consommée. De ce fait, nous supposons que l’alimentation traditionnelle des
Inuits contient une ou plusieurs formes de sélénium qui sont redirigées vers les cellules
sanguines où elles sont stockées. De plus, on se basant sur les données de Yamashita, nous
supposons aussi que le sélénium dans les cellules sanguines est sous forme de sélénonéine.
Enfin, au terme de cette étude il devient clair de mentionner que pour mieux caractériser le
statut en sélénium dans une population comme celle des Inuits du Nunavik, une spéciation
dans la fraction cellulaire sanguine s’impose. Cette dernière va nous permettre de mieux
comprendre l’effet que peut avoir une alimentation riche en sélénium sur la santé des Inuits
et donnera plus de détails quant aux interactions que peut avoir le sélénium avec les
différents contaminants environnementaux, principalement le mercure.

57
Références:
[1]. Barceloux, D.G. and D. Barceloux, Selenium. Clinical Toxicology, 1999. 37(2): p.
145-72.
[2]. Nelson, A.A., O.G. Fitzhugh and H.O. Calvery, Liver tumors following cirrhosis
caused by selenium in rats. Cancer Research, 1943. 3(4): p. 230-36.
[3]. Schwarz, K. and C.M. Foltz, Selenium as an integral part of factor 3 against dietary
necrotic liver degeneration. Journal of the American Chemical Society, 1957. 79(12): p.
3292-93.
[4]. Rotruck, J.T., et al., Selenium: biochemical role as a component of glutathione
peroxidase. Science, 1973. 179(4073): p. 588-90.
[5]. Flohe, L., W.A. Gunzler and H.H. Schock, Glutathione peroxidase: a selenoenzyme.
FEBS Lett, 1973. 32(1): p. 132-34.
[6]. Loscalzo, J., Keshan disease, selenium deficiency, and the selenoproteome. N Engl
J Med, 2014. 370(18): p. 1756-60.
[7]. Cheng, Y.Y. and P.C. Qian, The effect of selenium-fortified table salt in the
prevention of Keshan disease on a population of 1.05 million. Biomed Environ Sci, 1990.
3(4): p. 422-28.
[8]. Rayman, M.P., Selenium and human health. Lancet, 2012. 379(9822): p. 1256-68.
[9]. Wen, H. and J. Carignan, Reviews on atmospheric selenium: Emissions, speciation
and fate. Atmospheric Environment, 2007. 41(34): p. 7151-65.
[10]. ATSDR Agency For Toxic Substances And Disease Registry, A.G.U.S.,
Toxicological profiles for selenium. 2003, [http://www.atsdr.cdc.gov/toxprofiles/tp92.pdf
Last accessed: December 15, 2014].
[11]. Liu, G., et al., Comparative on causes and accumulation of selenium in the tree-
rings ambient high-selenium coal combustion area from Yutangba, Hubei, China. Environ
Monit Assess, 2007. 133(1-3): p. 99-103.
[12]. Schutz, D.F. and K.K. Turekian, The investigation of the geographical and vertical
distribution of several trace elements in sea water using neutron activation analysis.
Geochimica et Cosmochimica Acta, 1965. 29(4): p. 259-313.
[13]. Alexander, J., Chapter 52 - Selenium, in Handbook on the Toxicology of Metals
(Fourth Edition). 2015: San Diego. p. 1175-208.
[14]. Gore, F., J. Fawell and J. Bartram, Too much or too little? A review of the
conundrum of selenium. J Water Health, 2010. 8(3): p. 405-16.
[15]. WHO, Guidelines for drinking-water quality. 2011, Geneva: World Health
Organization. 564.
[16]. Johnson, C.C., F.M. Fordyce and M.P. Rayman, Symposium on 'Geographical and
geological influences on nutrition': Factors controlling the distribution of selenium in the
environment and their impact on health and nutrition. Proc Nutr Soc, 2010. 69(1): p. 119-
32.
[17]. Reilly, C. and S.E.L. SpringerLink, Selenium in food and health. 2006, New York.
[18]. Floor, G.H. and G. Romà N-Ross, Selenium in volcanic environments: A review.
Applied Geochemistry, 2012. 27(3): p. 517-31.
[19]. Brown, K.M. and J.R. Arthur, Selenium, selenoproteins and human health: a review.
Public Health Nutr, 2001. 4(2B): p. 593-99.

58
[20]. Kryukov, G.V., et al., Characterization of mammalian selenoproteomes. Science,
2003. 300(5624): p. 1439-43.
[21]. Bock, A., et al., Selenocysteine: the 21st amino acid. Mol Microbiol, 1991. 5(3): p.
515-20.
[22]. Rayman, M.P., The importance of selenium to human health. The Lancet, 2000.
356(9225): p. 233-41.
[23]. Cone, J.E., et al., Chemical characterization of the selenoprotein component of
clostridial glycine reductase: identification of selenocysteine as the organoselenium moiety.
Proc Natl Acad Sci U S A, 1976. 73(8): p. 2659-63.
[24]. Labunskyy, V.M., D.L. Hatfield and V.N. Gladyshev, Selenoproteins: molecular
pathways and physiological roles. Physiol Rev, 2014. 94(3): p. 739-77.
[25]. Dolph L. Hatfield, I.S.C.T., Selenocysteine tRNA[Ser]Sec isoacceptors as central
components in selenoprotein biosynthesis in eukaryotes, in Selenium in Biology and
Human Health. 1994, Springer New York. p. 25-44.
[26]. Hatfield, D.L., et al., Selenocysteine incorporation machinery and the role of
selenoproteins in development and health. Prog Nucleic Acid Res Mol Biol, 2006. 81: p.
97-142.
[27]. Lee, B.J., et al., Identification of a selenocysteyl-tRNA(Ser) in mammalian cells that
recognizes the nonsense codon, UGA. J Biol Chem, 1989. 264(17): p. 9724-27.
[28]. Xu, X.M., et al., Biosynthesis of selenocysteine on its tRNA in eukaryotes. PLoS
Biol, 2007. 5(1): p. e4.
[29]. Yuan, J., et al., RNA-dependent conversion of phosphoserine forms selenocysteine
in eukaryotes and archaea. Proc Natl Acad Sci U S A, 2006. 103(50): p. 18923-27.
[30]. Guimaraes, M.J., et al., Identification of a novel selD homolog from eukaryotes,
bacteria, and archaea: is there an autoregulatory mechanism in selenocysteine metabolism?
Proc Natl Acad Sci U S A, 1996. 93(26): p. 15086-91.
[31]. Berry, M.J., et al., Recognition of UGA as a selenocysteine codon in type I
deiodinase requires sequences in the 3' untranslated region. Nature, 1991. 353(6341): p.
273-76.
[32]. Seeher, S., Y. Mahdi and U. Schweizer, Post-transcriptional control of selenoprotein
biosynthesis. Curr Protein Pept Sci, 2012. 13(4): p. 337-46.
[33]. Chavatte, L., B.A. Brown and D.M. Driscoll, Ribosomal protein L30 is a
component of the UGA-selenocysteine recoding machinery in eukaryotes. Nat Struct Mol
Biol, 2005. 12(5): p. 408-16.
[34]. Miniard, A.C., et al., Nucleolin binds to a subset of selenoprotein mRNAs and
regulates their expression. Nucleic Acids Res, 2010. 38(14): p. 4807-20.
[35]. Budiman, M.E., et al., Eukaryotic initiation factor 4a3 is a selenium-regulated RNA-
binding protein that selectively inhibits selenocysteine incorporation. Mol Cell, 2009.
35(4): p. 479-89.
[36]. Schrauzer, G.N., Selenomethionine: a review of its nutritional significance,
metabolism and toxicity. J Nutr, 2000. 130(7): p. 1653-56.
[37]. Schrauzer, G.N., The nutritional significance, metabolism and toxicology of
selenomethionine. Adv Food Nutr Res, 2003. 47: p. 73-112.
[38]. Qawwad, H.K., et al., Se75-selenomethionine incorporation into human plasma
proteins and erythrocytes. Metabolism, 1966. 15(7): p. 626-40.
[39]. Thomson, C.D., Selenium. 2013: p. 186-92.

59
[40]. Yamashita, Y., T. Yabu and M. Yamashita, Discovery of the strong antioxidant
selenoneine in tuna and selenium redox metabolism. World J Biol Chem, 2010. 1(5): p.
144-50.
[41]. Yamashita, M., et al., Identification and determination of selenoneine, 2-selenyl-N
alpha , N alpha , N alpha -trimethyl-L-histidine, as the major organic selenium in blood
cells in a fish-eating population on remote Japanese Islands. Biol Trace Elem Res, 2013.
156(1-3): p. 36-44.
[42]. Cheng, W.H., et al., Cellular glutathione peroxidase knockout mice express normal
levels of selenium-dependent plasma and phospholipid hydroperoxide glutathione
peroxidases in various tissues. J Nutr, 1997. 127(8): p. 1445-50.
[43]. Lei, X.G., W.H. Cheng and J.P. McClung, Metabolic regulation and function of
glutathione peroxidase-1. Annu Rev Nutr, 2007. 27: p. 41-61.
[44]. Weiss, S.S. and R.A. Sunde, Selenium regulation of transcript abundance and
translational efficiency of glutathione peroxidase-1 and -4 in rat liver. Biochem J, 2001.
357(Pt 3): p. 851-58.
[45]. Cheng, W., et al., Selenium-dependent cellular glutathione peroxidase protects mice
against a pro-oxidant-induced oxidation of NADPH, NADH, lipids, and protein. FASEB J,
1999. 13(11): p. 1467-75.
[46]. Ho, Y.S., et al., Mice deficient in cellular glutathione peroxidase develop normally
and show no increased sensitivity to hyperoxia. J Biol Chem, 1997. 272(26): p. 16644-51.
[47]. D'Autreaux, B. and M.B. Toledano, ROS as signalling molecules: mechanisms that
generate specificity in ROS homeostasis. 2007. 8(10): p. 813-24.
[48]. Lubos, E., J. Loscalzo and D.E. Handy, Glutathione peroxidase-1 in health and
disease: from molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal,
2011. 15(7): p. 1957-97.
[49]. Wingler, K. and R. Brigelius-Flohe, Gastrointestinal glutathione peroxidase.
Biofactors, 1999. 10(2-3): p. 245-49.
[50]. Chu, F.F., J.H. Doroshow and R.S. Esworthy, Expression, characterization, and
tissue distribution of a new cellular selenium-dependent glutathione peroxidase, GSHPx-
GI. J Biol Chem, 1993. 268(4): p. 2571-76.
[51]. Florian, S., et al., Cellular and subcellular localization of gastrointestinal glutathione
peroxidase in normal and malignant human intestinal tissue. Free Radic Res, 2001. 35(6):
p. 655-63.
[52]. Yoshimura, S., et al., Tissue specific expression of the plasma glutathione
peroxidase gene in rat kidney. J Biochem, 1991. 109(6): p. 918-23.
[53]. Hoffmann, P.R., et al., The selenoproteome exhibits widely varying, tissue-specific
dependence on selenoprotein P for selenium supply. Nucleic Acids Res, 2007. 35(12): p.
3963-73.
[54]. Papp, L.V., et al., From selenium to selenoproteins: synthesis, identity, and their
role in human health. Antioxid Redox Signal, 2007. 9(7): p. 775-806.
[55]. Wolin, M.S., Plasma glutathione peroxidase activity is potentially a key regulator of
vascular disease-associated thrombosis. Circulation, 2011. 123(18): p. 1923-24.
[56]. Scheerer, P., et al., Structural basis for catalytic activity and enzyme polymerization
of phospholipid hydroperoxide glutathione peroxidase-4 (GPx4). Biochemistry, 2007.
46(31): p. 9041-49.

60
[57]. Imai, H. and Y. Nakagawa, Biological significance of phospholipid hydroperoxide
glutathione peroxidase (PHGPx, GPx4) in mammalian cells. Free Radic Biol Med, 2003.
34(2): p. 145-69.
[58]. Takebe, G., et al., A comparative study on the hydroperoxide and thiol specificity of
the glutathione peroxidase family and selenoprotein P. J Biol Chem, 2002. 277(43): p.
41254-58.
[59]. Conrad, M., et al., Physiological role of phospholipid hydroperoxide glutathione
peroxidase in mammals. Biol Chem, 2007. 388(10): p. 1019-25.
[60]. Ursini, F., et al., Dual function of the selenoprotein PHGPx during sperm
maturation. Science, 1999. 285(5432): p. 1393-96.
[61]. Conrad, M., et al., The nuclear form of phospholipid hydroperoxide glutathione
peroxidase is a protein thiol peroxidase contributing to sperm chromatin stability. Mol Cell
Biol, 2005. 25(17): p. 7637-44.
[62]. Yoo, S.E., et al., Gpx4 ablation in adult mice results in a lethal phenotype
accompanied by neuronal loss in brain. Free Radic Biol Med, 2012. 52(9): p. 1820-27.
[63]. Imai, H., et al., Early embryonic lethality caused by targeted disruption of the
mouse PHGPx gene. Biochem Biophys Res Commun, 2003. 305(2): p. 278-86.
[64]. Saito, Y. and K. Takahashi, Characterization of selenoprotein P as a selenium
supply protein. Eur J Biochem, 2002. 269(22): p. 5746-51.
[65]. Hill, K.E., et al., Deletion of selenoprotein P alters distribution of selenium in the
mouse. J Biol Chem, 2003. 278(16): p. 13640-46.
[66]. Schomburg, L., et al., Gene disruption discloses role of selenoprotein P in selenium
delivery to target tissues. Biochem J, 2003. 370(Pt 2): p. 397-402.
[67]. Krol, M.B., J. Gromadzinska and W. Wasowicz, SeP, ApoER2 and megalin as
necessary factors to maintain Se homeostasis in mammals. Journal of Trace Elements in
Medicine and Biology, 2012. 26(4): p. 262-66.
[68]. Olson, G.E., et al., Megalin mediates selenoprotein P uptake by kidney proximal
tubule epithelial cells. J Biol Chem, 2008. 283(11): p. 6854-60.
[69]. Burk, R.F. and K.E. Hill, Selenoprotein P: an extracellular protein with unique
physical characteristics and a role in selenium homeostasis. Annu Rev Nutr, 2005. 25: p.
215-35.
[70]. St, G.D., et al., Insights into the role of deiodinases from studies of genetically
modified animals. Thyroid, 2005. 15(8): p. 905-16.
[71]. Gereben, B., et al., Cellular and molecular basis of deiodinase-regulated thyroid
hormone signaling. Endocr Rev, 2008. 29(7): p. 898-938.
[72]. Silva, J.E. and P.R. Larsen, Potential of brown adipose tissue type II thyroxine 5'-
deiodinase as a local and systemic source of triiodothyronine in rats. J Clin Invest, 1985.
76(6): p. 2296-305.
[73]. Dentice, M., et al., The FoxO3/type 2 deiodinase pathway is required for normal
mouse myogenesis and muscle regeneration. J Clin Invest, 2010. 120(11): p. 4021-30.
[74]. Arner, E.S. and A. Holmgren, Physiological functions of thioredoxin and
thioredoxin reductase. Eur J Biochem, 2000. 267(20): p. 6102-09.
[75]. Jeong, W., et al., Identification and characterization of TRP14, a thioredoxin-related
protein of 14 kDa. New insights into the specificity of thioredoxin function. J Biol Chem,
2004. 279(5): p. 3142-50.
[76]. Jeong, W., et al., Roles of TRP14, a thioredoxin-related protein in tumor necrosis
factor-alpha signaling pathways. J Biol Chem, 2004. 279(5): p. 3151-59.

61
[77]. Ahsan, M.K., et al., Redox regulation of cell survival by the thioredoxin
superfamily: an implication of redox gene therapy in the heart. Antioxid Redox Signal,
2009. 11(11): p. 2741-58.
[78]. Weissbach, H., et al., Peptide methionine sulfoxide reductase: structure, mechanism
of action, and biological function. Arch Biochem Biophys, 2002. 397(2): p. 172-78.
[79]. Jakupoglu, C., et al., Cytoplasmic thioredoxin reductase is essential for
embryogenesis but dispensable for cardiac development. Mol Cell Biol, 2005. 25(5): p.
1980-88.
[80]. Peng, X., et al., Sec-containing TrxR1 is essential for self-sufficiency of cells by
control of glucose-derived H2O2. 2014. 5: p. e1235.
[81]. Sun, Q.A., et al., Redox regulation of cell signaling by selenocysteine in
mammalian thioredoxin reductases. J Biol Chem, 1999. 274(35): p. 24522-30.
[82]. Conrad, M., et al., Essential role for mitochondrial thioredoxin reductase in
hematopoiesis, heart development, and heart function. Mol Cell Biol, 2004. 24(21): p.
9414-23.
[83]. Nalvarte, I., A.E. Damdimopoulos and G. Spyrou, Human mitochondrial
thioredoxin reductase reduces cytochrome c and confers resistance to complex III
inhibition. Free Radic Biol Med, 2004. 36(10): p. 1270-78.
[84]. Kim, M.R., et al., Involvements of mitochondrial thioredoxin reductase (TrxR2) in
cell proliferation. Biochem Biophys Res Commun, 2003. 304(1): p. 119-24.
[85]. Choksi, S., et al., A HIF-1 target, ATIA, protects cells from apoptosis by
modulating the mitochondrial thioredoxin, TRX2. Mol Cell, 2011. 42(5): p. 597-609.
[86]. Li, C., et al., CERKL interacts with mitochondrial TRX2 and protects retinal cells
from oxidative stress-induced apoptosis. Biochim Biophys Acta, 2014. 1842(7): p. 1121-
29.
[87]. Soerensen, J., et al., The Role of thioredoxin reductases in brain development. PLoS
ONE, 2008. 3(3): p. e1813.
[88]. Sun, Q.A., et al., Selenoprotein oxidoreductase with specificity for thioredoxin and
glutathione systems. Proc Natl Acad Sci U S A, 2001. 98(7): p. 3673-78.
[89]. Su, D., et al., Mammalian selenoprotein thioredoxin-glutathione reductase. Roles in
disulfide bond formation and sperm maturation. J Biol Chem, 2005. 280(28): p. 26491-98.
[90]. Lu, J. and A. Holmgren, The thioredoxin antioxidant system. Free Radical Biology
and Medicine, 2014. 66(0): p. 75-87.
[91]. Kim, H.Y. and V.N. Gladyshev, Methionine sulfoxide reductases: selenoprotein
forms and roles in antioxidant protein repair in mammals. Biochem J, 2007. 407(3): p. 321-
29.
[92]. Lee, B.C., et al., Functions and evolution of selenoprotein methionine sulfoxide
reductases. Biochim Biophys Acta, 2009. 1790(11): p. 1471-77.
[93]. Boschi-Muller, S., et al., The enzymology and biochemistry of methionine sulfoxide
reductases. Biochim Biophys Acta, 2005. 1703(2): p. 231-38.
[94]. Lee, B.C., et al., MsrB1 and MICALs regulate actin assembly and macrophage
function via reversible stereoselective methionine oxidation. Mol Cell, 2013. 51(3): p. 397-
404.
[95]. Jia, Y., et al., Involvement of MsrB1 in the regulation of redox balance and
inhibition of peroxynitrite-induced apoptosis in human lens epithelial cells. Exp Eye Res,
2012. 100: p. 7-16.

62
[96]. Stadtman, E.R., Protein oxidation and aging. Free Radic Res, 2006. 40(12): p. 1250-
58.
[97]. Petit, N., et al., Selenoprotein N: an endoplasmic reticulum glycoprotein with an
early developmental expression pattern. Hum Mol Genet, 2003. 12(9): p. 1045-53.
[98]. Rederstorff, M., A. Krol and A. Lescure, Understanding the importance of selenium
and selenoproteins in muscle function. Cell Mol Life Sci, 2006. 63(1): p. 52-59.
[99]. Castets, P., et al., Selenoprotein N in skeletal muscle: from diseases to function. J
Mol Med (Berl), 2012. 90(10): p. 1095-107.
[100]. Castets, P., et al., Satellite cell loss and impaired muscle regeneration in
selenoprotein N deficiency. Hum Mol Genet, 2011. 20(4): p. 694-704.
[101]. Jurynec, M.J., et al., Selenoprotein N is required for ryanodine receptor calcium
release channel activity in human and zebrafish muscle. Proc Natl Acad Sci U S A, 2008.
105(34): p. 12485-90.
[102]. Arbogast, S., et al., Oxidative stress in SEPN1-related myopathy: from
pathophysiology to treatment. Ann Neurol, 2009. 65(6): p. 677-86.
[103]. Han, S.J., et al., Characterization of mammalian selenoprotein o: a redox-active
mitochondrial protein. PLoS One, 2014. 9(4): p. e95518.
[104]. Horibata, Y. and Y. Hirabayashi, Identification and characterization of human
ethanolaminephosphotransferase1. J Lipid Res, 2007. 48(3): p. 503-08.
[105]. Loflin, J., et al., Selenoprotein W during development and oxidative stress. J Inorg
Biochem, 2006. 100(10): p. 1679-84.
[106]. Gu, Q.P., et al., Selenoprotein W accumulates primarily in primate skeletal muscle,
heart, brain and tongue. Mol Cell Biochem, 2000. 204(1-2): p. 49-56.
[107]. Beilstein, M.A., et al., Selenoprotein W of rat muscle binds glutathione and an
unknown small molecular weight moiety. J Inorg Biochem, 1996. 61(2): p. 117-24.
[108]. Jeong, D., et al., Selenoprotein W is a glutathione-dependent antioxidant in vivo.
FEBS Lett, 2002. 517(1-3): p. 225-28.
[109]. Chung, Y.W., et al., Antioxidative role of selenoprotein W in oxidant-induced
mouse embryonic neuronal cell death. Mol Cells, 2009. 27(5): p. 609-13.
[110]. Jeon, Y.H., et al., Inhibition of 14-3-3 binding to Rictor of mTORC2 for Akt
phosphorylation at Ser473 is regulated by selenoprotein W. Biochim Biophys Acta, 2013.
1833(10): p. 2135-42.
[111]. Park, Y.H., Y.H. Jeon and I.Y. Kim, Selenoprotein W promotes cell cycle recovery
from G2 arrest through the activation of CDC25B. Biochim Biophys Acta, 2012. 1823(12):
p. 2217-26.
[112]. Novoselov, S.V., et al., Selenoprotein H is a nucleolar thioredoxin-like protein with
a unique expression pattern. J Biol Chem, 2007. 282(16): p. 11960-68.
[113]. Panee, J., et al., Selenoprotein H is a redox-sensing high mobility group family
DNA-binding protein that up-regulates genes involved in glutathione synthesis and phase II
detoxification. J Biol Chem, 2007. 282(33): p. 23759-65.
[114]. Wu, R.T., et al., Selenoprotein H suppresses cellular senescence through genome
maintenance and redox regulation. J Biol Chem, 2014. 289(49): p. 34378-88.
[115]. Korotkov, K.V., et al., Mammalian selenoprotein in which selenocysteine (Sec)
incorporation is supported by a new form of Sec insertion sequence element. Mol Cell
Biol, 2002. 22(5): p. 1402-11.
[116]. Ferguson, A.D., et al., NMR structures of the selenoproteins Sep15 and SelM reveal
redox activity of a new thioredoxin-like family. J Biol Chem, 2006. 281(6): p. 3536-43.

63
[117]. Reeves, M.A., F.P. Bellinger and M.J. Berry, The neuroprotective functions of
selenoprotein M and its role in cytosolic calcium regulation. Antioxid Redox Signal, 2010.
12(7): p. 809-18.
[118]. Qiao, X., et al., Galectin-1 is an interactive protein of selenoprotein M in the brain.
Int J Mol Sci, 2013. 14(11): p. 22233-45.
[119]. Chen, P., et al., Different forms of selenoprotein M differentially affect Abeta
aggregation and ROS generation. Int J Mol Sci, 2013. 14(3): p. 4385-99.
[120]. Labunskyy, V.M., D.L. Hatfield and V.N. Gladyshev, The Sep15 protein family:
roles in disulfide bond formation and quality control in the endoplasmic reticulum. IUBMB
Life, 2007. 59(1): p. 1-05.
[121]. Kumaraswamy, E., et al., Structure-expression relationships of the 15-kDa
selenoprotein gene. Possible role of the protein in cancer etiology. J Biol Chem, 2000.
275(45): p. 35540-47.
[122]. Korotkov, K.V., et al., Association between the 15-kDa selenoprotein and UDP-
glucose:glycoprotein glucosyltransferase in the endoplasmic reticulum of mammalian cells.
J Biol Chem, 2001. 276(18): p. 15330-36.
[123]. Labunskyy, V.M., et al., A novel cysteine-rich domain of Sep15 mediates the
interaction with UDP-glucose:glycoprotein glucosyltransferase. J Biol Chem, 2005.
280(45): p. 37839-45.
[124]. Kasaikina, M.V., et al., Roles of the 15-kDa selenoprotein (Sep15) in redox
homeostasis and cataract development revealed by the analysis of Sep 15 knockout mice. J
Biol Chem, 2011. 286(38): p. 33203-12.
[125]. Shchedrina, V.A., et al., Selenoprotein K binds multiprotein complexes and is
involved in the regulation of endoplasmic reticulum homeostasis. J Biol Chem, 2011.
286(50): p. 42937-48.
[126]. Dikiy, A., et al., SelT, SelW, SelH, and Rdx12: genomics and molecular insights
into the functions of selenoproteins of a novel thioredoxin-like family. Biochemistry, 2007.
46(23): p. 6871-82.
[127]. Grumolato, L., et al., Selenoprotein T is a PACAP-regulated gene involved in
intracellular Ca2+ mobilization and neuroendocrine secretion. FASEB J, 2008. 22(6): p.
1756-68.
[128]. Fairweather-Tait, S.J., Bioavailability of selenium. Eur J Clin Nutr, 1997. 51 Suppl
1: p. S20-23.
[129]. Van Dael, P., et al., Selenium absorption and retention from a selenite- or selenate-
fortified milk-based formula in men measured by a stable-isotope technique. Br J Nutr,
2001. 85(2): p. 157-63.
[130]. Bjornstedt, M., S. Kumar and A. Holmgren, Selenodiglutathione is a highly efficient
oxidant of reduced thioredoxin and a substrate for mammalian thioredoxin reductase. J Biol
Chem, 1992. 267(12): p. 8030-34.
[131]. Boehler, C.J., A.M. Raines and R.A. Sunde, Deletion of thioredoxin teductase and
tffects of telenite and telenate toxicity in Caenorhabditis elegans. PLoS ONE, 2013. 8(8): p.
e71525.
[132]. Suzuki, K.T., C. Doi and N. Suzuki, Metabolism of 76Se-methylselenocysteine
compared with that of 77Se-selenomethionine and 82Se-selenite. Toxicology and Applied
Pharmacology, 2006. 217(2): p. 185-95.

64
[133]. Esaki, N., et al., Selenocysteine lyase, a novel enzyme that specifically acts on
selenocysteine. Mammalian distribution and purification and properties of pig liver
enzyme. J Biol Chem, 1982. 257(8): p. 4386-91.
[134]. Pinto, J., et al., Chemopreventive mechanisms of α-keto acid metabolites of
naturally occurring organoselenium compounds. 2011. 41(1): p. 29-41.
[135]. Lu, J. and A. Holmgren, Selenoproteins. J Biol Chem, 2009. 284(2): p. 723-27.
[136]. Kobayashi, Y., et al., Selenosugars are key and urinary metabolites for selenium
excretion within the required to low-toxic range. Proceedings of the National Academy of
Sciences, 2002. 99(25): p. 15932-36.
[137]. Ohta, Y., et al., Speciation analysis of selenium metabolites in urine and breath by
HPLC- and GC-inductively coupled plasma-MS after administration of selenomethionine
and methylselenocysteine to rats. Chemical Research in Toxicology, 2009. 22(11): p. 1795-
801.
[138]. Zhang, J.X., et al., Association of glutathione peroxidase-1 (GPx-1) rs1050450
Pro198Leu and Pro197Leu polymorphisms with cardiovascular risk: a meta-analysis of
observational studies. J Geriatr Cardiol, 2014. 11(2): p. 141-50.
[139]. Voetsch, B., et al., Promoter polymorphisms in the plasma glutathione peroxidase
(GPx-3) gene: a novel risk factor for arterial ischemic stroke among young adults and
children. Stroke, 2007. 38(1): p. 41-49.
[140]. Baez-Duarte, B.G., et al., Glutathione peroxidase 3 serum levels and GPX3 gene
polymorphisms in subjects with metabolic syndrome. Arch Med Res, 2014. 45(5): p. 375-
82.
[141]. Alanne, M., et al., Variation in the selenoprotein S gene locus is associated with
coronary heart disease and ischemic stroke in two independent Finnish cohorts. Hum
Genet, 2007. 122(3-4): p. 355-65.
[142]. Stranges, S., et al., Effects of selenium supplementation on cardiovascular disease
incidence and mortality: secondary analyses in a randomized clinical trial. Am J Epidemiol,
2006. 163(8): p. 694-99.
[143]. Rees, K., et al., Selenium supplementation for the primary prevention of
cardiovascular disease. Cochrane Database Syst Rev, 2013. 1: p. D9671.
[144]. Xun, P., et al., Longitudinal association between toenail selenium levels and
measures of subclinical atherosclerosis: the CARDIA trace element study. Atherosclerosis,
2010. 210(2): p. 662-67.
[145]. Flores-Mateo, G., et al., Selenium and coronary heart disease: a meta-analysis. Am J
Clin Nutr, 2006. 84(4): p. 762-73.
[146]. Rayman, M.P., et al., Effect of supplementation with high-selenium yeast on plasma
lipids: a randomized trial. Ann Intern Med, 2011. 154(10): p. 656-65.
[147]. Akbaraly, T.N., et al., Plasma selenium and risk of dysglycemia in an elderly French
population: results from the prospective Epidemiology of Vascular Ageing Study. Nutr
Metab (Lond), 2010. 7: p. 21.
[148]. Park, K., et al., Toenail selenium and incidence of type 2 diabetes in U.S. men and
women. Diabetes Care, 2012. 35(7): p. 1544-51.
[149]. Laclaustra, M., et al., Serum selenium concentrations and diabetes in U.S. adults:
National Health and Nutrition Examination Survey (NHANES) 2003-2004. Environ Health
Perspect, 2009. 117(9): p. 1409-13.
[150]. Bleys, J., A. Navas-Acien and E. Guallar, Serum selenium and diabetes in U.S.
adults. Diabetes Care, 2007. 30(4): p. 829-34.

65
[151]. Algotar, A.M., et al., Selenium supplementation has no effect on serum glucose
levels in men at high risk of prostate cancer. J Diabetes, 2013. 5(4): p. 465-70.
[152]. Algotar, A.M., et al., No effect of selenium supplementation on serum glucose
levels in men with prostate cancer. Am J Med, 2010. 123(8): p. 765-68.
[153]. Stranges, S., et al., Effects of long-term selenium supplementation on the incidence
of type 2 diabetes: a randomized trial. Ann Intern Med, 2007. 147(4): p. 217-23.
[154]. Lippman, S.M., et al., Effect of selenium and vitamin E on risk of prostate cancer
and other cancers: the Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA,
2009. 301(1): p. 39-51.
[155]. Rayman, M.P. and S. Stranges, Epidemiology of selenium and type 2 diabetes: can
we make sense of it? Free Radic Biol Med, 2013. 65: p. 1557-64.
[156]. Wood, S.M., et al., beta-Carotene and selenium supplementation enhances immune
response in aged humans. Integr Med, 2000. 2(2): p. 85-92.
[157]. Kiremidjian-Schumacher, L., et al., Selenium and immunocompetence in patients
with head and neck cancer. Biol Trace Elem Res, 2000. 73(2): p. 97-111.
[158]. Kiremidjian-Schumacher, L., et al., Supplementation with selenium and human
immune cell functions. II. Effect on cytotoxic lymphocytes and natural killer cells. Biol
Trace Elem Res, 1994. 41(1-2): p. 115-27.
[159]. Rayman, M.P., The importance of selenium to human health. Lancet, 2000.
356(9225): p. 233-41.
[160]. Campa, A., et al., Mortality risk in selenium-deficient HIV-positive children. J
Acquir Immune Defic Syndr Hum Retrovirol, 1999. 20(5): p. 508-13.
[161]. Beck, M.A., Selenium and vitamin E status: impact on viral pathogenicity. J Nutr,
2007. 137(5): p. 1338-40.
[162]. Broome, C.S., et al., An increase in selenium intake improves immune function and
poliovirus handling in adults with marginal selenium status. Am J Clin Nutr, 2004. 80(1):
p. 154-62.
[163]. Burbano, X., et al., Impact of a selenium chemoprevention clinical trial on hospital
admissions of HIV-infected participants. HIV Clin Trials, 2002. 3(6): p. 483-91.
[164]. Hurwitz, B.E., et al., Suppression of human immunodeficiency virus type 1 viral
load with selenium supplementation: a randomized controlled trial. Arch Intern Med, 2007.
167(2): p. 148-54.
[165]. Etminan, M., et al., Intake of selenium in the prevention of prostate cancer: a
systematic review and meta-analysis. Cancer Causes Control, 2005. 16(9): p. 1125-31.
[166]. Brinkman, M., et al., Are men with low selenium levels at increased risk of prostate
cancer? Eur J Cancer, 2006. 42(15): p. 2463-71.
[167]. Peters, U. and Y. Takata, Selenium and the prevention of prostate and colorectal
cancer. Mol Nutr Food Res, 2008. 52(11): p. 1261-72.
[168]. Clark, L.C., et al., Effects of selenium supplementation for cancer prevention in
patients with carcinoma of the skin. A randomized controlled trial. Nutritional Prevention
of Cancer Study Group. JAMA, 1996. 276(24): p. 1957-63.
[169]. Zhuo, H., A.H. Smith and C. Steinmaus, Selenium and lung cancer: a quantitative
analysis of heterogeneity in the current epidemiological literature. Cancer Epidemiol
Biomarkers Prev, 2004. 13(5): p. 771-78.
[170]. Amaral, A.F., et al., Selenium and bladder cancer risk: a meta-analysis. Cancer
Epidemiol Biomarkers Prev, 2010. 19(9): p. 2407-15.

66
[171]. Parizek, J. and I. Ostadalova, The protective effect of small amounts of selenite in
sublimate intoxication. Experientia, 1967. 23(2): p. 142-43.
[172]. Kerper, L.E., N. Ballatori and T.W. Clarkson, Methylmercury transport across the
blood-brain barrier by an amino acid carrier. Am J Physiol, 1992. 262(5 Pt 2): p. R761-65.
[173]. Aschner, M. and J.L. Aschner, Mercury neurotoxicity: mechanisms of blood-brain
barrier transport. Neurosci Biobehav Rev, 1990. 14(2): p. 169-76.
[174]. Kajiwara, Y., et al., Methylmercury transport across the placenta via neutral amino
acid carrier. Arch Toxicol, 1996. 70(5): p. 310-14.
[175]. Aberg, B., et al., Metabolism of methyl mercury (203Hg) compounds in man. Arch
Environ Health, 1969. 19(4): p. 478-84.
[176]. Ganther, H.E., et al., Selenium: relation to decreased toxicity of methylmercury
added to diets containing tuna. Science, 1972. 175(4026): p. 1122-24.
[177]. Khan, M.A. and F. Wang, Mercury-selenium compounds and their toxicological
significance: toward a molecular understanding of the mercury-selenium antagonism.
Environ Toxicol Chem, 2009. 28(8): p. 1567-77.
[178]. Yang, D., et al., Selenium and mercury in organisms: Interactions and mechanisms.
Environmental Reviews, 2008. 16(NA): p. 71-92.
[179]. Reid, R.S. and D.L. Rabenstein, Nuclear magnetic resonance studies of the solution
chemistry of metal complexes. XVII. Formation constants for the complexation of
methylmercury by sulfhydryl-containing amino acids and related molecules. Canadian
Journal of Chemistry, 1981. 59(10): p. 1505-14.
[180]. Sugiura, Y., Y. Tamai and H. Tanaka, Selenium protection against mercury toxicity:
high binding affinity of methylmercury by selenium-containing ligands in comparison with
sulfur-containing ligands. Bioinorg Chem, 1978. 9(2): p. 167-80.
[181]. Magos, L., M. Webb and A.R. Hudson, Complex formation between selenium and
methylmercury. Chem Biol Interact, 1979. 28(2-3): p. 359-62.
[182]. Naganuma, A. and N. Imura, Bis(methylmercuric) selenide as a reaction product
from methylmercury and selenite in rabbit blood. Res Commun Chem Pathol Pharmacol,
1980. 27(1): p. 163-73.
[183]. Koeman, J.H., et al., Mercury and selenium in marine mammals and birds. Sci Total
Environ, 1975. 3(3): p. 279-87.
[184]. Endo, T., et al., Distribution of total mercury, methyl mercury and selenium in pod
of killer whales (Orcinus Orca) stranded in the northern area of Japan: comparison of
mature females with calves. Environ Pollut, 2006. 144(1): p. 145-50.
[185]. Gaskin, D.E., et al., Changes in mercury levels in harbor porpoises from the Bay of
Fundy, Canada, and adjacent waters during 1969-1977. Arch Environ Contam Toxicol,
1979. 8(6): p. 733-62.
[186]. Smith, T.G. and F.A.J. Armstrong, Mercury and selenium in ringed and bearded
seal tissues from Arctic Canada. ARCTIC, 1978. 31(2): p. 75-84.
[187]. Itano, K., et al., Mercury and selenium levels in striped dolphins caught off the
Pacific Coast of Japan. Agricultural and Biological Chemistry, 1984. 48(5): p. 1109-16.
[188]. Meador, J.P., et al., Comparison of elements in bottlenose dolphins stranded on the
beaches of Texas and Florida in the Gulf of Mexico over a one-year period. Arch Environ
Contam Toxicol, 1999. 36(1): p. 87-98.
[189]. Martoja, R. and J.P. Berry, Identification of tiemannite as a probable product of
demethylation of mercury by selenium in cetaceans. A complement to the scheme of the
biological cycle of mercury. 1980. 30(1): p. 7-10.

67
[190]. Nigro, M. and C. Leonzio, Intracellular storage of mercury and selenium in different
marine vertebrates. Marine Ecology Progress Series, 1996. 135: p. 137-43.
[191]. Lailson-Brito, J., et al., Mercury-selenium relationships in liver of Guiana dolphin:
the possible role of Kupffer cells in the detoxification process by tiemannite formation.
PLoS ONE, 2012. 7(7): p. e42162.
[192]. Palmisano, F., N. Cardellicchio and P.G. Zambonin, Speciation of mercury in
dolphin liver: A two-stage mechanism for the demethylation accumulation process and role
of selenium. Marine Environmental Research, 1995. 40(2): p. 109-21.
[193]. Yoneda, S. and K.T. Suzuki, Equimolar Hg-Se complex binds to selenoprotein P.
Biochem Biophys Res Commun, 1997. 231(1): p. 7-11.
[194]. Ganther, H.E., Modification of methylmercury toxicity and metabolism by selenium
and vitamin E: possible mechanisms. Environ Health Perspect, 1978. 25: p. 71-76.
[195]. Delights, N.: [http://www.deliceboreal.com/en/nunavik/map.php Last accessed:
December 08, 2014].
[196]. Canada Statistics, Aboriginal peoples in Canada: First nations people, Métis and
Inuit:National Household Survey,2011.
[197]. Canada, S., ed. Aboriginal Peoples in Canada: First Nations People, Métis and Inuit.
2011, [http://www12.statcan.gc.ca/nhs-enm/2011/as-sa/99-011-x/99-011-x2011001-fra.pdf
Last accessed: December 28, 2014].
[198]. Lemire, M., et al., Local country food sources of methylmercury, selenium and
omega-3 fatty acids in Nunavik, Northern Quebec. Sci Total Environ, 2014. 509-510: p.
248-59.
[199]. Blanchet C, Rochette L. Nutrition Inuit Health Survey 2004, Qanuippitaa? How are
we? Québec: Institut National de la Santé Publique du Québec (INSPQ) and Nunavik
Regional Board of Health and Social Services (NRBHSS); 2008. Nutrition and food
consumption among the Inuit of Nunavik; pp. 1–87.
[200]. AANDC, Canadian Arctic Contaminant Assessment Report III — Mercury in
Canada's North. Aboriginal Affairs and Northern Development Canada (AANDC),
Northern Contaminants Program (2012), [http://emrlibrary.gov.yk.ca/INAC_Mercury_
Report.pdf Last accessed: January 3, 2015].
[201]. Dallaire, R., et al., Exposure to organochlorines and mercury through fish and
marine mammal consumption: associations with growth and duration of gestation among
Inuit newborns. Environ Int, 2013. 54: p. 85-91.
[202]. Boucher, O., et al., Prenatal methylmercury, postnatal lead exposure, and evidence
of attention deficit/hyperactivity disorder among Inuit children in Arctic Quebec. Environ
Health Perspect, 2012. 120(10): p. 1456-61.
[203]. Boucher, O., et al., Prenatal exposure to methylmercury and PCBs affects distinct
stages of information processing: an event-related potential study with Inuit children.
Neurotoxicology, 2010. 31(4): p. 373-84.
[204]. Boucher, O., et al., Neurophysiologic and neurobehavioral evidence of beneficial
effects of prenatal omega-3 fatty acid intake on memory function at school age. Am J Clin
Nutr, 2011. 93(5): p. 1025-37.
[205]. Lucas, M., et al., Gestational age and birth weight in relation to n-3 fatty acids
among Inuit (Canada). Lipids, 2004. 39(7): p. 617-26.
[206]. Dewailly, E., et al., n-3 Fatty acids and cardiovascular disease risk factors among
the Inuit of Nunavik. Am J Clin Nutr, 2001. 74(4): p. 464-73.

68
[207]. Valera, B., et al., Influence of n-3 fatty acids on cardiac autonomic activity among
Nunavik Inuit adults. Int J Circumpolar Health, 2011. 70(1): p. 6-18.
[208]. Ayotte, P., et al., Relation between methylmercury exposure and plasma
paraoxonase activity in inuit adults from Nunavik. Environ Health Perspect, 2011. 119(8):
p. 1077-83.
[209]. Alkazemi, D., et al., New insights regarding tissue Se and Hg interactions on
oxidative stress from plasma IsoP and IsoF measures in the Canadian Inuit population. J
Lipid Res, 2013. 54(7): p. 1972-79.
[210]. Rochette, L. and C. Blanchet, Qanuippitaa? How are we? - Methodological report.
2007: Montreal, Qc, Canada. p. 328.
[211]. Li, Y., et al., Full quantification of selenium species by RP and AF-ICP-qMS with
on-line isotope dilution in serum samples from mercury-exposed people supplemented with
selenium-enriched yeast. Journal of Analytical Atomic Spectrometry, 2011. 26(1): p. 224-
29.
[212]. Association, F.D.N., Prescriptions générales concernant la compétence des
laboratoires d'étalonnages et d'essais: General requirements for the competence of testing
and calibration laboratories. Normes françaises. 2000: AFNOR.
[213]. Plecko, T., et al., Determination and distribution of human plasma selenoproteins.
1999. 363(5-6): p. 517-19.
[214]. Jitaru, P., et al., Challenges in the accurate speciation analysis of selenium in
humans: first report on indicative levels of selenoproteins in a serum certified reference
material for total selenium (BCR-637). 2010. 15(6): p. 343-50.
[215]. Steinbrenner, H. and H. Sies, Selenium homeostasis and antioxidant selenoproteins
in brain: Implications for disorders in the central nervous system. Archives of Biochemistry
and Biophysics, 2013. 536(2): p. 152-57.
[216]. Traulsen, H., et al., Selenoprotein P protects low-density lipoprotein against
oxidation. Free Radic Res, 2004. 38(2): p. 123-28.
[217]. Whitin, J.C., et al., Extracellular glutathione peroxidase is secreted basolaterally by
human renal proximal tubule cells. Am J Physiol Renal Physiol, 2002. 283(1): p. F20-28.
[218]. Suzuki, K.T. and Y. Ogra, Metabolic pathway for selenium in the body: speciation
by HPLC-ICP MS with enriched Se. Food Additives & Contaminants, 2002. 19(10): p.
974-83.
[219]. Hong, Y.S., Y.M. Kim and K.E. Lee, Methylmercury exposure and health effects. J
Prev Med Public Health, 2012. 45(6): p. 353-63.
[220]. Chapman, L. and H.M. Chan, The influence of nutrition on methyl mercury
intoxication. Environ Health Perspect, 2000. 108 Suppl 1: p. 29-56.
[221]. Lemire, M., et al., Selenium and mercury in the Brazilian Amazon: opposing
influences on age-related cataracts. Environ Health Perspect, 2010. 118(11): p. 1584-89.
[222]. Rayman, M.P., H.G. Infante and M. Sargent, Food-chain selenium and human
health: spotlight on speciation. Br J Nutr, 2008. 100(2): p. 238-53.
[223]. Lemire, M., et al., No evidence of selenosis from a selenium-rich diet in the
Brazilian Amazon. Environ Int, 2012. 40: p. 128-36.
[224]. Jette, M., A health profile of the Inuit of Nunavik: report of the Sante Quebec
Health Survey (1992). Int J Circumpolar Health, 1998. 57 Suppl 1: p. 630-35.
[225]. Kaňa, A.N., et al., Software solution for post-column isotope dilution liquid
chromatography-inductively coupled plasma mass spectrometry. Analytical Letters, 2013.
46(15): p. 2430-43.

69
[226]. Valera, B., E. Dewailly and P. Poirier, Cardiac autonomic activity and blood
pressure among Nunavik Inuit adults exposed to environmental mercury: a cross-sectional
study. Environ Health, 2008. 7: p. 29.
[227]. Garcia-Sevillano, M.A., T. Garcia-Barrera and J.L. Gomez-Ariza, Development of a
new column switching method for simultaneous speciation of selenometabolites and
selenoproteins in human serum. J Chromatogr A, 2013. 1318: p. 171-79.
[228]. Jitaru, P., et al., Simultaneous speciation analysis of glutathione peroxidase,
selenoprotein P and selenoalbumin in human serum by tandem anion exchange-affinity
HPLC and on-line isotope dilution ICP-quadrupole MS. Anal Bioanal Chem, 2008. 391(2):
p. 661-69.
[229]. Hinojosa Reyes, L., et al., Determination of selenium in biological materials by
isotope dilution analysis with an octapole reaction system ICP-MS. Journal of Analytical
Atomic Spectrometry, 2003. 18(1): p. 11-16.
[230]. Letsiou, S., et al., High-throughput quantification of selenium in individual serum
proteins from a healthy human population using HPLC on-line with isotope dilution
inductively coupled plasma-MS. Proteomics, 2010. 10(19): p. 3447-57.
[231]. Spina, A., et al., Anthropometric indices and selenium status in British adults: the
U.K. National Diet and Nutrition Survey. Free Radic Biol Med, 2013. 65: p. 1315-21.
[232]. Golubkina, N.A. and G.V. Alfthan, The human selenium status in 27 regions of
Russia. J Trace Elem Med Biol, 1999. 13(1-2): p. 15-20.
[233]. Kim, Y.J., et al., Serum selenium level in healthy Koreans. Biol Trace Elem Res,
2009. 131(2): p. 103-09.
[234]. Lyons, G.H., et al., Selenium in Australia: selenium status and biofortification of
wheat for better health. J Trace Elem Med Biol, 2005. 19(1): p. 75-82.
[235]. Karita, K. and T. Suzuki, Fish eating and variations in selenium and mercury levels
in plasma and erythrocytes in free-living healthy Japanese men. Biol Trace Elem Res,
2002. 90(1-3): p. 71-81.
[236]. Arnaud, J., et al., Serum selenium determinants in French adults: the SU.VI.M.AX
study. Br J Nutr, 2006. 95(2): p. 313-20.
[237]. Alfthan, G., et al., Effects of nationwide addition of selenium to fertilizers on foods,
and animal and human health in Finland: From deficiency to optimal selenium status of the
population. J Trace Elem Med Biol, 2014. 31(0): p. 142-47.
[238]. Jitaru, P., et al., A systematic approach to the accurate quantification of selenium in
serum selenoalbumin by HPLC-ICP-MS. Anal Chim Acta, 2010. 657(2): p. 100-07.
[239]. Jitaru, P., et al., Speciation analysis of selenoproteins in human serum by solid-
phase extraction and affinity HPLC hyphenated to ICP-quadrupole MS. Journal of
Analytical Atomic Spectrometry, 2008. 23(3): p. 402-06.
[240]. Hinojosa Reyes, L., et al., Quantitative speciation of selenium in human serum by
affinity chromatography coupled to post-column isotope dilution analysis ICP-MS. Journal
of Analytical Atomic Spectrometry, 2003. 18(10): p. 1210-16.
[241]. Roman, M., et al., Plasma selenoproteins concentrations in type 2 diabetes mellitus-
-a pilot study. Transl Res, 2010. 156(4): p. 242-50.
[242]. Rayman, M.P., Selenoproteins and human health: insights from epidemiological
data. Biochim Biophys Acta, 2009. 1790(11): p. 1533-40.
[243]. Reeves, M.A. and P.R. Hoffmann, The human selenoproteome: recent insights into
functions and regulation. Cell Mol Life Sci, 2009. 66(15): p. 2457-78.

70
[244]. Lye, E., et al., Blood total mercury concentrations in the Canadian population:
Canadian Health Measures Survey cycle 1, 2007-2009. Can J Public Health, 2013. 104(3):
p. e246-51.
[245]. Mortensen, M.E., et al., Total and methyl mercury in whole blood measured for the
first time in the U.S. population: NHANES 2011-2012. Environ Res, 2014. 134: p. 257-64.
[246]. Park, J.H., et al., Total mercury concentrations in the general Korean population,
2008-2011. Regul Toxicol Pharmacol, 2014. 70(3): p. 681-86.
[247]. Fok, T.F., et al., Fetal methylmercury exposure as measured by cord blood mercury
concentrations in a mother-infant cohort in Hong Kong. Environ Int, 2007. 33(1): p. 84-92.
[248]. Birgisdottir, B.E., et al., Essential and toxic element concentrations in blood and
urine and their associations with diet: results from a Norwegian population study including
high-consumers of seafood and game. Sci Total Environ, 2013. 463-464: p. 836-44.
[249]. Lemire, M., et al., Elevated blood selenium levels in the Brazilian Amazon. Sci
Total Environ, 2006. 366(1): p. 101-11.
[250]. Nielsen, A.B., M. Davidsen and P. Bjerregaard, The association between blood
pressure and whole blood methylmercury in a cross-sectional study among Inuit in
Greenland. Environ Health, 2012. 11: p. 44.
[251]. Canada, H., Report on human biomonitoring of environmental chemicals in Canada,
results of the Canadian Health Measures Survey Cycle 1 (2007–2009). 2010,
[http://www.hc-sc.gc.ca/ewh-semt/alt_formats/hecs-sesc/pdf/pubs/contaminants/chms-
ecms/report-rapport-eng.pdf Last accessed: December 28, 2014].
[252]. Jain, R.B. and Y.S. Choi, Normal reference ranges for and variability in the levels
of blood manganese and selenium by gender, age, and race/ethnicity for general U.S.
population. Journal of Trace Elements in Medicine and Biology, 2015. 30: p. 142-52.
[253]. Batariova, A., et al., Whole blood selenium content in healthy adults in the Czech
Republic. Sci Total Environ, 2005. 338(3): p. 183-88.
[254]. Kłapcińska, B., et al., Selenium levels in blood of Upper Silesian population. 2005.
108(1-3): p. 1-15.
[255]. Chen, C., et al., The roles of serum selenium and selenoproteins on mercury toxicity
in environmental and occupational exposure. Environ Health Perspect, 2006. 114(2): p.
297-301.
[256]. Hansen, J.C., B. Deutch and H.S. Pedersen, Selenium status in Greenland Inuit. Sci
Total Environ, 2004. 331(1-3): p. 207-14.
[257]. Dumont, E., F. Vanhaecke and R. Cornelis, Selenium speciation from food source
to metabolites: a critical review. Anal Bioanal Chem, 2006. 385(7): p. 1304-23.
[258]. Larsen, E.H., et al., Speciation and bioavailability of selenium in yeast-based
intervention agents used in cancer chemoprevention studies. J AOAC Int, 2004. 87(1): p.
225-32.
[259]. Lemes, M., et al., Methylmercury and selenium speciation in different tissues of
beluga whales (Delphinapterus leucas) from the western Canadian Arctic. Environ Toxicol
Chem, 2011. 30(12): p. 2732-38.
[260]. Yamashita, M., et al., Selenoneine, a novel selenium-containing compound,
mediates detoxification mechanisms against methylmercury accumulation and toxicity in
zebrafish embryo. Mar Biotechnol (NY), 2013. 15(5): p. 559-70.













![VFinalEurotoolbox 14-15 -brochure June -c- trad.c[1]-2 · Communication in the Arts Faculty of the University of Porto. She has participated in a workshop of Illustration Techniques](https://img.pdfslide.fr/doc/110x75/5f71a3c2049d746e9c7685aa/vfinaleurotoolbox-14-15-brochure-june-c-tradc1-2-communication-in-the-arts.jpg)















