Embed Size (px)
Citation preview

Patrick Gagnon
Dynamique de répartition et mécanismes de défense des macrophytes de la zone infralittorale rocheuse dominée par
l’oursin vert, Strongylocentrotus droebachiensis
Thèse présentée
à la Faculté des études supérieures de lʹUniversité Laval pour l’obtention
du grade de Philosophiae Doctor (Ph.D.)
Département de biologie FACULTÉ DES SCIENCES ET DE GÉNIE
UNIVERSITÉ LAVAL QUÉBEC
DÉCEMBRE 2003
© Patrick Gagnon, 2003

À mes très chers parents, Patricia Bourget et Richard Gagnon,
ainsi qu’à Nellie…

I
Sommaire
Le but de mon étude était de caractériser la dynamique de répartition des
macrophytes de la zone infralittorale rocheuse de l’archipel de Mingan, au nord du
golfe du Saint‐Laurent, puis d’examiner les mécanismes de défense permettant à
certains macrophytes dominants de résister au broutage de l’oursin vert,
Strongylocentrotus droebachiensis. La position de la limite inférieure des ceintures de
laminaires variait grandement dans le temps et d’un site à l’autre et était
principalement contrôlée par le broutage de l’oursin. En zone dénudée, une grande
stabilité temporelle caractérisait la distribution et l’abondance de la phaéophycée
pérenne Agarum cribrosum dont l’agrégation en plaques résilientes à de faibles
perturbations favorisait le recrutement algal. L’évitement du broutage de l’oursin par
les juvéniles d’A. cribrosum reposait sur leur association avec la phaéophycée annuelle
Desmarestia viridis. La répulsion de l’oursin par D. viridis était principalement
attribuable au mouvement de balayage de l’algue, résultat de l’effet combiné de sa
délicate morphologie et de l’action des vagues.
Auteur: Patrick Gagnon
Directeur: John H. Himmelman

II
Résumé
Dans le nord du golfe du Saint‐Laurent, lʹoursin vert, Strongylocentrotus droebachiensis,
est lʹherbivore dominant en zone infralittorale rocheuse. Son impact sur la structure et
la dynamique des communautés de macrophytes est toutefois peu connu. Le but de
mon étude était de caractériser la dynamique de répartition des macrophytes de la
zone infralittorale rocheuse de l’archipel de Mingan, puis d’examiner les mécanismes
de défense permettant à certains macrophytes dominants de résister au broutage de
l’oursin. Des observations et expériences sur huit sites sur des périodes couvrant
jusqu’à deux ans ont indiqué que la position de la limite inférieure des ceintures de
laminaires variait grandement dans le temps et entre les sites et était principalement
contrôlée par le broutage de l’oursin. Lʹabrasion par les glaces et la formation de
barrières algales limitant le déplacement des oursins ont vraisemblablement affecté
cette variation. En zone dénudée, une grande stabilité temporelle caractérisait la
distribution et l’abondance de la phaéophycée pérenne Agarum cribrosum.
L’agrégation d’A. cribrosum en plaques résilientes à de faibles perturbations favorisait
le recrutement algal, dont la rhodophycée Ptilota serrata. L’évitement du broutage de
l’oursin par les juvéniles d’A. cribrosum reposait sur leur association avec la
phaéophycée annuelle Desmarestia viridis. Pour les juvéniles évoluant sous un couvert
de D. viridis, le coût d’une perte de croissance était nettement compensé par le
bénéfice d’une réduction du broutage. Des expériences en laboratoire ont établi que la
répulsion de l’oursin par D. viridis était principalement due au mouvement de
balayage de l’algue, résultat de l’effet combiné de sa délicate morphologie et de
l’action des vagues puis, dans une faible mesure, de sa composition chimique. Les
attaques de l’oursin sur D. viridis augmentaient avec la densité de l’oursin mais
diminuaient avec l’augmentation du mouvement de l’eau.
Auteur: Patrick Gagnon
Directeur: John H. Himmelman

III
Abstract
In the northern Gulf of St. Lawrence, the green sea urchin, Strongylocentrotus
droebachiensis, is the dominant herbivore in the rocky subtidal zone. Its impact on the
structure and dynamics of algal communities is, however, poorly known. The
objective of my study was to characterize the dynamics in the distribution and
abundance of macrophytes in the rocky subtidal zone in the Mingan Islands, and to
examine the mechanisms of defense that allow dominant macrophytes to resist urchin
grazing. Observations and experiments at eight sites over periods of up to two years
indicated that the position of the lower limit of the kelp beds varied markedly
through time and among sites and was largely controlled by urchin grazing. Ice
souring and the formation of algal barriers that reduced urchin displacement likely
affected this variation. In the barrens zone, there was a high temporal stability in the
distribution and abundance of the perennial phaeophyte Agarum cribrosum. The
aggregation of A. cribrosum into patches resilient to small disturbances enhanced algal
recruitment, notably that of the rhodophyte Ptilota serrata. The avoidance of urchin
grazing by juveniles of A. cribrosum was related to their association with the annual
phaeophyte Desmarestia viridis. For juveniles, the cost of a reduced growth under a
D. viridis canopy was clearly outweighed by the benefit of reduced urchin grazing.
Laboratory experiments established that the repulsion of urchins by D. viridis was
primarily due to the sweeping motion of the alga resulting from the interaction
between its delicately‐branched morphology and wave action and, to a lesser extent,
by its chemical make‐up. Urchin attacks on D. viridis increased with urchin density,
but decreased with increased water motion.
Auteur: Patrick Gagnon
Directeur: John H. Himmelman

IV
Avant‐propos
Cette thèse comporte six chapitres. Les chapitres I et VI correspondent
respectivement aux introduction et conclusion générales. Les chapitres II, III, IV et V
(corps de la thèse) font état de mes travaux de recherche dans l’archipel de Mingan et
ont été rédigés en anglais, sous forme d’articles scientifiques dont voici les références:
Chapitre II: Gagnon P., Himmelman J.H. & Johnson L.E. (sous presse). Temporal
variation in community interfaces: kelp bed boundary dynamics
adjacent to persistent urchin barrens. Mar. Biol.
Chapitre III: Gagnon P., Himmelman J.H. & Johnson L.E. Spatial and temporal
stability in algal assemblages on urchin barrens in the northern Gulf of
St. Lawrence.
Chapitre IV: Gagnon P., Himmelman J.H. & Johnson L.E. (2003). Algal colonization
in urchin barrens: defense by association during recruitment of the
brown alga Agarum cribrosum. J. Exp. Mar. Biol. Ecol. 290: 179‐196.
Chapitre V: Gagnon P., St‐Hilaire‐Gravel L.V., Himmelman J.H. & Johnson L.E.
Multiple plant defenses against herbivores: the interplay of chemical,
morphological, and hydrodynamic factors.
J’ai conçu et réalisé toutes les expériences et observations de terrain et de
laboratoire, effectué la totalité des analyses statistiques (avec l’appui de statisticiens
pour les analyses plus complexes), puis rédigé les différents chapitres avec l’aide de
mes codirecteurs John Himmelman et Ladd Johnson. Les résultats apparaissant dans
cette thèse ont été présentés lors des congrès et ateliers suivants:
Gagnon P., Himmelman J.H. & Johnson L.E. (2003). Sweeping or using acid: how
does Desmarestia viridis deter urchin attacks? 32nd Benthic Ecology Meeting,
Groton, Connecticut, USA.

V
Gagnon P., Himmelman J.H. & Johnson L.E. (2000). Seaweed survival in barren
zones: a case of associational refuge for juvenile Agarum cribrosum.
29th Benthic Ecology Meeting, Wilmington, North Carolina, USA.
Gagnon P., Himmelman J.H. & Johnson L.E. (2000). Utilisation du couvert de
Desmarestia viridis par les juvéniles d’Agarum cribrosum pour une protection
contre l’herbivorie. 68ème Congrès International de l’ACFAS (Association
Canadienne Française pour l’Avancement des Sciences), Montréal, Québec,
Canada.
Gagnon P., Himmelman J.H. & Johnson L.E. (2000). Utilisation du couvert de
Desmarestia viridis par les juvéniles d’Agarum cribrosum pour une protection
contre l’herbivorie. 25ème Congrès annuel de la Société Québécoise pour
l’Étude Biologique du Comportement, Rimouski, Québec, Canada.
Gagnon, P. (1999). Importance du refuge d’association pour l’établissement des
macrophytes de la communauté infralittorale. Forum Québécois en Sciences
de la Mer, Mont‐Joli, Québec, Canada.
La description du générateur de vagues que j’ai conçu et fabriqué pour réaliser
certaines de mes expériences apparaît en annexe du chapitre V, puis dans l’article
suivant rédigé simultanément à mes travaux de thèse:
Gagnon P., Wagner G. & Himmelman J.H. (2003). Use of a wave tank to study the
effects of water motion and algal movement on the displacement of the sea
star Asterias vulgaris towards its prey. Mar. Ecol. Prog. Ser. 258: 125‐132.
Je remercie mon directeur, John Himmelman, ainsi que mon codirecteur,
Ladd Johnson, pour la confiance témoignée et la latitude accordée depuis mes tous
premiers pas à l’initiation à la recherche, jusqu’à la fin de cette aventure doctorale.
Leur disponibilité, leur support et leurs nombreuses recommandations scientifiques
pendant la rédaction de ma thèse ont été particulièrement appréciés et la rigueur de
leur encadrement très formatrice. Merci également à Gilles Houle, Jeremy McNeil et

VI
Edwin Bourget pour leur grande ouverture et pour toutes ces discussions profitables,
puis au Conseil de Recherche en Sciences Naturelles et Génie du Canada (CRSNG),
au Fonds pour la formation de Chercheurs et l’Aide à la Recherche du Québec
(FCAR), au Ministère des Pêches et Océans du Canada (MPO), à Québec‐Océan
(GIROQ) et au Département de biologie de l’Université Laval pour leur généreuse
contribution monétaire.
J’exprime toute ma gratitude aux personnes qui ont participé, de près ou de
loin, à la réalisation de mon étude. Merci à mes précieux assistants de recherche pour
toutes ces heures passées au laboratoire et pour ces interminables plongées dans des
eaux glaciales et agitées: Philippe Gauthier, Valérie Messier, Brigitte Laberge,
François Durocher, Simon‐Pierre Gingras, David Drolet, Damien Aiello,
Miles Thompson, Erasto Marcano, Isabelle Deschênes, Marc‐Olivier Nadon et
Catherine Vallières. Un merci tout particulier à Oliver D’Amours, Giselle Wagner,
Louis Vincent St‐Hilaire‐Gravel et Mark Dionne pour leur énergie sans borne, leur
enthousiasme contagieux et la qualité du travail accompli, à mes collègues de
laboratoire Jean‐François Raymond, Julien Gaudette, Carlos Gaymer, Martin Guay et
Thierry Gosselin pour les étés de terrains exceptionnels passés en leur compagnie à
Havre‐Saint‐Pierre, puis à Benoît Roberge et Nancy Dénommée de Parcs Canada
pour le temps consacré à partager avec moi le fruit de leurs observations sur la
formation hivernale des glaces dans l’archipel de Mingan.
Un gros merci à Jean‐Marc Daigle, ainsi qu’à son frère Gaétan, puis à
Eva Bessac pour leur exceptionnelle patience à m’aider dans mes analyses et à me
transmettre, pendant d’innombrables heures, une partie de leur grande connaissance
de ce monde fascinant que constituent les statistiques.
Jamais je n’exprimerai assez de reconnaissance envers ma mère Patricia, mon
père Richard, mon frère Gabriel, puis ma sœur Sophie qui ont été de tous les instants
présents. Votre sensibilité, vos encouragements, votre support et votre aide à tous les
niveaux ont nettement dépassé tout ce que je ne pouvais même oser espérer. Pour
leur accueil si chaleureux à Havre‐Saint‐Pierre, je remercie grandement Michelle

VII
Maloney et Clément Cormier dont la gentillesse n’a d’égal que la grandeur de leur
cœur. Merci aussi à mes bons amis Louis‐Philippe, Geneviève, Gaétan, Olivier, Paul,
Jean‐Claude, Roger, Stéphane, Mélanie, Hugo, Catherine et Annie‐Claude pour leurs
encouragements soutenus et leur compréhension. Vous étiez bien présents à mes
côtés malgré la distance…
Mais par‐dessus tout, j’offre mes plus chaleureux remerciements à ma très
chère Nellie, l’étoile de ma vie et ma confidente, pour avoir brillamment dirigé la
barque lors des moments de brouillard et de grandes vagues et pour son
extraordinaire patience, sa douceur, son entrain, sa compréhension et son support
moral et affectif inconditionnels… une femme tout à fait exceptionnelle que j’admire
profondément.
Merci à toutes et à tous!

VIII
Table des matières
Sommaire ..............................................................................................................................I
Résumé ............................................................................................................................ II
Abstract .......................................................................................................................... III
Avant‐propos .......................................................................................................................... IV
Table des matières ................................................................................................................. VIII
Chapitre I. Introduction générale...................................................................................... 1
Chapitre II Temporal variation in community interfaces: kelp bed boundary dynamics adjacent to persistent urchin barrens ...................................... 11
Résumé ..................................................................................................................................... 12 Abstract .................................................................................................................................... 13 Introduction ............................................................................................................................. 14 Materials and methods........................................................................................................... 18
Study sites ...................................................................................................................... 18 Temporal variability in A. esculenta beds and urchin populations........................ 18 Effect of urchins on algal recruitment in the barrens .............................................. 21 Limitation of upward movements of urchins by A. cribrosum ............................... 22 Statistical analysis ......................................................................................................... 23
Results....................................................................................................................................... 24 Temporal variability in the lower limit of kelp beds............................................... 24 Structure of kelp beds................................................................................................... 27 Structure of urchin populations.................................................................................. 30 Effect of urchins on algal recruitment in the barrens .............................................. 34 Limitation of upward movements of urchins by A. cribrosum ............................... 36
Discussion ................................................................................................................................ 36

IX
Chapitre III. Spatial and temporal stability in algal assemblages on urchin
barrens in the northern Gulf of St. Lawrence .......................................... 44
Résumé ..................................................................................................................................... 45 Abstract .................................................................................................................................... 46 Introduction ............................................................................................................................. 47 Materials and methods........................................................................................................... 49
Temporal variability in abundance and distribution of A. cribrosum.................... 49 Temporal variability in the size of A. cribrosum stands........................................... 50 Distribution of algae and urchins near A. cribrosum ................................................ 50 Effect of A. cribrosum canopy on recruitment and abundance of algae ................ 51 Statistical analysis ......................................................................................................... 51
Results....................................................................................................................................... 53 Temporal variability in abundance and distribution of A. cribrosum.................... 53 Temporal variability in the size of A. cribrosum stands........................................... 56 Distribution of algae and urchins near A. cribrosum ................................................ 56 Effect of A. cribrosum canopy on recruitment and abundance of algae ................ 56
Discussion ................................................................................................................................ 63
Chapitre IV. Algal colonization in urchin barrens: defense by association during recruitment of the brown alga Agarum cribrosum ..................... 67
Résumé ..................................................................................................................................... 68 Abstract .................................................................................................................................... 69 Introduction ............................................................................................................................. 70 Materials and methods........................................................................................................... 72
Study sites ...................................................................................................................... 72 Distribution of juvenile A. cribrosum and urchins near D. viridis........................... 73 Protection provided by D. viridis ................................................................................ 74 Growth of juvenile A. cribrosum.................................................................................. 75 Seasonal cycle of D. viridis ........................................................................................... 76 Statistical analysis ......................................................................................................... 76
Results....................................................................................................................................... 78 Distribution of juvenile A. cribrosum and urchins near D. viridis........................... 78 Protection provided by D. viridis ................................................................................ 81 Growth of juvenile A. cribrosum.................................................................................. 85 Seasonal cycle of D. viridis ........................................................................................... 85
Discussion ................................................................................................................................ 88

X
Chapitre V. Multiple plant defenses against herbivores: the interplay of
chemical, morphological, and hydrodynamic factors............................. 93
Résumé ..................................................................................................................................... 94 Abstract .................................................................................................................................... 95 Introduction ............................................................................................................................. 96 Materials and methods........................................................................................................... 99
Response of urchins to odors of Desmarestia viridis ............................................... 100 Mechanical repulsion of urchins by Desmarestia viridis......................................... 101 Effect of urchin density on attack success on Desmarestia viridis ......................... 103 Field observations ....................................................................................................... 103 Statistical analysis ....................................................................................................... 103
Results..................................................................................................................................... 105 Response of urchins to odors of Desmarestia viridis ............................................... 105 Mechanical repulsion of urchins by Desmarestia viridis......................................... 108 Effect of urchin density on attack success on Desmarestia viridis ......................... 111 Field observations ....................................................................................................... 111
Discussion .............................................................................................................................. 111 Appendix 5.1.......................................................................................................................... 118
Chapitre VI. Conclusion générale ................................................................................... 119
Dynamique de répartition des macrophytes .......................................................... 120 Mécanismes de défense des macrophytes............................................................... 121 Portée de l’étude ......................................................................................................... 122 Nouvelles questions et hypothèses .......................................................................... 124
Bibliographie ......................................................................................................................... 127

Chapitre I
Introduction générale

2
Toute communauté écologique est fondamentalement dynamique (hétérogène dans
le temps et l’espace) car les organismes qui la composent sont en constante interaction
(entre eux et avec les propriétés changeantes de leur environnement physico‐
chimique) et s’affectent dans leur distribution et leur abondance relatives (Connell,
1961; Paine, 1966; Holling, 1973; Schoener, 1974; Branch, 1975; Menge, 1976; Sousa,
1984; Sih et al., 1985). La stabilité d’une communauté réfère à sa capacité de
conservation de son état (composition) malgré la force exercée par une perturbation
(Holling, 1973; Krebs, 2001). Les perturbations (aléatoires ou cycliques) sous forme de
variations dans les conditions physiques (e.g., feu, vent, vagues), d’invasions
d’espèces compétitrices et d’ennemis naturels peuvent déplacer l’état d’équilibre
(avant perturbation) d’une communauté vers un état stable alternatif
(Connell & Slatyer, 1977; White, 1979; Ebeling et al., 1985; Chapman & Johnson, 1990;
Vadas et al., 1990; Dayton et al., 1992; Trowbridge, 1995; Ruiz et al., 1997;
Underwood, 1999). Cet état d’équilibre (climax) est l’aboutissement d’un patron de
colonisation et d’extinction des espèces (succession) non saisonnier, directionnel et
continu (Odum, 1969; Begon et al., 1996) dirigé par un complexe de processus
interactifs (e.g., prédation, compétition, immigration et émigration) parmi lesquels
certains en neutralisent d’autres (Odum, 1969; Underwood & Jernakoff, 1981;
Witman, 1987; Hendrix et al., 1988; Davidson, 1993; Benedetti‐Cecchi, 2000;
Duffy & Hay, 2000). Une communauté n’est donc jamais à un point d’équilibre exact
car l’état de celle‐ci se retrouve dans un flux constant (Lewontin, 1969; Odum, 1969;
Holling, 1973; Sutherland, 1974; Gray, 1977).
L’herbivorie peut exercer un impact considérable sur la structure et la
dynamique des communautés végétales (Ogden & Lobel, 1978; Witman, 1987;
Lindroth, 1989; Weis & Berenbaum, 1989; Brawley, 1992; Vadas & Elner, 1992;
Vasquez & Buschmann, 1997; Zamora et al., 1999; Duffy & Hay, 2001) tel que
démontré par l’effet dramatique d’herbivores sur la distribution et l’abondance des
organismes végétaux (Jones & Kain, 1967; Breen & Mann, 1976b; Dean et al., 1984;
Carpenter, 1986; Lewis, 1986; Morrison, 1988; Lindroth, 1989; Duffy & Hay, 2000).
Dans une communauté où le recrutement et la survie des végétaux sont limités par

3
l’herbivorie, la réduction de l’abondance des herbivores par une perturbation
(e.g., tempêtes, maladies et prédation) peut provoquer le déclenchement d’une
succession végétale secondaire menant à l’établissement d’une communauté dont la
richesse et la biomasse spécifiques diffèrent grandement de celles de la communauté
maintenue par les herbivores (Paine & Vadas, 1969; Pearse & Hines, 1979; Miller,
1985; Lindroth, 1989; Brown & Gange, 1992; Sousa & Connell, 1992;
Leinaas & Christie, 1996; Scheibling et al., 1999). L’impact d’un herbivore sur une
communauté végétale est non seulement fonction de son abondance, mais également
de sa taille par rapport aux organismes végétaux, de ses interactions avec les
compétiteurs et les prédateurs, ainsi que de l’abondance, de la productivité
(recrutement ou croissance) et de la richesse spécifique des organismes végétaux
(Brown & Ewel, 1987; Sousa & Connell, 1992). L’état d’une communauté végétale
repose donc en partie sur un complexe d’interactions plantes‐herbivores à l’intérieur
desquelles l’exploitation des ressources végétales par un herbivore particulier est
déterminée par les conditions environnementales (e.g., température, vents, courants),
la disponibilité de la ressource, puis les attributs individuels des plantes
(e.g., digestibilité et valeur énergétique) et de l’herbivore (e.g., degré de mobilité,
préférences alimentaires) (Emlen, 1966; MacArthur & Pianka, 1966; Menge, 1972;
Charnov, 1976).
On distingue par ailleurs deux composantes dans le comportement de quête
alimentaire d’un animal (herbivore): (1) le comportement qui lui permet d’entrer en
contact avec sa proie (une plante) et (2) sa réponse à cette source de nourriture une
fois le contact établi (Chapman & Underwood, 1992). Comme les plantes ne peuvent
se déplacer et fuir en cas de besoin, leur vulnérabilité aux attaques d’un herbivore est
particulièrement élevée et le contact entre l’herbivore et la plante est donc facilité par
rapport à une proie mobile. À ce titre, les herbivores sont d’importants agents de
sélection contre lesquels les plantes ont développé des adaptations structurales et
métaboliques remarquables qui diminuent cette vulnérabilité en modulant la réponse
alimentaire de l’herbivore (Lindroth, 1989; Weis & Berenbaum, 1989; Hay & Fenical,
1992; Coley & Barone, 1996; Duffy & Hay, 2001). Les plantes utilisent l’un ou l’autre

4
de deux mécanismes de résistance à l’herbivorie: la tolérance et l’évitement
(Duffy & Hay, 1990; Rosenthal & Kotanen, 1994). Une plante peut tolérer la présence
d’un herbivore grâce à un taux de croissance supérieur (surcompensatoire) au taux de
broutage de l’herbivore, à une activité photosynthétique accrue, à l’utilisation de
produits de réserves et au retard de certaines étapes de sa phénologie (McNaughton,
1983; Lindroth, 1989; Davidson, 1993; Tiffin, 2000). Pour éviter un herbivore, une
plante peut (1) produire des défenses structurales (e.g., aiguilles, épines, tissus
calcifiés, Steneck, 1985; Myers & Bazely, 1991) et chimiques (e.g., métabolites
secondaires comprenant terpènes, alkaloïdes et phénols, Hay & Fenical, 1988;
Karban & Myers, 1989; Schappert & Shore, 1999; Cetrulo & Hay, 2000; Pavia & Toth,
2000; Becerra et al., 2001; Schnitzler et al., 2001) permettant de repousser l’herbivore
ou de réduire son taux d’alimentation ou (2) utiliser un refuge temporel
et/ou bénéficier d’un refuge spatial réduisant les probabilités de contact avec
l’herbivore (Weis & Berenbaum, 1989).
Un cas particulier de refuge spatio‐temporel, connu sous le nom de refuge
(ou défense) par association, survient lorsqu’une plante échappe à l’herbivorie en
s’associant à d’autres plantes (ou animaux) plus ou moins attirantes pour l’herbivore
(Tahvanainen & Root, 1972; Littler et al., 1987; Pfister & Hay, 1988; Duffy & Hay,
1990; Kerr & Paul, 1995). L’association peut accroître la compétition pour les
ressources (e.g., espace, lumière, nutriments) et engendrer une perte de croissance
pour l’une ou l’autre des espèces végétales impliquées (Hay, 1986; McAuliffe, 1986;
Pfister & Hay, 1988; Forseth et al., 2001). Pour l’espèce protégée, cette perte est
cependant compensée par le gain de protection permettant la survie. La majorité des
études sur les mécanismes de défense des plantes impliquent des individus adultes.
Or, les juvéniles sont particulièrement vulnérables aux attaques des herbivores car, en
général, leurs attributs structurels et chimiques répulsifs ne sont pas assez développés
pour procurer une défense efficace contre les herbivores (Davidson, 1993;
Van Alstyne et al., 1999) et ils ne disposent pas d’une taille refuge (sensu Paine, 1976;
Lubchenco & Gaines, 1981) pouvant gêner le broutage ou minimiser l’impact de

5
pertes tissulaires. La défense par association peut donc s’avérer déterminante pour la
survie des juvéniles et l’évitement de l’extinction locale d’une espèce.
L’oursin vert, Strongylocentrotus droebachiensis, peuple les fonds des régions
arctiques‐boréales des océans Atlantique et Pacifique (Mortensen, 1943; Jensen, 1974;
Bazhin, 1998). Bien qu’on le retrouve de la limite des marées basses jusqu’à 300 m de
profondeur, il est plus commun en zone peu profonde, à moins de 50 m de
profondeur (Jensen, 1974; Scheibling & Hatcher, 2001) et généralement associé à des
substrats de type rocheux (e.g., roche‐mère, blocs, cailloux, Himmelman, 1986;
Scheibling & Raymond, 1990). L’oursin vert est principalement herbivore mais
consomme également des carcasses animales ainsi que d’autres invertébrés et des
proies bactériennes (Himmelman & Steele, 1971; Lawrence, 1975; Duggins, 1981;
Johnson & Mann, 1982; Briscoe & Sebens, 1988; Nestler & Harris, 1994). Il tolère, sans
mal apparent, des périodes de jeûne prolongé (jusqu’à un mois, Garnick, 1978). Il
utilise la chémodétection à distance pour détecter sa nourriture et affiche un haut
degré de sélectivité (préférence) alimentaire (Vadas, 1977; Garnick, 1978; Larson et al.,
1980; Sloan & Campbell, 1982; Mann et al., 1984; Himmelman & Nédélec, 1990; Prince
& LeBlanc, 1992). Cette sélectivité est cependant fortement influencée par des facteurs
environnementaux tels que la disponibilité des ressources, l’action du courant et des
vagues, la température, la compétition et la prédation (Lawrence, 1975; Vadas, 1977;
Himmelman, 1984; Vadas, 1990; Scheibling, 1996). Le taux d’alimentation individuel
de S. droebachiensis fluctue de façon saisonnière, habituellement avec un pic en
période estivale et un creux en période hivernale (Vadas, 1977; Larson et al., 1980;
Himmelman, 1984; Meidel & Scheibling, 1998). La production et la maturation des
tissus gonadiques débutent généralement à l’automne et l’expulsion des gamètes
survient en début de printemps, vraisemblablement en conjonction avec
l’augmentation rapide de la production phytoplanctonique (Himmelman, 1975;
Falk‐Petersen & Lonning, 1983; Starr et al., 1990; Meidel & Scheibling, 1998).
Dans le nord‐ouest de l’Atlantique, S. droebachiensis exerce une forte influence
sur la distribution et l’abondance des macrophytes (Vadas & Elner, 1992). Le

6
développement rapide (quelques mois) d’assemblages végétaux luxuriants à la suite
d’exclusions manuelles et naturelles de l’oursin témoigne de son impact négatif sur le
recrutement et la survie algale (Himmelman et al., 1983; Scheibling, 1986; Keats et al.,
1990). Néanmoins, les portions côtières rocheuses peu profondes exposées aux
vagues supportent habituellement de vastes et compactes bandes végétales dominées
par les laminaires, communément appelées ceintures de laminaires (Vadas & Elner,
1992; Lobban & Harrison, 1994; Graham & Wilcox, 2000; Tegner & Dayton, 2000).
L’importance écologique de ces ceintures est énorme car en plus de figurer parmi les
systèmes les plus productifs au monde (Mann, 1973), elles procurent un habitat à
plusieurs espèces pour le recrutement, l’alimentation et la reproduction, de même
qu’un refuge contre les prédateurs et l’action des vagues et des courants
(Himmelman et al., 1983; Himmelman & Lavergne, 1985; Vadas & Elner, 1992;
Bologna & Steneck, 1993; Vasquez, 1993; Bégin, 2002; Gagnon et al., 2003a). Des
fluctuations substantielles au sein des populations d’oursins pourraient donc avoir de
profondes répercussions (positives ou négatives) sur les communautés végétales
(e.g., ceintures de laminaires) et, en bout de ligne, sur la structure du reste de la
communauté infralittorale. Tel est le cas sur la côte atlantique de la Nouvelle‐Écosse:
la recrudescence épisodique d’un pathogène (amibe) provoque l’effondrement massif
des populations d’oursins et résulte en l’expansion rapide des ceintures de laminaires
(Miller & Colodey, 1983; Scheibling & Stephenson, 1984; Scheibling, 1986;
Scheibling & Hennigar, 1997).
Dans l’archipel de Mingan, au nord du golfe du Saint‐Laurent, S. droebachiensis
domine la zone infralittorale rocheuse peu profonde où sa densité est, en général, très
élevée (plus de 300 individus m‐2, Himmelman & Nédélec, 1990). Cette zone se
caractérise par un paysage subdivisé en deux communautés végétales qui diffèrent
nettement en terme de biomasse et de richesse spécifiques. L’une d’elles, la zone des
laminaires, occupe les premiers mètres d’eau et est composée d’un dense assemblage
végétal dominé par Alaria esculenta et, de façon moins importante, par d’autres
espèces du groupe des laminaires (e.g., Laminaria spp., Sacchoriza dermatodea).
À l’inverse, la zone dénudée, attenante à la zone des laminaires, est composée d’une

7
mosaïque de plaques végétales parsemant de vastes tapis d’algues calcaires
(e.g., Lithothamnion sp., Clathromorphum sp.) résistantes au broutage (Steneck, 1983;
Steneck, 1985; Steneck, 1986). L’abondance de l’oursin y est nettement plus élevée que
dans la zone des laminaires. La transition entre les deux communautés végétales est
habituellement abrupte et coïncide avec la présence de fronts d’oursins compacts à la
limite inférieure des ceintures de laminaires.
Les phaéophycées Agarum cribrosum et Desmarestia viridis sont les seules
macroalgues charnues communément rencontrées en zone dénudée, ce qui soulève
d’intéressantes questions quant aux attributs et mécanismes leur permettant de
croître dans cet environnement que l’herbivorie par l’oursin rend si hostile à la
végétation. La laminaire pérenne, A. cribrosum, possède un large crampon, un stipe
semi‐rigide et une large fronde ondulée. On la retrouve principalement en petits
agrégats (1‐10 m2), bien qu’elle forme à l’occasion de larges bandes (>500 m2),
particulièrement en bordure de la limite inférieure des ceintures de laminaires.
Sa croissance est lente comparativement aux autres espèces de laminaires
(Vadas, 1968). La présence de composés chimiques répulsifs dans les tissus
d’A. cribrosum (e.g, phénols) expliquerait sa faible consommation par l’oursin vert
(Vadas, 1977; Larson et al., 1980; Himmelman & Nédélec, 1990). Si les adultes
d’A. cribrosum apparaissent peu vulnérables à la prédation par l’oursin, des
observations exploratoires de la distribution des juvéniles suggèrent que ces derniers
le sont grandement. Peu de juvéniles sont localisés aux endroits facilement accessibles
par l’oursin (e.g., aires ouvertes). Ils semblent pour la plupart confinés à l’intérieur
d’agrégats de conspécifiques adultes et sous le couvert de D. viridis. Le recrutement et
la survie des juvéniles d’A. cribrosum pourrait donc dépendre d’associations algales
intra‐ et interspécifiques contre le broutage de l’oursin (e.g., refuge par association).
À l’inverse, D. viridis est une espèce annuelle dotée d’un petit crampon, d’un
stipe hautement flexible et d’une large fronde extrêmement ramifiée. La production et
l’accumulation par D. viridis d’acide sulfurique à l’intérieur de vacuoles spécialisées
la rend très acide (pH 0.5‐0.9, McClintock et al., 1982; Sasaki et al., 1999). L’utilité

8
défensive d’une telle acidité contre l’herbivorie (défense chimique) a été suggérée à
maintes reprises chez plusieurs espèces de Desmarestiales (dont D. viridis), sans
toutefois avoir fait l’objet d’investigations rigoureuses (Anderson & Velimirov, 1982;
Dayton, 1985a; Thompson, 1988; Himmelman & Nédélec, 1990; Lobban & Harrison,
1994). Par ailleurs, des observations in situ ont indiqué que l’oursin semble éviter le
contact avec des frondes de D. viridis agitées par les vagues mais qu’il s’agrège
rapidement, en eau calme, sur des frondes immobiles (Himmelman & Steele, 1971;
Himmelman, 1984; Konar, 2000). Le mouvement de balayage de l’algue causé par
l’action des vagues pourrait donc également contribuer à maintenir l’oursin à
distance (défense mécanique). Une investigation de la réponse comportementale de
S. droebachiensis face à D. viridis permettrait de déterminer la contribution d’une
défense chimique et/ou mécanique à la capacité répulsive de l’algue contre l’oursin.
Aucune étude n’a encore quantifié l’impact de l’oursin vert sur la structure et
la dynamique de répartition naturelle des assemblages algaux dans le nord du golfe
du Saint‐Laurent. Certes, les quelques études réalisées dans (ou près de) cette région
appuient le concept de sélectivité alimentaire chez l’oursin et soulignent son effet
négatif sur le recrutement et la survie algale (Himmelman, 1969; Himmelman et al.,
1983; Himmelman, 1984; Himmelman & Nédélec, 1990; Keats et al., 1990). Cependant,
(1) la nature ponctuelle de ces études, (2) l’étroitesse des échelles spatiales impliquées,
(3) les approches expérimentales utilisées (perturbations anthropiques) et (4) le
manque de relativisation de l’impact de l’oursin sur la structure et la dynamique des
communautés de macrophytes par rapport à l’impact d’autres facteurs (e.g., abrasion
par les glaces, force des courants, compétition intra‐ et interspécifique) ne permettent
pas de dresser de portrait rigoureux des mécanismes écologiques qui alimentent la
dynamique communautaire naturelle et, encore moins, de quantifier cette dernière.
Plusieurs interrogations fondamentales ont motivé la tenue de la présente
étude sur les interactions algues‐oursins dans le nord du golfe du Saint‐Laurent.
Notamment, quels patrons de variation temporelle caractérisent la structure et la
distribution des communautés de macrophytes et des populations d’oursins?

9
Ces patrons s’appliquent‐ils uniformément à l’ensemble des localités ou varient‐ils
plutôt d’un endroit à l’autre? Dans quelle mesure le broutage de l’oursin affecte‐il la
stabilité des communautés végétales et quelle est son importance par rapport à
d’autres facteurs? Finalement, quels sont les mécanismes qui permettent à certains
macrophytes de résister au broutage de l’oursin? En l’absence de réponses à ces
questions, notre connaissance des facteurs et processus écologiques qui façonnent cet
écosystème est limitée, d’où la nécessité d’entreprendre l’examen attentif des
interactions qu’entretiennent l’oursin vert et les communautés de producteurs
primaires.
L’objectif de mon étude est double: (1) caractériser la dynamique de répartition
des macrophytes de la zone infralittorale et identifier les principaux facteurs à
l’origine de cette dynamique dans l’archipel de Mingan, puis (2) examiner les
mécanismes défensifs utilisés par certains macrophytes dominants contre le broutage
de l’oursin vert. Deux chapitres caractérisent la dynamique de répartition des
assemblages algaux, l’un au voisinage de l’interface entre la zone des laminaires et la
zone dénudée, l’autre dans la zone dénudée, puis deux autres chapitres examinent les
interactions entre l’oursin vert et les phaéophycées D. viridis et A. cribroum.
Mes travaux résultent d’une combinaison d’observations et d’expériences effectuées
en milieu naturel en utilisant le scaphandre autonome (SCUBA), puis en laboratoire, à
l’aide de dispositifs et d’appareils décrits dans la littérature scientifique
(e.g., labyrinthe en Y) ou que j’ai conçus afin de contrôler adéquatement les
paramètres de certaines expériences qui ne pouvaient être réalisées en milieu naturel
(e.g., générateur de vagues oscillatoires). Le chapitre II examine les facteurs affectant
la stabilité temporelle de la limite inférieure des ceintures de laminaires en accordant
une attention toute particulière aux impacts du broutage de l’oursin vert. Le
chapitre III évalue la stabilité temporelle de la distribution et de l’abondance des
assemblages algaux de la zone dénudée dominés par A. cribrosum. Le chapitre IV teste
l’hypothèse d’une défense par association permettant aux juvéniles d’A. cribrosum
d’éviter le broutage de l’oursin vert. Finalement, le chapitre V examine les

10
mécanismes par lesquels D. viridis résiste aux attaques de l’oursin en évaluant
l’importance relative d’une défense chimique et mécanique.

Chapitre II
Temporal variation in community interfaces: kelp bed boundary dynamics adjacent to persistent urchin barrens

12
Résumé
Nous avons examiné, pendant deux ans, les facteurs qui affectent la stabilité
temporelle de la limite inférieure des ceintures de laminaires (Alaria esculenta) sur
cinq sites infralittoraux dans l’archipel de Mingan, dans le nord du golfe du
Saint‐Laurent. La position de la limite inférieure des ceintures variait beaucoup entre
les sites et dans le temps et était principalement contrôlée par l’oursin vert,
Strongylocentrotus droebachiensis, qui s’agrégeait en de denses (jusqu’à
500 individus m‐2) fronts de broutage en marge inférieure des ceintures. Ces fronts
avançaient sur les laminaires le plus rapidement pendant l’été (à des taux aussi élevés
que 2.5 m mois‐1), et il semblait y avoir une biomasse seuil d’oursins d’environ
5 kg m‐2 en‐dessous de laquelle les fronts ne pouvaient réduire substantiellement la
limite des ceintures. Les fronts étaient principalement composés de gros individus,
tandis que les plus petits oursins dominaient en zone dénudée, au‐dessous des
ceintures. À l’un des sites, nous avons noté de grands changements saisonniers dans
les densités globales d’oursins avec des augmentations et des baisses importantes
pendant l’été et l’hiver, respectivement. Une expérience comportant l’exclusion
d’oursins a indiqué que le recrutement algal en zone dénudée était de plus de cent
fois supérieur en absence qu’en présence d’oursins. La laminaire Agarum cribrosum
limitait grandement les mouvements de l’oursin et la stabilité la plus élevée d’une
ceinture sur l’un des sites semblait liée au remplacement graduel d’A. esculenta, dans
la partie inférieure de la ceinture, par une large bande d’A. cribrosum. Nous
proposons que des perturbations causées par des facteurs abiotiques (e.g., abrasion
par les glaces et mouvement de l’eau) déclenchent d’importants changements locaux
dans les densités d’oursins qui, à leur tour, déterminent principalement les limites de
distribution des ceintures de laminaires dans cette région de l’Atlantique où la zone
dénudée est un état communautaire dominant.

13
Abstract
We examined, over 2 years, factors affecting the temporal stability of the lower limit
of kelp beds (Alaria esculenta) at five subtidal sites in the Mingan Islands, northern
Gulf of St. Lawrence. The position of the lower limit of the beds varied markedly
among sites and over time and was largely controlled by the green sea urchin,
Strongylocentrotus droebachiensis, that formed dense (up to 500 individuals m‐2) feeding
fronts at the lower edge of the beds. These aggregations advanced over the kelp most
rapidly during the summer (at rates as high as 2.5 m month‐1), and there appeared to
be a threshold urchin biomass of ~5 kg m‐2 below which the fronts could not
substantially reduce the limit of the beds. The fronts consisted mainly of large
individuals, whereas smaller urchins predominated in the barrens zone below the
kelp beds. At one site, we recorded large seasonal shifts in overall urchin densities,
with large increases and decreases during the summer and winter, respectively.
An urchin exclusion experiment indicated that algal recruitment in the barrens was
two orders of magnitude greater in the absence than in the presence of urchins.
The kelp Agarum cribrosum greatly restricted urchin movements, and the greater
temporal stability of the kelp bed at one site appeared related to the gradual
replacement of A. esculenta in the lower kelp bed by a large stand of A. cribrosum.
We propose that perturbations by abiotic factors (e.g., ice scouring and water motion)
trigger important but localized changes in urchin densities which, in turn, largely
determine the limits of kelp bed distribution in this region of the Atlantic where
urchin barrens are a persistent community state.

14
Introduction
Kelp beds are highly diverse shallow coastal communities found in temperate and
polar zones worldwide. They are organized around the physical structure and
primary productivity provided by large brown algae of the order Laminariales
(Tegner & Dayton, 2000). In addition to being among the most productive systems on
earth (Mann, 1973), kelp beds provide habitat to many species for recruitment,
feeding, reproduction, and refuge from predators and from strong water motion
(e.g., Bernstein et al., 1981; Himmelman et al., 1983; Himmelman & Lavergne, 1985;
Duggins et al., 1990; Vadas & Elner, 1992; Vasquez, 1993; Balch & Scheibling, 2000;
Gagnon et al., 2003a). Although the light environment and substratum type are
primary factors determining where kelp beds develop, many other factors, such as
sedimentation, nutrients, salinity, water motion, and ice scour can modify local
patterns of occurrence (see review in Dayton, 1985b; but also Hooper, 1981;
Cowen et al., 1982; Ebeling et al., 1985; Keats et al., 1985; Seymour et al., 1989;
Sivertsen, 1997; Kawamata, 2001). The most important factor in many regions is
grazing by sea urchins (see review in Lawrence, 1975; but also Paine & Vadas, 1969;
Pearse & Hines, 1979; Hagen, 1983; Dean et al., 1984; Himmelman, 1984; Dayton,
1985a; Witman, 1987; Leinaas & Christie, 1996; Sivertsen, 1997;
Vasquez & Buschmann, 1997). Although urchins and kelp can co‐occur at small
spatial scales (i.e., within the foraging distance of urchins), high densities of urchins
usually completely denude areas of fleshy macrophytes, including kelp. Such areas,
referred to as urchin barrens (Lawrence, 1975), have both low diversity (but see
Bégin, 2002) and low productivity, although the abundance and diversity of coralline
algae can be quite high (Steneck, 1983; 1986; Chapman & Johnson, 1990;
Scheibling & Hatcher, 2001). Urchin barrens can be both extensive and persistent
(Breen & Mann, 1976b; Chapman, 1981; Hagen, 1983; Himmelman et al., 1983; Miller,
1985; Scheibling, 1986; Leinaas & Christie, 1996; Sivertsen, 1997), and natural and
experimental removals of urchins demonstrate that kelp can rapidly colonize barrens
when urchins are absent (e.g., Pearse & Hines, 1979; Himmelman et al., 1983;
Scheibling, 1986; Witman, 1987; Keats et al., 1990; Scheibling et al., 1999).

15
In the northwestern North Atlantic, the green sea urchin, Strongylocentrotus
droebachiensis, severely limits the distribution of kelp in rocky subtidal environments
and often forms dense feeding aggregations at the lower edge of kelp beds. This
phenomenon is reported in the Gulf of Maine (Sebens, 1985; Witman, 1985; 1987),
Newfoundland (Himmelman, 1984; Keats et al., 1985; Keats et al., 1990), and the
St. Lawrence Estuary (Himmelman et al., 1983; Himmelman & Lavergne, 1985).
In Nova Scotia (the most extensively studied region), urchin fronts can destroy kelp
beds (primarily Laminaria longicruris) at rates as high as 4 m month‐1, and as long as
the urchins remain sufficiently abundant, fleshy macroalgae cannot recolonize
(Breen & Mann, 1976a; b; Chapman, 1981; Miller, 1985; Scheibling, 1986; Scheibling et
al., 1999). As outlined by Scheibling et al. (1986; 1999), mass mortality of
S. droebachiensis caused by a pathogenic amoeba (see review in Scheibling, 1988) is the
only known natural mechanism for the large‐scale shift in community state from
urchin barrens to kelp beds. Such mortalities have been reported a number of times
over the last decades and were always followed by rapid kelp colonization
(Miller & Colodey, 1983; Scheibling & Stephenson, 1984; Miller, 1985; Scheibling,
1986; Scheibling & Hennigar, 1997; Scheibling et al., 1999). Although lobsters, crabs,
sea stars, wolffish, sea gulls and eider ducks are all predators of S. droebachiensis in the
northwestern North Atlantic (Himmelman & Steele, 1971), they have never been
shown to regulate urchin populations (see review in Scheibling, 1996).
Two such community states are found on shallow bedrock platforms in
wave‐exposed areas in the Mingan Islands, northern Gulf of St. Lawrence: (1) kelp
beds dominated by Alaria esculenta (but also including Laminaria spp., Saccorhiza
dermatodea, and Agarum cribrosum) in shallow water, and (2) urchin barrens
dominated by S. droebachiensis. Within the kelp bed, urchin densities are low, and
algal and invertebrate biomass is high (Bégin, 2002). In the barrens, urchins often
attain densities of 300 m‐2 (Himmelman & Nédélec, 1990; Gagnon et al., 2003b), and
the main algae are crustose coralline red algae (Lithothamnion sp. and Clathromorphum
sp.). The only common fleshy macroalgae in the barrens are the brown algae Agarum
cribrosum and Desmarestia viridis and the red alga Ptilota serrata. In most locations

16
A. cribrosum is found near the lower limit of the kelp zone where it usually forms
small patches (1‐10 m2), but it occasionally covers larger areas (>500 m2). Desmarestia
viridis, a filamentous annual, also forms patches in urchin barrens, but the patches are
typically small (1‐5 m2) and are only present from winter to early fall (Gagnon et al.,
2003b). Both A. cribrosum and D. viridis are thought to be chemically defended from
urchin grazing (Vadas, 1977; Larson et al., 1980; Himmelman & Nédélec, 1990;
Lobban & Harrison, 1994) although more recent work suggests that tactile contact
induced by water movement is more important, particularly for the finely‐branched
D. viridis (Gagnon et al., 2003b). Ptilota serrata often grows under patches of
A. cribrosum but can also form extensive turfs at greater depths.
Mass mortalities of urchins, as occur in Nova Scotia, has never been reported
in the northern Gulf. Thus, the urchin barrens appear to be a persistent community
state. The transition between kelp beds and barrens is distinct as the kelp generally
abruptly disappear where fronts of urchins begin (Fig. 2.1). The factors controlling the
stability of this conspicuous interface are poorly understood. Qualitative observations
over 20 years (Himmelman and coworkers) suggest that its position (i.e., its depth
and distance from shore) varies over time from site to site (with wave exposure and
slope). Although the kelp‐barrens interface represents an equilibrium in an important
herbivore‐plant interaction, there are no long‐term observations of how it fluctuates
over time, or an understanding of how such changes are affected by other
environmental factors (e.g., ice scour, dislodgment by waves and currents,
competition). A necessary first step is a detailed examination of the spatial and
temporal dynamics of this interface relative to the demographic changes in both the
kelp bed and the urchin population.
Here, we examine factors affecting the temporal stability of the lower limit of
kelp beds in the Mingan Islands. Specifically, we (1) record temporal variations in the
position of the lower limit of kelp beds, (2) measure temporal changes in the structure
(density and size‐distribution) of kelp beds and urchin fronts in the transition zone,

17
Fig. 2.1. A grazing front of the green sea urchin, Strongylocentrotus droebachiensis (2 to 3 layers thick), at a depth of 3‐4 m at the lower edge of a subtidal kelp (Alaria esculenta) bed in the Mingan Islands in June 2000. The urchins are actively consuming kelp blades and leaving only the bases of stipes in their path.

18
(3) examine the relationship between urchin biomass in grazing fronts and the rate
of change in the position of the lower limit of kelp beds, (4) determine the impact
of urchins on the recruitment of kelp in the barrens zone, and (5) assess the
effectiveness of A. cribrosum stands in limiting upward movements of urchins into the
shallower kelp zone dominated by A. esculenta.
Materials and methods
Study sites
Our study was conducted during the summer and fall of 2000, 2001, and 2002 at
five subtidal sites on four of the Mingan Islands, northern Gulf of St. Lawrence
(Fig. 2.2). All sites were gently‐sloping bedrock platforms and had kelp beds in
shallow water (to a maximal depth of 6 m) and urchin barrens in deeper water. At the
beginning of the study, the kelp beds at the sites were composed primarily of
Alaria esculenta and the transition from kelp beds to barrens was abrupt. The sites
varied markedly in exposure to waves and tidal currents due to differences in
orientation and topography. After initial qualitative surveys, two sites, Île à Firmin
East and Île du Havre, were selected for intensive study due to striking differences in
algal density within the kelp beds (kelp were more dense at the former site).
The Île à Firmin East site was exposed to strong wave action and tidal currents,
whereas the Île du Havre site had less exposure to waves and had moderate tidal
currents. The three other sites, Île Niapiskau, Île à Firmin West, and Petite île au
Marteau, were studied less intensively and served to provide information on the
generality of the temporal patterns observed at the two intensively studied sites.
We did not make measurements during other seasons due to the remote location of
the field sites, unpredictable weather conditions, and ice cover in winter.
Temporal variability in A. esculenta beds and urchin populations
We evaluated community shifts by monitoring changes over 2 years in (1) the

19
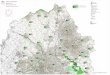
Fig. 2.2. Location of study sites at Île Niapiskau, Île à Firmin (West and East), Île du Havre, and Petite île au Marteau, in the Mingan Islands, northern Gulf of St. Lawrence, eastern Canada.

20
absolute position of the lower limit of the kelp bed, and (2) the density and
size‐distribution of kelp and urchins in the kelp‐barrens transition zone. For each
of the two intensively studied sites, we delineated 12 permanent 16‐m‐long transects
that ran perpendicular to the kelp‐barrens transition and parallel to one another.
The transects were spaced at 3‐m intervals and thus covered 33 m of shoreline.
The ends of each transect were tied to two 33‐m‐long benchmark lines (one located in
the kelp bed and the other in the barrens) running parallel to the shoreline and
anchored by bolts (3 on each line) permanently set into holes drilled into the
substratum. The transect lines were removed between sampling periods.
The absolute position of the kelp‐barrens interface was determined by
measuring the distance (to the nearest 1 cm) from the deep end of each transect up to
the interface. To measure the abundance and size of urchins and kelp, we placed
quadrats along each transect line in both seaward and landward directions from the
kelp‐barrens interface. Quadrats were positioned by placing the edge of a quadrat
(25 x 25 cm) at distances of 0.0 (i.e., at the interface), 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 4.0 and
5.0 m in each direction (this arrangement placed the two quadrats at 0.0 m
immediately adjacent to one another). In each quadrat we counted the number of
urchins in three size classes (<2 cm, 2‐4 cm, and >4 cm in test diameter). We also
categorized each quadrat according to the density of kelp (0‐10, 10‐30, 30‐50,
and >50 individuals per quadrat) and their length (<50 cm, 50‐100 cm, and >100 cm).
Sampling was done six times over a 2‐year period: June, August, and October 2000,
June and August 2001, and June 2002. We could not, however, take all measurements
each time. Bad weather precluded the measurement of the density and
size‐distribution of kelp and urchins in October 2000, and time considerations limited
the quadrat samplings in 2001 and 2002 to 5 distances (0.0, 0.5, 1.0, 3.0, and 5.0 m) in
each direction from the kelp‐barrens interface.
At the other three sites, we simply measured the absolute position of the
kelp‐barrens interface along 20 m of shoreline. At each site, a transect line was placed
along the lower limit of the kelp bed (two positions were permanently marked

21
by bolts set into the substratum), and on each observation date (August 2000,
June and August 2001, and June 2002), we recorded the perpendicular distance to the
lower limit of the kelp bed at 5‐m intervals along the transect line. The line was
removed between sampling dates.
Should grazing by urchins cause the reduction of kelp beds, the rate of kelp
bed regression should be positively correlated with urchin biomass in the grazing
fronts. To test this hypothesis, we examined the data on the position of the lower limit
of kelp beds and urchin densities acquired from the 2‐year monitoring at Île à Firmin
East and Île du Havre sites. For each site, we first determined the monthly rate
of change (advance or regression) in the mean position of the lower limit of the kelp
bed (the average over the twelve transect lines) by comparing their position at the
beginning and at the end of each of four time periods (between June and August 2000
and 2001, and between August and June 2000‐2001 and 2001‐2002). For each period,
we then calculated the urchin biomass within the first meter of the edge of the
barrens (i.e., the front). We used the average of the quadrats placed at 0.0, 0.5, and
1.0 m from the edge and converted densities to biomass using a regression
(Wet weight (g) = 0.000908 x Diameter (mm)2.804; r2 = 0.99) obtained from a sample of
144 urchins (ranging in test diameter from 0.8 to 6.1 cm) collected at the Île du Havre
site. Test diameters used for each size class (<2 cm, 2‐4 cm, and >4 cm) were 1, 3, and
5 cm, respectively, and densities were the average of the urchins sampled at the
beginning and end of each period. We then examined the relationship between
urchin biomass in the grazing fronts and monthly rate of changes in the position
of the lower limit of kelp beds at each site.
Effect of urchins on algal recruitment in the barrens
To quantify the impact of urchins on algal recruitment in the barrens,
we experimentally manipulated urchins using exclosures. The experiment consisted
of three treatments: 1. Grazed (no manipulation) to quantify algal recruitment under
natural urchin densities; 2. Ungrazed (exclosure and urchins manually removed)
to quantify algal recruitment in the absence of urchins; and 3. Fence control

22
(exclosure, but urchins not manually removed) to test for effects of the exclosure
beyond those associated with the presence of grazers.
For the Ungrazed and Fence control treatments, we constructed exclosures that
consisted of a 0.75 x 0.75‐m fence of 60‐cm‐wide strips of hardware cloth (1‐cm mesh
size) supported by a PVC frame. The fences were anchored to the substratum with
nylon ropes connecting the top of each corner to eye‐bolts set into the bedrock.
Urchins were able to climb the fences but with visits every 2‐3 d to hand remove
them, we were able to limit greatly their penetration inside the fences. Fence controls
were identical except that urchins were never removed after starting the experiment.
We selected 12 experimental areas (each marked with a bolt in the center) in
June 2000, spaced at 3.8‐m intervals 3 to 4 m below the lower limit of the kelp bed at
Île du Havre (near the transects at this location). The areas were grouped into four
blocks of three adjacent areas that were then randomly assigned the three treatments.
The experiment began on 3 July 2000, when we removed the urchins from all the
areas (releasing them at least 3 m from the areas) and then wire‐brushed the rock
surface to remove any fleshy macroalgae present (e.g., rare recruits of Laminaria sp.
and A. cribrosum), thereby increasing the homogeneity of the initial conditions of the
experimental units. The experiment ended 44 d later at which time we quantified the
density of kelp recruits in each area using two quadrats (25 x 25 cm) placed on the
north and south sides of the center bolt in each area. We averaged the data from the
two quadrats in each area before statistically comparing densities of kelp recruits
among treatments.
Limitation of upward movements of urchins by A. cribrosum
During the first year of the study, changes were seen in the kelp‐barrens transition
that were related to an observed increase in the abundance of Agarum cribrosum at the
Île du Havre site. To assess the effectiveness of mature stands of A. cribrosum in
limiting the upward movement of urchin fronts into the kelp (Alaria esculenta) zone,
we conducted an experiment to compare the rate of penetration of urchins in the
absence and presence of adult A. cribrosum. The two treatments consisted of

23
A. cribrosum removal (stipes cut off at the holdfasts) and a control (no manipulation).
Experimental units consisted of eight 4‐m2 areas spaced at 3.8‐m intervals along the
lower edge of a large (over 500 m2) mature stand of A. cribrosum located near the
transect lines at the Île du Havre site. Each area, 1‐m wide and extending 4 m up into
the A. cribrosum zone, was marked with bolts at the lower and upper edge. The areas
were grouped into four blocks of two adjacent areas to which the two treatments
were randomly assigned. The experiment began on 27 June 2001 with the removal of
all adult and juvenile A. cribrosum for the removal treatment and the removal of all
urchins (released >25 m away) from all experimental units. The movement of urchins
into the experimental units was then recorded on 6 dates over the next 20 d (June 28,
29 and 30 and July 4, 10 and 17). On each date we counted the number of urchins in
three size classes (<2 cm, 2‐4 cm, and >4 cm) in quadrats (40 x 40 cm) placed at 0, 1, 2,
and 3 m from the edge of the original A. cribrosum zone in each area. To provide an
indication of the overall urchin densities, we then averaged the densities of urchins
from the 1‐, 2‐ and 3‐m distances in each area on each date before comparing
treatments.
Statistical analysis
Temporal changes in the density of sea urchins in the kelp‐barrens transition zones
of Île à Firmin East and Île du Havre were analyzed using a repeated measures
ANOVA (Hand & Taylor, 1987; Crowder & Hand, 1990) with the factors, Time
(June 2000, July 2000, June 2001, July 2001, and June 2002) and Site (Île à Firmin East
and Île du Havre) with one nested element (transect replicate within island). Since
variances did not vary over time within the islands, and the correlation in the data
over time was similar between the islands, we used a pooled covariance structure in
the ANOVA (Proc Mixed, Type = CS, SAS Institute Inc., 1999). We applied the
analysis to the raw data as the data for each time were normally distributed.
We examined the relationship between urchin biomass in grazing fronts and
rate of change in the position of the lower limit of kelp beds at Île à Firmin East and
Île du Havre using regression analysis (Draper & Smith, 1981; Proc Reg, SAS Institute

24
Inc., 1999). We applied the analysis to the raw data as the data for each site were
homoscedastic and normally distributed. We evaluated the effect of urchins on algal
recruitment in the barrens using a two‐way ANOVA (Zar, 1999) with the factors,
Block (four) and Treatment (Grazed, Ungrazed, Exclosure control) applied to the raw
data as they were homoscedastic and normally distributed. To test whether
Agarum cribrosum impeded the upward movement of urchin fronts, we carried out a
repeated measures ANOVA with the factors, Block (four), Treatment (with/without
canopy), and Time (after 1, 2, 3, 7, 13, and 20 d) with one nested element (treatment
within block). Since we detected fluctuations in the variances over time, we modeled
the covariance structure with a heterogeneous compound symmetry structure
(Proc Mixed, Type = CSH, SAS Institute Inc., 1999). We analyzed the raw data as the
data for each time were normally distributed.
In the above analyses, normality was verified using Shapiro‐Wilk’s statistic
(SAS Institute Inc., 1999) and homoscedasticity by examining the graphical
distribution of the residuals and by applying Levene test (Snedecor & Cochran, 1989).
To detect differences among levels within a factor, we used least‐square means
multiple comparisons tests (LS means, SAS Institute Inc., 1999). A significance level
of 0.05 was used for all statistical tests.
Results
Temporal variability in the lower limit of kelp beds
During the 2‐year study, the lower limit of kelp beds showed greater temporal
variability at Île à Firmin East than at Île du Havre (Fig. 2.3). At Île à Firmin East, the
kelp bed retreated steadily during the first year and by August 2001 had been pushed
back by 11.9 ± 5.0 (SD) m (equivalent to a loss of 0.9 m month‐1). Nearly 40 % of this
loss occurred between June and August 2001 (2.5 m month‐1). There was a strong
recruitment event during the winter of 2001‐2002, and the kelp bed reinvaded
a distance of 13.7 ± 4.8 m, exceeding by 1.8 ± 2.0 m the initial lower limit. In contrast,

25
0
10
20
30
40Jun 2000 Aug 2000
Aug 2001 Jun 2002
Île à Firmin EastOct 2000
0
10
20
30
40
Île du HavreJun 2000 Aug 2000 Oct 2000
0 6 12 18 24 30
Aug 2001
0 6 12 18 24 30
Jun 2002
Alongshore distance (m)
0
10
20
30
40Jun 2001
0 6 12 18 24 300
10
20
30
40Jun 2001
Barrens
Kelp bed
(8.1±1.9) (11.6±1.9) (13.7±1.6)
(15.6±4.6) (20.5±4.2) (6.7±1.7)
(8.7±1.5) (9.4±1.9)
(9.6±1.9) (9.1±2.2)
(12.1±1.2)
(8.2±2.7)
0
0
0
0
Fig. 2.3. Changes in the position of the kelp‐barrens interface at Île à Firmin East and Île du Havre between June 2000 and June 2002. The depth across the grid (from 40 to 0 m along the y‐axis) was from 3 to 5 m at Île à Firmin East, and from 4 to 6 m at Île du Havre. Values in parenthesis represent the mean distance (± SD) of the kelp interface relative to the benchmark transect in the urchin barrens (0 m). Horizontal dashed lines indicate the mean distance of the kelp bed on the first sampling date (June 2000).

26
the kelp‐barrens interface at Île du Havre was relatively stable over the 2‐year period:
the greatest regression was just 3.5 ± 1.3 m, with a maximal rate of loss
(1.3 m month‐1) between August and October 2000. Some recolonization occurred
during the first winter so that the lower limit in June 2001 was only 0.9 ± 1.7 m above
the lower limit of the bed in the previous June. The bed extended deeper during the
second year, and, as for the Île à Firmin East site, slightly exceeded the initial lower
limit by the end of the study. Thus, although monthly changes in the position of the
lower limit of the kelp bed were much greater at Île à Firmin East than at Île du
Havre, the net displacement of the lower limit of the kelp bed over 2 years was
similar at the two sites.
Measurements of the lower limit of the kelp beds at the three less‐intensely
studied sites also showed a high degree of variability among sites and over time. At
two sites, Île Niapiskau and Petite île au Marteau, there was a substantial regression
of the kelp bed (over a distance of 3.8 ± 1.1 m and 5.9 ± 1.4 m, respectively) during the
first 12 months, as observed at Île à Firmin East. In the second year, there was little
change at Petite île au Marteau as the lower limit of the bed in June 2002 was only
1.0 ± 1.3 m below the lower limit in August 2001. However, as seen at Île à Firmin
East, substantial recolonization appeared to occur at Île Niapiskau during the winter
of 2001‐2002. We were unable to locate reference points at this site in June 2002 due to
the kelp cover, but noted that the lower limit of the kelp bed was well below that of
the previous summer. In contrast to all other sites, the third site, Île à Firmin West,
showed a major extension of the kelp bed during the first winter (7.1 ± 1.2 m) and
then retreated during the summer season (4.4 ± 1.9 m). We were also unable to find
the reference points at this site in 2002, but again observed another major extension of
the kelp bed during the second winter. There appeared to have been a major
disturbance over the winter 2001‐2002 as cobbles and boulders covered with small
kelp were scattered over the bottom. Also, there were denuded areas at shallower
depths in the kelp bed that were likely due to ice scour. None of these sites showed
the stability in the kelp‐barrens interface seen at the Île du Havre site.

27
Structure of kelp beds
The 2‐year monitoring of the density and size of kelp in the transition zone indicated
that kelp in the lower 5 m of the bed were always more abundant (Table 2.1) and
larger (Table 2.2) at Île à Firmin East than at Île du Havre. At Île à Firmin East, there
were >50 kelp plants in almost half of the quadrats at the beginning of the study.
However, densities decreased sharply during the summers of 2000 and 2001 as the
kelp bed retreated to a shallower depth (Fig. 2.3). The major increase in June 2002
relative to August 2001 was due to the newly recruited kelp that caused a major
extension of the kelp bed (Fig. 2.3). The occurrence of >100‐cm plants was relatively
high at all times (13‐32 %). Alaria esculenta always dominated, but Laminaria digitata
and Saccorhiza dermatodea were also common and patches of the green alga
Spongomorpha arcta, ulvoid algae, and the brown alga Scytosiphon lomentaria were
present on surfaces between kelp holdfasts, especially in June of each year.
In contrast, at Île du Havre, there were <10 kelp plants in most quadrats at the
beginning of the study. Densities decreased further over time and were extremely
low by June 2002. This decrease in kelp density was accompanied by a general
decrease in kelp size. For example, the occurrence of plants >50 cm in length was
initially only 29 % but decreased to 5 % by the end of the study. The decline in both
kelp density and length was due to a shift from A. esculenta to Agarum cribrosum in the
lower kelp bed (in mature stands, A. cribrosum rarely exceeds 1 m in length, and
individuals are more dispersed than in a bed of A. esculenta; P. Gagnon, personal
observation). By June 2002, the lowest portion of the bed was a mature stand
of A. cribrosum that extended up to 8 m from the lower limit of the A. esculenta zone.
Above the A. cribrosum stand, a luxuriant kelp bed of large A. esculenta still flourished.
There were also a few large Laminaria longicruris (often >2 m in length), but L. digitata
and S. dermatodea were virtually absent, and S. arcta, ulvoid algae, and S. lomentaria
were scarce. We never observed any major recruitment events at this site.

28
Table 2.1. Proportion of quadrats (out of 48) containing different density classes of kelp (0‐10, 10‐30, 30‐50, and >50 individuals per 25 x 25‐cm quadrat) on different dates from June 2000 to June 2002 for the lower 5 m of the kelp bed zone (quadrats placed at 0.0, 1.0, 3.0 and 5.0 m) at Île à Firmin East and Île du Havre.
Île à Firmin East Île du Havre
0‐10 10‐30 30‐50 >50 0‐10 10‐30 30‐50 >50
Jun 2000 27 12 15 46 61 35 0 4
Aug 2000 54 27 8 11 90 10 0 0
Jun 2001 50 17 21 12 92 4 2 2
Aug 2001 94 6 0 0 98 2 0 0
Jun 2002 21 19 27 33 100 0 0 0

29
Table 2.2. Proportion of quadrats (out of 48 25 x 25‐cm quadrats) containing different size classes of kelp (<50, 50‐100, and >100 cm in length) on different dates from June 2000 to June 2002 for the lower 5 m of the kelp bed zone (quadrats placed at 0.0, 1.0, 3.0 and 5.0 m) at Île à Firmin East and Île du Havre.
Île à Firmin East Île du Havre
<50 cm 50‐100 cm >100 cm <50 cm 50‐100 cm >100 cm
Jun 2000 73 13 14 71 21 8
Aug 2000 73 12 15 82 10 8
Jun 2001 74 4 22 97 1 2
Aug 2001 58 10 32 91 5 4
Jun 2002 51 36 13 95 5 0

30
Structure of urchin populations
The 2‐year monitoring of the density and size‐structure of urchin populations in the
transition zone showed substantial changes at each site as well as major differences
between sites. There was an overall decline in urchin abundance by approximately
50 % during the course of the study at both sites (Fig. 2.4). At Île à Firmin East, this
trend was seasonally reversed with dramatic increases during the summer. In
contrast, no seasonal variation was seen at Île du Havre, and a massive decline in
densities (47 %) between August 2000 and June 2001 accounted for most of the
change during the study.
The sampling at different distances from the kelp‐barrens interface (into the
barrens and into the kelp bed) showed that urchin densities consistently peaked in
the first meter below the edge of the bed at both sites (Fig. 2.5). There was almost
always a distinct front of urchins, often 2‐3 layers thick. Urchin abundance fell off
much more rapidly within the kelp bed than in the urchin barrens although at times,
there were substantial densities (>100 m‐2) within the first 2 m, and occasionally even
far into the kelp bed (e.g., Île du Havre in June 2001). The bell‐shaped distribution
around the kelp‐barrens interface flattened with time, particularly at Île à Firmin East,
associated with the overall decrease in urchin numbers (see above).
The analysis of the size‐structure of urchins in the transition zone showed that
mean size generally increased over time at both sites, but urchins were always larger
at Île à Firmin East (Fig. 2.6). In the kelp bed above the interface, urchins were
generally larger with the largest size class often comprising more than 50 % of the
sample and the smallest size class usually accounting for <25 %. Most urchins in the
front and in the kelp bed were medium (2‐4 cm) to large (>4 cm) in size (Fig. 2.6)
whereas small urchins (<2 cm) were far more common in the urchin barrens,
particularly at Île du Havre. At Île à Firmin East, the proportion of small urchins in
the overall area decreased during the first year (to nearly nil in the summer of 2001)
and then increased by June 2002. Urchin biomass in the grazing front (in the first

31
0
50
100
150
200
250 Île à Firmin East
Île du Havre
Jun 00 Aug 00 Jun 01 Aug 01 Jun 02
abaa
bc
cdde
dede e
f
Fig. 2.4. Changes in the mean density (+ SE) of the green sea urchin, Strongylocentrotus droebachiensis, within 5 m of the lower limit of kelp beds at Île à Firmin East and Île du Havre between June 2000 and June 2002. Values not sharing the same letter are different (LS means, P < 0.05).

32
0100200300400500600
Île à Firmin East Île du Havre
Distance to the kelp bed edge (m)
0100200300400500600
0100200300400500600
0100200300400500600
0100200300400500600
Jun 2000
Aug 2000
Jun 2001
Aug 2001
Jun 2002
Jun 2000
Aug 2000
Jun 2001
Aug 2001
Jun 2002
Kelp bed Barrens Kelp bed Barrens
Edge Edge
2 222 4 66 644 6 4
Fig. 2.5. Changes in the densities of the green sea urchin, Strongylocentrotus droebachiensis, within 5 m of the lower limit of kelp beds at Île à Firmin East and Île du Havre between June 2000 and June 2002. Data are means (+ SE) for quadrats (n = 12) placed at different distances inside and outside of the kelp‐barrens interface (0.0, 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 4.0 and 5.0 m in June and August 2000, and at 0.0, 0.5, 1.0, 3.0, and 5.0 m in June and August 2001, and June 2002).

33
Medium
Large
Jun 2000
Aug 2000
Île à Firmin East
Jun 2001
Aug 2001
Jun 2002
Île du Havre
Distance to the kelp bed edge (m)
Small
Kelp bed Barrens0-1 3 535
Jun 2000
Aug 2000
Jun 2001
Aug 2001
Jun 2002
1-0
5.5 2.0 2.5
7.7 3.6 2.7
7.6 2.7 5.1
10.2 5.5 6.3
3.2 3.3 2.4
5.6 2.8 1.7
3.7 3.6 1.4
4.5 2.3 2.6
4.6 3.1 2.2
4.8 2.9 2.5
0.6 0.5 3.6
1.1 3.5 5.9
0.7 0.1 2.7
0.7 3.2 5.2
0.2 0.4 0.5
1.8 1.0 2.6
2.5 3.4 2.8
6.2 3.5 1.8
3.2 2.4 2.3
0.8 3.3 1.2
Edge
Fig. 2.6. Changes in the proportions of small (<2 cm test diameter), medium‐sized (2‐4 cm) and large (>4 cm) urchins, Strongylocentrotus droebachiensis, within 5 m of the lower limit of kelp beds at Île à Firmin East and Île du Havre between June 2000 and June 2002. Data are means for quadrats (n = 12) placed at 0.0, 0.5, and 1.0 m (averaged and designated as 0‐1 m interval), and at 3.0 and 5.0 m inside and outside of the kelp‐barrens interface. Values above the circle plots represent urchin biomass (kg m‐2) estimated from a regression equation relating fresh weight to test diameter (n = 144) collected at Île du Havre.

34
meter below the kelp edge) was initially similar (∼5.5 kg m‐2) at the two sites (Fig. 2.6).
Although the biomass increased almost twofold over the following 14 months at
Île à Firmin East (to August 2001), it always remained less than the initial value
at Île du Havre. Thus, during most of the study, the grazing potential was higher at
Île à Firmin East than at Île du Havre.
Our data did not support the hypothesis that the rate of kelp bed regression is
positively correlated with urchin biomass in the grazing fronts. We found no
significant relationship between the biomass of urchins in the grazing fronts and the
rate of change in the position of the lower limit of kelp beds at the two intensively
studied sites, either when averaged for the entire site (Fig. 2.7) or when examined
within a site using transects as replicates (data not shown). Nevertheless, we noted a
tendency towards increased kelp regression with increased urchin biomass at
Île à Firmin East (r2 = 0.33, P = 0.42). There appeared to be a threshold biomass of
urchins of ~5 kg m‐2 (estimated from a regression equation of the pooled data from
both sites; i.e, when kelp edge displacement (m) = 0 = 1.79 ‐ 0.37 x Urchin biomass
(kg m‐2), r2 = 0.29, P = 0.17) above which the beds receded. As urchin biomass was
usually just below 5 kg m‐2 (June 2000 being the only exception) at Île du Havre, this
might explain why changes in the position of the kelp bed interface varied little
during the study (but see below). Overall, these results indicate that the variability in
the position of the kelp‐barrens interface is a more complex function than simply
being related to the urchin biomass in the grazing front (e.g., related to kelp density,
wave exposure, tendency of urchins to feed).
Effect of urchins on algal recruitment in the barrens
The 44‐d exclusion of urchins from experimental areas in the urchin barrens
demonstrated that urchins strongly limited kelp recruitment as recruitment was
>100 times greater in the Ungrazed treatment (232.0 ± 66.5 [SE] recruits m‐2) than in
the Grazed treatment (2.0 ± 2.0; LS means, P = 0.0053). Early colonists, such as
diatoms, ulvoid algae, and Spongomorpha arcta, also flourished in the absence of

35
-3
-2
-1
0
1
2
0 2 4 6 8
Urchin biomass (kg m-2)
10
Fig. 2.7. Relationship between the rate of change in the position of the lower limit of kelp beds and urchin biomass in the grazing fronts at Île à Firmin East (filled circles) and Île du Havre (open circles) between June 2000 and June 2002. Positive rates indicate kelp bed expansion and negative rates kelp bed regression.

36
urchins. The similarity between the Fence control (4.0 ± 2.3) and Grazed treatments
(LS means, P = 0.97) demonstrated that the manipulation did not affect recruitment
beyond the effect of grazing by urchins.
Limitation of upward movements of urchins by A. cribrosum
The experimental removal of mature Agarum cribrosum showed that dense stands of
kelp can greatly reduce shoreward movement of urchins. In the areas where
A. cribrosum was not removed, the density of urchins in the lower A. cribrosum zone
increased gradually over the first 3 d of the experiment but then remained low for the
duration of the experiment (<34 m‐2, Fig. 2.8). In contrast, in the areas where the
A. cribrosum canopy was removed, the density of urchins increased (particularly
between 3 and 7 d), and after 20 d was more than four times greater than the first day.
The final urchin density (after 20 d) was three times greater in the removal than in the
non‐removal areas. During the experiment, we observed dense urchin aggregations at
the outer edge of the intact A. cribrosum areas (up to 356 urchins m‐2, SE = 177.1)
during periods of calm wave conditions. Conditions were especially calm during the
last 3 d of the study, and we observed groups of urchins at the edge of the bed
anchoring the blades of A. cribrosum to the bottom. These aggregations likely
accounted for the large increase in the density of urchins in the intact A. cribrosum
areas as recorded on Day 20.
Discussion
Although large seaweeds generally dominate shallow rocky subtidal communities on
northwestern North Atlantic coasts (Chapman & Johnson, 1990) grazing by the green
sea urchin, Strongylocentrotus droebachiensis, nevertheless severely limits the
distribution of kelp (Himmelman & Steele, 1971; Breen & Mann, 1976a; b;
Chapman, 1981; Himmelman et al., 1983; Himmelman, 1984; Miller, 1985; Scheibling,
1986; Witman, 1987; Keats et al., 1990; Scheibling et al., 1999). Our urchin exclusion
experiments showing that urchins dramatically limited the recruitment of both kelp

37
a
a
a
bbbc
ccdcd
d
de
e
0
20
40
60
80
100
120
140
Intact A. cribrosum
A. cribrosum removed
Time (d)
1 2 3 7 13 20
Fig. 2.8. Changes, over a 20‐d period, in the mean density (+ SE) of the green sea urchin, Strongylocentrotus droebachiensis, in 1 x 4‐m areas along the lower edge of a mature stand of the kelp Agarum cribrosum where the kelp was removed and in 1 x 4‐m areas where A. cribrosum was left intact. Values not sharing the same letter are different (LS means, P < 0.05).

38
and ephemeral algae in the barrens is consistent with this body of knowledge. Thus,
as seen elsewhere, urchin grazing is a primary factor controlling algal distribution in
the shallow rocky subtidal zone of the Mingan Islands.
Most previous studies have focused on the negative impact of the green sea
urchin on primary productivity, but these one‐time observations have only provided
limited information about interactions controlling changes in kelp bed boundaries
(but see Scheibling et al., 1999). Here we have shown that the kelp‐barrens interface is
dynamic and can shift over weeks to months. Moreover, we have demonstrated that
these changes in the lower limit of kelp beds are site‐specific. For example, between
August 2000 and June 2001, three sites experienced a regression in the kelp‐barrens
interface of 4 to 7 m, one site experienced no change, and one site showed an advance
of 7 m. This variation was not clinal as the two most extreme sites (Île à Firmin East
and West) were the closest together. Substantial temporal variation was also seen,
especially at Île à Firmin East where there was a continuous regression of the
kelp bed from 2000 to 2001, followed by massive kelp recolonization in 2002.
We had hypothesized that changes in the interface would be related to changes
in the urchin population. Whereas the massive recruitment at Île à Firmin East
occurred when urchin densities were at their lowest (August 2001 to June 2002), the
movement of the interface did not track variations in urchin densities. For example,
in spite of a 44 % decrease in urchin density during the winter of 2000‐2001, the kelp
bed continued to regress. However, rates of regression were always highest during
the summer (up to 2.5 m month‐1), when urchin numbers were increasing. Because
urchins feeding rates typically peak in summer (Vadas, 1977; Larson et al., 1980;
Himmelman, 1984; Himmelman & Nédélec, 1990; Meidel & Scheibling, 1998), their
increased voraciousness likely enhanced the effect of their increased density.
The rates of kelp bed regression that we observed were similar to those seen along the
coast of Nova Scotia (up to 4.0 m month‐1, Breen & Mann, 1976b; Mann, 1977;
Scheibling et al., 1999). Overall, our data suggest that the effect of urchins was
non‐linear at Île à Firmin East and that kelp could only colonize when the urchin

39
density was below ~100 urchins m‐2. Possibly, the low number of urchins reduced
their ability to weigh down and graze on the kelp. Biomass offers another measure of
grazing pressure, and Breen and Mann (1976a; b; and later supported by Scheibling
et al. 1999) postulated that a threshold urchin biomass of ~2 kg m‐2 causes kelp
regression in Nova Scotia, a value less than half the threshold that we estimated for
the Mingan Islands (~5 kg m‐2). The difference might be attributable to the higher kelp
density in the Mingan Islands, or to the morphology or chemistry of the dominant
kelps in the two systems, Laminaria longicruris in Nova Scotia and Alaria esculenta in
the Mingan Islands. Another possibility is that our sites were more exposed to wave
action or tidal currents than the sites in Nova Scotia, as increased wave action likely
decreases the rate at which urchins move and attack algae (Kawamata, 1998;
Chapter 5). The greater exposure of the site at Île à Firmin East to waves and currents,
compared to the site at Île du Havre, could have contributed to the increased
variability in the lower limit of the kelp bed at this site. If urchins at Île à Firmin East
experienced less predictable flow forces, they may have been able to move rapidly
upwards during periods of low wave action. Likewise, unexpected periods of high
wave action could have dislodged urchins and permitted a massive expansion of
the kelp bed. Finally, perhaps neither density nor biomass are individually the best
measure of grazing pressure, and the best may be some combination of the two.
The large decrease in urchin density at Île à Firmin East during the winter
of 2001‐2002 was accompanied by an increased proportion of small urchins. Because
of the much lower ingestion rate of small compared to large urchins (the rate for
<15‐mm urchins is 10‐fold less than for 50‐mm urchins, Larson et al., 1980;
Himmelman, 1984; Minor & Scheibling, 1997; Meidel & Scheibling, 1999;
Scheibling & Hatcher, 2001), the overall grazing pressure may have been too low to
prevent colonization by kelp. Several observations from Île du Havre support this
idea. First, urchin densities were always >100 m‐2 yet no massive colonization was
observed (nevertheless, the greatest extension of the kelp bed did occur when urchin
densities declined by 47 % in the winter of 2000‐2001). On the other hand, in spite of
the constant high urchin densities, there was never a substantial regression of the

40
kelp bed, even though kelp density was always lower and the plants smaller in the
lower kelp bed compared to Île à Firmin East, which should have facilitated the
advance of the front (Scheibling et al., 1999). Our data support the hypothesis of a
threshold urchin biomass as the grazing front at Île du Havre remained below
5 kg m‐2 throughout most of the study and was apparently insufficient to cause
a substantial regression of the interface there.
Another major difference between the two sites was the difference in kelps that
formed the beds. Alaria esculenta was the dominant kelp at Île à Firmin East, whereas
we observed a shift from A. esculenta to Agarum cribrosum at the lower edge of the bed
at Île du Havre during the study as reflected by the gradual decline in mean density
and height of the kelp. Agarum cribrosum is probably more resistant to grazing than
A. esculenta given the low preference of the urchin for this alga in both field and
laboratory feeding studies (Himmelman & Steele, 1971; Vadas, 1977; Larson et al.,
1980; Himmelman, 1984; Himmelman & Nédélec, 1990). This low preference may be
partially due to chemical defenses, such as phenolics (Vadas, 1977; Larson et al., 1980;
Himmelman & Nédélec, 1990). The tough stipe of A. cribrosum, which is fairly rigid
and holds the blade above the bottom, may also contribute to its resistance to grazing.
The greater resistance of A. cribrosum compared to A. esculenta is further shown by its
ability to form patches within the barrens (Vadas, 1968; 1977; Keats et al., 1982;
Himmelman, 1984). Moreover, as shown by our A. cribrosum removal trials, the
presence of A. cribrosum can greatly reduce the upward movement of urchins. Thus,
in certain situations, A. cribrosum appears to form a border at the lower edge of the
kelp zone that limits incursions by urchins and in turn protects the more vulnerable
A. esculenta growing at shallower depths from urchins. This effect on urchins may be
unique to A. cribrosum because of its distinct grazing resistant characteristics but
could also be a general characteristic of any alga capable of forming a dense stand.
The mechanism by which A. cribrosum reduces movements of urchins remains
to be determined. The alga might ”block” the movement of urchins, by forming
a barrier or repulsing the urchins, as seen for some other macroalgae

41
(Velimirov & Griffiths, 1979; Himmelman, 1984; Dayton, 1985a; Vasquez & McPeak,
1998; Konar, 2000; Gagnon et al., 2003b), or urchins might simply stop once they
encounter a potential food source. A slowing of the advance of urchins by
A. cribrosum was suggested by our observations at Île du Havre. Although the urchin
front was clearly defined in June 2000 (high numbers at the lower edge of the kelp
bed), the front became less pronounced over time as A. cribrosum gradually replaced
A. esculenta. Because most of the urchins aggregated at the lower edge of the
A. cribrosum, fewer urchins penetrated to the A. esculenta in shallower water. The
greater temporal variability in the position of the lower limit of the kelp bed at
Île à Firmin East, compared to Île du Havre, may thus have been related to the
absence of A. cribrosum at this site.
We observed substantial declines in urchin abundance over time at both Île du
Havre and Île à Firmin East, but events varied between sites. At Île du Havre,
densities declined abruptly between August 2000 and June 2001, coincident with an
increase in the average size, suggesting a demographic change (such as mortality or
emigration). In contrast, at Île à Firmin East, we observed a more gradual temporal
decline along with a seasonal pattern. The seasonal pattern was likely due to the
migration of urchins into deeper water during the winter, perhaps to avoid ice scour
(see below) or increased wave action as suggested for urchins populations in the
St. Lawrence Estuary (Himmelman & Lavergne, 1985), and their return the following
spring. The gradual decline in density by >50 % between June 2000 and June 2002 is
more difficult to explain. The decrease in the first year coincided with an increase in
average size, as also seen at Île du Havre. A number of studies show that the growth
rate of S. droebachiensis increases with size (Swan, 1961; Thompson, 1984;
Raymond & Scheibling, 1987; Russell et al., 1998). Possibly, the small urchins at
Île à Firmin East benefited from the drop in large urchins that were either killed or
moved away due to ice scour. The reduced intraspecific competition and increased
food availability (with the new growth of kelp in the barrens) should have favored
increased growth and thus increases in the average size. However, the change during

42
the second year did not suggest a density‐dependent response to resources as a shift
to smaller urchins accompanied the decrease in numbers.
We documented striking differences between the two intensively studied sites,
the most striking being the dynamics of the kelp‐barrens interface and the general
decline in urchin abundance. Unfortunately, we did not expect such extreme
variation (e.g., the establishment of A. cribrosum at the Île du Havre site) and thus did
not sufficiently replicate sampling for making definitive statements regarding the
causes of these differences. Still, we postulate that A. cribrosum can play a pivotal role
in determining the dynamics of the kelp‐barrens interface by providing a
herbivore‐resistant fringe at the lower edge of the kelp bed. The distribution of
A. cribrosum in shallow waters may, in turn, be controlled by ice scour, which occurs
frequently during winter and early spring along rocky shores in the St. Lawrence
Estuary and the Gulf of St. Lawrence (Archambault & Bourget, 1983;
Bergeron & Bourget, 1984; Bourget et al., 1985), and along the coast of Newfoundland
(Hooper, 1981; Keats et al., 1985). We saw evidence of ice scour at Île à Firmin East
during the winter of 2000‐2001, as five of the six 10‐cm steel bolts set into the bedrock
as markers for the benchmark transects were found bent in June 2001. There was also
evidence of ice scour (although less severe) in the winter of 2001‐2002, as two
out of six bolts were bent (the damaged bolts found in June 2001 had been replaced).
These observations are consistent with those of Parks Canada personnel
(Benoît Roberge and Nancy Dénommée, personal communications) who recorded
a solid ice sheet between the islands for only two weeks in 2001 (20 February to
6 March), whereas there was a solid cover for two months in 2002 (18 January to
17 March) (there is less ice scouring when the cover is solid). In contrast, ice scour
appeared to have been less at Île du Havre, as only three out of six bolts were bent
during the winter of 2000‐2001 (all three in the shallower part of the kelp bed) and
only a single bolt during the 2001‐2002 winter. If the condition of the bolts did reflect
less ice scouring at Île du Havre, decreased disturbance may explain the greater
survival of A. cribrosum at this site. The drop in urchin density observed at
Île à Firmin East during winter of 2000‐2001 is harder to explain but may reflect

43
migration to greater depths due to winter conditions. Severe ice scouring appears to
have occurred at the Île à Firmin West site during the winter of 2001‐2002 as there
were numerous denuded areas in the kelp community at shallower depths. More
detailed observations are needed, however, to relate ice conditions to urchin
aggregation and dispersion below the kelp bed.
Our study is the first to examine factors affecting the temporal stability in the
lower limit of kelp beds in the northern Gulf of St. Lawrence. We provide evidence
that sea urchin grazing plays a pivotal role in determining the lower limit of kelp.
However, our results also show that the intensity of urchin‐kelp interactions is both
site and time specific. For example, the tendency of an increased rate of kelp bed
regression with increased urchin biomass was only seen at one of the intensively
studied site (Île à Firmin East). We propose that perturbations by abiotic factors
(e.g., ice scouring and water motion) triggering important but localized changes in
urchin abundance are of primary importance for controlling kelp beds in the northern
Gulf, rather than the large‐scale effects of biotic factors as observed in other regions
(e.g., predation and disease, Estes et al., 1978; Simenstad et al., 1978; Pearse & Hines,
1979; Duggins, 1980; Miller, 1985; Scheibling, 1986; Scheibling & Hennigar, 1997;
Scheibling et al., 1999). These perturbations would allow kelp beds to re‐establish
over the barrens during periods of reduced grazing. To increase our global
understanding of the dynamics of such subtidal macroalgal communities, additional
long‐term studies are needed to examine the temporal stability of algal assemblages
in the barrens zone, as well as to investigate the mechanisms by which a few fleshy
macroalgae (e.g., A. cribrosum and Desmarestia viridis) withstand intensive grazing by
urchins.

Chapitre III
Spatial and temporal stability in algal assemblages on urchin barrens in the northern Gulf of St. Lawrence

45
Résumé
L’algue brune Agarum cribrosum est la seule grande macrophyte pérenne charnue
communément retrouvée en zone dénudée dans le nord‐ouest de l’Atlantique. Nous
avons examiné la stabilité spatiale et temporelle des agrégats d’A. cribrosum ainsi que
leur impact sur le recrutement algal dans l’archipel de Mingan, dans le nord du golfe
du Saint‐Laurent. Les agrégats étaient très stables dans l’espace et dans le temps avec
que de faibles variations entre les sites étudiés. L’abondance (pourcentage de
recouvrement) d’A. cribrosum dans deux aires de 144 m2 s’est accrue de 6.5‐11.4 % en
2 ans et la plupart des changements d’abondance sont survenus en marge des
agrégats. L’aire de la surface de petits (<13 m2) agrégats d’A. cribrosum augmentait de
~1.8 % mois‐1 bien que des augmentations hivernales marquées (>95 %) sont
survenues, principalement dues à la fusion d’agrégats adjacents en de plus grands
agrégats. Les agrégats d’A. cribrosum favorisent le recrutement algal tel qu’indiqué
par l’abondance des juvéniles d’A. cribrosum et de l’algue rouge Ptilota serrata, plus de
cent fois supérieure à l’intérieur qu’à l’extérieur des agrégats. L’élimination
expérimentale de la canopée d’A. cribrosum (portions de 1 m2) en bordure d’agrégats
n’a eu aucun effet à long terme sur l’abondance d’A. cribrosum: en 14 mois, l’algue
avait à nouveau colonisé la plupart des aires initialement fauchées malgré le
recrutement de la laminaire Alaria esculenta. Les agrégats d’A. cribrosum semblent
donc résilients à de petites perturbations mais pourraient dépendre des brouteurs
pour supprimer les espèces compétitrices. À l’inverse des juvéniles d’A. cribrosum,
l’abondance de P. serrata a diminué rapidement après l’élimination de la canopée
pour n’être, à la fin de l’étude, qu’à 17 % de la valeur observée dans le traitement avec
canopée. Nos résultats démontrent que les agrégats d’A. cribrosum sont une
importante composante temporelle et spatiale en zone dénudée, en dépit du
broutage intensif typique de l’oursin. Le maintien et l’expansion des agrégats
d’A. cribrosum et de la flore associée semblent dépendre d’interactions positives
(facilitation) avec les adultes d’A. cribrosum, eux‐mêmes capables de repousser
l’oursin.

46
Abstract
The brown alga Agarum cribrosum is the only large, perennial, fleshy macrophyte
commonly found on urchin‐dominated barrens in the northwestern North Atlantic.
We examined the spatial and temporal stability of A. cribrosum stands and their
impact on algal recruitment in the Mingan Islands, northern Gulf of St. Lawrence.
The stands were highly stable in space and time, with only small variations between
the study sites. The abundance (percentage cover) of A. cribrosum in two 144‐m2 areas
increased by 6.5‐11.4 % over a 2‐year period, and most changes in abundance
occurred at the edge of the stands. The surface area of small (<13 m2) single stands of
A. cribrosum typically increased by ∼1.8 % month‐1, although marked increases (>95 %)
occurred during winter, largely because adjacent stands merged into larger, single
stands. Mature stands of A. cribrosum appear to enhance algal recruitment, as juvenile
A. cribrosum and the understory red alga Ptilota serrata were orders of magnitude
more abundant inside than outside the stands. The experimental removal of the
A. cribrosum canopy (1‐m2 portions) at the border of stands had no long‐term effect on
the abundance of A. cribrosum as within 14 months, the alga had recolonized most
of the cleared areas in spite of recruitment of the kelp Alaria esculenta. Thus,
A. cribrosum stands appear resilient to small‐scale perturbations but may require
grazers to suppress competitors. In contrast to juvenile A. cribrosum, the abundance
of P. serrata rapidly decreased after canopy removal and by the end of the study had
fallen to just 17 % of the value seen in control areas with an intact canopy. Our results
demonstrate that A. cribrosum stands are a temporally and spatially important
component of urchin barrens in spite of the heavy grazing that typically occurs there.
Maintenance and expansion of A. cribrosum stands and associated flora appear
to depend on positive interactions (facilitation) with self‐defended adults
of A. cribrosum.

47
Introduction
Herbivory and competition often reduce the recruitment and survival of marine algae
and both these types of negative interactions are thought to play key roles in
structuring marine communities (Randall, 1961; Lawrence, 1975; Lubchenco &
Gaines, 1981; Harrold & Reed, 1985; Carpenter, 1986; Lewis, 1986; Morrison, 1988).
Less attention has been paid to the potential positive effects of grazing‐resistant algae
on other algal species that are vulnerable to grazing (e.g., by interfering with the
foraging of herbivores and providing microhabitats of increased algal recruitment
and survival) (but see Hay, 1986; Pfister & Hay, 1988; Gagnon et al., 2003b) in spite
of the increasing recognition of the importance of positive interactions (facilitation) in
structuring communities (Bertness & Callaway, 1994; Callaway, 1995;
Bruno & Bertness, 2001).
This bias is evident in polar and temperate regions, where herbivory
(Pearse & Hines, 1979; Dayton et al., 1984; Keats et al., 1990; Sivertsen, 1997;
Scheibling et al., 1999; Gagnon et al., in press) and competition (Dayton et al., 1984;
Johnson & Mann, 1988; Paine, 1990; Reed, 1990; Leinaas & Christie, 1996) can strongly
affect the abundance and distribution of macrophytes, although abiotic factors such
as light availability, water motion, nutrients, sedimentation, salinity and ice scour are
clearly important as well (see review in Dayton, 1985b; but also Hooper, 1981;
Dean & Jacobsen, 1984; Gerard, 1984; Ebeling et al., 1985; Keats et al., 1985). Intensive
hebivory has been well documented in the subtidal zone in the northwestern North
Atlantic where the green sea urchin, Strongylocentrotus droebachiensis, severely limits
the distribution of kelp beds (Breen & Mann, 1976a; b; Chapman, 1981; Miller, 1985;
Scheibling, 1986; Scheibling et al., 1999; Gagnon et al., in press). Kelps are often
restricted to shallow water refuges but rapidly extend to deeper water when urchins
are naturally or experimentally removed (Himmelman et al., 1983; Scheibling, 1986;
Keats et al., 1990; Gagnon et al., in press).
As elsewhere, the intensive grazing of S. droebachiensis has led to the formation
of extensive urchin barrens in the Mingan Islands (northern Gulf of St. Lawrence,

48
Canada) that generally lack fleshy macrophytes and are instead dominated by
calcified red algae (Gagnon et al., in press). The perennial kelp, Agarum cribrosum, and
the annual filamentous alga Desmarestia viridis, are the only two brown algae
regularly found on these barrens. Agarum cribrosum has a large holdfast, a fairly rigid
stipe, and a large crinkled frond that becomes thicker and tougher with age. Urchins
have a low preference for this alga which is thought to be due to chemical deterrents
(e.g., phenolics, Vadas, 1968; 1977; Larson et al., 1980; Himmelman, 1984;
Himmelman & Nédélec, 1990). Agarum cribrosum is most common on topographical
high points and in areas with strong currents, which suggest that these situations
provide advantages. Although A. cribrosum is most often found in small stands
(1‐10 m2) on urchin barrens, it sometimes forms large stands (>500 m2) at the lower
edge of shallow beds of the kelp Alaria esculenta in exposed areas
(P. Gagnon, personal observation). Elsewhere, A. cribrosum has been shown to be
competitively inferior to other kelp species and is thought to require some level of
grazing to suppress competitors (Vadas, 1968).
Distinct invertebrate assemblages are associated with stands of A. cribrosum
(Bégin, 2002) with urchin densities much lower than in the surrounding barrens, and
A. cribrosum has been shown to restrict urchin movements (Gagnon et al., in press).
This reduction in herbivore density and activity could influence the flora associated
with stands of A. cribrosum, but there is no information on abundance or dynamics of
other algal species on urchin barrens. Moreover, we know little about the temporal or
spatial stability of this conspicuous and important component of the urchin barrens.
In this study, we examine the temporal variability in the abundance and distribution
of A. cribrosum and associated algae on urchin barrens in the Mingan Islands.
Specifically, we (1) quantify temporal changes in the abundance and distribution
of A. cribrosum over large areas, (2) examine temporal variability in the size of
A. cribrosum stands, (3) quantify the distribution of juvenile A. cribrosum,
Ptilota serrata and urchins in the vicinity of stands of A. cribrosum, and (4) assess the
impact of an A. cribrosum canopy on the recruitment and abundance of algae.

49
Materials and methods
Our study was conducted from 1998 to 2002 (with observations during the summer
and fall) in the subtidal zone on the eastern side of Île aux Goélands, and on the
western side of Petite île au Marteau, in the Mingan Islands, northern Gulf of
St. Lawrence (Fig. 2.2). At each site, the bottom is a gently‐sloped bedrock platform
which is covered by dense kelp bed, primarily Alaria esculenta, to a depth of 3‐5 m.
Below this shallow kelp bed an urchin barrens begins and extends a considerable
distance (~200 m from the shore at Île aux Goélands and 150 m at Petite île au
Marteau) to a depth of ~10 m, where a vertical wall begins. The only fleshy
macrophytes commonly found on the barrens at the two sites were the brown algae
Agarum cribrosum and Desmarestia viridis although small patches of the red
alga Ptilota serrata were also present, especially at greater depths.
Temporal variability in abundance and distribution of A. cribrosum
To evaluate the variability in the abundance and distribution of Agarum cribrosum, we
monitored changes in 12 x 12‐m (144 m2) areas in the barrens at the Île aux Goélands
and Petite île au Marteau sites. These sites were selected after exploratory surveys
based on the similarity in stand configuration, i.e., distinct stands of A. cribrosum
separated by large areas without fleshy algae. Although the abundance
of A. cribrosum was somewhat higher at these sites than on most urchin barrens in this
region, these sites were selected because the clear boundaries of the stands increased
our ability to detect changes in the abundance and distribution of A. cribrosum, and
the proximity of the sites to our laboratory facilitated sampling.
At each site, we permanently marked the corners of the sampling area with
bolts set into the bedrock. We subdivided each area into an 8 x 8 sampling grid of
64 1.5 x 1.5‐m contiguous plots using the following procedure. We first delineated
two facing sides of the area with 12‐m benchmark lines tied to bolts and marked at
1.5‐m intervals. A 1.5 x 12‐m section was then defined by attaching two 12‐m transect
lines to the benchmark lines and then subdivided the section into 8 plots using
a 1.5 x 1.5‐m quadrat positioned using the marks at 1.5‐m intervals on the transect

50
lines. Transect lines were then shifted to the adjacent 1.5 x 12‐m section and the
sampling repeated until the entire area was sampled. Within each plot, the same
diver estimated the percent cover of A. cribrosum at 5 times over a 2‐year period
(June 2000, August 2000, June 2001, August 2001, and June 2002). Benchmarks and
transect lines were removed between sampling periods.
Temporal variability in the size of A. cribrosum stands
To evaluate changes in the size of stands of Agarum cribrosum over time, we
monitored the surface area of 12 haphazardly‐selected stands of adult A. cribrosum
near the permanent 12 x 12‐m area at the Île aux Goélands site. For each stand, we
marked its position with a bolt set into the substratum at the edge of the stand, and
then recorded (to the nearest 1 cm) (1) the longest distance across the stand (A), and
(2) the longest distance running perpendicular to the first axis (B) based on the area
covered by the canopy. Given that most of the stands were ellipsoidal in shape, we
approximated the surface area (S) of each stand using the equation S = π (A/2) (B/2).
Sampling was done 5 times over a 2‐year period (July 1999, May 2000, August 2000,
June 2001 and July 2001). Initial size of stands ranged from 1.6 to 12.8 m2.
Distribution of algae and urchins near A. cribrosum
To determine whether stands of adult Agarum cribrosum are associated with fewer
urchins and increased algal recruitment, we compared the abundance of juvenile
A. cribrosum (i.e., <10 cm long), Ptilota serrata and urchins at the center of the stand
with that at different distances from the edge of stands. Six haphazardly‐chosen
A. cribrosum stands (average width of 2.4 m [SD = 0.6] and separated by at least 4 m)
were sampled on the eastern side of Île aux Goélands in July 2000. For each stand, we
counted the numbers of juvenile A. cribrosum and urchins and estimated the percent
cover of P. serrata in 25 x 25‐cm quadrats placed east and west of the point which
we considered was the center of the stand, and also at 0, 50 and 100 cm from the
eastern and western edges of the stand. The data from the quadrats in two directions
were pooled in the analysis.

51
Effect of A. cribrosum canopy on recruitment and abundance of algae
To assess the impact of Agarum cribrosum on the recruitment and abundance of algae
on urchin barrens, we monitored changes in the density of juvenile A. cribrosum and
percent cover of Ptilota serrata in the absence and presence of a canopy of adult
A. cribrosum over 14 months. The two treatments consisted of 1‐m2 areas where
A. cribrosum was experimentally removed (cut off at the holdfast) and controls
where the A. cribrosum were left intact. For this experiment, we used two large
(>50 m2) oblong stands of A. cribrosum that ran parallel to the shore at 3‐4 m in depth
at Île aux Goélands. In August 1998 we selected 16 1‐m2 plots, spaced at 1‐2 m
intervals along the lower edges of the stands, and marked each with a bolt in the
center. The plots were grouped into 8 blocks of two adjacent 1‐m2 areas, and one of
the two was randomly chosen for the canopy removal treatment. Stands were then
sampled 5 times: August 1998 (1 d after canopy removal), October 1998, June 1999,
August 1999 and October 1999. At each time, we estimated the number of juvenile
A. cribrosum and the percent cover of P. serrata in each plot using two quadrats
(25 x 25 cm) placed on the north and south sides of the center bolt. Following a heavy
recruitment of the kelps A. cribrosum and Alaria esculenta during the winter of
1998‐1999, we were not able to separate juveniles and adults in 1999 due to logistic
constraints. We averaged the data from the two quadrats in each area before
comparing species abundance among treatments.
Statistical analysis
We used a repeated measures ANOVA (Hand & Taylor, 1987; Crowder & Hand,
1990) to examine temporal changes in the percent cover of A. cribrosum in the
8 x 8 observation plots on the barrens with the factors Site (Île aux Goélands
and Petite île au Marteau) and Time (5 sampling dates), and with one nested element
(Plots within Site). To take into account the spatial dependency among the 64 plots
at each site, we modeled the covariance structure with an exponential correlation
structure (Proc Mixed, type = SP(exp)(longitude latitude), SAS Institute Inc., 1999).
This structure assumed that the correlation between plots decreased with the distance

52
separating them. Note that the information obtained for the 64 dependent plots was
the same as for 6 independent plots (Lettenmaier, 1976). The temporal dependency
was modeled with a first order autoregressive structure (Proc Mixed,
type = SP(pow)(month), SAS Institute Inc., 1999) common to both sites. Since no
transformation corrected the lack of normality in the data on some observation dates,
the ANOVA was also run with the rank transformed data. As both analyses gave
similar results, we presented the results from the analysis of the raw data
as suggested by Conover (1980).
We analyzed temporal changes in the size of stands of A. cribrosum using
a repeated measures ANOVA with the factor Time (5 sampling periods). Because four
of the pairs of stands had merged together by June 2001 (and remained merged until
the end of our study), the surface area of four stands was given a value of nul (0) in
the analysis in June and July 2001. The surface area of each of the pairs of stands that
had merged was attributed to the larger stand at the beginning of the study
(this minimized the increase in size). Since variances fluctuated over time,
we modeled the covariance structure with a specific heterogeneous compound
symmetry structure (Proc Mixed, type = CSH, SAS Institute Inc., 1999). We applied
the analysis to the raw data as the data for each sampling date were normally
distributed.
To evaluate the effect of the distance from stands of Agarum cribrosum on the
abundance of juvenile A. cribrosum, Ptilota serrata and urchins, we ran one‐way
ANOVAs (Zar, 1999) with the factor Distance (center of the stand, and 0, 50, and
100 cm from the edge of the stand). The analyses were applied to the raw data for
urchins whereas numbers of juvenile A. cribrosum were square‐root transformed to
correct for heteroscedasticity. Since no transformations corrected for the
heteroscedasticity detected in the raw data on the percent cover of P. serrata, the
ANOVA was also applied to the rank transformed data. As both analyses gave
similar results, we presented the raw data (Conover, 1980).

53
To examine temporal changes in the density of A. cribrosum juveniles and
P. serrata in the absence and presence of a canopy of A. cribrosum, we conducted
repeated measures ANOVAs with the factors Treatment (with/without canopy),
Stand (2 stands of A. cribrosum), Block (8 blocks of two treatments), and Time
(August and October 1998, and June, August, and October 1999), and with one nested
element (Block within Stand). Since variances were stable over time within the stands,
and the correlation in the data over time was similar between the treatments, we used
a pooled covariance structure in the ANOVA (Proc Mixed, type = CS, SAS Institute
Inc., 1999). We applied the analysis to the raw numbers on A. cribrosum as the data for
each time were normally distributed, whereas the data for the percent cover of
P. serrata were square‐root transformed to obtain normality for each observation date.
In the above analyses, normality was verified using Shapiro‐Wilk’s statistic
(SAS Institute Inc., 1999) and homoscedasticity by examining the graphical
distribution of the residuals and by applying Levene test (Snedecor & Cochran, 1989).
To detect differences among levels within a factor, we used least‐square means
multiple comparisons tests (LS means, SAS Institute Inc., 1999). A significance level
of 0.05 was used for all statistical tests.
Results
Temporal variability in abundance and distribution of A. cribrosum
The 2‐year monitoring of the 12 x 12‐m observation areas on the barrens indicated
only small overall increases in the abundance of Agarum cribrosum at both sites
(Table 3.1, Fig. 3.1). At Île aux Goélands, the abundance of A. cribrosum decreased
slightly during the first two months but increased steadily (~0.7 % month‐1) after
August 2000 and was 11.4 % greater at the end than at the beginning of the study
(LS means, P < 0.0001). At Petite île au Marteau, more substantial increases and
decreases occurred during the study, including a ~25 % increase between August
2000 and June 2001, but the percentage cover of A. cribrosum at the end of the study
was 6.5 % greater than at the beginning.

54
Table 3.1. Summary of the repeated measures ANOVA (mixed linear model) showing the effect of Site (Île aux Goélands and Petite île au Marteau) and Time (June and August 2000, June and August 2001, and June 2002) on the percent cover of A. cribrosum in 12 x 12‐m areas of urchin barrens (as averaged from 64 plots of 1.5 x 1.5 m at each site).
Source of variation
(fixed effects)
df
(numerator)
df
(denominator)
F‐value
P
Site 1 126 0.33 0.56
Time 4 504 42.26 <0.0001
Site x Time 4 504 12.37 <0.0001

55
0
20
40
60
80
100
Jun 2000 Aug 2000
Petite île au Marteau
Île aux Goélands
Jun 2001 Aug 2001 Jun 2002
aba
bc
d
ABBC
DC
******
** *
Fig. 3.1. Changes, over 2 years, in the mean percent cover (± SE) of kelp Agarum cribrosum on urchin barrens at Île aux Goélands and Petite île au Marteau. Values not sharing the same letter (small letters for Petite île au Marteau, capitals for Île aux Goélands) are different (LS means, P < 0.05). Asterisks are from statistical comparisons between islands in the increase or decrease in percent cover during 4 successive sampling periods (Group contrasts, *P < 0.05, **P < 0.01, ***P < 0.001).

56
The distribution of A. cribrosum appeared more stable at Île aux Goélands
(Fig. 3.2). The general gradual increase in A. cribrosum was due to colonization along
the edges of stands which were around one large area of barrens (in the upper
right corner on Fig. 3.2). Agarum cribrosum was more patchily distributed at Petite île
au Marteau and the observed fluctuations in abundance occurred across the grid.
Temporal variability in the size of A. cribrosum stands
During the first year of the 2‐year monitoring of the size of stands of
Agarum cribrosum, there was no significant change in area (LS means, P = 0.22).
In contrast, stand area increased greatly (13 % month‐1) between August 2000 and
July 2001 (Fig. 3.3; LS means, P = 0.019) due primarily to the merging of stands after
incremental increases at the margins of stands (all the stands merged with other
stands; four of the mergers occurring between stands we were monitoring whereas
the remaining four stands merged with stands that we were not monitoring).
The 17.1 % increase in size of stands in the month following this massive increase
(June to July 2001) was not statistically significant.
Distribution of algae and urchins near A. cribrosum
An inverse pattern of abundance of algae and urchins occurred inside and around
stands of adult Agarum cribrosum (Fig. 3.4). Numbers of juvenile A. cribrosum and
percentage cover of P. serrata were orders of magnitude greater inside than outside
the stands, whereas urchin densities were ≥5 times less inside than outside the stands.
No differences in the abundance of urchins and algae were seen at different distances
sampled from the edge of the stands (sampling at 0, 50 and 100 cm, Fig. 3.4),
consistent with the idea that A. cribrosum stands provided areas with decreased
urchins and favored algal recruitment or survival.
Effect of A. cribrosum canopy on recruitment and abundance of algae
The 14‐month monitoring of changes in the numbers of juvenile Agarum cribrosum

57
Petite île au MarteauÎle aux Goélands
Jun 2000
Aug 2000
Jun 2001
Aug 2001
Jun 2002
0
10
20
30
40
50
60
70
80
90
100
Percent cover (%)
Fig. 3.2. Changes, over 2 years, in the distribution and abundance of kelp Agarum cribrosum in a 12 x 12‐m zone of urchin barrens at Île aux Goélands and Petite île au Marteau. Dashed lines represent the position of transect lines used to divide each zone into a sampling grid of 64 1.5 x 1.5‐m plots.

58
0
5
10
15
20
Jul 1999 May 2000 Aug 2000 Jun 2001 Jul 2001
a
a
bbb
Fig. 3.3. Changes, over 2 years, in the mean surface area (+ SE) of stands of adult kelp Agarum cribrosum. The arrow indicates the time at which we first observed mergers between the stands. Values not sharing the same letter are different (LS means, P < 0.05).

59
Centerof stand
0 50 100Distance (cm)
0
50
100
150
200
250
0
10
20
30
40
50
0
100
200
300
400
500 S. droebachiensis
Juvenile A. cribrosum
P. serrata
b
a a
a
a
b b b
a
b b b
Edge
OutsideInside
Fig. 3.4. Mean density and percent cover (+ SE) of juvenile kelp Agarum cribrosum, the red alga Ptilota serrata, and the green sea urchin, Strongylocentrotus droebachiensis, at different distances from the center of stands of adult A. cribrosum. For juvenile A. cribrosum, the data have been back‐transformed from the transformed data used in the analysis. Values not sharing the same letter are different (LS means, P < 0.05).

60
and percentage cover of Ptilota serrata in the absence and presence of a canopy
of A. cribrosum indicated that the canopy had little or no effect on the recruitment of
A. cribrosum but had a large effect on the abundance of P. serrata (Table 3.2, Fig. 3.5).
The density of juvenile A. cribrosum did not differ between treatments except in
June 1999 when densities were almost double in the presence of a canopy.
Unfortunately, as we did not distinguish between adult and juveniles during this and
subsequent samplings, part of this difference is due to the inclusion of the adult
plants that formed the canopy itself. However, given that densities of adults are
typically near 50 m‐2 and the generally high recruitment of both A. cribrosum and
Alaria esculenta (see below) that occurred between October 1998 and June 1999, we
attribute this difference primarily to higher recruitment in treatments with an adult
canopy. Regardless, this difference did not persist and the two treatments were
statistically indistinguishable by the end of the season.
In contrast, differences in the abundance of P. serrata were apparent from the
beginning and persisted until the end. One day after the removal of the canopy, the
mean percent cover of P. serrata fell to less than 50 % of that seen in areas with an
A. cribrosum canopy although this difference was not significant (LS means, P = 0.10).
In the treatment with a canopy, the cover of P. serrata generally increased during the
study and by October 1999 was 53 % greater than at the beginning of the study
(Fig. 3.5). Although there was not a statistical difference between the first and last
date of the study, cover was significantly greater on the penultimate sampling date
(LS means, P = 0.0054). In contrast, in the areas with the canopy removed the cover
of P. serrata remained low (<5.3 %) throughout the study and was only 17 % of that
in control areas by the end of the study (LS means, P = 0.0003; Fig. 3.5).
In both areas with and without a canopy, the percent cover of other
non‐calcified algae (e.g., ulvoid algae, diatoms, and the red algae Phycodrys rubens
and Porphyra sp.) was always low (<5 %). An exception was a substantial recruitment
of the kelp Alaria esculenta between October 1998 and June 1999 that appeared to be

61
Table 3.2. Summary of repeated measures ANOVA (mixed linear model) showing the effect of Stand (two stands), Treatment (with or without a canopy of Agarum cribrosum), and Time (August and October 1998, and June, August, and October 1999) on the mean density of A. cribrosum and percent cover of Ptilota serrata in the A. cribrosum canopy‐removal experiment. Data on P. serrata were square‐root transformed.
Source of variation (fixed effects)
df (numerator)
df (denominator)
F‐value
P
Agarum cribrosum Stand 1 12 0.97 0.34 Treatment 1 12 4.88 0.047 Stand x Treatment 1 12 0.89 0.36 Time 4 47 23.12 <0.0001 Stand x Time 4 47 5.24 0.0014 Treatment x Time 4 47 1.84 0.14 Stand x Treatment x Time 4 47 0.14 0.96 Ptilota serrata Stand 1 12 2.61 0.13 Treatment 1 12 13.85 0.0029 Stand x Treatment 1 12 0.17 0.69 Time 4 47 2.08 0.10 Stand x Time 4 47 1.60 0.19 Treatment x Time 4 47 4.18 0.0056 Stand x Treatment x Time 4 47 0.10 0.98

62
0
50
100
150
200
250
300
350 A. cribrosum
With A. cribrosum
Without A. cribrosum
0
5
10
15
20
25
30
35
40 P. serrata
Aug 1998 Oct 1999Aug 1999Jun 1999Oct 1998
cc
a a
b
cc
b
a
bc
BC
BC
B
A
AB
C D CDD D
Fig. 3.5. Changes, over 14 months, in the mean density (+ SE) of kelp Agarum cribrosum and percent cover of the red alga Ptilota serrata in the absence and presence of a canopy of A. cribrosum. The arrow indicates the time from which the sampling was conducted without discrimination between juvenile and adult stages of A. cribrosum. Values not sharing the same letter (small letters for A. cribrosum, capitals for P. serrata) are different (LS means, P < 0.05).

63
greater in the presence (93.0 m‐2, SD = 69.4) than absence (65.1 m‐2, SD = 24.2) of
an A. cribrosum canopy. This difference was not significant (LS means, P = 0.39),
but A. esculenta recruits completely disappeared from areas with a canopy by
August 1999 whereas they were still common in treatments where the canopy was
removed (38.0 m‐2, SD = 18.3). At the end of the study (October 1999), A. esculenta was
absent in both treatments.
Discussion
Agarum cribrosum, the only large, perennial, fleshy macrophyte and the only kelp
species commonly found on urchin‐dominated barrens in coastal areas of the
northwestern North Atlantic (Tremblay & Chapman, 1980; Keats et al., 1982;
Himmelman & Lavergne, 1985; Novaczek & McLachlan, 1989), appears to play a
considerable role in determining the structure of the barrens community. A diverse
and distinct community of invertebrates is associated with A. cribrosum, and its
presence can increase the recruitment and growth of a number of invertebrate species
including the mussel Mytilus edulis and the polychaete Spirorbis borealis (Bégin, 2002).
In contrast, the effect of A. cribrosum on the subtidal flora has not been investigated,
and the spatial and temporal stability of A. cribrosum stands is unknown over any
spatial or temporal scale. Here, we documented a relatively high degree of temporal
stability in the abundance and distribution of A. cribrosum as well as a tight
association between A. cribrosum and other fleshy macroalgae in the Mingan Islands.
Our 2‐year monitoring of A. cribrosum in the 12 x 12‐m observation areas at
Île aux Goélands and Petite île au Marteau showed that A. cribrosum can be
a dominant feature of urchin barrens. It had a percent cover of >45 % throughout the
study and was capable of increasing in abundance in spite of the intensive grazing by
urchins (Gagnon et al., in press). The increases were due to gradual increases in size
of currently existing stands, rather than to the formation of new stands, with
expansion or recession occurring along the edge of stands and no change in the
central areas. Our observations of individual stands in an adjacent area at
Île aux Goélands are consistent with these larger scale trends as the area covered by

64
A. cribrosum increased both at stand margins and by the merging of stands. Thus,
established stands appear to be persistent (sensu Dayton et al., 1984) features of the
landscape (with a tendency towards a general increase in size through time) although
they have the potential for substantial short‐term decreases and increases in size.
Much of our knowledge about the ecology of A. cribrosum comes from the
pioneering work of Vadas (1968) in the San Juan Islands, northeastern Pacific.
He found that (1) the growth of A. cribrosum is greatest during late fall, winter, and
early spring (when water temperatures are at their lowest), (2) A. cribrosum can
become established during the fall and winter seasons, when the majority of potential
competitors (other kelp species) are unable to develop, and (3) A. cribrosum is
competitively inferior to other kelp species and can only become established when
urchins (S. droebachiensis and Strongylocentrotus franciscanus) are present. Our
experimental removal of the A. cribrosum canopy are partially consistent with these
observations. We too observed the greatest recruitment during the winter months,
but in contrast to previous work, we observed that A. cribrosum was able to recover
in spite of colonization by other kelp species (e.g., Alaria esculenta). The explanation
for this resilience is not totally clear. When A. esculenta first appeared in June 1999, the
density of A. esculenta recruits was less than half that of A. cribrosum, giving
A. cribrosum a numerical advantage if competition was important. A more likely
explanation is selective herbivory by the green sea urchin, which consumes
A. cribrosum at a much lower rate than preferred algae, such as species of Alaria and
Laminaria (Vadas, 1977; Larson et al., 1980; Himmelman, 1984; Himmelman
& Nédélec, 1990). Consistent with this idea are our observations from a concurrent
study in which A. cribrosum replaced A. esculenta (Gagnon et al., in press). This shift in
algal dominance occurred at the lower edge of an A. esculenta bed where urchins had
formed a grazing front. Over two years, the lower portion of a large
A. esculenta‐dominated zone was gradually replaced by A. cribrosum. In the present
study, the gradual decrease in the density of A. cribrosum from June to October 1999
to the levels at the beginning of the study may have been due to loss to grazers.
Alternatively, it was possibly also due to the loss of individuals from intraspecific

65
competition for resources (e.g., space, light and nutrients). Overall, our observations
indicate that A. cribrosum is resilient to punctual, small‐scale (~1 m) disturbances,
which should permit A. cribrosum to recover from disturbances by ice scour,
a common phenomenon in Newfoundland and the Gulf of St. Lawrence (Hooper,
1981; Bergeron & Bourget, 1984; Keats et al., 1985; Gagnon et al., in press).
The spatial and temporal stability of A. cribrosum stands should favor
associations with algae or algal stages which are more vulnerable to grazing or to
dislodgment by currents and waves. Our observation that A. cribrosum juveniles and
the red alga Ptilota serrata recruited within the stands of adult A. cribrosum, where
urchins densities are reduced, suggests that A. cribrosum stands provide understory
algae with protection from urchin grazing. However, our experiments only partially
support this idea. Recruitment and recovery of A. cribrosum did not require the
presence of an adult canopy although it appears that recruit densities of both
A. cribrosum and A. esculenta were higher in the presence of the canopy during
the initial phases of colonization (i.e., during the winter period). Alternatively, the
association of recruitment of A. cribrosum with adult stands could be more linked to
a higher density of gametophytes under the canopy. The removal of the canopy
would then not have had an immediate effect on recruitment as there may have been
a bank of gametophytes already present when the experiment was initiated.
In contrast, there was a clear positive effect of A. cribrosum on P. serrata
as removal of the canopy lead to a rapid and persistent decrease in its abundance.
More detailed investigations are needed to determine the exact mechanism by which
A. cribrosum might enhance the recruitment of P. serrata. Reduced grazing is certainly
a strong possibility, and previous studies have shown that mechanical abrasion from
wave‐induced movement of algal fronds repulses urchins (Konar, 2000;
Gagnon et al., 2003b). Whether the stiffer and more upright fronds of A. cribrosum
provide the same type of protection remains to be seen. Alternatively, it is possible
that P. serrata is more abundant under A. cribrosum because the canopy prevents
it from being broken and dislodged by waves and currents. Ptilota serrata is often

66
loosely attached (P. Gagnon, personal observation), and algal canopies have been
shown to reduce flow (Eckman et al., 1989; Ackerman & Okubo, 1993). This
mechanism could explain the decrease in the abundance of P. serrata immediately
after the removal of the A. cribrosum canopy. The positive effects of stands of
A. cribrosum on the recruitment of algae on urchin barrens did not apply
to Desmarestia viridis, the other common brown alga of this habitat. This annual
macroalga was not associated with stands of A. cribrosum and appears to colonize the
barrens by recruiting in the winter when urchin grazing activity is low and then
relying on mechanical repulsion of urchins once it reaches a larger size (Chapter 5).
A critical remaining question is how stands of A. cribrosum are initially formed.
We have previously shown that juvenile A. cribrosum are avidly consumed by
urchins, but protected to a degree when associated with D. viridis (Gagnon et al.,
2003b). However, as D. viridis is an annual, this protection is ephemeral. It remains
unknown how juvenile stages can survive for longer periods outside stands of adult
plants. Perhaps an exceptional year of massive recruitment is required, but only
longer term and larger scale studies are likely to be able to answer this question.
Our study is the first to examine the temporal changes in algal assemblages on
urchin barrens in the northwestern North Atlantic. We have shown that the spatial
and temporal stability of stands of A. cribrosum can be high, contrary to the low
stability of kelps which are readily grazed by the green sea urchin (Breen & Mann,
1976b; Scheibling et al., 1999; Gagnon et al., in press). This stability may be due, in
part, to the enhanced recruitment of juveniles in adult stands of A. cribrosum. These
stands also increase algal diversity by increasing the abundance of P. serrata. As
demonstrated for D. viridis (Gagnon et al., 2003b), this situation represents a case of
facilitative interaction (Connell & Slatyer, 1977; Callaway, 1995; Bruno & Bertness,
2001) that permits algal recruitment and survival on urchin barrens. More studies are
needed to determine how A. cribrosum and D. viridis withstand urchin grazing during
juvenile stages and to estimate the relative contribution of adult A. cribrosum and
D. viridis in maintaining algal assemblages in this intensively‐grazed environment.

Chapitre IV
Algal colonization in urchin barrens: defense by association during recruitment of the brown alga Agarum cribrosum

68
Résumé
Nous avons examiné le processus par lequel les juvéniles de la laminaire
Agarum cribrosum échappent au broutage de l’oursin vert, Strongylocentrotus
droebachiensis, dans la zone rocheuse infralittorale dénudée de l’archipel de Mingan,
dans le nord du golfe du Saint‐Laurent. Le recrutement le plus élevé de juvéniles
d’A. cribrosum avait lieu sous la canopée de la grande phaéophycée filamenteuse
Desmarestia viridis, où les densités d’oursins étaient considérablement réduites
comparativement à l’aire environnante. Ces patrons de distribution semblaient liés au
mouvement de balayage de D. viridis, bien que les courants peuvent modifier le
mouvement de va‐et‐vient de l’algue en poussant la canopée dans une direction
spécifique, permettant alors aux oursins d’envahir les aires non balayées. La densité
de juvéniles d’A. cribrosum sous les plants de D. viridis augmentait avec la taille du
plant et avec l’accroissement de la proximité au crampon. Les juvéniles d’A. cribrosum
vivant sous une canopée de D. viridis affichaient un taux de croissance légèrement
inférieur aux juvéniles dépourvus d’une canopée mais cette perte de croissance était
nettement compensée par le gain de protection contre le broutage de l’oursin.
La période pendant laquelle D. viridis fournit une protection aux juvéniles est de
l’ordre de quelques mois car D. viridis est une espèce annuelle qui disparaît en début
d’automne. Cette défense des juvéniles d’A. cribrosum par association avec D. viridis
est possiblement une étape dans la succession menant à la formation de plaques
matures d’A. cribrosum.

69
Abstract
We investigated the process whereby juveniles of the kelp Agarum cribrosum escape
grazing by the green sea urchin, Strongylocentrotus droebachiensis, on urchin barrens in
the rocky subtidal zone in the Mingan Islands, northern Gulf of St. Lawrence. The
highest recruitment of juvenile A. cribrosum occurred under the canopy of the large
filamentous phaeophyte Desmarestia viridis, where urchin densities were markedly
reduced, compared to the surrounding area. These patterns of distribution appeared
to be related to the wave‐induced sweeping motion of D. viridis, although currents
may modify the back and forth motion of the alga by pushing the canopy towards a
specific direction, thereby allowing urchins to invade the non‐swept areas.
The density of juveniles of A. cribrosum under D. viridis plants increased with plant
size and increasing proximity to the holdfast. Living under D. viridis slightly reduced
the growth rate of the A. cribrosum juveniles, but this loss in growth was clearly
outweighed by the gain in protection from sea urchin grazing. The time scale over
which D. viridis provides protection to the juveniles is in the order of months, as
D. viridis is an annual alga that disappears in early autumn. This defensive
association of juvenile A. cribrosum with D. viridis is possibly a successional step
leading to the formation of mature stands of A. cribrosum.

70
Introduction
Plants exhibit two general resistance strategies against herbivory: tolerance and
avoidance (Rosenthal & Kotanen, 1994). Whereas mature plants can use both
strategies, immature plants cannot usually tolerate a substantial loss of tissue to
herbivores and therefore must avoid being eaten, either by producing defenses or by
escaping in space or time (Rosenthal & Kotanen, 1994). A plant defense is any
attribute that negatively influences the feeding behavior or abundance of potential
herbivores (OʹDowd & Williamson, 1979). The production of chemical deterrents and
of grazing resistant structures are defenses used to reduce the foraging success
of herbivores (Paul & Hay, 1986; Hay & Fenical, 1988; Van Alstyne, 1988; Lucas et al.,
2000; Schnitzler et al., 2001). Spatial and temporal escapes are also a well known
phenomenon. One common example is defense by association, which occurs when a
palatable species exploits micro‐sites with lowered herbivory created by the presence
of an unpalatable species (Tahvanainen & Root, 1972; Atsatt & OʹDowd, 1976;
OʹDowd & Williamson, 1979; Hay, 1986; Brown & Ewel, 1987; Pfister & Hay, 1988).
Although this strategy might lead to competitive costs (e.g., lowered growth rates) for
the protected species, the prevention of local extinction is an obvious advantage
(Hay, 1986).
Sea urchins are often the dominant herbivore in marine rocky habitats and
may strongly influence the structure of algal assemblages by reducing the growth and
survival of algae (see review by Lawrence, 1975; but also Paine & Vadas, 1969;
Dean et al., 1984; Bulleri et al., 1999; Rose et al., 1999). In eastern Canada, intensive
grazing by the green sea urchin, Strongylocentrotus droebachiensis, often leads to the
formation of subtidal barrens with sparse algal cover (Breen & Mann, 1976b;
Scheibling et al., 1999). Such barrens are extensive in shallow areas of the Mingan
Islands, northern Gulf of St. Lawrence, where urchin densities often attain
300 individuals m‐2 (Himmelman & Nédélec, 1990), and where only a few algal
species can withstand the intensive grazing (e.g., the red alga Ptilota serrata and
crustose corallines Lithothamnion sp. and Clathromorphum sp.).

71
Of the larger brown algae, only two species, Agarum cribrosum and
Desmarestia viridis, commonly occur on urchin barrens. Agarum cribrosum is a
perennial alga which has a large holdfast, a semi‐rigid stipe, and a large crinkled
frond which becomes thicker and tougher with age (Vadas, 1968). Its low palatability
to urchins is thought to be due to chemical deterrents in the tissues (e.g., phenolics,
Vadas, 1977; Larson et al., 1980; Himmelman & Nédélec, 1990). Vadas (1968) also
indicates that A. cribrosum is most often found in areas with strong currents. In the
Mingan Islands, we have noted that A. cribrosum is usually found in small patches
(1‐10 m2) within urchin barrens and is more common on topographical high points. In
contrast, D. viridis is an annual alga with a small holdfast, a highly flexible stipe and a
large, profusely‐branched frond which sweeps back and forth over the bottom with
even slight wave action. The sweeping motion appears to repel urchins as they
rapidly aggregate on the alga during calm conditions (Himmelman & Steele, 1971;
Himmelman, 1984; Konar, 2000; Chapter 5). Many species of Desmarestia, including
D. viridis, produce sulfuric acid (Eppley & Bovell, 1958; McClintock et al., 1982;
Thompson, 1988), and numerous studies infer that this might also deter grazers
(Anderson & Velimirov, 1982; Dayton, 1985a; Himmelman & Nédélec, 1990; Lobban
& Harrison, 1994). However, the only experimental evidence supporting this
hypothesis is a recent study on Desmarestia munda (Pelletreau & Muller‐Parker, 2002).
The exact mechanism by which D. viridis repels urchins remains unclear.
Early stages of algae are often particularly vulnerable to herbivores, because of
their small size and more delicate structure (Van Alstyne et al., 1999). We have a poor
understanding of the factors permitting the survival of juvenile A. cribrosum and
D. viridis. The rapid growth of juvenile D. viridis in late winter and early spring, when
feeding rates of the urchins are low (Larson et al., 1980; Himmelman, 1984), may
permit it to reach the size where the sweeping action (or acid content) provides
resistance to grazing (thus an escape in size, sensu Paine, 1976; Lubchenco & Gaines,
1981). On the other hand, juvenile A. cribrosum (defined here as individuals
measuring <10 cm in length) appear to be unable to survive on their own as they
rarely occur in open areas on urchin barrens. However, we have often observed

72
juvenile A. cribrosum under stands of adult A. cribrosum and also under canopies of
D. viridis, which suggests that recruitment under these algae may represent a type of
defense by association. Such a relationship could provide a mechanism whereby
patches of A. cribrosum can become established.
In the present study we investigate this potential associational plant defense to
determine how A. cribrosum might colonize urchin barrens. Specifically, we
(1) quantify the distribution of the urchins and juvenile A. cribrosum in the vicinity of
D. viridis, (2) determine whether the size of the D. viridis plant influences these
distributions, (3) measure the degree of vulnerability of juvenile A. cribrosum to
urchin grazing, (4) examine whether there is a competitive cost for juvenile
A. cribrosum in being associated with D. viridis, and (5) assess the time scale involved
in these interactions.
Materials and methods
Study sites
Our study was conducted during the summer and fall of 1997, 1998 and 1999 in the
subtidal zone of Île aux Goélands, a small (<1 km2) exposed island in the Mingan
Islands, northern Gulf of St. Lawrence (Fig. 2.2). On the bedrock platforms on both
eastern and western sides of the island, dense kelp beds (primarily Alaria esculenta)
occur to a depth of 3 m, and then urchin barrens dominate gently sloping platforms
that extend to ∼200 m from shore. These platforms end at ∼10 m in depth, where
vertical walls begin. On the western platforms, Desmarestia viridis is abundant and
Agarum cribrosum scarce, and on the eastern platforms, D. viridis is less abundant and
A. cribrosum more abundant. Urchin predators, such as lobsters (Homarus americanus)
and wolffish (Anarhichas lupus) have never been observed on the platforms, and the
other conspicuous herbivores are small limpets and chitons which are micro‐grazers
and generally less abundant than urchins (Gaymer et al., 2001; P. Gagnon, personal
observation).

73
Distribution of juvenile A. cribrosum and urchins near D. viridis
We first examined the impact of the sweeping action of Desmarestia viridis in July and
August of 1997 by comparing the abundance of urchins and juvenile
Agarum cribrosum (<10 cm in length) inside and outside the area swept by individual
D. viridis plants. This was done in the vicinity of nine haphazardly‐chosen,
averaged‐sized (50‐60 cm in length) D. viridis plants. Each plant was at least 2 m from
other plants (holdfast to holdfast). For each plant we counted the numbers of urchins
and A. cribrosum juveniles in two quadrats (25 x 25 cm) placed next to the holdfast
(on the north and south sides, respectively) and in two quadrats placed 25 cm outside
of the area swept by the plant (again on the north and south sides). Six of the plants
were on the eastern side of Île aux Goélands (three on each of two sampling dates)
and the other three on the western side (one sampling date).
Current and wave conditions cause the sweeping action of D. viridis and may
in turn affect the distribution of urchins and A. cribrosum juveniles. We therefore
measured current conditions on the two sides of Île aux Goélands using Aanderaa
RCM4 current meters. The current meters were anchored at 1 m above the bottom
(4.4 m in depth on the eastern side and 5.3 m on the western side) with rigid supports
and recorded at 15‐min intervals from 26 June to 19 August of 1999. A plastic conical
fence below the current meters prevented crabs and urchins from climbing onto the
apparatus. Wave action was not quantified but is generally higher on the western
side of the island due to the predominant winds from the west (meteorological
airport services from Havre‐Saint‐Pierre Airport), except during occasional storms
from the east.
Because D. viridis can measure up to 145 cm in length at Île aux Goélands, we
also examined the effect of the size of D. viridis on the abundance of juvenile
A. cribrosum and urchins. We applied the same sampling procedure described above
around ten haphazardly‐selected medium‐sized (50‐70 cm) and large‐sized
(110‐130 cm) D. viridis, and observations were carried out on the western side of the

74
island on six dates in July 1998 (for each plant, data from the north and south sides of
the holdfast were pooled in the analysis).
To determine if juvenile A. cribrosum and urchins were uniformly distributed
within the area swept by D. viridis, we estimated their abundance at different
distances from D. viridis holdfasts (sampled in July and August of 1997 on the eastern
side of Île aux Goélands). We recorded the distance from the holdfast for every
A. cribrosum juvenile located within a 40‐cm radius of the holdfast. Densities were
subsequently calculated for concentric 10‐cm wide zones (numbers were adjusted for
the increasing area of the concentric zones moving away from the holdfast). Because
urchins were numerous, we measured their densities in 10‐cm sections along 15‐cm
wide belt transects running north and south from the holdfast to a distance of 60 cm
(the data from the two directions were pooled in the analysis). For these
measurements, we haphazardly selected five 50‐70‐cm‐long D. viridis plants. As our
observations showed that radius of the average area swept during wave action was
31 cm (SE = 2.8), we considered areas beyond 30 cm as outside the swept area.
Protection provided by D. viridis
We investigated the effectiveness of the Desmarestia viridis canopy in providing
protection for juvenile Agarum cribrosum from urchin grazing using two approaches.
First, in July and August 1997, we conducted an experiment on the eastern side of the
island to compare the rate at which the urchins made contact with juveniles
A. cribrosum in the absence and presence of a D. viridis canopy. For each of four trials,
we identified ten juveniles under each of four haphazardly selected 55‐65‐cm
D. viridis plants (a total of 160 juvenile A. cribrosum under 16 D. viridis) by placing
a numbered stainless steel washer next to each juvenile. We then removed two of the
four D. viridis by cutting their stipes and returned after 24, 48 and 72 h to score
whether each juvenile was in contact with an urchin.
Second, in July and August of 1998, we performed a second experiment on the
western side that was similar to the trials in 1997, except that we quantified the

75
changes in the total length of A. cribrosum juveniles in the absence or presence of
a D. viridis canopy. For each of three trials, we identified and measured the length of
ten juveniles under each of six haphazardly selected 50‐60‐cm D. viridis plants. Then,
after randomly removing three plants, we measured the length of the juveniles after
24, 48, 72 and 96 h. Subsequently, we expressed length as a percentage of initial
length for each juvenile. Mean initial length was almost the same in the treatments
with (4.9 cm, SE = 0.3) and without (4.8 cm, SE = 0.2) cover (⏐t⏐ = 0.07, df = 141,
P = 0.94).
Growth of juvenile A. cribrosum
We evaluated the effect of a Desmarestia viridis canopy on the growth of
Agarum cribrosum juveniles by comparing the changes in length of juveniles in the
presence and absence of a canopy, in areas where urchins were excluded using
fences. We first identified five A. cribrosum juveniles under the cover of each of eight
haphazardly‐selected D. viridis plants (55‐65 cm in length) on the western side of the
island. Then, after removing four of the D. viridis plants (chosen at random), we
quantified the changes in total length of the juveniles at 5‐d intervals over a 39‐d
period (3 July to 11 August 1998). Prior to 3 July, we constructed fences (2 x 2 m with
60‐cm‐high walls) around each of the eight experimental areas to exclude urchins.
The fences were constructed of 0.5‐cm mesh wire supported by PVC pipes and were
bolted to holes drilled in the bedrock. We first removed all the urchins from all fences
at the start of the experiment and then made daily visits to remove urchins that had
climbed onto the cage walls. For the analysis we calculated the length of each plant as
a percentage of initial length. Individual A. cribrosum were approximately 4‐cm‐tall at
the beginning of the experiment and tended to be slightly taller in the treatment with
(4.4 cm, SE = 0.6) than without (3.5 cm, SE = 0.5) cover (⏐t⏐ = 1.22, df = 28, P = 0.23).
The data on five A. cribrosum juveniles from each treatment were lost during the
study and were thus excluded from the analyses. Although two D. viridis plants in
the cover treatment broke away in the latter part of the experiment, they were

76
replaced (within 24 h) by attaching other D. viridis plants (with cable ties) to bolts
next to the holdfasts of the original plants.
Seasonal cycle of D. viridis
Because Desmarestia viridis is an annual alga and can only provide a certain period of
protection to Agarum cribrosum, we documented the decline in size of identified
D. viridis plants on the western side of the island in both 1998 (15 haphazardly chosen
individuals observed on 16 July, 23 August, and 25 October) and 1999 (28 plants
observed on 9 June, 2 and 18 August, and 26 October).
Statistical analysis
Split‐plot ANOVAs (Montgomery, 1991) were used to analyze the distribution of
juvenile Agarum cribrosum and urchins near Desmarestia viridis with the factors,
Side (western/eastern), Location (inside/outside), and Direction (north/south) with
one nested element (plant replicate within side). The analyses were applied to the raw
numbers of juvenile A. cribrosum whereas the data on numbers of urchins were
square‐root transformed to correct for heteroscedasticity.
To assess the impact of the size of D. viridis cover on the distribution of
juvenile A. cribrosum and urchins, we performed split‐plot ANOVAs with the factors,
Size (medium/large) and Location (inside/outside) with one nested element (plant
replicate within size). The data on numbers of juveniles were square‐root transformed
to obtain normality and homoscedasticity. Since no transformations corrected for the
heteroscedasticity detected in the raw data on numbers of urchins, the ANOVA was
also applied to the rank transformed data. Because both analyses gave similar results,
we present results from the former (raw data) following the suggestion of Conover
(1980).
We evaluated the effect of the distance from the D. viridis holdfast on the
density of juvenile A. cribrosum and urchins using one‐way ANOVAs with the factor
Distance (0‐10, 10‐20, 20‐30 and 30‐40 cm from the holdfast for juvenile A. cribrosum;

77
0‐10, 10‐20, 20‐30, 30‐40, 40‐50 and 50‐60 cm from the holdfast for urchins) applied to
the raw data for numbers of juvenile A. cribrosum, and to the square‐root transformed
data for numbers of urchins (to correct for heteroscedasticity).
To analyze the frequency of contact of urchins with juveniles, we applied
a repeated measures ANOVA (Hand & Taylor, 1987; Crowder & Hand, 1990) with
the factors, Treatment (with/without canopy) and Time (after 24, 48 and 72 h).
We treated this analysis as the particular case of the generalized linear models
(McCullagh & Nelder, 1989) that assumes a binomial distribution of the response
variable (dichotomous response, with or without contact). Thus, we did not test for
normality and homoscedasticity in the data, and no binomial variation was detected.
A second repeated‐measures ANOVA was carried out to test whether a D. viridis
cover protected juvenile A. cribrosum from urchin grazing with the factors, Treatment
(with/without canopy) and Time (after 24, 48, 72, and 96 h). Since we detected an
increase in the variance over time for each treatment and the correlation in the data
over time was different from one treatment to another, we modeled the covariance
structure with a specific heterogeneous compound symmetry structure (Proc Mixed,
Type = CSH, SAS Institute Inc., 1999). We applied the analysis to the raw data as the
data for each day were normally distributed.
A randomization (permutation) test (Legendre & Legendre, 1998) was used to
analyze the effect of a D. viridis cover on the growth of juvenile A. cribrosum during
the overall sampling period. Thus, we compared the result of a standard Student’s
t‐test obtained from our field sampling with the 95th percentile of the distribution of
Student’s t‐test generated by any possible randomized combination of our sampling
results.
In the above analyses, normality was verified using Shapiro‐Wilk’s statistic
(SAS Institute Inc., 1999) and homoscedasticity by examining the graphical
distribution of the residuals and by using Levene test (Snedecor & Cochran, 1989).
Differences between levels within a factor were identified using least‐square means
multiple comparisons tests (LS means, SAS Institute Inc., 1999), with sequential

78
Bonferonni correction of probabilities (Rice, 1989; Peres‐Neto, 1999). We used
a significance threshold of 0.05 for all statistical tests.
Results
Distribution of juvenile A. cribrosum and urchins near D. viridis
The 1997 sampling inside and outside of the cover of Desmarestia viridis plants
showed inverse patterns for the densities of juvenile Agarum cribrosum and urchins
(Fig. 4.1). Juvenile A. cribrosum were six times more abundant inside than outside the
cover (P < 0.0001, Table 4.1), whereas urchins were more than two times more
abundant outside than inside the cover (P < 0.0001, Table 4.1). In this analysis, we
pooled the data from different dates on the eastern side, as a preliminary analysis
revealed no difference between the two sampling dates (F1,18 = 0.03, P = 0.87 for
juveniles; F1,18 = 0.64, P = 0.43 for urchins), and then compared them to the data from
the single sampling date on the western side.
The densities (individuals m‐2) of both A. cribrosum juveniles and urchins were
significantly greater on the western than on the eastern side of the island
(66.7 ± 73.4 [SD] vs. 41.3 ± 45.0 for juveniles, P = 0.041; 168.0 ± 70.1 vs. 81.3 ± 83.7 for
urchins, P = 0.0032). Although densities of both A. cribrosum juveniles and urchins
were similar at both sides of the island outside the cover (LS means, P = 0.80 and
P = 0.19, respectively), they were greater at the western side inside the cover
(LS means, P = 0.031 and P < 0.001, respectively). Moreover, the distribution of
urchins in the vicinity of D. viridis varied between sides, there being significant
interactions among factors (Location x Direction, Side x Location, and
Side x Direction; Table 4.1). At the western side of the island, more urchins were
found on the south than on the north side of D. viridis plants (LS means, P = 0.045),
and juveniles tended to be more abundant on the north than on the south side of the
D. viridis plants, although not significantly (80.0 ± 96.0 [SD] vs. 53.3 ± 47.1; LS means,

79
0
20
40
60
80
100
120
140
160
180
A. cribrosum S. droebachiensis
P < 0.0001
P < 0.0001Inside D. viridis
Outside D. viridis
Fig. 4.1. Mean density (+ SE) of juvenile kelp Agarum cribrosum and of the green sea urchin, Strongylocentrotus droebachiensis, inside and outside the zone swept by the filamentous phaeophyte Desmarestia viridis. P‐values are for comparisons between the two zones. For the urchin, the data have been back‐transformed from the transformed data used in the analysis.

80
Table 4.1. Summary of split‐plot ANOVAs showing the effect of the Side of the island (eastern or western sides), Location (inside or outside a canopy of Desmarestia viridis) and Direction (north or south of D. viridis) on the mean density of juvenile kelp Agarum cribrosum and of the green sea urchin, Strongylocentrotus droebachiensis. Data on urchins were square‐root transformed.
Source of variation df MS F‐value P Juvenile Agarum cribrosum Side 1 5134.22 6.24 0.041 Plant(Side) 7 822.86 0.46 0.85 Location 1 52921.00 29.37 <0.0001 Direction 1 2048.00 1.14 0.30 Location x Direction 1 6883.56 3.82 0.064 Side x Location 1 7523.56 4.17 0.054 Side x Direction 1 910.22 0.51 0.49 Side x Location x Direction 1 512.00 0.28 0.60 Error 21 1802.16 Corrected total 35 Urchins Side 1 237.19 19.37 0.0032 Plant(Side) 7 12.25 1.89 0.12 Location 1 269.43 41.63 <0.0001 Direction 1 19.11 2.95 0.10 Location x Direction 1 60.02 9.27 0.0061 Side x Location 1 83.67 12.93 0.0017 Side x Direction 1 34.16 5.28 0.032 Side x Location x Direction 1 2.74 0.42 0.52 Error 21 6.47 Corrected total 35

81
P = 0.76). Such differences were not detected on the eastern side of the island where
densities of juvenile algae and urchins were similar on north and south sides
of D. viridis (LS means, P = 0.62 and P = 0.76, respectively).
Whereas tidal currents close to the bottom generally showed a similar pattern
on both sides of the island, current speeds (summer 1999) were higher on the western
(6.79 cm s‐1) than on the eastern (4.70 cm s‐1) side (⏐t⏐ = 21.69, df = 10306, P < 0.0001),
and the strongest currents on the western side were oriented towards the
north‐northeast with a peak at 15° (Fig. 4.2).
In addition to confirming the inverse distributions of A. cribrosum juveniles and
urchins inside and outside the cover of D. viridis, the sampling around medium‐sized
and large plants revealed that there were higher densities of juveniles under large
D. viridis (LS means, P = 0.015; Fig. 4.3). The density of juveniles outside the cover did
not vary with plant size (LS means, P = 0.92; Fig. 4.3). No effect of D. viridis size was
detected for urchin densities (split‐plot ANOVA, F1,18 = 1.25, P = 0.28; Fig. 4.3).
The more detailed sampling in the vicinity of the D. viridis plants revealed an
effect of distance from the holdfast on the densities of both A. cribrosum juveniles
(ANOVA, F3,12 = 7.58, P = 0.0042) and urchins (ANOVA, F5,20 = 7.03, P = 0.0006).
The data indicated that A. cribrosum juveniles were most abundant at a distance of
10‐20 cm from the holdfast (Fig. 4.4). The juveniles were also less abundant just
outside the swept zone (30‐40 cm) than within the swept zone as a whole (Group
contrast test, F1,12 = 14.43, P < 0.01). Again, urchin densities were much greater outside
than inside the swept area (Group contrast test, F1,20 = 31.77, P < 0.0001) and within the
swept zone tended to be least abundant closest to the holdfast (0‐20 cm).
Protection provided by D. viridis
In the 1997 experiment, the observations over three consecutive days showed that the
number of Agarum cribrosum juveniles in contact with urchins increased more rapidly

82
Current direction (% frequency)
Western side Eastern side
Current speed (cm s-1)
T (°C) = 5.98 (SE=1.88)Salinity (PPT) = 31.25 (SE=0.29)
T (°C) = 6.33 (SE=1.93)Salinity (PPT) = 31.06 (SE=0.34)
510152025 90
180
270
360
90
180
270
360
510152025
90
180
270
360
2810 46 90
180
270
360
2810 46
Fig. 4.2. Frequencies distribution (%) for current direction and speed from a current meter located at 1 m above the bottom on the western and eastern sides of Île aux Goélands during the summer of 1999.

83
A. cribrosum
0
10
20
30
40
50
60
70
50-70 cm D. viridis
110-130 cm D. viridis
a
a
�b
b
Inside Outside
c c
0
50
100
150
200
250
300
b
a
b
S. droebachiensis
Fig. 4.3. Mean density (+ SE) of juvenile Agarum cribrosum and of the green sea urchin, Strongylocentrotus droebachiensis, inside and outside the zone swept by medium‐sized (50‐70 cm) and large (110‐130 cm) Desmarestia viridis. For juveniles, the data have been back‐transformed from the transformed data used in the analysis. Values not sharing the same letter are significantly different (LS means, P < 0.05).

84
0-10 10-20 20-30 30-40 40-50 50-600
20
40
60
80
100
120
140
160
180
Swept zone Non-swept zone
Distance from the D. viridis holdfast (cm)
A
A
B
AB
B
A
ab
a
b
ab
S. droebachiensis
A. cribrosum
Fig. 4.4. Mean density (+ SE) of juvenile kelp Agarum cribrosum and of the green sea urchin, Strongylocentrotus droebachiensis, at different distances from the holdfast of the filamentous phaeophyte Desmarestia viridis (A. cribrosum not sampled beyond 30‐40 cm). The limit of the area swept by D. viridis was 31 cm. For the urchins, the data have been back‐transformed from the transformed data used in the analysis. Values not sharing the same letter (small letters for juveniles, capitals for urchins) are significantly different (LS means, P < 0.05).

85
in the absence than in the presence of a Desmarestia viridis canopy (Fig. 4.5). At the
end of the experiment, over 90 % of the juveniles in the canopy‐removal treatment
had had contact with urchins compared to <15 % in the control treatment.
The 1998 experiment, which quantified changes in length of juvenile
A. cribrosum over four consecutive days in the absence and presence of a D. viridis
canopy, showed a 53 % loss in length when the canopy was removed compared to
a non significant loss (6 %) with the canopy present (Fig. 4.6). The decrease in frond
length in the absence of the cover appeared linear with time, and the difference
between treatments became significant after 3 d (LS means, P < 0.01; Fig. 4.6). At the
end of the study, 34 % of the juveniles with the removed canopy had been completely
grazed, compared to only 3 % when the canopy was left intact. The predicted time to
disappearance of A. cribrosum juveniles, based on the rates of loss recorded over 4 d
(and assuming that urchins continue to graze at the same rate), would be an order
of magnitude less in the presence (7.6 d) than in the absence (71.4 d) of D. viridis.
Growth of juvenile A. cribrosum
Although the mean growth (% increase in size) of Agarum cribrosum juveniles over the
39‐d period was greater in the absence (106.4 %, [SE = 12.6]) than in the presence
(77.4 %, [SE = 7.5]) of a Desmarestia viridis cover, the difference was not significant
(P = 0.14; n = 4).
Seasonal cycle of D. viridis
The measurements of Desmarestia viridis plants in 1998 and 1999 showed a rapid
decrease in size during the summer. Mean length decreased by 78 % between 16 July
and 23 August in 1998 (equivalent to a 2 % decrease per day) and by 91 % between
9 June and 18 August in 1999 (1.3 % per day). In both years, no D. viridis sporophytes
were observed during dives in late October. Thus, Agarum cribrosum juveniles that
developed under D. viridis became fully exposed to urchins during September and
October.

86
24 48 720
20
40
60
80
100
Time (h)
Without D. viridis
With D. viridis
a
b
c
d
d d
Fig. 4.5. Mean percentage (+ SE) of juvenile kelp Agarum cribrosum in contact with the green sea urchin, Strongylocentrotus droebachiensis, over 72 h in the presence or absence of a cover of the filamentous phaeophyte Desmarestia viridis. Values not sharing the same letter are significantly different (LS means, P < 0.05).

87
a
24 48 72 960
20
40
60
80
100With D. viridis
Without D. viridis
Time (h)
aaa
a
a
b
b
Fig. 4.6. Length as a percentage of initial length (± SE) of juvenile kelp Agarum cribrosum over 96 h, when growing under a cover of the filamentous phaeophyte Desmarestia viridis and when the cover has been removed. Values not sharing the same letter are significantly different (LS means, P < 0.05).

88
Discussion
The plant‐herbivore interaction described in this study represents an unusual marine
plant defense association in which the annual alga Desmarestia viridis provides a
short‐term shelter for juvenile stages of the kelp Agarum cribrosum from grazing by
the green sea urchin, Strongylocentrotus droebachiensis. The repulsion of sea urchins
by the movement of algal fronds (induced by water motion) has been previously
noted for D. viridis (Himmelman & Steele, 1971; Himmelman, 1984; Konar, 2000),
Desmarestia ligulata (Dayton, 1985a), and the kelp Laminaria pallida
(Velimirov & Griffiths, 1979). We showed that the cover of D. viridis had a strong
impact on the spatial distribution of both juvenile A. cribrosum and urchins, but in
opposite ways. Juvenile A. cribrosum were almost always restricted to the area swept
by D. viridis plants, and urchins were mostly located outside the swept areas.
The variations observed between our two study sites indicate that the relationship
between the repulsive effect of D. viridis on urchins and the recruitment
of A. cribrosum is sensitive to several factors. Although urchin densities outside of
areas covered by D. viridis were similar on the western and eastern sides of
Île aux Goélands, urchins were better able to invade the area swept by D. viridis on
the western side. Should grazing by urchins be responsible for the distribution of
A. cribrosum juveniles, one might predict that the densities of juveniles under
D. viridis would have been lower on the western than on the eastern side of the island
(urchins were more abundant under the alga on the western side). However, the
inverse was observed. This seemingly contradicting pattern could be due to such
factors as changes in urchin behavior related to temporal changes in water movement
(e.g., sampling during a calm period) or to higher levels of A. cribrosum recruitment
on the western side.
Variation was also seen at finer spatial scales. Urchins on the western side of
Île aux Goélands were more abundant on the south side of D. viridis, whereas
A. cribrosum juveniles tended to be most common on the north side, a pattern
consistent with the idea that recruitment success is related to grazing intensity.

89
Again, the hydrodynamic conditions might explain this variation. The strongest
subtidal currents were observed on the western side of the island and were usually
oriented in a northerly direction. The speed and orientation data suggest there may
have been prolonged periods of unidirectional flow that may have dampened the
wave‐induced oscillations in shallow water. If so, one would expect the deterrence of
urchins by D. viridis to be strongest in the northerly direction. Green sea urchins are
highly mobile animals (maximum 2.6 m h‐1, P. Gagnon, unpublished data) and
readily shift their foraging behavior in response to other organisms and to physical
conditions (Garnick, 1978; Larson et al., 1980; Johnson & Mann, 1982; Mann et al.,
1984; Scheibling & Hamm, 1991). Thus, they could have temporarily attacked the
D. viridis plants from the southern side during periods when currents push the
canopy towards the north. These results indicate the importance of considering
the behavioral repertoire of mobile consumers relative to variations in the small‐scale
physical environment.
The probability of contact between urchins and juvenile A. cribrosum appears
to be influenced by the nature of the D. viridis canopy itself. Larger D. viridis plants
appear to provide better protection from urchins as higher densities of juvenile
A. cribrosum were found beneath them. We did not detect an effect of D. viridis size on
the density of urchins, suggesting that density alone does not always predict the
potential effect of herbivory. The enhanced recruitment under large plants was
possibly because the larger plants interfered more with the urchin’s foraging.
Alternatively, larger plants with their greater swept areas may provide a buffer zone
with moderate levels of grazing around a more highly protected central zone. This
idea is supported by the micro‐distributional data that showed higher densities
of juveniles, and corresponding lower densities of urchins, in the center of the swept
area. Increased algal recruitment in areas swept by larger plants has also been
reported for the kelp Laminaria pallida in South Africa (Velimirov & Griffiths, 1979).
The increase was attributed to decreased disturbance of the bottom by the fronds,
because the stiffer stipes of the large plants held the fronds away from this zone.
Although the stipes of D. viridis do not appear to stiffen with size, the branches near

90
the holdfast appear to move less than the outer branches (possibly because wave
action is attenuated). In contrast, all branches of small plants move back and forth
with wave action. The increased number of A. cribrosum juveniles near the holdfasts
of larger D. viridis might similarly be due to a reduced level of frond abrasion.
The damage to a plant from a herbivore is a function of (1) the likelihood that
the herbivore will come into contact with the plant, (2) the probability that the contact
leads to a portion of the plant being eaten, and (3) the loss in fitness due to damage
from the herbivore (Lubchenco & Gaines, 1981). During the 1997 experiment, the
probability of contact between an urchin and an A. cribrosum juvenile was 6 times
greater in the absence than in the presence of a D. viridis canopy. Furthermore, in the
1998 experiment tissue loss was 9 times greater in the absence than in the presence of
a D. viridis canopy. Thus, a D. viridis cover reduces both the probability of contact
with urchins and also tissue loss from urchin grazing. Extension of our tissue loss
curves, assuming the levels of interactions are similar over a longer period, suggests
that A. cribrosum juveniles would survive an order of magnitude longer in the
presence than in the absence of a D. viridis canopy. Van Alstyne (1999) showed that
many herbivores prefer the juvenile stages of algae, perhaps because chemical
or morphological defenses are less well developed. Possibly juvenile A. cribrosum
undergo chemical or structural changes to increase resistance to grazing as they age.
Adult A. cribrosum are not preferred foods of urchins (Vadas, 1977; Larson et al., 1980;
Himmelman & Nédélec, 1990). Thus, an association with D. viridis may provide
A. cribrosum juveniles with an escape from grazing during the critical period when
their defenses are not yet well developed.
In addition to affecting urchin behavior, macrophyte canopies can also reduce
(1) flow (Ackerman & Okubo, 1993; Hurd et al., 1997), and thus particle transport
(Eckman et al., 1989) and nutrient exchange (Gerard, 1982), and (2) light availability
(Gerard, 1984; Reed & Foster, 1984). Reduced light and nutrients could reduce growth
rates of plants growing under an algal canopy. For example, a cover of the brown
alga Sargassum filipendula has been shown to decrease the growth of the associated

91
red alga Gracilaria tikvahiae (Pfister & Hay, 1988). Vadas (1968) has shown that growth
of A. cribrosum is sensitive to reduced light intensities, which is consistent with our
39‐d experiment in which the growth rate of juvenile A. cribrosum was 29 % less
under a D. viridis canopy than with the canopy removed. This decrease was not
significant but might have been if the experiment had run for a longer period or
sample size was increased. Nevertheless, on urchin barrens, the costs of loss in
growth associated with living under a canopy are clearly outweighed by the benefits
of reduced urchin grazing.
Most studies of associational plant defenses have described the results of
long‐term interactions between species. In contrast, the association of A. cribrosum
with D. viridis is short‐lived because of the annual life cycle of D. viridis.
Our observations in 1998 and 1999 show that the length of D. viridis plants decreases
by about 1.5 % daily during the summer, and that all the plants are gone by late
October. Thus, the protection provided by D. viridis diminishes over the summer and
disappears in the autumn. Nevertheless, two factors may favor the survival of the
remaining juveniles in the autumn. The first is the decrease in feeding rates
of the urchins during late summer and autumn (Vadas, 1977; Larson et al., 1980;
Meidel & Scheibling, 1998). The second is that juveniles may switch from an escape to
a defensive strategy once they have reached a size where their own defenses are
developed (e.g., more polyphenolics, tougher tissues, and a size where the sweeping
action of its own blade might deter urchin attacks).
The interaction that we examined appears to represent a successional step
leading to the formation of mature stands of A. cribrosum. Thus, D. viridis colonizes
and grows rapidly during the winter, and then provides areas of decreased urchin
grazing where vulnerable A. cribrosum juveniles can become established. Then, the
A. cribrosum juveniles may be able to survive on their own when D. viridis disappears
in the autumn. Such an interaction would represent successional facilitation
(sensu Connell & Slatyer, 1977), which has been receiving increasing recognition as a
key process structuring communities (Walker et al., 1986; Wood & del Moral, 1987;

92
Callaway, 1995; Bruno & Bertness, 2001). Long‐term studies are required to determine
the survival rate of young A. cribrosum once the D. viridis cover disappears.
Such studies should also evaluate the influence of such factors as topographical
heterogeneity, ice scouring (which may destroy or disperse urchins in localized areas)
and wave action.

Chapitre V
Multiple plant defenses against herbivores: the interplay of chemical, morphological, and hydrodynamic factors

94
Résumé
Les plantes disposent d’un large éventail de mécanismes pour repousser les
herbivores. Même si les défenses chimiques sont bien documentées dans
les communautés terrestres et marines, peu d’études ont examiné les interactions
possibles entre la répulsion chimique et les autres types de défense. Nous avons
réalisé des expériences en laboratoire afin de déterminer si la résistance de
Desmarestia viridis (algue brune fortement acide et délicatement branchue) au
broutage de l’oursin s’explique par ses propriétés chimiques ou par une défense
mécanique attribuable à sa morphologie. Lors d’expériences de chémodétection en
utilisant un labyrinthe en Y, les oursins étaient attirés par des frondes de D. viridis
dans certains essais, mais repoussés dans d’autres (la température variait entre les
différents essais). Au contraire, les oursins étaient ni attirés, ni repoussés par leur
nourriture préférée, la laminaire Alaria esculenta. Des tests dans un générateur de
vagues ont démontré que l’agrégation des oursins près ou sur D. viridis diminuait
avec l’augmentation du mouvement de balayage provoqué par l’augmentation du
mouvement de l’eau. Cette habileté de D. viridis à repousser les attaques de l’oursin
décroissait drastiquement lorsque la densité de l’oursin atteignait un niveau critique.
Ces résultats sont comparables à nos observations de terrain à l’effet que les oursins
attaquent facilement D. viridis lorsque les conditions sont calmes mais évitent le
contact avec l’algue lorsqu’agitée par l’action des vagues, mêmes modérées.
Contrairement aux spéculations littéraires que l’acidité des Desmarestiales procure
une protection contre le broutage, nos résultats démontrent que la composition
chimique de D. viridis est à elle seule ni nécessaire, ni suffisante pour protéger l’algue
contre les oursins. Nous démontrons que la protection de D. viridis contre les
herbivores dépend principalement de l’interaction entre sa morphologie branchue et
l’action des vagues.

95
Abstract
Plants rely on a wide array of mechanisms to deter herbivores. Although chemical
defenses are well documented in terrestrial and marine communities, few studies
have examined possible interactions between chemical deterrence and other types of
defense. We conducted laboratory experiments to determine if the resistance of the
highly acidic, delicately‐branched, brown alga Desmarestia viridis to urchin grazing is
explained by its chemical properties or by mechanical defenses related to its
morphology. In Y‐maze chemodetection experiments, urchins were attracted by
D. viridis fronds in some trials but repelled in others (temperature varied among
trials). In contrast, urchins were neither attracted to nor repelled by their preferred
food, the kelp Alaria esculenta. Trials in a wave tank showed that the aggregation of
urchins near or on D. viridis decreased as the sweeping motion of D. viridis increased
with increased water motion. This ability of D. viridis to repel urchin attacks
decreased markedly when urchin density attained a critical level. These results are
consistent with field observations that urchins readily attack D. viridis under calm
conditions but avoid contact with the alga when it is moved about even by moderate
wave action. Contrary to speculation in the literature that the acidic nature of
Desmarestiales provides protection from grazers, our results show that the chemical
make‐up of D. viridis alone is neither necessary nor sufficient to protect the alga from
urchins. We show that the protection of D. viridis from herbivores depends primarily
on the interaction between its highly‐branched morphology and wave action.

96
Introduction
Herbivory is ubiquitous in terrestrial and marine communities, and the substantial
increase in studies on plant‐herbivore interactions over the last three decades
(see reviews in Lubchenco & Gaines, 1981; Hay & Fenical, 1988; Lindroth, 1989;
Coley & Barone, 1996; Zamora et al., 1999) has lead to a better understanding of
processes regulating populations and communities (Dayton, 1985b; Karban & Myers,
1989; Sousa & Connell, 1992; Callaway et al., 2000; Schmitz et al., 2000). In response to
herbivory, plants have evolved a variety of defenses including spatial or temporal
escapes (Atsatt & OʹDowd, 1976; Hay, 1986), morphological defenses (Duffy & Hay,
1990; Hay et al., 1994; Lucas et al., 2000), and chemical deterrents (Hay & Fenical,
1988; Paul & Van Alstyne, 1992). Most recent studies have focused on chemical
defenses (e.g., Pavia & Toth, 2000; Taylor et al., 2002), perhaps to the neglect of other
mechanisms and the possibility of multiple plant defenses (Hay et al., 1994).
Herbivory can be intense in marine benthic communities (Lubchenco &
Gaines, 1981), especially in tropical reefs (Carpenter, 1986; Lewis, 1986) and nearshore
temperate habitats (Breen & Mann, 1976b; Scheibling et al., 1999; Gagnon et al., in
press) where intensive grazing by fishes and sea urchins can lead to the formation of
“barren zones” (Lawrence, 1975). Obvious mechanical defenses, such as calcified
tissues, have long been recognized as a key adaptation to living in heavily grazed
environments (Steneck, 1986), but more recent studies have demonstrated that
chemical defenses are also common in marine plants and can be highly effective in
deterring herbivores (Duffy & Hay, 1990; Bolser & Hay, 1996; Schnitzler et al., 2001).
Brown algae of the order Desmarestiales, common in temperate coastal waters
(Peters et al., 1997), are unusual because of their production and storage of high
concentrations of sulfuric acid (H2SO4) within cell vacuoles, resulting in extremely
low internal pH (down to ~0.5 pH, McClintock et al., 1982). A number of studies have
suggested this trait chemically repulses grazers (Anderson & Velimirov, 1982;
Dayton, 1985a; Himmelman & Nédélec, 1990). This idea, although attractive, has only
recently received experimental support: the addition of sulfuric acid to an agar‐based

97
food source reduced urchin feeding rates in the laboratory (Pelletreau
& Muller‐Parker, 2002). However, field observations show that urchins feed readily
on Desmarestia sp. under certain environmental conditions (Himmelman & Steele,
1971; Cowen et al., 1982; Konar, 2000). Thus, laboratory studies might not reflect how
grazers respond to acidic species of Desmarestia, as they do not incorporate important
environmental factors. Water motion and temperature, urchin density and motivation
to feed likely affect the behavior of urchins (Garnick, 1978; Bernstein et al., 1983;
Mann et al., 1984; Vadas et al., 1986; Kawamata, 1998) and could modify the efficacy
of chemical defenses.
In the Mingan Islands, northern Gulf of St. Lawrence, the green sea urchin,
Strongylocentrotus droebachiensis, dominates bedrock platforms, and, as in many other
regions, its grazing leads to the formation of urchin barrens (Lawrence, 1975). This
community state is strikingly different from adjacent shallow zones where urchins
are scarce and kelp are abundant (primarily Alaria esculenta). The causal nature of this
pattern is well established both in the Mingan Islands (Gagnon et al., in press) and
elsewhere (e.g., Pearse & Hines, 1979; Himmelman et al., 1983; Witman, 1987;
Scheibling et al., 1999): removal of urchins leads to rapid colonization by kelp and
other algae. Desmarestia viridis is an exception to this pattern, being one of the few
fleshy macrophytes that can establish on urchin barrens. It is an annual alga that
occurs exclusively on barrens, either individually or in small groups (Fig. 5.1). It has
a small holdfast, a thin highly‐flexible stipe, and a profusely‐branched, filamentous
frond that can measure up to 1.5 m in length.
The occurrence of this fleshy macrophyte in an intensively‐grazed
environment, along with its known high acidity (McClintock et al., 1982), suggests
that it deters grazing using chemical defenses. Several observations support this idea.
First, D. viridis is only consumed at low rates when offered to urchins in the
laboratory (Himmelman & Nédélec, 1990). Second, field experiments comparing the
degree to which urchins move towards monospecific bundles of various algae reveal

98
Fig. 5.1. An isolated plant of Desmarestia viridis (~60 cm in length) surrounded by a dense aggregation of the green sea urchin, Strongylocentrotus droebachiensis, at 5 m in depth on a gently‐sloped bedrock platform in the Mingan Islands in June 2000.

99
that urchins are not attracted to D. viridis (Himmelman & Nédélec, 1990) when other
algal species are present. Third, the consumption rate of D. viridis is generally much
lower than for preferred algae such as the kelps A. esculenta and Laminaria spp.
(Vadas, 1977; Larson et al., 1980; Himmelman, 1984; Himmelman & Nédélec, 1990).
Although the evidence for chemical deterrence is convincing, other
possibilities have not been adequately explored. The delicate structure of D. viridis
permits the frond to sweep back and forth over the bottom with even slight wave
action, and observations and experiments indicate that urchins are much less
abundant under a D. viridis canopy (Himmelman, 1984; Konar, 2000; Gagnon et al.,
2003b) as well as for other algae (Velimirov & Griffiths, 1979; Dayton, 1985a).
Although the low number of urchins under Desmarestia spp. has been interpreted as a
response to the chemicals in the fronds (e.g., ʺacid broomingʺ sensu Dayton, 1985a),
the repulsive effect may be due simply to the mechanical contact between algal
fronds and urchins (Konar, 2000). This is likely the case for the kelp Laminaria pallida
(Velimirov & Griffiths, 1979) that has no known chemical defenses.
Here, we test whether the ability of D. viridis to resist urchin grazing depends
on the interaction between its morphology, hydrodynamics and urchin abundance, or
relies on its chemical make‐up. Our objectives are to (1) compare the degree to which
urchins are attracted towards D. viridis and the preferred alga A. esculenta,
(2) quantify the accumulation of urchins on and around D. viridis under different
levels of wave action, and (3) determine whether urchin density affects the degree to
which urchins accumulate on D. viridis.
Materials and methods
Our study was conducted during the summers of 1999 and 2001 at Havre‐Saint‐Pierre
in the Mingan Islands in the northern Gulf of St. Lawrence, eastern Canada
(50°13´6˝N, 63°41´12˝W). Experiments were performed in the laboratory with the
green sea urchin, Strongylocentrotus droebachiensis (2 to 6 cm in diameter), and the

100
algae Desmarestia viridis and Alaria esculenta, which were all collected at depths of
3‐8 m around Île aux Goélands and Île du Havre (Fig. 2.2), two islands located
approximately 5 km from the laboratory, and then maintained in tanks continually
supplied with seawater pumped from 10 m in depth.
Response of urchins to odors of Desmarestia viridis
To examine whether Desmarestia viridis chemically repulses Strongylocentrotus
droebachiensis, we compared urchin displacement towards chemical odors released by
D. viridis and the preferred alga, Alaria esculenta, in a Y‐maze (Castilla & Crisp, 1970).
The experiment included eight treatments of which six were conducted between
3‐6 °C: (1) a control for asymmetries in the maze (flowing seawater only); (2) a control
for the presence of conspecifics (6 urchins measuring 4 cm in test diameter); and
treatments with algal stimuli, (3) 75‐g piece of an A. esculenta frond; (4) 250‐g piece of
an A. esculenta frond; (5) 75‐g piece of a D. viridis frond; (6) 250‐g piece of a D. viridis
frond. The two other treatments, (7) a control for asymmetries in the maze (flowing
seawater only) and (8) 250‐g piece of a D. viridis frond, were conducted at higher
temperatures (10‐11 °C) as temperature can affect the displacement and
chemodetection ability of the green sea urchin (Mann et al., 1984). Our system did not
allow us to control water temperature, but we could take advantage of natural
variations.
Each trial began with the addition of the stimulus to one arm (45 cm long and
17 cm wide) of the Y‐maze followed 30 s later by the addition of 10 urchins
(4‐cm diameter) to the basal portion of the Y‐maze (20 cm long and 17 cm wide). This
sequence allowed odors to spread towards the mixing zone by the flow of seawater
(2.4 L min‐1) running through perforations positioned at 0.5 cm above the bottom in
the end of each arm. The water from the two arms mixed just below the point where
the arms joined and left the maze through perforations at 5.0 cm above the bottom at
the end of the basal portion, therefore maintaining a 5‐cm depth throughout the
maze.

101
The displacement of the urchins into the arms of the maze was evaluated every
10 min during a 40‐min period. Once observed in an arm, urchins were removed to
avoid interference with other urchins. At the end of each trial, those urchins that had
remained in the basal arm were considered to have not made a choice. Time and
logistic considerations limited us to 4 trials of the same treatment per day. Each
treatment was repeated 12 times (= 3 blocks x 4 trials) between 23 June and 17 August
of 1999. To eliminate possible biases due to asymmetries in the maze or in the
laboratory (e.g., light), we added the stimulus to a randomly‐selected arm of the maze
at the start of each day and then alternated the arm into which the stimulus was
added for subsequent trials. Between trials, the maze was drained and wiped with
a cotton towel.
Mechanical repulsion of urchins by Desmarestia viridis
To examine the degree to which the wave‐induced movement of Desmarestia viridis
fronds mechanically repulses sea urchins, we quantified the success of attack by
urchins on D. viridis or an algal mimic (defined here as the number of urchins in
contact with the alga or the mimic) in five treatments in a wave tank: (1) in the
absence of D. viridis and waves (control), (2) in the presence of a cotton mop without
waves to measure the tendency of urchins to aggregate on a chemically‐neutral,
D. viridis‐like mimic, (3) in the presence of D. viridis without waves to quantify the
vulnerability of D. viridis in the absence of wave action, (4) in the presence of D. viridis
with waves that caused a 5‐cm sweep of the fronds, and (5) in the presence of
D. viridis with waves which produced a 10‐cm sweep of the fronds. Sweep was
defined as the marginal extension of the area touched by the plant, and not the total
displacement of any particular part of the alga.
The wave tank has been described previously (see Gagnon et al., 2003a; but
also Appendix 5.1). A pulley system allowed us to control the back and forth water
motion and thus the amplitude of the sweeping action of D. viridis. The tank bottom
measured 1.94 x 1.22 m and was completely covered with concrete tiles with their
surface sculpted with undulations, holes and cracks to simulate the irregularities of

102
the natural substratum. Each corner of the tank was illuminated by a 150 W floodlight
positioned 90 cm above the bottom. For treatments with D. viridis or the algal mimic,
either a new plant (50‐60 cm in length) or a mop head with 50‐cm long cotton
filaments was attached with a plastic cable tie to an eye‐bolt at the center of the tank.
For the treatments with water motion, the system was set up to generate
23 wave cycles min‐1, equivalent to the wave frequency under moderate winds
(P. Gagnon, personal observation). To manipulate the displacement of the algal frond,
the height of the water column was adjusted to alter the strength of the waves
(deeper water produced stronger waves).
Urchins (2 to 6 cm in test diameter) were collected in July 2001 near
Île aux Goélands, and then haphazardly split into 5 groups of 275 individuals that
were fed every third day with the kelp Alaria esculenta until the end of the
experiments. The trials were run over 5 consecutive days, starting 2 d after
collection (sea water temperature: 1.5 to 5.5 °C), and the 5 groups of urchins were
subjected each day to one of the 5 treatments in a Latin square design.
Desmarestia viridis plants were freshly collected from near Île aux Goélands the
morning of each trial and were kept continually immerged in plastic buckets
provided with a constant inflow of seawater to avoid the release of chemical
compounds induced by stagnation of the water or by exposition of the fronds to air.
Mop heads were soaked for 24 h to minimize possible release of odors from the
cotton filaments. To explore possible site differences, a second group of urchins was
collected two weeks later near Île du Havre and subjected to an identical second
series of trials. Unexpectedly, water temperatures changed significantly between the
two experiments, and thus the effect of site could not be separated from an effect of
temperature.
In each of 50 trials, the urchins were first evenly distributed over the entire
tank bottom at a density of 116 m‐2, equivalent to an intermediate density of urchins
on barrens in the Mingan Islands (Gagnon et al., 2003b; Gagnon et al., in press). After
20, 40 and 60 min, we counted the number of urchins in contact with D. viridis or the

103
algal mimic and the number of urchins on the walls of the tank. We interpreted the
number of urchins on the walls as an index of the tendency of urchins to move within
the tank. For those treatments with water motion, the urchins were allowed 1 min to
attach themselves to the bottom before the motor was turned on. Prior to each new
trial, the tank was drained, cleaned and refilled with fresh seawater.
Effect of urchin density on attack success on Desmarestia viridis
We explored the importance of urchin density in determining the frequency of urchin
attacks on Desmarestia viridis by quantifying attacks under three densities, 78 m‐2,
136 m‐2 and 194 m‐2. These trials were run with water motion that produced a 5‐cm
sweep of the D. viridis fronds. On each of six sampling days (in late July and in early
August), urchins (2 to 6 cm in diameter) were collected near Île aux Goélands and
split into 3 groups to provide the necessary densities. The urchins were maintained in
running seawater for 3‐5 h prior to the trials. The procedure was as described above
(in the treatment with D. viridis with a 5‐cm sweep), except that each group of urchins
was used for just one trial (order chosen at random). Water temperatures varied
between 3 and 7 °C.
Field observations
To compare observations in our wave tank with those in the field, we counted the
numbers of urchins in contact with Desmarestia viridis plants at times of contrasting
hydrodynamic conditions at Île aux Goélands. The urchins in contact with
8 haphazardly‐chosen D. viridis plants (37‐61 cm in length) were sampled when water
motion was moderate (slack tide and ~29 km h‐1 winds), and again 4 d later when
there was low water motion (slack tide and ~9 km h‐1 winds). These conditions caused
the plants to sweep a distance of 10‐15 and <2 cm, respectively.
Statistical analysis
We used a three‐way ANOVA to analyze the tendency of the urchins to move
towards various chemical stimuli with the factors of Block (3), Treatment (8), and

104
Arm (3). We did not test for normality and homoscedasticity in the data as we treated
the analysis as a particular case of the generalized linear models (McCullagh
& Nelder, 1989; Proc Genmod, SAS Institute Inc., 1999) that assumed a binomial
distribution of the response variable (presence or absence of urchins in each arm).
Repeated measures ANOVAs (Hand & Taylor, 1987) with two nested elements
(Day of sampling within Site where urchins were collected and Group of urchins
within Site where urchins were collected) were used to examine the effect of water
motion and algal movement on (1) the attack success of urchins on Desmarestia viridis
and (2) the tendency of urchins to move within the wave tank. The factors were Site
(urchins collected from Île aux Goélands or Île du Havre), Day (10 sampling days),
Group (10 groups of urchins), Treatment (4 for the analysis on the attack success and
5 for the tendency of urchins to move) and Time (after 20, 40 and 60 min). Since
variances in the first analysis were similar over time within any of the treatments and
the correlation in the data over time was similar for all treatments, we used a pooled
covariance structure in the ANOVA (Proc Mixed, Type = CS, SAS Institute Inc., 1999).
However, because we detected fluctuations in the variances over time in the second
analysis, we modeled the covariance structure with a heterogeneous compound
symmetry structure (Proc Mixed, Type = CSH, SAS Institute Inc., 1999). We applied
the first analysis to the square‐root transformed data to obtain normality in the
distribution of the data for each observation time, whereas the second analysis was
applied to the raw data, which were normally distributed for each observation time.
The effects of urchin density on (1) the attack success of urchins on D. viridis
and (2) the tendency of urchins to move within the wave tank were analyzed using
repeated measures ANOVAs with the factors Block (6), Density (low, intermediate
and high urchin density), and Time (after 20, 40 and 60 min). Since we detected
fluctuations in the variances over time in both analyses, we modeled the covariance
structure with a heterogeneous compound symmetry structure (Proc Mixed,
Type = CSH, SAS Institute Inc., 1999). We applied both analyses to the raw data as the
data for each observation time were normally distributed.

105
In the above analyses, normality was verified using Shapiro‐Wilk’s statistic
(SAS Institute Inc., 1999) and homoscedasticity by examining the graphical
distribution of the residuals and by using Levene test (Snedecor & Cochran, 1989).
To detect differences among levels within a factor, we used least‐square means
multiple comparisons tests (LS means, SAS Institute Inc., 1999). A significance
threshold of 0.05 was used for all statistical tests.
Results
Response of urchins to odors of Desmarestia viridis
Our Y‐maze experiment examining the chemotaxic responses showed that under
certain circumstances urchins discriminated odors (Tables 5.1 and 5.2). The similar
numbers of urchins in the two arms in the control treatment (without stimuli at two
temperatures) indicated that there were no substantial asymmetries in the system
(Table 5.2). However, the smaller proportion of the urchins that remained in the basal
portion of the maze at the higher temperature (19.2 % at 10.3 °C versus 41.7 % at
4.7 °C; LS means, P = 0.0002) suggested that urchin displacements increased with
increasing temperature. There was no evidence of an effect of conspecifics (Table 5.2),
but the lower proportion of urchins that remained in the basal portion compared to
the test without conspecifics at 4.7 °C (LS means, P = 0.0054) suggested that odors
from conspecifics may also increase the tendency of urchins to move.
The response to chemical odors from algae provided variable results
depending on the algal species. Urchins had no reaction to Alaria esculena, but they
were either repelled or attracted to Desmarestia viridis, depending on the exact
treatment. Nearly 1.5 times as many urchins moved into the arm containing 75 g
of D. viridis as into the empty arm (LS means, P = 0.022; Table 5.2), demonstrating an
attraction. In contrast, the response to larger pieces of D. viridis (250 g) was the
opposite, and there was even greater repulsion at a higher temperature.

106
Table 5.1. Summary of the three‐way ANOVA (generalized linear model) examining the effect of Block (3 blocks of 4 replicates each), Treatment (8 stimuli), and Arm (3) on the chemotaxic response of urchins Strongylocentrotus droebachiensis to various odors (the algae Desmarestia viridis and Alaria esculenta and other conspecifics) in the Y‐maze experiment.
Source of variation df χ2 P
Block 2 1.22 0.57
Treatment 7 8.22 0.31
Block x Treatment 14 5.77 0.97
Arm 2 120.46 <0.0001
Block x Arm 4 3.61 0.46
Treatment x Arm 14 150.74 <0.0001
Block x Treatment x Arm 28 51.68 0.0042

107
Table 5.2. Directional response of the urchin Strongylocentrotus droebachiensis to various chemical stimuli in a Y‐maze choice chamber. P‐values are from statistical comparisons between the proportions of urchins that moved to the stimulus arm and the empty arm or to the two empty arms in the control tests without stimulus (LS means, P < 0.05).
Treatment Proportion of urchins (%)
Stimulus
Quantity
(g)
Sea water
temperature
(SD) (°C)
Basal
portion
Stimulus
arm
Empty
arm
P
None ‐ 4.7 (1.3) 41.7 26.7 31.6 0.41
None ‐ 10.3 (1.0) 19.2 40.0 40.8 0.89
6 urchins ‐ 5.3 (0.5) 5.0 45.8 49.2 0.59
A. esculenta 75 5.5 (0.7) 17.5 45.8 36.7 0.16
A. esculenta 250 5.0 (0.0) 20.0 42.5 37.5 0.41
D. viridis 75 5.8 (0.7) 30.0 41.7 28.3 0.022
D. viridis 250 3.8 (1.7) 28.3 25.0 46.7 0.0005
D. viridis 250 11.1 (0.6) 10.8 25.8 63.4 <0.0001

108
Mechanical repulsion of urchins by Desmarestia viridis
Our wave tank trials showed that the number of urchins in contact with Desmarestia
viridis decreased sharply with an increase in the amplitude of the movement of the
alga (Table 5.3; Fig. 5.2). In trials with urchins from Île aux Goélands, the overall
number of urchins (the mean for the three 20‐min intervals) attacking a motionless
D. viridis was more than an order of magnitude greater than for D. viridis with a 5‐cm
sweep, and two orders of magnitude greater than for D. viridis with a 10‐cm sweep.
Under still conditions, D. viridis was attacked much more than the algal mimic
(LS means, P = 0.0029), suggesting that urchins were attracted to D. viridis. Our
measurements at 20‐min intervals indicated that the cumulative number of urchins
on D. viridis increased only slightly after the first 20 min and then showed no further
change with an increase in wave action and the sweeping motion of the alga (Fig. 5.2).
Although the same pattern applied to the trials with urchins from Île du Havre, the
attacks on both mimic and natural D. viridis were much greater (Fig. 5.2). The overall
number of urchins in contact with both non‐sweeping D. viridis and the algal mimic
was 2 to 3‐fold greater for urchins from Île du Havre than for urchins from Île aux
Goélands (LS means, P < 0.0001). Whereas urchins from Île aux Goélands were
virtually unable to attack sweeping fronds of D. viridis, those from Île du Havre had
some success in attacking D. viridis with a 5‐cm sweep. The lower water temperatures
during the trials with urchins from Île aux Goélands (3.6 ± 1.4 °C) relative to those
with urchins from Île du Havre (8.0 ± 0.9 °C) could have contributed to the
differences seen between the two groups of urchins.
The numbers of urchins on the walls of the tank indicated that urchin
displacement varied markedly among treatments (Table 5.3; Fig. 5.2). The numbers of
urchins on the walls were similar in all three treatments without waves, but
decreased markedly with increasing water motion (Fig. 5.2). Urchins from both Île
aux Goélands and Île du Havre were >9 times more numerous on the walls in the
trials with non‐sweeping D. viridis than in the trials with 10‐cm‐sweeping D. viridis.

109
Table 5.3. Summary of repeated measures ANOVAs (mixed linear model) showing the effect of the Site where the urchins Strongylocentrotus droebachiensis were collected (Île aux Goélands and Île du Havre), Day of sampling (10 days), the Group of urchins being tested (10 groups), Treatment (4 in the first analysis and 5 in the second), and Time (observations at 20, 40 and 60 min) on the number of urchins in contact with Desmarestia viridis (or algal mimic) and on the walls of the wave tank. Data from the first analysis were square‐root transformed whereas the raw data were used in the second analysis.
Source of variation (fixed effects)
df (numerator)
df (denominator)
F‐value
P
In contact with D. viridis Site 1 16 77.66 <0.0001 Day(Site) 8 16 0.97 0.49 Group(Site) 8 16 0.90 0.54 Treatment 3 16 74.42 <0.0001 Time 2 64 49.48 <0.0001 Site x Treatment 3 16 7.80 0.0020 Site x Time 2 64 2.13 0.13 Treatment x Time 6 64 3.56 0.0042 Site x Treatment x Time 6 64 1.26 0.29
On the walls of the tank Site 1 24 0.26 0.62 Day(Site) 8 24 1.84 0.12 Group(Site) 8 24 0.90 0.53 Treatment 4 24 22.89 <0.0001 Time 2 80 132.50 <0.0001 Site x Treatment 4 24 0.18 0.94 Site x Time 2 80 0.78 0.46 Treatment x Time 8 80 15.65 <0.0001 Site x Treatment x Time 8 80 1.31 0.25

110
0
10
20
30
40
50
20 min
40 min
60 min
0
20
40
60
80
100
In contact with D. viridis
On the walls of the tank
No alga
or waves
Mimic
no
sweep
D. viridis
no
sweep
D. viridis
5-cm
sweep
D. viridis
10-cm
sweep
c
b
d d3.5 (1.0)
10.2 (1.9)
0.6 (0.2) 0.1 (0.1)
8.4 (1.2)
32.9 (3.7)
9.6 (1.3)
0.8 (0.3)
a
bb
d
48.9 (5.9) 53.4 (5.1)40.8 (5.5)
13.8 (1.9)
4.5 (0.5)
A AA
B
B
57.2 (7.9)
55.3 (7.3)
38.6 (4.2)
16.9 (2.9)
4.1 (0.6)
A
A
A
B
B
No alga
or waves
Mimic
no
sweep
D. viridis
no
sweep
D. viridis
5-cm
sweep
D. viridis
10-cm
sweep
Île aux Goélands Île du Havre
Fig. 5.2. Mean number (+ SE) of urchins Strongylocentrotus droebachiensis in contact with Desmarestia viridis or an algal mimic, and on the walls of the tank, during three consecutive 20‐min intervals in trials in a wave tank with urchins from two sites examining the effect of the amplitude of the sweeping motion of D. viridis. Mean values for the three 20‐min intervals (± SE) are shown above bars. Treatments not sharing the same letters are different (LS means tests, P < 0.05).

111
The measurements at 20‐min intervals indicated that numbers tended to level off
rapidly with increasing water motion (Fig. 5.2), and urchins were attached most
firmly at the end of trials in the 10‐cm sweep treatment (P. Gagnon, personal
observation). Finally, the absence of significant effects of either Day(Site)
or Group(Site) in the analysis on the effect of water motion and algal movement
indicated the repeated use of the same 5 groups of urchins did not affect the ability
of urchins to attack D. viridis or climb onto the walls (Table 5.3).
Effect of urchin density on attack success on Desmarestia viridis
Our wave tank trials at a constant 5‐cm sweep of Desmarestia viridis showed that the
number of urchins attacking plants increased with urchin density, but not in a linear
fashion (Table 5.4; Fig. 5.3). The numbers (means for the three 20‐min intervals) were
roughly the same at low and intermediate densities (LS means, P = 0.83), but were
more than 2‐ to 3‐fold greater at the highest density. The number of urchins attacking
D. viridis increased more rapidly during the high‐density trials than at either low or
intermediate densities. Indeed, the number of urchins attacking after 20 min in the
high‐density treatment was nearly identical to that attained after 60 min at low and
intermediate densities (Fig. 5.3).
Field observations
Our field observations of urchins in contact with Desmarestia viridis plants indicated
that the urchins were much more abundant on plants with a small (<2 cm) than
a larger (10‐15 cm) sweep, with 17.8 (SD = 2.9) and 0 urchins per plant, respectively,
a pattern highly consistent with our observations in the wave tank experiments. Most
of the urchins in contact with D. viridis had filaments of D. viridis in their jaws.
Discussion
Most studies of marine algal defenses against herbivory have described the effect of

112
Table 5.4. Summary of repeated measures ANOVAs (mixed linear model applied to raw data) showing the effect of Block (6), Density of urchins Strongylocentrotus droebachiensis (low, intermediate and high), and Time (observations at 20, 40 and 60 min) on the number of urchins in contact with Desmarestia viridis.
Source of variation (fixed effects)
df (numerator)
df (denominator)
F‐value
P
Block 5 10 0.86 0.54 Density 2 10 9.12 0.0056 Time 2 30 34.54 <0.0001 Density x Time 4 30 6.15 0.0010

113
0
10
20
30
40
Low Intermediate High
a
b
ccc
c
ccd d
Urchin density
7.2 (1.2) 6.4 (1.2)
19.1 (2.5)
20 min
40 min
60 min
Fig. 5.3. Mean number (+ SE) of urchins Strongylocentrotus droebachiensis in contact with Desmarestia viridis during three consecutive 20‐min intervals in trials in a wave tank at low (78 m‐2), intermediate (136 m‐2) and high (194 m‐2) urchin densities. Mean values for the three 20‐min intervals (± SE) are shown above bars. Bars not sharing the same letters, and treatments not bracketed by the same horizontal line (at the bottom) are different (LS means tests, P < 0.05).

114
single defensive traits, therefore neglecting possible additive or synergistic effects of
multiple defenses (Hay, 1996). Our study of the highly acidic brown alga Desmarestia
viridis to grazing by the green sea urchin, Strongylocentrotus droebachiensis, showed
that the chemical make‐up of the alga provides only limited protection from grazing.
Rather, the survival of D. viridis in intensively‐grazed subtidal environments depends
on the interaction between its delicate, finely‐branched structure and the physical
environment. Waves cause fronds to sweep back and forth over the bottom and
maintain urchins at a distance. This finding contradicts the perception that chemical
defenses are paramount in protecting acidic Desmarestiales species.
Sea urchins are highly selective feeders that have been reported to detect food
over a distance of several meters (Vadas, 1977; Garnick, 1978; Larson et al., 1980;
Mann et al., 1984). Multiple‐algae choice experiments have suggested that
Desmarestia spp. are avoided by urchins (Himmelman, 1984; Pelletreau & Muller‐
Parker, 2002). However, feeding experiments and field observations show that
urchins will consume Desmarestia species in the absence of alternative algal foods
(Himmelman & Steele, 1971; Cowen et al., 1982; Himmelman & Nédélec, 1990; Konar,
2000). Our Y‐maze study demonstrated that D. viridis can even attract urchins,
although this was not the case when there was a large quantity of the alga.
Unexpectedly, we found no chemotaxic response of urchins to odors of the preferred
food, Alaria esculenta.
Temperature is likely a key factor affecting the foraging behavior of the green
sea urchin as it appears to affect both its ability to respond to odors and the rate of
displacement. In the Mingan Islands, seawater temperatures usually oscillate around
6 °C during the summer (Gagnon et al., 2003b), although they rise to above 10 °C for
several days on two or three occasions each year (J. Himmelman and coworker’s
personal observations). Urchin behavior varied with temperature in the control trials
in our Y‐maze study, and we observed increased displacements at higher
temperatures as reported for green sea urchins (Mann et al., 1984). The two
treatments in which urchins avoided odors of D. viridis were those conducted near

115
the two summer temperature extremes for the Mingan Islands (3.8 and 11.1 °C).
At 5.0‐5.8 °C, urchins were attracted to D. viridis, but not to their preferred food,
Alaria esculenta. Repulsion of urchins by D. viridis at higher temperatures could be
related to increased acid release, but this is not likely the case at low temperatures.
Avoidance of algae by green sea urchins at low temperatures was reported by
Mann et al. (1984). Attractiveness towards a preferred alga (Laminaria longicruris)
declined with temperature and urchins were even repulsed at 2 °C. These
inexplicable results might be related to changes in the behavior and physiology of the
urchin at low temperatures.
Although there is indication that water motion can affect the behavioral
repertoire of animals (e.g., displacement and feeding rates, Rochette et al., 1994;
Kawamata, 1998; Gagnon et al., 2003a; Siddon & Witman, 2003), including behaviors
related to predation, herbivory, and competition (Cowen et al., 1982; Witman, 1987;
Menge et al., 1994), our study is the first to manipulate the hydrodynamic
environment to evaluate the possible deterrence of urchin attacks by wave‐induced
movement of algal fronds. Our wave tank trials demonstrated that the number of
urchins attacking a motionless D. viridis plant was orders of magnitude greater than
for plants moved about by waves. Moreover, the attack success of urchins on the alga
was inversely related to the amplitude of the wave‐induced sweeping motion of the
plants. This is consistent with our field observations that urchins attacks on D. viridis
are much greater during calmer conditions. Thus, both our laboratory and field
observations show that wave‐induced movements of D. viridis plays a major role in
protecting it from urchin grazing.
Although the repulsion of urchins appears to be due primarily to the indirect
effect of water motion causing sweeping of algal fronds, the urchins could also be
responding directly to water motion. This hypothesis is supported by the drop in the
number of urchins on the walls of the tank with increasing water motion. In the trials
with D. viridis, urchins were ~9 times more abundant on the walls at the end of the
trials with no sweep than in the trials with a 10‐cm sweep. Moreover, urchins did not

116
move after the first 20 min in the trials with the highest water motion. Current speeds
peaked at ~0.15 and 0.30 m s‐1 for 5‐cm and 10‐cm sweeping D. viridis, respectively
(as measured with a Doppler current meter), indicating that S. droebachiensis stops
moving at current speeds >0.30 m s‐1. This threshold current speed is less than half
than that reported in studies of effects of current speed on Strongylocentrotus nudus
(~0.70 m s‐1, Kawamata, 1998). Thus, water motion protects D. viridis from attacks by
S. droebachiensis first because hydrodynamic forces (e.g., drag, lift and acceleration
forces) decrease urchin movement and second due to the repulsion of urchins by the
sweeping of fronds. Given this hierarchical relationship, the deterrence of urchins by
the sweeping of algal fronds may only be important at intermediate flow speeds
when urchins are still moving.
The formation of dense aggregations appears to be an important part of the
foraging strategy of S. droebachiensis (Garnick, 1978; Bernstein et al., 1983; Vadas et al.,
1986), and can lead to the formation of grazing fronts than can advance through kelp
beds at a rate of up to 4.0 m month‐1. However, this only appears to occur when
urchins attain a critical biomass (Breen & Mann, 1976b; Scheibling et al., 1999;
Gagnon et al., in press). Our experiments demonstrate the key role of density in the
interaction between urchins and macroalgae. The relation of attack success to urchin
density is not linear but involves a threshold density, as with the formation of grazing
fronts. Our wave tank trials at different current speeds suggest that the critical
density increases with increased wave conditions.
In the Mingan Islands, D. viridis occurs exclusively on urchin barrens. Our field
observations indicate that urchins readily accumulate on, and consume D. viridis
under still conditions, but that urchins avoid this alga when the fronds move back
and forth with wave action. Thus, D. viridis is less well defended in the absence of
waves, and the chemical composition of D. viridis alone seems to provide little
protection from grazing. Why D. viridis produces acid remains unclear. Perhaps,
small amounts of acid are released when the fronds are swept across surfaces and act
synergistically with water motion to repel grazers, or the acid is more effective

117
against less voracious grazers (e.g., amphipods). The chemical defense may also only
be effective when more palatable algae are present. Alternatively, the primary role of
acid may not be a grazer deterrent, but rather act as an allelochemical or anti‐fouling
substance as shown for sulfuric acid and other chemicals in various species
interactions (Stoecker, 1978; Hay, 1996). Finally, D. viridis occurs in many regions
(Peters et al., 1997), and its capacity to produce acid might have evolved in response
to factors in other communities, and may not be a trait which provides advantages in
the northern Atlantic region.
Our study is the first to examine the interaction between chemical,
morphological and hydrodynamic factors in providing an alga with protection
against herbivory. We show that the survival of D. viridis in an intensively‐grazed
environment largely depends on its delicate structure. This contrasts with the
strategy of producing heavily calcified tissues as used by the coralline algae
commonly found on urchin barrens (Steneck, 1986). Beyond the documentation
of this novel plant defense, we have shown how the direct and indirect effects of
water motion can modulate species interactions, further providing evidence of the
importance of hydrodynamic factors in structuring of marine communities (Lissner,
1983; Witman & Dayton, 2001; Gagnon et al., 2003a). Finally, we demonstrate the
need to integrate a variety of biotic and abiotic variables, including algal chemistry
and morphology, water motion and temperature, and the abundance of predators, in
studies of algal defenses.

118
Appendix 5.1
Wave tank system. a) The rotational force generated by a 2‐HP electric motor is transmitted through a series of pillow blocks, axles, pulleys, and belts. (b) A perforated bar welded to the last axle (enlarged in the upper‐left corner, reverse view) is connected to the nearest of two plywood panels hinged to the bottom of the wave tank. Two metal bars connecting the top of the first panel with the top of the second causes the two panels to move back and forth in parallel producing oscillating waves.

Chapitre VI
Conclusion générale

120
Mon étude augmente notre connaissance des facteurs et mécanismes qui contrôlent
les communautés végétales de la zone infralittorale rocheuse de l’archipel de Mingan.
D’une part, elle constitue l’examen le plus détaillé de l’évolution temporelle de la
distribution et de l’abondance des macrophytes et quantifie pour la première fois
l’impact du broutage de l’oursin vert, Strongylocentrotus droebachiensis, sur la structure
et la régulation des assemblages algaux. D’autre part, elle identifie certains
mécanismes de défense qui permettent aux juvéniles d’Agarum cribrosum et à
Desmarestia viridis, les deux macrophytes dominants en zone dénudée, de résister au
broutage de l’oursin vert.
Dynamique de répartition des macrophytes
Le premier volet de l’étude démontre que la distribution et l’abondance des
macrophytes varient grandement selon (1) la communauté (zone des laminaires et
zone dénudée), (2) la période de l’année et (3) la localité (site) considérées. Les plus
grandes fluctuations surviennent dans la communauté des laminaires; pendant que
certaines ceintures reculent de plusieurs mètres en quelques mois, d’autres avancent
de façon tout aussi substantielle. Une large part de cette variabilité est attribuable au
broutage de l’oursin vert qui limite, de façon prononcée, le recrutement et la
succession algale, tel que démontré lors de l’exclusion d’oursins en zone dénudée. La
concordance entre la position des fronts d’oursins et la position de la limite inférieure
des ceintures tout au long de l’étude indique que l’effet du broutage se répercute à
l’échelle côtière en contrôlant la pénétration en profondeur des ceintures de
laminaires. Les retraits de ceintures les plus marqués surviennent pendant l’été,
lorsque les fronts de broutage s’intensifient, alors que la recolonisation a lieu
principalement en hiver, lorsque les populations d’oursins aux abords des ceintures
s’atténuent. L’efficacité d’un front de broutage à détruire une ceinture de laminaires
tend à augmenter avec la biomasse d’oursins dans le front. Cette efficacité serait
toutefois affectée par la composition végétale des ceintures. En effet, mes résultats
indiquent que la progression ascendante d’un front est retardée au contact d’agrégats
d’Agarum cribrosum et suggèrent fortement que l’établissement de larges bandes

121
d’A. cribrosum en marge inférieure des ceintures de laminaires augmente la stabilité
de ces dernières. Mes observations suggèrent que l’abrasion par les glaces en hiver
provoque de grandes fluctuations dans les populations d’oursins (par mortalité ou
dispersion des oursins).
En zone dénudée, la stabilité temporelle de la distribution et de l’abondance
des macrophytes charnus peut être étonnamment élevée malgré la forte abondance
d’oursins, comme c’est notamment le cas d’A. cribrosum. Les agrégats d’A. cribrosum
persistent pendant des périodes de plusieurs années (au moins deux ans), avec même
un léger taux d’accroissement. La résilience élevée des agrégats aux perturbations à
fine échelle (m) en fait un habitat relativement stable qui favorise le recrutement
algal, comme en témoigne l’abondance plus élevée de la rhodophycée Ptilota serrata et
des juvéniles d’A. cribrosum à l’intérieur qu’à l’extérieur des agrégats. Le recrutement
accru sous la canopée d’A. cribrosum s’expliquerait en partie par une réduction du
broutage de l’oursin dont l’abondance sous la canopée est faible (les oursins sont
majoritairement localisés à l’extérieur des agrégats). Les agrégats d’A. cribrosum
affectent donc positivement le reste de la communauté végétale en contribuant à
l’augmentation de la survie algale en zone dénudée.
Mécanismes de défense des macrophytes
Le second volet de l’étude démontre que la survie algale en zone dénudée repose
fondamentalement sur la modulation des interactions macrophytes‐oursins par les
propriétés hydrodynamiques du milieu. L’examen de la distribution des juvéniles
d’Agarum cribrosum en zone dénudée révèle qu’ils sont nettement plus nombreux sur
les aires balayées par les plants de Desmarestia viridis qu’en zone ouverte.
L’abondance des juvéniles sous un plant de D. viridis augmente avec la taille du plant
et décroît avec l’augmentation de la distance par rapport au centre du plant. Au
contraire, l’oursin est plus abondant à l’extérieur que sous un plant de D. viridis et son
abondance augmente avec la distance par rapport au centre du plant. Ce patron de
distribution serait principalement attribuable au mouvement de balayage de D. viridis
causé par les vagues, lequel mouvement empêcherait la pénétration des oursins sous

122
la canopée, résultant en une diminution du broutage de l’oursin sous la canopée et,
par conséquent, en un taux de survie accru des juvéniles. Les juvéniles sont
vulnérables au broutage de l’oursin; en moins de quatre jours, un juvénile dépourvu
d’un couvert de D. viridis subit une perte moyenne de plus de 50 % de sa longueur,
alors que cette perte n’est que de 3 % avec couvert. Même si la croissance des
juvéniles sous D. viridis est légèrement diminuée par rapport à leur croissance en
zone ouverte, cette perte est nettement compensée par le gain de protection contre le
broutage découlant de l’association. Mes résultats indiquent donc que la défense par
association avec D. viridis augmente considérablement la survie des juvéniles
d’A. cribrosum. Cette association défensive éphémère (D. viridis disparaît à l’automne)
est proposée comme étape de succession menant à la formation d’agrégats adulte
d’A. cribrosum.
L’investigation, en milieu contrôlé, de la réponse comportementale de l’oursin
face à D. viridis révèle que la propriété répulsive de l’algue est davantage liée à
l’interaction entre ses attributs morphologiques et l’action des vagues, qu’à sa
composition chimique. En l’absence de vagues, les oursins s’agrègent rapidement sur
l’algue immobile, alors qu’ils l’évitent en eau agitée, lorsque l’algue balaie le substrat.
L’intensité des attaques, telle que déterminée par le nombre d’oursins en contact avec
l’algue, diminue avec l’augmentation de la force des vagues et augmente avec
l’abondance des oursins. Le déplacement de l’oursin et l’intensité de ses attaques
contre D. viridis semblent augmenter avec l’accroissement de la température. L’oursin
n’est repoussé par l’odeur de D. viridis qu’à des températures correspondant aux
extrêmes estivaux dans l’archipel de Mingan. Ces résultats, couplés aux observations
in situ d’agrégats d’oursins s’alimentant aisément sur D. viridis en eau calme,
indiquent que la composition chimique de l’algue (e.g., acide sulfurique) ne contribue
que faiblement à la défense de l’algue contre le broutage de l’oursin.
Portée de l’étude
Mon étude contribue de plusieurs façons à l’avancement de notre connaissance de
l’écologie des communautés végétales marines et des interactions plantes‐herbivores:

123
1) Elle identifie l’oursin vert, Strongylocentrotus droebachiensis, comme facteur‐clé dans
la régulation de la distribution et de l’abondance des macrophytes dans l’archipel
de Mingan et fournit la première preuve quantitative que d’importantes
fluctuations saisonnières surviennent dans les populations d’oursins et de
macrophytes, tant au niveau de leur abondance que de leur structure de taille;
2) Elle est la seule étude ayant quantifié la dynamique de répartition naturelle des
assemblages algaux infralittoraux dans le golfe du Saint‐Laurent (et l’une des rares
dans le nord‐ouest de l’Atlantique). Ce genre d’étude est essentiel à notre
compréhension du fonctionnement de cet écosystème dans lequel les assemblages
algaux sont d’une grande importance pour le reste de la communauté;
3) Elle identifie les interactions positives (e.g., refuge par association) comme
mécanisme important pour la survie algale infralittorale et fournit, du coup, un
argument de poids aux écologistes de plus en plus nombreux à reconnaître l’impact
significatif de telles interactions dans la structure et la régulation des
communautés;
4) Elle souligne l’importance d’examiner plusieurs facteurs biotiques (e.g., attributs
morphologiques et chimiques des plantes et abondance de l’herbivore) et de
mesurer l’impact de variations dans les paramètres physiques (e.g., température,
conditions hydrodynamiques) dans les études d’interactions plantes‐herbivores;
5) Elle fournit la première preuve expérimentale quantitative que l’hydrodynamisme
peut moduler les interactions plantes‐herbivores en milieu marin en exerçant à la
fois un effet direct et indirect sur la quête alimentaire d’un herbivore.
La plupart des modèles élaborés pour tenter d’expliquer la structure des
communautés ont accordé une grande importance aux interactions négatives telles
que la prédation et la compétition (e.g., zones intertidale et infralittorale). Cet intérêt
scientifique élevé pour les interactions négatives a relégué en arrière‐plan les
interactions positives dont la reconnaissance de l’importance, en tant que mécanisme
structurant les communautés, s’est accrue au cours des dernières années

124
(e.g., marais salés). Les interactions positives, telles que le mutualisme et le
commensalisme, s’opèrent par l’entremise d’espèces fondatrices dont la présence
améliore la qualité d’un habitat (pour un ou plusieurs autres organismes de la
communauté) en modifiant ses propriétés physico‐chimiques ou en augmentant
l’hétérogénéité de l’environnement, créant ainsi des refuges spatiaux contre les stress
environnementaux ou la prédation. Dans un environnement où la prédation est
élevée et le recrutement des proies est faible, les interactions positives peuvent jouer
un rôle important dans la structure des communautés et contribuer à augmenter la
diversité spécifique.
Mon étude s’inscrit bien dans ce récent mouvement de reconnaissance du rôle
communautaire important que peuvent jouer les interactions positives. En effet, mes
résultats indiquent que l’oursin vert exerce une prédation considérable sur les
communautés algales infralittorales de l’archipel de Mingan, résultant en une baisse
marquée de l’abondance de plusieurs espèces (e.g., laminaires). Néanmoins, les
formes adultes (résistantes au broutage de l’oursin) des phaéophycées
Agarum cribrosum et Desmarestia viridis contribuent au maintien de la diversité
végétale en zone dénudée. Les agrégats d’A. cribrosum et les plaques de D. viridis
augmentent l’hétérogénéité spatiale de la zone dénudée et forment des micro‐habitats
répulsifs à l’oursin vert et à l’intérieur desquels le recrutement algal (e.g., juvéniles
d’A. cribrosum et Ptilota serrata) est nettement plus élevé qu’en milieu ouvert. À ce
titre, A. cribrosum et D. viridis semblent représenter deux espèces fondatrices, l’une
(A. cribrosum) exerçant un effet positif sur une période potentielle de plusieurs années
(espèce pérenne), l’autre (D. viridis) sur une période de quelques mois (espèce
annuelle). Peu étudiées, les interactions algales positives, telles que décrites ici dans
l’archipel de Mingan (e.g., refuge par association), pourraient s’avérer d’une
importance sous‐estimée dans la structure des communautés infralittorales marines.
Nouvelles questions et hypothèses
Mes travaux ont soulevé d’intéressantes questions et généré de nouvelles hypothèses
qui méritent un examen attentif:

125
1) Les patrons de variation observés dans la distribution et l’abondance des
macrophytes sont‐ils représentatifs des patrons à plus grande échelle? La réponse à
cette question passe impérativement par le recours à une méthode plus
performante que la plongée sous‐marine utilisée dans le cadre de cette étude et ne
pouvant suffire à quantifier de tels patrons que sur de courtes distances
(e.g, une centaine de mètres) au prix d’investissements considérables en temps et
efforts de plongée. L’utilisation de la cartographie infralittorale par imagerie
aérienne, bien que plus onéreuse que la plongée, s’avère extrêmement prometteuse
(des percées majeures dans ce domaine sont en cours) et se justifie par la réduction
substantielle du temps et des efforts d’échantillonnage ainsi que par l’élargissement
considérable des échelles spatiales et temporelles (i.e., sur des centaines de
kilomètres et pendant des décennies). La distribution en faible profondeur de la
plupart des macrophytes et la clarté habituelle de l’eau dans le nord du golfe du
Saint‐Laurent favorisent l’utilisation des techniques de photographies aériennes;
2) L’abrasion hivernale et printanière par les glaces exerce un impact notable sur la
structure des communautés intertidales dans le golfe et l’estuaire du Saint‐Laurent.
Mes résultats indiquent que l’effet des glaces dans l’archipel de Mingan s’étend à
des profondeurs pouvant dépasser 5 m et que l’établissement de la laminaire
Agarum cribrosum survient principalement en hiver. Il serait souhaitable de mener
d’autres études afin de tester rigoureusement les deux hypothèses selon lesquelles
l’abrasion par les glaces (1) contrôle les populations d’oursins en causant la
mortalité de ces derniers ou en provoquant leur migration en zones plus profondes
et (2) facilite l’établissement d’A. cribrosum en zone dénudée;
3) La présence de Desmarestia viridis en zone dénudée augmente nettement la survie
des juvéniles d’A. cribrosum en procurant à ces derniers un refuge éphémère contre
le broutage de l’oursin. Une investigation sur une plus longue période que celle
impliquée dans cette étude (i.e., >5 mois), incluant le suivi de juvéniles avant et
après la période hivernale, est requise afin d’évaluer l’importance de ce refuge par
association dans la formation d’agrégats adultes d’A. cribrosum;

126
4) Mes résultats indiquent que le taux de retrait des ceintures de laminaires tend à
augmenter avec la biomasse d’oursins dans les fronts de broutage. Il semble
toutefois que cette relation ne s’applique pas à l’ensemble des sites étudiés et
qu’elle puisse même changer de façon saisonnière. D’autres suivis temporels de la
position de la limite inférieure des ceintures de laminaires et de la structure des
fronts d’oursins, incluant une taille échantillonnale accrue et des mesures
automnales et hivernales, sont requis afin de mieux caractériser le comportement
de cette relation;
5) L’exploitation commerciale des stocks naturels d’oursins dans l’archipel de Mingan
est‐elle possible et si oui, comment procéder? Mes résultats indiquent qu’une telle
pratique devrait cibler les fronts de broutage aux abords des ceintures de
laminaires où l’abondance et la taille des oursins sont supérieures
comparativement à tout autre endroit du littoral. Ceci favoriserait la récolte rapide
des individus les plus susceptibles de rencontrer les standards commerciaux
(e.g., taille du test et index gonado‐somatique). D’autre part, l’élevage de stocks
d’oursins en mer (sea ranching) pourrait s’effectuer par l’adition d’oursins à
l’intérieur d’ouvertures pratiquées dans de larges agrégats d’A. cribrosum.
Les oursins seraient alors contenus par cette barrière algale naturelle, nourris par
l’apport anthropique de macrophytes et retirés à l’approche de l’hiver afin de
prévenir d’éventuelles pertes d’oursins causées par l’abrasion par les glaces.

127
Bibliographie
Ackerman J.D. & Okubo A. (1993). Reduced mixing in a marine macrophyte canopy. Funct. Ecol. 7: 305‐309.
Anderson R.J. & Velimirov B. (1982). An experimental investigation of the palatability of kelp bed algae to the sea urchin Parechinus angulosus Leske. Mar. Ecol. 3: 357‐373.
Archambault D. & Bourget E. (1983). Importance du régime de dénudation sur la structure et la succession des communautés intertidales de substrat rocheux en milieu subarctique. Can. J. Fish. Aquat. Sci. 40: 1278‐1292.
Atsatt P.R. & OʹDowd D.J. (1976). Plant defense guilds. Many plants are functionally interdependent with respect to their herbivores. Science 193: 24‐29.
Balch T. & Scheibling R.E. (2000). Temporal and spatial variability in settlement and recruitment of echinoderms in kelp beds and barrens in Nova Scotia. Mar. Ecol. Prog. Ser. 205: 139‐154.
Bazhin A.G. (1998). The sea urchin genus Strongylocentrotus in the seas of Russia. In: Mooi R. & Telford M. (Eds.), Echinoderms: San Francisco. Balkema, Rotterdam, pp. 563‐566.
Becerra J.X., Venable D.L., Evans P.H. & Bowers W.S. (2001). Interactions between chemical and mechanical defenses in the plant genus Bursera and their implications for herbivores. Amer. Zool. 41: 865‐876.
Bégin C. (2002). Influence de la couverture algale sur les communautés dʹinvertébrés benthiques dans le nord du Golfe du St‐Laurent. M.Sc. Thesis, Université Laval, Québec.
Begon M., Harper J.L. & Townsend C.R. (1996). Ecology. Blackwell Science, Oxford. Benedetti‐Cecchi L. (2000). Predicting direct and indirect interactions during
succession in a mid‐littoral rocky shore assemblage. Ecol. Monogr. 70: 45‐72. Bergeron P. & Bourget E. (1984). Effet du froid et des glaces sur les peuplements
intertidaux des régions nordiques particulièrement dans lʹestuaire du Saint‐Laurent. Oceanis 10: 279‐304.
Bernstein B.B., Williams B.E. & Mann K.H. (1981). The role of behavioral responses to predators in modifying urchinsʹ (Strongylocentrotus droebachiensis) destructive grazing and seasonal foraging patterns. Mar. Biol. 63: 39‐49.
Bernstein B.B., Schroeter S.C. & Mann K.H. (1983). Sea urchin (Strongylocentrotus droebachiensis) aggregating behavior investigated by a subtidal multifactorial experiment. Can. J. Fish. Aquat. Sci. 40: 1975‐1986.
Bertness M.D. & Callaway R. (1994). Positive interactions in communities. Trends Ecol. Evol. 9: 191‐193.
Bologna P.A.X. & Steneck R.S. (1993). Kelp beds as habitats for American lobster Homarus americanus. Mar. Ecol. Prog. Ser. 100: 127‐134.

128
Bolser R.C. & Hay M.E. (1996). Are tropical plants better defended? Palatability and defenses of temperate vs. tropical seaweeds. Ecology 77: 2269‐2286.
Bourget E., Archambault D. & Bergeron P. (1985). Effet des propriétés hivernales sur les peuplements épibenthiques intertidaux dans un milieu subarctique, lʹestuaire du Saint‐Laurent. Nat. Can. 112: 131‐142.
Branch G.M. (1975). Mechanisms reducing intraspecific competition in Patella spp.: migration, differentiation and territorial behaviour. J. Anim. Ecol. 44: 575‐600.
Brawley S.H. (1992). Mesoherbivores. In: John D.M., Hawkins S.J. & Price J.H. (Eds.), Plant‐animal interactions in the marine benthos. Oxford University Press, Oxford, pp. 235‐263.
Breen P.A. & Mann K.H. (1976a). Changing lobster abundance and the destruction of kelp beds by sea urchins. Mar. Biol. 34: 137‐142.
Breen P.A. & Mann K.H. (1976b). Destructive grazing of kelp by sea urchins in eastern Canada. J. Fish. Res. Bd. Can. 33: 1278‐1283.
Briscoe C.S. & Sebens K.P. (1988). Omnivory in Strongylocentrotus droebachiensis (Müller) (Echinodermata: Echinoidea): predation on subtidal mussels. J. Exp. Mar. Biol. Ecol. 115: 1‐24.
Brown B.J. & Ewel J.J. (1987). Herbivory in complex and simple tropical successional ecosystems. Ecology 68: 108‐116.
Brown V.K. & Gange A.C. (1992). Secondary plant succession: how is it modified by insect herbivory? Vegetatio 101: 3‐13.
Bruno J.F. & Bertness M.D. (2001). Habitat modification and facilitation in benthic marine communities. In: Bertness M.D., Gaines S.D. & Hay M.E. (Eds.), Marine community ecology. Sinauer Associates, Sunderland, pp. 201‐218.
Bulleri F., Benedetti‐Cecchi L. & Cinelli F. (1999). Grazing by the sea urchins Arbacia lixula L. and Paracentrotus lividus Lam. in the Northwest Mediterranean. J. Exp. Mar. Biol. Ecol. 241: 81‐95.
Callaway R.M. (1995). Positive interactions among plants. Bot. Rev. 61: 306‐349. Callaway R.M., Kikvidze Z. & Kikodze D. (2000). Facilitation by unpalatable weeds
may conserve plant diversity in overgrazed meadows in the Caucasus Mountains. Oikos 89: 275‐282.
Carpenter R.C. (1986). Partitioning herbivory and its effects on coral reef algal communities. Ecol. Monogr. 56: 345‐363.
Castilla J.C. & Crisp D.J. (1970). Responses of Asterias rubens to olfactory stimuli. J. Mar. Biol. Assoc. UK. 50: 829‐847.
Cetrulo G.L. & Hay M.E. (2000). Activated chemical defenses in tropical versus temperate seaweeds. Mar. Ecol. Prog. Ser. 207: 243‐253.
Chapman A.R.O. (1981). Stability of sea urchin dominated barren grounds following destructive grazing of kelp in St. Margaretʹs Bay, eastern Canada. Mar. Biol. 62: 307‐311.

129
Chapman A.R.O. & Johnson C.R. (1990). Disturbance and organization of macroalgal assemblages in the Northwest Atlantic. Hydrobiologia 192: 77‐121.
Chapman M.G. & Underwood A.J. (1992). Foraging behaviour of marine benthic grazers. In: John D.M., Hawkins S.J. & Price J.H. (Eds.), Plant‐animal interactions in the marine benthos. Oxford University Press, New York, pp. 289‐317.
Charnov E.L. (1976). Optimal foraging, the marginal value theorem. Theor. Popul. Biol. 9: 129‐136.
Coley P.D. & Barone J.A. (1996). Herbivory and plant defenses in tropical forests. Annu. Rev. Ecol. Syst. 27: 305‐335.
Connell J.H. (1961). The influence of interspecific competition and other factors on the distribution of the barnacle Chthamalus stellatus. Ecology 42: 710‐723.
Connell J.H. & Slatyer R.O. (1977). Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 111: 1119‐1144.
Conover W.J. (1980). Practical Nonparametric Statistics. John Wiley & Sons, New York.
Cowen R.K., Agegian C.R. & Foster M.S. (1982). The maintenance of community structure in a central California giant kelp forest. J. Exp. Mar. Biol. Ecol. 64: 189‐201.
Crowder M.J. & Hand D.J. (1990). Analysis of repeated measures. Chapman & Hall, London.
Davidson D.W. (1993). The effects of herbivory and granivory on terrestrial plant succession. Oikos 68: 23‐35.
Dayton P.K. (1985a). The structure and regulation of some South American kelp communities. Ecol. Monogr. 55: 447‐468.
Dayton P.K. (1985b). Ecology of kelp communities. Annu. Rev. Ecol. Syst. 16: 215‐245. Dayton P.K., Tegner M.J., Parnell P.E. & Edwards P.B. (1992). Temporal and spatial
patterns of disturbance and recovery in a kelp forest community. Ecol. Monogr. 62: 421‐445.
Dayton P.K., Currie V., Gerrodette T., Keller B.D., Rosenthal R. & Ven Tresca D. (1984). Patch dynamics and stability of some California kelp communities. Ecol. Monogr. 54: 253‐289.
Dean T.A. & Jacobsen F.R. (1984). Growth of juvenile Macrocystis pyrifera (Laminariales) in relation to environmental factors. Mar. Biol. 83: 301‐311.
Dean T.A., Schroeter S.C. & Dixon J.D. (1984). Effects of grazing by two species of sea urchins (Strongylocentrotus franciscanus and Lytechinus anamesus) on recruitment and survival of two species of kelp (Macrocystis pyrifera and Pterygophora californica). Mar. Biol. 78: 301‐313.

130
Draper N.R. & Smith H. (1981). Applied regression analysis. John Wiley & Sons, New York.
Duffy J.E. & Hay M.E. (1990). Seaweed adaptations to herbivory. BioScience 40: 368‐375.
Duffy J.E. & Hay M.E. (2000). Strong impacts of grazing amphipods on the organization of a benthic community. Ecol. Monogr. 70: 237‐263.
Duffy J.E. & Hay M.E. (2001). The ecology and evolution of marine consumer‐prey interactions. In: Bertness M.D., Gaines S.D. & Hay M.E. (Eds.), Marine community ecology. Sinauer Associates, Sunderland, pp. 131‐157.
Duggins D.O. (1980). Kelp beds and sea otters: an experimental approach. Ecology 61: 447‐453.
Duggins D.O. (1981). Sea urchins and kelp: the effects of short term changes in urchin diet. Limnol. Oceanogr. 26: 391‐394.
Duggins D.O., Eckman J.E. & Sewell A.T. (1990). Ecology of understory kelp environments. II. Effects of kelps on recruitment of benthic invertebrates. J. Exp. Mar. Biol. Ecol. 143: 27‐45.
Ebeling A.W., Laur D.R. & Rowley R.J. (1985). Severe storm disturbances and reversal of community structure in a southern California kelp forest. Mar. Biol. 84: 287‐294.
Eckman J.E., Duggins D.O. & Sewell A.T. (1989). Ecology of understory kelp environments. I. Effects of kelps on flow and particle transport near the bottom. J. Exp. Mar. Biol. Ecol. 129: 173‐187.
Emlen J.M. (1966). The role of time and energy in food preference. Am. Nat. 100: 611‐617.
Eppley R.W. & Bovell C.R. (1958). Sulfuric acid in Desmarestia. Biol. Bull. 115: 101‐106. Estes J.A., Smith N.S. & Palmisano J.F. (1978). Sea otter predation and community
organization in the western Aleutian Islands, Alaska. Ecology 59: 822‐833. Falk‐Petersen I.‐B. & Lonning S. (1983). Reproductive cycles of two closely related sea
urchin species, Strongylocentrotus droebachiensis (O. F. Müller) and Strongylocentrotus pallidus (G. O. Sars). Sarsia 68: 157‐164.
Forseth I.N., Wait D.A. & Casper B.B. (2001). Shading by shrubs in a desert system reduces the physiological and demographic performance of an associated herbaceous perennial. J. Ecol. 89: 670‐680.
Gagnon P., Wagner G. & Himmelman J.H. (2003a). Use of a wave tank to study the effects of water motion and algal movement on the displacement of the sea star Asterias vulgaris towards its prey. Mar. Ecol. Prog. Ser. 258: 125‐132.
Gagnon P., Himmelman J.H. & Johnson L.E. (2003b). Algal colonization in urchin barrens: defense by association during recruitment of the brown alga Agarum cribrosum. J. Exp. Mar. Biol. Ecol. 290: 179‐196.

131
Gagnon P., Himmelman J.H. & Johnson L.E. (in press). Temporal variation in community interfaces: kelp bed boundary dynamics adjacent to persistent urchin barrens. Mar. Biol.
Garnick E. (1978). Behavioral ecology of Strongylocentrotus droebachiensis (Müller) (Echinodermata: Echinoidea). Aggregating behavior and chemotaxis. Oecologia 37: 77‐84.
Gaymer C.F., Himmelman J.H. & Johnson L.E. (2001). Distribution and feeding ecology of the seastars Leptasterias polaris and Asterias vulgaris in the northern Gulf of St. Lawrence, Canada. J. Mar. Biol. Assoc. UK. 81: 827‐843.
Gerard V.A. (1982). In situ water motion and nutrient uptake by the giant kelp Macrocystis pyrifera. Mar. Biol. 69: 51‐54.
Gerard V.A. (1984). The light environment in a giant kelp forest: influence of Macrocystis pyrifera on spatial and temporal variability. Mar. Biol. 84: 189‐195.
Graham L.E. & Wilcox L.W. (2000). Algae. Prentice‐Hall, Upper Saddle River. Gray J.S. (1977). The stability of benthic ecosystems. Helgoländer wiss. Meeresunters.
30: 427‐444. Hagen N.T. (1983). Destructive grazing of kelp beds by sea urchins in Vestfjorden,
northern Norway. Sarsia 68: 177‐190. Hand D.J. & Taylor C.C. (1987). Multivariate analysis of variance and repeated
measures. Chapman & Hall, London. Harrold C. & Reed D.C. (1985). Food availability, sea urchin grazing, and kelp forest
community structure. Ecology 66: 1160‐1169. Hay M.E. (1986). Associational plant defenses and the maintenance of species
diversity: turning competitors into accomplices. Am. Nat. 128: 617‐641. Hay M.E. (1996). Marine chemical ecology: whatʹs known and whatʹs next? J. Exp.
Mar. Biol. Ecol. 200: 103‐134. Hay M.E. & Fenical W. (1988). Marine plant‐herbivore interactions: the ecology of
chemical defense. Annu. Rev. Ecol. Syst. 19: 111‐145. Hay M.E. & Fenical W. (1992). Chemical mediation of seaweed‐herbivore interactions.
In: John D.M., Hawkins S.J. & Price J.H. (Eds.), Plant‐animal interactions in the marine benthos. Oxford University Press, New York, pp. 319‐337.
Hay M.E., Kappel Q.E. & Fenical W. (1994). Synergisms in plant defenses against herbivores: interactions of chemistry, calcification, and plant quality. Ecology 75: 1714‐1726.
Hendrix S.D., Brown V.K. & Gange A.C. (1988). Effects of insect herbivory on early plant succession: comparison of an English site and an American site. Biol. J. Linn. Soc. 35: 205‐216.
Himmelman J.H. (1969). Some aspects of the ecology of Strongylocentrotus droebachiensis in eastern Newfoundland. M.Sc. Thesis, Memorial University of Newfoundland, St. Johnʹs.

132
Himmelman J.H. (1975). Phytoplankton as a stimulus for spawning in three marine invertebrates. J. Exp. Mar. Biol. Ecol. 20: 199‐214.
Himmelman J.H. (1984). Urchin feeding and macroalgal distribution in Newfoundland, eastern Canada. Nat. Can. 111: 337‐348.
Himmelman J.H. (1986). Population biology of green sea urchins on rocky barrens. Mar. Ecol. Prog. Ser. 33: 295‐306.
Himmelman J.H. & Steele D.H. (1971). Foods and predators of the green sea urchin Strongylocentrotus droebachiensis in Newfoundland waters. Mar. Biol. 9: 315‐322.
Himmelman J.H. & Lavergne Y. (1985). Organization of rocky subtidal communities in the St. Lawrence Estuary. Nat. Can. 112: 143‐154.
Himmelman J.H. & Nédélec H. (1990). Urchin foraging and algal survival strategies in intensely grazed communities in eastern Canada. Can. J. Fish. Aquat. Sci. 47: 1011‐1026.
Himmelman J.H., Cardinal A. & Bourget E. (1983). Community development following removal of urchins, Strongylocentrotus droebachiensis, from the rocky subtidal zone of the St. Lawrence Estuary, eastern Canada. Oecologia 59: 27‐39.
Holling C.S. (1973). Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 4: 1‐23.
Hooper R. (1981). Recovery of Newfoundland benthic marine communities from sea ice. Proc. Int. Seaweed Symp. 8: 360‐366.
Hurd C.L., Stevens C.L., Laval B.E., Lawrence G.A. & Harrison P.J. (1997). Visualization of seawater flow around morphologically distinct forms of the giant kelp Macrocystis integrifolia from wave‐sheltered and exposed sites. Limnol. Oceanogr. 42: 156‐163.
Jensen M. (1974). The Strongylocentrotidae (Echinoidea), a morphologic and and systematic study. Sarsia 57: 113‐148.
Johnson C.R. & Mann K.H., 1982. Adaptations of Strongylocentrotus droebachiensis for survival on barren grounds in Nova Scotia. In: Lawrence J.M. (Ed.), International Echinoderms Conference. Balkema, A. A., Tampa Bay, pp. 277‐283.
Johnson C.R. & Mann K.H. (1988). Diversity, patterns of adaptation, and stability of Nova Scotian kelp beds. Ecol. Monogr. 58: 129‐154.
Jones N.S. & Kain J.M. (1967). Subtidal algal colonization following the removal of Echinus. Helgoländer wiss. Meeresunters. 15: 460‐466.
Karban R. & Myers J.H. (1989). Induced plant responses to herbivory. Annu. Rev. Ecol. Syst. 20: 331‐348.
Kawamata S. (1998). Effect of wave‐induced oscillatory flow on grazing by a subtidal sea urchin Strongylocentrotus nudus (A. Agassiz). J. Exp. Mar. Biol. Ecol. 224: 31‐48.

133
Kawamata S. (2001). Adaptive mechanical tolerance and dislodgement velocity of the kelp Laminaria japonica in wave‐induced water motion. Mar. Ecol. Prog. Ser. 211: 89‐104.
Keats D.W., South G.R. & Steele D.H. (1982). The occurence of Agarum cribrosum (Mert.) Bory (Phaeophyta, Laminariales) in relation to some of its competitors and predators in Newfoundland. Phycologia 21: 189‐191.
Keats D.W., South G.R. & Steele D.H. (1985). Algal biomass and diversity in the upper subtidal at a pack‐ice disturbed site in eastern Newfoundland. Mar. Ecol. Prog. Ser. 25: 151‐158.
Keats D.W., South G.R. & Steele D.H. (1990). Effects of an experimental reduction in grazing by green sea urchins on a benthic macroalgal community in eastern Newfoundland. Mar. Ecol. Prog. Ser. 68: 181‐193.
Kerr J.N.Q. & Paul V.J. (1995). Animal‐plant defense association: the soft coral Sinularia sp. (Cnidaria, Alcyonacea) protects Halimeda spp. from herbivory. J. Exp. Mar. Biol. Ecol. 186: 183‐205.
Konar B. (2000). Seasonal inhibitory effects of marine plants on sea urchins: structuring communities the algal way. Oecologia 125: 208‐217.
Krebs C.J. (2001). Ecology. Benjamin Cummings, San Francisco. Larson B.R., Vadas R.L. & Keser M. (1980). Feeding and nutritional ecology of the sea
urchin Strongylocentrotus drobachiensis in Maine, USA. Mar. Biol. 59: 49‐62. Lawrence J.M. (1975). On the relationships between marine plants and sea urchins.
Oceanogr. Mar. Biol. Ann. Rev. 13: 213‐286. Legendre P. & Legendre L. (1998). Numerical ecology. Elsevier, Amsterdam. Leinaas H.P. & Christie H. (1996). Effects of removing sea urchins (Strongylocentrotus
droebachiensis): stability of the barren state and succession of kelp forest recovery in the east Atlantic. Oecologia 105: 524‐536.
Lettenmaier D.P. (1976). Detection of trends in water quality data from records with dependent observations. Water Resour. Res. 12: 1037‐1046.
Lewis S.M. (1986). The role of herbivorous fishes in the organization of a Caribbean reef community. Ecol. Monogr. 56: 183‐200.
Lewontin R.C. (1969). The meaning of stability. Brookhaven Symp. Biol. 22: 13‐24. Lindroth R.L. (1989). Mammalian herbivore‐plant interactions. In: Abrahamson W.G.
(Ed.), Plant‐animal interactions. McGraw‐Hill, New York, pp. 163‐206. Lissner A.L. (1983). Relationship of water motion to the shallow water distribution
and morphology of two species of sea urchins. J. Mar. Res. 41: 691‐709. Littler M.M., Littler D.S. & Taylor P.R. (1987). Animal‐plant defense associations:
effects on the distribution and abundance of tropical reef macrophytes. J. Exp. Mar. Biol. Ecol. 105: 107‐121.
Lobban C.S. & Harrison P.J. (1994). Seaweed ecology and physiology. Cambridge University Press, Cambridge.

134
Lubchenco J. & Gaines S.D. (1981). A unified approach to marine plant‐herbivore interactions. I. Populations and communities. Annu. Rev. Ecol. Syst. 12: 405‐437.
Lucas P.W., Turner I.M., Dominy N.J. & Yamashita N. (2000). Mechanical defences to herbivory. Ann. Bot.‐London. 86: 913‐920.
MacArthur R.H. & Pianka E.R. (1966). On optimal use of a patchy environment. Am. Nat. 100: 603‐609.
Mann K.H. (1973). Seaweeds: their productivity and strategy for growth. Science 182: 975‐981.
Mann K.H. (1977). Destruction of kelp‐beds by sea‐urchins: A cyclical phenomenon or irreversible degradation? Helgol. wiss. Meeresunters. 30: 455‐467.
Mann K.H., Wright J.L.C., Welsford B.E. & Hatfield E. (1984). Responses of the sea urchin Strongylocentrotus droebachiensis (O. F. Müller) to water‐borne stimuli from potential predators and potential food algae. J. Exp. Mar. Biol. Ecol. 79: 233‐244.
McAuliffe J.R. (1986). Herbivore‐limited establishment of a Sonoran Desert tree, Cercidium microphyllum. Ecology 67: 276‐280.
McClintock M., Higinbotham N., Uribe E.G. & Cleland R.E. (1982). Active, irreversible accumulation of extreme levels of H2SO4 in the brown alga, Desmarestia. Plant. Physiol. 70: 771‐774.
McCullagh P. & Nelder J.A. (1989). Generalized linear models. Chapman & Hall, New York.
McNaughton S.J. (1983). Compensatory plant growth as a response to herbivory. Oikos 40: 329‐336.
Meidel S.K. & Scheibling R.E. (1998). Annual reproductive cycle of the green sea urchin, Strongylocentrotus droebachiensis, in differing habitats in Nova Scotia, Canada. Mar. Biol. 131: 461‐478.
Meidel S.K. & Scheibling R.E. (1999). Effects of food type and ration on reproductive maturation and growth of the sea urchin Strongylocentrotus droebachiensis. Mar. Biol. 134: 155‐166.
Menge B.A. (1972). Foraging strategy of a starfish in relation to actual prey availability and environmental predictability. Ecol. Monogr. 42: 25‐50.
Menge B.A. (1976). Organization of the New England rocky intertidal community: role of predation, competition, and environmental heterogeneity. Ecol. Monogr. 46: 355‐391.
Menge B.A., Berlow E.L., Blanchette C.A., Navarrete S.A. & Yamada S.B. (1994). The keystone species concept: variation in interaction strength in a rocky intertidal habitat. Ecol. Monogr. 64: 249‐286.
Miller R.J. (1985). Succession in sea urchin and seaweed abundance in Nova Scotia, Canada. Mar. Biol. 84: 275‐286.

135
Miller R.J. & Colodey A.G. (1983). Widespread mass mortalities of the green sea urchin in Nova Scotia, Canada. Mar. Biol. 73: 263‐267.
Minor M.A. & Scheibling R.E. (1997). Effects of food ration and feeding regime on growth and reproduction of the sea urchin Strongylocentrotus droebachiensis. Mar. Biol. 129: 159‐167.
Montgomery D.C. (1991). Design and analysis of experiments. John Wiley & Sons, New York.
Morrison D. (1988). Comparing fish and urchin grazing in shallow and deeper coral reef algal communities. Ecology 69: 1367‐1382.
Mortensen T.H. (1943). A monograph of the Echinoidea. Reitzels, C. A., Copenhagen. Myers J.H. & Bazely D. (1991). Thorns, spines, prickles, and hairs: are they stimulated
by herbivory and do they deter herbivores? In: Tallamy D.W. & Raupp M.J. (Eds.), Phytochemical induction by herbivores. John Wiley, New York, pp. 325‐344.
Nestler E.C. & Harris L.G. (1994). The importance of omnivory in Strongylocentrotus droebachiensis (Muller) in the Gulf of Maine. In: David B., Guille A., Féral J.‐P. & Roux M. (Eds.), Echinoderms through time. Balkema, Rotterdam, pp. 813‐818.
Novaczek I. & McLachlan J. (1989). Investigations of the marine algae of Nova Scotia XVII: vertical and geographic distribution of marine algae on rocky shores of the Maritime Provinces. Proc. N. S. Inst. Sci. 38: 91‐143.
OʹDowd D.J. & Williamson G.B. (1979). Stability conditions in plant defense guilds. Am. Nat. 114: 379‐383.
Odum E.P. (1969). The strategy of ecosystem development. Science 164: 262‐270. Ogden J.C. & Lobel P.S. (1978). The role of herbivorous fishes and urchins in coral
reef communities. Env. Biol. Fish. 3: 49‐63. Paine R.T. (1966). Food web complexity and species diversity. Am. Nat. 100: 65‐75. Paine R.T. (1976). Size‐limited predation: an observational and experimental
approach with the Mytilus‐Pisaster interaction. Ecology 57: 858‐873. Paine R.T. (1990). Benthic macroalgal competition: complications and consequences. J.
Phycol. 26: 12‐17. Paine R.T. & Vadas R.L. (1969). The effects of grazing by sea urchins,
Strongylocentrotus spp., on benthic algal populations. Limnol. Oceanogr. 14: 710‐719.
Paul V.J. & Hay M.E. (1986). Seaweed susceptibility to herbivory: chemical and morphological correlates. Mar. Ecol. Prog. Ser. 33: 255‐264.
Paul V.J. & Van Alstyne K.L. (1992). Activation of chemical defenses in the tropical green alga Halimeda spp. J. Exp. Mar. Biol. Ecol. 160: 191‐203.
Pavia H. & Toth G.B. (2000). Inducible chemical resistance to herbivory in the brown seaweed Ascophyllum nodosum. Ecology 81: 3212‐3225.

136
Pearse J.S. & Hines A.H. (1979). Expansion of a central California kelp forest following the mass mortality of sea urchins. Mar. Biol. 51: 83‐91.
Pelletreau K.N. & Muller‐Parker G. (2002). Sulfuric acid in the phaeophyte alga Desmarestia munda deters feeding by the sea urchin Strongylocentrotus droebachiensis. Mar. Biol. 141: 1‐9.
Peres‐Neto P.R. (1999). How many statistical tests are too many? The problem of conducting multiple ecological inferences revisited. Mar. Ecol. Prog. Ser. 176: 303‐306.
Peters A.F., van Oppen M.J.H., Wiencke C., Stam W.T. & Olsen J.L. (1997). Phylogeny and historical ecology of the Desmarestiaceae (Phaeophyceae) support a southern hemisphere origin. J. Phycol. 33: 294‐309.
Pfister C.A. & Hay M.E. (1988). Associational plant refuges: convergent patterns in marine and terrestrial communities result from differing mechanisms. Oecologia 77: 118‐129.
Prince J.S. & LeBlanc W.G. (1992). Comparative feeding preference of Strongylocentrotus droebachiensis (Echinoidea) for the invasive seaweed Codium fragile ssp. tomentosoides (Chlorophyceae) and four other seaweeds. Mar. Biol. 113: 159‐163.
Randall J.E. (1961). Overgrazing of algae by herbivorous marine fishes. Ecology 42: 812.
Raymond B.G. & Scheibling R.E. (1987). Recruitment and growth of the sea urchin Strongylocentrotus droebachiensis (Muller) following mass mortalities off Nova Scotia, Canada. J. Exp. Mar. Biol. Ecol. 108: 31‐54.
Reed D.C. (1990). The effects of variable settlement and early competition on patterns of kelp recruitment. Ecology 71: 776‐787.
Reed D.C. & Foster M.S. (1984). The effects of canopy shading on algal recruitment and growth in a giant kelp forest. Ecology 65: 937‐948.
Rice W.R. (1989). Analyzing tables of statistical tests. Evolution. 43: 223‐225. Rochette R., Hamel J.‐F. & Himmelman J.H. (1994). Foraging strategy of the asteroid
Leptasterias polaris: role of prey odors, current and feeding status. Mar. Ecol. Prog. Ser. 106: 93‐100.
Rose C.D., Sharp W.C., Kenworthy W.J., Hunt J.H., Lyons W.G., Prager E.J., Valentine J.F., Hall M.O., Whitfield P.E. & Fourqurean J.W. (1999). Overgrazing of a large seagrass bed by the sea urchin Lytechinus variegatus in Outer Florida Bay. Mar. Ecol. Prog. Ser. 190: 211‐222.
Rosenthal J.P. & Kotanen P.M. (1994). Terrestrial plant tolerance to herbivory. Trends Ecol. Evol. 9: 145‐148.
Ruiz G.M., Carlton J.T., Grosholz E.D. & Hines A.H. (1997). Global invasions of marine and estuarine habitats by non‐indigenous species: mechanisms, extent, and consequences. Amer. Zool. 37: 621‐632.

137
Russell M.P., Ebert T.A. & Petraitis P.S. (1998). Field estimates of growth and mortality of the green sea urchin, Strongylocentrotus droebachiensis. Ophelia 48: 137‐153.
SAS Institute Inc. (1999). SAS/STAT Userʹs Guide, Version 8. SAS Institute Inc, Cary, NC.
Sasaki H., Kataoka H., Kamiya M. & Kawai H. (1999). Accumulation of sulfuric acid in Dictyotales (Phaeophyceae): taxonomic distribution and ion chromatography of cell extracts. J. Phycol. 35: 732‐739.
Schappert P.J. & Shore J.S. (1999). Cyanogenesis, herbivory and plant defense in Turnera ulmifolia on Jamaica. Écoscience 6: 511‐520.
Scheibling R. (1986). Increased macroalgal abundance following mass mortalities of sea urchins (Strongylocentrotus droebachiensis) along the Atlantic coast of Nova Scotia. Oecologia 68: 186‐198.
Scheibling R.E. (1988). Microbial control of sea urchins: Achillesʹ heel or Pandoraʹs box? In: Burke R.D., Mladenov P.V., Lambert P. & Parsley R.L. (Eds.), Echinoderm biology. Balkema, Rotterdam, pp. 745‐754.
Scheibling R.E. (1996). The role of predation in regulating sea urchin populations in eastern Canada. Oceanol. Acta 19: 421‐430.
Scheibling R.E. & Stephenson R.L. (1984). Mass mortality of Strongylocentrotus droebachiensis (Echinodermata: Echinoidea) off Nova Scotia, Canada. Mar. Biol. 78: 153‐164.
Scheibling R.E. & Raymond B.G. (1990). Community dynamics on a subtidal cobble bed following mass mortalities of sea urchins. Mar. Ecol. Prog. Ser. 63: 127‐145.
Scheibling R.E. & Hamm J. (1991). Interactions between sea urchin (Strongylocentrotus droebachiensis) and their predators in field and laboratory experiments. Mar. Biol. 110: 105‐116.
Scheibling R.E. & Hennigar A.W. (1997). Recurrent outbreaks of disease in sea urchins Strongylocentrotus droebachiensis in Nova Scotia: evidence for a link with large‐scale meteorologic and oceanographic events. Mar. Ecol. Prog. Ser. 152: 155‐165.
Scheibling R.E. & Hatcher B.G. (2001). The ecology of Strongylocentrotus droebachiensis. In: Lawrence J.M. (Ed.), Edible sea urchins: biology and ecology. Elsevier, Amsterdam, pp. 271‐306.
Scheibling R.E., Hennigar A.W. & Balch T. (1999). Destructive grazing, epiphytism, and disease: the dynamics of sea urchin‐kelp interactions in Nova Scotia. Can. J. Fish. Aquat. Sci. 56: 2300‐2314.
Schmitz O.J., Hambäck P.A. & Beckerman A.P. (2000). Trophic cascades in terrestrial systems: a review of the effects of carnivore removals on plants. Am. Nat. 155: 141‐153.

138
Schnitzler I., Pohnert G., Hay M. & Boland W. (2001). Chemical defense of brown algae (Dictyopteris spp.) against the herbivorous amphipod Ampithoe longimana. Oecologia 126: 515‐521.
Schoener T.W. (1974). Resource partitioning in ecological communities. Science 185: 27‐39.
Sebens K.P. (1985). Community ecology of vertical rock walls in the Gulf of Maine, U.S.A.: small‐scale processes and alternative community states. In: Moore P.G. & Seed R. (Eds.), The ecology of rocky coasts. Hodder and Stoughton, London, pp. 346‐371.
Seymour R.J., Tegner M.J., Dayton P.K. & Parnell P.E. (1989). Storm wave induced mortality of giant kelp, Macrocystis pyrifera, in Southern California. Estuar. Coast. Shelf S. 28: 277‐292.
Siddon C.E. & Witman J.D. (2003). Influence of chronic, low‐level hydrodynamic forces on subtidal community structure. Mar. Ecol. Prog. Ser. 261: 99‐110.
Sih A., Crowley P., McPeek M., Petranka J. & Strohmeier K. (1985). Predation, competition and prey communities: a review of field experiments. Annu. Rev. Ecol. Syst. 16: 269‐311.
Simenstad C.A., Estes J.A. & Kenyon K.W. (1978). Aleuts, sea otters, and alternate stable‐state communities. Science 200: 403‐411.
Sivertsen K. (1997). Geographic and environmental factors affecting the distribution of kelp beds and barren grounds and changes in biota associated with kelp reduction at sites along Norwegian coast. Can. J. Fish. Aquat. Sci. 54: 2872‐2887.
Sloan N.A. & Campbell A.C. (1982). Perception of food. In: Jangoux M. & Lawrence J.M. (Eds.), Echinoderm nutrition. Balkema, A. A., Rotterdam, pp. 3‐23.
Snedecor G.W. & Cochran W.G. (1989). Statistical methods. Iowa State University Press, Ames.
Sousa W.P. (1984). The role of disturbance in natural communities. Annu. Rev. Ecol. Syst. 15: 353‐391.
Sousa W.P. & Connell J.H. (1992). Grazing and succession in marine algae. In: John D.M., Hawkins S.J. & Price J.H. (Eds.), Plant‐animal interactions in the marine benthos. Oxford University Press, New York, pp. 425‐441.
Starr M., Himmelman J.H. & Therriault J.‐C. (1990). Direct coupling of marine invertebrate spawning with phytoplankton blooms. Science 247: 1071‐1074.
Steneck R.S. (1983). Escalating herbivory and resulting adaptive trends in calcareous algal crusts. Paleobiology 9: 44‐61.
Steneck R.S. (1985). Adaptations of crustose coralline algae to herbivory: patterns in space and time. In: Toomey D.F. & Nitecki M.H. (Eds.), Paleoalgology: contemporary research and applications. Springer‐Verlag, Heidelberg, pp. 352‐366.

139
Steneck R.S. (1986). The ecology of coralline algal crusts: convergent patterns and adaptative strategies. Annu. Rev. Ecol. Syst. 17: 273‐303.
Stoecker D. (1978). Resistance of a tunicate to fouling. Biol. Bull. 155: 615‐626. Sutherland J.P. (1974). Multiple stable points in natural communities. Am. Nat. 108:
859‐873. Swan E.F. (1961). Some observations on the growth rate of sea urchins in the genus
Strongylocentrotus. Biol. Bull. 120: 420‐427. Tahvanainen J.O. & Root R.B. (1972). The influence of vegetational diversity on the
population ecology of a specialized herbivore, Phyllotreta cruciferae (Coleoptera:Chrysomelidae). Oecologia 10: 321‐346.
Taylor R.B., Sotka E. & Hay M.E. (2002). Tissue‐specific induction of herbivore resistance: seaweed response to amphipod grazing. Oecologia 132: 68‐76.
Tegner M.J. & Dayton P.K. (2000). Ecosystem effects of fishing in kelp forest communities. ICES J. Mar. Sci. 57: 579‐589.
Thompson R.J. (1984). Partitioning of energy between growth and reproduction in three populations of the green sea urchin Strongylocentrotus droebachiensis. Adv. Invertebr. Reprod. 3: 425‐432.
Thompson T.E. (1988). Acidic allomones in marine organisms. J. Mar. Biol. Assoc. UK. 68: 499‐517.
Tiffin P. (2000). Mechanisms of tolerance to herbivore damage: what do we know? Evol. Ecol. 14: 523‐536.
Tremblay C. & Chapman A.R.O. (1980). The local occurrence of Agarum cribrosum in relation to the presence or absence of its competitors and predators. Proc. N. S. Inst. Sci. 30: 165‐170.
Trowbridge C.D. (1995). Establishment of the green alga Codium fragile ssp. tomentosoides on New Zealand rocky shores: current distribution and invertebrate grazers. J. Ecol. 83: 949‐965.
Underwood A.J. (1999). Physical disturbances and their direct effect on an indirect effect: responses of an intertidal assemblage to a severe storm. J. Exp. Mar. Biol. Ecol. 232: 125‐140.
Underwood A.J. & Jernakoff P. (1981). Effects of interactions between algae and grazing gastropods on the structure of a low‐shore intertidal algal community. Oecologia 48: 221‐233.
Vadas R.L. (1968). The ecology of Agarum and the kelp bed community. Ph.D. Thesis, University of Washington, Seattle.
Vadas R.L. (1977). Preferential feeding: an optimization strategy in sea urchins. Ecol. Monogr. 47: 337‐371.
Vadas R.L. (1990). Comparative foraging behavior of tropical and boreal sea urchins. In: Hughes R.N. (Ed.), Behavioural mechanisms of food selection. Springer‐Verlag, Berlin, pp. 479‐514.

140
Vadas R.L. & Elner R.W. (1992). Plant‐animal interactions in the north‐west Atlantic. In: John D.M., Hawkins S.J. & Price J.H. (Eds.), Plant‐animal interactions in the marine benthos. Oxford University Press, Oxford, pp. 33‐60.
Vadas R.L., Wright W.A. & Miller S.L. (1990). Recruitment of Ascophyllum nodosum: wave action as a source of mortality. Mar. Ecol. Prog. Ser. 61: 263‐272.
Vadas R.L., Elner R.W., Garwood P.E. & Babb I.G. (1986). Experimental evaluation of aggregation behavior in the sea urchin Strongylocentrotus droebachiensis. A reinterpretation. Mar. Biol. 90: 433‐448.
Van Alstyne K.L. (1988). Herbivore grazing increases polyphenolic defenses in the intertidal brown alga Fucus distichus. Ecology 69: 655‐663.
Van Alstyne K.L., Ehlig J.M. & Whitman S.L. (1999). Feeding preferences for juvenile and adult algae depend on algal stage and herbivore species. Mar. Ecol. Prog. Ser. 180: 179‐185.
Vasquez J.A. (1993). Effects on the animal community of dislodgment of holdfasts of Macrocystis pyrifera. Pac. Science 47: 180‐184.
Vasquez J.A. & Buschmann A.H. (1997). Herbivore‐kelp interactions in Chilean subtidal communities: a review. Rev. Chil. Hist. Nat. 70: 41‐52.
Vasquez J.A. & McPeak R.H. (1998). A new tool for kelp restoration. Calif. Fish Game 84: 149‐158.
Velimirov B. & Griffiths C.L. (1979). Wave‐induced kelp movement and its importance for community structure. Bot. Mar. 22: 169‐172.
Walker L.R., Zasada J.C. & Chapin III F.S. (1986). The role of life history processes in primary succession on an Alaskan floodplain. Ecology 67: 1243‐1253.
Weis A.E. & Berenbaum M.R. (1989). Herbivorous insects and green plants. In: Abrahamson W.G. (Ed.), Plant‐animal interactions. McGraw‐Hill, New York, pp. 123‐162.
White P.S. (1979). Pattern, process, and natural disturbance in vegetation. Bot. Rev. 45: 229‐299.
Witman J.D. (1985). Refuges, biological disturbance, and rocky subtidal community structure in New England. Ecol. Monogr. 55: 421‐445.
Witman J.D. (1987). Subtidal coexistence: storms, grazing, mutualism and the zonation of kelps and mussels. Ecol. Monogr. 57: 167‐187.
Witman J.D. & Dayton P.K. (2001). Rocky subtidal communities. In: Bertness M.D., Gaines S.D. & Hay M.E. (Eds.), Marine community ecology. Sinauer Associates, Sunderland, pp. 339‐366.
Wood D.M. & del Moral R. (1987). Mechanisms of early primary succession in subalpine habitats on Mount St. Helens. Ecology 68: 780‐790.
Zamora R., Hodar J.A. & Gomez J.M. (1999). Plant‐herbivore interaction: beyond a binary vision. In: Pugnaire F.I. & Valladares F. (Eds.), Handbook of functional plant ecology. Marcel Dekker, Inc., New York, pp. 677‐718.

141
Zar J.H. (1999). Biostatistical analysis. Prentice‐Hall, New Jersey.