Embed Size (px)
Citation preview

BIOCHIMIE, 1973, 55, 189-194.
Etude de l'effectif ribosomique du foie chez la souris normale et chez la souris naine.
Jacque l ine MELLET ( ' ) . Service de Phgsiologie cellulaire, Institut de Biologie mol~culaire
Universit~ Paris VII, 2 place Jussieu, 75005 Paris. et Laboratoire d'Endocrinologie, Facultd des Sciences Pharmaceutiques
et Biologiques, Paris- Luxembourg. (21-11-1972).
Summary. - - Secretion rates of TSH and STH of adenohypophysis in genetically dwarf mice are found to be very low. Many metabolic troubles are due to this deficiency, thus we observed a decrease of protein synthesis in ribosomes isolated from the liver.
The present publication deals with the polysomal profil from d~varf miee's livers before and after treatment with STH and T3. There is a loveer proportion polysomes/ monosomes in dwarf mice than in normal mice. In dvearf mice T3 treatement does not change this state. On the other hand STH injection restores polysome/monosome pro- portions to normal state.
INTRODUCTION.
Les effets phys io logiques et b iochimiques des iodothyronines et de l ' ho rmone somatot rope ont 6t6 en g~n6ral observ6s chez des animaux priv6s de s6er6tion hormona le par ablation ch i rurg ica le de la thyroSde on de l 'hypophyse , puts soumis l ' in ject ion de Pun ou de l 'autre type d 'hormone. On a 6tabli ehez le rat op6r6, puts traitS, que ces hormones sont h l 'o r ig ine d 'une s t imulat ion de la biosynth~se des prot6ines et des R~A [1-6], [7-10]. Les modali t6s des effets act ivateurs, qui ont d~jh fait l 'objet de nombreux t ravaux, ne sont pas encore ent i6rement 81ucid6es. Apr6s hypophysec- tomie ou thyro idec tomie , on observe d 'une par t une d iminu t ion d 'act ivi t6 de la f rac t ion <( enzyme pH 5 >> cytoplasmique, laquelle cont ient les tRNA, les enzymes de charge des tRNA et cer ta ins fac- teurs prot6iques qui in te rv iennen t dans la tra- duct ion - - et d 'aut re par t une r6duct ion dans l 'apt i tude qu 'ont les r ibosomes isol6s h synth6tiser des prot6ines. L ' in jec t ion d 'hormone thy ro id ienne r6tablit complbtement l 'act ivi t6 <( E pH 5 ~ et par- t ie l lement celle des polyr ibosomes [4].
Une d iminut ion du nombre de r ibosomes est observ6e aprbs ablation de l 'nne ou l 'aut re des deux glandes endocr ines [11, 1~]. On constate apr6s in ject ion de somato t rophine une augmenta- t ion de la vitesse d ' incorpora t ion de l ' ac ide oro- t ique marqu6 dans le RNA r ibosomique [11] ou dans ses pr6curseurs [13, 14]. De plus, la st imula- t ion de l 'act ivi t6 de la RNA polym6rase, celle no tamment qui est d6pendante du Mg ~÷, est mise en
(*) Adresse actuelle : Laboratoire de Biochimie g6n~- rule et compar~e, Coll~ge de France, 75005 Paris.
6vidence sur les noyaux isol6s apr6s in ject ion de chacune des deux hormones [11, 12]. Plus r6cem- ment, les RNA polym6rases nucl6aires A, B et C ont 6t6 isol6es du foie d ' an imaux hypophysec to- mis6s et leurs activit6s sp6cifiques test6es [15]. Apr6s inject ion d 'ho rmone somatolrope, on observe p r inc ipa lement une augmentat ion de la teneur en RNA polym6rase A nucl6olaire, tandis que la 3,5,3 ' - t r i iodo-L-thyronine agit fi la fois en augmentant la teneur en RNA polym6rase et en s t imulant l 'activit6 sp6cifique de cette enzyme.
I1 nous a sembl6 que l '6tude du profil polyso- mique pr6sentai t un cer ta in int6r6t dans la mesure off il permet d ' app roehe r l '6qui l ibre qui existe entre tous les facteurs qui in t e rv iennen t au niveau de la t raduc t ion lots de la synth~se des prot6ines.
D'apr6s les t ravaux r6alis6s ant6r ieurement , il semble que l ' hypophysec tomie modifie le profil polysonfique [12] et que la thyro ' idectomie soit sans effet. Tontefois ees r6sultats sont souvent d ' in terpr~ta t ion d61icate car il appara i t difficile de rappor te r h une senle ho rmone les effets que l 'on peut observer sur des an imaux op6r6s ou trait6s. En effet, par suite des r~gulations cons6- cutives h l 'ablat ion de la thyro'ide ou de l 'hypo- physe, des modif icat ions dans les taux de nom- breuses s6cr6tions endocr ines se manifestent . Ainsi, chez l ' an imal thyr6opr ive , on eonstate une d iminut ion de la somato t raphine [16, 18], alors que l ' hypophysec tomie re tent i t sur l ' ensemble des m~- tabolismes : non seulement les diverses st imulines an t6hypophysa i res sont supprim6es, mats ]es hor- mones de la pos thypophyse font 6galement d~faut.
Nous avons donc pens6 que la souris g6n6tique- ment naine DW ÷÷ [19] const i tuai t sans doute un

190 J. Me l l e t .
meil leur mat6riel d '6tude puisque, chez cet animal, les s6cr6tions d 'hormone de croissance et d 'hor- mone thyr6otrope sont p ra t iquement inexis tantes [20, 261, tandis que d 'une par t les autres s t imulines ne sont que par t ie l lement r6duites I20, 22] et que, d 'autre part, la neurohypophyse est fonctionnelle.
Nous avons entrepr is des recherches sur la bio- synthbse des prot6ines par les fract ions subcellu- laires isol6es de ces animaux : la vitesse d ' incor- porat ion de leucine marqu6e est ralent ie et le taux max imum est diminu6.
Le pr6sent t ravai l concerne l '6tude comparat ive de la r6par t i t ion des r ibosomes en polysomes, s tructures repr6sentat ives des r ibosomes engag6s sur les RNA messagers. I1 est apparu que si l'effec- tif global en RNA r ibosomique cytoplasmique sem- ble peu affect6 par l '6tat de d6ficience hormonale , par contre la d is t r ibut ion des r ibosomes en unit6s polysomiques, analys6s en gradient con t inu de saccharose, est profond6ment modifi6e. De plus, le t ra i tement des souris naines par l ' in jec t ion quo- t id ienne de l 'hormone de croissance ovine r6tablit in t6gralement le profil polysomique.
I. - - MATI~RIEL ET MI~THODES.
A) Choix et t ra i tement des animaux.
Les exp6riences comparat ives entre les deux lots de souris naines et normales sont effectu6es avec des souris de m6me ~ge, celui-ci va r ian t de 2 h 3 roots.
Un nombre 6gal de souris de chaque sexe 6tait utilis6 dans nos exp6riences de fa~on h teni r eompte d'6ventuelles var ia t ions li6es au caract6re sexuel.
La quanti t6 d 'hormone inject6e par vote sous- cutan6e pour le t ra i tement des souris naines est calcu16e en se r6f6raut au poids de 1'animal : 1'hor- mone de croissance est adminis t r6e h raison de 300 i~g/100 g de poids. Un lot t6moin recoit des inject ions sous-cutan6es de s6rum physiologique seul.
B) Milieux.
Le tampon utilis6 pour la pr6para t ion des homo- g6nats et pour la pr6para t ion des gradients conti- nus et d iscont inus est le tampon TKM (Tris 50 mM KC1 25 raM, MgCI~ 6 mM h pH 7.5.) addi t ionn6 de mercapto6thanol 6 mM.
C) Preparation des fract ions subcellulaires.
Preparat ion de l 'homogdnat.
Apr~s avoir tu~ les souris par d~capitation, le foie est pr61ev6, pes6, 6minc6, lay6 plusieurs fois
pour ~l iminer au max inmm les lipides et ]'h6mo- globine, puts homog~n~is6 au Potter avec le piston loose (10 ¢ al ler-retour >>) en pr6sence du tampon, en respectant les propor t ions suivantes : 1 g de foie/1,5 ml de tampon TKM pH 7,5 h 0 °.
Les phases successives de la pr6para t ion de l 'homog~nat ont 6t6 r6alis6es h la temperature de la glace fondante et reprodui tes de facon iden- tique dans le temps pour chacune des souris naines et normales ; h cet effet, "~ ehaque exp6- r ienee 4 foies de souris naines sont utilis6s (en- viron 1,25 g) et homog6n~is6s dans un volume de tampon convenable , 1,9 ml. Un poids 6quivalent de foie de souris normale est homog6n6is6 dans les m6mes condit ions.
hnm6dia tement apr6s l 'homog~n6isation, on effectue les divers f rac t ionnements par ul t racen- t r i fugat ion diff~rentielle.
Frac t ionnement par ul tracentri fugation diff~- rentielle.
La fract ion F I (fractions con~tenant les poly- somes totaux) est constitu6e par le surnageant pos tmi tochondr ia l - - les noyaux et les mitochon- dries sont pr6alablenlent 61linings par deux centri- fugations successives de l 'homog6nat dans une Sorval r6frig~r6e, la premiere centr i fugat ion est effectu6e h 1 000 g (10'), la seconde h 25 000 (109 - - Le surnageant l impide est pr61ev6 par aspirat ion h l 'a ide d 'une seringue st6rile en 6vitant la touche l ip idique de surface et la zone profonde et t rouble qui avoisine le culot.
Frac t ionnement des r ibosomes totaux en deux pools :
Pool des r ibosomes libres = F II
Pool des r ibosomes li~s {t l 'ergastoplasme = F I I I
2 ml de surnageant pos tmi tochondr ia l sont d6- pos~s h la surface du gradient d i scont inu de sac- charose (0,SM h 2 M) dans le tampon TKM. Apr~s centr i fugat ion 24 h h 40 000 t / m (rotor 40) dans une Spinco h 4°C, la eouche superficielle du gra- dient est aspir6e et 61imin6e.
La fract ion F III qui cont ient les r ibosomes li~s aux membranes ergatoplasmiques est h la surface de la couche inf6rieure de saccharose 2 M, elle est pr61ev~e h la pipette Pasteur et holnog6n6is~e.
Dans cer tains cas, F I I I sera d6barrass~e des membranes par t ra i tement au d6soxycholate h 10 p. cent (concentra t ion 2 p. cent finale), puts puri- fi6e par une centr i fugat ion de deux heures "h 200.000 g dans une Spinco h 4 °. Le culot est mis en suspension dans le tampon TKM dans un volume
BIOCHIMIE, 1 9 7 3 , 55, n ° 2.

E f f e c t i f r ibosomique du foie chez la souris naine. 191
de 1,5 ml (les trois quarts du volume de surnageant pos tmi tochondr ia l in i t ia lement d6pos6 sur gra- dient discont inu) .
La fract ion F III qui cont ient les r ibosomes libres est constitude par le culot. Celui-ci lay6 en surface trois lois ~ I 'eau distill6e ",k 0 ° est extrait du fond du tube ~ l 'aide d 'un piston fin en 6vitant tout contact avec les patois du tube, (RNase), puis homog6n6is6 au Potter dans un volmne de tampon TKM de 0,5 ml (volume 6gal au quart du volume de surnageant pos tmi tochondr ia l iu i t ia lement d6pos6 sur gradient continu) .
Nature des fractions subcellulaires.
L'absorpt ion /~ 260 m.~ et 280 m,~ est lue an spec- trophotonl6tre Beckmann pour chacune des frac- t ions F I, F I[, F I I I apr~s di lut ion au 1/50 dans le tampon TKM.
DO 260 m~ On vdrifie le rappor t -- 1,25 h 1,3'2
DO 280 iu,~ pour tons les surnageants pos tmi tochondr iaux 6tu- di6s.
Darts l 'dtude qui suit, la concent ra t ion en RNA de chaque fract ion est exprim6e par concen t ion en UDO h 260 m,~ ou A260.
• Calcul du rendement.
La quanti t6 des ribosomes recueil l is dans cha- cune des fract ions : f ract ion F II : r ibosomes non lids ~ l 'ergastoplasme, fract ion F I I I : repr6sentant les r ibosomes lids $ l 'ergastoplasme, a 6t6 calcu16e d'apr6s les op6rations de f rac t ionnement .
On remarque entre les deux soucbes de souris une faible diff6rence, la quantit6 de r ibosomes li6s aux membranes ergastoplasmiques est 48 p. cent pour les souris normales, 49 p. cent pour les souris haines, tandis que les r ibosomes non lids reprd- sentent respect ivement 28 p. cent et 23 p. cent de la densit6 optique h 260 ~ du surnageant postmi- tochondr ia l (tableau I).
Etude comparative des profils de polysome de role.
Cette dtude est faite en d6posant le surnageant pos tmi tochondr ia l sur gradient cont inu de saccha- rose 15 p. cent-40 p. cent TKM ; les condi t ions de pr6formation des gradients sont celles de gradients l in6aires pr6par6s ~ par t i r de deux solutions de saccharose en tampon T K M - p H 7,4 /~ 15 p. cent et 40 p. cent (poids de saccharose /poids final, dans des tubes Sw 25 et sons un volume final de 20 ml).
Apr6s lecture de la DO 260 m,:x, les concentra- t ions des surnageants pos tmi tochondr iaux des sou-
ris naines et normales sont ajustdes avec le tam- pon d 'homogdn6isat ion h 60 A260/ml, puis addi- t ionn6es de ddsoxycholate de sodimn h la surface de chaque gradient sons le volume de 0,25 ml, soit 15 A26 o dquivalents par tube SW 25 1. Un t6moin est r6alis6 en d6posant un m6me volume de tam- pon ayant servi ~ la pr6para t ion de l 'homogdnat ; un second t6moin est fair en ajoutant prdalable- ment au surnageant pos tmi tochondr ia l de la RNase pancr6at ique 50 7. /t 1 mg /ml : elle d6grade les polysomes en disomes 116 S et monosomes (80 S).
La centr i fugat ion est r6alis6e ~ 28 000 tpm rotor SW 25 dans une Spinco L 50 r6frigdrde ~ 4 °, pen- dant 2 h 50.
La lecture de l ' absorpt ion ~ 260 irr~ est faite au spectrophotom6tre Beckmann et enregistr6e auto- matiquement , le l iquide arr ive de facon cont inue dans une cuve ~ c i rcula t ion de trajet optique 1 cm, l ' aspi ra t ion est faite ~ l 'aide d 'une pompe Perpex aprbs in t roduc t ion de l 'aiguille dans le fond du tube - - la vitesse d 'aspi ra t ion est de 137 ml par heure.
Les quantitds de polysomes et monosomes recueill is sont d6termin6es en in t6grant la fonct ion DO 260 m~x f(x), (x) ~tant le volume total de l iquide aspir6 h chaque ins tant depuis le d6but de la lec- ture.
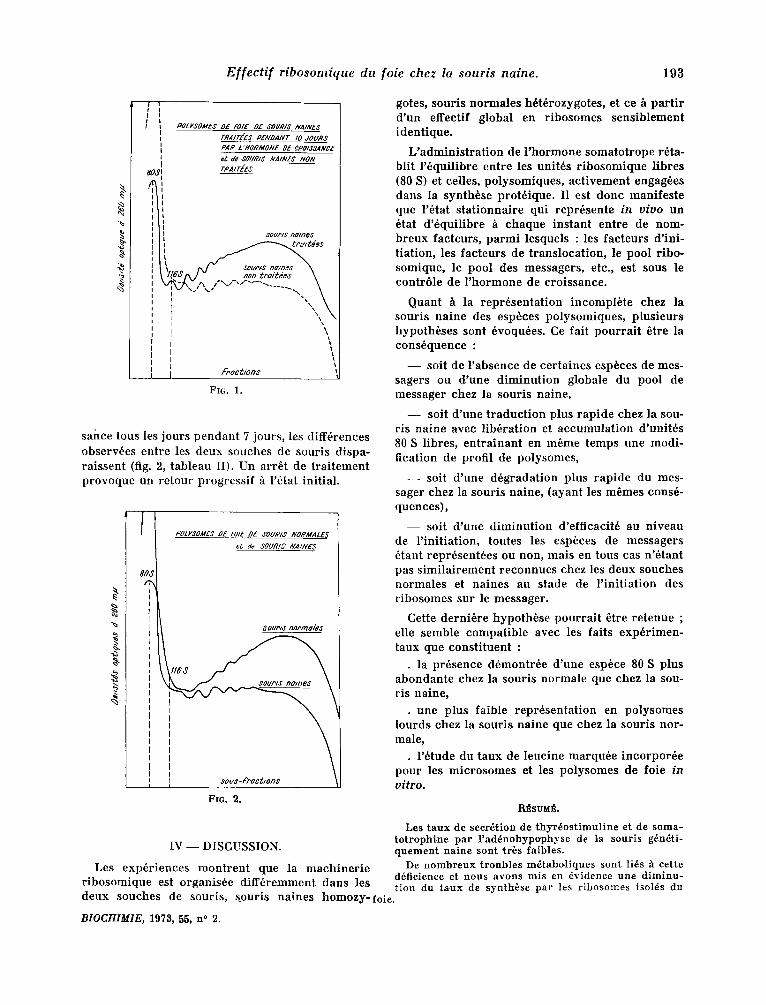
Pour calculer le rappor t polysomes/monosomes, on fair le rappor t des surfaces comprises sons les courbes reprdsentant respect ivement les deux esp6ces (fig. 1, tableau II).
III - - Rt~SULTATS.
Souris normales et souris haines non trait~es.
Le surnageant pos tmi tochondr ia l est ident ique pour les deux souches ; en par t icu l ie r si l 'on con- sidbre l ' absorp t ion h 260 rn~, on volt que h 1 g de tissu frais correspond une densit6 optique h 260 m~ de 115 UDO pour le surnageant postmitochon- drial, qu ' i l s'agisse du foie de souris normales ou de souris naines.
C'est en tenant compte de ce rdsultat que le ren- dement de toutes les fract ions r ibosomiques, calcn- 16 dans un premier temps par rappor t au surna- geant pos tmi tochondr ia l , a 6t6 rapport6 h u n m6me poids d 'organe frais (r6sultats tableau I).
I1 apparai t que le r endement en ribosomes l ibres cor respondants ~ 1 g de foie est pen diffdrent chez les souris normales : 32 UDO 260 m~, et chez les souris n a i n e s : 26.5 UDO 260 ~ ; il e n e s t de
BIOCHIMIE, 1973, 55, n ° 2.

192 J. Mellet .
m 6 m e p o u r le r e n d e m e n t en r i b o s o m e s li6s aux m e m b r a n e s e r g a s t o p l a s m i q u e s (54 UDO 260 m~ c o n t r e 56.35 UDO 260) et p o u r le r e n d e m e n t en r i b o s o m e s t o t aux (87 et 82.8).
Si 1'on cons id~ re que ces r e n d e m e n t s ref l6 tent les quan t i t6s de r i b o s o m e s li6s et non li6s h la m e m b r a n e e r g a s t o p l a s m i q u e c o n t e n u clans 1 g de foie [r6f. 11] on peu t c o n c l u r e que la c o m p o s i t i o n g loba le du fo ie en r i b o s o m e s to taux est peu diff6- r en te p o u r les d e u x souches de sour is .
72 p. cen t de l ' a b s o r p t i o n h 260 m~ du surna- geant p o s t m i t o c h o n d r i a l sont dus aux r i b o s o m e s chez la sour i s n o r m a l e , 68 p. cen t chez la sour i s na ine , e f fec t i f r i b o s o m i q u e g loba l qui se r6pa r t i t en d e u x f r a c t i o n s : l ' une est l ib re , l ' au t r e est l i6e aux m e m b r a n e s e rga s top l a smiques .
Mais les quan t i t6s r e l a t i ve s de p o l y s o m e s et monoso ines son t s i g n i f i c a t i v e m e n t d i f f6 ren tes (fig. 1) p u i s q u e le r a p p o r t quan t i t 6 de p o l y s o m e s / q u a n - tit6 n m n o s o m e s est de 4,8 p o u r les sour i s n o r m a l e s et 2,68 p o u r les sour i s na ines ( tab leau II) .
- - Si l ' on c o m p a r e la r e p r 6 s e n t a t i o n de l ' e spbce p o l y s o m i q u e en t r e les d e u x souehes , l ' 6 q u i p e m e n t p o l y s o m i q u e de la sour i s n a i n e est 87 p. c e n t de ce lui de la sour i s no rma le .
- - P a r c o n t r e l ' e sp~ce 80 S qu i r e p r 6 s e n t e les r i b o s o m e s f ibres ou engag6s senls sur le m e s s a g e r est p lus a b o n d a m m e n t r ep r6sen t6e chez les sour i s naines .
Aprbs t r a i t e m e n t c h r o n i q u e de la sour i s na ine pa r i n j ec t i on sous-cu tan6e d ' h o r m o n e de c ro is -
TABLEAU ][.
Rendement du [ract ionnement de surnageant postmitochondrial (SPM) en ribosomes non li~s, li~s et totaux (F I, F II, F I l I ) chez les Souris nor- males et les Souris naines.
Fraction F II
( R i b o s o m e s non li6s)
Fraction F I I I
(Ribosomes li~s)
Fraction F I (Ribosomes
totaux)
Rendement du SPM
Souris normales Souris haines
25 p. cent ( 27 p. cent t 28 p. cent 32 p. cent
23 p. cent { 25p. cent i 23 p. cent 21 p. c en t .
Ribosomes libres contenus dans 32 A.,~o 26.45 A~o 1 g de foie frais (moyenne)
45 p. cent t Rendement du SPM 47 p. cent i 48 p. cent
48 p. cent
R i b o s o m e s li6s c o n t e n u s dans 54 A.26o 1 g de foie ( m o y e n n e )
Rendement du SPM 76 p. cent
48 p. cent { 49 p, cent I 49 p. cent 51 p. cent
56.35 A~,~
R i b o s o m e s totaux contenus dans 1 g de foie
87 A.26,, [82,8 A26,,
TABLEAU II.
Comparaison entre les qaantit~s de ribosomes tolaux e t l a r~partition des ribosomes en polysomes et monosomes, en [onction de l'~tat hormonal des souris.
Polysomes
Monosomes Rdt du gradient en ribosomes :
(Polysomes ,-]- monosomes)
Souris haines Souris aaiues trait~es 7 j par STM
2,7 t 4,6 2,5 2,68 4,8 { 4,8
2,85 5
6 5 p ' c e n t I c e n t ~ p ' c e n t ! 53p . cent 62 p. cent 65 p. p. cen t ( 70 p. c e n t
Souris normales
4,55 t 4,76 4,75~ 5
50 p. cent i 52 p. cent i 52 p. cent 52 p. cent
BIOCHIMIE, 1973, 55, n ° 2.
Effectif ribosomique du foie chez la souris naine. 193
POLY~OMf~ DE .rOlE DE ~OMRI~ .'VAINE,.~
TRAITEES PENDANT I0 JOUR~ PAR L'HORMONE OE CROI..e..eANPE
e~ de ~OU, q/~ ,~¢AINES NON TR, dlT[E~
~vOUPIS ng/neg
\ \ \
Fpac'~ion~o
FIG. 1.
\
sance tous le s jours pendan t 7 jours, les diff6rences observ~es entre les deux souches de souris dispa- raissent (fig. 2, tableau II). Un arr6t de t ra i tement provoque un retour progressif ~ l '6tat init ial .
POLYSOME.~ DE FOIE DE SOUR/~ HORMALE~
c t de 30URI,.~ NAINES
~OUr/,S normale~
\ , \
I ~ous-Praa~/ona i
Fro. 2.
IV - - DISCUSSION•
Les experiences mont ren t que la machiner ie
gotes, souris normales h6t6rozygotes, et ce ~ par t i r d 'un effectif global en r ibosomes sensiblement identique.
L ' admin i s t r a t ion de l ' ho rmone somatotrope r6ta- blit l '6qui l ibre entre les unit6s r ibosomique i ibres (80 S) et celles, polysomiques, act ivement engag6es dans la synth6se prot6ique. II est donc manifeste que l '6tat s ta t ionnai re qui repr6sente in oivo un 6tat d '6qui l ibre ~ chaque ins tan t entre de nom- breux facteurs, pa rmi lesquels : leg facteurs d ' in i - tiation, les facteurs de t ranslocat ion, le pool r ibo- somique, le pool des messagers, etc., est sous le contr61e de l 'hormone de croissance.
Quant ~ la repr6senta t ion incompl6te chez la souris na ine des esp6ces polysomiques, p lus ieurs hypoth6ses sont 6voqu6es. Ce fait pour ra i t 6tre la cons6quence :
- - soit de l 'absence de certaines esp6ces de mes- sagers ou d 'une d iminu t ion globale du pool de messager chez la souris naine,
- - soit d 'une t raduc t ion plus rapide chez la sou- ris na ine avec l ib6rat ion et accumula t ion d 'uni t6s 80 S libres, en t r a inan t en m6me temps une modi- fication de profil de polysomes,
- - soit d 'une d6gradat ion plus rapide du mes- sager chez la souris naine, (ayant les mfmes cons6- quences),
- - soit d 'une d iminu t ion d'efficacit6 au niveau de l ' in i t ia t ion , toutes les esp6ces de messagers 6tant repr6sent6es ou non, mais en tous cas n '6 tant pas s imi la i rement reconnues chez les deux souches normales et na ines au stade de l ' in i t i a t ion des ribosomes sur le messager.
Cette derni6re hypoth6se pour ra i t 6tre re tenue ; elle semble compatible avec les faits exp6rimen- taux que const i tuent :
. la pr6sence d6montr6e d 'une esp6ce 80 S plus abondante chez la souris normale que chez la sou- ris naine,
• une plus faible repr6senta t ion en polysomes lourds chez la souris na ine que chez la souris nor- male,
• l '6tude du taux de leucine marqu6e incorpor6e pour les microsomes et les polysomes de foie in vitro.
P ~ S U M ~ .
Les taux de secr6tion de thyr6ostimuline et de soma- totrophine par l'adSnohypophyse de la souris g~n6ti- quement naine sont tr~s faibles.
De nombreux troubles m6tab~liques sont li6s h cette d6ficience et nous avons mis en 6vidence une diminu-
r ibosomique est organis6e diff6remment dans les tion du ta.ux de synth6se par les ribosomes isolds du deux souches de souris, s.ouris na ines homozy-foie.
BIOCHIMIE, 1973, 55, n ° 2.

1 9 4 J . M e l l e l .
Le pr6sent .travail eomporte l '6tude du profil polyso- mique du foie de la sour is na ine avan t et apr6s ¢raite- men t par l ' h o r m o n e somato t rope et 1.a t r i iodothyronine . Chez la sour is na ine la propor t ion p o l y s o m e s / m o n o - somes est p lus faihle que chez la sour is normale . Le t r a i t e m e n t de la sour i s na ine par :la t r i i odo thyron ine ne modifie pas cet 6tat. L ' in jec t ion d ' h o r m o n e somato- trope au eont ra i re r6tabl i t le rappor t h une va leur nor- male.
BI~BLIOGRAPHIE.
1. Tara, J. R., Ernster , L., Lindberg, O., Arrhenius , E., Ped'ersen, S. ~ Hedman, R. (1963) Biochem. J., 86, 408-428.
2. Tata, J. R. (19~6) Progr. Nucleic. Acid. Res. Mol. Biol., 5, 1911.
3. Tara, J. R. (196,8) in Regulatory mechan i sms for protein synthes is in m a m m a l i a n cells (Pietro, A., Lambt~rg, H. R. ~ Kenneys , F. T., eds.), Pro- ceedings of Thi rd Ket ter ing S y m p o s i u m 299, Aca- demic Press, New York.
4. Corr~ze, C1. ~ Nunez, J. (1,9~6~) Bull . Soc. Chim. Biol., 5, 909-917.
5. Korner, A. (1960) Biochem. J., 74, 462-471. 6. Korner, A. (1959) Biochem. J., 73, 61-71. 7. Korner , A. (1961) Biochem. J., 81, 292-297. 8. Jefferson, L. S. ~ Korner, A. (19.67) Biochem. J., 104,
826. 9. Jefferson, L. S. & Korner, A. (1!9,6!9,) Biochem. J., 111,
703. t0. Clemens, H. J. ~ Korner , A. (197'0) Bioehem. J., 119,
629.
11. Tata, J. R. ~ Widnell , C. C. (196,6) Biochem. J., 98, 604-620.
12. Widnel l , C. C. a Tata, J. R. (1966) Biochem. J., 98, 621-627.
13. Jackson, C. D. & Selle, B. H. (19675 Biochem. Bio- phys. Acta, 142, 419-429.
14. Korner, A. (19.65) Recent Progr. Hormones Res., 21, 205.
15. Smuckter , E. A. a Tara, J. R. (1971) Nature, 234, 37-39.
16. Lewis, U. J., Cheerer, E. V. a Vander laan , W. P. (1965) Endocrinology, 76, 362.
17. Daughaday , W. H., P eake, G. T., Birge, C. A. Marisz, I. K. (1967) Intern. Symp. Growth Hor- mone, Milan, 238, Excerpta Medica, Ams te rdam.
18. E'scobar del Rey, F., Morreale de Escobar, G., Jolin, T. a Lopez Quijada, C. (1968) Endocrinology, 83, 41.
19. Snell, G. D. (1929) Proc. Nal. Acad. Sc. U.S., 15, 733.
20. Smith, P. E. ~ MacDowell, E. C. (1930) Anat . Record, 46, 249.
21. Rennets , E. G. & MacMitt, W. (1958) Anat. Record, 131, 5,.91.
22. Elf, tman , H. ~ ~Vegelius, O. (19.59.) Anat. Record, 135, 43.
23. Peterson, IL R. (1959) Anat. Record, 135, 43. 24. Or tman, R. (1'9'56) J. Morphol., 99, 419. 25. Lewis, U. J., Cheerer, E. V. a Vanderl.aan, W. P.
(1.965) Endocrinology, 76, 210. 26. Bartke, A. (1964) Anat. Record, 149, 225. 27. Van Tol, A. (1970) Biochem. Biophys. Acla, 219,
227-230. 28. Blobel, G. ~ Van Potter, R. (1967) J. Mol. Biol., 26,
279-301.
BIOCHIMIE, 1973, 55, n ° 2.





























