Embed Size (px)
Citation preview

République Algérienne Démocratique et Populaire
Université d’Oran Es-sénia
Faculté des sciences
Département de biologie
Laboratoire de Microbiologie Appliquée
Mémoire pour l’obtention du diplôme de magister en microbiologie
Thème :
Etude du potentiel des bactéries lactiques
pour leur utilisation en industrie laitière
Présenté par :
Mlle RABAH noura
Composition de jury :
Président : Pr. HENNI .D.E
Examinateur : Pr. KIHAL .M
: Dr. GUESSAS.B
: Dr. HEDADJI.M
Encadreur : SAIDI. N
2009-2010

Remerciement :
Ce travail a été réalisé au niveau du laboratoire de microbiologie appliquée du
département de biologie, Faculté des Sciences Es-senia.
Je tien a remercier vivement :
Pr. HENNI. D. directeur du laboratoire de phytopharmacie, qui a bien vouloir présider le jury.
Dr. SAIDI. N. pour l’encadrement et les conseils scientifiques qu’il a apporté à mon travail.
Pr. KIHEL. M. le directeur du laboratoire de microbiologie appliquée pour l’attention et l’aide
précieuse qu’il m’a prodigué et d’avoir jugé ce travail.
Dr. GUESSAS. B. et Dr. HADADJI. M. d’avoir accepté d’examiner mon travail.
Tous ceux qui m’ont aidé durant la réalisation de ce travail.

Sommaire :
1. Introduction ..................................................................................................... 1
2. Analyse bibliographique ................................................................................ 4
2.1Les bactéries lactiques .................................................................................... 5
2. 2. Classification des bactéries lactiques ......................................................................5
2.2.1. Streptococcus ........................................................................................................5
2.2.2. Lactococcus .........................................................................................................5
2.2.3. Pediococcus ..........................................................................................................6
2.2.4. Leuconostoc ............................................................................................... 6
2.2.4. Lactobacillus ............................................................................................... 9
3.2. Les bactériocines ........................................................................................... 11
2.3.1. Définition ..................................................................................................... 11
2.3.2. Nomenclature ............................................................................................. 11
2.3.3. Bactériocines et antibiotiques ..................................................................... 12
2.3.4. Classification des bactériocines .................................................................. 14
2.3.5. Génétique des bactériocines .................................................................................16
2.3.6. Biosynthèse et excrétion ........................................................................................19
2.3.7. Mode d’action ........................................................................................................20
2.3.8. Immunité de la bactérie productrice .......................................................................22
2.3.9. Purification des bactériocines ................................................................................22
2.3.10. Rôle écologique des bactériocines ......................................................................24
2.3.11. L’application des bactériocines dans le secteur alimentaire .................................24
3. Matériel et méthodes ...................................................................................... 26
3.1. Matériel biologique et conditions de culture ................................................... 28
3.1.1. Laits ........................................................................................................... 28
3.1.2. Milieux et conditions de culture ................................................................... 28
3.1.3. Conservation des souches ......................................................................... 28
3.2. Méthodes ....................................................................................................... 29
3.2.1. Isolement des lactocoques ......................................................................... 29
3.2.2. Recherche de la catalase ........................................................................... 29

3.2.3. Identification des souches purifiées ............................................................ 30
a- Test de l’arginine dihydrolase (ADH) ................................................................ 30
b- Le type fermentaire........................................................................................... 30
c- Température de croissance .............................................................................. 31
d- Croissance à 0%, 4%, 6,5% de NaCl et à pH 9,6 ............................................ 31
e- Test de lait bleu de Shermann .......................................................................... 31
f- Fermentation des sucres .................................................................................. 31
3.2.4. Cinétique d’acidification .............................................................................. 32
3.2.5. Antagonisme direct sur milieu solide ......................................................... 33
3.2.6. Détection de la substance inhibitrice dans le surnageant de culture .......... 33
3.2.7. Caractérisation de la substance inhibitrice ................................................. 33
a-Action des protéases ......................................................................................... 33
b-Stabilité thermique ............................................................................................. 35
4. Résultat et discussion ............................................................................... 36
4.1. Résultat des tests macroscopiques ............................................................... 36
4.2 . Etude microscopique ................................................................................... 36
4.3. Test de catalase ............................................................................................ 40
4.4. Test de NaCl et de pH ................................................................................. 40
4.5. Test du lait bleu de shermann ....................................................................... 40
4.6. Test du profil fermentaire ............................................................................... 42
4.7. Test de l’arginine déshydrogénase 42
4.8. Test de fermentation des sucres ................................................................... 42
4.9. Cinétique d’acidification ................................................................................. 48
4.10. Test d’antagonisme sur milieu solide ........................................................... 48
4.11. Recherche de l’activité inhibitrice dans le surnageant de culture ................ ..53
4.12. Identification des agents inhibiteurs des souches testées ........................... ...53
5. Discussion ...................................................................................................... …57
Conclusion et perspectives .............................................................................. …61
Références………………………………………………………………………………… 62
Annexe

Résumé :
Les bactéries lactiques jouent un rôle important dans la fermentation des
aliments. Elles peuvent synthétiser des substances antibactériennes nommées
bactériocines qui sont utilisées dans la bioconservation des aliments.
L’objectif de notre travail est l’isolement, la purification et l’identification de
lactocoques qui on un pouvoir antagoniste.
Différentes méthodes basées sur les critères morphologiques, physiologiques
et biochimiques on été utilisées.
Nous avons isolé à partir de lait de chamelle, 30 souches de bactéries
lactiques et de les classer en plusieurs espèces dont Lactococcus lactis est la plus
dominantes.
18 souches ont été retenues après avoir éliminé l’effet de l’acidité, pour leurs
activités bactéricides, notamment vis-à-vis des bactéries pathogènes :
Staphylococcus aureus et Escherichia coli. Cette activité antibactérienne pourrai être
due à une substance excrétée dans le surnagent de culture des isolats. Ces agents
inhibiteurs ont présenté une sensibilité aux traitements thermiques et enzymatiques.
Mots clés : bactéries lactiques, lactocoques, industrie alimentaire, bioconservation,
effet inhibiteur, bactériocines, bactéries pathogènes.

Liste des tableaux :
Tableau 1: principaux caractéristiques des lactocoques. (DeRoissart, 1986)
Tableau 2: principaux caractères des pédiocoques (DeRoissart, 1986)
Tableau 3: principaux caractères des leuconostoques (DeRoissart, 1986)
Tableau 4 : les principaux caractéristiques des trois groupes de Lactobacillus
Tableau 5. différences entre bactériocines et antibiotiques (Cleveland et
al., (2001)
Tableau 6: aspect macroscopique des colonies et l’aspect microscopique des
souches purifiées et leurs caractères biochimiques
Tableau 7: profil fermentaire des isolats
Tableau 8: présentation de l’identification des souches isolées.
Tableau 9: la variation du pH et du degré Dornic des souches 10B, 6C et 2 en
fonction du temps
Tableau 10: diamètre des zones d’inhibition provoqué par les souches actives sur
les souches pathogène en cm
.

Liste des figures :
Figure 1 : séquence et structure de lantibiotiques de type A (Nisine), B (Mersacidine)
et d’un lantibiotique « two-peptides (Lacticine 3147 A1 et A2) respectivement
Figure 2 : représentation schématique des opérons de la coaguline (Le Merrec et al.,
2000) et de la nisine (Siergers et Etian, 1995)
Figure 3 : aspect macroscopique des colonies en surface (A). Aspect d’une culture
pure dans un milieu liquide M17(B)
Figure 4 : aspect microscopique à l’agrandissement G100 après coloration de Gram
de la souche 11B.
Figure 5 : résultat du test de bleu de méthylène
Figure 6 : aspect des souches ADH positives sur milieu M16 BCP, la couleur mauve
représente la présence de l’ADH
Figure 7 : résultat du test des sucres. 1. souche 1B, 2. souche 11A
Figure 8: la variation du pH des souches 10B, 6C et 2 en fonction du temps
Figure 9: variation du D° des souches 10B, 6C et 2 en fonctio n du temps
Figure 10: test d’antagonisme direct sur milieu solide en cm
Figure 11: diamètre des zones d’inhibitions en centimètre des souches testées vis-à-
vis des souches pathogènes
Figure 12 : zones d’inhibitions provoquées par le surnagent de culture des souches
testées

1
1. Introduction

2
La recherche sur les bactéries lactiques et leur caractérisation ont
énormément modifié la fabrication des produits laitiers fermentés. La capacité de
manipuler et de contrôler ces microorganismes a atteint maintenant un tel niveau
qu’il était inimaginable d’y penser il y a quelques années (Eijsink, 2002).
Ces recherches ont permis aux microbiologistes et aux industriels de choisir
les meilleures souches et d'améliorer la productivité, la qualité, et la sûreté des
produits finaux. La caractérisation des bactéries lactiques a favorisé le
développement de souches bactériennes bien définies, maintenant connues sous le
nom de levains lactiques qui jouent un rôle technologique fondamental en
transformation laitière (Saidi et al., 2002).
La grande méfiance des consommateurs vis-à-vis des additifs alimentaires
comme les conservateurs chimiques utilisés pour augmenter la durée de vie de
certains aliments ainsi que l’utilisation de traitement thermique souvent préjudiciable
aux propriétés organoleptiques et nutritionnelles d’aliments sensibles à la chaleur,
sans oublier la résistance qui accompagne l’utilisation massive des antibiotiques, ont
conduit les chercheurs à l'exploitation de nouvelles souches, telle que les bactéries
lactiques, possédant des activités biologiques particulières comme la production de
bactériocines, substances inhibitrices pour des applications de biopréservation
(Deegan et al., 2006; Galvez et al., 2007; Guinane et al., 1989 ; Vermeiren et al.,
2004).
Les bactériocines les plus étudiées sont celles des bactéries à Gram
négatives comme les colicines des entérobactéries (Cleveland et al., 2001), et les
bactériocines des bactéries lactiques (Neetles et barefoot, 1993), seule la nisine est
reconnue généralement comme efficace. Cette substance saine et sans danger
(GRAS : generally reconised as safe) est acceptée par l’organisation mondiale de la
santé comme étant un conservateur alimentaire dans plus de 40 pays incluant les
USA (Cleveland et al., 2001).
De ce fait, les bactéries lactiques sont largement impliquées dans la
fabrication de produits laitiers fermentés (PLF), qui jouent un rôle majeur dans leurs
conservations et elles contribuent à l’inhibition des germes contaminants (Guessas,
2007).

3
Le Laboratoire de Microbiologie Appliquée vise à la caractérisation d’activités
technologiques importantes des bactéries lactiques et leur exploitation dans le
secteur agroalimentaire et par la suite parapharmaceutique.
Etant donné tous ces intérêts, le Laboratoire de Microbiologie Appliquée a
constitué, depuis plusieurs années, une collection de souches locales de bactéries
lactiques provenant de différents laits (Saidi et al., 2002) et ainsi que des souches de
Bifidobacterium (Hadadji et al., 2005).
Cette étude s’inscrit dans le cadre d’un axe de recherche du Laboratoire de
Microbiologie Appliquée du département de Biologie, Faculté des Sciences,
Université d’Oran. Ce mémoire comprend quatre parties:
1. une synthèse bibliographique
2. une partie matériel et méthodes qui décrit toutes les techniques qui se
rapportent aux objectifs de ce thème.
3. les résultats de ce mémoire qui sont répartis en deux volets.
Volet 1: occupe une place centrale et principale au sein de notre laboratoire.
La tache consiste à isoler, identifier, selon des tests phénotypiques, de nouvelles
souches locales de bactéries lactiques isolées à partir de différents laits crus. Les
souches ainsi identifiées sont ensuite caractérisées selon des critères
technologiques. Il a pour objectif de constituer et gérer une banque de souches.
Volet 2: La caractérisation de nouvelles souches de bactérie lactique
productrices de bactériocines et de développer les connaissances nécessaires à leur
exploitation en transformation fromagère et pour la production de bioingrédients à
activité antimicrobienne.
4. Une discussion qui traite les différents résultats obtenus en comparaison
avec d’autres travaux de recherche.
L’objectif de notre travail est :
- L’isolement de lactocoques à partir de lait de chamelle ;
- La recherche des substances antibactérienne ;
- L’identification des bactéries lactiques productrice de bactériocines.

4
2. Analyse bibliographique

5
2.1. Les bactéries lactiques
Les bactéries lactiques regroupent un ensemble d’espèces hétérogènes dont
le trait commun est la production d’acide lactique. Elles appartiennent à divers
genres comme Bifidobacterium, Enterococcus, Lactobacillus, Lactococcus,
Leuconostoc, Pediococcus, Streptococcus, Aerococcus, et Carnobacterium.
Elles interviennent dans l’industrie laitière et dans la fermentation de
nombreux autres produits alimentaires : salaison des légumes, boulangerie etc.
(Hugenholtz et Kleerebezem, 1999). Les bactéries lactiques contribuent à la texture,
à la saveur des aliments et à la production de composés aromatiques (Callewaert, et
al., 2000). Leur pouvoir antagoniste résulte aussi d’une compétition vis-à-vis du
substrats et, si les conditions de développement sont favorables pour l’élaboration de
bactériocines comme la nisine (Kleerebezem, 2004; Lubelski, 2008).
2. 2. Classification des bactéries lactiques
2.2.1. Streptococcus
Appartenant à la famille des Streptococcaceae. Les espèces de ce genre sont
des cellules ovoïdes, sphériques ou quelques fois allongées en fuseaux en paires ou
en chainettes. Les streptocoques lactiques se distinguent par leur capacité de croitre
à 45°C (Garvie et Farrow).
2.2.2. Lactococcus
Le groupe de lactocoques correspond aux streptocoques mésophiles de la
flore lactique. Elles sont généralement microaérophiles et se développent à 30°C.
Leur fermentation des sucres est homolactique et donne de l’acide lactique comme
produit final (Tableau1).
Les lactocoques peuvent se trouver dans les aliments comme les produits
laitiers, elles sont utilisées dans l’industrie agroalimentaire, les espèces les plus
importantes sont: Lactococcus lactis et Lactococcus cremoris (Stiles et Holzapfel,
1997).

6
2.2.3. Pediococcus
Les pédiocoques sont des bactéries microaérophiles à besoin nutritifs
complexes. Leurs développements nécessite plusieurs facteurs de croissances,
plusieurs espèces sont acidophiles, elles sont saprophytes et contaminent les
produits végétaux, ce sont des agents de dégradation en brasserie. Elles peuvent
aussi provoquer des altérations (viscosité) dans les produits végétaux (olives).
Parfois, on les utilise comme des levains lactiques pour les charcuteries (Pèrez,
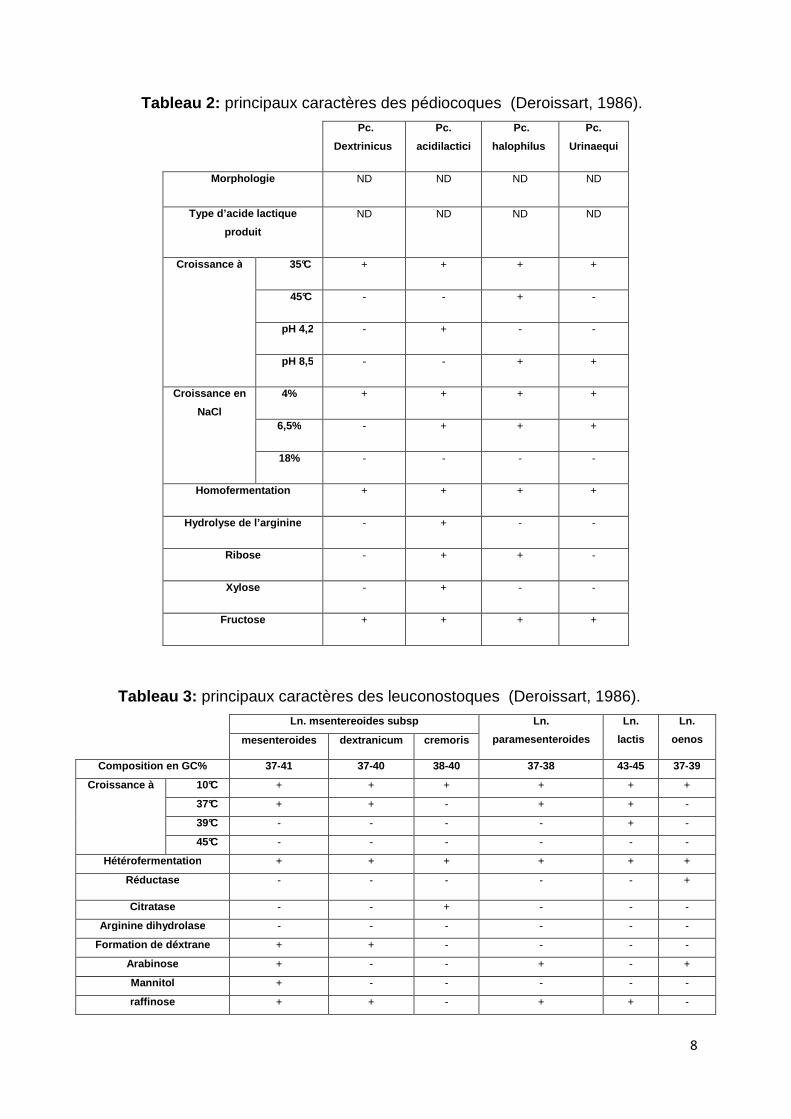
2005). Les différents caractères de ces bactéries sont résumés dans le tableau 2.
2.2.4. Leuconostoc
Les leuconostoc ont de grandes exigences sur le plan nutritionnel vis-à-vis
des vitamines et des acides aminés. Elles sont classées en quatre espèces :
-Ln. mesenteroides.
-Ln. paramesenteroides.
-Ln. lactis.
-Ln. oenos.
Ce genre comporte quelques souches qui ont la faculté de produire, à partir
du saccharose, une capsule polysaccharidique épaisse. Elles sont responsables de
contaminations et d’altérations de divers produits tels que les boissons acides et
sucrés. Elles sont utiles dans certains fromages car elles facilitent leur ouverture par
la production de CO2 (Kihel, 1996).

7
Tableau 1: principaux caractéristiques des lactocoques. (Deroissart, 1986).
1 2 3
ADH + - +
CO2 - - +
Acétoine - - +
culture 10°c + + +
culture 37°c + + +
culture 40°c + - +
culture 45°c - - -
croissance en pH 9,6 + - +
résistance 30 mn à 63°c - - -
culture 4% de NaCl + - +
Fermentation des sucres
Arabinose - - -
Galactose - - -
Glucose + + +
Maltose + + +
Raffinose + - -
Saccharose - - -
Tréhalose + - -
1. Lactococcus lactis subsp. lactis
2. Lactococcus lactis subsp cremoris
3. Lactococcus lactis subsp. lactis biovar diacetylactis
8
Tableau 2: principaux caractères des pédiocoques (Deroissart, 1986).
Pc.
Dextrinicus
Pc.
acidilactici
Pc.
halophilus
Pc.
Urinaequi
Morphologie ND ND ND ND
Type d’acide lactique
produit
ND ND ND ND
Croissance à
35°C + + + +
45°C - - + -
pH 4,2 - + - -
pH 8,5 - - + +
Croissance en
NaCl
4% + + + +
6,5% - + + +
18% - - - -
Homofermentation + + + +
Hydrolyse de l’argin ine - + - -
Ribose - + + -
Xylose - + - -
Fructose + + + +
Tableau 3: principaux caractères des leuconostoques (Deroissart, 1986). Ln. msentereoides subsp Ln.
paramesenteroides
Ln.
lactis
Ln.
oenos mesenteroides dextranicum cremoris
Compos ition en GC% 37-41 37-40 38-40 37-38 43-45 37-39
Croissance à
10°C + + + + + +
37°C + + - + + -
39°C - - - - + -
45°C - - - - - -
Hétérofermentation + + + + + +
Réductase - - - - - +
Citratase - - + - - -
Arginine dihydrolase - - - - - -
Form ation de déxtrane + + - - - -
Arabinose + - - + - +
Mannitol + - - - - -
raffinose + + - + + -

9
2.2.5. Lactobacillus
Les bactéries appartenant à ce genre sont des bâtonnets souvent allongés, à
Gram positives, parfois groupés en paires ou en chaines. Elles sont catalase
négatives, microaérophiles. Chez les lactobacilles, le métabolisme des sucres produit
de l’acide lactique comme produit final ou homofermentaires, d’autres produisent, à
coté de l’acide lactique, de l’éthanol, du CO2 et d’acides volatils (Siegumfeldt et al.,
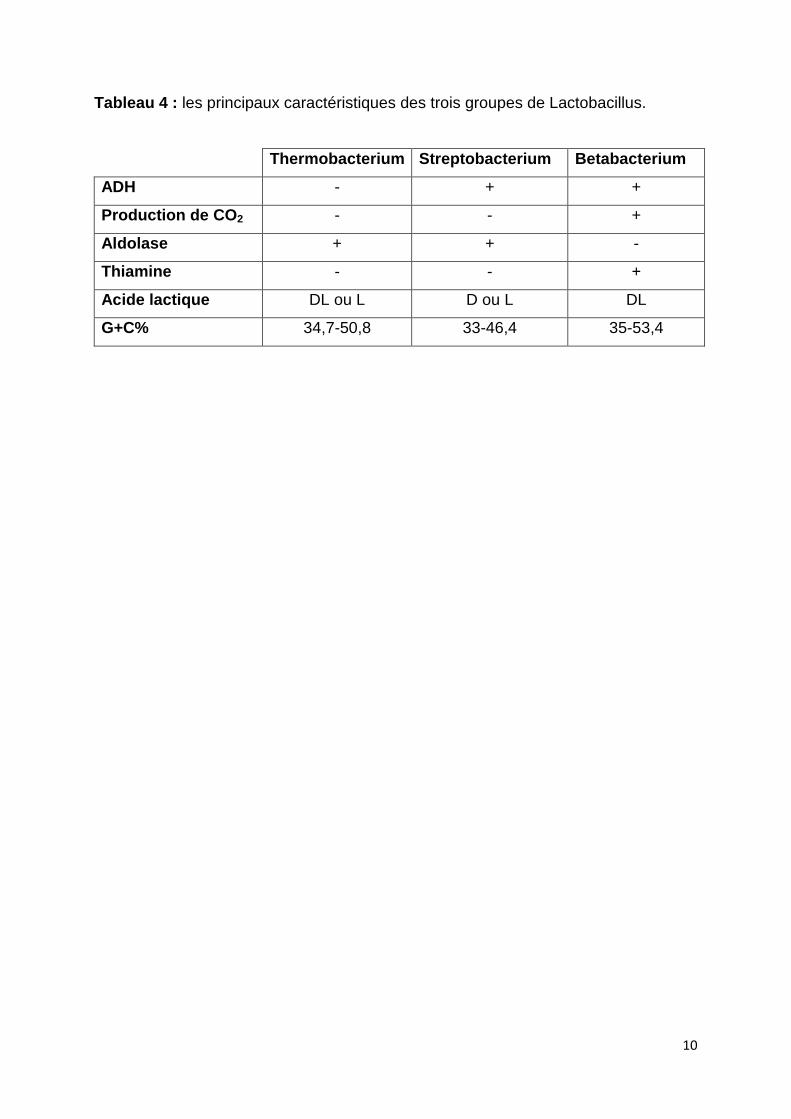
2000). Ce genre est subdivisé en trois groupes (Tableau 4):
- Thermobacterium
Il comprend des bactéries homofermentaires thermophiles qui se développent
à 45°C mais pas à 15°C, les espèces les plus fréque ntes dans l’alimentation (lait,
yaourts, fromages) sont Lb. helveticus, Lb. jugurti, Lb. bulgaricus, Lb. lactis, Lb.
acidophilus, etc. (Hammes et Hertel, 2006).
- Streptobacterium
Il regroupe les lactobacilles homofermentaires mésophiles qui se développent
à 15°C. Il comporte les espèces Lb. curvatus, Lb. sake, Lb. graminis, etc (Bottazzi,
1988).
- Betabacterium
Ce groupe comporte les bacilles hétérofermentaires. Les espèces les plus
fréquentes dans l’alimentation sont : Lb. kefir, Lb. fructivorans, Lb. hilgardii, etc.
Les lactobacilles sont très utilisées en laiteries, fromagerie. Leur rôle est
également important dans les végétaux fermentés. Les caractéristiques des trois
groupes sont résumées dans le tableau 4 (De Roissart et Sharpe, 1994).
10
Tableau 4 : les principaux caractéristiques des trois groupes de Lactobacillus.
Thermobacterium Streptobacterium Betabacterium
ADH - + +
Production de CO 2 - - +
Aldolase + + -
Thiamine - - +
Acide lactique DL ou L D ou L DL
G+C% 34,7-50,8 33-46,4 35-53,4

11
Les bactériocines
2.3.1. Définition
La première définition des bactériocines est celle donnée aux colicines et
stipulait qu’une bactériocine est une substance inhibitrice caractérisée par une
composante active majeure de nature protéique, une activité létale intra spécifique et
elle s’adsorbe à un récepteur spécifique. Cette définition reste non appropriée du
faite du grand nombre de bactériocines pour lesquelles au moins un de ces critères
n’est pas vérifié.
La définition la plus valable est celle donnée par Jack et al., (1995) qui
considèrent les bactériocines comme étant toute molécule de type bactériocine-like
directement produite par synthèse protéique ribosomale sous forme de prépeptide ou
de précurseur de polypeptides générant après clivage le propeptide ou la protéine
active.
2.3.2. Nomenclature
Pour ne pas donner des appellations différentes à la même bactériocine
comme pédiocine PA-1 et pédiocine AcH ou à des variants naturels de la même
bactériocine comme coaguline et pédiocine, les auteurs recommandent de ne pas
attribuer un nom à une bactériocine nouvellement découverte et d’utiliser le terme
BLIS (bacteriocin-like inhibitory substance) suivi de la désignation de la souche
productrice. Quand le séquençage prouve que la nouvelle substance diffère des
bactériocines déjà décrites, l’appellation doit se baser sur le genre ou de préférence
sur l’espèce productrice. Quand la substance présente des différences mineurs avec
une bactériocine connue, et que ces différence ne provoquent pas de grands
changements au niveau de la structure secondaire ou le spectre d’activité, alors elle
doit être considérée comme un variant naturel de la bactériocine déjà connue.
Selon Jack et al., (1995), les trois lettres initiales des bactériocines doivent
être utilisées pour nommer les gènes de l’opéron comme par exemple nis pour les
gènes de la nisine : nis A de la structure, nis I d’immunité, T, E, F et G : gène de
transport ; P gène codant pour la protéase, B, C, D et M : gènes de modification post-
traductionnelle ; R, K et Q : gènes régulateurs.
12
2.3.3. Bactériocines et antibiotiques
La confusion entre bactériocines et antibiotiques dans la littérature, a
contribué à limiter leur utilisation dans les applications alimentaires. Il est donc
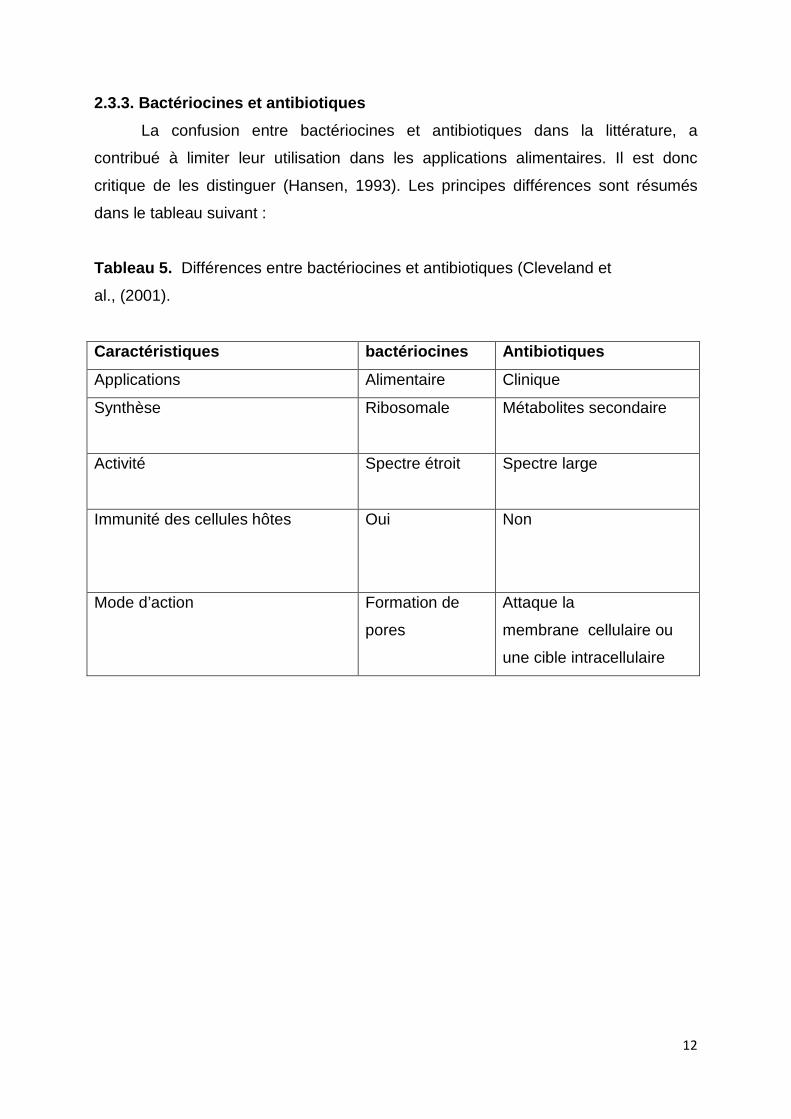
critique de les distinguer (Hansen, 1993). Les principes différences sont résumés
dans le tableau suivant :
Tableau 5. Différences entre bactériocines et antibiotiques (Cleveland et
al., (2001).
Caractéristiques bactériocines Antibiotiques
Applications Alimentaire Clinique
Synthèse Ribosomale
Métabolites secondaire
Activité Spectre étroit
Spectre large
Immunité des cellules hôtes Oui
Non
Mode d’action Formation de
pores
Attaque la
membrane cellulaire ou
une cible intracellulaire

13
2.3.4. Classification des bactériocines
Les bactériocines produites par les bactéries lactiques sont réparties en
quatre classes, comme proposé par Klaenhammer (1993). Ces quatre classes sont :
Classe I : Les lantibiotiques : peptides de taille inférieure à 5 kDa, stables à la
chaleur et qui contiennent des acides aminés inhabituels soufrés formés
postraductionnellement, c’est acides aminés sont : la lanthionine, la β-méthyl
lanthionine, la déhydrobutyrine et la déhydroalanine. Ils peuvent être divisés en deux
types :
Sous-Classe Ia : Cette classe comprend des peptides cationiques hydrophobes
allongés contenant jusqu’à 34 acides aminés.
Sous-Classe Ib : Elle comprend les peptides globulaires chargés négativement ou
sans charge nette et contenant jusqu’à 19 acides aminés (Mc Auliffe et al., 2001;
Twomey et al., 2002). Certains lantibiotiques sont par ailleurs constitués de deux
peptides agissant ensemble pour avoir une activité comme la lacticin 3147. Les
séquences et les structures d’un lantibiotique de chaque type se trouvent dans la
figure1.
Classe II : Peptides de taille inférieure à 10 kDa, stables à la chaleur, ne contenant
pas d’acides aminés modifiés. Cette classe est divisée en trois sous-classes :
Sous-Classe IIa : Contient entre 27 et 48 acides aminés et une partie N-terminale
hydrophobe contenant la séquence consensus YGNGV ainsi qu’un pont disulfure. La
partie C-terminale est moins conservée avec un caractère hydrophobe (Fimland et
al., 2000 ; Richard, 2006). Elles ont toutes une activité contre Listeria
monocytogenes. Certaines bactériocines de cette sous-classe contiennent
également un deuxième pont disulfure dans leur domaine C-terminale qui semble
être important dans la stabilisation de la structure tertiaire (Eijsink et al., 1998 ;
Fimland, 2000 ; Drider, 2006 ; Richard, 2006).
Sous-Classe IIb : Comprend les bactériocines ayant besoin de deux peptides pour
avoir une activité.
Sous-Classe IIc : Contient les bactériocines ne pouvant pas être classées dans les
autres sous-classes.
Classe III : Leurs protéines sont de tailles supérieures à 30 kDa et sensibles à la
chaleur. La structure et le mode d’action de ces bactériocines diffèrent complètement
des autres bactériocines produites par les bactéries lactiques. Cette classe ne
regroupe que quatre bactériocines : l’helveticin J produite par Lactobacillus

14
helveticus A, l’enterolysin A produite par Enterococcus faecium, la zoocin A produite
par Spreptococcus zooepidemicus et la millericin B produite par Streptococcus milleri
(Nilsen et al., 2003 ; Papagianni, 2003 ; Nigutova et Paik, 2007).
Classe IV : Peptides requérant une partie carbohydratée ou lipidique pour avoir une
activité. Aucune bactériocine de cette classe n’a été décrite.
Figure 1 : séquence et structure de lantibiotiques de type A (Nisine), B (Mersacidine)
et d’un lantibiotique « two-peptides (Lacticine 3147 A1 et A2) respectivement.
nisine mersacidi ne
Lactacine A1 LactacineA 2

15
2.3.5. Génétique des bactériocines
L’utilité des bactériocines dans les domaines d’application pratique notamment
comme additifs alimentaires a poussé les chercheurs à caractériser les éléments
génétiques impliqués dans leurs synthèses (Jack et al., 1995).
La biosynthèse des bactériocines nécessite plusieurs gènes organisés en
opéron, comportant les gènes de structure, d’immunité, de translocation, de
maturation et de régulation. Cependant le support génétique on diffère.
Le déterminant génétique des bactériocines des bactéries à Gram positives
est fréquemment plasmidique (Klaenhammer, 1993). En effet de nombreux
arguments sont en faveur d’une localisation plasmidique des opérons codant pour
les bactériocines dont on peut citer les études de curage plasmidique et la
comparaison des profils des souches productrices et des souches non productrices
(Stoddard et al., 1992).
Staphylococcus epidermis, productrice de la PeP5, possède au moins cinq
plasmides. Des oligonucléotides spécifique à PeP5 sont capables de s’hybrider au
plasmide PED503 mais pas à l’ADN chromosomique et aux quatre autres plasmides
(Kaletta et Entian, 1989). En plus le séquençage d’une région du plasmide en
question montre qu’il contient le gène de structure PeP5 et les autres gènes liés qui
forment probablement un opéron.
Les études de curage plasmidique réalisées sur Bacillus coagulans I4
génèrent des dérivés non producteurs. Ceci suggère une localisation plasmidique
des gènes de production de la coaguline (Le Merrec et al., 2000). La Lactococcine et
la lactacine 481 sont codées respectivementbpar des plasmides de 55 kb et 70 kb
(Piard et al., 1993 ; Klaenhammer, 1993).
D’autre bactériocines sont codées par des gènes localisés sur l’ADN
chromosomique, comme c’est le cas de la subtiline produite par Bacillus subtilis
(Banerjec et Hansen, 1988). La mersacidine produite par Xanthomonas campresis
(Heu et al., 2001). Toutefois, le déterminant génétique des bactériocines peut être
aussi un transposant c’est le cas de la lactacine F (Muriana et Kklaenhammer, 1991)
et la nisine (Horn et al., 1991) et de la bactériocine produite par Listeria innocua 743
(Klamokoff et al., 2001).
Les déterminants génétiques responsables de la production des bactériocines
sont en général groupés sous forme de plusieurs unités transcriptionnelles arrangées
en opérons. Dans la plupart des cas, le premier gène de cette organisation

16
polycistronique code pour la protéine structurale. Les produits des gènes associés
sont impliqués dans la régulation de la transcription, les modifications post-
traductionnelles, le transport de la protéine vers l’extérieur de la cellule et l’immunité
de la bactérie productrice.
Le nombre de gène dans chaque opéron diffère d’une bactériocine à une
autre. L’opéron de la coaguline comprend quatre gènes :coa A, coa B, coa C et coa
D qui codent respectivement la pré-coaguline et les protéines d’immunités, de
transport et de clivage du précurseur (Le Merrec et al., 2000). L’organisation de la
nisine est plus compliquée, son opéron comprend 11 gènes : nis A de structure, nis B
de maturation post-traductionnelle, nisT, nis C de transport, nis P de clivage du
précurseur et les gènes d’immunité nisI, nisR, nisK, nisF, nisE et nisG (Siergers et
Etian, 1995), Figure 2.

17
Coa A B C D
Opéron coaguline
Nis B A T C I P R K F E G
Opéron nisine
Protéines de structure Modification post-traductionnelle
Clivage du prépeptide
Protéine d’immunité
Transport du prépeptide
Figure 2 : représentation schématique des opérons de la coaguline (Le Merrec et
al., 2000) et de la nisine (Siergers et Etian, 1995).

18
2.3.6. Biosynthèse et excrétion
Bien que les bactériocines sont rébosomiquement synthétisées, le transcrit qui
en résulte doit être modifier pour devenir actif. Les gènes qui codent pour les
enzymes facilitant les modifications sont généralement à proximité des gènes de
structure. La modification la plus importante est connue chez les lantibiotiques
(Cleveland et al., 2001). Lan B, protéines membranaire transcrite par les producteurs
de la nisine, modifie enzymatiquement ces bactériocines après leur transport vers
l’extérieur de la cellule (Engelke et al., 1992). Lan C participe aussi dans la formation
d’un pont thio-ester au niveau de ces lantibiotiques (Kleerebezem et Ouadri, 2001).
La modification post-traductionnelle des lantibiotiques incluse la formation de
nombreux acides aminés inhabituels.
La formation de ces acides aminés ait lieu après l’addition des groupes
cystéines-thiol. En tout, plus d’une douzaine d’acide aminés inhabituels sont fondus
au niveau des lantibiotiques.
Le prépeptide des non lantibiotiques est aussi modifié par clivage de la
séquence leader (Diep et al., 1996 ; Ehrmann et al., 2000). Ces modifications sont
nécessaires pour la sécrétion et le transport à travers la membrane cellulaire.
Les précurseurs peptidiques des non lantibiotiques comme la pediocine AcH
(Bukhtiyarova et al., 1994) et le lantibiotique Pep 5 (Sahl et al., 1987 ; Weil et al.,
1990) sont isolés et détectés sur gel. Ceci supporte la règle générale qui dit que les
bactériocines de faible poids moléculaire des bactéries a gram positives sont
synthétisées ribosomiquement sous forme de précurseur en premier lieu. Ces
précurseur apparaissent non biologiquement actifs et contiennent un propeptide C-
terminal qui, après avoir subit plusieurs modifications post-traductionnelles, est clivé
de la séquence N-terminale pour donner une molécule antimicrobienne mature (de
Vos et al., 1993).
Malgré les détails fournis par les évènements post-traductionnels ayant lieu
durant la maturation des non lantibiotiques, plusieurs phénomènes restent mal
connus. Pour cette raison un modèle pour la pediocine Ach a été proposé
(Bukhtiyarova et al., 1994).
Ce modèle est basé sur des observations de pediocine AcH prédit que
puisque la pédiocine active déstabilise probablement la membrane plasmique, la
translocation doit avoir lieu sous forme de pré-pediocine. L’endopeptidase impliqué

19
dans la séparation du peptide leader du propeptide au niveau d’un cite de clivage
spécifique est désigné peptidase leader (Jack et al., 1995).
Peu d’information concernant les propriétés de ou de(s) enzyme(s)
impliquée(s) dans la transformation de la prépediocine en pediocine AcH. L’enzyme
semble être mieux activée à un pH = 5 ou plus (Biswas et al., 1991 ; Ray et al., 1993)
alors que son activité est inhibée par un inhibiteur acide et par le β-mercaptoethanol
qui rompe les ponts disulfure (Ray et al., 1993).
Des études récentes montre que les fonctions de translocation
transmembranaires et l’activité peptidase sont portées par la même protéine
(Bukhtiyarova et al., 1994). Une étude séparée apporte que cette protéine a
uniquement un rôle de transport et possède une homologie structurale avec des
protéines ATPasiques transporteuses à travers les membranes (MarugG et al.,
1992).
La plupart des protéines de la classe I et II sont secrétées à l’extérieur des
cellules par un système transporteur ABC. Ce transporteur est formé par un domaine
hydrophobe intégré dans la membrane, d’un domaine de fixation d’ATP du coté C-
terminal situé à la face interne de la membrane plasmique et d’un domaine
protéolytique du coté N-terminal responsable du clivage de la séquence leader.
L’unique exception est fournie par une faible proportion de la classe II qui est
externalisée par la voie sec-dépendante (séquence leader N-terminale de type sec)
(Clevelande et al., 2001).
Le substrat Nis A est le prépeptide non biologiquement actif qui sera
déshydraté par Nis B et cyclisé par Nis C avant sa translocation par l’ABC
transporteur Nis T est le responsable de clivage de la séquence signal par la
protéase Nis P. Ces modifications conduiront au peptide biologiquement actif. La
nisine interagira avec l’histidine kinase Nis K, ce qui induira la phosphorylation du
régulateur de réponse Nis R et l’activation de la transcription des gènes nécessaires
à la production de la nisine. La protection de la cellule vis-à-vis de la nisine est
réalisée par deux mécanismes : la lipoprotéine d’immunité Nis I et l’ABC transporteur
formé par Nis G, Nis E et Nis F
2.3.7. Mode d’action
L’étude du mode d’action comporte deux aspects distincts :
-La cinétique des interactions physiques, entre les bactériocines et les cellules
sensibles, caractérisée par l’adsorption de la bactériocines.

20
-L’étude des liaisons biochimiques spécifiques sur les cellules qui se
manifestent par la perte de l’intégrité cellulaire et par conséquence le relargage de
certains composés intracellulaire (Atrich, 1994).
Deux types d’interaction sont connus lors de la fixation de la bactériocine sur
les membranes des cellules cibles (i) une interaction directe dépendante du potentiel
membranaire avec les structures phospholipidiques de la membrane cible. Il en
résulte une désorganisation membranaire. Ce type d’interaction est connu chez les
lantibiotiques comme la nisine (Moll et al., 1999). (ii) une interaction avec des
récepteurs protéiques spécifiques, c’est le cas des non lantibiotiques (Ennahar et al.,
2000). Exemple la pediocine AcH produite par Pediococcus acidilactis H (Bhunia et
al., 1991).
L’activité bactéricide des non lantibiotiques contre les cellules sensibles est
produite principalement par la déstabilisation de la fonction membranaire. Ces
derniers peuvent aussi cibler des composants intracellulaires comme l’ADN, l’ARN et
les enzymes pour tuer leurs cellules sensibles (Martinez et al., 2000).
Les bactériocines sont des molécules chargées positivement, ayant des
portions hydrophobes qui interagissent électrostatiquement avec des groupements
phosphates chargés négativement des membranes des cellules cibles (Chen et al.,
1997).
L’interaction de la nisine avec les composants membranaires des cellules
cibles est considérée comme une étape cruciale dans leur mode d’action, le degré de
cette association dépend du type de lipide en présence. Il a été démontré que
l’activité in vitro de la nisine, qui est de nature cationique, est plus efficace en
présence d’un haut pourcentage de composants lipidiques anionique (Garcia Gacera
et al., 1993).
Les lantibiotiques agissent par la formation de pores au niveau des cellules
cibles provoquant une dissipation des potentiels transmembranaires et/ou du
gradient du pH ce qui résulte une libération du matériel intracellulaire. Des études
ultérieures suggèrent que la formation de pores par la nisine dépend du potentiel
membranaire et du gradient du pH des cellules cibles ( Montville et Bruno, 1994).
La fixation des bactériocines sur les cellules cibles est facilitée par la
présence des molécules ancrées au niveau de la membrane qui augmente l’efficacité
de l’action de la bactériocine. Le lipide II est une molécule ancré au niveau de la
membrane plasmique jouant le rôle de peptidoglycane. La fixation de la mersadicine

21
sur cette molécule inhibe la synthèse des peptidoglycanes (Breukink et al., 1999 ;
Brotz et al., 1998).
Les lantibiotiques et les non lantibiotiques affectent tous les deux la barrière
de la perméabilité membranaire par la formation de canaux ou pores.
2.3.8. Immunité de la bactérie productrice
Les bactéries productrices pouvant être sensibles à leur propre bactériocine,
elles se préimunissent à l'aide d'une protéine qualifiée «d'immunité» (Abee, 1995).
Le seul modèle élaboré pour les bactéries lactiques a été très récemment proposé
par Venema et al (1995).
La protéine d'immunité, une protéine possédant un large domaine
transmembranaire pourrait interagir avec le récepteur potentiel de la bactériocine et
empêcherait ainsi l'insertion de cette dernière dans la membrane. Alternativement ou
de façon complémentaire, la protéine d'immunité pourrait également interagir
directement avec la bactériocine. la protéine d'immunité a été trouvée de façon
largement majoritaire dans le cytoplasme (Ouadri et al., 1995).
2.3.9. Purification des bactériocines
Puisque les bactériocines sont secrétées dans le milieu de culture, la
purification de ces molécules actives est un moyen indispensable à leur
caractérisation. En effet, la plupart des approches de purification commencent par la
concentration de surnageant de culture comme la précipitation aux sels (Holo et al.,
1991 ; Jeorjer et Klaenhammer, 1986), aux acides (Hasting et al., 1991) la
concentration par aspiration ou encore l’extraction par les acides organiques (Oscariz
et Pisabarro, 2000).
Ces produits bien que nécessaire pour la réduction du volume à traiter ne
donne pas en pratique un haut degré de purification. Par conséquent de nombreux
étapes de chromatographies sont nécessaires pour avoir une purification significative
(Vennema et al., 1997).
Certaines méthodes de purification se basent sur la nature hydrophobe de la
bactériocine comme la cereine 7 produite par Bc 7 qui est purifiée par une méthode
d’extraction par le butanol (Oscariz et Bisabarro, 2000).

22
La purification partielle de l’entomocine 9 produite par B thuringiensis ssp.
entomocidus HD9 consiste d’abord a précipiter les protéines au sulfate d’ammonium,
l’échantillon obtenu est en suite soumis à une filtration sur gel de sephacryl suivie
d’une chromatographie échangeuse d’anions (Cherif et al., 2003).
Vennema et ces collaborateurs propose une méthode de purification rapide et
efficace pour les bactériocines de faible poids moléculaire hydrophobes et
cationiques tel que la lactococcin B et la pediocine PA-1. Ces deux bactériocines
sont purifiées par une précipitation par l’éthanol, une dissolution dans l’eau
déminéralisée puis une isoélectrofocalisation suivie d’une ultrafiltration (Vennema et
al., 1997).
L’adsorption puis la désorption des bactériocines sur les bactéries
productrices, en fonction de pH, constituant un moyen rapide de purification initiale
de ces molécules. Cette procédures a été reprise au cours de la purification partielle
de la bacillocine 490 produite par Bacillus licheniformis 490 /5 (Maritirani et al.,
2002).
Malgré la mise en évidence ces derniers années de plusieurs bactériocines,
leur purification complète reste limitée à certains cas. Les difficultés rencontrées lors
des purifications résident dans :
(i) Les milieux de culture des bactéries sont généralement complexes et
comportent une concentration élevée de peptides et des protéines comparées à celle
de bactériocines produites qui ne présentent qu’environ 1mg par litre de culture
(Rammelsberg et al., 1990).
(ii) Les bactériocines forment un groupe chimique qui semble hétérogène
d’où la nécessité de tester plusieurs technique de purification (Van Laak et al., 1992).
Certains détergents utilisés dans les milieux de culture comme le Tween 80
(dans le milieu MRS) interfèrent avec la précipitation de ces molécules en raison de
leur caractère hydrophobe. Ce phénomène a été signalé par Muriana et
Klaenhammer (1991). Ces auteurs ont noté que le film formé à la surface lors de la
précipitation de la lactacin F avec le sulfate d’ammonium, contient 97% de l’activité.
(iii) La tendance des bactériocines à s’associer entre elles ou avec d’autres
protéines nécessite souvent l’utilisation d’agent dissociant tel que l’urée ou le SDS
(Muriana et Klaenhammer, 1991).

23
Il est ainsi recommandé de quantifier l’activité spécifique des bactériocines à
chaque étape de purification et de modifier ou d’éliminer les étapes conduisant à des
pertes excessives (tagg et al., 1988).
2.3.10. Rôle écologique des bactériocines
Les bactériocines sont produites lorsque la densité cellulaire est élevée et les
réserves alimentaires se trouvent rares. Ceci est démontré par Chao et Levin, (1981)
qui ont montré que les conditions d’invasion des souches productrices de colicine
sont plus large dans un environnement structuré ou les réserves alimentaires sont
faibles tel qu’une boite de Pétri que dans un environnement non structuré.
Le spectre d’activité, centré généralement sur des espèces rapprochées
phylogénétiquement, suggère un rôle des bactériocines dans l’écologie microbienne.
Les bactériocines servent comme des « anti-concurrents » permettant
l’invasion des souches productrices au sein d’une communauté microbienne. Elles
ont aussi un rôle défensif en interdisant l’envahissement des autres souches ou
espèces dans leur niche écologique ou encore en limitant l’avancée des souches
voisines (Riley et al., 2002).
La régulation de l’expression des bactériocines peut être enfin incluse dans le
processus de « quorum-sensing ». En effet, toute une panoplie de gènes codant
pour les activités de pathogénicité et autre, est régulée par la densité cellulaire ce qui
représente un avantage écologique.
La cereine 7 produite par la souche Bc7 est représentée en large partie sous
forme d’agrégats dans une solution aqueuse. Ceci indique que les agrégats sont
mieux maintenus par des interactions hydrophobes. Ces agrégats ont pour rôle de
prévenir la diffusion et la perte d’antibiotiques et son maintien tout autour de la
population bactérienne productrice (Oscariz et Pissabarro, 2000).
2.3.11. L’application des bactériocines dans le sec teur alimentaire
Les bactériocines purifiées sont considérée comme un additif alimentaire.
Jusqu’à présent, seule la nisine, un lantibiotique, est acceptée comme additif
alimentaire (E234) (Guinane et al., 2005).
Les bactériocines peuvent également être appliquées sous la forme d’un
concentré obtenu après fermentation par la souche productrice.

24
Cette préparation sera considérée comme un ingrédient fermenté. Elle
contiendra la bactériocine mais également d’autres métabolites microbiens tels que
l’acide lactique. La pédiocine, une bactériocine de la classe IIa, est commercialisée
sous cette forme sous le nom ALTA 2341. Des essais ont été récemment fait avec la
lacticine 3147, un lantibiotique (Deegan et al., 2006 ; Galvez et al., 2007).
Un autre mode d’application des bactériocines consiste en leur immobilisation
sur les cellules productrices, dans des gels ou des films telle que la gélatine, la
cellulose, les protéines de soja, des films de polysaccharides, etc. La bactériocine
sera alors libérée dans le produit au cours de la conservation (Luchansky et al., 2004
; Deegan et al., 2006 ; Ghalfi, 2006 ; Galvez et al., 2007).

27
3. Matériel et méthodes

28
3.1. Matériel biologique et conditions de culture
3.1.1. Laits
L’échantillon de lait de chamelle étudié provient de la région de Béchar. Le
trayon est lavé soigneusement à l’eau tiède. Les premiers jets sont éliminés, puis 100
ml de lait sont recueillis dans des flacons stériles qui sont immédiatement étiquetés et
placés dans la glacière (4°C) pour être rapidement tran sporté au laboratoire pour
l’analyse microbiologique et l’isolement de bactéries lactiques.
Une autre partie des souches étudiées proviennent d’une collection de
bactéries lactiques du laboratoire de microbiologie appliquée. Nous avons utilisé
également des souches pathogènes de références pour évaluer la substance
antibactérienne proviennent du laboratoire central d’analyse du CHU d’Oran.
3.1.2. Milieux et conditions de culture
Les milieux de culture de base utilisés sont des milieux M17 (Terzaghi et
Sandine, 1975) soit liquides, soit solides par addition d’agar (0.7%) pour les géloses
molles et 1.6% pour les milieux solides décrits par la Fédération Internationale du Lait
(FIL, 1981). Le lait écrémé stérile additionné de 1% d’extrait de levure est utilisé pour
les cultures bactériennes. L’incubation des bactéries lactiques se fait à 30° C pendant
48h.
3.2. Méthodes
3.2.1. Isolement des lactocoques
L’échantillon de lait est mis dans un tube à raison de 10 ml puis incubé à 30°C
jusqu’à coagulation par les bactéries lactiques endogènes.
Les ensemencements sont réalisés sur milieu M17 (Terzaghi et Sandine, 1975)
en utilisant les dilutions décimales obtenues à partir de la suspension mère.
100 µl sont ensemencés sur milieu solide pour avoir des colonies bien
séparées. Après incubation (30°C, 24 à 48h), un examen microscopique est effectué,
il permet d’observer la morphologie des cellules, leurs tailles, éventuellement leur
mode de regroupement. Il peut être fais soit à l’état frais ou après coloration simple ou
coloration de Gram sur frottis fixé. La coloration de Gram nous permet une séparation

29
entre deux grands groupes : les bactéries à Gram positives des bactéries à Gram
négatives.
Cette différence de coloration provienne de différences structurales de la paroi
bactérienne. Les bactéries à Gram positives, à paroi rigide gardant la coloration
violette du violet de gentiane après l’action de l’alcool, alors que les bactéries à Gram
négatives, à paroi plus mince, perdent cette coloration après action de l’alcool et sont
recolorées en rose par la fuchsine. Ce critère est l’un des bases de la classification
des bactéries.
Après coloration de Gram. La forme des cellules et leur mode d’association
sont notés.
3.2.2. Recherche de la catalase
Cette méthode est basée sur la réaction suivante :
Catalase
2H2O2 2 H2O2 +1/2O2
La recherche de la catalase est effectuée par addition d’une goutte de
peroxyde d’oxygène à une colonie étalée sur une lame. Le dégagement de bulles
d’air indique que l’isolat est catalase positive; dans le cas contraire, la souche est
catalase négative.
Les isolats à Gram+ et catalase- sont repiqués de façon alternée sur milieu M17
liquide ou solide jusqu’à purification. A chaque fois, 7 ou 10 colonies bien isolées sont
prélevées du milieu M17 solide et transférées sur M17 liquide et vice versa. La pureté
de la souche est vérifiée par une observation microscopique; l’aspect des colonies
(forme, couleur, taille) et l’aspect caractéristique de la culture des bactéries lactiques
en milieu liquide.
3.2.3. Identification des souches purifiées
Afin de classer les isolats en genre, en espèce et en sous espèces, les tests
biochimique s’avère nécessaires pour mener à bien cette tache.
a- Test de l’arginine dihydrolase (ADH)
La mise en évidence de cette enzyme est intéressent pour la caractérisation
des bactéries lactiques. Le rôle de cette enzyme est de libérer l’ammoniac à partir de

30
l’arginine. Pour effectuer ce test on utilise le milieu M16 BCP de Thomas (1973), ce
milieu contient un indicateur de pH, le pourpre de bromocrésol, si la couleur du milieu
vire au jaune puis vers le violet ceci indique la présence de l’enzyme et la dégradation
de l’arginine. Si elle reste jaune cela veut dire que la bactérie est ADH négative.
b- Le type fermentaire
Ce test permet d’apprécier le type de métabolisme par le quel le substrat
carboné est transformé ; Par définition, l’hétérofermentation est la capacité des
bactéries lactiques à produire des molécules différentes du lactate telles que le CO2
l’acétate et l’éthanol à partir de la dégradation des sucres. Il consiste à mettre en
évidence le gaz (CO2) produit.
L’ensemencement des souches à tester s’effectue dans un milieu liquide M17
contenant le glucose comme source de carbone, menu au préalable d’une cloche de
Durham. Le bouillon est, par la suite, mis à incuber à 30°C, les tubes sont observés
dans un délai de 3 ou 5 jours en fonction de l’aspect du milieu (trouble) et la
production du gaz. (Garvie, 1984; Schillinger et Lücke, 1989).
c- Température de croissance
C’est un critère important car il permet de distinguer entre les souches
mésophiles des thermophiles. Après inoculation en milieu M17 liquide avec une
culture pure, les tubes sont incubés pendant sept jours à 10°C et 24 à 48 heures à
45°C.
qLa thermorésistance des souches est testée par leur incubation en bain marie
pendant 30mn à 63-65°C (Stiles et Holtzapfel, 1997; Klein et al., 1998). En suite, les
tubes sont portés à incuber pendant 24 à 48h. Au bout de ces délais, la croissance
est appréciée par l’examen des milieux (présence de trouble).
d- Croissance à 4%, 6,5% de NaCl et à pH 9,6
L’habilité à croître sur milieu M17 en présence de NaCl à différentes
concentrations (4% et 6.5%) et à pH de 9.6 a été observé pendant 2 à 3 jours
d’incubation à 30°C. Les lactocoques ne poussent pas en pr ésence de 6.5% de NaCl;
ce test permet de séparer les lactocoques (streptocoques lactiques), des
entérocoques (Devriese et al., 1993).

31
e- Test de lait bleu de Shermann
Une série de tubes à essais de 7ml de lait stérilisé à 0,1% et à 0,3% de bleu de
méthylène est ensemencé par des cultures pures puis incubés durant une période de
24 à 48 heures à 30°C. Seuls les entérocoques peuvent se développer dans le lait à
0,3% de bleu de méthylène.
f- Fermentation des sucres
La fermentation de 14 sucres a été testée pour l’ensemble des souches qui on
donné un effet inhibiteur sur les souches pathogènes utilisées a fin de les identifier. Le
milieu de base utilisé est le bouillon MRS (sans le glucose et l’extrait de viande)
additionné de pourpre de bromocrésol comme indicateur de pH (MRS-BCP) (Mannu
et al., 2000) supplémenté de 1% des sucres suivants: mannitol(1), saccharose(2),
lévulose(3), sucrose(4), rhamnose(5), galactose(6), xylose(7), esculine(8), sorbitol(9),
arabinose(10), maltose(11), fructose(12), lactose(13) glucose(14) et raffinose(15).
Les solutions de sucres ont été préparées dans 10 ml d’eau distillée contenant
3 g des différents sucres. Ces préparations sont stérilisées par chauffage au bain
Marie à 100°C pendant 10 min puis conservées au réfrigé rateur (Joffin et Leyral,
1996). Ce test est résumé dans les étapes suivantes :
(i) Dans des tubes à hémolyse stériles, on mit 1ml de MRS BCP.
(ii) On récupère les culots après centrifugation des cultures jeunes des
souches étudiées après lavage par le tampon phosphate.
(iii) On ajoute 0,1ml de chaque sucre dans les tubes précédents.
(iv) On ensemence le contenu des tubes avec 0,1ml des souches et en
dernier on y ajoute une goutte d’huile de paraffine stérile pour l’anaérobiose.
(v) L’incubation se fait à 30°C pendant 24 à 48 h.
Le virage de la couleur du milieu vers le jaune indique que la souche a dégradé le
sucre (résultat positif), si non le résultat est négatif.
3.2.3. Conservation des souches
La conservation des souches pures a été faite selon deuxméthodes: à court et
à long terme.
La conservation à court terme des souches pures est effectuée sur milieu M17
solide incliné. Après croissance à la température optimale, les cultures sont

32
maintenues à +4°C et leur renouvellement se fait par repiquage toutes les 4
semaines. A long terme, les souches sont conservées dans une solution contenant
70% de lait écrémé (enrichie par 0.5g/l d'extrait de levure et 0.5g/l de glucose) et 30%
de glycérol à 40% à -20°C (Samelis et al., 1994).
3.2.4. Cinétique d’acidification
L’acide lactique produit par les souches isolées est mesuré à l’aide d’un pH
mètre puis titré par le NaOH N/9 en présence d’un indicateur de pH (la
phénolphtaléine 1%). Cette acidité est exprimée en degré Dornic (D°). Un degré
Dornic correspond à l’acidité apporté par 0,1g d’acide lactique dans un litre de lait.
Chaque souche est ensemencée dans un tube à 10 ml de lait écrémé stérile
puis incubés à 30°C pendant 24h. Dés que le lait coagule , on mesure le pH et on note
la souche la plus acidifiante, la moins acidifiante et qui acidifié le lait moyennement.
Dans 3 flacons contenant 100 ml de lait écrémé stérile, on transfère 3ml de lait
coagulé de chaque tube sélectionnés puis on homogénéise. Chaque flacon est réparti
dans des tubes stériles à raison de 10ml de lait par tube puis en mis dans l’incubateur.
La quantité d’acide lactique est calculée après différents temps (2h, 4h, 6h, 8h, 18h et
24h).
3.2.5. Antagonisme direct sur milieu solide
Il s’agit de détecter des zones d’inhibition après Co-culture entre les souches à
tester avec une souche indicatrice. Le test est effectué selon la méthode directe (Paik
et al ., 1997). Les souches à tester sont ensemencées avec un multipoint sur milieu
M17 tamponné pour éviter l’effet de l’acidité, après incubation à 30°C, 7ml de gélose
molle inoculée par 45 µl de la souche pathogène (E.Coli et Staphylococcus pyogenes
séparément) sont étalés en surcouche puis incubés 24h à 30°C. Les boites sont
examinées s’il ya présence ou non de zones d’inhibition. La présence d’une zone
d’inhibition correspond à un arrêt de la multiplication de la souche indicatrice dans la
partie de la gélose contenant une (des) substance(s) antimicrobienne(s) produite(s)
par la souche testée. Le diamètre des zones d’inhibition est ensuite mesuré.
3.2.6. Détection de la substance inhibitrice dans l e surnageant de culture
La méthode utilisée est celle de la diffusion de surnagent de culture à partir des
puits décrite par Tagg et Mc Given, (1971). Les souches qui on donné des zones

33
d’inhibition sur milieu solide sont cultivées dans le milieu M17 liquide à 30°C. Après
24h, les cultures sont collectées puis centrifugées à 5000 tpm pendant 7 mn pour
récupérer les surnagent. Cinq ml de gélose molle préalablement inoculés par 45 µl de
la souche indicatrice sont ajoutés en surcouche dans une boite de Pétri contenant le
milieu M17 solide. Après solidification, des puits de 10mm de diamètre sont creusés
dans la gélose et leurs fonds scellés par 20 µl de gélose molle liquéfiée à 45°C. Deux
cent µl de chaque surnagent de culture à tester sont placé dans chaque puits.
Les boites sont laissées 30mn dans la température ambiante pour permettre la
diffusion de surnagent dans la gélose molle. Les diamètres des zones d’inhibition sont
mesurés après 24h d’incubation à 30°C.
3.2.7. Caractérisation de la substance inhibitrice
a- Action des protéases
Afin de déterminer la nature chimique de la substance inhibitrice secrétée par
les souches actives, le surnagent de culture de ces souches est traité par les
enzymes suivants : la trypsine et la chymotrypsine à raison de 1mg par ml de
surnagent, puis incubés 1h à 37°C. L’activité résiduelle des surnagent traités est
testée par la méthode de diffusion à partir des puits.
b- Stabilité thermique
L’effet de la température sur l’agent antimicrobien est étudié de la manière
suivante : 1ml de chaque surnagent est placé dans un tube Eppendorfs puis incubés
pendant 30 mn à 120°C. L’activité résiduelle est testé e par la méthode de diffusion à
partir des puits en utilisant le surnagent natif comme témoin.

34
4. Résultats et discussion

35
Etude morphologique
4.1. Résultat des tests macroscopiques
Les observations macroscopiques nous a permis de décrire les colonies
obtenues sur milieu solide (la taille, la couleur, la forme et le contour). Le repiquage
successif sur milieu M17 solide a révélé des colonies rondes avec une couleur
blanchâtre et d’un pourtour régulier (Figure 3(A)). .
En milieu liquide, elle se base sur l’observation du trouble fumeux surmonté
d’une zone claire. Il s’accentue au fur et à mesure de la purification des souches, ceci
traduit le caractère microaerophile des bactéries lactiques (Figure 3(B)). .
4.2 . Etude microscopique
La caractérisation microscopique est basée sur la coloration de Gram. Nous
n’avons gardé que les bactéries à Gram positives et en forme de coque (Figure 5),
leur mode d’association est résumé dans le tableau 6.

36
A B
Figure 3 : aspect macroscopique des colonies en surface (A). Aspect d’une culture
pure dans un milieu liquide M17(B).
Figure 4 : aspect microscopique à l’agrandissement G1000 après coloration de Gram
de la souche 11B.
37
Tableau 6: aspect macroscopique des colonies et l’aspect microscopique des
souches purifiées et leurs caractères biochimiques.
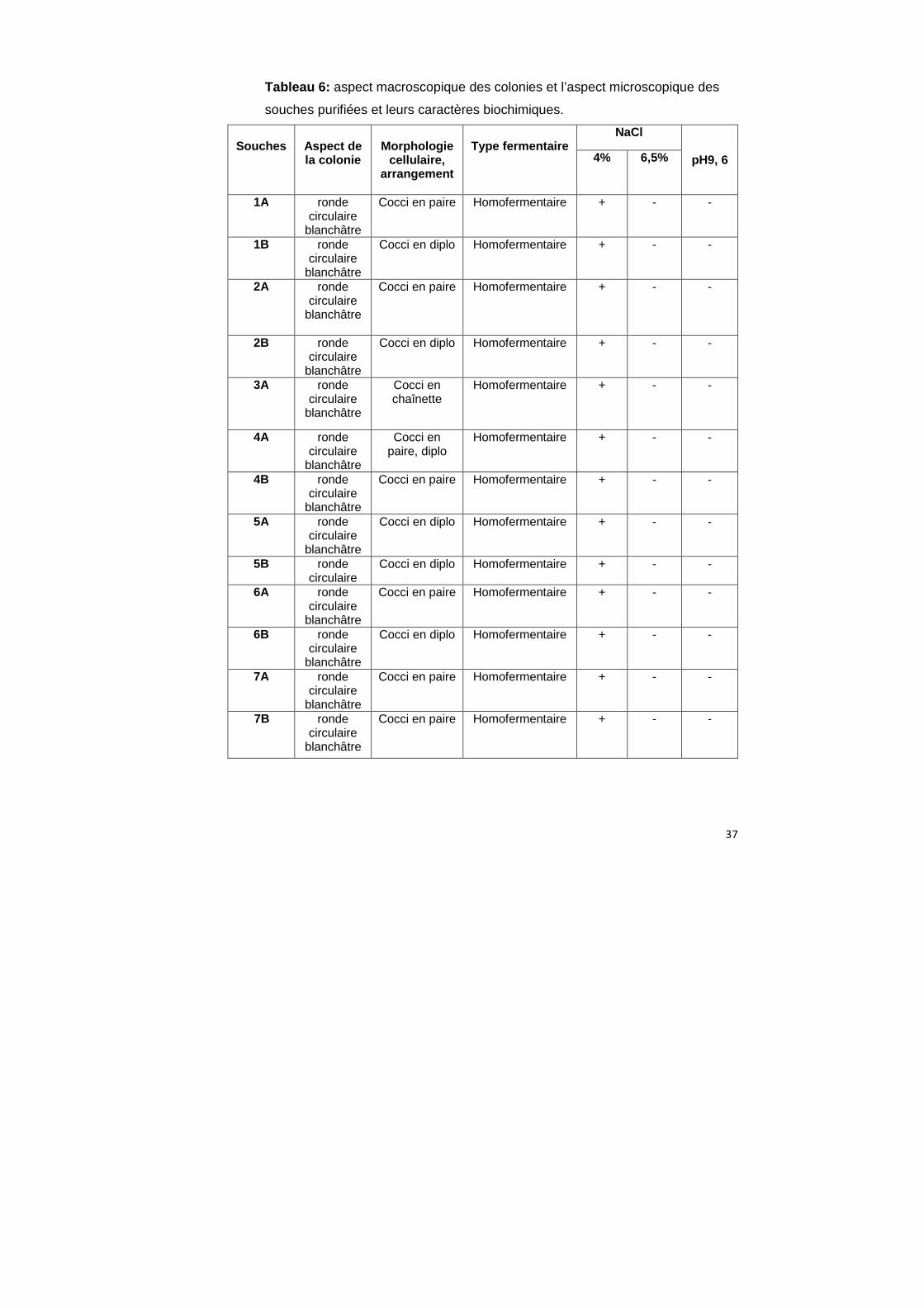
Souches
Aspect de la colonie
Morphologie
cellulaire, arrangement
Type fermentaire
NaCl
pH9, 6 4% 6,5%
1A ronde circulaire
blanchâtre
Cocci en paire Homofermentaire + - -
1B ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
2A
ronde circulaire
blanchâtre
Cocci en paire Homofermentaire + - -
2B ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
3A ronde circulaire
blanchâtre
Cocci en chaînette
Homofermentaire + - -
4A ronde circulaire
blanchâtre
Cocci en paire, diplo
Homofermentaire + - -
4B ronde circulaire
blanchâtre
Cocci en paire Homofermentaire + - -
5A ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
5B ronde circulaire
Cocci en diplo Homofermentaire + - -
6A ronde circulaire
blanchâtre
Cocci en paire Homofermentaire + - -
6B ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
7A ronde circulaire
blanchâtre
Cocci en paire Homofermentaire + - -
7B ronde circulaire
blanchâtre
Cocci en paire Homofermentaire + - -
38
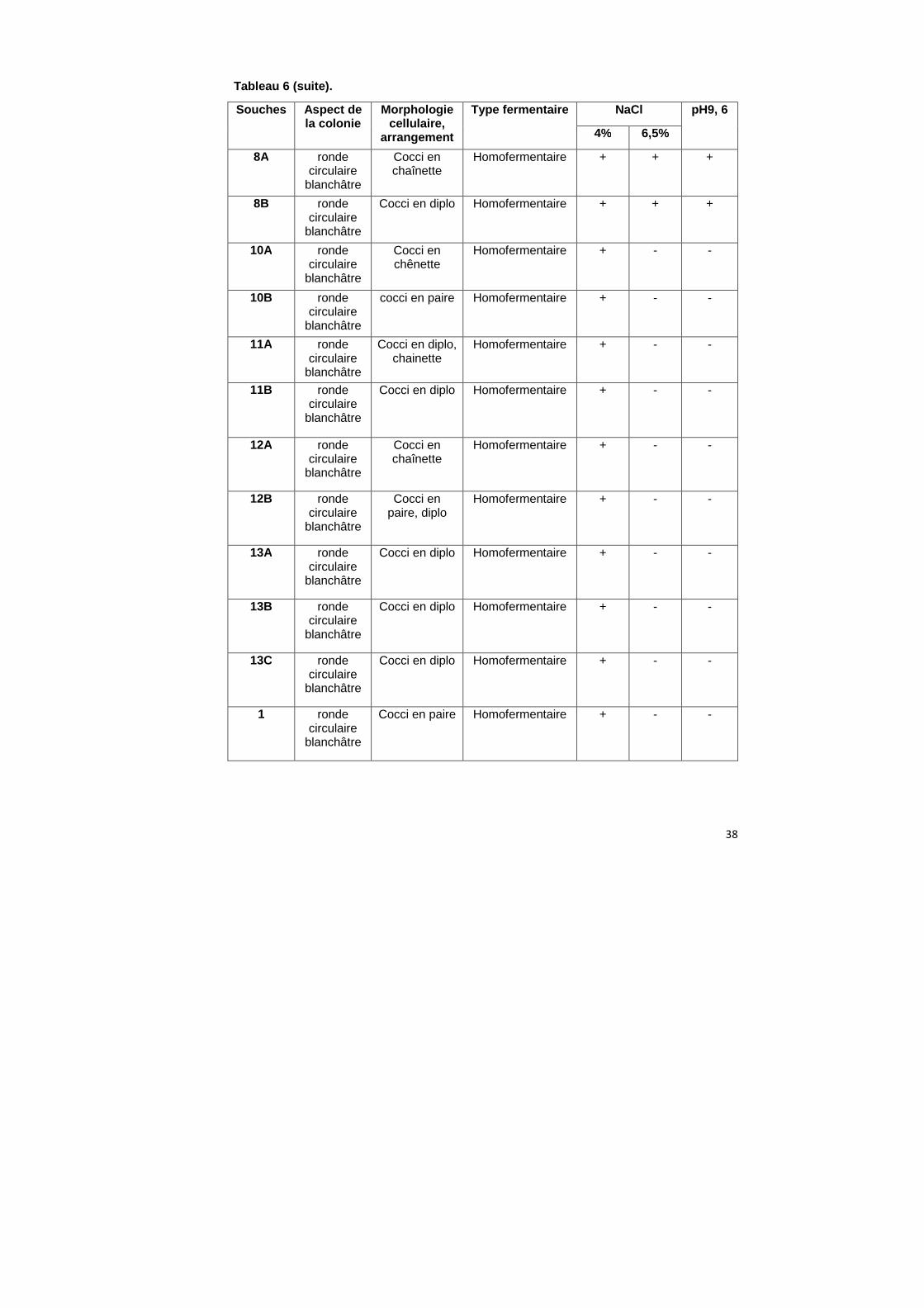
Tableau 6 (suite).
Souches
Aspect de la colonie
Morphologie cellulaire,
arrangement
Type fermentaire NaCl pH9, 6
4% 6,5%
8A
ronde circulaire
blanchâtre
Cocci en chaînette
Homofermentaire + + +
8B ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + + +
10A ronde circulaire
blanchâtre
Cocci en chênette
Homofermentaire + - -
10B ronde circulaire
blanchâtre
cocci en paire Homofermentaire + - -
11A ronde circulaire
blanchâtre
Cocci en diplo, chainette
Homofermentaire + - -
11B ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
12A ronde circulaire
blanchâtre
Cocci en chaînette
Homofermentaire + - -
12B ronde circulaire
blanchâtre
Cocci en paire, diplo
Homofermentaire + - -
13A ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
13B ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
13C ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
1 ronde circulaire
blanchâtre
Cocci en paire Homofermentaire + - -

39
Tableau 6 (suite).
Souches
Aspect de la colonie
Morphologie cellulaire,
arrangement
Type fermentaire NaCl Ph9,6
4% 6,5%
2
ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
3
ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
4 ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
5
ronde circulaire
blanchâtre
Cocci en diplo Homofermentaire + - -
6
ronde
circulaire blanchâtre
Cocci en chaînette
Homofermentaire
+
-
-

40
4.3. Test de catalase
La recherche de la catalase se fait par la mise en contact des colonies avec
quelques gouttes d’eau oxygénée à 10V. Le dégagement gazeux traduit l’activité
positive de cette enzyme. Lorsqu’on a un milieu liquide, on ajoute 1ml d’H2O2 à 3% à
une culture de 18 à 24h. Nous avons éliminé toutes les souches catalase positives.
.
4.4. Test de NaCl et de pH
Ce test est réalisé dans le but de différencier les lactocoques des
entérocoques, Il nécessite l’emploi de deux milieux M17 liquide : l’un contenant
respectivement 6,5% et 4% de NaCl et l’autre à un pH 9,6 ; dans lesquels on a
ensemencé les souches à tester. Après culture, nous avons remarqué que toutes les
souches testées n’ont pas pu pousser en présence de 6,5% de NaCl et dans le milieu
M17 à pH 9,6 sauf la souche 8A et 8B.
4.5. Test du lait bleu de shermann
Nous avons ensemencé les souches isolées dans deux séries de tubes : la
première avec un pourcentage de 1% en bleu de méthylène et l’autre avec 3% de
bleu de méthylène. Le principe de ce test est basé sur le mode respiratoire des
souches ensemencées, si elles sont aérobies (cas des entérocoques) elles vont tirés
leurs oxygène depuis le bleu de méthylène présent dans le lait, donc ce dernier va
perdre sa couleur, si c’est le cas contraire, cas des lactocoques qui sont
microaérophiles, elles vont utiliser une petite quantité d’oxygène qui ne peut pas
changer la couleur du lait donc il reste bleu. La figure 5 montre la souche 8A qui a
réduit le bleu de méthylène 3%, ceci signifie que cette souche est une entérocoque.
La souche 5B a donné une réaction négative (bleu de méthylène à 3%). Les souches
10B, 10A et 7B on donné une réaction négative avec le bleu de méthylène 1%.

41
8A (+) 5B(-) 10B(-) 10A(-) 7B(-)
3% en bleu de méthylène 1% en ble u de méthylène
Figure 5 : résultat du test de bleu de méthylène.
(+) : réduction du bleu de méthylène.
(-) : résultat négatif.

42
4.6. Test du profil fermentaire
Dans des tubes à essais, menus de cloches de Durham et contenant le milieu
M17 pH 7,1, on a ensemencé les souches à tester. Les souches hétérofermentaires
vont produire, en plus de l’acide lactique, l’acide acétique et le CO2. Ce gaz libéré va
être mis en évidence par la poussée de la cloche de Durham vers le haut. Nous avons
remarqué que toutes les souches étudiées sont homofermentaire (Tableau 6).
4.7. Test de l’arginine déshydrogénase
La production de l’acide lactique acidifie le milieu de culture (M16 BCP), qui
contient un indicateur de pH, et la couleur de ce dernier va virer vers le jaune. Les
bactéries possédant l’ADH vont realcalinisé le milieu et sa couleur reviendra mauve,
les souches qui ne possèdent pas cette enzyme leur milieu va rester jaune.
Les souches testées sont ensemencées par un multipoint, après incubation à
30°C, nous avons remarqué que les isolats 6A, 6B et 6 n ’ont pas une arginine
déshydrogénase alors que les autres sont ADH positives (Figure 6).
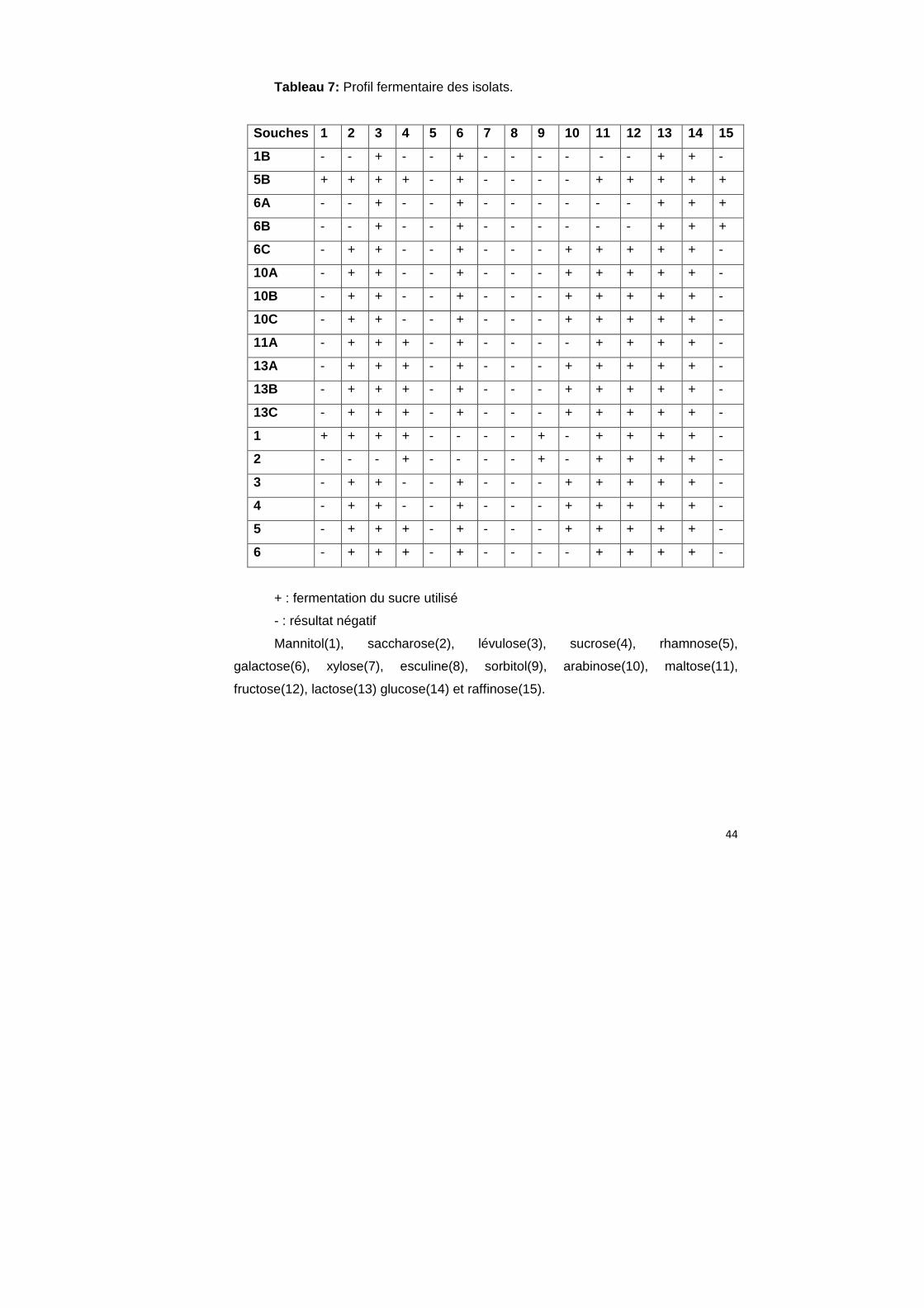
4.8. Test de fermentation des sucres
La fermentation des sucres est un test utilisé afin d’identifier les espèces.
Seules les souches qui on un effet inhibiteur sont identifiées. Les sucres utilisés sont :
mannitol(1), saccharose(2), lévulose(3), sucrose(4), rhamnose(5), galactose(6),
xylose(7), esculine(8), sorbitol(9), arabinose(10), maltose(11), fructose(12),
lactose(13) glucose(14) et raffinose(15). Les résultats obtenus sont résumés dans le
tableau 7.

43
Figure 6 : aspect des souches ADH positives sur milieu M16 BCP
Présence d’une couleur mauve indique la présence de l’enzyme de l’ADH
Souches ensemencées
en multipoint
44
Tableau 7: Profil fermentaire des isolats.
Souches 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
1B - - + - - + - - - - - - + + -
5B + + + + - + - - - - + + + + +
6A - - + - - + - - - - - - + + +
6B - - + - - + - - - - - - + + +
6C - + + - - + - - - + + + + + -
10A - + + - - + - - - + + + + + -
10B - + + - - + - - - + + + + + -
10C - + + - - + - - - + + + + + -
11A - + + + - + - - - - + + + + -
13A - + + + - + - - - + + + + + -
13B - + + + - + - - - + + + + + -
13C - + + + - + - - - + + + + + -
1 + + + + - - - - + - + + + + -
2 - - - + - - - - + - + + + + -
3 - + + - - + - - - + + + + + -
4 - + + - - + - - - + + + + + -
5 - + + + - + - - - + + + + + -
6 - + + + - + - - - - + + + + -
+ : fermentation du sucre utilisé
- : résultat négatif
Mannitol(1), saccharose(2), lévulose(3), sucrose(4), rhamnose(5),
galactose(6), xylose(7), esculine(8), sorbitol(9), arabinose(10), maltose(11),
fructose(12), lactose(13) glucose(14) et raffinose(15).

45
15 14 13 12 11 10 9 8 7 6 5 4 3 2 1 T
A
15 14 13 12 11 10 9 8 7 6 5 4 3 2 1 T
B
Figure 7 : résultat du test des sucres.
A : souche 1B,
B : souche 11A.
T : témoin (MRS BCP sans sucre)
Mannitol(1), saccharose(2), lévulose(3), sucrose(4), rhamnose(5), galactose(6),
xylose(7), esculine(8), sorbitol(9), arabinose(10), maltose(11), fructose(12),
lactose(13) glucose(14) et raffinose(15).

46
Tableau 8: présentation de l’identification des souches isolées.
Souches Espèce
1B Lc. cremoris
6A Lc. cremoris
6B Lc .cremoris
6C Lc. lactis
10A Lc. lactis
10B Lc. lactis
10C Lc .lactis
11A Lc. hordiniaea
13A Lc. lactis
13B Lc. lactis
13C Lc. lactis
2 Lc. lactis
3 Lc. lactis
4 Lc. lactis
5 Lc. garviaea
6 Lc. cremoris

47
4.9. Cinétique d’acidification
L’acidification est le rôle principal des bactéries utilisées comme ferments.
Celle- ci a différents buts:
- la coagulation du lait en facilitant l’action de l’enzyme de la présure et
l’augmentation de la synérèse du caillé ;
- la participation aux propriétés rhéologiques du produit final ;
- l’inhibition de la croissance des bactéries nuisibles (Weber et Broich, 1986).
La nature du lait a une influence sur la croissance et l’activité acidifiante des
souches (Casalta et al.1995).
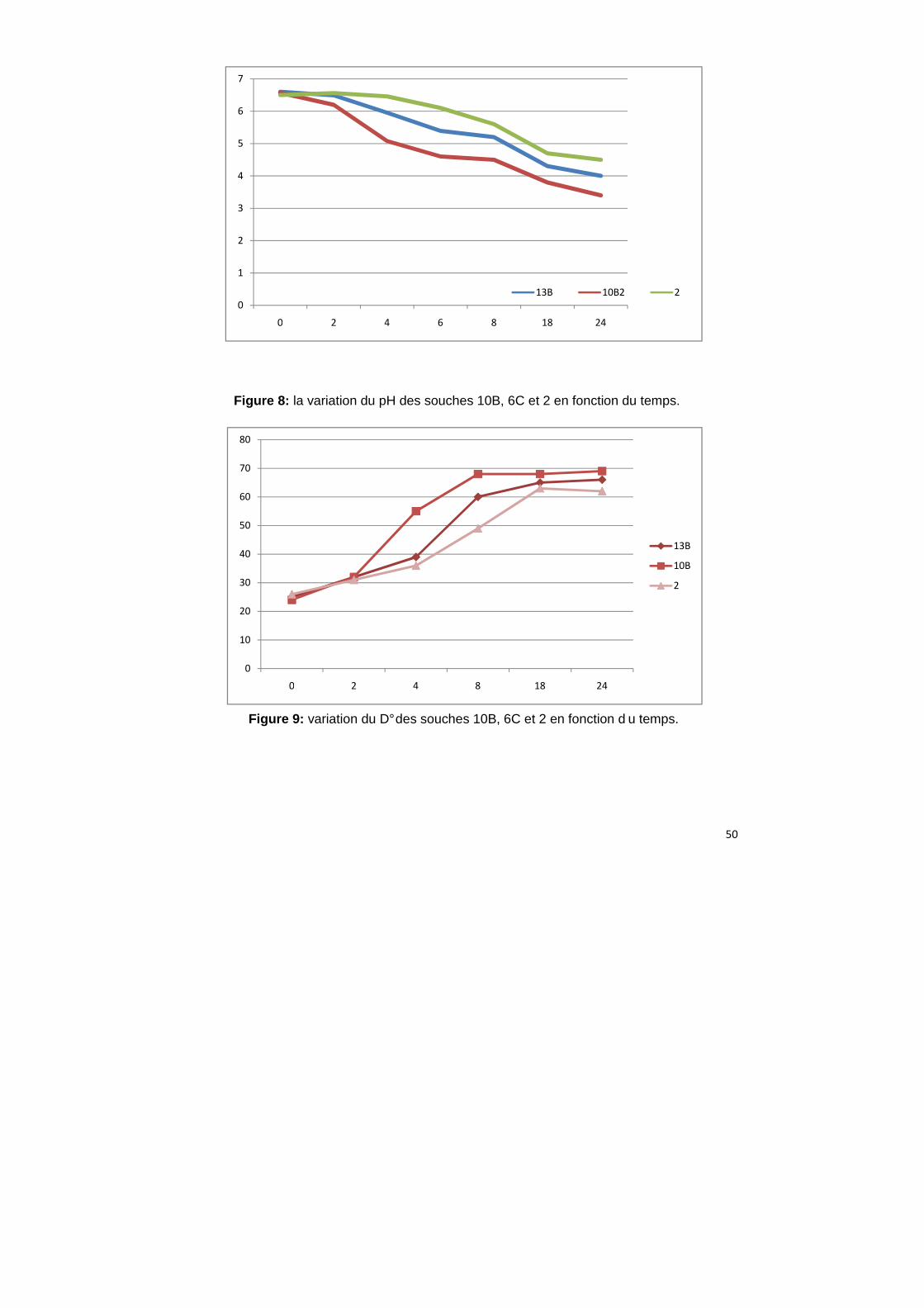
La mesure de l’activité acidifiante à 30°C montre que la nature du lait a une
influence sur la vitesse d’acidification de Lactococcus lactis (Casalta et al.1995).
Parmi les bactéries isolées à partir de lait cru de chamelle, on a choisi 3
souches : 10B qui est fortement acidifiante, la souche 13B qui est moyennement
acidifiante et la souche 2 faiblement acidifiante afin de suivre leur cinétique
d’acidification, le pH a diminué progressivement en fonction du temps (Figure 8,
Tableau 9). Par contre le degré Dornic a augmenté à cause de l’augmentation de
l’acide lactique présent dans le milieu de culture (Figure 9).
En effet, Les ferments présentant une capacité d’acidification élevée sont
adaptés à la fabrication de produits laitiers fermentés, les autres ferments, à capacité
acidifiante plus faible, semblent plus adaptés à la fabrication fermière.
4.10. Test d’antagonisme sur milieu solide
Dans le but de détecter les souches productrices de substance(s) inhibitrice(s),
nous avons réalisé une Co-culture sur milieu solide entre chaque souche et les
souches pathogènes (E. coli, Staphylococcus aureus) qui ont été isolés et identifiés à
partir de produits biologiques contaminés au Laboratoire central d’analyses médicales
de CHU d’Oran. Après 24h, nous avons sélectionné que les souches ayant un effet
inhibiteur sur la croissance des souches pathogènes, cet effet est traduit par
l’apparition des zones claires autours des colonies (Figure 10).

48
En général, le diamètre d’inhibition est plus large lorsqu’on utilise Sc. aureus comme
bactérie indicatrice, alors que les isolats 1, 4, 5 et 6 ont donné une activité plus élevé
sur les deux bactéries pathogènes.

49
Tableau 9: la variation du pH et du degré Dornic des souches 10B, 6C et 2 en
fonction du temps.
10B 6C 2
pH V °D pH V °D pH V °D
0h 6,6 2,5 25 6,57 2,4 24 6,5 2,6 26
2h 6,49 3,2 32 6,2 3,8 38 6,56 3,1 31
4h 5,95 3,9 39 5,08 5,5 55 6,46 3,6 36
6h 5,39 4,8 48 4,6 4,3 43 6,1 4 4
8h 5,2 6 60 4,5 6,8 68 5,6 4,9 49
18h 4,5 6,5 65 4,2 6,8 68 4,7 6,3 63
24h 4,4 6,6 66 4 6,9 69 4,6 6,5 65
D° : degré dornic
V : volume du NaOH coulé
50
Figure 8: la variation du pH des souches 10B, 6C et 2 en fonction du temps.
Figure 9: variation du D° des souches 10B, 6C et 2 en fonction d u temps.
0
1
2
3
4
5
6
7
0 2 4 6 8 18 24
13B 10B2 2
0
10
20
30
40
50
60
70
80
0 2 4 8 18 24
13B
10B
2

51
zone d’inhibition
Figure 10: test d’antagonisme direct sur milieu solide.
52
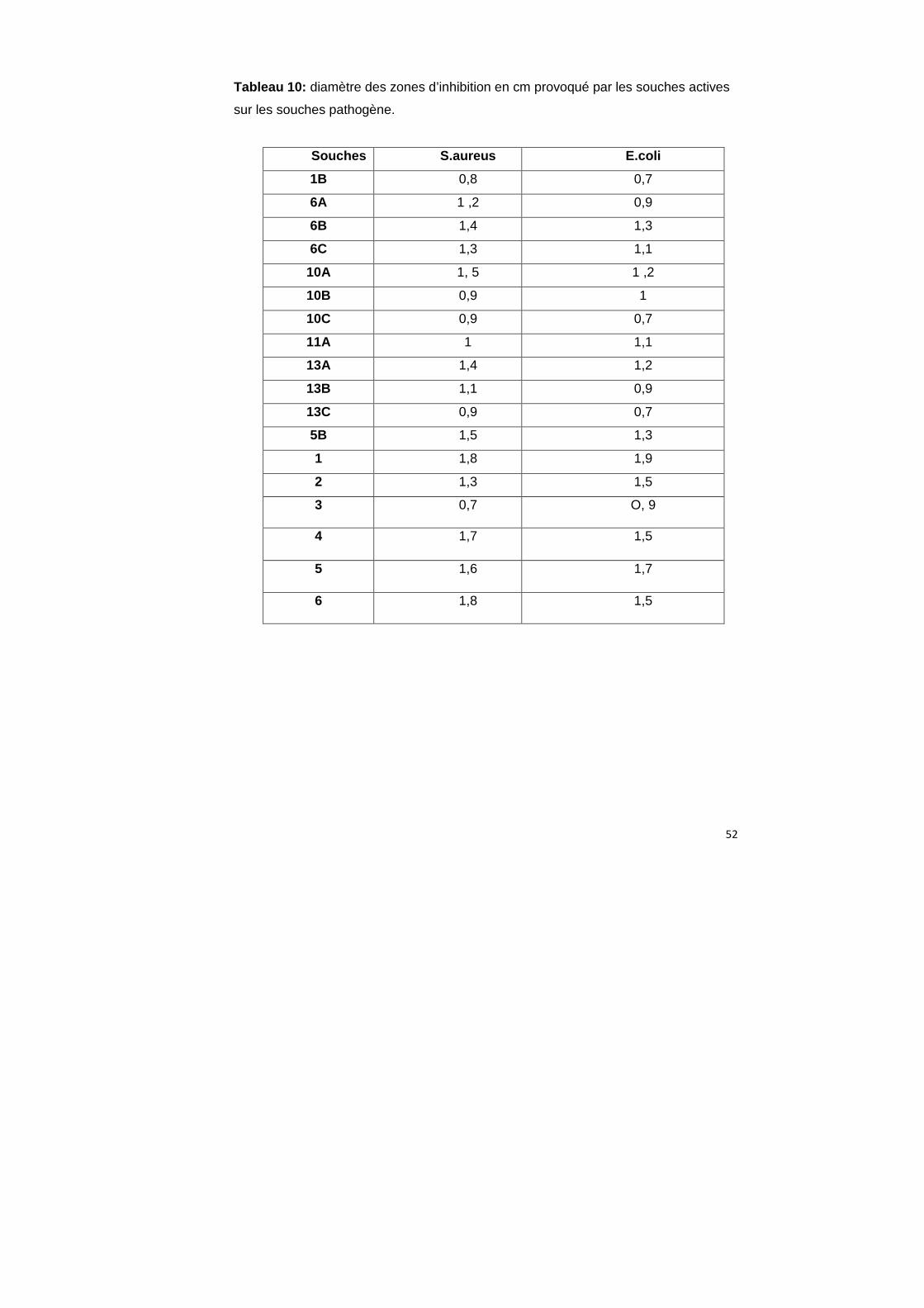
Tableau 10: diamètre des zones d’inhibition en cm provoqué par les souches actives
sur les souches pathogène.
Souches S.aureus E.coli
1B 0,8 0,7
6A 1 ,2 0,9
6B 1,4 1,3
6C 1,3 1,1
10A 1, 5 1 ,2
10B 0,9 1
10C 0,9 0,7
11A 1 1,1
13A 1,4 1,2
13B 1,1 0,9
13C 0,9 0,7
5B 1,5 1,3
1 1,8 1,9
2 1,3 1,5
3 0,7 O, 9
4 1,7 1,5
5 1,6 1,7
6 1,8 1,5
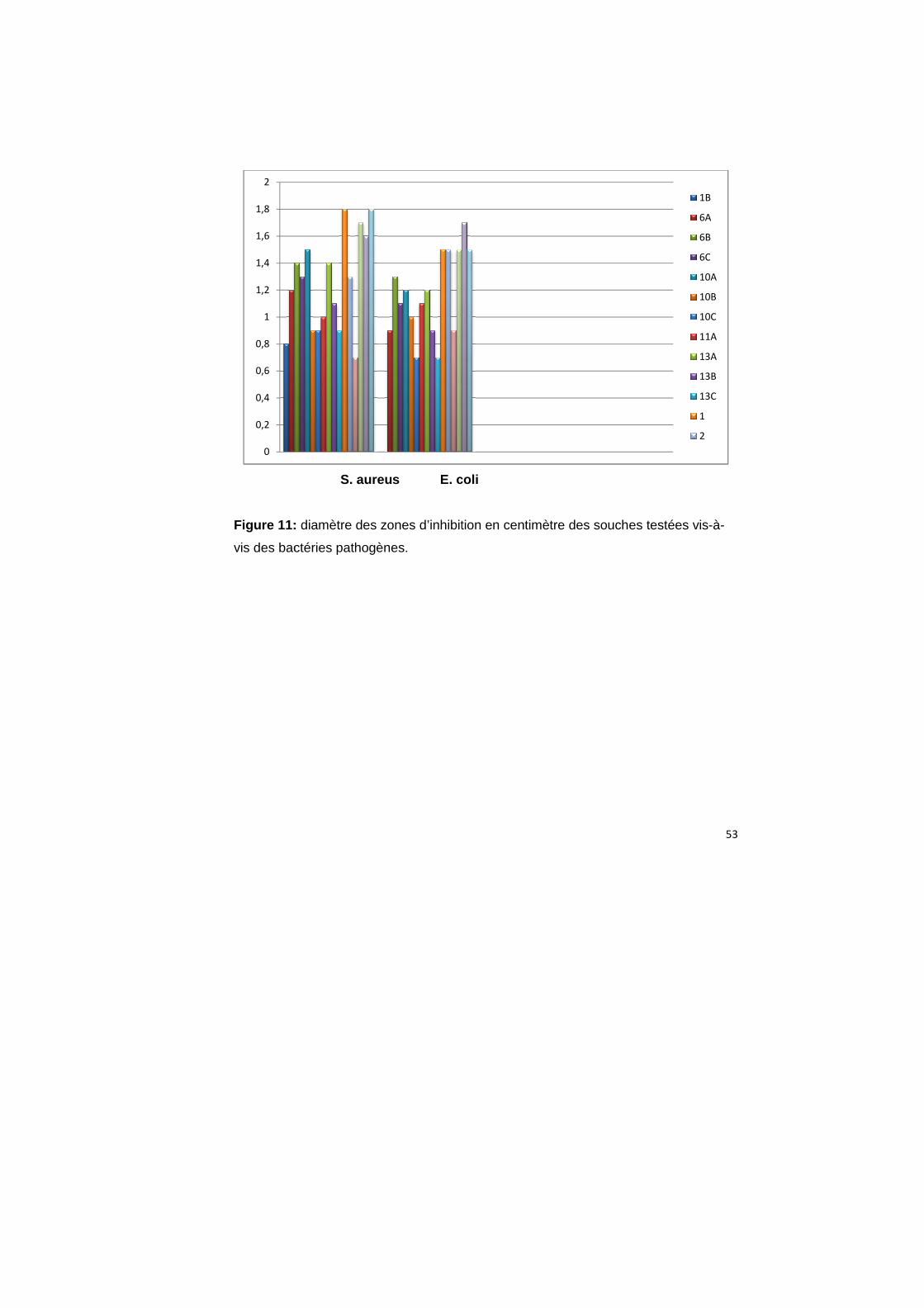
Figure 11 :
vis des bactéries
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
S. aureus E. coli
: diamètre des zones d’inhibition en centimètre des souches testées vis
bactéries pathogènes.
coli
d’inhibition en centimètre des souches testées vis
53
d’inhibition en centimètre des souches testées vis-à-
1B
6A
6B
6C
10A
10B
10C
11A
13A
13B
13C
1
2

54
4.11. Recherche de l’activité inhibitrice dans le s urnageant de culture
L’agent inhibiteur a été recherché dans le surnagent de culture. Après 24h,
nous avons collecté les différents milieux de culture puis sont centrifugés et testés par
la méthode de diffusion à partir des puits. Après un délai de 24h on a remarqué que
l’activité inhibitrices des souches est présente dans leur surnagent, ce qui confirme
que la substance inhibitrice est secrétée à l’extérieur de la cellule (Figure 12).
4.12. Identification des agents inhibiteurs des sou ches testées
Pour être sûr que la zone d’inhibition remarquée après le test de diffusion à
partir des puits est due à une bactériocine, nous avons effectué plusieurs tests qui
sont : l’effet de la température, de la trypsine et de la chymotrypsine sur le surnagent
de culture des différents souches. Après incubation nous avons remarqué que
l’activité inhibitrice a disparue, on peut dire que l’activité inhibitrice est due à une
substance de nature protéique et on peut dire que se sont des protéines de type
bactériocine-like.
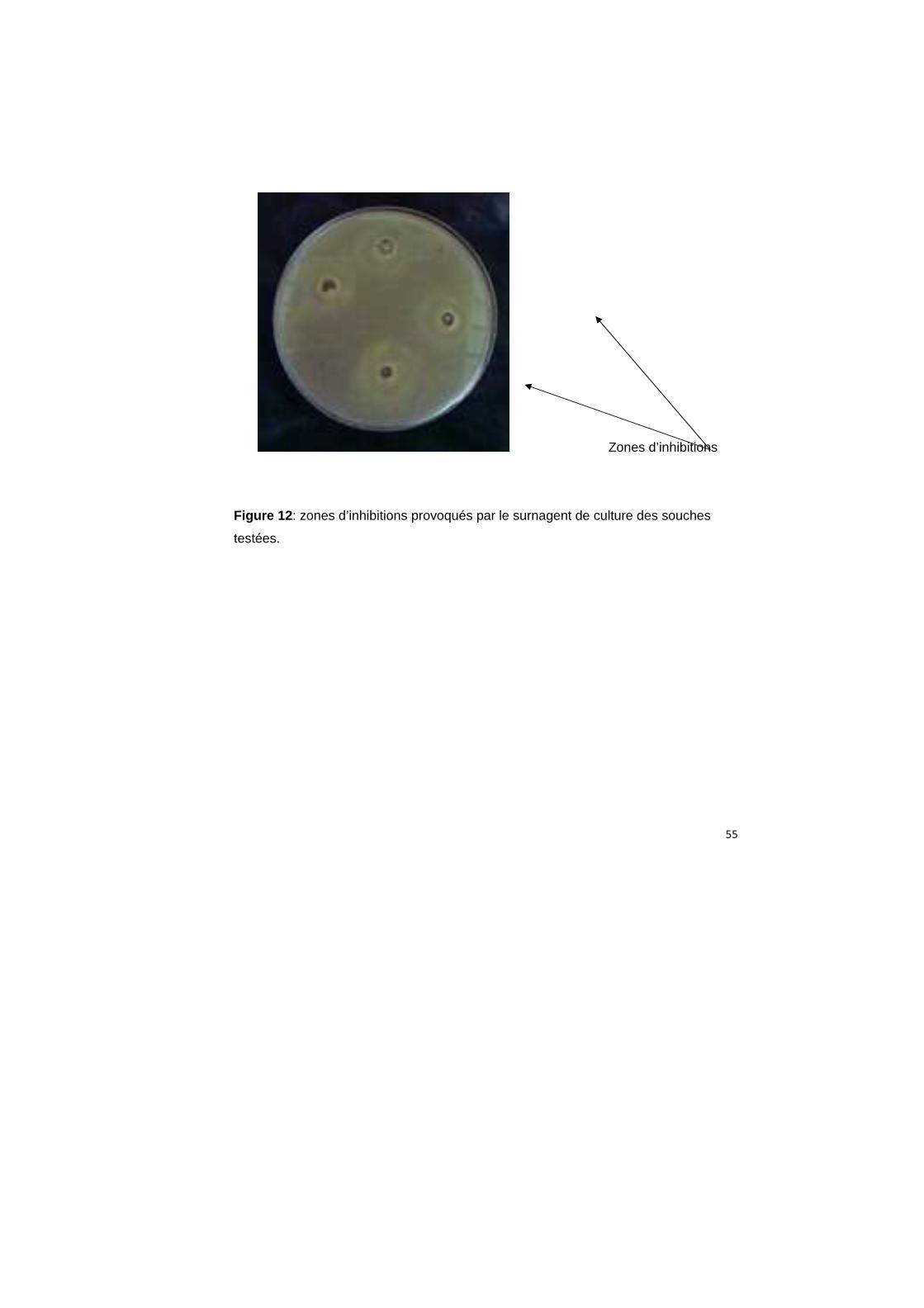
55
Zones d’inhibitions
Figure 12 : zones d’inhibitions provoqués par le surnagent de culture des souches
testées.

56
5. Discussion et Perspectives

57
Les laits algériens pourraient constituer une source intéressante d’isolement de
bactéries lactiques pourvues de propriétés industrielles importantes et en plus
possédant une biodiversité de l'information génétique.
L’identification phénotipique des bactéries nous permet d’identifier rapidement
et simultanément un nombre élevée des souches bactériennes et d’étudier leurs
propriétés biotechnologiques. Elle présente un pouvoir discriminant qui s’étant du
groupe à l’espèce et dans une moindre mesure à la sous-espèces et à la souche
(Curk et al.,1994).
La caractérisation phénotypique des lactocoques est peu claire (Drouault et
al., 2002), et beaucoup de souches sont mal classifiées aux niveaux du genre et de
l'espèce. Les méthodes standard employées pour la caractérisation du phénotype
donnent des réponses ambiguës avec des souches de lactocoques,
Cette caractérisation peut être erronés par:
la variation du phénotype par la présence ou l’absence d’un plasmide
codant pour des fonctions métaboliques;
la variation de la taille de l’inoculum et la durée d’incubation;
des profils différents entre des souches récemment isolées et celles qui
sont conservées depuis longtemps;
l’expression différente des caractères phénotypiques selon la
température, la composition du milieu de culture, pour cela les chercheurs,utilise la
taxonomie moléculaire.
Dans le présent travail, pour différencier les entérocoques, des lactocoques
(qui est impossibles sur milieu sélectif) nous nous somme basés sur l'incapacité des
souches de Lactococcus de se développer à pH 9,6 et en présence de 6,5% de NaCl.
Il est ainsi possible que les habitats normaux, y compris le lait cru, puissent
héberger de nouvelles souches intéressantes de lactocoques avec l'application
potentielle de produire de nouveaux produits laitiers fermentés (Wouters et al., 2002).
Parmi les lactococcus, deux sous-espèces sont principales: la sous espèce
lactis, et la sous espèce cremoris., Bien qu'elles montrent différentes caractéristiques,
elles ont plusieurs attributs biochimiques en commun (Pritchard, et Coolbear, 1993;

58
Desmazeaud et Cogan, 1996). Des souches de Lactococcus lactis subsp. cremoris
ont été obtenus à partir des échantillons de lait cru algérien (Saidi et al., 2002; Badis
et al., 2003), marocain, chinois, yougoslave et ukrainien (Samaržija et al., 2001).
Certaines substances inhibitrices ont été caractérisées comme étant des
protéines antimicrobiennes et appelées bactériocines. Les bactéries lactiques sont
connues pour leur aptitude à produire des composés antibactériens leur permettant de
se développer préférentiellement dans divers écosystèmes. Parmi les substances
synthétisées, des peptides, dénommés bactériocines qui sont produits puis exportés à
l'extérieur des cellules productrices.
Ces bactériocines font l'objet depuis une dizaine d'années d'un nombre
croissant d'études, ainsi qu'en témoigne l'abondante littérature scientifique s'y
rapportant (Jack et al., 1995). En arrière plan de ces nombreux travaux figurent divers
applications potentielles dans les produits laitiers, carnés et marins.
Chez les bactéries lactiques, touts les genres comportent des souches
productrices de bactériocines. Si jusque vers la fin des années 1980, seuls les
lactobacilles, les lactocoques et les pédiocoques avaient révélé la présence de
bactériocines (Klaenhammer, 1988), dans ces dernières années, la découverte de
nombreuses souches de Leuconostoc et de carnobactéries présentant des activités
antagonistes (Cenatiempo et al., 1995 ; Hastings et al., 1991 ; Héchard et al., 1992 ;
Felix et al., 1994). On peut considérer que la majorité de bactériocines décrites à ce
jour correspond à des Bactériocines de bactéries lactiques (Worobo et al, 1994;
Quadri et al., 1994).
Parmi les 30 souches purifiées, nous avons sélectionné 18 souches qui ont
donné un effet inhibiteur vis-à-vis les bactéries suivantes : Staphylococcus pyogène et
E. coli sur milieu solide, Par la suite, nous avons recherché l’effet inhibiteur des
souches productrice dans leurs surnagent de culture, ceci est réalisé par la méthode
de diffusion à partir des puits. Nous avons remarqué que cette activité inhibitrice est
présente dans le surnagent.
Sherwitz et al., (1986) ont montré qu’on peut s’assuré qu’il s’agit bien d’une
bactériocine si on observe la disparition des zones d’inhibition après action d’une
enzyme protéolytique, donc elles devraient être sensibles à l’action de ces enzymes

59
(Upreti et Hinsdill, 1973; Tagg et Wannamaker, 1978; Barefoot et Klaenhammer,
1983; Mehta et al., 1984; Klaenhammer, 1993).
Le surnagent des différentes souches sont traités par deux enzymes la trypsine
et la chymotrypsine. L’activité inhibitrice a disparue après ces traitements, les
mêmes résultats sont trouvés par Holo, (1991) qui on purifié la Lactococcin A, isolée à
partir de Lactococcus lactis subsp. Cremoris.
L’activités des bactériocines produites par huit souches de bactéries lactiques
isolées à partir de lait fermenté du Burkina Faso ont été perdues après traitement
avec l’α-chymotrypsine, la trypsine, et la pepsine, tandis que traitement avec de la
lipase, catalase, l'amylase n'a pas affecté l'activité de la bactériocine (Kozak et al.,
1978).
La zone d’inhibition est en générale plus large lorsque on utilise
Staphylococcus aureus car les bactéries lactiques sont surtouts actives sur les
bactéries pathogènes à Gram positives (O’sullivan et al., 2002) et que Les bactéries à
Gram positives sont généralement plus sensibles à l’effet bactéricide des bactéries
lactiques (Biswas et al., 1991).
La vitesse d’acidification est un critère important en technologie. On cherche le
plus souvent des souches dites « rapide » c’est-à-dire qui acidifie rapidement le lait.
Dans le cas des lactocoques, il a été démontré que cette propriété est étroitement liée
à la présence de plasmides dans la cellule : Lac+ (souche capable d’utiliser le
lactose), Prt+ (protéolytique) (Mc KAY, 1983 ; Mc KAY et Baldwin, 1975).
En effet, nous avons remarqué que l’activité inhibitrice et l’acidité apparaissent
très variables et sans relation apparente. Les deux souches 10B et 2 (fortement
acidifiante et faiblement acidifiante respectivement) ne présentent pas la même action
vis-à-vis des bactéries pathogènes. Ces résultats sont les même que ceux de (Labioui
et al., 2005).
On fin on peut dire que cet effet inhibiteur est du à une protéine de type
bacteriocin-like.

61
Conclusion
Les bactéries lactiques sont largement utilisées dans l’industrie agro-
alimentaire afin d’exploiter leur pouvoir acidifiant et aromatisant (fabrication de
yaourts et de fromages). Ces bactéries on la capacité de synthétiser des substances
à effet inhibiteur, ce qui a attiré l’attention des chercheurs dans les derniers années
afin de les utiliser comme bioconservateurs et d’augmenter la duré de conservation
des denrées alimentaires.
A partir de lait cru de chamelle, 30 souches sont isolées et identifiées.
L’espèce la plus dominante est Lactococcus lactis. Le test d’antagonisme direct sur
milieu solide nous a permis de mettre en valeur le pouvoir inhibiteurs des isolats
purifiés. 18 souches sont actives a l’encontre de bactéries pathogène (E. coli et de
Staphylococcus aureus), cette activité est présente dans leur surnagent de culture.
Parmis les 18 isolats, 1, 4, 5 et 6 ont présentée une activité inhibitrice plus élevée. La
sensibilité vis-à-vis des traitements thermiques et l’action des enzymes
protéolytiques nous permis de dire que ces substances sont de nature bactériocine-
like.
Des alternatives peuvent être proposées pour terminer ce travail :
- La purification partielle ou complète de ces agents inhibiteurs
- L’analyse de leurs structures chimiques
- L’identification du déterminant génétique
- L’étude de leurs effets sur les cellules cancéreuses ou bien leurs utilisations
dans le domaine de la biotechnologie.

62
References:
Abee, T., 1995. Pore-forming bacteriocins of Gram+ bacteria and self-protection
mechanisms of producer organisms. FEMS. Microbiol. Lett., 129 : 1-9.
Atrich, A., 1994. Etude des bactériocines produites par deux souches de
Lactobacillus plantarum caractérisation, purification, structure chimique et mode
d’action. Thèse de doctorat. Institut national polytechnique de Lorraine., p. 1-175.
Badis,A., Guetarni, D., Moussa-Boudjema, B., Henni, D.E., Tornadijo, M.E., et
Kihel, M., 2004. Identification of cultivable of lactic acid bacteriaisolated from algerian
raw goat’s milk and evaluation of their technologicalproperties. Food. Microbiol. 3:72-
78.
Banerjec, S. and Hansen, J.N., 1988. Structure and expression of the gene
encoding the precursor of subtilin, a small protein antibiotic. J. B. Chem., 263: 9508-
9514.
Barefoot, S.F. et Klaenhammer, T.R., 1983 . Detection and activity of lactacin B, a
bacteriocin produced by Lactobacillus acidophilus. Appl. Environ. Microbiol. 45: 1808-
1815.
Bhunia, A.K., Johnson, M.C., Ray, B. and Kalchayana nd, N. 1991 . Mode of
action of pediocin AcH from Pediococcus acidilactis H on sensitive bacterial strains.
J. APP. Bacteriol., 70: 25-33.
Biswas, S.R., Ray, B. 1991. Ifluence of grouthconditions on the production of a
bacteriocin, pediocin AcH from Pediococcus acitilactici H. App. Environ. Microbiol.,
57 : 1265-1267.
Botazzi, V., 1988. An introduction to rod-shaped lactic bacteria. Biochimie. 34:109-
113.
Breukink, E., Wiedmen, I., Van Kraaij, C., Kuipers, O.P., Sahl, H. and de Kruijff,
B., 1999. Use of the cell wall precursor lipid II by aspore forming peptide antibiotic.
Sciences., 256:159-164.
Brotz, H., Bierbaum, G., Leopard, K., Reynolds, P.E . and Sahl, H.,G. 1998. Role
of lipid-bound peptidoglycane synthetsis by targeting lipid II. Antimicrobiol. Agents
Chemother., 42 : 154-160.

63
Budde, B.B., Hornbaek, T., Jacobsen. T., Barkholt, V. and Koch, A.G., 2003.
Leuconostoc carnosum 4010 has the potential for use as protective culture for
vacum-packed meats: culture isolation, bacteriocin identification, and meat
application experiments. Int. J. Food Microbiol. 83: 171-184.
Bukhtiyarova, M. and Yang Ray, B., 1994. Analysis of pediocin AcH gene cluster
from plasmide pSMB 74 and its expression in a pediocin negative Pediococcus
acidilactici strain. App. Environ. Microbiol., 60: 3405-3408.
Callewaert, T. et De Vuyst, L., 2000. Bacteriocin production with
L. amylovorus DCE471 is improved and stabilized by fed-batch fermentation. Appl.
Environ. Microbiol., 66(2), 606-613.
Carr, F.G., Chill, D., Maida, N,. 2002 . The lactic acid bacteria: a literature survey.
Crit. Rev. Microbiol., 28: 281-370.
Cenatiempo, Y., Biet, F., Fremaux, C., Héchard, Y, Robichon, D., 1995. Whal's
new in bacteriocins of Carnobacterium, Leuconostoc and other lactic. Can. J.
Microbiol. 33 :22-27.
Chen, Y.S., Srionnual, S., Onda, T and Yanagida, F. , 1997. Effects of prebiotic
oligosaccharides and trehalose on growth and production of bacteriocins by lactic
acid bacteria. Lett. Appl. Microbiol., 45: 190-193.
Cherif, A., Chehemi, S., Limem, F., Hansen, B.M., H endriksen, N.B., Daffonchio,
D., and Boudabous, A., 2003. Detection and characterisation of the nouvel
bacteriocin entomocin 9, and safety evaluation of its producer, Bacillus thuringiensis
ssp Entomocidus HD9. J. Appl. Environ. Microbiol., 95:990-1000.
Cleveland, J., Montville, T.J., Nes, I.F., and chik indas, M.L., 2001. Bacteriocins:
safe, natural antimicrobials for food preservation. J. Food. Microbiol., 71 :1-20.
Curk, M.C., Hubert, J.C. and Bringel, F., 1994. Lactobacillus paraplantarum sp.
Nov., anew species related to Lactobacillus plantarum, Int. J. Syst. Bacteriol. 46:595-
598.
De Roissart, H.B. Sharpe, E. M., 1994. Taxonomie, métabolisme, croissance et
génétique des bactéries lactiques. Vol. Bactéries lactiques (tome 1). Edit. Lorica
(Chemain de saint-gorges). France. P: 25-204.
de Vos, W.M., Mulder, J.W.M., Siesen, R.J., Hugenho ltz, J. and Kuipers, O.,
1993. Properties of nisin Z and distributions of its gene, nis Z, in Lactococcus lactis.
App. Environ. Microbiol., 59:213-218.

64
Deegan, L.H., Cotter, P.D., Hill, C. and Ross P., 2 006. Bacteriocins: biological tools
for bio-preservation and shelf-life extension. Int. Dairy J., 16: 1058-1071.
Desmazeaud, M. et Cogan, T.M., 1996. Role of cultures in cheese repening. Int:
Cogan, T.M., Acoolas, J-P. (Eds), Dairy Starter Cultures. VCH Puplishers, Inc., New
York, pp. 207-231.
Diep, D., Salehian, Z., Holo, H. and Nes, I.F., 200 7. Common mechanisms of target
cell recognition and immunity for class II bacteriocin. Proc. Natl Acad. Sci., 104:
2384-2389.
Drider, D., 2006. The continuing story of class IIa bacteriocin. Microbiol. Mol. Biol.
Rev., 70(2), 564-582.
Drouault, S., Just, C ., Marteau, P. and Corthier, G., 2002. Oral treatment with Lactococcus
lactis expressing lipase enhances lipid digestion in pigswith induced pancreatic insufficiency.
Appl. Environ. Microbiol. 68: 3166-3168.
Ehrmann, M.A., Remiger, A., Eijsink, V.G. and Vogel , R.F., 2000. A gene cluster
encoding plantaricin beta and other bacteriocin-like peptides in Lactobacillus
plantarum TMW1., Biochim. Biophy. Acta., 1490:355-361.
Eijsink, V.G., 1998. Comparative studies of class IIa bacteriocins of lactic acid
bacteria. Appl. Environ. Microbiol., 64(9): 3275-3281.
Eijsink, V.G., 2002. Production of class II bacteriocins by lactic acid bacteria; an
example of biological warfare and communication. Antonie Van Leeuwenhoek.,
81(1-4), 639-654.
Engelke, G., Gutweski-Echel, Z., Hammelmann, M. et Entian, K.D., 1992.
Biosynthese of the lantibiotic nisin : genomic organisation and membrane localisation
of the NisB protein. Appl. Environ. Microbiol. 58: 3730-7342.
Ennahar, S., Sashihara, T., Sonomoto, K. and Ishiza ki, A., 2000. Class IIa
bacteriocins: biosynthesis, structure and activity. FEMS Microbiol. Rev., 24: 85-106.

65
Felix, J.V., Papathanasopoulos, M.A., Smith, A.A., von Holy, A. and Hastings,
J.W. 1994. Characterization of teucocin B-Tall a: a bacteriocin from Leuconostoc
cetnosum Tall a isolated from meal. Curr Microbiol. 29: 207-212.
Fimland G., 2000. A C-terminal disulfide bridge in pediocin-like bacteriocins renders
bacteriocin activity less temperature dependent and is a major determinant of the
antimicrobial spectrum. J. Bacteriol., 182: 2643-2648.
Galvez, A., Abriouel, H., Lopez, R.L. and Ben Omar, N., (2007). Bacteriocin-based
strategies for food biopreservation. Int. J. Food. Microbiol., 120(1-2): 51-70.
Garcia Gacera, M.J., Elferink, M.J.L., Dierssen, A. J.M. and Koning, W.N., 1993.
In vitro pore forming activity of the lantibiotic nisin: role of proton motive force and
lipid composition. Eur. J. Bioch., 212:417-422.
Garvie., E. I., 1986. Taxonomy and identification of bacteria important in cheese
fermented dairy products. In advances in the microbiology and biochemistry and
fermented milk. F. L. Davies., B. A.
Ghalfi, H., 2006. Bacteriocin activity by L. curvatus CWBI-B28 to inactivate Listeria
monocytogenes in cold-smoked salmon during 4 degrees C storage. J. Food
Protect., 69: 1066-1071.
Guessas, B., 2007. Les particularités métaboliques des bactéries lactiques isolées
du lait cru de chèvre dans le biocontrole de Staphylococcus aureus. Thèse de
doctorat. Université d’Oran Es-senia.
Guinane, C.M., Cotter, P.D., Hill, C. and Ross, P., 2005. A review: microbial
solutions to microbial problems; lactococcal bacteriocins for the control of undesirable
biota of food. J. Appl. Microbiol., 98: 1316-1325.
Hadadji, M., Benama, R., Saidi, N., Henni, D. and K ihel, M., 2005 . Identification of
cultivable Bifidobacterium species isolated from boast-fed infants faeces in west
Algeria. Afr. J. Biotechnol. 4(5): 422-430.
Hammes, W.P. et Hortel. C., 2006. The genera Lctobacillus and and
Carnobacterium. Chap.1. 2. 10. In procariotes. 4: 320-403.

66
Hansen, J.N., 1993. Antibiotic synthetized by post translation modification. Annu.
Rev. Microbiol., 47:535-549.
Hastings, J.W., Sailer, M., Roy, K.L., Vederas, J.C . and Stiles, M.E. 1991.
Characterisation of leucucin A.UAL 187 and cloning of the bacteriocin gene from
Leuconostoc gelidium . J. Bacteriol., 173:7491-7500.
Héchard, Y., Pelletie,r C., Cenatiempo, Y. and Frèr e, J., 2001. Analysis of
sigma(54)-dependent genes in Enterococcus faecalis: a mannose PTS permease is
involved in sensitivity to a bacteriocin, mesentericin Y105. Microbiology, 147: 1575-
1580.
Héchard, Y., Dérijard, B., Letellier, F., Cenatiemp o, Y., 1992. Characterization and
purification of mesentericin Yl05, an anti-Listeria bacteriocin from Leuconostoc
mesenteroides. J. Gen. Microbiol., 138: 2725-2731.
Heu, S., Oh, J., Kang, Y., Ryu, S., Cho, K. and Cho , M., 2001. Gly gen
cloning, expression and purification of glycinercine A produced by Xanthomonas
campestri pv. Glycines 8ra. App. Environ. Microbiol. 67: 4105-4110.
Holo, H., Nilsen, C. and Nes, I.F., 1991. Lactococcin A, a new bacteriocin from
Lactococcus lactis subsp. cremoris: isolation and characterization of the protein and
gene. J. Bacteriol., 173 :3879-3887.
Horn, N., Swindell, S., Dodd, H.M. and Gasson, M.J. , 1991. Nisin biosynthesis
genes are encodedby novel conjugative transposon. Mol. Gen. Genet., 228: 129-135.
Hugenholtz ,J., and Kleerebezem, M., 1999. Metabolic engineering of lactic acid
bacteria: overview of the approaches and results of pathway rerouting involved in
food fermentations. Curr. Opin. Biotechnol., 10(5), 492-497.
International Dairy Federation., 1981. The composition of ewe’sand goat’s milk.
Document., 140: 5-19.
Jack, R.W., Tagg, J.R and Ray, B., 1995. Bacteriocin of Gram positive bacteria.
Microbiol. Rev. 59 :171-200.
Joerger, M.C. and Klaenhammer, T.R.., 1986. Characterization and purification of
helveticin J and evidence for a chromosomally determined bacteriocin produced by
Lactobacillus he/veticus 481. J. Bacteriol.,167: 439-446.

67
Joffin, J. N. and Layral, G., 1996. Microbiologie technique. Centre regional de
documentation pédagogique d’Aquitaine Bordeaux., p :219-223.
Jeorjer 2003.
Kaletta, C., et Entian, K.D.,1989. Nisin, a peptide antibiotic : cloning and sequencing
of the nisA gene and post-translation processing of its peptide product. J. Bacteriol.
171:1597-1601.
Kihel, M., 1996. Etude de la production du dioxyde de carbone par Leuconostoc
mesenteroides élément d’application en technologie fromagère ype fromage bleu.
Thèse de docteur d’état, université d’Oran Es-senia.
Klaenhammer, T.R., 1993. Genetics of bacteriocins produced by lactic acid bacteria.
FEMS. Microbiol. Rev., 12(1-3): 39-85.
Klaenhammer, T.R., 1988. Bacteriocins of lactic acid bacteria. Biochimie. 70: 337-
349.
Klamokoff, M.L., Banerjee, S.K., Cyr, T., Hefford, M.A. and Gleesson, T., 2001.
Identification of a new plasmide-encoded sec-dependet bacteriocin produced by
Listeria innocua 743. Appl. Environ. Microbiol. 67 : 4041-4047.
Kleerebezem, M. and Ouadri, L. E., 2001. Peptide pheromone-dependent
regulation of antimicrobial peptide production in Gram positive bacteria: a case of
multicellular behavior. Peptide., 22: 1579-1596.
Kleerebezem, M., 2004. Quorum sensing control of lantibiotic production; nisin and
subtilin autoregulate their own biosynthesis. Peptides, 25: 1405-1414.
Klein, G., Pack, A., Bonaparte, C. and Reuter, G., 1998. Taxonomy and physiology
of probiotic lactic acid bacteria. Int. J. Food. Microbiol. 41: 103-125.
Kozak, W., Barddowski, J. and Dobrzanski, W.T.,1978 . Lactostrepcins acid
bacteriocins produced by lactic streptococci. J. Dairy Res. 45: 247-257.
Labioui, H., Almoualdi, L., Al Yachioui, M. et Ouhs sin, M., 2005. Sélection de
souches de bactéries lactiques antibactériennes. Bull. Soc. Pharma. Bordeaux. 144 :
273-250.

68
Le Merrec, C., Hyronimus, B., Bressollier, P. and V erneuil, B. 2000 . Biochemical
and genetic characterisation of coagulin, a new antilisterial bacteriocin in the pediocin
family of bacteriocins, produced by Bacillus coagulans 14. App. Environ. Microbiol.,
66 :5231-5220.
Lubelski, J., 2008. Biosynthesis, immunity, regulation, mode of action and
engineering of the model lantibiotic nisin. Cell. Mol. Life Sci., 65: 455-476.
Luchansky, J.B. and Call, J.E., 2004. Evaluation of nisin-coated cellulose casings
for the control of L. monocytogenes inoculated onto the surface of commercially
prepared frankfurters. J. Food Prot., 67: 1017-1021.
Mannu, L.G., Riu, R., Comunian, M. C. F. and Ozzi, M. F., 2002. A preliminary
study of lactic acid bacteria in why starter culture and industrial piconiro sardo ewes
milk cheese: PCR- identification and evolution during roping. Int. J. Dairy. 12: 17-26.
Maritirani, L., Vaarcamonti, M., Naclerio, G. and d e Felice, M., 2002. Purification
and partial characterization of bacillocin 490, a novel bacteriocin produced by
thermophilic strain of Bcillus licheniformis. Microbiol. Cell. Factories., 1:1-5.
Martinez, B., Rodriguez, A. and Suarez, A.E., 2000. Lactococcin 972, abacteriocin
that inhibits septum in Lactococci. Microbiology., 146: 949-955.
MarugG, J.D., Gonzalez, C.F., Kunka, B.S., Ledeboer , A.M., Pucci, M.J.,
Toonem, M.Y., Walker, S.A., Zoetmudler, L.C.M. and Vandenbergh, P.A. 1992.
Cloning, expression and nucleotide sequence of genes involved in the production of
pediocin P A-1, abacteriocin from Pediococcus acidilactici PCA 1.O. App. Environ.
Microbiol., 58:2360-2367.
Mc KAY et Baldwin, J., 1975. Stabilisation of lactose metabolism in Streptococcus
lactis C2. Appl. Environ. Microbiol. 36:360-367.
Mc KAY, 1983. Functional properties of plasmids in lactic streptococci. J. Microbiol.
Serol. 49:259-247.
Mc Auliffe, O. and Hill, C., 2001. Lantibiotics: structure, biosynthesis and mode of
action. FEMS. Microbiol. Rev., 25: 285-308.

69
Mehta, A.M., Patel, K.A. and Dave, P.J., 1984 . Purification and some properties of
an inhibitory protein isolated from Lactobacillus acidophilus AR1. Milchwissenschaft
39: 86-89.
Moll, G.N., Konings, W.N. and Driessen, A.J.M., 19 99. Bacteriocins: mechanism
of membrane insertion and pore formation. Antonie Van Leeuwenhoek., 76:185-198.
Montville, T.J. and Bruno. M. E. C., 1994. Evidence that dissipation of proton
motive force is a common mechanism of action for bacteriocins and other
antimicrobial proteins. Int. Food. Microbiol., 24:53-74.
Muriana, P.M. and Kklaenhammer, T.R. 1991. Purification and partial
characterization of lactacin F a bacteriocin produced by Lactobacillus acidophilus
11088. App. Environ. Microbiol., 57 :114-121.
Neetles, C.G. and barefoot, S.F., 1993. Biochemical and genetic characterization of
bacteriocins of associated lactic acid bacteria. J. Food. Prot., 56:338-356.
Nigutova, K. et Paik, D., 2007. Production of enterolysin A by rumen Enterococcus
faecalis strain and occurrence of enlA homologues among ruminal Gram+ cocci. J.
Appl. Microbiol., 102(2) : 563-569.
Nilsen, T., Nes, I.F., and Holo, H., 2003. Enterolysin A, a cell wall-degrading
bacteriocin from Enterococcus faecalis LMG2333. Appl. Environ. Microbiol., 69(5):
2975-2984.
Nilsen, T., Nes I.F. and Holo, H., 2003. Enterolysin A, a cell wall-degrading
bacteriocin from Enterococcus faecalis LMG 2333. App. Environ. Microbiol., 69 :
2975-2987.
Okereck, A. and Montiville, T.J., 1992. Nisin dissipates the proton motive force of
the obligate anaerobic Clostridium sporogenes PA 3679. App. Environ. Microbiol.,
58 :2463-2467.
Oscariz, J.C. and Pisabarro, A.G., 2000. Characterization and mechanism of cerein
7, anew bacteriocin produced by Bacillus cereus with a broad spectrum of activity.
FEMS Microbiol. Lett. 178 :337-341.
O’sullivan, L., Ross, R. P. and Hill, C., 2002. Potential of bacteriocin producing
lactic acid bacteria for improvements in food safety and quality. Biochimie. 84: 593-
604.

70
Paik, H. D., Bae, S.S. and Pan, J. G., 1997. Identification and partial
characterization of thochicin, a new bacteriocin produced by Bacillus thurengiensis
subsp thochigiensis. J. Indust. Microbiol. Biotech. 19: 294-298.
Papagianni, M., 2003. Ribosomally synthesized peptides with antimicrobial
properties: biosynthesis, structure, function and applications. Biotechnol. Adv., 21(6):
465-499.
Pèrez, Guerra, N., 2005 . Fed-batch pediocin production by Pediococcus acidilactici
NRRL B-5627 on whey. Biotechnol. Appl. Biochem., 42:17-23.
Piard, J.C., Kuipers, O.P., Rollema ,H.S., Desmazea ud, M. and de Vos, W.M.
1993. Structure, organisation and expression of lct gene of lactacin 481, a new
lantibiotic produced by Lactococcus lactis. J. Biol. Chem., 268 :16361-16368.
Pritchard, G. C. and Coolbear, T., 1993. The physiology and biochemistry of the
proteolytic system in lactic acid bacteria. FEMS. Microbiol. Rev. 12: 179-206.
Quadri, L.E.N., Saile,r M., Roy, K.L., Vederas, J.C . and Stiles, M.E., 1994.
Chemical and genetic organization of bacteriocins produced by Camobacterium
piscicola LV17B. J. Biol. Chem. 269: 12204-12211.
Ouadri, L.E.N., Sailer, M., Terebiznek, M. Roy, K.L ., Vederas, J.C. and Stiles,
M.E. 1995. Characterization of the protein conferring immunity to the antimicrobial
peptide carnobacteriocin B2 and expression of carnobacteriocin B2 and BM1. J.
Bacteriol., l77: 1144-1151.
Rammelsberg, M., Muller, E. and Radler, F., 1990. Caseicin 80: purification and
characterization of a new bacteriocin from Lactibacillus. Csei. Arch. Microbial.,
154:249-252.
Ray, B., Moltagh, A. and Johnson, M.C. 1993. Processing of prepediocin in
Pediococcus acidilactici . FEMS Microbiol. Rev., 12:119-123.
Richard C., 2006. Evidence on correlation between number of Disulfide Bridge and
toxicity of class IIa bacteriocins. Food. Microbiol., 23(2) : 175-183.
Sahl, H.J., Kordel, M. and Benz, R., 1987. Voltage dependent $depolarization of
bacterial membrane and artificial lipid bilayers by peptid antibiotic nisin.
Arch.Microbiol., 149:120-124.

71
Saidi, N., Guessas, B., Bensalah, F., Badis, A., Ha dadji, M., Henni, D.E., Prevost,
H., and Kihel, M., 2002. Caractérisation des bactéries lactiques isolées du lait cru
de chèvre des régions arides.J. Alg. Reg. Arides. 1 : 1-11.
Samaržija, D., Antumac, N., and Havranek, J.L., 200 1. Taxonomy, physiology and
growth of Lactococcus lactis. a review. Mljekarstvo 51: 1: 35-48.
Samelis, J., Maurogenakis, F. and Metaxopoloulos, J ., 1994. Characterization of
lactic acid bacteria isolated from naturally fermented greek dry salami. Int. J. Food.
Microbiol. 23: 179-196.
Schillinger, U. and Lücke, F.K.,1989. Identification of Lactobacilli from meat and
meeat products. J. Food. Microbiol. 4: 199-208.
Sherwitz , M.E., Feher, A. V. and Cintas, L. M., 19 86. Thuricin: the bacteriocin
produced by Bacillus thurengiensis. J. Invert. Pathol., 53: 206-216.
Siegumfeldt, H., Rechinger, K.B., and Jacobsen, M., 2000. Dynamic changes of
intracellular pH in individual lactic acid bacterium cells in response to a rapid drop in
extracellular pH. Appl. Environ. Microbiol., 66: 2330-2335.
Siergers, K. and Etian, K.D., 1995. Genes involved in immunity to the lantibiotics
nisin produced by Lactococcus lactis 6F3. App. Environ. Microbiol., 61 : 1082-1089.
Stiles, M.E. and Holzapfel, W., 1997. Lactic acid bacteria of foods and their current
taxonomy. Int. J. Food Microbiol., 36(1), 1-29.
Stoddard, W., Petzel, J.P., Van Belkum, M.J., Kok, J. and Mc Kay, L.L., 1992.
Moleculer analyses of the lactococcin A gene cluster from Lactococcus lactis subsp.
lactis biovar diacetylactis WM4. App.Environ. Microbiol., 58:1952-1961.
Tagg, J.R., Dajani, A.S. and Wannmar, K., 1988. Production and isolation of
reuterin, agrowthinhibitor produced by Lactobacillus reuterin, a growth inhibitor
produced by Lactobacillus reterii. Antimicrobiol. Agent. Chemothe., 32 :1954-1958.
Terzaghi, B. E. and S andine, W.E., 1975. Improved medium for lactic streptococci
and their bacteriophages.Appl. Environ. Microbiol. 29: 807-813.
Thomas, D., 1973. Agar medium for differentiation of Streptococcus cremoris from
the other bacteria. N. Z. J. Dairy. Sci. Technol. 8: 70-71.

72
Twomey D., Ryan M., Meaney B. and Hill C., 2002. Lantibiotics produced by lactic
acid bacteria: structure, function and applications. Antonie van Leeuwenhoek., 82:
165-185.
Upreti, G.C. and Hinsdill., R.D., 1973. Production and mode of action of lactocin
27: bacteriocin from a homofermentative Lactobacillus. Antimicrob. Agents
Chemother. 7: 145-149.
Van Laak, R.L.M. J., Schillinger, U. and holzapfel, W., 1992. Characterization and
partial purification of bacteriocin produced Leuconostoc carnosom LA 44A. Int. J.
Food. Microbiol., 57 :114-121.
Venema, K., Venema, G. and Kok, J. 1995 . Lactococcal bacteriocins: mode of
action and immunity. Trends Microbiol., 3: 299-304.
Vennema, K., Chikindas, M.L., Seegers, J.F.M.L. Haa ndrikman, A. J., Leenhouts,
K. J., Venema J. and Kok, J., 1997. Rapid and efficient purification method for
small, hydrophobic, cationnic, bacteriocins: purification of lactococcin B and pediocin
PA-1. App. Environ. Microbiol., 89 : 361-369.
Vermeiren, L., Devlieghere, F. and Debevere, J., 20 04. Evaluation of meat born
lactic acid bacteria as protective cultures for the biopreservation of cooked meat
products. Int. J. Food Microbiol., 96(2): 149-164.
Weil, H.P., Beck-Sickinger, A.G., Metzger, J., Stev anovic, S. and Sahl, H.G.,
1990. Biosynthesis of the lantibiotic Pep5: isolation and characterization of a peptide
containing dehydroamino acids. Eur. J. Biochem. 194: 217-223.
Worobo, R.W., Henkel, T., Sailer, M., Roy, K., Vede ras, J.C., Stiles M.E., 1994.
Characteristics and genetic determinants of a hydrophobic peptide bacteriocin,
carnobacteriocin A, produced by Carnobacterium piscicots LV17A. Microbilgy.,
140:517-526.
Wouters, J.T.M., Ayad, E.H.E. et Smit G., 2002. Microbs from raw milk for
fermented dairy products. Int. J. 12:91-109.

Annexes
Milieux de culture :
Milieux MRS (De man Rogossa et sharp, 1960) : est utilisé pour la culture des
lactobacilles.
ingrédients Piods g/l
Extrait de viande 10g
Extrait de levure 2g
Glucose 20g
Tween 80 1mg
Citrate de sodium 2g
Acétate de sodium 1g
Sulfate de manganèse 0,0g5
Phosphate potassique 2g
Eau distillée 1000ml
Ph 7,2
Autoclavage 120°C pendant 20 min
Milieu MRSBCP (KIHEL, 1996) : utilisé pour l’étude du profil fermentaire.
ingrédients Piods en g/l
Protéose peptone 10g
Extrait de levure 5g
Tween 80 1mg
Citrate de sodium 2g
Acétate de sodium 1g
Sulfate de manganèse 0,0g5
Phosphate potassique 2g
Eau distillée 1000ml
pH 7,2
On ajoute à ce milieu un indicateur de pH qui est le pourpre de bromocrésole a
raison de O,O5 g/l. On ajuste le pH à 6,9 puis on l’autoclave 20min à 120°C.
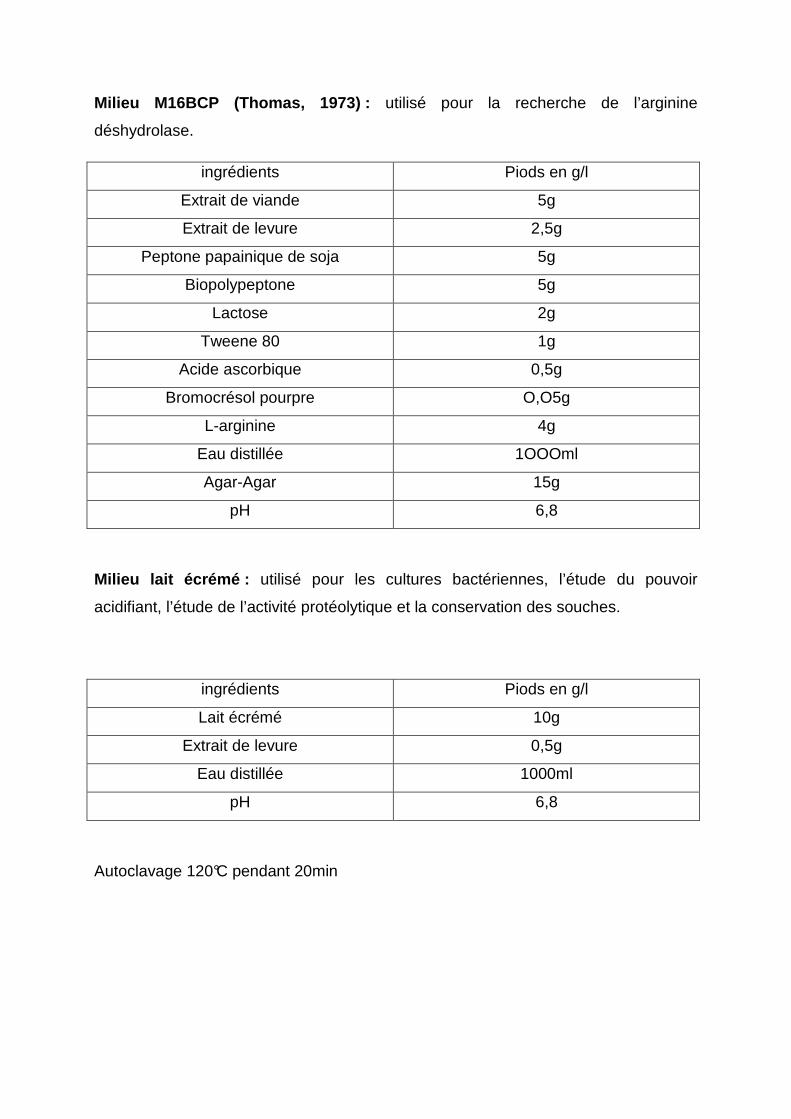
Milieu M16BCP (Thomas, 1973) : utilisé pour la recherche de l’arginine
déshydrolase.
ingrédients Piods en g/l
Extrait de viande 5g
Extrait de levure 2,5g
Peptone papainique de soja 5g
Biopolypeptone 5g
Lactose 2g
Tweene 80 1g
Acide ascorbique 0,5g
Bromocrésol pourpre O,O5g
L-arginine 4g
Eau distillée 1OOOml
Agar-Agar 15g
pH 6,8
Milieu lait écrémé : utilisé pour les cultures bactériennes, l’étude du pouvoir
acidifiant, l’étude de l’activité protéolytique et la conservation des souches.
ingrédients Piods en g/l
Lait écrémé 10g
Extrait de levure 0,5g
Eau distillée 1000ml
pH 6,8
Autoclavage 120°C pendant 20min