Embed Size (px)
Citation preview

ÉCOLE NATIONALE VETERINAIRE D’ALFORT
Année 2005
INFLUENCE DES FACTEURS ENVIRONNEMENTAUX
SUR LA REPRODUCTION DE LA JUMENT
THESE
Pour le
DOCTORAT VETERINAIRE
Présentée et soutenue publiquement devant
LA FACULTE DE MEDECINE DE CRETEIL
Le……………
par
Vincent, Pierre, André DRUET Né le 30 juillet 1980 à Saint Germain-en-Laye (Yvelines)
JURY
Président : M. Professeur à la Faculté de Médecine de CRETEIL
Membres
Directeur : Mme CHASTANT-MAILLARD Sylvie Maître de conférences à l’Ecole Nationale Vétérinaire d’Alfort
Assesseur : Mme BLANCHARD Géraldine Professeur à l’Ecole Nationale Vétérinaire d’Alfort

14 septembre 2005
LISTE DES MEMBRES DU CORPS ENSEIGNANT Directeur : M. le Professeur COTARD Jean-Pierre
Directeurs honoraires : MM. les Professeurs MORAILLON Robert, PARODI André-Laurent, PILET Charles Professeurs honoraires: MM. BORDET Roger, BUSSIERAS Jean, LE BARS Henri, MILHAUD Guy, ROZIER Jacques, THERET Marcel
DEPARTEMENT DES SCIENCES BIOLOGIQUES ET PHARMACEUTIQUES (DSBP)
Chef du département : M. BOULOUIS Henri-Jean, Professeur - Adjoint : M. DEGUEURCE Christophe, Professeur
-UNITE D’ANATOMIE DES ANIMAUX DOMESTIQUES Mme CREVIER-DENOIX Nathalie, Professeur* M. DEGUEURCE Christophe, Professeur Mlle ROBERT Céline, Maître de conférences M. CHATEAU Henri, AERC -UNITE DE PATHOLOGIE GENERALE , MICROBIOLOGIE, IMMUNOLOGIE Mme QUINTIN-COLONNA Françoise, Professeur* M. BOULOUIS Henri-Jean, Professeur -UNITE DE PHYSIOLOGIE ET THERAPEUTIQUE M. BRUGERE Henri, Professeur * Mme COMBRISSON Hélène, Professeur M. TIRET Laurent, Maître de conférences -UNITE DE PHARMACIE ET TOXICOLOGIE Mme ENRIQUEZ Brigitte, Professeur * M. TISSIER Renaud, Maître de conférences M. PERROT Sébastien, Maître de conférences -DISCIPLINE : BIOCHIMIE M. MICHAUX Jean-Michel, Maître de conférences
- UNITE D’HISTOLOGIE , ANATOMIE PATHOLOGIQUE M. CRESPEAU François, Professeur * M. FONTAINE Jean-Jacques, Professeur Mme BERNEX Florence, Maître de conférences Mme CORDONNIER-LEFORT Nathalie, Maître de conférences - UNITE DE VIROLOGIE M. ELOIT Marc, Professeur * Mme LE PODER Sophie, Maître de conférences -DISCIPLINE : PHYSIQUE ET CHIMIE BIOLOGIQUES ET MEDICALES M. MOUTHON Gilbert, Professeur -DISCIPLINE : GENETIQUE MEDICALE ET CLINIQUE Melle ABITBOL Marie, Maître de conférences contractuel -DISCIPLINE : ETHOLOGIE M. DEPUTTE Bertrand, Professeur -DISCIPLINE : ANGLAIS Mme CONAN Muriel, Ingénieur Professeur agrégé certifié
DEPARTEMENT D’ELEVAGE ET DE PATHOLOGIE DES EQUIDES ET DES CARNIVORES (DEPEC) Chef du département : M. FAYOLLE Pascal, Professeur - Adjoint : M. POUCHELON Jean-Louis , Professeur
-UNITE DE MEDECINE M. POUCHELON Jean-Louis, Professeur* Mme CHETBOUL Valérie, Professeur M. BLOT Stéphane, Maître de conférences M. ROSENBERG Charles, Maître de conférences Melle MAUREY Christelle, Maître de conférences contractuel - UNITE DE CLINIQUE EQUINE M. DENOIX Jean-Marie, Professeur * M. AUDIGIE Fabrice, Maître de conférences Mme CARSTANJEN Bianca, Maître de conférences contractuel Mme GIRAUDET Aude, Professeur contractuel Melle VIREVIALLE Hameline, Maître de conférences contractuel
- UNITE DE REPRODUCTION ANIMALE Mme CHASTANT-MAILLARD Sylvie, Maître de conférences* (rattachée au DPASP) M. NUDELMANN Nicolas, Maître de conférences M. FONTBONNE Alain, Maître de conférences M. REMY Dominique, Maître de conférences (rattaché au DPASP) M. DESBOIS Christophe, Maître de conférences Melle CONSTANT Fabienne, AERC (rattachée au DPASP) Melle LEDOUX Dorothée, Maître de conférences Contractuel (rattachée au DPASP)
- UNITE DE PATHOLOGIE CHIRURGICALE M. FAYOLLE Pascal, Professeur * M. MAILHAC Jean-Marie, Maître de conférences M. MOISSONNIER Pierre, Professeur Mme VIATEAU-DUVAL Véronique, Maître de conférences Mlle RAVARY Bérangère, AERC (rattachée au DPASP) M. ZILBERSTEIN Luca, Maître de conférences contractuel M. HIDALGO Antoine, Maître de conférences contractuel - UNITE DE RADIOLOGIE Mme BEGON Dominique, Professeur* Mme STAMBOULI Fouzia, Maître de conférences contractuel -UNITE D’OPHTALMOLOGIE M. CLERC Bernard, Professeur Melle CHAHORY Sabine, Maître de conférences contractuel - UNITE DE PARASITOLOGIE ET MALADIES PARASITAIRES M. CHERMETTE René, Professeur * M. POLACK Bruno, Maître de conférences M. GUILLOT Jacques, Professeur Mme MARIGNAC Geneviève, Maître de conférences contractuel - DISCIPLINE : ALIMENTATION M. PARAGON Bernard, Professeur M. GRANDJEAN Dominique, Professeur Mme BLANCHARD Géraldine, Professeur contractuel
DEPARTEMENT DES PRODUCTIONS ANIMALES ET DE LA SANTE PUBLIQUE (DPASP) Chef du département : M. CERF Olivier, Professeur - Adjoint : M. BOSSE Philippe, Professeur
-UNITE DES MALADIES CONTAGIEUSES M. BENET Jean-Jacques, Professeur* M. TOMA Bernard, Professeur Mme HADDAD/ H0ANG-XUAN Nadia, Maître de conférences Mme DUFOUR Barbara, Maître de conférences -UNITE D’HYGIENE ET INDUSTRIE DES ALIMENTS D’ORIGINE ANIMALE M. BOLNOT François, Maître de conférences * M. CARLIER Vincent, Professeur M. CERF Olivier, Professeur Mme COLMIN Catherine, Maître de conférences M. AUGUSTIN Jean-Christophe, Maître de conférences - DISCIPLINE : BIOSTATISTIQUES M. SANAA Moez, Maître de conférences
- UNITE DE ZOOTECHNIE, ECONOMIE RURALE M. COURREAU Jean-François, Professeur* M. BOSSE Philippe, Professeur Mme GRIMARD-BALLIF Bénédicte, Professeur Mme LEROY Isabelle, Maître de conférences M. ARNE Pascal, Maître de conférences M. PONTER Andrew, Maître de conférences - UNITE DE PATHOLOGIE MEDICALE DU BETAIL ET DES ANIMAUX DE BASSE-COUR M. MILLEMANN Yves, Maître de conférences* Mme BRUGERE-PICOUX Jeanne, Professeur M. MAILLARD Renaud, Maître de conférences M. ADJOU Karim, Maître de conférences
Mme CALAGUE, Professeur d’Education Physique * Responsable de l’Unité AERC : Assistant d’Enseignement et de Recherche Contractuel

A Monsieur le Professeur
Professeur à la faculté de médecine de Créteil,
Qui nous a fait l’honneur d’accepter la présidence de notre jury de thèse.
Hommage respectueux.
A Madame le Docteur CHASTANT-MAILLARD
Maître de conférences à l’Ecole Nationale Vétérinaire d’Alfort,
Qui m’a fait l’honneur de diriger et corriger ce travail.
Qu’elle trouve ici l’expression de ma reconnaissance et de mon plus profond respect.
A Madame le Docteur BLANCHARD
Professeur à l’Ecole Nationale Vétérinaire d’Alfort,
Pour l’attention qu’elle a portée à l’examen de ce travail.
Qu’elle trouve ici l’expression de mes sincères remerciements.

INFLUENCE DES FACTEURS ENVIRONNEMENTAUX
SUR LA REPRODUCTION DE LA JUMENT
NOM : DRUET
Prénom : Vincent
Résumé : Parmi les facteurs pouvant influencer la reproduction de la jument, les facteurs
environnementaux jouent un rôle important. La présence d’un étalon favorise la détection des
chaleurs et améliore la fertilité des juments en stimulant leurs sens. Au sein du troupeau,
l’établissement de la hiérarchie peut diminuer la fertilité des plus dominées. Si l’action de la
température ou de l’hygrométrie reste encore floue, on connaît mieux l’influence de la
photopériode dont les caractéristiques sont transmises au système nerveux central grâce à la
sécrétion épiphysaire de mélatonine. Elle régule le caractère saisonnier de la cyclicité de la
jument, influence le temps de gestation et le moment du poulinage. La lune n’a quant à elle
pas d’influence avérée. Le statut nutritionnel de la jument influe également sur ses capacités
reproductrices. L’état corporel doit être surveillé, l’embonpoint ou la maigreur étant nuisibles
à la fertilité et à la fécondité. Il importe d’alimenter ses juments en quantité suffisante quand
leurs besoins augmentent, pendant la fin de gestation et la lactation, et ce de manière
équilibrée : des protéines de bonnes qualité, des acides gras essentiels, une complémentation
minérale et vitaminée adaptée. Il est essentiel de préserver les juments de toute forme de
stress, celui-ci étant responsable d’une diminution de la production de gonadotrophines et par
conséquent d’une diminution de la fertilité et de la fécondité.
Mots clés : jument, reproduction, environnement, photopériode, mélatonine, alimentation, état
corporel, leptine, stress, équidé, cheval
Jury :
Président : Pr.
Directeur : Dr. CHASTANT-MAILLARD
Assesseur : Pr. BLANCHARD
Adresse de l’auteur :
56, bd de Lozère
91120 PALAISEAU
FRANCE

INFLUENCE OF ENVIRONMENTAL FACTORS
ON THE MARE’S REPRODUCTION
SURNAME : DRUET
Given name : Vincent
Summary : Between all factors that may influence mare’s reproduction, environmental factors
play an important role. The presence of a stallion enhances heat detection and increases
mares’ fertility by stimulating their senses. Inside the herd, establishment of hierarchy may
decrease the fertility of the more dominated. The influence of temperature and bad weather is
unknown but we know how photoperiod acts, giving informations to the central nervous
system via the epiphysis’ secretion of melatonin. It regulates the influence of the season on
the mare’s cycle, gestation length and time of birth. The moon is not reported to have any
influence. The mare’s nutritional status also have an influence on her reproduction capacities.
The body condition have to be monitored, excessive or very poor body condition are both
detrimental to fertility and fecundity. It’s important to provide enough food when the mares’
requirements increase, during late gestation and lactation, and a balanced diet : high quality
proteins, essentials fatty acids, and an adapted mineral and vitamins supplement. Finally, it’s
very important to prevent mares from every kind of stress, because it may lead to an impaired
production of gonadotrophins resulting in a lower fertility.
Keywords : mare, reproduction, environment, photoperiod, melatonin, nutrition, body
condition, leptin, stress, equine, horse Jury :
President : Pr.
Director : Dr. CHASTANT-MAILLARD
Assessor : Pr. BLANCHARD Author’s address :
56, bd de Lozère
91120 Palaiseau
FRANCE

- 1 -
Table des matières Table des matières…………………………………………………………………………… 1 Table des illustrations…………………………………………………………………...……4 Introduction…………………………………………………………………………..……….6
I. Facteurs sociaux……………………………………………………………….……...7 1. Influence de la présence du mâle………………………………………………...7
1.1. Influence de l’ouïe et de l’olfaction…………………….....…………………7
1.2. Influence de la vue et de l’olfaction…………………………………….……8
2. Influence de l’allaitement d’un poulain…………………..……………………11
3. Influence de la présence d’autres chevaux…..………………………………...12
3.1. Conséquences de la structure hiérarchique…………………………………12
3.2. Conséquences inhérentes au regroupement d’individus…………………....13
II. Facteurs climatiques……………………………………………….………………15
1. Influence de la température…………………….………………………………15 2. Influence de l’hygrométrie……...………………………………………………16
3. Influence de la photopériode……………………..………………………..……16
3.1. Mise en évidence du rôle de la mélatonine………...…………………....….16
3.2. Influence de la photopériode sur la saisonnalité de la jument……………...18
3.3. Influence de la photopériode et de la saison sur la gestation et le moment du poulinage………………………….………………………………………..….20
4. Influence de la lune……………………….…………………………………..…22

- 2 -
III. Alimentation……………………………………………………………………..23
1. Relation entre la saison et l’alimentation………………..……..………….23
2. Influence de l’état corporel…………...……………………………………..23
2.1. Estimation de l’état corporel…..………………………………………...23
2.2. Modifications associées aux variations de l’état corporel………...….….24
2.2.1. Influence sur la cyclicité……………………………………..…...24
2.2.2. Influence sur la gestation……………………………………..…..28
2.2.3. Influence sur la lactation……………………………………….....28
2.3. Déterminisme……………..………………………………………….….29
2.3.1. Rôle général de la leptine…………………………………………29 2.3.2. La leptine dans l’espèce équine…...………………………………29
3. Influence du niveau alimentaire, de la qualité des nutriments et de l’équilibre nutritionnel…………………….………………………………..33
3.1. Influence du niveau alimentaire.……..……………………….......……..33
3.2. Influence des apports protéiques...………..……………………………..35
3.2.1. Apport quantitatif……………….……………………………...….35
3.2.2. Apport qualitatif…………….………………………………...…...36
3.3. Influence de l’apport lipidique………..……………………………...….37
3.4. Influence de la composition minérale de la ration...…………………….37
3.5. Influence de l’apport vitaminique………..…………………...…………38
3.6. Reproduction et substances indésirables présentes dans les aliments.......38
3.6.1. Plantes susceptibles d’interférer avec la reproduction...………..…38
3.6.2. Les phytœstrogènes…………………………..………………........40
3.6.3. Les mycotoxines…………………………...…………………...…40
3.6.4. Un cas particulier d’influence de l’alimentation sur la reproduction de la jument : les avortements du Kentucky……………..…41

- 3 -
IV. Influence de la conduite d’élevage……………………………………………...42
1. Détection et gestion des chaleurs……...…………………………………….42
2. Le logement………………..…………………………………………………43
3. Le stress……………..………………………………………………………..44
3.1. Physiologie du stress – Interaction avec l’axe reproducteur...……..…44
3.2. Influence du stress chez la jument………..….………………………..47
3.2.1 Influence du stress sur la cyclicité de la jument………….…….47
3.2.2. Influence du stress sur la gestation chez la jument……..…..…48
3.2.3. Influence du stress sur la mise bas chez la jument..…….….…48
Conclusion……………………………………………………………………………………49 Annexe…………..……………………………………………………………………………50
Bibliographie…………………………………………………………………………...…....52

- 4 -
Table des illustrations Figures Figure 1 : Concentrations veineuses en ocytocine hypophysaire chez une jument en œstrus en contact uniquement auditif avec un étalon (d’après Irvine et Alexander [64])……….…...9 Figure 2 : Concentrations veineuses en ocytocine (OT) hypophysaire chez une jument en œstrus en contact tactile et auditif avec un étalon (d’après Irvine et Alexander [64])……..…………………………………………………………………….......10 Figure 3 : Evolution du taux de progestérone de dix juments comparée aux variations de la durée d’éclairement (d’après Guillaume et al. [48]) …………………………………...17 Figure 4 : Evolution de la sécrétion de mélatonine par une ponette sur 24 heures lors d’une nuit longue ou d’une nuit courte (d’après Guillaume et al. [48]) …………………..…17 Figure 5 : Schéma général de la transmission de l’information lumineuse de la rétine aux gonadotrophines (d’après Guillaume et al. [48]) ………………..……………..……..…19 Figure 6 : Effet de différentes photopériodes expérimentales sur la stimulation de l’activité ovarienne chez la jument (d’après Guillaume et Palmer [64]) ………………….…21 Figure 7 : Représentation des aires à palper pour évaluer l’état corporel de son cheval ([72] d’après Henneke et al. [57]).……………………………………………………………25 Figure 8 : Schéma général des rôles de la leptine dans la régulation du métabolisme (d’après Hossner [61])………………………………………………………….…………….31 Figure 9 : Schéma général des effets de la leptine sur la fonction de reproduction (d’après Hossner [61])……………………………………………………………………......31 Figure 10 : Hypothèses sur des boucles de régulation par lesquelles la leptine pourrait moduler la fonction de reproduction en général (d’après Bruneau et al. [18])….......32 Figure 11 : Schéma général des possibles interactions de la leptine avec les cellules de l’épithélium mammaire (d’après Bonnet et al. [13])…………………….……..………….32 Figure 12 : Les différents sites d’action du stress et leur relation avec la fonction de reproduction (d’après Armstrong [3])……………………………………………….…….46

- 5 -
Tableaux Tableau 1 : Maladies et germes responsables de troubles de la reproduction chez la jument (d'après [53])…..……………………..……………………………………………….14 Tableau 2 : Notation de l’état corporel selon l’INRA. (d’après Martin-Rosset [76]).....…....26 Tableau 3 : Notation de l’état corporel (d’après [57, 72])…………..……………………….27 Tableau 4 : Plantes abortives et/ou tératogènes chez la jument (d’après [73])…..………..…39

- 6 -
Introduction Le cheval est depuis peu passé du statut d’animal de rente à celui d’animal de loisir, mais l’objectif de l’éleveur reste inchangé : obtenir un poulain par jument et par an, aidé en cela par le fait que la jument est une des rares femelles à ne pas subir d’anœstrus de lactation. Dans le cas des chevaux de rente, la contrainte est supérieure ; en effet, plus le produit naît précocement dans l’année, plus il sera apte à satisfaire les futures exigences de ses éleveurs : le cheval destiné à la boucherie sera plus lourd que ses semblables nés plus tard dans la saison. De même les poulains destinés aux courses sont d’autant plus mâtures qu’ils sont nés tôt dans l’année, ce qui leur donne une avance physique et une période d’entraînement plus longue, les courses étant basées sur l’âge en années pleines. Il est important de savoir en effet qu’officiellement l’âge d’un cheval est le même qu’il soit né le 01 janvier ou le 31 décembre. Si l’on tient compte de ces enjeux économiques, on comprend mieux l’intérêt d’avancer la saison de reproduction de la jument le plus tôt possible dans l’année tout en tenant compte des contraintes administratives (saison de reproduction démarrant au 15 février) et de la physiologie de la jument. En effet, cette dernière entre en inactivité ovarienne (« anœstrus ») vers le mois d’octobre (elle présente parfois tout de même des chaleurs mais elles sont anovulatoires), et elle en sort après une période de transition (fin d’hiver - début de printemps) vers fin avril, ce qui n’arrange pas les éleveurs qui recherchent des produits les plus précoces possibles. L’éleveur cherche de plus à améliorer la fertilité et la fécondité de ses juments afin de remplir l’objectif d’un poulain par jument et par an. Classiquement on définit deux grandes catégories de facteurs pouvant influencer la fonction de reproduction de la jument [39] : les facteurs extrinsèques (environnementaux) et les facteurs intrinsèques (propres à l’animal). Ces derniers regroupent l’âge, la race de la jument, la génétique et les affections congénitales ou acquises dont elle peut souffrir. Les premiers sont, pour les plus couramment incriminés, la photopériode, l’alimentation et la température. S’il paraît évident qu’une jument âgée ou atteinte d’une affection de l’appareil reproducteur sera moins féconde, on pense beaucoup moins au rôle de l’environnement qui, par le biais de divers récepteurs, peut modifier la fonction ovarienne de la jument, sa capacité à concevoir un embryon, à mener une gestation à terme, à mettre bas et à allaiter. L’objet de ce travail est donc de montrer quels sont les facteurs environnementaux ayant une influence positive ou négative sur la reproduction de la jument, comment ils agissent et comment les gérer au mieux afin de remplir les objectifs des éleveurs. Nous envisagerons dans ce but successivement l’influence des facteurs sociaux au travers des relations que peuvent avoir les juments avec leurs congénères, des conditions climatiques et lumineuses, de l’alimentation et de la conduite d’élevage.

- 7 -
I. Facteurs sociaux
A l’état naturel, le cheval est un animal grégaire. La structure sociale est de type « harem » permanent : à un groupe de juments est associé un mâle adulte et leur progéniture (foals et yearlings). D’autre part il existe des bandes de mâles célibataires qui n’ont pas accès à la reproduction. Certains individus restent quant à eux solitaires, ce sont souvent les mâles âgés ou malades [25].
On peut donc légitimement se demander si les conditions de vie imposées par les techniques d’élevage traditionnel, se caractérisant par une relative pauvreté de relations sociales, et ce pour des raisons tout à fait compréhensibles (prévention de traumatismes sur des chevaux de prix et/ou dont l’utilisation est source de revenus, prévention de la dissémination de germes…), ne perturbent pas en plus des comportements sociaux normaux la physiologie des individus, et plus particulièrement la reproduction de la jument.
1. Influence de la présence du mâle
Alors que le mâle est omniprésent dans la structure hiérarchique normale du harem chez les chevaux sauvages, il est en général absent des élevages français qui possèdent en moyenne deux à trois poulinières (en moyenne 2,4 juments saillies par éleveur en 2000, toutes races confondues [71]) mais pas d’entier.
Il convient donc d’étudier le rôle précis du mâle lorsqu’il est présent et de déterminer si son absence peut influencer le potentiel reproducteur des juments. Son implication quand à la propagation de maladies sexuellement transmissibles sera envisagé dans le paragraphe traitant de l’influence du regroupement d’individus.
L’étalon stimule les différents sens de la jument de façon concomitante, que ce soit
l’ouïe, l’odorat, le toucher ou bien la vue [64].
1.1. Influence de l’ouïe et du toucher Comme l’ont montré les travaux de Irvine et Alexander [64], le hennissement du mâle
semble être pour la femelle un critère de choix de son partenaire sexuel lorsque celle-ci en a plusieurs à sa disposition. Les auteurs pensent que l’intensité du son émis est un facteur déterminant dans le choix de la femelle.
Mais le rôle de l’audition ne s’arrête pas là, la perception auditive de l’étalon s’accompagnant chez la jument d’une modification des profils hormonaux. Selon Irvine et Alexander [64] chaque hennissement d’un mâle situé à 50 m d’une jument ne pouvant ni le voir ni le sentir s’ensuit d’une décharge d’ocytocine (fig. 1) qui stimule la production de gonadotrophines et les contractions utérines.
La sensibilité tactile de la jument semble encore plus efficace. Les décharges d’ocytocine sont encore plus importantes (fig.2) lorsque le mâle est autorisé à courtiser la jument en lui touchant les flancs et la région périnéale du bout du nez vérifiant par là même qu’elle est prête à l’accepter.
Lors du coït, la stimulation tactile de la vulve, du clitoris, du vagin et du col de l’utérus est également à l’origine d’une décharge d’ocytocine. L’insémination artificielle reproduit ces mêmes stimuli à ceci près qu’elle ne provoque pas de distension du vagin.

- 8 -
Le premier rôle attribué à ces décharges d’ocytocine est l’augmentation de la pulsatilité de LH permettant ainsi d’avancer la première ovulation de la saison de reproduction. En effet, il a été montré chez le rat [88] et chez l’homme [62] que l’administration d’ocytocine favorise la libération d’hormone lutéinisante par l’hypophyse. On peut alors imaginer que la stimulation répétée par un étalon d’une jument en fin de période de transition pourrait permettre de gagner ainsi un cycle ou deux supplémentaires. L’effet de l’ocytocine sur la sécrétion de FSH n’a pas fait l’objet d’aussi nombreuses études que pour la LH, mais il semblerait qu’elle favorise également sa libération [36].
Le deuxième rôle attribué à la décharge ocytocique, et celui-ci est bien avéré chez la jument, est la stimulation des contractions de l’utérus et des oviductes, augmentant ainsi la probabilité de rencontre des spermatozoïdes et de l’ovocyte. Elle favorise également le nettoyage de l’utérus que ce soit au sortir de l’hiver après la phase d’inactivité ovarienne ou bien après l’involution utérine ayant suivi la gestation précédente, participant ainsi à la résistance de l’utérus aux infections éventuelles.
Enfin, l’ocytocine est supposée participer au renforcement des liens sociaux à l’intérieur du troupeau [64].
Bien que les juments ovulent spontanément et que la majorité soit capable d’avoir une
cyclicité normale sans stimulation par un mâle, certaines ont néanmoins des problèmes pour ovuler ou pour maintenir leur corps jaune. Ces difficultés sont souvent associées à une stimulation insuffisante des ovaires par les gonadotrophines dont la sécrétion est favorisée par la décharge d’ocytocine. Les pics sanguins provoqués par les stimulations de l’étalon peuvent être reproduits [64] par l’administration d’ocytocine exogène ce qui entraîne aussi la stimulation de l’utérus et des oviductes. Mais pour obtenir les très hautes doses retrouvées au niveau du cerveau ou de l’hypophyse, il faudrait administrer des quantités d’ocytocine (figure 2) incompatibles avec un fonctionnement correct du tractus génital [64]. On ne peut donc pas reproduire pharmacologiquement l’influence de l’étalon.
1.2. Influence de la vue et de l’olfaction
La vue est le dernier sens mis à contribution lors de la reproduction mais aussi le plus
difficile à étudier. En effet, il n’est pas aisé d’isoler son rôle propre de celui des autres sens. Bien souvent, la jument sent ou même entend l’étalon avant même de le voir.
Ainsi pour des raisons aussi bien pratiques qu’éthiques (son étude spécifique nécessiterait la destruction des aires corticales responsables de l’audition et de l’olfaction), aucune étude n’a pu être envisagée chez la jument.
Bien qu’aucune étude concernant spécifiquement le cheval n’ait été menée, l’influence des stimuli olfactifs et en particulier des phéromones notamment sur la fertilité est très fortement suspectée. Elles seraient de plus responsables de la synchronisation des chaleurs chez des juments vivant à proximité l’une de l’autre.
L‘effet de ces mêmes phéromones a été démontré chez la truie d’où la commercialisation de sprays réputés améliorer la fertilité des femelles. Un effet similaire a été rapporté chez certains singes [64].
Chez l’étalon, le signe du flehmen nous suggère qu’il est sensible à son environnement olfactif. Les travaux de Ginther (cité par [64]) font suspecter un rôle de l’olfaction mais sans en expliquer la teneur exacte.
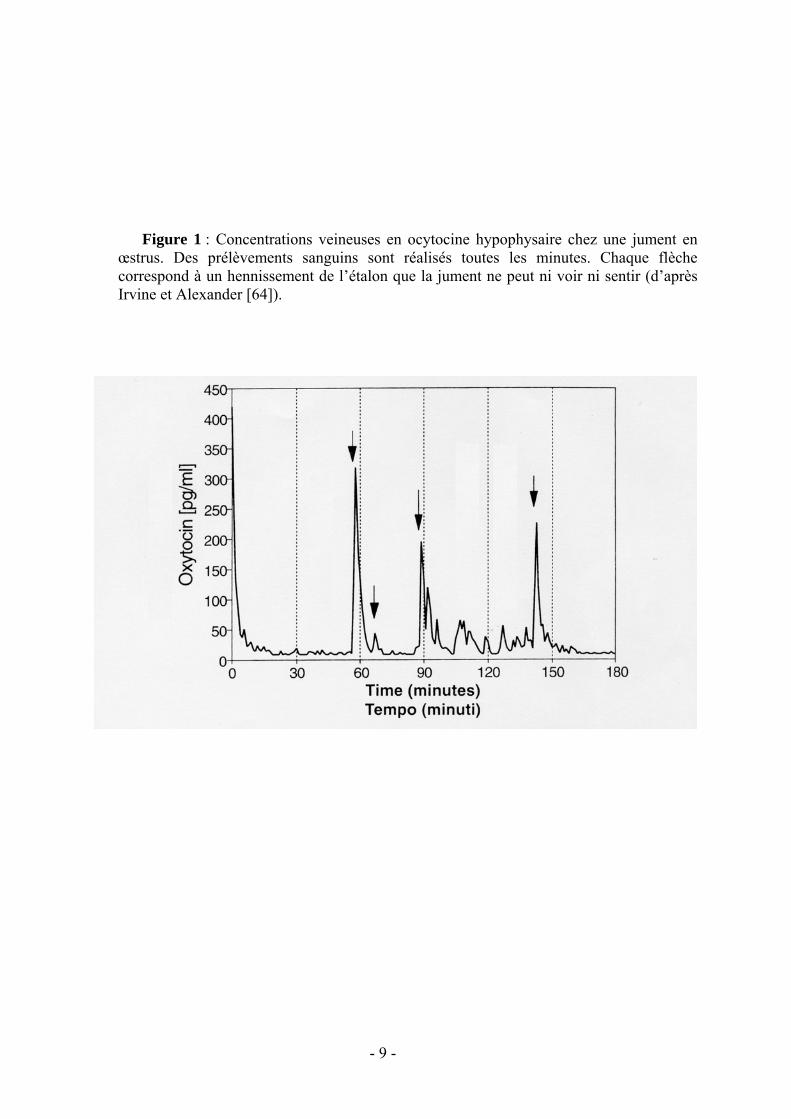
- 9 -
Figure 1 : Concentrations veineuses en ocytocine hypophysaire chez une jument en
œstrus. Des prélèvements sanguins sont réalisés toutes les minutes. Chaque flèche correspond à un hennissement de l’étalon que la jument ne peut ni voir ni sentir (d’après Irvine et Alexander [64]).
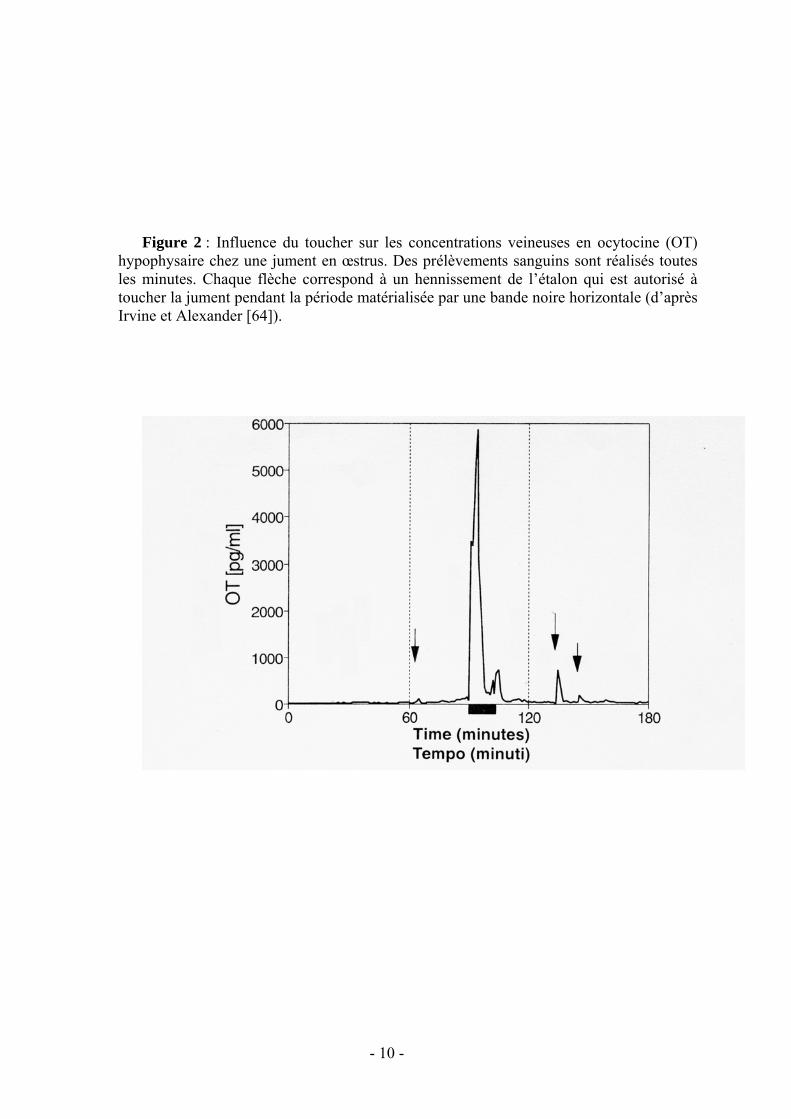
- 10 -
Figure 2 : Influence du toucher sur les concentrations veineuses en ocytocine (OT)
hypophysaire chez une jument en œstrus. Des prélèvements sanguins sont réalisés toutes les minutes. Chaque flèche correspond à un hennissement de l’étalon qui est autorisé à toucher la jument pendant la période matérialisée par une bande noire horizontale (d’après Irvine et Alexander [64]).

- 11 -
L’absence totale de mâle comme c’est souvent le cas en élevage traditionnel n’entrave
donc pas la reproduction normale de la jument mais sa présence semble favoriser la fertilité et la fécondité de la jument. C’est pourquoi la présence d’un entier jouant le rôle de souffleur pour la détection des chaleurs ou sur le lieu de l’insémination artificielle peut se révéler bénéfique.
2. Influence de l’allaitement d’un poulain Parmi les individus constitutifs du troupeau on trouve bien entendu les poulains
nouveau-nés. Et même si, comme évoqué plus haut, la jument ne souffre pas d’anoestrus post-partum comme la vache ou d’anoestrus de lactation comme la truie, il convient de s’interroger sur l’influence du nouveau produit sur la gestation à venir.
L’objectif de l’éleveur étant d’obtenir de sa poulinière un produit par an, la jument
commence une nouvelle gestation alors qu’elle allaite depuis environ un mois, et ce pour encore 5 mois. Cette lactation auto-entretenue par la succion du poulain a plutôt des effets négatifs, aussi bien sur la capacité à concevoir qu’à mener à terme cette nouvelle gestation.
La lactation est extrêmement exigeante pour la jument et demande un suivi
nutritionnel adapté. La toute première fonction à pâtir d’un déficit alimentaire est la reproduction. Une jument sous-alimentée donnera donc la priorité à l’alimentation de sa progéniture au détriment de la reprise de sa cyclicité, ce qui peut expliquer que certaines juments ne poursuivent une gestation qu’une année sur deux.
De la même façon il faut tenir compte de la production laitière de la jument. D’après Van Niekerk F.E. et Van Niekerk C.H. [107], les chaleurs de lait sont plus discrètes voire absentes chez les fortes productrices (type juments de trait) par rapport aux juments de format plus réduit (trotteuses notamment). Il faut donc penser à adapter l’alimentation des fortes laitières en conséquence.
Ginther et al. [45] ont fait de leur côté une observation du même ordre : les juments dont la tétée est limitée (par séparation d’avec leur foal entre 8h et 17h le jour suivant la mise bas) voient leur chaleurs de lait survenir significativement plus tôt.
Henneke et Kreider [55] confirment ce résultat : les juments dont le contact avec leur poulain est limité à une heure toutes les six heures retombent en chaleur en moyenne quinze jours plus tôt que les juments témoins qui pâturent librement avec leur progéniture. De plus, ce protocole a permis à des juments particulièrement sensibles au stress de la lactation de remplir deux ans de suite contrairement à leur rythme habituel d’un an sur deux. Cette étude a de plus cherché à vérifier si un tel protocole n’avait pas de répercussions sur la santé du poulain : les paramètres relevés (GMQ, poids à 180 jours, état corporel) n’ont pas montré d’influence néfaste d’un tel régime.
Cette relation lactation-alimentation-reproduction est également soulignée par l’observation récurrente d’une inactivité ovarienne hivernale plus longue chez les juments ayant allaité l’été précédent [49].
Enfin, la lactation déprime la sécrétion de progestérone par le corps jaune. Et cette diminution du taux de progestérone peut compromettre la survie du nouveau conceptus une fois la nouvelle gestation entamée [108].

- 12 -
3. Influence de la présence d’autres chevaux
3.1. Conséquences de l’établissement d’une hiérarchie Les poulinières sont le plus souvent regroupées entre elles. Il faut donc considérer la
hiérarchie qui s’établit au sein du troupeau de juments. La structure fondamentale de cette hiérarchie est linéaire : si A domine B, et si B domine C, alors A domine C. Mais elle peut présenter à différents niveaux des relations triangulaires, c’est-à-dire que A domine B, B domine C, mais C domine A.
La prise en compte de cette hiérarchie prend toute son importance dans la gestion de
l’alimentation. En effet, dans le cadre par exemple d’un régime alimentaire volontairement restrictif, les juments dominantes n’en seront que modérément affectées et les dominées plus sévèrement que prévu. Dans le cadre d’un régime type flushing, steaming ou bien alimentation d’une jument en lactation, les juments dominées ne bénéficieront pas de l’intégralité de leur ration et ne pourront couvrir leurs besoins. Cette dualité dominante/dominée n’est donc pas à négliger et peut avoir de réelles conséquences. Il est toujours préférable de séparer si possible les juments au moins le temps du repas. Les affinités interindividuelles peuvent atténuer certains effets de la hiérarchie puisque ces affinités sont indépendantes de la position relative dans le sociogramme : une jument de rang élevé peut être la compagne d’une jument très dominée, cette dernière ayant accès plus facilement à l’aliment si elle se place à côté de sa compagne [25].
Le regroupement des juments en période de transition permet une certaine
synchronisation du début de la saison ovulatoire, de même que des cycles ultérieurs si les juments restent confinées dans un espace restreint. Ce phénomène de mimétisme montre à nouveau l’influence des stimulations sensorielles et en particulier ici des phéromones. Mais une fois de plus les relations hiérarchiques peuvent influer : dans le cas d’une détection des chaleurs effectuée en liberté par un étalon vasectomisé, la jument dominante aura tendance à évincer la dominée de l’entourage du mâle d’où une mauvaise détection des chaleurs [25].
Chez la vache, des changements de rang social sont responsables selon Dobson et Smith [31] d’une amélioration (ou d’une détérioration si la vache descend dans la hiérarchie) de nombreux paramètres : intervalle part-nouvelle fécondation écourté (de 143 à 97 jours), nombre moyen d’insémination nécessaire à l’obtention d’une gestation diminué (de 2,2 à 1,6), production laitière augmentée, moindre nombre de cellules présentes dans le lait.
Enfin, au moment du poulinage, une dominante peut s’approprier le poulain d’une
jument de rang inférieur, surtout si elle est primipare. Il devient ensuite difficile de faire accepter le poulain par sa vraie mère et on peut perdre le poulain. Ce « vol » de progéniture est également décrit chez les ovins surtout lors de naissances synchrones [25].

- 13 -
3.2. Conséquences inhérentes au regroupement d’individus
Un autre problème inhérent à la proximité de congénères est la transmission de maladies contagieuses. L’exemple le plus flagrant et le plus répandu est celui de la rhinopneumonie dans sa forme abortive. Sa forte contagiosité peut être responsable de véritables épidémies avec pour conséquence une perte économique substantielle causée par l’absence quasi-totale de produits l’année en question. D’où l’importance de la vaccination systématique de l’intégralité des troupeaux de poulinières en respectant le protocole « poulinière » qui prône trois injections à 5, 7 et 9 mois de gestation, et ce même si la protection conférée par le vaccin n’est pas de 100 %. Une unique injection annuelle est largement insuffisante. La rhinopneumonie n’est qu’un exemple parmi tant d’autres, on peut penser aussi à la salmonellose, la leptospirose, l’artérite virale…
Le regroupement d’individus augmente également le risque de traumatismes dus à une mésentente.
Enfin, le contact occasionné par la saillie de l’étalon favorise la dissémination de maladies sexuellement transmissibles (métrite contagieuse, streptococcose…).
Les maladies potentiellement responsables de troubles reproducteurs chez la jument sont répertoriées dans le tableau 1.
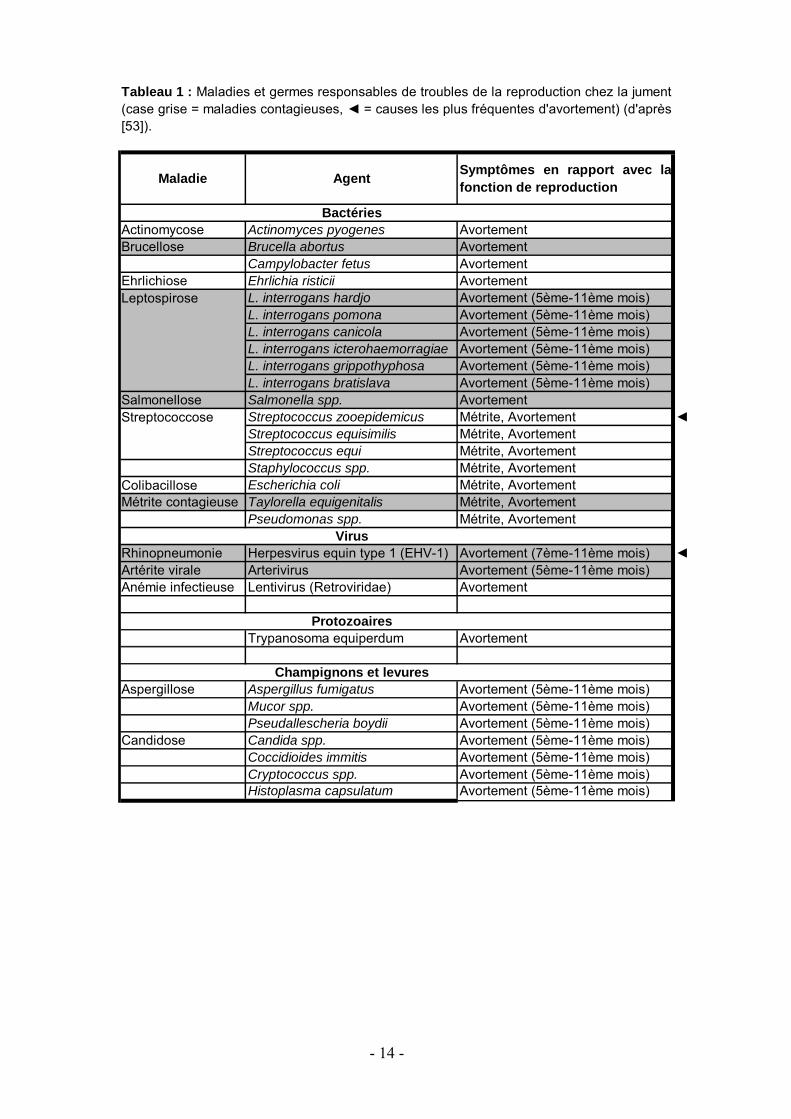
- 14 -
BactériesActinomycose Actinomyces pyogenes AvortementBrucellose Brucella abortus Avortement
Campylobacter fetus AvortementEhrlichiose Ehrlichia risticii Avortement
L. interrogans hardjo Avortement (5ème-11ème mois)L. interrogans pomona Avortement (5ème-11ème mois)L. interrogans canicola Avortement (5ème-11ème mois)L. interrogans icterohaemorragiae Avortement (5ème-11ème mois)L. interrogans grippothyphosa Avortement (5ème-11ème mois)L. interrogans bratislava Avortement (5ème-11ème mois)
Salmonellose Salmonella spp. AvortementStreptococcus zooepidemicus Métrite, Avortement ◄Streptococcus equisimilis Métrite, AvortementStreptococcus equi Métrite, AvortementStaphylococcus spp. Métrite, Avortement
Colibacillose Escherichia coli Métrite, AvortementMétrite contagieuse Taylorella equigenitalis Métrite, Avortement
Pseudomonas spp. Métrite, AvortementVirus
Rhinopneumonie Herpesvirus equin type 1 (EHV-1) Avortement (7ème-11ème mois) ◄Artérite virale Arterivirus Avortement (5ème-11ème mois)Anémie infectieuse Lentivirus (Retroviridae) Avortement
ProtozoairesTrypanosoma equiperdum Avortement
Champignons et levuresAspergillose Aspergillus fumigatus Avortement (5ème-11ème mois)
Mucor spp. Avortement (5ème-11ème mois)Pseudallescheria boydii Avortement (5ème-11ème mois)
Candidose Candida spp. Avortement (5ème-11ème mois)Coccidioides immitis Avortement (5ème-11ème mois)Cryptococcus spp. Avortement (5ème-11ème mois)Histoplasma capsulatum Avortement (5ème-11ème mois)
Leptospirose
Streptococcose
Tableau 1 : Maladies et germes responsables de troubles de la reproduction chez la jument(case grise = maladies contagieuses, ◄ = causes les plus fréquentes d'avortement) (d'après[53]).
Maladie Agent Symptômes en rapport avec lafonction de reproduction

- 15 -
II. Facteurs climatiques
Dans des conditions naturelles, chez les chevaux comme chez la plupart des mammifères des régions tempérées ou froides, une synchronisation des naissances est observée. Les poulinages ont lieu pendant la saison la plus favorable pour la survie du jeune, généralement à la fin du printemps. En outre, pour la plupart des animaux vivant dans ces zones, à l’approche du froid hivernal, l’organisme privilégie l’énergie alimentaire disponible pour les fonctions de survie (thermorégulation, résistance aux maladies) au détriment de la fonction de reproduction. Les équidés n’échappent pas à cette règle : en automne, la jument entre en inactivité et le comportement sexuel de l’étalon est diminué. La majorité des juments ont alors une période sans ovulation qui débute dans l’hémisphère Nord vers le mois d’octobre et s’achève à la fin du mois d’avril. Cette phase d’inactivité, dont la durée dépend de l’âge de la jument et de son état physiologique, est appelée inactivité ovarienne saisonnière. Ce terme est plus exact que celui d’anœstrus car la jument peut manifester des comportements d’œstrus toute l’année mais à cette saison ils ne seront pas suivis d’ovulation.
Outre le déterminisme du rythme annuel de reproduction de la jument, la saison influe également sur la longueur relative de l’œstrus et du diœstrus au cours de l’avancement de la saison de reproduction (réduction du temps d’œstrus de 10 à 5 jours et augmentation du diœstrus de 14 à 20 jours) [45] ; elle joue également sur les taux hormonaux et sur le retour en chaleurs des juments après la mise bas [107] : les juments qui mettent bas très tôt dans l’année peuvent ne pas présenter de chaleurs de lait et ne reprendre leur cyclicité que quelques semaines à quelques mois plus tard.
Pour comprendre le rôle de la saison, il faut se rendre compte qu’il s’agit en réalité de plusieurs facteurs interdépendants les uns des autres mais ayant chacun leur rôle propre. Le changement de saison s’accompagne de modifications de température, d’hygrométrie, de photopériode (longueur relative du jour par rapport à la nuit) et d’alimentation.
1. Influence de la température
Plusieurs études (citées par [94]) ont suggéré l’influence de la température ambiante
notamment sur l’apparition des premières chaleurs suivies d’une ovulation, mais aucune n’a pu distinguer son rôle de celui d’autres facteurs tels que l’alimentation ou la photopériode avec lesquels elle est étroitement liée.
L’expérience de Guerin et Wang [47] est à ce jour la seule étude à tenter d’isoler le facteur température (en relevant les données météorologiques pendant 10 ans sur un même élevage ne pratiquant pas de photostimulation). Ils suggèrent l’influence d’une température minimum (environ 9°C) au cours de la journée qui, une fois dépassée, favoriserait l’entrée en période ovulatoire. Mais cette hypothèse d’une température minimum à une phase donnée de la journée (éventuellement au moment où la jument est la plus sensible à la lumière c’est-à-dire 9,5 à 10 heures après la tombée de l’obscurité) reste à explorer plus en détail.
D’autres observations suggèrent également que les températures froides allongent la phase de transition et retardent la date de la première ovulation [48].
La température agirait principalement au printemps [48], pendant la phase de transition, sur l’axe hypothalamo-hypophysaire en faisant vraisemblablement intervenir très partiellement la prolactine selon un mécanisme encore non élucidé.

- 16 -
Bien que le bon sens et les premières études suggèrent que l’arrivée des beaux jours et le réchauffement qui les accompagnent favorisent la sortie de la phase de transition, le rôle exact et surtout le mode d’action précis de la température ambiante fait toujours l’objet de suppositions et reste à étudier.
2. Influence de l’hygrométrie
Chez la brebis, il a été démontré [64] expérimentalement que la pluie pendant 6 heures
durant les jours 14 à 17 du cycle pouvait diminuer significativement le taux d’ovulation (de 45 %).
Chez le cheval, bien que suspectée, l’influence de la pluie n’a jamais été démontrée. 3. Influence de la photopériode Il s’agit de la longueur relative du jour par rapport à la nuit, quantifiée par la durée de
l’éclairement diurne. L’influence de la photopériode est un facteur relativement facile à étudier car la photopériode est le facteur le plus constant d’une année sur l’autre contrairement à la température ou encore l’alimentation qui, elles, fluctuent significativement d’une année sur l’autre [64].
Son influence sur la reproduction animale est connue depuis longtemps dans plusieurs espèces animales. Chez la jument, son influence a été bien étudiée par l’équipe de Ginther [38, 40, 45]. On définit ainsi des espèces à jours longs, c’est-à-dire dont la période d’activité sexuelle s’éveille avec l’allongement des jours et le raccourcissement des nuits comme le cheval (fig. 3), et des espèces dites à jours courts dont l’éveil sexuel se fait à la période inverse, comme les ovins.
3.1. Mise en évidence du rôle de la mélatonine
C’est au début des années 80, avec les premières expériences de pinéalectomie chez la
jument qu’un pas supplémentaire a pu être franchi dans la compréhension de l’influence de la photopériode sur la reproduction de la jument.
Les signaux photopériodiques sont perçus par des cellules spécialisées de la rétine [48] et traduits en signal endocrinien par la glande pinéale qui se caractérise par une sécrétion presque exclusivement nocturne [64] (fig.4) de mélatonine dans le sang et le liquide céphalorachidien.
La mélatonine constitue une des clefs du contrôle neuro-endocrinien de la reproduction chez les mammifères. Elle a fait l’objet de nombreuses études chez la jument pendant ces vingt dernières années où son rôle d’intermédiaire entre la perception de la photopériode et l’activité ovarienne a été démontrée.
La première mise en évidence du rôle fonctionnel de la mélatonine a été réalisée chez des juments pinéalectomisées [48]. Chez ces juments, le début de l’activité de reproduction n’est pas avancé par des photopériodes artificielles et les juments pinéalectomisées pendant l’hiver ont une acticité ovarienne cyclique significativement plus tardive que les juments témoins pendant la deuxième saison après la chirurgie.
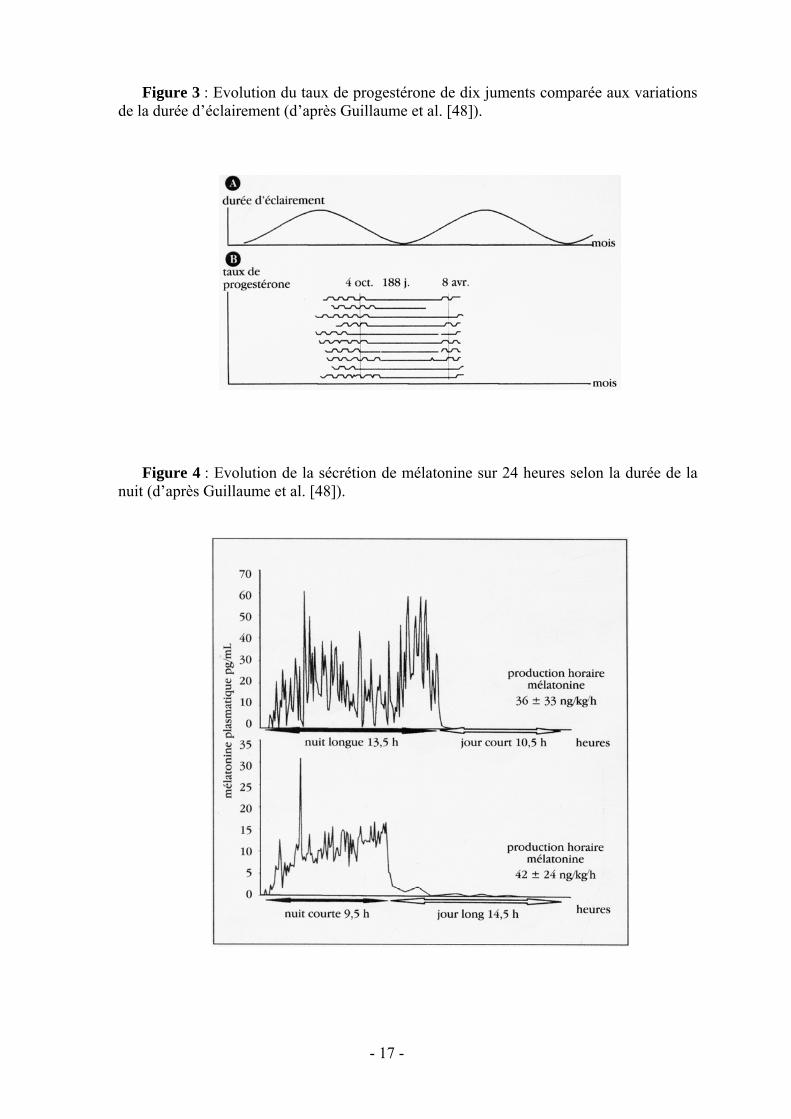
- 17 -
Figure 3 : Evolution du taux de progestérone de dix juments comparée aux variations de la durée d’éclairement (d’après Guillaume et al. [48]).
Figure 4 : Evolution de la sécrétion de mélatonine sur 24 heures selon la durée de la
nuit (d’après Guillaume et al. [48]).

- 18 -
Le rôle de la mélatonine dans le transfert du signal photopériodique a été démontré par l’administration de mélatonine exogène [50, 51], chez des juments photostimulées, 4 heures avant le début des nuits courtes. Cette administration supprime l’effet stimulant des nuits courtes. De façon similaire, des juments en inactivité ovarienne saisonnière ne répondent pas à la photopériode stimulante si la mélatonine est administrée toutes les deux heures pendant une période de douze heures qui inclut les neuf heures d’obscurité.
La photopériode et le rythme de sécrétion de la mélatonine entraînent le rythme annuel de reproduction sans avoir d’action directe sur les gonades. Malgré l’abondante documentation sur les effets directs de la photopériode, le site d’action sur l’axe hypothalamo-hypophysaire de la mélatonine n’a pas été étudié chez le cheval. Dans d’autres espèces, il a été démontré in vivo que la mélatonine n’influe pas directement sur la sécrétion de gonadolibérine (GnRH) mais qu’elle agirait par l’intermédiaire d’interneurones. Chez le cheval, des récepteurs spécifiques de la mélatonine ont été découverts dans la pars tuberalis, dans l’éminence médiane et dans le noyau suprachiasmatique (fig. 5) [48].
On ne sait donc pas de quelle manière la longueur du jour et le taux de mélatonine induisent une modification de la fréquence et de l’amplitude des pulses de GnRH. Est-ce le jour qui s’allonge et le taux de mélatonine qui décroît qui stimulent un taux de GnRH intrinsèquement faible ou bien est-ce un jour qui raccourcit et une augmentation du taux de mélatonine qui inhibent une pulsatilité intrinsèquement élevée ?
Cette question reste encore sans réponse mais de nombreux auteurs à la recherche d’intermédiaire entre la mélatonine et l’axe hypothalamo-hypophysaire pencheraient plutôt pour la deuxième hypothèse. Les opioïdes notamment seraient responsables (via les β-endorphines endogènes par exemple) d’une inhibition de cette même pulsatilité comme le montre sa levée par l’injection de naloxone. Un rôle inhibiteur semblable est recherché dans la famille des catécholamines avec des molécules comme la dopamine [48].
Bien que ne comprenant pas le mécanisme intime de son action, il est néanmoins
possible de jouer sur la photopériode pour hâter la sortie de la jument de son inactivité ovarienne saisonnière.
3.2. Influence de la photopériode sur la saisonnalité de la jument
Tout d’abord il convient de préciser le rôle exact joué par cette photopériode. Il faut
savoir qu’un état réfractaire à la photopériode stimulante s’installe lorsque le traitement lumineux débute en été. Les juments manifestent alors une inactivité ovarienne hivernale mais sous jours artificiellement longs [48].
Inversement, un état réfractaire à la photopériode inhibitrice s’installe lorsque la jument est artificiellement maintenue sous jours courts, le traitement ayant débuté en hiver. Dans ce cas les juments manifestent une activité ovarienne en été mais sous jours artificiellement courts.
Dans ces deux situations, la phase de cyclicité est à peu près synchrone avec celle des juments sous photopériode naturelle. Cette installation d’un état réfractaire aux deux traitements lumineux est une mise en évidence indirecte de l’existence d’un rythme endogène circannuel et limite le rôle de la photopériode à un « synchroniseur » de ce rythme endogène.

- 19 -
Figure 5 : Schéma général de la transmission de l’information lumineuse de la rétine
aux gonadotrophines chez la jument (d’après Guillaume et al. [48]).

- 20 -
De nombreuses études [50, 51] ont permis d’établir le protocole de stimulation lumineuse le plus efficace pour obtenir une avancée de la saison de reproduction. La stimulation maximale de l’activité ovarienne (= délai le plus court obtenu entre le début du traitement et la première ovulation) est obtenue par l’application de 14 h 30 de lumière et 9 h 30 d’obscurité (fig. 6) ; moins de 13 heures et plus de 16 heures de lumière ne sont pas stimulantes. Une seule heure d’éclairement artificiel peut suffire à condition qu’elle soit appliquée à un moment particulier : 9h30 après la tombée de la nuit. Une phase photosensible est ainsi définie.
Un tel protocole, pour donner des résultats optimaux, doit être commencé en décembre, soit environ deux mois avant la date de la première ovulation escomptée [38, 44, 48, 75, 93]. Cependant, le temps de réponse varie considérablement d’une jument à l’autre (2 mois ± 10 j [44, 75, 93]). Pratiquement, on commence le traitement le 1er décembre (le démarrer plus tôt, en novembre, n’est pas plus efficace [93]) en éclairant les juments à l’aide d’une ampoule au néon d’une puissance de 200 W [44, 93] quelques heures avant le lever du soleil et quelques heures après la tombée de la nuit de façon à couvrir la période d’éclairement escomptée (entre 13 et 16 h).
3.3. Influence de la photopériode et de la saison sur la gestation et le moment du poulinage
L’influence saisonnière semble représenter la principale source de variation de la
longueur de la gestation [28]. Les juments saillies tôt dans la saison ont des gestations plus longues par rapport aux juments saillies plus tard dans l’année. Ainsi la gestation est de 8 à 10,4 jours plus longue, selon les études, pour les juments poulinant au printemps par rapport aux juments poulinant à l’automne [22, 60]. Ces constatations ont pu être reproduites expérimentalement de manière artificielle en faisant varier la photopériode : un groupe de juments exposées à 16 h d’éclairement pendant le dernier tiers de gestation a vu diminuer de 10 jours le délai entre ovulation et mise bas [22, 58].
C’est donc l’augmentation du temps d’éclairement pendant le dernier tiers de la
gestation qui en détermine la durée, le moment du part se trouvant ainsi soumis à une influence saisonnière. Par ailleurs, le fait que les poulinages se déroulent en général la nuit suggère l’existence d’autres facteurs intervenant sur le déclenchement de la mise bas.
En effet, environ 80 % des juments poulinent la nuit [22], la proportion étant plus marquée en été qu’en hiver, ce qui laisse supposer que la lumière est un facteur influant sur le déclenchement de la mise bas. Son déterminisme précis reste à explorer.

- 21 -
Figure 6 : Effet de différentes photopériodes expérimentales sur la stimulation de
l’activité ovarienne chez la jument. Le degré de stimulation est estimé en fonction du nombre de jours qui séparent le début du traitement (1er décembre) de la première ovulation. En moyenne, les juments stimulées ovulent 1 mois avant les autres. (d’après Guillaume et Palmer [51]).

- 22 -
4. Influence de la lune Il s’agit pour certains d’une évidence : la pleine lune serait responsable d’une
augmentation des naissances aussi bien chez l’homme que chez la jument ou bien d’autres espèces encore. La lune aurait, semble-t-il, une influence à la fois sur la fertilité et sur le jour de la mise bas.
Les études les plus nombreuses et portant sur les plus gros effectifs ont été menées chez l’homme. Malheureusement les résultats ne sont pas univoques. Tandis que l’une proclame une influence de la lune sur l’ovulation, la fertilité ou la date des naissances, l’autre réfute toute influence de quelque nature que ce soit [26, 43, 52, 85, 98].
Chez les bovins, Bohy [11, 12] a tenté d’étudier l’incidence de notre satellite sur la reproduction et en a conclu que l’ovulation des vaches semble plus fréquente pendant le deuxième quartier du cycle lunaire, mais aucun lien entre les dates de naissance et le cycle lunaire n’a pu être statistiquement établi.
La seule étude menée chez le cheval l’a été par Kollerstrom et Power [67], et elle conclut à une absence d’effet dans les conditions de leur expérience, notant que compte tenu de certains aspects astronomiques de révolution lunaire et solaire, une nouvelle étude se voulant plus pertinente devrait être menée sur 8 ans pour obtenir des résultats plus probants. Ils soulignent qu’une influence de la lune chez une espèce dont le cycle dure 21 jours environ alors que le cycle lunaire est de 28 jours est très peu probable, et que s’il fallait vraiment chercher quelque chose on pourrait se pencher sur une éventuelle influence de la lumière lunaire sur le système neuroendocrinien.
On ne peut donc rien conclure de définitif quant à l’influence lunaire sur la reproduction équine; dans le doute, mieux vaut pour le vétérinaire ne rien prévoir les soirs de pleine lune…

- 23 -
III. Alimentation
L’alimentation joue un rôle fondamental pour le bon fonctionnement de l’appareil reproducteur. La fonction de reproduction est en effet la première à pâtir d’un défaut de gestion alimentaire et ce souvent de manière insidieuse car sans autre signe apparent et de manière différée dans le temps. De plus la photostimulation évoquée juste avant est illusoire et sans effet sur une jument maigre. Ceci souligne donc l’influence du niveau alimentaire, mais il va de soi que la qualité de la ration a aussi son importance.
1. Relation entre la saison et l’alimentation. En élevage extensif, l’alimentation est elle-même dépendante de la saison, aussi bien
en terme de quantité que de qualité. L’hiver, les pâtures sont au repos, les chevaux sont nourris essentiellement à base de foin et d’aliments complémentaires. Dès le printemps, les animaux sont de nouveaux lâchés dans les pâtures avec souvent l’herbe pour seul aliment. La quantité et la qualité de cette herbe n’est pas non plus constante au cours de la saison : l’herbe jeune de printemps est riche en eau et en azote puis elle s’enrichit en cellulose et s’appauvrit en protéines.
La qualité du fourrage (herbe, foin, luzerne…) n’est pas non plus la même d’une année sur l’autre. Elle est le reflet des conditions climatiques au moment de la pousse aussi bien qu’au moment de la récolte et de la gestion des sols par son récoltant.
Il est donc recommandé de faire des analyses régulières de son fourrage afin d’adapter au mieux la ration.
2. Influence de l’état corporel Conséquence directe mais différée du niveau alimentaire, l’état corporel joue
beaucoup sur les capacités reproductrices de la jument. Si la maigreur extrême semble avoir des conséquences quasi-évidentes, l’embonpoint est tout aussi préjudiciable. Les travaux récents sur la leptine permettent de donner quelques explications.
2.1. Estimation de l’état corporel
Que ce soit dans le cas qui nous intéresse ici pour ajuster le régime de la jument ou
bien d’une façon générale pour bien alimenter un équidé, il est important d’être capable d’évaluer s’il est maigre, bien alimenté ou encore trop gras.
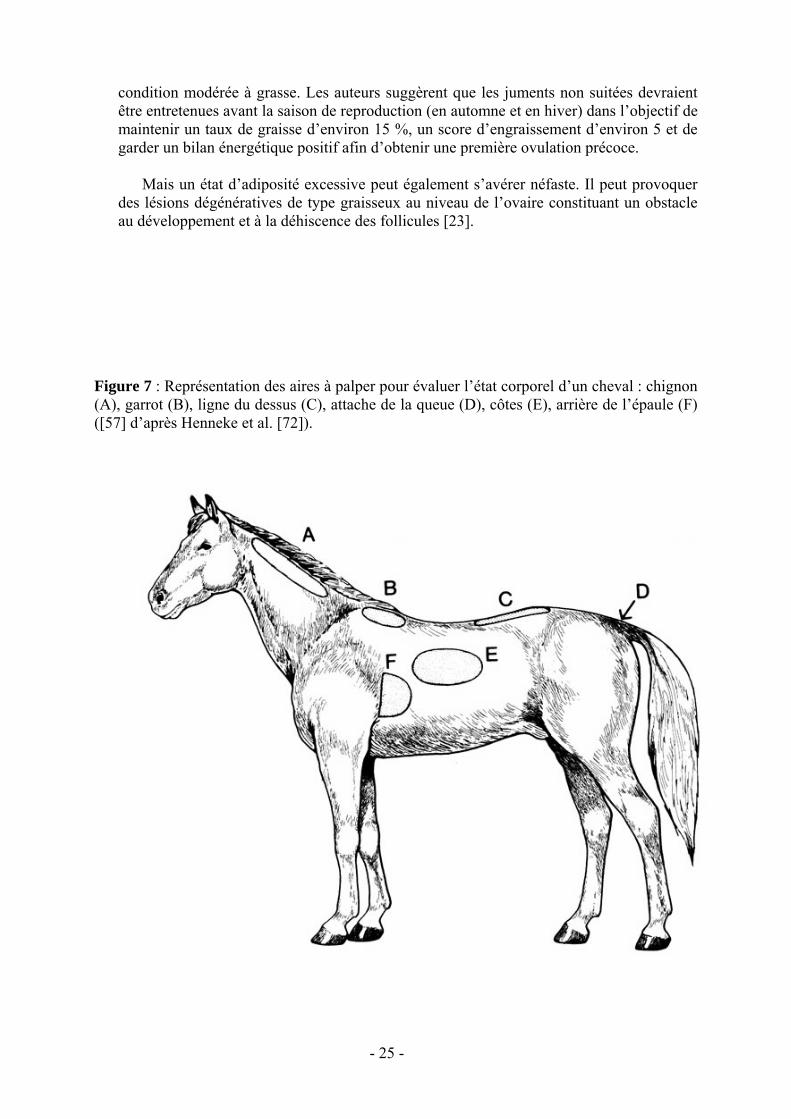
Pour évaluer l’état corporel, des critères visuels ont été définis. C’est Henneke et al. [57, 72] qui les premiers (1983) ont défini six régions anatomiques qu’il convient d’observer afin d’évaluer au mieux l’état d’embonpoint d’un cheval (fig. 7). Il s’agit du chignon (bord supérieur de l’encolure), du garrot, de la ligne du dos (ligne des processus épineux), de l’arrière de l’épaule, des côtes et de l’attache de la queue. En 1990, l’INRA [76] publie lui aussi un indice d’évaluation de l’état corporel des chevaux de sport et de selle et ajoute à cette liste un septième site qui est la croupe (tableau 2). Ces sites présentent l’avantage d’être relativement sensibles aux changements quantitatifs de la part de masse grasse corporelle. L’évaluation visuelle s’avérant insuffisante pour un bilan précis, elle s’accompagne systématiquement d’une palpation manuelle dont le but est d’estimer l’importance, la consistance et la forme des dépôts adipeux sous-cutanés dans ces régions.

- 24 -
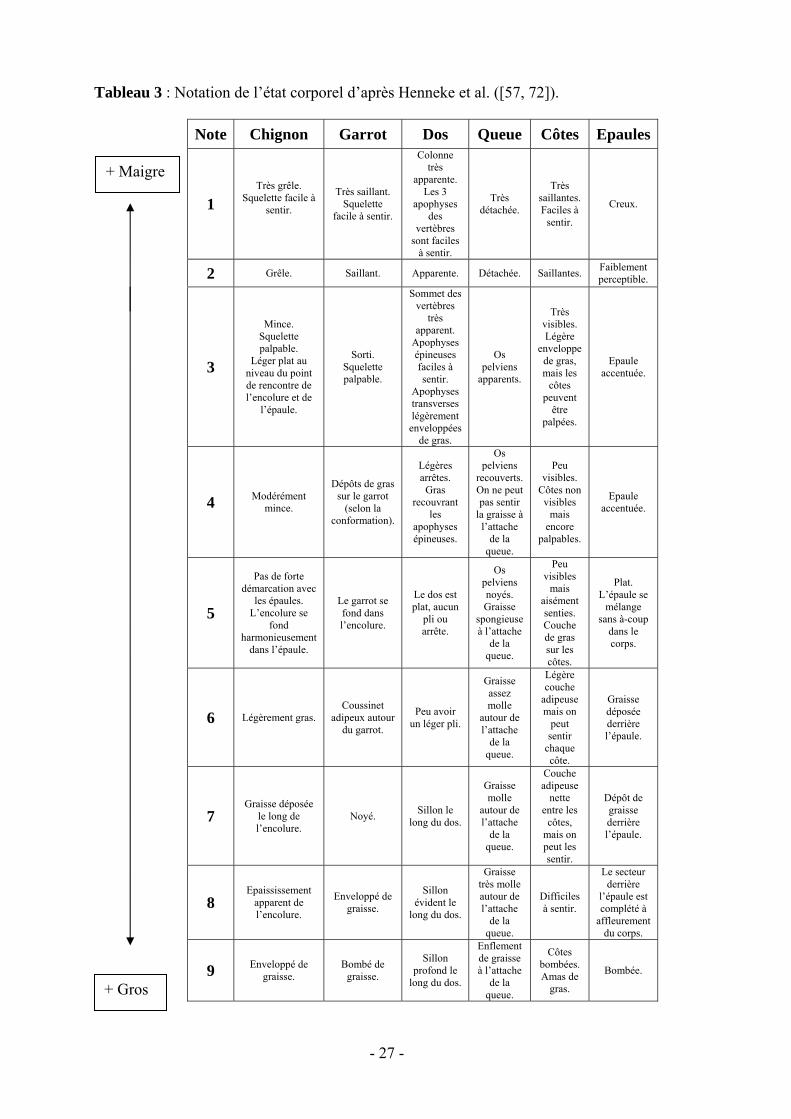
Si l’on suit l’échelle proposée par Henneke et al. (tableau 3), on assigne au cheval une note d’état corporel située entre 1 et 9 selon les résultats de l’observation visuelle et de la palpation. En allant de 1 à 9, les différents états correspondent respectivement à pauvre, très maigre, maigre, légèrement maigre, modéré, modérément charnu, charnu, gros, extrêmement gros. L’INRA propose quant à elle une échelle allant de 0 à 5 (tableau 2), la note 3 correspondant à un embonpoint normal.
Cette évaluation de l’état corporel prend toute son importance chez les poneys pour
qui les normes alimentaires utilisables chez les chevaux surévaluent les besoins d’entretien. Il convient d’ajuster leur alimentation donc de façon plus ou moins empirique faute de référence.
Enfin, on peut estimer le pourcentage de masse grasse corporelle de l’animal par
échographie [68]. La mesure s’effectue à mi chemin entre la pointe de la fesse (tuber ischiae) et le sommet de la hanche (tuber sacrale) à 5 cm de la colonne vertébrale. L’épaisseur de tissu adipeux (X en cm) relevée permet une estimation du pourcentage de gras corporel (Y en %) selon l’équation Y= 4.7 + 8.64 X [57].
2.2. Modifications associées aux variations de l’état corporel
Des variations de l’état corporel de la jument sont rapportées classiquement au cours
de son cycle [76]. On note en général une perte d’état en fin de gestation et début de lactation puis une reprise une fois passé le premier mois de lactation. Ces variations d’état corporel sont à mettre en relation avec des variations de poids : la jument prend du poids avec la croissance du fœtus durant les quatre derniers mois de gestation ; elle perd 10 à 14 % de son poids vif à la mise bas (fœtus, enveloppes, liquides) et peut encore en perdre en début de lactation [76].
2.2.1. Influence sur la cyclicité
Une observation faite depuis longtemps laissait déjà supposer une influence de l’état
corporel : l’inactivité ovarienne est plus longue chez les juments ayant allaité l’été précédent, c’est-à-dire dont l’état corporel a été plus ou moins fortement déprimé par les dépenses énergétiques que demande la lactation. A l’inverse, la phase d’inactivité ovarienne se trouve écourtée si les juments prennent du poids en début de printemps [49].
Henneke et al. [56] ont montré que l’intervalle entre la mise bas et la première ovulation est significativement plus long (2 jours pour la première ovulation, environ 10 jours pour la seconde) chez les juments qui ont un score d’engraissement inférieur à 5 comparées à celles dont le score d’engraissement est supérieur à 5. Il est également intéressant de noter que les jeunes juments (2 à 5 ans), qui sont physiologiquement moins grasses que leurs congénères plus âgées (toutes espèces confondues, un jeune individu contient plus d’eau et moins de graisse qu’un individu âgé), entrent en inactivité ovarienne plus tôt dans la saison contrairement aux dernières qui continuent leurs cycles plus ou moins longtemps au cours de l’hiver [42].
La relation entre l’apport énergétique et l’état d’engraissement pour les performances
reproductrices des juments non gestantes a été évalué par Kubiak et al. [68]. Ils constatent qu’un apport énergétique élevé (150 % des apports recommandés par le NRC à des juments n’ayant pas encore repris leur cyclicité en Mars) écourte la phase de transition chez des juments à état d’engraissement faible mais ne profite pas aux juments de
- 25 -
condition modérée à grasse. Les auteurs suggèrent que les juments non suitées devraient être entretenues avant la saison de reproduction (en automne et en hiver) dans l’objectif de maintenir un taux de graisse d’environ 15 %, un score d’engraissement d’environ 5 et de garder un bilan énergétique positif afin d’obtenir une première ovulation précoce.
Mais un état d’adiposité excessive peut également s’avérer néfaste. Il peut provoquer
des lésions dégénératives de type graisseux au niveau de l’ovaire constituant un obstacle au développement et à la déhiscence des follicules [23].
Figure 7 : Représentation des aires à palper pour évaluer l’état corporel d’un cheval : chignon (A), garrot (B), ligne du dessus (C), attache de la queue (D), côtes (E), arrière de l’épaule (F) ([57] d’après Henneke et al. [72]).

- 26 -
Tableau 2 : Notation de l’état corporel chez le cheval selon l’INRA. (d’après Martin-
Rosset [76]).
- 27 -
Tableau 3 : Notation de l’état corporel d’après Henneke et al. ([57, 72]).
Note Chignon Garrot Dos Queue Côtes Epaules
1 Très grêle.
Squelette facile à sentir.
Très saillant. Squelette
facile à sentir.
Colonne très
apparente. Les 3
apophyses des
vertèbres sont faciles
à sentir.
Très détachée.
Très saillantes. Faciles à
sentir.
Creux.
2 Grêle. Saillant. Apparente. Détachée. Saillantes. Faiblement perceptible.
3
Mince. Squelette palpable.
Léger plat au niveau du point de rencontre de l’encolure et de
l’épaule.
Sorti. Squelette palpable.
Sommet des vertèbres
très apparent.
Apophyses épineuses faciles à sentir.
Apophyses transverses légèrement enveloppées
de gras.
Os pelviens
apparents.
Très visibles. Légère
enveloppe de gras, mais les
côtes peuvent
être palpées.
Epaule accentuée.
4 Modérément mince.
Dépôts de gras sur le garrot
(selon la conformation).
Légères arrêtes.
Gras recouvrant
les apophyses épineuses.
Os pelviens
recouverts. On ne peut pas sentir
la graisse à l’attache
de la queue.
Peu visibles.
Côtes non visibles
mais encore
palpables.
Epaule accentuée.
5
Pas de forte démarcation avec
les épaules. L’encolure se
fond harmonieusement
dans l’épaule.
Le garrot se fond dans l’encolure.
Le dos est plat, aucun
pli ou arrête.
Os pelviens noyés. Graisse
spongieuse à l’attache
de la queue.
Peu visibles
mais aisément senties. Couche de gras sur les côtes.
Plat. L’épaule se
mélange sans à-coup
dans le corps.
6 Légèrement gras. Coussinet
adipeux autour du garrot.
Peu avoir un léger pli.
Graisse assez molle
autour de l’attache
de la queue.
Légère couche
adipeuse mais on
peut sentir
chaque côte.
Graisse déposée derrière l’épaule.
7 Graisse déposée
le long de l’encolure.
Noyé. Sillon le long du dos.
Graisse molle
autour de l’attache
de la queue.
Couche adipeuse
nette entre les
côtes, mais on peut les sentir.
Dépôt de graisse derrière l’épaule.
8 Epaississement
apparent de l’encolure.
Enveloppé de graisse.
Sillon évident le
long du dos.
Graisse très molle autour de l’attache
de la queue.
Difficiles à sentir.
Le secteur derrière
l’épaule est complété à
affleurement du corps.
9 Enveloppé de graisse.
Bombé de graisse.
Sillon profond le
long du dos.
Enflement de graisse à l’attache
de la queue.
Côtes bombées. Amas de
gras.
Bombée.
+ Maigre
+ Gros

- 28 -
2.2.2. Influence sur la gestation
Le taux de gestation chute significativement si la jument est sous-nourrie (correspondant à une note d’état corporel inférieure à 5 sur l’échelle de Henneke [56] ou à 2,5 sur celle de l’INRA [7, 8]). Cette diminution est majorée en cas de lactation concomitante.
L’état corporel joue sur le nombre de cycles nécessaires à l’obtention d’une gestation, car la sous-nutrition diminue non seulement le taux de conception (de 50 % au bout de trois cycles) mais également les chances de survie de l’embryon (75 % de mortalité embryonnaire contre 0 % chez des juments nourries pour couvrir leurs besoins (ce résultat exceptionnel, 0 %, est du à la petite taille de l’effectif)) [56].
Selon Henneke et al., la gestation ne semble pas influencée par l’état corporel (maigre
ou trop gros) [56] : le poids du foal à la naissance, sa taille ou bien sa circonférence cardiaque ne sont pas modifiés. Ce n’est pas l’avis de Wolter qui soutient qu’une suralimentation forte et prolongée augmente quelque peu le poids du nouveau-né et augmente légèrement le temps de gestation [115].
Il ne faut néanmoins pas chercher à engraisser coûte que coûte une jument constatée
trop maigre surtout après le part car la quantité d’aliment nécessaire pour obtenir un gain de poids dans une période de telles dépenses énergétiques dues à la lactation augmente de façon significative le risque de coliques.
L’obésité peut quant à elle poser de nombreux problèmes, et tout d’abord des
inconvénients comme des ennuis locomoteurs : un surpoids permanent majoré par celui d’une gestation sollicite énormément l’appareil locomoteur d’où l’apparition de boiteries. Ces gênes locomotrices, si elles sont marquées, peuvent devenir un véritable handicap pour la nutrition ou l’abreuvement avec des répercutions à terme inévitables sur la santé de la jument et/ou de son fœtus. Ensuite un problème se pose au moment de la mise bas. La jument obèse voit sa cavité pelvienne tapissée de dépôts adipeux qui rétrécissent parfois de façon significative la filière pelvienne empêchant le part de se dérouler normalement, augmentant ainsi le risque de dystocie.
2.2.3. Influence sur la lactation
Une jument maigre en lactation donne la priorité à l’allaitement de son poulain au
détriment d’une reprise éventuelle de sa cyclicité. Mais en cas de dénutrition sévère, la production laitière peut s’en trouver affectée au point de ne plus suffire à l’alimentation correcte d’un jeune en croissance.
L’obésité est aussi responsable d’une diminution de la production de lait. Une hypothèse avancée [69] serait que la jument obèse voit son appétit déprimé par les fortes réserves énergétiques qu’elle possède, par conséquent la production laitière s’en ressentirait.
L’état corporel joue donc essentiellement sur la quantité de lait produite mais ne semble pas influer de manière notable sur sa composition [69] du moins quantitative (pourcentages relatifs protéines/glucides/lipides).

- 29 -
Au final, l’INRA formule quelques recommandations pratiques [76]. A l’automne, la jument doit être en bon état (note d’état corporel de 3.5 sur 5) pour être alimentée à un moindre coût pendant l’hiver dans le cas des juments lourdes ou de loisir, ou pour permettre aux juments de course d’avoir une production laitière proche du potentiel génétique et sevrer un poulain ayant un développement maximum. Au poulinage, la note d’état corporel devrait être de 2.5 au minimum quel que soit le type de jument, mais d’autant plus élevée que la date de mise bas est précoce dans l’hiver. Au terme du premier mois de lactation la jument devrait avoir retrouvé une note d’état corporel de 3.
2.3. Déterminisme
Les interactions entre l’alimentation et la cyclicité font intervenir une hormone
découverte récemment [18, 59], la leptine. Cette protéine de 16 kD apparentée à la famille des cytokines est synthétisée et sécrétée principalement par les adipocytes, majoritairement par le tissu adipeux sous-cutané abdominal, plus que par le tissu adipeux intra-abdominal [18].
2.3.1. Rôle général de la leptine
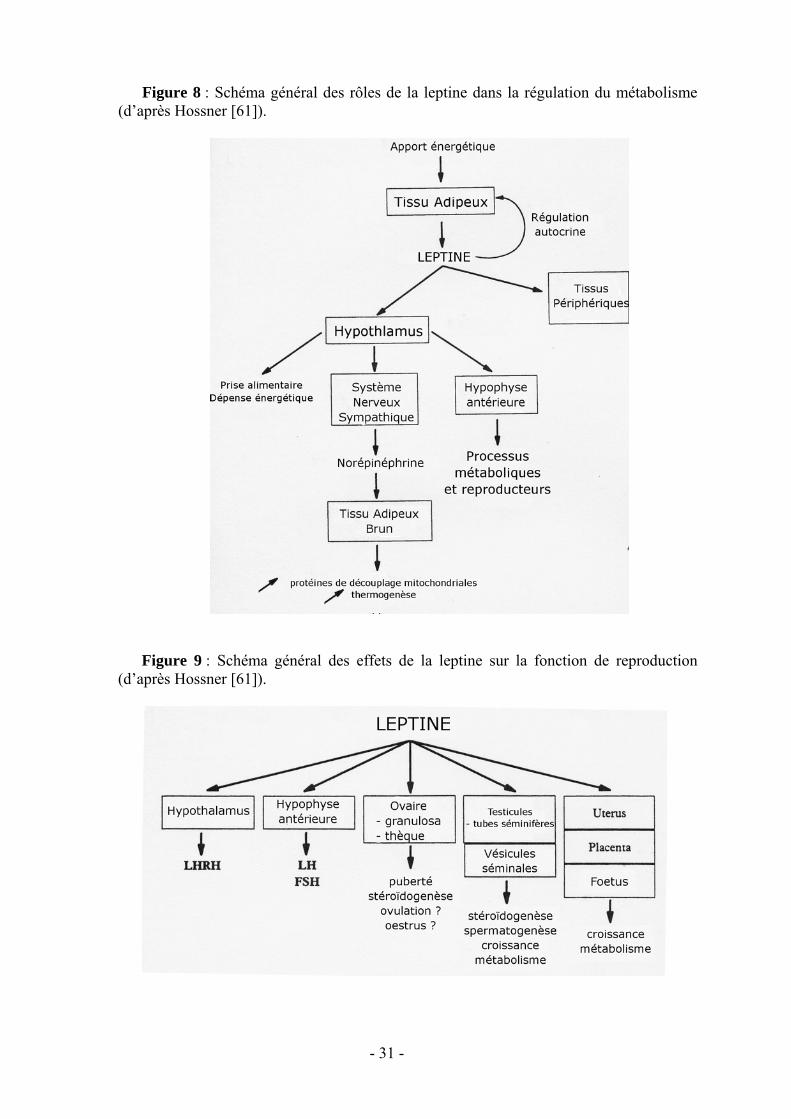
La leptine agit sur l’homéostasie générale (fig. 8) en réduisant la prise de nourriture,
en augmentant la dépense énergétique et la thermogenèse ce qui conduit à une perte de poids corporel. Son action s’exerce au moins en partie au niveau du système nerveux central. C’est un facteur satiétogène et brûleur de calories qui agit, entre autres, en inhibant l’expression hypothalamique du neuropeptide Y (NPY), ce dernier agissant lui-même comme stimulateur de l’appétit.
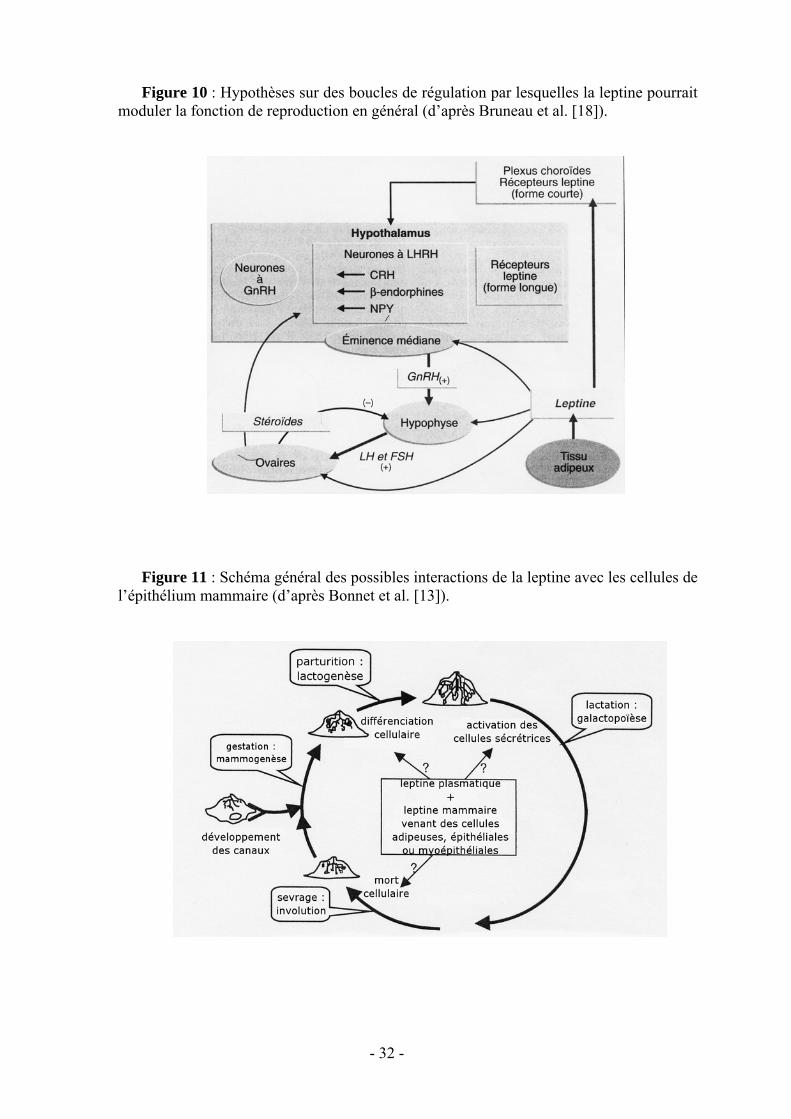
Les études sur son mode d’action [18] et sur ses récepteurs ont permis d’établir qu’elle agit aussi bien sur l’hypothalamus où elle règle la sécrétion pulsatile de la LHRH et module sa sensibilité à l’œstradiol, sur l’hypophyse où elle module la sécrétion des gonadotrophines que directement sur les gonades (fig. 9 et 10). La leptine pourrait donc constituer un signal nécessaire au système nerveux central pour déclencher la puberté et, à l’âge adulte, intervenir dans la décision d’ovuler, en fonction de l’état des réserves adipeuses. Il est intéressant de noter l’existence de taux plasmatiques élevés de leptine chez les jeunes individus reflétant une certaine résistance à la leptine. Celle-ci permettrait de maintenir un bon niveau de prise alimentaire nécessaire à la croissance et d’empêcher la survenue trop précoce de la puberté [59].
Sa sécrétion corrélée à la masse graisseuse corporelle et à la taille des adipocytes lui confère un rôle d’adipostat capable de renseigner l’hypothalamus de la femelle sur l’état des réserves énergétiques à long terme et sur leur compatibilité avec la menée à terme d’une gestation et d’une lactation ultérieure [59].
La leptine semblerait également agir sur la mammogenèse, la lactogenèse ainsi que sur la sécrétion du colostrum [13] (fig. 11). On évoque même des rôles de la leptine sur le nouveau-né, au moins au niveau de son tractus intestinal puisqu’on y a retrouvé des récepteurs à la leptine.
2.3.2. La leptine dans l’espèce équine
Les études menées chez l’homme et le singe laissent à penser que la leptine joue un
rôle plus ou moins important selon l’espèce. Sa découverte relativement récente, chez la souris, fait que peu d’études se sont penchées sur son implication dans l’espèce équine. Elle a cependant bel et bien été mise également en évidence chez le cheval [41, 42].

- 30 -
Cependant, les travaux de Gentry et al. [41, 42] chez le cheval ont montré une variation saisonnière du taux de leptine plasmatique : la concentration moyenne augmente de juillet à septembre et diminue rapidement d’octobre à décembre. On note également une variation circadienne du taux de leptine plasmatique. Le taux circulant est plus important chez les juments grasses comparé aux juments maigres, il est aussi plus marqué chez les juments adultes que chez les jeunes. Mais la présence de variations importantes au sein même des juments grasses souligne le fait que d’autres facteurs que le taux de masse grasse régulent son expression. En effet on sait déjà que le cortisol et l’insuline [97] sont des stimulants possibles de son expression, et que les agonistes β-adrénergiques, l’AMPc et les thiazolidinediones (classe de médicaments pour diabétiques de type 2 augmentant la sensibilité des cellules à l’insuline) la freinent [59].
La leptine semble donc être l’une des clefs de la relation nutrition-reproduction. Sa découverte récente soulève pour le moment plus de questions auxquelles il faut encore répondre qu’elle ne donne de réponses. Notamment son implication dans l’espèce équine reste à investiguer.
Au final, si l’on s’écarte des recommandations faites par l’INRA [76] concernant l’état
corporel de la jument, on constate que les effets négatifs sont bien plus importants que les hypothétiques avantages. Le déterminisme de cette gestion à long terme de la nutrition reste à préciser chez la jument.
- 31 -
Figure 8 : Schéma général des rôles de la leptine dans la régulation du métabolisme (d’après Hossner [61]).
Figure 9 : Schéma général des effets de la leptine sur la fonction de reproduction
(d’après Hossner [61]).
- 32 -
Figure 10 : Hypothèses sur des boucles de régulation par lesquelles la leptine pourrait moduler la fonction de reproduction en général (d’après Bruneau et al. [18]).
Figure 11 : Schéma général des possibles interactions de la leptine avec les cellules de
l’épithélium mammaire (d’après Bonnet et al. [13]).

- 33 -
3. Influence du niveau alimentaire, de la qualité des nutriments et de l’équilibre nutritionnel
Selon Wolter [113, 114, 115], la malnutrition est un des responsables majeurs de
l’infertilité de la jument dont l’importance est largement sous-estimée, d’où une prévention insuffisante. Cette tendance au sous-diagnostic est principalement due au fait que l’infertilité nutritionnelle est souvent différée par rapport à la période de sous-alimentation.
Nous envisagerons donc ici les particularités de la jument gestante ou allaitante. Il s’agit d’étudier en quoi les besoins de production que constituent l’élaboration d’un fœtus ou la production laitière diffèrent des simples besoins d’entretien et en quoi le non respect de ces exigences alimentaires peut déprécier le potentiel reproducteur de la jument.
3.1. Influence du niveau alimentaire
Les besoins nutritionnels totaux d’une jument poulinière sont constitués de ses besoins d’entretien, des besoins de gestation et/ou de lactation et éventuellement des besoins de croissance pour les pouliches poulinant pour la première fois à quatre mais surtout à trois ans [76]. L’annexe récapitule les besoins de la jument aux différents stades de sa vie de reproductrice.
Ce n’est vraiment que durant les quatre derniers mois de gestation que les besoins nutritionnels de la mère diffèrent vraiment des besoins d’entretien [76, 81]. En effet, pendant les sept premiers mois de gestation, le gain de poids des juments, lié à la croissance du fœtus, est faible. Il est maximal au cours des trois mois précédant le poulinage et représente 8 à 10 pourcent du poids vif de la jument mesuré 24 heures après la mise bas [76]. La croissance du fœtus se répartit, du huitième au onzième mois, à raison de 18, 19, 23 et 23 % du poids du poulain nouveau-né [Meyer et Ahlswede (1976) cités par 65].
Afin de satisfaire cette augmentation des besoins, une supplémentation est nécessaire, surtout si l’état corporel de la jument est jugé insuffisant, et en particulier si pour des raisons économiques une restriction des apports alimentaires a été effectuée en hiver dans le cas des animaux élevés pour leur viande [76]. Cette supplémentation pallie les déficits que la simple augmentation du niveau d’ingestion du fourrage n’arrive pas à compenser. Elle permet aussi d’ajuster les apports en vitamines et oligo-éléments. En pratique, on ajoute progressivement de 1 à 3 kg de complément de fin de gestation du huitième au onzième mois.
Un apport énergétique en excès par rapport à la couverture des dépenses n’affecte pas sensiblement le poids du poulain à la naissance, mais se traduit par un engraissement des juments [65]. Une restriction alimentaire modérée par l’apport de 80 % environ des besoins n’a aucune influence sur le poids des poulains à la naissance, leur vitalité ou la santé de la mère [65]. Banach et Evans [cités par 65] ont distribué à des juments de selle pendant les trois derniers mois précédant le poulinage un régime couvrant 55 % des recommandations énergétiques du NRC, le besoin azoté restant largement couvert. Ils n’ont observé aucune variation du poids à la naissance du poulain ou de la durée de gestation. En revanche, les juments ont perdu 900 g/j de masse corporelle.
Il est donc possible de sous-alimenter fortement les juments en fin de gestation à condition d’apporter suffisamment d’azote, de minéraux et de vitamines.. La jument est capable de puiser amplement dans ses réserves lipidiques corporelles pour assurer le développement du fœtus. Cette restriction énergétique n’est possible évidemment que si les juments sont en bon état corporel au début de la période de restriction [65]. De plus si

- 34 -
les juments sont grasses, on peut craindre un syndrôme hyperlipémique se traduisant par de la diarrhée, de l’anorexie, de la somnolence, de l’œdème ventral pouvant conduire à la mort des sujets [39, 81]. La lipolyse est alors excessive et le foie est dépassé devant l’excès de triglycérides (stéatose hépatique) synthétisés laissant s’installer l’acidose métabolique.
Les apports recommandés pour une jument en début de lactation sont évalués d’après
la quantité d’énergie exportée dans le lait [65, 35, 76, 81] et sont récapitulés dans l’annexe. La connaissance des besoins à mi-lactation est limitée et les juments retournent souvent en pâture à cette période [35].
Un accroissement des apports énergétiques se traduit le plus souvent par une
augmentation de la production laitière et par un gain de poids de la jument [65]. Plusieurs auteurs [35, 102] rapportent les effets bénéfiques d’une balance énergétique positive : stimulation de l’activité ovarienne et sortie plus précoce de la phase inactivité ovarienne saisonnière. Une observation du même ordre est effectuée par Henneke et Kreider [55] qui constatent une première ovulation post-partum plus précoce lorsque la tétée du poulain est limitée. Néanmoins une suralimentation énergétique chez des juments non allaitantes n’aurait pas d’effet favorable sur le taux de fécondation [65]. Un apport énergétique supérieur aux apports recommandés n’apporte donc qu’un bénéfice limité à mettre en balance avec le coût d’une telle supplémentation. On peut la réserver aux juments allaitantes dont l’état corporel est jugé insuffisant et à celles qui poulinent tard en saison afin de hâter la nouvelle fécondation.
Les effets d’une sous-alimentation énergétique ne font pas l’unanimité. Une sous-
alimentation modérée semble être responsable d’une diminution de la production laitière [65]. Henneke, Potter et Kreider [cités par 65] ne rapportent pas d’influence des apports énergétiques sur les performances reproductrices des juments en bonne condition alors que Van Niekerk et Van Heerden [102] affirment qu’une sous-alimentation énergétique retarderait l’apparition des chaleurs de lait et augmenterait le taux de mortalité embryonnaire si elle se poursuit après la fécondation.
D’un point de vue pratique, Doreau et al. [32] ont montré que la jument en début de lactation est capable d’accroître son niveau d’ingestion si on lui fournit un fourrage de qualité moyenne à volonté, limitant ainsi à 5 % de la matière sèche ingérée l’apport de concentrés. Le choix du rapport fourrage/concentrés n’est donc pas une donnée fixe et immuable mais le résultat d’un choix de l’éleveur compte tenu de ses stocks de fourrage (ou des pâtures disponibles), de la qualité de ceux-ci et de son budget alloué aux concentrés. L’intérêt des concentrés est d’apporter ce que les fourrages ne fournissent pas ou alors en quantité insuffisante.
Retenons au final que les performances reproductrices de la jument sont optimales
lorsque chez une jument en bon état corporel, que les apports recommandés sont respectés et que, bien que la jument soit capable de mobiliser ses réserves adipeuses, la sous nourrir ne peut que nuire à ses performances [35].
Il est important de noter que toutes ces adaptations du niveau alimentaire doivent se
faire sans changement brutal de régime sous peine d’engendrer des désordres digestifs. Il existe cependant un cas particulier où un changement soudain d’alimentation est
parfois préconisé [20, 80, 83, 110] : les gestations gémellaires. La restriction alimentaire imposée consiste à arrêter la distribution d’aliments complets et de céréales et à n’apporter qu’un peu de foin de prairie. On associe généralement ce traitement à un confinement de

- 35 -
la jument au box en permanence. Un tel régime est réputé favoriser la résorption d’un des deux embryons et nécessite un suivi échographique régulier de la jument afin de rétablir le plus vite possible un régime adapté à la gestation. Le taux de succès accordé à cette méthode est d’environ 60 % [20, 80]. Mais certains auteurs [20, 83] pensent que la réussite de ce protocole n’est en fait due qu’à la résorption embryonnaire naturelle responsable chez la jument de l’élimination d’une des vésicules dans 85 % des cas pour peu que la gestation soit unilatérale. On sait en effet qu’en cas de gestation bilatérale il n’y a pas de résorption spontanée à attendre [20]. Cette technique est donc très contraignante et sans réelle efficacité prouvée, elle augmente le risque de perdre les deux vésicules même si le suivi échographique est fait régulièrement.
Un niveau alimentaire bien adapté bénéficie à la fois à la sécrétion lactée et à la
fertilité mais ces derniers sont aussi grandement tributaires de l’équilibre de la ration.
3.2. Influence des apports protéiques
3.2.1. Apport quantitatif La valeur azotée des aliments est exprimée en g de MADc (Matière Azotée Digestible
cheval) qui correspond à une évaluation de la quantité et de la qualité des matières azotées effectivement digérées par le cheval. Le système MADc évalue le coefficient d’utilisation digestive des matières azotées apportées dans la ration.
L’étude de Meyer et Ahlswede [cités par 65] fournit une base quantitative pour estimer le besoin supplémentaire engendré par la gestation. Le fœtus se développe rapidement à partir du huitième mois. La quantité de protéines fixée est successivement de 14, 22, 23 et 31 g au cours des 8ème, 9ème, 10ème et 11ème mois pour un poids d’environ 60 kg à la naissance. Il faudrait majorer ces besoins d’environ 20 % pour tenir compte des quantités fixées parallèlement dans les annexes fœtales et dans les tissus maternels [Meyer et Ahlswede (1976) cités par 65].
Les besoins azotés de lactation sont calculés à partir de la quantité de protéines exportées par jour et du rendement de l’utilisation métabolique des matières azotées digestibles pour cette synthèse [65] (60 % des MADc apportées par l’alimentation seront effectivement utilisées par la jument pour la croissance du poulain). Les apports recommandés sont récapitulés dans l’annexe.
La jument est capable de mobiliser des protéines de son organisme pour couvrir ses
dépenses azotées lorsque l’apport alimentaire est insuffisant. Cela permet des économies d’aliment pendant la période hivernale [65]. Cependant, en l’absence de données précises sur l’influence d’un déficit protéique de la ration sur les performances reproductrices de la jument, il est plus prudent de limiter le recours à cette mobilisation. En effet il a déjà été montré que le taux de progestérone circulant dépend du taux protéique de la ration [81]. On ne connaît pas de seuil minimum. Les travaux de Web et al. [cités par 34] ainsi que ceux de Doreau et al. [34] semblent indiquer qu’aucune diminution des performances reproductrices n’est à mettre en relation avec une restriction azotée. Le taux de fécondation reste le même que lorsque les besoins sont couverts [34] mais l’on note une tendance à l’augmentation de l’intervalle poulinage-première ovulation [65] ainsi qu’une diminution du taux de fécondation lors de la chaleur de poulinage [65].
Au bilan, on ne connaît pas précisément les effets d’une restriction azotée sur la fertilité. Les données connues suggèrent un effet négatif [65].

- 36 -
Concernant la jument allaitante, la couverture des besoins en comparaison avec un régime pauvre en azote favoriserait la teneur en matières azotées du lait et la production laitière [34, 81] et donc par là même la croissance des poulains, au moins durant le premier mois car au-delà l’ingestion d’herbe par le poulain lui permet de compenser ce déficit [34].
Peu de données concernent l’influence d’un excès azoté sur la jument reproductrice. Wolter [115] suggère qu’un excès protéique favoriserait l’intoxication de l’embryon par les produits issus de la dégradation de l’azote tel que l’ammoniac. Quoiqu’il en soit, un apport excédentaire est un non-sens économique pour l’éleveur.
3.2.2. Apport qualitatif
Les besoins précis du cheval en acides aminés indispensables ne sont pas connus à
l’heure actuelle. On suppose simplement qu’à l’instar des autres espèces animales il n’est pas capable de synthétiser, ou à une vitesse insuffisante, les neufs acides aminés dits, de ce fait, indispensables : leucine, isoleucine, valine, méthionine, phénylalanine, thréonine, lysine, tryptophane et histidine [65].
Ce sont essentiellement les travaux de l’équipe de Van Niekerk [103, 104, 105, 106, 107, 108, 109] qui ont fait avancer les connaissances sur ce point chez le cheval. Ces études montrent que les protéines de bonne qualité (riches en thréonine, méthionine, isoleucine, leucine, lysine et arginine) stimulent la sécrétion de LH et de FSH permettant ainsi une sortie de la phase de transition plus précoce (entre deux semaines et un mois plus tôt).
Des protéines de bonne qualité entraînent un meilleur taux de conception (les juments supplémentées remplissent dès leur premier cycle contre une moyenne de 2,5 pour les autres) et une diminution des chaleurs anormales. Elles préviennent les pertes embryonnaires précoces (7,5 % de mortalité embryonnaire précoce chez les juments supplémentées contre 35,7 % pour les autres) en augmentant le taux de progestérone circulant généré par le corps jaune primaire par stimulation de la sécrétion de LH.
Elles influent peu sur le délai d’apparition des chaleurs de lait. Du fait de sa production de protéine riches en acides aminés indispensables, la jument
en lactation devrait avoir des besoins accrus. Or on ne dispose d’aucune expérience à ce sujet si ce n’est qu’une supplémentation de la ration en lysine ou en méthionine entraîne une augmentation des teneurs sanguines correspondantes [Rogers et al. cités par 34 et 65].
L’utilisation d’azote non protéique (urée, ammoniac) par le cheval est sujette à
controverse [81]. Quand bien même le cheval serait capable d’utiliser une faible partie de l’azote non protéique de sa ration cela ne suffirait pas à combler les besoins d’une jument en lactation ou d’un poulain en croissance. L’urée, bien que tolérée par le cheval, n’a aucun intérêt nutritionnel chez le cheval. La production laitière quantitative de la jument est d’ailleurs diminuée lorsque la majeure partie de l’apport azoté est fait sous forme d’urée.
Dans la gestion de l’alimentation de la jument, il est important de noter que l’apport
protéique de la ration est différent du pourcentage de protéines présentes dans celle-ci. C’est tout l’intérêt du système MADc qui tient compte de la quantité de protéines effectivement utilisables par la jument en fonction du type d’aliment fourni. Les recommandations formulées dans l’annexe 1 tiennent compte du coefficient d’utilisation digestive et du rendement métabolique (encore appelé valeur biologique).

- 37 -
D’un point de vue pratique, une ration pour jument au dernier tiers de sa gestation constituée d’un mauvais foin et d’avoine ne fournira pas quantitativement et qualitativement assez de protéines (ni de calcium).
3.3. Influence de l’apport lipidique
L’incorporation de lipides à la ration de la jument reproductrice peut sembler
intéressante, particulièrement lorsque les besoins énergétiques sont élevés, en fin de gestation ou en début de lactation.
Les chevaux acceptent facilement l’addition de matières grasses à leur ration pourvu que celles-ci ne soient pas rances [76]. Un délai de 15 à 20 jours est nécessaire pour une bonne adaptation du cheval à l’utilisation de cette forme d’énergie [9]. En général, on note une légère diminution de la prise alimentaire associée à un régime riche en graisses (10% en comparaison d’une alimentation naturelle contenant 2 % de matières grasses) [29, 76].
Concernant la jument, l’addition de lipides à la ration de juments en bon état général ne semble pas hâter la reprise de leur cyclicité [76]. L’incorporation de matières grasses (huile de maïs riche en acide linoléique chef de file des ω-6) au régime des juments allaitantes permet une vitesse de croissance de leurs foals plus importante par rapport aux foals des juments non supplémentées [76].
Les travaux de Davidson et al. [29] indiquent une légère tendance à l’amélioration des performances reproductrices de la jument (diminution du délai d’apparition des chaleurs de lait, augmentation du taux de fécondation) lors d’incorporation de matières grasses (soja riche en acide α linolénique chef de file des ω-3) à la ration bien que ces résultats ne soient pas significatifs.
Ce régime s’accompagne également d’une augmentation du taux de matières grasses du lait et d’une vitesse de croissance des foals plus rapide lors de leur première semaine de vie.
Les besoins quantitatifs des équidés en acides gras essentiels ne sont pas connus à l’heure actuelle. Néanmoins, du point de vue qualitatif, le NRC [81] recommande l’incorporation dans toute ration d’au moins 0,5 % d’acide linoléique.
3.4. Influence de la composition minérale de la ration
Les effets sur la reproduction de la jument d’un déficit en tel ou tel minéral sont très
mal connus [76]. Nous savons seulement que les besoins en certains minéraux évoluent en fonction du stade reproducteur de la jument. Il s’agit bien évidemment du calcium et du phosphore nécessaires à la minéralisation du squelette du fœtus et dont les besoins augmentent pendant le dernier tiers de la gestation et la lactation. Les besoins en potassium, sodium et magnésium augmentent aussi aux mêmes périodes mais dans une moindre proportion [81, 95].
Les besoins de la jument en fonction du stade de son cycle sont récapitulés dans l’annexe.
Seuls les effets d’un défaut ou d’un excès d’iode sont un peu mieux connus chez la jument. La carence d’une ration en iode est favorisée par l’ingestion de plantes renfermant des thiocyanates (crucifères), de protéines de soja insuffisamment traitées par la chaleur, une carence ou un excès en cobalt, une carence en vitamine A, une eau riche en calcium ou en magnésium, ou encore par la vie en zone montagneuse [23]. Un défaut d’iode peut entraîner l’apparition de cycles anormaux chez la jument [81]. Les poulains issus de mères carencées naissent goitreux, faibles et sont plus sensibles à la mortinatalité [15]. Il faut cependant se garder d’une complémentation à l’aveugle car les symptômes d’un excès

- 38 -
d’iode (au-delà de 40 mg par jour) sont plus ou moins semblables à ceux d’une carence. Il faut donc bien identifier la nature du trouble avant de corriger la teneur en iode de la ration dans un sens ou dans l’autre [15].
On pense également qu’un déficit en cuivre augmenterait l’incidence de rupture de l’utérus [18b] et l’artère utérine chez les juments âgées [81].
Voss et Pickett [111] ont étudié l’influence d’un apport minéral supérieur aux recommandations du NRC sur les capacités reproductrices de la jument. Aucune conséquence n’a été notée sur le nombre d’inséminations artificielles par cycle, le pourcentage de juments pleine par cycle, sur la fécondation ou sur le poids des juments. Vingt ans plus tard, Ott et Asquith [84] confirment ces résultats.
Wolter [115] affirme que le phosphore, le manganèse et le zinc jouent un rôle important dans la fertilité de la jument en stimulant la production anté-hypophysaire de gonadotrophines. Malheureusement aucune étude n’étaye ces propos. Brochart [17] rapporte quand à lui un rôle du cobalt et du sélénium chez les ruminants. Le cobalt permettrait d’augmenter le taux protéique du lait chez la brebis ; le sélénium favoriserait le taux d’ovulations doubles chez la vache. Cobalt et sélénium seraient de plus tous les deux tératogènes à forte dose.
Les besoins précis de la jument sont donc mal connus ainsi que les effets d’éventuelles carences ou excès. D’un point de vue pratique il convient de toujours fournir un complément minéral sous forme d’un bloc à lécher qui permettra de compenser les éventuels déficits de la ration. Le risque d’atteindre les doses toxiques en pratiquant de la sorte est négligeable.
3.5. Influence de l’apport vitaminique
Les besoins en vitamines des juments reproductrices sont également mal connus. Seule la vitamine A semblerait avoir une action directe sur l’activité reproductrice de la jument bien que ceci soit également controversé [81]. La supplémentation en β-carotène (à raison de 100 mg/j) de juments en stabulation permettrait de renforcer l’expression des chaleurs, d’augmenter le taux de conception et de diminuer le taux de mortalité embryonnaire précoce.
Cette supplémentation serait sans effets sur des juments bénéficiant de fourrages déjà riches en carotène comme l’herbe ou la luzerne [81]. Un excès prolongé de vitamine A chez la jument gestante serait quand à lui responsable de malformations fœtales [81].
3.6. Reproduction et substances indésirables présentes dans les aliments
3.6.1. Plantes susceptibles d’interférer avec la reproduction
Outre les plantes responsables d’intoxications graves chez le cheval capables d’altérer
la fertilité et d’entraîner la mort du fœtus chez une jument fortement débilitée, certaines plantes sont connues pour être tératogènes et pour leur capacité à causer l’avortement des juments [73]. Ces plantes sont répertoriées dans la première partie du tableau 4.
D’autres plantes listées dans la deuxième partie du même tableau sont connues pour être abortives et altérer la fertilité du bétail. Elles peuvent théoriquement être à l’origine des mêmes symptômes chez la jument [73].
Cette liste est loin d’être exhaustive mais peu d’études ont été consacrées à l’influence des plantes toxiques sur la fonction de reproduction en général et chez le cheval en particulier.
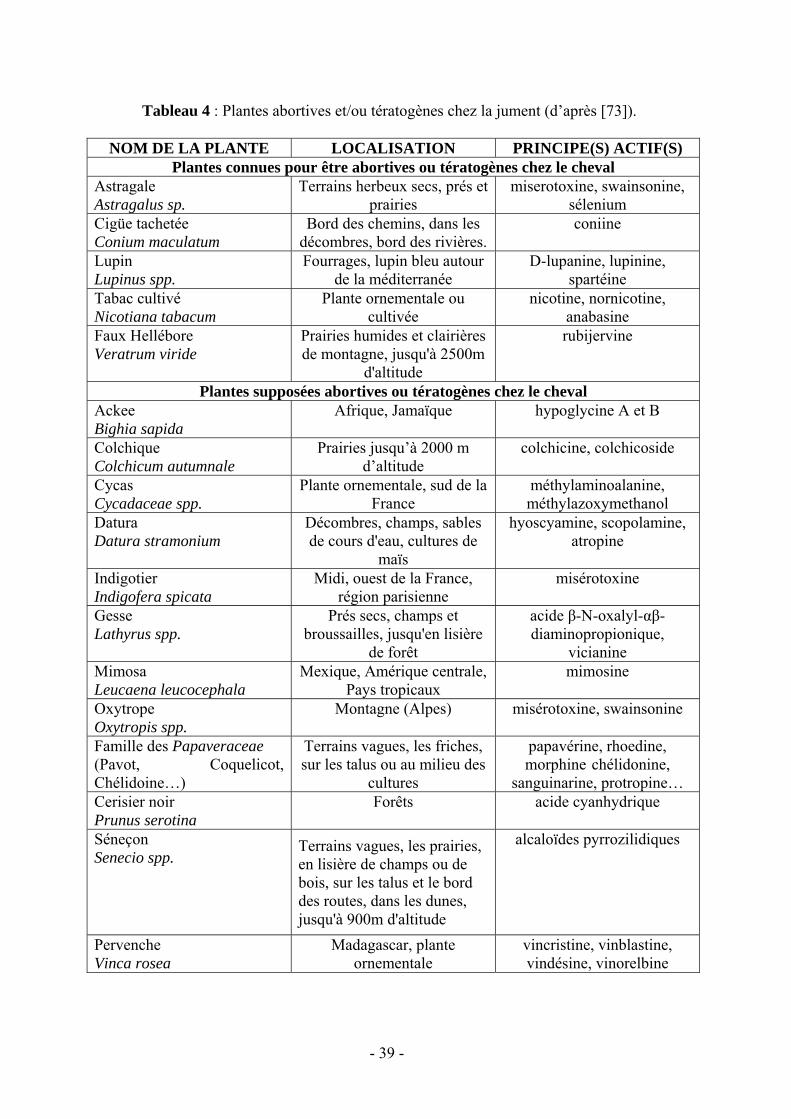
- 39 -
Tableau 4 : Plantes abortives et/ou tératogènes chez la jument (d’après [73]).
NOM DE LA PLANTE LOCALISATION PRINCIPE(S) ACTIF(S) Plantes connues pour être abortives ou tératogènes chez le cheval
Astragale Astragalus sp.
Terrains herbeux secs, prés et prairies
miserotoxine, swainsonine, sélenium
Cigüe tachetée Conium maculatum
Bord des chemins, dans les décombres, bord des rivières.
coniine
Lupin Lupinus spp.
Fourrages, lupin bleu autour de la méditerranée
D-lupanine, lupinine, spartéine
Tabac cultivé Nicotiana tabacum
Plante ornementale ou cultivée
nicotine, nornicotine, anabasine
Faux Hellébore Veratrum viride
Prairies humides et clairières de montagne, jusqu'à 2500m
d'altitude
rubijervine
Plantes supposées abortives ou tératogènes chez le cheval Ackee Bighia sapida
Afrique, Jamaïque hypoglycine A et B
Colchique Colchicum autumnale
Prairies jusqu’à 2000 m d’altitude
colchicine, colchicoside
Cycas Cycadaceae spp.
Plante ornementale, sud de la France
méthylaminoalanine, méthylazoxymethanol
Datura Datura stramonium
Décombres, champs, sables de cours d'eau, cultures de
maïs
hyoscyamine, scopolamine, atropine
Indigotier Indigofera spicata
Midi, ouest de la France, région parisienne
misérotoxine
Gesse Lathyrus spp.
Prés secs, champs et broussailles, jusqu'en lisière
de forêt
acide β-N-oxalyl-αβ-diaminopropionique,
vicianine Mimosa Leucaena leucocephala
Mexique, Amérique centrale, Pays tropicaux
mimosine
Oxytrope Oxytropis spp.
Montagne (Alpes) misérotoxine, swainsonine
Famille des Papaveraceae (Pavot, Coquelicot, Chélidoine…)
Terrains vagues, les friches, sur les talus ou au milieu des
cultures
papavérine, rhoedine, morphine chélidonine,
sanguinarine, protropine… Cerisier noir Prunus serotina
Forêts acide cyanhydrique
Séneçon Senecio spp.
Terrains vagues, les prairies, en lisière de champs ou de bois, sur les talus et le bord des routes, dans les dunes, jusqu'à 900m d'altitude
alcaloïdes pyrrozilidiques
Pervenche Vinca rosea
Madagascar, plante ornementale
vincristine, vinblastine, vindésine, vinorelbine

- 40 -
3.6.2. Les phytoestrogènes
Il s’agit de substances à activité oestrogéniques que l’on retrouve dans certaines plantes : trèfles, luzerne, graines de soja, dactyle, fétuque [53, 79].
Leur influence chez la jument est méconnue. Woodley et al. [112] ont étudié l’influence d’un traitement à base d’oestradiol sur la
lutéolyse chez la jument. Ils n’ont noté aucune modification quand au délai d’apparition de celle-ci. Ils notent cependant qu’une dose quotidienne de 10 mg d’oestradiol limite la croissance folliculaire et inhibent l’ovulation.
De plus amples travaux sont nécessaires afin de déterminer quelles sont les doses potentiellement absorbables par les chevaux et leur réelle influence sur la jument.
3.6.3. Les mycotoxines
Les mycotoxines sont des substances produites par une grande variété de moisissures
se développant sur différents types d’aliments bruts ou transformés. Les mycotoxines constituent un groupe de substances présentant des activités mutagènes, cancérogènes, tératogènes, immunogènes et œstrogéniques [53]. Elles ne sont pas transmissibles d’animal à animal et leur ingestion ne répond à aucun traitement antibiotique. La plupart sont toxiques mais pas létales aux doses auxquelles on les trouve dans l’environnement [82]. Elles peuvent en revanche être plus concentrées dans les aliments secs ou composés, les toxines étant restées alors que les moisissures les ayant sécrété ont disparu.
Deux champignons sont principalement en cause lors d’avortement chez la jument : - Claviceps purpurea : ce mycélium se loge dans le caryopse du seigle, de l’orge
ou du froment et y développe une excroissance. Elle sécrète des substances alcaloïdes qui s’avèrent être de puissants abortifs [53] : ergonovine, metilergonovine, ergotamine, ergocristine, ergocornine, ergocriptine et bromocriptine [24].
- Acremonium coenophialum : ce champignon endophyte parasite principalement la fétuque mais aussi parfois l’ivraie ou le ray-grass. Les toxines produites par le mycélium qui ont été identifiées sont la pyrrolizidine, pyrrolopyrazine et l’ergovaline [96].
Les principaux symptômes rencontrés chez la jument ayant ingéré un fourrage
contaminé sont [27, 86, 96] : un allongement du temps de gestation (entre deux semaines et trois mois), une absence de développement de la mamelle accompagnée d’agalactie, une augmentation du taux de mortalité de la mère et du foal du à la diminution de la sécrétion de lait, une diminution des taux sériques de progestérone et de prolactine [78] et une augmentation du taux sérique de 17β-oestradiol, dystocie (rotation de 90° ou 180°, poulain plus large). Les problèmes semblent concerner principalement la fin de gestation, aucune incidence sur le premier tiers de la gestation n’a été démontrée [116].
Le mécanisme d’action retenu [27] serait l’activation par les alcaloïdes des récepteurs dopaminergiques [24]. De plus les alcaloïdes peuvent franchir la barrière placentaire, gagner le fœtus où ils pourront jouer leur rôle hépatotoxique [24].
Le traitement à envisager lors de suspicion de pâturage sur une parcelle infectée est bien entendu le retrait des animaux de cette même parcelle. Les taux de prolactine et de progestérone retrouvent des valeurs normales en deux à trois semaines [78].
Un traitement médical peut être associé en cas d’intoxication grave. L’effet des phénotiazines est controversé [27]. L’utilisation des antagonistes dopaminergiques semble

- 41 -
être le plus intéressant. La dompéridone est une molécule de choix car dépourvue chez le cheval d’effets secondaires neuroleptiques [27].
3.6.4. Un cas particulier d’influence de l’alimentation sur la reproduction de la jument : les avortements du Kentucky
Durant la saison de reproduction 2001, aux Etats-Unis, dans le Kentucky, plus de la
moitié des juments mises à la reproduction la saison précédente ont avorté. Face à l’ampleur du désastre économique dans ce berceau de l’élevage du pur-sang américain, de nombreuses investigations ont été menées [54, 79, 82, 96] concernant ce qui est appelé le « Mare Reproductive Loss Syndrome » (MRLS). Tour à tour, plusieurs hypothèses ont été écartées : les mycotoxines [82, 96], les phytoestrogènes [79].
Assez rapidement les études épidémiologiques ont mis en relation le MLRS avec une forte concentration géographique de cerisiers et de chenilles [54]. Les conditions climatiques particulières des mois précédents auraient particulièrement favorisé la recrudescence de ces chenilles. D’où une première hypothèse imaginant les chenilles comme vecteurs de substances cyanogènes présentes en grande quantité dans les cerisiers et responsables des avortements. Mais la reproduction expérimentale [54] des avortements n’ayant pu être faite par administration de mandelonitrile, précurseur libérant les radicaux cyanogènes dans le cerisier, cette hypothèse a été abandonnée.
L’hypothèse de travail la plus récente concerne directement les chenilles [99] (Malacosoma americanum). Ce serait l’ingestion des poils urticants, agissant comme vecteurs de bactéries, qui serait responsable des avortements observés. Les embols septiques auraient la capacité de migrer à travers les tissus du cheval. Le fœtus serait particulièrement sensible à ce matériel pathogène du fait d’une moindre résistance immunitaire et serait d’autant plus réceptif que le stade de gestation est avancé [99]. Des avortements ayant pu être reproduits expérimentalement 32 heures après l’exposition à ces chenilles, cette hypothèse semble la plus probable [99]. La lutte se porte donc maintenant sur la prophylaxie à l’égard de ces chenilles.
Un rationnement rigoureux de son cheptel reproducteur est donc nécessaire à la bonne
gestion de l’élevage. Les effets d’une sous-alimentation ne sont pas tous connus mais il semble qu’il n’y ait que des inconvénients à attendre. Les bénéfices éventuels d’un apport supérieur aux recommandations sont quant à eux à mettre en balance avec le coût engendré par ce supplément. Le suivi de l’état corporel des juments semble être un relativement bon indicateur du statut nutritionnel.

- 42 -
IV. Influence de la conduite d’élevage
Bien que pouvant être considéré comme un aspect environnemental de la jument, nous avons écarté de cette étude le côté médicamenteux de la conduite d’élevage. De nombreux traitements, hormonaux pour la plupart, permettent d’optimiser la gestion du cycle de la jument. On retrouve ici la progestérone souvent administrée (Janssen, REGUMATE ND, Issy-les-Moulineaux) en début de gestation pour soutenir un corps jaune supposé ou avéré déficient avant le relais placentaire, des prostaglandines, les corticoïdes ou bien la mélatonine (commercialisée uniquement chez les ovins : CEVA, MELOVINE ND, Libourne). La plupart des traitements administrés visent à induire un cycle chez les juments ne venant pas en chaleurs (progestérone ± œstrogènes puis prostaglandines) ou à maîtriser le cycle existant : induction de l’ovulation (gonadotrophines ou analogues), induction de la lutéolyse pour un retour en chaleur plus précoce (à visée reproductrice ou pour éviter qu’il ne tombe pendant une compétition). Il est aussi possible d’induire l’expulsion du contenu utérin (à l’aide d’ocytocine) que ce soit pour faciliter l’évacuation des enveloppes fœtales ou du nouveau-né lui-même en prenant garde si l’on décide d’induire le part à ce que la jument soit bien à terme, le risque de mettre au monde un prématuré étant assez élevé.
1. Détection et gestion des chaleurs
Une détection de qualité présente trois avantages : - déterminer avec précision le début de la période ovulatoire, c’est-à-dire la sortie de
la phase de transition, - déterminer le moment précis auquel pratiquer l’insémination artificielle ou bien le
moment de faire saillir la jument, - détecter une éventuelle gestation dans le cas où la jument ne revient pas en
chaleur dans les quinze jours suivant sa dernière ovulation. La présentation d’un souffleur à la jument (ou d’une jument au souffleur), peut
entraîner des comportements atypiques [70], surtout quand les stimulations sont très fréquentes. Ainsi des juments amenées de façon répétée à la barre et soumises à des stimulations sexuelles en dehors de la période d’œstrus, peuvent prendre l’habitude d’exprimer des comportements de rejet et les présenter même en période d’œstrus, rendant plus délicate l’interprétation de la barre. A l’inverse, d’autres juments vont devenir indifférentes aux attentions de l’étalon et ne montrer aucun signe d’œstrus ou de diœstrus. Enfin, des comportements d’œstrus sont possibles en l’absence d’activité folliculaire ce qui ne facilite pas le diagnostic [42]. Notons au passage que le souffleur aussi peut présenter des comportements « anormaux » : au fur et à mesure de sa carrière il devient de moins en moins réceptif aux juments en chaleurs et il peut passer à coté de juments faiblement expressives.
La prévention de ces réactions inhabituelles passe par le respect du caractère de la jument; les contacts avec le souffleur doivent être les plus doux possibles.
La détection précoce et suivie des chaleurs conditionne le taux de réussite de
l’insémination ou de la saillie. En effet, en dehors de la fertilité intrinsèque de la jument, une mauvaise gestion du moment de l’insémination ou de la saillie par rapport à l’ovulation peut compromettre les chances de succès [19]. Une insémination au-delà de 6-8 heures post-ovulation s’accompagne d’une augmentation significative du taux de mortalité embryonnaire [19].

- 43 -
Une bonne gestion des chaleurs nécessite donc un partenariat étroit entre le propriétaire de la jument, l’étalonnier, l’inséminateur et le vétérinaire.
2. Le logement Les bâtiments d’élevage doivent répondre aux mêmes exigences que les bâtiments
destinés aux chevaux en général en matière de matériaux de construction, de facilité d’entretien et de nettoyage, de stockage des aliments et du fourrage, d’élimination des déjections… Mais certains aménagements peuvent améliorer le confort de la jument [70] :
- la stalle est peu recommandée car elle ne permet pas à la jument de bouger. Le
poulinage en stalle est à proscrire. - Le boxe est le type de logement le plus fréquent. Sa surface minimale doit être de
3x3 m avec une hauteur de 3 à 3,40 m. Pour la jument suitée, la surface monte à 4x4 m. La porte doit mesurer 1,5 m de large.
- En stabulation libre où cohabitent plusieurs juments, chacune doit disposer de 10 m2 minimum, 15 m2 pour les juments suitées. Lorsque les juments sont trop nombreuses, les blessures peuvent devenir fréquentes, notamment pour les poulains. Ce type de logement est donc à réserver aux juments calmes, par petits lots.
- Le pré avec abri est réservé aux chevaux robustes. La surveillance des juments pour la détection des chaleurs ou la prévision du moment du poulinage est rendue d’autant plus difficile que la parcelle est grande. Comme dans le cas de la stabulation libre et comme précisé plus haut, la distribution de l’aliment peut s’avérer délicate et entraîner l’amaigrissement des juments soumises au profit des plus dominantes. Cependant la mise au pré des juments pendant la belle saison peut avoir un effet bénéfique stimulant de l’ovulation : la mise en pâture est associée à un exercice physique minimum suspecté responsable d’une augmentation de la sécrétion de prolactine [21] sensée influencer la croissance folliculaire et hâter la première ovulation de la saison. Enfin, ce type de logement ne permet pas une bonne détection des problèmes qui peuvent faire suite au part : coliques de la mère (la torsion du colon est favorisée par la vacuité qui suit la mise bas) ou du foal, poulain faible ou malajusté, déviations angulaires…
La mise au pré peut soulever un problème dans le cas de parcelles situées sous des lignes électriques haute tension. L’influence des champs magnétiques générés par ces lignes a fait l’objet de quelques études chez l’homme [10], le rat [1, 63, 90] ou la vache [2, 77, 89]. Les résultats ne sont pas univoques. Néanmoins une étude menée chez des vaches en Suède pâturant normalement (les autres expériences consistent à enfermer les animaux étudiés dans des cages magnétiques) est assez représentative de ce qui peut affecter les chevaux [2]. Ce travail conclut à l’absence d’effet sur le taux de réussite en insémination artificielle. En cas de doute sur des juments pâturant sur ce type de parcelles, on peut simplement les changer de pâture si l’on en a la possibilité ou bien procéder par élimination et écarter d’abord toutes les autres causes d’infertilité.

- 44 -
Il est intéressant de noter que le logement, selon l’option choisie, influe sur le comportement alimentaire de l’animal. Le cheval en pâture régulera lui-même son niveau d’ingestion fourragère tandis qu’au box une évaluation précise des besoins sera nécessaire afin d’éviter au cheval de se retrouver de longues périodes sans fourrage, ce qui favoriserait l’ennui du cheval et prédisposerait au développement de stéréotypies. A l’inverse, laisser du fourrage ad libitum peut entraîner un certain gaspillage qu’il conviendra de limiter. On note donc à quel point la conduite de l’élevage est une chose complexe et que tous les facteurs dépendent les uns des autres. La conduite d’élevage est un tout qui doit être mûrement réfléchi en fonction des objectifs et des contraintes logistiques (pâtures, bâtiments, main d’œuvre).
3. Le stress
Il va sans dire que l’espèce équine, de par sa nature de proie, est constamment sur le
qui-vive et donc très sensible au stress. Le mot stress regroupe un grand nombre d’entités qui sont autant de stimuli capables de perturber l’homéostasie de l’organisme [87] : stress alimentaire, thermique, psychologique, physique, métabolique…
Les stress thermique et alimentaire ont été traité précédemment. Sauf mention contraire nous traiterons ici du stress physique ou psychologique voire métabolique. Nous nous efforcerons de préciser les mécanismes biologiques du stress au sens large et de voir en quoi il peut influencer la reproduction.
3.1. Physiologie du stress – Interaction avec l’axe reproducteur
Le signe le plus couramment mis en évidence comme marqueur de stress, quelque soit
sa nature, est l’augmentation du taux de glucocorticoïdes circulants [74]. En fait, le stress entraîne la stimulation de l’axe hypothalamo-hypophysaire en favorisant la libération de CRH (Corticotrophin Releasing Factor) et d’AVP (Arginin Vasopressin). Le CRH entraîne la sécrétion d’ACTH par l’hypophyse et par voie de conséquence la libération de cortisol par les glandes surrénales. Ce pic de cortisol est atteint en deux heures [31]. Si l’animal est soumis à un stress d’une durée suffisante (environ 48 h), on peut même noter une hypertrophie et une hyperplasie des glandes cortico-surrénales [74].
Le cortisol serait donc le médiateur principal du stress et de nombreux sites d’actions ont pu être mis en évidence [3] en particulier au niveau de l’hypophyse et de l’hypothalamus le rendant en cela responsable de la baisse d’activité ovarienne due à la diminution de la sécrétion de LH.
Néanmoins tout n’est pas si simple puisque paradoxalement certains stress ponctuels qui devraient théoriquement stimuler la sécrétion de cortisol stimulent la sécrétion d’œstradiol et de LH [5, 6, 64]. En effet, il a été démontré chez l’homme et le rat [87] que l’exposition à un stress de courte durée correspond à une augmentation du taux plasmatique de LH. La réponse de l’organisme est en fait biphasique avec une phase initiale de stimulation et si le stress perdure ou atteint un seuil d’intensité suffisant, même un court instant, s’ensuit une phase d’inhibition [87].

- 45 -
Toutes ces modifications de l’homéostasie générale et des profils hormonaux peuvent influencer l’axe reproducteur à différents niveaux (fig. 12) :
- Le cerveau : il s’agit plus particulièrement de l’influence du stress sur la
sécrétion hypothalamique de GnRH. L’injection d’un inhibiteur du CRF (Corticotrophin Releasing Factor) dans le ventricule du cerveau d’un rat a mis en évidence [68] le rôle du système nerveux central en contrecarrant l’effet du stress sur la sécrétion de LH. Cette action centrale a, entre autres, pour intermédiaires des peptides opioïdes endogènes, des catécholamines (noradrénaline), le neuropeptide Y (NPY) et l’acide gamma aminobutyrique (GABA) [31].
- L’hypophyse : l’ACTH a été reconnue responsable de la diminution directement
au niveau de l’hypophyse de la sécrétion de LH suite à la stimulation par la GnRH [3, 37, 87].
- Les gonades : il est généralement accepté [87] que le stress puisse agir
directement sur les gonades bien qu’aucun mécanisme précis n’ait pu être élucidé. On attribue aux corticoïdes la capacité d’abaisser la sensibilité des gonades aux gonadotrophines. Il est bon de noter que les effets du stress sur la fonction gonadique adulte sont réversibles [87] : la fonction de reproduction retrouve ses droits dès que l’on ôte la source de stress. Il n’en est pas de même si le stress est appliqué pendant la phase précoce du développement fœtal, c’est ce que l’on appelle le syndrome du stress prénatal. Il est responsable, du moins chez le rat, de comportements sexuels anormaux chez le mâle et d’une moindre fertilité chez la femelle. De telles études n’ont pas été menées chez le cheval mais il convient de garder ces faits présents à l’esprit lors de la gestion de ses poulinières.
- Les autres organes : ils n’appartiennent pas au tractus génital mais leur
perturbation entraîne des changements métaboliques pouvant indirectement altérer l’axe hypothalamo-hypophysaire ; il s’agit entre autres du foie, de la thyroïde…
Bien que lors d’un stress il y ait systématiquement augmentation des taux de cortisol
et de catécholamines circulants, leur rôle dans la diminution de sécrétion de GnRH ou de LH, c’est-à-dire toute action directement sur le système nerveux central, semble limitée puisque ces effets perdurent après surrénalectomie. De plus le cortisol et les catécholamines circulantes ne passent pas la barrière hémato-céphalique [36].
Tout stress s’accompagne également [5] de la libération de cytokines à visée pro
inflammatoire, d’une diminution du taux d’acide ascorbique circulant et d’une augmentation des concentrations en hormone thyroïdienne [74] (plutôt chez les jeunes animaux après un stress psychologique aigu, et plutôt chez les animaux âgés lors de stress physique), et d’une diminution du taux de progestérone [5].
Les effets de l’administration de fortes doses de corticoïdes peuvent s’expliquer en
partie par le fait que la dexaméthasone et les glucocorticoïdes naturels peuvent se lier aux récepteurs de la progestérone [4]. Mais ceci reste très controversé [30, 31].
Mais le cortisol, marqueur principal du stress, n’aurait pas que des effets délétères. Les cytokines et le cortisol auraient un rôle physiologique au cours de la maturation

- 46 -
folliculaire et de l’ovulation. Certains auteurs [14] proposent une action régulatrice sur FSH et LH à ces périodes ; d’autres constatent une augmentation du cortisol plasmatique pendant le pro œstrus ou l’oestrus ou bien une augmentation du cortisol libre intra folliculaire en période pré ovulatoire et proposent pour cette hormone un rôle de régulateur de l’inflammation et réparateur après l’ovulation. On comprendrait mieux alors le rôle inhibiteur sur l’ovulation de fortes doses de corticoïdes exogènes qui entraveraient les phénomènes inflammatoires physiologiques.
Enfin les glucocorticoïdes sont connus pour avoir une action inhibitrice sur le système
immunitaire. Il a été démontré une diminution d’activité des lymphocytes T [74] via une suppression de la production de lymphokines. Il peut s’en suivre une plus grande sensibilité aux infections et en particulier aux endométrites pouvant être responsable d’une baisse notable de la fertilité.
Figure 12 : Les différents sites d’action du stress et leur relation avec la fonction de reproduction (d’après Armstrong [3]).

- 47 -
3.2. Influence du stress chez la jument
Les études menées sur les autres mammifères montrent que le stress ou l’administration de corticoïdes inhibent le fonctionnement du corps jaune, retardent voire bloquent le comportement d’œstrus (en diminuant la production de gonadotrophines et la sensibilité de l’ovaire à ces mêmes hormones d’où une moindre production d’œstrogènes [30]) et inhibent l’ovulation comme explicité plus haut.
Relativement peu d’études se sont penchées sur la question chez le cheval en confrontant la jument aux situations potentiellement stressantes ; la plupart de la bibliographie traite de l’effet des injections de corticoïdes.
3.2.1 Influence du stress sur la cyclicité de la jument
L’équipe de Baucus s’est intéressée à l’influence du transport des juments sur leur cycle [6]. En effet, l’étude de ce type de stress est particulièrement importante chez le cheval car les juments sont amenées à beaucoup se déplacer pour se faire saillir (essentiellement chez les purs-sangs où l’insémination est interdite) ou inséminer et enfin pour le suivi du cycle [6] ou de la gestation par le vétérinaire, contrairement aux bovins par exemple où ce sont l’inséminateur et le vétérinaire qui se déplacent.
Le stress causé par le transport a déjà été mis en cause chez la vache et la brebis : retard d’ovulation, suppression du comportement d’œstrus, modification de la durée des chaleurs [6]. Il a été également reconnu comme stressant chez le cheval car responsable d’une élévation du cortisol sérique et d’une diminution du taux de progestérone [5, 101].
L’étude de Baucus et al. n’a permis de montrer aucune influence du transport sur aucun des paramètres précités chez les autres espèces. L’auteur conclut que des conditions différentes d’expérimentation pourraient avoir des conséquences différentes, à savoir un temps de transport plus long ou la même expérience durant une phase du cycle différente. Néanmoins cette étude a l’avantage de reproduire des conditions couramment rencontrées : les transports pour saillie ou insémination artificielle dépassent rarement les 12 heures. Par contre la qualité de la conduite, fluide ou bien heurtée, joue probablement sur l’intensité du stress perçu par la jument mais est-ce suffisant pour observer une différence ?
Il faut également souligner que le cheval est une espèce habituée en général depuis son plus jeune âge à être transportée. Il serait dès lors intéressant de comparer avec un groupe de juments n’ayant jamais été transportées.
Une autre étude [14] a cherché déterminer si une altération passagère de la santé des
juments pouvait entraîner une modification de leur fonction de reproduction. Au final, aucun des paramètres étudiés (durée de la phase de croissance folliculaire, durée de la phase terminale de croissance folliculaire, nombre d’ovulations, taux plasmatique de LH et de stéroïdes) n’a été modifié au cours de l’expérience. Toutefois il ne faut pas oublier qu’en cas de forte altération de l’état de santé, la fonction de reproduction, considérée comme un luxe, est la première touchée, et donc une pathologie plus invalidante que celle mimée au cours de l’expérience pourrait avoir de plus graves conséquences.

- 48 -
3.2.2. Influence du stress sur la gestation chez la jument
Une autre étude toujours menée par Baucus et al. [5] a tenté d’étudier l’influence du transport sur la mortalité embryonnaire précoce entre les jours 16 et 22 ainsi que 32 et 38 de gestation. Une fois de plus, bien que l’expérience se soit avérée stressante pour les juments (augmentation du taux de cortisol circulant, diminution du taux sanguin d’acide ascorbique, diminution du taux sanguin de progestérone), le nombre de juments ayant subi une résorption embryonnaire n’était pas plus important pour les juments transportées que pour les juments témoins.
Une des limites de ce type d’expérience est qu’il n’a pas été possible de distinguer si les juments ayant avorté pendant le transport étaient les plus stressées du lot ou non.
Enfin il serait intéressant de reconduire cette étude vers 60 à 90 jours post ovulation, période vers laquelle le corps jaune secondaire laisse la place au placenta pour la sécrétion de progestérone. Une diminution physiologique du taux circulant à cette période majorée par le stress pourrait avoir de plus amples conséquences.
3.2.3. Influence du stress sur la mise bas chez la jument
Rossdale [91] attribue aux méthodes d’élevage (en particulier le confinement en box
de la jument, en présence d’observateurs pendant le poulinage) les déviations comportementales de la jument au moment de la mise bas : poulinage le jour, signes de première étape non suivis par la deuxième, mais aussi parturition debout et lever immédiatement après l’expulsion du fœtus. Rossdale et Short [92] précisent qu’en présence permanente d’observateurs en période prépartum, les juments poulinent aussi bien le jour que la nuit. De façon plus générale, des retards dans le déclenchement de la mise bas ont été attribués à la présence de l’homme et à son interférence, aussi bien qu’à la présence de groupes de juments spectatrices.
Ce fait laisse présumer un certain contrôle « psychique » exercé par la jument sur le déclenchement de la mise bas, une volonté de pouliner quand l’activité humaine fait une pause, la nuit.
En conclusion, beaucoup de situations ont été reconnues comme stressantes pour le
cheval (isolement en boxe, transport…) mais aucune n’a pu être considérée comme responsable d’une baisse significative des capacités reproductrices, contrairement aux administrations exogènes de corticoïdes. Il semble que les effets de ces stress soient sensiblement différents des effets induits par l’administration de corticoïdes à forte dose, qui eux compromettent gravement l’ovulation. Les cytokines, présentes à des concentrations plus élevées en cas de stress pourraient être en partie responsables de cette différence. L’administration de corticoïdes est bien loin de mimer avec précision ce qui se passe chez l’individu puisque nous l’avons vu, l’action centrale du stress (notamment sur la sécrétion de GnRH et de LH) n’est pas le fait du cortisol mais des opioïdes endogènes, de la noradrénaline, du NPY et du GABA.
Le stress a peut-être comme chez la femme [16] un rôle de potentialisation d’un autre trouble affectant la reproduction (potentialisation d’un défaut nutritionnel par un stress psychologique).
Les effets du stress sur la reproduction de la jument restent donc à préciser. Il convient néanmoins de tout mettre en œuvre pour éviter aux juments toute source de stress car dans tous les cas il n’y a que des inconvénients.

- 49 -
Conclusion
L’optimisation du potentiel reproducteur prend toute son importance chez la jument compte tenu des enjeux économiques présents dans la filière équine. La gestion de l’environnement joue à ce titre un rôle important.
Cette étude souligne le rôle prépondérant d’un plan nutritionnel raisonné, adapté au
stade physiologique de la jument. La tradition, très importante dans le milieu du cheval, n’a plus sa place, l’alimentation des reproducteurs et de leurs produits devant être le fruit d’une démarche raisonnée.
La photostimulation s’avère être le moyen le plus efficace pour avancer la saison de
reproduction des juments et ainsi satisfaire les exigences des éleveurs qui veulent obtenir un poulain, le plus précoce possible, par jument et par an. Elle permet ainsi de gagner environ deux mois sur les juments non traitées.
Mais l’obtention d’une bonne fertilité et d’une bonne fécondité passe avant tout pour
une gestion cohérente de son élevage en respectant au maximum les besoins physiologiques (alimentation, logement…) et psychologiques (limitation du stress, relations sociales inter et intraspécifiques…).

- 50 -
Annexe
Tableau récapitulatif des apports recommandés pour une jument de 500 kg à
chaque stade de son cycle (* : à moduler en fonction de l'état corporel de la jument et
de son stade de gestation/lactation; ** : Ca/P optimum = 1,5; ? : les connaissances
actuelles ne permettent pas de répondre) (d'après [76, 81]).
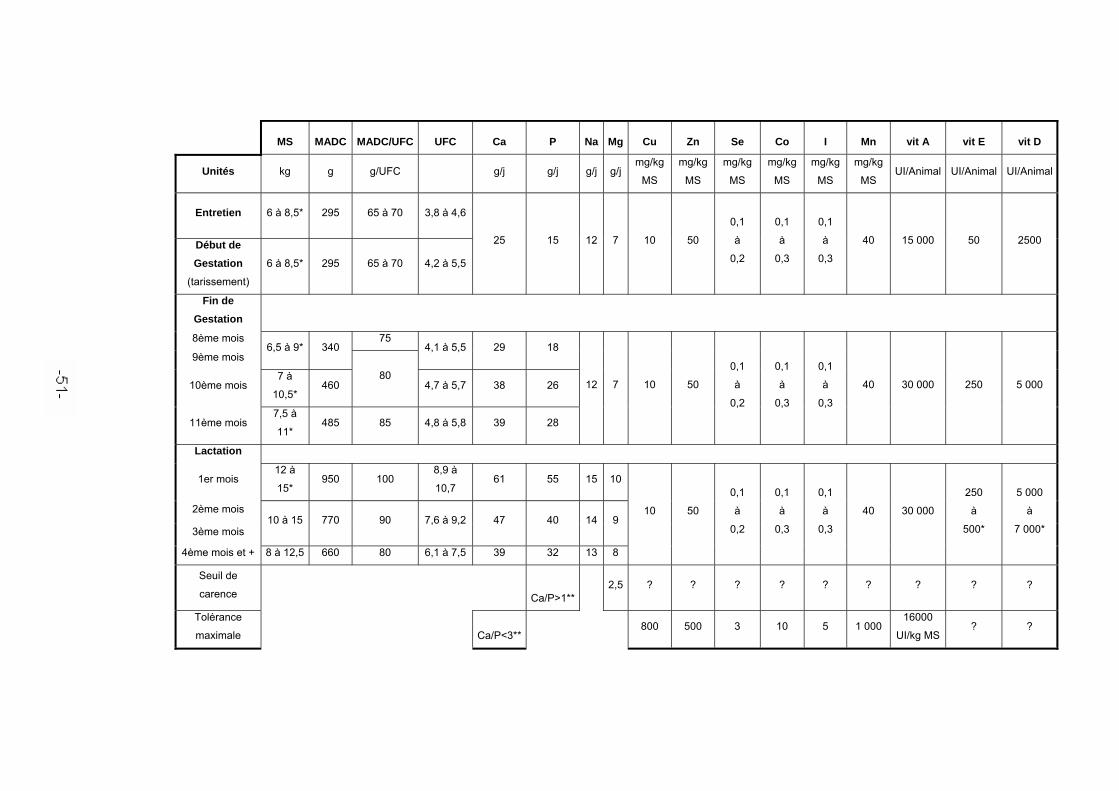
51
MS MADC MADC/UFC UFC Ca P Na Mg Cu Zn Se Co I Mn vit A vit E vit D
Unités kg g g/UFC g/j g/j g/j g/j mg/kg
MS
mg/kg
MS
mg/kg
MS
mg/kg
MS
mg/kg
MS
mg/kg
MS UI/Animal UI/Animal UI/Animal
Entretien 6 à 8,5* 295 65 à 70 3,8 à 4,6
Début de Gestation
(tarissement) 6 à 8,5* 295 65 à 70 4,2 à 5,5
25 15 12 7 10 50
0,1
à
0,2
0,1
à
0,3
0,1
à
0,3
40 15 000 50 2500
Fin de Gestation
8ème mois 75
9ème mois 6,5 à 9* 340 4,1 à 5,5 29 18
10ème mois 7 à
10,5* 460
80 4,7 à 5,7 38 26
11ème mois 7,5 à
11* 485 85 4,8 à 5,8 39 28
12 7 10 50
0,1
à
0,2
0,1
à
0,3
0,1
à
0,3
40 30 000 250 5 000
Lactation
1er mois 12 à
15* 950 100
8,9 à
10,7 61 55 15 10
2ème mois
3ème mois 10 à 15 770 90 7,6 à 9,2 47 40 14 9
4ème mois et + 8 à 12,5 660 80 6,1 à 7,5 39 32 13 8
10 50
0,1
à
0,2
0,1
à
0,3
0,1
à
0,3
40 30 000
250
à
500*
5 000
à
7 000*
Seuil de
carence Ca/P>1** 2,5 ? ? ? ? ? ? ? ? ?
Tolérance
maximale Ca/P<3** 800 500 3 10 5 1 000
16000
UI/kg MS? ?

52
Bibliographie 1. AL-AKHRAS MA., ELBETIEHA A., HASAN MK., AL-OMARI I., DARMANI H., ALBISS B. (2001) Effects of extremely low frequency magnetic field on fertility on adult male and female rats. Bioelectromagnetics, 22 (5) : 340-4 2. ALGERS B. and HENNICHS K. (1985) The effect of exposure to 400 kV transmission lines on the fertility of cows. A retrospective cohort study. Prev. Vet. Med., 3 : 351-361 3. ARMSTRONG D.T. (1986) Environmental Stress and Ovarian Function. Biol. Reprod., 34 : 29-39 4. ASA C.S. and GINTHER O.J. (1982) Glucocorticoid suppression of oestrus, follicles, LH and ovulation in the mare. J. Reprod. Fert., Suppl.32 : 247-251 5. BAUCUS K.L., RALSTON S.L., NOCKELS C.F., McKINNON A.O., SQUIRES E.L. (1990) Effect of transportation on early embryonic death in mares. J. Anim. Sci., 68 : 345-351 6. BAUCUS K.L., SQUIRES E.L., RALSTON S.L., McKINNON A.O., NETT T.M. (1990) Effect of transportation on the oestrus cycle of hormones in mares. J. Anim. Sci., 68 : 419-426 7. BAUDOIN N. et ARNAUD G. (1999) L’état corporel des chevaux. Les Haras Nationaux, Fiches Techniques, Alimentation. 8. BAUDOIN N. et ARNAUD G. (2003) Notation de l’état corporel des chevaux de selle et de sport, Guide pratique. Ed. Institut de l’élevage, 40p. 9. BENOIT P. (2004) Alimentation du cheval athlète – Actualités et perspectives. Cours de 3ème cycle professionnel “Sciences cliniques des équidés” Ecole Nationale Vétérinaire d’Alfort. 10. BLAASAAS KG., TYNES T., LIE RT. (2004) Risk of selected birth defects by maternal residence close to power lines during pregnancy. Occup. Environ. Med., 61 (2) : 174-176 11. BOHY A. (1986) Incidence des cycles lunaires sur la reproduction dans l’espèce bovine. Thèse Méd. Vét., Lyon, n°2, 120 p. 12. BOHY A. (1992) Cycles lunaires et vêlages. Courrier des lecteurs, Point Vétérinaire, 24 (147) : 7-8 13. BONNET M., DELAVAUD C., LAUD K., GOURDOU I., LEROUX C., DJIANE J., CHILLIARD Y. (2002) Mammary leptin synthesis, milk leptin and their putative physiological roles. Reprod. Nutr. Dev., 42 : 399-413 14. BRIANT C., HERVE V., GUILLAUME D. Cortisol, stress et reproduction chez la jument. Les Haras Nationaux, 26ème journée d’étude, 1er mars 2002, 101-110 15. BRIDGET D., BARBER W.P., WILLIAMS D.G. (1975) The effect of excess dietary iodine on pregnant mares and foals. Vet. Rec., 97, 93-95

53
16. BRINGER J., LEFEBVRE P., RENARD E.(1999) Nutrition et fonction ovarienne., Med. Sc., 15 : 197-203 17. BROCHART M. (1972) Oligo-éléments et fertilité., Ann. Nutr. Aliment., 26(3) : suppl.B 493-520 18. BRUNEAU G., VAISSE C., CARATY A., MONGET P. (1999) La leptine : une clé pour la reproduction. Med. Sc., 15 : 191-196 19. BRUYAS J.F. (2004) Mortalité embryonnaire chez la jument : facteurs de risque et conduite à tenir. 3ème cycle professionnel court dominante pathologie équine module reproduction Ecole Nationale Vétérinaire de Nantes, tome 3 : pathologie gynécologique, 91-103 20. BRUYAS J.F., BATTUT I., FIENI F., TAINTURIER D. (1997) Gestation gémellaire chez la jument : une cause majeure d’avortement. Le Point Vétérinaire, 28 (183) : 1261-1271 21. CARNEVALE E.M., HERMENET M.J., GINTHER O.J. (1997) Age and pasture effects on vernal transition in mares. Theriogenology, 47: 1009-1018 22. CAROFF G. (1995) Le poulinage et la période périnatale: particularités du déterminisme et aspects comportementaux. Thèse Méd. Vét., Alfort, n°110, 115 p. 23. CHODKOWSKI G.A.T. (1985) Alimentation et troubles de la fécondité de la jument. Thèse Méd. Vét., Toulouse, n°125, 124 p. 24. COPETTI M.V., SANTURIO J.M., BOECK A.A.P., SILVA R.B., BERGERMAIER L.A., LUBECK I., LEAL A.B.M., ALVES S.H., FERREIRO L. (2002) Agalactia in mares fed with grain contaminated with Claviceps purpurea., Mycopathologia, 154 (4) : 199-200 25. COUGOUILLE-GAUFFRETEAU B. (1983) Etude du comportement des chevaux : interactions sociales et sexuelles, conséquences pratiques pour la conduite de l’élevage. 9ème journée d’étude de la recherche chevaline, 43-62 26. CRISS TB., MARCUM JP. (1981) A lunar effect on fertility. Soc. Biol., 28 (1-2) : 75-80 27. CROSS D.L., REDMOND L.M., STRICKLAND J.R. (1995) Equine fescue toxicosis : signs and solutions, J. Anim. Sci., 73, 899-908 28. DAVID F. (2002) Contribution à l’élaboration d’un carnet de clinique : la période périnatale pour la jument et son poulain. Thèse Méd. Vét., Lyon, n°171, 308 p. 29. DAVIDSON K.E., POTTER G.D., GREENE L.W., EVANS J.W., McMULLAN W.C. (1991) Lactation and reproductive performance of mares fed added dietary fat during late gestation and early lactation, J. Eq. Vet. Sci., 11 (2), 111-115 30. DOBSON H., GHUMAN S., PRABHAKAR S., SMITH R. (2003) A conceptual model of the influence of stress on female reproduction. Reproduction, 125, 151-163 31. DOBSON H., SMITH R.F. (2000) What is stress, and how does it affect reproduction ? Anim. Reprod. Sci., 60-61, 743-752

54
32. DOREAU M., BOULOT S. (1989) Recent knowledge on mare milk production : a review., Livest. Prod. Sci., 22, 213-235 33. DOREAU M., BOULOT S., MARTIN-ROSSET W., DUBROEUCQ H. (1988) L’alimentation de la jument., Rev. Alim. Anim., 30-34 34. DOREAU M., BRUHAT J.P., MARTIN-ROSSET W. (1988) Effets du niveau des apports azotes chez la jument en début de lactation., Ann. Zootech., 37 (1), 21-30 35. DOREAU M., MARTIN-ROSSET W., BOULOT S. (1988) Energy requirements and the feeding of mares during lactation : a review, Liv. Prod. Sci., 20, 53-68 36. EVANS J.J., ROBINSON G., CATT J. (1989) Gonadotrophin-releasing activity of neurohypophysial hormones : I. Potential for modulation of pituitary hormone secretion. J. Endocrinol., 122, 96-106 37. EVANS M.J., LOY R.G., TAYLOR T.B., BARROWS S.P. (1982) Effects of exogenous steroids on serum FSH and LH, and on follicular development in cyclic mares. J. Reprod. Fert., Suppl. 32, 2005-212 38. FREEDMAN L.J., GARCIA M.C., GINTHER O.J. (1979) Influence of ovaries and photoperiod on reproductive function in the mare. J. Reprod. Fert., Suppl. 27, 79-86 39. GABILLOT F. (1993) Alimentation et pathologie chez la poulinière et le foal. Thèse Méd. Vét., Lyon, n°15, 91 p. 40. GARCIA M.C., FREEDMAN L.J., GINTHER O.J. (1979) Interaction of seasonal and ovarian factors in the regulation of LH and FSH secretion in the mare. J. Reprod. Fert., Suppl. 27, 103-111 41. GENTRY L.R., THOMPSON Jr D.L., GENTRY Jr G.T., DAVIS K.A., GODKE R.A. (2002) High versus low body condition in mares: interactions with responses to somatotropin, GnRH analog, and dexamethasone. J. Anim. Sci., 80 : 3277-3285 42. GENTRY L.R., THOMPSON Jr D.L., GENTRY Jr G.T., DAVIS K.A., GODKE R.A., CARTMILL J.A. (2002) The relationship between body condition, leptin, and reproductive and hormonal characteristics of mares during the seasonal anovulatory period. J. Anim. Sci., 80 : 2695-2703 43. GHIANDONI G., SECLI R., ROCCHI M.B., UGOLINI G. (1998) Does lunar position influence the time of delivery ? A statistical analysis. Eur. J. Obstet. Gynecol. Reprod. Biol., 77 (1) : 47-50 44. GINTHER O.J. (1993) Reproductive Biology of the Mare. Second Edition. Ed. Equiservices, 642p. 45. GINTHER O.J., WHITMORE H.L., SQUIRES E.L. (1972) Characteristics of oestrus, dioestrus, and ovulation in mares and effects of season and nursing. Am. J. Vet. Res., 33 (10), 1935-1939

55
46. GLADE M.J. (1991) Dietary yeast culture supplementation of mares during late gestation and early lactation – effects on milk production, milk composition, weight gain and linear growth of nursing foals, J. Eq. Vet. Sci., 11 (2), 89-95 47. GUERIN M.V., WANG X.J. (1994) Environmental temperature has an influence on timing of the first ovulation of seasonal estrus in the mare. Theriogenology, 42 : 1053-1060 48. GUILLAUME D., DAELS P-F., NAGY P. (2000) L’inactivité ovarienne saisonnière chez la jument, 1- Aspects physiologiques. Prat. Vet. Eq., 32 (128) : 375-382 49. GUILLAUME D., DUCHAMP G., BRUNEAU B., NAGY P. L’alimentation est le facteur prédominant de l’installation de l’inactivité ovarienne hivernale de la jument. Les Haras Nationaux, 27ème journée d’étude, 7 mars 2001, 3-11 50. GUILLAUME D., PALMER E. (1991) Effect of oral melatonin on the first ovulation after ovarian inactivity in mares under artificial photoperiod. J. Reprod. Fert., Suppl. 44, 249-257 51. GUILLAUME D., PALMER E. (1992) Lumière, mélatonine et reproduction chez la jument. Ann. Zootech., 41 : 263-269 52. GUILLON P., GUILLON D., LANSAC J., SOUTOUL JH., BERTRAND P., HORNECKER JP. (1986) Birth, fertility, rhythms and lunar cycle. A statistical study of 5927 births. J. Gynecol. Obstet. Biol. Reprod., 15 (3) : 265-271 53. HANZEN C. (2004) Les avortements chez les ruminants et les espèces equine et porcine. [en ligne], [http://www.fmv.ulg.ac.be/oga/dloads/Doc2Notes/Ch23.doc] (consulté le 30 août 2005) 54. HARKINS J.D., DIRIKOLU L., SEBASTIAN M., CRUTCHFIELD J., TROPPMANN A., BOYLES J., McDOWELL K., LONG W., HENNING J., FITZGERALD T., TOBIN T. (2002) Cherry trees, plant cyanogens, caterpillars, and mare reproductive loss syndrome : toxicological evaluation of a working hypothesis., First Workshop on Mare Reproductive Loss Syndrome, 68-74 55. HENNEKE D.R., KREIDER J.L. (1979) Effects of restrictive suckling on postpartum reproductive performance in mares. Am. J. Vet. Res., 40 (9) : 1281-1284 56. HENNEKE D.R., POTTER G.D., KREIDER J.L. (1984) Body condition during pregnancy and lactation and reproductive efficiency of mares. Theriogenology, 21 (6) : 897-909 57. HENNEKE D.R., POTTER G.D., KREIDER J.L., YEATES B.F. (1983) Relationship between condition score, physical measurements and body fat percentage in mares. Equine. Vet. J., 15 (4), 371-372 58. HODGE S.L., KREIDER J.L., POTTER G.D., HARMS P.G., FLEEGER J.L. (1982) Influence of photoperiod on the pregnant and postpartum mare. Am. J. Vet. Res., 43 (10) : 1752-1755 59. HOUSEKNECHT K.L., BAILE C.A., MATTERI R.L., SPURLOCK M.E. (1998) The biology of leptin : a review. J. Anim. Sci., 76 : 1405-1420 60. HOWELL C.E., ROLLINS W.C. (1951) Environmental sources of variation in the gestation length of the horse. J. Anim. Sci., 10 : 789-796

56
61. HOSSNER K.L. (1998) Cellular, molecular and physiological aspects of leptin : potential application in animal production. Can. J. Anim. Sci., 78 : 463-472 62. HULL M.L., REID R.A., EVANS J.J. et al. (1995) Preovulatory oxytocin administration promotes the onset of the luteinizing hormone surge in human females. Human. Reprod., 10, 2266-2269 63. HUUSKONEN H., SAASTAMOINEN V., KOMULAINEN H., LAITINEN J., JUUTILAINEN. (2001) Effects of low-frequency magnetic fields on implantation in rats. Reprod. Toxicol., 15 (1) : 49-59 64. IRVINE C.H.G., ALEXANDER S.L. (1997) The role of environmental factors in reproduction in the mare. Ippologia, 8 (1) : 55-73 65. JARRIGE R., MARTIN-ROSSET W. (1984) Le cheval : reproduction, alimentation, sélection, exploitation., ed. INRA, 689p. 66. JUHASZ J., NAGY P., HUSZENICZA G., SZIGETI G., REICZIGEL J., KULCSAR M. (1997) Long term exposure to T-2 fusarium mycotoxin fails to alter luteal function, follicular activity and embryo recovery in mares., Equine Vet. J. Suppl., 25, 17-21 67. KOLLERSTROM N., POWER C. (2000) The influence of the lunar cycle on fertility on two Thoroughbred studfarms. Equine Vet. J., 32 (1) : 75-77 68. KUBIAK J.R., CRAWFORD B.H., SQUIRES E.L., WRIGLEY R.H., WARD G.M. (1986) The influence of energy intake and percentage of body fat on the reproductive performance of non-pregnant mares. Theriogenology, 27 : 587-598 69. KUBIAK J.R., EVANS J.W., POTTER G.D., HARMS P.G., JENKINS W.L. (1991) Milk yield and composition in the multiparous mare fed to obesity. Eq. Vet. Sci., 11 (3) : 158-162 70. LEMOINE M. (2003) Facteurs influençant les résultats de fécondité en élevage équin. Etude dans le département de l’Ain, de la Loire et du Rhône. Thèse Méd. Vét., Lyon, n°18, 145 p. 71. Les Haras Nationaux – Direction du Développement – Observatoire économique et social du cheval (2002) Le cheval en France en 2002, 7p. 72. Le Site Cheval. Etat Corporel. [en ligne]. [http://www.le-site-cheval.com] (consulté le 7 mars 2005) 73. LEWIS L.D. (1995) Feeding and care of the horse (second edition), Ed. Williams & Wilkins, 446 p. 74. MAL M.E., FRIEND T.H., LAY D.C., VOGELSANG S.G., JENKINS M.S and O.C. (1991) Physiological response of mares to short term confinement and social isolation. Eq. Vet. Sci., 11 (2) : 96-102 75. MALINOWSKI K., JOHNSON A.L., SCANES C.G. (1985) Effects of interrupted photoperiods on the induction of ovulation in anœstrous mares. J. Anim. Sci., 61 (4), 951-955

57
76. MARTIN-ROSSET W. (1990) L’alimentation des chevaux., ed. INRA, 232p. 77. MARTIN F.B., BENDER A., STEUERNAGEL G., ROBINSON R.A., REVSBECH R., SORENSEN D.K., WILLIAMSON N., WILLIAMS A. (1986) Epidemiologic study of Holstein dairy cow performance and reproduction near a high-voltage direct-current power-line. J. Toxicol. Environ. Health., 19 (3) : 303-324 78. McCANN J.S., CAUDLE A.B., THOMPSON F.N., STUEDEMANN J.A., HEUSNER G.L., THOMPSON D.L. (1992) Influence of endophyte-infected tall fescue on serum prolactin and progesterone in gravid mares., J. Anim. Sci., 70, 217-223 79. McDOWELL K., ALLMAN R., HENNING J. (2002) Phytoestrogens and estrogenic activity in white clover samples from no-loss and high-loss fields during mare reproductive loss syndrome., First Workshop on Mare Reproductive Loss Syndrome, 63-65 80. MERKT H., JUNGNICKEL S., KLUG E. (1982) Reduction of early twin pregnancy to single pregnancy in the mare by dietetic means. J. Reprod. Fert., Suppl. 32, 451-452 81. National Research Council (1989) Nutrient Requirements of Horses (Fifth revised edition) Ed. Nat. Acad. Press., 112 p. 82. NEWMAN K. (2002) Review of mycotoxins as a possible cause of mare reproductive loss syndrome., First Workshop on Mare Reproductive Loss Syndrome, 66-68 83. OSBORNE M. (2001) Diagnosis and management of twin pregnancy in the mare. Irish Vet. J., Vol. 54 (10), 520-522 84. Ott. E.A., ASQUITH R.L. (1994) Trace mineral supplementation of broodmares., J. Eq. Vet. Sci., 14 (2), 93-100 85. PERITI E., BIAGIOTTI R. (1994) Lunar phases and incidence of spontaneous deliveries. Our experience. Minerva. Ginecol., 46 (7-8) : 429-433 86. PUTNAM M.R., BRANSBY D.I., SCHUMACHER J., BOOSINGER T.R., BUSH L., SHELBY R.A., VAUGHAN J.T., BALL D., BRENDEMUEHL J.P. (1991) Effects of the fungal endophyte Acremonium coenophialum in fescue on pregnant mares and foal viability., Am. J. Vet. Res., 52 (12), 2071-2074 87. RIVIER C., RIVEST S. (1991) Effect of stress on the activity of the hypothalamic-pituitary-gonadal axis : peripheral and central mechanisms. Biol. Reprod., 45 : 523-532
88. ROBINSON G, EVANS J.J., CATT K.J. (1992) Oxytocin stimulates LH production by the anterior pituitary gland of the rat. J. Endocrinol., 132 : 277-283 89. RODRIGUEZ M., PETITCLERC D., BURCHARD J.F., NGUYEN D.H., BLOCK E. (2004) Blood melatonin and prolactin concentrations in dairy cows exposed to 60 Hz electric and magnetic fields during 8 h photoperiods. Bioelectromagnetics, 25 (7) : 508-515

58
90. ROMMEREIM D.N., ROMMEREIN R.L., SIKOV M.R., BUSCHBOM R.L., ANDERSON L.E. (1990) Reproduction, growth, and development of rats during chronic exposure to multiple fields strengths of 60-Hz electric fields. Fundam. Appl. Toxicol., 14 (3) : 608-621 91. ROSSDALE P.D. (1968) Abnormal perinatal behaviour in the thoroughbred horse. Br. Vet. J., 124 : 540-552 92. ROSSDALE P.D., SHORT R.V. (1967) The time of foaling of thoroughbred mares. J. Reprod. Fert.,13 : 341-343 93. SCRABA S.T., GINTHER O.J. (1985) Effects of lighting programs on onset of the ovulatory season in mares. Theriogenology, 24 (6), 667-679 94. SHARP D.C., GINTHER O.J. (1975) Stimulation of follicular activity and oestrus behaviour in anoestrus mares with light and temperature. J. Anim. Sci., 41 (5) : 1368-1372 95. SCHRYVER H.F., OFTEDAL O.T., WILLIAMS J., SODERHOLM L.V., HINTZ H.F. (1986) Lactation in the horse : the mineral composition of mare milk., J. Nutr., 116 (11), 2142-2147 96. SCHULTZ C.L., BUSH L.P. (2002) The potential role of ergot alkaloids in mare reproductive loss syndrome., First Workshop on Mare Reproductive Loss Syndrome, 60-63 97. SPICER L.J. (2001) Leptin : a possible metabolic signal affecting reproduction. Dom. Anim. Endoc., 21 : 251-270
98. STROLEGO F., GIGLI C., BUGALHO A. (1991) The influence of lunar phases on the frequency of deliveries. Minerva. Ginecol., 43 (7-8) : 359-363 99. TOBIN T. (2004) The mare reproductive loss syndrome and the eastern tent caterpillar II : a toxicokinetic/clinical evaluation and a proposed pathogenesis : septic penetrating setae., Intern. J. Appl. Res. Vet. Med., 2 (2), 142-158 100. VAGNER I. (2003) Conduite diagnostique et thérapeutique lors de gémellité chez la jument: étude de terrain. Thèse Méd. Vét, Nantes, n°126, 143 p. 101. VAN NIEKERK C.H., MORGENTHAL J.C. (1982) Fetal loss and the effect of stress on plasma progestagen levels in pregnant Thoroughbred mares. J. Reprod. Fert., Suppl. 32, 453-457 102. VAN NIEKERK C.H., VAN HEERDEN J.S. (1972) Nutrition and ovarian activity of mares early in the breeding season. J. S. Afr. Vet. Ass., 43 (4), 351-360 103. VAN NIEKERK F.E., VAN NIEKERK C.H. (1997) The effect of dietary protein on reproduction in the mare. I. The composition and evaluation of the digestibility of dietary protein from different sources. J. S. Afr. Vet. Ass., 68 (3) : 78-80 104. VAN NIEKERK F.E., VAN NIEKERK C.H. (1997) The effect of dietary protein on reproduction in the mare. II. Growth of foals, body mass of mares and serum protein concentration of mares during the anovulatory, transitional and pregnant periods. J. S. Afr. Vet. Ass., 68 (3) : 81-85

59
105. VAN NIEKERK F.E., VAN NIEKERK C.H. (1997) The effect of dietary protein on reproduction in the mare. III. Ovarian and uterine changes during the anovulatory, transitional and ovulatory periods in the non-pregnant mare. J. S. Afr. Vet. Ass., 68 (3) : 86-92 106. VAN NIEKERK F.E., VAN NIEKERK C.H. (1997) The effect of dietary protein on reproduction in the mare. IV. Serum progestagen, FSH, LH and melatonin concentrations during the anovulatory, transitional and ovulatory periods in the non-pregnant mare. J. S. Afr. Vet. Ass., 68 (4) : 114-120
107. VAN NIEKERK F.E., VAN NIEKERK C.H. (1998) The effect of dietary protein on reproduction in the mare. V. Endocrine changes and conception during the early post partum period. J. S. Afr. Vet. Ass., 69 (3) : 81-88 108. VAN NIEKERK F.E., VAN NIEKERK C.H. (1998) The effect of dietary protein on reproduction in the mare. VI. Serum progestagen concentrations during pregnancy. J. S. Afr. Vet. Ass., 69 (4) : 143-149 109. VAN NIEKERK F.E., VAN NIEKERK C.H. (1998) The effect of dietary protein on reproduction in the mare. VII. Embryonic development, early embryonic death, foetal losses and their relationship with serum progestagen. J. S. Afr. Vet. Ass., 69 (4) : 150-155 110. VIOLLET G. (1986) La Gémellité chez la jument. Thèse Méd. Vét., Lyon, n°27, 66 p. 111. VOSS J.L., PICKETT B.W. (1974) Effect of nutritional supplement on pregnancy rate in nonlactating mares., J. Am. Vet. Med. Assoc., 165 (8), 702-703 112. WOODLEY S.L., BURNS P.J., DOUGLAS R.H., OXENDER W.D. (1979) Prolonged interovulatory interval after oestradiol treatment in mares., J. Reprod. Fertil. Suppl., 27, 205-209 113. WOLTER R. (1975) Alimentation et fertilité chez la jument. Prat. Vet. Eq., 7 (4) : 161-168 114. WOLTER R. (1992) Le rationnement pratique des juments et des poulains. Prat. Vet. Eq., 24 (1) : 39-52 115. WOLTER R. (1999) Alimentation du Cheval. 2ème édition. Ed. France Agricole, 478 p. 116. YOUNGBLOOD R.C., FILIPOV N.M., RUDE B.J., CHRISTIANSEN D.L., HOPPER R.M., GERARD P.D., HILLS N.S., FITZGERALD B.P., RYAN P.L. (2004) Effects of short-term early gestational exposure to endophyte-infected tall fescue diets on plasma 3?4-dihydroxyphenyl acetic and fetal development in mares., J. Anim. Sci., 82 (10), 2919-2929





























