Embed Size (px)
Citation preview

INFLUENCE DU RÉGIME DES FEUX SUR LES MODIFICATIONS À COURT TERME ET LA STABILITÉ À LONG TERME
DE LA FLORE D'UNE GARRIGUE DE QUERCUS COCCIFERA
Louis TRABAUD
Centre d'Ecologie Fonctionnelle et Evolutive. CNRS. Montpellier*
Dans la région méditerranéenne, le feu est un facteur de perturbation qui a façonné les communautés et les paysages. La connaissance de son action sur les végétaux méditerranéens permettra de comprendre le passé et la mise en place de la végétation et de la flore, mais aussi de connaître leur devenir (maintien ou disparition) selon que les incendies seront dans l'avenir plus ou moins nombreux : certaines espèces et certaines communautés risquent de disparaître, d'autres d'apparaître, et des changements surviendront selon que les incendies seront trop ou pas assez fréquents.
Les travaux sur les effets du feu, et plus particulièrement son influence sur les écosystèmes de type méditerranéen, ont été très nombreux en Californie, en Australie et en Afrique du Sud. Dans le Bassin de la Méditerranée, de telles recherches n'ont débuté que depuis une décennie environ: en Espagne (Casal et al. , 1986, 1990; Tarrega & Luis, 1987), en Italie (De Lillis & Testi, 1990;Mazzoleni & Pizzolongo, 1990), et en Grèce (Papanastasis, 1977 ; Arianoutsou & Margaris, 1981 ; Arianoutsou, 1984; Thanas et al. , 1989). En Israël, Naveh (1974,1975) a seulement donné un aperçu général des effets du feu sur la végétation, mais ne décrit pas les processus de réponse des végétaux.
En France, la dynamique de la végétation après incendie a été surtout étudiée dans les garrigues calcaires du Bas-Languedoc (Trabaud, 1970, 1977, 1983; Trabaud & Lepart, 1980; Trabaud et al. , l985a et b) dans huit types de communautés dominantes et représentatives de la région. Après incendie, la végétation retourne assez rapidement (une dizaine d'années) à son état initial . Cette tendance vers un retour à l'état d'origine a été aussi constatée en Provence calcaire (Abbas et al. , 1984; Barbéro et al. , 1987) ainsi que dans les Pyrénées orientales siliceuses (Pradon et al. , 1984).
Toutefois, aucune des études citées ne considère l'influence de mises à feu répétées pendant une longue période. Lors d'une analyse antérieure, portant sur les premiers résultats de brûlages expérimentaux réalisés sur une garrigue de Quercus coccifera (Chêne kermès), Trabaud (1974, 1984) et Trabaud & Lepart (1981) avaient présenté les changements qui apparaissaient au cours des six premières années. Au point de vue qualitatif, la flore gardait une bonne stabilité.
* Adresse : Centre Louis Emberger, Route de Mende, B.P. 505 1 , F-34033 Montpellier Cedex.
Rev. Eco/. ( Terre Vie) , vol. 47, 1 992.
- 209 -

Sur le plan quantitatif, par contre, la répétition des mises à feu produisait des changements structuraux : plus leur nombre était élevé moins la stratification du système était complexe. En outre, il y avait une diminution importante des végétaux ligneux et un accroissement des végétaux herbacés, en particulier avec les mises à feu d'automne.
Les résultats présentés dans cet article concernent les modifications de la composition ftoristique obtenues au cours d'une expérience qui a duré une vingtaine d'années sur une garrigue de Q. coccifera soumise à différents régimes de mises à feu. Le but était de savoir si la répétition des brûlages entraînait des changements (et lesquels ?) parmi les espèces vivant dans la communauté.
DISPOSITIF EXPÉRIMENTAL ET MÉTHODES UTILISÉES
Pour analyser finement et comprendre parfaitement l'action du feu sur les végétaux et la végétation, il est nécessaire d'utiliser des brûlages contrôlés. En effet, seule une expérimentation entreprise sur des communautés végétales bien étudiées avant le feu permet de bien connaître le stade initial et de le comparer aux résultats obtenus par la suite.
Les objectifs de l'expérience étaient d'analyser : - le développement de la végétation par rapport à l'état initial, afin de
déterminer la résistance au feu des espèces présentes ; - l'impact sur la végétation, et les espèces végétales, de mises à feu répétées
selon diverses fréquences ; - les effets sur la végétation, et les espèces, de la date (saison) des mises à
feu, en vue de déterminer si les conditions saisonnières qui se traduisent par des variations du comportement phénologique et physiologique des plantes, sont en relation avec les effets du feu, et si elles modifient les réponses des espèces.
L'expérience a duré 19 ans : de 1969 à 1987 inclus .
SITUA TION DE LA ZONE ÉTUDIÉE
L'expérimentation a été faite sur une colline au lieu-dit Puech-du-Mas-duJuge (commune de Saint Gély-du-Fesc) à 10 km au nord de Montpellier (43° 41' N, 3° 48' E). La communauté végétale choisie est une garrigue de Quercus coccifera (Cocciferetum Br-BI. 1924, sous-association brachypodietosum Br-BI. 1935 (Braun-Blanquet et al. , 1952). Les parcelles sont situées sur le versant sud-ouest de la colline, constituée par du calcaire lacustre de l 'Eocène moyen.
Le climat est méditerranéen humide selon la classification d'Emberger ( 1942, 1971). Pour la période de dix-neuf ans ( 1969-1987), la moyenne annuelle des températures fut de 13,2 oc ; la moyenne des minimales du mois le plus froid (janvier) fut égale à 2,4 oc ; tandis que celle des maximales du mois le plus chaud (juillet) atteignit 28,0 oc. La pluviosité moyenne, pour cette même période, fut de 919 mm.
Ces terrains ont été cultivés il y a longtemps, puis furent abandonnés vers le milieu du XIXe siècle et devinrent des terrains de parcours pour les troupeauxd'ovins. Ils ont alors évolué vers une garrigue à cause des incendies répétés, allumés délibérément par les bergers pour « régénérer » les pâturages ou ouvrir la végétation afin de permettre le passage des animaux. Le dernier feu connu eut lieu au mois d'août 1951.
- 210 -

Depuis 1 969, année au cours de laquelle débuta l 'expérimentation, la zone d'étude fut complètement protégée de toute action anthropique et de tout pâturage. Au début de l'expérience, la garrigue apparaissait sous la forme d'une broussaille dense de Chêne kermès, de 1 à 1 , 50 rn de hauteur, recouvrant 80 â 1 00 % de la surface du sol. La strate des buissons était constituée par Q. coccifera (80 % de recouvrement environ), Dorycnium pentaphyl/um ( 10 %), Genista scorpius (5- 1 0 %), Teucrium chamaedrys ; la strate herbacée était dominée presque exclusivement par Brachypodium retusum ( 10-20 %), accompagné de Rubiaperegrina, Carex hallerana, C. humilis (la nomenclature est celle de Tutin et al. , 1 964- 1 980) .
PRO TOCOLE DES MISES À FEU
Les dates de mise à feu ont été choisies en tenant compte de certains critères phénologiques de l'espèce dominante, le Chêne kermès.
La première série de mises à feu eût lieu après le démarrage végétatif printanier de Q. coccifera ; les pousses de l'année, ainsi que les nouvelles feuilles, étaient alors largement développées et les fleurs étaient apparues. Le chêne se trouvait donc dans un état d'activité photosynthétique maximal. Les brûlages furent effectués fin mai-début juin selon les conditions météorologiques. Ils sont appelés « feux de printemps » (P) dans cet article.
Une seconde série de mises à feu fut effectuée au début de l'automne, après l'aoûtement des rameaux de l'année ; la végétation, au moins en apparence, semblait être au repos . Ces brûlages ont toujours débuté dans la première quinzaine de septembre, et ils sont appelés ici « feux d'automne » (A).
La fréquence des mises à feu a tenu partiellement compte du « rythme » utilisé par les bergers (tous les cinq à huit ans selon l'embroussaillement de la végétation) : un groupe de parcelles a été brûlé tous les six ans (6), un autre groupe tous les trois ans (3) et un dernier groupe tous les deux ans (2) . Ce plan d'expérience permet donc de comparer les effets de deux facteurs : la saisonnalité et la périodicité des feux. Six traitements différents sont alors obtenus en combinant les saisons et les fréquences.
Des parcelles « témoins » (T), non brûlées, furent intégrées au dispositif ; elles ont servi de référence pour comparer les changements survenus dans la végétation.
Cinq répétitions furent prévues pour l'ensemble des facteurs combinés, ce qui donne un total de 35 parcelles élémentaires de 50 m2 (5 x 1 0 rn) chacune. Celles-ciétaient séparées par des allées de 2 rn de large.
RÉCOLTE ET ANAL YSE DES DONNÉES
La liste de toutes les espèces présentes sur chaque parcelle a été dressée régulièrement tous les ans au mois de mai, avant les mises à feu de printemps, pendant les 1 9 ans qu'a duré l'expérimentation. Une analyse de la variance a été réalisée pour juger l'effet de la saison et de la fréquence des mises à feu sur le nombre de présences des espèces observées au cours du temps.
Pour quantifier et caractériser les changements, entraînés par la saisonnalité et la périodicité des brûlages, apparaissant dans le comportement des végétaux, deux indices ont été calculés, pour chaque espèce et pour chaque traitement. Le premier, appelé Indice de présence (lp), tient compte seulement des observations
- 21 1

effectuées au début et à la fin de la période d'expérimentation ; il caractérise en quelque sorte la« persistance dynamique » des espèces, puisqu'il permet d'apprécier si une espèce est encore présente en fin d'expérience, si son degré de présence s'est accru ou à diminué, ou si elle est nouvellement apparue.
lp = I Pf/ I Po1 1
où x est le nombre de répétitions par traitement où l'espèce est observée, Pf, la présence observée à la fin de la période d'expérimentation, et Po, la présence observée au début (origine) de la période d'expérimentation.
Le second, appelé Indice de changement (le), tient compte de toutes lesobservations réalisées annuellement au cours de la totalité de la période d'observation. Il caractérise les changements (fluctuations) ou le degré de permanence des espèces au cours du temps ; il est calculé par deux équations différentes selon que les espèces étaient présentes ou non au début de l'expérience. Pour les espèces présentes à l'origine l'expression est :
le = L NPtl L Nt1 1
où NPt est le nombre de précences totales observées au cours de l'expérimentation, et Nt, le nombre total d'années d'observation.
le est égal à L NPtl L Nt - 1, pour les espèces apparaissant au cours de 1 1
l'expérimentation, Nt - 1 étant le nombre total d'années d'observation moins une, puisque cette formule ne s'applique qu'à des espèces ayant apparu au cours de l'expérimentation, donc absentes à l'origine. Toutefois, ces espèces peuvent, ou auraient pu, apparaître dès la deuxième année.
Quel que soit le moment d'apparition, plus les valeurs du deuxième indice sont élevées, plus les espèces sont restées présentes longtemps, ou ont été présentes plusieurs fois au cours du temps.
L'utilisation combinée de ces deux indices a permis de classer les espèces en plusieurs catégories selon leurs réponses : envahissantes, favorisées, résistantes, persistantes, sensibles, disparaissantes, etc . . . Seules les espèces observées plus de deux fois ont été considérées pour effectuer les calculs.
RÉSULTATS
Deux aspects permettent de caractériser l'évolution de la compositiOn floristique de la garrigue de Chêne kermès brûlée selon les différents traitements. Ce sont (a) l'évolution de la richesse floristique, et (b) le comportement des espèces par rapport aux mises à feu. En outre, les moyens de dissémination et de survie des végétaux apportent des indications quant à leurs capacités à résister ou à occuper l'espace.
VARIA TIONS DE LA COMPOSITION FLORISTIQ UE
Le nombre d'espèces présentes dans les traitements n'est nullement affecté par la « fréquence des mises à feu » pour l'ensemble des observations effectuées
- 2 1 2 -
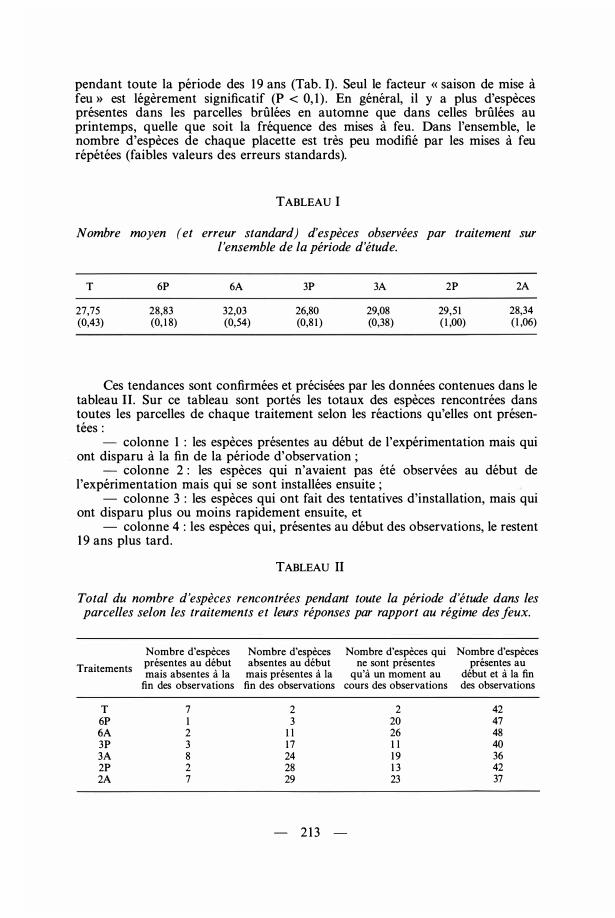
pendant toute la période des 1 9 ans (Tab. 1). Seul le facteur « saison de mise àfeu » est légèrement significatif (P < 0, 1 ) . En général, il y a plus d'espèces présentes dans les parcelles brûlées en automne que dans celles brûlées au printemps, quelle que soit la fréquence des mises à feu. Dans l'ensemble, le nombre d'espèces de chaque placette est très peu modifié par les mises à feu répétées (faibles valeurs des erreurs standards).
TABLEAU 1 Nombre moyen (et erreur standard) d'espèces observées par traitement sur
l 'ensemble de la période d'étude.
T
27,75 (0,43)
6P
28,83 (0, 1 8)
6A
32,03 (0,54)
3P
26,80 (0,8 1 )
3A
29,08 (0,38)
2P
29,5 1 ( 1 ,00)
2A
28,34 ( 1 ,06)
Ces tendances sont confirmées et précisées par les données contenues dans le tableau Il. Sur ce tableau sont portés les totaux des espèces rencontrées dans toutes les parcelles de chaque traitement selon les réactions qu'elles ont présentées :
- colonne 1 : les espèces présentes au début de l'expérimentation mais qui ont disparu à la fin de la période d'observation ;
- colonne 2 : les espèces qui n'avaient pas été observées au début de l'expérimentation mais qui se sont installées ensuite ;
- colonne 3 : les espèces qui ont fait des tentatives d'installation, mais qui ont disparu plus ou moins rapidement ensuite, et
- colonne 4 : les espèces qui, présentes au début des observations, le restent 1 9 ans plus tard.
TABLEAU Il
Total du nombre d'espèces rencontrées pendant toute la période d'étude dans les parcelles selon les traitements et leurs réponses par rapport au régime des feux.
Nombre d'espèces Nombre d'espèces Nombre d'espèces qui Nombre d'espèces
Traitements présentes au début absentes au début ne sont présentes présentes au mais absentes à la mais présentes à la qu'à un moment au début et à la fin
fin des observations fin des observations cours des observations des observations
T 7 2 2 42 6P 1 3 20 47 6A 2 I l 26 48 3P 3 17 I l 40 3A 8 24 1 9 36 2P 2 28 1 3 42 2A 7 29 23 37
- 2 1 3 -

Ce tableau permet les constatations suivantes : 1 . Le nombre d'espèces présentes au début de l'expérimentation et qui ont
disparu est plus important dans la vétégation non brûlée que dans celle brûlée tous les six ou trois ans ; cela peut être dû à la fermeture de la végétation au cours des ans. Ce qui est remarquable c'est le petit nombre d'espèces qui se sont installées, ou qui ont fait des tentatives d'installation, mais qui n'ont pu persister dans la végétation non brûlée.
2. Pour les trois premières catégories d'espèces (les trois premières colonnes),la saison des mises à feu exerce une influence sur le nombre d'espèces observées, qui est significativement supérieur (P < 0,05) pour les brûlages d'automne. Ainsi, les mises à feu d'automne provoquent un accroissement des espèces qui apparaissent, qui disparaissent ou qui tentent de s'installer. Au contraire, pour la dernière catégorie (quatrième colonne, celle des espèces « persistantes), ce sont les feux de printemps qui entraînent un nombre légèrement supérieur (P < 0, 1) d'espèces permanentes, effet auquel s'ajoute celui de la fréquence qui entraîne une diminution de ce type d'espèces avec les mises à feu tous les 2 ou 3 ans .
Toutefois, malgré ces variations, la composition floristique de la garrigue garde une bonne stabilité puisque l'ensemble des espèces persistantes constitue 53, 1 % de la totalité des espèces rencontrées tout au long de la période d'étude.
COMPORTEMENT DES ESPÈCES
Si 56,9 % des espèces recensées dans les parcelles témoins persistent du début à la fin de l'expérimentation, l'absence de feu déclenche toutefois quelques modifications. Ainsi, le degré de présence de Phillyrea angustifo/ia, P. /atifo/ia, et Rhamnus a/aternus augmente fortement (Tab. III), ainsi, bien qu'à un degré moindre, celui de Clematis j/ammula et Lonicera imp/exa. Ce sont des espèces de port arborescent comme les trois premières ou lianiformes comme les deux dernières, plus fréquemment rencontrées dans les taillis ou bois âgés. Plus surprenant est l'accroissement du degré de présence de Thymus vulgaris et Ononis minutissima, plantes plutôt considérées comme héliophiles. Il en est de même pour Euphorbia nicaeensis et Festuca spadicea qui s'installent dans cette végétation non brûlée. Festuca spadicea est une plante extrêmement envahissante dans les parcelles expérimentales et elle accroît fortement son recouvrement dans les parcelles incendiées.
D'autre part, le degré de présence de certaines espèces diminue quelque peu, c'est le cas pour Asphode/us racemosus, Brachypodium phoenicoides, Carex humilis, Euphorbia characias, Cistus salvifolius. Ce sont des espèces vivant surtout dans des formations de garrigue, ayant une taille assez importante, et qui résistent à la fermeture de la végétation. Au contraire, Fumana ericoides, Galium corrudaefolium, Hippocrepis comosa, Sanguisorba minor, Sedum sediforme, Silene italica, et Stachys officina/is, qui sont des plantes de lumière et ont une taille plus petite que les précédentes, disparaissent complètement. Enfin, Biscutella levigata a tenté de s'installer, mais n'a pas pu persister.
Selon les saisons et les fréquences de mises à feu, les espèces font preuve de comportements différents. Certaines ne réagissent apparemment pas ; d'autres, en revanche, ont tendance à accroître leur degré de présence, d'autres à le diminuer, d'autres enfin à disparaître. L'ordre des espèces dans le tableau III est fonction de leurs réponses vis-à-vis du feu, à savoir : réaction favorable, aucune réaction
- 2 1 4 -
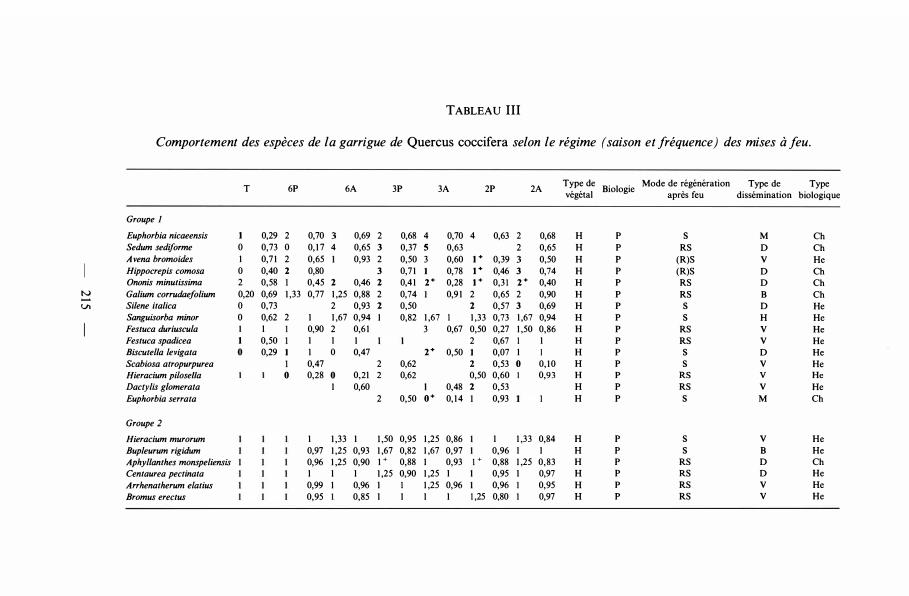
TABLEAU III
Comportement des espèces de la garrigue de Quercus coccifera selon le régime (saison et fréquence) des mises à feu.
T 6P 6A 3P 3A 2P 2A Type de
Biologie Mode de régénération Type de Type
végétal après feu dissémination biologique
Groupe 1 Euphorbia nicaeensis 1 0,29 2 0,70 0,69 2 0,68 4 0,70 4 0,63 2 0,68 H p s M Ch Sedum sediforme 0 0,73 0 0, 1 7 4 0,65 3 0,37 5 0,63 2 0,65 H p RS D Ch Avena bromoides 1 0,7 1 2 0,65 0,93 2 0,50 3 0,60 1+ 0,39 0,50 H p (R)S v He Hippocrepis comosa 0 0,40 2 0,80 3 0,7 1 0,78 1+ 0,46 3 0,74 H p (R)S D Ch Ononis minutissima 2 0,58 0,45 2 0,46 2 0,4 1 2+ 0,28 1+ 0,31 2+ 0,40 H p RS D Ch
N Galium corrudaefolium 0,20 0,69 1 ,33 0,77 1 ,25 0,88 2 0,74 0,9 1 2 0,65 2 0,90 H p RS B Ch Vl Silene italica 0 0,73 2 0,93 2 0,50 2 0,57 3 0,69 H p s D He
Sanguisorba minor 0 0,62 2 1 1 ,67 0,94 0,82 1 ,67 1 1 ,33 0,73 1 ,67 0,94 H p s H He Festuca duriuscu/a 1 0,90 2 0,6 1 3 0,67 0,50 0,27 1 ,50 0,86 H p RS v He Festuca spadicea 1 0,50 1 1 1 2 0,67 1 1 H p RS v He Biscutel/a levigata 0 0,29 1 0 0,47 2+ 0,50 0,07 1 H p s D He Scabiosa atropurpurea 0,47 2 0,62 2 0,53 0 0,1 0 H p s v He Hieracium pi/ose/la 0 0,28 0 0,21 2 0,62 0,50 0,60 0,93 H p RS v He Dactylis g/omerata 1 0,60 0,48 2 0,53 H p RS v He Euphorbia serra/a 2 0,50 o+ 0,1 4 0,93 H p s M Ch
Groupe 2 Hieracium murorum 1 1 ,33 1 1 ,50 0,95 1 ,25 0,86 1 1 ,33 0,84 H p s v He Bupleurum rigidum 0,97 1 ,25 0,93 1 ,67 0,82 1 ,67 0,97 0,96 1 1 H p s B He Aphyllanthes monspeliensis 0,96 1 ,25 0,90 1 + 0,88 1 0,93 1 + 0,88 1 ,25 0,83 H p RS D Ch Centaurea pectinata 1 1 1 1 ,25 0,90 1 ,25 1 0,95 1 0,97 H p RS D He Arrhenatherum e/atius 0,99 1 0,96 1 1 1 ,25 0,96 0,96 0,95 H p RS v He Bromus erectus 0,95 0,85 1 1 1 1 1,25 0,80 0,97 H p RS v He
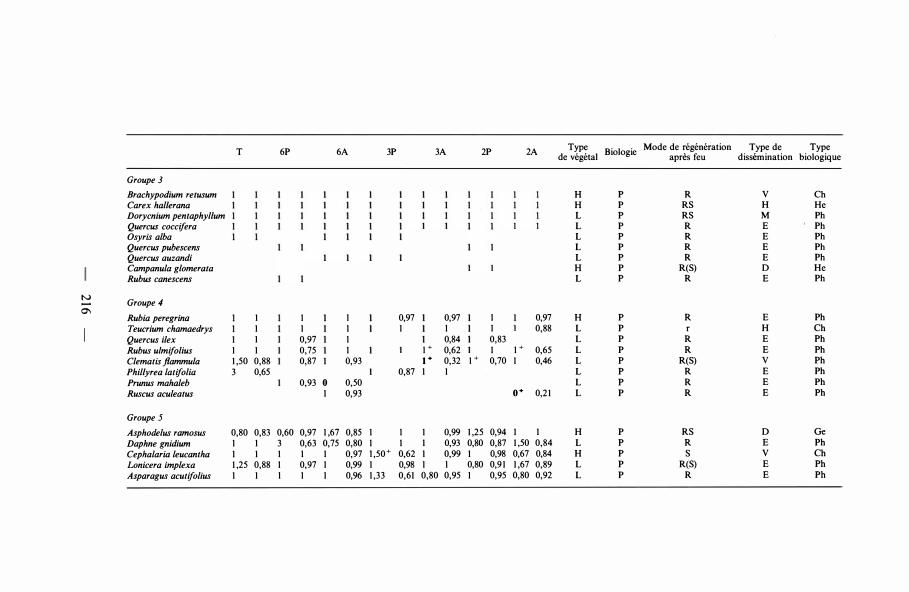
T 6P 6A 3P 3A 2P 2A Type Biologie Mode de régénération Type de Type de végétal après feu dissèmination biologique
Groupe 3 Brachypodium retusum H p R v Ch Carex hallerana H p RS H He Dorycnium pentaphyllum L p RS M Ph Quercus coccifera L p R E Ph Osyris alba L p R E Ph Quercus pubescens L p R E Ph Quercus auzandi L p R E Ph Campanula glomerata H p R(S) D He Rubus canescens L p R E Ph
N Groupe 4 ......
0'1 Rubia peregrino 1 1 1 0,97 0,97 1 0,97 H p R E Ph Teucrium chamaedrys 1 1 1 1 1 1 0,88 L p r H Ch Quercus ilex 1 0,97 1 0,84 0,83 L p R E Ph Rubus ulmifolius 1 1 0,75 1 1 + 0,62 1 1 + 0,65 L p R E Ph C/ematis jlammula 1 ,50 0,88 0,87 0,93 t+ 0,32 1 + 0,70 1 0,46 L p R(S) v Ph Phillyrea /atifo/ia 3 0,65 0,87 1 1 L p R E Ph Prunus maha/eb 0,93 0 0,50 L p R E Ph Ruscus aculeatus 1 0,93 o+ 0,2 1 L p R E Ph
Groupe 5 Asphode/us ramosus 0,80 0,83 0,60 0,97 1 ,67 0,85 1 1 1 0,99 1 ,25 0,94 1 1 H p RS D Ge Daphne gnidium 1 1 3 0,63 0,75 0,80 1 1 1 0,93 0,80 0,87 1 ,50 0,84 L p R E Ph Cephalaria /eucantha 1 1 1 1 1 0,97 I,5o+ 0,62 1 0,99 1 0,98 0,67 0,84 H p s v Ch Lonicera imp/exa 1 ,25 0,88 1 0,97 1 0,99 1 0,98 1 1 0,80 0,9 1 1 ,67 0,89 L p R(S) E Ph Asparagus acutifolius 1 1 1 1 1 0,96 1 ,33 0,61 0,80 0,95 1 0,95 0,80 0,92 L p R E Ph
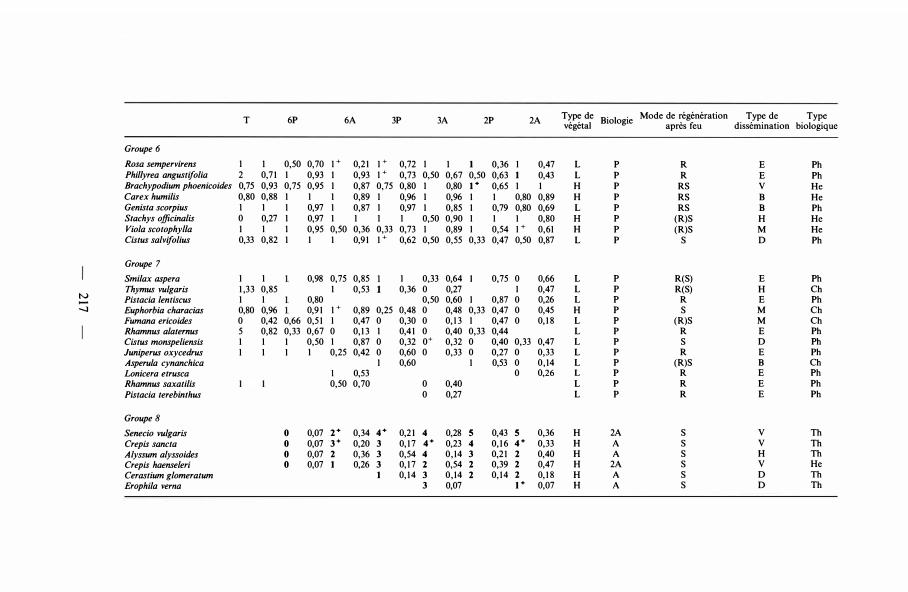
T 6P 6A 3P 3A 2P 2A Type de Biologie Mode de régénération Type de Type végétal après feu dissémination biologique
Groupe 6 Rosa sempervirens 1 1 0,50 0,70 1 + 0,2 1 1 + 0,72 1 1 1 0,36 1 0,47 L p R E Ph Phillyrea angustifo/ia 2 0,7 1 1 0,93 1 0,93 1 + 0,73 0,50 0,67 0,50 0,63 1 0,43 L p R E Ph Brachypodium phoenicoides 0,75 0,93 0,75 0,95 1 0,87 0,75 0,80 1 0,80 1+ 0,65 1 1 H p RS v He Carex humilis 0,80 0,88 1 1 1 0,89 1 0,96 1 0,96 1 1 0,80 0,89 H p RS B He Genista scorpius 1 1 1 0,97 1 0,87 1 0,97 1 0,85 1 0,79 0,80 0,69 L p RS B Ph Stachys officina/is 0 0,27 1 0,97 1 1 1 1 0,50 0,90 1 1 1 0,80 H p (R)S H He Viola scotophylla 1 1 1 0,95 0,50 0,36 0,33 0,73 1 0,89 1 0,54 1 + 0,61 H p (R)S M He Cistus sa/vifolius 0,33 0,82 1 1 1 0,9 1 1 + 0,62 0,50 0,55 0,33 0,47 0,50 0,87 L p s D Ph
Groupe 7 Smilax aspera 1 1 0,98 0,75 0,85 1 0,33 0,64 0,75 0 0,66 L p R(S) E Ph Thymus vulgaris 1 ,33 0,85 1 0,53 0,36 0 0,27 1 0,47 L p R(S) H Ch
N Pistacia lentiscus 1 1 0,80 0,50 0,60 1 0,87 0 0,26 L p R E Ph -.1 Euphorbia characias 0,80 0,96 0,9 1 1 + 0,89 0,25 0,48 0 0,48 0,33 0,47 0 0,45 H p s M Ch
Fumana ericoides 0 0,42 0,66 0,5 1 1 0,47 0 0,30 0 0, 1 3 1 0,47 0 0, 1 8 L p (R)S M Ch Rhamnus alaternus 5 0,82 0,33 0,67 0 0,1 3 1 0,4 1 0 0,40 0,33 0,44 L p R E Ph Cistus monspe/iensis 1 1 1 0,50 1 0,87 0 0,32 o+ 0,32 0 0,40 0,33 0,47 L p s D Ph Juniperus oxycedrus 1 1 1 1 0,25 0,42 0 0,60 0 0,33 0 0,27 0 0,33 L p R E Ph Asperula cynanchica 1 0,60 1 0,53 0 0,14 L p (R)S B Ch L.onicera etrusca 1 0,53 0 0,26 L p R E Ph Rhamnus saxatilis 0,50 0,70 0 0,40 L p R E Ph Pistacia terebinthus 0 0,27 L p R E Ph
Groupe 8 Senecio vu/garis 0 0,07 2+ 0,34 4+ 0,2 1 4 0,28 5 0,43 5 0,36 H 2A s v Th Crepis sancta 0 0,07 ·3+ 0,20 3 0,1 7 4+ 0,23 4 0,1 6 4+ 0,33 H A s v Th Alyssum a/yssoides 0 0,07 2 0,36 3 0,54 4 0,14 3 0,2 1 2 0,40 H A s H Th Crepis haense/eri 0 0,07 1 0,26 3 0,1 7 2 0,54 2 0,39 2 0,47 H 2A s v He Cerastium glomeratum 1 0,1 4 3 0,14 2 0,1 4 2 0,1 8 H A s D Th Erophi/a verna 3 0,07 t+ 0,07 H A s D Th
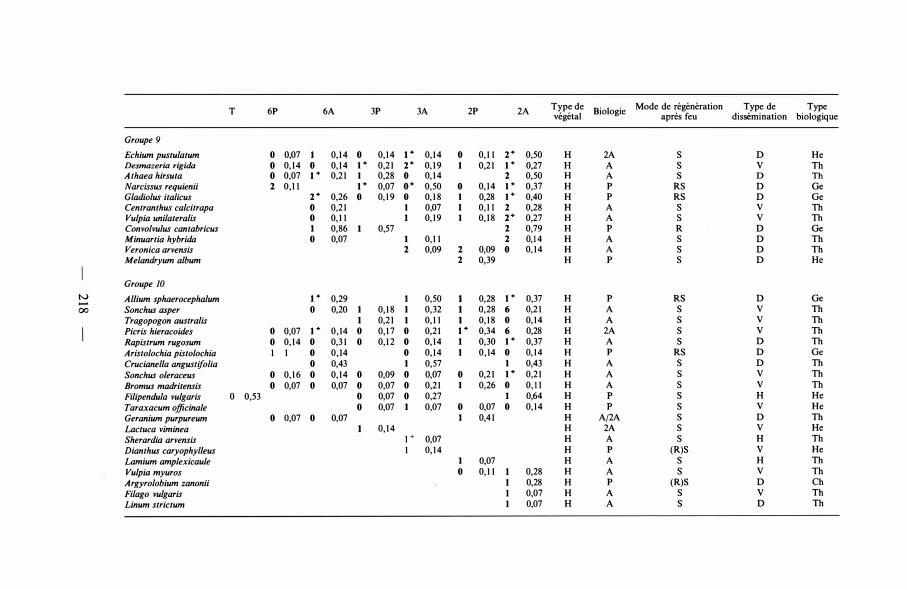
T 6P 6A 3P 3A 2P 2A Type de Biologie Mode de régénération Type de Type végétal après feu dissémination biologique
Groupe 9 Echium pustu/atum 0 O,o7 1 0,1 4 0 0, 1 4 1+ 0,1 4 0 0, 1 1 2+ 0,50 H 2A s 0 He Desmazeria rigida 0 0, 1 4 0 0, 1 4 1+ 0,2 1 2+ 0, 1 9 1 0,2 1 1+ 0,27 H A s v Tb Athaea hirsuta 0 0,07 1+ 0,2 1 1 0,28 0 0,1 4 2 0,50 H A s 0 Tb Narcissus requienii 2 0,1 1 1+ 0,07 o• 0,50 0 0,1 4 1+ 0,37 H p RS 0 Ge G/adiolus italicus 2+ 0,26 0 0, 1 9 0 0, 1 8 1 0,28 1+ 0,40 H p RS 0 Ge Centranthus calcitrapa 0 0,2 1 1 O,o7 1 0, 1 1 2 0,28 H A s v Tb Vu/pia uni/ateralis 0 0, 1 1 1 0, 1 9 1 0, 1 8 2+ 0,27 H A s v Th Convolvulus cantabricus 1 0,86 0,57 2 0,79 H p R 0 Ge Minuartia hybrida 0 0,07 1 0, 1 1 2 0,14 H A s 0 Tb Veronica arvensis 2 0,09 2 0,09 0 0, 1 4 H A s 0 Tb Melandryum album 2 0,39 H p s 0 He
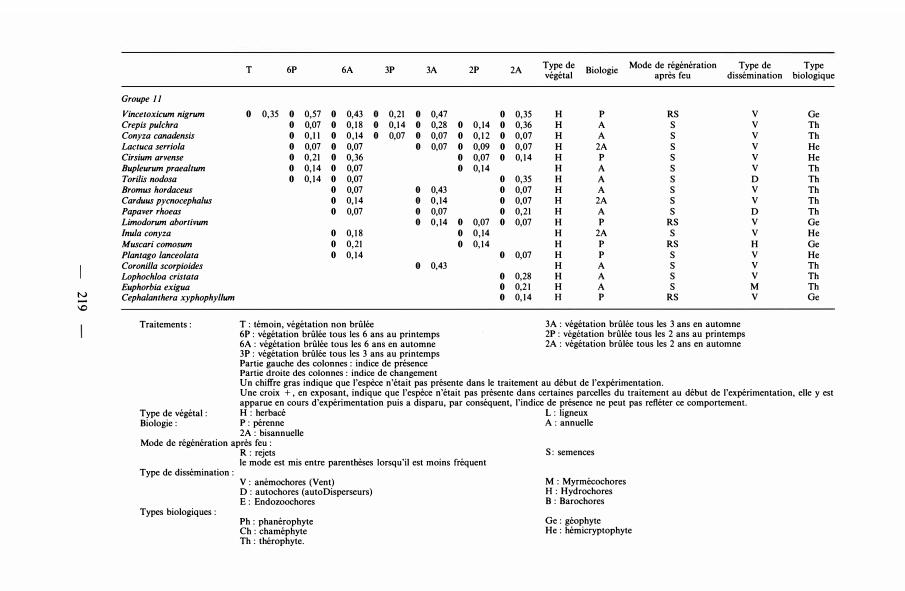
Groupe 10 tv Allium sphaerocepha/um 1+ 0,29 1 0,50 0,28 1+ 0,37 H p RS 0 Ge 00 Sonchus asper 0 0,20 1 0,1 8 1 0,32 0,28 6 0,2 1 H A s v Tb
Tragopogon australis 1 0,2 1 1 0, 1 1 0, 1 8 0 0,1 4 H A s v Tb Picris hieracoides 0 0,07 1+ 0,14 0 0,1 7 0 0,2 1 1+ 0,34 6 0,28 H 2A s v Tb Rapistrum rugosum 0 0, 1 4 0 0,3 1 0 0, 1 2 0 0,1 4 1 0,30 1+ 0,37 H A s 0 Tb Aristolochia pisto/ochia 1 1 0 0,14 0 0,1 4 1 0,14 0 0, 1 4 H p RS 0 Ge Crucianella angustifolia 0 0,43 1 0,57 1 0,43 H A s 0 Tb Sonchus o/eraceus 0 0,1 6 0 0,1 4 0 0,09 0 0,07 0 0,2 1 1+ 0,2 1 H A s v Tb Bromus madritensis 0 0,07 0 0,07 0 0,07 0 0,2 1 1 0,26 0 0,11 H A s v Tb Filipendu/a vu/garis 0 0,53 0 0,07 0 0,27 1 0,64 H p s H He Taraxacum officinale 0 0,07 1 0,07 0 0,07 0 0, 1 4 H p s v He Geranium purpureum 0 0,07 0 0,07 1 0,4 1 H A/2A s 0 Tb Lactuca viminea 0,1 4 H 2A s v He Sherardia arvensis 1 + 0,07 H A s H Tb Dianthus caryophylleus 1 0, 1 4 H p (R)S v He Lamium amplexicaule 1 0,07 H A s H Tb Vu/pia myuros 0 0, 1 1 0,28 H A s v Tb Argyro/obium zanonii 0,28 H p (R)S 0 Ch Fi/ago vu/garis 0,07 H A s v Th Linum strictum 0,07 H A s 0 Tb
N ...... \0
Groupe 1 1 Vincetoxicum nigrum Crepis pulchra Conyza canadensis Lactuca serriola Cirsium arvense Bup/eurum praealtum Torilis nodosa Bromus hordaceus Carduus pycnocephalus Papaver rhoeas Limodorum abortivum /nu/a conyza Muscari comosum Plantago lanceo/ata Coronilla scorpioides Lophoch/oa cristata Euphorbia exigua Cephalanthera xyphophyllum
Traitements :
Type de végétal : Biologie:
T 6P 6A 3P 3A
0 0,35 0 0,57 0 0,43 0 0,21 0 0,47 0 0,07 0 0,1 8 0 0,1 4 0 0,28 0 0,1 1 0 0,14 0 0,07 0 0,07 0 0,07 0 0,07 0 0,07 0 0,21 0 0,36 0 0,1 4 0 0,07 0 0,1 4 0 0,07
0 0,07 0 0,43 0 0,1 4 0 0,14 0 0,07 0 0,07
0 0,14 0 0,1 8 0 0,21 0 0,1 4
0 0,43
T : témoin, végétation non brûlée 6P: végétation brûlée tous les 6 ans au printemps 6A : végétation brûlée tous les 6 ans en automne 3P : végétation brûlée tous les 3 ans au printemps Partie gauche des colonnes : indice de présence Partie droite des colonnes : indice de changement
2P
0 0 0,14 0 0 0,1 2 0 0 0,09 0 0 0,07 0 0 0,1 4
0 0 0 0
0 0,07 0 0 0,14 0 0,1 4
0
0 0 0
2A
0,35 0,36 0,07 0,07 0, 1 4
0,35 0,07 0,07 0,21 0,07
0,07
0,28 0,2 1 0, 1 4
Type de Biologie Mode de régénération Type de végétal après feu dissémination
H p RS H A s H A s H 2A s H p s H A s H A s H A s H 2A s H A s H p RS H 2A s H p RS H p s H A s H A s H A s H p RS
3A : végétation brûlée tous les 3 ans en automne 2P : végétation brûlée tous les 2 ans au printemps 2A : végétation brûlée tous les 2 ans en automne
v v v v v v D v v D v v H v v v M v
Un chiffre gras indique que l'espèce n'était pas présente dans le traitement au début de l'expérimentation.
Type biologique
Ge Th Th He He Th Th Th Th Th Ge He Ge He Th Th Th Ge
Une croix +, en exposant, indique que l'espèce n'était pas présente dans certaines parcelles du traitement au début de l'expérimentation, elle y est apparue en cours d'expérimentation puis a disparu, par conséquent, l'indice de présence ne peut pas refléter ce comportement. H : herbacé L : ligneux P : pérenne A : annuelle 2A : bisannuelle
Mode de régénération après feu :
Type de dissémination :
Types biologiques :
R: rejets le mode est mis entre parenthèses lorsqu'il est moins fréquent
V : anémochores (Vent) D : autochores (autoDisperseurs) E : Endozoochores
Ph : phanérophyte Ch : chaméphyte Th : thérophyte.
S: semences
M : Myrmécochores H : Hydrochores B : Barochores
Ge : géophyte He : hémicryptophyte

apparente, tendance à la diminution de leur degré de présence, et enfin disparition, tout d'abord pour les espèces observées dans les parcelles dès le début de l'expérimentation, et ensuite, pour celles qui sont apparues en cours d'expérimentation. L'utilisation du deuxième indice, concernant le degré de changement au cours du temps, permet de distinguer les tendances évolutives à l'intérieur d'une même catégorie d'espèces ayant le même indice de présence. Selon leurs tendances, les espèces ont ainsi été réparties en 1 1 groupes d'importances très diverses.
A) Espèces favorisées par le feu
Groupe 1. La plupart sont des espèces rencontrées au moins une fois dans un traitement dès le début de l'expérimentation mais dont l'indice de présence croît fortement, et/ou qui apparaissent nouvellement dans certains traitements et y persistent. Ce sont des plantes qui s'installent grâce aux conditions créées par les mises à feu. En général, une fois en place dans les parcelles où elles n'existaient pas avant les brûlages, elles s'y maintiennent à demeure. Elles sont donc favorisées par le feu.
Groupe 2. Ces espèces étaient toutes présentes dans tous les traitements au début de l'expérimentation. Leur degré de présence croît, mais moins fortement que pour le groupe précédent. Lorsqu'elles apparaissent dans un traitement, elles s'installent très rapidement et persistent : leur incide de changement est élevé. Ce sont aussi des végétaux favorisés par le feu.
B) Espèces apparemment indifférentes à l 'action du feu
Groupe 3. Toutes ces espèces sont vivaces et appartiennent au cortège floristique de la garrigue de Q. coccifera (Braun-Blanquet et al. , 1 952), qu'elles caractérisent floristiquement et physionomiquement. Le feu ne semble apparemment pas avoir d'effet sur elles : quelles que soient la fréquence et l'époque des brûlages, elles sont présentes à la fin de la période considérée, et leur nombre de présences n'a pas changé par rapport à celui d'origine. L'indice de changement est aussi égal à 1 , montrant que ces végétaux sont toujours présents tout au long de la période d'observation.
Ces espèces résistent bien au feu. Et, quoique celui-ci ne semble pas avoir d'effet sur elles, il n'est pas impossible qu'en dehors des parcelles délimitées le feu favorise leur extension au détriment d'autres espèces.
Ont été incluses dans ce groupe Osyris alba, Quercus pubescens, Q . auzandi, Campanula glomerata, Rubus canescens, qui ont les valeurs des deux indices égales à 1 , mais ne sont pas suffisamment représentées dans l 'ensemble des parcelles . Si elles avaient été présentes dans un plus grand nombre de traitements, il se peut que leur comportement eut été différent et qu'il aurait pu être mieux précisé.
Groupe 4. C'est un groupe très voisin du précédent. Les espèces présentent exactement le même comportement en ce qui concerne l'indice de présence : elles ont persisté tout au long de la période ; toutefois, l'indice de changement, dans un des traitements quel qu'il soit, est inférieur à 1 : à un moment quelconque, généralement au printemps suivant un brûlage, l'espèce n'a pas été observée ; elles sont donc légèrement plus sensibles au feu que les espèces du groupe précédent.
- 220 -

Groupe 5. Les espèces incluses dans ce groupe ont un comportement indéterminé. Parfois, le feu semble favoriser leur extension ; parfois, au contraire, leur degré de présence est diminué, et cela indépendamment de la fréquence ou de la saison des mises à feu. Toutefois, l'indice de changement présente, au moins dans un traitement, des valeurs inférieures à 1 . Il semblerait qu'Asparagus acutifolius soit sensible plus particulièrement aux feux d'automne.
C) Espèces défavorisées par l 'action du feu
Groupe 6. Ces espèces sont sensibles à l'action du feu. Généralement, des mises à feu tous les 6 ans n'entraînent pas une diminution du nombre de fois où elles sont présentes sur les parcelles. Mais si les fréquences des mises à feu sont plus nombreuses (tous les 2 ou 3 ans), l'indice de présence diminue. Cette diminution est d'autant plus marquée que les brûlages sont plus nombreux et allumés en automne. Cependant, à la différence de celles du groupe suivant, les espèces ne disparaissent pas complètement quels que soient le nombre et l'époque des mises à feu. Cistus salvifolius semble être l'espèce la plus sensible.
Groupe 7. Les plantes incluses dans ce groupe sont très sensibles à l'action du feu puisqu'elles peuvent disparaître complètement. Les indices de changement ont, généralement, de faibles valeurs, indiquant une persistance de courte durée. Les cas de disparition sont d'autant plus nombreux que la fréquence des brûlages est élevée ( 1 cas seulement de disparition totale lorsque les feux sont allumés tous les 6 ans ; 1 1 cas pour un feu tous les trois ans et 9 cas lorsque les brûlages ont lieu tous les 2 ans) . L'époque des mises à feu a un effet marqué sur les disparitions, puisque les feux d'automne provoquent 76,2 % des cas de disparition.
D) Espèces apparaissant nouvellement et s'installant
Les espèces, qui constituent les groupes qui vont suivre, sont généralement étrangères au cortège ftoristique normal de la garrigue de Q. coccifera. Elles n'étaient pas présentes dans les parcelles avant le début de l'expérimentation. Ce sont toutes les espèces herbacées ; aucune n'est ligneuse. Leur venue a été indirectement favorisée par le feu qui a créé des espaces momentanément vides de végétation. Elles se maintiennent grâce à cette action ; en l'absence de feu elles disparaissent.
Groupe 8. Ce groupe est constitué par des espèces dont l'indice de présence est caractérisé par de fortes valeurs, principalement lorsque les brûlages sont allumés fréquemment (tous les 2 ou 3 ans) . Elles ne persistent que grâce à l'action répétée du feu, sinon elles disparaîtraient.
Groupe 9. Le comportement des végétaux de ce groupe est très comparable à celui du groupe précédent. Les différences résident en des valeurs plus faibles des indices de présence et de changement, indiquant soit une moins grande capacité de dissémination, soit une moindre résistance aux feux, soit une persistance moins grande. Quelques tentatives d'installation peuvent même avoir avorté.
Groupe 10. Les nouvelles espèces incluses dans ce groupe présentent un comportement indéterminé : tantôt l'indice de présence est égal à 1 , tantôt il est
- 22 1

égal à 0 ; les individus apparaissent ou disparaissent. Ces espèces n'arrivent réellement pas à s'installer et les mises à feu fréquentes ne favorisent pas leur installation. Cependant, 58,7 % des tentatives réussies ou des échecs d'installation ont eu lieu avec des mises à feu d'automne.
E) Espèces apparaissant nouvellement mais ne s 'installant pas
Groupe 11. Toutes les espèces de ce groupe, nouvellement apparues, ont été observées à un moment donné au cours de l 'expérimentation, mais étaient absentes lors de la dernière période d'observation ; elles n'ont donc pas pu s'installer. Ici aussi, les tentatives d'installation sont les plus nombreuses avec les brûlages d'automne, certaines espèces n'apparaissant qu'à la faveur de ces mises à feu. Les seules Orchidées observées lors de l 'expérimentation n'apparaissent que dans ce groupe.
MODES DE DISSÉMINA TION ET DE RÉSISTANCE
Le comportement des espèces par rapport au régime des feux est déterminé par leur capacité à résister mais aussi à se disséminer pour occuper l 'espace brûlé. Il est possible de caractériser les groupes d'espèces établis par les modes de dissémination qu'utilisent les végétaux qui les constituent. Certains modes permettent, plus que d'autres ou plus rapidement, une conquête de l'espace. Ce sont certaines grandes catégories (au nombre de 6) de dissémination parmi celles proposées par Molinier & Muller ( 1 938) qui ont été utilisées. Toutefois, les plantes emploient bien souvent plusieurs types de dissémination en même temps ; aussi, pour chaque espèce observée, le moyen qui a été pris en compte est celui considéré par ces deux auteurs comme le principal, mais en y apportant des précisions plus restrictives qu'il faut souligner brièvement. Les anémochores (symbolisés par V dans le tableau III) sont des végétaux dont le Vent est le principal agent transporteur, leurs diaspores ne pouvant être transportées pratiquement que par lui . Les autochores (D pour autoDisperseurs) sont des espèces qui assurent elles-mêmes la dissémination de leurs diaspores par divers moyens tels que crochets, tiges ou capsules projetantes. Les végétaux dont les diaspores sont transportées par les animaux ont été divisées en deux catégories : les Endozoochores (E) transportés par les animaux après ingestion, et les Myrmécochores (M) transportés exclusivement par les fourmis. Les Hydrochores (H) ont leurs diaspores entraînées par l'eau, et les Barochores (B) sont ceux dont les diaspores se détachent et tombent à terre de leur propre poids.
Les groupes 1 et 2 (Tab. IV) caractérisés par un accroissement de l 'indice de présence, possèdent de forts pourcentages de végétaux anémochores et autochores ; ce sont des modes de dissémination permettant une rapide dispersion et favorisant les invasions. L'absence d'endozoochores est à noter.
Les groupes 3, 4 et 5 qui possèdent les espèces les plus résistantes et qui ont persisté tout au long de la période des observations, sont caractérisés par de forts pourcentages (presque exclusifs) d'endozoochores. Ce sont les espèces les plus fréquentes, les plus dominantes et caractérisant physionomiquement ce type de garrigue. Il faut noter aussi l'absence de plantes barochores.
Le groupe 6, dans lequel coexistent les espèces dont l'indice de présence décroît, est caractérisé par la présence des six modes de dissémination sans prédominance de l'un d'entre eux.
- 222 -

TABLEAU IV
Répartition (en pourcentages) des modes de dissémination pour les groupes d'espèces identifiés.
Modes de dissémination 2 3 4 5 6 7 8 9 10 I l
Anémochores (V) 40,00 50,00 1 1 , 1 1 12,50 20,00 12,50 50,00 27,27 50,00 77,78 Autochores (D) 33,33 33,33 1 1 , 1 1 20,00 12,50 8,33 33,33 72,73 35,00 1 1 , 1 1 Endozoochores (E) 55,56 75,00 60,00 25,00 58,33 Myrmécochores (M) 1 3,33 1 1 , 1 1 12,50 1 6,67 5,56 Hydrochores (H) 6,67 1 1 , 1 1 12,50 12,50 8,33 16,67 1 5,00 5,56 Barochores (B) 6,67 1 6,67 25,00 8,33
Le groupe 7, chez lequel les espèces ont tendance à disparaître lorsque les mises à feu sont trop fréquentes, est caractérisé par un fort pourcentage d'endozoochores, mais surtout par l'absence totale d'anémochores. Aucune des espèces participant à ce groupe n'a la possibilité de se disséminer par le vent. Est-ce une des raisons de leur propension à disparaître ?
Enfin, les derniers groupes 8, 9, 10 et 1 1 , constitués par des espèces généralement étrangères à la communauté, et qui envahissent ou tentent d'envahir les espaces brûlés, sont caractérisés par des pourcentages élevés de plantes anémochores et autochores (comme pour les deux premiers groupes) . Ces modes de dissémination permettent de rapides colonisations sur des distances relativement grandes. Il faut aussi souligner, chez ces quatre groupes, l'absence totale, ou quasi totale, de végétaux endozoochores, barochores et myrmécochores. Ce fait est facilement compréhensible puisque ces plantes sont des espèces qui n'étaient pas présentes avant les mises à feu et qui vont s'introduire à la faveur des brûlages. L'absence de la myrmécochorie est, toutefois, étrange car les fourmis peuvent transporter des diaspores sur d'assez longues distances et permettre une large dispersion, même après feu (Troumbis & Trabaud, 1987) .
Bien que ne permettant pas de caractériser précisément la régénération des végétaux après incendie (Trabaud, 1 987), les types biologiques de Raunkiaer ( 1 905), fréquemment utilisés en biologie, peuvent contribuer à donner un aperçu des formes de végétaux qui sont soit favorisés, soit résistants, soit défavorisés par le feu.
L'absence de phanérophytes, géophytes et thérophytes est un trait des groupes 1 et 2 (Tab. V) chez lesquels les espèces tendent à accroître leur degré de présence ou à envahir lès parcelles brûlées. Ces deux groupes sont constitués essentiellement par des hémicryptophytes et accessoirement des chaméphytes, formes végétales de petite taille. Les espèces sont toutes des herbacées pérennes, ayant de bonnes capacités de dissémination et de régénération à la fois par semences et par rejets.
Les groupes 3 à 7 caractérisés soit par leur résistance (ou indifférence) au régime des feux soit par leur faiblesse ou même leur disparition à cause des feux, ne possèdent pas (ou pratiquement pas) de géophytes ou de thérophytes. Par contre, ils ne sont pratiquement constitués que par des phanérophytes (pourcentages tous supérieurs à 50 % ) . Ce sont des végétaux de bonne taille, pérennes, soit
- 223 -
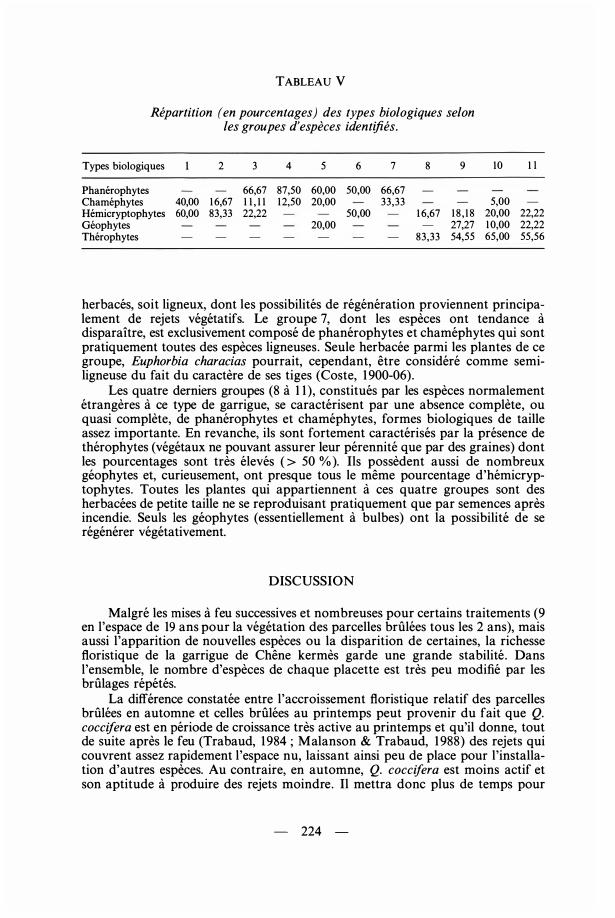
TABLEAU V
Répartition (en pourcentages) des types biologiques selon les groupes d'espèces identifiés.
Types biologiques 2 3 4 5 6 7 8 9
Phanérophytes 66,67 87,50 60,00 50,00 66,67 Chaméphytes 40,00 1 6,67 1 1 , 1 1 12,50 20,00 33,33 Hémicryptophytes 60,00 83,33 22,22 50,00 1 6,67 1 8 , 1 8 Géophytes 20,00 27,27 Thérophytes 83,33 54,55
10 I l
5,00 20,00 22,22 1 0,00 22,22 65,00 55,56
herbacés, soit ligneux, dont les possibilités de régénération proviennent principalement de rejets végétatifs. Le groupe 7, dont les espèces ont tendance à disparaître, est exclusivement composé de phanérophytes et chaméphytes qui sont pratiquement toutes des espèces ligneuses. Seule herbacée parmi les plantes de ce groupe, Euphorbia characias pourrait, cependant, être considéré comme semiligneuse du fait du caractère de ses tiges (Coste, 1 900-06) .
Les quatre derniers groupes (8 à 1 1 ), constitués par les espèces normalement étrangères à ce type de garrigue, se caractérisent par une absence complète, ou quasi complète, de phanérophytes et chaméphytes, formes biologiques de taille assez importante. En revanche, ils sont fortement caractérisés par la présence de thérophytes (végétaux ne pouvant assurer leur pérennité que par des graines) dont les pourcentages sont très élevés ( > 50 % ). Ils possèdent aussi de nombreux géophytes et, curieusement, ont presque tous le même pourcentage d'hémicryptophytes. Toutes les plantes qui appartiennent à ces quatre groupes sont des herbacées de petite taille ne se reproduisant pratiquement que par semences après incendie. Seuls les géophytes (essentiellement à bulbes) ont la possibilité de se régénérer végétativement.
DISCUSSION
Malgré les mises à feu successives et nombreuses pour certains traitements (9 en l'espace de 19 ans pour la végétation des parcelles brûlées tous les 2 ans), mais aussi l'apparition de nouvelles espèces ou la disparition de certaines, la richesse ftoristique de la garrigue de Chêne kermès garde une grande stabilité . Dans l'ensemble, le nombre d'espèces de chaque placette est très peu modifié par les brûlages répétés.
La différence constatée entre l'accroissement ftoristique relatif des parcelles brûlées en automne et celles brûlées au printemps peut provenir du fait que Q. coccifera est en période de croissance très active au printemps et qu'il donne, tout de suite après le feu (Trabaud, 1 984 ; Malanson & Trabaud, 1 988) des rejets qui couvrent assez rapidement l'espace nu, laissant ainsi peu de place pour l'installation d'autres espèces. Au contraire, en automne, Q. coccifera est moins actif et son aptitude à produire des rejets moindre. Il mettra donc plus de temps pour
- 224 -

réoccuper l'espace, ce qui permettra à d'autres espèces de s'installer, au moins momentanément. Les annuelles, entre autres, seront plus nombreuses au moment des observations.
Ces présences, multiples mais temporaires d'espèces annuelles qui essayent de s'installer à la faveur des brûlages sont parfaitement bien visibles dans le tableau Il. Ainsi, le nombre d'espèces qui, absentes au début de l'expérimentation sont présentes à la fin, croît avec la fréquence des mises à feu et plus particulièrement celles d'automne : l'action des feux récurrents entretient la présence de ces espèces. Il en est de même, mais à un moindre degré, pour les espèces qui ne sont présentes qu'à un certain moment au cours de l'expérimentation. Pour ces dernières, les valeurs sont plus élevées dans le cas de mises à feu tous les 6 ans ; ainsi une trop forte fréquence des brûlages les élimine, à moins qu'il ne s'agisse d'espèces extrêmement fugaces se renouvelant constamment.
Les feux amènent peu de disparitions d'espèces pérennes. Toutefois, les disparitions sont liées à la fréquence et à la saison des mises à feu ; ce sont ceux de printemps qui sont les moins nocifs.
La fermeture de la végétation entraîne une disparition d'espèces aussi nombreuse que les mises à feu d'automne tous les 2 ou 3 ans. Au fur et à mesure que la garrigue avance en âge (au début de l'expérimentation elle était âgée de 1 8 ans après un feu sauvage, et elle avait 37 ans à la fin), et qu'elle devient plus dense et plus haute, les espèces apparues au cours des stades antérieurs ne peuvent plus supporter les nouvelles conditions de milieu et disparaissent. De nouvelles espèces, appartenant surtout à des stades plus mûrs de cette communauté ou à des communautés plus évoluées dans la dynamique forestière progressive, vont alors s'installer ou accroître leur nombre de présences. La plupart d'entre elles (tels les Phyllyrea et Rhamnus) possèdent des baies charnues qui sont grandement disséminées par les oiseaux comme le pense Debussche ( 1 985, 1 988).
Toutefois, la stabilité de la flore est assurée par un ensemble important d'espèces (plus de 53 % pour la totalité des traitements) qui sont présentes du début jusqu'à la fin de la période des observations . Ce sont les végétaux permanents, dominants et caractéristiques de ce type de garrigue. Dans ce cas aussi, la fréquence et la saison des mises à feu déterminent l'importance de ces plantes résistantes, les feux de printemps étant les moins nocifs.
La fréquence des feux une fois tous les 6 ans est celle qui entraîne le moins de modifications. Les espèces qui envahissent les parcelles puis s'y maintiennent sont des plantes herbacées pérennes ayant des moyens de dissémination (vent, autodispersion) leur permettant de se disperser facilement. Les végétaux qui résistent (ou sont apparemment indifférents aux feux) sont toujours des pérennes soit ligneux soit herbacés, se régénérant préférentiellement par voie végétative ; tandis que ceux qui sont sensibles au feu ou disparaissent, présentent une plus grande variété de types morphologiques pouvant se régénérer par plusieurs moyens, mais possédant peu ou pas d'espèces anémochores. Leur capacité de régénération par voie végétative est faible et la trop forte fréquence des feux les élimine. Selon Keeley & Zedler ( 1 978), les végétaux ligneux de petite taille (buissons) qui se régénèrent végétativement résistent beaucoup mieux lorsque les feux sont très fréquents (courts intervalles), alors que ceux qui ne se reproduisent que par voie sexuée ont besoin de plus longs intervalles entre les mises à feu. Le comport,mentde nombreux végétaux observés lors de notre expérimentation confirme ce modèle . Toutefois, certaines espèces ne se régénérant que par rejets de souches ne peuvent pas supporter des fréquences de mises à feu trop rapprochées et
- 225 -

disparaissent donc (en particulier certaines espèces du groupe 7). Ces végétaux ne remplissent donc pas les conditions du modèle théorique proposé.
Les espèces « étrangères » et absentes de la communauté à l'origine qui tendent de s'installer à la faveur des brûlages sont toutes herbacées, principalement des annuelles ou bisannuelles ne se reproduisant que par semences et utilisant des moyens de dissémination (vent, autochorie) très efficaces. Il est bien connu (Keeley, 198 1 ) que les annuelles sont aptes à coloniser très rapidement tous les espaces perturbés (quelle que soit la perturbation), ce qui entraîne la très grande présence des annuelles dans la végétation fréquemment brûlée. Malgré leur présence en abondance, ces espèces (de petite taille) ne sont jamais dominantes et ne participent guère à la phytomasse.
Les espèces « envahissantes », qu'elles appartiennent aux catégories déjà présentes au début de l'expérimentation ou qu'elles soient apparues après, utilisent des moyens très « actifs » de dissémination : vent et autodispersion, tandis que celles qui « résistent » ou qui disparaissent utilisent plutôt des modes de dissémination « passifs » : transport par les animaux après passage ou non dans l'appareil digestif. Ceci peut être un handicap de dispersion pour ces dernières.
Enfin, les espèces qui colonisent les espaces brûlés sont préférentiellement des hémicryptophytes. Celles qui résistent passivement ou disparaissent sont plutôt des phanérophytes, tandis que les thérophytes constituent la masse de la cohorte des plantes nouvelles envahissantes.
La composition floristique de la garrigue de Chêne kermès est très stable dans son ensemble : après les brûlages, elle est quasi identique à celle qui existait à l'origine. Les espèces caractéristiques dominantes sont constamment présentes. L'apparition, ou la disparition, de certaines d'entre elles n'entraîne qu'un changement floristique minime à l'intérieur de la communauté. L'influence de la saison des mises à feu est beaucoup plus importante que leur fréquence : l'accroissement temporaire de la richesse floristique et la disparition de certaines espèces sont plus particulièrement dus aux mises à feu d'automne.
Cette relative stabilité provient du fait que la majorité des espèces présentes dans les parcelles avant les feux se régénère essentiellement par voie végétative (rejets), tandis que les espèces « étrangères » envahissantes n'apparaissent qu'à la faveur des brûlages sous la forme d'individus issus de semences, ne constituant jamais de peuplements denses ni une importante phytomasse ; ils sont, la plupart du temps, éliminés par les espèces d'origine.
Cette étude confirme et surtout précise les tendances déjà entrevues au cours des précédents travaux (Trabaud, 1 974, 1 984 ; Trabaud & Lepart, 1 98 1 ) . Les végétaux qui préexistaient réoccupent rapidement l'espace ouvert par le feu et empêchent toute modification du milieu pouvant permettre l'arrivée de nouvelles plantes. Ce phénomène apparaît dans beaucoup de situations (Connell & Slatyer, 1 977) : par reproduction végétative, les espèces dominantes peuvent persister très longtemps. Les espèces nouvellemment apparues, principalement des annuelles, disparaissent rapidement du fait de la concurrence interspécifique dès que l'influence du feu s'atténue. Il y a disparition de certaines espèces mais la plupart des dominantes demeurent. L'action du feu entraîne seulement un changement de structure : une disparition des strates hautes ( > 1 rn) dominées par des végétaux ligneux au bénéfice des strates basses ( < 1 rn) plus denses et floristiquement plus diversifiées (Trabaud, 1984) .
Cette étude apporte enfin de nombreux renseignements sur le comportement des espèces de la garrigue de Q. coccifera vis-à-vis du régime (saison et fréquence)
- 226 -

des feux : favorisées, indifférentes ou résistantes, sensibles, etc . . . Cependant, ses résultats peuvent être appliqués à d'autres communautés, car les végétaux rencontrés au cours de l 'étude existent dans de nombreuses autres phytocénoses de la région méditerranéenne. Ses résultats peuvent, aussi, contribuer à la conservation et à la protection de certaines espèces, connaissant leur degré de résistance ou de sensibilité, soit par une utilisation du feu, soit par une interdiction des incendies, selon les écosystèmes dans lesquels elles vivent.
RÉSUMÉ
Une expérimentation a été réalisée dans une garrigue de Quercus coccifera du sud de la France afin d'analyser les effets des fréquences et des saisons de mises à feu sur les réactions des végétaux. Le régime des brûlages consistait en des feux de fin de printemps ou de début d'automne, soit tous les 6 ans, les 3 ans ou les 2 ans. L'expérimentation a débuté en 1 969 et a duré pendant 19 ans (jusqu'en 1987) . Chaque année, au printemps, des listes floristiques ont été établies pour toutes les parcelles .
Selon leurs réactions au régime des feux, les espèces ont été classées en 1 1 groupes correspondant à leur comportement à savoir : espèces favorisées par le feu (dont l'indice de présence augmente), espèces apparemment indifférentes (persistantes), espèces défavorisées (indice de présence décroissant ou espèces disparaissant parfois) ou espèces étrangères capables ou non d'installation (envahissantes ou occasionnelles) . La plupart des espèces dont l'indice de présence augmente sont des herbacées pérennes, les indifférentes ou celles qui ont tendance à décroître ou à disparaître sont des ligneuses pérennes ; après feu elles se régénèrent toutes par rejets et/ou semences. Les espèces envahissantes ou occasionnelles sont principalement des annuelles ou bisannuelles se reproduisant seulement par graines, sauf chez les géophytes. La dissémination par anémochorie et autochorie caractérise les espèces dont le degré de présence s'accroît ; chez les indifférentes c'est l 'endozoochorie qui prédomine, tandis que les espèces dont le degré de présence tend à décroître ou à disparaître ne présentent pas de mode de dissémination caractéristique. Les espèces envahissantes et occasionnelles sont surtout anémochores ou autochores . Les réactions des végétaux sont aussi considérées en fonction des types biologiques.
Malgré les changements qui apparaissent l'ensemble de la composition floristique de la garrigue est resté pratiquement inchangé, traduisant une remarquable stabilité .
SUMMARY
This paper summarizes the results of a 19 years study of the effects on vegetation of the burning frequency ( every six th, third, or second year), and of the season of burning (late spring or early autumn), in a Quercus coccifera « garrigue » of southern France. The species present each spring on all the experimental plots were recorded.
Depending on the species' responses to the various fire regimes, plants can be allocated to five major ecological groups, i .e. : fire-favoured (increasing) species,
- 227 -

indifferent (persistent) species, disavantaged (decreasing or disappearing) species, alien settlers (permanent invaders), and occasionals (temporary settlers). Most of these groups can in turn be further divided in sub-groups ( I l altogether) .
Most of the fire-favoured species are herbaceous perennials, whereas indifferent and disavantaged species are woody perennials ; ali of them propagate after fire by sprouts and/or seeds. Most new settlers and occasionals are annuals or biannuals reproducing by seeds, except for the geophytes .
Dispersal also varies among the five major groups : wind dispersal and autochory characterize the fire-favoured species, and animal dispersal (endozoochory) the indifferent species. Dispersal is very variable in disavantaged species. New settlers and occasionals almost exclusively disperse by wind or autochory.
As for the !ife forms, hemicryptophytes and chamaephytes predominate among fire-favoured species, phanerophytes among indifferent and disavantaged species, and therophytes among new settlers.
On the whole, repeated burning did not change much the fioristic composition of the Quercus coccifera « garrigue » studied : more than 53 % of the species present at the beginning of the experiment were still present 1 9 years later, whatever the fire regime.
REMERCIEMENTS
L'auteur remercie tous les techniciens qui l'ont assisté tout a long des multiples mises à feu : Messieurs H. Bastide, M. Beltran, C. Collin, M . Grandjanny, R. Marican, F. Rouvier, P. Souillac, J. Trauchessec. Il ne saurait oublier N. Denelle pour son aide lors des déterminations d'espèces ou d'échantillons difficiles.
RÉFÉRENCES
ABBAS, H., BARBERO, M. & LOISEL, R. ( 1 984). - Réflexions sur le dynamisme actuel de la régénération du pin d'Alep dans les pinèdes incendiées en Provence calcaire. Ecologia Mediterrranea, 10 : 85- 104.
ARIANOUTSOU, M. (1 984). - Post-tire successional recovery of a phryganic (east Mediterranean) ecosystem. Acta Oecologica, Oecologia Plantarum, 5 : 387-394.
ARIANOUTSOU, M. & MARGARIS, N.S. ( 1981) . - Early stages of regeneration after tire in a phryganic ecosystem (east Mediterranean). I. Regeneration by seed germination. Biologie Ecologie Méditerranéennes, 8 : 1 1 9- 128.
BARBERO, M. , BONIN, G., LOISEL, R., MIGLIORETTI, F. & QUÉZEL, P. ( 1 987). - Incidence of exogenous factors on the regeneration of Pinus halepensis after tire. Influence of tire on the stability of Mediterranean forest ecosystems. Ecologia Mediterranea, 1 3 : 5 ! -56.
BRAUN-BLANQUET, J. , ROUSSINE, N. & NÈGRE, R. ( 1 952). - Les groupements végétaux de la France méditerranéenne. Centre National de la Recherche Scientifique, Paris.
CASAL, M., BASANTA, M. & GARCIA Novo, F. ( 1 986). - Sucesion secundaria de la vegetacion herbacea tras el incendio del matorral bajo replobacion forestal de Pinus. Boletin, Sociedad Espafiola de Historia Natural (Biologia), 82 : 25-34.
CASAL, M., BASANTA, M., ÜONZALEZ, F., MONTERO, R., PEREIRAS, J . & PUENTES, A. ( 1990). Post-tire dynamics in experimental plots of shrubland ecosystems in Galicia (NW Spain) . In : Pire in Ecosystem Dynamics, J.G. Goldammer & M .J. Jenkins (eds). SPB Academie Publishing, The Hague, pp. 33-42.
CONNELL, J .H. & SLATYER, R.O. ( 1 977). - Mechanisms of succession in natural communities and their role in community stability and organization. American Naturalist, I l l , 1 1 19- 1 1 44.
COSTE, H. ( 1900-06). - Flore descriptive et illustrée de la France. Paris.
228

DEBUSSCHE, M. ( 1 985). - Rôle des oiseaux disséminateurs dans la germination des graines de plantes à fruits charnus en région méditerranéenne. Acta Oecologica, Oeco/ogia Plantarum, 6 : 365-374.
DEBUSSCHE, M. ( 1 988). - La diversité morphologique des fruits charnus en Languedoc méditerranéen : relations avec les caractéristiques biologiques et la distribution des plantes et avec les disséminateurs. A cta Oecologia, Oecologia Generalis, 9 : 37-52.
DE LILLIS, M. & TEST!, A. ( 1990). - Post-l'Ire dynamics in a disturbed mediterranean community in central ltaly. In : Fire in Ecosystem Dynamics, J.G. Goldamrner & M .J. Jenkins (eds). SPB Academie Publishing The Hague, pp. 53-62.
EMBERGER, L. ( 1942). - Un projet de classification des climats au point de vue phytogéographique. Bulletin de la Société d'Histoire Naturelle de Toulouse, 78 : 1 59- 1 80.
EMBERGER, L. ( 1 97 1 ) . - Considérations complémentaires au sujet des recherches bioclimatologiques et phytogéographiques-écologiques. In : Travaux de Botanique et d'Ecologie. Masson, Paris, pp. 29 1 -30 1 .
KEELEY, J.E. ( 198 1 ) . - Reproductive cycles and fire regimes. In : Fire Regimes and Ecosystem Properties. USDA Forest Service, General Technical Report W0-26 : 23 1 -277.
KEELEY, J.E. & ZEDLER, P.H. ( 1 978). - Reproduction of chaparral shrubs after fire : a comparison of sprouting and seeding strategies. American Mid/and Naturalist, 99 : 142- 1 6 1 .
MALANSON, G.P. & TRABAUD, L. ( 1 988). - Vigour of post-fire resprouting b y Quercus coccifera L. Journal of Eco/ogy, 76 : 3 5 1 -365.
MAZZOLENI, S . & PIZZOLONGO, P. ( 1 990) . - Post-fire regeneration patterns of mediterranean shrubs in the Campania region, southern Italy. In : Fire in Ecosystem Dynamics, J.G. Goldammer & M.J. Jenkins (eds) . SPB Academie Publishing The Hague, pp. 43-5 1 .
MOLINIER, R . & MULLER, P. ( 1 938). - La dissémination des espèces végétales. Revue Générale de Botanique, 50 : 1 - 1 78 .
NAVEH, Z. ( 1 974). - Effects of fire in the Mediterranean region. In : Fire and Ecosystems. T.T. Kozlowski & C.E. Ahlgren (eds). Academie Press, New York, pp. 40 1 -434.
NAVEH, Z. ( 1 975). - The evolutionary significance of fire in the Mediterranean region. Vegetatio, 29 : 1 99-208.
PAPANASTASIS, V.P. ( 1 977). - Fire ecology and management of phrygana communities in Greece. Environmental consequences of fire and fuel management in Mediterranean ecosystems. USDA Forest Service, General Technical Report W0-3 : 476-482.
PRODON, R. , FoNs, R. & PETER, A.M. ( 1 984). - L'impact du feu sur la végétation, les oiseaux et les micro-mammifères dans diverses formations méditerranéennes des Pyrénées orientales : premiers résultats. Revue d'Ecologie {Terre et Vie) , 39 : 129- 1 58 .
RAUNKIAER, C . ( 1 905). - Types biologiques pour la géographie botanique. Danske Videnskabernes Selskabs Forhandl. , 5 : 347-437.
TARREGA, R. & LUIS-CALABUIG, E. ( 1987). - Effects of fire on structure, dynamics and regeneration of Quercus pyrenaica ecosystems. Influence of Fire on the Stability of Mediterranean Forest Ecosystems. Ecologia Mediterranea, 1 3 : 79-86.
THANOS, C., MARCOU, S., CHRISTODOULAKIS, D. & TANNITSAROS, A. ( 1989). - Early post-fire regeneration in Pinus brutia forest ecosystem of Samos island (Greece). Acta Oeco/ogica, Oecologia Plantarum, 10 : 79-94.
TRABAUD, L. ( 1970). - Quelques valeurs et observations sur la phyto-dynamique des surfaces incendiées dans le Bas-Languedoc. Naturalia Monspelensia, 2 1 : 23 1 -242.
TRABAUD, L. ( 1974). - Experimental study of the effects of prescribed burning on a Quercus coccifera L. garrigue. Annual Tai/ Timbers Fire Eco/ogy Conference, 1 3 : 97- 1 29.
TRABAUD, L. ( 1 977). - Comparison between the effect of prescribed tires and wild tires on the global quantitative development of the kermes scrub oak (Quercus coccifera) garrigues. Symposium on Environmental Consequences of Fire & Fuel Management in Mediterranean Ecosystems. US Forest Service, General Technica/ Report W0-3, Washington, 272-282.
TRABAUD, L. ( 1 983). - Evolution après incendie de la structure de quelques phytocénoses méditerranéennes du Bas-Languedoc (Sud de la France). Annales des Sciences Forestières, 40 : 1 77- 195 .
TRABAUD, L. ( 1984). - Changements structuraux apparaissant dans une garrigue de Chêne kermès soumise à différents régimes de feux contrôlés. Acta Oecologica, Oecologia App/icata, 5 : 127- 143 .
229

TRABAUD, L. ( 1 987). - Fire and survival traits of plants. In : Ro/e of Fire in Ecological Systems. L. Trabaud (ed.). SPB Academie Publishing, The Hague, pp. 65-89.
TRABAUD, L. & LEPART, J. ( 1980). - Diversity and stability in garrigue ecosystems after fire. Vegetatio, 43 : 49-57.
TRABAUD, L. & LEPART, J. ( 198 1) . - Floristic changes in a Quercus coccifera L. garrigue according to different fire regimes. Vegetatio, 46 : 105- 1 16 .
TRABAUD, L. , GROSMAN, 1. & WALTER, T. ( 1985). - Recovery of burnt Pinus ha/epensis Mill. forest. 1. Understorey and litter phytomass development after wildfire. Forest Eco/ogy andManagement, 12 : 269-277.
TRABAUD, L., MICHELS, C. & GROSMAN, J. ( 1985). - Recovery of burnt Pinus ha/epensis Mill. forest. Il. Pine reconstitution after wildfire. Forest Eco/ogy and Management, 1 3 : 1 67- 1 79.
TROUMBIS, A. & TRABAUD, L. ( 1 987). - Dynamique de la banque de graines de deux espèces de cistes dans les maquis grecs. Acta Oeco/ogica, Oeco/ogia Plan/arum, 8 : 1 67- 1 79.
TUTIN, T.G., HEYWOOD, V.H., BURGES, N.A., MOORE, D.M., VALENTINE, D.H., WALTERS, S.M. & WEBB, D.A. ( 1 964- 1980). - Flora Europea. Cambridge University Press.
230





























