Embed Size (px)
Citation preview

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
24/09/2013CHOUCHA Anis L2BMCPPr A.AUTILLO-TOUATI12 pages
INTRODUCTION: DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
1/12
Plan A. Présentation et programme de l'année
I. Présentation II. Programme de l'année
B. Des agents thérapeutiques ciblant les interactions protéines-protéines I. Introduction II. Diversité des interactions protéines-protéines III. Intérêt des inhibiteurs d'interaction Protéine-Protéine ('IA P-P)
C. Comment identifier les interactions protéine-protéine à l'échelle du protéome entier ? I. La technique du “double hybride” II. Le FRET
D. Interaction p53 avec Mdm2 : cible thérapeutique en oncologie. I. La protéine P53 et sa régulation
II. Comment restaurer la voie de p53 dans les cellules cancéreuses surexprimant Mdm2 ?

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
A. Présentation et programme de l'année
I. Présentation
1ersemestre• 20h de cours magistraux• 2 ECTS (Système Européen de Transfert et Cumul des crédits)• Contrôle des connaissances :
- 3 QR (question rédactionnelle) courtes de 10min chacune
- épreuve de 30min
- noté sur 20
- note éliminatoire si inférieure à 10
• ENT : plan et schémas des cours ou power point
2ème semestre
• 20h de cours magistraux
+ 6h de TP 1 ED où la présence est obligatoire
• 3 ECTS (Système Européen de Transfert et Cumul des crédits)
• Contrôle des connaissances :
- examen de fin d'année
- QR courtes (problème expérimental transversal)
- épreuve de 30min
- noté sur 14 + note des ED sur 6
• ENT- plan et schémas des cours ou power point
- problèmes résolus (annales, épreuves de 45min)
II. Programme de l'année
1er semestre
• Anomalies de structure et de fonction des constituants cellulaires et pathologies concernées
• Interaction et métabolisme - voie de signalisation - expression génique et pathologies
2ème semestre :
• Communication intercellulaire, signalisation et pathologie concernées
• Méthodes d'explorations fonctionnelles et imagerie 1 ED
2/12

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
B. Des agents thérapeutiques ciblant les interactions protéine-protéine
I. Introduction
Il existe environ 100 000 protéines dans l'organisme. Une protéine n'agit jamais seule, elle interagit avec d'autres protéines afin d'assurer sa fonction.
Interactome : Étude et recensement des interactions protéiques dans une cellule ou un organisme vivant. On étudie le champ d'interaction des protéines.
Exemple : la clathrine qui interagit avec diverses protéines associées à l'endocytoseCette étude des interactions protéiques débouchent sur de nouvelles stratégies thérapeutiques et notamment dans le traitement des cancers.
II. Diversité des interactions protéines-protéines1. Diversité structurale
L'interaction a lieu entre homo ou hétéro oligomères.
• Homo : chaînes identiques
• Hétéro : chaînes différentes
L'association est isologue ou hétérologue.
• Isologue : surfaces de contact sont identiques (homo oligomères)
• Hétérologues : surfaces de contact sont différentes (homo 1 hétéro oligomères)
Remarque : La surface de l'interaction (nombre d'acides aminés) est importante pour la stratégie thérapeutique.
2. Diversité fonctionnelle
L'interaction protéine-protéine (IPP) est obligatoire si :
• Les protéines ne sont pas stables indépendamment
• Les protéines ne sont pas fonctionnelles indépendamment
Ainsi, l'interaction est nécessaire à la stabilité et la fonction.
Exemple : gros complexe protéique (ADN polymérase, ARN polymérase, ribosomes...).
L'interaction protéine-protéine est non obligatoire si :
• Les protéines sont stables indépendamment
3/12

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
• Les protéines sont fonctionnelles indépendamment
L'interaction est responsable d'une action, d'une voie de signalisation.
Exemple : complexe antigène-anticorps, enzyme-inhibiteur, complexe de signalisation intracellulaire...
3. Diversité dynamique
L'interaction protéine-protéine permanente :• N'existe qu'au sein d'un complexe
• Les interactions protéine-protéine obligatoires sont généralement permanentes
L'interaction protéine-protéine transitoire :
• S'associe et se dissocie in vivo; mais il peut exister des associations fortes, qui requièrent un signal moléculaire pour altérer l’affinité mutuelle des composants du complexe.
Exemple : La PKA (protéine kinase AMPc dépendant) : la fixation de l’AMPc sur la sous-unité régulatrice (R) de la kinase rompt son interaction avec la sous-unité catalytique ce qui libère son domaine catalytique et lui permet de phosphoryler d'autres protéines.
Les interactions protéine-protéine non obligatoires peuvent être transitoires ou permanentes
Exemple : maladies neurodégénératives (Parkinson, Alzheimer...) caractérisées par une accumulation dans les neurones de complexes protéiques qui forment des agrégats. La stratégie thérapeutique consiste à les dissocier.
Comment fonctionnent les interfaces protéines-protéines au sein des complexes moléculaires ?Les surfaces d'interactions sont dans de nombreux cas larges (typiquement 1300 à 3000 Å)Sur le plan biochimique : compactes, hydrophobe et relativement planes. Point important : on note la présence d'acides aminés essentiels pour l'interaction (hot spot), (mis en évidence par cristallographie et mutagenèse dirigée)Les hot spots sont présents sur les 2 côtés de l'interface. Les régions sont hautement complémentaires et les mécanismes de reconnaissance sont extrêmement précis, avec des résidus hydrophobes d'une surface s'engageant précisément dans des "puits" présents sur la face opposée. La mutation de quelques AA suffit à empêcher la formation du complexe.
III. Intérêt des inhibiteurs d'interaction Protéine/Protéine ('IA P-P)
1) La très forte complémentarité des interfaces P-P permet de développer des inhibiteurs hautement spécifiques, évitant ainsi l'inhibition de cibles cellulaires non spécifiques.
2) Les complexes protéiques ciblés par ces inhibiteurs devraient être moins sujets à des mutations leur conférant une résistance contrairement aux protéines qui fixent des inhibiteurs au niveau de leur site actif. Ex : Inhibition de la fixation d'une cellule à la matrice en jouant sur la séquence RGD de l'intégrine (région qui constitue un site de fixation pour cette protéine)
4/12

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
3) De fait, une simple mutation au niveau d'une interface nécessite une mutation complémentaire sur l'autre interface pour restaurer l'interaction. Comme une double mutation simultanée n'est guère possible, il est hautement improbable que la protéine ciblée acquière une résistance à l'inhibiteur.
Donc efficacité théorique Puissante de ces inhibiteurs d'interaction P-P C. Comment identifier les interactions protéines-protéines à l'échelle du protéome entier ?
1. Mise en évidence d'interactions in vitro (non traité dans ce cours)
2. Mise en évidence d'interactions in vivo (2 techniques principales)
I La technique dite du “double hybride”: la modularité des facteurs de transcription, comme principe élémentaire du double hybride dans la levure.
Cette technique requiert l'utilisation de 2 domaines d'un facteur de transcription :
Domaine de fixation à l'ADN DF + Domaine d'activation de la transcription DA, capable d'activer la machinerie basale de la transcription.
Principe du test du double hybride :
• • On choisit une protéine appât (celle dont on veut identifier les interacteurs) qui va être fusionnée au domaine de fixation à l'ADN DF d'un facteur de transcription.• Les protéines proies (interacteurs potentiels) sont fusionnées au domaine d'activation de la transcription (DA) d'un facteur de transcription.• Les protéines fusions sont exprimées dans des cellules de levure contenant un gène rapporteur
5/12

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
dont l'expression est placée sous le contrôle du site de fixation pour le domaine de fixation à l'ADN (DF).
Remarque: On construit la protéine chimérique “appât + DF” par transfection avec la séquence nucléotidique de l'appât et celle de DF que l'on ajoute. Idem pour la protéine chimérique “proie + DA”
• Lorsque la protéine proie Y est capable d'interagir avec la protéine appât X, le domaine d'activation DA se retrouve à proximité du promoteur du gène rapporteur et la transcription a lieu.
Remarques: S'il n'y a pas d'interactions protéiques, il n'y a pas d'activation du gène rapporteur. Le test permet de visualiser l'activation de la transcription du gène dans les levures.
Quelles interactions sont identifiées par un crible double hybride ?
• Interactions permanentes
• Interactions transitoires dont les interactions enzyme-substrat (exemple : 10 à 40% des interactions Kinase-substrat). Ces interactions transitoires ne sont visibles qu'à l'expérimentation in vivo.
• Interactions qui n'existent pas physiologiquement → beaucoup de faux positifs car:
- L'appât auto activateur : il y a activation de la transcription en absence d’interacteur (pas besoin du domaine transcriptionnel pour induire la transcription du gène rapporteur)
- La proie collante: la proie interagit avec un très grand nombre d'appâts de façon non spécifique.
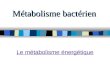
II Le FRET (Fluorescence Resonance Energy Transfer) Principe :
• La molécule « donneur » porte un fluorochrome dont la longueur d'onde d'émission correspond à la longueur d'onde d'excitation du fluorochrome diffèrent du premier placé sur la molécule « accepteur ». • La molécule « donneur » présente un domaine susceptible de se lier spécifiquement à un domaine de la molécule accepteur. Lorsque les conditions biologiques sont favorables, ces deux molécules interagissent, permettant le rapprochement des 2 fluorochromes. • Si les paramètres de distance (la distance maximale entre les deux fluorochromes est de 20Å) et d'orientation (géométrie) entre les 2 fluorochromes est correct, un transfert d'énergie pourra se produire et conduire à des modifications analysables de la fluorescence.
6/12
BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
Remarques : S'il n'y pas interaction entre la molécule « accepteur » et la molécule « donneur », il n'y a pas de transfert d'énergie.
èLe FRET a lieu in vivo sur des cellules de culture ou des tranches de tissus vivants ce qui en fait une méthode trés intéressante.
7/12

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
D. Interaction p53 avec Mdm2 : cible thérapeutique en oncologie.
I. La protéine P53 et sa régulation1. La protéine p53
La protéine p53 est la protéine régulatrice du cycle cellulaire, on l'appelle la « gardienne du génome ». Son rôle très important évite la transmission d'anomalies génétiques lors du cycle cellulaire. S'il y a une anomalie, p53 entraîne un blocage du cycle cellulaire. Elle détecte les anomalies génétiques et active la transcription des gènes impliquée dans la réparation de l’ADN.
• S'il y a réparation, le cycle continue.
• Si la réparation est impossible, il y a induction de l’apoptose par p53.
P53 participe à la senescence mitotique: vieillissement en relation avec le nombre de réplications cellulaires.
Dans les cancers, p53 est très fréquemment mutée (50% des cas), ce qui en fait une molécule attractive car elle peut induire l’apoptose. Elle n'assure plus sa fonction de « gardienne du génome » et entraîne la poursuite du cycle cellulaire même en cas d'anomalies génétiques détectées, il n'y a pas d'apoptose.
Pourquoi est-elle inactive ?
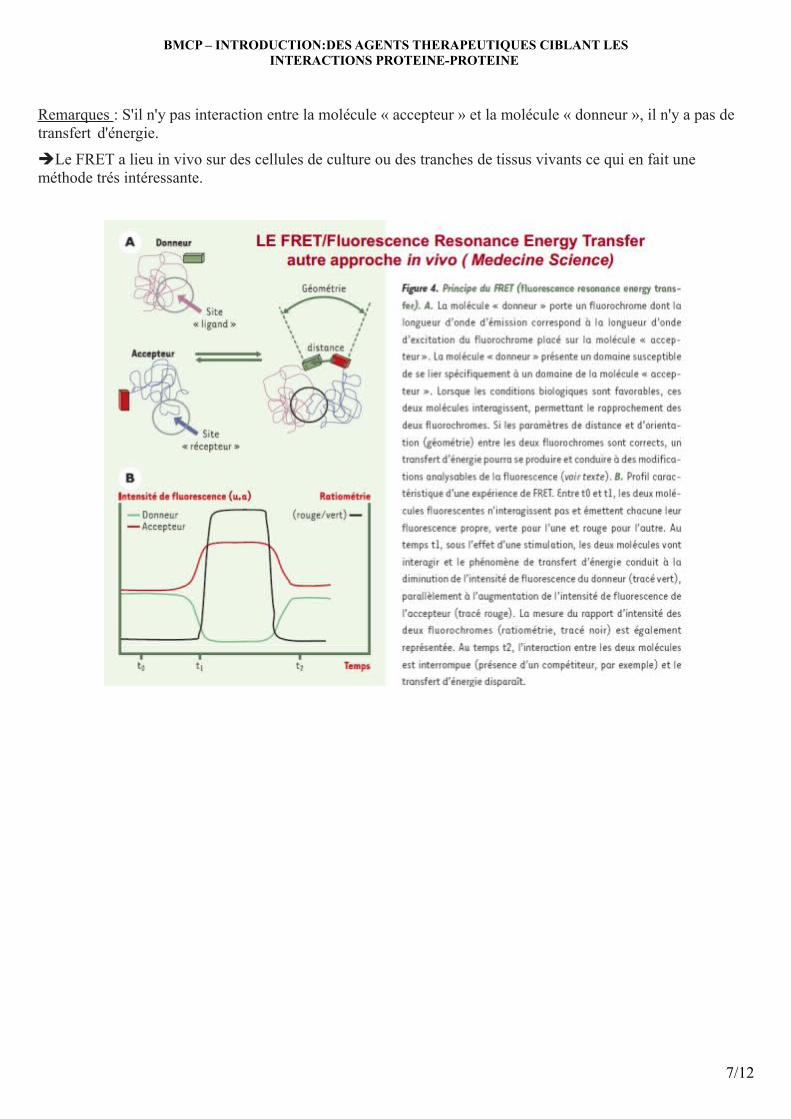
P53 interagit avec de façon préférentielle avec la protéine Mdm2 (Mouse double minutes) qui est un régulateur négatif qui réprime l'activité de la p53. Mdm2 est surexprimée dans de nombreux cancers avec une p53 sauvage ce qui implique une dégradation accrue de la p53.
Comment restaurer la voie de la p53 pour induire l’apoptose p53 dépendante dans les cellules cancéreuses surexprimant Mdm2 ?
Par de petits peptides ou de petites molécules chimiques qui bloquent l’IA p53-Mdm2.
2. Régulation de p53P53 est un facteur de transcription capable de réguler la transcription de Mdm2 et ce dernier est un régulateur négatif de p53 (c'est une boucle d'autorégulation). Si il y a stress de la cellule, alors il y synthèse de p53 qui va induire la transcription de Mdm2. Remarque : Mdm2 est une ubiquitine ligase.La protéine p53 est un FRT tout à fait banalequi peut être phosphorylé en Nter et Cter.
8/12

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
Dans les conditions normales, p53 est inactive. Elle existe sous une forme repliée: l'extrémité Cter se replie sur le site de fixation à l'ADN. Pour qu'elle soit active, il faut qu'elle se déplie pour que le domaine de fixation à l'ADN soit libéré. Elle passe à la forme active en réponse à un stress génotoxique. Le taux basal de p53 est tres faible, si il n’y a pas de stress géntoxique et sa durée de vie est très courte (20min) puis elle est dégradée par le protéasome.
L'activation implique la phosphorylation de la p53 sur son extrémité N Terminale (modification structurale) et la perte de l'interaction avec Mdm2 et tétramérisation de la p53. La phosphorylation de Mdm2 inhibe l'interaction p53-Mdm2, ce qui active la p53.
Note importante : bien connaître cette boucle de régulation : Mdm2 induit une ubiquitinylation de p53 suivie d'une dégradation de celle-ci dans le protéasome alors que réciproquement, la p53 induit la synthèse de Mdm2
L'activation de p53 entraîne son interaction avec l'ADN et l'arrêt du cycle cellulaire, la transcription des gènes de réparation de l'ADN, voir l'apoptose. La protéine p53 est instable, sa durée de vie est très courte (< 20min), ce qui la rend ainsi presque indétectable dans les conditions normales. Pour être dégradée par des protéases, p53 passe du noyau au cytoplasme.
Remarque : La protéine p53 a été détectée initialement dans les cancers.
3. Régulation de p53 par Mdm2
La protéine Mdm2 se fixe sur la partie NH2 terminale de p53 et induit sa dégradation.
9/12
BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
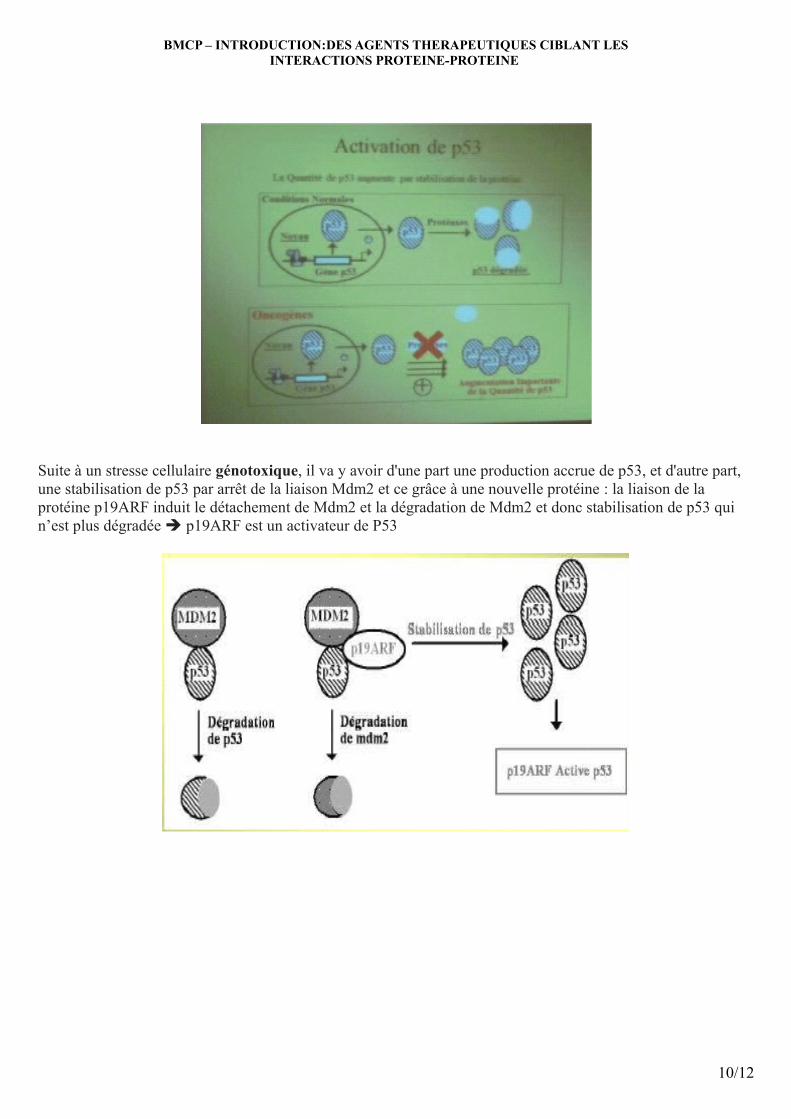
Suite à un stresse cellulaire génotoxique, il va y avoir d'une part une production accrue de p53, et d'autre part, une stabilisation de p53 par arrêt de la liaison Mdm2 et ce grâce à une nouvelle protéine : la liaison de la protéine p19ARF induit le détachement de Mdm2 et la dégradation de Mdm2 et donc stabilisation de p53 qui n’est plus dégradée è p19ARF est un activateur de P53
10/12

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
II Comment restaurer la voie de p53 dans les cellules cancéreuses sur exprimant Mdm2 ?
Les Nutlins sont des petites molécules antagonistes de la liaison p53 Mdm2.
NB: aujourd'hui en 2013, d'autres molécules que les nutlins qui peuvent bloquer ces interaction Mdm2/P53 ont été découvertes.Les Nutlins se lient de manière sélective dans le compartiment de liaison de p53 à la surface de Mdm2 (Mdm2 ne peut plus se fixer). Elles miment l'interaction des 3 résidus d'acides aminés critiques (Leu, Trp, Phe è Nom de ces 3 acides aminés pas à savoir) du peptide p53.
1. Effets des Nutlins sur les cellules tumorales p53 +/+ et surexprimant Mdm2 Cellules tumorales p53 +/+ : cellules cancéreuses ayant un p53 sauvage inactif car sur expression Mdm2
Propriétés des Nutlins :• traversent les membranes cellulaires• se lient à la cavité hydrophobique (un hot spot) dédiée à p53 située sur la molécule Mdm2 au niveau des 3 acides aminés particuliers évoqués plus haut• blocage de la liaison entre p53 et Mdm2• libération de p53 de son régulateur négatif Mdm2• activation de p53
Conséquences :
• stabilisation et accumulation de p53 au niveau du noyau
• activation des différents gènes cibles de p53 avec
- arrêt du cycle cellulaire en phase G1 et G2
- apoptose dépendante des caspases
- senescence des cellules tumorales
11/12

BMCP – INTRODUCTION:DES AGENTS THERAPEUTIQUES CIBLANT LESINTERACTIONS PROTEINE-PROTEINE
Test effectués :In vitro sur lignées de cellules cancereuses
In vivo : souris Nudes
2. La protection des cellules normales lors d'une chimiothérapie pour un cancer p53 -/- Les Nutlins pourraient protéger les cellules normales en prolifération durant une chimiothérapie antimitotique (qui agissent habituellement sur des cellules malades et saines à la fois) d'un cancer avec une p53-/-. Pas d’effets des Nutl ins sur les ce l lules tumorales .
Le pré traitement avec un Nutlin devrait activer la voie p53 et ainsi induire l'arrêt du cycle cellulaire en phase G1 et G2 dans les cellules normales qui ne passeraient pas en phase mitotique, ce qui diminuerait leur sensibilité aux agents chimio thérapeutiques antimitotiques.
A è les cellules normales ont une P53+/+ ce qui leur permet d’arrêter le cycle cellulaire. Ainsi, ces cellules seraient partiellement protégées de la cytotoxicité du paclitaxel (un inhibiteur mitotique fréquemment utilisé en chimiothérapie)B è les cellules mutantes pour P53 ne peuvent provoquer l’arrêt du cycle cellulaire, ce qui les rend insensibles […]
Le power point du cours et les différents schémas sont sur l'ENT
12/12