Embed Size (px)
Citation preview


Journal of Apicultural Science
Editorial Board
Wojciech SkowronekTeresa Szczêsna
Research Institute of Pomology and Floriculture, Apiculture Division,ul. Kazimierska 2, 24-100 Pu³awy, Poland.E-mail: [email protected]
Associate Board
Jerzy Woyke - breeding and geneticsJaros³aw Prabucki - biologyJerzy Wilde - technologyAndrzej Pidek - economicsZdzis³aw Gliñski - diseasesBoles³aw Jab³oñski - pollinationZdzis³aw Wilkaniec - pollination insectsHelena Rybak-Chmielewska - technology and chemistry of bee products
International Scientific Board
Kaspar Bienefeld, Hohe Neuendorf, GermanyStefan Fuchs, Oberursel, GermanyJan Kopernický, Liptovský Hrádok, SlovakiaNikolai Krivcov, Rybnoje, RussiaJurgis Racis, Akademija, LithuaniaBurkhard Schricker, Berlin, GermanyEnikõ Mátray Szalai, Gödöllõ, HungaryVladimir Veselý, Dol, Czech Rep.
Published byResearch Institute of Pomology and Floriculture, Apiculture Division,ul. Kazimierska 2, 24-100 Pu³awy, Poland, e-mail: [email protected]
Apicultural Research Association,ul. Kazimierska 2, 24-100 Pu³awy, Poland.

Journal of Apicultural Science is the continuation of former scientific journalPszczelnicze Zeszyty Naukowe published by the same organizations.
Under this name 44 volumes of the journal have been published.
Technical staffZygmunt Zydorowicz
Publication funded by the Committee for Scientific Research.Wydawnictwo pisma dofinansowywane przez Komitet Badañ Naukowych.

Salus Apis mellifera
2nd EUROPEAN SCIENTIFIC APICULTURAL
CONFERENCE
REPORTS AND PAPERS
Balatonlelle, HungarySeptember, 11-13 2002

Content
László Békési, Enikõ Mátray Szalai - Experiments on hygienic behaviourof honey bees . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
Janusz Bratkowski, Jerzy Wilde - Effect of changing of colony structureon trapped pollen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
Wit Chmielewski - Biological criteria for accepting of bee bread as a mitefood by Tyrolichus casei Oudemans (Acarina: Acaridae) . . . . . . . . . . . . . 21
Bo¿ena Chuda-Mickiewicz, Jaros³aw Prabucki, Jerzy Samborski - Onsetof oviposition in honeybee queens kept in boxes with non-free flyingbees . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
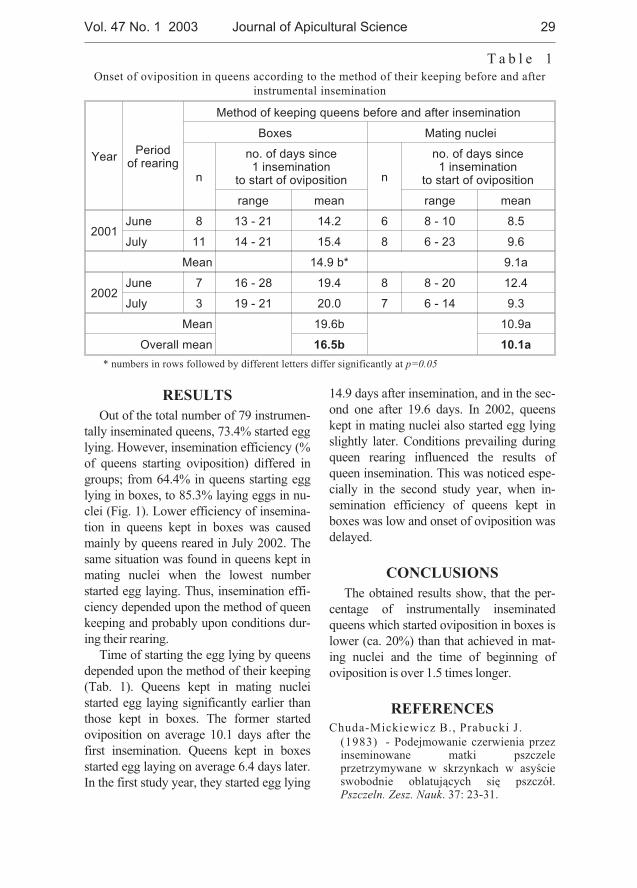
Jarmila Drobna, Vladimir Ptacek - Experience with using bumblebeesas pollinators in the regeneration of the genetic resources of someforage legumes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
Assegid Garedew, Erik Schmolz, Ingolf Lamprech - The antimicrobialactivity of honey of the stingless bee Trigona spp. . . . . . . . . . . . . . . . . . . . 37
József Nyéki, Tibor Szabó, Zoltán Szabó - Flowering phenologyand fertility of sour cherry (Prunus cerasus L.) cultivars selectedin Hungary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 51
Levente Nyárs - Situation and perspective of the Hungarian bee-keeping . . . . . . 59
Vladimir Ptacek - Deliberate production of young queens in Bombus terrestris L.(Hymenoptera, Apoidea) colonies reared in laboratory . . . . . . . . . . . . . . . 67
Miklós Sárospataki, Judit Novák, Viktória Molnár - Distribution and relativeabundance of bumble bees (Bombus and Psithyrus) in Hungary . . . . . . . . 73
REVIEW ARTICLES
Richard A. Baker, Wit Chmielewski - How old are bees?- A look at the fossil record . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 79
Pál Benedek - Bee pollination of fruit trees: recent advancesand research perspectives I. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87
Pál Benedek - Bee pollination of fruit trees: recent advancesand research perspectives II. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 95
Dorothea Kauhausen, Roland Keller - Methods of classificationof honeybee races using wing characters - a review . . . . . . . . . . . . . . . . . 103
Vladimir Ptacek - Renovation of J. G. Mendel’s apiary . . . . . . . . . . . . . . . . . . . 109
Vladimír Veselý - Programm of the fight against resistant Varroa mitesin Czech Republic . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 113
Vol. 47 No. 1 2003 Journal of Apicultural Science 3

Spis treœci
László Békési, Enikõ Mátray Szalai - Badania nad zachowaniemhigienicznym pszczó³ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
Janusz Bratkowski, Jerzy Wilde - Wp³yw zmiany struktury rodzinpszczelich na iloœæ odebranych obnó¿y py³kowych. . . . . . . . . . . . . . . . . . . 11
Wit Chmielewski - Biologiczne kryteria akceptacji pierzgi pszczelejjako pokarmu roztoczy przez Tyrolichus casei Oudemans(Acarina: Acaridae) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
Bo¿ena Chuda-Mickiewicz, Jaros³aw Prabucki, Jerzy Samborski -Rozpoczynanie czerwienia przez matki pszczele przetrzymywanew skrzynkach z pszczo³ami bez mo¿liwoœci oblatywania siê pszczó³ . . . . 27
Jarmila Drobna, Vladimir Ptacek - Doœwiadczenie z u¿yciem trzmielijako zapylaczy w odnawianiu zasobów genetycznych kilkustr¹czkowych roœlin pastewnych . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
Assegid Garedew, Erik Schmolz, Ingolf Lamprech - Aktywnoœæprzeciwdrobnoustrojowa miodu pszczó³ bez¿¹d³ych Trigona spp. . . . . . . . 37
József Nyéki, Tibor Szabó, Zoltán Szabó - Biologia kwitnienia i zapylaniewiœni (Prunus ceraus L.) odmian wyselekcjonowanych na Wêgrzech . . . . 51
Levente Nyárs - Stan obecny i perspektywy wêgierskiego pszczelarstwa . . . . . . 59
Vladimir Ptacek - Rozwa¿ania nad mo¿liwoœci¹ wychowu m³odych matektrzmiela ziemnego - Bombus terrestris L. (Hymenoptera, Apoidea)w rodzinach hodowanych w laboratorium . . . . . . . . . . . . . . . . . . . . . . . . . 67
Miklós Sárospataki, Judit Novák, Viktória Molnár - Wystêpowanietrzmieli (Bombus) i trzmielców (Psithyrus) na Wêgrzech . . . . . . . . . . . . . 73
ARTYKU£Y PRZEGL¥DOWE
Richard A. Baker, Wit Chmielewski - Jak stare s¹ pszczo³y?- spojrzenie na rejestr skamienia³oœci . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 79
Pál Benedek - Zapylanie drzew owocowych przez pszczo³y:ostatnie postêpy i perspektywy badañ I. . . . . . . . . . . . . . . . . . . . . . . . . . . . 87
Pál Benedek - Zapylanie drzew owocowych przez pszczo³y:ostatnie postêpy i perspektywy badañ II. . . . . . . . . . . . . . . . . . . . . . . . . . . 95
Dorothea Kauhausen, Roland Keller - Metody oceny przynale¿noœcirasowej pszczó³ za pomoc¹ cech morfologicznych skrzyde³ - przegl¹d . . 103
Vladimir Ptacek - Odtworzenie pasiek J. G. Mendla . . . . . . . . . . . . . . . . . . . . . 109
Vladimír Veselý - Program walki z odpornymi populacjami roztoczyVarroa w Czechach . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 113
4

EXPERIMENTS ON HYGIENIC BEHAVIOUROF HONEY BEES
L á s z l ó B é k é s i , E n i k õ M á t r a y S z a l a i 1
Szent István University, Faculty of Veterinary Science, Department of Parasitology and Zoology,H-1400 Budapest P.O.Box 2 Hungary
1Institute for Small Amimal Research, Dept. of Honey-bee Breeding and Biology,H-2101 Gödöllo, Méhészet, Hungary, e-mail [email protected]
S u m m a r y
It has been a long-debated question, if the colony is able to get rid of any agent of disease bythemselves, without the help from outside factors. This is the reason, why the researchers followwith keen attention the hereditary hygienic behaviour of immune-mechanism among the beecolonies. Many experts are of the opinion that this form of behaviour has an important role inthe defence against infectious and parasitic diseases.
We performed our experiments in the apiary of the Szent Istvan University, Gödöllo, (apiary1), in a private apiary (apiary 2), and in the apiary of the Institute for Small Animal Research(apiary 3).
The differences in hygienic behaviour between colonies of the three apiaries can be seen inTables 1-3. The average cleaning success was 74% in the first, and 72% in the second apiary,after 24 hours. In the third apiary, where the cleaning instinct as breeding trait was evaluated,the cleaning success was the highest, 86%, with 19.7% coefficient of variation. The results inthe first two apiaries are nearly the same, in the third apiary, due to the freezing method, weresignificantly lower in the same colonies than the result of the piercing method.
In these experiments the very close results of observation for 24 and 48 hours show that thetime factor has a little role in this behavioural pattern. In the case of the freezing method, thehygienic behaviour of colonies was nearly the same.
On the base of our experiments, we recommend the application of the pin piercing method fortesting the hygienic behaviour. The systematic testing would be advisable in colonies kept forbreeding and for queen rearing.
Keywords: hygienic behaviour, cleaning success, frosty method, pin piercing method.
INTRODUCTIONThe new scientific results and experi-
ences in the bee management have eluci-dated the pathological and epidemic reasonsof many honeybee diseases. In the earlierdays general view was that one part of theagent, causing illness, was connected to onedefinite territory, (supposedly had endemicorigin), but nowadays, due to the very in-tensive international traffic, much of thembecame wide spread. This perception hasarisen anxiety throughout the world, and re-sulted in serious consequences. Defence
against the infectious honeybee diseases isgoing on in many countries using treatmentwith medicines (chemotherapy), and inmany countries there are very strict officialprescriptions and blockades against the in-fected apiaries combined with colony exter-mination with official compensation.
This defence has resulted in a very dif-ferent outcome. A perfect solution for pre-venting the spreading or for defence againstthe bee diseases have not been developedup to now. The growing number of healthproblems due to high concentration of
Vol. 47 No. 1 2003 Journal of Apicultural Science 5

apiaries in Hungary, as well as differentmethodical troubles, arising from the differ-ent types of hives, make the Hungarian api-arists unquiet and disturbed about the honeyproduction in our country.
It has been a long-debated question if thecolony is able to get rid of any agent of dis-ease by themselves without the help fromoutside factors. This is the reason, why theresearchers follow with keen attention thehereditary hygienic behaviour of im-mune-mechanisms among the bee colonies.Many experts are of the opinion that thisform of behaviour has an important role inthe defence against the infectious and para-sitic diseases (Bailey and Ball 1991).
Kefuss et al. (1996) advice to apply apractical method for checking the hygienicbehaviour in the colonies. Their work stim-ulated our research team to test the applica-bility of this well known, but in Hungaryhardly tested method, in our apiaries.
Rothenbulher (1964) was the first whoproved that selection could be a tool inbreeding much more resistant colonies. Herecognised that the hygienic behaviour ischecked by two recessive alleles. By one al-lele the bee recognises the problem, and isable to open the sealed cell, the other allelecontrols the instinct of clearing away thedead larvae. Both alleles are necessary for ahygienic colony. In heterozygous popula-tions, the appearance of dominant alleles di-minishes the success of this character. Inthis case only one part of bees has hygienicinstinct, or follows one or the other behav-ioural form.
Harbo (1995) found significant connec-tion between hygienic behaviour of threeweeks old workers, and the pest.
Spivak (1996) infected larvae of hy-gienic and non-hygienic colonies by Varroamite and found differences only by heavierinfection. There were differences in thenumber of chewed or scratched larvae at thebottom of the hives. Among these the hy-gienic colonies. have an advantage.
MATERIALS AND METHODSWe performed our experiments in the
apiary of the Szent Istvan University,Gödöllo, (apiary 1), in a private apiary (api-ary 2), and in the apiary of the Institute forSmall Animal Research (apiary 3). In api-ary 1, and 2. 10 and 11 colonies were cho-sen by random in July, and from each ofthem one sealed comb was carved out con-taining larvae. In an area probably not con-taining empty cells, a space of 5 5 cm wasmarked out by help of a model. This frag-ment contained about 100 cells. By using asterile injection pin, in the marked fragmenteach cell was pierced in its centre to kill thelarvae, then the comb fragments weremarked and taken back to their originalplace. After 24 and 48 hours, the numbersof cells that were opened and cleaned,opened but not cleaned, and not openedwere counted.
Thirty pollen collecting bees were takenfrom each colony to be tested for Nosema.The bees were homogenised in a mortar,adding previously a little water to them.Then they were allowed to dry on a slideand were fixed above flame. Dyeing with0.4% methylene blue lasted 15 minutes, thecounter-dyeing with 0.6% fuchsine solutiononly a few seconds. The assessment wasbased on the number of spores in three lev-els of infection.
Testing of the third apiary was in Juneand August. This testing was different fromthe previous ones, because 5 5 cm larvaecontaining, sealed comb fragments werecarved out and frozen during 24 hours in arefrigerator. These fragments were subse-quently put and fixed into the covered, lar-vae containing combs of 10 colonies.(Kefuss et al. 1996) The experiment wasrepeated in the same colonies by piercingwith a pin.
Other data were also collected, e.g. ofthe number of honeycombs with larvae,type of hive, age of queens, signs of illness,etc.
6

For the comparison of the results,Staatgraf (Advanced Procedures) programanalysis was used.
RESULTSThe results of the experiments can be
seen in Table 1. This table also contains theresults of Nosema testing. For statisticalcomparison the term of cleaning successwas introduced, which was equal to 100 mi-nus the number of the cleaned cells,counted after 24 and 48 hours.
The application of the chi test failed toreveal differences between the first and thesecond apiary in the 24 hour test but a sig-nificant difference was between colonies(P<0.01) for cleaning success. In the thirdapiary a significant difference was found(P<0.05) between the results from the meth-ods piercing and freezing.
DISCUSSIONThe differences in the hygienic behav-
iour between colonies of the three apiariescan be seen in Tables 1-3. The averagecleaning success after 24 hours was 74% inthe first, and 72% in the second apiary. Inthe third apiary, where the cleaning instinctas breeding trait was evaluated, the cleaningsuccess was the highest, 86%, with 19.7%coefficient of variation. The results in thefirst two apiaries are nearly the same, in thethird apiary, results due to the freezingmethod were significantly lower in thesame colonies than the result of the piercingmethod. (P<0.05).
In these experiments the very close re-sults of the observation for 24 and 48 hoursshow that the time factor has a little role inthis behavioural pattern. Fewer hygieniccleaning bees can clean the dead cells butduring longer time, and for this reason weaccepted the 24 hours observation. Withinapiaries the treatments show significant dif-ferences, but the differences between apiar-ies did not reach the level of significance.
In the least hygienic two colonies of one
of the apiaries a very serious pest was dis-covered thus verifying the statement ofHarbo (1995). Nosema infection seemedstrong in apiaries, tested in the beginning ofsummer, but there seemed to be no connec-tion between the cleaning instinct and theinfection.
In the case of the frosty method, the hy-gienic behaviour of the colonies was nearlythe same. Application of this method ismore complicated, needs more time, andthe result does not give more information inspite of the fact that the bees should dis-cover the dead larvae in a covered not in-jured honeycomb. In this method thestrange smell of the little fragment of thetreated honeycomb could be disadvanta-geous, as could perhaps be the low temper-ature, as well as the arising vapour. All thismeans more work for the bees, because onboth sides of the honeycomb dead cells canbe found, the evaluation is difficult, cannotguarantee the same number of cells(100-100 cells) on both sides of the honey-comb fragment. At piercing a lot of larvaeare injured, which also means work for thecleaning bees.
The number of the injured cells could bediminished by cutting or piercing the hon-eycomb according to the shape of the cellse.g. by following the shape of rhombus.A freshly disinfected model containing 100pins can be prepared for piercing,. An im-portant point for the comparison is that thetest in an apiary should be performed on thesame date because the number of the youngbees changes in different periods of theyear. The place of testing should also be thesame, because the instinct of cleaning isstronger in the middle of the honeycombthan farther away from it.
Based on our experiments we recom-mend the application of the pin piercingmethod for testing the hygienic behaviour.The systematic testing would be advisablein colonies kept for breeding and for queenrearing.
Vol. 47 No. 1 2003 Journal of Apicultural Science 7
8
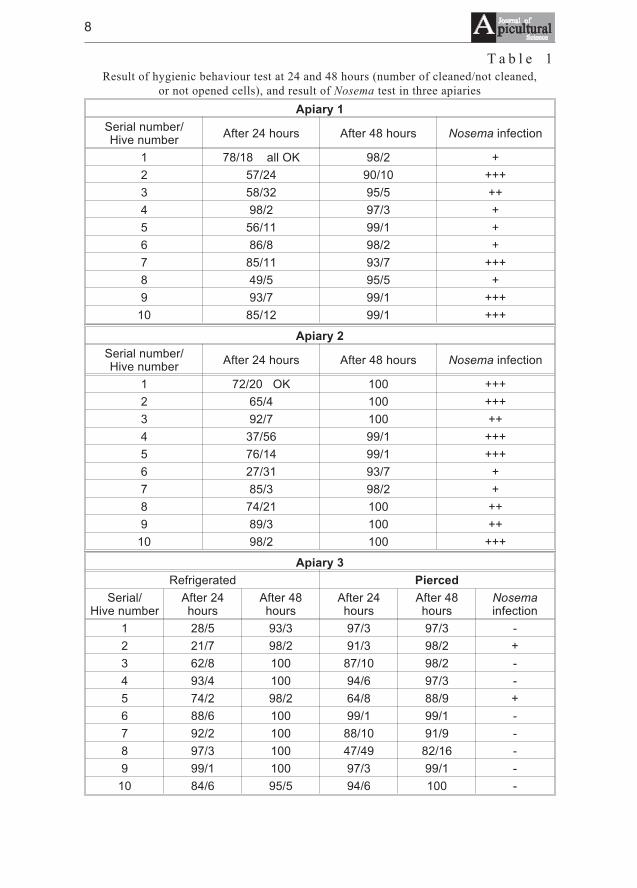
Apiary 1
Serial number/Hive number After 24 hours After 48 hours Nosema infection
1 78/18 all OK 98/2 +
2 57/24 90/10 +++
3 58/32 95/5 ++
4 98/2 97/3 +
5 56/11 99/1 +
6 86/8 98/2 +
7 85/11 93/7 +++
8 49/5 95/5 +
9 93/7 99/1 +++
10 85/12 99/1 +++
Apiary 2
Serial number/Hive number After 24 hours After 48 hours Nosema infection
1 72/20 OK 100 +++
2 65/4 100 +++
3 92/7 100 ++
4 37/56 99/1 +++
5 76/14 99/1 +++
6 27/31 93/7 +
7 85/3 98/2 +
8 74/21 100 ++
9 89/3 100 ++
10 98/2 100 +++
Apiary 3
Refrigerated Pierced
Serial/Hive number
After 24hours
After 48hours
After 24hours
After 48hours
Nosemainfection
1 28/5 93/3 97/3 97/3 -
2 21/7 98/2 91/3 98/2 +
3 62/8 100 87/10 98/2 -
4 93/4 100 94/6 97/3 -
5 74/2 98/2 64/8 88/9 +
6 88/6 100 99/1 99/1 -
7 92/2 100 88/10 91/9 -
8 97/3 100 47/49 82/16 -
9 99/1 100 97/3 99/1 -
10 84/6 95/5 94/6 100 -
T a b l e 1Result of hygienic behaviour test at 24 and 48 hours (number of cleaned/not cleaned,
or not opened cells), and result of Nosema test in three apiaries

ACKNOWLEDGEMENTThanks are extended to Dr. György
Csaba Head of Dept. for his useful advice,Dr. Arpad Tóth and György Patocska forallowing the experiment in his apiary.
Without the help and stimulation of ouralready deceased colleague, this study couldnot have come into existence.
REFERENCESBailey L. and Ball B. V. (1991) - Honey
bee pathology. Sidcup, Kent, UK, HarcourtBrace Jovanovich.
Harbo J.R. (1995) - Observations onhygienic behavior and resistance tochalkbrood. American Bee Journal,135(12) 828.
Kefuss J., Taber S. III, Vanpoucke J.and Rey F. (1996) - A practical methodto test for disease resistance in honey bees.American Bee Journal, 136(1) 31-32.
Rothenbuler, W (1964) - Behaviourgenetics of nest cleaning in honey bees.Americ. Zoologoist. 4: 113-123
Spivak, M. (1996) - Honey bee hygienicbehavior and defense against Varroajacobsoni. Apidologie, 27(4) 245-260.
Spivak, M. (1996) - Response of hygienichoney bees to Varroa jacobsoni mites.Resistant Pest Management, 8(1) 42-44.
BADANIA NAD ZACHOWANIEM HIGIENICZNYM PSZCZÓ£
B é k é s i L . , S z a l a i E . M .
S t r e s z c z e n i e
Od d³u¿szego czasu prowadzone s¹ rozwa¿ania czy rodzina pszczela jest w stanie przypomocy czynników zewnêtrznych chroniæ siê przed czynnikami chorobotwórczymi. To jestpowodem, dla którego badacze przywi¹zuj¹ wiele uwagi do dziedziczenia siê zachowaniahigienicznego i odpornoœci na choroby pszczó³. Wielu ekspertów uwa¿a, ¿e to zachowanie
Vol. 47 No. 1 2003 Journal of Apicultural Science 9
Fig. 2 Testing the hygienic behaviour of beecolonies, by the method of freezinga brood comb section. The piece ofcomb was fixed by wire. The beescleaned only small cells after 24 hours.In the majority of the cells they didnot recognize the dead larvae, or thecells had already been opened, but thedead larvae were not removed yet
Fig. 1 Testing the hygienic behaviour of beecolonies by the method of pin piercing.The result of testing after 24 hours: inthe middle area all cells are cleaned,a few pierced cells remained in theoff-centre parts which were not recog-nised by bees

spe³nia wa¿n¹ rolê w obronie przed infekcj¹ pszczó³ przez choroby paso¿ytnicze.Nasze badania wykonaliœmy w pasiece Uniwersytetu Szent Istvan w Gödöllo, (pasieka 1),
w prywatnej pasiece (pasieka 2) i w pasiece Instytutu Drobnego Inwentarza (pasieka 3).Ró¿nica w zachowaniu higienicznym miêdzy rodzinami trzech badanych pasiek (tabele 1-3).
W pasiece 3, w której zachowanie higieniczne jest przedmiotem selekcji, czêstotliwoœæoczyszczania komórek by³a najwy¿sza (86%). W dwóch pierwszych pasiekach wyniki by³yzbli¿one ale zawsze ni¿sze w przypadku czerwiu zabijanego przez zamra¿anie ni¿ przezprzek³uwanie. W doœwiadczeniach tych podobne wyniki uzyskano po 24 i 48 godzinachwskazuj¹, ¿e czynnik czasu ma niewielki wp³yw na wynik ostateczny.
Na podstawie naszych doœwiadczeñ polecamy w badaniach behavioru higienicznegozabijanie czerwiu poprzez przek³ucie szpilk¹. Ten sposób powinien byæ stosowany w pasiekachprowadz¹cych selekcjê pszczó³.
S³owa kluczowe: zachowanie higieniczne, usuwanie czerwiu, zamra¿anie, przek³uwanieszpilk¹.
10

EFFECT OF CHANGING OF COLONY STRUCTUREON TRAPPED POLLEN
J a n u s z B r a t k o w s k i , J e r z y W i l d e
Apiculture Division, Warmia and Mazury University, S³oneczna 48, 10-710 Olsztyn, Poland,e-mail: [email protected]
S u m m a r yThe goal of the experiment was to establish the effect of the changing of the structure of bee
colonies on the amount of trapped pollen. Colonies were divided into 5 groups: the group PTC(pollen trapping - control), the group PTF (pollen trapping and transferring colonies onto combfoundation), the group PTN (pollen trapping and nuclei building from brood and bees), thegroup PTI (pollen trapping and isolation of queens). The total production was established thatconsisted of harvested honey, trapped pollen and calculated honey (trapped pollen, wax, beesand brood taken from colonies expressed as kgs of honey).
Pollen trapping method applied in the group PTN (8.5 kg) was the most efficient during theexperiment. The average amount of collected pollen was the largest by a highly dignificantdegree in comparison with the other groups: PTC (3.6 kg), PTI (5.2 kg) and PTF (5.7 kg).
The highest honey yield was harvested from the control colonies (PTC - 16.3 kg) and thegroup transferred onto comb foundation (PTF - 14.4 kg), while the lowest one from the colonieswhere nuclei were created (PTN - 7.9 kg) and those differences were confirmed statistically(p=0.01).
Regulation of colony structure through taking brood and bees out in order to create nuclei(group PTN) as well as caging queens (group PTI) decreased significantly honey production incomparison with the control colonies (group PTC).
It can be said that the lower honey yield and the significant increase of pollen production incolonies of the group PTN were the result of the applied technology of regulation of colonystructure.
Keywords: honey, pollen, pollen trapping, total production, brood, bees, queens.
INTRODUCTIONPollen has been used for feeding honey-
bee colonies (Bobrzecki et al. 1994),bumblebees (Maciejewska, Wilkaniec1998) and its therapeutic effect on the hu-man body has been demonstrated as well(Dro¿d¿, Gwizdek 1986, Stojko et al.1997). The great interest in therapeuticproperties (Taber 1984) of pollen as wellas its usefulness for feeding honeybee colo-nies (Bobrzecki et al. 1994) recommendthe aplication of pollen trapping in the api-aries. It has been proved, that this kind ofbee management increases the gross in-come from an apiary (Cichoñ, Wilde2002, Marcinkowski 1994, Nelson et al.
1987, Wilde, Cichoñ 1997).Flow conditions, strength and condition
of colony as well as applied pollen trappingtechnology are the main factors that decideabout the amount of trapped pollen(Bobrzecki, Wilde 1991, Bratkowskiet al. 1999, Bratkowski, Wilde 2002,Duff, Furgala 1986, Nelson et al. 1987).In foreign cuntries with a longer growingseason and richer pollen flows ca 13 kg ofpollen loads per one colony are obtained(Nelson et al. 1987), while under Polishclimatic and weather conditions it is sup-posed to be about 2 kg (Bratkowski,Wilde 1998).
The installation of a pollen trap is the
Vol. 47 No. 1 2003 Journal of Apicultural Science 11

simplest pollen trapping method. A10-day-cycle of pollen trapping installationhas been recommended in order to avoidmaking pollen trapping difficult to bees:pollen is trapped for 5 days and after thatperiod it should be stopped for the next 5days (Polišèuk 1984). Duff and Furgala(1986) also applied a 14-day-cycle but theyobtained a lower pollen production (from1.4 to 2.9 kg) in comparison with traps ap-plication lasting the whole season. Betterresults in the amount of trapped pollen havebeen obtained by interference in the struc-ture of the honeybee colony (Wilde et al.1994a).
The goal of the experiment was to estab-lish the effect of the changing of the struc-ture of bee colonies on the amount oftrapped pollen.
MATERIALS AND METHODSThe experiment was carried out in
1997-1999. Colonies of Apis melliferacarnica Pollm. were used in the experimentin 1997, 1998 and 1999, respectively: 48,43 and 49, and were divided into 5 experi-mental groups. In the group PTC (pollentrapping, control) traditional bee manage-ment method were applied and pollen wastrapped. In the group PTF (pollen trapping,foundation) all combs were replaced byframes with comb foundation as soon as asthe eggs appeared in queen-cell cups.Honey and brood combs were given to thecolonies of the same group that did notshow symptoms of the swarm mood. In thegroup PTN (pollen trapping, nuclei) a partof combs with capped brood and some beescrowding its surface were used to make thenuclei. The nuclei were delivered with egglaying queens and pollen traps were in-stalled. Their trapped pollen was includedinto productivity of the colonies from whichthey were built (Wilde et al. 1994b). In thegroup PTI (pollen trapping, isolation)queens were put into the Zander cages for 2
weeks as soon as they started laying eggsinto queen cell cups. The queens got easy intouch with bees because of a peace of queenexcluder installed on the cage sides.
Productivity of bee coloniesTop pollen traps were used in the experi-
ments (Bobrzecki et al. 1989), that were in-stalled on the bottom of the hives (Wilde,Bobrzecki 1989). Their functional partswere provided by plastic bars with roundholes 5 mm in diameter. Pollen traps wereinstalled at the hive bottom at the beginningof winter rape blooming, and taken off inthe end of July on the following respectivedates over the study years: 10th May and30th July in 1997, 17th May and 27th July in1998, and 20th May and 27th July in 1999.The weight of pollen loads was determinedindividually for every single colony in theeach group.
The amount of harvested honey wasmeasured twice in each year for two mainhoney flow periods, after blooming of win-ter rape and of buckwheat (Bornus et al.1974, Wilde, Siuda 1996).
Converted production consisted of addi-tional bee products expressed as kgs ofhoney which was evaluated as honey valueunits as follows: 1kg of bees = 2.5 kg ofhoney, 10 dm2 of sealed brood = 4 kghoney, 1 kg of wax = 3.5 kg of honey(Bornus 1973, Bornus et al. 1974), 1 kgof pollen loads = 4 kg of honey (Wilde,Siuda 1996). The total production was ex-pressed as kgs of honey and it consisted ofcentrifuged honey and converted produc-tion (Bornus et al. 1974, Wilde, Siuda1996).
Average mean (�) was the main numeri-cal value for analized characters. The col-lected data were evaluated statisticallyusing the computer programmeSTATISTICA. The estimation of signifi-cance of differences was made by varianceanalysis and the least significant differences(LSD) method.
12

RESULTSInfluence of method of bee colony regula-tion on pollen production
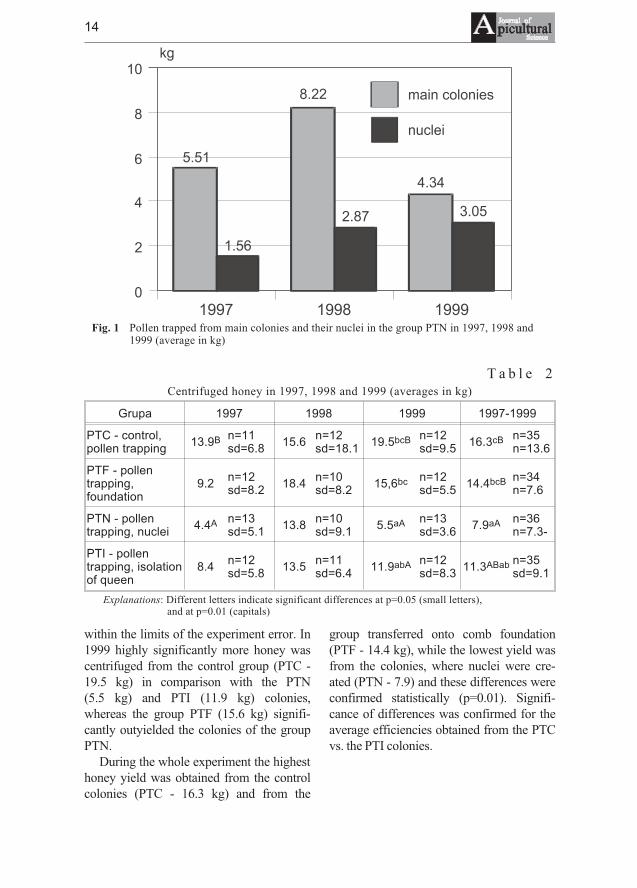
In 1997 the amount of pollen loads, con-cerning pollen trapped from nuclei, trappedin the group, where nuclei were created(PTN - 7.1 kg) was high significantly morein comparison with the control group (PTC- 3.0 kg - tab. 1), and significantly higherthan amount of pollen collected from thegroup, in which queens were kept in cages(PTI - 3.6 kg). In 1998 the biggest amountof pollen loads was trapped again from thecolonies of the groups PTN and PTI, 11.1and 7.6 kg, respectively. The differences forthe amount of trapped pollen in the groupwith nuclei vs. the control group (PTC - 3.5kg) and vs. the colonies transferred ontocomb foundation (PTF - 6.2 kg) werehighly significant and they were significantcompared with the group where queenswere kept in cages (PTI - 7.6 kg). As in theprevious years, in the season of 1999 thelargest pollen yield was obtained from thenuclei group (PTN - 7.4 kg). This value wassignificantly higher compared with thegroups where traditional bee management(PTC - 4.3 kg) was applied and queens
were caged (PTI - 4.4 kg).In the whole period of the experiment
the creating of nuclei (PTN - 8.5 kg) wasthe most efficient pollen trapping method.The average amount of collected pollenwas, to a highly significant degree, the larg-est of all the other groups. The pollen pro-duction of the rest of the colonies was verysimilar, and statistical differences appearedonly between the colonies transfred ontocomb foundation (PTF - 5.7 kg) and thecontrol colonies (PTC - 3.6 kg).
In 1997, 1998 and 1999 nuclei were cre-ated in the group of PTN: 10, 11 and 18, re-spectively. In 1997, the average productionobtained from one new colony was only 1.6kg, whereas in the last year it increased toan average of 3.1 kg of pollen loads (fig. 1).
Influence of bee colony structure regula-tion on honey yield
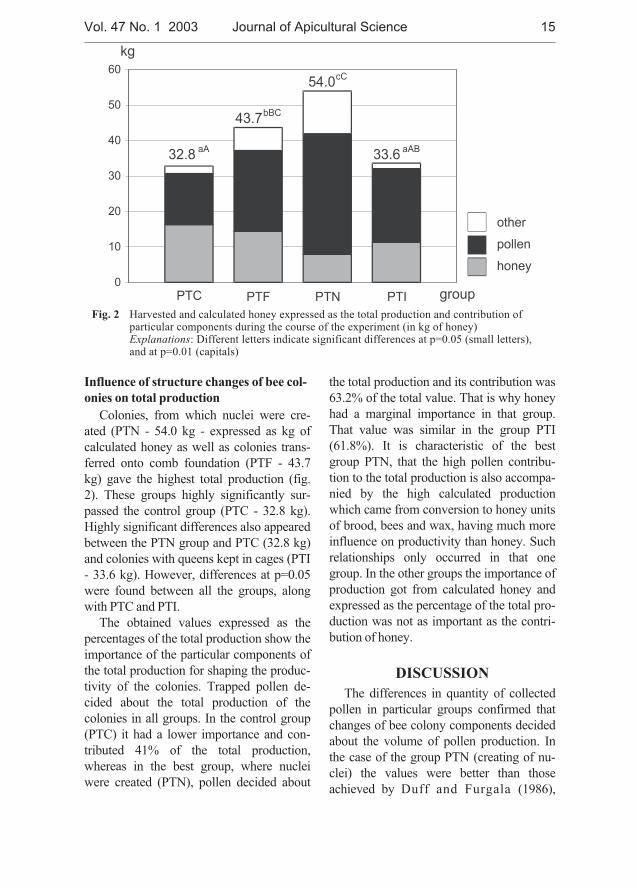
In 1997, 13.9 kg of honey was harvestedfrom control colonies, which was highlysignificantly more than from the colonies inwhich which nuclei were created (PTN -4.4 kg - tab. 2). In 1998, the total honeyyield ranged from 13.5 kg (group PTI) to18.4 kg (group PTF) in the season, but thedifferences between all the groups came
Vol. 47 No. 1 2003 Journal of Apicultural Science 13
Grupa 1997 1998 1999 1997-1999
PTC - control,pollen trapping 3.0aA n=11
d=1.7 3.5aA n=12sd=1.5 4.3aA n=12
sd=2.4 3.6aA n=35sd=2.1
PTF - pollentrapping, foundation 5.4 n=12
sd=3.9 6.2abA n=10sd=3.0 5.4a n=12
sd=2.1 5.7bA n=34sd=3.4
PTN - pollentrapping, nuclei 7.1bB* n=13
sd=4.1 11.1cB* n=10sd=5.8 7.4bB* n=13
sd=3.1 8.5bB n=36sd=5.1
PTI - pollentrapping, isolationof queen
3.6a n=12sd=1.2 7.6b n=11
sd=4.0 4.4aAn=12
sd=1.65.2abA n=35
sd=3.2
* - including pollen trapped from nuclei,Explanations: Different letters indicate significant differences at p=0.05 (small letters),
and at p=0.01 (capitals)
T a b l e 1Pollen trapped in 1997, 1998 and 1999 as well as in the course of the experiment
(averages in kg)
within the limits of the experiment error. In1999 highly significantly more honey wascentrifuged from the control group (PTC -19.5 kg) in comparison with the PTN(5.5 kg) and PTI (11.9 kg) colonies,whereas the group PTF (15.6 kg) signifi-cantly outyielded the colonies of the groupPTN.
During the whole experiment the highesthoney yield was obtained from the controlcolonies (PTC - 16.3 kg) and from the
group transferred onto comb foundation(PTF - 14.4 kg), while the lowest yield wasfrom the colonies, where nuclei were cre-ated (PTN - 7.9) and these differences wereconfirmed statistically (p=0.01). Signifi-cance of differences was confirmed for theaverage efficiencies obtained from the PTCvs. the PTI colonies.
14
Fig. 1 Pollen trapped from main colonies and their nuclei in the group PTN in 1997, 1998 and1999 (average in kg)
Grupa 1997 1998 1999 1997-1999
PTC - control,pollen trapping 13.9B n=11
sd=6.8 15.6 n=12sd=18.1 19.5bcB n=12
sd=9.5 16.3cB n=35n=13.6
PTF - pollentrapping,foundation
9.2 n=12sd=8.2 18.4 n=10
sd=8.2 15,6bc n=12sd=5.5 14.4bcB n=34
n=7.6
PTN - pollentrapping, nuclei 4.4A n=13
sd=5.1 13.8 n=10sd=9.1 5.5aA n=13
sd=3.6 7.9aA n=36n=7.3-
PTI - pollentrapping, isolationof queen
8.4 n=12sd=5.8 13.5 n=11
sd=6.4 11.9abA n=12sd=8.3 11.3ABab n=35
sd=9.1
Explanations: Different letters indicate significant differences at p=0.05 (small letters),and at p=0.01 (capitals)
T a b l e 2Centrifuged honey in 1997, 1998 and 1999 (averages in kg)
Influence of structure changes of bee col-onies on total production
Colonies, from which nuclei were cre-ated (PTN - 54.0 kg - expressed as kg ofcalculated honey as well as colonies trans-ferred onto comb foundation (PTF - 43.7kg) gave the highest total production (fig.2). These groups highly significantly sur-passed the control group (PTC - 32.8 kg).Highly significant differences also appearedbetween the PTN group and PTC (32.8 kg)and colonies with queens kept in cages (PTI- 33.6 kg). However, differences at p=0.05were found between all the groups, alongwith PTC and PTI.
The obtained values expressed as thepercentages of the total production show theimportance of the particular components ofthe total production for shaping the produc-tivity of the colonies. Trapped pollen de-cided about the total production of thecolonies in all groups. In the control group(PTC) it had a lower importance and con-tributed 41% of the total production,whereas in the best group, where nucleiwere created (PTN), pollen decided about
the total production and its contribution was63.2% of the total value. That is why honeyhad a marginal importance in that group.That value was similar in the group PTI(61.8%). It is characteristic of the bestgroup PTN, that the high pollen contribu-tion to the total production is also accompa-nied by the high calculated productionwhich came from conversion to honey unitsof brood, bees and wax, having much moreinfluence on productivity than honey. Suchrelationships only occurred in that onegroup. In the other groups the importance ofproduction got from calculated honey andexpressed as the percentage of the total pro-duction was not as important as the contri-bution of honey.
DISCUSSIONThe differences in quantity of collected
pollen in particular groups confirmed thatchanges of bee colony components decidedabout the volume of pollen production. Inthe case of the group PTN (creating of nu-clei) the values were better than thoseachieved by Duff and Furgala (1986),
Vol. 47 No. 1 2003 Journal of Apicultural Science 15
Fig. 2 Harvested and calculated honey expressed as the total production and contribution ofparticular components during the course of the experiment (in kg of honey)Explanations: Different letters indicate significant differences at p=0.05 (small letters),and at p=0.01 (capitals)

the latter investigators having trapped 9.6kg of pollen loads on average. However,Nelson et al. (1987) collected an averageof 13 kg of pollen for 3 months, using, likethe previous authors, a method based onlyon the installation of traps. Such a methodapplied in this study (pollen trapping with-out additional operations on bee colony -group PTC) did not allow the obtaining ofcomparable efficient results. However, thechances are that pollen production will beincreased through making smoller the di-ameter of pollen bars wholes. Wilde et al.(1994a) produced on average 8.1 kg of pol-len loads without changing the structure ofbee colonies, using plate with meshes 4.8mm in diameter. Bieñkowska andPohorecka (1996) proved, that the widerdiameter of the holes in the plate from 5.0mm to 4.8 mm caused the 50% increase inpollen trapped. Taking into considerationthat relationship, our result obtained in thegroup PTC (control) agrees with dates ofWilde et al. (1994a). However, pollen trap-ping and traditional management of beecolonies was less efficient as evaluatedagainst the other methods, but it was statis-tically confirmed in comparison with thegroups PTN (p=0.01) and PTF (p=0.05).
In 1998, in the group made up of nuclei(group PTN) 11.1 kg of pollen loads wastrapped in the authors’ experiment whileWilde et al. (1994a) obtained with thatmethod only 3.9 kg of pollen. However,Wilde and Bratkowski (1996) increasedthe efficiency of that method to 7.4 kg,which is a value close to the average for thewhole experiment of this study (PTN -8.5 kg). Wilde et al. (1994b) stressed theinfluence of new nuclei on the productionobtained, the amount of pollen trapped be-ing similar to that in the colonies in whichthe nuclei were built. Such results can beexplained by stimulation of egg laying bythe queens and by the positive correlationbetween pollen collection by bees andreared brood (Crailsheim et al. 1992,
1996, Fewell, Bertram 1999), which wasespecially manifest in the last year ofinvestigations.
The relationship between trapped pollenand honey yield showed that the applyingof colony structure regulation methods mayhave caused a decrease in pollen productionespecially under unfavourable climatic andflow conditions. In 1999 the results con-firmed such a finding and that the year wasconsidered as cold and rainy. In the groupsPTF, PTN and PTI not more that 1kg ofpollen on average was trapped in the secondpart of season, while in the group PTC(control) that production was 2.7 kg. It wasespecially interesting that colonies trans-ferred onto comb foundation (group PTF)responded like this in each year which sug-gested a very negative influence of thismethod on pollen collection. Comparingcollection of honey and pollen in particularparts of the season it may be also noticedthat high rate of pollen trapping was con-nected with a low honey yield, while lowpollen production was associated with ahigh honey yield, which is consistent withresults obtained by Polišèuk (1984).
On the basis of honey yield in investi-gated colonies it was found that appliedtechnologies of pollen trapping decreasedhoney output which is consistent with thefindings by Wild et al. (1994a). Silmilar re-lationship for pollen production vs. honeyyield was found by Wilde andBratkowski (1996). Alongside the groupwith nuclei, the group in which the queenswere kept in cages also showed lowerhoney production in comparison with thegroup from which pollen was trapped andtraditional bee manegement methods wereapplied, the relevant differences being sta-tistically valid. Nelson et al. (1987) foundonly once in the course of 3 years a nega-tive effect of pollen trapping on the honeyyield. Wilde et al. (1994a) explained thedecrease in honey production observed inthe group, in which nuclei were made, by
16

the weakening of the colonies rather than bypollen trapping only. In our experiment sig-nificantly lower amounts of honey werecentrifuged from the colonies from whichbrood and bees were taken out comparedwith honey yielded by the group PTC,which confirms a negative effect of remov-ing brood and bees on honey yield. Thus,when trapping plenty of pollen and usingnuclei we have to take into consideration asignificant decrease in honey production.A increase from 9.5 kg (1997) to 14 kg(1999) in honey centrifuged was found inthe group PTN in comparison with thegroup PTC (control), which is equivalent tothe increase in honey yield, 0.74 and 1.89kg, respectively, for every kg of trappedpollen. Caging queens (group PTI) alsocaused significant decrease in honey yield(p=0.05) in comparison with the group PTC(control), but the amount of trapped pollenwas at one time the smallest during the ex-periment. The observed relationships be-tween pollen and honey production in thosetwo last groups in comparision with thecontrol group allows the conclusion thatsuch kind of regulation of colony structuredecides about the efficiency of pollentrapping.
Pollen trapping increased to a large ex-tent the total production in the group PTN -63.2%, despite a significant decrease incentrifuged honey, compared with that inthe control group. Our results were superiorto those obtained by Pidek (1988), whonoticed a decrease of only about 15.4%,Nelson et al. (1987), in whose study thedecrease in output was about 26% andPolišèuk (1984), who got production moreabout 33%. It should be said, that they didnot regulate structure of colonies, fromwhich pollen was trapped, but they com-pared their productivity with colonies with-out pollen traps.
Biological conditions of bee colonieswere very important and decided about dif-ferent reaction of bees to applied methods
(Page, Fondrk 1995). A permanentswarm mood was a very negative symptomin the group of PTF, which was observed ineach colony transferred onto comb founda-tion. We could suppose that the reason forthis phenomenon was removing of brood,which changed the biological relationshipsin colonies (Pankiw et al. 1998, Seeley1989) and disturbed pheromons interactionbetween brood, queen and bees (Fewell,Bertram 1999, Hrassnigg, Crailsheim1998, Winston et al. 1991). Pettis et al.(1997) demonstrated that brood phero-mones and of queen in relation withqueen’s pheromones decreased about 50%the number of queen cells. Among colonieswith caged queens (group PTI) bees madequen cells after queens were placed intocages. Such behaviour was observed byMuszyñska (1987). We may draw theconclusion, that queen cells were the resultof the lack of a spread of the queen sub-stance (Seeley 1989). Creating of nucleiwas the only guarantee to eliminate theswarm mood, because in the group PTNthere were no symptoms of swarming inany of the years (Wilde, Bratkowski1996). Such behaviour caused a necessityfor an additional inspection of bee coloniesand for more frequent visits to the apiary towhich increased costs.
CONCLUSIONSRegulation of colony structure thrugh
taking brood and bees out in order to createnuclei (group PTN) as well as cagingqueens (group PTI) decreased significantlyhoney production in comparison with thatin the control colonies (group PTC). Lowerhoney yield and significant increase of pol-len production in colonies of the group PTNwere the result of the applied technology ofregulation of colony structure.
Taking brood and bees out in order tocreate nuclei and pollen trapping in thisgroup was the most efficient method of the
Vol. 47 No. 1 2003 Journal of Apicultural Science 17

intensification of the production. Results ofpollen production obtained from nuclei aswell as elimination of swarm mood in colo-nies influenced the effectiveness of thatmethod.
Pollen trapping in colonies transferredonto comb foundation (group PTF) and incolonies with caged queens (group PTI)have the limited practical application be-cause of a high costs of labour resultingfrom additional colony inspections to de-velop removed brood, liquidation of swarmmood, destroying of queen cells and look-ing for the queen.
REFERENCESBieñkowska M., Pohorecka K. (1996) -
Efekty pozyskiwania py³ku w zale¿noœci odwielkoœci otworów we wk³adce str¹caj¹cejobnó¿a py³kowe. Pszczeln. Zesz. Nauk., 40:95-101.
Bobrzecki J., £utowicz Z., Wilde J.(1989) - Udoskonalony powa³kowypo³awiacz py³ku. Pszczelarstwo, 40 (5):16-18.
Bobrzecki J., Wilde J. (1991) - Wp³ywpozyskiwania obnó¿y trzema rodzajamipo³awiaczy py³ku na rozwój iprodukcyjnoœæ rodzin. Pszczeln. Zesz.Nauk., 35: 19-27.
Bobrzecki J., Wilde J., Krukowski R.(1994) - Wp³yw podkarmianiapobudzaj¹cego py³kiem kwiatowym narozwój i produkcyjnoœæ rodzin. Acta Acad.Agricult. Techn. Olst., Zootechnica, 39:193-203.
Bornus L. (1973) - Ocena wartoœciu¿ytkowej selekcjonowanych pszczó³.Instytut Sadownictwa i Kwiaciarstwa,Pu³awy. Instrukcja: 1-6.
Bornus L., Bobrzecki J., Bojarczuk Cz.,Gromisz M., Kalinowski J., Król A.,Nowakowski J., Ostrowska W.,WoŸnica J., Zaremba J. (1974) -Badania nad miêdzyrasowymi mieszañcamipszczo³y miodnej. I. Przydatnoœæ u¿ytkowamieszañców miêdzyrasowych pszczo³ymiodnej w warunkach przyrodniczychPolski. Pszczeln. Zesz. Nauk., 18: 1-51.
Bratkowski J., Siuda M., Wilde J.(1999) - Plantacje gryki jako po¿ytekpy³kowy i nektarowy w gospodarcepasiecznej. Biul. Nauk., 4: 107-110.
Bratkowski J., Wilde J. (1998) -Charakterystyka pszczelarzy producentówmiodu i py³ku. V KonferencjaNaukowo-Promocyjna: Lepsza ¿ywnoœæ.26-28.06. Olsztyn ART: 112-115.
Bratkowski J., Wilde J. (2002) -Technologie zwiêkszaj¹ce pozyskiwanieobnó¿y py³kowych i poprawiaj¹cedochodowoœæ pasiek. Biul. Nauk., 18 (5):113-122.
Cichoñ J., Wilde J. (2002) -Pozyskiwanie obnó¿y py³kowych jakosposób na poprawienie op³acalnoœciprodukcji pasiecznej. Biul. Nauk., 18 (5):144-150.
Crailshaim K., Schneider L.H.W.,Hrassnigg N., Bühlmann G., BroschU., Gmeinbauer R., Schöffmann B.(1992) - Pollen consumption andutilization in worker honeybees (Apismellifera carnica): dependence onindividual age and function. J. InsectPhysiol. 38 (6): 409-419.
Dro¿d¿ E., Gwizdek E. (1986) - Py³ekkwiatowy jako od¿ywka regeneracyjna. VMiêdzynarodowe Sympozjum ApiterapiiApimondia. Kraków: 23-26.
Duff S.R., Furgala B. (1986) - Pollentrapping honey bee colonies in Minnesota.Part II: Effect on foraging activity, honeyproduction, honey moisture content andnitrogen content of adult workers. Am. BeeJ., 126 (11): 755-758.
Fewell J.H., Bertram S.M. (1999) -Division of labor in a dynami environment:response by honeybees (Apis mellifera) tograded changes in colony pollen store.Behav. Ecol. Sociobiol., 46: 171-179.
Hrassnigg N., Crailsheim K. (1998) -The influence of brood on the pollenconsumption of worker bees (Apis melliferaL.). J. Insect Physiol., 44: 393-404.
Maciejewska M., Wilkaniec Z. (1998) -Gaining bumble bee families by a modifiedlaboratory method. Pszczeln. Zesz. Nauk.,42 (1): 313-320.
18

Marcinkowski J. (1994) - Mo¿liwoœcipoprawy efektywnoœci prowadzenia pasiekiprzez wzbogacenie wyboru produktówpasiecznych. Pszczeln. Zesz. Nauk., 38:91-99.
Muszyñska J. (1987) - The relationsshipbetween the biological structure ofhoneybee colonies and their winteringsuccess. Pszczeln. Zesz. Nauk., 31: 41-53.
Nelson D.L., Mckenna D., Zumwalt E.(1987) - The effect of continous pollentraping on sealed brood, honey productionand gross income in Northern Alberta. Am.Bee J., 127 (8): 648-650.
Page R.E., Fondrk M.K. (1995) - Theeffect of colony-level selection on thesocial organization of honey bee (Apismellifera L.) colonies: colony-levelcomponents of pollen hoarding. Behav.Ecol. Sociobiol. 36: 135-144.
Pankiw T., Page R.E., Fondrk M.K.(1998) - Brood pheromone stimulatespollen foraging in honey bees (Apismellifera L.), Behav. Ecol. Sociobiol. 44:193-198.
Pettis S.J., Higo H.A., Pankiw T.,Winston M.L. (1997) - Queen rearingsuppression in the honey bee - evidence fora fecundity signal. Insectes soc., 44:311-322.
Pidek A. (1988) - Wp³yw pozyskiwaniapy³ku na rozwój i produkcyjnoœæ rodzinpszczelich oraz efekty ekonomiczne pasiek.Pszczeln. Zesz. Nauk., 32: 197-213.
Polišèuk W.P. (1984) - Sbory pylcy wperiod medosbora. Pèelovodstvo, 61 (11):12-13.
Seeley T.D. (1989) - The honey beecolony as a superorganism. Am. Sci., 77:546-553.
Stojko A., Rzepecka-Stojko A., StojkoJ. (1997) - Zastosowanie standaryzowa-nych obnó¿y pszczelich jako czynnikaos³onowego w badaniu embriotoksycznoœcikwasu acetylosalicylowego. XXXIVNaukowa Konferencja Pszczelarska.Pu³awy 12-13 marzec: 62.
Taber S. (1984) - Pollen and pollentrappig. Am. Bee J. 124 (7): 512-513.
Wilde J., Bobrzecki J. (1989) -U¿ytkowanie powa³kowego po³awiaczapy³ku. Pszczelarstwo, 40 (5): 14-15.
Wilde J., Bratkowski J. (1996) -Pozyskiwanie py³ku w rodzinach zzastosowaniem ró¿nych metod gospodarkipasiecznej. 33. Naukowa KonferencjaPszczelarska. Pu³awy. Materia³y konferen-cyjne: 90-91.
Wilde J., Cichoñ J. (1997) - Op³acalnoœæpozyskiwania obnó¿y py³kowych w Polsce.IV Krajowa Konferencja Apidologiczna.8-9. kwiecieñ, WSP Bydgoszcz: 96-106.
Wilde J., Krukowski R., Bobrzecki J.(1994a) - Porównanie metod gospodarkipasiecznej umo¿liwiaj¹cych zwiêkszenieiloœci pozyskiwanych obnó¿y py³kowych.Acta Acad. Agricult. Techn. Olst.Zootechnica, 39: 181-191.
Wilde J., Krukowski R., Bobrzecki J.(1994b) - Wp³yw metod tworzenianowych rodzin na rozwój i produkcyjnoœæmacierzaków. Acta Acad. Agricult. Techn.Olst. Zootechnica, 39: 123-133.
Wilde J., Siuda M. (1996) - Usefulnessevaluation of honey bee with shortpostcapping period for beekeepingmanagement. Proceedings of theInternational conference “Environmentalfactors and bee productivity andhealthiness” Dotnuva-Academija,: 58-63.
Winston M.L., Higo H.A., Colley S.J.,Pankiw T., Slessor K.N. (1991) - Therole of queen mandibular pheromone andcolony congestion in honey bee (Apismellifera L.) reproductive swarming(Hymenoptera: Apidae). J. Insect. Behav.,4: 649-660.
Vol. 47 No. 1 2003 Journal of Apicultural Science 19

WP£YW ZMIANY STRUKTURY RODZIN PSZCZELICHNA ILOŒÆ ODEBRANYCH OBNÓ¯Y PY£KOWYCH
B r a t k o w s k i J . , W i l d e J .
S t r e s z c z e n i e
Celem badañ by³o ocenienie wp³ywu zmian struktury rodzin pszczelich na iloœæ odebranychobnó¿y py³kowych. Doœwiadczenie przeprowadzono w latach 1997-1999. Rodziny podzielonona piêæ grup doœwiadczalnych. W grupie PTC (kontrolna) stosowano tradycyjne metodygospodarki pasiecznej oraz odbierano obnó¿a py³kowe. W grupie PTF wszystkie plastryzastêpowano ramkami z wêz¹. W grupie PTN tworzono odk³ady, i zak³adano im równie¿po³awiacze py³ku. W grupie PTI matki umieszczano na 2 tygodnie w klateczkach Zandera
Produkcja ca³kowita rodzin zosta³a wyra¿ona w kilogramach miodu i sk³ada siê z mioduodwirowanego i miodu przeliczeniowego. W ca³ym okresie doœwiadczenia tworzenie odk³adów(PTN - 8,5 kg) by³o najbardziej wydajn¹ metod¹ produkcji py³ku. Œrednia masa pozyskanegopy³ku by³a wysoko istotnie wy¿sza w porównaniu z pozosta³ymi grupami. Produkcja wpozosta³ych grupach by³a zbli¿ona, a ró¿nice istotne stwierdzono jedynie pomiêdzy rodzinamigrupy przesiedlanej na wêzê (PTF - 5,7 kg) a grup¹ kontroln¹ (PTC - 3,6 kg).
W tym czasie najwiêcej miodu odwirowano od rodzin grupy kontrolnej (PTC - 16,3 kg) igrupy, w której wymieniano gniazdo na wêzê (PTF - 14,4 kg), zaœ najmniej od rodzin, odktórych tworzono odk³ady (PTN - 7,9 kg) i ró¿nice te potwierdzono statystycznie (p=0,01).Istotnoœæ ró¿nic przy p=0,05 wyst¹pi³a pomiêdzy œrednimi wydajnoœciami uzyskanymi odrodzin grup PTC i PTI.
W doœwiadczeniu najwy¿sz¹ produkcjê ca³kowit¹ uzyskano od rodzin, w których tworzonoodk³ady (grupa PTN - 54,0 kg) i przesiedlano na wêzê (PTF - 43,7 kg). Grupy te wysokoistotnie przewy¿szy³y grupê kontroln¹ (PTC - 32,8 kg). Wysoko istotne ró¿nice wyst¹pi³yrównie¿ pomiêdzy grup¹ PTN a PTC (32,8 kg) i rodzinami, w których matki izolowano wklateczkach (PTI - 33,6 kg).
Mo¿na, zatem stwierdziæ, ¿e regulowanie struktury rodzin pszczelich przez odbieranieczerwiu i pszczó³, w celu wykonywania odk³adów (grupa PTN) oraz izolowanie matek (grupaPTI) obni¿a³o istotnie produkcjê miodu w porównaniu z rodzinami grupy kontrolnej (grupaPTC). Obni¿enie produkcji miodu a istotny wzrost produkcji py³ku w rodzinach grupy PTNby³y wynikiem zastosowanej metody regulowania struktury rodziny.
Odbieranie czerwiu i pszczó³ w celu tworzenia odk³adów oraz pozyskiwanie od nich obnó¿ypy³kowych by³o najbardziej wydajn¹ metod¹ intensyfikacji produkcji.
S³owa kluczowe: miód, py³ek, pozyskiwanie py³ku, produkcja ca³kowita, czerw pszczeli,pszczo³y, matki pszczele.
20

BIOLOGICAL CRITERIA FOR ACCEPTING OF BEEBREAD AS A MITE FOOD BY Tyrolichus casei Oudemans
(Acarina: Acaridae)
W i t C h m i e l e w s k i
Department of Bee Products, Apiculture Division, Research Institute of Pomology and Floriculture,Kazimierska 2, 24-100 Pulawy, Poland.
Fax: +48 81 8864209; e-mail: [email protected]
S u m m a r y
The object of the study was to confirm the possibility of the development of Tyrolichus caseiOudemans on bee bread and the acceptance of this product by mites as evidence that these pestsmight threaten the provisions of honeybees.
Monocultures of the species were set up from specimens collected from hive debris and werekept in the laboratory at temperature ca. +20°C, RH - about 85%; bee bread was used as mitefood. Biological experiments were conducted under the same conditions.
Some live parameters (average) of T. casei were as follows: embryonic development - 3.4days, complete life history - 14.5 days, mortality of juvenile instars - 10.0%, frequency offemales - 52.2%, longevity of adults - 101.8 days, oviposition period - 34.3 days, fecundity offemales per lifespan - 426.3 eggs.
These results show a comparatively high biological potential for raising T. casei on bee breadand confirm a working hypothesis as concern the attractiveness and effectiveness of this kind ofmedium in rearing acarid mites. Evidence is presented that cheese mites are potential pests ofstored hive products.
Keywords: Acarina, bee bread, biology, pests, Tyrolichus casei.
INTRODUCTIONTyrolichus casei Oudemans is a cosmo-
politan acarid species commonly known asa cheese mite and pest of various storedfood products. Apart from cheese and otherstored food it has been observed as a pestinfesting grain, damp flour, dog-food,sometimes on ergot of rye and in insect col-lections. There are also records of this spe-cies under the bark of old tree stumps and ina mouse nest (Boczek 1980, Hughes1976, Olsen et al. 1982, Robertson 1946,Smiley 1991, Türk, Türk 1957,Zakhvatkin 1941). It belongs to a groupof allergenic mite species causing humandermatitis (Henschel 1929, Rosicky et al.1979)
In apiaries T. casei was observed as an
inhabitant of beehives colonizing hive de-bris, old honeycombs and as a pest infestingstored bee products and the provisions ofbees, first of all bee bread and pollen(Chmielewski 1991 1992, 2001, Grobov1981). As the results show from earlierstudies on cheese mites fed bee- -collectedpollen (pollen pellets), they accept thisproduct as a suitable nourishment for popu-lation increase (Chmielewski 1994).
The purpose of this study was the experi-mental confirmation of the possibility ofcheese mite development on bee bread andthe acceptance of this product as evidencethat these pests might threaten the provi-sions of honey bees.
Vol. 47 No. 1 2003 Journal of Apicultural Science 21
MATERIAL AND METHODSMonocultures of the species were set up
from live specimens collected from hive de-bris which were put into rearing cages andkept in the laboratory at temperature ca.+20°C, RH - about 85%; bee bread takenfrom honeycombs was used as mite food.Biological experiments were conducted un-der the same conditions.
Observations of mite development beganon 100 newly deposited eggs (10 rearingcages x 10 one-day-old eggs) and finishedafter the eclosion of adults. Advances in thedevelopment cycle and mortality of speci-mens were recorded every 1-2 days.
Longevity and fecundity of mites wereexamined during observations of 25 pairsformed of freshly emerged adults (one pair= 1 female + 1 male), which were placedinto rearing cages supplied with smallpieces of bee bread (each pair was put intoseparate cage). Observations were made ev-ery 2-3 days until the natural death of all
specimens.More particulars and descriptions of pro-
cedures used in similar studies on someacaroid mite species fed bee bread andbee-collected pollen, were published in ear-lier papers (Chmielewski 1978, 1983,1994, 1995, 1998, 2000, 2002).
RESULTS AND DISCUSSIONLife parameters of T. casei obtained in
the present studies conducted on bee bread,showed that this kind of nourishment wasacceptable by these mites.
Comparison of these results with thebionomics of cheese mite obtained onbee-collected pollen, i.e. pollen loads, underthe same temperature and humidity(Chmielewski 1994) proved that both ofthese media are very effective and bringabout a favourable population increase.However some biological data weredifferentiated.
22
Character Bee bread(present results)
Pollen loads(Chmielewski 1994)
Embryonic development (days) 3.4(3-7) 5.7(3-10)
Complete life history (days) 14.5(9-19) 18.5(14-22)
Mortality of juvenile stages (%) 10.0(0-30) 49.3(10-90)
Frequency of females (%) 52.2(40-62) 47.2(30-90)
Longevity of adults (days) 101.8(28-216) 85.6(30-205)
Oviposition period (days) 34.3(19-53) 63.9(16-129)
Nonfecundity general (days) 39.6(4-137) 19.2(3-123)
Fecundity of femalesper life-span (eggs) 426.3(236-570) 443.9(93-820)
Productivity of femaleper oviposition day (eggs) 12.4(1-35) 7.4(1-25)
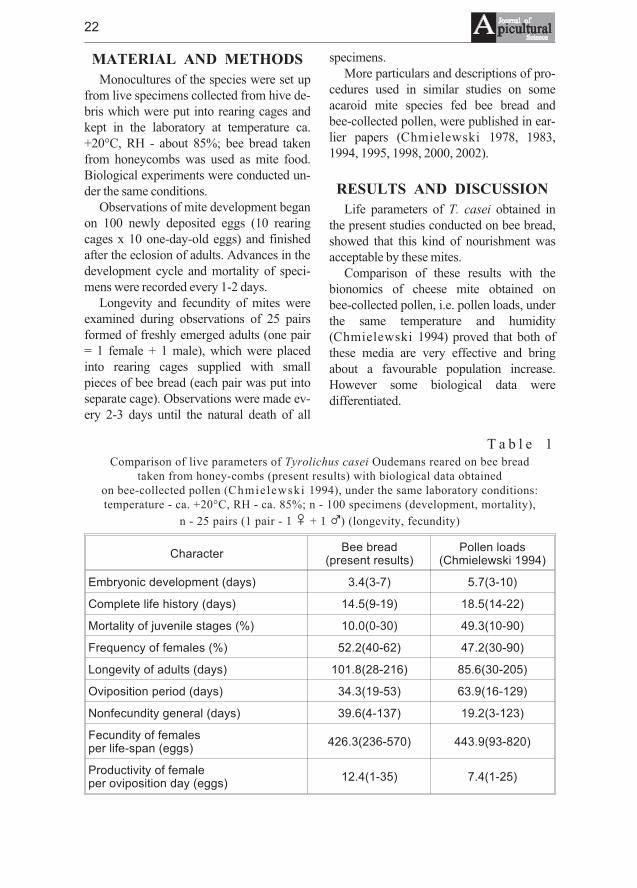
T a b l e 1Comparison of live parameters of Tyrolichus casei Oudemans reared on bee bread
taken from honey-combs (present results) with biological data obtainedon bee-collected pollen (Chmielewski 1994), under the same laboratory conditions:temperature - ca. +20°C, RH - ca. 85%; n - 100 specimens (development, mortality),
n - 25 pairs (1 pair - 1 � + 1 �) (longevity, fecundity)

Such parameters as embryonic develop-ment (3.4 days) and complete developmentcycle (14.5 days) of mites obtained on beebread were shorter than on pollen (5.7 and18.5 days, respectively). The ovipositionperiod on bee bread (34.3 days) was alsosignificantly shorter than on pollen (63.9days), but productivity of females fed beebread (average 426.3 eggs) was slightlylower than on pollen pellets (443.9 eggs perlife-span). Viability of T. casei fed beebread expressed as a percentage of speci-mens finishing their development cycle asimagines (90.0%) and their longevity(101.8 days) was significantly higher thanviability of mites reared on pollen (eclosionof adults - 50.7% and their average longev-ity - 85.6 days) (Table 1).
A comparison of the bionomics of T.casei fed on bee bread with biological dataof other mite species (Acarus farris(Oudemans), Acarus immobilis Griffiths,Acarus siro L., Glycyphagus domesticus(De Geer)) obtained earlier, on the samekind of food, and under similar conditions,shows that biological parameters of cheesemite are evidently greater than the same pa-rameters of other examined acaroids(Chmielewski 1983, 2002).
CONCLUSIONSThe results of these experiments show a
comparatively high biological potential forraising T. casei on bee bread and for itspopulation increase on this medium.
Calculated biological parameters con-firm a working hypothesis concerning theattractiveness of this kind of food for thisspecies.
It also provides evidence that cheesemite is a potential pest of some stored hiveproducts (bee bread, bee-collected pollen).
Bee bread might be recommended as anuseful, effective medium for rearing acaridmites under laboratory conditions for exper-imental and educational purposes.
ACKNOWLEDGEMENTSAuthor would like to thank Richard A.
Baker (University of Leeds, UK) for checkof the English version of manuscript in re-spect of language correctness.
REFERENCESBoczek J. (1980) - Zarys akarologii
rolniczej. PWN, Warszawa, 355pp.
Chmielewski W. (1978) - P³odnoœæ irozwój niektórych gatunków roztoczyulowych na py³ku. Pszczeln. Zesz. Nauk.,22: 173-180.
Chmielewski W. (1983) - Pierzga jakopokarm rozkruszków z rodzaju Acarus(Acarina: Acaridae). Zesz. Problem. Post.Nauk Roln., 252: 171-178.
Chmielewski W. (1991) - Stored productmites (Acaroidea) in Polish beehives. (In:)Dusbabaek, F. and V. Bukva (eds.),Academia, Prague and SPB Academicpublishing bv, The Hague, vol. 1: 615-619.
Chmielewski W. (1992) - Sk³ad gatun-kowy i liczebnoœæ akarofauny w osypienaturalnym zimuj¹cych rodzin pszczelich.Pszczeln. Zesz. Nauk., 36: 74-90.
Chmielewski W. (1994) - Sk³adanie jaj irozwój rozkruszków (Acaroidea) na py³kuzbieranym przez pszczo³y. Pszczeln. Zesz.Nauk., 38: 131-142.
Chmielewski W. (1995) - Pollen loads asa food of stored product mites (Acaroidea).Pszczeln. Zesz. Nauk., 39(1): 169-176.
Chmielewski W. (1998) - Some biolo-gical data of Lepidoglyphus destructor(Schr.) (Acari: Glycyphagidae) - results oflaboratory rearing on pollen pellets.).Pszczeln. Zesz. Nauk., 42(2): 23-24.
Chmielewski W. (2000) - Michaelopuscorticalis (Mich.) (Acari: Acaridae),arboreal mite species found also in beehivedebris - biology on pollen pellets. Pszczeln.Zesz. Nauk., 46(2): 115-121.
Chmielewski W. (2001) - Sk³ad gatun-kowy szkodliwej akaro-entomofaunyœwie¿ych obnó¿y py³kowych. Materia³y zXXXVII Naukowej KonferencjiPszczelarskiej, Pu³awy, 2001: 13-14.
Vol. 47 No. 1 2003 Journal of Apicultural Science 23

Chmielewski W. (2002) - Life parametersof Glycyphagus domesticus (De Geer)(Acarina: Glycyphagidae) settling storedapples - results of laboratory culture onbee-bread. J. Fruit. Ornam. Plant Res., 10:155-163 .
Grobov O.F. (1981) - Akaridievyje klešèi- vrediteli kormovych zapasov pchel.Veterinaria, 1: 55-57.
Henschel J. (1929) - ReizphysiologischeUntersuchungen der Käselmilbe Tyrolichuscasei (Oudemans). Z. Ergl. Physiol., 9:802-837.
Hughes A.M. (1976) - The mites of storedfood and houses. Tech. Bull. Minist. Agric.,Fish. and Food, No 9, 2nd ed. HerMajesty’s Stationery Office, London,400pp.
Olsen A.R., Potter R.W., Bryce J.R.(1982) - A key for the identification ofthe adults of common food-contaminatingacarid mites and discussion of slidemounting technique. Food and DrugAdmin. Lab. Inform. Bull., 2588: 1-6.
Rosicky B., Cerny V., Daniel, M.,Dusbabek F., Palicka P., Samsinak K.(1979) - Roztoci a klistata skodici zdravicloveka. CSAV, Academia, Praha, 1208pp.
Smiley R.L. (1991) - Mites (Acari). (In:)J.R. Gorham (Ed.) Insect and mite pests infood. USDA & USDH&HS, U.,S.Government Printing Office Washington:vol.1: 3-44; vol. 2: 483-526.
Türk E., Türk F. (1957) - Systematik undÖkologie der Tyroglyphiden Mitteleuropas.Beitr. Syst. Ökol. 1: 1-233.
Zakhvatkin A.A. (1941) - Paukoobraznyje,Tiroglyphoidnyje klešèi (Tyroglyphoidea),Fauna SSSR, 6(1), Moskva-Leningrad,475pp.
BIOLOGICZNE KRYTERIA AKCEPTACJI PIERZGIPSZCZELEJ JAKO POKARMU ROZTOCZY PRZEZ
Tyrolichus casei Oudemans (Acarina: Acaridae)
C h m i e l e w s k i W .
S t r e s z c z e n i e
Rozkruszek serowiec, Tyrolichus casei Oudemans, powszechnie znany jako szkodnik wieluprzechowywanych produktów spo¿ywczych, jest tak¿e mieszkañcem uli pszczelich, gdziezwykle kolonizuje osyp gromadz¹cy siê na dennicach i pora¿a zapasy pokarmu swychgospodarzy zgromadzone w plastrach (pierzgê, py³ek, miód); pojawia siê te¿ niekiedyw pracowniach pszczelarskich i magazynach pasiecznych na sk³adowanych w nich produktach.
Czyste gatunkowo hodowle rozkruszka za³o¿ono na bazie osobników doros³ych wybranych zosypu zimuj¹cych pszczó³ i prowadzono w laboratorium - w temperaturze ±20°C, w wilgotnoœciwzglêdnej powietrza ok. 85%, w klateczkach hodowlanych u¿ywanych przez akarologóww tego typu badaniach biologicznych; jako pokarm podawano roztoczom pierzgê pszczel¹.
Doœwiadczenia biologiczne prowadzono w tych samych warunkach. Obserwacje rozwojuroztoczy rozpoczynano na 100 œwie¿o z³o¿onych jajach (10 jedno-dniowych jaj x 10 komórekhodowlanych) i koñczono po wylêgu osobników doros³ych. Zaawansowanie cyklurozwojowego sprawdzano co 1-2 dni. D³ugowiecznoœæ i p³odnoœæ roztoczy by³y badane w tokuobserwacji 25 par utworzonych ze œwie¿o wylêgniêtych imagines (jedna para = 1 samica + 1samiec), które umieszczano w klateczkach hodowlanych zaopatrzonych w ma³e kawa³ki pierzgipszczelej (ka¿d¹ parê trzymano w oddzielnej komórce hodowlanej). Kontrolê doœwiadczeñ,
24

a tak¿e w miarê potrzeby uzupe³nianie lub wymianê pokarmu, prowadzono co 2-3 dni a¿ donaturalnej œmierci wszystkich osobników.
Uzyskane parametry ¿yciowe T. casei (œrednie dane) przedstawiaj¹ siê nastêpuj¹co:Rozwój embrionalny (dni) - 3.4 (3-7)Ca³kowity rozwój osobniczy (dni) - 14.5 (9-19)Œmiertelnoœæ osobników w stadiach m³odocianych (%) - 10.0 (0-30)Frekwencja samic (%) - 52.2 (40-62)D³ugoœæ ¿ycia osobników doros³ych (dni) - 101.8 (28-216)Okres sk³adania jaj (dni) - 34.3 (19-53)Bezp³odnoœæ ogó³em (dni) - 39.6 (4-137)P³odnoœæ samic w ci¹gu ca³ego ¿ycia (jaja) - 426.3 (236-570)Produktywnoœæ samic na dzieñ p³odny (jaja) - 12.4 (1-35)
Wyniki te wskazuj¹ na stosunkowo wysoki potencja³ biologiczny T. casei na pierzdzepszczelej i potwierdzaj¹ za³o¿enia hipotezy roboczej odnoœnie do atrakcyjnoœci i skutecznoœcitego rodzaju po¿ywki w hodowli rozkruszków. S¹ one tak¿e jednym z dowodów na to, ¿erozkruszek serowiec nale¿y do potencjalnych szkodników przechowywanych produktówpasiecznych stanowi¹c dla nich realne zagro¿enie, w tym zw³aszcza dla pierzgi.
S³owa kluczowe: Acarina, biologia, pierzga, szkodniki, Tyrolichus casei.
Vol. 47 No. 1 2003 Journal of Apicultural Science 25

ONSET OF OVIPOSITION IN HONEYBEE QUEENS KEPTIN BOXES WITH NON-FREE FLYING BEES
B o ¿ e n a C h u d a - M i c k i e w i c z , J a r o s ³ a w P r a b u c k i ,J e r z y S a m b o r s k i
Department of Apiculture, Agricultural University Doktora Judyma 20, 71-466 Szczecin, Poland,e-mail: [email protected]
S u m m a r y
The aim of the paper was to examine the start of egg lying by instrumentally inseminatedqueens kept both before and after insemination with non-free flying bees in closed boxes in aroom without air-conditioning.
The experiment was carried out on queens reared from 1-day-old larvae in June and July.Sealed queen cells were introduced 1-2 days before queens emerging to boxes 130×115×70 mmsettled with about 50 bees. Each box was provided with a comb, candy and a syringe with water.Queens were instrumentally inseminated twice at the age of the 7th and 9th day with 4 µl ofsemen. After the first insemination they were introduced to new boxes with about 250 bees andcandy containing 5% pollen. Control group consisted of queens kept in trapezoid mating nucleiwith about 400 bees. These queens were also instrumentally inseminated twice at the same ageand with the same dose of semen.
It was found that queens kept in boxes started egg lying. However, insemination efficiencywas worse than that in queens kept in nuclei. Queens in nuclei started egg lying on average 10days after the first insemination, whereas those in boxes about 1.5 times later.
Keywords: honeybee queen, instrumental insemination, oviposition, box.
INTRODUCTIONThe employment of instrumental insemi-
nation in the process of rearing honeybeequeens has substantially contributed to theprogress in beekeeping, i.e. to the introduc-tion of highly productive and docile beelines. By their appropriate crossbreeding,one obtains hybrids that produce signifi-cantly more honey then the starting material(Prabucki and Chuda-Mickiewicz1998, 2000, 2002). A considerable portionof inseminated honeybee queens producedby breeding and reproductive apiaries con-sists of queens without verified oviposition(Troszkiewicz, personal communica-tion). These queens are malevolently ac-cepted by bees and their introduction intocolonies requires the employment of suit-able methods (Wilde et al. 2002). This
situation indisposes beekeepers for intro-ducing them to apiaries. Woyke andJasiñski (1979 1980) recommend to keepqueens with non-verified oviposition fortwo days more, after insemination, in boxes(with a capacity of 793 ml each) settledwith 250-350 bees. Studies of Chuda--Mickiewicz and Prabucki (1993)showed that inseminated queens kept in theapiary in boxes accompanied by about 250free flying bees started egg lying in thesame time as those kept in trapezoid matingnuclei.
The aim of the study was to examine theonset of oviposition by queens kept bothbefore and after insemination with bees inclosed boxes placed in a room.
Vol. 47 No. 1 2003 Journal of Apicultural Science 27
MATERIAL AND METHODSExperiment was carried out in 2001 and
2002 on Carniolan (Apis mellifera carnica)queens bred in 2 rearing series. Queenswere reared from 1-day-old larvae in aqueenless colony, the first series was inJune and the second in July. On 1-2 daysprior to queens emerging, queen cells wereintroduced into boxes, 130x115x70 mm,settled with about 50 bees., A slice of combwas secured in the boxes to the ceiling, nextto which a syringe with water was intro-duced through an opening in the ceiling,and a candy lying on a shelf above box
bottom. Boxes were kept in a room withoutair-conditioning both before and after queeninsemination at 18-20°C. Queens were in-strumentally inseminated twice at the age of7 and 9 days, each time with 4 µl of semen.After the first insemination, they were intro-duced into new boxes settled with about250 bees and provided with candy contain-ing 5% pollen addition. Control group con-sisted of queens kept in trapezoid matingnuclei with about 400 bees. These queenswere also instrumentally inseminated twiceon the 7th and the 9th day of life with 4 µl ofsemen. The obtained data were analysedstatistically with the use of Student’s t-test.
28
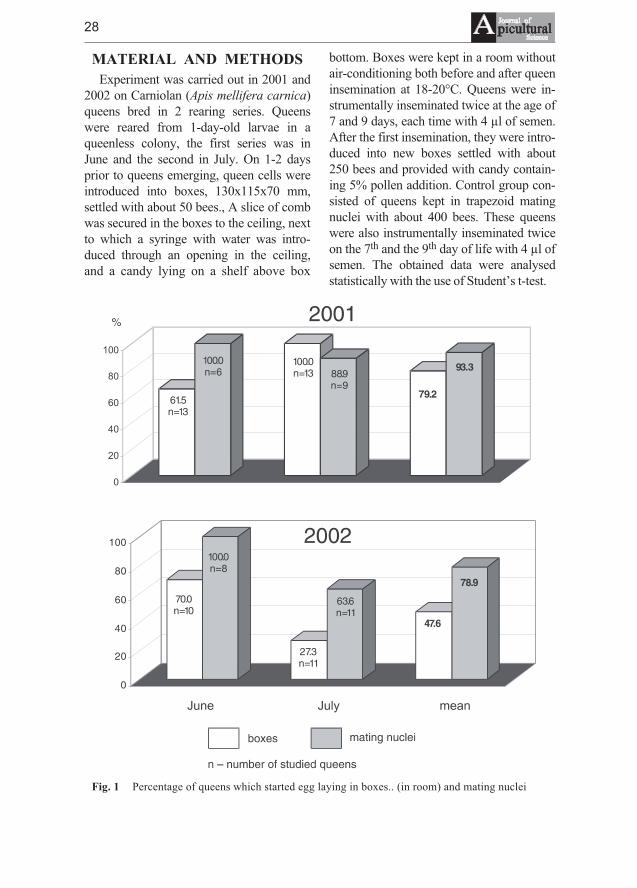
Fig. 1 Percentage of queens which started egg laying in boxes.. (in room) and mating nuclei
RESULTSOut of the total number of 79 instrumen-
tally inseminated queens, 73.4% started egglying. However, insemination efficiency (%of queens starting oviposition) differed ingroups; from 64.4% in queens starting egglying in boxes, to 85.3% laying eggs in nu-clei (Fig. 1). Lower efficiency of insemina-tion in queens kept in boxes was causedmainly by queens reared in July 2002. Thesame situation was found in queens kept inmating nuclei when the lowest numberstarted egg laying. Thus, insemination effi-ciency depended upon the method of queenkeeping and probably upon conditions dur-ing their rearing.
Time of starting the egg lying by queensdepended upon the method of their keeping(Tab. 1). Queens kept in mating nucleistarted egg laying significantly earlier thanthose kept in boxes. The former startedoviposition on average 10.1 days after thefirst insemination. Queens kept in boxesstarted egg laying on average 6.4 days later.In the first study year, they started egg lying
14.9 days after insemination, and in the sec-ond one after 19.6 days. In 2002, queenskept in mating nuclei also started egg lyingslightly later. Conditions prevailing duringqueen rearing influenced the results ofqueen insemination. This was noticed espe-cially in the second study year, when in-semination efficiency of queens kept inboxes was low and onset of oviposition wasdelayed.
CONCLUSIONSThe obtained results show, that the per-
centage of instrumentally inseminatedqueens which started oviposition in boxes islower (ca. 20%) than that achieved in mat-ing nuclei and the time of beginning ofoviposition is over 1.5 times longer.
REFERENCESChuda-Mickiewicz B., Prabucki J.
(1983) - Podejmowanie czerwienia przezinseminowane matki pszczeleprzetrzymywane w skrzynkach w asyœcieswobodnie oblatuj¹cych siê pszczó³.Pszczeln. Zesz. Nauk. 37: 23-31.
Vol. 47 No. 1 2003 Journal of Apicultural Science 29
Year Periodof rearing
Method of keeping queens before and after insemination
Boxes Mating nuclei
n
no. of days since1 insemination
to start of oviposition n
no. of days since1 insemination
to start of oviposition
range mean range mean
2001June 8 13 - 21 14.2 6 8 - 10 8.5
July 11 14 - 21 15.4 8 6 - 23 9.6
Mean 14.9 b* 9.1a
2002June 7 16 - 28 19.4 8 8 - 20 12.4
July 3 19 - 21 20.0 7 6 - 14 9.3
Mean 19.6b 10.9a
Overall mean 16.5b 10.1a
* numbers in rows followed by different letters differ significantly at p=0.05
T a b l e 1Onset of oviposition in queens according to the method of their keeping before and after
instrumental insemination

Prabucki J., Chuda-Mickiewicz B.(1998) - Results of the middle Europeanbee improvement in Western Pomerania.Fol. Univ. Agric. Stetin. 185 Zootechnica(36):27-37.
Prabucki J., Chuda-Mickiewicz B.(2000) - Doskonalenie towaroweu¿ytkowanych w Polsce ras pszczó³. Fol.Univ. Agric. Stetin. 210 Zootechnica(39):131-142.
Prabucki J., Chuda-Mickiewicz B.(2002) - Honey yield of Polishcommercial lines of middle European bee(Apis mellifera mellifera L.) and theircrossbreed with other races. 5th
International Conference the Black BeeApis mellifera mellifera, September 2-6,2002, Wierzba, Poland, Programe andsummaries:22.
Wilde J., Wilde M., Gogolewska E.(2002) - £atwe i skuteczne sposobywymiany matek pszczelich. Pszczelnictwo53(4): 4-6.
Woyke J., Jasiñski Z. (1979) - Numberof worker bees necessary to attendedinstrumentally inseminated queen kept inan incubator. Apidologie 10(2):149-155.
Woyke J., Jasiñski Z. (1980) - Influenceof number of attendant workers on theresults of instrumental insemination ofhoneybees queens kept in roomtemperature. Apidologie 11(2):173-180.
ROZPOCZYNANIE CZERWIENIA PRZEZ MATKI PSZCZELEPRZETRZYMYWANE W SKRZYNKACH Z PSZCZO£AMI
BEZ MO¯LIWOŒCI OBLATYWANIA SIÊ PSZCZÓ£
C h u d a - M i c k i e w i c z B . , P r a b u c k i J . , S a m b o r s k i J .
S t r e s z c z e n i e
Doœwiadczenie przeprowadzono w 2001 i 2002 roku na matkach wychowywanychz 1-dniowych larw w czerwcu i lipcu. Mateczniki na 1-2 dni przed wygryzieniem siê matekpoddawano do zasiedlonych ok. 50 pszczo³ami skrzynek o wymiarach 130x115x70 mm.W skrzynkach umieszczony by³ plasterek woszczyny, ciasto miodowo-cukrowe i strzykawkaz wod¹. Skrzynki do czasu inseminacji jak i po unasiennieniu matek przetrzymywanow nieklimatyzowanym pomieszczeniu. Matki inseminowano dwukrotnie w wieku 7 i 9 dni, 4µlnasienia. Po pierwszej inseminacji poddawano je do nowych skrzynek z ok. 250 pszczo³amii ciastem miodowo-cukrowym z 5% dodatkiem py³ku kwiatowego. Grupê kontroln¹ stanowi³ymatki przetrzymywane w ulikach weselnych z ok. 400 pszczo³ami. Matki te równie¿inseminowano dwukrotnie w 7 i 9 dniu ich ¿ycia, 4µl nasienia.
Stwierdzono, ¿e matki przetrzymywane w skrzynkach podejmuj¹ czerwienie. Skutecznoœæunasieniania by³a jednak gorsza ni¿ u matek przetrzymywanych w ulikach weselnych. Okresrozpoczêcia czerwienia by³ zró¿nicowany. W ulikach matki przystêpowa³y do sk³adania jajœrednio po 10 dniach od pierwszego unasieniania, natomiast w skrzynkach ok. 1,5 raza póŸniej.
S³owa kluczowe: matka pszczela, unasienianie, rozpoczynanie czerwienia, skrzynka.
30

EXPERIENCE WITH USING BUMBLEBEESAS POLLINATORS IN THE REGENERATION OF THEGENETIC RESOURCES OF SOME FORAGE LEGUMES
J a r m i l a D r o b n a 1 , V l a d i m i r P t a c e k 2
1Research Institute of Plant Production, Bratislavska 122, 92168 Piestany, Slovak Republic,e-mail: [email protected]
2Masaryk University, Faculty of Science, Kotlarska 2, 61137 Brno, Czech Republic,e-mail: [email protected]
S u m m a r y
In our study the effectiveness of the bumble bees as pollinators of several species of thegenera Trifolium L. (T. rubens, T. ambiguum, T. hybridum, T. repens, T. pratense, T. medium),Medicago L. (M. romanica, M. polychroa, M. falcata, M. sativa) and Lotus corniculatus wasevaluated over a 5-year period at the Research Institute of Plant Production, Piestany. Thosespecies were propagated in technical isolation separately or in combinations using Bombusterrestris for pollination. In the years 2000, 2001 Bombus pascuorum and B. lapidarius wereused for pollination of T. pratense and M. falcata. The results showed that bumblebees couldserve as effective pollinators of some perennial forage legumes with the aim to obtain sufficientamount of seeds for their maintenance in a gene bank. The existing differences in attractivenessbetween various plant genera may influence the amount of harvested seed in the case of theirpresence under the same coverage. Better seed yields per plant were obtained in Medicago incomparison to Trifolium species when grown together in the same enclosure. The effect ofBombus species on the seed weight of T. pratense was not significant. However, additionalresearch is required to improve our knowledge in the reproductive biology and pollinationrequirements inclusive specific demands on the insect agent.
Keywords: forage legumes, multiplication, pollination, Bombus spp.
INTRODUCTIONRegeneration of germplasm is one of the
most critical processes involved in the con-servation of plant genetic resources and inthe gene bank management. Even under op-timum ex situ storage conditions, the viabil-ity of seeds declines and the geneticdiversity is lost. Thus, the monitoring of vi-ability and timely regeneration of seedsmust be the prior activity of all gene banks(Richards 2001).
Maintenance of perennial forage le-gumes, which mostly need insect cross--pollination and are self-incompatible, ismore difficult and costly than that of self--pollinated crops. The complexity and
expense of regeneration are higher for spe-cies, which are insect pollinated.
Regeneration of sufficient number ofplants in isolation is a possible solution tomaintain the genetic integrity of increasedperennial forage legumes. Principally, thereare three possible ways of isolation toachieve it: technical, space isolation andmanual pollination.
Manual pollination being thorough andprecise it is also time consuming with highdemand for labour. On the other hand, natu-ral pollinators are more efficient and effec-tive in producing quality germplasm thanpollinating by hand.
Isolation in space may be possible, if
Vol. 47 No. 1 2003 Journal of Apicultural Science 31

specific areas for the multiplication of ge-netic resources are reserved. Risk arises inthe case of forage crops as cross-pollinationwith wild-growing species and possiblywith cultivated species grown on neigh-bouring fields may occur. Also the inter- aswell as intra-specific competition for polli-nating insects may play its negative role ifspace isolation is used.
The possible solution how to maintainthe genetic integrity of increased forage le-gumes seems to be through the regenerationof sufficient number of plants in technicalisolation. The use of screened cages withspecially designed hives for honey, leafcutteror bumble bees for several insect-pollinatedcrop species is now common (Ellis et al.1981, Ptacek 1987, 1999, Richards1995). From several insect species bumblebees, Bombus spp. are recognized as effi-cient and effective pollinators of many cropspecies. Bumblebees have some obviousadvantages over honeybees for pollinatingplant germplasm in cages. Under limitedspace condition various Bombus species canpollinate successfully even those plant spe-cies which do not belong among their fa-vourite food sources in the open. Ptacek(1983), Ptacek et al. (1984) obtained satis-factory results in cage pollination of alfalfasecured not only by B. lucorum and B.terrestris, but also by B. lapidarius, B.pratorum and B. sylvarum. To save on re-sources, different crop species may beplanted under one enclosure (Engels andRao 1995).
The aim of this study was (1) to obtainthe basic information on agronomics andpollination requirements of some perennialforage legumes, (2) to compare the possibil-ities to pollinate various species of foragelegumes in one enclosure, (3) to find outthose Bombus species which are the mostsuitable for pollination of particular foragelegumes, (4) to improve the regenerationprocedures for the concerned crops.
METHODSThe effectiveness of bumble bees as
pollinators of several species of genusTrifolium L. (T. repens L., T. pratense L.,T. medium L., T. rubens L., T. ambiguumL., T. hybridum L.,), Medicago L.(M. romanica Prodan, M. polychroaGrossh., M. falcata L., M. sativa L.) andLotus corniculatus L. was evaluated overa 5-year period at the Research Institute ofPlant Production, Piestany. These specieswere multiplied in technical isolation sepa-rately or in combinations. In the years 1997,1998, and 1999 Bombus terrestris colonieswere used for pollination, in the followingyears, also B. pascuorum and B. lapidariuswere used for pollination of T. pratense andM. falcata.
Plants were planted out individually on2 m x 3 m beds and the first or the secondcropping season year was used for seedsproduction. Isolators were assembled andinstalled in the time of flower bud forma-tion, and hives with bumblebees were putinto cages at the beginning of flowering.The flowering period lasted 6 to 8 weeks,depending on the particular plant species.
Colonies of bumblebees were preparedin laboratory conditions. Given the re-stricted space and consequently limitedfood sources from plants on beds, smallerunits were prepared. In each of them a lay-ing queen or workers and brood in larvalstage were present. In order to minimise ini-tial stress after release, the colonies wassupplied with a portion of pollen dough. In-side the isolators sugar solution was sup-plied ad libitum.
RESULTS AND DISCUSSIONIn 1997 seed production per plant of
10 white clover (Trifolium repens L.) vari-eties grown outside and in enclosures werecompared. The varieties grown in isolatorsyielded more seed in 7 of all cases (Fig. 1),
32
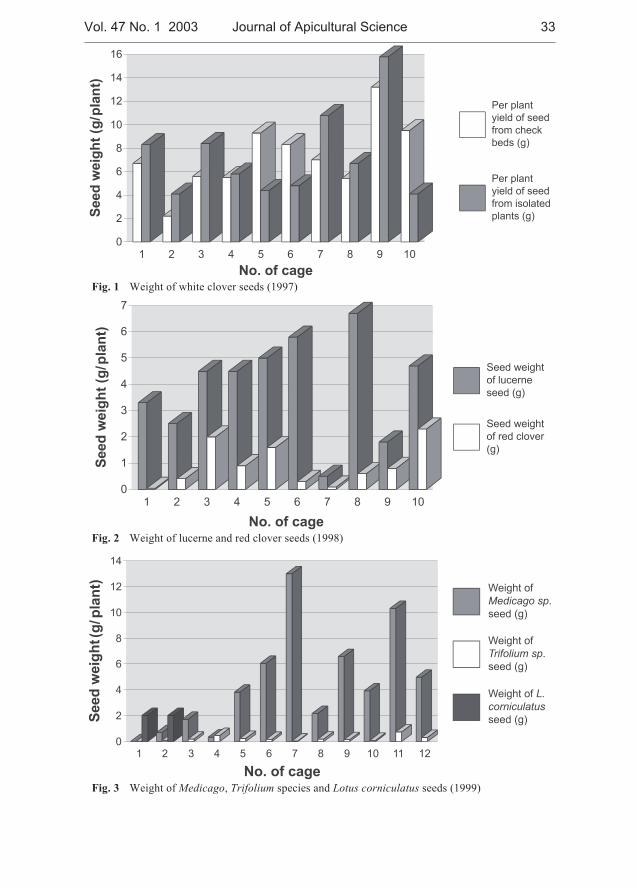
Vol. 47 No. 1 2003 Journal of Apicultural Science 33
Fig. 2 Weight of lucerne and red clover seeds (1998)
Fig. 3 Weight of Medicago, Trifolium species and Lotus corniculatus seeds (1999)
Fig. 1 Weight of white clover seeds (1997)
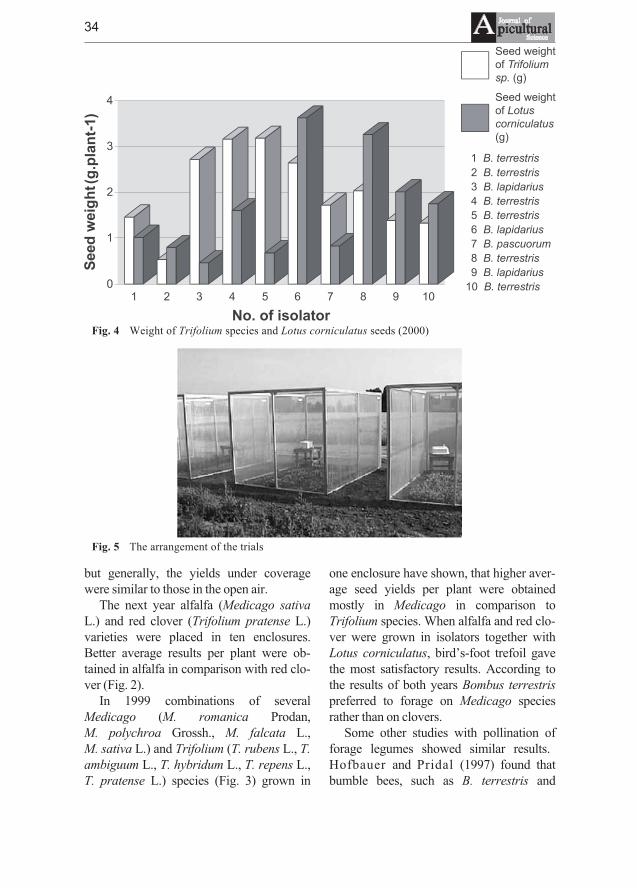
but generally, the yields under coveragewere similar to those in the open air.
The next year alfalfa (Medicago sativaL.) and red clover (Trifolium pratense L.)varieties were placed in ten enclosures.Better average results per plant were ob-tained in alfalfa in comparison with red clo-ver (Fig. 2).
In 1999 combinations of severalMedicago (M. romanica Prodan,M. polychroa Grossh., M. falcata L.,M. sativa L.) and Trifolium (T. rubens L., T.ambiguum L., T. hybridum L., T. repens L.,T. pratense L.) species (Fig. 3) grown in
one enclosure have shown, that higher aver-age seed yields per plant were obtainedmostly in Medicago in comparison toTrifolium species. When alfalfa and red clo-ver were grown in isolators together withLotus corniculatus, bird’s-foot trefoil gavethe most satisfactory results. According tothe results of both years Bombus terrestrispreferred to forage on Medicago speciesrather than on clovers.
Some other studies with pollination offorage legumes showed similar results.Hofbauer and Pridal (1997) found thatbumble bees, such as B. terrestris and
34
Fig. 4 Weight of Trifolium species and Lotus corniculatus seeds (2000)
Fig. 5 The arrangement of the trials

B. lucorum operate less effectively in isola-tors on red clover than B. lapidarius,B. pascuorum or B. sylvarum do. Brodie(1996) compared several species of bum-blebees in the open and observed thatB. terrestris never foraged on T. pratense.T. pratense was foraged in her study onlyby B. pascuorum. Also Ptacek (1987)found that B. pascuorum and B. sylvarumwere more effective on Trifolium pratenseand B. terrestris on alfalfa.
Basically on these statements, togetherwith B. terrestris also B. pascuorum andB. lapidarius were used as pollinatingagents in the cages with red clover next twoyears.
In 2000, combinations of L. corniculatuswith T. pratense or T. repens were evalu-ated. There were no considerable differ-ences between particular forage legumes inseed weight, but the higher average seedyield per plant was obtained in Trifoliumspecies. In contrast to the previous findings,no negative effect of Bombus species onseed weight of T. pratense (Fig. 4) was ob-served. Only in some cases red clover polli-nated by B. lapidarius and B. pascuorumgave better seed yield than by B. terrestris.According to the results both plant speciesseem to be similarly attractive for any of thebumblebee species used for pollination. In2001 mainly B. lapidarius was used for pol-lination of T. pratense and M. falcatagrown separately that time. In that yearhigher seed yield was obtained in redclover.
The simple evaluation of the results thatwe were able to do so far does not allowmuch definite conclusions. Obviously, thetechnical isolation can be used for regenera-tion of genetic sources because it brings atleast as high seed yields as the pollinationoutside. The comparison of plants on thelevel of the genera, especially in the case ofTrifolium and Medicago, hides the possibleinterspecific differences. Nevertheless, itcan be concluded, that B. terrestris might
not be a suitable pollinator in cases whereMedicago together with Trifolium with longcorolla tubes should be grown in one enclo-sure. Trifolium species can be grown withLotus species regardless of the bumblebeespecies. Using of some “non terrestris” spe-cies may be convenient for enabling thepollination “made-to-measure”.
CONCLUSIONSAccording to the results from this study
bumblebees can serve as pollinating agentsof some perennial forage legumes in enclo-sures and thus ensure the sufficient amountof seeds for the maintenance of a givenplant population in gene banks. Growingmore than one plant species under the samecoverage in order to lower the expenses ispossible. However, remarkable differencesin seed yields between Trifolium spp. andMedicago spp. grown together show thatthe process itself has to be studied more ac-curately and from the point of view both ofthe plants and the bees. Additional knowl-edge of the reproductive biology and polli-nation requirements of plants in theenclosures would be helpful, as well as theimprovement of bumblebee management inorder to allow the choice among severalspecies.
REFERENCESBrodie, L. (1996) - Bumblebee foraging
preferences: Differences between speciesand individuals. A thesis submitted as partof the requirements for the Degree of B. Sc.(Hons.) in Ecology at the University ofAberdeen.
Ellis, M.D., Jackson, G.S., Skrdla,W.H., Spencer, H.C. (1981) - Use ofhoneybee for controlled interpollination ofplant germplasm collections. Hortsci., 16:488-491.
Engels, J.M.M., Rao, R.R. (1995) -Regeneration of seed crops and their wildrelatives. Proc. of a Consult Mtg.,ICRISAT, Hyderabat, India. IPGRI, 167pp.
Vol. 47 No. 1 2003 Journal of Apicultural Science 35

Hofbauer, J., Pridal, A. (1997) - Use oflaboratory reared bumble bees forregeneration of genetic resources. Newmethods and knowledge used for thedocumentation and characterisation ofgenetic resources. Proc. Conf., Troubsko:105-108.
Ptacek V. (1983) - Pollination of alfalfa inenclosures. Rep. 28th Alfalfa Improv.Conf., Univ. Calif., Davis, Calif.,: 67-68.
Ptacek V., Krejci F., Babinec J. (1984)- Pollinators in breeding lucerne. Proc.
Eucarpia (Medicago sativa) session, Brno :273-276.
Ptacek V. (1987) - Technical isolation inresearch and breeding of legumes. Sbornikved. praci, OSEVA, VSUP Troubsko, (10):117-128. (Czech, En.sum.)
Ptacek V. (1999) - Different bee speciesfor pollination in enclosure. Insectpollination in enclosure. Proceedings of thespecialist’s meeting held in Soesterberg,The Netherlands, 30 Sept. to 3 Oct.:133-137.
Richards K.W. (1995) - The alfalfaleafcutter bee, Megachile rotundata: apotential pollinator of some annual forageclovers. J. Agr. Res. 34:115-121.
Richards K.W. (2001) - Pollinationconcerns in the conservation of plantgenetic resources.Proc. 8th PollinationSymp. Acta Hort. 561:191-209.
DOŒWIADCZENIE Z U¯YCIEM TRZMIELI JAKOZAPYLACZY W ODNAWIANIU ZASOBÓW GENETYCZNYCH
KILKU STR¥CZKOWYCH ROŒLIN PASTEWNYCH
D r o b n a J . , P t a c e k V .
S t r e s z c z e n i e
W 5-letnich badaniach (1997-2001) prowadzonych w Naukowym Instytucie ProdukcjiRoœlinnej w Piestanach (Czechy) oceniano efektywnoœæ trzmieli jako zapylaczy kilku gatunkówkoniczyny (Trifolium rubens, T. ambiguum, T. hybridum, T. repens, T. pratense, T. medium),lucerny (Medicago romanica, M. polychroa, M. falcata, M. sativa) i komonicy ro¿kowej -Lotus corniculatus. Roœliny uprawiano pod izolatorami oddzielnie lub w ró¿nych kombinacjachu¿ywaj¹c do ich zapylania trzmiela ziemnego (Bombus terrestris). W latach 2000-2001 dozapylania T. pratense i M. falcata u¿yto dodatkowo trzmiela rudego (Bombus pascuorum)i kamiennika (B. lapidarius). Badania wykaza³y, ¿e trzmiele mog¹ byæ wykorzystane jakoefektywne zapylacze trwa³ych pastewnych roœlin motylkowych dla uzyskania dostatecznejiloœci nasion na potrzeby banku genów.
S³owa kluczowe: pastewne roœliny motylkowe, zapylanie, Bombus spp.
36

THE ANTIMICROBIAL ACTIVITY OF HONEYOF THE STINGLESS BEE Trigona spp.
A s s e g i d G a r e d e w 1 , * E r i k S c h m o l z 1 ,I n g o l f L a m p r e c h t 2
1 Free University of Berlin, Institute of Zoology, Königin-Luise-Strasse 1-3, D - 14195 Berlin2 Free University of Berlin, Institute of Animal Physiology, Ehrenbergstrasse 26 - 28, D-14195 Berlin
* Corresponding author: e-mail: [email protected]
S u m m a r y
Stingless bees (Trigona spp.) produce a special honey which is less viscous and darker thanhoney of the honeybees (Apis spp.) and has a strong acid flavour. Trigona bee honey is highlyprized in Ethiopia for medicinal use as a panacea for many ills. The antimicrobial activities oftwo samples of Trigona spp. honey obtained from Ethiopia and honeybee honey from Germanywere tested in vitro against four fungal and six bacterial species.
Fungi were generally less sensitive than the bacteria or not sensitive at all to the treatments.Only Aspergillus niger and Penicillium chrysogenum responded slightly to one honey sample ata higher concentration. The bacteria reacted differently to the honey samples tested, with norelation to their Gram reaction. The minimal inhibitory concentrations of catalase treatedhoneys from stingless bees were higher than that of the non-treated samples for most bacterialspecies, indicating that hydrogen peroxide contributes to the antibacterial activity. Apis honey’shydrogen peroxide played no essential role in the antimicrobial activity against most bacteriatested. Concentrations below a certain value resulted in a microbial lawn of some bacteriaaround the inhibition zone indicating that the use of honey against infections at lowerconcentrations could be dangerous by encouraging the growth of bacteria.
Keywords: stingless bee, stingless bee honey, antimicrobial activity, non-peroxide activity,Trigona spp.
INTRODUCTIONTherapy with bee products (apitherapy)
is a worldwide old tradition, used for thou-sands of years but replaced by antibiotics inmodern times by all but a few individualspreferring traditional or natural remedies orthose with no access to antibiotics (Molanet al. 1988). It is, however, reviving cur-rently, apart from others, due to the increas-ing report of resistance of bacteria againstantibiotics (Greenwood 1995, Molan andBrett 1998). In addition to the drug resis-tance of bacteria, the side-effects of somepharmaceutical products may give rise to anaversion to synthetic drugs (Thompson1976, Kauffman 1991) and to an
increasing interest in the use of alternativetherapies. Although honey has been used asa medicine since ancient times, its effective-ness as a remedy has been revealed to bedue to its antibacterial activity only a cen-tury ago (Molan 2001). The antibacterialactivities of honey were considered to bedue to: (I) osmotic effects - owing to thehigh osmolarity of honey whereby water isdrawn away from microorganisms, reduc-ing their ability to survive; II) acidity - ashoney is acidic with a pH value between 3.2and 4.9 which inhibits growth of manypathogens whose optimal pH of growth isnot acidic; (III) hydrogen peroxide - whichis produced in the honey due to conversion
Vol. 47 No. 1 2003 Journal of Apicultural Science 37

of glucose to gluconic acid by glucoseoxidase, an enzyme produced by thehypopharyngeal gland of the bees; (IV)phytochemical factors - non peroxide anti-bacterial factors believed to be the variouscomplex phenols and organic acids oftenreferred to as flavonoids. These latter com-plex compounds do not breakdown easilyunder heat or light treatment and providesome types of honey with antibacterial ac-tivities even after these honeys were ex-posed to the mentioned factors, whichusually destroy enzymes (Molan 2001).
The typical honey investigated by mostresearchers is produced by Apis spp. Thishoney is undoubtedly the most widely andmassively collected and used by people fordifferent purposes. But regionally, espe-cially in the tropics and subtropics, there areother honeys made by different bee species,which are sometimes collected in substan-tial quantities (Krell 1996). One group ofbees that produces a considerable amount ofhoney and hence can be used in beekeepingare the stingless bees.
Stingless bees are eusocial insects thatbelong to the family Apidae and thesubfamily Meliponinae and lack a func-tional sting. Some species defend aggres-sively by biting the intruder - since theypossess well developed mandibles, emittinga caustic liquid from the mouth, releasingunpleasant odours, and irritating by crawl-ing in to the eyes, ears etc., of a predator(Ruttner 1992). Like the honeybees,stingless bees store honey and pollen(Michener 1974). The two important gen-era of Meliponinae that produce largeamounts of honey and hence are used instingless beekeeping are Melipona andTrigona (Crane 1992). The Melipona spe-cies are restricted to central and southAmerica whereas Trigona species occur inall the tropical continental regions (Amanoet al. 2000, Wille 1979).
Trigona spp. occur in Ethiopia at me-dium altitudes of up to 2300 m above sea
level and are about 10 mm long (Fichtland Adi 1994). They construct their nestsin underground cavities of naturally aban-doned ant nests, termite mounds, and cavi-ties under plant roots at a depth of about1m. The nest consists of thimble-sized ovalshaped honey/pollen pots placed around theone cell thick brood area of the combs thatare arranged horizontally, unlike the verti-cal combs of honeybees.
Stingless bees produce a special honeyknown in Ethiopia as Tazma honey. TheTazma honey is less viscous and darkerthan Apis honey and has a stronger acid fla-vour. In addition to that it also possesses astronger bacteriostatic effect (Krell 1996).Ripened honey of the stingless bees con-tains 30 to 35% water unlike Apis honey,with about 17.1% (Ruttner 1992). Thoughthe water content of stingless bee honey ishigh, it does not ferment in the nest presum-ably due to the abundant resin chemicals,which impart dark colour to it, and hydro-gen peroxide (http://www.sibexlink.com.my/g15magazine/g15mag_vol3jan_science.html). This honey is often highly prized lo-cally for medicinal use as a panacea formany ills, especially by the poor people ofrural areas with less access to modern medi-cine. Due to its strong sour taste it is inedi-ble and used only for medicinal purpose totreat cough, stomach disturbance, sorethroat, tonsillitis, stomach and intestinal ul-cers, cold, disease of the mouth and mucusmembrane, and as wound dressing.Stingless bee honey is considered in folkmedicine to be more powerful than honey-bee honey for use as a “natural” cure fortreating common diseases (Vit 2001).
The yield of honey from a colony rarelyexceeds 1 to 2 litres. It is sold on marketand is more expensive than Apis honeymainly due to its medicinal value and alsothe labour intensiveness of its harvestingprocess.
The purpose of our present in vitro in-vestigation is to elucidate if the Trigona
38

honeys collected from Ethiopia haveantimicrobial effects against different bacte-rial and fungal strains.
MATERIALS AND METHODSHoney source
Two samples of Trigona spp. honeywere obtained from two regions in Ethiopia,(I) Honey B from Bahir Dar (11° 35’ N and37° 28’ E) Northwest Ethiopia, at an alti-tude of 1830 m above sea level; (II) HoneyT from Temben (13° 53’ N and 39° 53’E)Northern Ethiopia, at an altitude of 1500 mabove sea level. The vegetation in both re-gions is categorized as Ethiopian undiffer-entiated woodland (White 1983). Honeysamples were bought from Tazma expertsand stored in a refrigerator, for one year,until needed for laboratory assay.
For comparison of the activity ofTrigona honey with that of honeybeehoney, a sample of honey was obtainedfrom the research beehives of the Instituteof Zoology, Free University of Berlin andnamed here as Honey D.
The pH of all the three honey sampleswas measured using a WTW Multi 340i pHmeter (Wissenschaftlich-Technische Werk-stätten GmbH, Weilheim, Germany) afterdiluting them 1:1 with distilled water.
Biological material
Bioassays of the antimicrobial activitiesof the different honey samples were per-formed using (I) four fungal species: theyeast Saccharomyces cerevisiae (DSM211) and three filamentous fungi,Aspergillus niger (DSM 737), Penicilliumchrysogenum (DSM 844) and Trichodermaviride (DSM 63065); (II) four species ofGram positive bacteria: Bacillus subtilis(DSM 347), Micrococcus luteus (DSM348), Bacillus megaterium (DSM 90), Ba-cillus brevis (DSM 5609); and (III) twospecies of Gram negative bacteria: Esche-richia coli (DSM 31), Pseudomonassyringae (DSM 5176).
Growth media
Cultivation of all bacterial cultures wasdone on Standard I nutrient broth (MerckLot VL 630582) and/or Standard I nutrientagar (Merck, Lot VL 694681). The yeastwas cultivated on a medium composed of10 g Oxoid agar (Agar BacteriologicalNo. 1, Lot 817706-2), 1 g yeast extract(Sigma, CAS No. 8013-01-2), 2 g glucose(Merck), 0.5 g peptone (Sigma, Lot128H0184), 10 ml Na-phosphate buffer(1M, pH 7) in 1 l distilled water. This samemedium without agar was used as a nutrientbroth for the yeast.
Trichoderma viride was cultivated onmalt extract peptone agar (MEPA) com-posed of 30 g malt extract (Sigma, Lot41k0181), 3 g soya peptone (Hy soy T,Sigma, Lot 128 H0184), 15 g Oxoid agar(Agar Bacteriological No. 1, Lot 817706-2)in 1 l distilled water. Aspergillus niger andPenicillium chrysogenum were cultivatedon potato dextrose agar (PDA) composed of20 g glucose (Merck), 15 g Oxoid agar(Agar Bacteriological No.1, Lot 817706-2),in 1 l potato infusion obtained by boiling200 g scrubbed and sliced potato in 1 l wa-ter for 1h and passed through a fine sieve.
All types of media were sterilized byautoclaving at 121°C and 15 psi for 15 min.
Bioassay
Preparation of the honey samplesThe different honey samples mentioned
were serially diluted to 1:1 (50%), 1:5(20%), 1:10 (10%), 1:20 (5%), 1:50 (2%),and 1:100 (1%), with distilled water andused for the determination of totalantimicrobial activity. The totalantimicrobial activity of the honey samplesis assigned the letter T followed by a sub-script of the letter for the name of the corre-sponding honey samples B, T, D for honeysfrom Bahir Dar, Temben, and Germany, re-spectively. Therefore the total antimicrobialactivity of these honey samples is hereafterreferred to as TB, TT, TD. By analogy the
Vol. 47 No. 1 2003 Journal of Apicultural Science 39

corresponding non-peroxide (NP)antimicrobial activities of these honey sam-ples are hereafter referred to as NPB, NPT,NPD, respectively.
The honey samples were treated withcatalase to remove H2O2 and to elucidatetheir non-peroxide activity. Treatment wasdone by a 1:1 mixing of the honey samplewith catalase solution of 20,000 units ml-1
modified slightly after Molan and Russell(1988). The catalase solution was preparedby dissolving a 13,300 units/mg solidcatalase from bovine liver (Sigma, Lot 32k7031, E.C. 1. 11. 1. 6) in distilled water. Thecatalase treated honey, which is diluted 1:1due to the treatment, was diluted further to1:5, 1:10, 1:20, and 1:50 and used in thePetridish bioassay.
Petridish Bioassay
Bacteria and yeastAn isolated pure colony of an overnight
grown strain was picked carefully using asterile transfer loop, inoculated to a nutrientbroth in an Erlenmeyer flask and grownovernight at 30°C. About 50 µl of the over-night culture was inoculated to a 20 ml solu-tion of nutrient broth and grown further forabout 3 to 5 h until an O.D. of 0.6 (546 nm)was achieved (method slightly modified af-ter Faye and Wyatt 1980). The suspensionwas then diluted 1:50 with the correspond-ing nutrient broth in order to prepare thestandard inoculum.
The sterilized nutrient agar was cooled to48°C, 5 ml of the standard inoculum wasmixed with the 1 l nutrient agar and pouredin to plastic Petridish of � = 85 mm, 10 mlin each. When the agar was solidified 3holes were bored per Petridish using a corkborer of � = 9 mm. Each hole was thenfilled with 50 µl honey of a certain dilutionand the Petridishes were placed in a refrig-erator for 24 h, giving the honey enoughtime to diffuse. Finally, the plates were re-moved from the refrigerator, incubated at30°C for 24 to 48 h and the inhibition zones
were measured. Each concentration of allhoney samples was investigated in triplePetridishes with three holes per Petridish,i.e., n = 9.
Filamentous fungiAn isolated pure colony of fungal cul-
ture which was grown for 72 h on solid me-dium and started to sporulate was scrubbedup using a sterile transfer loop and placedin 5 ml sterile distilled water in a test tube.The hyphae were disintegrated by addingsterile glass beads and shaking vigorouslyfor 1 to 2 min, in order to get a uniformsuspension. The 5 ml suspension wasadded to a 1 l agar solution at 48°C and theprocedure above was followed further. Theplates were taken out of the refrigerator andleft at room temperature (ca. 25°C) for 72 hand the inhibition zones were measured.
The control experiments for both thebacterial and fungal cultures were done byinoculating the cultures with sugar syrup(Lyle’s Golden syrup, UK) of dilutionssimilar to the honey samples. Sugar syruphas similar osmolarity to honey (Efem etal. 1992) and could reveal if the osmolarityof honey plays a significant role in thebacteriostatic/bactericidal activity of honey.The control experiments in the case of thenon-peroxide activity tests were done usinga catalase solution of 20,000 units ml-1 indistilled water. Each concentration of allhoney samples was investigated in triplePetridishes with three holes per Petridish,i.e., n = 9.
Turbidimetric bioassay
Though the agar well diffusion tech-nique is convenient to use, it is less sensi-tive than the turbidimetric method indisplaying the activity of an antimicrobialagent. The main reasons for its insensitivityis that the honey sample is further dilutedby the agar medium surrounding the welland that the extent of the clear zone ob-tained is not directly proportional to the ac-tivity of the sample, because outlying
40

colonies could grow before an inhibitoryconcentration diffuses to them (James etal. 1972, Molan et al. 1988). In contrast tothe agar well diffusion technique that dis-plays cumulative result of incubation after24 or 48 h, the turbidimetric method allowsfor kinetic investigations and displays theactivity of the antimicrobial agents with in-cubation period “live”. It demonstrates theactivity of the antimicrobial agents that maynot completely inhibit growth but prolongthe cell cycle time and hence could not beobserved by the Petridish bioassay method.For this reason the antimicrobial activitiesof the two stingless bee honeys were addi-tionally investigated using this method.Since the 1:1 and 1:5 diluted honey sampleswere too turbid, it was impossible to inves-tigate their antibacterial activities spectro-photometrically.
An actively growing bacterial culturewas diluted with nutrient broth to an O.D. == 0.6 (546 nm) and 100 µl of the suspensionwas added to an Erlenmeyer flask with sidenose (for O.D. measurement) that contains10 ml nutrient broth with 1:10, 1:20, 1:50,or 1:100 diluted honey. The culture wasthen incubated at 30°C by measuring theO.D. at 2 h intervals for 6 to 8 h. Nutrientbroth with the corresponding dilution ofhoney was used for blanking thespectrophotometer during each measure-ment. The control experiments were bacte-rial cultures with no honey and alsobacterial cultures inoculated with sugarsyrup/golden syrup with concentrations cor-responding to that of the honey samples.Each concentration was investigated sixtimes (n = 6).
Statistical analysis
Results are presented as mean ± S.D.values. Statistical tests were performed us-ing the two-tailed student’s test, Tukey’stest, two-way ANOVA and � = 0.05 wasconsidered as the critical value.
RESULTSThe two honey samples from Trigona
species were darker than the honey fromApis mellifera and also more acidic, withpH values of 3.2 and 3.45 for honeys Band T, respectively, compared to the higherpH of 4.9 for honey D.
The control experiments with sugarsyrup did not show any inhibitory action onany bacterial or fungal species tested exceptfor B. brevis, where a remarkable inhibitionzone was observed at 100% and 50% con-centration with inhibition zone diameters of12.3±0.3 and 10.7±3.5 mm, respectively.
The responses of a specific microorgan-ism to the different honey samples (Tukey’stest) and to the two categories (total- andnon-peroxide activities) of the same honeysample (two-tailed t-test) were significantlydifferent in some cases and not in others, asseen in Table 1. The total activities of bothhoneys B and T (TB and TT) were superiorto the non-peroxide activities (NPB andNPT) against the bacteria B. brevis, M.luteus, and E. coli at all concentrationstested, though the activities of only 50%concentrations of the different honey sam-ples are displayed in Table 1. In addition tothat 50% TB exhibited stronger antibacterialactivity against B. megaterium contrary toNPB which encouraged lawn growth of thisbacterium. There was no significant differ-ence (two-tailed t-test) in the antibacterialactivity of 50% TT and NPT againstB. megaterium, both showing weak inhibi-tory activities. Micrococcus luteus, thoughinhibited with 50% TB and TT, showeda very dense microbial lawn due to incuba-tion with the corresponding concentrationsof hydrogen peroxide devoid samples, i.e.,50% NPB and NPT. Escherichia coli wasinhibited strongly with 50% concentrationsof hydrogen peroxide containing honeys Band T, but not with hydrogen peroxide freehoney samples. Though TB and NPB
showed slight differences in the strength of
Vol. 47 No. 1 2003 Journal of Apicultural Science 41
42
TB NPb TT NPT TD NPD
B. brevis 21.1±0.7a 13.3±1.5a 16.2±1.7a 7.0±1.0a 7.3±2.3a 10.3±2.1a
B. megaterium 11.0±0.8b l.g. 3.0±0.4b 5.3±1.2a 7.0±1.0a 6.0±1.0a
B. subtilis 14.8±3.5b 20.7±5.0a 12.4±0.9a 29.0±3.6b 42.3±9.3b 46.7±4.2b
M. luteus 10.8±1.1b l.g. 33.9±3.0c l.g. 42.0±2.0b 41.7±2.1b
E. coli 13.1±0.8b 0.0 27.6±7.7c 0.0 21.7±2.9c 20.3±0.6c
P. syringae 12.5±0.5b 15.7±6.4a 4.1±0.0b 7.3±2.3a 7.2±0.8a 10.0±2.0a
S. cerevisae 0.0 0.0 0.0 0.0 0.0 0.0
A. niger 5.0±1.0c 4.7±1.2b 0.0 0.0 0.0 0.0
P. chrysogenum 3.7±0.6c 4.0±1.0b 0.0 0.0 0.0 0.0
T. viride 0.0 0.0 0.0 0.0 0.0 0.0
T a b l e 2Comparison of responses of the different species of bacteria and fungi to treatment with
50% dilution of each honey sample represented by the mean diameter ± S.D. of theinhibition zone in mm. Statistical significance test was done across species (along rows)
using One way ANOVA and the Tukey's test (� = 0.05, n = 9) for each honey sample.Values with in a row that possess the same letter do not have significant differences.
Identical letters at the values along columns do not designate any statistical relation. l. g.denotes the promotion of lawn of growth
Organism TB TT TD NPB NPT NPD
B. brevis 21.1±0.7a 16.2±1.7ab 7.3±2.3c 13.3±1.5b 7.0±1.0c 10.3±2.1cb
B. megaterium 11.0±0.8a 3.0±0.4b 7.0±1.0b l.g. 5.3±1.2b 6.0±1.0b
B. subtilis 14.8±3.5a 12.4±0.9a 42.3±9.3b 20.7±5.0a 29.0±3.6b 46.7±4.2b
M. luteus 10.8±1.1a 33.9±3.0b 42.0±2.0b l.g. l.g. 41.7±2.1b
E. coli 13.1±0.8a 27.6±7.7c 21.7±2.9c 0.0 0.0 20.3±0.6c
P. syringae 12.5±0.5a 4.1±0.0b 7.2±0.8c 15.7±6.4a 7.3±2.3c 10.0±2.0ac
S. cerevisae 0.0 0.0 0.0 0.0 0.0 0.0
A. niger 5.0±1.0a 0.0 0.0 4.7±1.2a 0.0 0.0
P. chrysogenum 3.7±0.6a 0.0 0.0 4.0±1.0a 0.0 0.0
T. viride 0.0 0.0 0.0 0.0 0.0 0.0
T a b l e 1Antimicrobial activity of 50% dilution of different honey samples elucidated by the
inhibition zone diameter (mm), mean ±S.D. First letters denote the total activity (T) ornon-peroxide activity (NP) and the subscripts indicate the origin of each sample; B: BahirDar, T: Temben (both from Ethiopia), D: Germany. One way ANOVA and Tukey's tests
(� = 0.05, n = 9) were performed across honey samples for each microbial species (along arow but not a column). Values within a row that possess the same letter do not have
significant difference. l.g. denotes the promotion of lawn of growth
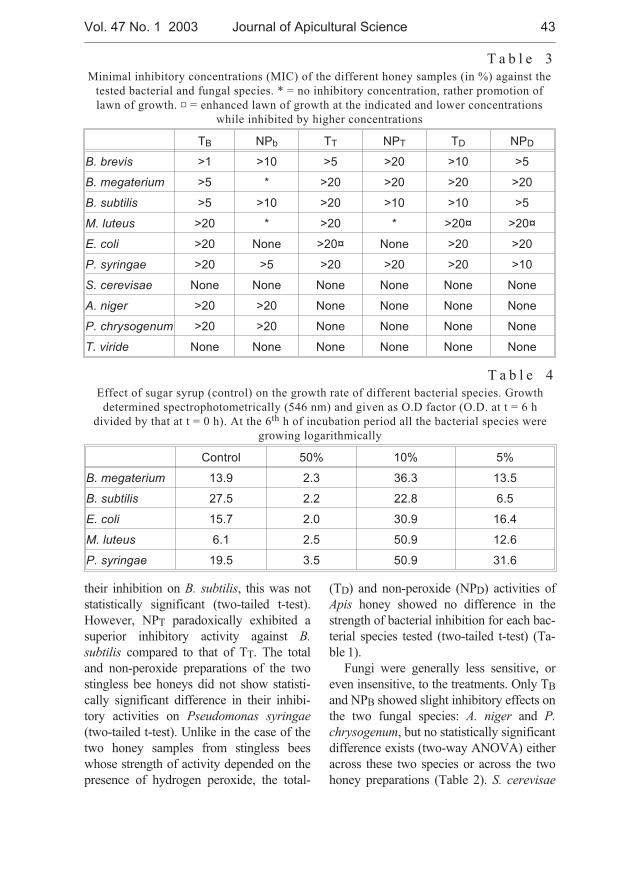
their inhibition on B. subtilis, this was notstatistically significant (two-tailed t-test).However, NPT paradoxically exhibited asuperior inhibitory activity against B.subtilis compared to that of TT. The totaland non-peroxide preparations of the twostingless bee honeys did not show statisti-cally significant difference in their inhibi-tory activities on Pseudomonas syringae(two-tailed t-test). Unlike in the case of thetwo honey samples from stingless beeswhose strength of activity depended on thepresence of hydrogen peroxide, the total-
(TD) and non-peroxide (NPD) activities ofApis honey showed no difference in thestrength of bacterial inhibition for each bac-terial species tested (two-tailed t-test) (Ta-ble 1).
Fungi were generally less sensitive, oreven insensitive, to the treatments. Only TB
and NPB showed slight inhibitory effects onthe two fungal species: A. niger and P.chrysogenum, but no statistically significantdifference exists (two-way ANOVA) eitheracross these two species or across the twohoney preparations (Table 2). S. cerevisae
Vol. 47 No. 1 2003 Journal of Apicultural Science 43
Control 50% 10% 5%
B. megaterium 13.9 2.3 36.3 13.5
B. subtilis 27.5 2.2 22.8 6.5
E. coli 15.7 2.0 30.9 16.4
M. luteus 6.1 2.5 50.9 12.6
P. syringae 19.5 3.5 50.9 31.6
T a b l e 4Effect of sugar syrup (control) on the growth rate of different bacterial species. Growth
determined spectrophotometrically (546 nm) and given as O.D factor (O.D. at t = 6 hdivided by that at t = 0 h). At the 6th h of incubation period all the bacterial species were
growing logarithmically
TB NPb TT NPT TD NPD
B. brevis >1 >10 >5 >20 >10 >5
B. megaterium >5 * >20 >20 >20 >20
B. subtilis >5 >10 >20 >10 >10 >5
M. luteus >20 * >20 * >20¤ >20¤
E. coli >20 None >20¤ None >20 >20
P. syringae >20 >5 >20 >20 >20 >10
S. cerevisae None None None None None None
A. niger >20 >20 None None None None
P. chrysogenum >20 >20 None None None None
T. viride None None None None None None
T a b l e 3Minimal inhibitory concentrations (MIC) of the different honey samples (in %) against the
tested bacterial and fungal species. * = no inhibitory concentration, rather promotion oflawn of growth. ¤ = enhanced lawn of growth at the indicated and lower concentrations
while inhibited by higher concentrations
and T. viride are insensitive to any of thehoney samples. The response of the differ-ent bacterial species to the treatment with acertain honey sample is not dependent onthe Gram reaction of the bacteria, both theGram positive and Gram negative bacteriaresponding to the treatments. The responsesof the bacterial species are rather dependent
on the type of honey sample and individualorganisms within a category, Gram positiveor Gram negative, rather than the Gram re-action (Tables 1 and 2).
In case of the three species of Bacillusand M. luteus there were dense bacterialgrowth zones next to the inhibition zoneswith the honey samples B and T even at a
44
Fig. 2 The effect of stingless bee honey (Honey T) on the growth rate of different bacterial speciesdetermined spectrophotometrically. The OD factor is obtained by dividing the OD at tt withthe OD at t0. The different symbols represent different concentrations of honey
Fig. 1 The effect of stingless bee honey (Honey B) on the growth rate of different bacterialspecies determined spectrophotometrically. The OD factor is obtained by dividing theOD at tt with the OD at t0. The different symbols represent honey concentrations
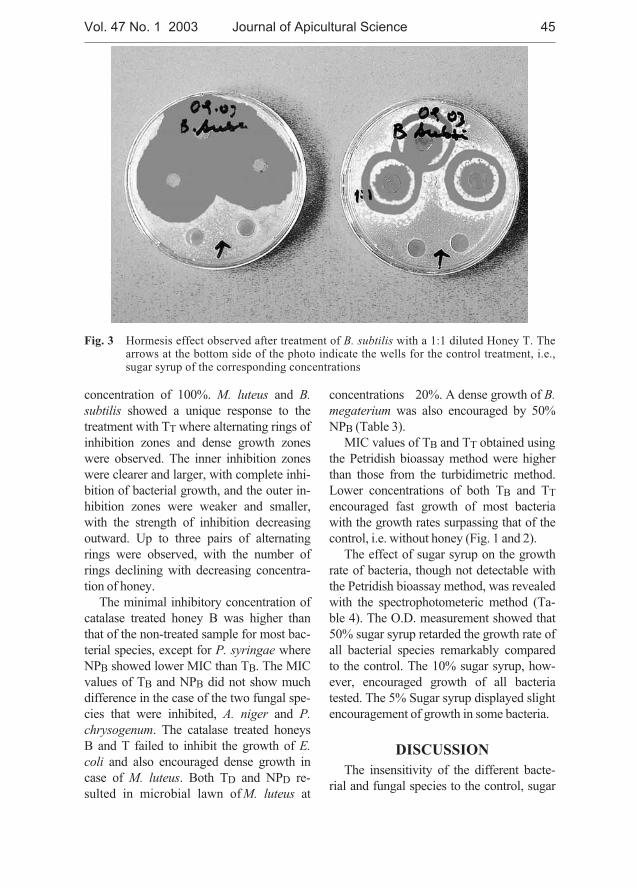
concentration of 100%. M. luteus and B.subtilis showed a unique response to thetreatment with TT where alternating rings ofinhibition zones and dense growth zoneswere observed. The inner inhibition zoneswere clearer and larger, with complete inhi-bition of bacterial growth, and the outer in-hibition zones were weaker and smaller,with the strength of inhibition decreasingoutward. Up to three pairs of alternatingrings were observed, with the number ofrings declining with decreasing concentra-tion of honey.
The minimal inhibitory concentration ofcatalase treated honey B was higher thanthat of the non-treated sample for most bac-terial species, except for P. syringae whereNPB showed lower MIC than TB. The MICvalues of TB and NPB did not show muchdifference in the case of the two fungal spe-cies that were inhibited, A. niger and P.chrysogenum. The catalase treated honeysB and T failed to inhibit the growth of E.coli and also encouraged dense growth incase of M. luteus. Both TD and NPD re-sulted in microbial lawn of M. luteus at
concentrations 20%. A dense growth of B.megaterium was also encouraged by 50%NPB (Table 3).
MIC values of TB and TT obtained usingthe Petridish bioassay method were higherthan those from the turbidimetric method.Lower concentrations of both TB and TT
encouraged fast growth of most bacteriawith the growth rates surpassing that of thecontrol, i.e. without honey (Fig. 1 and 2).
The effect of sugar syrup on the growthrate of bacteria, though not detectable withthe Petridish bioassay method, was revealedwith the spectrophotometeric method (Ta-ble 4). The O.D. measurement showed that50% sugar syrup retarded the growth rate ofall bacterial species remarkably comparedto the control. The 10% sugar syrup, how-ever, encouraged growth of all bacteriatested. The 5% Sugar syrup displayed slightencouragement of growth in some bacteria.
DISCUSSIONThe insensitivity of the different bacte-
rial and fungal species to the control, sugar
Vol. 47 No. 1 2003 Journal of Apicultural Science 45
Fig. 3 Hormesis effect observed after treatment of B. subtilis with a 1:1 diluted Honey T. Thearrows at the bottom side of the photo indicate the wells for the control treatment, i.e.,sugar syrup of the corresponding concentrations

syrup, is an indication that the osmolarity ofhoney does not play an important role in theinhibition of growth of these microorgan-isms, but higher concentrations may pro-long the cell cycle time. Hence, the otherantibacterial factors of honey, i.e. hydrogenperoxide activity, non-peroxide activitymainly due to the flavonoids of honey, acid-ity, and lysozyme (Bogdanov andBlumer 2001) could be responsible for theinhibition of growth of these microorgan-isms. Bacillus brevis, however, is sensitiveto and inhibited by the high osmolarity ofsugar syrup indicating that, apart from oth-ers, the high osmolarity of honey could in-hibit the growth of this bacterium.
The stronger total antibacterial activitiesof stingless bee honeys against some bacte-rial species compared to the non-peroxideactivities are indications that hydrogen per-oxide plays a significant role in the bacteri-cidal/bacteriostatic action of these honeysamples. This may be due to the fact thathoney from stingless bees possesses consid-erable amount of hydrogen peroxide thatcould be, at least partially, responsible forpreventing the fermentation of this honeywhich otherwise would have taken placedue to its higher amount of water, 30 to35%. Catalase treated honey B resulted inan enhancement of growth of B. mega-terium and M. luteus though these bacteriawere strongly inhibited by the non-treatedhoney samples. The most likely explanationfor this could be that they are inhibited byhydrogen peroxide in honey in case of thetotal activity, and that the non-peroxidecomponents are below inhibitory concentra-tions. Removal of hydrogen peroxide elimi-nates the inhibitory factor and hencebacterial growth could be encouraged eitherdue to the higher sugar concentration, pro-viding them with excess energy and nutri-tional sources, or the lower concentration ofthe phytochemical components below a cer-tain critical level. The presence of certainbacteriostatic/bactericidal chemicals in the
growth medium at a concentration lowerthan a critical inhibitory level enhances thegrowth of the organism that otherwisewould have been inhibited by higher con-centrations, a phenomenon known ashormesis (Edward et al. 1998). This effectof hormesis was also clearly observed in thecase of treatment of M. luteus with 50%NPT, though 50% TT showed a very stronginhibitory action with an inhibition zone ofnearly 34 mm diameter. The activities ofboth types of stingless bee honeys againstE. coli were only due to the presence of hy-drogen peroxide, whose removal resulted inthe normal growth of this bacterium withoutany detectable inhibition zone.
The formation of concentric rings of in-hibition- and dense growth zones could alsobe explained based on the phenomenon ofhormesis, with the different components ofhoney (mainly the phytochemical compo-nents with varying diffusion potentials) act-ing as hormesis factors in the differentzones.
Both stingless bee honeys showed ahigher non-peroxide activity against thebacterium B. subtilis than total activity,though it was statistically insignificant forhoney B. This is in contrast to most litera-ture and difficult to explain. It was, how-ever, pointed out by Burdon (1995) thathydrogen peroxide, at a concentration be-low a certain critical level, could stimulatethe division and proliferation of many celltypes in mammalian tissue and plays a rolein wound healing. This chemical acts at var-ious points in the mechanisms that controlcell growth and differentiation probably byoxidising proteins involved, and thus caus-ing a change in the conformation of the pro-tein molecules. Apart from this evidence onmammalian cells, a positive role of hydro-gen peroxide supporting the growth of bac-teria was not yet reported.
The comparable antimicrobial activity ofcatalase treated and non-treated samples ofhoneys T and B on some bacteria
46

demonstrates that the activities are not dueto hydrogen peroxide. The lack of hydrogenperoxide based activity and the insensitivityto sugar syrup demonstrates that the activityof stingless bee honeys could be mainly dueto the phytochemical (flavonoids),lysozyme, organic acid components(Bogdanov and Blumer 2001) and prob-ably due to its strong acidity. Conclusiveevidence has been demonstrated by Molanand Russell (1988) that the antibacterialactivity of some New Zealand honeys wasnot only due to hydrogen peroxide butmainly to the antibacterial agents of plantorigin. The authors also concluded that inhoneys of higher antibacterial activities,non-peroxide activity is by far the largestpart of the total activity.
The Apis honey tested did not show hy-drogen peroxide based activity. One possi-ble reason could be that the honey was ripesince it was stored for one year. Storage ofthe honey for one year was important sinceits activity had to be compared withTrigona honey that was also stored for oneyear. The enzyme glucose oxidase is practi-cally inactive in ripe honey, no more re-plenishing the degraded hydrogen peroxideand hence lower concentration of the latterwhich hardly inhibits bacteria (Bogdanovand Blumer 2001, http://privatewww.essex.ac.uk/~islamic/ilm/health-fit/honey_2.html).This is because the acidity produced in theaction of the enzyme drops the pH to a valuetoo low for the enzyme to work any more. Inaddition to that the activity of water (aw) of aripe honey is very low inhibiting the process(Bogdanov and Blumer 2001). The lackhydrogen peroxide activity demonstrates thatthe antibacterial activity of this honey ismainly due to the phytochemical compo-nents, or lysozyme. The acidity of this honey(pH = 4.9) is not strong enough to inhibit thegrowth of most bacteria as it is further di-luted and weakened in the growth medium.Osmolarity also does not play a role sincethe bacteria tested did not respond to the
osmolarity of sugar syrup, except B. brevis.The pH and osmolarity of different NewZealand honeys tested against various or-ganisms did not inhibit bacterial growth(Molan and Russel 1988).
CONCLUSIONSThe traditional use of Trigona honey as a
panacea against different illnesses is ratio-nal if the infection to be treated is caused bybacteria, not by fungi, since only one of thehoney samples showed a minor antifungaleffect against two of the four fungal speciestested, though reports exist that honey in-hibits the growth of fungi.
The enhancement of growth of somebacterial species by lower concentrations ofhoney indicates that the use of honey atlower concentrations as a means ofapitherapy could be dangerous. However, ifused undiluted, Trigona honey offers manypossibilities as a broad spectrum-healingagent against both Gram positive and Gramnegative bacteria. Especially the use of thishoney as a wound dressing agent may beeffective since its dilution by body fluid isless intensive than for internal administra-tion.
Though it is premature to conclude atthis level that stingless bee honey can beused as a panacea, as claimed by the localpeople, it can be ascertained that it has abroad spectrum of antimicrobial action in-hibiting the growth of both Gram positiveand Gram negative bacteria. Further re-search in this field may help to recognizeand use indigenous knowledge to help alle-viate the ever-increasing report of resistanceof pathogenic bacteria to current antibiotics.
ACKNOWLEDGMENTSWe would like to express our gratitude
to Dr Lars Podsiadlowski, Mrs. Beate Bach,Mrs. Christine Bergmann and Mrs. GudrunWelge for their support and advice with thelaboratory work. We also thank the DAAD
Vol. 47 No. 1 2003 Journal of Apicultural Science 47

(Deutscher Akademischer Austauschdienst)for the financial support of A.G.
REFERENCESAmano K., Nemoto T., Heard T. A.
(2000) - What are stingless bee, and whyand how to use them as crop pollinators? Areview. Japan Agricul. Res. Quarterly, 34(3).
Bogdanov S., Blumer P. (2001) -Natürliche antibiotische Eigenschaften desHonigs. Schweizerische Bienen Zeitung,124(2):18-21.
Calabrese E. J. and Baldwin L. A.(1998) - Hormesis as a BiologicalHypothesis. Environ. Health Perspectives106, Supplement 1.
Crane E. (1992) - The past and presentstatus of beekeeping with stingless bees.Bee World, 73:29-42
Efem S.E.E., Udoh K.T., Iwara C.I.(1992) - The antimicrobial spectrum ofhoney and its clinical significance.Infection, 20(4):227-229.
Faye I.; Wyatt GR. (1980) - Thesynthesis of antibacterial proteins inisolated fat body from Cecropia silkmothpupae. Experientia, 36:1325
Fichtl R., Adi A. (1994) - HoneybeeFlora of Ethiopia. Weikersheim. MargrafVerlag, pp. 510.
Greenwood D. (1995) - Sixty years on:antimicrobial drug resistance comes of age.Lancet 346 (supplement 1).
- http://privatewww.essex.ac.uk/~islamic/ilm/health-fit/honey_2.html
- http://www.sibexlink.com.my/g15magazine/g15mag_vol3jan_science.html
James O.B.O’L, Segroe W., VenturaA.K. (1972) - Some antibacterialproperties of Jamaican honey. W. IndianMed. J., 21:7-17.
Kauffman G.B. (1991) - ChemophobiaChemistry in Britain. June 512-516.
Krell R. (1996) - Value added productsfrom Beekeeping. FAO Agricul. Serv. Bull.No.24.
Michener C.D. (1974) - The SocialBehaviour of the Bees; a ComparativeStudy. Cambridge, Mass, Harvard Univ.Press, pp. 404
Molan P. (2001) - Why honey is effectiveas a medicine, II. The scientific explanationof its effects. Bee World, 82 (1):22-40.
Molan P., Brett M. (1998) - Honey has apotential as a dressing for wounds infectedwith MRSA. The second Australian woundmanagement association conference.Brisbane, Australia.
Molan P.C., Russell K.M. (1988) -Non-peroxide antibacterial activity in someNew Zealand honeys. J. apic. Res.,27(1):62-67.
Molan P.C., Smith I.M. Reid G.M.(1988) - A comparison of theantibacterial activities of some NewZealand honey. J. api. Res., 27(4):252-256.
Ruttner F. (1992) - Naturgeschichte derHonigbienen. Ehrenwirth Verlag München,pp. 357.
Thompson W.A.R. (1976) - Herbs thatheal. Journal of the Royal College ofGeneral Practitioners, 26:365-370.
Wille A. (1979) - Phylogeny andrelationships among the genera andsubgenera of the Stingless bees(Meliponinae) of the world. Revisita deBiologia Tropical, 27(2):241-277.
White F. (1983) - The vegetation ofAfrica. A descriptive memo to accompanythe Unesc/AETFAT/UNSO Vegetationmap of Africa. United nations. UNESCO,Switzerland, pp. 356.
Vit P. (2001) - Stingless bee honey and thetreatment of cataracts, in: Munn P. andJones R. (eds.) Honey and healing, IBRA,Cardiff UK, pp. 49.
48

AKTYWNOŒÆ PRZECIWDROBNOUSTROJOWAMIODU PSZCZÓ£ BEZ¯¥D£YCH Trigona spp.
G a r e d e w A . , S c h m o l z E . , L a m p r e c h t I .
S t r e s z c z e n i e
Pszczo³y bez¿¹d³e (Trigona spp.) wytwarzaj¹ charakterystyczny miód o niskiej lepkoœci,zdecydowanie bardziej ciemny i kwaœny w stosunku do miodu pszczo³y miodnej. W Etiopiimiód Trigona spp. jest dro¿szy ze wzglêdu na jego zastosowanie jako panaceum na wieleschorzeñ.
W pracy oznaczono aktywnoœæ antymikrobow¹ dwóch próbek miodu pszczó³ bez¿¹d³ychz Etiopii, i dla porównania, aktywnoœæ jednej próbki miodu pszczo³y miodnej z Niemiec.Aktywnoœæ miodu okreœlono testami in vitro w stosunku do czterech gatunków grzybówi szeœciu gatunków bakterii.
Grzyby generalnie s¹ du¿o mniej wra¿liwe na dzia³anie miodu ni¿ bakterie. Tylko Aspergillusniger i Penicillium chrysogenum w niewielkim stopniu reagowa³y na jedn¹ z dwóch próbekmiodu Trigona spp. i tylko przy jego wy¿szych stê¿eniach. Miód oddzia³ywa³ na bakterie bezzwi¹zku z ich reakcj¹ na test Grama. Minimalne stê¿enie miodu Trigona spp., które hamowa³orozwój bakterii (MIC), by³o wy¿sze po zadzia³aniu na miód katalaz¹, od tych próbek, które nieby³y traktowane katalaz¹ (nie zosta³ roz³o¿ony nadtlenek wodoru). Œwiadczy to o wp³ywieakumulacji nadtlenku wodoru na aktywnoœæ antymikrobow¹ tego miodu. W przypadku miodupszczo³y miodnej (próbka porównawcza miodu niemieckiego) akumulacja nadtlenku wodorunie odgrywa³a g³ównej roli w aktywnoœci antybakteryjnej w reakcji przeciw wiêkszoœci u¿ytychw doœwiadczeniu szczepów bakterii.
S³owa kluczowe: pszczo³y bez¿¹d³e, miód pszczó³ bez¿¹d³ych, aktywnoœæprzeciwdrobnoustrojowa, aktywnoœæ nie-nadtlenkowa Trigona spp.
Vol. 47 No. 1 2003 Journal of Apicultural Science 49

FLOWERING PHENOLOGY AND FERTILITYOF SOUR CHERRY (Prunus cerasus L.)CULTIVARS SELECTED IN HUNGARY
J ó z s e f N y é k i 1 , T i b o r S z a b ó 2 , Z o l t á n S z a b ó 1
1University of Debrecen, Centre of Agricultural Sciences,H-4032 Debrecen, Böszörményi út 138., Hungary,
Phone: (36) 52-508-477, e-mail: [email protected] and Extension Centre for Fruit Growing, H-4244 Újfehértó, Vadastag 2.
Phone: (36) 42-290-822, Fax: (36) 42-290-822
S u m m a r y
Experiments were conducted between 1972 and 2002 at three sites in Hungary. At Érd 97,Helvécia 10, and Újfehértó 3 cultivars of sour cherry were studied in variety collections.Observations were made on the flowering phenology (start, main time, end and length of thebloom period), on the flowering dynamics (the rate of the open flowers counted every day), onthe fruit set following self- and open-pollination and on the effect of association of varieties inthe orchards (choice, rate and placement of pollinisers).
Based on the results the rate of the overlap of the blooming times were calculated andvarieties were assigned into five bloom time groups according to their main bloom. Self-fertilityconditioned by natural self pollination was studied and good pollinisers were chosen (sweet,sour and duke cherry varieties) for the self-sterile and partially self-fertile varieties.
The necessity of bee pollination was proved by different pollination methods: naturalself-pollination, artificial self-pollination, open pollination.
Keywords: fertility, flowering, pollination, sour cherry.
INTRODUCTIONSour cherry production is an important
industry and it is also an export item inHungary. All except one of the registeredand available 22 varieties are of Hungarianorigin. The assortment is rather rich, more-over, regional selection and pedigree breed-ing promise currently new varieties.
Fertilisation in the Pándy meggy, thenleading variety, has concerned the growerssince the 19th century. Maliga (1942) sur-veyed the relevant literature. Self-fertilityand its relation to fruit set obtained by openpollination was assessed in several Hungar-ian and foreign varieties by Nyéki (1974).All variants of Pándy meggy proved to beself-incompatible, and mutually inter-in-compatible. Nyéki et al. (1992) called the
attention to the fact that the self-incompati-ble Pándy meggy cannot be fertilised byseveral self-fertile varieties (Debrecenibõtermõ, Kántorjánosi, Újfehértói fürtös), i.e.they are inter-incompatible.
Sugar content of nectaries is high in sourcherry flowers; thus honeybees are at-tracted. Earlier observations stated the ben-eficial effect of bee activity in both self--incompatible as well as self-fertile variet-ies. i.e. fruit set increased substantially(Benedek et al. 1990).
In the present paper first of all the vari-ability of fruit set depending on self- or al-ternatively open pollination in sour cherryvarieties was analysed.
The objectives of the studies were to de-termine the blooming time, self and open
Vol. 47 No. 1 2003 Journal of Apicultural Science 51

pollination of Hungarian sour cherryvarieties.
METHODSObservations of blooming dynamics fol-
lowed the method applied by Nyéki (1974)on flowers distributed to the four quarters ofthe heavens on the end of branches, a sam-ple comprising 100-500 flowers per variety.Starting with the beginning of bloom, everyday at the same time (between 10 and 12a.m.) the number of flowers according totheir stage of development from the budphase, opening and petal shedding wasregistered.
Flowers are considered to be open if theanthers and the pistil are easily recognisedfrom above, and the stigma is green or yel-low. The stigma of fading flowers startedbrowning. Those dynamic observationserved as a basis of determining the start ofblooming (with the first open flowers), themain blooming period (when more than50% of flowers opened) and the day ofmain bloom (when the number of openflowers reached the maximum) and the endof bloom (when the last flower faded).
According to the start of blooming vari-eties are grouped into 5 categories (early,medium early, intermediate, medium late,late).
Studies of fertilisation were done accord-ing to the method of Rudloff &Schanderl (1950).
Autogamy was investigated on 20-30 cmlong branches of different orientation (N, E,S, W) covered still at bud stage with parch-ment paper bags. 5-10 isolated units com-prising 100-400 flowers represented eachvariety. Paper bags were eliminated 3-4days after the blooming period ended. Theeffect of artificial self pollination orgeitonogamy was checked on isolatedbranches prepared as in the case ofautogamy, except that the fully open flow-ers were hand pollinated once with
previously collected and stored (at +4°C)pollen of the same variety, subsequently,the bags were restored until the end ofblooming period as in the case of autogamy.
Open pollinated flowers were observed,equally, on 5-10 branches 1.5-2 meterabove ground level of four different orienta-tions and comprising 100-500 flowers pervariety.
Fruit set was registered and compared inall treatments at 1-2 weeks before fruitripening.
RESULTS AND DISCUSSIONBlooming time
Sour cherry belongs to the group of me-dium early-blooming fruit trees but it is thelatest among the stone fruits. At the start ofblooming cultivated varieties cover a periodof 4-7 days, during some seasons excep-tionally also 10 days. Most authors distin-guish three blooming time groups in sourcherry varieties. Present observations de-rived from three growing sites, 101 variet-ies, and a long period of years (1972-2002),which allowed the establishment of fiveblooming time groups. In Table 1 the prop-agated varieties are shown separately fromthe most important Hungarian and foreignvarieties.
Neighbouring blooming time groups dif-fer by 1-2 days only, depending on the sea-son. In order to achieve considerable (morethan 20%) fruit set a self-incompatible vari-ety should be pollinated by a compatiblepolliniser, when their blooming period isoverlapping each other by 70%, at least. Ina system of 3 blooming time groups variet-ies belonging to the same group should becombined, whereas with 5 groups the over-lap also of the neighbouring groups mayprove to be sufficient for safe fruit set. Syn-chronous blooming or the extent of overlap-ping, however, may change yearly.
Variants of the variety Pándy differ inblooming time, thus Pándy 48 is medium
52
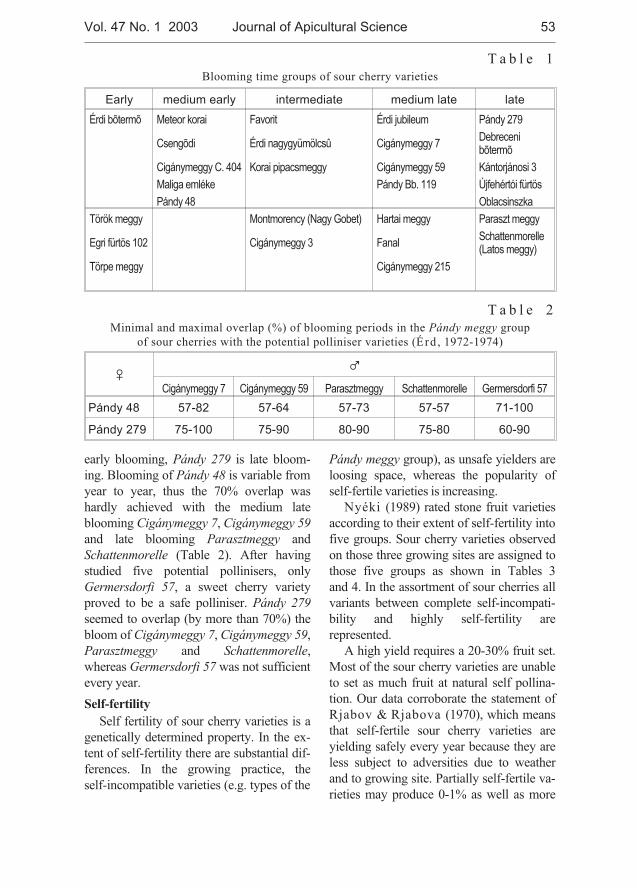
early blooming, Pándy 279 is late bloom-ing. Blooming of Pándy 48 is variable fromyear to year, thus the 70% overlap washardly achieved with the medium lateblooming Cigánymeggy 7, Cigánymeggy 59and late blooming Parasztmeggy andSchattenmorelle (Table 2). After havingstudied five potential pollinisers, onlyGermersdorfi 57, a sweet cherry varietyproved to be a safe polliniser. Pándy 279seemed to overlap (by more than 70%) thebloom of Cigánymeggy 7, Cigánymeggy 59,Parasztmeggy and Schattenmorelle,whereas Germersdorfi 57 was not sufficientevery year.
Self-fertilitySelf fertility of sour cherry varieties is a
genetically determined property. In the ex-tent of self-fertility there are substantial dif-ferences. In the growing practice, theself-incompatible varieties (e.g. types of the
Pándy meggy group), as unsafe yielders areloosing space, whereas the popularity ofself-fertile varieties is increasing.
Nyéki (1989) rated stone fruit varietiesaccording to their extent of self-fertility intofive groups. Sour cherry varieties observedon those three growing sites are assigned tothose five groups as shown in Tables 3and 4. In the assortment of sour cherries allvariants between complete self-incompati-bility and highly self-fertility arerepresented.
A high yield requires a 20-30% fruit set.Most of the sour cherry varieties are unableto set as much fruit at natural self pollina-tion. Our data corroborate the statement ofRjabov & Rjabova (1970), which meansthat self-fertile sour cherry varieties areyielding safely every year because they areless subject to adversities due to weatherand to growing site. Partially self-fertile va-rieties may produce 0-1% as well as more
Vol. 47 No. 1 2003 Journal of Apicultural Science 53
Early medium early intermediate medium late late
Érdi bõtermõ Meteor korai Favorit Érdi jubileum Pándy 279
Csengõdi Érdi nagygyümölcsû Cigánymeggy 7 Debrecenibõtermõ
Cigánymeggy C. 404 Korai pipacsmeggy Cigánymeggy 59 Kántorjánosi 3
Maliga emléke Pándy Bb. 119 Újfehértói fürtös
Pándy 48 Oblacsinszka
Török meggy Montmorency (Nagy Gobet) Hartai meggy Paraszt meggy
Egri fürtös 102 Cigánymeggy 3 Fanal Schattenmorelle(Latos meggy)
Törpe meggy Cigánymeggy 215
T a b l e 1Blooming time groups of sour cherry varieties
��
Cigánymeggy 7 Cigánymeggy 59 Parasztmeggy Schattenmorelle Germersdorfi 57
Pándy 48 57-82 57-64 57-73 57-57 71-100
Pándy 279 75-100 75-90 80-90 75-80 60-90
T a b l e 2Minimal and maximal overlap (%) of blooming periods in the Pándy meggy group
of sour cherries with the potential polliniser varieties (Érd , 1972-1974)
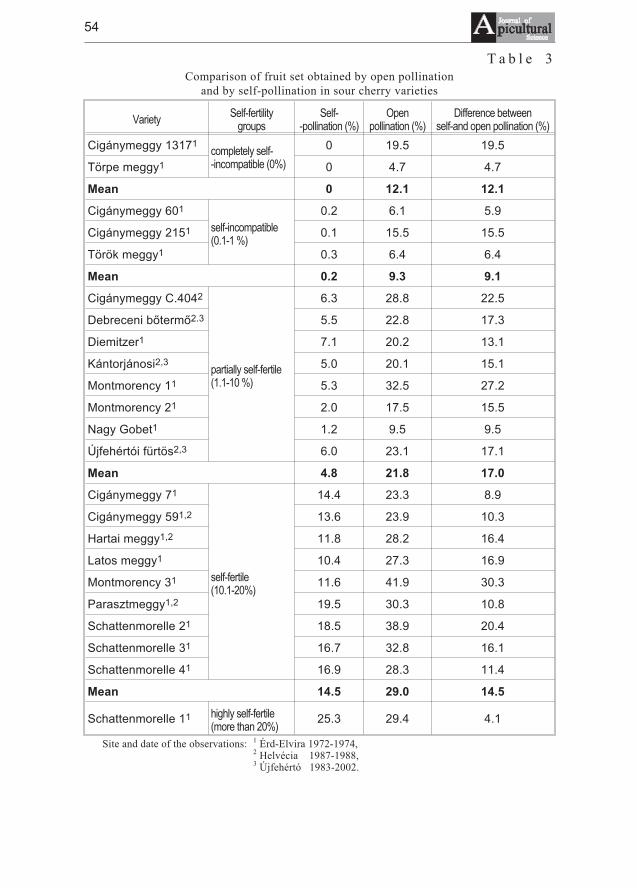
54
Variety Self-fertilitygroups
Self--pollination (%)
Openpollination (%)
Difference betweenself-and open pollination (%)
Cigánymeggy 13171completely self--incompatible (0%)
0 19.5 19.5
Törpe meggy1 0 4.7 4.7
Mean 0 12.1 12.1
Cigánymeggy 601
self-incompatible(0.1-1 %)
0.2 6.1 5.9
Cigánymeggy 2151 0.1 15.5 15.5
Török meggy1 0.3 6.4 6.4
Mean 0.2 9.3 9.1
Cigánymeggy C.4042
partially self-fertile(1.1-10 %)
6.3 28.8 22.5
Debreceni bõtermõ2.3 5.5 22.8 17.3
Diemitzer1 7.1 20.2 13.1
Kántorjánosi2,3 5.0 20.1 15.1
Montmorency 11 5.3 32.5 27.2
Montmorency 21 2.0 17.5 15.5
Nagy Gobet1 1.2 9.5 9.5
Újfehértói fürtös2,3 6.0 23.1 17.1
Mean 4.8 21.8 17.0
Cigánymeggy 71
self-fertile(10.1-20%)
14.4 23.3 8.9
Cigánymeggy 591,2 13.6 23.9 10.3
Hartai meggy1,2 11.8 28.2 16.4
Latos meggy1 10.4 27.3 16.9
Montmorency 31 11.6 41.9 30.3
Parasztmeggy1,2 19.5 30.3 10.8
Schattenmorelle 21 18.5 38.9 20.4
Schattenmorelle 31 16.7 32.8 16.1
Schattenmorelle 41 16.9 28.3 11.4
Mean 14.5 29.0 14.5
Schattenmorelle 11 highly self-fertile(more than 20%)
25.3 29.4 4.1
Site and date of the observations: 1 Érd-Elvira 1972-1974,2 Helvécia 1987-1988,3 Újfehértó 1983-2002.
T a b l e 3Comparison of fruit set obtained by open pollination
and by self-pollination in sour cherry varieties
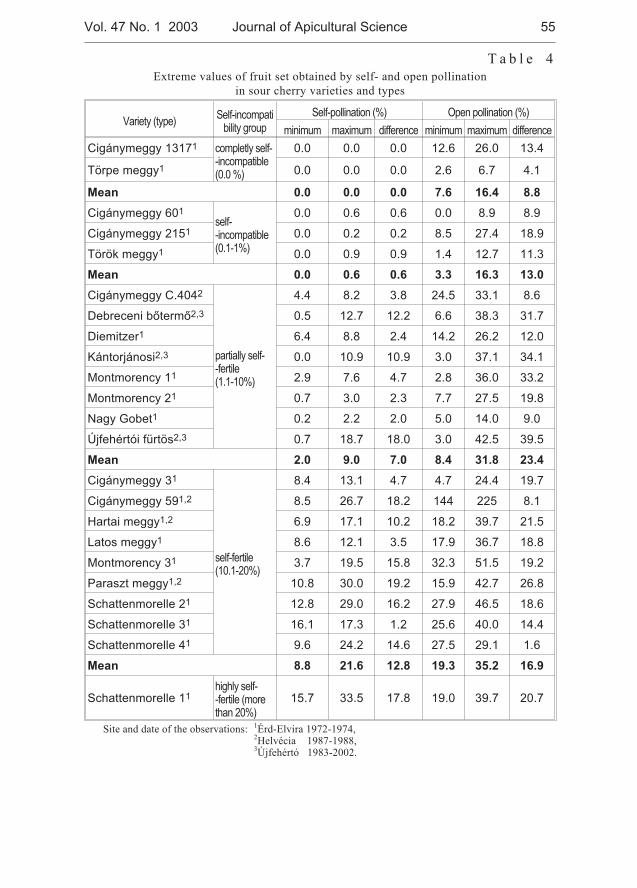
Vol. 47 No. 1 2003 Journal of Apicultural Science 55
Variety (type) Self-incompatibility group
Self-pollination (%) Open pollination (%)
minimum maximum difference minimum maximum difference
Cigánymeggy 13171 completly self--incompatible(0.0 %)
0.0 0.0 0.0 12.6 26.0 13.4
Törpe meggy1 0.0 0.0 0.0 2.6 6.7 4.1
Mean 0.0 0.0 0.0 7.6 16.4 8.8
Cigánymeggy 601self--incompatible(0.1-1%)
0.0 0.6 0.6 0.0 8.9 8.9
Cigánymeggy 2151 0.0 0.2 0.2 8.5 27.4 18.9
Török meggy1 0.0 0.9 0.9 1.4 12.7 11.3
Mean 0.0 0.6 0.6 3.3 16.3 13.0
Cigánymeggy C.4042
partially self--fertile(1.1-10%)
4.4 8.2 3.8 24.5 33.1 8.6
Debreceni bõtermõ2,3 0.5 12.7 12.2 6.6 38.3 31.7
Diemitzer1 6.4 8.8 2.4 14.2 26.2 12.0
Kántorjánosi2,3 0.0 10.9 10.9 3.0 37.1 34.1
Montmorency 11 2.9 7.6 4.7 2.8 36.0 33.2
Montmorency 21 0.7 3.0 2.3 7.7 27.5 19.8
Nagy Gobet1 0.2 2.2 2.0 5.0 14.0 9.0
Újfehértói fürtös2,3 0.7 18.7 18.0 3.0 42.5 39.5
Mean 2.0 9.0 7.0 8.4 31.8 23.4
Cigánymeggy 31
self-fertile(10.1-20%)
8.4 13.1 4.7 4.7 24.4 19.7
Cigánymeggy 591,2 8.5 26.7 18.2 144 225 8.1
Hartai meggy1,2 6.9 17.1 10.2 18.2 39.7 21.5
Latos meggy1 8.6 12.1 3.5 17.9 36.7 18.8
Montmorency 31 3.7 19.5 15.8 32.3 51.5 19.2
Paraszt meggy1,2 10.8 30.0 19.2 15.9 42.7 26.8
Schattenmorelle 21 12.8 29.0 16.2 27.9 46.5 18.6
Schattenmorelle 31 16.1 17.3 1.2 25.6 40.0 14.4
Schattenmorelle 41 9.6 24.2 14.6 27.5 29.1 1.6
Mean 8.8 21.6 12.8 19.3 35.2 16.9
Schattenmorelle 11highly self--fertile (morethan 20%)
15.7 33.5 17.8 19.0 39.7 20.7
Site and date of the observations: 1Érd-Elvira 1972-1974,2Helvécia 1987-1988,3Újfehértó 1983-2002.
T a b l e 4Extreme values of fruit set obtained by self- and open pollination
in sour cherry varieties and types
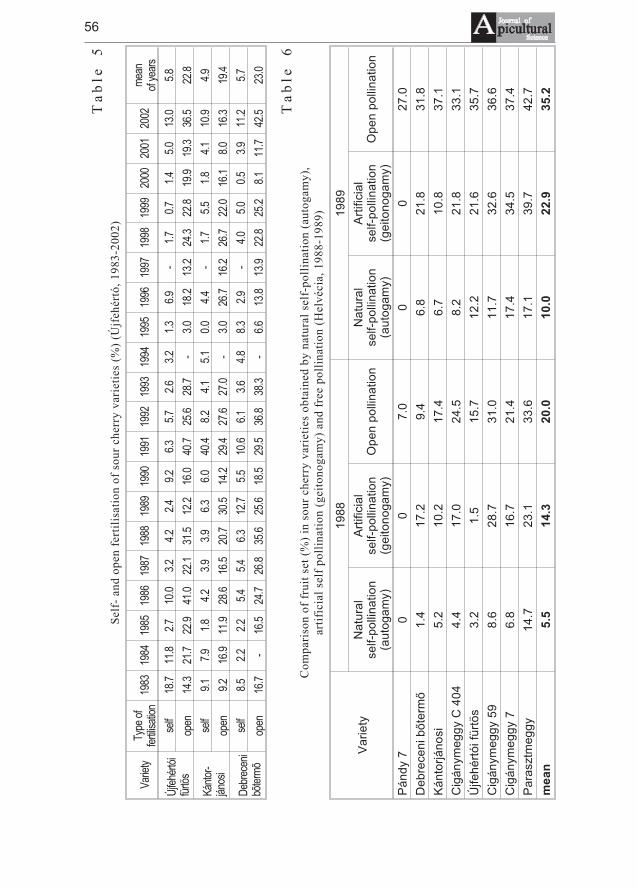
56
Ta
ble
5S
elf-
and
open
fert
ilis
atio
nof
sour
cher
ryva
riet
ies
(%)
(Újf
ehér
tó,
1983
-200
2)
Varie
tyTy
peof
ferti
lisat
ion
1983
1984
1985
1986
1987
1988
1989
1990
1991
1992
1993
1994
1995
1996
1997
1998
1999
2000
2001
2002
mea
nof
year
s
Újfe
hértó
ifü
rtös
self
18.7
11.8
2.7
10.0
3.2
4.2
2.4
9.2
6.3
5.7
2.6
3.2
1.3
6.9
-1.
70.
71.
45.
013
.05.
8
open
14.3
21.7
22.9
41.0
22.1
31.5
12.2
16.0
40.7
25.6
28.7
-3.
018
.213
.224
.322
.819
.919
.336
.522
.8
Kánt
or-
jáno
sise
lf9.
17.
91.
84.
23.
93.
96.
36.
040
.48.
24.
15.
10.
04.
4-
1.7
5.5
1.8
4.1
10.9
4.9
open
9.2
16.9
11.9
28.6
16.5
20.7
30.5
14.2
29.4
27.6
27.0
-3.
026
.716
.226
.722
.016
.18.
016
.319
.4
Deb
rece
nibõ
term
õse
lf8.
52.
22.
25.
45.
46.
312
.75.
510
.66.
13.
64.
88.
32.
9-
4.0
5.0
0.5
3.9
11.2
5.7
open
16.7
-16
.524
.726
.835
.625
.618
.529
.536
.838
.3-
6.6
13.8
13.9
22.8
25.2
8.1
11.7
42.5
23.0
Ta
ble
6C
ompa
riso
nof
frui
tse
t(%
)in
sour
cher
ryva
riet
ies
obta
ined
byna
tura
lse
lf-p
olli
nati
on(a
utog
amy)
,ar
tifi
cial
self
poll
inat
ion
(gei
tono
gam
y)an
dfr
eepo
llin
atio
n(H
elvé
cia,
1988
-198
9)
Va
rie
ty
19
88
19
89
Na
tura
lse
lf-p
olli
na
tion
(au
tog
am
y)
Art
ifici
al
self-
po
llin
atio
n(g
eito
no
ga
my)
Op
en
po
llin
atio
nN
atu
ral
self-
po
llin
atio
n(a
uto
ga
my)
Art
ifici
al
self-
po
llin
atio
n(g
eito
no
ga
my)
Op
en
po
llin
atio
n
Pá
nd
y7
00
7.0
00
27
.0
De
bre
cen
ibõ
term
õ1
.41
7.2
9.4
6.8
21
.83
1.8
Ká
nto
rjá
no
si5
.21
0.2
17
.46
.71
0.8
37
.1
Cig
án
yme
gg
yC
40
44
.41
7.0
24
.58
.22
1.8
33
.1
Újfe
hé
rtó
ifü
rtö
s3
.21
.51
5.7
12
.22
1.6
35
.7
Cig
án
yme
gg
y5
98
.62
8.7
31
.01
1.7
32
.63
6.6
Cig
án
yme
gg
y7
6.8
16
.72
1.4
17
.43
4.5
37
.4
Pa
rasz
tme
gg
y1
4.7
23
.13
3.6
17
.13
9.7
42
.7
me
an
5.5
14
.32
0.0
10
.02
2.9
35
.2

than 10% fruit set; variability of their yieldis much higher than that of the self-fertilevarieties (Table 4). Maximum fruit set ofpartially self-fertile varieties may amount4.5 times their minimum set, whereas only2.5 times in self-fertile varieties.
The high yearly variation of fruit sets isshown in Table 5. In the Pándy meggy typeof varieties being partially self-fertile theyields show multiple differences dependingon the season. Selfed flowers yield nearly0% fruit, and the maximum yields did notreach 20% in any of them. As a mean of 19years there were three varieties with similarrates of fruit set. Earlier observations(Nyéki, 1974) proved the beneficial effectof geitonogamy on the rate of fruit set in re-lation to natural self pollination orautogamy even in self-fertile varieties. Inpartially self-fertile varieties the advantageof geitonogamy was much less expressedthan in the self-fertile group. However, nat-ural autogamy allowed a higher variation infruit set than did geitonogamy within theclone. Table 6 proves that withgeitonogamy (hand pollination with pollenof the same clone) fruit set increased by afactor of 2-3 in relation to natural self-polli-nation or autogamy. Self-incompatible vari-eties (e.g. Pándy 7) did not set fruit evenafter geitonogamous hand pollination.
Open pollinationFruit set on open pollinated flowers var-
ied substantially according to growing siteas well as to the season, which proves thehigh susceptibility of sour cherry to ecolog-ical adversities. Most decisive is the influ-ence of weather on fruit set during theblooming period. Nyéki (1989) stated thatfruit set of open pollinated Pándy meggytypes is low and seasonally highly variable.
Tables 3 and 4 shows fruit sets of the va-rieties assigned to five self-fertility groups.Within the group of Cigánymeggy there arevarieties, which are very poorly (e.g.Cigánymeggy 60), or well fertilised (e.g.
Hartai meggy, Paraszt meggy). Some fa-vourable seasons may produce high(26-27%) fruit sets even in self- -incompati-ble varieties (e.g. Cigánymeggy 215,Cigánymeggy 1317). At present, only thebest fertilised Cigánymeggy types are ad-mitted to be propagated (Cigánymeggy 7,Cigánymeggy 59). Some varieties (e.g.Montmorency 3) may set fruit at rates morethan 50%.
As was mentioned, self-incompatibleand partially self-incompatible varietiesyield more variably not only after self polli-nation but also with free pollination in rela-tion to the self-fertile varieties.
Nyéki (1989) stated that the higher theself-fertility of a variety the higher yieldsare expected also with open pollination.A linear correlation exists between the rateof self-fertility and the productivity at openpollination at P=0.01 level. It is also provedby data appearing on Table 3. Completelyand highly self-incompatible varieties setfruit with open pollination at rates of 12.1%and 9.3%, whereas self-fertile and highlyself-fertile sour cherry varieties producedthe highest fruit set: 29.0% and 29.4%.
CONCLUSIONS1. Self-incompatible and partially self-
-incompatible sour cherry varieties mustbe planted with associated polliniservarieties. Appropriate polliniser varietiesare to be chosen from groups of synchro-nous blooming period. If three bloomingtime groups are considered, the partnersshould belong to the same group,whereas from the five groups, they maybe taken from the neighbouring bloom-ing time groups too.
2. The rate of self- and open fertilisation ishighly variable depending on the grow-ing site as well as on the season. Varia-tion of fruit set (and yield) is a good deallower in self-fertile varieties than inself-incompatible and partially self-fer-
Vol. 47 No. 1 2003 Journal of Apicultural Science 57

tile varieties. Consequently, the safestyields are expected in self-fertile variet-ies.
3. Artificial self-pollination (geitonogamy)resulted in 2-3 times higher rates of fruitset in relation to natural self-pollination(autogamy), except in completelyself-incompatible varieties. Highest fruitset is expected on open blooming flow-ers visited by bees. The advantage ofopen pollination is clearly evident inself-fertile and completely self-fertilevarieties too. Consequently, it should bestated that bee pollination increased theyield not only in self-incompatible vari-eties but also in self-fertile sour cherryplantations.
REFERENCESBenedek P., Nyéki J., Szabó Z. (1990)
- Cseresznye- és meggyfajtákméhmegporzást befolyásoló tulajdonságai.Kertgazdaság 22(5):1-23.
Maliga P. (1942) - Adatok a Pándy meggyvirágbiológiájához. Magyar Kir. KertészetiAkad. Közl. 8: 3-5.
Nyéki J. (1974) - Meggyfajták virágzásaés termékenyülése. Kandidátusi értekezés.Kertészeti Egyetem, Budapest (Kézirat).
Nyéki J. (1989) - Csonthéjas gyümölcsûekvirágzása, termékenyülése. Akadémiaidoktori értekezés. MTA, Budapest(Kézirat).
Nyéki J., Szabó Z., Salgim, S. S. (1992)- Self and cross compatibility of
Hungarian sour cherry cultivars. Acta Agr.Hung. 41(1-2): 11-14.
Rjabov, I. N., Rjabova, A. N. (1970) -Samoopylenie i perekrestnoe opylenie ucheresni, visni i vihsne-cheresni. TrudiiGosud. Nikit. Botan. Sada 4: 53-153.
Rudloff C. F., Schanderl H. (1950) -Die Befruchtungsbiologie derObstgewächse und ihre Anwendung in derPraxis. Stuttgart, Ulmer-Verlag.
BIOLOGIA KWITNIENIA I ZAPYLANIE WIŒNI(Prunus ceraus L.) ODMIAN WYSELEKCJONOWANYCH
NA WÊGRZECH
N y é k i J . , S z a b ó T . , S z a b ó Z .
S t r e s z c z e n i e
Doœwiadczenia prowadzono w latach 1972 - 2002 w trzech miejscowoœciach: Erd, Helveciai Ujfeherto, w których badano odmiany wiœni uprawiane w ró¿nych zestawach odmianowych.Obserwacje obejmowa³y: biologiê kwitnienia (czas rozpoczêcia, okres trwania, najwiêkszenasilenie), dynamikê kwitnienia (liczba otwieranych codziennie kwiatów) oraz zawi¹zywanieowoców i wp³yw na to s¹siedztwa odmian znajduj¹cych siê w sadzie.
Opieraj¹c siê na uzyskanych wynikach oceniono stopieñ nak³adania siê okresów kwitnieniaró¿nych odmian, a na podstawie okresu najwiêkszego nasilenia kwitnienia wszystkie odmianypodzielono na 5 grup. Badano samop³odnoœæ i zdolnoœæ do naturalnego samozapylania siê orazwybrano dobre zapylacze dla odmian samosterylnych i czêœciowo samosterylnych.
Potrzebê zapylenia przez pszczo³y oceniono stosuj¹c ró¿ne sposoby zapylania; naturalnesamozapylanie, sztuczne samozapylanie i wolne zapylanie.
S³owa kluczowe: zap³odnienie, kwitnienie, zapylanie, wiœnia.
58

SITUATION AND PERSPECTIVEOF THE HUNGARIAN BEE-KEEPING
L e v e n t e N y á r s
Research and Information Institute for Agriculture Economics,1093 Budapest IX. Zsil u. 3-5. and 1355, P.O. Box 5, Tel.: (+36 1) 476 6071, e-mail: [email protected]
S u m m a r y
Apiculture constitutes a sector of Hungarian agriculture, which has not been big yetperspective in respect of its development possibilities. The ratio of apiculture shared 0.5 percentof the gross production value of agriculture, and 1.03 percent of that of animal husbandry in2000. Due to its natural conditions the whole territory of Hungary is suitable for apiculture. Thelatter is a complex activity: along honey production, the pollinating activity of bees must not beneglected (positive externality). The participants of the sector are mainly small-scale producers.Concentration due to economic reasons will be increased in the future. Concerning the quantityHungary cannot compete with large honey exporting countries therefore Hungary has topenetrate the EU honey markets with highly processed and branded products.
Keywords: honey production, cost-income situation, production structure, honey export,EU accession, service organisations.
INTRODUCTIONAccording to FAO statistics, the world
honey production was 1 262 thousand met-ric tons in 2001. Global output increased by30 percent from 1979 to 1989, which subse-quently stabilised in the following decade,with an increase of 3 percent from 1989 to1999. The world’s major producing regionis Asia, which is followed by Europe,Northern and Central America. In the con-text of the world trade, China is the majorexporter and the European Union is the ma-jor import market.
The honey production of the world be-comes more concentrated from step to step.Either the price competition or the produc-tion and the trade of the special honey prod-ucts make influence on market. Traditions,natural and economic conditions reflectwide variations of honey in the large honeyproducing countries, namely China, the CIScountries, USA, EU, Argentina andMexico.
METHODSIncreasing competitiveness gives much
more emphasis to the cost reduction, stand-ardisation and intensive production. Thecharacteristics of concentration can also bedetermined in this sector by the increasingnumber of bee colonies and decreasingnumber of honeybee-keeping farms. In cer-tain countries, like USA, Canada and Mex-ico, the demand for pollination withhoneybees increases. The participants of thesector also have to meet the customers’ de-mand; this concerns mainly those honeyproducts that originate from GMO cropproduction. The honeybee species and theirtreatments determine mainly the honeyproduction.
In Hungary, the annual honey productionoscillates about 16 thousand metric tons.According to the Hungarian Central Statis-tical Office (2001a), 68 per cent of the pro-duced honey is black locust, 19 per cent isblossom, 6 per cent is sunflower, 5 per centis rape and 2 per cent is other type of honey.
Vol. 47 No. 1 2003 Journal of Apicultural Science 59
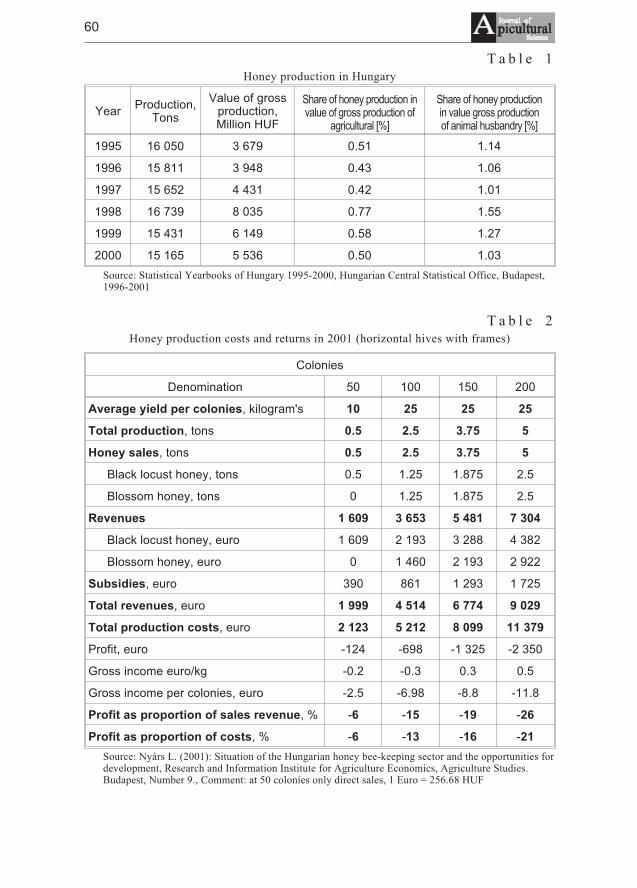
60
Year Production,Tons
Value of grossproduction,Million HUF
Share of honey production invalue of gross production of
agricultural [%]
Share of honey productionin value gross productionof animal husbandry [%]
1995 16 050 3 679 0.51 1.14
1996 15 811 3 948 0.43 1.06
1997 15 652 4 431 0.42 1.01
1998 16 739 8 035 0.77 1.55
1999 15 431 6 149 0.58 1.27
2000 15 165 5 536 0.50 1.03
Source: Statistical Yearbooks of Hungary 1995-2000, Hungarian Central Statistical Office, Budapest,1996-2001
T a b l e 1Honey production in Hungary
Colonies
Denomination 50 100 150 200
Average yield per colonies, kilogram's 10 25 25 25
Total production, tons 0.5 2.5 3.75 5
Honey sales, tons 0.5 2.5 3.75 5
Black locust honey, tons 0.5 1.25 1.875 2.5
Blossom honey, tons 0 1.25 1.875 2.5
Revenues 1 609 3 653 5 481 7 304
Black locust honey, euro 1 609 2 193 3 288 4 382
Blossom honey, euro 0 1 460 2 193 2 922
Subsidies, euro 390 861 1 293 1 725
Total revenues, euro 1 999 4 514 6 774 9 029
Total production costs, euro 2 123 5 212 8 099 11 379
Profit, euro -124 -698 -1 325 -2 350
Gross income euro/kg -0.2 -0.3 0.3 0.5
Gross income per colonies, euro -2.5 -6.98 -8.8 -11.8
Profit as proportion of sales revenue, % -6 -15 -19 -26
Profit as proportion of costs, % -6 -13 -16 -21
Source: Nyárs L. (2001): Situation of the Hungarian honey bee-keeping sector and the opportunities fordevelopment, Research and Information Institute for Agriculture Economics, Agriculture Studies.Budapest, Number 9., Comment: at 50 colonies only direct sales, 1 Euro = 256.68 HUF
T a b l e 2Honey production costs and returns in 2001 (horizontal hives with frames)
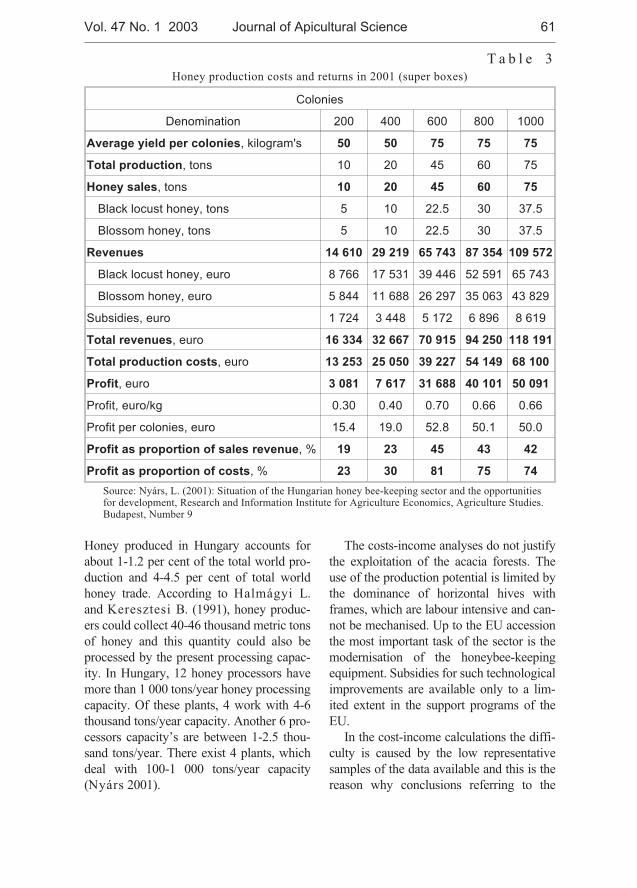
Honey produced in Hungary accounts forabout 1-1.2 per cent of the total world pro-duction and 4-4.5 per cent of total worldhoney trade. According to Halmágyi L.and Keresztesi B. (1991), honey produc-ers could collect 40-46 thousand metric tonsof honey and this quantity could also beprocessed by the present processing capac-ity. In Hungary, 12 honey processors havemore than 1 000 tons/year honey processingcapacity. Of these plants, 4 work with 4-6thousand tons/year capacity. Another 6 pro-cessors capacity’s are between 1-2.5 thou-sand tons/year. There exist 4 plants, whichdeal with 100-1 000 tons/year capacity(Nyárs 2001).
The costs-income analyses do not justifythe exploitation of the acacia forests. Theuse of the production potential is limited bythe dominance of horizontal hives withframes, which are labour intensive and can-not be mechanised. Up to the EU accessionthe most important task of the sector is themodernisation of the honeybee-keepingequipment. Subsidies for such technologicalimprovements are available only to a lim-ited extent in the support programs of theEU.
In the cost-income calculations the diffi-culty is caused by the low representativesamples of the data available and this is thereason why conclusions referring to the
Vol. 47 No. 1 2003 Journal of Apicultural Science 61
Colonies
Denomination 200 400 600 800 1000
Average yield per colonies, kilogram's 50 50 75 75 75
Total production, tons 10 20 45 60 75
Honey sales, tons 10 20 45 60 75
Black locust honey, tons 5 10 22.5 30 37.5
Blossom honey, tons 5 10 22.5 30 37.5
Revenues 14 610 29 219 65 743 87 354 109 572
Black locust honey, euro 8 766 17 531 39 446 52 591 65 743
Blossom honey, euro 5 844 11 688 26 297 35 063 43 829
Subsidies, euro 1 724 3 448 5 172 6 896 8 619
Total revenues, euro 16 334 32 667 70 915 94 250 118 191
Total production costs, euro 13 253 25 050 39 227 54 149 68 100
Profit, euro 3 081 7 617 31 688 40 101 50 091
Profit, euro/kg 0.30 0.40 0.70 0.66 0.66
Profit per colonies, euro 15.4 19.0 52.8 50.1 50.0
Profit as proportion of sales revenue, % 19 23 45 43 42
Profit as proportion of costs, % 23 30 81 75 74
Source: Nyárs, L. (2001): Situation of the Hungarian honey bee-keeping sector and the opportunitiesfor development, Research and Information Institute for Agriculture Economics, Agriculture Studies.Budapest, Number 9
T a b l e 3Honey production costs and returns in 2001 (super boxes)

whole sector cannot be drawn. Conse-quently, these data are only for information.For the analyses static models are applied.Calculations were made in two differentcategories. In one category of horizontalhives with frames the calculations refer tohoneybee-keeping farms with 50, 100, 150,200 bee colonies. In the other category ofhives with supers (boxes) honey-bee-keeping with 200, 400, 600, 800, 1000colonies was referred to. Table 2 and table 3provides estimates of the revenues, costsand returns from operating horizontal hiveswith frames and super boxes. Yields weredetermined by categories. Pricing is an im-portant issue in achieving sales and profit-ability. The price depends on what kind ofmarketing channels the honey is sold to.The minimum acceptable price shouldcover all production, marketing, transporta-tion and labour costs. Competitor prices arean important consideration in pricing honeyfor sale to consumers. Advantages of farmgate sales include: cash payments from con-sumers without incurring transportationcost, opportunity to develop products forniche markets. The potential to increasesales is limited and producers may not beable to market all of the product throughtheir outlet. As a result, producers needother marketing channels for honey thatcannot be sold at the farm gate. In ourmodel, the honeybee-keeping farm with 50colonies sold honey to the consumers
directly. Based on estimates otherbeekeeping farmers sold their honey topackers and honey agents. Operating withhorizontal boxes, revenues cannot hold thewage costs and the depreciation of equip-ment and vehicles. Due to the low yields,the production is showing loss. In the Hun-garian honeybee-keeping sector, the pro-ducers do not account for their wages andtheir contributions.
Real incomes were generated at honey-bee-keeping farms provided with hives withsuper boxes.
According to profitability calculations,the optimal production scale is about 600bee colonies per apiary in the Hungarianhoney bee-keeping sector. Larger stockswith higher yield resulted in higher profit-ability indicators; however, the high risksinvolved in large yields (risks of animalhealth) cannot be ignored (Nyárs L.,2001).
RESULTSThe production structure and specific in-
dicators of the honeybee keeping by stocksize do not show the real situation. The par-ticipants of the sector are mainly small--scale producers and due to the special taxallowances the ownership of bee colonies isdistributed among the family members. Thetotal number of beekeepers in Hungary wasapproximately 19 thousand in 1991 (Ta-ble 4).
62
Denomination 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000
Number ofbeekeepers 19 923 19 013 17 598 16 970 16 887 15 372 15 677 16 672 17 087 16 579
Number of beecolonies (thousand) 716 725 674 646 669 604 642 690 806 840
Bee colonies/beekeeper 36 38 38 38 39 39 41 41 47 51
Source: Honey Product Council
T a b l e 4Number of beekeepers and bee colonies in Hungary
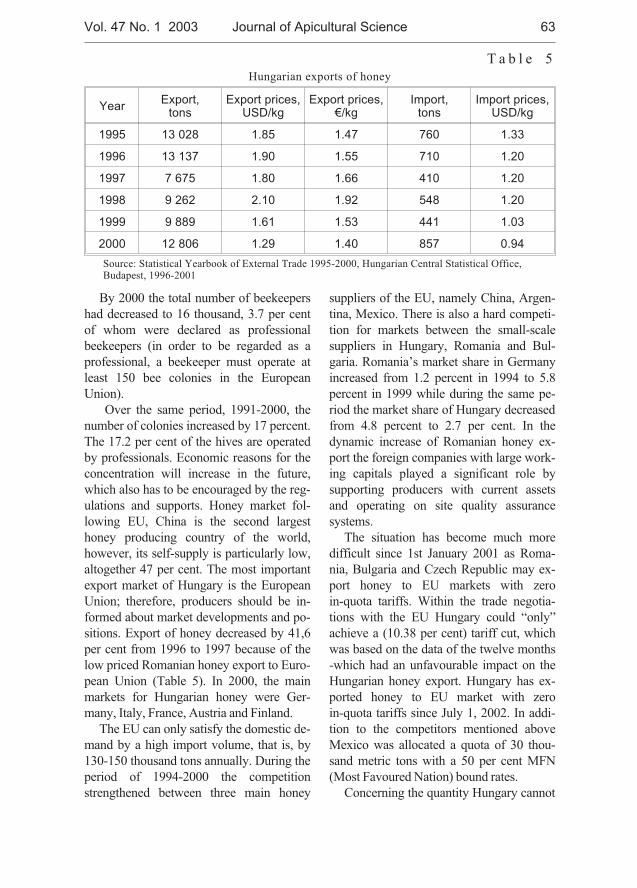
By 2000 the total number of beekeepershad decreased to 16 thousand, 3.7 per centof whom were declared as professionalbeekeepers (in order to be regarded as aprofessional, a beekeeper must operate atleast 150 bee colonies in the EuropeanUnion).
Over the same period, 1991-2000, thenumber of colonies increased by 17 percent.The 17.2 per cent of the hives are operatedby professionals. Economic reasons for theconcentration will increase in the future,which also has to be encouraged by the reg-ulations and supports. Honey market fol-lowing EU, China is the second largesthoney producing country of the world,however, its self-supply is particularly low,altogether 47 per cent. The most importantexport market of Hungary is the EuropeanUnion; therefore, producers should be in-formed about market developments and po-sitions. Export of honey decreased by 41,6per cent from 1996 to 1997 because of thelow priced Romanian honey export to Euro-pean Union (Table 5). In 2000, the mainmarkets for Hungarian honey were Ger-many, Italy, France, Austria and Finland.
The EU can only satisfy the domestic de-mand by a high import volume, that is, by130-150 thousand tons annually. During theperiod of 1994-2000 the competitionstrengthened between three main honey
suppliers of the EU, namely China, Argen-tina, Mexico. There is also a hard competi-tion for markets between the small-scalesuppliers in Hungary, Romania and Bul-garia. Romania’s market share in Germanyincreased from 1.2 percent in 1994 to 5.8percent in 1999 while during the same pe-riod the market share of Hungary decreasedfrom 4.8 percent to 2.7 per cent. In thedynamic increase of Romanian honey ex-port the foreign companies with large work-ing capitals played a significant role bysupporting producers with current assetsand operating on site quality assurancesystems.
The situation has become much moredifficult since 1st January 2001 as Roma-nia, Bulgaria and Czech Republic may ex-port honey to EU markets with zeroin-quota tariffs. Within the trade negotia-tions with the EU Hungary could “only”achieve a (10.38 per cent) tariff cut, whichwas based on the data of the twelve months-which had an unfavourable impact on theHungarian honey export. Hungary has ex-ported honey to EU market with zeroin-quota tariffs since July 1, 2002. In addi-tion to the competitors mentioned aboveMexico was allocated a quota of 30 thou-sand metric tons with a 50 per cent MFN(Most Favoured Nation) bound rates.
Concerning the quantity Hungary cannot
Vol. 47 No. 1 2003 Journal of Apicultural Science 63
Year Export,tons
Export prices,USD/kg
Export prices,�/kg
Import,tons
Import prices,USD/kg
1995 13 028 1.85 1.47 760 1.33
1996 13 137 1.90 1.55 710 1.20
1997 7 675 1.80 1.66 410 1.20
1998 9 262 2.10 1.92 548 1.20
1999 9 889 1.61 1.53 441 1.03
2000 12 806 1.29 1.40 857 0.94
Source: Statistical Yearbook of External Trade 1995-2000, Hungarian Central Statistical Office,Budapest, 1996-2001
T a b l e 5Hungarian exports of honey

compete with the large honey exportingcountries. Therefore, Hungary has to pene-trate the EU honey markets with highly pro-cessed and branded products. In the EUmember states there are purchasing, pack-aging and marketing co-operatives in thehoney sector. It is important for Hungary aswell, because an export price increase is notpossible with a bulk product. At present, theinterest of the Hungarian honey processorsand commercial companies lies in packag-ing and exporting the honey in barrels. Thelow producer prices do not favour the pro-ducers and the retailers transfer the price re-ductions to the producers.
In Hungary 90 percent of the honey pro-duction is for human consumption andbased on the estimations the remaining 10percent of the honey production is used bythe industry (baking industry, sweets indus-try, pharmaceutical industry and cosmetics)and social programmes. The honey industryalso produces beeswax, which is used inmaking candles and cosmetics. As for thesales of honey there are several marketingchannels. The wholesalers purchase 10-13thousand metric tons of honey from the pro-ducers each year depending on the fluctua-tion of the honey production; that is, themost significant part of the total productionannually. The nominal capacity of the Hun-garian honey processing plants is about 40thousand metric tons, which is double of thehighest production level of the last tenyears. Most of the commercial companies(all the large ones) operate their own honeyprocessing plants. This is the reason why atpresent the capacities of honey processingexceed the level of production in Hungary.
The regulation system of the Hungarianhoneybee keeping is harmonized with theEU regulations; concerning in particular theanimal health and the food hygiene legisla-tion. After the EU accession the presentHungarian market order will be replaced bythe market orders of the EU Member States.The harmonisation process requires
national programmes with accurate dataregister system. The national programmeshave to include both the current and thefixed costs (European Commission (2001b)). If the sector is not prepared for the EUaccession the Hungarian beekeepers willnot be eligible for the EU subsidies. The in-terest groups and professional bodies to-gether with the participants involved inhoney production will have to preparejointly their annual work plan based on therequirements defined by the regulation.
The national development plans and themeasures have had to contain also plans formonitoring the implementation and for theevaluation since the use of the financial re-sources from the EU budget is regulated.The needed information of the sector hasnot been provided for the preparation of EUaccession since the data of the Honey Pro-duce Council and that of the Central Statis-tical Office did not correspond to eachother. At present there are only estimatesconcerning the marketing channels used bythe producers for sales (direct sales to con-sumers, wholesaler, and retailers, sales tothe industry).
CONCLUSIONSHoney beekeeping is an integrated agri-
cultural activity. In addition to the honeyproduction the pollination by bees is also asignificant positive impact externality. InHungary pollination is considered as a sig-nificant part of the technology and for thatreason crop production and horticulture aresupposed to pay for this service. In the USAand Canada these activities are done bycontracts.
Within the Hungarian bee-keeping themost important task is the mechanisation ofhoney production; and in this way its pres-ent labour demand can be lowered. It isonly for stand sizes of 200-300 bee coloniesthat developments will pay off and tolerateinterest rates, as well as it is these stand
64

sizes that permit the financing of productionfor the following year. The major obstacleto development is the lack of capital andcredit.
The development of the sector could beencouraged by establishing service organi-sations and penetrating both the export andthe domestic markets, and branded productswith quality assurance certifications in con-sumer-friendly packages produced by pro-ducer’s organisations since bee keepingfarms with less than 100 bee colonies can-not be competitive on the market. Besidesthe activities listed above, the service or-ganisation could also get involved in jointmaterial purchase (medicines, sugar,apicultural implements), as in the case ofbig quantities the service organisation isable to achieve considerable price conces-sions. Service organisations will be opera-ble only if backed by capital-intensiveproducers’ communities or marketing com-panies.
REFERENCESApiculture in Hungary in 2000 -
Hungarian Central Statistical Office.Budapest, (2001a).
Bogenfrst F. et al. (1998) - The NaturalCompetitiveness of Hungary’s MainAgriculture Products. An Internationalcomparison. Integration StrategicWork-group. Budapest. pp. 54-56.
Erdsz Ferencn et al. (1998) - TheEconomic Competitiveness of Hungary’sMain Agricultural Products. Advatages andDrawbacks. Integration StrategicWork-group. Budapest. pp. 66-71.
European Commission (2001 a) - TheAgricultural Situation in the EuropeanUnion 1998 Report. Brussels. Luxembourg.1999.
European Commission (2001 b) - Reportfrom the Commission to the Council andthe European Parliament on theimplementation of Council Regulation (EC)No. 1221/97 laying down general rules forthe application of measures to improve theproduction and the marketing of honey.Brussels. 2001. http://europa.eu.int/comm/agriculture/markets/honey/index_en.htm
Halmágyi L. and Keresztesi, B. (1991)- The bee-pasture (A mhlegel). AkadmiaiKiad. Budapest. pp. 307.
Hartmut W. (1998) - Market andProduction Structure of the HungarianAgriculture and Food Export. Research andInformation Institute for AgricultureEconomics. Agriculture Studies. Budapest.Number 1. pp. 55-59.
Kartali J. et al. (1998) - The ExpectedCompetitiveness of Hungary’sFood-Industry Products in an ExpandedEurope. Integration Strategic Work-group.Budapest. pp. 97-103.
Nyárs L. (2001) - Situation of theHungarian honeybee-keeping sector and theopportunities for development, Researchand Information Institute for AgricultureEconomics, Agriculture Studies. Budapest.Number 9. 102 p.
Nyárs L. (2002) - Situation of Honeyproduction in Hungary, and its role in EUHoney consumption. Budapest.Gazdálkodás. 2002. Number 1. pp. 53-59.
Statistical Yearbook of External Trade1995-2000 - Hungarian CentralStatistical Office. Budapest. 1996-2001.
Statistical Yearbooks of Hungary1995-2000 - Hungarian CentralStatistical Office. Budapest. 1996-2001.
Vol. 47 No. 1 2003 Journal of Apicultural Science 65

STAN OBECNY I PERSPEKTYWYWÊGIERSKIEGO PSZCZELARSTWA
N y á r s L .
S t r e s z c z e n i e
Pszczelarstwo jest czêœci¹ rolnictwa na Wêgrzech, które obecnie nie ma mo¿liwoœcirozwojowych. Udzia³ pszczelarstwa w ca³kowitej wartoœci produkcji rolniczej w roku 2000wynosi³ 0,3%, a w produkcji zwierzêcej 1,03%. Warunki naturalne sprawiaj¹, ¿e ca³eterytorium Wêgier jest przydatne do produkcji pszczelarskiej. Na aktualne znaczeniepszczelarstwa patrzeæ nale¿y nie tylko ze wzglêdu na produkcjê miodu, doceniana winna byædzia³alnoœæ pszczó³ jak zapylaczy roœlin. Udzia³owcami w produkcji pszczelniczej s¹ g³ówniemali producenci. Ze wzglêdów ekonomicznych w przysz³oœci bêdzie nastêpowa³a koncentracjaprodukcji. Wêgry ze wzglêdu na wielkoœæ nie s¹ w stanie konkurowaæ z wielkimi eksporteramimiodu jednak dostarczaj¹ na rynki UE wysoko przetworzone markowe produkty.
S³owa kluczowe: produkcja miodu, koszty produkcji, struktura produkcji, eksport miodu,Unia Europejska, organizacja obs³ugi.
66

DELIBERATE PRODUCTION OF YOUNG QUEENS INBombus terrestris L. (Hymenoptera, Apoidea) COLONIES
REARED IN LABORATORY
V l a d i m i r P t a c e k
Masaryk University of Brno, Faculty of Science, Kotlarska 2, 611 37 Brno, Czech Republic
S u m m a r y
Two possible ways of obtaining young queens in laboratory were tested: to rear them ingroups of workers, and to make the regular colonies queenless in comparatively early stage oftheir development.
In 1997 four groups of four workers in each did not accept young diploid larvae either fromtheir maternal colony or from another one. Workers, which already have established dominancein the groups, destroyed the inserted brood and laid their own eggs. The method still should betested under different conditions, because, exceptionally, larger groups of workers did acceptalien brood and raised queens from it.
From 7 colonies deliberately made queenless at the stage of 20 - 30 workers 6 did raise youngqueens (20-40 per colony) from remaining brood. One colony produced males only. Youngqueens reared on this way behaved according to the queen-model and were able to start normalcolonies.
Keywords: bees, Bombus terrestris, bumble bees, queen rearing.
INTRODUCTIONAccording to the scheme presented first
by Duchateau (1991), in laboratory onlysome colonies produce queens. They arereared from a portion of diploid broodavailable in a colony when it reaches aphysiological stage called as the “switchpoint”. No other young queens emerge thenin such colony as its old queen turns to lay-ing haploid eggs only. However a large pro-portion of colonies do not produce queensat all. After having reared several genera-tions of workers, they turn to mail produc-ers for the rest of their life. However, therearing methods of bumble bees will not becomplete without having a possibility torear young queens intentionally from anycolony or even any queen.
Since the first laboratory colonies ofB. terrestris were reared from queens ac-companied by honeybee workers (Ptacek,
1983, 1985), and the queens showed a be-haviour partially similar to that of honeybee queens, it seems to be logical to use themodel of a queenless honey bee colony forraising emergency queens also in the caseof the mentioned bumble bee species.
MATERIALS AND METHODSTwo possible ways of obtaining youngqueens in laboratory were tested:
1. To rear them in groups of workers takenfrom regular colonies,
2. To make the colonies queenless in com-paratively early stage of their develop-ment.In the pilot trial in1997 four groups of
four workers in each (taken from a queenright colony) were given young larvae orig-inating either from their own queen (twogroups) or from the alien ones (the othertwo groups).
Vol. 47 No. 1 2003 Journal of Apicultural Science 67

In the second case the old queens wereremoved from colonies at the stage of 20-30workers with the expectation that at least apart of the remaining diploid brood will bereared into young queens. In the years 1999- 2001 the influence of queen-absence wastested on the total of 7 colonies.
RESULTS AND DISCUSSIONThe worker groups with 4 individuals in
each did not accept young larvae for furtherfeeding. The larvae were left do die.
The second method brought better re-sults. From the 7 colonies made queenless6 did raise young queens. In one colonysolely males were produced, probably be-cause it had reached the switch point at thestage of removing its queen. We were notable to register the accurate numbers of de-veloped queens but guessingly they werefrom 20 to 40 per a hive.
To be suitable for affected queen pro-duction a colony should have much broodin the stage of eggs and young larvae(Fig. 1). After the old queen is removed,a part of the diploid brood is fed as queens.
Such larvae enlarge in size noticeably(Fig. 2). Also their cocoons are much largerthen those of workers and males (Fig. 3).Cocoons can then be removed from a col-ony and treated in the already known way(Ptacek 2000). According to our experi-ence the queens raised from them behavenormally and are able to start and developstandard colonies.
Evaluating the gained experience weshould consider the second method - re-moving the queen - to be more successful. Itgives queens from nearly any colony. Onthe other hand we do not want to put asidethe first method - workers with addedbrood, either. In our trials, which were onlyvery simple, maybe the age of workersand/or their number in groups might playtheir role, as well as the age of the broodgiven to them. Workers in groups used byus already had established dominance be-fore they were given the alien brood whichalso might contribute to its rejection. Forthe possibility of further improvement ofthis method speaks also the fact, that lateron, when we by chance gave young broodto queenless but more numerous colonies,
68
Fig. 1 The colony suitable for affected queen production has brood in all stages and round 20workers. Queen (right in the picture) can be removed

some of them exceptionally did rear younglarvae from it (Fig. 4). Further experimentsin the field surely will bring more explana-tory results.
The brood from a certain “breeder”queen can be obtained easily by keeping it
in a small colony with several workers ofhers. When such a colony is given a clusterof male cocoons from any colony, thequeen covers it by egg cells within severaldays. This brood can than be taken off forraising young queens.
Vol. 47 No. 1 2003 Journal of Apicultural Science 69
Fig. 3 The young queen larvae (left in the lower corner) of the third experimental colony at theend of heir larval development. Some of them have just finished their cocoons
Fig. 2 Some of the former worker larvae of another queen less colony have been raised into youngqueens (the large ones in the middle and lower part of the picture)

CONCLUSIONSAccording to the gained experience it
seems to be possible to rear new queens innearly any colony of B. terrestris L. by re-moving the old queen at the stage when thecolony has 20 -30 workers.
Further research is necessary in the fieldof producing queens by deliberately madegroups of workers.
Similar responses of workers to the ab-sence of queen as they were observed in B.terrestris can be expected also in other bum-blebee species.
ACKNOWLEDGEMENTSThe trials and their presentation were
possible thanks to the support of theMasaryk University grant No. CEZ:J07/98:143100010 and the GACR grantNo: 203/98/0453.
REFERENCESDuchateau, M. J. (1991) - Regulation of
colony development in bumblebees. - Proc.5th. Symp. Poll., Acta Horticulturae,288:139-143.)
Ptacek V. (1983) - Stimulation of broodrearing by bumble bees (Hymenoptera,Bombidae) by means of contacts withhoney bees (Apis mellifera L.).(Preliminary communication). Sbornik ved.Praci, VSUP Troubsko, (8):157-161.(Czech, En., Ru. sum.)
Ptacek V. (1985) - Testing three methodsof rearing bumble bees (Hymenoptera,Bombinae). Sbornik ved. Praci VSUPTroubsko, 9: 56-57. (Czech, En., Ru. sum.)
Ptacek V. - (2000) Obtaining and overwintering young bumble bee(Hymenoptera, Bombinae) queens. In:Insect pollination in greenhouses.Proceedings of the specialist's meeting heldin Soesterbeg, The Netherlands, 30 Sept.- 2Oct. 1999. Issued: Utrecht, TheNetherlands: Apimondia, ICPBR--Pollination Section., 2000., s. 55-57. ISBN90-805899-1-8.
70
Fig. 4 One of the group of queen-less workers (beyond the experiments), which did accept alienbrood and raised queens (now in cocoons) from it

ROZWA¯ANIA NAD MO¯LIWOŒCI¥ WYCHOWU M£ODYCHMATEK TRZMIELA ZIEMNEGO - Bombus terrestris L.
(Hymenoptera, Apoidea) W RODZINACH HODOWANYCHW LABORATORIUM
P t a c e k V .
S t r e s z c z e n i e
Oceniano dwie metody otrzymywania m³odych matek w warunkach laboratoryjnych:wyprowadzanie ich w grupach robotnic i w rodzinach osieroconych we wzglêdnie wczesnymstadium ich rozwoju.
W 1997 roku robotnice trzmieli (po cztery w ka¿dej grupie) nie przyjmowa³y m³odychdiploidalnych larw pochodz¹cych zarówno z ich matecznej rodziny, jak i z obcych. Robotniceprzejmuj¹c dominacjê w grupach, zniszczy³y wstawiany czerw i sk³ada³y w³asne jaja. Badaniapowinny byæ jeszcze kontynuowane, poniewa¿ okaza³o siê, ¿e wiêksze grupy robotnicprzyjmowa³y obcy czerw i wyprowadza³y z niego m³ode matki.
Z 7 umyœlnie osieroconych rodzin w stadium 20-30 robotnic 6 wyprowadzi³o m³ode matki(20-40 na rodzinê), za wyj¹tkiem jednej, która produkowa³a tylko samce.
S³owa kluczowe: pszczo³y, Bombus terrestris, trzmiele, wychów matek.
Vol. 47 No. 1 2003 Journal of Apicultural Science 71

DISTRIBUTION AND RELATIVE ABUNDANCEOF BUMBLE BEES (Bombus and Psithyrus) IN HUNGARY
M i k l ó s S á r o s p a t a k i , J u d i t N o v á k ,V i k t ó r i a M o l n á r
Department of Zoology and Ecology, Szent István University, 2103 Gödöllõ, Páter K. u. 1.,e-mail: [email protected]
S u m m a r y
The distribution data of the Bombus and Psithyrus species occurring in Hungary were collatedinto a database. Over 6,500 occurrence data represented 25 Bombus and 6 Psithyrus species.Our summarised data covered the 42% of the UTM squares in Hungary. Three of the bumblebee species (B. distinguendus, B. elegans and B. serrisquama) had only occurrence data frombefore 1953 so these species are probably extinct in Hungary. Regarding the relative occurrencefrequency of the species, we found that the Hungarian bumble bee fauna has a great percentageof rare species. We found that most of the bumble bee species had shown a declining relativedistribution in the last 50 years.
Keywords: bumblebee, Bombus, cucoobumblebee, Psithyrus, distribution, Hungary.
INTRODUCTIONIn addition to the well known honeybee
(Apis mellifera) there are about 600 wildbee species in Hungary. They have a veryimportant role both in agricultural and natu-ral habitats with pollinating several plantspecies. The best known and most com-monly observed group of wild bees is thebumble bee group (Bombus Latr. andPsithyrus Lep.). These species are wide-spread in the Palearctic region (Prys--Jones & Corbet 1987). The bumble beefauna of the Carpathian basin is rich ascompared with other parts of Europe(Móczár 1953). Several plant species needbumble bee pollination, because these spe-cies have unique pollination quality (largebody size, buzz pollination etc). In the lastfew decades the number and distribution ofwild bees showed a decreasing tendency allover the Europe and North America (Kwak1996, Kwak et al. 1996, Westrich 1996,Williams 1996, Kearns & Thomson2001). So it is very important from a nature
conservation point of view as well to collectall of the available information about spe-cies distribution and frequency of bumblebee species (Sárospataki et al. 2002).
MATERIALS AND METHODSWe gathered together the distribution
data of bumble bees from different insectcollections (Hungarian Natural History Mu-seum, Natural History Museum of Bakony,private collections of Zsolt Józan and PálBenedek) and also from the available Hun-garian scientific literature (for the list ofpublications see Sárospataki et al. 2002).We collected approximately 13-14 thou-sands data, and after the erasing of the du-plicates the database consists of about 5200records.
The method of UTM- (Universal Trans-verse Mercator) mapping was used for visu-alise our data. This method covers thewhole surface of the Earth with a uniformnet, consists of 100x100 km, 10x10 km andeven smaller units (Bácsatyai 1997). In
Vol. 47 No. 1 2003 Journal of Apicultural Science 73
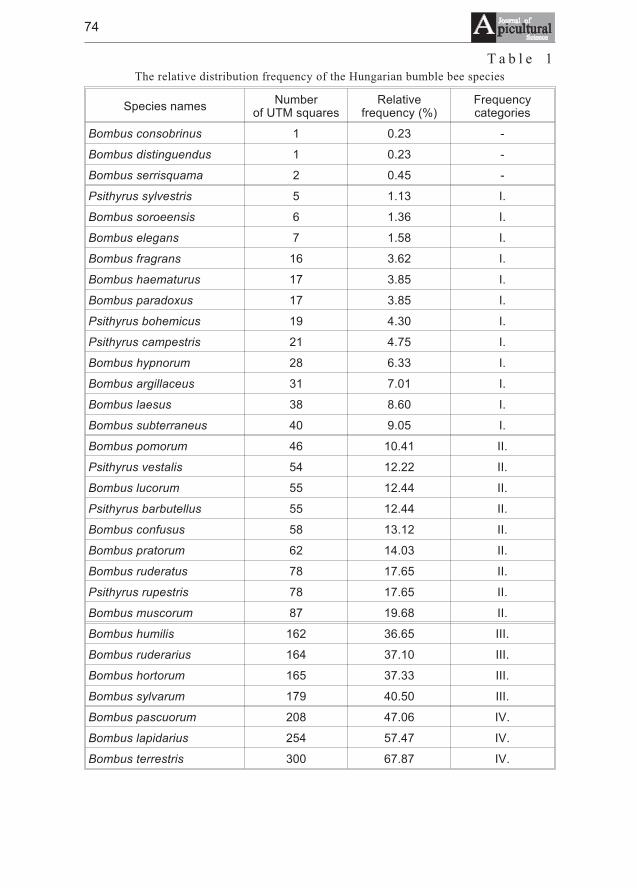
74
T a b l e 1The relative distribution frequency of the Hungarian bumble bee species
Species names Numberof UTM squares
Relativefrequency (%)
Frequencycategories
Bombus consobrinus 1 0.23 -
Bombus distinguendus 1 0.23 -
Bombus serrisquama 2 0.45 -
Psithyrus sylvestris 5 1.13 I.
Bombus soroeensis 6 1.36 I.
Bombus elegans 7 1.58 I.
Bombus fragrans 16 3.62 I.
Bombus haematurus 17 3.85 I.
Bombus paradoxus 17 3.85 I.
Psithyrus bohemicus 19 4.30 I.
Psithyrus campestris 21 4.75 I.
Bombus hypnorum 28 6.33 I.
Bombus argillaceus 31 7.01 I.
Bombus laesus 38 8.60 I.
Bombus subterraneus 40 9.05 I.
Bombus pomorum 46 10.41 II.
Psithyrus vestalis 54 12.22 II.
Bombus lucorum 55 12.44 II.
Psithyrus barbutellus 55 12.44 II.
Bombus confusus 58 13.12 II.
Bombus pratorum 62 14.03 II.
Bombus ruderatus 78 17.65 II.
Psithyrus rupestris 78 17.65 II.
Bombus muscorum 87 19.68 II.
Bombus humilis 162 36.65 III.
Bombus ruderarius 164 37.10 III.
Bombus hortorum 165 37.33 III.
Bombus sylvarum 179 40.50 III.
Bombus pascuorum 208 47.06 IV.
Bombus lapidarius 254 57.47 IV.
Bombus terrestris 300 67.87 IV.
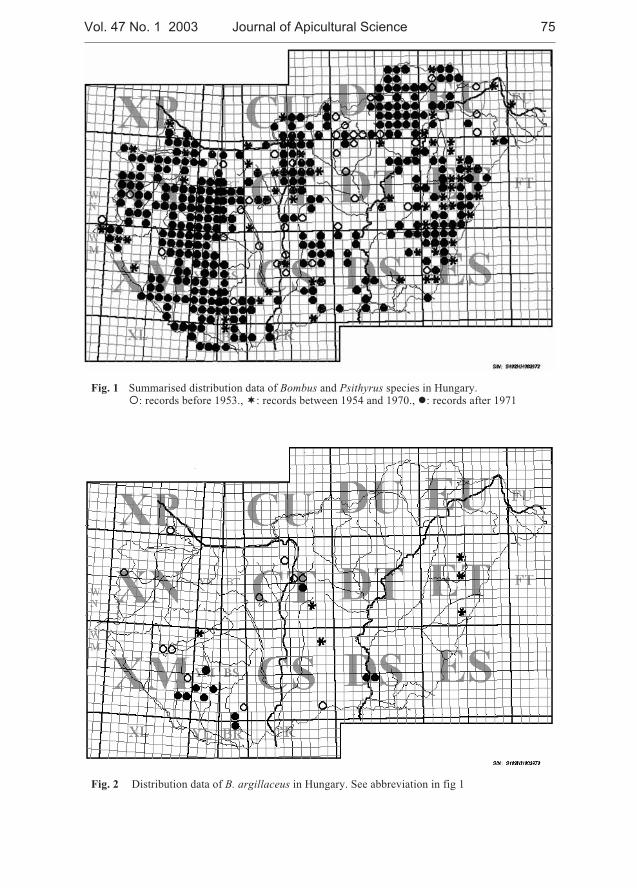
Vol. 47 No. 1 2003 Journal of Apicultural Science 75
Fig. 1 Summarised distribution data of Bombus and Psithyrus species in Hungary.�: records before 1953., �: records between 1954 and 1970., �: records after 1971
Fig. 2 Distribution data of B. argillaceus in Hungary. See abbreviation in fig 1

our work we used the 10x10 km units formapping the distribution data of the species.We used the “BioTér” software package forcreating the maps (Dévai et al. 2000). Onthe maps we separated the data into threecategories on the basis of collection date:before 1953, between 1953 and 1970, after1970.
The relative distribution frequency ofspecies was measured by the percentage ofall the UTM squares covered by thedatabase.
RESULTS AND DISCUSSIONOur database contains the distribution
data of 25 Bombus and 6 Psithyrus species(table 1). From the 1052 UTM squares ofHungary our data cover 442 squares (fig. 1).This is 42%, so the coverage is higher thanin other similar publications on other ani-mal groups in Hungary (Dévai &Miskolczi 1987, Bakó & Korsós 1999).Two examples of species maps are pub-lished here (fig. 2 and 3): one rare species
(B. argillaceus, the only protected species)and one common species (B. terrestris, oneof the most well-known and commonspecies).
Three species (B. distinguendus B.elegans and B. serrisquama) can be seen asextinct from Hungary, because we foundoccurrence data there only before 1953. Wehave only one data on B. consobrinus. Themore detailed observation of this only onecollection site is needed to find out the con-sistency of this population in Hungary.
The relative frequencies (table 1) showabout the half of the Hungarian species tobe rare. However, only one species(B. argillaceus) is protected in Hungary.The high percentage of rare species in theHungarian bumble bee fauna shows thatmuch more species need official protection.
It is very important to consider some diffi-culties of the UTM mapping methods:
1. We have a lot of UTM squares (58%)without any data. The absence of datacan mean not only the absence of the
76
Fig. 3 Distribution data of B. terrestris in Hungary. See abbreviation in fig 1

species, but frequently the absence of theresearch in the given area as well.
2. The distribution of data sometimesshows the site preference of the collec-tors.
3. The common species are probablyunderrepresented in the database,because collectors usually have a prefer-ence for rare, “more interesting” species(Móczár 1953). That is shown by therather low relative frequencies of themost common species, which probablycan be found almost all UTM squares ofHungary (Józan, personal communica-tion).
4. From similar consideration the rare spe-cies are overrepresented in the database,in other words they are more scarce thanthe data shows. This underlines the needof protection of these species.
ACKNOWLEDGEMENTWe are grateful for everybody who
helped as to collect the occurrence data ofbumble bees, namely: Zsolt Józan and PálBenedek (making available their privatecollections) as well as Lajos Zombory(Hungarian Natural History Museum) andCsaba Kutasi (Natural History Museum ofBakony). The work was partly financed bythe grants of the Oak Association for NatureConservation. During the study MiklósSársopataki had a Bolyai János ResearchGrant.
REFERENCESBakó B., Korsós Z.(1999) -A magyarországi
herpetofauna U. T. M. -térképezésénekfelhasználási lehetõségei. - ÁllattaniKözlemények, 84:43–52.
Bácsatyai L. (1997) - Vetülettan –Mûszaki Kiadó, Budapest. pp. 157–167.
Corbet S. A. (1996) - Why bumble beesare special. – In: Matheson A. (ed.).Bumble bees for pleasure and profit.International Bee Research Association,Cardiff, UK, pp. 1–12.
Dévai Gy. & Miskolczi M. (1987) -Javaslat egy új környezetminõsítõértékelési eljárásra a szitakötõk hálótérképszerinti elõfordulási adatai alapján. – ActaBiol. Debrecina 20:33–54.
Dévai Gy., Harangi J. & Miskolczi M.(2000) - Biotér 2.0 (Biotikaihálótérképezõ program), Debrecen.
Kearns C. A. & Thomson J.D. (2001) -The natural history of Bumblebees. –University Press of Colorado, Boulder,Colorado.
Kwak M. M. (1996) - Bumble bees athome and at school. – In: Matheson A.(ed.). Bumble bees for pleasure and profit.International Bee Research Association,Cardiff, UK, pp.12–24.
Kwak M. M., Velterop O. & BoerrigterE. J. M. (1996) - Insect diversity and thepollination of rare plant species. – In:Matheson A., Buchmann S. L., O’Toole C.,Westrich P. & Williams I. H. (eds.):Conservation of bees. Academic Press,London. pp.115–125.
Móczár M. (1953) - Magyarország és akörnyezõ területek dongóméheinek(Bombus Latr.) rendszere és ökológiája. –Ann. Hist-Natur. Musei Nation. Hung.4:131–159.
Osborn J. I. & Williams I. H. (1996) -Bumble bees as pollinators of crops andwild flowers. – In: Matheson A. (ed.).Bumble bees for pleasure and profit.International Bee Research Association,Cardiff, UK, pp. 24–33.
Prys-Jones O. E. & Corbet S. A. (1987)- Bumblebees. – Cambridge University
Press, Cambridge.
Sárospataki M., Novák J. & Molnár V.(2002) - Hazai poszméh- ésálposzméhfajok (Hymenoptera: Apidae,Bombus és Psithyrus) UTM-térképezése ésaz adatok természetvédelmifelhasználhatósága. Állattani Közlemények,in press.
Westrich P. (1996) - Habitat requirementsof central European bees and the problemof partial habitats. – In: Matheson A.,Buchmann S. L., O’Toole C., Westrich P.& Williams I. H. (eds.): Conservation ofbees. Academic Press, London. pp. 1–17.
Vol. 47 No. 1 2003 Journal of Apicultural Science 77

Williams I. H. (1996) - Aspects of beediversity and crop pollination in theEuropean Union. – In: Matheson A.,Buchmann S. L., O’Toole C., Westrich P.& Williams I. H. (eds.): Conservation ofbees. Academic Press, London. pp. 63–81.
WYSTÊPOWANIE TRZMIELI (Bombus)I TRZMIELCÓW (Psithyrus) NA WÊGRZECH
S á r o s p a t a k i M . , N o v á k J . , M o l n á r V .
S t r e s z c z e n i e
Opracowano ponad 6 500 danych dotycz¹cych wystêpowania 25 gatunków trzmieli (Bombus)i 6 gatunków trzmielców (Psithyrus) na Wêgrzech, które pokrywaj¹ 42% kwadratów mapyUTM. Trzy gatunki trzmieli (B. distinguendus, B. elegans i B. serrisquama) uznano za wymar³e,poniewa¿ dane o ich wystêpowaniu pochodz¹ sprzed 1953 roku. Pozosta³a fauna trzmieliwêgierskich nale¿y w znacznym procencie do gatunków rzadkich. Stwierdzono wzglêdneobni¿enie wystêpowania trzmieli na przestrzeni ostatnich 50 lat.
S³owa kluczowe: trzmiel, Bombus, trzmielec, Psithyrus, rozmieszczenie, Wêgry.
78

HOW OLD ARE BEES?- A LOOK AT THE FOSSIL RECORD
R i c h a r d A . B a k e r 1 , W i t C h m i e l e w s k i 2
1 School of Biology, University of Leeds, Leeds LS2 9JT, UK2 Apiculture Division, Research Institute of Pomology and Floriculture,
ul. Kazimierska 2, 24-100 Pu³awy, Poland
S u m m a r y
How old are bees? Which came first, bees or flowers? When did bees first develop socialbehaviour? Fossil bees are found in amber and copal but are quite rare as inclusions except forProplebeia in amber from the Dominican Republic. Amber occurs throughout the world and inEurope the main deposits are in the Baltic region, Germany, Romania and Sicily. Baltic amberis approximately 40 million years old (Myr). Amber belonging to the Cretaceous period is foundin several parts of the world and is about 120 Myr. Lebanese amber is the oldest and dates backto the early Cretaceous period some 135 Myr. At the time there were tropical rain forests wherethe resins from these trees preserved the insect fossils, which we have today. Copal bycomparison is “modern”, less than 2 Myr and is mainly from Africa, Colombia and theDominican Republic.
“The oldest fossil bee” was described by Michener and Grimaldi in 1988 and is nowreferred to as Cretotrigona prisca. It is from the Cretaceous amber of New Jersey and believedto be 96 to 74 Myr before the present. Rasnitsyn and Michener (1991) questioned the date ofthis amber and Rasnitsyn believes that late Eocene Baltic amber is where the oldest fossil beesare to be found. He is still strongly opposed to the Cretaceous origin of bees. Prior to thediscovery of Cretotrigona, the oldest fossil bees known were all from late Eocene Baltic amber,about 40 Myr. Engel (2001) has written a comprehensive monograph on Baltic amber bees andfirmly believes bees originated in the Cretaceous and that the oldest possible age for them is 125Myr. This is after the origin of flowering plants. No evidence exists (Engel 2001) to suggestbees arose prior to the origin and early diversification of flowering plants. Nests are also knownfrom the fossil record and provide some additional information for dating the origin of bees.
Photographs of bees in Baltic and Dominican amber are displayed and included bees from theMuseum of Amber Inclusions, University of Gdañsk; the Natural History Museum, Kraków; theMuseum of Earth, Warsaw and the Natural History Museum, London.
The paper is review of the present state of knowledge and attempt to answer the questionsraised at the beginning of this presentation. Relevant references are given.
Keywords: amber, Apis, bees, fossils, museum collections.
INTRODUCTIONFossil bees are rare. They are found
mainly in Baltic amber which is about 30 to40 million years old (myBP = million yearsbefore present). Amber occurs throughout
the world and in Europe the main depositsare in the Baltic region, Germany, Romaniaand Sicily. Copal by comparison is less than2 million years old and is mainly from Af-rica, Columbia and the Dominican
Vol. 47 No. 1 2003 Journal of Apicultural Science 79
REVIEW ARTICLES

Republic. Bees in amber are “probably bi-ased toward bees that used resin in nestconstruction” (Michener 2000).
Predatory sphecid wasps (Sphecoidea)are generally believed to be the ancestors ofpresent day bees (Apoidea). The oldest ofthese wasps date from the early Cretaceous.Why they changed from being predatory tocollecting pollen and nectar is not known.
The genus Apis Linnaeus 1758 also has afossil record. Apis mellifera L. fossils havebeen found as far back as 1.5 to 2 myBP.Specimens of honey bees from the copal ofEast Africa in the Natural History Museumin London are described by Zeuner andManning (1976).
RESULTS AND DISCUSSIONThe Oldest Bee
The oldest fossil bee, that is the earliestbee in the fossil record, is a worker de-scribed by Michener and Grimaldi(1988) from the late Cretaceous (80 myBP)amber of New Jersey, USA. It is referred toas Trigona prisca (Michener etGrimaldi) and is a stingless bee (Apidae,Meliponinae). Engel (2000) has redescribedthe fossil and transferred it to a new genusCretotrigona. However a “cloud hangs over
the dating for the piece of amber”(Michener 2000).
Rasnitsyn (see: Rasnitsyn andMichener 1991) has challenged the datingof the amber in which Trigona prisca hasbeen found and believes (personal commu-nication) the oldest body fossils of bees arethose from late Eocene Baltic amber andopposes the idea of the Cretaceous age ofTrigona. Prior to this report, the oldest beesknown were found in late Eocene Balticamber (40 myBP). Several species from theextinct genus Electrapis have been de-scribed from Baltic amber. These bees areconsidered to be social because: 1) manyspecimens have been found together in am-ber and 2) the presence of pollen collectingapparatus (Poinar 1992).
Specimens of amber with bee inclusionscan be found in museums throughout Eu-rope including Gdañsk, Hamburg,Kaliningrad, Kraków, London, Paris andWarsaw (see captions to photographs (Ta-ble 1; Figs 1-6)).
Fossil Honey BeesNo honey bees are known from before
the Oligocene. T.D.A. Cockerell (1866--1948) was one of the earliest workers onfossil honey bees and the first to describe a
80
Fig. 1 Apoidea (Hymenoptera) in Baltic amber. Museum of Amber Inclusions,University of Gdañsk (photo. E. Sontag).
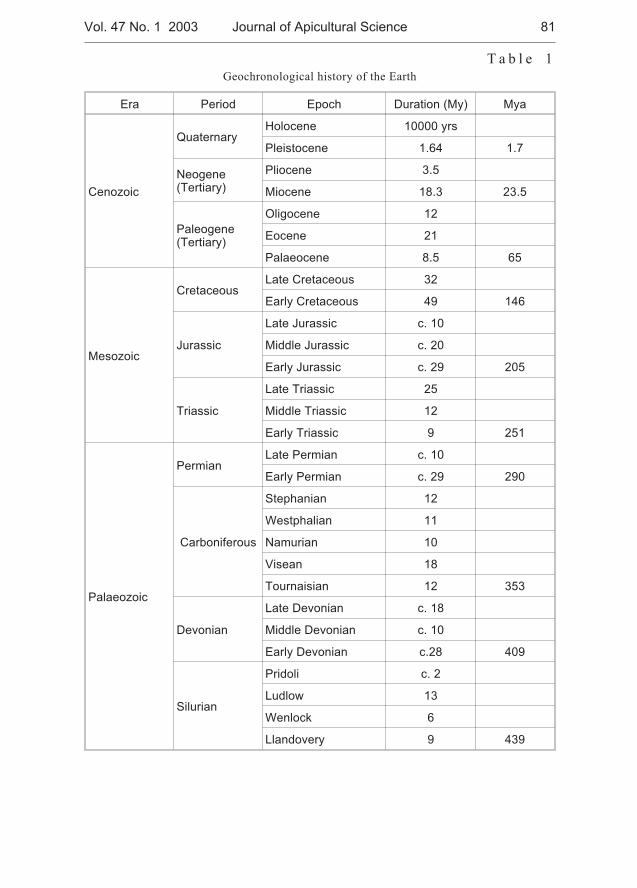
Vol. 47 No. 1 2003 Journal of Apicultural Science 81
Era Period Epoch Duration (My) Mya
Cenozoic
QuaternaryHolocene 10000 yrs
Pleistocene 1.64 1.7
Neogene(Tertiary)
Pliocene 3.5
Miocene 18.3 23.5
Paleogene(Tertiary)
Oligocene 12
Eocene 21
Palaeocene 8.5 65
Mesozoic
CretaceousLate Cretaceous 32
Early Cretaceous 49 146
Jurassic
Late Jurassic c. 10
Middle Jurassic c. 20
Early Jurassic c. 29 205
Triassic
Late Triassic 25
Middle Triassic 12
Early Triassic 9 251
Palaeozoic
PermianLate Permian c. 10
Early Permian c. 29 290
Carboniferous
Stephanian 12
Westphalian 11
Namurian 10
Visean 18
Tournaisian 12 353
Devonian
Late Devonian c. 18
Middle Devonian c. 10
Early Devonian c.28 409
Silurian
Pridoli c. 2
Ludlow 13
Wenlock 6
Llandovery 9 439
T a b l e 1Geochronological history of the Earth
82
Fig. 2 Apidae (Hymenoptera) (492) in Baltic amber. Natural History Museum, Kraków(photo. W. Krzemiñski).
Fig. 3 Electrapis krishnorum Engel (Apidae, Hymenoptera) (TG 5589). Museum of the Earth,Warsaw (photo. J. Kupryjanowicz).

Vol. 47 No. 1 2003 Journal of Apicultural Science 83
Fig. 5 Apidae (Hymenoptera) (In. 64560) in Dominican amber. Natural History Museum, London(photo. P. J. Evennett).
Fig. 4 Proplebeia dominicana (Wille et Chandler) (Apidae, Hymenoptera) in Dominican amber(photo. J. Kupryjanowicz).

fossil species of the genus Apis. In 1907 hedescribed Apis henshawi Cockerell. Theholotype of this, “the most famous of thefossil honey bees” (Engel 1998), is to befound in the Museum of Comparative Zool-ogy at Harvard University.
Zeuner and Manning (1976) referredto 17 fossil honey bees but now only 8 or 9fossil species are recognized. Of the to-dayliving species of honey bee, only one hasbeen found in the fossil record (Engel1998). This is Apis mellifera L. and isknown from the Pleistocene-Holocene, lessthan 2 myBP. Zeuner and Manning(1976) refer to specimens in the NaturalHistory Museum in London from the Pleis-tocene in East African copal.
Cockerell (1909) examined a piece of“amber” said to be from the coast offYarmouth, England containing twoA. mellifera L. and discusses their antiquity,suspecting the bees to be in a piece of copal(not amber) and not of English origin.
Engel (1998), lists the fossil taxa, pro-vides a revision and proposes a phylogenyfor Apis.
Which Came First - Bees or FlowerPlants?
Recent studies have shown that angio-sperm (Angiospermae) developed early inthe Cretaceous and by the end of this periodthere was a great deal of diversity in flower-ing plants. Bees probably arose before themiddle Cretaceous at a time when angio-sperms were becoming more abundant.
However there is a possibility that beesarose earlier and made use of pollen col-lected from gymnosperm plants(Gymnospermae) and may therefore havearisen before the angiosperms becamecommon.
It is probably true to say that bees aroseat the same time as or after the floweringplants.
CONCLUSIONSThis brief review of fossil bees high-
lights recent work and illustrates the needfor further on their biology and geology, in-cluding taxonomy and dating.
84
Fig. 6 Proplebeia sp. (Apidae, Hymenoptera) in Dominican amber. Natural History Museum,London (Baker, Chmielewski and Evennett - 2002 (in press) - photo. P. J. Evennett).

ACKNOWLEDGEMENTSThe authors would like to thank Peter
Evennett, Victoria Edmonds, Wies³awKrzemiñski, Janusz Kupryjanowicz, An-drew Ross and El¿bieta Sontag for permis-sion to use their photographs for illustrationof our presentation.
REFERENCESBaker R.A., Chmielewski W., Evennett
P.J. (2002) - Museum collections ofinsects and mites in amber - documentationand photography. Acta ZoologicaCracoviensia (special issue: Fossil Insects)45: (in press).
Cockerell T.D.A. (1907) - A fossil honeybee. Entomologist, 40:227-229.
Cockerell T.D.A. (1909) - SomeEuropean fossil bee. Entomologist, 42:313-317.
Engel M.S. (1998) - Fossil honey bees andevolution in the genus Apis (Hymenoptera:Apidae). Apidologie, 29:265-281.
Engel M.S. (2000) - A new interpretationof the oldest fossil bee (Hymenoptera:Apidae). Novitates, The American Museumof Natural History, number 3296:1-11.
Engel M.S. (2001) - A monograph of theBaltic amber bees and evolution of theApoidea (Hymenoptera). Bull. Am. Mus.Nat. Hist., No. 259:1-192.
Kupryjanowicz J. (2001) - Stawonogi wbursztynie baltyckim i ich fotograficznyobraz. (In:) B. Kosmowska-Ceranowicz(Red.) Bursztynowy skarbiec. Cz.I. Katalogkolekcji Tadeusza Giecewicza w zbiorachMuseum Ziemi PAN w Warszawie.Oficyna Wydawnicza Sadyba, Warszawa:19-97.
Michener C.D. (2000) - The bees of theworld. John Hopkins University, Baltimore.1-913.
Michener C.D., Grimaldi D.A. (1988) -The oldest fossil bee: apoid history,evolutionary stasis, and antiquity of socialbehaviour. Proc. Natl. Acad. Sci., USA.,85:6424-6426.
Poinar G.O. (1992) - Life in amber.Stanford University Press, Stanford, USA.1-350.
Rasnitsyn A.P., Michener C.D. (1991)- Miocene fossil bumble bee from theSoviet Far East with comments on thechronology and distribution of fossil bees(Hymenoptera: Apidae). Ann. Ent. Soc.America, 84(6):583-589.
Zeuner F.E., Manning F.J. (1976) - Amonograph on fossil bees. Bull. Brit. Mus.(Nat. Hist.) Geology, 27(3):1-268.
JAK STARE S¥ PSZCZO£Y?- SPOJRZENIE NA REJESTR SKAMIENIA£OŒCI
B a k e r R . A . , C h m i e l e w s k i W .
S t r e s z c z e n i e
Przedstawiono przegl¹d obecnego stanu wiedzy na temat skamienia³oœci owadówpszczo³owatych z pszczo³¹ miodn¹ w³¹cznie, na tle geochronoligcznego uk³adu dziejów Ziemi.Omówiono nazewnictwo najstarszej kopalnej pszczo³y stwierdzonej w zarejestrowanychdotychczas skamienia³oœciach i przedyskutowano kontrowersje w odniesieniu do wieku tegoznaleziska w œwietle literatury. Zestawiono wykaz wa¿niejszego piœmiennictwa na tematznalezisk tego typu i stwierdzenia skamienia³oœci (fosylia) Apis mellifera. Podano wykaz kilkueuropejskich muzeów, które posiadaj¹ zbiory bursztynów zawieraj¹cych inkluzje pszczele.
S³owa kluczowe: Apis, bursztyn, pszczo³y, skamienia³oœci, zbiory muzealne.
Vol. 47 No. 1 2003 Journal of Apicultural Science 85

BEE POLLINATION OF FRUIT TREES:RECENT ADVANCES AND RESEARCH PERSPECTIVES I.
P á l B e n e d e k
University of West Hungary, Faculty of Agricultural and Food Sciences,H-9201 Mosonmagyaróvár, Vár 4., Hungary
S u m m a r y
Intense research was carried out on the bee pollination of temperate zone deciduous fruit treesduring the past decade. Much progress was achieved to explore the flower characters of a greatnumber of cultivars affecting honeybee activity at flowers and their pollinating efficiency.Flower characters were found to be consequently different among apple and pear cultivars inconsecutive years but in case of some stone fruits differences between consecutive years weregreater than between cultivars in given years. The necessity of bee pollination was clearlydemonstrated both at self-sterile and at self-fertile fruit cultivars. The rate of flower constancyof honeybee foragers was different at different fruit tree species but the role of competing plantswas found to be much less deleterious than stated in the literature earlier. However, a number ofquestions arose partly from the results of latest pollination research and partly from practicalexperiences in commercial plant production. These indicate several research tasks to understandand to solve the problems possibly in the near future. The questions concentrate on theeffectiveness of bee visits in the pollination of individual fruit crops and their different cultivarsand on the reliable estimate of the overall amount of bees as well as on the control of beedensity during the flowering period of fruit orchards for optimal, controlled honeybeepollination. Much less effort was made to manage native wild bees for fruit tree pollination,however, some mason bees seem to be promising for this purpose in Europe, The Far East andNorth America.
Keywords: insect pollination, honeybees, temperate zone deciduous fruit crops.
INTRODUCTIONAlmost all temperate zone fruit tree spe-
cies need insect pollination, the only excep-tion is the horse chestnut, which is windpollinated, but partly benefits insect pollina-tion, too (Pesson and Louveaux 1984).The role of insects in fruit crop pollinationhas been recognized long ago. The informa-tion accumulated on this field has been re-viewed by Free (1993) in thecomprehensive handbook “Insect pollina-tion of crops (second edition)” some tenyears ago. After the appearance of this bookfruit pollination research has remained in-tense. Important new information accumu-lated in this field, however, several new
problems arose that needs further research.The aim of this paper is to give an outlookonto the state of knowledge in this field andto point out the need for further research.
RESULTSImproving the efficiency of honeybeesin crop pollination
A number of techniques have beenknown to increase the pollinating efficiencyof honeybees by increasing the bee visita-tion to target crops, increasing the ratio ofpollen collecting foragers, improving theirpollen dispersal, and so on (Free 1993).
However, to increase the bee visitationof the target crop is rather a hard task, hence
Vol. 47 No. 1 2003 Journal of Apicultural Science 87

it depends on several characteristics of thecrop and of cultivated or wild plants grownin the surroundings, on the food demand ofthe honeybee colonies, and of course, on theweather. This may be the explanation why anumber of different methods, neither theapplication of bee attractants, nor sequentialintroduction of colonies to the floweringcrop, nor feeding bees, nor diluting the nec-tar with overhead irrigation, nor interplantalfalfa increased the number of honeybeesand increased the yield significantly at seedonion fields (Mayer and Lunden 2001).
On the other hand, Winston andSlessor (1993) reviewing their results onthe recognition and the use of the queenmandibular pheromone (QMP), predictedthat its most significant commercial appli-cation would be in crop pollination. Thisstatement was based on good results ob-tained in field experiments. Spraying flow-ering crops increased the number ofhoneybees foraging on apple, pear, cran-berry, and blueberry (Currie et al. 1992a,1992b, 1994, Hugo et al. 1995, MacKen-zie and Averill 1992). The effect of QMP,however, is greatly dependent on dosageand partly on crop. Namely, 0.1 queenequivalents/ha was ineffective, but 1000was effective on apple and pear (Currie etal. 1992b), the same dosage failed to in-crease bee visits on sweet cherry (Naumannet al. 1994), while 100 was the most effec-tive on cranberry and 100 was as effectiveas 1000 on blueberry (Currie et al. 1992a).In other instances, with pear and apple,QMP application increased only fruit sizeswithout actual yield increase, and this stillresulted in higher profits (Currie et al.1992b, Naumann et al. 1994). On he con-trary, on sweet cherry a negative correlationarose between bee visits and fruit size.These experiences, however, are not consis-tent enough, and therefore further researchis needed on this topic, not only in the USbut also in other countries with differentweather conditions. Also, the profitability
of the method should be analysed, as itsacceptance in the practice depends on itsprofitability. There is one other, cheaperkind of application of QMP in crop pollina-tion, because the pollination value ofqueenless “disposable” pollination units canbe improved when QMP is added, whichmay increase the foraging activity; more-over should brood be present, even pollencollecting activity is increased (Currie etal. 1994). Other kinds of bee attractants(e.g. Beeline, Be-Q, Beehere, lavender oil,citral, geraniol, etc.), however, did not in-crease the bee visitation at a number of dif-ferent crops (Ambrose et al. 1995, Neiraet al. 1994, Ortiz-Sanchez 1993, Singand Sinha 1996, Zvedenok 1996).
As in-hive pollen transfer is recognizedto be an important factor in pollen dispersalamong compatible but self-sterile orself-fertile but readily cross-fertilizingcultivars of some crops (e.g. Dag andKammar 2001, DeGrandi-Hoffman etal. 1984, DeGrandi-Hoffman and Mar-tin 1993, 1995, McLaren et al. 1996), themethods increasing pollen exchange be-tween foragers of the honeybee coloniescan be regarded to be very important. Fol-lowing the pioneer work of Free et al.(1991), several kinds of hive entrance de-vices were used lately to brush pollen fromincoming and deposit on departing bees(Hatjina 1996, Eijnde and Velde 1995,Szalai 2000). This type of research shouldbe continued in order to develop the mosteffective devices without harming foragersand without decreasing germination capac-ity of the pollen.
However, the mechanism and the effec-tiveness of in-hive pollen exchange are notadequately studied and its role in crop polli-nation is probably underestimated. Latestresearch on the fruit set of large sin-gle-cultivar blocks of apple indicates thatin-hive pollen exchange (or possibly carry-ing pollen on the body hairs of bees fromone foraging trip to the next) could be much
88

more important in the effective pollinationof orchards than believed so far, becauseeven large and very large single-cultivarblocks in apple plantations can effectivelybe pollinated in lack of pollinizer cultivarsinside the block (Bla�ek 1996, Benedekunpublished). Therefore, the in-hive pollentransfer among bees needs to be studiedthoroughly with fruit trees and other cropplants.
Recently, the role of electrostatic chargesfor non-contact pollen detachment betweenpollinating honeybees and flowers is con-firmed, and this can be utilized for supple-mentary pollination in commercialproduction (Vaknin et al. 2000), this newapproach, however, needs further researchto base commercial implementation.
A need of controlled honeybee pollinationAs successful pollination may be a limit-
ing factor of a good yield, growers wouldlike to know how many honeybee (or bum-blebee) colonies are needed and how to or-ganize them for optimal pollination at thefield. We can give general instructionsbased on experiments and/or measurementsabout timing the placement and the arrange-ment of the location of the bee colonies in-side or round the target crop, but thenumber of bee colonies needed is muchmore difficult to define on a reliable basis.
On the other hand, the recommendationson the stocking rates of bee colonies neededto pollinate the crop plants are based on as-sumptions rather than on experimental re-sults. There is very little information in theliterature how to estimate the necessarynumber of bee visits/flower or the necessarybee density/per unit area of optimal pollina-tion or about the optimal set of variouscrops (the examples are sunflower, cotton,hairy vetch as well as fruit plantations tosome extent: see in Free 1993). This kindof information would, however, be very im-portant in the practice, because not only theinadequate activity of pollinating insects,
but their excessive activity can also be dis-advantageous to the yield.
The unpleasant consequences ofoversetting, numerous but too small fruits,poor quality, the need of laborious and ex-pensive thinning, are well know for somefruit trees in the practice, for peach and nec-tarine for example. Lately, both insufficientand excessive visiting of bees (honeybeesand bumblebees) for pollination of straw-berry have been found to be undesirable,both causing faulty pollination under cover.A density of 10 - 15 thousands of workerbees/1000 sq. metres in plastic tunnels orglasshouses is recommended, but neitherthe lower nor the higher densities are desir-able (Lieten 1993). Similar results wereobtained with cucumber; fruit set and thenumber of filled seeds were the higherwhen there were some 0.5 honeybee colo-nies per some 2000 plants, reduced numberof bees was resulted in lower yields, on thecontrary overpopulation caused so strongcompetition for flowers (for pollen) whichreduced the yield (Cervancia and Forbes1993). In case of sunflower crops some 10thousand bees/hectare was found to be opti-mal for pollination in Hungary, however,increased bee visitation did not affect theyield (Benedek 1972). Similarly, addi-tional extra hives at flowering pear in-creased bee numbers in the orchard, but notthe fruit set in New Zealand (McLaren etal. 1996).
For this reason, it is important to explorethe relationship between the number of beevisit/flower and the set. In case of pumpkin,for example, latest results show that some10 insect visits are needed o that enoughpollen grains are deposited onto the stigmaneeded for a good set (Masierowska andWien 2000). In case of squash each plantrequires at least one honeybee visit duringthe optimum pollination time (that is be-tween 06.00 and 09.00 in this case)(Al-Fattah 1991), and some 5-6 bee-vis-its/flower are needed for maximum
Vol. 47 No. 1 2003 Journal of Apicultural Science 89

productivity of raspberry (Oliveira et al.1991). Based on experimental results 6-12bee visits were necessary in Hungary duringthe life of a single apple flower to set a fruit(Benedek et al. 1989). Both fruit set andquality were related to the number of beevisits/flower in experiments with apple inCanada, but an other experiment gave in-conclusive result due to overall poor fruitset caused by bad weather (Brault et al.1995).
In fact, the present practice to move cer-tain number of bee colonies to the targetcrop just prior to, or at the start of flower-ing, and to leave them on site until petalfall, is an inadequate solution to crop polli-nation problems. This can be exemplifiedby a sequential introduction of honeybeecolonies to pear pollination; namely, plac-ing additional bee colonies in pear orchardsat 50 % flowering resulted in more bees vis-iting the flowers for at least one day whichsignificantly increased fruit set in 10 out ofthe 14 experimental orchards (Mayer1994). However, growers need some kindof method, as simply as possible, to decideif they should do something with bees dur-ing the flowering period of crops. A simplesolution was suggested in Hungary. Takingsome factors, the number of bee visits perflower necessary to set a fruit, the length ofthe receptive period of the flowers, the re-quired ratio of fruit set and the patters of thedaily bee activity into account, 3-6 bee vis-its per 50 opening flowers during a ten min-utes period was recommended as theoptimal intensity of bee visitation in appleorchards to get a good crop (Benedek1996). The growers were recommended tocontrol the intensity of bee pollination dur-ing the flowering period and to move addi-tional colonies to the orchard immediately,when the bee visitation was much lowerthan the proposed optimal level thus avoid-ing insufficient, low sets. At other in-stances, however, excessive bee visitationwas decreased during the flowering period
to avoid unwanted oversetting at somecrops. This kind of bee pollination manage-ment should be implemented during theflowering period of crops, which wouldlead to controlled bee pollination in theagriculture.
Consequently, intense research is neededto base controlled bee pollination experi-mentally for those crop plants, of differentfruits first of all, where lower and higherthan optimal sets are equally undesirable forprofitability reasons. Growers should berecommended to carry out simple observa-tions or should be given sophisticated, butuser-friendly computer simulations, as thewell-known PC-REDAPOL for example,which can reliably predict the apple yield(DeGrandi-Hoffman et al. 1995). Thisenables growers to decide what to do duringthe flowering period for the optimal pollina-tion of their insect pollinated crops. Theproblem is especially crucial in the newtype, high density orchards with semi-dwarfor dwarf fruit trees being the productive lifemuch shorter, 10 to 15 years at most, thanof traditional, large crown fruit plantations,which can stand and produce fruit for de-cades. During the short productive period ofhigh density orchards no single year withbad yield or with bad fruit quality can besuffered, because the investment cost of thistype of orchards is very high and also theircultivation is much more expensive than ofa traditional plantation, consequently theproduction must be profitable each year oftheir short productive life. Accordingly,pollination must not be an unstable elementof their management system.
There is a host of factors affecting thegeneral behaviour of bees towards the re-wards that different crops and competingother plants, flowering simultaneously offerto them. Other environmental factors affectalso the foraging behaviour, and conse-quently the pollinating efficiency of beesvisiting the flowers of crop plants (more ofsocial than of solitary bees). Weather and
90

competitor plants seem to have the most de-cisive influence on bee activity. The effectof weather conditions is fairly well known,but the effect of competing plant species iscontradictory and confusing. Lately, evenmassive appearance of flowering dandelion,that is known to have very strong competi-tive effect on honeybees in fruit orchards,failed to affect bee visitation in pear or-chards, neither sour cherry attracted beesfrom pear (Benedek et. al. 1998a). Thereare some other indications that the effect ofcompetitor plants is not sufficiently ex-plored, accordingly this item needs furtherstudies to understand why certain plant spe-cies are strong competitors in some casesand no similar effect can be recognized atother instances.
CONCLUSIONSBased on the above discussion there
seem to be a number of topics that greatlyneeded intense research in the near future toimprove the knowledge on as well as thetechnology of the insect pollination of cropplants cultivated in temperate zone regions.These are partly related to the pollinationrequirements of selected crops and to man-aging some solitary wild bee species forcrop pollination. The problems, however,concentrate on the effectiveness of bee vis-its in the pollination of individual fruit crops(and their different cultivars) and on the re-liable estimate of the stocking rates of beesfor crop pollination as well as on the man-agement of actual bee density during theflowering period of fruit orchards for opti-mal controlled bee pollination. The follow-ing topics are proposed as the subject offurther research in the coming years:1. Improving the efficiency of honeybees in
crop pollination— The effect of the queen mandibular
pheromone sprays is to be furtherstudied to increase the pollination ef-ficiency of honeybees at target crops
in flower.— Further research is needed on the
mechanism and the effectiveness ofin-hive pollen exchange among hon-eybees during crop plant pollination.
— Further research is required to de-velop the most effective devices in-creasing pollen exchange betweenforagers of honeybee colonies forpollination.
2. A need of controlled honeybee pollina-tion
— Intense research is needed to basecontrolled bee pollination experi-mentally for those crop plants, of dif-ferent fruits first of all, where lowerand higher than optimal sets areequally undesirable for profitabilityreasons. Growers should be recom-mended to carry out simple observa-tions or should be given sophisticated,but user- friendly computer simula-tions to manage flower visiting beepopulations at an optimal level dur-ing the flowering period of insect--pollinated crop plants.
— Further studies are needed on the ef-fect of competing plant species tounderstand why certain plant speciesare strong competitors in some casesand no similar effect can be recog-nized at other instances.
REFERENCESAl-Fattah M.A.A. (1991) - Behaviour of
honeybee foragers and their influence aspollinators on summer squash flowers(Cucurbita pepo L.). Bull. FacultyAgriculture, Univ. Cairo, 42: 1169-1186.
Ambrose J.T., Schultheis J.R., BambaraS.B., Mangum, W. (1995) - Anevaluation of selected commercial beeattractants in the pollination of cucumbersand watermelons. Am. Bee J., 135(4):267-272.
Vol. 47 No. 1 2003 Journal of Apicultural Science 91

Benedek P. (1996) - Insect pollination offruit crops. In: Nyéki J.- Soltész M.editors.: Floral biology of temperate-zonefruit trees and small fruits. Akad. Kiadó,Budapest: 287-340.
Benedek P. (2000) - Some flowercharacters, variety features andenvironmental effects affecting beepollination of pear (Pyrus communis L.): areview of the results of latest research.Internat. J. Horticultural Science, 6(3):59-66.
Benedek P., Béres I., Nyéki J. (1998a)- Competition between pear flowers,flowering weeds and other fruit trees forhoneybee pollination. Acta Horticulturae,475: 417-426.
Benedek P., Manninger S., Nagy B.(1972) - The number of colonies and thedensity of honeybees in sunflower fields inrelation to the pollination of the crop. Z.ang. Ent., 71: 385-389.
Benedek P., Ruff J., Nagy Cs., Nyéki J.(2000c) - Flower constancy of honeybees(Apis mellifera L.) to blooming pearplantations. Internat. J. HorticulturalScience, 6(3): 81-85.
Benedek P., Szabó T., Nyéki J. (2000f)- The effect of the limitation of insect
pollination period on the fruit set and yieldof quince cultivars (Cydonia oblongaMill.). Internat. J. Horticultural Science,6(3): 103-108.
Bla�ek J. (1996) - Pollination ofsingle-cultivar blocks of apple cv. GoldenDelicious. Acta Horticulturae, 423: 95-102.
Brault A.M., Oliveira D. de, Marceau J.(1995) - Optimization of apple orchardpollination by honey bees in easternCanada. Canadian Honey Council Res.Symposium Proceeding 1995: 55-104.
Cervancia C.R., Forbes M.F. (1993) -Density of bees (Apis mellifera L.) neededfor effective pollination of cucumber(Cucumis sativus L.). Philippine J. Science,122: 129-131.
Currie R.W., Winston M.L., SlessorK.N. (1992a) - Effect of synthetic queenmandibular pheromone sprays on honeybee (Hymenoptera: Apidae) pollination ofberry crops. J. Economic Entomology, 85:1300-1306.
Currie R.W., Winston M.L., SlessorK.N. (1994) - The effect of honey bee(Apis mellifera L.) synthetic queenmandibular compound on queenless“disposable” pollination init. Am. Bee J.,134(3): 200-202.
Currie R.W., Winston M.L., SlessorK.N., Mayer D.F. (1992b) - Effect ofsynthetic queen mandibular pheromonesprays on pollination of fruit crops byhoney bees (Hymenoptera: Apidae). J.Economic Entomology, 85: 1293-1299.
Dag A., Kammar Y. (2001) -Comparison between the effectiveness ofhoney bee (Apis mellifera) and bumble bee(Bombus terrestris) as pollinators ofgreenhouse sweet pepper (Capsicumannuum). Am. Bee J., 141: 447-448.
DeGrandi-Hoffman G., HoopingarnerR., Baker K. (1984) - Pollen transfer inapple orchards: tree-to-tree or bee-to-bee?Bee World, 65: 126-133.
DeGrandi-Hoffman G., Martin J.H.(1993) - The size and distribution of thehoney bee (Apis mellifera L.)cross-pollinating population on male-sterilesunflowers (Helianthus annuus L.). J.Apicultural Res., 32: 135-142.
DeGrandi-Hoffman G., Martin J.H.(1995) - Does a honey bee (Apismellifera) colony’s foraging population onmale-sterile sunflowers (Helianthusannuus) affect the amount of pollen onnestmates foraging on male-steriles. J.Apicultural Res., 34: 109-114.
Eijnde J. van der, Velde H. van der(1995) - Improvement of pollen exchangebetween honeybees (Apis mellifera) byusing hive entrance bristles. ActaHorticulturae, 423: 79-81.
Free J.B. (1993) - Insect pollination ofcrop. Second edition. Academic Press,London.
Free J.B., Paxton R.J., WaghchoureE.S. (1991) - Increasing the amount offoreign pollen carried by honey beeforagers. J. Apicult. Res., 30: 132-136.
92

Hatjina F. (1996) - The use of ‘temporaryconfinement’ and ‘pollen transfer device’to increase pollination potential of honeybees. PhD Thesis, University of WalesCollege of Cardiff, UK: XII + 208 pp.(Abstract in: Apiciltural Abstracts, 48(4):412; Item number 1477/1997).
Hugo H., Winston M.L., Slessor K.N.(1995) - Mechanisms by which honey bee(Hymenoptera: Apidae) queen pheromonesprays enhance pollination. Ann. Entomol.Soc. Am., 88: 366-373.
Lieten F. (1993) - Vruchtmisvorming bijaarbeinen. Overbevlieing. Fruitteelt, 6(2):27-29.
MacKenzie K.E., Averill A.L. (1992) -A new honey bee attractant, the queenmandibular pheromone. Cranberries, 56:13-14.
Masierowska M.L., Wien H.C. (2000) -Flowering and fruit set of two pumpkin(Cucrbita pepo L.) cultivars under fieldconditions. Apiacta, 35: 97-105.
Mayer D.F. (1994) - Sequentialintroduction of honey bee colonies for pearpollination. Acta Horticulturae, 367:267-269.
Mayer D.F., Lunden J.D. (2001) -Honey bee management and wild bees forpollination of hybrid onion seed. ActaHorticulturae, 581: 275-278.
McLaren G.F., Fraser J.A., Grant J.E.(1996) - Some factors influencing fruitset in ‘Sundrop’ apricot. N.Z.J. CropHrticult. Science, 24: 55-63.
Naumann K., Winston M.L., SlessorK.N., Smirle M.J. (1994) - Synthetichoney bee (Hymenoptera: Apidae) queenmandibular gland pheromone applicationsaffect pear and sweet cherry pollination. J.Economic Entomology, 87: 1595-1599.
Neira C., Barriga G.J., Munaca B.N.(1994) - Caracrerísticas de los frutos deRubus idaeus L., asociadas al uso de dosatrayentes y un replente. Agro Sur, 22(1):77-82.
Oliveira D. de, Gingras J., Chagnon M.(1991) - Honey bee visits and pollinationof red raspberries. Acta Horticulturae, 288:415-419.
Ortiz-Sánchez F.J. (1993) - Evaluationof the efficiency of Beehere as a honeybee(Apis mellifera Linnaeus, 1758) attractantto marrow crop (Cucurbita pepo Linnaeus,1753) cultivated in greenhouse conditionsin southern Spain (Hymenoptera: Apidae).Entomofauna, 14(28): 465-469.
Singh P.P., Sinha S.N. (1996) - Effect ofBee-Q on honeybee visits and seed yield ofhybrid sunflower. Seed Research, 24(2):151-153.
Szalai Z. (2000) - Application of pollenmixing device in front of the hive entrance.International J. of Horticultural Science,6(3): 151-152.
Vaknin Y., Gan-Mor S., Bechar S.,Ronen B., Eisikowitch D. (2000) -The role of electrostatic forces inpollination. Plant Sys. Evol., Special issue:Pollen and pollination, 222(1-4): 133-142.
Winston M.J., Slessor K.N. (1993) -Applications of queen honey beemandibular pheromone for beekeeping andcrop pollination. Bee World, 74: 111-128.
Zvedenok A.P. (196) - [The onion seedcrop can be improved]. Kartofel’ IOvoshchi, No.4: 29.
ZAPYLANIE DRZEW OWOCOWYCH PRZEZ PSZCZO£Y:OSTATNIE POSTÊPY I PERSPEKTYWY BADAÑ I.
Vol. 47 No. 1 2003 Journal of Apicultural Science 93

B e n e d e k P .
S t r e s z c z e n i e
Podczas minionej dekady prowadzono wiele szczegó³owych badañ dotycz¹cych zapylaniaprzez pszczo³y drzew owocowych strefy umiarkowanej. Wielki postêp zosta³ osi¹gniêty kiedyodkryto, ¿e kwiaty wielu odmian posiadaj¹ cechy oddzia³uj¹ce na aktywnoœæ pszczó³ miodnychna kwiatach i ich wydajnoœæ jako zapylaczy. Odnalezione cechy kwiatów by³y zdecydowanieró¿ne w przypadku odmian jab³oni i gruszy w poszczególnych latach ale w przypadkuniektórych odmian gruszy ró¿nice miêdzy kolejnymi latami by³y wiêksze ni¿ miêdzyodmianami w danych latach. Koniecznoœæ zapylania przez pszczo³y zosta³a wyraŸnie wykazanazarówno w przypadku obcopylnych jaki i samopylnych odmian owoców. Stopieñ wiernoœcikwiatowej zbieraczek pszczo³y miodnej by³ ró¿ny na poszczególnych gatunkach drzewowocowych ale rola po¿ytków konkurencyjnych okaza³a siê byæ mniej szkodliwa ni¿stwierdzano to wczeœniej w literaturze. Jednak¿e, pewna liczba pytañ powsta³a czêœciowow wyniku najnowszych badañ nad zapylaniem i czêœciowo z praktycznych doœwiadczeñw polowej produkcji roœlin. Pytania te wskazuj¹ kilka zagadnieñ do badania, by zrozumieæi rozwi¹zaæ problemy prawdopodobnie w najbli¿szej przysz³oœci. Koncentruj¹ siê one naskutecznoœci wizyt pszczó³ w zapylaniu poszczególnych upraw owoców i ró¿nych ich odmianoraz na wiarygodnej ocenie ca³kowitej liczby pszczó³ jak równie¿ na kontroli zagêszczeniapszczó³ podczas okresu kwitnienia sadów owocowych dla optymalnego, kontrolowanegozapylania ich przez pszczo³y. Du¿o mniej wysi³ku zrobiono, by wykorzystaæ rodzime dzikiepszczo³y samotnice dla zapylania drzew owocowych, jednak¿e, niektóre murarki wydaj¹ siê byæodpowiednie do tego w Europie, na Dalekim Wschodzie i Ameryce Pó³nocnej.
S³owa kluczowe : zapylanie przez owady, pszczo³y miodne, plonowanie owoców w strefieumiarkowanej.
94

BEE POLLINATION OF FRUIT TREES:RECENT ADVANCES AND RESEARCH PERSPECTIVES II.
P á l B e n e d e k
University of West Hungary, Faculty of Agricultural and Food Sciences,H-9201 Mosonmagyaróvár, Vár 4., Hungary.
S u m m a r y
Intense research was carried out on the bee pollination of temperate zone deciduous fruit treesduring the past decade. Much progress was achieved to explore the flower characters of a greatnumber of cultivars affecting honeybee activity at flowers and their pollinating efficiency.Flower characters were found to be consequently different among apple and pear cultivars inconsecutive years but in case of some stone fruits differences between consecutive years weregreater than between cultivars in given years. The necessity of bee pollination was clearlydemonstrated both at self-sterile and at self-fertile fruit cultivars. The rate of flower constancyof honeybee foragers was different at different fruit tree species but the role of competing plantswas found to be much less deleterious than stated in the literature earlier. However, a number ofquestions arose partly from the results of latest pollination research and partly from practicalexperiences in commercial plant production. These indicate several research tasks to understandand to solve the problems possibly in the near future. The questions concentrate on theeffectiveness of bee visits in the pollination of individual fruit crops and their different cultivarsand on the reliable estimate of the overall amount of bees as well as on the control of beedensity during the flowering period of fruit orchards for optimal, controlled honeybeepollination. Much less effort was made to manage native wild bees for fruit tree pollination,however, some mason bees seem to be promising for this purpose in Europe, The Far East andNorth America.
Keyworlds: insect pollination, honeybees, temperate zone deciduous fruit crops.
INTRODUCTIONAlmost all temperate zone fruit tree spe-
cies need insect pollination, the only excep-tion is the horse chestnut, which is windpollinated, but partly benefits insect pollina-tion, too. The role of insects in fruit croppollination has been recognized long ago.The information accumulated on this fieldhas been reviewed by Free (1993) in thecomprehensive handbook “Insect pollina-tion of crops (second edition)” some tenyears ago. After the appearance of this bookfruit pollination research has remained in-tense. Important new information accumu-lated in this field, however, several newproblems arose that needs further research.
The aim of this paper is to give an outlookonto the state of knowledge in this field andto point out the need for further research.
RESULTSFlowers characters and other conditionsaffecting bee activity
Pollination requirements of temperatezone fruit crops, their pollinating insects,the rewards to attract bees and the foragingbehaviour of bees on their flowers as wellas the effect of bee pollination on the yieldis fairly well known (Free 1993). Lately,intensive studies were carried out on varietyfeatures affecting bee activity,
There were some few indications in the
Vol. 47 No. 1 2003 Journal of Apicultural Science 95

literature that some differences betweencultivars, for example the relative positionof petals and stamens, or the nectar and pol-len production of flowers had a definite in-fluence on bee behaviour and on thepollinating efficiency of bees at some in-stances (see in Free 1993), this problem,however, has not been not adequately stud-ied and no more than sporadic informationwas available on very few cultivars of someselected fruit species, first if all of apple andpear. For this reason lately large series ofcultivars were inspected for various fruitspecies, first of apple (Benedek et al.1989b, Benedek and Nyéki 1996a,Devary-Nejad et al. 1993), thereafter ofapricot (Benedek et al. 1991b, 1995),peach and nectarine (Benedek et al.1991a, Benedek and Nyéki 1996b),sweet and sour cherry (Benedek et al.1990, 1996) and plum (Szabó et al. 1989,Benedek et al. 1994). Differences weredetected among cultivars in flower sizes, inthe number of stamens/flower, in the rela-tive position of petals to stamens and to pis-tils, in the nectar content of flowers, in thesugar concentration in nectars, in the anthersize and in the pollen production, and fi-nally in the intensity of bee visitation at theflowers. In case of apricot, flower charac-teristics varied considerably depending onthe year and site, greater differences oc-curred at the same cultivar in consecutiveyears than between cultivars in given years.Similarly, no consistent differences werefound in the bee visitation at a number ofalmond cultivars in India; floral preferencesvaried from year to year (Thakur et al.1995). Contrarily, at the rest of the men-tioned fruit species, the differences in theflower characteristics of cvs were consistentin different years, and consequently, alsotheir bee visitation differed. In the case ofpear, the opening order of flowers withininflorescence was different; cvs fell intothree types of opening order which also dif-fered in the average number of bee visits
per flower; this was explained by the differ-ent number of flowers and differences in thelongevity of flowers within the inflores-cence of the tree types (Dibuz el al. 1997,1998).
In other studies the nectar content andthe bee visitation of several almond (Abrol1995), peach (Nyéki et al. 2000), pear(Benedek et al. 2000a) and quincecultivars were compared (Benedek et al.2000d, 2000e); cultivars with higher sugarconcentration, i.e. with higher caloric re-wards usually attracted more foraging in-sects compared to other cultivars of thesame fruit species, except pear (Abrol1995, Benedek et al. 1997, 1998b, 2000e,Benedek 2000). Nectar production of pearis generally regarded to be very poor in theliterature (Free 1993); in fact, it has beenshown lately that pear flowers could pro-duce a plenty of nectar, but the amount ofnectar was extremely dependent on theweather (Benedek et al. 2000a). However,pear nectar always contains very littleamount of sugar (Benedek et al. 2000a)which is not attractive to bees at all (Free1993). That is why honeybees collect al-most exclusively pollen on pear, even withhigh nectar production (Benedek et al.1997, 1998b, Benedek and Nyéki 1997a,Benedek 2000).
Comparing the nectar production and thebee visitation of fruit tree flowers at the spe-cies level, the amount of nectar/flower wasthe highest at sour cherry, followed by apri-cot, apple, plum, peach and nectarine; pearalways produce very small but sometimescan produce very high amount of nectar(Benedek and Nyéki 1997a). The meansugar concentration of pear nectar was al-ways very low, regardless the amount, andthe sugar concentration of nectar was thehighest at sour cherry, followed by apple,plum, peach and nectarine (Benedek andNyéki 1997a). Mean intensity of bee visi-tation was also different according to thefruit species, and again, it was clearly
96

related to the sugar concentration of thenectar (except pear), but the amount of nec-tar had no effect on the bee visitation at thespecies level (Benedek and Nyéki 1997a).
Different factors, weather conditionsfirst of all, can restrict pollinating insects towork the flowers of fruit trees during theirblooming period. In spite of this, very littleinformation has been available in the litera-ture on the rate of limitation that affectedthe fruit set and yield. For this, reason ex-periments were made on this issue latelywith apple (Benedek et al. 1989a,Benedek and Nyéki 1996c, 1997b), withpear (Benedek et al. 2000b), with quince(Benedek et al. 2000f), with sour cherry(Benedek et al. 1990), with plum (Szabóet al. 1989, Benedek et al. 1994) and withapricots (Benedek et al. 2000b). Summa-rizing the results of relevant experiments,the main conclusion was, that in the case ofself-sterile fruit species and cultivars (apple,pear, quince, some plums, some sour cher-ries) even partial limitation of the effectiveduration of the bee pollination period sig-nificantly reduced the fruit set and the yield;in the case of self-fertile fruits (some plums,some sour cherries, some apricots), on theother hand, the effect of partial limitationwas usually small, and the complete (or in-complete but strong) limitation resulted in astrong reduction of yields; it means that notonly self-sterile, but also self-fertile fruitcultivars definitely need insect pollination(Benedek and Nyéki 1995, 1996d,Benedek et al. 2000b).
Lately flower constancy of honeybees atfruit trees was investigated by analysing thepollen loads of pollen gatherer honeybeescaptured at fruit tree flowers (Benedek andNagy 1995, Benedek et al. 2000c); the fi-delity of bees was fairly high for pear, but itwas smaller for apricot, much smaller forapple and even smaller for sour cherry.Competing plants influenced the flowerconstancy of honeybees and the influencewas greatly different for fruit species; it was
very little at pear (Benedek et al. 1998a,Benedek 2000). High fidelity of pollengatherers was also observed towardscultivars, since bees returning to the hiveshad 80-90% of their loads from only onecultivar out of the 5 or 6 in an almond or-chards (Vezvaei and Jackson 1997,DeGrandi-Hoffman et al. 1992). How-ever, bees with high amount of pollen froma single cultivar usually had some pollenfrom other cultivars, too (DeGrandi--Hoffman et al. 1992).
The gene flow, as investigated withisozyme markers, was quite restricted in al-mond orchards in Australia, being the stron-gest between neighbouring halves of cross--compatible pairs of trees (Jackson andClarke 1991, Vezvaei and Jackson1997). However, the nut set on branches ad-jacent to compatible pollen sources did notdiffer significantly from the set on branchesadjacent to the trees of the same cultivar inone other experiment in the US(DeGrandi-Hoffman et al. 1992).
Managing wild bees and other insects forfruit tree pollination
Managing and utilizing wild bees aspollinators attract more and more interest,because some wild bees are relatively easyto manage and they posses several advan-tages as pollinators compared to honeybees.Wild bees, some solitary bees and bumble-bees, however, have been exploited for fieldcrop pollination and for insect pollinationunder cover because (Richards 1993,Cane 1997, Ruijter 1997) because themanaged species are not specialised visitorsof the Rosaceae plant family.
Managing mason bees for fruit treepollination: It is a fairly new developmentto try managing different mason bees forpollination. In temperate regions, Osmiacornuta seems to be the most promising ma-son bee to this purpose. Its normal seasonalactivity starts very early spring, and coin-cides well with the flowering period of fruit
Vol. 47 No. 1 2003 Journal of Apicultural Science 97

trees (Bosch et al. 1993). There are someserious parasites that can restrict its popula-tion size (Bosch 1992, Bosch et al. 1993).Pollen collected for provisions show thatthe species prefers to collect Prunus pollen,and switches to Malus pollen when it ismore readily available (Márquez et al.1994). It is calculated that during almondflowering, each female potentially visit asmuch as 9.5 to 23 thousand flowers, andtherefore 3 females/tree are sufficient for al-mond pollination (Bosch 1994a). Cocoonswere exposed to different overwinteringand incubating temperatures trying to ma-nipulate the time of emergence but with lit-tle success so far (Bosch and Blas 1994).Suitable nesting material was searched for(Bosch 1994b, 1995), but further researchis needed. Osmia cornuta has been tried tobe used for blackberry pollination in plastictunnel with success (Pinzauti et al. 1997).Very probably it will be suitable to pollinateother fruit crops too, hence this issue alsoneeds further studies. Some other masonbees, Osmia lignaria propinqua, O.californica, O. Montana, were also success-fully managed for mobile orchard pollina-tion in the US (Torchio 1991), andO. cornifrons for Japanese pear and applepollination in the Far East (Maeta et al.1993, DooHyun et al. 1996, Sekita2001). There are some additional proposalsto manage some other solitary bees for croppollination; Andrena flavipes - which alsooccurs in Europe - was proposed to managein India (Abrol 1993) and Anthophorapilipes villosula in Japan (Batra 1994).
An other European early spring masonbee species, Osmia rufa, closely related toO. cornuta, is regarded as unsuccessful fororchard pollination because this species ex-ploits much wider range of pollen sourcesand no pollen preference exists towardsfruit trees (Ricciardelli d’Albore et al.1994).
CONCLUSIONSBased on the above discussion there
seem to be a number of topics that greatlyneeded intense research in the near future toimprove the knowledge on as well as thetechnology of the insect pollination of cropplants cultivated in temperate zone regions.These are partly related to the pollinationrequirements of selected crops and to man-aging some solitary wild bee species forcrop pollination. The problems, however,concentrate on the effectiveness of bee vis-its in the pollination of individual fruit crops(and their different cultivars) and on the re-liable estimate of the stocking rates of beesfor crop pollination as well as on the man-agement of actual bee density during theflowering period of fruit orchards for opti-mal controlled bee pollination. The follow-ing topics are proposed as the subject offurther research in the coming years:1. Flowers characters and other conditions
affecting bee activity— Further studies are needed on the
flower constancy of honeybees onfruit trees as related to their pollinat-ing efficiency.
— The pollinating efficiency of honey-bees on self-incompatible fruit treespecies should be re-evaluated by in-vestigating the gene flow withisozyme markers.
2. Managing wild bees for orchard pollina-tion
— Further research is needed to developrearing techniques for commercialusage of the European orchard ma-son bee, Osmia cornuta, for fruit treepollination. One other closely relatedspecies, Osmia rufa, might be suc-cessful for a number of crops undercover.
98

REFERENCESAbrol D.P. (1993) - New pollinator bee.
Current Res. - Univ. Agric. Sci.(Bangalore), 22(9/10): 130.
Abrol D.P. (1995) - Energetics of nectarproduction in some almond cultivars as apredictor for floral choice by honeybeesApis cerena indica F. and Apis mellifera L.(Hymenoptera: Apidae). Proc IndianNational Science Academy, Part B,Biological Sciences, 61: 285-289.
Batra S.W.T. (1994) - Anthophora pilipesvillosula Sm. (Hymenoptera:Anthophoridae), a manageable Japanesebee that visits blueberry and apples duringcool, rainy, spring weather. Proc. Entomol.Soc., Washington, 96: 98-119.
Benedek P. (in press) - Bee pollination offruit trees and the plant protection practice.In: Kozma P., Nyéki J., Soltész M. editors:Floral biology, pollination and fertilisationin temperate zone fruit species and grape.Akadémiai Kiadó, Budapest
Benedek P., Kocsisné Molnár G., NyékiJ.(2000a) - Nectar production of pear(Pyrus communis L.) cultivars. Internat. J.Horticultural Science, 6(3): 65-75.
Benedek P., Nagy Cs. (1995) -Honeybee flower constancy on some fruittree species. Horticultural Science, 27(3/4):38-42.
Benedek P., Nyéki J. (1995) - Role ofbee pollination in the fruit set and yield ofself-fertile and self-sterile apple, sourcherry and plum cultivars. HorticulturalScience, 27: (3-4): 34-37.
Benedek P., Nyéki J. (1996a) -Pollinating efficiency of honeybees onapple cultivars as affected by their flowercharacteristics. Horticultural Science, 28:(1-2): 40-47.
Benedek P., Nyéki J., (1996b) - Studieson the bee pollination of peach andnectarine. Acta Horticulturae, No. 374:169-176.
Benedek P., Nyéki J. (1996c) -Relationship between the duration of insectpollination and the yield of some applecultivars. Horticultural Science, 2(3-4):93-96.
Benedek P., Nyéki J. (1996d) - Fruit setof selected self-sterile and self-fertile fruitcultivars as affected by the duration ofinsect pollination. Acta Horticulturae, No.423: 57-63.
Benedek P., Nyéki J. (1997a) -Considerations on the nectar productionand the honeybee visitation of fruit treeflowers. Horticultural Science, 29(3-4):117-122.
Benedek P., Nyéki J. (1997b) - Yield ofselected apple cultivars as affected by theduration of bee pollination. ActaHorticulturae, No. 437: 207-211.
Benedek P., Nyéki J., Lukács Gy.(1989a) - A méhmegporzás intenzításánakhatása az alma kötõdésére és termésére.Kertgazdaság, 21(3): 8-26.
Benedek P., Nyéki J., Soltész M., ErdõsZ., Skola I., Szabó T., Amtmann I.,Bakcsa F., Kocsisné Molnár G.,Vadas Z., Szabó Z. (2000b) - Theeffect of the limitation of insect pollinationperiod on the fruit set and yield oftemperate-zone fruit tree species. Internat.J. Horticult. Science, 6(1): 91-95.
Benedek P., Nyéki J., Szabó Z. (1990)- Cseresznye és meggyfajtákméhmegporzást befolyásoló tulajdonságai.Kertgazdaság, 22(5): 1-23.
Benedek P., Nyéki J., Szabó Z. (1991a)- Õszibarack fajták méhmegporzást
befolyásoló tulajdonságai. Kertgazdaság,23(1): 40-58.
Benedek P., Nyéki J., Szabó Z. (1991b)- Kajszifajták méhmegporzást befolyásoló
tulajdonságai. Kertgazdaság, 23(2): 27-39.
Benedek P., Nyéki J., Szabó Z. (1995)- Bee pollination of apricot: variety featuresaffecting bee activity. Acta Horticulturae,No. 384: 329-332.
Benedek P., Nyéki J., Szabó Z. (1996)- Features affecting bee pollination of sweetand sour cherry varieties. ActaHorticulturae, 410: 121-126.
Bosch J. (1992) - Parasitism in wild andmanaged populations of the almondpollinator Osmia Bosch, J. (1994a): Thenesting behaviour of the mason bee Osmiacornuta (Latr.) with special reference t itspollination potential (Hymenoptera,Megachilidae). Apidologie, 25: 84-93.
Vol. 47 No. 1 2003 Journal of Apicultural Science 99

Bosch J. (1993) - Parasitism in wild andmanaged populations of the almondpollinator Osmia cornuta Latr.(Hymenoptera, Megachilidae). J. Apicult.Res., 31: 77-82.
Bosch J. (1994b) - Osmia cornuta Latr.,(Hymenoptera: Megachilidae) as apotentialpollinator in almond orchards. J. Appl.Entomol., 117: 151-157.
Bosch J. (1995) - Comparison of nestingmaterials for the orchard pollinator Osmiacornuta (Hymenoptera: Megachilidae).Entomologia Generalis, 19: 285-289.
Bosch J., Blas M. (1994) - Effect ofover-wintering and incubation temperatureson adult emergence in Osmia cornuta Latr.(Hymenoptera, Megachilidae). Apidologie,25: 265-277.
Bosch J., Blas M., Lacasa A. (1993) -Una abeja solitaria de interés agrícola. VidaApícola, No. 59:36-37, 39, 41, 43.
Cane J.H. (1997) - Ground-nesting bees:the neglected pollinator resource foragriculture. Acta Horticulturae, 437:309-323.
Davary-Nejad G.H., Szabó Z., Nyéki J.,Benedek P. (1993) - Almafajtákvirág-tulajdonságai és méhmegporzása.Kertgazdaság, 25(2): 72-88
DeGrandi-Hoffman G., Martin J.H.(1995) - Does a honey bee (Apismellifera) colony’s foraging population onmale-sterile sunflowers (Helianthusannuus) affect the amount of pollen onnestmates foraging on male-steriles. J.Apicultural Res., 34: 109-114.
DeGrandi-Hofman G., Mayer D., TerryL., DongHui L. (1995) - Validation ofPC-REDAPOL: fruit set prediction modelfor apples. J. Economic Entomology, 88:965-972.
DeGrandi-Hoffman G., Thorp R., LoperG., Eisikowitch D. (1992) -Identification and distribution ofcross-pollinating honey-bees on almonds. J.Appl. Ecol., 29: 238-246.
Dibuz E., Benedek P., Soltész M.,Nyéki J., (1997) - Bee visitation andfruit set of pear as affected by the openingsequence of flowers in the inflorescence.Horticultural Science, 29(3-4): 129-136.
Dibuz E., Benedek P., Soltész M.,Nyéki J. (1998) - Relationship betweenthe type of inflorescence and the beepollination of pear cultivars. ActaHorticulturae, 475: 223-229.
DooHyun C., JinSoo K., JaeTak Y.,BooSull C. (1996) - [Effects of Osmiacornifrons on fruit setting in Fuji appletrees]. RDA J. Agricultural Science, CropProtection, 38(2): 382-386. (In Korean withEnglish summary).
Free J.B. (1993) - Insect pollination ofcrop. Second edition. Academic Press,London.
Jackson J.F., Clarke G.R. (1991) - Geneflow in an almond orchard. Theoretical andApplied Genetics, 82: 169-173.
Maeta Y., Goukon K., Tezuka T. (1993)- [Utilization of Osmia cornifrons as a
pollinator of Japanese pears (Hymenoptera,Megachilidae)]. Chugoku Kontylu, No.7:1-12. (In Japanese with English summary).
Márquez J., Bosch J., Vincens N.(1994) - Pollens collected by wild andmanaged populations of the potentialorchard pollinator Osmia cornuta (Latr.)(Hym., Megachilidae). J. Appl. Entomol.,117: 52-359.
Nyéki J., Szabó Z., Benedek P., SzalayL. (2000) - Nectar production andpollination in peach. Internat. J.Horticultural Science, 6(3): 123-126.
Pinzauti M., Lazzarini D., Felicioli A.(1997) - Preliminary investigations ofOsmia cornuta Latr. (Hymenoptera,Megachilidae) as potential pollinator forblackberry (Rubus fruticosus L.) underconfined environment. Acta Horticulturae,437: 329-333.
Richards K.W. (1993) - Non-Apis bees ascrop pollinators. Rev. Suisse Zoologie, 100:807-822.
Riciardelli d’Albore G., Quaranta M.,Pinzauti M. (1994) - Studiomicroscopico della dieta Osmia cornigeraRossi, in Umbria. In: Atti XVII CongressoNazionale Italiano de Entomologia (Udine,Italy, 13-18 Giugno 1994). AacademiaNazionale Italina di Entomol. Soc. Italiano:843-846.
100

Ruijter A. de (1997) - Commercialbumblebee rearing and its implications.Acta Horticulturae, 437: 261-269.
Ruijter A. de, Eijnde J. van der, StehenJ. van der (1997) - Diseases and pestsfound in bumblebee rearing. Apidologie,28: 222-225.
Sekita N. (2001) - Managing Osmiacornifrons to pollinate apples in AomoriPrefecture, Japan. Acta Horticulturae, 561:303-307.
Szabó Z., Nyéki J., Benedek P. (1989)- A mézelõ méhek tevékenységeszilvafákon, szerepük a megporzásban és agyümölcskötodésben. Kertgazdaság, 21(1):53-70.
Thakur S.S., Mishra R.C., Kumar J.,(1995) - Floral preference of honey beesto different cultivars of almond (Prunusamygdalus Batsch.). Indian Bee J., 54(4):163-165.
Torchio P.F. (1991) - Use of Osmialignaria propinqua (Hymenoptera:Megachilidae) as a mobile pollinator oforchard crops. Environmental Entomology20: 590-596.
Vezvaei A., Jackson J.F. (1997) - Geneflow by pollen in an almond orchard asdetermined by isozyme analysis ofindividual kernel and honey bee pollenloads. Acta Horticulturae, 437: 75-81.
ZAPYLANIE DRZEW OWOCOWYCH PRZEZ PSZCZO£Y:OSTATNIE POSTÊPY I PERSPEKTYWY BADAÑ II.
B e n e d e k P .
S t r e s z c z e n i e
Podczas minionej dekady prowadzono wiele szczegó³owych badañ dotycz¹cych zapylaniaprzez pszczo³y drzew owocowych strefy umiarkowanej. Wielki postêp zosta³ osi¹gniêty kiedyodkryto, ¿e kwiaty wielu odmian posiadaj¹ cechy oddzia³uj¹ce na aktywnoœæ pszczó³ miodnychna kwiatach i ich wydajnoœæ jako zapylaczy. Odnalezione cechy kwiatów by³y zdecydowanieró¿ne w przypadku odmian jab³oni i gruszy w poszczególnych latach ale w przypadkuniektórych odmian gruszy ró¿nice miêdzy kolejnymi latami by³y wiêksze ni¿ miêdzyodmianami w danych latach. Koniecznoœæ zapylania przez pszczo³y zosta³a wyraŸnie wykazanazarówno w przypadku obcopylnych jaki i samopylnych odmian owoców. Stopieñ wiernoœcikwiatowej zbieraczek pszczo³y miodnej by³ ró¿ny na poszczególnych gatunkach drzewowocowych ale rola po¿ytków konkurencyjnych okaza³a siê byæ mniej szkodliwa ni¿stwierdzano to wczeœniej w literaturze. Jednak¿e, pewna liczba pytañ powsta³a czêœciowow wyniku najnowszych badañ nad zapylaniem i czêœciowo z praktycznych doœwiadczeñw polowej produkcji roœlin. Pytania te wskazuj¹ kilka zagadnieñ do badania, by zrozumieæi rozwi¹zaæ problemy prawdopodobnie w najbli¿szej przysz³oœci. Koncentruj¹ siê one naskutecznoœci wizyt pszczó³ w zapylaniu poszczególnych upraw owoców i ró¿nych ich odmianoraz na wiarygodnej ocenie ca³kowitej liczby pszczó³ jak równie¿ na kontroli zagêszczeniapszczó³ podczas okresu kwitnienia sadów owocowych dla optymalnego, kontrolowanegozapylania ich przez pszczo³y. Du¿o mniej wysi³ku zrobiono, by wykorzystaæ rodzime dzikiepszczo³y samotnice dla zapylania drzew owocowych, jednak¿e, niektóre murarki wydaj¹ siê byæodpowiednie do tego w Europie, na Dalekim Wschodzie i Ameryce Pó³nocnej.
S³owa kluczowe : zapylanie przez owady, pszczo³y miodne, plonowanie owoców w strefieumiarkowanej.
Vol. 47 No. 1 2003 Journal of Apicultural Science 101
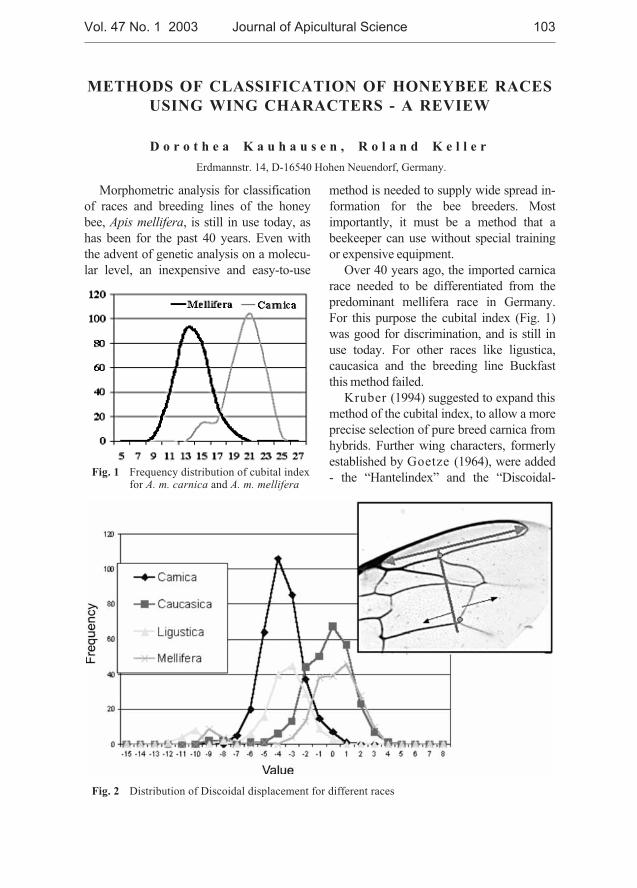
METHODS OF CLASSIFICATION OF HONEYBEE RACESUSING WING CHARACTERS - A REVIEW
D o r o t h e a K a u h a u s e n , R o l a n d K e l l e r
Erdmannstr. 14, D-16540 Hohen Neuendorf, Germany.
Morphometric analysis for classificationof races and breeding lines of the honeybee, Apis mellifera, is still in use today, ashas been for the past 40 years. Even withthe advent of genetic analysis on a molecu-lar level, an inexpensive and easy-to-use
method is needed to supply wide spread in-formation for the bee breeders. Mostimportantly, it must be a method that abeekeeper can use without special trainingor expensive equipment.
Over 40 years ago, the imported carnicarace needed to be differentiated from thepredominant mellifera race in Germany.For this purpose the cubital index (Fig. 1)was good for discrimination, and is still inuse today. For other races like ligustica,caucasica and the breeding line Buckfastthis method failed.
Kruber (1994) suggested to expand thismethod of the cubital index, to allow a moreprecise selection of pure breed carnica fromhybrids. Further wing characters, formerlyestablished by Goetze (1964), were added- the “Hantelindex” and the “Discoidal-
Vol. 47 No. 1 2003 Journal of Apicultural Science 103
Fig. 2 Distribution of Discoidal displacement for different races
Fig. 1 Frequency distribution of cubital indexfor A. m. carnica and A. m. mellifera
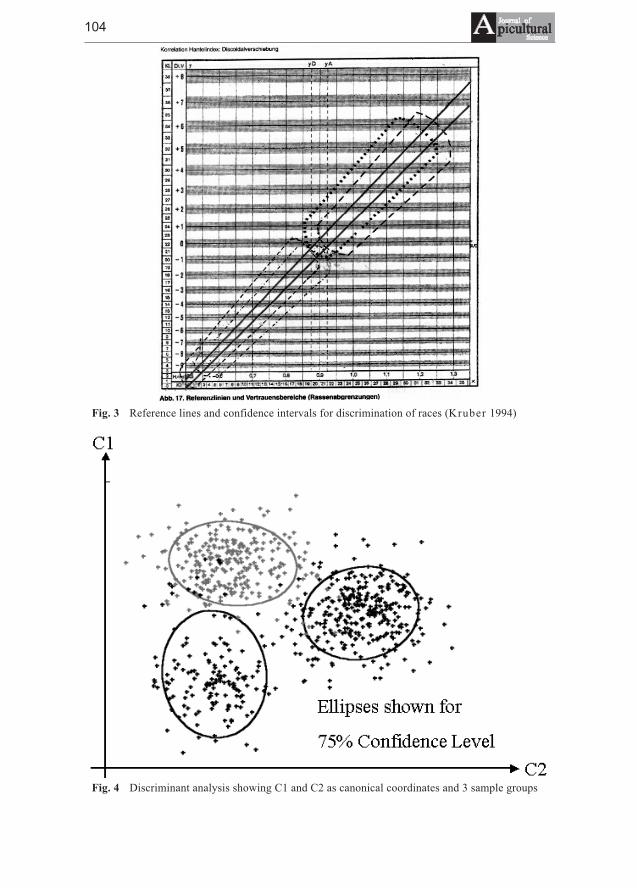
104
Fig. 4 Discriminant analysis showing C1 and C2 as canonical coordinates and 3 sample groups
Fig. 3 Reference lines and confidence intervals for discrimination of races (Kruber 1994)

verschiebung” (Fig. 2). The results of thesetwo characters represented in a diagram,would allow better differentiation betweencarnica and mellifera, and their hybrids(Fig. 3). With other races, like A. m.ligustica or A. m. caucasica, however, a dif-ferentiation is still not better than with thecubital index.
A major step forward was undertaken byProf. F. Ruttner (1988), who used 15wing characters, 13 size characters, lengthof cover hair and 2 characters of thetomentum as well as 5 characters of pig-mentation. These 36 different characterswere used in a statistical method called“discriminant analysis” which reduces themany characters to a few “best canonicalfactors” from which two or three dimen-sional distribution plots could be made. Apriori predictions were made by choosing
certain confidence levels expressed in dis-tance from the group centroid (Fig. 4).Using this method, it was possible forRuttner to classify among twenty four dif-ferent subspecies of Apis mellifera. Thepreparation and measurement of sampleswas quite tedious - nearly a whole weekwas required for one single sample oftwenty working bees from a colony.
In the following years, various develop-ment in using digitizers linked with a com-puter for faster data taking went under way.Video cameras soon took the place of digi-tizing tablets, and finally slide scanners(Fig. 5), which did not need optical lenses.Since measurements of wing characterswere the most precise, with a high repeat-ability, focus was set on these, and the otherbody characters were neglected.
Vol. 47 No. 1 2003 Journal of Apicultural Science 105
Fig. 6 Position of group centroids using only wing characters (introduced by DuPraw 1964)
Fig. 5 Slide scanner with PC

Using only the wing characters, intro-duced by DuPraw (1964), gives a fairlygood discrimination between mellifera,carnica, ligustica and caucasica, to take afew of the common Central European sub-species (Fig. 6). DuPraw took the positionof 18 vein intersections on the forewing,drew a standard wing diagram and mea-sured 13 angles and two size characters.
An attempt was made by Kauhausen--Keller and Keller (1994) to improve thisdiscrimination by adding characters of theassociated hind wing - with success. How-ever, the hind wing is quite difficult to han-dle, and to mount on a slide.
A latest and simplest method of usingwing characters was to discard the measur-ing and calculating of wing characters likeDuPraw, and using pure geographic coordi-nates of the centres of the vein intersectionson the forewing (Fig. 6). Also the scanningand digitizing techniques were improved tosuch a degree that easy handling and precise
measurements were possible. The smallhind wing can bee neglected. Sufficient dis-crimination can be achieved among thementioned subspecies (Fig. 7). Using thismethod, the determination of racial purity isas simple to handle as when determiningthe cubital index.
CONCLUSIONSWing characters have proven to be the
most reliable in measurement, and easiest tohandle. During mounting, the wing does notchange in size or structure, as it is withother body parts. Also, the geographic coor-dinates seem to exhibit a wide variability,correlated with different races. Hence, forthe determination of race or racial purity (ordegree of hybridization), this methodproves to be a quick and reliable techniquewhich can also be used by bee breeders.
Keywords: morphological characters, wing, discriminant analysis, honeybee races.
REFERENCESDuPraw E.J. (1964) - Non Linnean
taxonomy. Nature 202 (4935):849-852.
Goetze G. (1964) - Die Honigbiene innatürlicher und künstlicher Zuchtauslese.Paul Parey, Hamburg.
Kauhausen-Keller D., Keller R. (1994)- Morphometrical control of pure race
breeding in the honey bee (Apis melliferaL.). Apidologie, 25, 133-143.
Kruber W. (1994) - Hantelindex undDiskoidalverschiebung. Die biene, 1:8-18,2:62-77, 3:125-135.
106
Fig. 7 Location of geographic coordinates on the forewing

Ruttner F. (1988) - Biogeography andTaxonomy of Honeybees. Springer, Berlin.
METODY OCENY PRZYNALE¯NOŒCI RASOWEJ PSZCZÓ£ZA POMOC¥ CECH MORFOLOGICZNYCH SKRZYDE£
- PRZEGL¥D
K a u h a u s e n D . , K e l l e r R .
S t r e s z c z e n i e
Pomiar cech morfologicznych pszczó³ do oceny ich przynale¿noœci rasowej prowadzony jestod ponad 40 lat. Ras¹ dominuj¹c¹ w Niemczech jest pszczo³a kraiñska. Od pszczo³yœrodkowoeuropejskiej ³atwo j¹ odró¿niæ wartoœci¹ indeksu kubitalnego (rys. 1), trudniej odinnych ras (w³oska, kaukaska). Do przeprowadzenia pe³nej analizy morfologicznejwykonywano dotychczas pomiar 36 ró¿nych cech, jest to jednak bardzo pracoch³onne. Okaza³osiê, ¿e do dobrego odró¿nienia wystarczy pomiar 18 cech na skrzydle pszczo³y. Wykonanie tychpomiarów na ekranie komputera i poddanie ich analizie dyskryminacji przy u¿yciu specjalnegoprogramu komputerowego pozwala na stosunkowo szybkie i ³atwe odró¿nienie ras i liniipszczó³.
S³owa kluczowe: cechy morfologiczne, skrzyd³o, analiza dyskryminacji, rasy pszczó³.
Vol. 47 No. 1 2003 Journal of Apicultural Science 107

RENOVATION OF J. G. MENDEL'S APIARY
V l a d i m i r P t a c e k
Masaryk University, Faculty of Science, Kotlarska 2, 611 37 Brno, Czech Republice-mail: [email protected]
As is well known, the discoverer of ge-netic laws - J. G. Mendel - was also an ac-tive scientist in apiculture. He tried to findout the best way of wintering productivecolonies, conducted experiments with mat-ing queens and selected drones in a cage,and compared productivity and charactersof different honey bee races. Mendel evenobserved and wintered a colony of Trigonasting-less bees. He presented his resultsmainly on meetings of the committee of theMoravian Beekeeper's Union, of which hewas a member. Mediated information abouthis results occurred in the beekeeping mag-azine Vèela Brnìnská (Die Honigbiene vonBrünn).
Mendel (1822-1884) lived in the city ofBrno in the southeastern part of what is nowthe Czech Republic. As an abbot of the OldBrno Augustinian Monastery he had a lot ofduties but nevertheless he built a nice apiarywithin the area of the abbey. It was con-structed according to the latest knowledge
of Mendel's time and has two manipulationrooms and the space for placing 15 hives.Behind the house there is a cellar for indoorwintering of colonies. Three original hivesand two copies made in early 1960s remainin the bee-house till now. Unfortunately, theoriginals are mostly rotten. The building it-self, the front wall of the cellar as well asthe fence surrounding the area needs funda-mental renovation, urgently.
On the 180th anniversary of Mendel'sbirth an excellent exhibition called “The ge-nius of genetics” was opened in the OldBrno Abbey on May 15, 2002, and an inter-national workshop of top scientists in thefield took place on this occasion.
Because Mendel was an active memberof the beekeeper's union, the beekeepers ofBrno have taken part in the renovation ofthe apiary, which naturally belongs to Men-del's heritage. The plan is for it to becomean educational site showing the basicknowledge about the life of the honey bee,the kinds of hives from Mendel's time tillnow, as well as the various ways ofbeekeeping. As a beginning, colonies inmodern magazine hives were placed in thearea surrounding the bee house, which itselfhas to be reconstructed first.
The beekeepers of Brno will be gratefulfor any material support in their activities.For any (urgently needed) sponsorship con-tact please the author of the article.
Keywords: Mendel, apiary, apiculture.
ACKNOWLEDGEMENTSThe presentation was possible thanks to
the support of the Masaryk University ofBrno, grant No. CEZ: J07/98:143100010.
Vol. 47 No. 1 2003 Journal of Apicultural Science 109
Fig. 1 J. G. Mendel (July 20,1822 - Jan. 9,1884)

110
Fig. 3 The bee room inside with the old hives (right), all is waiting for conservation andrenovation
Fig. 2 Moravian hive from Mendel's time

ODTWORZENIE PASIEK J. G. MENDLA
P t a c e k V .
S t r e s z c z e n i e
Odkrywca praw genetycznych J. G. Mendel by³ tak¿e aktywnym badaczem pszczelarskim.Wyniki dwóch obserwacji przedstawia³ g³ównie na spotkaniach Morawskiego Zwi¹zkuPszczelarzy. Swoj¹ pasiekê, w postaci pawilonu mieszcz¹cego 15 uli posiada³ w Brnie naterenie klasztoru Augustynów. Podczas obchodzonej w roku 2002, 180 rocznicy urodzinMendla, pszczelarze z Brna podjêli decyzjê o odtworzenie jego pasieki w kszta³cie w jakimistnia³a za jego czasów.
S³owa kluczowe: Mendel, pasieka, pszczelarstwo.
Vol. 47 No. 1 2003 Journal of Apicultural Science 111
Fig. 5 Mendel's experimental garden. Right is situated the entrance to the exhibition
Fig. 4 The modern hives making the apiary living again

PROGRAMM OF THE FIGHT AGAINST RESISTANTVARROA MITES IN CZECH REPUBLIC
V l a d i m í r V e s e l ý
Bee Research Institute at Dol, 252 66 Libèice n. Vlt, Czech Republic
S u m m a r y
Prevention and control of resistant populations of Varroa mites is based on the monitoring ofthe occurrence and on the measures in foci and their protection zones. Monitoring is carried outon the basis of field experiments, where natural fall and efficacy of acrinathrin, fluvalinate andamitraz are compared. Control measures are legalized. They include limitation of movement ofbee colonies, rotation of preparations and increased care in winter time, especially using aerosolform of application. The objective of the measures is interruption of the generation sequence ofresistant mites.
Keywords: resistance, Varroa mite, cross field experiments, pyrethroids, acrinathrin,fluvalinate, amitraz, aerosol administration, generation sequence.
The territory of Czech Republic is likeother European countries threatened withresistant populations of Varroa mites(Milani 2001). But hitherto only individualfoci of resistance to pyrethroids were re-corded and this phenomenon was found insouthern and southeastern border regions.Regardless of this fact we have introducedcertain organization measures (Veselý2002).
The Czech programm of prevention andcontrol of resistant populations consists oftwo parts: First, monitoring of the occur-rence and second, measures in occurrencefoci and their protection zones. Monitoringis carried out on the basis of field cross ex-periments where the efficacy of acrinathrinand fluvalinate at late summer treatment(tab.1) and the efficacy of fluvalinate andamitraz at autumnal treatment (tab. 2) ofbee colonies are compared. Besides that,there is compared the efficacy of carrierswith long time effectiveness in the first daysof exposition with the previous natural fallof mites. The monitoring action is com-pleted by the declaration of the foci and 5km protection zones.
Cross experiments are established on fol-lowing principles: The experimental groupis to include at least 8 bee colonies andhives are to be equipped by double nettpads to prevent the bringing out of mites bybees. This equipment enables daily moni-toring of the fall of mites. Bee colonies ofthis experimental group are divided intoodd and even colonies. Into odd colonies atlate summer experiments we put long timeacting strips with the content of acrinathrin(Gabon PA 92). At autumnal experimentsodd bee colonies are treated by fumigationby amitraz (Varidol FUM). Into even beecolonies at late summer experiments we putlong time acting strips with the content offluvalinate (Gabon PF 90). At autumnal ex-periments we treat these colonies byfluvalinate fumigation (MP-10 FUM). Atlate summer experiments after 15 days wechange the strips mutually between odd andeven colonies. At autumnal experiments wecarry out the second treatment by fumiga-tion after 4 - 7 days. This treatment is recip-rocal, that means odd bee colonies aretreated by fluvalinate and even bee coloniesby amitraz. After the finish of late summer
Vol. 47 No. 1 2003 Journal of Apicultural Science 113
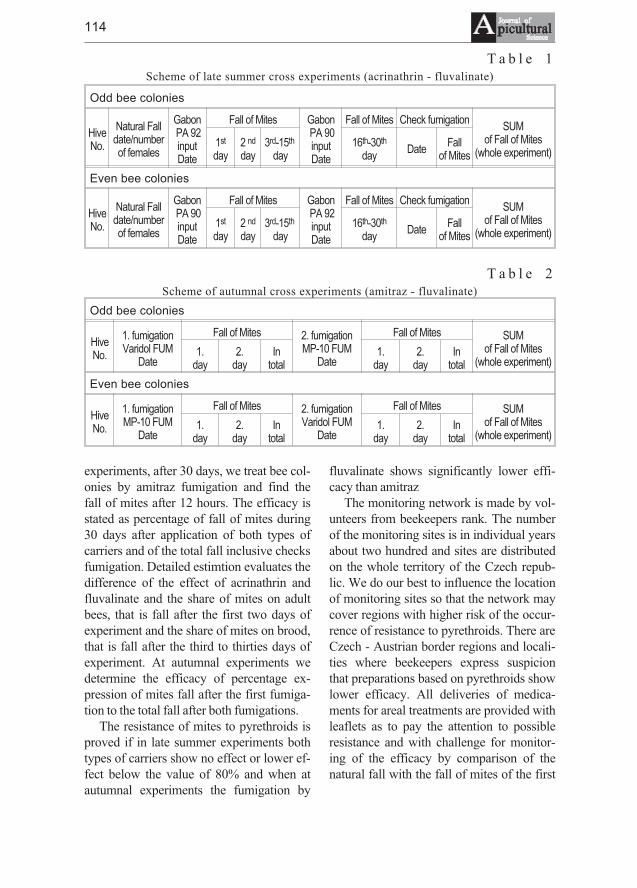
experiments, after 30 days, we treat bee col-onies by amitraz fumigation and find thefall of mites after 12 hours. The efficacy isstated as percentage of fall of mites during30 days after application of both types ofcarriers and of the total fall inclusive checksfumigation. Detailed estimtion evaluates thedifference of the effect of acrinathrin andfluvalinate and the share of mites on adultbees, that is fall after the first two days ofexperiment and the share of mites on brood,that is fall after the third to thirties days ofexperiment. At autumnal experiments wedetermine the efficacy of percentage ex-pression of mites fall after the first fumiga-tion to the total fall after both fumigations.
The resistance of mites to pyrethroids isproved if in late summer experiments bothtypes of carriers show no effect or lower ef-fect below the value of 80% and when atautumnal experiments the fumigation by
fluvalinate shows significantly lower effi-cacy than amitraz
The monitoring network is made by vol-unteers from beekeepers rank. The numberof the monitoring sites is in individual yearsabout two hundred and sites are distributedon the whole territory of the Czech repub-lic. We do our best to influence the locationof monitoring sites so that the network maycover regions with higher risk of the occur-rence of resistance to pyrethroids. There areCzech - Austrian border regions and locali-ties where beekeepers express suspicionthat preparations based on pyrethroids showlower efficacy. All deliveries of medica-ments for areal treatments are provided withleaflets as to pay the attention to possibleresistance and with challenge for monitor-ing of the efficacy by comparison of thenatural fall with the fall of mites of the first
114
Odd bee colonies
HiveNo.
Natural Falldate/numberof females
GabonPA 92inputDate
Fall of Mites GabonPA 90inputDate
Fall of Mites Check fumigation SUMof Fall of Mites
(whole experiment)1st
day2 nd
day3rd-15th
day16th-30th
day Date Fallof Mites
Even bee colonies
HiveNo.
Natural Falldate/numberof females
GabonPA 90inputDate
Fall of Mites GabonPA 92inputDate
Fall of Mites Check fumigation SUMof Fall of Mites
(whole experiment)1st
day2 nd
day3rd-15th
day16th-30th
day Date Fallof Mites
T a b l e 1Scheme of late summer cross experiments (acrinathrin - fluvalinate)
Odd bee colonies
HiveNo.
1. fumigationVaridol FUM
Date
Fall of Mites 2. fumigationMP-10 FUM
Date
Fall of Mites SUMof Fall of Mites
(whole experiment)1.
day2.
dayIn
total1.
day2.
dayIn
total
Even bee colonies
HiveNo.
1. fumigationMP-10 FUM
Date
Fall of Mites 2. fumigationVaridol FUM
Date
Fall of Mites SUMof Fall of Mites
(whole experiment)1.
day2.
dayIn
total1.
day2.
dayIn
total
T a b l e 2Scheme of autumnal cross experiments (amitraz - fluvalinate)

two days with the exposition to medica-ments.
Measures in resistance foci and in pro-tection zones are as follows: ban on trans-port of bee colonies into not resistantregions, ban on use of pyrethroids in all ap-plication forms, substitution of pyrethroidsby formic acid in season time, increasednumber of winter treatments by amitraz inthe aerosol form, winter debris investigationand early spring treatment of sites wheremites in winter debris were found and thisin quantity more that one mite per bee col-ony. Such colonies are treated repeatedly byamitraz in the interval of 10 days and assoon as possible that means if daily temper-ature achieves without larger deviations20°C, by evaporating sheets with formicacid. Methodical Instruction of the StateVeterinary Administration legalized allthese measures. The target is to reach an in-terruption of the generation sequence of
resistant mites by full destruction or by thedecisive reduction of their population.
All efforts connected with the imple-mentation of the programm are justified.Combination of acrinathrin in late summerand amitraz in autumnal and winter treat-ment proved to be highly effective and hy-gienically flawless without negative sideeffects.
REFERENCESMilani, N. (2001) - The resistance to
chemotherapy in parasites and pathogens ofthe honeybee. In: Proceedings ofEuroconference on molecular mechanismsof disease tolerance in honeybees, Kralupy,Oct. 17-19, 2000. Bee Research Institute,Dol, s. 117 - 131.
Veselý, V. (2002) - Kontrola rezistenceroztoèù Varroa v roce 2001.Vèelaøství 55(9): 195-198.
PROGRAM WALKI Z ODPORNYMI POPULACJAMIROZTOCZY Varroa W CZECHACH
V e s e l ý V .
S t r e s z c z e n i e
Zapobieganie i zwalczanie odpornych populacji roztoczy Varroa oparte jest o monitoringwystêpowania i o pomiary ognisk i ich stref ochronnych. Monitoring prowadzony jest w oparciuo doœwiadczenia terenowe, gdzie porównuje siê osyp naturalny i skutecznoœæ akrynatryny,fluvalinatu i amitrazu. Zabiegi zwalczania s¹ zalegalizowane. Obejmuj¹ one przemieszczanierodzin pszczelich, rotacjê (przemienne stosowanie) preparatów i zwiêkszon¹ opiekê w sezoniezimowym, w szczególnoœci stosowanie preparatów w formie aerosolu. Celem zabiegów jest
przerwanie nastêpstwa pokoleñ odpornych roztoczy.
S³owa kluczowe: odpornoœæ, roztocze Varroa, doœwiadczenia terenowe, pyretroidy,akrynatryna, fluvalinat, amitraz, forma aerozolowa aplikacji, nastêpstwopokoleñ, Czechy.
Vol. 47 No. 1 2003 Journal of Apicultural Science 115

Vol. 47 No. 1 2003 Journal of Apicultural Science 117
JOURNAL OF APICULTURAL SCIENCE (notes for authors)
Journal of Apicultural Science is a scientific journal intended to publish original papers in thefield of the broad-sense beekeeping science.
Manuscripts submitted in English and Polish (authors from Poland) should be concise and to thepoint without unnecessary circumlocutions and stylistic figures, written in a simple language.They must not exceed 30,000 characters.
The manuscripts should be sent to: Research Institute of Pomology and Floriculture, ApicultureDivision, 24-100 Pu³awy, ul. Kazimierska 2, Poland
Each submitted manuscript is reviewed. The editor reserves the right to choose the reviewer/s andto make stylistic changes and other minor corrections that do not alter the scientific content ofthe paper. The authors are obliged to make the first proof reading of the manuscript. Once thecorrected proofs are returned the authors bear the full responsibility for the content and thelanguage of the paper.
The following arrangement of the paper is required:
The title of the paper should be as short as possible, clear and consistent with the subject. No ab-breviations are allowed in the title save for the authors of taxonomic descriptions.
The surnames of authors are preceded by full first names. If there is more than one first name aninitial of the second name is given. Below are full names and addresses of institutions atwhich the work was done. If authors’ affiliations do not clearly follow from the order ofnames and institutions the names and the institutions should be marked by the same charactere.g a numeral.
A brief summary with keywords - not exceeding 2,000 characters. It should give a short ac-count of the paper’s content.
The introduction gives an account of the subject with a synthetic discussion of the relevant litera-ture. Citations of literature in the text are made by quoting the author’s name followed by yearof publication. If more than two co-authors are cited only the first author is quoted followedby “et al.”. An exaggerated number of literature references should be avoided. The objectiveof the study should be defined at the end of introduction.
Methods - briefly states the protocols used and describes the experiment materials clearly enoughfor the reader to be able to reproduce faithfully a similar study.
Results should be reported very concisely without detailed discussion of data contained in the ta-bles. An emphasis should be put on the significance of differences or the phenomena de-scribed. The same data must not be simultaneously contained in tables and figures (graphs).
The discussion is welcome only when it is necessary to confront one’s own data with those ofother authors. It is redundant in other papers.
The conclusions should generalize the results obtained. They should not be the summary of thepaper or the recommendations for commercial beekeeping.
The list of references comprises original publications directly relevant to the subject of paperquoted in the main text body. The references should be arranged according to names of au-thors, the same authors should be placed chronologically. They should be written in the fol-lowing style: Sakofski F., Koeniger N., Fuchs S. (1990) - Seasonality of honey bee colonyinvasion by Varroa jacobsoni Oud. Apidologie, 21(6):547-550.

118
The extended summary with paper title, initials and full surnames of authors and with keywords- not to exceed one and a half page of text. It gives a brief account of methods, results andconclusions. It is necessary to prepare this summary in Polish.
Keywords derived either from the title or from the main text ought to be informative of the sub-ject. They should be placed at the end of summaries. The keywords, not to exceed five, shouldbe in the first case and in singular if possible.
Tables presented in paper ought to be self-explanatory i.e. it should be comprehensible withoutmaking references to the main text. The headings must not contain symbols which are notgenerally known and the wording of headings must be concise. Tables and figures should beaccompanied with descriptions. Tables - should be designed in such a way as to stand reduc-tion to publication size (125 x 200 mm) without compromising legibility.
Figures (drawings, graphs) should be prepared on white paper and have a size of 125 mm x 200mm. Larger illustrations will be reduced to publication size of which the due allowanceshould be made (if scaled down to required size small print may become illegible). The fig-ures must be identified by placing a number on the back e.g. Fig. 1, Fig. 2 etc. Graphs canalso be submitted as MS Excel 2000 files or in the TIF format scanned at 300 dpi resolutionwith a hardcopy attached.
Photos should be submitted with the number in the reverse and transparencies (slides) shouldhave the number on the frame. If the original photo cannot be supplied a grayscale TIF filescanned at 300 dpi resolution is acceptable.
The manuscripts should be submitted in the MS Word 2000 format. Should other word processingpackage be used they ought to be exported to the RTF or TXT formats and accompanied withone hardcopy containing legible formatting details (bold, italics, special characters).Hand-made corrections on the printout are allowed; they should be highlighted in red.



![Journal of Membrane Science · of 1021m2 g-1and pore volume of 0.40cm3 g [38]), high resistance to heat, (430–540°C [37,39]), mechanical pressure and water adsorption [40,41]](https://img.pdfslide.fr/doc/110x75/5ecce269bfe251793f43740e/journal-of-membrane-science-of-1021m2-g-1and-pore-volume-of-040cm3-g-38-high.jpg)

























