Embed Size (px)
Citation preview

Le bivalve Montanaria Spriestersbach, 1909 : habitat et morphologie d'une nouvelle esphce emsienne de la Formation de York River de GaspC, Quebec1
SYLVAIN DESBIENS Commission gdologique du Canada, MI1, rue Booth, Ottawa, ON KIA OE8, Canada
Rep le 6 avril 1993 RCvision acceptte le 24 novembre 1993
I Une nouvelle espkce emsienne du bivalve rhtnan Montanaria Spriestersbach, 1909 est dCcrite h partir de spCcimens de la Formation de York River de la region de GaspC. C'est la premikre mention du genre en AmCrique. Cette espkce endoben- thique, M. honguedoensis nov. sp., s'agglomkrait dans les aires finement sablonneuses de baies ou lagunes saumstres rat- tachkes h un environnement de saimentation deltaique. Elle Ctait confrontCe h un benthos acide qui rongeait profondkment la region du crochet de la population adulte. La diffusion du Montanaria vers Gaspt s'explique par la dimension restreinte de 1'ocCan Iapttus emsien, et elle coincide avec une migration de bivalves, de 1'Europe de l'ouest vers le nord-est de 1'Amk- rique du Nord, au DCvonien prCcoce et moyen. La famille monogtntrique des Montanariidae est rCviste et son affiliation taxonomique est discutCe. Les caracteristiques dentaires et ligamentaires du Montanaria indiquent un niveau d'organisation intermkdiaire ?I ceux des actinodontoides primitifs et des hCttrodontes.
A new Emsian species of the Rhenish bivalve Montanaria Spriestersbach, 1909 is described from specimens that were col- lected in the York River Formation, Gaspt area. It is reported for the first time in America. This endobenthic species, M. honquedoensis n.sp., developed communities in areas of fine sand embayments or in brackish lagoons related to a sedi- mentation in a deltaic environment. The acidic benthos in which it lived corroded deeply the beak in the mature population. The diffusion of Montanaria towards Gasp6 is explained by the restricted dimension of the Emsian Iapetus Ocean, and it coincides with the bivalve migration, from the western Europe to the northeast of North America, from Early to Middle Devonian. The monogenic family of Montanariidae is revised and its taxonomic affiliation is discussed. The dental and ligamental features of Montanaria indicate an intermediate level of organization between primitive actinodontoids and heterodonts.
[Journal translation] Can. 1. Earth Sci. 31, 381 -392 (1994)
1 Introduction La partie infkrieure des Grbs de Gasp6 incluse dans la
Formation de York River (McGerrigle 1950) contient une faune diverse mais peu connue de bivalves du DCvonien infC- rieur (Praguien?-Ernsien). Cette faune n'a pas CtC rCCtudiCe depuis Billings (1874) et Clarke (1907, 1908), mais elle est illustree en partie dans Desbiens (1991). Une nouvelle espbce de Montanaria, M. honguedoensis, est dCcrite dans ce travail. Les donnCes fauniques, taphonomiques, ~Cdirnentologiques et stratigraphiques permettent de reconstituer son habitat de f a ~ o n
I prkcise. C'est la premibre mention du genre en AmCrique, et la signification palc5obiogCographique de sa prCsence en Gasptsie
1 est bribvernent discutte.
I Contexte stratigraphique et skdimentaire La Formation de York River silicoclastique recouvre en
concordance les sCdirnents de plate-forme carbonatCe des Calcaires supCrieurs de Gaspt (LespCrance 1980; Lavoie et al. 1990) et passe, vers le haut, aux sCdirnents a dominance flu- viatile de la Formation de Battery Point (Cant et Walker 1976; Rust 1981; Rust et al. 1989; Lawrence 1986; Lawrence et Williams 1987; Lawrence et Rust 1989). Elle est essentielle- ment formCe d'une alternance de niveaux de grbs de granulo- mCtrie fine a rnoyenne et de niveaux d'argilite et de microgrbs (McGerrigle 1950; Brisebois 198 1). Desbiens (199 1) associe la Formation de York River 2 la progradation, du sud-est vers le nord-ouest, d'un complexe deltaique reliC aux prernibre? phases importantes de 1'orogCnbse acadienne en GaspCsie. A I'extrCmitC est de la ptninsule (fig. l), sa puissance varie de 310 rn au cap du Petit-Gasp6 (Desbiens 1991) h un peu plus de
1. Contribution CGC 16293. Printed in Canada 1 Irnpnrnt au Canada
1600 m dans le secteur de la rivibre York (Sikander 1976). La carte rCcente de Brisebois (1981) donne la distribution rCgionale des unit& stratigraphiques .
Localisation des niveaux fossilif&res Le M. honguedoensis n'est abondant qu'au sommet de la
Formation de York River, B proximitk stratigraphique de la Formation de Battery Point. L'espbce est commune dans cer- taines couches qui affleurent le long de la route 197 et h l'anse 2 Brillant, respectivement au nord et au sud-est de la ville de Gasp6 (fig. 1). La position stratigraphique des niveaux fossili- fbres de la route 197 est donnCe a la fig. 3. Cette coupe est situte 12,3 km au sud-ouest de Rivibre-au-Renard et 4,4 km au nord du tron~on de la route 132 qui borde la baie de Gasp6 (fig. 1, A). Les sptcirnens de l'anse h Brillant ont CtC rCcoltCs au sommet d'un lit de grbs fin, 61,5 rn stratigraphiques sous le sommet des affleurements situCs au nord de l'embouchure de la rivibre de l'anse B Brillant (fig. 1, B). De rares spCcimens mal conservCs de Montanaria sp. ont Cgalement CtC collec- tionnCs le long de la rivibre Saint-Jean (fig. 1, C), 9,5 km B l'ouest de son intersection avec la route 132.
Biostratigraphie La faune h brachiopodes appalachiens qui caracttrise la
Zone B, Etymothyris trouve, dans la Formation de York River de la region de GaspC, son dCveloppernent le plus typique (Boucot et Johnson 1967; Desbiens 1991). Le genre Etymo- thyris apparait dans la Formation d'Indian Cove, au sommet des Calcaires suptrieurs de Gasp6 (LespCrance 1980), et il est prCsent jusqu'au sommet de la Formation de York River (Desbiens 1991; fig. 2). La Formation de Battery Point ne contient pas de brachiopodes permettant une datation prCcise.
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.

(I) (0 W W w a
FORMATIONS ZONES DE ZONES DE
.= 2 BRACHIOPODES SPORES w 2 vstatuP/angi l
w douglastowneIISe- - BATTERY eurypterota
0 z Q POINT a n n u l a t u s -
w u s e x t a n t i i 0 u ---- 7-- - - -- - - - - -7- - - - --
n A-' W
YORK ~l: ,u
-7- RIVER E t y m o t h y r i s
-- - - - -7. - - - - - O(O
\
W [ T ~ W
-?- 2 5 INDIAN p o l y g o n a l i s -
6; ~ ' W W COVE R e n s s e l a e r i a e m s i e n s i s ;:z:; -
382 CAN. J. EARTH SCI. VOL. 31, 1994
FIG. 2. Tableau de corrklation montrant la position stratigraphique relative des lits fossilifires discutCs dans le texte (voir fig. 1). La base de la Zone i Etymothyris est tirke de Lespkrance (1980). L'extention
FIG. 1. Localisation gkographique des lits fossilifires discutks dans des zones de spores est modifike de Richardson et McGregor (1986). le texte. A, lits de la route 197; B, lit de l'anse i Brillant; C, lits de Prag., Praguien; Sieg., SiegCnien; Eif., Eifklien. la rivikre Saint-Jean.
Montanaria lata Dahmer et Plethomytilus bilsteinensis Roemer La Zone B Etymothyris, reprCsentCe dans 1 '~ ta t de New York du Harz (Dahmer 1922, p. 260) est notoire. Un assemblage 1 par la Formation d7Esopus, est corrtlte avec le Praguien tardif benthique 1 B 2 dans la terminologie de Boucot (1975) est (Johnson 1979) ou la fin du SiegCnien et (ou) 1'Emsien (Boucot indiquk. et Johnson 1967; Boucot et al. 1967, 1969; Dutro 1981). La Les Montanaria de la rivibre Saint-Jean sont incomplets et rkpartition stratigraphique du genre Montanaria en Allemagne indtterminables B I'espbce. 11s ont kt6 collectionnCs avec quel- (grbs de Kahleberg et Remscheider Schichten; Haffer 1959) ques brachiopodes fragmentaires (Etymothyris gaspensis supporte un Age emsien pour la partie supCrieure de la Forma- (Clarke), u Spirifer a gaspensis Billings et Protoleptosrrophia tion de York River. blainvifIei (Billings)) dans des lits de conglomtrat (clastes
Les quelques Cchantillons de spores de la Formation de d'argilite) intercalts ?I des niveaux de g b s ?i stratifications York River CtudiCs par McGregor (1973, 1977) suggbrent obliques, ce qui denote un transporl important et un mtlange Cgalement une corrClation avec le SiegCnien tardif et (ou) faunique. I'Emsien, mais aucune es@ce distinctive de la Zone a Emphani- sporites annulatus - Carnarozonotriletes sextantii (Emsien prC- Habitat du Montanaria honguedoensis coce - debut tardif; assignation de Richardson et McGregor Les niveaux fossilifbres de la route 197 sont associCs B un 1986, p. 13) n'a CtC identifiCe dans la formation (D.C. horizon d,une puissance de 50 d'argilite, microgrbs et grbs McGregOr, communication ~ersonnelle, 1993). McGregor de granulomttrie fine trbs fine qui prennent une couleur brun (1977) place le contact Emsien-Eifdien dans la partie rougeitre distinctive en s9altkrant (fig. 3). Le sommet de cet su~~r i eu re de la or mat ion de Battery Point (fig. 2), B horizon coyncide avec la base de la Formation de Battery l'intkrieur de la Zone B Grandispora douglastownense - Point. Les lits B bivalves sont trouvCs au sommet de sCquences ~ n c ~ s ~ o r a eurypterota de ~ichardson et McGregor (1986). peu Cpaisses (1 - 10 m) oh la granul~mCtrie et 17Cpaisseur de
la stratification augmentent vers le haut. Certaines sCquences Faune des lits B Montanaria prtsentent une dCcroissance granulomktrique au sommet. Leur
Le M. honguedoensis et un Ambonychiidae CdentC et B sur- base est formCe d'argilite silteuse, et le passage au grbs supt- face unie assign6 provisoirement au Plethomytilus ,, sp. sont rieur est progressif. Une bioturbation intense oblitbre gCnC- les fossiles les plus communs des lits fossilifires de la route ralement les structures ddimentaires. De la pyrite dissCminCe 197 et de l'anse B Brillant. Les Palaeoneilo spp., Lingula sp. et d'abondants debris de vtgCtaux macCrCs et masses informes et Mutationella sp. sont Cgalement prCsentes, cette dernibre de matibres organiques (inertinite, vitrinite et bitumes solides; Ctant plus abondante dans les lithologies silteuses. Des restes Bertrand 1987) sont visibles au microscope dans le grbs de poissons (placodermes et acanthodiens), quelques ostra- (jusqu'8 15% des lames minces). Des strates similaires B celles codes et de rares fragments d'euryptkrides complbtent la de la route 197 sont visibles B l'anse Brillant. Notons l'exis- faune. Les bivalves sont souvent articulCs et en position de vie tence, B ce site, de quelques lits de grbs avec racines de plantes et ils forment localement des agglomCrations denses dans des et d'un lit de charbon de 20 cm. Des s6quences positives (N fining lits de grbs argileux de granulomCtrie fine B trbs fine. upward sequences D) atteignant 20 m d'kpaisseur, de grbs B
La composition faunique et l'absence de brachiopodes arti- stratifications obliques dkbutant par des surfaces de ravine- culCs autres que le Mutationella sp. suggbrent des eaux sau- ment conglomCratiques, sont prtsentes proximite stratigra- mdtres peu profondes. Les sCdiments c6tiers siegtnien et phique des niveaux fossilifbres. emsien de 1'Eifel contiennent des associations fauniques com- Les sCquences fossilifires de la route 197 se rapprochent des I parables (voir Fuchs 1982). En particulier, l'analogie des lits sCquences deltayques de baies interdistributaires E (q minor canadiens B Montanaria honguedoensis avec les bancs B sand spit ,,) et J (<< bay mouth sequence *) d'Eliott (1974, 1986)
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.

DESBIENS 383
CHENAUX
FLUVIATILES ?
M-
CHENAL
DISTRIBUTEUR
DELTAIQUE
PATRONS FAUNE LITHOLOGIQUES ----- ----- 8 BIVALVES ----- ARGITE ----- ---- G G A S T ~ R O P O D E S ----- ----- MICROGRES
L LINGULES
M Mutat lonel la nov. sp. G R ~ S
0 OSTRACODES
E EURYPTERIDES SURFACE D'EROSION F POISSONS CONGLOMERATIOUE
() PRESENCE STRATIF ICATION OBLIQUE
EN FOSSES GRANULOMCTRIE --s= STRATIF ICATION OBLIQUE IM-S'T'F'M'G'C?
PEU INCLINEE m t 4
w u
PETITES RIDES LL Z;j.W
4 e urn" 8 ,%z&gg
) , BIOTURBATION INTENSE c1g +u Iaz 2s Z E - 2
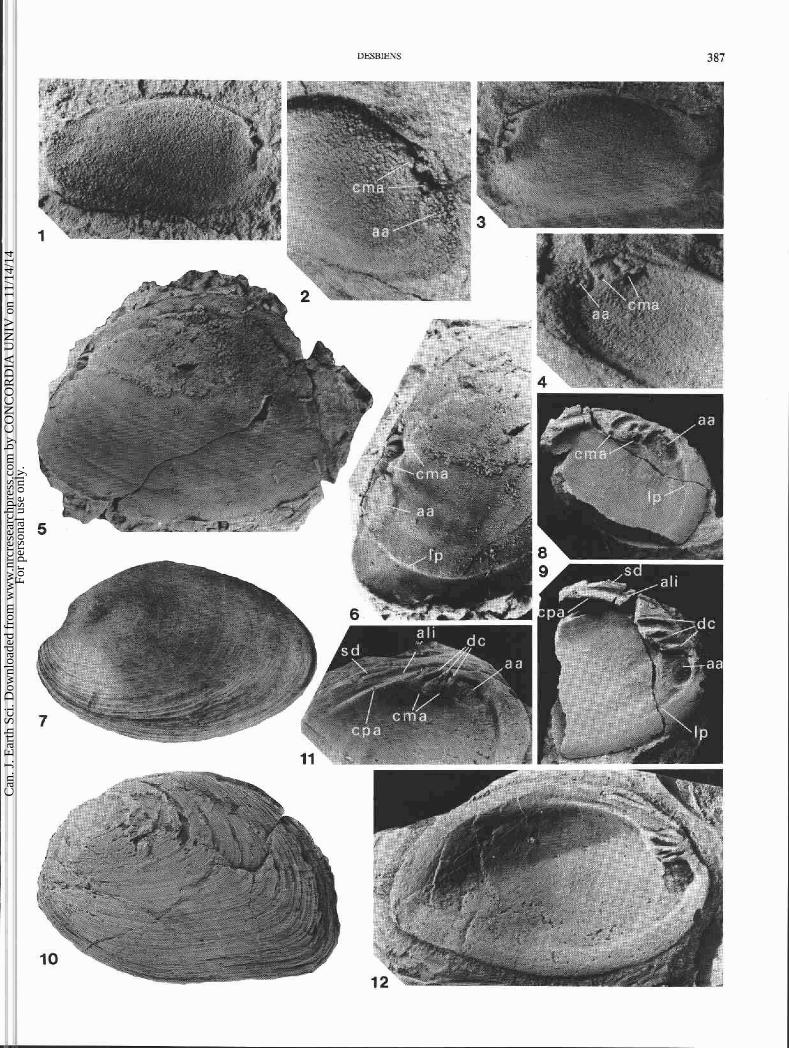
FIG. 3. Environnement de skdimentation, faune et position stratigraphique des lits fossiliferes de la coupe de la route 197. Les Cpaisseurs stratigraphiques ont CtC mesurCes B partir de la base de la Formation de York River. La faune est donnee dans un ordre dkcroissant d'abondance, except6 pour les groupes entre parenthkses. La presence seule est alors signal6e. (A) deux individus du Montanaria honguedoensis nov. sp. en position de vie dans la partie infkrieure d'un lit de grks fin. Une pavCe de valves i convexit6 vers le haut de la mkme espkce est prCsente au sornrnet du lit. (B) coupe polie d'un lit montrant deux niveaux avec d'abondantes M. honguedoensis articulks. Noter les structures gCop6tales B 11int6rieur de certains specimens (flkches). Les Cchantillons de lits fossiliferes (A) et (B) sont conservCs dans la collection de sp6cimens types de la Commission g6ologique du Canada i Ottawa (respectivement GSC-107466 et GSC-107467).
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.

384 CAN, 1. EARTH SCI. VOL. 31, 1994
et de certaines stquences silicoclastiques d'environnement marin littoral de De Raff et al. (1977). Similairement, Lawrence (1986), Lawrence et Rust (1989) et Rust et al. (1989) attri- buent B la Formation de York River de l'anse B Brillant un environnement stdimentaire de baies et (ou) lagunes situtes en marge de petits chenaux distributeurs deltaiques. La faune et le contexte stratigraphique et stdimentaire gtntral renforcent ces interprttations .
L'ensemble des donntes indiquent que le M. honguedoensis habitait des baies (ou lagunes) saumdtres rattachees B un envi- ronnement de stdimentation deltaique. I1 s'agglomerait (fig. 3B) dans les aires finement sablonneuses de faible bathymttrie, quelques centimbtres sous la surface du benthos (fig. 3A).
Les phCnomknes de dissolution Des marques de corrosion ptri-umbonale prononctes affec-
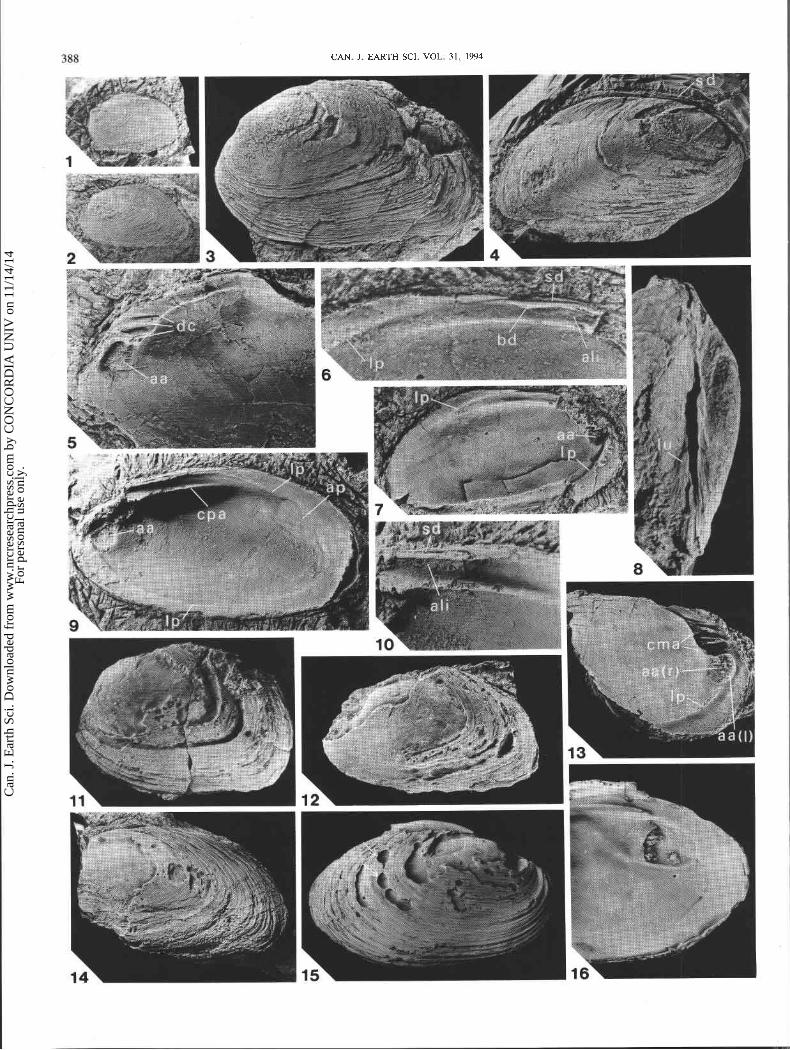
tent plus de 90% des valves du M. honguedoensis dont la longueur dtpasse 5 cm (planche 1, fig. 10; planche 2, fig. 3, 4, 11, 12 et 14). Les petits sptcimens en sont exempts. Cette corrosion affecte aussi bien les individus articults qu'in- articules, et les surfaces internes sont intactes. Elle est donc contemporaine de la vie des individus.
La dissolution est le principal agent destructeur des tests bio- carbonat& sous l'interface eau marine - stdiment (Driscoll 1970, fig. 4; Aller 1982). Cette dissolution, favoriste par la bioturbation, est induite par des concentrations tlevtes de C02 provenant de matibres organiques en dtcomposition (Aller 1982; McCall et Tevesz 1983; Brett et Baird 1986). La forte stratification haline des eaux saumkres c6tibres B circula- tion restreinte favorise un enrichissement benthique en C02 (Remane 197 1).
Des traces de dissolution ptri-umbonale sont communes chez les Unionacea (Tevesz et Carter 1980; Johnston et Good- body 1989; cette Ctude, planche 2, fig. 15) et les corbulidts et corbiculidts dul~aquicoles et d'eau saumltre actuels (Lewy et Samtleben 1979; Kat 1982). Ce phtnombne est gtntrale- ment expliqut par l'effet corrosif de l'eau douce acide sur les parties anciennes de la coquille qui ne sont plus prottgtes par le ptriostracum (Dechaseaux 1952, p. 267; Tevesz et Carter 1980). Les archives paltontologiques contiennent des rtft- rences au mCme phtnombne (Dechaseaux 1952; Noe-Nygaard et al. 1987; Johnston et Goodbody 1989). Suite B l'usure du ptriostracum, une dissolution excessive menant B la perfora- tion de la coquille peut Ctre ltthale (Lewy et Samtleben 1979; Kat 1982; Hunter 1990). Kat (1982) a document6 un taux Clevt de mortalit6 infectieuse dans une population dul~aqui- cole de Corbiculafluminea (Miiller) sujette B la corrosion. La prtsence de couches de conchiolines dans la coquille des Unionacea et certaines espbces de Corbiculidae, Corbulidae et Solenidae permet de rtsister B la dissolution (Lewy et Samtleben 1979; Tevesz et Carter 1980; Kat 1982, 1983, 1985). Les bivalves d'eau douce ont, en outre, la capacitt de rtgtntrer leur coquille perforte (Wilbur et Watabe 1963; Wilbur 1964; Beedham 1965). Des surfaces de rtgtnerations irrtgulibres atteignant 1,5 cm de diambtre et des calus internes sont frt- quents dans la rtgion des empreintes adductrices des popula- tions d'Elliptio complanata (Lightfoot) et de Lampsilis radiata siliquoidea (Barnes) de la rivibre Outaouais prbs d'Ottawa (S. Desbiens, observations personnelles, planche 2, fig. 16). L'eau de mer gCne par contre la cicatrisation (Wagge et Mittler 1953) en entravant la formation de la membrane orga- nique qui est B l'origine de la rtgtntration (Wilbur 1964).
Les contraintes chimiques auxquelles ttait confront6 le M. hhoguedoensis devaient &tre nefastes pour une partie de la population 6 g k (voir la degradation avancie des sgcirnens de la planche 2, fig. 11, 12 et 14). Par contre, l'epaisseur exceptionelle de sa coquille (planche 1, fig. 8) lui confirait certainement un avantagc adaptif con* la corrosion. L'acidit6 du biotope occupt par le M. honguedoensis rtsultait de l'abon- dance des matibres organiques (phytoclastes) en dtcompo- sition dans un environnement de stdimentation saumltre B circulation restreinte ou semi-restreinte.
PalCobiogCographie Boucot (1963, 1975, 1982; Boucot et al. 1967) a signal6 la
prtsence du brachiopode rhtnan Rhenorensselaeria Kegel dans la faune appalachienne de la Formation de York River et dtduit qu'i l'Emsien, la limite biogtographique des royaumes de l'ancien monde rhtno-hercynien et des Amtriques de l'est ttait situte a proximitt de Gaspt. Les tchanges fauniques ont tgalement affectts les bivalves (Desbiens 1991). A l'exttrieur de Gaspt, le Montanaria n'est connu avec certitude que de strates emsiennes des Berghishes Land et du Harz. Mention- nons Cgalement les prtsences, dans la Formation de York River, de Carydium sp., Paracyclas rugosa (Goldfuss), Proso- coelus sp., Schysodus? aff. circularis (Beushausen) et Gonio- phora (Goniophora) aff. exilis (Dreverman). Ces incursions rhtnanes en Gasptsie renforcent l'image d'un octan Iapttus emsien de dimension restreinte (Scotese et McKerrow 1990, fig. 14). Au Dtvonien moyen, ttendant leur dispersion B l'est des tats Unis, les bivalves de l'ancien monde europten sont devenus des Cltments importants des faunes de 1'Hamilton new-yorkais (Bailey 1978, 1983).
DkpBt des spkcimens Les sptcimens types et illustrts sont conservts B la Commis-
sion gtologique du Canada B Ottawa (prtfixe GSC) et au Geologisch - Paleontologisch Institut de Gottingen (prkfixe GPIG) .
PHYLUM Mollusca Cuvier CLASSE Bivalvia (Bonnani) Linnt (=Pelecypoda Goldfuss)
Remarque La classification suprafamiliale de Pojeta (1971, 1987) est
utilide.
SOUS-CLASSE Heteroconchia Hertwig ORDRE Actinodontoida Douvillt
FAMILLE Montanariidae Scarlato et Starobogatov
Genre type Montanaria Spriestersbach, 1909 (Spriestersbach et Fuchs
1909).
Diagnose hende'e Coquilles tquivalves, intquilattrales, subisomyaires et inttgri-
pallites. Lunule peu marqute ou absente. Absence d'escut- chton. Plateau d'articulation robuste supportant un ligament opisthodbte enchlsd profondtment entre les marges des valves. Pas de nymphe ligamentaire. Dentition radiale sous-umbonale sans prolongements pseudo-lattraux. Dents souvent divistes par des sillons longitudinaux. Lattrales absentes.
Montanaria Spriestersbach, 1909
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.

DESBIENS 385
Esptce type I Pleurophorus devonicus Beushausen, 1884 par designation
subskquente de Haffer 1959, p. 166.
Strates types Schalker Schichten (Grbs de Kahleberg; Emsien) de la
rCgion de Bocksberg dans le Harz en Allemagne.
Diagnose kmendke ~ ~ u i v a l v e , fortement inkquilattral, moyennement convexe et
de forme ovale plus ou moins allongCe. Crocket anttrieur non saillant. Lunule petite et peu marquCe, quelquefois indistincte. Prosopon de fines stries concentriques d'accroissement. Plateau d'articulation robuste. Ligament opisthodbte enchbse pro- fondtment entre les marges des valves : rtsilium interne situt dans un creux allongt parallble au bord dorsal, et couches liga- mentaires suptrieures dans un sillon dorso-marginal. Denti- tion cardinale variable : typiquement une B trois dents droites et deux ou trois dents gauches souvent bifides ou munies de sillons. Laterales absentes. Empreinte adductrice anttrieure ovale, bien imprimte; empreinte postkrieure un peu plus grosse et superficielle, habituellement non visible. Deux cicatrices musculaires accessoires du pied au-dessus de l'empreinte adductrice anttrieure. Ligne palltale simple.
Distribution stratigraphique et gi5ographique En AUemagne, le Montanaria est prCsent dans les Remscheider
Schichten des Berghishes Land (Massif schisteux rhCnan; Emsien) et dans les Schalker et nessigi Schichten (Grb de Kahleberg; Emsien) du Harz (Spriestersbach et Fuchs 1909; Spriestersbach 1919, 1925, 1942; Vietor 1919; Dahmer 1922, 1942, 1946; Haffer 1959; Stoltidis 1971). Le genre est tgale- ment prtsent dans la Formation de York River de la rtgion de Gasp6 au QuCbec.
Esptces incluses Montanaria devonica (Beushausen, 1884; = M. ovata
Spriestersbach, 1909), M. elongata Spriestersbach, 1909, M. curta Spriestersbach, 1909, M. anulifera Spriestersbach, 1909, M. lata Dahmer, 1922 et M. honguedoensis nov. sp.
Esptces possibles Montanaria elliptica Khalfin, 1948 et Montanaria sp.
Quenstedt , 1926.
Discussion Comme le note Chavan (1967), les figures d'Haffer (1959)
et le pibtre ttat de conservation du lectotype du M. devonica (supra citato, planche 12, fig. 5 et 6, rCillustrt ici, planche 1, fig. 1-4) donnent une image incomplbte des particularitts de la charnibe du Montanaria. Haffer (1959) signale une ou deux dents cardinales droites et deux ou trois dents cardinales gauches chez l'espbce type. De plus, trois cardinales droites sont attributes au M. lata par Dahrner (1922, p. 259, planche 17, fig. 4). Une troisibme cardinale est Cgalement prCsente sur certaines valves droites du M. honguedoensis. Chavan (1967, 1969) prCte au genre une latCrale anttrieure droite (AIII, dans la notation de F. Bernard), mais Khalfin (1974), Haffer (1959) et Spriestersbach (1919) insistent sur l'absence de dents latt- rales chez le Montanaria. Les descriptions et illustrations de Beushausen (1884, p. 105, planche 3, fig. 8), Spriestersbach (Spriestersbach et Fuchs 1909, pp. 42-47, planche 5, fig. 12- 14, planche 6, fig. 1 - 12, et planche 7, fig. 1 -4) et Dahmer (1922, pp. 258-260, planche 14, fig. 2, et planche 17, fig. 1 - 5) ne permettent pas de corroborer la prtsence d'une telle
lattrale (AIII). L'absence de lattrales est ici confirmte chez le M. honguedoensis. En outre, des incisions ou sillons dentaires longitudinaux plus ou moins profonds (Haffer 1959, p. 65, Spriestersbach et Fuchs 1909, planche 5, fig. 14, et planche 6, fig. 3a-7, cette ttude, planche 1, fig. 8, 9, 11 et 12 et planche 2, fig. 13) amplifient la variabilitt de l'appareil cardi- nal du Montanaria. Les dents sont souvents bifides (planche 1, fig. 8, 9, 11 et 12) et, dans quelques cas, de petites pibces dentaires isoltes sont observtes. Des valves du M. ovata (=M. devonica; Haffer 1959) prtsentant une division dentaire anormalement tlevte ont t t t illustrtes par Spriestersbach et Fuchs (1909, planche 6, fig. 5 et 7). Selon Haffer (1959), ces cas extremes sont exceptionnels.
Spriestersbach (Spriestersbach et Fuchs 1909, p. 43, planche 5, fig. 14) a correctement interprttt l'aire concave postCro- dorsale du plateau d'articulation comme le site d'insertion du ligament. L'extension du ligament du Montanaria (Haffer 1959, p. 164, fig. 14 et planche 14, fig. 4 (partie)) a par contre t t t surestimte. Un sptcimen du M. honguedoensis (planche 2, fig. 10) indique une arta ligamentaire interne (ali) ne s'tten- dant que sur la moitit anttrieure de l'aire dorso-concave. Cette arta, qui prtsente quelquefois de fines stries obliques, correspond B l'emplacement des couches ligamentaires fibreuses. Un mince sillon dorsal (sd, fig. 4A, planche 1, fig. 9 et 11, et planche 2, fig. 6 et 10) surmontant un petit bourrelet (bd) servaient de support aux couches suptrieures, probablement lamellaires, du ligament. Un sillon similaire est visible au bord dorsal de la valve droite du M. ovata (= M. devonica) illustrte par Spriestersbach et Fuchs (1909, planche 6, fig. 5a) et Spriestersbach (1919, planche 3, fig. 3). Le petit bourrelet dorsal du M. honguedoensis reprtsente peut-Ctre l'amorce d'une nymphe ligamentaire, mais cette structure est invisible de l'exttrieur (fig. 4b). Elle n'est donc pas considtrte ici comme une nymphe au sens stricte.
La nature sub-isomyaire des adducteurs est discernable sur un moule interne gauche du M. lata de Dahmer (1922, planche 17, fig. 1) et sur une valve droite du M. honguedoensis (planche 2, fig. 9). La position des muscles accessoires du pied est similaire chez les espbces M. devonica (planche 1, fig. 2 et 4), M. lata (planche 1, fig. 6) et M. honguedoensis (planche 1, fig. 8 , 9 et 1 1, planche 2, fig. 13) : deux cicatrices musculaires accessoires (cma) surmontent l'empreinte adduc- trice antkrieure (aa).
Outre les espbces allemandes et canadienne, deux espbces ont Ctt assigntes au Montanaria. Le M. elliptica du Dtvonien inftrieur de 1'Altai (Khalfin 1948 (p. 375, planche 36, fig. 51), 1974 (p. 94, fig. 41) est trop peu connu pour un diagnostique gCntrique dtcisif. Les illustrations de Quenstedt (1926, planche 2, fig. 28 et 29) du Montanaria sp. de la Formation de Grey Hoek (Emsien - Eiftlien; Friend 1961 ; Orvig 1969) du Spitsbergen sont peu significatives, mais le schtma de la charnibre (Quenstedt 1926, p. 65, fig. 4) correspond assez bien B celle du Montanaria (fig. 4A).
Afiliation taxonomique L'affiliation taxonomique du Montanaria est controversa-
ble, et plusieurs sous-classes ont CtC utilistes pour le carac- ttriser. Dans la classification de Newel1 (1965), celui-ci est habituellement assign6 aux Heterodonta, mais des relations phylogCniques avec les Palaeoheterodonta ont tgalement Ctt suggCrCes. Dans celle de Pojeta (1971, 1987), la litteratwe assigne le Montanaria B des familles d'Heteroconchia ou d'Isofilibranchia.
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.
CAN. J. EARTH SCI. VOL. 3 1, 1994
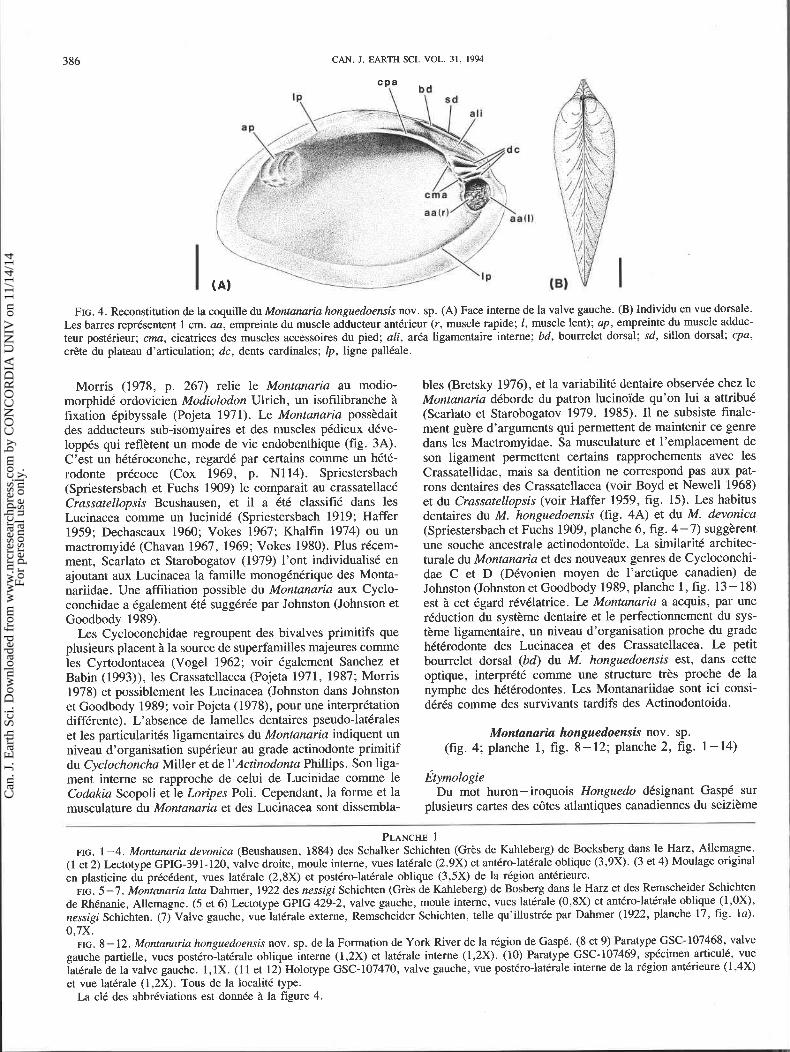
FIG. 4. Reconstitution de la coquille du Montanaria honguedoensis nov. sp. (A) Face interne de la valve gauche. (B) Individu en vue dorsale. Les barres reprksentent 1 cm. aa, empreinte du muscle adducteur antkrieur (r, muscle rapide; 1, muscle lent); up, empreinte du muscle adduc- teur postCrieur; cma, cicatrices des muscles accessoires du pied; ali, arka ligamentaire interne; bd, bourrelet dorsal; sd, sillon dorsal; cpa, cr2te du plateau d'articulation; dc, dents cardinales; Ip, ligne palltale.
Morris (1978, p. 267) relie le Montanaria au modio- bles (Bretsky 1976), et la variabilitt dentaire observte chez le morphidt ordovicien Modiolodon Ulrich, un isofilibranche i Montanaria dtborde du patron lucinoide qu'on lui a attribut fixation tpibyssale (Pojeta 1971). Le Montanaria possbdait (Scarlato et Starobogatov 1979, 1985). I1 ne subsiste finale- des adducteurs sub-isomyaires et des muscles pCdieux dive- ment gubre d'arguments qui permettent de maintenir ce genre loppts qui reflbtent un mode de vie endobenthique (fig. 3A). dans les Mactromyidae. Sa musculature et l'emplacement de C'est un htttroconche, regard6 par certains comme un httt- son ligament permettent certains rapprochements avec les rodonte prtcoce (Cox 1969, p. N114). Spriestersbach ~rassatellidae, mais sa dentition ne correspond pas aux pat- (Spriestersbach et Fuchs 1909) le comparait au crassatellact rons dentaires des Crassatellacea (voir Boyd et Newel1 1968) Crassatellopsis Beushausen, et il a t t t classifit dans les et du Crassatellopsis (voir Haffer 1959, fig. 15). Les habitus Lucinacea comme un lucinidt (Spriestersbach 1919; Haffer dentaires du M. honguedoensis (fig. 4A) et du M. devonica 1959; Dechaseaux 1960; Vokes 1967; Khalfin 1974) ou un (Spriestersbach et Fuchs 1909, planche 6, fig. 4 -7) suggkrent mactromyidt (Chavan 1967, 1969; Vokes 1980). Plus rtcem- une souche ancestrale actinodontoide. La similaritt architec- ment, scarlato et Starobogatov (1979) l'ont individualist en ajoutant aux Lucinacea la famille monogCntrique des Monta- nariidae. Une affiliation possible du Montanaria aux Cyclo- conchidae a tgalement t t t suggtrte par Johnston (Johnston et Goodbody 1989).
Les Cycloconchidae regroupent des bivalves primitifs que plusieurs placent i la source de superfamilles majeures comme les Cyrtodontacea (Vogel 1962; voir tgalement Sanchez et Babin (1993)), les Crassatellacea (Pojeta 1971, 1987; Morris 1978) et possiblement les Lucinacea (Johnston dans Johnston et Goodbody 1989; voir Pojeta (1978), pour une interprttation difftrente). L'absence de lamelles dentaires pseudo-lattrales et les particularitts ligamentaires du Montanaria indiquent un niveau d'organisation suptrieur au grade actinodonte primitif du Cyclochoncha Miller et de l'dctinodonta Phillips. Son liga- ment interne se rapproche de celui de Lucinidae comme le Codakia Scopoli et le Loripes Poli. Cependant, la forme et la musculature du Montanaria et des Lucinacea sont dissembla-
turale du Montanaria et des nouveaux genres de Cycloconchi- dae C et D (Dtvonien moyen de l'arctique canadien) de Johnston (Johnston et Goodbody 1989, planche 1, fig. 13 - 18) est ?i cet tgard rtvtlatrice. Le Montanaria a acquis, par une riduction du systbme dentaire et le perfectionnement du sys- tbme ligamentaire, un niveau d'organisation proche du grade htttrodonte des Lucinacea et des Crassatellacea. Le petit bourrelet dorsal (bd) du M. honguedoensis est, dans cette optique, interprttt comme une structure trbs proche de la nymphe des htttrodontes. Les Montanariidae sont ici consi- dQQ comrne des survivants tardifs des Actinodontoida.
Montanaria honguedoensis nov. sp. (fig. 4; planche 1, fig. 8-12; planche 2, fig. 1-14)
~ t ~ r n o l o ~ i e Du mot huron-iroquois Honguedo dtsignant GaspC sur
plusieurs cartes des c6tes atlantiques canadiennes du seizibme
PLANCHE 1 FIG. 1-4. Montanaria devonica (Beushausen, 1884) des Schalker Schichten (Grks de Kahleberg) de Bocksberg dans le Harz, Allemagne.
(1 et 2) Lectotype GPIG-391-120, valve droite, moule interne, vues latCrale (2.9X) et antkro-latkrale oblique (3,9X). (3 et 4) Moulage original en plasticine du prkcCdent, wes latkrale (2,8X) et posttro-latkrale oblique (3,5X) de la rkgion antkrieure.
FIG. 5 -7. Montanaria lata Dahmer, 1922 des nessigi Schichten (Grks de Kahleberg) de Bosberg dans le Harz et des Remscheider Schichten de RhCnanie, Allemagne. (5 et 6) Lectotype GPIG 429-2, valve gauche, moule interne, vues latCrale (0,8X) et anttro-laterale oblique (1 ,OX), nessigi Schichten. (7) Valve gauche, vue latCrale externe, Remscheider Schichten, telle qu'illustrCe par Dahmer (1922, planche 17, fig. la). 0,7X.
FIG. 8 - 12. Montanaria honguedoensis nov. sp. de la Formation de York River de la rCgion de GaspC. (8 et 9) Paratype GSC-107468, valve gauche partielle, vues postkro-latkrale oblique interne (1,2X) et latCrale interne (1,2X). (10) Paratype GSC-107469, sptcimen articulC, vue latkrale de la valve gauche. 1,lX. (1 1 et 12) Holotype GSC-107470, valve gauche, vue postCro-latkrale interne de la region antkrieure (1,4X) et vue latkrale (1,2X). Tous de la localitk type.
La clC des abbrkviations est donnke i la figure 4.
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.
388 CAN. 1. EARTH SCI. VOL. 31, 1994
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.

DESBIENS 389
sibcle (Ganong 1934; Ganong et Layng 1964). Refire a la Plateau d'articulation posttro-cardinal robuste, longitudi- localisation des strates types de l'espbce. nalement concave, muni d'une cr6te (ou tpaississement) inft-
Diagnose Espbce ovale B subrhomboidale, au crocket anttrieur sub-
terminal et B coquille Cpaisse. Inflexion marqute des stries d'accroissement le long de la pente postumbonale. Cr6te rtsul- tant d'un Cpaississement B la base du plateau d'articulation posttrocardinal. Petit bourrelet, au-dessus de l'arta ligamen- taire interne, supportant un sillon ligamentaire dorso-marginal invisible de I'extCrieur. Plateau dentaire long, portant deux ou trois dents cardinales, parfois bifides, par valve. Empreinte adductrice anttrieure rugueuse et bien imprimte. Cicatrice adductrice posttrieure unie et superficielle, tronqute antt- rieurement . Strates types
Formation de York River de la rtgion de Gaspt. Les strates du sud de la baie de Gaspt situtes immtdiatement au nord de l'embouchure de la rivibre de l'anse B Brillant (fig. 1, B; 320020m.E., 5398260m.N.) constituent la localitt type; celles de la route 197 (fig. 1, A et fig. 3; 304080rn.E., 5419500m.N.), une localit6 de rtftrence compltmentaire.
Mat&riel Holotype GSC-107470, paratypes GSC : 107468, 107469,
107473, 107475, 107477, 107480 (localitt type) et 107474 ainsi que 107476 (route 197); sptcimens GSC illustrts : 107471, 107472, 107478 et 107479 (route 197); tchantillons de lits fossilifires GSC-107466 et GSC-107467 (route 197); 59 sptcimens articults, 60 valves droites et 5 1 valves gauches, le quart provenant de l'anse a Brillant et le reste de la route 197. - - Description
Coquille tquivalve t r b tpaisse Cjusqu'B 5 mm), fortement intquilattrale et de convexitt moyenne. Longueur atteignant 10,5 cm. Contour ovoide B subrhomboi'dal. Ligne postCro- cardinale doucement arquCe et bord posttrieur bien arrondi, s'infltchissant rapidement vers l'avant B l'angle postkro- ventral. Partie arribre du bord inftrieur subrectiligne et con- tour ventral de la rtgion anttrieure uniformtment recourbt. Crochet prosogyre anttrieur non saillant. Bord faiblement concave sur une courte distance sous le crochet (planche 1, fig. 12). Lobe anttrieur court, formant 5 B 10% de la longueur de la coquille. Prosopon de fines stries d'accroissement con- centriques s'incurvant rapidement B angle droit le long de la pente postumbonale. Petite lunule circulaire peu marquee (planche 2, fig. 8, lu), souvent indistincte. Absence d'es- cutchton.
. rieure plus ou moins protminente qui atteint le centre longitudinal de la coquille (cpa, fig. 4A; planche 1, fig. 9, 11 et 12; planche 2, fig. 9). Ligament opisthodbte profondtment enchbd entre les marges suptrieures des valves. Empreinte interne du ligament (ali, planche 2, fig. 10) circonscrite B la moitit anttrieure doucement surtltvte de l'aire dorso-concave du plateau d'articulation. Mince bourrelet dorsal (bd) suppor- tant un sillon ligamentaire marginal (sd), au-dessus de 1'arCa ligamentaire interne. Ce sillon, invisible de l'exttrieur, se pro- longe sur une courte distance en arribre du bourrelet dorsal, vers l'extention suptrieure de la ligne palltale (planche 2, fig. 6 et 7). Bourrelet des petits sptcimens plus protminent. Fines stries obliques parfois visibles sur l'arta ligamentaire interne et sur le cBtC du bourrelet dorsal (planche 1, fig. 9). Faible baillement du bord suptrieur des valves des gros indivi- dus, correspondant B l'emplacement du ligament (fig. 4B). Plateau dentaire dtveloppt, tpais, surmontant une faible cavitt umbonale. Valve gauche caracttriste par trois dents cardinales allongtes divergeant vers l'arribre. Deux ou trois cardinales droites. Dent occupant une position intermidiaire habituellement plus dtveloppte. Dent mtdiane gauche typi- quement bifide (planche 1, fig. 8, 9, 11 et 12) ou ornte de sil- lons. Dents inftrieure et intermtdiaire droite plus rarement bifides. Dent cardinale posttrieure droite peu B non dtvelop- pte; fossette gauche correspondante superficielle ou absente. Lattrales manquantes. La formule suivante, baste sur la nota- tion de Boyd et Newel1 (1968), rtsume les caracttristiques dentaires de l'espbce :
posttrieur VD (1) (0) I(,) 0 I(,) (0) anttrieur
oh 1 indique une dent, 0 une fossette dentaire, , des sillons ou incisions dentaires longitudinaux, () l'absence possible, VD la valve droite et VG la valve gauche.
Empreinte du muscle adducteur antCrieur ovale, bien imprimte, avec axe long subvertical : trace de l'adducteur rapide fortement rugueuse, CtalCe sur la majeure partie de l'empreinte (planche 2, fig. 13, aa(r)); portion de la cicatrice correspondant B l'adducteur lent restreinte B une mince bande anttrieure fusiforme et unie (planche 2, fig. 13, aa(1)). Empreinte de l'adducteur posttrieur un peu plus grosse, unie et superficielle, de contour ovalaire tronqut B l'avant, avec axe long inclinC vers l'arribre (planche 2, fig. 9, up). Forte cicatrice musculaire accessoire du pied B la base du plateau
-
PLANCHE 2 FIG. 1 - 14. Montanaria honguedoensis nov. sp. de la Formation de York River de la region de Gasp& (1) GSC-107471, valve gauche, w e
laterale externe. 2,2X. (2) GSC-107472, valve gauche, vue laterale externe. 1,4X. (3) Paratype GSC-107473, spkcimen articulk, vue laterale de la valve gauche. 0,9X. (4) Paratype GSC-107474, specimen articult, vue laterale de la valve droite. 1,3X. (5) Paratype GSC-107475, valve droite partielle, vue latkrale interne. 1,4X; (6 et 7) Paratype GSC-107476, valve gauche partielle, agrandissement de la partie superieure interne (4,4X) et vue laterale interne (1,7X). (8) Paratype GSC-107473, specimen articult, vue dorso-anterieure oblique. 1,2X. (9 et 10) Paratype GSC-107477, valve droite partielle, vue laterale interne (1,3X) et agrandissement de la region dorso-antkrieure (2,7X). (11) GSC-107478, specimen articule, vue laterale externe de la valve gauche. 0,9X. (12) GSC-107479, valve gauche partielle, vue laterale externe. 1,OX. (13) Paratype GSC-107480, moule interne partiel d'une valve droite, vue laterale. 1,3X. (14) GSC-107481, valve gauche partielle, vue latkrale externe. 0,8X. Les specimens des figures 1, 2, 4, 6 et 7 proviennent du lit (B) de la coupe de la route 197 (fig. 3); ceux des figures 3, 5 et 8- 13 de la localit6 type.
FIG. 15 et 16. Lampsilis radiata siliquoidea (Barnes, 1823), recent, baie Britannia, rivikre Outaouais prks d'Ottawa en Ontario. GSC-107482, specimen teratologique, valve droite, vues laterale externe (0,8X) et laterale interne (1,OX) de la region posterieure. Noter la presence de calus internes au pourtour de la cicatrice adductrice postkrieure (fig. 16) et les perforations externes correspondantes (flkches, fig. 15).
La clt des abbrtviations est donnee i la figure 4.
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.

390 CAN. J. EARTH SCI. VOL. 31, 1994
dentaire (cicatrice d'un muscle protracteur), au-dessus de l'empreinte de l'adducteur anterieur; une seconde plus petite (cicatrice d'un muscle rktracteur?), juste sous la dent sup6 rieure gauche (planche 1, fig. 8 et 11). Ligne pallkale intCgri- pallike se prolongeant dorsalement au-dessus de l'empreinte de l'adducteur postCrieur (planche 2, fig. 6, 7 et 9), en direc- tion du sillon ligamentaire dorsal (sd). Trbs faible sinuositk du tracC pallCal juste sous l'adducteur postkrieur. Aire interne ventrale des valves faiblement dCprimCe le long de la ligne palltale.
Cornparaison avec les autres esp&ces du genre Montanaria Le M. honguedoensis se rapproche du M. devonica et de son
synonyme junior M. ovata. Faute d'illustrations rCcentes, les comparaisons avec l'espbce type se limitent ti quelques dCtails qui justifient la distinction spCcifique du materiel canadien. Le plateau dentaire du M. honguedoensis est plus allongC et sa limite infkrieure mieux dCfinie que chez le M. devonica. Le plateau postkro-cardinal de cette dernibre est un peu moins robuste et sa base moins prokminente. Les figures de Spriesters- bach et Fuchs (1909) rtvblent un contour variable comparable 2 celui du M. honguedoensis. Le lectotype du M. lata (planche 1, figs. 5 et 6) possbde des dents non bifides comparativement plus courtes, faiblement recourbkes, un plateau dentaire plus haut et un plateau postCro-cardinal peu proCminent. L'in- flexion postumbonale des stries d'accroissement de cette espke est rCgulibre (planche 1, fig. 7) et son bord infkrieur uniform6 ment courbC. Le contour du M. elongata et du M. curta et le prosopon du M. anulifera sont distinctifs.
Remerciements D. Brisebois a dCcouvert les niveaux fossilifires de la
route 197. Les spCcimens ont CtC collectionnCs au cours des 6 th 1987 et 1988, dans le cadre d'un projet de doctorat dirigt par P.J. LespCrance. Ce dernier a fourni l'appui financier aux travaux de 1987 par l'entrernise d'une subvention de dCpenses courantes du Conseil de recherches en sciences naturelles~t en gCnie du Canada (CRSNG A-17897). Le ministbre de 1'Ener- gie et des Ressources du Qukbec a fourni l'appui logistique aux travaux de 1988. H. Jahnke a pr6tC des spCcimens types du Geologisch-Paleontologisch Institut de Gottingen. Cette contribution a CtC rCalisCe lors d'un stage postdoctoral ti la Commission gCologique du Canada ti Ottawa. Le support du CRSNG sous forme de bourses a CtC grandement apprCciC. Que toutes les personnes et organismes ci-haut mentionnCs re~oivent l'expression de mes sincbres remerciements.
Aller, R.C. 1982. Carbonate dissolution in nearshore terrigenous muds: the role of physical and biological reworking. Journal of Geology, 90 : 79-95.
Bailey, J.B. 1978. Provincialism and migration in Lower and Middle Devonian pelecypods. Palaeogeography, Palaeoclimatology, Pala- eoecology, 23 : 119- 130.
Bailey, J.B. 1983. Middle Devonian Bivalvia from the Solsville Member (Marcellus Formation), central New York State. Bulletin of the American Museum of Natural History, Vol174 : 193 -326.
Beedham, G.E. 1965. Repair of the shell in species of Anodontu. Proceedings of the Zoological Society of London, 145 : 107 - 124.
Bertrand, R. 1987. Maturation thermique et potentiel pCtroligkne $es sCries post-taconiennes du nord-est de la GaspCsie et de 1'Ile d'Anticosti (Canada). These de doctorat, UniversitC de Neuchltel, Neuchltel.
Beushausen, L. 1884. Beitrage zur Kenntniss des Oberharzer Spiri- ferensandsteins und seiner Fauna. Abhandlungen zur Geologischen
Spezialkarte von Preussen und den Thiiringischen Staaten, 6(1). Billings, E. 1874. Palaeozoic fossils. Vol. 2, partie 1. Commission
gkologique du Canada, Ottawa. Boucot, A.J. 1963. The Globithyrid facies of the Lower Devonian.
Senckenbergiana Lethaea, 44 : 79 - 84. Boucot, A. J. 1975. Evolution and extinction rate controls. Develop-
ments in palaeontology and stratigraphy. Vol. 1. Elsevier Scien- tific Publishing Company, Inc., New York.
Boucot, A.J. 1982. Ecostratigraphic framework for the Lower Devonian of the North American Appohimchi Subprovince. Neues Jahrbuch fiir Geologie und Palaontologie, Abhandlung, 163 : 81 - 121.
Boucot, A.J., et Johnson, J.G. 1967. Paleogeography and correlation of Appalachian province Lower Devonian sedimentary rocks. Tulsa Geological Society Digest, 35 : 33-87.
Boucot, A.J., Curnming, L.M., et Jaeger, H. 1967. Contributions to the age of the Gasp6 Sandstone and Gasp6 Limestone. Commission gtologique du Canada, Ctude no 67-25.
Boucot, A.J., Johnson, J.G., et Talent, J.A. 1969. Early Devonian brachiopod zoogeography. Geological Society of America Special Paper no 1 19.
Boyd, D.W., et Newell, N.D. 1968. Hinge grades in the evolution of Crassatellacean bivalves as revealed by Permian genera. Ameri- can Museum Novitates, no 2328.
Bretsky, S.S. 1976. Evolution and classification of the Lucinidae (Mollusca; Bivalvia). Palaeontographica Americana, 8(50) : 2 15 - 337.
Brett, C.E., et Baird, G.C. 1986. Comparative taphonomy: a key to paleoenvironmental interpretation based on fossil preservation. Palaios, 1 : 207 -227.
Brisebois, D. 1981. Gtologie de la rCgion de GaspC. Ministkre de 1'Energie et des Ressources du Quebec, DPV-824.
Cant, D.J., et Walker, R.G. 1976. Development of a braided-fluvial facies model for the Devonian Battery Point Sandstone, QuCbec. Revue canadienne des sciences de la Terre, 13 : 102- 119.
Chavan, A. 1967. Sur les HCtCrodontes du Dtvonien rhCnan (Mol- lusques PClCcypodes). Compte rendu sommaire des sCances de la SocietC gCologique de France, fascicule 2. pp. 4-46.
Chavan, A. 1969. Superfamily Lucinacea Fleming, 1828. Dans Treatise on invertebrate paleontology. Vol. 2, partie N. Mollusca 6, Bivalvia. Editeurs : R.C. Moore et C. Teichert. Geological Society of America, Inc., Boulder, et University of Kansas, Lawrence. pp. N491 -N518.
Clarke, J.M. 1907. Some new Devonic fossils. New York State Museum, Bulletin no 107. pp. 153-291.
Clarke, J.M. 1908. Early Devonic history of New York and eastern North America. New York State Museum, Memoir no 9, partie 1.
Cox, L.R. 1969. Evolutionary history of Bivalvia. Dans Treatise on invertebrate paleontology. Vol. 2, partie N. Mollusca 6, Bivalvia. Editeurs : R.C. Moore et C. Teichert. Geological Society of America, Inc., Boulder, et University of Kansas, Lawrence. pp. N109-N118.
Dahmer, H.G. 1922. Studien iiber die Fauna des Oberharzer Kahle- bergsandsteins. 11. Jahrbuch der Preussischen Geologischen Landesanstalt, 40, partie 2 : 161 -306.
Dahmer, H.G. 1942. Zur Kenntnis der Beziehungen zwischen der Oberharzer und der bergisch-sauerlandischen Fazies der Oberko- blenz-Schichten (Unter-Devon). Senckenbergiana, 25 : 99- 103.
Dahmer, H.G. 1946. Revidiertes Verzeichnis der Versteinerungen des Oberharzer Kahleberg-Sandsteins (Unter-Devon). Senckenber- giana, 27 : 167 - 187.
Dechaseaux, C. 1952. Classy des Lamellibranches. Dans Trait6 de palContologie, Tome 2. Editeur : J. Piveteau. Masson et Cie, Paris. pp. 220-364.
Dechaseaux, C. 1960. Bivalves fossiles. Dans Trait6 de zoologic. Anatornie, systkmatique, biologie. Tome 5, fascicule 2. Editeur : P.P. GrassC. Masson et Cie, Paris. pp. 2134-2164.
De Raff, J.F.M., Boersma, J.R., et VAN Gelder, A. 1977. Wave- generated structures and sequences from a shallow marine succes-
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.

DESBIENS 391
sion, Lower Carboniferous, County Cork, Ireland. Sedimentology, 24 : 451 -483.
Desbiens, S. 1991. Le complexe deltaique de la Formation de York River (DCvonien infkrieur) de la rCgion de Gasp6 : palCoCcologie et biostratigraphie. Thbse de Ph. D., UniversitC de Montreal, MontrCal.
Driscoll, E.G. 1970. Selective bivalve shell destruction in marine environments, a field study. Journal of Sedimentary Petrology, 40 : 898 - 905.
Dutro, J.T., Jr. 1981. Devonian brachiopod biostratigraphy of New York State. Dans Devonian biostratigraphy of New York. Part 1. Editeurs : W.A. Oliver Jr. et G. Klapper. International Union of Geological Sciences, Subcommission on Devonian Stratigraphy, Antwerpen. pp. 67 - 82.
Eliott, T. 1974. Interdistributary bay sequences and their genesis. Sedimentology, 21 : 61 1 -622.
Eliott, T. !986. Deltas. Dans Sedimentary environments and facies. 2" Cd. Editeur : H.G. Reading. Blackwell Scientific Publications, Ltd., Oxford. pp. 113- 154.
Friend, P.F. 1961. The Devonian stratigraphy of north and central Vestspitsbergen. Proceedings of the Yorkshire Geological Society, 33 : 77-118.
Fuchs, G. 1982. Upper Siegenien and Lower Emsien in the Eifel Hills. Courier Forschungsinstitut Senckenberg, 55 : 229-256.
Ganong, W.F. 1934. Crucial maps in the early cartography and place- nomenclature of the Atlantic coast of Canada. VI. The voyages of Jacques Cartier. Transactions of the Royal Society of Canada, sCrie 3, section 2, 28 : 149-294.
Ganong, W.E , et Layng, T.E. 1964. Crucial maps in the early cartog- raphy and place-nomenclature of the Atlantic coast of Canada. Royal Society of Canada, Special Publications, no 7.
Haffer, J. 1959. Der Schlossbau Friih-Heterodonter Lamelli- branchiaten aus dem rheinischen Devon. Palaeontographica, Abteilung A, Palaeozoology -Stratigraphic, 112 : 133 - 192.
Hunter, R.D. 1990. Shell dissolution rate in freshwater pulmonate snails: influence of periostracum and water chemistry. Nautilus, 104 : 87-91.
Johnson, J.G. 1979. Devonian brachiopod biostratigraphy. Dans The Devonian system. Editeurs : M.R. House, C.T. Scrutton et M.G. Basset. Special Papers in Palaeontology no 23 : 291 -306.
Johnston, P.A., et Goodbody, Q.H. 1989. Middle Devonian bivalves from Melville Island, Arctic Canada. Dans Devonian of the world. Vol. III. Paleontology, paleoecology, and biostratigraphy. Editeurs : N.J. McMillan, A.F. Embry et D.J. Glass. Proceedings of the Second International Symposium on the Devonian System, Calgary, aoiit 1987, Canadian Society of Petroleum Geologists, Calgary. pp. 337-346.
Kat, P.W. 1982. Shell dissolution as a significant cause of mortality for Corbicula fluminea (Bivalvia: Corbiculidae) inhabiting acidic waters. Malacological Review, 15 : 129- 134.
Kat, P.W. 1983. Conchiolin layers among the Unionidae and Margari- tiferidae (Bivalvia): microstructural characteristics and taxonomic implications. Malacologia, 24 : 298 - 3 1 1.
Kat, P.W. 1985. Convergence in bivalve conchiolin layer rnicrostruc- ture. Malacological Review, 18 : 97 - 106.
Khalfin, L.L. 1948. Faouni i stratigrafiia devonskilch otlojenii Gornogo Altaia. Izvestiya Tomskogo Politekhnicheskogo Instituta, 65(1). [En russe.]
Khalfin, L.L. 1974. Classification key of the Early Devonian Pelecypoda of the Gorny Altai. Sibirskii Nauchno-Issledovatel'skii Institut Geologii Geofiziki i Mineral'nogo Syr'ya 164. [En russe.]
Lavoie, D., TassC, N., et Asselin, E. 1990. Lithostratigraphic frame- work of the Upper Gasp6 Limestones (Early Devonian) in eastern GaspC basin, Quebec. Dans Recherches en cours, partie B. Com- mission gkologique du Canada, Ctude no 90-ID. pp. 25 -35.
Lawrence, D.A. 1986. Sedimentology of the Lower Devonian Bat- tery Point Formation, eastern GaspC Peninsula, Quebec, Canada. Thkse de Ph. D., Bristol University, Bristol.
Lawrence, D.A., et Rust, B.R. 1989. The Devonian clastic wedge of
eastern Gasp6 and the acadian Yrogeny. Dans Devonian of the world. Vol. 1. Sedimentation. Editeurs : N.J. McMillan, A.F. Embry et D.J. Glass. Proceedings of the Second International Symposium on the Devonian System, Calgary, aoQt 1987. Cana- dian Society of Petroleum Geologists, Calgary. pp. 53 -64.
Lawrence, D. A., et Williams, B.P. J. 1987. Evolution of drainage systems in response to Acadian deformation: the Devonian Battery Point Formation, eastern GaspC. Society of Economic Paleontolo- gists and Mineralogists Special Publication no 39. pp. 287-300.
LespCrance, P.J. 1980. Calcaires SupCrieurs de, GaspC. Les aires types et le prolongement ouest. Ministhe de 1'Energie et des Res- sources du QuCbec, DPV-595.
Lewy, Z., et Samtleben, C. 1979. Functional morphology and palaeontological significance of the conchiolin layers in corbulid pelecypods. Lethaia, 12 : 341 -351.
McCall, P.L., et Tevesz, M.J.S. 1983. Soft-bottom succession and the fossil record. Dans Biotic interactions in recent and fossil ben- thic communities. Editeurs : M.J. S. Tevesz et P.L. McCall. Topics in geobiology. Vol. 3. Plenum Press, New York. pp. 157-194.
McGerrigle, H.W. 1950. La gkologie de l'est de GaspC. Ministihe des Mines du QuCbec, RG 35.
McGregor, D.C. 1973. Lower and Middle Devonian spores of east- ern GaspC, Canada. I. Systematics. Palaeontographica, Abteilung B, Palaeophytologie, 142 : 1-77.
McGregor, D.C. 1977. Lower and Middle Devonian spores of east- ern GaspC, Canada. 11. Biostratigraphy. Palaeontographica, Abteilung B, Palaeophytologie, 163 : 11 1 - 142.
Morris, N.J. 1978. The infaunal descendants of the Cycloconchidae: an outline of the evolutionary history and taxonomy of the Hetero- conchia, superfamilies Cycloconchacea to Chamacea. Philosophi- cal Transactions of the Royal Society of London, Series B, Biological Sciences, 284 : 259 - 275.
Newell, N.D. 1965. Classification of the Bivalvia. American Museum Novitates, no 2206.
Noe-Nygaard, N., Surlyk, F., et Piasecki, S. 1987. Bivalve mass mortality caused by toxic dinoflagellate blooms in a Berriasian- Vallangian lagoon, Bornholm, Denmark. Palaios, 2 : 263-273.
@wig, T. 1969. Vertebrates of the Wood Bay Group and the position of the Emsian-Eifelian boundary in the Devonian of Vestspitsber- gen. Lethaia, 2 : 273-328.
Pojeta, J. , Jr. 1971. Review of Ordovician pelecypods. United States Geological Survey Professional Paper no 695.
Pojeta, J., Jr. 1978. The origin and early diversification of pelecy- pods. Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences, 284 : 225-246.
Pojeta, J., Jr. 1987. Class Pelecypoda. Dans Fossil invertebrates. Editeurs : R.S. Boardman, A.H. Cheetham et A.J. Rowell. Black- well Scientific Publications, Ltd., Oxford. pp. 386-435.
Quenstedt, W. 1926. Mollusken aus den Redbay- und Greyhook- Schichten. Resultater av de Norske StatsunderstGttede Spitsber- genekspeditioner (Skrifter om Svalbard og Ishavet), Oslo, l(11).
Remane, A. 1971. Ecology of brackish water. Partie 1. Dans Biology of brackish water. 2" Cd. rCvisCe. Editeurs : A. Remane et C. Schlieper. John Wiley & Sons, Inc., New York. pp. 1-210.
Richardson, J.B., et McGregor, D.C. 1986. Silurian and Devonian spore zones of the Old Red Sandstone Continent and adjacent regions. Commission gCological du Canada, bulletin no 364.
Rust, B.R. 1981. Alluvial deposits and tectonic style: Devonian and Carboniferous successions in easJern GaspC. Dans Sedimentation and tectonics in alluvial basins. Editeur : A.D. Miall. Geological Association of Canada Special Paper no 23. pp. 49-76.
Rust, B.R., Lawrence, D.A., et Zaitlin, B.A. 1989. The sedimentol- ogy and tectonic significance of Devonian and Carboniferous ter- restrial successions in GaspC, Quebec. Atlantic Geology, 25 : 1-13.
Sanchez, T.M., et Babin, C. 1993. Un insolite mollusque bivalve, Catamarcaia n. g., de 1'Arenig (Ordovicien infkrieur) d'Argen- tine. Comptes rendus de 1'Acadtmie des sciences de Paris, sCrie 2, 316: 265-271.
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.

392 CAN. J . EARTH SCI. VOL. 31, 1994
Scarlato, O.A., et Starobogatov, Ya.1. 1979. General evolutionary patterns and the system of the class Bivalvia. Trudy Zoologi- cheskogo Instituta, Akademiya Nauk SSSR, 80: 5-38. [En russe.]
Scarlato, O.A., et Starobogatov, Ya.1. 1985. General evolutionary patterns and the system of the class Bivalvia. Harvard University, Department of Mollusks, Special Occasional Publication no 5. [Traduction anglaise de Scarlato et Starobogatov 1979.1
Scotese, C.R., et McKerrow, W.S. 1990. Revised world maps and introduction. Dans Palaeozoic palaeogeography and biogeography. Editeurs : W.S. McKerrow et C.R. Scotese. Geological Society, London, Memoir no 12. pp. 1-21.
Sikander, A.H. 1976. Lithological observation on Grande Grkve, York River and Battery Point Formations, and hydrocarbon possi- bilities in eastern GaspC, Quebec. Ministhe de 1'Energie et des Ressources du QuCbec, GM-33690.
Spriestersbach, J. 1919. Die Stellung von Montanaria Spriestersbach und Crassatellopsis Beushausen. Jahrbuch der Preussischen Geo- logischen Landesanstalt, 39 : 32-40.
Spriestersbach, J. 1925. Die Oberkoblenzschichten des Bergischen Landes und Sauerlandes. Jahrbuch der Preussischen Geologischen Landesanstalt, 45 : 367 -450.
Spriestersbach, J. 1942. Lenneschiefer (Stratigraphie, Fazies und Fauna). Abhandlungen des Reichsamts f i r Bodenforschung (Ger- many), 203.
Spriestersbach, J. , et Fuchs, A. 1909. Die Fauna der Remscheider Schichten. Abhandlungen der Koniglich Preussischen Geolog- ischen Landesanstalt, 58.
Stoltidis, I. 1971. Ostrakoden aus dem Unterdevon des Bergischen
Landes (Rheinisches Schiefergebirge) . Decheniana, 124 : 1 - 38. Tevesz, J.S., et Carter, J.G. 1980. Environmental relationships of
shell form and structure of unionacean bpalves. Dans Skeletal growth of aquatic organisms. Chapitre 8. Editeurs : D.C. Rhoads et R.A. Lutz. Plenum Press, New York. pp. 295-322.
Vietor, W. 1919. Der Koblenzquarzit, seine Fauna, Stellung und linksrheinische Verbreitung. Jahrbuch der Koniglich Preussischen Geologischen Landesanstalt, 37 : 317-476.
Vogel, K. 1962. Muscheln mit Schlossziihnen an dem spanischen Kambrium und ihre Bedeutung fiir die Evolution der Lamelli- branchiaten. Akademie der Wissenschaften und der Literatur, Abhandlungen der Mathematisch-Natunvissenschaftlichen Klasse (Mainz), no 4, pp. 197 -244.
Vokes, H.E. 1967. Genera of the Bivalvia: a systematic and biblio- graphic catalogue. Bulletins of American Paleontology, 51(232): 109-394.
Vokes, H.E. 1980. Genera of the Bivalvia: a systematic and biblio- graphic catalogue (revised and updated). Paleontological Research Institution, Ithaca.
Wagge, L.E., et Mittler, T. 1953. Shell regeneration in some British molluscs. Nature (London), 171 : 528-529.
Wilbur, K.M. 1964. Shell folfnation and regeneration. Dans Physiol- ogy of Mollusca. Vol. 1. Editeurs : K.M. Wilbur et C.M. Yonge. Academic Press, New York. pp. 243-282.
Wilbur, K.M., et Watabe, N. 1963. Experimental studies on calcifi- cation in molluscs and the alga Coccolithus huxleyi. Annals of the New York Academy of Sciences, 109 : 82- 112.
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
CO
NC
OR
DIA
UN
IV o
n 11
/14/
14Fo
r pe
rson
al u
se o
nly.