Embed Size (px)
Citation preview

L’évolution de la biodiversité au cours des temps géologiques.
Préparation au concours de l’agrégation, Université Pierre & Marie Curie.
Leçon de démonstration secteur C.
Proposition de plan par : Mathieu Rodriguez, Pr. Agrégé SV-STU & doctorant iSTeP-ENS.
Adresse mail : [email protected]
Introduction :
*Actuellement, du fait de l’action de l’homme, se déroule une crise biologique : les
espèces disparaissent à un rythme anormal.
*La plus grande crise de l’histoire de la vie est l’apparition de l’oxygène dans
l’atmosphère, il y a 1.8 Ga. Seules les bactéries pourpres, ainsi que les autres bactéries dont
elles étaient le symbiote, étaient capables de lutter contre l’action toxique de l’oxygène via
leur chaîne respiratoire. Elles ont donc été sélectionnées. Limiter cependant le sujet aux temps
phanérozoïques, en raison de la présence d’organismes à « corps dur » fossilisables. Quitte à
ignorer 85 % de l’histoire de la vie!
*L’échelle des temps géologiques est évènementielle, ses subdivisions correspondant à
des crises de +/- grande amplitude. Il y a eu 5 grandes crises biologiques majeures dans
l’histoire du vivant au cours du Phanérozoïque.
*Définition de la biodiversité (terme défini en 1992 à la conférence de Rio de Janeiro):
Le terme « biodiversité » est la contraction de « diversité biologique ». L’étude de la
biodiversité rend compte de la variabilité du vivant, depuis l’échelle moléculaire jusqu’aux
interactions entre espèces. En pratique l’unité de la biodiversité est l’espèce.
*Problématique : L’estimation actuelle de la biodiversité présente des incertitudes (ex.
la diversité des arthropodes, en particulier les insectes tropicaux): qu’en est-il pour le passé ?
Comment la biodiversité a-t-elle évolué au cours des temps géologiques ? Quels sont les
facteurs et les mécanismes qui contrôlent la biodiversité ?

1) Estimer la biodiversité dans les temps géologiques : méthodes et limites.
La biodiversité du passé est inscrite dans les roches, qui constituent les archives de la vie
sur Terre. Mais ces données sont-elles représentatives de l’histoire de la biodiversité ?
a) Quelle unité pour estimer la biodiversité ?
L’utilisation des genres et des familles introduit un biais : un genre (ou une famille)
peut comprendre 50 espèces, un autre une seule espèce. Pour une approche quantitative de la
biodiversité, il est préférable d’utiliser l’espèce comme unité de mesure. Se pose alors la
question de la définition de l’espèce en paléontologie. Le critère de définition principal d’une
espèce vivante est l’interfécondité entre individus de la même espèce, et la production d’une
descendance elle-même interféconde… Critère difficilement applicable sur des fossiles !
Deux animaux peuvent appartenir à une même espèce sur le plan morphologique, mais
différer sur le plan génomique. Les limites d’une espèce en paléontologie sont parfois floues
et arbitraires. Ex. Les différentes espèces de globigérines, espèce de foraminifère asexuée
dont l’évolution est graduelle, sont définies selon le diamètre de leur Test. Autre exemple : le
cœlacanthe, dont la morphologie actuelle est proche des fossiles du paléozoïque, a
probablement beaucoup évolué sur le plan génomique sans que cela ne se répercute sur son
apparence.
b) Quels sont les biais à prendre en compte lorsqu’on estime la biodiversité à l’échelle
des temps géologiques ?
Les aléas de la fossilisation : seuls les tissus durs (squelette, dents, bois) se fossilisent
bien. Par conséquent, il existe peu de fossiles d’organismes constitués de tissus mous, comme
les méduses, les végétaux, et les micro-organismes. Des comptages ont permis de montrer que
seul 1% du plancton se retrouve dans le sédiment au fond de l’océan, en raison de leur
dissolution (lors du passage de la CCD) : les espèces qui y sont le plus sensibles ne sont donc
pas fossilisées. Il y a donc une forme de tri sélectif. Il y a de plus des périodes et des lieux où
les processus de fossilisation étaient plus ou moins efficaces. De plus, des sites fossilifères
peuvent subir une importante érosion, et ne pas être conservés au cours du temps - c’est ce qui
expliquerait en partie la très faible quantité de sites disponibles pour étudier la crise Permo-
Trias. Des méthodes d’extrapolation, qui postulent que le rapport entre organismes
fossilisables et non fossilisables est constant au cours du temps pour un environnement donné,
permettent de s’affranchir, dans une certaine mesure, des biais liés à la fossilisation, lorsque
couplées au principe d’actualisme.
D’autre part, l’étude de la biodiversité en géologie ne prend en compte que la diversité
des fossiles, qui n’image pas nécessairement la biodiversité réelle : par exemple, la
distribution des espèces actuelles montre que les espèces les plus nombreuses sont
bactériennes, que l’on retrouve très rarement à l’état fossile ! Sans compter d’autre biais,
d’ordre méthodologique : il y a en effet des corrélations entre l’évolution du nombre
d’espèces au cours du temps, le nombre de spécialistes pour les différents âges géologiques et

la superficie des affleurements pour chaque âge. Est-ce parce qu’un âge comporte beaucoup
d’espèces qu’il y a plus de spécialistes pour l’étudier ? Ou est-ce parce qu’il y a plus de
spécialistes que plus d’espèces ont été découvertes pour cet âge ? De même, plus la surface
d’affleurement d’un âge est grande, plus il y a de spécialistes…
c) Les courbes de l’évolution de la biodiversité au cours du temps
En vertu de tous ces biais, la courbe d’évolution de la biodiversité est-elle
représentative de la réalité passée ? Et si les variations qu’elle met en évidence sont
discutables, peut-on vraiment parler de crises ?
Les courbes d'évolution de la biodiversité ne représentent certainement pas la réalité...
Mais cela reste une forme de "signal" qui mérite d'être étudié en ayant à l'esprit tous les biais
qui le composent! Il existe deux courbes de l’évolution de la biodiversité : une concernant les
familles terrestres, l’autres les familles marines. Celles-ci n’ont pas la même allure, car, bien
sûr, au Paléozoïque, la vie terrestre n’était pas encore complètement installée ; de plus, les
espèces marines et terrestres n’ayant pas le même écosystème, elles réagiront différemment
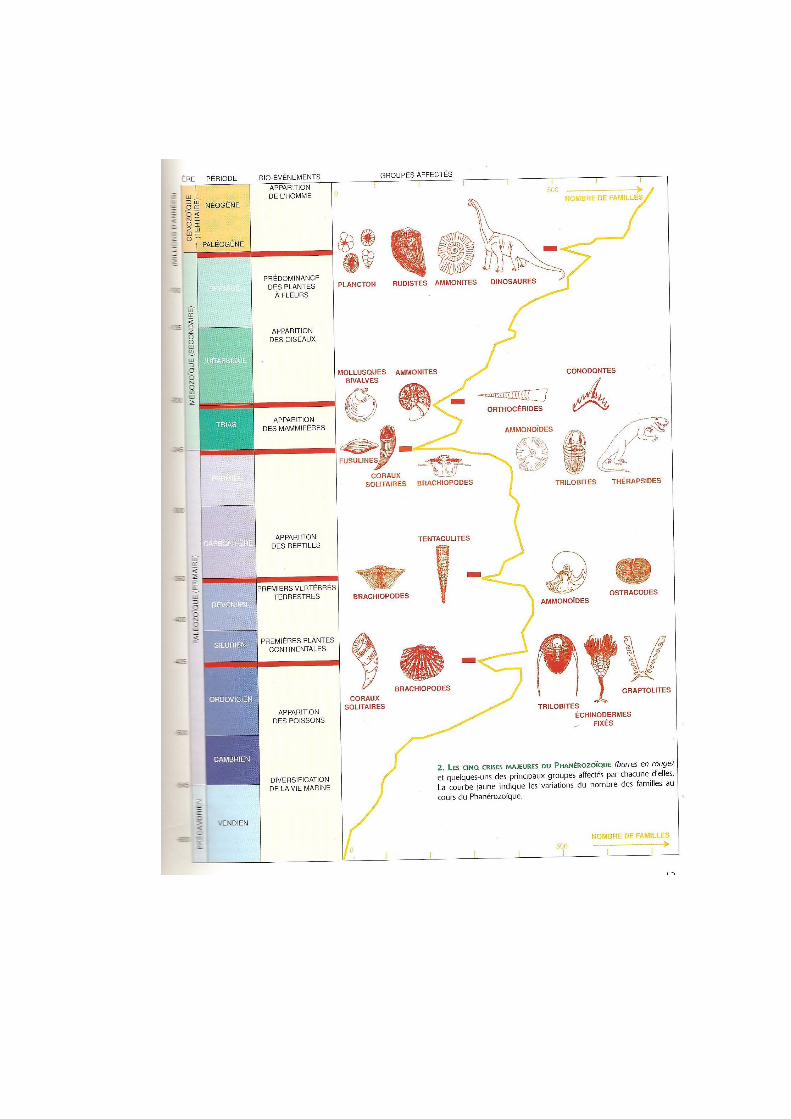
selon la nature du stress imposé. Quoiqu’il en soit, la courbe affiche clairement 5 grandes
crises majeures au cours du Phanérozoïque. Ces 5 grandes crises ont eu lieu : 1) à
l’Ordovicien supérieur (445 Ma), 2) au Dévonien supérieur (375 Ma), 3) aux limites Permo-
Trias(P/T) (250 Ma, la plus importante de toutes avec près de 95% des espèces éteintes), 4) et
Trias-Jurassique (T/J) (200 Ma), et enfin 5) Crétacé-Tertiaire (K/T) (65 Ma). Ces crises
définissent les divisions majeures de l’échelle des temps géologiques.
A l’aide d’un tableau bilan, associer chaque crise à la disparition de certains
groupes représentatifs (ex. les dinosaures non-aviens pour la crise K-T), et préciser le
% d’espèces disparues.
Cependant, après chaque crise, la biodiversité augmente. Quelles sont les causes des
extinctions, et comment la biodiversité finit-elle par reprendre ses droits ?
2) Les crises biologiques.
Mise en évidence de la crise K/T, à partir des microfossiles (ex. d’affleurement en France,
la plage de Bidart au Pays Basque) : les couches du Maastrichtien sup. sont riches en
Globotruncanidés et en Hétérohélécidés, qui disparaissent dans les couches du Danien, au
profit des Globigérinidés. A partir de l’étude simple de deux lames minces nous mettons en
évidence la disparition d’espèces et leur renouvellement après une crise. Quelles sont les
causes de ces remaniements de la biodiversité ?
a) A la recherche des causes des crises biologiques
La crise biologique actuelle, conséquence des activités humaines, le montre bien : les
espèces biologiques disparaissent en réponse aux changements de leur milieu de vie, auxquels
elles ne sont pas adaptées. Ici, nous partons à la recherche des grands évènements naturels
susceptibles d’avoir modifié l’environnement des espèces et causé leur extinction, à l’époque
des grandes crises. Plusieurs causes, parfois contemporaines, sont identifiées :

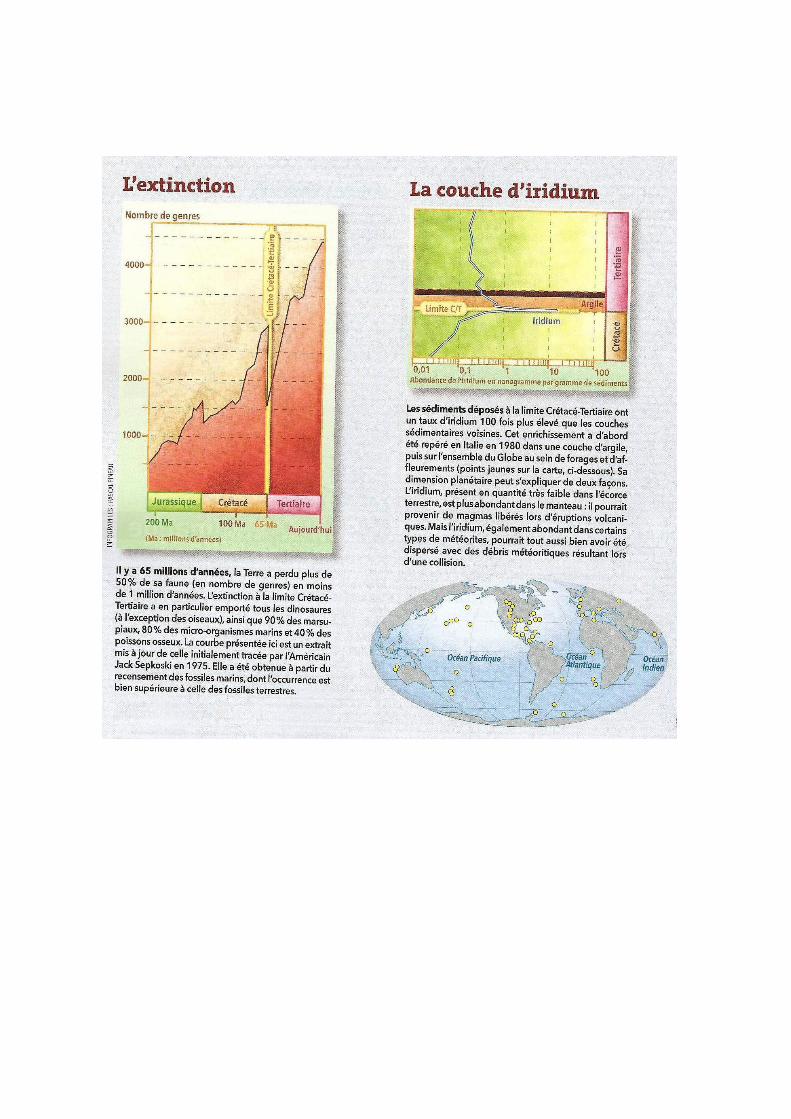
Les météorites : des traces d’impact de météorites ont été trouvées pour les
crises K/T, P/T, T/J. L’exemple le plus connu est le cas du cratère du Chicxulub, au
Mexique, qui serait une des causes possibles de la crise K/T. En plus de ce cratère, la
limite K/T se caractérise stratigraphiquement par une concentration anormalement
élevée en Iridium, un élément caractéristique des météorites. L’anomalie en Iridium
est mondiale. La limite K/T est également enrichie en spinelle nickélifère, et en
minéraux choqués. Les impacts de météorite dégagent de la poussière dans
l’atmosphère, entraînant une baisse des températures ; ils génèrent des tsunamis si
l’impact a lieu dans l’océan ; ils entraînent la vaporisation de plusieurs éléments
contenus dans les roches ou la biomasse (composés destructeurs de la couche
d’Ozone, Soufre contenu dans le gypse (diminution de la T°), CO2 contenu dans les
carbonates (augmentation de la T°).
Les trapps : De grands épanchements volcaniques sont mis en place de façon
contemporaine aux plus grandes crises : les trapps du Deccan pour la crise K/T (2.4.
106 km
3); les trapps de Sibérie pour la crise P/T (>3. 10
6 km
3 initialement); la province
magmatique de l’Atlantique central pour la crise T/J (2. 106 km
3). Les datations radio-
chronologiques sont capitales pour relier une province magmatique à la crise
correspondante. Ces éruptions importantes relarguent elles aussi des poussières dans
l’atmosphère, du soufre, du CO2. L’altération des trapps consomme du CO2.
(Le Soufre entraîne une diminution de la T° sur 100 ans env. ; le CO2 entraîne
une augmentation de la T° sur 1000 ans env.)
Les variations du niveau marin (NM) et les glaciations : il existe une
corrélation entre les crises et les variations du niveau marin. Les zones de plate-forme
continentales sont parmi celles qui concentrent la plus grande biodiversité, et sont
donc les plus sensibles en terme d’extinction. Par exemple, au Permien moyen, le
niveau des mers était l’un des plus hauts jamais atteints, environ 200 m au dessus du
niveau actuel. Le niveau marin chuta progressivement de 280 à 250 Ma d’environ 250
m, émergeant ainsi la plupart des domaines de plate-forme continentale, où était
concentré l’essentiel de la biodiversité. Les espèces marines ont donc été plus
affectées que les espèces continentales lors de la crise Permo-Trias. Les espèces fixes
(ex. crinoïdes) ont été plus affectées que les espèces mobiles. La baisse du NM
entraîne une compétition biologique accrue sur la plate forme.
Une glaciation est retenue comme la principale cause de la crise
Ordovicienne : les changements climatiques, les variations du NM consécutives, et
l’arrivée d’eaux appauvries en oxygène sur le plateau continental lors de la
déglaciation auraient été fatals à de nombreuses espèces.
La tectonique des plaques : les migrations des continents favorise le
développement des eaux océaniques profondes et froides (moins soumises aux
changements), modifie la composition chimique des océans et en perturbe les
courants ; elle provoque des collisions et la formation d’isthmes, lesquels favorisent
les migrations et les compétitions entre les espèces.

b) La durée des crises
Les causes invoquées pour les grandes crises sont souvent catastrophiques, et impliquent
une disparition rapide des espèces. Cependant, la disparition des Ichtyosaures (reptiles marins,
n'appartenant pas aux dinosaures(!)) au Crétacé sup. précède la crise et ne peut être attribuée
à l’action conjuguée de la météorite du Yutacan et des trapps du Deccan. De plus, on peut se
demander si l’enregistrement fossile, dont nous avons déjà exposé certains biais, permet de
discriminer entre une extinction en masse graduelle ou instantanée. L’enregistrement fossile
est en effet discontinu dans le temps. Lorsqu’une espèce est présente à la base et au sommet
d’une série sédimentaire, on en conclut qu’elle vivait pendant tout l’intervalle de temps
enregistré par la série. La répartition stratigraphique des occurrences étant aléatoire, se pose le
problème de la signification de la dernière occurrence d’une espèce, laquelle ne reflète
probablement pas le moment de la véritable extinction de cette espèce. Ainsi, des extinctions
apparentes dans l’enregistrement fossile anticipent les véritables extinctions, et donnent
l’impression d’extinctions graduelles. Ce problème est en partie résolu lorsqu’on attribue à

chaque occurrence d’espèce sur un affleurement donné un intervalle de confiance calculé à
partir de tous les spécimens découverts dans le monde. Pour le cas des ammonites de la baie
de Biscaye (voir doc.) l’extinction apparaît alors instantanée à la limite K/T. Cet artefact de
l’enregistrement sédimentaire porte le nom d’effet Signor-Lipps.
c) Crises et extinctions : étude comparée et synthèse
Les crises touchent des milieux écologiques aux vulnérabilités différentes : certaines
espèces disparaissent, d’autres sont fortement atteintes sans pour autant disparaître, d’autres
enfin ne sont qu’effleurées. De façon générale, on remarque une intensité différente des
extinctions entre le domaine continental et le domaine marin, une meilleure résistance du
benthos profond que du benthos littoral (les récifs sont ravagés par chaque crise). Les espèces
endémiques et spécialisées sont les plus fragiles, tout comme celles de grande taille.
La biodiversité semble avoir toujours été contrôlée par la dispersion ou le regroupement
des continents, les variations climatiques, les variations du niveau marin pour les extinctions
délimitant les étages et les époques…et par des évènements catastrophiques comme les
grandes éruptions volcaniques, ou les météorites pour les grandes extinctions délimitant les
ères. Il s’agit principalement de facteurs liés au fonctionnement interne de la planète, auxquels
se rajoutent les causes extra-terrestres, comme les météorites. Les crises sont pluri-
factorielles et sont la conséquence d’un enchaînement de processus catastrophiques plus que
d’une seule cause isolée.
3) Les processus de renouvellement de la biosphère.
Une extinction en masse fait évoluer le mode vivant et sa biodiversité par les
redéploiements et les reconquêtes des milieux redevenus plus hospitaliers à l’issue de la crise.
a) Les différentes stratégies de survie à une crise biologique.
On distingue diverses catégories d’organismes selon leur réaction à la crise : les
exterminés (ceux qui disparaissent pour de bon), les opportunistes, et les survivants.
Les espèces « désastres », ou opportunistes : Ces formes de vie, rares avant la crise,
profitent de la crise pour se développer, avant de régresser puis de disparaître à la fin de la
période néfaste. Par exemple, après la crise du Permo- Trias, l’ensemble des fonds marins
était dévasté et appauvri en oxygène. Ces conditions ont favorisé la prolifération d’espèces
opportunistes, favorisées par ce genre de milieu car elles n’ont pas de concurrence écologique.
Ce sont généralement des espèces rudérales.
Les espèces survivantes ont des potentialités ou des comportements divers. Les
espèces pré-adaptées sont celles qui possèdent un caractère qui leur permet de s’adapter aux
conditions de la crise. Par exemple, l’endothermie a permis aux mammifères de passer la crise
K/T. Mais de façon générale, ce sont les espèces généralistes du point de vue de leurs
exigences écologiques, i.e. qui supportent de grandes variations de conditions du milieu
(ressources alimentaires, salinité, température…) qui traversent le mieux les crises. A la
différence des espèces opportunistes, ces espèces survivent mieux une fois de retour à des

conditions de vie normales. Un cas intriguant est celui des espèces Lazare, qui semblent
disparaître pendant la crise et réapparaissent au terme de celle-ci. Cet effet implique
l’existence, durant la crise, de refuges non repérés dans l’enregistrement géologique. Des
milieux de transition (marécages, eaux saumâtres) pourraient jouer le rôle de foyer de
recolonisation après les crises.
Les survivants les plus importants du point de vue de la biodiversité sont ceux qui
initient de nouvelles radiations après la crise : les géniteurs. Les géniteurs sont peux
nombreux pour un groupe donné, qui subit pendant la crise un goulot d’étranglement.
L’histoire des Ammonoïdes illustre bien cet effet, ce groupe ayant été ponctuellement décimé
par des crises pour s’éteindre définitivement à la crise K/T. Nous avons vu que les espèces de
petite taille sont favorisées en cas de crise.
b) Les mécanismes des radiations évolutives
La libération des niches écologiques suite à la disparition des espèces permet à de
nouvelles espèces de se développer, en l’absence de compétition écologique. La libération des
niches s’accompagne d’une réorganisation des réseaux trophiques. On assiste alors à une
augmentation rapide du nombre d’espèces, et à des diversifications morpho-anatomiques
rapides. Il s’agit d’un renouvellement profond et imprévisible de la lithosphère. Ainsi,
mammifères et oiseaux ont pu se diversifier au Tertiaire, suite à la disparition des dinosaures
non-aviens.
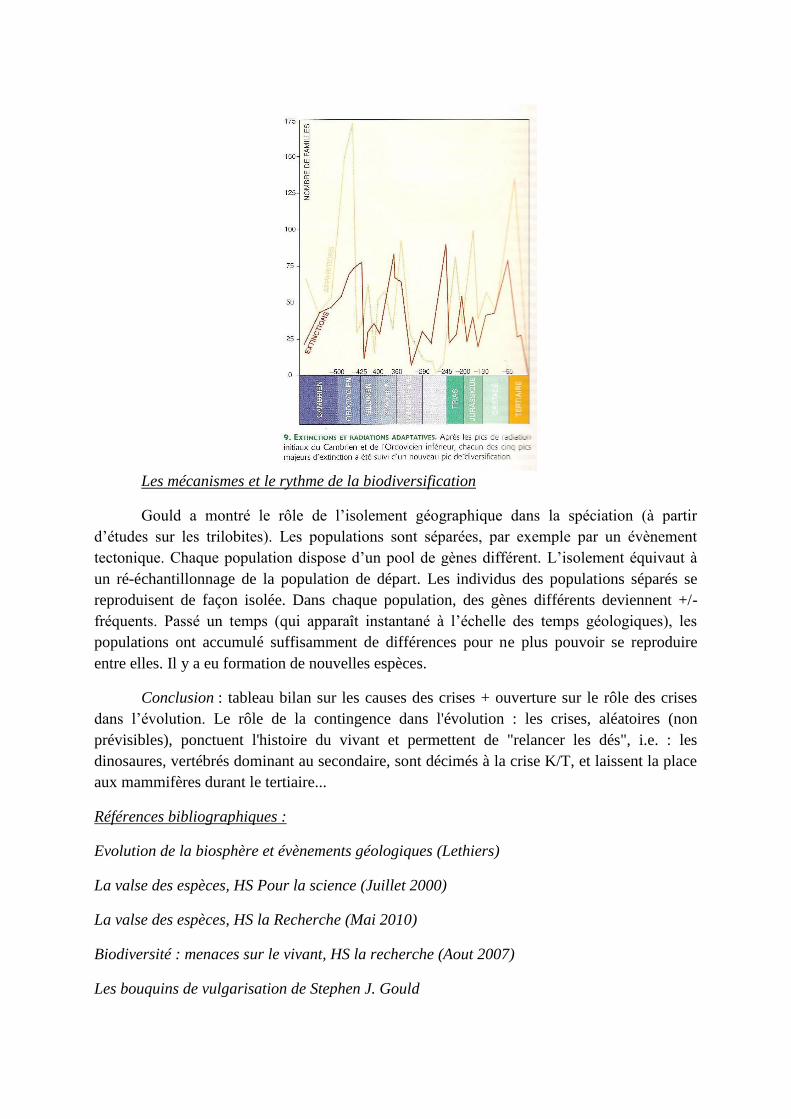
Les mécanismes et le rythme de la biodiversification
Gould a montré le rôle de l’isolement géographique dans la spéciation (à partir
d’études sur les trilobites). Les populations sont séparées, par exemple par un évènement
tectonique. Chaque population dispose d’un pool de gènes différent. L’isolement équivaut à
un ré-échantillonnage de la population de départ. Les individus des populations séparés se
reproduisent de façon isolée. Dans chaque population, des gènes différents deviennent +/-
fréquents. Passé un temps (qui apparaît instantané à l’échelle des temps géologiques), les
populations ont accumulé suffisamment de différences pour ne plus pouvoir se reproduire
entre elles. Il y a eu formation de nouvelles espèces.
Conclusion : tableau bilan sur les causes des crises + ouverture sur le rôle des crises
dans l’évolution. Le rôle de la contingence dans l'évolution : les crises, aléatoires (non
prévisibles), ponctuent l'histoire du vivant et permettent de "relancer les dés", i.e. : les
dinosaures, vertébrés dominant au secondaire, sont décimés à la crise K/T, et laissent la place
aux mammifères durant le tertiaire...
Références bibliographiques :
Evolution de la biosphère et évènements géologiques (Lethiers)
La valse des espèces, HS Pour la science (Juillet 2000)
La valse des espèces, HS la Recherche (Mai 2010)
Biodiversité : menaces sur le vivant, HS la recherche (Aout 2007)
Les bouquins de vulgarisation de Stephen J. Gould










![Synthèse et caractérisation d'une nouvelle phase Nd5Pt3[substitution,MEB,cristallographie,diffractionX,alliage ternaire,néodyme,platine,iridium]](https://img.pdfslide.fr/doc/110x75/55720900497959fc0b8bdd21/synthese-et-caracterisation-dune-nouvelle-phase-nd5pt3substitutionmebcristallographiediffractionxalliage-ternaireneodymeplatineiridium.jpg)


















