Embed Size (px)
Citation preview

UNIVERSITE FELIX HOUPHOUET BOIGNYUFR Biosciences
Laboratoire de Biotechnologies
ANNEE UNIVERSITAIRE : 2013-2014
LICENCE III DE BIOCHIMIE
UE : SYNTHESE, INTERET ET VALORISATION DES
METABOLITES SECONDAIRES
ECUE: BIOSYNTHESE DES METABOLITES SECONDAIRES
(COURS)
Dr. SORO Yadé René

INTRODUCTION
Les métabolites secondaires sont des composés naturels synthétisés par tous les types
d’organismes vivants (les plantes, les champignons, les bactéries, les algues et les animaux). Ils
sont considérés par certains auteurs comme ne jouant pas à priori un rôle fondamental dans la vie
de l’organisme producteur. Cependant, différentes classes de ces molécules constituent pour les
plantes des moyens de défense contre les prédateurs, d’interaction entre plantes, de pollinisation
etc. Pour l’homme également, les métabolites secondaires constituent les principes bioactifs de
plusieurs épices, plantes médicinales, aromatiques, colorantes et des aliments fonctionnels. Ces
propriétés suscitent évidemment un réel intérêt pour la détection, l’isolement et la caractérisation
en vue d’une meilleure utilisation de ces métabolites.
Des milliers de métabolites secondaires ont déjà été caractérisés. Certains sont très répandus
tandis que l’existence d’autres est limitée à quelques centaines d’espèces ou même à un genre et
parfois une seule espèce. Pour une même espèce la composition chimique des métabolites
secondaires peut aussi varier selon qu’elle pousse dans des endroits différents, même si on ne le
voit pas du point de vue anatomique. On distingue classiquement plusieurs catégories de
métabolites secondaires en fonction de leur nature biochimique et de leur origine biosynthétique.
La classification proposée ici correspond à ce que proposent la plupart des auteurs (malgré
quelques variantes). Elle repartit les milliers de molécules de ce type déjà isolées en deux
principaux groupes qui sont :
- les métabolites secondaires contenant l’azote (N) (amines, alcaloïdes)
- les métabolites secondaires sans azote (composés phénoliques et terpènoïdes)
Cette classification présente évidemment des limites, car un certain nombre de composés
d’origines biosynthétiques mixtes (exemple : la condensation d’un groupement isoprénoïde avec
une structure de type « composé phénolique » pour la synthèse des tocophérols ou des
furocoumarines) ne trouvent pas forcément une place logique, de même que le groupe des
hétérosides qui pose un certain nombre de problèmes car certains n’y incluent pas les dérivés
glycosylés des composés aromatiques.

ALCALOÏDES
Définition
Les alcaloïdes sont des substances naturelles azotés agissant comme des alcalis (de l’arabe al
kaly = soude et du grec eidos = aspect), qui précipitent avec des réactifs iodométalliques (réactif
de Dragendorff) et qui sont pour la plupart biologiquement actifs en raison de leurs propriétés.
En effet, Ils représentent les principes actifs de nombreuses plantes médicinales ou toxiques
connues parfois depuis l'Antiquité. A l’état naturel, ils sont généralement salifiés par des acides
organiques (citrates, malates, tartrates, …). Groupe de substances naturelles d’intérêt
thérapeutique le plus important en termes de nombre, de diversité structurale et de l’éventail de
leurs activités pharmacologiques, cette famille de métabolites secondaires a été particulièrement
étudiée du fait des enjeux économiques qui leur sont associés.
RépartitionSelon les alcaloïdes concernés la distribution peut être large ou restreinte. Dans le cas d’une
distribution large, un même alcaloïde peut se retrouver dans des familles botaniques qui sont très
éloignées. (Ex : le caféier (Rubiacée), le théier (Ternstromiacée), le kolatier (sterculiacée),
synthétisent tous une même molécule qui est la caféine). Dans le cas de la distribution restreinte,
un alcaloïde se retrouve uniquement au niveau d’une famille, d’un genre et même d’une espèce.
(Ex : l’hyoscyamine qu’on trouve dans toutes les plantes de la famille des Solanacées, la
thébaïne est restreinte au genre Papaver, La morphine est uniquement synthétisée au niveau de
l’espèce Papaver somniferum).
La teneur en alcaloïdes est variable selon les organes. Pour le Cinchona par exemple, l’extraction
de la quinine se fait à partir de l’écorce car il n’y a pas de quinine dans les feuilles. Chez le
caféier, seules les graines contiennent la caféine. Les alcaloïdes les plus toxiques sont présents en
très faible quantité, de l’ordre du ppm.
Structure/diversitéLes alcaloïdes forment un groupe hétérogène, du point de vue de la structure, des propriétés
chimiques et des effets biologiques qu'ils manifestent. Il existe ainsi une très grande diversité de
sous-familles d’alcaloïdes, qui ont été classés en fonction de leurs origines biosynthétiques et de
la nature des hétérocycles azotés. Selon les origines biosynthétiques on distingue ainsi :
- Les alcaloïdes vrais
- Les proto-alcaloïdes

- Les pseudo-alcaloïdes.
Cependant, certains alcaloïdes complexes font exception et associent dans leurs structures des
noyaux azotés dérivés d’acides aminés à des résidus terpènes (exemple : alcaloïdes indoliques
monoterpéniques de la pervenche de Madagascar).
Alcaloïdes vrais :Ils ont une structure souvent complexe avec l’azoté (N) inclus dans un hétérocycle. Leur origine
biosynthétique est un acide aminé dont la nature va déterminer le noyau de base de l’alcaloïde.
Ils sont doués de propriétés pharmacologiques significatives. (exemples : la strychnine est un
dérivé du tryptophane et la nicotine dérive de l’acide aspartique.
Dioscorine Nicotine
Pseudo-alcaloïdes :
Ils présentent le plus souvent toutes les caractéristiques des alcaloïdes vrais sauf que les acides
aminés ne constituent pas leur origine biosynthétique. Ils dérivent dans la majorité des cas
d’isoprénoïdes et sont désignés dans ce cas alcaloïdes terpéniques comme l’aconitine (alcaloïde
diterpénique) de l’aconit mais certains proviennent du métabolisme de l’acétate (exemple : la
coniine ).
Coniine Cryptophorine
Proto-alcaloïdes :
Ce sont des amines simples dont l’azote n’est pas inclus dans un hétérocycle. Ils sont élaborés in
vivo à partir d’acides aminés (Exemples : la mescaline et la sérotonine).

Mescaline Sérotonine
Constitution chimique et classification
Les processus biosynthétiques conduisant aux alcaloïdes sont maintenant assez bien connus pour
envisager, en fonction des précurseurs, une classification biogénétique relativement cohérente
qui tend à s'imposer. On distingue ainsi les :
- Pyrrolidines (hygrine)
- Pyridines (nicotine)
- Benzoisoquinoléine (morphine)
- Pipéridines (coniine, pelletierine)
- Tropanes (atropine, hyoscyamine)
- Quinoléines (quinine)
- Isoquinoléiques (papavérine)
- Aporphines (boldine)
- Quinolizidines ou norlupinane (spartéine, lupanine)
- Indoles ou benzopyrroles (yohimbine)
- Imidazoles ou glyoxalines (pilocarpine)
- Purines ou imidazole + pyrimidine (caféine)
- Quinazolines (péganine)
- Tropolone (colchicine)
Il existe également des alcaloïdes stéroïdiques et des alcaloïdes terpènoïques.
BiogénèseLes alcaloïdes sont élaborés dans tous les êtres vivants (végétaux, microorganismes et animaux)
à partir de précurseurs du métabolisme primaire qui, dans la majorité des cas, sont des acides
aminés. Ainsi, les alcaloïdes indoliques et la quinine dérivent du tryptophane, les tropaniques de
l'ornithine, les isoquinoléiques de la phénylalanine. Toutefois, d'autres unités métaboliques sont
simultanément incorporées, comme le mévalonate, précurseur des terpènes, dans la plupart des
alcaloïdes indoliques. Certains ont une autre origine, comme la coniine qui dérive d'un motif
polyacétique (−CH2−CO−)n. les grandes lignes de la biosynthèse des alcaloïdes indiquent que la

biosynthèse s'effectue par une suite de réactions enzymatiques. Cependant, certaines réactions
intermédiaires restent hypothétiques et des mécanismes chimiques restent à préciser. Les
exemples à titre d’illustration sont choisis parmi les mieux connus et les plus caractéristiques.
Biosynthèse d’un alcaloïde vrai : la nicotine
La nicotine est un alcaloïde de nicotiana. Sa biosynthèse commence par celle de ses deux
précurseurs (l’acide quinoléique et l’ion pyrrolidium). L’acide quinoléique est synthétisé à partir
du glycéraldéhyde Phosphate et de l’acide aspartique par la série de réactions suivantes (a).
a) + + Pi
L’ion pyrrolidium est issu de après les réactions suivantes (b)
b)
La condensation de ces deux composés aboutit à la formation de la nicotine au terme des
réactions suivantes (c).
c)
Biosynthèse d’un proto-alcaloïde : la coniineLa coniine est un alcaloïde de la grande cigüe. La biosynthèse de sa structure de base se fait par
la condensation de quatre unités malonyles. L’azote est ensuite apporté par la lysine.
L’incorporation de l’azote est suivie par la fermeture du cycle pour donner un hétérocycle
comme indiqué ci-dessous.
4 Malonyl CoA + lysine

-conicéine coniine
Biosynthèse d’un pseudo-alcaloïde : la mescalineLa mescaline est un alcaloïde du peyolt. Il a comme précurseur la tyrosine (un acide aminé
aromatique et hydroxylé). Il est obtenu au terme d’une série de réactions de décarboxylation,
d’oxydation et de méthylation.

TERPENOÏDES
Définition
Les Terpènoïdes ou isoprénoïdes sont des composés organiques synthétisés par la condensation
d’unités isopréniques. Ils sont présents chez tous les organismes vivants, remplissent des rôles
physiologiques très divers et constituent la famille la plus large et la plus diversifiée des produits
naturels. Plusieurs molécules de la saveur et des arômes comme le menthol, le géraniol, le
caryophyllène sont formées de deux ou trois unités isopréniques. Cependant, les molécules
formées de 4, 6, 8 unités isopréniques et plus sont aussi des composés bioactifs. Les isoprénoïdes
regroupent donc à la fois des molécules de faibles poids moléculaires, volatiles et composants
principaux d’huiles essentielles et des molécules polymérisées de haut poids moléculaires
comme le caoutchouc.
Origine de l’unité isoprèneChez tous les êtres vivants, l’unité de base de la biosynthèse des terpènoïdes est l’isopentényle
diphosphate (IPP = isopentényle pyrophosphate) et son isomère le diméthylallyle-diphosphate
(DMAPP). Deux voies de biosynthèse conduisent à ces unités de base à 5 carbones.
Ce sont :
- la voie de l’acétate/mévalonate
- la voie du methyl-erythrytol-phosphate (MEP).
a) voie de l’acétate/mévalonateLa plus ancienne connue. Elle débute par la condensation de 3 unités acétyl CoA, une série de
phosphorylations puis une décarboxylation qui aboutit à l’IPP. Celui-ci est finalement isomérisé
pour donner le diméthylallyl-diphosphate (DMAPP). Elle se déroule dans le cytosol.
TERPENOÏDES
Définition
Les Terpènoïdes ou isoprénoïdes sont des composés organiques synthétisés par la condensation
d’unités isopréniques. Ils sont présents chez tous les organismes vivants, remplissent des rôles
physiologiques très divers et constituent la famille la plus large et la plus diversifiée des produits
naturels. Plusieurs molécules de la saveur et des arômes comme le menthol, le géraniol, le
caryophyllène sont formées de deux ou trois unités isopréniques. Cependant, les molécules
formées de 4, 6, 8 unités isopréniques et plus sont aussi des composés bioactifs. Les isoprénoïdes
regroupent donc à la fois des molécules de faibles poids moléculaires, volatiles et composants
principaux d’huiles essentielles et des molécules polymérisées de haut poids moléculaires
comme le caoutchouc.
Origine de l’unité isoprèneChez tous les êtres vivants, l’unité de base de la biosynthèse des terpènoïdes est l’isopentényle
diphosphate (IPP = isopentényle pyrophosphate) et son isomère le diméthylallyle-diphosphate
(DMAPP). Deux voies de biosynthèse conduisent à ces unités de base à 5 carbones.
Ce sont :
- la voie de l’acétate/mévalonate
- la voie du methyl-erythrytol-phosphate (MEP).
a) voie de l’acétate/mévalonateLa plus ancienne connue. Elle débute par la condensation de 3 unités acétyl CoA, une série de
phosphorylations puis une décarboxylation qui aboutit à l’IPP. Celui-ci est finalement isomérisé
pour donner le diméthylallyl-diphosphate (DMAPP). Elle se déroule dans le cytosol.
TERPENOÏDES
Définition
Les Terpènoïdes ou isoprénoïdes sont des composés organiques synthétisés par la condensation
d’unités isopréniques. Ils sont présents chez tous les organismes vivants, remplissent des rôles
physiologiques très divers et constituent la famille la plus large et la plus diversifiée des produits
naturels. Plusieurs molécules de la saveur et des arômes comme le menthol, le géraniol, le
caryophyllène sont formées de deux ou trois unités isopréniques. Cependant, les molécules
formées de 4, 6, 8 unités isopréniques et plus sont aussi des composés bioactifs. Les isoprénoïdes
regroupent donc à la fois des molécules de faibles poids moléculaires, volatiles et composants
principaux d’huiles essentielles et des molécules polymérisées de haut poids moléculaires
comme le caoutchouc.
Origine de l’unité isoprèneChez tous les êtres vivants, l’unité de base de la biosynthèse des terpènoïdes est l’isopentényle
diphosphate (IPP = isopentényle pyrophosphate) et son isomère le diméthylallyle-diphosphate
(DMAPP). Deux voies de biosynthèse conduisent à ces unités de base à 5 carbones.
Ce sont :
- la voie de l’acétate/mévalonate
- la voie du methyl-erythrytol-phosphate (MEP).
a) voie de l’acétate/mévalonateLa plus ancienne connue. Elle débute par la condensation de 3 unités acétyl CoA, une série de
phosphorylations puis une décarboxylation qui aboutit à l’IPP. Celui-ci est finalement isomérisé
pour donner le diméthylallyl-diphosphate (DMAPP). Elle se déroule dans le cytosol.

Figure : Biosynthèse de l’Isopentenyl phosphate (IPP) par la voie du mévalonate1: beta-cétothiolase; 2: 3-met-hydroxy-3-méthylglutaryl-CoA synthase ; 3: 3-met-hydroxy-3-methylglutaryl-CoAréductase; 4: mévalonate kinase; 5: 5-phosphomévalonate kinase; 6: 5-diphosphomévalonate décarboxylase;7 :Isopentényl diphosphate isomérase ;
b) voie du méthylérythritol phosphate (MEP)Encore nommée voie indépendante du mévalonate elle est spécifique des végétaux et se déroule
dans les plastes. Elle débute par la condensation d’une unité pyruvate (C3) avec une unité
glycéraldéhyde 3-phosphate (C3) et conduit au méthylérythritol phosphate (MEP) un composé
intermédiaire en C5. Plusieurs étapes enzymatiques conduisent ensuite à la synthèse de l’IPP.
Cette voie n’a été mise en évidence que récemment (fin des années 90), mais elle s’est
rapidement avérée la voie prépondérante pour la biosynthèse de la majeure partie des terpènes.
Ce « détail » a révélé toute son importance dans le cadre de projets visant à augmenter la
synthèse des composés terpéniques par transgenèse, par la modulation des niveaux d’expression
des enzymes de la voie du MEP.
Figure : Biosynthèse de l’Isopentenyl phosphate (IPP) par la voie du mévalonate1: beta-cétothiolase; 2: 3-met-hydroxy-3-méthylglutaryl-CoA synthase ; 3: 3-met-hydroxy-3-methylglutaryl-CoAréductase; 4: mévalonate kinase; 5: 5-phosphomévalonate kinase; 6: 5-diphosphomévalonate décarboxylase;7 :Isopentényl diphosphate isomérase ;
b) voie du méthylérythritol phosphate (MEP)Encore nommée voie indépendante du mévalonate elle est spécifique des végétaux et se déroule
dans les plastes. Elle débute par la condensation d’une unité pyruvate (C3) avec une unité
glycéraldéhyde 3-phosphate (C3) et conduit au méthylérythritol phosphate (MEP) un composé
intermédiaire en C5. Plusieurs étapes enzymatiques conduisent ensuite à la synthèse de l’IPP.
Cette voie n’a été mise en évidence que récemment (fin des années 90), mais elle s’est
rapidement avérée la voie prépondérante pour la biosynthèse de la majeure partie des terpènes.
Ce « détail » a révélé toute son importance dans le cadre de projets visant à augmenter la
synthèse des composés terpéniques par transgenèse, par la modulation des niveaux d’expression
des enzymes de la voie du MEP.
Figure : Biosynthèse de l’Isopentenyl phosphate (IPP) par la voie du mévalonate1: beta-cétothiolase; 2: 3-met-hydroxy-3-méthylglutaryl-CoA synthase ; 3: 3-met-hydroxy-3-methylglutaryl-CoAréductase; 4: mévalonate kinase; 5: 5-phosphomévalonate kinase; 6: 5-diphosphomévalonate décarboxylase;7 :Isopentényl diphosphate isomérase ;
b) voie du méthylérythritol phosphate (MEP)Encore nommée voie indépendante du mévalonate elle est spécifique des végétaux et se déroule
dans les plastes. Elle débute par la condensation d’une unité pyruvate (C3) avec une unité
glycéraldéhyde 3-phosphate (C3) et conduit au méthylérythritol phosphate (MEP) un composé
intermédiaire en C5. Plusieurs étapes enzymatiques conduisent ensuite à la synthèse de l’IPP.
Cette voie n’a été mise en évidence que récemment (fin des années 90), mais elle s’est
rapidement avérée la voie prépondérante pour la biosynthèse de la majeure partie des terpènes.
Ce « détail » a révélé toute son importance dans le cadre de projets visant à augmenter la
synthèse des composés terpéniques par transgenèse, par la modulation des niveaux d’expression
des enzymes de la voie du MEP.

Figure : Biosynthèse de l’IPP par la voie du MEP1 : 1-desoxyxylulose-5 P synthase ; 2 : 1-desoxyxylulose-5 P isoméroréductase ; 3 : 2-C-mét-D-érythritol-4 P-cytidyltransférase; 4: 4-diPcytidyl-2-C-mét-D-érythritol-kinase ; 5: 2-C-mét-D-érythritol 2,4 cyclodiPsynthase; 6 : 4-hydroxy-2-métbut-2-ènyl diP synthase;7 : IPP/DMAPP synthase; 8 : IPP/DMAPPisomérase
Biosynthèse des terpènoïdesLa synthèse des terpènoïdes comporte 4 étapes majeures. Un nombre restreint de réactions
enzymatiques est ainsi impliqué dans la formation des squelettes de ces composés à partir des
deux sous-unités en C5, l’IPP et le DMAPP. Il s’agit des réactions catalysées par les
prényltransférases, des terpènes synthases et des autres enzymes de modification.
a) les prényl transférases.Elles catalysent la synthèse des précurseurs des différents terpènoïdes par addition répétitive des
unités en C5 (IPP et DMAPP). Les produits de cette synthèse sont le diphosphate de géranyle, de
farnésyle ou de geranylgéranyle. On distingue les trans- et les cis-prényl transférases suivant la
stéréochimie de la réaction et la stéréochimie de la double liaison néoformée dans le produit de
la réaction. Ces enzymes effectuent un couplage tête-à-queue des précurseurs et permettent
d’obtenir tous les diphosphates de polyprényles avec des squelettes en C5n à partir des deux
sous-unités de base en C5. Au niveau des végétaux en particulier, la synthèse a lieu dans le
cytosol et le réticulum endoplasmique pour les sesqui- (C15), les tri- (C30) et les polyterpènes (>
C40). Les hémi- (C5), mono- (C10), di- (20) et tetraterpènes (40) sont synthétisés dans les
plastes.
Figure : Couplage des unités IPP par les prényltransférases
Figure : Biosynthèse de l’IPP par la voie du MEP1 : 1-desoxyxylulose-5 P synthase ; 2 : 1-desoxyxylulose-5 P isoméroréductase ; 3 : 2-C-mét-D-érythritol-4 P-cytidyltransférase; 4: 4-diPcytidyl-2-C-mét-D-érythritol-kinase ; 5: 2-C-mét-D-érythritol 2,4 cyclodiPsynthase; 6 : 4-hydroxy-2-métbut-2-ènyl diP synthase;7 : IPP/DMAPP synthase; 8 : IPP/DMAPPisomérase
Biosynthèse des terpènoïdesLa synthèse des terpènoïdes comporte 4 étapes majeures. Un nombre restreint de réactions
enzymatiques est ainsi impliqué dans la formation des squelettes de ces composés à partir des
deux sous-unités en C5, l’IPP et le DMAPP. Il s’agit des réactions catalysées par les
prényltransférases, des terpènes synthases et des autres enzymes de modification.
a) les prényl transférases.Elles catalysent la synthèse des précurseurs des différents terpènoïdes par addition répétitive des
unités en C5 (IPP et DMAPP). Les produits de cette synthèse sont le diphosphate de géranyle, de
farnésyle ou de geranylgéranyle. On distingue les trans- et les cis-prényl transférases suivant la
stéréochimie de la réaction et la stéréochimie de la double liaison néoformée dans le produit de
la réaction. Ces enzymes effectuent un couplage tête-à-queue des précurseurs et permettent
d’obtenir tous les diphosphates de polyprényles avec des squelettes en C5n à partir des deux
sous-unités de base en C5. Au niveau des végétaux en particulier, la synthèse a lieu dans le
cytosol et le réticulum endoplasmique pour les sesqui- (C15), les tri- (C30) et les polyterpènes (>
C40). Les hémi- (C5), mono- (C10), di- (20) et tetraterpènes (40) sont synthétisés dans les
plastes.
Figure : Couplage des unités IPP par les prényltransférases
Figure : Biosynthèse de l’IPP par la voie du MEP1 : 1-desoxyxylulose-5 P synthase ; 2 : 1-desoxyxylulose-5 P isoméroréductase ; 3 : 2-C-mét-D-érythritol-4 P-cytidyltransférase; 4: 4-diPcytidyl-2-C-mét-D-érythritol-kinase ; 5: 2-C-mét-D-érythritol 2,4 cyclodiPsynthase; 6 : 4-hydroxy-2-métbut-2-ènyl diP synthase;7 : IPP/DMAPP synthase; 8 : IPP/DMAPPisomérase
Biosynthèse des terpènoïdesLa synthèse des terpènoïdes comporte 4 étapes majeures. Un nombre restreint de réactions
enzymatiques est ainsi impliqué dans la formation des squelettes de ces composés à partir des
deux sous-unités en C5, l’IPP et le DMAPP. Il s’agit des réactions catalysées par les
prényltransférases, des terpènes synthases et des autres enzymes de modification.
a) les prényl transférases.Elles catalysent la synthèse des précurseurs des différents terpènoïdes par addition répétitive des
unités en C5 (IPP et DMAPP). Les produits de cette synthèse sont le diphosphate de géranyle, de
farnésyle ou de geranylgéranyle. On distingue les trans- et les cis-prényl transférases suivant la
stéréochimie de la réaction et la stéréochimie de la double liaison néoformée dans le produit de
la réaction. Ces enzymes effectuent un couplage tête-à-queue des précurseurs et permettent
d’obtenir tous les diphosphates de polyprényles avec des squelettes en C5n à partir des deux
sous-unités de base en C5. Au niveau des végétaux en particulier, la synthèse a lieu dans le
cytosol et le réticulum endoplasmique pour les sesqui- (C15), les tri- (C30) et les polyterpènes (>
C40). Les hémi- (C5), mono- (C10), di- (20) et tetraterpènes (40) sont synthétisés dans les
plastes.
Figure : Couplage des unités IPP par les prényltransférases

Les unités de base en C5 peuvent aussi être couplées en tête-à-tête et tête-à-milieu. Les différents
modes de couplage sont résumés ci-dessous :
Tête-à-queue Tête-à-tête Tête-à-milieu
Figure : modes de couplage des motifs isopréniques
b) Les terpènes synthases
Ces enzymes permettent de cycliser les substrats terpéniques acycliques caractérisés par des
motifs polyéniques 1,5. Elles réalisent la grande diversité des squelettes terpéniques. Les
structures de tous les isoprénoïdes peuvent ainsi être déduites à partir de concepts découlant
directement des connaissances acquises sur les mécanismes réactionnels en chimie organique sur
la chimie des carbocations : cyclisation de polyènes 1-5 induite soit par perte de diphosphate
comme pour la formation de l’alpha-pinène à partir du diphosphate de géranyle, soit par
protonation d’une double liaison ou de l’époxyde correspondant, réarrangements de Wagner-
Meerwein, et neutralisation du carbocation final par élimination d’un proton ou addition d’eau,
comme pour la biosynthèse des triterpènes. Ces mécanismes constituent la base de la règle
isoprénique biogénétique qui permet d’interpréter et de prévoir toutes les structures de
terpénoïdes.
c) Les autres modificationsCe sont les réactions d’isomérisations, d’oxydations, réductions etc. catalysées par les enzymes
correspondantes (isomérases, oxydases, réductases) qui vont achever la synthèse de certaines
molécules en leur conférant la structure et les fonctions définitives.
Les unités de base en C5 peuvent aussi être couplées en tête-à-tête et tête-à-milieu. Les différents
modes de couplage sont résumés ci-dessous :
Tête-à-queue Tête-à-tête Tête-à-milieu
Figure : modes de couplage des motifs isopréniques
b) Les terpènes synthases
Ces enzymes permettent de cycliser les substrats terpéniques acycliques caractérisés par des
motifs polyéniques 1,5. Elles réalisent la grande diversité des squelettes terpéniques. Les
structures de tous les isoprénoïdes peuvent ainsi être déduites à partir de concepts découlant
directement des connaissances acquises sur les mécanismes réactionnels en chimie organique sur
la chimie des carbocations : cyclisation de polyènes 1-5 induite soit par perte de diphosphate
comme pour la formation de l’alpha-pinène à partir du diphosphate de géranyle, soit par
protonation d’une double liaison ou de l’époxyde correspondant, réarrangements de Wagner-
Meerwein, et neutralisation du carbocation final par élimination d’un proton ou addition d’eau,
comme pour la biosynthèse des triterpènes. Ces mécanismes constituent la base de la règle
isoprénique biogénétique qui permet d’interpréter et de prévoir toutes les structures de
terpénoïdes.
c) Les autres modificationsCe sont les réactions d’isomérisations, d’oxydations, réductions etc. catalysées par les enzymes
correspondantes (isomérases, oxydases, réductases) qui vont achever la synthèse de certaines
molécules en leur conférant la structure et les fonctions définitives.
Les unités de base en C5 peuvent aussi être couplées en tête-à-tête et tête-à-milieu. Les différents
modes de couplage sont résumés ci-dessous :
Tête-à-queue Tête-à-tête Tête-à-milieu
Figure : modes de couplage des motifs isopréniques
b) Les terpènes synthases
Ces enzymes permettent de cycliser les substrats terpéniques acycliques caractérisés par des
motifs polyéniques 1,5. Elles réalisent la grande diversité des squelettes terpéniques. Les
structures de tous les isoprénoïdes peuvent ainsi être déduites à partir de concepts découlant
directement des connaissances acquises sur les mécanismes réactionnels en chimie organique sur
la chimie des carbocations : cyclisation de polyènes 1-5 induite soit par perte de diphosphate
comme pour la formation de l’alpha-pinène à partir du diphosphate de géranyle, soit par
protonation d’une double liaison ou de l’époxyde correspondant, réarrangements de Wagner-
Meerwein, et neutralisation du carbocation final par élimination d’un proton ou addition d’eau,
comme pour la biosynthèse des triterpènes. Ces mécanismes constituent la base de la règle
isoprénique biogénétique qui permet d’interpréter et de prévoir toutes les structures de
terpénoïdes.
c) Les autres modificationsCe sont les réactions d’isomérisations, d’oxydations, réductions etc. catalysées par les enzymes
correspondantes (isomérases, oxydases, réductases) qui vont achever la synthèse de certaines
molécules en leur conférant la structure et les fonctions définitives.

Figure : Synthèse des terpènoïdes
Structures et propriétés des terpènoïdes
La classification des terpènoïdes repose sur le nombre d’unités terpéniques. On distingue ainsi :
a) Monoterpènes (deux unités isoprène)Ce sont des composés à 10 carbones obtenus par la condensation de deux unités isoprènes en
géranyl PP suivie de l’action des terpènes cyclases sur celui-ci. Ils sont souvent volatiles,
aromatiques (sens olfactif), biologiquement actifs (bactériostatiques, signalisation plantes-
insectes) et largement présents dans les résines et les huiles essentielles (exemples du pinène
constituant majeur de l’essence de térébenthine et du menthol). On distingue des monoterpènes
linéaires, monocycliques et bicycliques. Exemples : L'ocimène du basilic, le myrcène du laurier,
le géraniol des géranium, le citronellol des roses, le limonène des citrons, le menthol des
menthes, le thymol du thym, le pinène du pin, le camphre du Cinnamum camphora (camphrier).
Ocimène Myrcène Linalol Géraniol Menthol
Figure : Synthèse des terpènoïdes
Structures et propriétés des terpènoïdes
La classification des terpènoïdes repose sur le nombre d’unités terpéniques. On distingue ainsi :
a) Monoterpènes (deux unités isoprène)Ce sont des composés à 10 carbones obtenus par la condensation de deux unités isoprènes en
géranyl PP suivie de l’action des terpènes cyclases sur celui-ci. Ils sont souvent volatiles,
aromatiques (sens olfactif), biologiquement actifs (bactériostatiques, signalisation plantes-
insectes) et largement présents dans les résines et les huiles essentielles (exemples du pinène
constituant majeur de l’essence de térébenthine et du menthol). On distingue des monoterpènes
linéaires, monocycliques et bicycliques. Exemples : L'ocimène du basilic, le myrcène du laurier,
le géraniol des géranium, le citronellol des roses, le limonène des citrons, le menthol des
menthes, le thymol du thym, le pinène du pin, le camphre du Cinnamum camphora (camphrier).
Ocimène Myrcène Linalol Géraniol Menthol
Figure : Synthèse des terpènoïdes
Structures et propriétés des terpènoïdes
La classification des terpènoïdes repose sur le nombre d’unités terpéniques. On distingue ainsi :
a) Monoterpènes (deux unités isoprène)Ce sont des composés à 10 carbones obtenus par la condensation de deux unités isoprènes en
géranyl PP suivie de l’action des terpènes cyclases sur celui-ci. Ils sont souvent volatiles,
aromatiques (sens olfactif), biologiquement actifs (bactériostatiques, signalisation plantes-
insectes) et largement présents dans les résines et les huiles essentielles (exemples du pinène
constituant majeur de l’essence de térébenthine et du menthol). On distingue des monoterpènes
linéaires, monocycliques et bicycliques. Exemples : L'ocimène du basilic, le myrcène du laurier,
le géraniol des géranium, le citronellol des roses, le limonène des citrons, le menthol des
menthes, le thymol du thym, le pinène du pin, le camphre du Cinnamum camphora (camphrier).
Ocimène Myrcène Linalol Géraniol Menthol

Limonène Thymol (thym)-Pinène (pin) Camphre 1,8-Cinéol
Figure : structures de monoterpènes
b) Sesquiterpènes (trois unités isoprène)Composés à 15 carbones assez universellement représentés chez les végétaux. On distingue
également des sesquiterpènes linéaires, monocycliques et polycycliques. Ils dérivent tous du
Farnesyl PP. Les composés cycliques résultent des réactions de cyclisation catalysées par les
sesquiterpènes synthases. Comme les monoterpènes, ils sont aussi volatiles et aromatiques.
(Exemples : le farnésol, un sesquiterpène linéaire de nombreuses huiles essentielles,
abondamment utilisé en parfumerie ; Le caryophyllène, un sesquiterpène bicyclique en partie
responsable du piquant du poivre).
Farnésol Caryophyllène Bêta-cadinène
c) Diterpènes (quatre unités isoprène)
Composés terpéniques à 20 carbones. Ils dérivent du géranylgéranyl PP. On retrouve parmi les
dérivés de diterpènes le collophane ou acide abiétique, le taxadiène et le phytol. Certains
diterpènes ont des propriétés médicinales importantes. On peut citer le guanacastapène, un
diterpène aux propriétés antibiotiques.
Taxadiène
Acide abiètique
Limonène Thymol (thym)-Pinène (pin) Camphre 1,8-Cinéol
Figure : structures de monoterpènes
b) Sesquiterpènes (trois unités isoprène)Composés à 15 carbones assez universellement représentés chez les végétaux. On distingue
également des sesquiterpènes linéaires, monocycliques et polycycliques. Ils dérivent tous du
Farnesyl PP. Les composés cycliques résultent des réactions de cyclisation catalysées par les
sesquiterpènes synthases. Comme les monoterpènes, ils sont aussi volatiles et aromatiques.
(Exemples : le farnésol, un sesquiterpène linéaire de nombreuses huiles essentielles,
abondamment utilisé en parfumerie ; Le caryophyllène, un sesquiterpène bicyclique en partie
responsable du piquant du poivre).
Farnésol Caryophyllène Bêta-cadinène
c) Diterpènes (quatre unités isoprène)
Composés terpéniques à 20 carbones. Ils dérivent du géranylgéranyl PP. On retrouve parmi les
dérivés de diterpènes le collophane ou acide abiétique, le taxadiène et le phytol. Certains
diterpènes ont des propriétés médicinales importantes. On peut citer le guanacastapène, un
diterpène aux propriétés antibiotiques.
Taxadiène
Acide abiètique
Limonène Thymol (thym)-Pinène (pin) Camphre 1,8-Cinéol
Figure : structures de monoterpènes
b) Sesquiterpènes (trois unités isoprène)Composés à 15 carbones assez universellement représentés chez les végétaux. On distingue
également des sesquiterpènes linéaires, monocycliques et polycycliques. Ils dérivent tous du
Farnesyl PP. Les composés cycliques résultent des réactions de cyclisation catalysées par les
sesquiterpènes synthases. Comme les monoterpènes, ils sont aussi volatiles et aromatiques.
(Exemples : le farnésol, un sesquiterpène linéaire de nombreuses huiles essentielles,
abondamment utilisé en parfumerie ; Le caryophyllène, un sesquiterpène bicyclique en partie
responsable du piquant du poivre).
Farnésol Caryophyllène Bêta-cadinène
c) Diterpènes (quatre unités isoprène)
Composés terpéniques à 20 carbones. Ils dérivent du géranylgéranyl PP. On retrouve parmi les
dérivés de diterpènes le collophane ou acide abiétique, le taxadiène et le phytol. Certains
diterpènes ont des propriétés médicinales importantes. On peut citer le guanacastapène, un
diterpène aux propriétés antibiotiques.
Taxadiène
Acide abiètique

Phytol
Figure : Structure de diterpènes
d) Triterpènes
Cette famille regroupe des composés dérivés d’une unité à 30 carbones le squalène, qui est lui-
même issu de la condensation de deux unités FPP. En fonction du nombre de cycles on distingue
les monoterpènes pentacycliques des monoterpènes stéroïdiens (tetracycliques). Bien que les
composés stéroïdiens soient largement représentés dans le monde animal, de nombreux
phytostérols sont spécifiques des végétaux. Les saponosides et les cardénolides sont des
hétérosides formés à partir d’un triterpène qui en constitue la génine (= fraction aglycone) et de
résidus glucidiques.
Castastérone Squalène
Figure : Structure de triterpènes
e) Tétraterpènes (caroténoïdes)
Ce sont des composés à 40C qui résultent de la condensation de deux unités diterpènes suivie de
diverses étapes de modifications. Parmi les tétraterpènes, on peut citer le bêta-carotène dont la
coupure donne d'abord le rétinal, puis sa forme réduite ou vitamine A. Le lycopène, contenu dans
la tomate est un produit très voisin des carotènes.
(A)
Phytol
Figure : Structure de diterpènes
d) Triterpènes
Cette famille regroupe des composés dérivés d’une unité à 30 carbones le squalène, qui est lui-
même issu de la condensation de deux unités FPP. En fonction du nombre de cycles on distingue
les monoterpènes pentacycliques des monoterpènes stéroïdiens (tetracycliques). Bien que les
composés stéroïdiens soient largement représentés dans le monde animal, de nombreux
phytostérols sont spécifiques des végétaux. Les saponosides et les cardénolides sont des
hétérosides formés à partir d’un triterpène qui en constitue la génine (= fraction aglycone) et de
résidus glucidiques.
Castastérone Squalène
Figure : Structure de triterpènes
e) Tétraterpènes (caroténoïdes)
Ce sont des composés à 40C qui résultent de la condensation de deux unités diterpènes suivie de
diverses étapes de modifications. Parmi les tétraterpènes, on peut citer le bêta-carotène dont la
coupure donne d'abord le rétinal, puis sa forme réduite ou vitamine A. Le lycopène, contenu dans
la tomate est un produit très voisin des carotènes.
(A)
Phytol
Figure : Structure de diterpènes
d) Triterpènes
Cette famille regroupe des composés dérivés d’une unité à 30 carbones le squalène, qui est lui-
même issu de la condensation de deux unités FPP. En fonction du nombre de cycles on distingue
les monoterpènes pentacycliques des monoterpènes stéroïdiens (tetracycliques). Bien que les
composés stéroïdiens soient largement représentés dans le monde animal, de nombreux
phytostérols sont spécifiques des végétaux. Les saponosides et les cardénolides sont des
hétérosides formés à partir d’un triterpène qui en constitue la génine (= fraction aglycone) et de
résidus glucidiques.
Castastérone Squalène
Figure : Structure de triterpènes
e) Tétraterpènes (caroténoïdes)
Ce sont des composés à 40C qui résultent de la condensation de deux unités diterpènes suivie de
diverses étapes de modifications. Parmi les tétraterpènes, on peut citer le bêta-carotène dont la
coupure donne d'abord le rétinal, puis sa forme réduite ou vitamine A. Le lycopène, contenu dans
la tomate est un produit très voisin des carotènes.
(A)

(B)Figure : Structures des tétraterpènes (A : beta-carotène ; B : lycopène)
f) Polyterpènes :Ce sont des polymères au-delà de 40 C ou 50 C comme le caoutchouc naturel. Le caoutchouc
naturel est un dérivé du latex (unités d’isoprène). Il provient de la coagulation du latex de
plusieurs plantes, principalement de l’hévéa (Hevea brasiliensis, famille des Euphorbiacées).
Toutes ses doubles liaisons sont Z (chaîne prioritaitre) et chaque molécule comporte de 1000 à
5000 motifs isoprèniques. Son isomère dont toutes les liaisons sont E est la gutta-percha, qui est
une matière dure et cassante.
Caoutchouc naturel gutta-percha
Figure : Structures de polyterpènes
(B)Figure : Structures des tétraterpènes (A : beta-carotène ; B : lycopène)
f) Polyterpènes :Ce sont des polymères au-delà de 40 C ou 50 C comme le caoutchouc naturel. Le caoutchouc
naturel est un dérivé du latex (unités d’isoprène). Il provient de la coagulation du latex de
plusieurs plantes, principalement de l’hévéa (Hevea brasiliensis, famille des Euphorbiacées).
Toutes ses doubles liaisons sont Z (chaîne prioritaitre) et chaque molécule comporte de 1000 à
5000 motifs isoprèniques. Son isomère dont toutes les liaisons sont E est la gutta-percha, qui est
une matière dure et cassante.
Caoutchouc naturel gutta-percha
Figure : Structures de polyterpènes
(B)Figure : Structures des tétraterpènes (A : beta-carotène ; B : lycopène)
f) Polyterpènes :Ce sont des polymères au-delà de 40 C ou 50 C comme le caoutchouc naturel. Le caoutchouc
naturel est un dérivé du latex (unités d’isoprène). Il provient de la coagulation du latex de
plusieurs plantes, principalement de l’hévéa (Hevea brasiliensis, famille des Euphorbiacées).
Toutes ses doubles liaisons sont Z (chaîne prioritaitre) et chaque molécule comporte de 1000 à
5000 motifs isoprèniques. Son isomère dont toutes les liaisons sont E est la gutta-percha, qui est
une matière dure et cassante.
Caoutchouc naturel gutta-percha
Figure : Structures de polyterpènes

LES POLYPHENOLS
Les molécules phénoliques sont des composés qui contiennent un groupe phénol (anneau aromatique
avec un groupe hydroxyle). Ils peuvent avoir un ou plusieurs autres substituants différents. Cette vaste
famille regroupe des molécules non azotées présentant des cycles aromatiques, le plus souvent solubles
dans l’eau et présentes sous forme de glycoconjugués. Elle comprend des petits composés
biologiquement actifs, volatils, des composés présents dans certaines huiles essentielles (en association
avec des terpènes à faible poids moléculaire) mais aussi des composés anti-nutritionnels comme les
tannins, et la lignine, un polymère de haut poids moléculaire formé de dérivés de l’acide cinnamique.
Comme les autres métabolites secondaires, chaque espèce produit un groupe spécifique de molécules.
Ils ont beaucoup de fonctions dans différentes plantes.
Phenylethylalcool(orchidées)
Vanilline(vanille)
Gingérols(Gingembre)
Pipérine (piment)
Figure : structures et source de quelques composés phénoliques
1. LES VOIES DE BIOSYNTHESE
Les principales voies de biosynthèse des composés phénoliques sont la voie des phénylpropanoïdes, la
voie du shikimate et une dernière voie non spécifique qui regroupe toutes les autres combinaisons.
a) la voie des phénylpropanoïdes
La majeure partie des composés aromatiques est constituée par la famille des phenylpropanoïdes, qui
dérivent eux-mêmes de la phénylalanine et de la tyrosine. La désamination de ces acides aminés par une
enzyme clé qui est la phénylalanine ammonium lyase (PAL), conduit à l’acide cinnamique et à l’acide
para-coumarique. L’acide cinnamique est un composé en C6-C3 (un noyau phényl + un acide
propénoïque) qui constitue un carrefour métabolique intermédiaire avec le métabolisme primaire.
L’acide para-coumarique est obtenu par action de la PAL sur la tyrosine suivi par l’action de la cinnamate
hydroxylase sur l’acide cinnamique. Il est précurseur de centaines de métabolites secondaires (lignines,
lignanes, subérines, flavonoïdes, coumarines).
LES POLYPHENOLS
Les molécules phénoliques sont des composés qui contiennent un groupe phénol (anneau aromatique
avec un groupe hydroxyle). Ils peuvent avoir un ou plusieurs autres substituants différents. Cette vaste
famille regroupe des molécules non azotées présentant des cycles aromatiques, le plus souvent solubles
dans l’eau et présentes sous forme de glycoconjugués. Elle comprend des petits composés
biologiquement actifs, volatils, des composés présents dans certaines huiles essentielles (en association
avec des terpènes à faible poids moléculaire) mais aussi des composés anti-nutritionnels comme les
tannins, et la lignine, un polymère de haut poids moléculaire formé de dérivés de l’acide cinnamique.
Comme les autres métabolites secondaires, chaque espèce produit un groupe spécifique de molécules.
Ils ont beaucoup de fonctions dans différentes plantes.
Phenylethylalcool(orchidées)
Vanilline(vanille)
Gingérols(Gingembre)
Pipérine (piment)
Figure : structures et source de quelques composés phénoliques
1. LES VOIES DE BIOSYNTHESE
Les principales voies de biosynthèse des composés phénoliques sont la voie des phénylpropanoïdes, la
voie du shikimate et une dernière voie non spécifique qui regroupe toutes les autres combinaisons.
a) la voie des phénylpropanoïdes
La majeure partie des composés aromatiques est constituée par la famille des phenylpropanoïdes, qui
dérivent eux-mêmes de la phénylalanine et de la tyrosine. La désamination de ces acides aminés par une
enzyme clé qui est la phénylalanine ammonium lyase (PAL), conduit à l’acide cinnamique et à l’acide
para-coumarique. L’acide cinnamique est un composé en C6-C3 (un noyau phényl + un acide
propénoïque) qui constitue un carrefour métabolique intermédiaire avec le métabolisme primaire.
L’acide para-coumarique est obtenu par action de la PAL sur la tyrosine suivi par l’action de la cinnamate
hydroxylase sur l’acide cinnamique. Il est précurseur de centaines de métabolites secondaires (lignines,
lignanes, subérines, flavonoïdes, coumarines).
LES POLYPHENOLS
Les molécules phénoliques sont des composés qui contiennent un groupe phénol (anneau aromatique
avec un groupe hydroxyle). Ils peuvent avoir un ou plusieurs autres substituants différents. Cette vaste
famille regroupe des molécules non azotées présentant des cycles aromatiques, le plus souvent solubles
dans l’eau et présentes sous forme de glycoconjugués. Elle comprend des petits composés
biologiquement actifs, volatils, des composés présents dans certaines huiles essentielles (en association
avec des terpènes à faible poids moléculaire) mais aussi des composés anti-nutritionnels comme les
tannins, et la lignine, un polymère de haut poids moléculaire formé de dérivés de l’acide cinnamique.
Comme les autres métabolites secondaires, chaque espèce produit un groupe spécifique de molécules.
Ils ont beaucoup de fonctions dans différentes plantes.
Phenylethylalcool(orchidées)
Vanilline(vanille)
Gingérols(Gingembre)
Pipérine (piment)
Figure : structures et source de quelques composés phénoliques
1. LES VOIES DE BIOSYNTHESE
Les principales voies de biosynthèse des composés phénoliques sont la voie des phénylpropanoïdes, la
voie du shikimate et une dernière voie non spécifique qui regroupe toutes les autres combinaisons.
a) la voie des phénylpropanoïdes
La majeure partie des composés aromatiques est constituée par la famille des phenylpropanoïdes, qui
dérivent eux-mêmes de la phénylalanine et de la tyrosine. La désamination de ces acides aminés par une
enzyme clé qui est la phénylalanine ammonium lyase (PAL), conduit à l’acide cinnamique et à l’acide
para-coumarique. L’acide cinnamique est un composé en C6-C3 (un noyau phényl + un acide
propénoïque) qui constitue un carrefour métabolique intermédiaire avec le métabolisme primaire.
L’acide para-coumarique est obtenu par action de la PAL sur la tyrosine suivi par l’action de la cinnamate
hydroxylase sur l’acide cinnamique. Il est précurseur de centaines de métabolites secondaires (lignines,
lignanes, subérines, flavonoïdes, coumarines).

(PAL)
(A) (B)
Figure : Structures de la Phe (A) et son dérivé l’acide cinnamique (B)
b) la voie du shikimate
En plus des phénylpropanoïdes, les composés aromatiques regroupent également des composés issus
d’étapes de biosynthèse situées en amont de la synthèse des acides aminés aromatiques par la voie du
shikimate. Cette voie du shikimate est très spécifique des végétaux et conduit à la synthèse des trois
acides aminés essentiels suivants : tryptophane, phénylalanine et tyrosine.
Figure : Biosynthèse du shikimate(1) : DAHP synthase, (2) :DHQ synthase, (3) : 3-DHQ dehydratase, (4) : shikimate deshydrogénase
Shikimate shikimate-3P 5-énoylpyruvylshikimate-3P

chorismate préphenate
4-hydroxyphenylpyruvate tyrosine
Figure : Biosynthèse de la tyrosine par la voie du shikimate1 : shikimate kinase, 2 : EPSP synthase, 3 : chorismate synthase, 4: rearrangement de Claisen, 5:deshydrogénase, 6 : transaminase.
c) Autres voies de synthèse
Certains composés aromatiques sont des composés mixtes formés par ajout de résidus terpènes
(exemple : formation du noyau furane des furocoumarines) ou par condensation de plusieurs unités C2
à partir du malonyl-CoA (cas du cannabinol). Par exemple, l’initiation de la voie de synthèse des
flavonoïdes à partir d’un phénylpropane et de 3 unités malonyl-CoA, est catalysée par la chalcone
synthase une enzyme de la famille des polykétide synthases.
Figure : structure du 3,4 cis-tétrahydocannabinol
2. DERIVES DE L’ACIDE CINNAMIQUE
a) Monolignols et lignification
Trois alcools dérivant de l’acide cinnamique ont un statut particulier. Ce sont : l’alcool coumarylique,
l’alcool coniférylique et l’alcool sinapylique. La polymérisation de différentes proportions de ces
composés via des liaisons C-O et C-C aboutit à la formation des lignines, des polymères à très haut poids
chorismate préphenate
4-hydroxyphenylpyruvate tyrosine
Figure : Biosynthèse de la tyrosine par la voie du shikimate1 : shikimate kinase, 2 : EPSP synthase, 3 : chorismate synthase, 4: rearrangement de Claisen, 5:deshydrogénase, 6 : transaminase.
c) Autres voies de synthèse
Certains composés aromatiques sont des composés mixtes formés par ajout de résidus terpènes
(exemple : formation du noyau furane des furocoumarines) ou par condensation de plusieurs unités C2
à partir du malonyl-CoA (cas du cannabinol). Par exemple, l’initiation de la voie de synthèse des
flavonoïdes à partir d’un phénylpropane et de 3 unités malonyl-CoA, est catalysée par la chalcone
synthase une enzyme de la famille des polykétide synthases.
Figure : structure du 3,4 cis-tétrahydocannabinol
2. DERIVES DE L’ACIDE CINNAMIQUE
a) Monolignols et lignification
Trois alcools dérivant de l’acide cinnamique ont un statut particulier. Ce sont : l’alcool coumarylique,
l’alcool coniférylique et l’alcool sinapylique. La polymérisation de différentes proportions de ces
composés via des liaisons C-O et C-C aboutit à la formation des lignines, des polymères à très haut poids
chorismate préphenate
4-hydroxyphenylpyruvate tyrosine
Figure : Biosynthèse de la tyrosine par la voie du shikimate1 : shikimate kinase, 2 : EPSP synthase, 3 : chorismate synthase, 4: rearrangement de Claisen, 5:deshydrogénase, 6 : transaminase.
c) Autres voies de synthèse
Certains composés aromatiques sont des composés mixtes formés par ajout de résidus terpènes
(exemple : formation du noyau furane des furocoumarines) ou par condensation de plusieurs unités C2
à partir du malonyl-CoA (cas du cannabinol). Par exemple, l’initiation de la voie de synthèse des
flavonoïdes à partir d’un phénylpropane et de 3 unités malonyl-CoA, est catalysée par la chalcone
synthase une enzyme de la famille des polykétide synthases.
Figure : structure du 3,4 cis-tétrahydocannabinol
2. DERIVES DE L’ACIDE CINNAMIQUE
a) Monolignols et lignification
Trois alcools dérivant de l’acide cinnamique ont un statut particulier. Ce sont : l’alcool coumarylique,
l’alcool coniférylique et l’alcool sinapylique. La polymérisation de différentes proportions de ces
composés via des liaisons C-O et C-C aboutit à la formation des lignines, des polymères à très haut poids

moléculaire qui, au-delà du renforcement des parois végétales, limitent fortement la digestibilité des
tissus lignifiés. La composition des lignines est différente entre les gymnospermes (100% coniférylique),
les dicotylédones (coniferylique + sinapylique) et les monocotylédones (coniferylique + sinapylique +
coumarylique). Ces différences ont un impact direct sur des propriétés du polymère telles que la
résistance vis-à-vis de procédés utilisés en papeterie pour purifier la cellulose. Contrairement à la
cellulose, la lignine n’est pas digérée par les polygastriques, elle constitue un facteur antinutritionnel par
exemple dans l’ensilage de maïs.
(A) (B) (C)
Figure : Structures des principaux monolignols (A : alcool coumarylique ; B : alcool conférylique ; C :alcool sinapylique
b). Coumarines
Elles proviennent de la cyclisation du résidu C3 des dérivés du cinnamate. Les dérivés du noyau de base
qu’est la coumarine sont appelés coumarines. Les coumarines sont des composés souvent synthétisés
en réponse à des attaques pathogènes. Plusieurs coumarines ont des propriétés bactériostatiques et
représentent donc des phytoalexines chez un certain nombre de plantes (ex : la scopolétine qui
s’accumule chez le tabac au cours de la réaction hypersensible). La coumarine est une molécule
aromatique (au sens olfactif). Elle est présente sous forme glycoconjuguée chez certaines graminées
(ex : la flouve odorante), mais c’est lorsque les tissus sont endommagés par la coupe que les
glycosidases libèrent la coumarine libre à l’origine de l’odeur de foin coupé.
Figure : Formation de la structure de base des coumarines
c). Flavonoïdes
Les flavonoïdes sont des dérivés phénylpropanoïdes solubles dans l’eau, souvent incolores ou jaunes
(sauf exceptions dont les anthocyanes). Ces composés sont des dérivés de la naringénine-chalcone, elle-
moléculaire qui, au-delà du renforcement des parois végétales, limitent fortement la digestibilité des
tissus lignifiés. La composition des lignines est différente entre les gymnospermes (100% coniférylique),
les dicotylédones (coniferylique + sinapylique) et les monocotylédones (coniferylique + sinapylique +
coumarylique). Ces différences ont un impact direct sur des propriétés du polymère telles que la
résistance vis-à-vis de procédés utilisés en papeterie pour purifier la cellulose. Contrairement à la
cellulose, la lignine n’est pas digérée par les polygastriques, elle constitue un facteur antinutritionnel par
exemple dans l’ensilage de maïs.
(A) (B) (C)
Figure : Structures des principaux monolignols (A : alcool coumarylique ; B : alcool conférylique ; C :alcool sinapylique
b). Coumarines
Elles proviennent de la cyclisation du résidu C3 des dérivés du cinnamate. Les dérivés du noyau de base
qu’est la coumarine sont appelés coumarines. Les coumarines sont des composés souvent synthétisés
en réponse à des attaques pathogènes. Plusieurs coumarines ont des propriétés bactériostatiques et
représentent donc des phytoalexines chez un certain nombre de plantes (ex : la scopolétine qui
s’accumule chez le tabac au cours de la réaction hypersensible). La coumarine est une molécule
aromatique (au sens olfactif). Elle est présente sous forme glycoconjuguée chez certaines graminées
(ex : la flouve odorante), mais c’est lorsque les tissus sont endommagés par la coupe que les
glycosidases libèrent la coumarine libre à l’origine de l’odeur de foin coupé.
Figure : Formation de la structure de base des coumarines
c). Flavonoïdes
Les flavonoïdes sont des dérivés phénylpropanoïdes solubles dans l’eau, souvent incolores ou jaunes
(sauf exceptions dont les anthocyanes). Ces composés sont des dérivés de la naringénine-chalcone, elle-
moléculaire qui, au-delà du renforcement des parois végétales, limitent fortement la digestibilité des
tissus lignifiés. La composition des lignines est différente entre les gymnospermes (100% coniférylique),
les dicotylédones (coniferylique + sinapylique) et les monocotylédones (coniferylique + sinapylique +
coumarylique). Ces différences ont un impact direct sur des propriétés du polymère telles que la
résistance vis-à-vis de procédés utilisés en papeterie pour purifier la cellulose. Contrairement à la
cellulose, la lignine n’est pas digérée par les polygastriques, elle constitue un facteur antinutritionnel par
exemple dans l’ensilage de maïs.
(A) (B) (C)
Figure : Structures des principaux monolignols (A : alcool coumarylique ; B : alcool conférylique ; C :alcool sinapylique
b). Coumarines
Elles proviennent de la cyclisation du résidu C3 des dérivés du cinnamate. Les dérivés du noyau de base
qu’est la coumarine sont appelés coumarines. Les coumarines sont des composés souvent synthétisés
en réponse à des attaques pathogènes. Plusieurs coumarines ont des propriétés bactériostatiques et
représentent donc des phytoalexines chez un certain nombre de plantes (ex : la scopolétine qui
s’accumule chez le tabac au cours de la réaction hypersensible). La coumarine est une molécule
aromatique (au sens olfactif). Elle est présente sous forme glycoconjuguée chez certaines graminées
(ex : la flouve odorante), mais c’est lorsque les tissus sont endommagés par la coupe que les
glycosidases libèrent la coumarine libre à l’origine de l’odeur de foin coupé.
Figure : Formation de la structure de base des coumarines
c). Flavonoïdes
Les flavonoïdes sont des dérivés phénylpropanoïdes solubles dans l’eau, souvent incolores ou jaunes
(sauf exceptions dont les anthocyanes). Ces composés sont des dérivés de la naringénine-chalcone, elle-

même issue de la condensation de trois résidus malonyl-CoA avec une molécule d’acide cinnamique. La
structure de base comporte deux cycles aromatiques à 6 carbones joints par un hétérocycle à oxygène.
Figure : synthèse de la structure de base des flavonoïdes
Les flavonoïdes constituent en eux même une famille de composés extrêmement vaste dont la variété
est essentiellement liée au degré d’hydroxylation/methylation/glycosylation de chacun des trois cycles
des molécules de base. Ils jouent des rôles physiologiques importants (interactions
Légumineuses/Rhizobium, filtres UV…). Les principales catégories de flavonoïdes sont définies par :
- la présence ou l’absence d’une double liaison entre les carbones 2 et 3 du cycle C, qui déterminent la
planéité de la molécule. Les flavones, flavonols et dérivés présentent une double liaison et sont des
molécules planes, contrairement aux flavanes, flavanones et dérivés
- la présence de fonctions cétones, alcools et méthoxy.
* Les anthocyanes
Les anthocyanes sont des composés colorés (orange, pourpre à bleu) contrairement aux autres
flavonoïdes qui absorbent en général essentiellement dans le spectre ultra violet. Ils sont largement
impliqués dans la coloration des pétales mais on les retrouve également dans de nombreux tissus
(exemple du chou rouge). Leur synthèse dans les organes foliaires est souvent activée par des stress
(froid, carences, sénescence…). Ces composés sont beaucoup utilisés comme colorants alimentaires et
présentent des propriétés anti-oxydantes.
Figure : Structure de la cyanidine (couleur des roses)
même issue de la condensation de trois résidus malonyl-CoA avec une molécule d’acide cinnamique. La
structure de base comporte deux cycles aromatiques à 6 carbones joints par un hétérocycle à oxygène.
Figure : synthèse de la structure de base des flavonoïdes
Les flavonoïdes constituent en eux même une famille de composés extrêmement vaste dont la variété
est essentiellement liée au degré d’hydroxylation/methylation/glycosylation de chacun des trois cycles
des molécules de base. Ils jouent des rôles physiologiques importants (interactions
Légumineuses/Rhizobium, filtres UV…). Les principales catégories de flavonoïdes sont définies par :
- la présence ou l’absence d’une double liaison entre les carbones 2 et 3 du cycle C, qui déterminent la
planéité de la molécule. Les flavones, flavonols et dérivés présentent une double liaison et sont des
molécules planes, contrairement aux flavanes, flavanones et dérivés
- la présence de fonctions cétones, alcools et méthoxy.
* Les anthocyanes
Les anthocyanes sont des composés colorés (orange, pourpre à bleu) contrairement aux autres
flavonoïdes qui absorbent en général essentiellement dans le spectre ultra violet. Ils sont largement
impliqués dans la coloration des pétales mais on les retrouve également dans de nombreux tissus
(exemple du chou rouge). Leur synthèse dans les organes foliaires est souvent activée par des stress
(froid, carences, sénescence…). Ces composés sont beaucoup utilisés comme colorants alimentaires et
présentent des propriétés anti-oxydantes.
Figure : Structure de la cyanidine (couleur des roses)
même issue de la condensation de trois résidus malonyl-CoA avec une molécule d’acide cinnamique. La
structure de base comporte deux cycles aromatiques à 6 carbones joints par un hétérocycle à oxygène.
Figure : synthèse de la structure de base des flavonoïdes
Les flavonoïdes constituent en eux même une famille de composés extrêmement vaste dont la variété
est essentiellement liée au degré d’hydroxylation/methylation/glycosylation de chacun des trois cycles
des molécules de base. Ils jouent des rôles physiologiques importants (interactions
Légumineuses/Rhizobium, filtres UV…). Les principales catégories de flavonoïdes sont définies par :
- la présence ou l’absence d’une double liaison entre les carbones 2 et 3 du cycle C, qui déterminent la
planéité de la molécule. Les flavones, flavonols et dérivés présentent une double liaison et sont des
molécules planes, contrairement aux flavanes, flavanones et dérivés
- la présence de fonctions cétones, alcools et méthoxy.
* Les anthocyanes
Les anthocyanes sont des composés colorés (orange, pourpre à bleu) contrairement aux autres
flavonoïdes qui absorbent en général essentiellement dans le spectre ultra violet. Ils sont largement
impliqués dans la coloration des pétales mais on les retrouve également dans de nombreux tissus
(exemple du chou rouge). Leur synthèse dans les organes foliaires est souvent activée par des stress
(froid, carences, sénescence…). Ces composés sont beaucoup utilisés comme colorants alimentaires et
présentent des propriétés anti-oxydantes.
Figure : Structure de la cyanidine (couleur des roses)

* Les isoflavones
Ce sont des composés pour lesquels le cycle B est relié au carbone 3 du cycle C. Ils se retrouvent en
particulier chez les légumineuses (exemple le la daïdzéine du soja dont les propriétés phytoestrogènes
sont particulièrement étudiées). Certains dérivés d’isoflavones sont des bactériostatiques et sont
spécifiquement induits lors d’infections par des organismes phytopathogènes. Exemples de nombreuses
phytoalexines des légumineuses (phaseolline (haricot), glyceolline (soja)).
Figure : structure de la phaseolline du haricot et de la daïdzéine du soja
* Les flavan-3-ols
Composés phagodéterrants, ils ont la propriété de faire précipiter les protéines et ils sont impliqués
dans la défense contre les pathogènes et les phytophages. Ces propriétés font qu’on assimile parfois aux
tanins ces molécules de faible poids moléculaire.
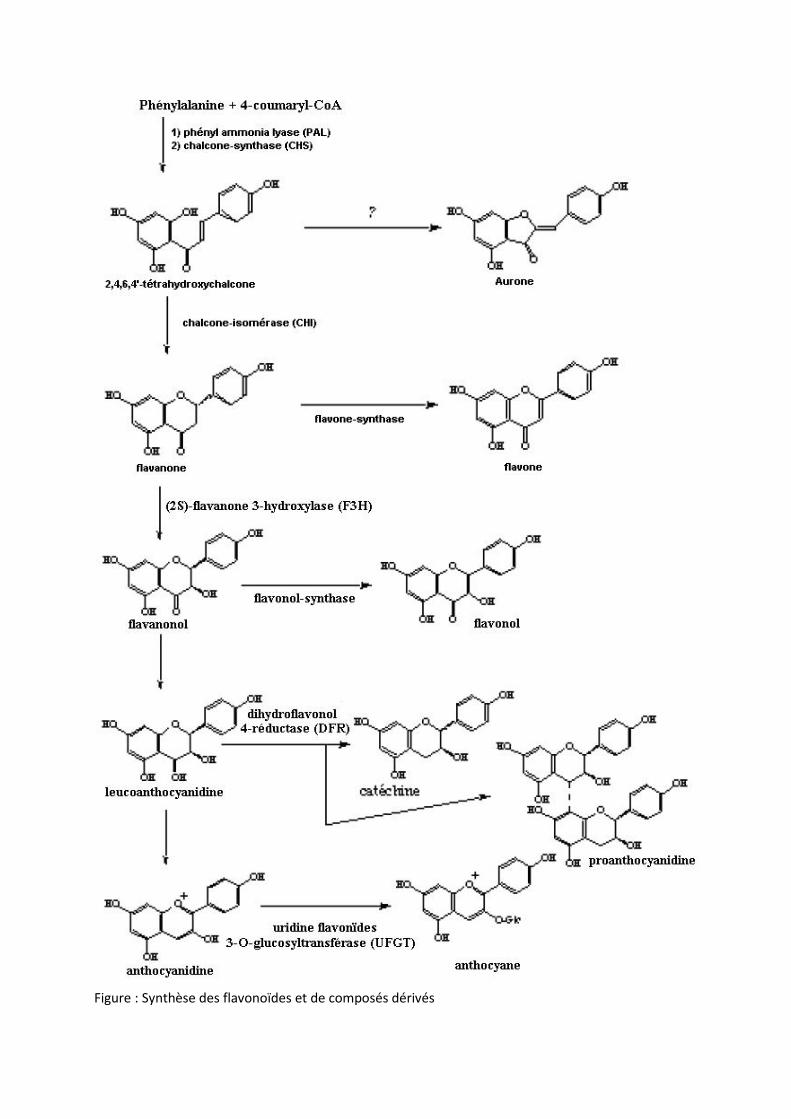
Figure : Synthèse des flavonoïdes et de composés dérivés

* Les Tanins non-hydrolysables = Proanthocyanidines
Ce sont des polymères de 2 à 7 résidus flavan-3-ols reliés (catéchine, épicatéchine ; on parle également
de tanins catéchiques) par des liaisons C-C non-hydrolyables. sont des composés phagodéterrants
capables de faire précipiter les protéines (en particulier celles de la salive ; astringence conférée par les
tanins par exemple dans le vin). Cette propriété provoque une baisse d’appétence chez les animaux et
surtout une diminution de la digestibilité des protéines (les oiseaux aiment le sorgho blanc mais
détestent le sorgho rouge à cause des tanins qui lui confèrent cette couleur. La précipitation par les
tanins des enzymes secrétées par les champignons phytopathogènes nécrotrophes est une propriété qui
peut contribuer à la résistance chez certaines plantes. La synthèse des tanins est dans certains cas
induite par la perception de stress. L’oxydation des tanins dans certaines conditions, permet de libérer
des monomères colorés d’anthocyanes (d’où l’appellation synonyme assez courante de
proanthocyanidines).
Figure : Structure des proanthocyanidines du sorgho rouge (couleur rouge du sorgho avec n=1-30)
3. Tanins hydrolysables
Ces composés phénoliques présentent des propriétés semblables à celles des proanthocyanidines mais
ce ne sont pas des flavonoïdes. Il s’agit d’esters d’acide gallique ou d’acide élagique (dérivé du
shikimate) et de glucose. On retrouve des gallotanins accumulés dans certaines galles de végétaux qui
étaient utilisées pour la fabrication d’encres. Les tanins libérés dans le vin par les tonneaux de chênes
sont des ellagitanins, contrairement à ceux apportés par la peau des baies de raisin qui sont des tanins
non-hydrolysables.
* Les Tanins non-hydrolysables = Proanthocyanidines
Ce sont des polymères de 2 à 7 résidus flavan-3-ols reliés (catéchine, épicatéchine ; on parle également
de tanins catéchiques) par des liaisons C-C non-hydrolyables. sont des composés phagodéterrants
capables de faire précipiter les protéines (en particulier celles de la salive ; astringence conférée par les
tanins par exemple dans le vin). Cette propriété provoque une baisse d’appétence chez les animaux et
surtout une diminution de la digestibilité des protéines (les oiseaux aiment le sorgho blanc mais
détestent le sorgho rouge à cause des tanins qui lui confèrent cette couleur. La précipitation par les
tanins des enzymes secrétées par les champignons phytopathogènes nécrotrophes est une propriété qui
peut contribuer à la résistance chez certaines plantes. La synthèse des tanins est dans certains cas
induite par la perception de stress. L’oxydation des tanins dans certaines conditions, permet de libérer
des monomères colorés d’anthocyanes (d’où l’appellation synonyme assez courante de
proanthocyanidines).
Figure : Structure des proanthocyanidines du sorgho rouge (couleur rouge du sorgho avec n=1-30)
3. Tanins hydrolysables
Ces composés phénoliques présentent des propriétés semblables à celles des proanthocyanidines mais
ce ne sont pas des flavonoïdes. Il s’agit d’esters d’acide gallique ou d’acide élagique (dérivé du
shikimate) et de glucose. On retrouve des gallotanins accumulés dans certaines galles de végétaux qui
étaient utilisées pour la fabrication d’encres. Les tanins libérés dans le vin par les tonneaux de chênes
sont des ellagitanins, contrairement à ceux apportés par la peau des baies de raisin qui sont des tanins
non-hydrolysables.
* Les Tanins non-hydrolysables = Proanthocyanidines
Ce sont des polymères de 2 à 7 résidus flavan-3-ols reliés (catéchine, épicatéchine ; on parle également
de tanins catéchiques) par des liaisons C-C non-hydrolyables. sont des composés phagodéterrants
capables de faire précipiter les protéines (en particulier celles de la salive ; astringence conférée par les
tanins par exemple dans le vin). Cette propriété provoque une baisse d’appétence chez les animaux et
surtout une diminution de la digestibilité des protéines (les oiseaux aiment le sorgho blanc mais
détestent le sorgho rouge à cause des tanins qui lui confèrent cette couleur. La précipitation par les
tanins des enzymes secrétées par les champignons phytopathogènes nécrotrophes est une propriété qui
peut contribuer à la résistance chez certaines plantes. La synthèse des tanins est dans certains cas
induite par la perception de stress. L’oxydation des tanins dans certaines conditions, permet de libérer
des monomères colorés d’anthocyanes (d’où l’appellation synonyme assez courante de
proanthocyanidines).
Figure : Structure des proanthocyanidines du sorgho rouge (couleur rouge du sorgho avec n=1-30)
3. Tanins hydrolysables
Ces composés phénoliques présentent des propriétés semblables à celles des proanthocyanidines mais
ce ne sont pas des flavonoïdes. Il s’agit d’esters d’acide gallique ou d’acide élagique (dérivé du
shikimate) et de glucose. On retrouve des gallotanins accumulés dans certaines galles de végétaux qui
étaient utilisées pour la fabrication d’encres. Les tanins libérés dans le vin par les tonneaux de chênes
sont des ellagitanins, contrairement à ceux apportés par la peau des baies de raisin qui sont des tanins
non-hydrolysables.

(A) (B)
Figure : Représentation acide gallique (A) et tanins galliques (B)
Figure : Voies de biosynthèse des composés phénoliques
Propriétés
-Défense contre les pathogènes
-Molécules de dissuasion alimentaire
-Attraction des pollinisateurs
-Protections des rayonnements UV
-Molécules qui donnent couleur, arômes, parfums aux plantes
-Rôle structurel ( ex. lignine, constituante du bois)
(A) (B)
Figure : Représentation acide gallique (A) et tanins galliques (B)
Figure : Voies de biosynthèse des composés phénoliques
Propriétés
-Défense contre les pathogènes
-Molécules de dissuasion alimentaire
-Attraction des pollinisateurs
-Protections des rayonnements UV
-Molécules qui donnent couleur, arômes, parfums aux plantes
-Rôle structurel ( ex. lignine, constituante du bois)
(A) (B)
Figure : Représentation acide gallique (A) et tanins galliques (B)
Figure : Voies de biosynthèse des composés phénoliques
Propriétés
-Défense contre les pathogènes
-Molécules de dissuasion alimentaire
-Attraction des pollinisateurs
-Protections des rayonnements UV
-Molécules qui donnent couleur, arômes, parfums aux plantes
-Rôle structurel ( ex. lignine, constituante du bois)

LES HETEROSIDES
INTRODUCTIONLes hétérosides sont des composés issus de la condensation d’un ose par son groupe réducteuravec une molécule non-glucidique appelé la génine ou encore l’aglycone. Avec les alcaloïdes,ils sont les substances les plus anciennement connues et constituent les principes actifs debeaucoup de drogues végétales. La littérature anglo-saxone parle de glycosides, terme qu’onpeut retrouver dans certaines traductions. L’origine biosynthétique des génines est trèshétérogène (composés phénoliques, certains monoterpènes, triterpènes, alcaloïdes terpéniques,composés soufrés…). Leur nature est donc très diverse en raison du mode de liaison entre lagénine et l’(les) ose(s) ainsi que de la structure des génines et celle du copule glucidique.
LES GENINESElles sont des molécules de structures très variées. Cependant la plupart sont des composésphénoliques, généralement toxiques et peu solubles qui sont ainsi détoxifiés et solubilisés. Lesterpènes donnent rarement des hétérosides. Dans ce groupe seuls les triterpènes se rencontrentfréquemment sous forme d’hétérosides (hétérosides cardiotoniques, saponosides...) même si onnote quelques glucosides (acide loganique, iridoïdes des gamétopales). Certains acides grasramifiés et hydroxylés se trouvent sous forme d’hétérosides (jalapine de la résine desconvolvulaceae). Les glucosinolates et les hétérosides cyanogénétiques qui peuvent libérer del’acide cyanhydrique (toxique pour la cellule) sont toujours liés à des oses. Les alcaloïdes neforment pas de dérivés glycosylés. Les gluco-alcaloïdes qui paraissent faire exception ont unegénine qui, bien qu’azotée se rattache aux stéroïdes.
LES OSESIls sont aussi divers. Le plus fréquent est le D-glucose. Les hétérosides qui résultent d’une liaisonavec le seul glucose sont les glucosides. Les autres hexoses rencontrés sont le fructose, galactose,sorbose, mannose). L’ose peut être aussi un pentose (arabinose, xylose, ribose), unméthylpentose (rhamnose) ou un desoxysucre (digitoxose, cymarose, digitalose). Les oses sontrattachés isoléméent à la génine en un point ou en plusieurs points. Ils peuvent formés une chaînede polyosidique longue de 2, 3, 4 parfois même 5-7 oses disposés les uns à la suite des autres.
D-apiose D-digitoxose D-digitalose L-sorboseFigure : structure de quelques oses des hétérosides
BIOSYNTHESELa partie osidique et la génine sont synthétisés isolement puis combinés entre elles. Cettecombinaison se fait parfois en utilisant une liaison osidique déjà existante par le jeu de

transférases. De façon générale, la combinaison ose-génine, puis la condensation éventuelle deplusieurs oses sont réalisées par les NDP-oses.Pour la biosynthèse du rutoside par exemple, la réaction est la suivante :Quercétine (quercitoside) + UDP-L-Rahmnose ==► rutine (rutoside)
DEGRADATIONPlusieurs enzymes assurant la dégradation des polyosides sont capables d’hydrolyser leshétérosides. Ce sont les -glucosidases, les -glucosidases. Cette hydrolyse est relativementspécifique et parfois partielle comme dans le cas du copule osidique du rutoside. Des enzymesspécifiques assurent la dégradation des hétérosides cyanogénétiques et des glucosinolates.
CLASSIFICATIONOn distingue quatre principaux groupes :- Les hétérosides à génines phénoliques- Les hétérosides cyanogènes- Les glucosinolates- Les saponosides
a) Les hétérosides à génine phénoliqueLes hétérosides de composés phénoliques sont les plus nombreux. Ils correspondent à lanécessité de solubiliser ces composés pour les détoxifier et les rendre physiologiquement actifs.C’est le cas des lignines, subérines qui transitent dans le cytosol et franchissent le plasmalemmesous forme osidique mais c’est également celui de nombreux colorants des fleurs et des fruits quis’accumulent dans la vacuole à l’état d’hétérosides. Par exemple le rutoside atteint 17 à 22% desboutons floraux du Sophora japonica, s’abaisse beaucoup durant la floraison puis devient nulledans le fruit.
Figure : rutoside
b) Les hétérosides cyanogènesCe sont des hétérosides dérivés d’acides aminés capables de libérer de l’acide cyanhydrique suiteaux actions successives de deux enzymes : une alpha-glucosidase qui libère la fractionglucidique, et une hydroxinitrile lyase qui libère l’acide cyanhydrique (toxique) + la fractionaglycone. Dans le cas de l’amygdaline, un hétéroside cyanogène présent dans les noyaux decertaines Rosacées, la fraction aglycone correspond à du benzaldéhyde responsable d’un arômecaractéristique d’amande amère. Les quantités d’acide cyanhydrique libérées peuvent poser desréels problèmes dans le cadre de l’alimentation animale et humaine. Les exemples classiquessont le sorgho et le manioc. Les jeunes pousses de sorgho contiennent des quantités importantes
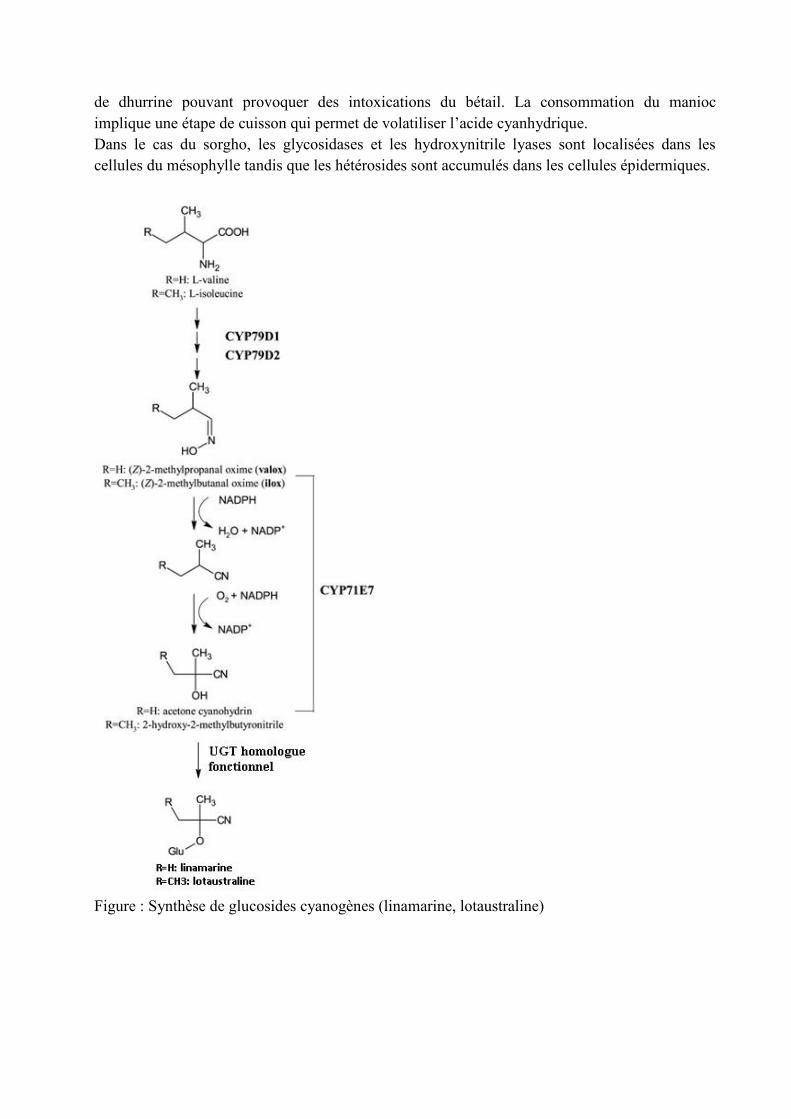
de dhurrine pouvant provoquer des intoxications du bétail. La consommation du maniocimplique une étape de cuisson qui permet de volatiliser l’acide cyanhydrique.Dans le cas du sorgho, les glycosidases et les hydroxynitrile lyases sont localisées dans lescellules du mésophylle tandis que les hétérosides sont accumulés dans les cellules épidermiques.
Figure : Synthèse de glucosides cyanogènes (linamarine, lotaustraline)

Figure : Hydrolyse d’un hétéroside cyanogène
c) Les glucosinolatesCe sont des hétérosides soufrés (thioglucosides) dont les génines sont dérivées d’acides aminés,relativement caractéristiques des plantes de l’ordre des Brassicales et responsables du goût âcrecaractéristique des choux, de la moutarde. La majorité de ces composés dérivent d’acides aminésspéciaux. Des mécanismes particuliers assurent l’allongement des chaînes suivis de réactionsd’oxydation, acylation, N-sulfonation etc. qui permettent la très grande variété desglucosinolates. L’hydrolyse de ces composés par des enzymes spécifiques des plantes qui lessynthétisent produit différents composés toxiques: l’isothiocyanate, le thiocyanate et du nitrile.Ces composés ont un effet goîtrogène lié à l’inhibition du métabolisme de l’iode dans lathyroïde. Ces composés sont également des phagodéterrants puissants bien que certains insectesspécialisés aient contourné le problème et même fait de la présence de glucosinolates undéterminant de leur comportement alimentaire et reproducteur.Remarque : la distribution taxonomique des glucosinolates est beaucoup plus restreinte que celledes hétérosides cyanogènes.
Figure : Structure des glucosinolates
d) Les saponosidesCe sont des hétérosides formés d’une génine de type triterpène appelée sapogénine, et degroupements osidiques. Ils ne se rencontrent que dans certains groupes voir un petit nombre defamilles ou une seule qu’ils caractérisent. Il s’agit de composés souvent amers, amphiphiles auxpropriétés tensioactives qui ont pu être mises à profit pour la confection de détergents. Cesmolécules ont également la propriété d’affecter la stabilité membranaire et ces composés sontimpliqués dans la résistance à des pathogènes (exemple de l’avécanine de l’avoine). Ondistingue :- les saponosides à génine triterpénique pentacyclique- les saponosides à génine stéroïde
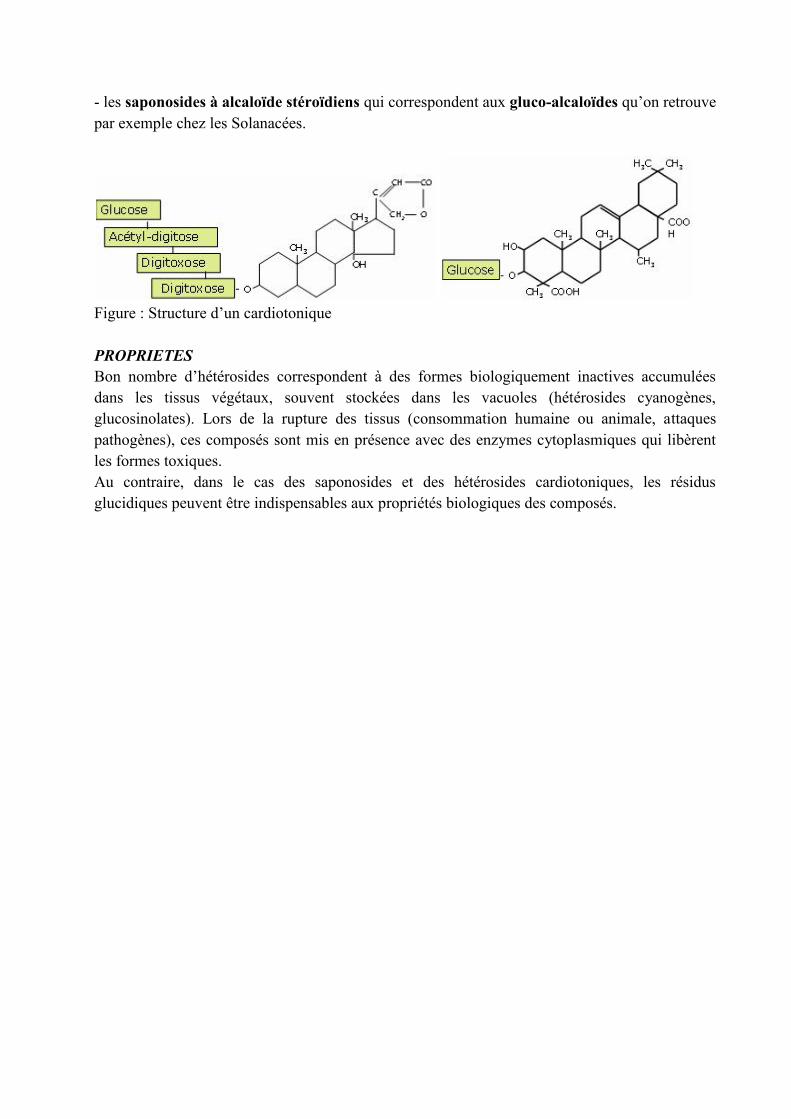
- les saponosides à alcaloïde stéroïdiens qui correspondent aux gluco-alcaloïdes qu’on retrouvepar exemple chez les Solanacées.
Figure : Structure d’un cardiotonique
PROPRIETESBon nombre d’hétérosides correspondent à des formes biologiquement inactives accumuléesdans les tissus végétaux, souvent stockées dans les vacuoles (hétérosides cyanogènes,glucosinolates). Lors de la rupture des tissus (consommation humaine ou animale, attaquespathogènes), ces composés sont mis en présence avec des enzymes cytoplasmiques qui libèrentles formes toxiques.Au contraire, dans le cas des saponosides et des hétérosides cardiotoniques, les résidusglucidiques peuvent être indispensables aux propriétés biologiques des composés.

B) SYNTHESE ET ACCUMULATION DES METABOLITES SECONDAIRES
Les métabolites secondaires sont pour la plupart des toxiques pour les cellules végétales ellemêmes. Leur accumulation à des teneurs pouvant atteindre plusieurs % de la masse de MSsuppose des mécanismes particuliers pour limiter leur toxicité. On peut citer :
1. La sécrétion extracellulaireElle concerne des métabolites secondaires peu solubles dans l’eau (essentiellement des terpènesmais pas uniquement) :- accumulation sous-cuticulaire des huiles essentielles au niveau de poils (trichomes) glanduleux(exemples nombreux chez les Lamiacées)- accumulation dans des poches à essence (exemple de la famille des Rutacées)- diffusion des composés volatiles qui traversent les structures cellulaires pour passer dans l’air.
2. La glycosylation et/ou la séquestration vacuolaire- nombreux composés phénoliques
3. L’accumulation intracellulaireLes métabolites sont conservés dans des tissus ou cellules très spécialisés- laticifères : exemple du latex riche en alcaloïdes chez le pavot- idioblastes : exemple de la synthèse des alcaloïdes chez la pervenche de Madagascar- trichomes
4. La compartimentationLa contention des enzymes responsable de la conversion d’un précurseur en un produit toxique(exemples de la juglone, des glucosinolates et des composés cyanogénétiques) est également unestratégie largement répandue.
Enfin, bon nombre de composés phénoliques s’accumulent dans l’apoplaste au niveau des paroisvégétales où ils jouent à la fois des rôles structuraux (lignine et pontages d’hémicelluloses) maiségalement dans la limitation de la progression des pathogènes.

C) MODES DE PRODUCTION DES METABOLITES SECONDAIRES
I. LES CELLULES VEGETALESLa production de molécules actives à partir de suspensions de cellules végétales est une des voiesexplorée et mise en œuvre dans quelques cas. Elle montre le potentiel immense de cette approcheautant en hémisynthèse qu’en synthèse entière. En pratique, elle nécessite la prise en compte dequelques précautions particulières. Il s’agit par exemple de travailler sur un matériel végétalstérile et dans des conditions aseptiques, de screener et stabiliser les lignées génétiquesintéressantes, de déterminer un milieu de culture approprié (macro et oligoéléments, vitamines…). Un équilibre hormonal adapté permet d’obtenir un cal et des cellules indifférenciées et decontrôler leur développement in vitro. Des éliciteurs sont également utilisés pour améliorer lesperformances de la culture.
Figure : Influence des hormones sur le développement des cellules végétales
La croissance cellulaire et la production de molécules actives sont confrontées à quelquesfacteurs limitants comme le transfert de masse (gaz-liquide), la résistance au cisaillement (≤ 100trs/min), l’homogénéisation du milieu, la sédimentation, l’attachement, le contact cellule-liquide… La plupart des métabolismes secondaires sont intracellulaires, intravacuolaires. Pour leurextraction, les membranes sont déstabilisées, l'efflux des métabolites est augmenté puis l'intégritémembranaire est restaurée.
La conjonction de tous ces facteurs fait que dans certains cas la culture in vitro donne un rapportde gain favorable comparé à la production au champ (2 :Nicotine/N. tabacum ; 5 : aciderosmarinic/ Coleus blumei). Mais, pour des composés comme la morphine et la vinblastine lesrendements (respectivement <<1 et 0) de la culture in vitro sont encore très faibles et même nulspar rapport à ceux du champ.
Tableau : Exemples de cellules végétales stabiliséesEspèce végétale (cals) Temps de doublement (jrs)N. tabaccumC. roseusQuassia amaraPsoralea cinerea
0.63/1.72.25.216

II. ALTERNATIVES A LA CULTURE CELLULAIRE
1. Plantes à traire
La diversité chimique des racines est moins étudiée et moins exploitée que celle des partiesaériennes, alors qu'une grande diversité de molécules y est produite, accumulée et excrétée (5 à30% du carbone fixé par la photosynthèse des plantes est restitué au sol par les racines). Latechnologie brevetée* PAT « Plantes à Traire »® permet d'accéder aux composés bio actifsracinaires en cultivant les plantes en hydroponie.C'est un procédé continu et non destructif, qui permet des récoltes répétitives de ces composés enréutilisant les mêmes plantes.
Figure : Principe de la technologie des plantes à traire
2. AutresLes tiges feuillées, les racines et la co-culture d’organes et/ou de cellules sont autant d’autrespossibilités. Ces nouvelles approches s’ajoutent au génie métabolique des végétaux qui vise àcontrôler le métabolisme des cellules pour la production de métabolites secondaires.

Relations entre métabolisme secondaire et métabolisme primaire
Les jonctions entre métabolisme primaire et secondaire sont nombreuses et parfois complexes.Les principales sont synthétisées dans le schéma suivant qui indique également l’originebiosynthétique des principales phytohormones. Les étapes enzymatiques les plus représentativessont :- la phénylalanine ammonia lyase (PAL) qui initie la synthèse de la plupart des composésphénoliques à partir d’un acide aminé- la chalcone synthase qui initie la synthèse des flavonoïdes à partir du malonyl CoA et d’unélement phénylpropane- la voie mévalonate indépendante de biosynthèse de l’isopentényl diphosphate (IPP), à l’originede la synthèse de plus grande partie des terpenoïdes.
Figure : Connexions entre les métabolismes primaire et secondaires





























