Embed Size (px)
Citation preview

INTERFACES• Interfaces fluides
– Membranes• Oléosomes (T. Chardot)• Construction d’une interface de phospholipides (M. Axelos)• Composition / Structure et dynamique membranaire (C. Tribet)
– Projets
• Interfaces ‘solides’– Parois II (Aguié)
• Monocouches orientées de cellulose + macromolécules pariétales Propriétés de surface: adhésion relation avec la structure
– Projets

Oléosomes OléosinesEnjeux
CognitifsCompréhension de la structure, et de l’assemblage des corps lipidiquesvégétaux oléagineux des zones tempéréesÉchelle intermédiaire molécule cellule
FinalisésProtéines amphiphiles (empaquetage de molécules hydrophobes)Lipochimie lipides à façon et dérivés pour la chimie des polymères
Le projet-Identifier décrire les différents composés du corps lipidique chez deuxorganismes modèles: A. thaliana et lalevure Y. lipolytica, comprendre leur assemblage -Etudier la dynamique de formation du corps lipidique(modèle levure) et la mobilisation ultérieure des lipides (modèle plante)
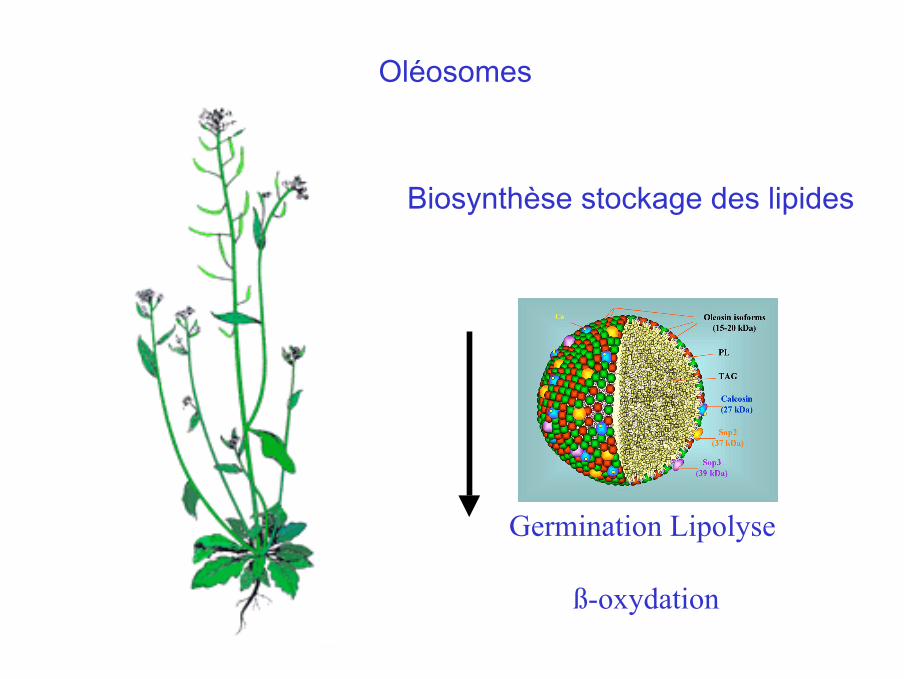
Biosynthèse stockage des lipides
Germination Lipolyse
ß-oxydation
Oléosomes

Expression, purification et propriétés d’oléosines àdifférentes interfaces
OléosomePurification des oléosomes
Les protéines de l’oléosomeInterfacialesIdentité
Les oléosinesExpression, purification, solubilitéInsertion à différentes interfaces
Perspectives
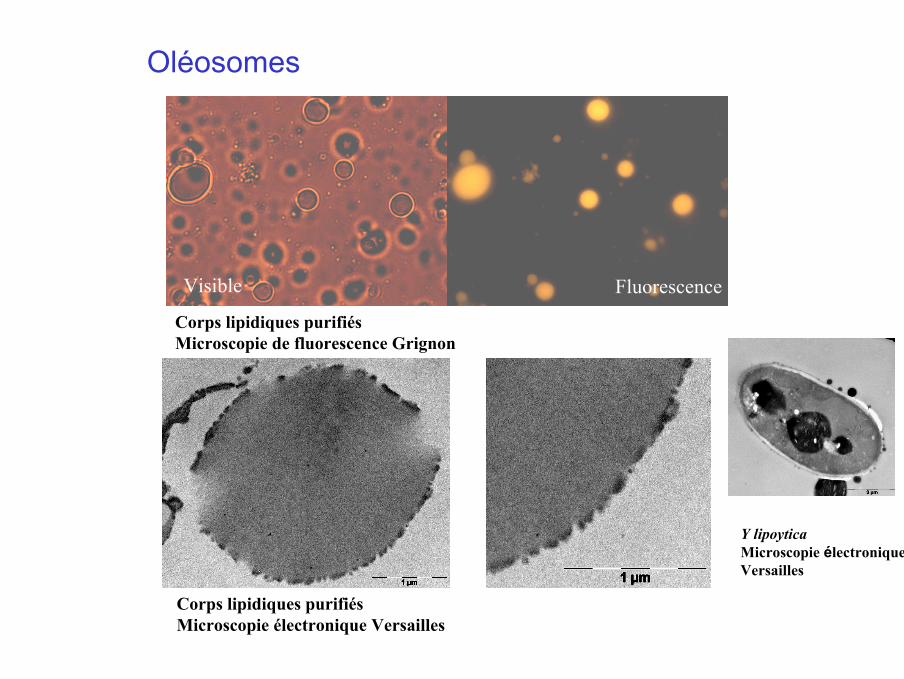
Oléosomes
Corps lipidiques purifiésMicroscopie électronique Versailles
Corps lipidiques purifiésMicroscopie de fluorescence Grignon
Visible Fluorescence
Y lipoyticaMicroscopie électroniqueVersailles
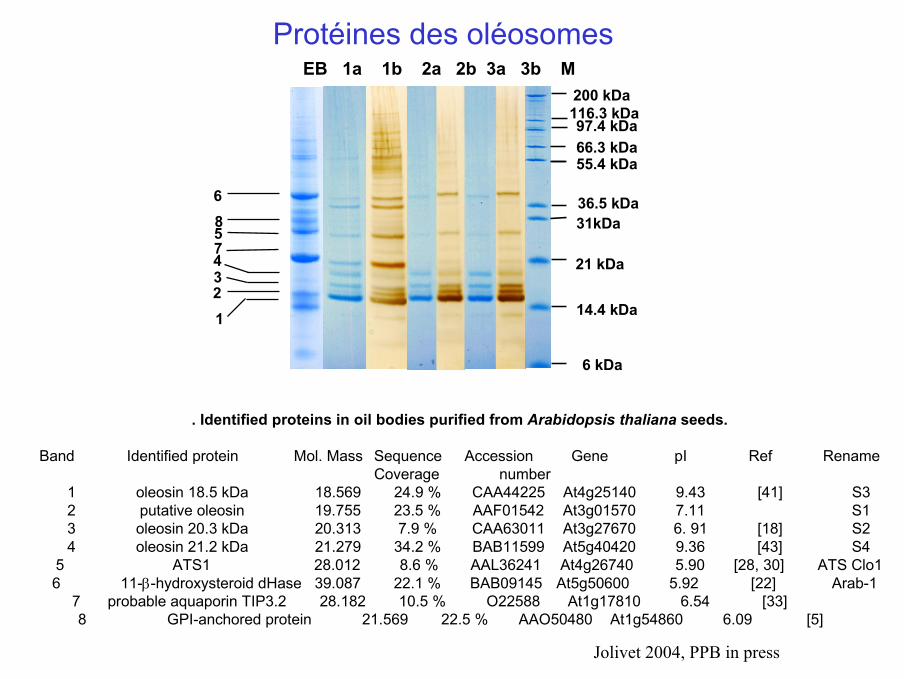
EB 1a 1b 2a 2b 3a 3b M
55.4 kDa
36.5 kDa31kDa
21 kDa
14.4 kDa
6 kDa
66.3 kDa
6
5
4321
97.4 kDa 116.3 kDa 200 kDa
8
7
Protéines des oléosomes
. Identified proteins in oil bodies purified from Arabidopsis thaliana seeds.
Band Identified protein Mol. Mass Sequence Accession Gene pI Ref Rename Coverage number
1 oleosin 18.5 kDa 18.569 24.9 % CAA44225 At4g25140 9.43 [41] S32 putative oleosin 19.755 23.5 % AAF01542 At3g01570 7.11 S13 oleosin 20.3 kDa 20.313 7.9 % CAA63011 At3g27670 6. 91 [18] S24 oleosin 21.2 kDa 21.279 34.2 % BAB11599 At5g40420 9.36 [43] S4
5 ATS1 28.012 8.6 % AAL36241 At4g26740 5.90 [28, 30] ATS Clo1 6 11-β-hydroxysteroid dHase 39.087 22.1 % BAB09145 At5g50600 5.92 [22] Arab-1
7 probable aquaporin TIP3.2 28.182 10.5 % O22588 At1g17810 6.54 [33]8 GPI-anchored protein 21.569 22.5 % AAO50480 At1g54860 6.09 [5]
Jolivet 2004, PPB in press
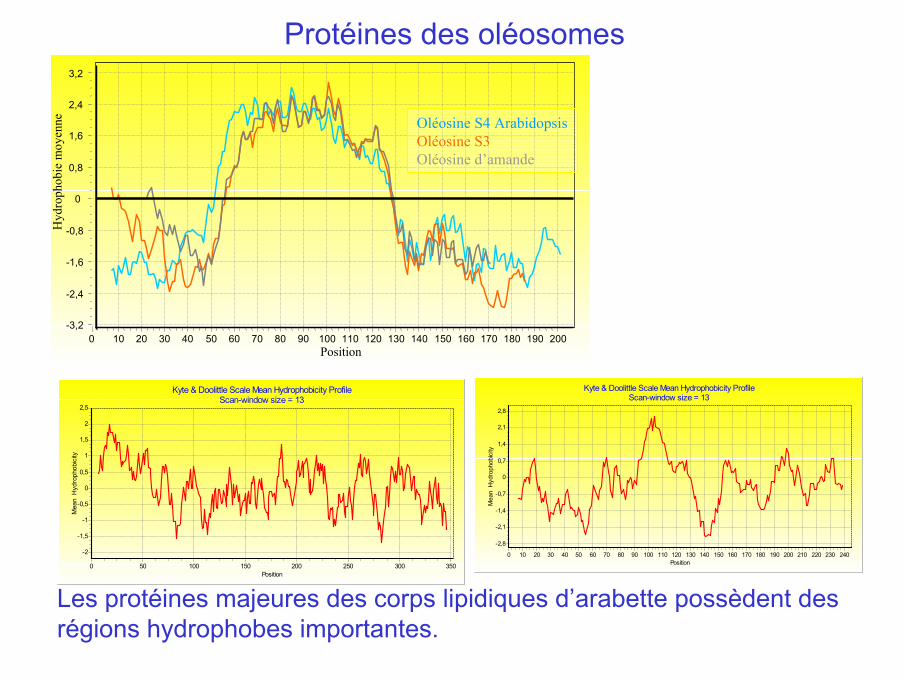
Kyte & Doolittle Scale Mean Hydrophobicity ProfileScan-window size = 13
Position2402302202102001901801701601501401301201101009080706050403020100
Mea
n H
ydro
phob
icity
2,8
2,1
1,4
0,7
0
-0,7
-1,4
-2,1
-2,8
Kyte & Doolittle Scale Mean Hydrophobicity ProfileScan-window size = 13
Position350300250200150100500
Mea
n H
ydro
phob
icity
2,5
2
1,5
1
0,5
0
-0,5
-1
-1,5
-2
Position2001901801701601501401301201101009080706050403020100
3,2
2,4
1,6
0,8
0
-0,8
-1,6
-2,4
-3,2
Hyd
roph
obie
moy
enne Oléosine S4 Arabidopsis
Oléosine S3Oléosine d’amande
Les protéines majeures des corps lipidiques d’arabette possèdent desrégions hydrophobes importantes.
Protéines des oléosomes
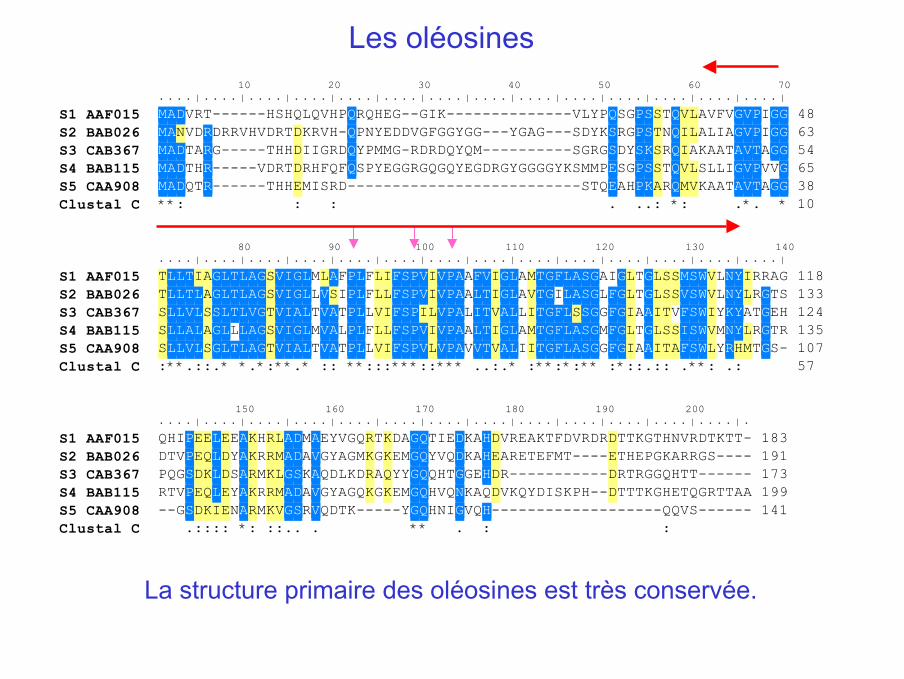
10 20 30 40 50 60 70. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
S1 AAF015 MADVRT------HSHQLQVHPQRQHEG--GIK--------------VLYPQSGPSSTQVLAVFVGVPIGG 48S2 BAB026 MANVDRDRRVHVDRTDKRVH-QPNYEDDVGFGGYGG---YGAG---SDYKSRGPSTNQILALIAGVPIGG 63S3 CAB367 MADTARG-----THHDIIGRDQYPMMG-RDRDQYQM----------SGRGSDYSKSRQIAKAATAVTAGG 54S4 BAB115 MADTHR-----VDRTDRHFQFQSPYEGGRGQGQYEGDRGYGGGGYKSMMPESGPSSTQVLSLLIGVPVVG 65S5 CAA908 MADQTR------THHEMISRD--------------------------STQEAHPKARQMVKAATAVTAGG 38Clustal C **: : : . ..: *: .*. * 10
80 90 100 110 120 130 140. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . |
S1 AAF015 TLLTIAGLTLAGSVIGLMLAFPLFLIFSPVIVPAAFVIGLAMTGFLASGAIGLTGLSSMSWVLNYIRRAG 118S2 BAB026 TLLTLAGLTLAGSVIGLLVSIPLFLLFSPVIVPAALTIGLAVTGILASGLFGLTGLSSVSWVLNYLRGTS 133S3 CAB367 SLLVLSSLTLVGTVIALTVATPLLVIFSPILVPALITVALLITGFLSSGGFGIAAITVFSWIYKYATGEH 124S4 BAB115 SLLALAGLLLAGSVIGLMVALPLFLLFSPVIVPAALTIGLAMTGFLASGMFGLTGLSSISWVMNYLRGTR 135S5 CAA908 SLLVLSGLTLAGTVIALTVATPLLVIFSPVLVPAVVTVALIITGFLASGGFGIAAITAFSWLYRHMTGS- 107Clustal C :**.::.* *.*:**.* :: **:::***::*** ..:.* :**:*:** :*::.:: .**: .: 57
150 160 170 180 190 200. . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | . . . . | .
S1 AAF015 QHIPEELEEAKHRLADMAEYVGQRTKDAGQTIEDKAHDVREAKTFDVRDRDTTKGTHNVRDTKTT- 183S2 BAB026 DTVPEQLDYAKRRMADAVGYAGMKGKEMGQYVQDKAHEARETEFMT----ETHEPGKARRGS---- 191S3 CAB367 PQGSDKLDSARMKLGSKAQDLKDRAQYYGQQHTGGEHDR-----------DRTRGGQHTT------ 173S4 BAB115 RTVPEQLEYAKRRMADAVGYAGQKGKEMGQHVQNKAQDVKQYDISKPH--DTTTKGHETQGRTTAA 199S5 CAA908 --GSDKIENARMKVGSRVQDTK-----YGQHNIGVQH-------------------QQVS------ 141Clustal C .:::: *: ::.. . ** . : :
La structure primaire des oléosines est très conservée.
Les oléosines

Évaluation de l’hydrophobie d’une molécule :
- Pour un émulsifiant de BPM :
HLB = 20 x Mhydrophile / Mtotal
si 3 ≤ HLB ≤ 6, bon émulsifiant eau dans huile
si 7 ≤ HLB ≤ 9, bon agent mouillant
si 10 ≤ HLB ≤ 18, bon émulsifiant huile dans eau
- Pour une protéine:
- Méthode de Kyte et Doolittle (1982) : calcul par fenêtre de x acides
aminés de l’hydropathie moyenne représentation graphique et
indice « GRAVY »
- Méthode de Nakai (1983) : hydrophobie = % d’acides aminés
hydrophobes contenus dans la protéine
- Hypothèse de Beisson (2001) sur les oléosines
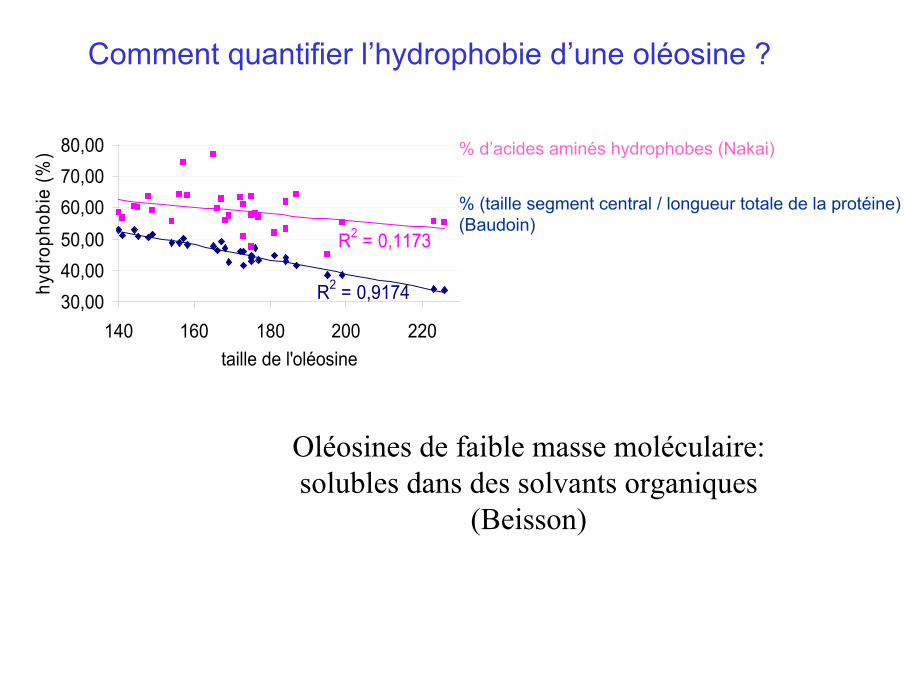
% d’acides aminés hydrophobes (Nakai)
% (taille segment central / longueur totale de la protéine)(Baudoin)
Comment quantifier l’hydrophobie d’une oléosine ?
R2 = 0,9174
R2 = 0,1173
30,0040,0050,00
60,0070,0080,00
140 160 180 200 220taille de l'oléosine
hydr
opho
bie
(%)
Oléosines de faible masse moléculaire:solubles dans des solvants organiques
(Beisson)

Domaines d’application des oléosines ?
hydro
phile
hydro
phob
e
Antimou
ssan
ts
émuls
ions e
au / h
uile
mouss
ants,
agen
ts de
mou
illage
émuls
ions h
uile /
eau
déter
gents
solub
ilisan
ts de
prod
uits o
rganiq
ues
Applications des surfactants (de Gennes et al., 2002)
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 8molarité en urée (M)
% d
e pr
otéine
s so
lubilis
ées
S1S2S3S4S5SM2SM3
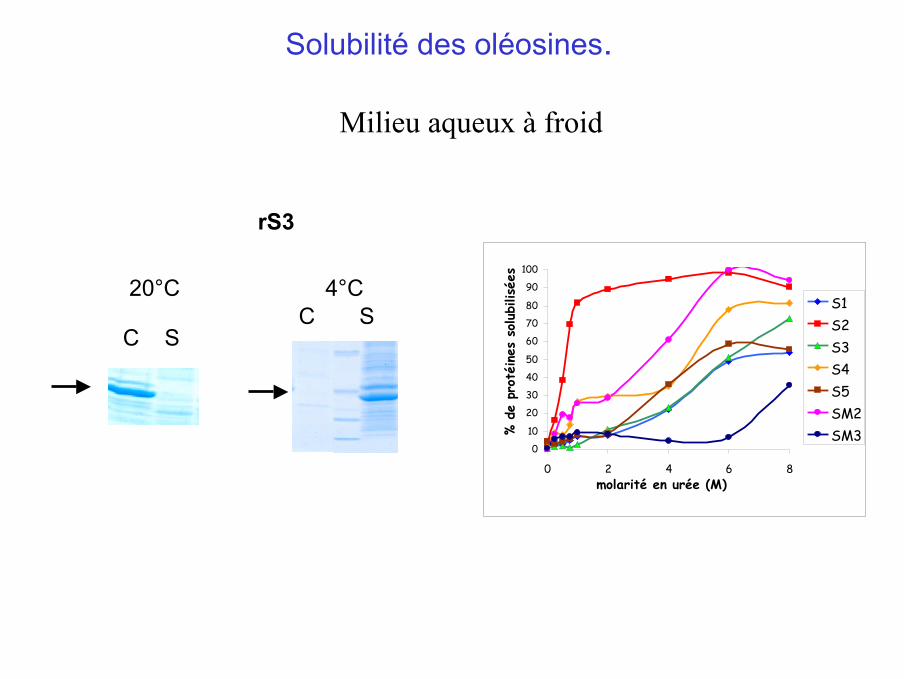
Solubilité des oléosines.
Milieu aqueux à froid
20°C 4°CC S
C S
rS3
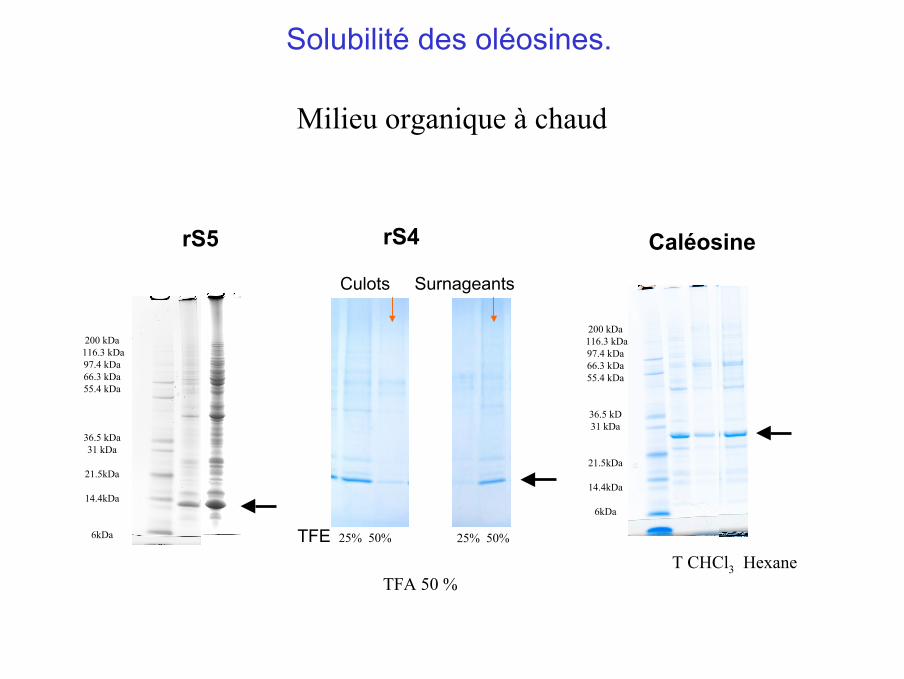
rS5
Milieu organique à chaud
Culots Surnageants
rS4
TFE 25% 50% 25% 50%
200 kDa 116.3 kDa97.4 kDa66.3 kDa55.4 kDa
36.5 kDa31 kDa
21.5kDa
14.4kDa
6kDa
Solubilité des oléosines.
T CHCl3 Hexane
200 kDa 116.3 kDa97.4 kDa66.3 kDa55.4 kDa
36.5 kD31 kDa
21.5kDa
14.4kDa
6kDa
Caléosine
TFA 50 %

S4S3
116.3
21.5
14.4
3136.5
55.466.397.4
200kDa 1 2 3
S4 S3
4 5
Les oléosine peuvent s’auto associer, via des motifs GxxxG ?
S1 S2 S3 S4 SM1 SM2 SM3GRGQG
GGYGG GQYEGGYGAG GYGGC
Leu Zipp1 ? GQLFGGVPIG GVPIGGSVIG GSVIG Leu Zipp2 ? GSVIG GTVIG
GTLIGGFLVG
GKEMC GKEMC GYFHG GYFHG
Statistical analysis of amino acid patterns in transmembrane helices: the GxxxG motif occurs frequently and in association with beta-branched residues at neighboring positions Alessandro Senes, Mark Gerstein and Donald M. Engelman J Mol.Biol (2000), 296(3), 921-936
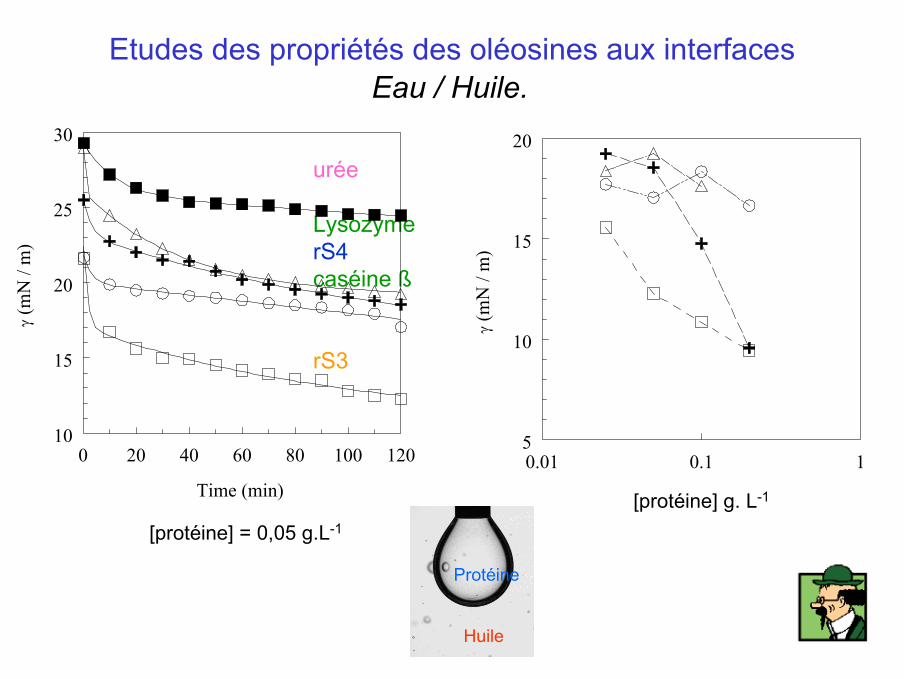
urée
Lysozyme rS4caséine ß
rS3
[protéine] = 0,05 g.L-1
Protéine
Huile
Etudes des propriétés des oléosines aux interfaces
0 20 40 60 80 100 120
Time (min)
10
15
20
25
30
γ (m
N /
m)
0.01 0.1 15
10
15
20
γ (m
N /
m)
[protéine] g. L-1
Eau / Huile.
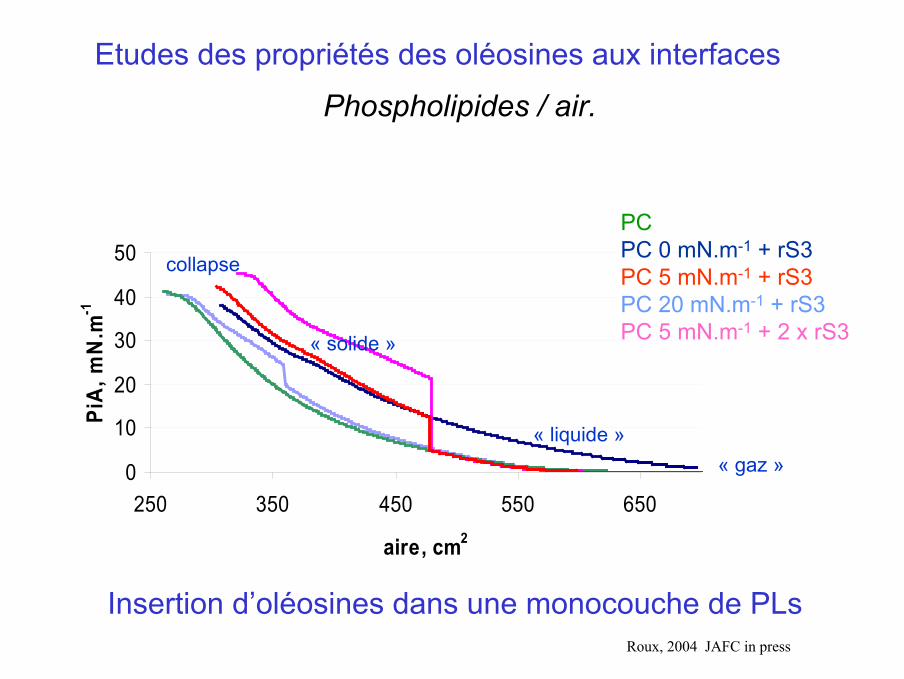
Etudes des propriétés des oléosines aux interfaces
0
10
20
30
40
50
250 350 450 550 650
aire, cm2
PiA
, mN
.m-1
PCPC 0 mN.m-1 + rS3PC 5 mN.m-1 + rS3PC 20 mN.m-1 + rS3PC 5 mN.m-1 + 2 x rS3
« gaz »« liquide »
« solide »
Insertion d’oléosines dans une monocouche de PLs
collapse
Phospholipides / air.
Roux, 2004 JAFC in press

Dépôt d’oléosines sur un film de PC plus ou moins comprimée
Dépôt d’oléosines rS3 sur un film
de PC à 0 mN.m-1 (« gaz »)
Dépôt d’oléosines rS3 sur un
film de PC à 5 ou 20 mN.m-1
(« liquide » ou « solide »)
4 m
m
5 mm
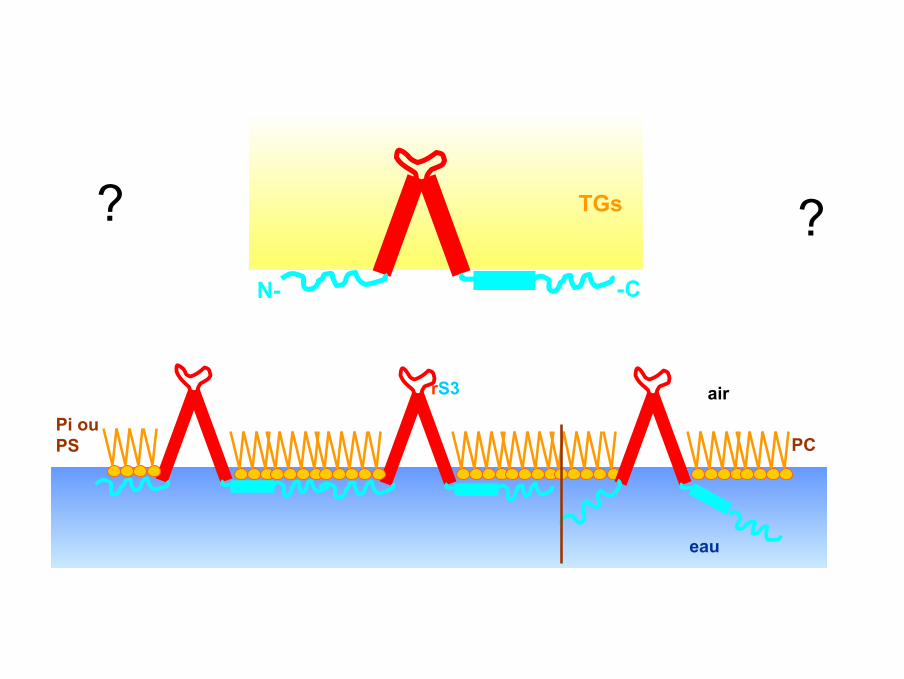
eau
PC
rS3
Pi ouPS
air
TGs
N- -C
? ?
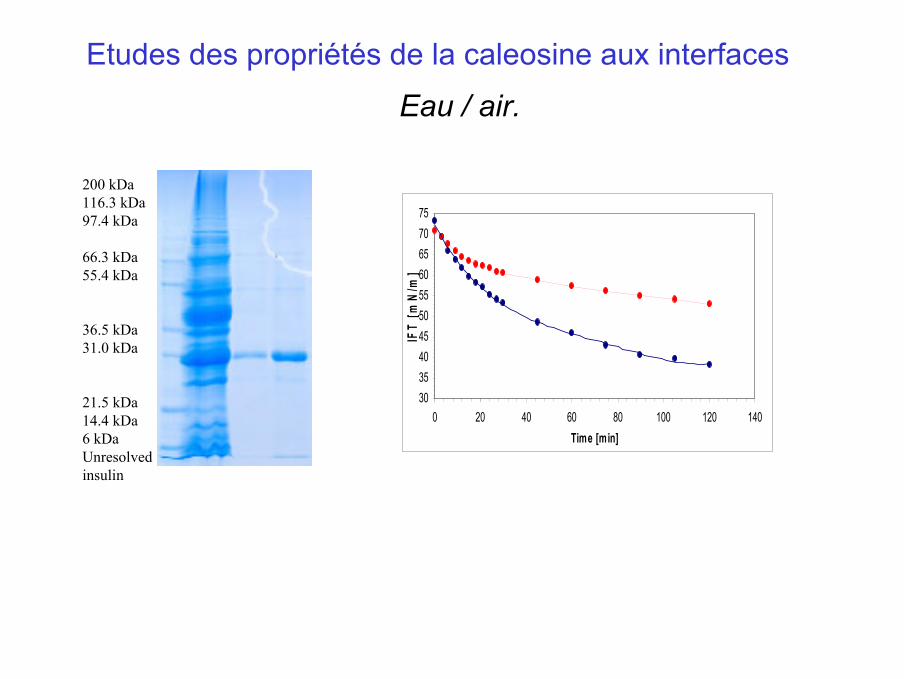
200 kDa116.3 kDa97.4 kDa
66.3 kDa55.4 kDa
36.5 kDa31.0 kDa
21.5 kDa14.4 kDa6 kDaUnresolvedinsulin
30354045505560657075
0 20 40 60 80 100 120 140Time [min]
IFT
[mN
/m]
Etudes des propriétés de la caleosine aux interfaces
Eau / air.
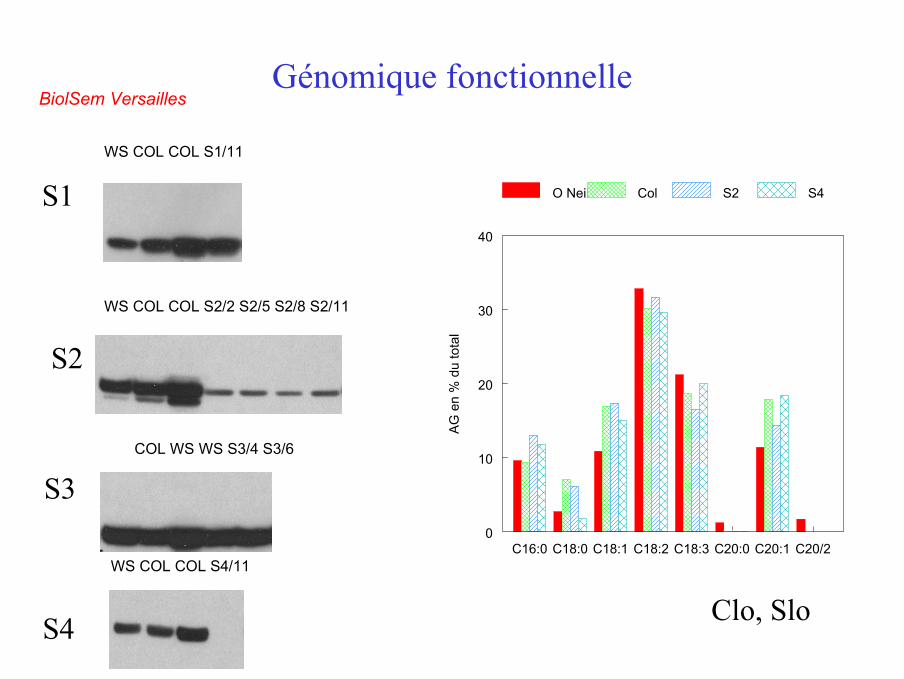
WS COL COL S2/2 S2/5 S2/8 S2/11
WS COL COL S1/11
S1
S2
S3
S4
COL WS WS S3/4 S3/6
WS COL COL S4/11
Génomique fonctionnelleBiolSem Versailles
Clo, Slo
C16:0 C18:0 C18:1 C18:2 C18:3 C20:0 C20:1 C20/20
10
20
30
40
AG
en
% d
u to
tal
O Neil Col S2 S4

Conclusions…
Purification d’oléosines recombinantes à l’homogénéité, en quantités.
Caractéristiques d’émulsifiants avec variation de l’hydrophobie en fonction de
l’isoforme étudiée émulsifiants à applications variées.
In vitro, stabilisation par des oléosines d’émulsions d’huile dans eau en synergie
avec des phospholipides. Capacité des oléosines à s’insérer à l’interface eau /
huile et à abaisser la tension de surface. Insertion dans une monocouche de
PLs à l’interface eau / air.
… et perspectivesDisponibilité en grandes quantités des oléosines études de structure en
solution dans des milieux modèles (protéines marquées, complexes ?), à
différentes des interfaces, reconstitution d’oléosomes, vectorisation.

Génétique végétale, Le Génétique végétale, Le MoulonMoulon
ISDISDInterfaces Systèmes dispersés, NantesInterfaces Systèmes dispersés, Nantes
Biologie des semences, VersaillesBiologie des semences, Versailles
M Zivy, L Negroni, M Davanture
M Axelos et M.H. Ropers
L Lepiniec, M Miquel
Chimie Biologique, GrignonChimie Biologique, Grignon E Roux, P Jolivet, S, D’Andrea, T Chardot
GDR AMVGDR AMV

Interactions phospholipides / polysaccharidesà l ’interface eau/air: construction d’un
modèle pour l’étude en réflectivité
M.H. Ropers
*Unité de Physico – Chimie des Macromolécules, INRA Nantes
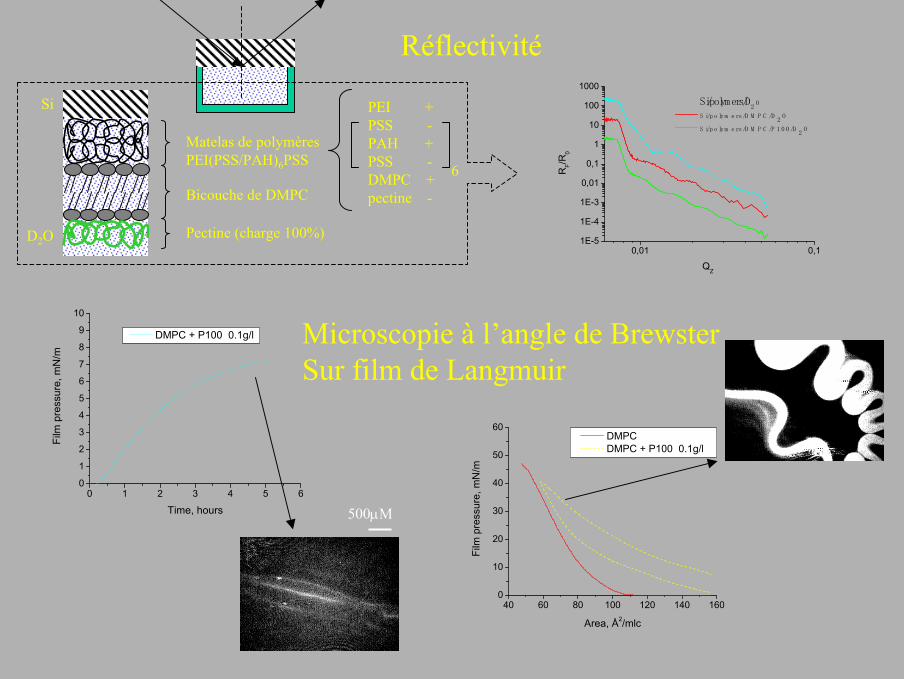
D2O
Si
Matelas de polymèresPEI(PSS/PAH)6PSS
Bicouche de DMPC
Pectine (charge 100%)
PEI +PSS -PAH +PSS -DMPC +pectine -
6
500µM
Microscopie à l’angle de BrewsterSur film de Langmuir
0 1 2 3 4 5 60
1
2
3
4
5
6
7
8
9
10
Film
pre
ssur
e, m
N/m
Time, hours
DMPC + P100 0.1g/l
40 60 80 100 120 140 1600
10
20
30
40
50
60
Film
pre
ssur
e, m
N/m
Area, Å2/mlc
DMPC DMPC + P100 0.1g/l
0,01 0,11E-5
1E-4
1E-3
0,01
0,1
1
10
100
1000
Si/polymers/D2 O
Si/polymers/DMPC/D2 O
Si/polymers/DMPC/P100/D2 O
RF/R
0
QZ
Réflectivité

Composition / Structure et dynamique membranaire
C. Tribet

Projet: « Oléosome » en cours d’élargissement
Approche in vitro: reconstitution des oléosomesStabilisation de gouttelettes par les # protéinesStructure locale des phospho autour des protéines (RMN)
Approche in vitro: sur interface modèle:Ségrégation entre les différents phospho, les différentes protéines, rôle des stérols ?Structure d’insertion des oléosines dans la couche de phosphoInteractions protéines/phospho: comparaison triblock
Méthodes: Corrélation de fluorescence, BAM, rhéologie interfaciale, réflectivité, Biacore
Approche in silico : modélisation (1post-doc)
INRA (UMR-CB Grignon, Nantes), ESPCI, + ?
Approche in planta: génétique: modulation du stockage des lipides de réservesmutants oléosomes de composition déterminée

Autres projets ?:
Interactions phospholipides/microfibrilles de cellulose ?
Interactions phospholipides/ protéines de réserve: CRPP – INRA Nantes

Interfaces Solides:
Véronique Aguié

Interfaces dans les parois II?
Signification?
Intérêts des modèles?
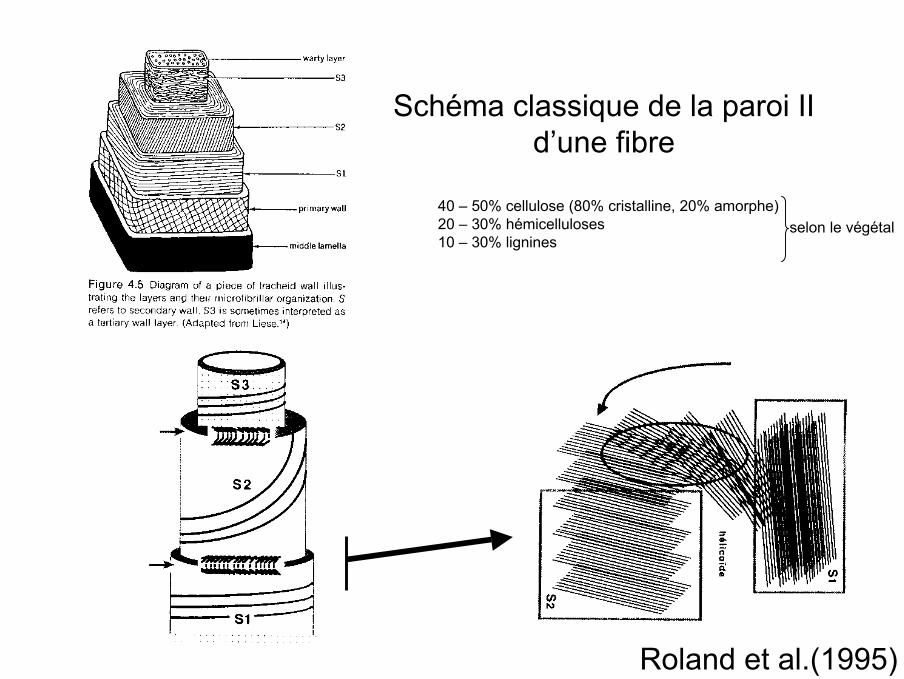
Schéma classique de la paroi IId’une fibre
Roland et al.(1995)
40 – 50% cellulose (80% cristalline, 20% amorphe)20 – 30% hémicelluloses10 – 30% lignines
selon le végétal
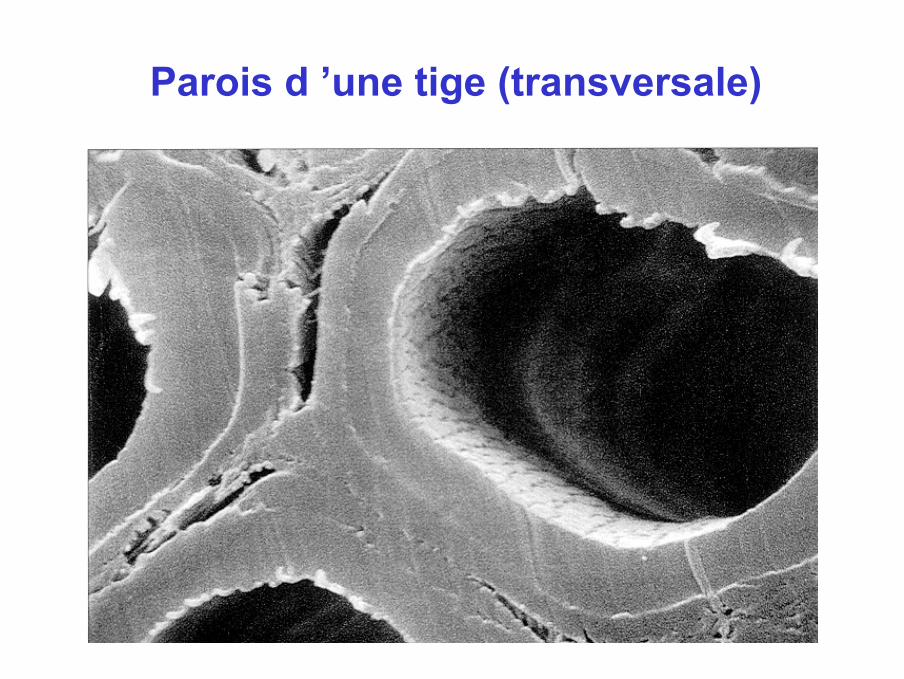
Parois d ’une tige (transversale)
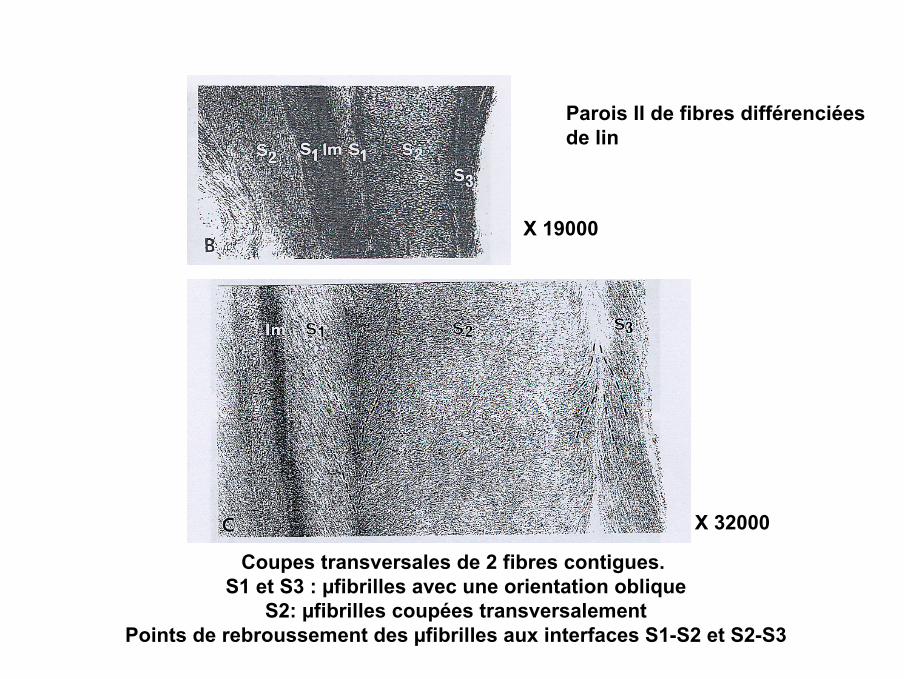
Parois II de fibres différenciéesde lin
X 19000
Coupes transversales de 2 fibres contigues. S1 et S3 : µfibrilles avec une orientation oblique
S2: µfibrilles coupées transversalementPoints de rebroussement des µfibrilles aux interfaces S1-S2 et S2-S3
X 32000

Signification des propriétés de surfacedans ces systèmes pariétaux?
⇒ Rôle des structures « 2D » parallèles/non parallèles sur:
- les propriétés de cohésion dans le plan et entre lesplans- la structure des autres polymères pariétaux- la réactivité (chimique, enzymatique)- la compréhension des interactions mises en jeu dansles zones interfaciales- sur les propriétés (3D) des tissus (mécaniques)

Réaliser des surfaces modèles planes de cellulose (/hémicelluloses, lignines)
orientée/non orientée

Intérêts des systèmes modèles « 2D » de composition connue
1) Mono-couches à partir de très peu de matière2) Interactions des macromolécules dans un espace confiné3) Rôle de l’orientation sur les propriétés dans le plan et
perpendiculairement au plan4) Adhésion entre couches5) Etalonnage (composition, géométrie) des propriétés de surface des
parois et fibres (adhésion, mouillage,…)

Conclusions
Construction de cette action transversale avance …
Interfaces de type ‘membrane’: deux projets bien identifiésUn autre à construire
Parois secondaires:Idées intéressantes à creuser
compétences en adhésion et mouillage ?












