Embed Size (px)
Citation preview

Méthodes de qualification des immobilisationschimiques d’une enzyme sur un support solide
Anne Belzil et Carmen Parent
Résumé : L’enzyme anhydrase carbonique humaine a été immobilisée sur un support polymérique thermoplastique envue de favoriser l’hydratation du CO2 gazeux en ions bicarbonate. L’immobilisation du catalyseur a été réalisée par unesérie de réactions chimiques qui permettent la liaison covalente de l’enzyme au support polyamide. Différentes concen-trations initiales d’enzymes ont été vérifiées soit 0,25; 0,50; 0,75; 1,00 et 2,00 mg/mL. Différentes techniques ont étéélaborées afin de qualifier l’attachement de l’enzyme. Une quantification du nombre de protéines liées fût déterminéepar un dosage Bradford des solutions protéiques résultantes de l’incubation du support avec l’enzyme anhydrase carbo-nique humaine de type II recombinante (ACHIIr). Un essai ELISA, quant à lui, a donné une évaluation protéique quali-tative nous permettant un suivi de la robustesse de la liaison de l’enzyme dans le temps. Les activités d’hydrolyse dup-nitrophényl acétate ainsi que l’hydratation du CO2 gazeux de l’anhydrase carbonique ont été évaluées respectivementpar spectrophotométrie et électrométrie. Le pourcentage d’enzymes actives suite à l’immobilisation fût mesuré grâce àune droite d’étalonnage en estérase. Des taux de transformation catalytique pour l’hydratation du CO2 gazeux ont étécalculés pour chaque type d’immobilisation.
Mots clés : immobilisation, enzyme, estérase, hydratation.
Abstract: Carbonic anhydrase was immobilized on a polymer thermoplastic to promote gaseous CO2 hydration intobicarbonate ions. Catalyst immobilization was realized through a series of chemical reactions enabling enzyme covalentbinding to polyamide support. Different initial enzyme concentrations of 0.25, 0.50, 0.75, 1, and 2 mg/ml were veri-fied. Different techniques were developed to assess enzyme attachment. The amount of bound protein was determinedusing Bradford dosage of proteins remaining in solution following CA II incubation with solid support. ELISA hasgiven a qualitative evaluation of the protein, enabling a follow up of enzyme binding robustness as a function of time. p-nitrophenyl acetate hydrolysis and CO2 hydration were assessed by spectrophotometry and electrometry, respectively.The percentage of active enzyme following immobilization was measured using an esterase linear model. Catalytictransformation rates for gaseous CO2 hydration were calculated for each type of immobilization.
Key words: immobilization, enzyme, esterase, hydration.
[Journal Translation] Belzil et Parent 77
Introduction
Dans son dernier rapport, le Groupe d’experts intergou-vernemental sur l’évolution du climat (GIEC 2001) démontreque la réduction des gaz à effet de serre est désormais un desdéfis auxquels est confrontée la communauté internationale.Le rapport GIEC indique une augmentation du CO2 de 31 %depuis 1750. La concentration actuelle de CO2 n’avait ja-mais atteint ce sommet au cours des 420 000 dernières an-nées. Le taux d’augmentation actuel est sans précédentdepuis au moins 20 000 ans.
Le dioxyde de carbone étant le principal gaz à effet deserre, de multiples avenues scientifiques et technologiquessont explorées ou reprises afin de mettre au point des procé-dés pour capter, séquestrer ou transformer à faible coût cegaz. Dans ce contexte, l’idée émise originalement par Mel-drum et Roughton (1933) d’utiliser l’anhydrase carboniquecomme catalyseur de ce procédé est actuellement reprise pardivers groupes parmi lesquels celui de Trachtenberg et Ge(2001) ainsi que celui de Bond et al. (2001).
L’utilisation des enzymes dans le contexte de la gestiondes gaz à effet de serre est envisagée par différents groupesde recherche (Bond et al. 2001; Jiang et al. 2002; Trachtenberget Ge 2001). L’immobilisation de l’anhydrase carbonique estattrayante comme technologie dans la transformation du CO2gazeux, que ce soit dans la gestion des gaz à effet de serre oudans le cadre de l’assainissement de l’air dans des environ-nements clos.
La réalisation d’un réacteur enzymatique visant à éliminerle CO2 gazeux des effluents industriels a été entreprise.L’enzyme, soit l’anhydrase carbonique humaine de type IIrecombinante (ACHIIr) immobilisée sur un support solidesert de catalyseur dans un réacteur de type colonne garnie.
Cet article présente les résultats des diverses méthodesd’analyse qui peuvent être utilisées pour contrôler la qualité
Biochem. Cell Biol. 83: 70–77 (2005) doi: 10.1139/O04-117 © 2005 CNRC Canada
70
Reçu le 5 mars 2004. Révision reçue le 13 septembre 2004.Accepté le 24 septembre 2004. Publié sur le site Web despresses scientifiques du CNRC à http://bcb.cnrc.ca le3 mars 2005.
A. Belzil1,2 et C. Parent. CO2 Solution Inc., 2300 rueEinstein, Parc Technologique du Québec Métropolitain,Sainte-Foy, QC G1P 4R1, Canada.
1. Auteur correspondant (courriel : [email protected]).2. Nouvelle adresse : 2300 rue Jean Perrin, Québec, QCG2C 1K8, Canada.

© 2005 CNRC Canada
Belzil et Parent 71
de l’enzyme immobilisée sur un support de polymère. Un telsupport peut être utilisé dans un réacteur de type colonnegarnie.
Étant donné que l’anhydrase carbonique permet la cata-lyse non seulement du gaz carbonique mais également celledu p-nitrophényl acétate, l’analyse de ces deux réactions aété effectuée. Les données recueillies, dans le cadre de ceprogramme, ont permis de déterminer l’état de la conforma-tion du site actif suite à l’immobilisation et aussi du degré demouillabilité du support. La qualification des immobilisa-tions chimiques est alors exprimée par une quantification dela masse d’enzymes liées, une évaluation de l’activité esté-rase et une mesure de l’activité d’hydratation du CO2 del’enzyme. L’intégration de ces différentes techniques au seind’une démarche industrielle de contrôle de la qualité etd’acceptation des lots d’enzymes immobilisées fera l’objetd’une future publication.
Matériel et méthodes
MatérielsL’anhydrase carbonique humaine de type II a été produite
au sein de l’entreprise CO2 Solution Inc. La ACHIIr a étéobtenue génétiquement par clonage. Elle avait un taux depurification supérieur à 90 % et une activité catalytiquecomparable à celle enregistrée pour l’anhydrase carboniquebovine commerciale dans les essais d’hydratation du CO2par la méthode décrite par le groupe Wilbur et Anderson(1948). Des anneaux Raschig de nylon 6,6 (6 mm de longpar 6 mm de large avec une lumière interne de 4 mm de dia-mètre) d’une surface moyenne de 220 mm2 constituent lesupport solide et provenaient de la compagnie GE PolymerShapes (Québec, Qc.). Les réactifs utilisés pour l’immobilisationchimique de l’enzyme provenaient de chez Sigma (Oakville,Ont.) (glutaraldéhyde, polyéthylènimide, acides aminés, NP40,caséine, p-nitrophényl acétate, orthophényl diamine et Tris).Les acides aminés utilisés étaient DL-acide aspartique, glycine,L-phénylanaline, D-leucine, L-arginine. Le méthanol, le chlorurede calcium et de sodium, l’acide chlorhydrique, le carbonateet bicarbonate de sodium et le phosphate de sodium mono-basique et dibasique provenaient de chez Lab-Mat (Québec,Qc.). L’acétonitrile a été acheté chez EM Science (Mont-Royal, Qc.). Les produits de quantification de protéines(Bio-assay Bradford) ont été achetés chez Bio-Rad (Missis-sauga, Ont.), ainsi que l’albumine bovine de sérum. Le gazcarbonique sous pression a été fourni par la compagniePraxair (Québec, Qc.). Les anticorps anti-ACH marqués à laperoxydase ont été achetés de la compagnie Research Dia-gnostics Inc. (Flanders, N.J.). Les mesures de spectrophoto-métrie ont été prises à l’aide d’un appareil Carry 100Bio deVarian. Les mesures des valeurs de pH pour électrométrieont été prises à l’aide du Sensorlink, et l’électrode était detype Électrodesure flow 8172BN de Orion.
Méthodes
Immobilisation chimique de l’enzymeLa technique d’immobilisation chimique a été développée
et adaptée à l’aide d’un système rotatif. L’utilisation d’un telsystème permet la réduction appréciable des réactifs d’oùune diminution des coûts de production.
Le support de nylon 6,6 doit d’abord être traité par une
solution acide (acide chlorhydrique 3,93 mol/L) pendant 1 hafin de générer des groupements aminés. La méthode uti-lisée était celle de Isgrove et al. (2001). La deuxième incu-bation du glutaraldéhyde (2,5 %) a été suivie de la liaisoncovalente de la ACHIIr à différentes concentrations (2; 1;0,75; 0,5 et 0,25 mg/mL). Les anneaux Raschig ont été lavésdans l’eau déionisée, et ensuite dans une solution de NaCl1 mol/L, puis lavés à nouveau trois fois à l’eau déioniséeavant d’être entreposés dans une solution de Tris 12 mmol/LpH 8,3. Tous les lavages entre les étapes ont été effectuésavec une eau déionisée.
Détermination de la concentration protéiqueLa quantité d’enzymes liées de façon covalente au support
a été déterminée selon deux méthodes. Une de celle-ci est undosage protéique dit Bradford pour la détermination de laquantité précise de protéines attachées au support. En sous-trayant la quantité de protéines résiduelles après immobilisa-tion et suite au premier lavage de la solution initiale, il aalors été possible d’estimer la quantité de ACHIIr sur cha-cun des anneaux Raschig. L’albumine de sérum bovine a étéutilisée comme standard dans la droite d’étalonnage.
La seconde méthode a permis un suivi quantitatif de laprotéine liée grâce à l’utilisation d’un anticorps anti-ACHhumaine marqué à la peroxydase. Cette technique, diteELISA, nous a permis une évaluation relative de la quantitéd’enzymes à la surface du support. Lors de l’utilisation dusupport activé dans les colonnes garnies, les essais ELISAont permis le suivi de la solidité du lien chimique soumisaux différents paramètres d’opération établis. Les anneauxRaschig ont été incubés, dans un premier temps, dans unesolution de caséine 1 % afin de bloquer les sites non spécifi-ques susceptibles de réagir avec l’anticorps anti-ACH. Suiteà un lavage dans un tampon phosphate 0,1 mol/L pH 7,4, lesanneaux Raschig ont été incubés en présence de 5 µg/mL del’anticorps anti-ACH marqué à la peroxydase pour 1 h à latempérature de la pièce. Suite à cinq lavages de tamponphosphate 0,1 mol/L pH 7,4 contenant NP40 à 0,5 %, lesubstrat orthophényl diamine a été ajouté à l’anneau Raschigsuite à sa dissolution dans l’eau nanopure selon les recom-mandations du fabricant. La réaction colorimétrique s’estdéveloppée durant 10 min sous une agitation constante. Lavaleur obtenue en spectrophotométrie à 490 nm a été estiméeen quantité protéique par rapport à une droite d’étalonnagede ACHIIr libre.
Détermination de l’activité estéraseLa méthode utilisée fût adaptée à partir de celle de Pocker
et Stone (1965). Celle-ci a été mise au point pour l’analysed’une enzyme immobilisée. Chaque anneau Raschig testé aété coupé en huit sections à l’aide d’un scalpel et placé dansune cuvette de spectrophotométrie de 4,5 mL. Le même lotd’enzymes ACHIIr utilisé pour l’immobilisation a été ajoutéà des cuvettes à raison de 0, 5, 10 et 20 µg afin d’établir unedroite d’étalonnage permettant l’évaluation de la quantitééquivalente d’enzymes actives liées de façon covalente surles anneaux Raschig.
Détermination de l’activité d’hydratation du CO2
Détermination du pourcentage de rendementLa technique pratiquée a été celle décrite par Wilbur et

© 2005 CNRC Canada
72 Biochem. Cell Biol. Vol. 83, 2005
Anderson (1948). La méthode a été modifiée afin de per-mettre la mesure d’activité de l’enzyme immobilisée sur unsupport solide. Le tampon utilisé a été le Tris 12 mmol/Ld’une valeur de pH de 8,3 à 25 °C (8,7 à 1 °C). L’essai a étéeffectué à 1 °C, le substrat provient d’une eau nanopure sa-turée de CO2, les essais ont été effectués avec 200 000 ppmde CO2 gazeux dans la chambre réactionnelle. Le CO2 ga-zeux passe à l’état aqueux pour former avec H2O l’acidecarbonique H2CO3 dissociée aussitôt en ions carbonate ethydrogène :
CO (g) CO (aq) + H O H CO
H CO H HCO
2 2 2 2 3
2 3+
3
↔ ↔
↔ + −
Comme l’indiquent les équations ci-dessus, la productiond’ions hydrogène entraîne une baisse de pH dans le milieu.L’utilisation du Tris comme tampon permet de lier les ionsH+ produits lors de la réaction d’hydratation du CO2 et ainside ralentir la réaction de chute de pH. De ce fait, il devientpossible de mesurer l’activité de l’enzyme.
Le temps de réaction requis pour que l’anneau Raschigcontenant ou non de l’enzyme passe du pH 8,7 à un pH de6,8 a été enregistré à l’aide du programme informatisé Sen-sorlink. Un pourcentage de rendement a été calculé à partirdu delta temps entre le support témoin négatif et celui avecenzyme. L’équation est la suivante :
[1] % de rendement = − ×[( )/ ]T T T0 1 0 100
où To est le temps requis au support témoin négatif pour unechute d’une valeur du pH maximale à une valeur de pH de6,8; et T1 est le temps requis au support avec enzymes pourune chute d’une valeur du pH maximale à une valeur de pHde 6,8.
Il convient ici de souligner les limites de cette méthode.L’enzyme libre atteint un maximum de 85 % compte tenudes contraintes de l’appareil (temps de réponse du systèmeet effet tampon du Tris). Ainsi, il n’est pas possibled’atteindre un rendement de 100 %. Lorsque l’enzyme estimmobilisée sur un support solide, celle-ci ne permet pasune répartition parfaite comme celle qui est retrouvée dansla chambre réactionnelle lors des essais avec l’enzyme libre.Il s’ensuit une diminution du rendement de l’activité.
Détermination du taux de transformation du substrat del’enzyme
La technique pratiquée pour la détermination du taux detransformation a été celle décrite précédemment, à l’exceptionde la solution de Tris utilisée, qui était d’une concentrationfinale de 6 mmol/L et son pH n’était pas ajusté. Cette modi-fication a permis de suivre la réaction à partir des valeurs depH de 10,0 jusqu’à des valeurs de 6,8. Les essais avec lesanneaux Raschig témoin négatif ont permis d’évaluer la vi-tesse réactionnelle naturelle de la transformation du CO2 dis-sous en ions bicarbonate et ions hydrogène. Le temps deréaction a été enregistré avec le même équipement que celuiqui a été utilisé pour l’évaluation du rendement.
Le taux de transformation du substrat (molécules de CO2)par unité de temps et par molécules d’enzymes a nécessité lamesure du temps requis pour baisser le pH d’une valeur de0,3 (évaluée par la méthode d’hydratation du CO2 deWilbur–Anderson) et d’une méthode de titrimétrie pour
quantifier le nombre de molécules H+ libérées lors de latransformation du CO2 dissous. Cette dernière méthode a étéadaptée de Hatch (1991). Cette méthode d’analyse a permisune évaluation plus juste de l’activité d’hydratation du CO2et de la valeur de pH offrant l’activité optimale de l’enzyme.La titrimétrie s’est effectuée sur une même quantité d’anneauxRaschig d’un même lot. Le titrage par HCl 0,1 mol/L d’unesolution Tris 6 mmol/L final contenant les anneaux Raschig,a permis de quantifier le nombre d’ions H+ générés corres-pondant à la baisse du pH. Le volume de solution titré doitêtre identique au volume total retrouvé dans les essaisd’hydratation du CO2 (Tris et eau saturée en CO2). Dans latitrimétrie, la baisse de pH est due à l’ajout de l’acide chlor-hydrique contrairement à l’essai d’hydratation du CO2, où lachute de pH est attribuable à la transformation du CO2 enions H+ et HCO3
–. Cette titrimétrie a donc tenu compte desions hydrogène pouvant être capturés par le tampon et lesupport lui-même lors de l’évaluation de l’activité réelled’hydratation du CO2.
Le temps requis pour chaque baisse de 0,3 de valeur depH pour l’échantillon témoin négatif (Tz) et celui de l’enzyme(Ty) a été noté ainsi que la quantité d’ions H+ (IH+) évaluéeen titrimétrie. Le taux de transformation de l’enzyme a étéobtenu par la soustraction des quotients de chacune des va-leurs obtenues suite à la division du nombre d’ions H+ requispour passer d’un pH à l’autre et du temps requis pour effec-tuer ce changement. En soustrayant le quotient des anneauxRaschig avec enzymes par le quotient des anneaux Raschigtémoin négatif, la quantité d’ions générés seulement parl’enzyme dans notre système est alors connu. En divisantpar la quantité d’enzymes actives estimées dans les essaisestérase et tenant compte du nombre d’anneaux Raschig uti-lisés, le taux de transformation (Tf) des molécules d’ions H+
par molécules d’enzymes par minute est alors obtenu.L’équation suivante a permis de connaître le taux de trans-
formation de l’enzyme par minute pour chaque valeur de pHet ainsi connaître également le pH auquel l’enzyme libre ouimmobilisée a la plus grande activité.
Le taux de transformation (Tf) de l’enzyme a été représentéen molécules d’ions H+ par minute par molécule d’enzymeet est égal à :
[2] Tf =IH IH /
Molécules d’enzymes actives
+/T Ty Z− +
où IH+ est la quantité de H+ générés de pH (X) à pH (X – 0,3);Ty est le temps requis à l’échantillon avec enzyme selonl’essai modifié de Wilbur–Anderson pour passer d’un pH àun pH de 0,3 unités inférieures. Le temps est exprimé en mi-nutes et TZ est le temps requis à l’échantillon sans enzymeselon l’essai modifié de Wilbur-Anderson pour passer d’unpH à un pH de 0,3 unités inférieures. Le temps est repré-senté en minutes.
StatistiquesLes données découlent de cinq expériences indépendantes.
Chaque expérience est analysée pour chaque qualificationpar trois à six essais. La déviation standard est déterminéepour chaque essai effectué.
© 2005 CNRC Canada
Belzil et Parent 73
Résultats
Les immobilisations à des concentrations de solutiond’enzymes différentes ont montré des efficacités variablesd’attachement de la ACHIIr, d’activités estérase et d’hydratationdu CO2 entre elles.
Quantification protéiqueLa quantification protéique a été effectuée selon deux mé-
thodes, soit par dosage indirect de protéines résiduelles ouune estimation par ELISA. Chacune de ces méthodes a sesavantages et apporte des outils dans l’évaluation ou le suivide nos immobilisations.
Quantification protéique par dosage indirect du surnageantLes résultats obtenus par quantification protéique suite au
dosage indirect de la solution résiduelle ont été confirméspar la technique d’absorption atomique de dosage de zincpar un laboratoire indépendant (données non montrées).L’anhydrase carbonique étant une métalloprotéine portant enson sein un atome de zinc, la détermination de la quantité dezinc à la surface du support suite à un traitement à l’acidenitrique a permis de valider la technique par dosage indirect.
Un essai colorimétrique de type Bradford sur la solutionrésiduelle suite à l’immobilisation ainsi que le premier la-vage a permis d’estimer la quantité de protéines non fixées.La quantité de protéines liées a été évaluée en soustrayant laquantité de protéines non liées à la quantité protéique de lasolution initiale.
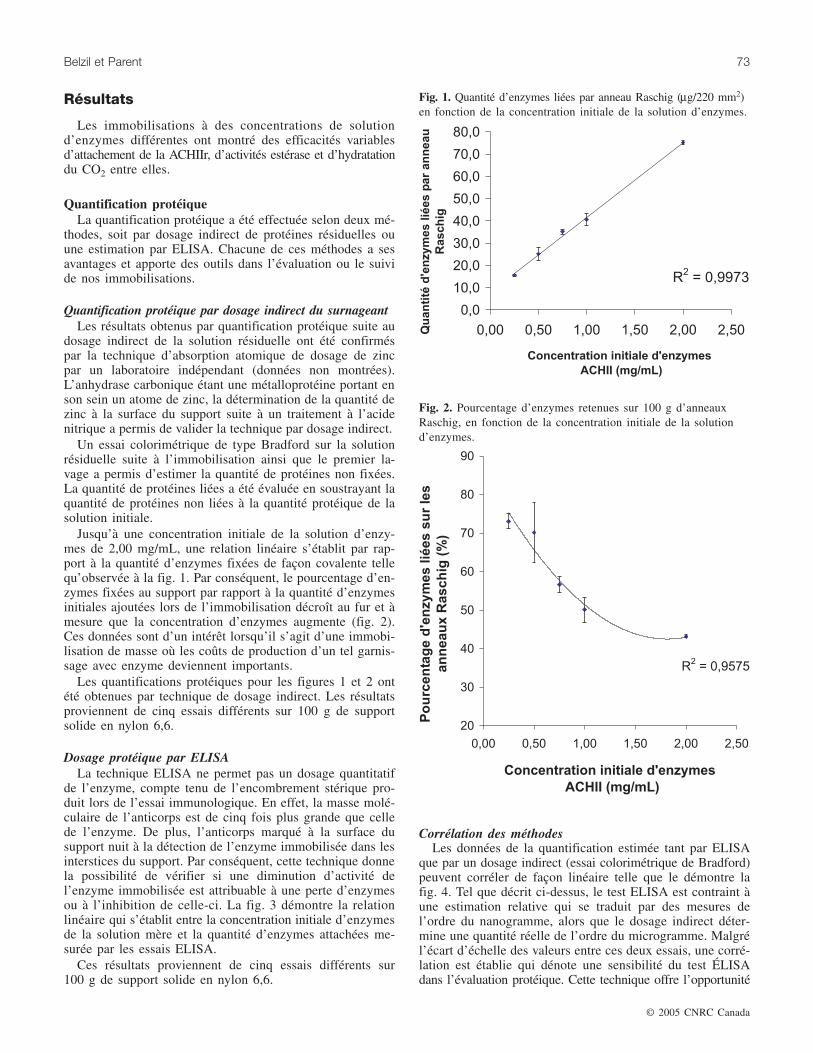
Jusqu’à une concentration initiale de la solution d’enzy-mes de 2,00 mg/mL, une relation linéaire s’établit par rap-port à la quantité d’enzymes fixées de façon covalente tellequ’observée à la fig. 1. Par conséquent, le pourcentage d’en-zymes fixées au support par rapport à la quantité d’enzymesinitiales ajoutées lors de l’immobilisation décroît au fur et àmesure que la concentration d’enzymes augmente (fig. 2).Ces données sont d’un intérêt lorsqu’il s’agit d’une immobi-lisation de masse où les coûts de production d’un tel garnis-sage avec enzyme deviennent importants.
Les quantifications protéiques pour les figures 1 et 2 ontété obtenues par technique de dosage indirect. Les résultatsproviennent de cinq essais différents sur 100 g de supportsolide en nylon 6,6.
Dosage protéique par ELISALa technique ELISA ne permet pas un dosage quantitatif
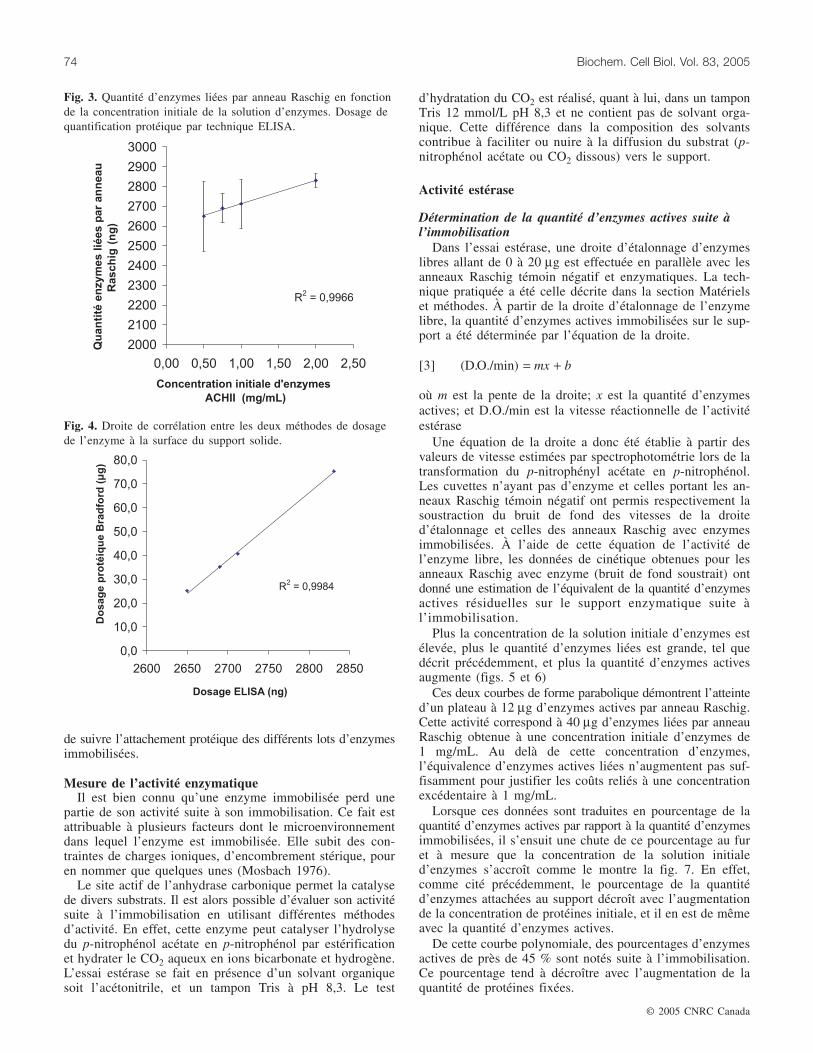
de l’enzyme, compte tenu de l’encombrement stérique pro-duit lors de l’essai immunologique. En effet, la masse molé-culaire de l’anticorps est de cinq fois plus grande que cellede l’enzyme. De plus, l’anticorps marqué à la surface dusupport nuit à la détection de l’enzyme immobilisée dans lesinterstices du support. Par conséquent, cette technique donnela possibilité de vérifier si une diminution d’activité del’enzyme immobilisée est attribuable à une perte d’enzymesou à l’inhibition de celle-ci. La fig. 3 démontre la relationlinéaire qui s’établit entre la concentration initiale d’enzymesde la solution mère et la quantité d’enzymes attachées me-surée par les essais ELISA.
Ces résultats proviennent de cinq essais différents sur100 g de support solide en nylon 6,6.
Corrélation des méthodesLes données de la quantification estimée tant par ELISA
que par un dosage indirect (essai colorimétrique de Bradford)peuvent corréler de façon linéaire telle que le démontre lafig. 4. Tel que décrit ci-dessus, le test ELISA est contraint àune estimation relative qui se traduit par des mesures del’ordre du nanogramme, alors que le dosage indirect déter-mine une quantité réelle de l’ordre du microgramme. Malgrél’écart d’échelle des valeurs entre ces deux essais, une corré-lation est établie qui dénote une sensibilité du test ÉLISAdans l’évaluation protéique. Cette technique offre l’opportunité
Fig. 1. Quantité d’enzymes liées par anneau Raschig (µg/220 mm2)en fonction de la concentration initiale de la solution d’enzymes.
Fig. 2. Pourcentage d’enzymes retenues sur 100 g d’anneauxRaschig, en fonction de la concentration initiale de la solutiond’enzymes.
© 2005 CNRC Canada
74 Biochem. Cell Biol. Vol. 83, 2005
de suivre l’attachement protéique des différents lots d’enzymesimmobilisées.
Mesure de l’activité enzymatiqueIl est bien connu qu’une enzyme immobilisée perd une
partie de son activité suite à son immobilisation. Ce fait estattribuable à plusieurs facteurs dont le microenvironnementdans lequel l’enzyme est immobilisée. Elle subit des con-traintes de charges ioniques, d’encombrement stérique, pouren nommer que quelques unes (Mosbach 1976).
Le site actif de l’anhydrase carbonique permet la catalysede divers substrats. Il est alors possible d’évaluer son activitésuite à l’immobilisation en utilisant différentes méthodesd’activité. En effet, cette enzyme peut catalyser l’hydrolysedu p-nitrophénol acétate en p-nitrophénol par estérificationet hydrater le CO2 aqueux en ions bicarbonate et hydrogène.L’essai estérase se fait en présence d’un solvant organiquesoit l’acétonitrile, et un tampon Tris à pH 8,3. Le test
d’hydratation du CO2 est réalisé, quant à lui, dans un tamponTris 12 mmol/L pH 8,3 et ne contient pas de solvant orga-nique. Cette différence dans la composition des solvantscontribue à faciliter ou nuire à la diffusion du substrat (p-nitrophénol acétate ou CO2 dissous) vers le support.
Activité estérase
Détermination de la quantité d’enzymes actives suite àl’immobilisation
Dans l’essai estérase, une droite d’étalonnage d’enzymeslibres allant de 0 à 20 µg est effectuée en parallèle avec lesanneaux Raschig témoin négatif et enzymatiques. La tech-nique pratiquée a été celle décrite dans la section Matérielset méthodes. À partir de la droite d’étalonnage de l’enzymelibre, la quantité d’enzymes actives immobilisées sur le sup-port a été déterminée par l’équation de la droite.
[3] (D.O./min) = +mx b
où m est la pente de la droite; x est la quantité d’enzymesactives; et D.O./min est la vitesse réactionnelle de l’activitéestérase
Une équation de la droite a donc été établie à partir desvaleurs de vitesse estimées par spectrophotométrie lors de latransformation du p-nitrophényl acétate en p-nitrophénol.Les cuvettes n’ayant pas d’enzyme et celles portant les an-neaux Raschig témoin négatif ont permis respectivement lasoustraction du bruit de fond des vitesses de la droited’étalonnage et celles des anneaux Raschig avec enzymesimmobilisées. À l’aide de cette équation de l’activité del’enzyme libre, les données de cinétique obtenues pour lesanneaux Raschig avec enzyme (bruit de fond soustrait) ontdonné une estimation de l’équivalent de la quantité d’enzymesactives résiduelles sur le support enzymatique suite àl’immobilisation.
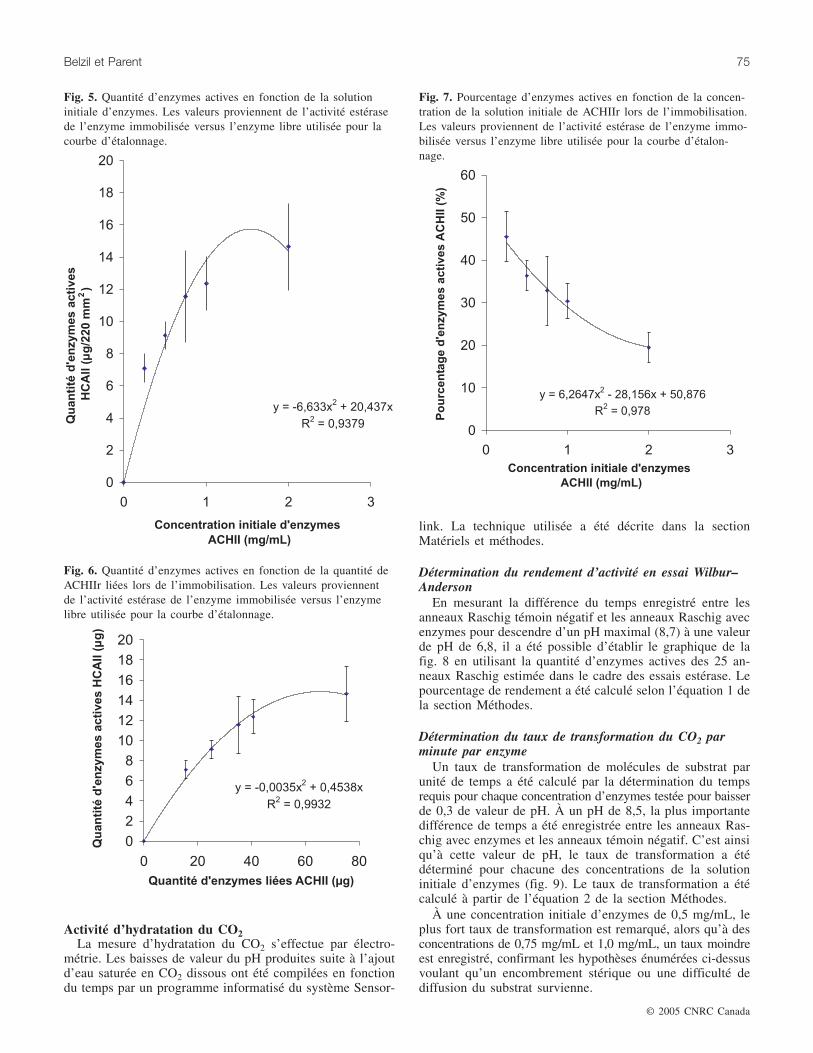
Plus la concentration de la solution initiale d’enzymes estélevée, plus le quantité d’enzymes liées est grande, tel quedécrit précédemment, et plus la quantité d’enzymes activesaugmente (figs. 5 et 6)
Ces deux courbes de forme parabolique démontrent l’atteinted’un plateau à 12 µg d’enzymes actives par anneau Raschig.Cette activité correspond à 40 µg d’enzymes liées par anneauRaschig obtenue à une concentration initiale d’enzymes de1 mg/mL. Au delà de cette concentration d’enzymes,l’équivalence d’enzymes actives liées n’augmentent pas suf-fisamment pour justifier les coûts reliés à une concentrationexcédentaire à 1 mg/mL.
Lorsque ces données sont traduites en pourcentage de laquantité d’enzymes actives par rapport à la quantité d’enzymesimmobilisées, il s’ensuit une chute de ce pourcentage au furet à mesure que la concentration de la solution initialed’enzymes s’accroît comme le montre la fig. 7. En effet,comme cité précédemment, le pourcentage de la quantitéd’enzymes attachées au support décroît avec l’augmentationde la concentration de protéines initiale, et il en est de mêmeavec la quantité d’enzymes actives.
De cette courbe polynomiale, des pourcentages d’enzymesactives de près de 45 % sont notés suite à l’immobilisation.Ce pourcentage tend à décroître avec l’augmentation de laquantité de protéines fixées.
Fig. 3. Quantité d’enzymes liées par anneau Raschig en fonctionde la concentration initiale de la solution d’enzymes. Dosage dequantification protéique par technique ELISA.
Fig. 4. Droite de corrélation entre les deux méthodes de dosagede l’enzyme à la surface du support solide.
© 2005 CNRC Canada
Belzil et Parent 75
Activité d’hydratation du CO2La mesure d’hydratation du CO2 s’effectue par électro-
métrie. Les baisses de valeur du pH produites suite à l’ajoutd’eau saturée en CO2 dissous ont été compilées en fonctiondu temps par un programme informatisé du système Sensor-
link. La technique utilisée a été décrite dans la sectionMatériels et méthodes.
Détermination du rendement d’activité en essai Wilbur–Anderson
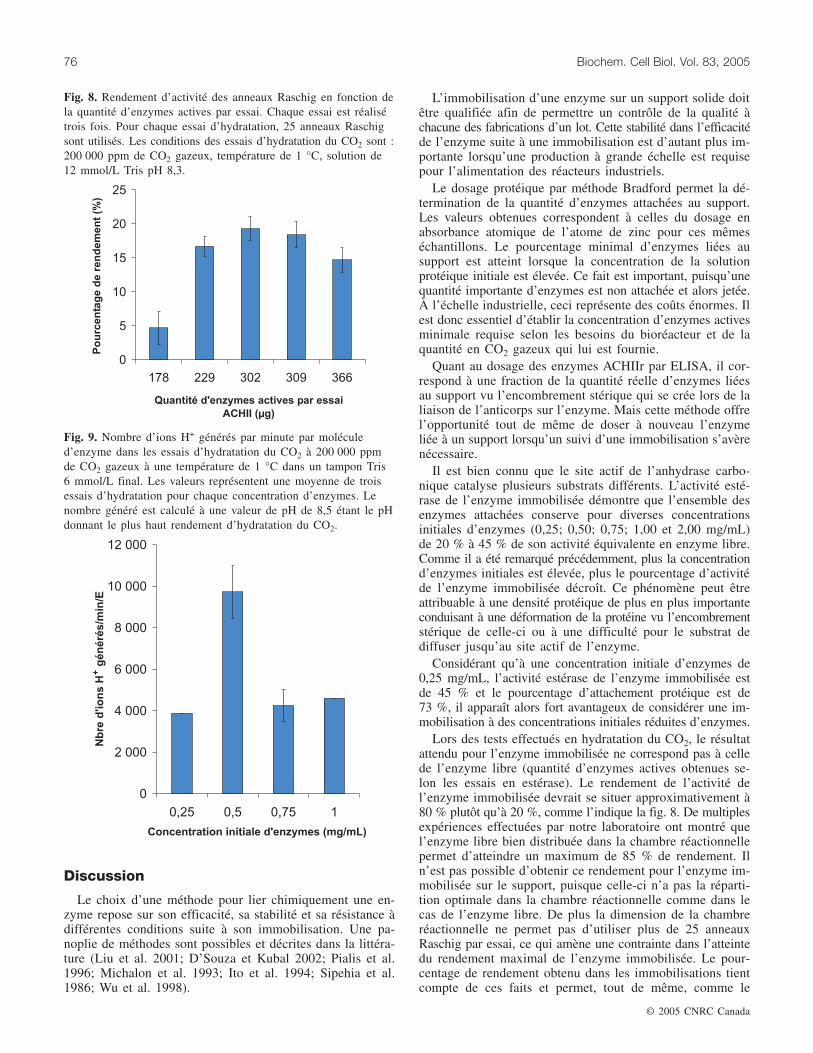
En mesurant la différence du temps enregistré entre lesanneaux Raschig témoin négatif et les anneaux Raschig avecenzymes pour descendre d’un pH maximal (8,7) à une valeurde pH de 6,8, il a été possible d’établir le graphique de lafig. 8 en utilisant la quantité d’enzymes actives des 25 an-neaux Raschig estimée dans le cadre des essais estérase. Lepourcentage de rendement a été calculé selon l’équation 1 dela section Méthodes.
Détermination du taux de transformation du CO2 parminute par enzyme
Un taux de transformation de molécules de substrat parunité de temps a été calculé par la détermination du tempsrequis pour chaque concentration d’enzymes testée pour baisserde 0,3 de valeur de pH. À un pH de 8,5, la plus importantedifférence de temps a été enregistrée entre les anneaux Ras-chig avec enzymes et les anneaux témoin négatif. C’est ainsiqu’à cette valeur de pH, le taux de transformation a étédéterminé pour chacune des concentrations de la solutioninitiale d’enzymes (fig. 9). Le taux de transformation a étécalculé à partir de l’équation 2 de la section Méthodes.
À une concentration initiale d’enzymes de 0,5 mg/mL, leplus fort taux de transformation est remarqué, alors qu’à desconcentrations de 0,75 mg/mL et 1,0 mg/mL, un taux moindreest enregistré, confirmant les hypothèses énumérées ci-dessusvoulant qu’un encombrement stérique ou une difficulté dediffusion du substrat survienne.
Fig. 5. Quantité d’enzymes actives en fonction de la solutioninitiale d’enzymes. Les valeurs proviennent de l’activité estérasede l’enzyme immobilisée versus l’enzyme libre utilisée pour lacourbe d’étalonnage.
Fig. 6. Quantité d’enzymes actives en fonction de la quantité deACHIIr liées lors de l’immobilisation. Les valeurs proviennentde l’activité estérase de l’enzyme immobilisée versus l’enzymelibre utilisée pour la courbe d’étalonnage.
Fig. 7. Pourcentage d’enzymes actives en fonction de la concen-tration de la solution initiale de ACHIIr lors de l’immobilisation.Les valeurs proviennent de l’activité estérase de l’enzyme immo-bilisée versus l’enzyme libre utilisée pour la courbe d’étalon-nage.
© 2005 CNRC Canada
76 Biochem. Cell Biol. Vol. 83, 2005
Discussion
Le choix d’une méthode pour lier chimiquement une en-zyme repose sur son efficacité, sa stabilité et sa résistance àdifférentes conditions suite à son immobilisation. Une pa-noplie de méthodes sont possibles et décrites dans la littéra-ture (Liu et al. 2001; D’Souza et Kubal 2002; Pialis et al.1996; Michalon et al. 1993; Ito et al. 1994; Sipehia et al.1986; Wu et al. 1998).
L’immobilisation d’une enzyme sur un support solide doitêtre qualifiée afin de permettre un contrôle de la qualité àchacune des fabrications d’un lot. Cette stabilité dans l’efficacitéde l’enzyme suite à une immobilisation est d’autant plus im-portante lorsqu’une production à grande échelle est requisepour l’alimentation des réacteurs industriels.
Le dosage protéique par méthode Bradford permet la dé-termination de la quantité d’enzymes attachées au support.Les valeurs obtenues correspondent à celles du dosage enabsorbance atomique de l’atome de zinc pour ces mêmeséchantillons. Le pourcentage minimal d’enzymes liées ausupport est atteint lorsque la concentration de la solutionprotéique initiale est élevée. Ce fait est important, puisqu’unequantité importante d’enzymes est non attachée et alors jetée.À l’échelle industrielle, ceci représente des coûts énormes. Ilest donc essentiel d’établir la concentration d’enzymes activesminimale requise selon les besoins du bioréacteur et de laquantité en CO2 gazeux qui lui est fournie.
Quant au dosage des enzymes ACHIIr par ELISA, il cor-respond à une fraction de la quantité réelle d’enzymes liéesau support vu l’encombrement stérique qui se crée lors de laliaison de l’anticorps sur l’enzyme. Mais cette méthode offrel’opportunité tout de même de doser à nouveau l’enzymeliée à un support lorsqu’un suivi d’une immobilisation s’avèrenécessaire.
Il est bien connu que le site actif de l’anhydrase carbo-nique catalyse plusieurs substrats différents. L’activité esté-rase de l’enzyme immobilisée démontre que l’ensemble desenzymes attachées conserve pour diverses concentrationsinitiales d’enzymes (0,25; 0,50; 0,75; 1,00 et 2,00 mg/mL)de 20 % à 45 % de son activité équivalente en enzyme libre.Comme il a été remarqué précédemment, plus la concentrationd’enzymes initiales est élevée, plus le pourcentage d’activitéde l’enzyme immobilisée décroît. Ce phénomène peut êtreattribuable à une densité protéique de plus en plus importanteconduisant à une déformation de la protéine vu l’encombrementstérique de celle-ci ou à une difficulté pour le substrat dediffuser jusqu’au site actif de l’enzyme.
Considérant qu’à une concentration initiale d’enzymes de0,25 mg/mL, l’activité estérase de l’enzyme immobilisée estde 45 % et le pourcentage d’attachement protéique est de73 %, il apparaît alors fort avantageux de considérer une im-mobilisation à des concentrations initiales réduites d’enzymes.
Lors des tests effectués en hydratation du CO2, le résultatattendu pour l’enzyme immobilisée ne correspond pas à cellede l’enzyme libre (quantité d’enzymes actives obtenues se-lon les essais en estérase). Le rendement de l’activité del’enzyme immobilisée devrait se situer approximativement à80 % plutôt qu’à 20 %, comme l’indique la fig. 8. De multiplesexpériences effectuées par notre laboratoire ont montré quel’enzyme libre bien distribuée dans la chambre réactionnellepermet d’atteindre un maximum de 85 % de rendement. Iln’est pas possible d’obtenir ce rendement pour l’enzyme im-mobilisée sur le support, puisque celle-ci n’a pas la réparti-tion optimale dans la chambre réactionnelle comme dans lecas de l’enzyme libre. De plus la dimension de la chambreréactionnelle ne permet pas d’utiliser plus de 25 anneauxRaschig par essai, ce qui amène une contrainte dans l’atteintedu rendement maximal de l’enzyme immobilisée. Le pour-centage de rendement obtenu dans les immobilisations tientcompte de ces faits et permet, tout de même, comme le
Fig. 8. Rendement d’activité des anneaux Raschig en fonction dela quantité d’enzymes actives par essai. Chaque essai est réalisétrois fois. Pour chaque essai d’hydratation, 25 anneaux Raschigsont utilisés. Les conditions des essais d’hydratation du CO2 sont :200 000 ppm de CO2 gazeux, température de 1 °C, solution de12 mmol/L Tris pH 8,3.
Fig. 9. Nombre d’ions H+ générés par minute par moléculed’enzyme dans les essais d’hydratation du CO2 à 200 000 ppmde CO2 gazeux à une température de 1 °C dans un tampon Tris6 mmol/L final. Les valeurs représentent une moyenne de troisessais d’hydratation pour chaque concentration d’enzymes. Lenombre généré est calculé à une valeur de pH de 8,5 étant le pHdonnant le plus haut rendement d’hydratation du CO2.

© 2005 CNRC Canada
Belzil et Parent 77
montre la fig. 8, une qualification selon la quantité d’enzymesactives retenues au support.
Un autre facteur intervient dans la détermination du ren-dement d’activité de l’enzyme immobilisée. Il s’agit dufaible transfert de matière entre le substrat, CO2 aqueux, etle support solide (Ramachandran et Chaudhari 1983) quipourrait aussi expliquer cet écart d’activité entre les deux es-sais. La difficulté pour le CO2 aqueux d’accéder au site actifde l’enzyme dépend de plusieurs facteurs tels que le carac-tère hydrophobe et le degré de mouillabilité du support. Cedernier joue un rôle prépondérant dans la capacité dusubstrat à atteindre le site actif de l’enzyme (Wang et al.2002). Les anneaux Raschig de nylon 6,6 sont constitués depolymère à caractère hydrophobe. Le CO2 se dissout dansl’eau et nécessite un environnement hydrophile pour migrerjusqu’à l’enzyme. Suite à l’hydrolyse du support, des grou-pements aminés et carboxylés sont créés contribuant ainsi àaccroître la mouillabilité du support. En améliorant la mouil-labilité du support, il sera possible de vérifier si la diffusiondu substrat, le CO2 dissous, est favorisée à la périphérie dusite actif.
La détermination du taux de transformation du CO2 parminute par molécule d’enzyme est un bon indicateur de lavitesse réelle de l’enzyme à différentes valeurs de pH. Parcette méthode, il est possible de déterminer la valeur de pHoptimale de catalyse pour un type d’immobilisation donnée.Dans le contexte de l’immobilisation de notre enzyme sur unsupport de nylon 6,6, la valeur du pH optimale (8,5) del’enzyme immobilisée est comparable à la valeur de pH del’enzyme libre soit 8,8 (données non montrées).
De plus, des tests de stabilité démontrent que l’activité del’enzyme immobilisée est conservée pour une période d’aumoins 1 an lorsque les supports avec enzymes sont entreposésà 4, 20 ou 37 °C en présence d’un agent bactériostatique(données non montrées). L’immobilisation de l’enzyme telleque décrite ici confère à l’enzyme un avantage non négli-geable si l’on tient compte du fait que le support avec en-zyme peut être réutilisé sur une très longue période de tempssans perte d’activité.
Toutes ces méthodes visant à contrôler la qualité de notreproduit, soit l’enzyme immobilisée sur un support solide,s’appliquent à différents types d’immobilisation (covalente,par absorption, cross-link, etc.) et à divers supports que cesoit des polymères synthétiques (polyéthylène, nylon,polystyrène) ou naturelles (cellulose, etc.). Toutes ces quali-fications d’une immobilisation d’enzymes permettent nonseulement le suivi de l’efficacité de la catalyse dans le tempsmais aussi de cerner les paramètres importants pour l’obtentionde la meilleure immobilisation : soit celle qui tient compted’une bonne efficacité et d’un coût moindre pour une pro-duction industrielle.
Remerciements
Nous remercions M. Pierre-Mathieu Charest de l’UniversitéLaval pour ses judicieux conseils et M. Richard Lacasse etMme. Lucie Labbé pour leur assistance technique et leurdévouement. Ce programme de recherche a été entièrementfinancé par CO2 Solution Inc.
RéférencesBond, G.M., Stringer, J., Brandvold, D.K., Simsek, F.A., Medina,
M.G., et Egeland, G. 2001. Development of integrated systemfor biomimetic CO2 sequestration using the enzyme carbonicanhydrase. Energy Fuels, 15 : 309–316.
D’Souza, S.F., et Kubal B.S. 2002. A cloth strip bioreactor withimmobilized glucoamylase. J. Biochem. Biophys. Methods, 51 :151–159.
GIEC (Groupe d’experts intergouvernemental sur l’évolution duclimat). 2001. Des changements climatiques : les éléments scienti-fiques. Rapport du Groupe de travail 1 du Groupe d’expertsintergouvernemental sur l’évolution du climat, Cambridge Uni-versity Press, Canbridge, R.-U.
Hatch, M.D. 1991. Carbonic anhydrase assay: strong inhibition ofthe leaf enzyme by CO2 in certain buffers. Anal. Biochem. 192 :85–89.
Isgrove, F.H., Williams, R.J., Niven, G.W., et Andrews, A.T. 2001.Enzyme immobilization on nylon-optimization and the steps usedto prevent enzyme leakage from the support. Enzyme Microb.Technol. 28(2–3) : 225–232.
Ito, Y., Fujii, H., et Imanishi, Y. 1994 Modification of lipase withvarious synthetic polymers and their catalytic activities in organicsolvant. Biotechnol. Prog. 10 : 398–402.
Jiang, Z., Wu, H., Xu, S., et Huang, S. 2002. Enzymatic conversionof carbon dioxide to methanol by dehydrogenases encapsulatedin sol-gel matrix. Fuel Chem. Div. Preprints, 47(1) : 306.
Liu, J.L., Wang, J.Q., Bachas, L.G., et Bhattacharyya, D. 2001.Activity studies of immobilized subtilisin on functionalized purecellulose-based membranes. Biotechnol. Prog. 17(5) : 866–871.
Meldrum, N.U., et Roughton, F.J.W. 1933. Carbonic anhydrase: itspreparation and properties. J. Physiol. 80 : 113–141.
Michalon, P., Roche, J., Couturier, R., Favre-Bonvin, G., etMarion, C. 1993. DNase activity of micrococcal endonucleasecovalently immobilized on nylon and polystyrene. EnzymeMicrob. Technol. 15 : 215–221.
Mosbach, R., Koch-Schmidt, A.C., et Mosbach, K. 1976. Immobi-lization of enzymes to various acrylic copolymer. MethodsEnzymol. 44 : 53–65.
Pialis, P., Hamann, M.C.J., et Savill, B.A. 1996. L-DOPA productionfrom tyrosinase immobilized on nylon 6,6. Biotechnol. Bioeng.51 : 141–147.
Pocker, Y., et Stone, J.T. 1965. The catalytic versatility of erythrocytecarbonic anhydrase. The enzyme-catalysed hydrolysis of p-nitrophenyl acetate. J. Am. Chem. Soc. 87 : 5497–5498.
Ramachandran, P.A., et Chaudhari, R.V. 1983. Three phase catalyticreactors. Dans Topics in chemical engineering. Gordon andBreach Science Publisher, USA. Vol. 2, pp. 15–17.
Sipehia, R., Chawla, A.S., et Chang, T.M.S. 1986. Enhanced albuminbinging to polypropylene beads via anhydrous ammonia gaseousplasma. Biomaterials, 7 : 471–473.
Trachtenberg, M.C., et Ge, J. 2001. Enzyme-based hollow fibercontained liquid membrane for CO2 removal. AIChE Conférences,Nouvelle Orléans, Louisianne, 10–14 Mars 2002. AIChE CustomerService, New York.
Wilbur, K.M., et Anderson, N.G. 1948. Electrometric and colorimetricdetermination of carbonic anhydrase. J. Biol. Chem. 176 : 147–154.
Wang, J., Buck, S.M., Even, M.A., et Chen, Z. 2002. Molecularresponses of proteins at different interfacial environmentsdetected by sum frequency generation vibrational spectroscopy.J. Am. Chem. Soc. 124(44) : 13302–13305.
Wu, C.-W., Lee, J.-G., et Lee, W.-C. 1998. Protein an enzymeimmobilization on non-porous microspheres of polystyrene.Biotechnol. Appl. Biochem. 27 : 225–230.
























![Rapport Immobilisations Corporelles[1]](https://img.pdfslide.fr/doc/110x75/577cdf441a28ab9e78b0d4a8/rapport-immobilisations-corporelles1.jpg)


