Embed Size (px)
Citation preview

© 1971 by Academic Press, Inc.
J. ULTRASTRUCTURE RESEARCH 37, 449-478 (1971) 449
Microf i laments et microtubules nucl6aires
et organisation ul t rastructurale des batonnets intranucl6aires
des neurones sympathiques ~
RAYMOND SE'ITE, JACQUES ESCAIG ET SUZANNE COUINEAU
Groupe de Recherches en Neurobiologie, Laboratoire d'Histologie I, FacultO de MOdecine, 13, Marseille (5e), France
et Laboratoire de Microscopie Electronique appliqude ?t la Biologie (C.N.R.S.), Bd Raspail, Paris 6 e, France
Refu le 10 mai 1971
Les inclusions nucl6aires des neurones sympathiques, assimilables aux (~ briton- nets intranucldaires >> sont prineipalement, sinon exclusivement, de nature pro- tdique. Elles offrent une extreme vari6t6 d'aspects dans les conditions habi- tuelles d'observation au microscope 61ectronique. En fait l'6tude de plusieurs centaines d'inclusions et l'emploi d'une platine d'observation goniom6trique permettent de montrer qu'elles sont constitu6es par deux 616ments fonda- mentaux : des microfilaments et des microtubules. Ces deux structures 616men- taires, consid6r6es comme des constituants nucl6aires normaux, donnent naissance ~ des inclusions filamenteuses et tubulo-filamenteuses. Ces derni6res se prdsentent soit sous la forme d 'un fuseau de microfilaments parall61es doubl6 par une couronne de microtubules, soit sous la forme de cristalloides tr~s com- plexes dans lesquels les microfilaments sont rang6s en couches (12 le plus sou- vent) ddcal6es de 60 °. Un corps granulaire satellite commun ~t toutes les in- clusions, une coque granulaire et un court faisceau tubulaire accessoire associ6s aux inclusions tubulo-filamenteuses ont 6t6 6galement mis en 6vidence.
Ce travail est discut6 dans le cadre des probl6mes pos6s par les prot6ines fibreuses neuronales et des hypoth6ses sont 6mises sur la dynamique des inter- relations entre les diff6rentes varidt6s d'inclusions.
Dans le noyau intercin6tique la microscopie 61ectronique a permis de mettre en
6vidence diverses entit6s morphologiques, et sans doute fonctionnelles, rangfies sous
la terminologie purement descriptive ~ d'inclusions nucl6aires >> (34, 38). L'un de nous
a montr6 qu'il en existe plusieurs vari6t6s dans les neurones sympathiques (34). Le pr6sent travail est consacr6 ~t l'6tude d'inclusions de grande taille, d6crites g6n6-
ralement sous le terme de ~ faisceau fibrillaire >> ou de ~ cristaUoide >> et assimil6es par
z Travail effectu~ avec l'aide mat6rielle et technique du C.N.R.S.

450 SEITE, ESCAIG ET COUINEAU
Siegesmund et coll. (36) aux << b~ttonnets intranucl6aires >~ mis en 6vidence dans divers neurones par les neurohistologistes du si6cle dernier.
A la suite des premieres observations de Siegesmund et coll. (36), on pouvait esp6rer
que les recherches ultrastructurales permettraient de r6pondre aux probl6mes con-
cernant la nature et la signification biologique de ce type d'inclusion. A l 'heure
actuelle, il est en r6alit6 tr6s difficile de fournir ces r6ponses pour au moins trois
raisons essentielles (34) :
- - ces inclusions sont consid6r6es comme 6tant rares ou m~me exceptionnelles par la
plupart des auteurs de telle sorte que les observations out 6t6 lc plus souvent for-
tuites; - - elles sont tr6s polymorphes et paraissent repr6senter des entit6s ultrastructurales
distinctes qu'il est difficile de classer;
- - elles seraient rencontr6es plus fr6quemment et parfois exclusivement au cours de
diff6rents 6tats pathologiques ce qui laisse un doute important quant ~t leur signi-
fication physiologique.
Des observations r6centes que nous avons pu faire sur des ganglions stellaires
soumis ~ une stimulation 61ectrique nous ont condui t / t reconsid6rer enti~rement cette
question 6nigmatique des bfitonnets intranucldaires de la cellule nerveuse dans le
double but de d6terminer leur organisation ultrastructurale et d 'appor ter un 616ment de r6ponse au probl~me de leur signification biologique.
Le problbme de leur organisation ultrastructurale, qui fait l 'objet du pr6sent
travail, sera envisag6/t partir de l '6tude syst6matique de plus de 15 000 sections de noyaux en microscopie 61ectronique ordinaire ou /l l 'aide d 'un microscope 61ectro-
nique 6quip6 d 'une platine d 'observat ion goniomdtrique. Ce travail compor tera
6galement une br~ve 6tude cytochimique.
MATERIEL ET METHODES
Nous avons utilis6 les ganglions stellaires de 23 Chats (dont deux femelles), vaccin6s, d 'un poids compris entre 3 Kg et 4 Kg. Ces animaux out 6t6 anesth6si6s au Nembutal par vole intra-veineuse it la dose de 25 mg/Kg apr~s une courte induction au fluothane. Les ganglions stellaires (soumis ou non it une stimulation 61ectrique) out 6t6 d6couverts et fix6s pendant quelques minutes par une solution ~, 4°C de glutarald6hyde it 2,5 % ~t pH 7,4, puis post-fix6s pendant uue heure par une solution de t6troxyde d'osmium 5, 2% 5 pH 7,4. L'inclusion a 6t6 faite en Epon ou dans un m61ange Epon-Araldite (2). Les coupes (blanches de 600/i 8019 A) out 6t6 contrastdes par l'ac6tate d'uranyl-citrate de plomb. Nous avons effectu6 des diges-
FIG. 1. Inclusion nucl6aire cristalloide sans localisation pr6fdrentielle par rapport au nucl6ole et /t l'enveloppe nucl6aire, x 30 000.
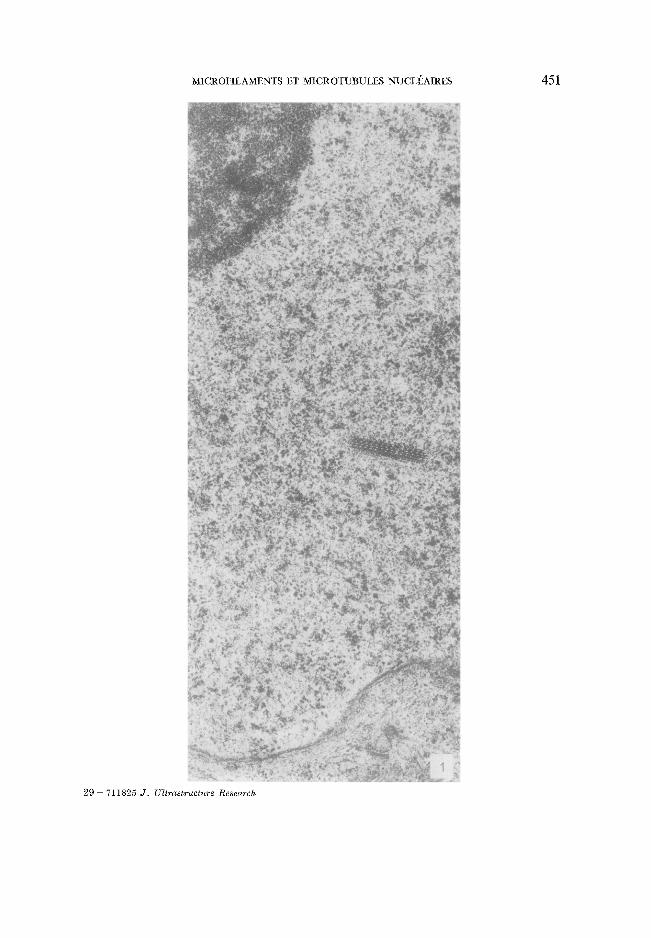
MICROFILAMENTS ET M I C R O T U B U L E S NUCLI~AIRES 451
29 - 711825 J . Ullrastructure Research

452 SEiTE, ESCAIG ET COUINEAU
tions enzymatiques aprOs double fixation et oxydation des coupes par l 'eau oxygOnOe ou l 'acide pOriodique sur du matOriel inclus en Epon selon la technique de Monneron et Bern- hard (25) ou en Epon-Araldi te selon la technique de Anderson et Andr6 (2). Les enzymes suivantes out 6t6 utilisOes :
- - Pronase (Worthington 2 x cristallisOe et Kaken Chemical, Tokyo) en solution aqueuse 5' 0,1% 5' p H 7,4 pendant 30 minutes 5' 4 heures (24);
- - Pepsine (Worthington 2 × cristaUisOe) en solution de 0,1% 5' 1% dans l 'acide chlor- hydrique 0,1 M pendant 1 5' 6 heures 5' 37°;
- - Trypsine (Worthington 2 x cristallisde) 5' 1% 5' p H 8 en prOsence de CaCI~; - - Ribonucldase (Worthington 2 × cristallisOe) en solution aqueuse 5' pH 6,8 pendant 1 5'
5 heures 5' 38 °.
La technique de Silverman et Glick (37), pour la raise en 6vidence des protOines par im- prOgnation des blocs 5' l 'acide phosphotungstique en solution aqueuse, a 6galemeut 6t6 appliquOe.
Les coupes out 6t6 examinOes aux microscopes EImiskop I et 101 de Siemens et EM 300 de Philips, ce dernier 6taut 6quip6 d 'une platine goniomOtrique qui permet d'effectuer une << bas- cule >~ de l 'objet de + 45 ° par rapport 5, la position d'origine de celui-ci. Grace 5' un nouveau dispositif d 'ant icontaminat ion, nous avons pu effectuer 6galement des observations sous des angles variant de + 60 °.
OBSERVATIONS
Ces obse rva t ions son t basses sur l 'Otude sys tdmat ique de 662 sect ions d ' i n c l u s i o n s
nuclOaires (15 616 sect ions de n o y au x ) observOes dans 43 gang l ions stellaires tOmoins
ou s o u m i s / t u n e s t imu la t i on 61ectrique.
I. ETUDE AU MICROSCOPE t~LECTRONIQUE ORDINAIRE
L ' inc idence de coupe 6 tant 6v ide rnment due au hasard , l 'Otude de ces sect ions d ' i n -
c lus ions nuclOaires n o u s a fou rn i u n e ex t reme variOt6 d ' images . Cer ta ines d ' en t r e elles
son t ana logues ou m~me iden t iques h des images d@rites dans d ' au t r e s terr i toires ou
dans les m~mes n e u r o n e s (34). Or, c o m m e n o u s le verrons , il est arriv6 f r d q u e m m e n t
que de telles images so ient interprdtOes de fagon erronOe et considdrdes gOnOralement
c o m m e des entitOs dist inctes. DOs lors il pa ra i t ind i spensab le , au prOalable, de d o n n e r
u n apergu de celles que n o u s avons observOes en microscopic 61ectronique o rd ina i r e
FIGs. 2/t 9. Images d'inclusions nuclOaires diffOrentes/t un triple point de vue. Loealisation : pros de l'enveloppe nucldaire (Figs. 2 h 4) juxtanucldolaire (Figs. 6 et 7) ou quelconque par rapport aux deux structures prOc6dentes (Figs. 5, 8 et 9). Aspect : trbs variable et pouvant 6tre interprdt6 comme fibril- laire (Fig. 2 et 6ventuellement Fig. 3), tubulaire (Fig. 4 et 6ventuellement 5 et 7) ce qui est en rOalit6 inexact, ou plus complexe (Figs. 6/t 9) avec une association de filaments et de tubules (Fig. 9). Taille : trOs variable. Noter la prOsence inconstante d'un court faisceau plus ou moins nettement tubulaire (Figs. 4, 8 et 9 -+), d 'un << corps satellite >> (Figs. 3 et 6 +--~) et d'une coque granulaire (Fig. 7 -+). Fig, 3 : ×20000;Fig. 6 : x 37 000; Figs. 2 , 4 , 5 , 7 : × 4 0 0 0 0 ; 8 e t 9 : xl00000.
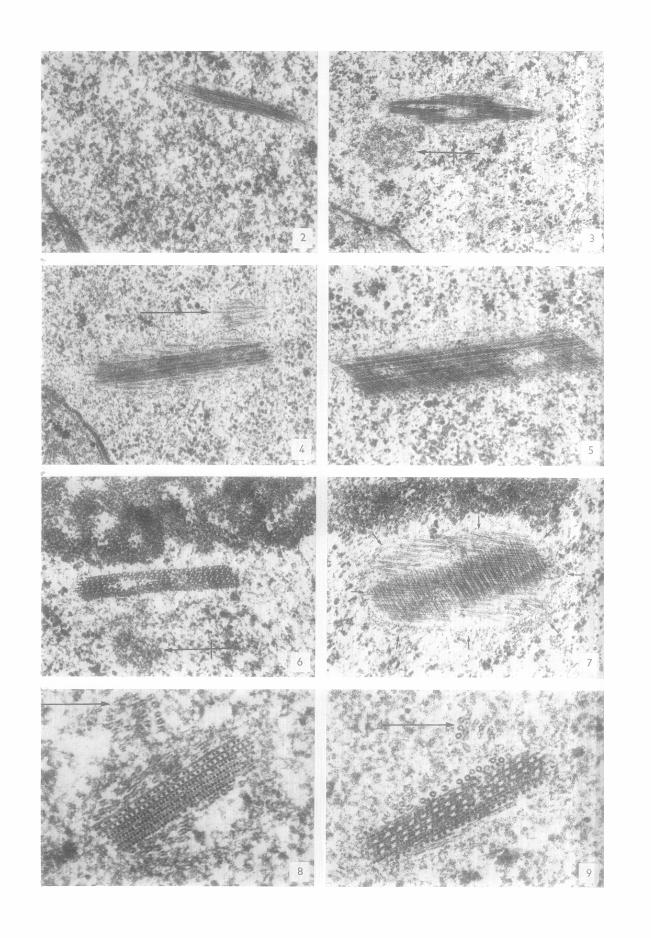

454 sz~xE, ESCAIG ET COUINEAU
(Figs. 1 /t 9), dans la mesure off le but principal de cette 6tude est d'en ddterminer la structure exacte.
Pour la commodit6 de l'expos6, nous distinguerons :
- - le corps central des inclusions; - - les structures assocides/t ce corps central.
A. Le corps central
Sur des coupes au hasard, la forme de ce corps central est tr~s variable : il peut
s'inscrire dans un fuseau (Figs. 2 et 3) ou dans des figures gdom6triques simples fr6- quemment rectangulaires (Figs. 4, 5, 6, 8 et 9).
Ses 616merits constitutifs paraissent ~tre soit des filaments dormant ~ l'inclusion 1'aspect de faisceau fibrillaire (Fig. 2), soit une association de filaments et de tubules, ces derniers n'6tant nettement d6finis que pour des incidences de coupes privil6gides (Fig. 9). Pour la majorit6 des inclusions, il est extr~mement difficile de lever l'incerti- tude et de se prononcer nettement en faveur de leur nature tubulaire ou filamenteuse
(Figs. 4, 5 et 7) ou encore de savoir si ces deux 61dments sont r6ellement associ6s
(Fig. 8). L'organisation interne de ce corps central peut se faire selon un groupement plus ou
moins parall61e des 616ments constitutifs (Fig. 4) ou au contraire selon un mode p6riodique qui leur donne nettement un aspect cristalloMe ou paracristallin (Figs. 5, 6, 8 et 9).
Darts cette partie de notre 6tude, il a 6t6 pratiquement impossible d'6tablir une iden- tit6 certaine entre deux images d'inclusions nucl6aires malgr6 l'6tude de plusieurs centaines de cas. De m~me, il n 'a pas 6t6 possible de savoir si filaments et tubules sont
toujours associ6s ou si, au contraire, il peut exister des inclusions purement filamen- reuses ou purement tubulaires.
B. Les structures assocides
En dehors des structures d'aspect plus ou moins franchement tubulaire qui sont rencontr6es parfois ~ la p6riph6rie du corps central (Fig. 7 ~t 9), nous avons pu noter la pr6sence, mais dans certains cas seulement :
- - d'une coque d'aspect finement granulaire qui para~t circonscrire le corps central (Fig. 7) et dont nous avions d6j~ not6 l'existence darts ces m~mes noyaux (34);
- - d'un court faisceau qui n 'apparai t tubulaire que pour certaines incidences privil6- gi6es (Fig. 4) et qui est situ6 h 0,1 # environ du corps central;
- - d'une structure que nous d6signons sous le terme de << corps granulaire satellite >>
(Figs. 3 et 6).

MICROFILAMENTS ET MICROTUBULES NUCLEAIRES 455
FI~. 10. Ext rac t ion partielle d ' une inclus ion juxta-nucl6olaire apr~s une digest ion de 30' par la pro- nase. Nucl. = nucl6ole, x 60 000.
L'ensemble constitu6 par le corps central et les structures associ6es pr6sente non seulement des diff6rences d'aspect et de forme, mais aussi des diff6rences de localisa- tion et de taille.
Quel que soit leur aspect, elles occupent une situation quelconque dans le nucl6o- plasme (Figs. 1, 5, 8 et 9). Certaines sont situ6es pr6s de l'enveloppe nucl6aire (Fig. 2 4), d'autres sont juxta-nucl6olaires (Figs. 6 et 7); leur orientation est 6galement quel- conque par rapport g ces deux structures.
Leur taille apparente est tr~s variable, car nous avons pu noter les valeurs extremes de 0,1 # et 4 #. En l'absence d'une s6rie complete de coupes s6ri6es d'une m~me in- clusion, il ne nous est pas possible de savoir, dans cette partie de notre 6tude, si ces valeurs extremes repr6sentent des variations rdelles de longueur des inclusions ou seulement des variations dues aux incidences de coupes, hypoth~se la plus vraisem- blable. En particulier, il n'est pas possible de considdrer la valeur extrame de 4 # comme 6tant la longueur maximum des inclusions filamenteuses en fuseau (Fig. 11), dans l'impossibilit6 o/1 nous sommes de savoir si la coupe est parfaitement longitudinale ou si l'inclusion est parfaitement rectiligne.
Ainsi il apparMt que, malgr6 l'6tude de plusieurs centaines d'inclusions nucl6aires, l'6tude en microscopie ~lectronique ordinaire ne permet pas, en raison de l'extrame diversit6 de leur aspect, de leur localisation et de leur taille, de d6finir exactement

456 SEITE~ ESCAIG ET COUINEAU
leur s tructure et la nature de leurs ElEments constitutifs. I1 est Egalement imposs ib le de
savoir si elles reprEscntent au tan t d'entitEs diffErentes ou si, au contraire , elles offrent
un certain nombre de caract6res communs . L ' impress ion est donc celle d ' un ensemble
tr6s d ispara te o/1 l ' on re t rouve non seulement la grande major i t6 des images d6crites
dans d 'au t res terr i toires pa r d ' au t res auteurs (cf. discussion), mais oh il appa ra i t
6galement un grand nombre d ' inclusions non observ6es jusqu ' ic i .
C. Etude cytochimique prdliminaire En ce qui concerne le p robl6me de la na ture de ces inclusions, nous r appor t e rons les
r6sultats que nous avons obtenus pa r la m6thode de Si lverman et Gl ick (37), con-
sid6r6e pa r ses auteurs comme spEcifique des protEines, et pa r diverses digest ions
enzymatiques selon la mEthode de M o n n e r o n et Bernhard (25) sur pi6ces incluses en
Epon ou de Ande r son et Andr6 (2) sur pi~ces incluscs en Epon-Ara ld i te .
- - L' imprEgnat ion des pibces pa r de l 'ac ide phospho tungs t ique en solut ion aqueuse
nous a donn6 un rEsultat positif . La densitE des inclusions nuclEaires est comparab le ,
pa r exemple, h celle de la matr ice des mi tochondr ies (dont les membranes ne sont pas
<< colorEes >0, des lysosomes ou des Mmat ies ; le nucl6ole pr6sente un contras te rela-
t ivement moins impor tan t .
- - La r ibonucl6ase utilisEe seule n ' a pas d 'effet sur les inclusions nuclEaires m~me
cn p ro longean t les temps d ' incuba t ion au-del/t de 2 heures.
- - La pronase nous a donne les r6sultats les plus rapides, sur tout sur des pi6ces in-
cluses en Epon-Ara ld i te . Son act ion est manifeste fi pa r t i r de 20 ~t 30 minutes (Fig. 10).
Au-del/ t elle p rovoque une trbs forte d iminu t ion de la densit6 de l ' ensemble des in-
clusions nuclEaires, mais il persiste une sorte de t r ame alors que d ' au t res s tructures ne
sont plus << color4es >> ou sont plus ou moins to ta lement extraites, comme les lyso-
somes. A 60' et 80', la d iminu t ion de contras te des inclusions nuclEaires s 'accentue (les
fibres de collag~ne sont alors dEtruites pour la p lupar t ) , mais l ' ex t rac t ion demeure
toujours incomplete . Au-del~t de ces temps, les coupes, et en par t icul ier le nucl6o-
plasme, sont trbs dElabrEs de telle sorte qu ' i l devient imposs ib le de savoir si les in-
clusions nuclEaires ont 6t6 effectivement extraites.
La r ibonuclEase appliquEe aprks la p ronase n ' a pas eu d 'ac t ion .
- - La pepsine nous a donne des rEsultats identiques, mais plus tardifs.
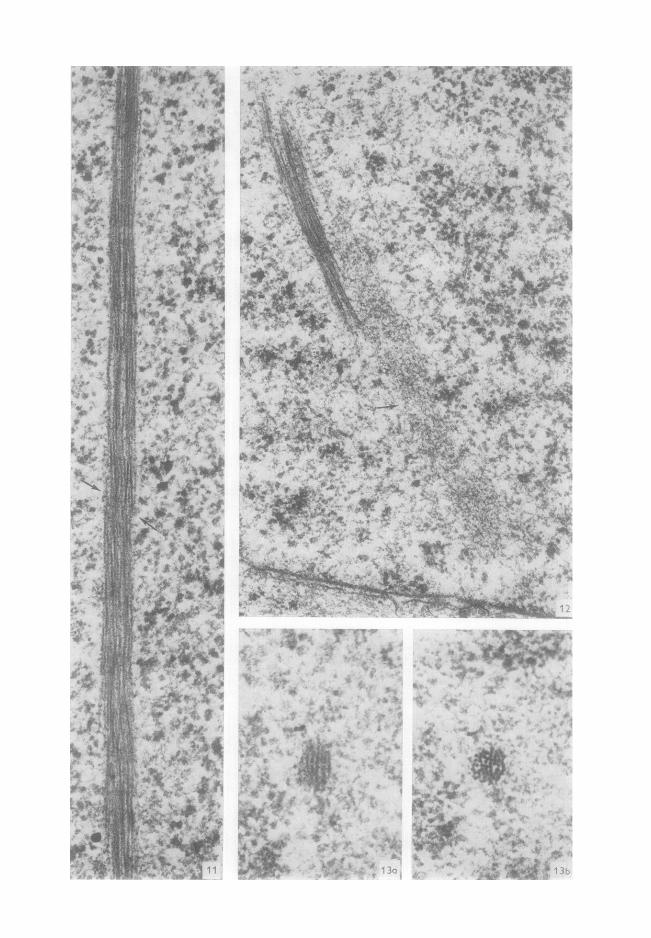
FIG. 11. Vue partielle d'une inclusion de 4/~ g aspect << fibrillaire >~. Noter la pr6sence de discontinuit~s (-+). x 70 000. FIG. 12. Rapport entre inclusion filamenteuse et << corps granulaire satellite >> (~) de forme allong6e. × 50 000.
FIG. 13. (a) Inclusion d'aspect << fibrillaire >> la coupe se trouvant dans les conditions normales d'ob- servation dans un plan perpendiculaire au faisceaux d'616ctrons. (b) La m&ne inclusion apr6s une bascule de 45 ° de la coupe. L'axe, autour duquel s'est fair cette bascule, est parall61e au c6t6 inf6rieur du clich6. L'inclusion est constitu6e exclusivement par des fila- ments parall61es, x 115 000.
J ~ ~
i~
i

458 SEITE, ESCAIG ET COU1NEAU
- - La t rypsine n ' a pas eu d ' ac t ion ou a m~me provoqu6, parfois , une augmenta t ion
de densit6 pa radoxa le de certaines structures telles que le nucl6ole.
En aucun cas, p a r cons6quent, nous ne sommes p a r v e n u s / t obteni r une extract ion
compl6 te , / t ~< l ' empor t e pi6ce >~, de ces inclusions nucl6aires sons Fact ion d 'enzymes
prot6olyt iques suivie ou non d 'une digest ion pa r la r ibonucl6ase.
II. ETUDE AU MICROSCOPE I~LECTRONIQUE EQUIPE D'UNE PLATINE D'OBSERVATION
GONIOMETRIQUE
Cette 6tude a eu p o u r objectif de ddfinir la nature et l ' o rgan isa t ion dans l 'espace
des 616ments consti tut ifs du corps central des inclusions et de savoir quels sont les
rappor t s , s ' il en existe, entre les diff6rentes images observdes dans les condi t ions
habituelles. L 'u t i l i sa t ion de la pla t ine d 'obse rva t ion goniom6tr ique nous a permis de
distinguer, pa rmi les inclusions ayan t la fo rme g6n6rale d ' un fuseau (Figs. 2, 3, 11 et
12), des inclusions consti tudes un iquement p a r des f i laments et des inclusions don t le
corps central c o m p o r t e / t la fois des f i laments et des tubules. Par ailleurs, elle nous a
permis de pr6ciser les symdtries d ' a r r angemen t dans les inclusions don t le corps central
est d ' aspec t cr is tal loide. Nous envisagerons ces trois cas successivement.
A. Inclusions filamenteuses en fuseau
Ce type d ' inc lus ion cor respond anx images des Figs. 2, 11, 12, 13. La plat ine gonio-
m6tr ique pe rme t d ' a m e n e r l ' axe de l ' inclusion para l l6 lement / t celui du faisceau d'dlec-
t rons et fourni t l ' image d 'une section qui serait s t r ic tement perpendicula i re ~t l 'axe de
l ' inclusion. Dans ces condi t ions, il appa ra i t (Fig. I3) que le corps central de ces in-
clusions est consti tu6 p a r un faisceau d 'une quaranta ine de f i laments plus ou moins
paral lNes. Chaque f i lament a u n diam6tre de 70/t 80 A environ. I1 faut r emarque r que
sur plus de 600 inclusions coup~.es selon une incidence quelconque, nous n ' av ions
j amais eu l 'occas ion d 'obse rver d i rectement de telles images.
Ces f i laments ne pr6sentent pas d ' a r r angemen t par t icul ier dans le p lan t ransversal
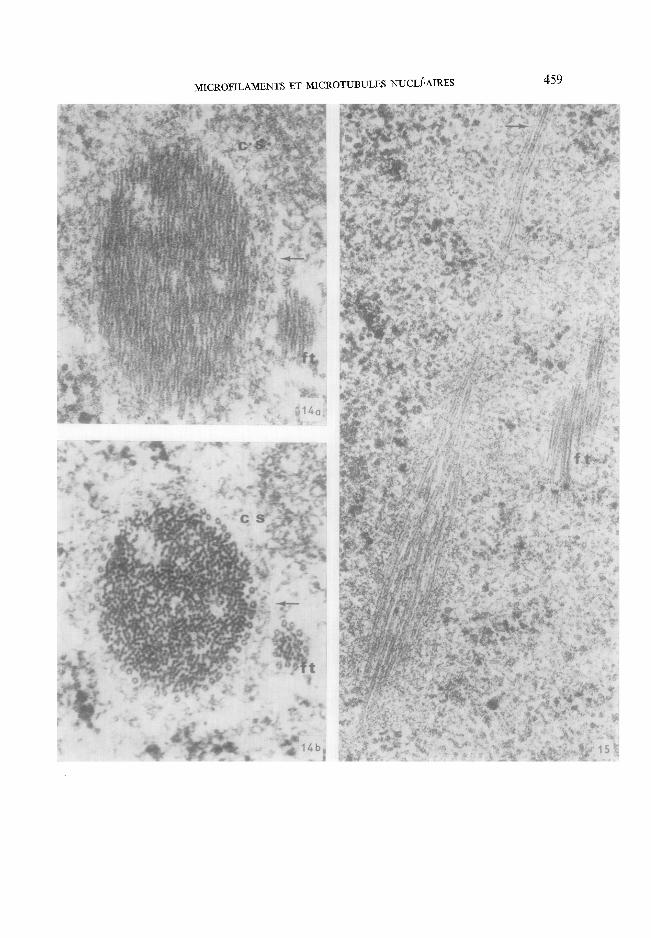
Fro. 14. (a) Sous cette incidence, il est difficile de dire si la zone centrale est tubulaire ou filamenteuse. Noter : une coque granulaire tr6s fine entourant l'inclusion; la pr&ence d'un court faisceau d'aspect tubulaire associ6 (ft); la pr6sence d'un << corps satellite >~ (cs) (b) Bascule de 45 ° qui permet de voir que le corps central de la m~me inclusion est constitu6 par de nombreux filaments parall61es/t arrangement le plus souvent irr6gulier, sauf dans la partie p6riph6- rique. I1 est circonscrit par une couronne (incompl6te ici) de tubules en une seule couche. Le faisceau associ6/t l'inclusion est nettement tubulaire (ft); l'aspect du << corps satellite >> (es) n'a pas vari6 et celui-ci peut 8tre consid6r6 comme tr& finement granulaire et grossi6rement sph6rique, x 80 000. FIG. 15. Section tangentielle d'une inclusion qui int6resse les tubules p6riph6riques, le << faisceau tubu- laire accessoire ~ (ft) et la coque granulaire. Cette image correspond tr6s vraisemblablement g une inclusion tubulo-filamenteuse analogue/t celle de la Fig. 14. Les tubules p6riph6riques forment un long fuseau, qui recouvre le corps central filamenteux et qui vase r6soudre progressivement dans le nucl6oplasme (-+). x 80 000.
©

460 SEITE, ESCAIG ET COUINEAU
(Fig. 13b). Sur des coupes grossi6rement longi tudinales , ces f i laments fo rment un
faisceau de 1 000 3, de diam6tre environ (Fig, 11). Leur longueur est diff ic i le / t d6ter-
miner avec cert i tude, comme nous l ' avons dit. Les mesures fournissent des valeurs qui
sont 6taldes entre 0,1/~ (Fig. 13a) et 4 # (Fig. 11). Nous re t iendrons donc, comme
valeur la plus p robab le , la valeur m a x i m u m observ6e soit 4/~.
Chaque f i lament pa ra l t atre plus cour t que le faisceau lui-m6me de telle sorte qu ' i l
pr6sente des discontinuit6s (Fig. 11) parfois tr6s 6videntes (Fig. 26).
C o m m e nous l ' avons vu, ce faisceau de f i laments est au contac t direct (Fig. 12) d ' un
corps satelli te qui est d ' a spec t f inement granula i re pa r r a p p o r t aux grains nucl6o-
plasmiques. Cette structure satellite, de fo rme ovalaire, don t le g rand axe est de 2/~
environ et le diam~tre de 0,2/~, est s i tu6e/ t une des extr6mitds du faisceau (Fig. 12).
Elle n 'es t pas toujours visible, mais l '6 tude de coupes s6ri6es tend/z mon t r e r que cette
absence n 'es t qu ' appa ren te et n 'es t due, tr6s vra isemblablement , qu'/~ l ' incidence de
coupe.
I1 est impor t an t de souligner que le corps centra l de ces inclusions est form6 ex- clusivement p a r des f i laments qui << ba ignent >) d i rectement dans le nucl6oplasme, sauf
dans le zone peu 6tendue o/1 ils sont accol6s au corps granula i re satellite.
Ces inclusions filamenteuses en fuseau sont le plus f r6quemment observ6es.
B. Inclusions tubulo-filamenteuses en fuseau
La plat ine goniom6tr ique en pe rmet t an t d 'obse rver des images qui correspondent /~
une section t ransversale de ce type d ' inclusion, mont re qu'el les sont consti tudes pa r
des f i laments de 7 0 / t 80 A de diam6tre environ, dispos6s de fagon quelconque au
centre et plus r6gul i6rement / t la p6riph6rie du corps central (Fig. 14 a et b), Ces fila-
ments sont au nombre de 800 environ.
A l ' inverse du cas pr6c6dent, ce groupe de f i laments n 'es t pas au contac t direct du
nucl6oplasme, mais il est entour6 pa r une couronne pdriph6rique de tubules dispos6s
rdgulibrement en une seule touche (Fig. 14b). Ces tubules sont au nombre de 60 en-
FIG. 16. Figures destin6es en particulier/~ montrer les diffbrents aspects pr6sent6s par une marne in- clusion selon l'angle de bascule (axe de bascule parall61e au bord inf6rieur du clich6). (a) 0 °. (b) 18 °. Sous cet angle les tubules p6riph6riques et le faisceau tubulaire accessoire sont tr6s nets et sont pratiquement parall61es au faisceau d'dlectrons. (c) 45 °. (d et f) Analyse sch6matique des Figs. 16 a (d) et 16c (f) permettant de voir que toutes les couches sont constitudes par des filaments et de comparer leur aspect en fonction de l'incidence du faisceau d'61ec- trons. Les filaments de la couche 1 sont perpendiculaires au faisceau sur le clich6 16 a alors qu'ils sont parall6les au faisceau sur la figure 16c. II apparaft ainsi que les lignes tr6s denses sont dues/t des fila- ments dispos6s r6guli6rement alors que les lignes de faible densit6 sont dues/t des filaments rdunis par paires (cf. 16c). (get h) + 45 °, g; + 30 °, h : axe de bascule toujours parall61e an bord inf6rieur du clich6. Aspect de la m~me inclusion qui a subi une rotation de 90 °dans le plan horizontal par rapport aux figures 16a, b et c. Remarquer la d6formation du corps granulaire satellite ce qui montre qu'il n'est pas strictement sph6rique dans ce cas. De plus ses dimensions paraissent plus r6duite que pr6c6demment, x 95 000.

~o
~o
ee
e
. e
e
ee
e
e
.o
e.
Ill
~11
• O
0
O6D
00
04
1'
O~
00,
O0
oo
oo
oq
Joo
oo
oo
OO
OO
OO
OO
Oq
~O
OO
OO
OO
O
O0
0o
oq
~o
oo
oo
OO
oo
~o
O0
00
40
OO
00
00
oO
~+:.:.
:.:.:~
:~x:~
:.:.:.:.
:.:.:.:.
:.x~>
:.x.:.x
.x.:.:+
x.:.:.:
.:.:̀....
..:.x.:
.:.:~:
.:.:~:
~:<
.̀.:.:.:.
~.x.:
<.:.:.
:.x+:
.:.:~:
+:~:
:.:~::.:
x~::~
:. oe
'e'.'
.'e'.'
e"O
'oo"
o#e"
~ee.
'oee
'~'e
oe...
.. J ]
llJJf
l I
OII
I~ III
v
Jill
O0
go
O0
go
go
go
O0
..
..
..
.
i,ii,,i
,,i

462 SEITE, ESCAIG ET COUINEAU
viron. Cette vari4t6 d'inclusions nucldaires est donc de nature tubulo-filamenteuse. L'ensemble des filaments et de la eouronne tubulaire est circonscrit par une sorte de
coque granulaire (Fig. 14). On peut noter 6galement la prdsence d 'un faisceau court
constitu4 par une quinzaine de tubules et d 'un corps satellite de 0,4/~ de diam6tre
environ. La forme circulaire et l 'aspect de ce corps satellite ne varient pas en fonct ion
de l ' inclinaison de la platine et c'est pourquoi nous le consid4rerons comme granulaire
et grossi6rement sphdrique.
Ici le corps granulaire satellite n 'est pas accol4 aux filaments, mais il est situ6 ~t
distance du corps central de l ' inclusion avec une localisation quelconque par rappor t
au faisceau tubulaire accessoire.
Le diam~tre de ces inclusions atteint 0,35 # environ. Un plan de coupe grossihre- ment parall~le ~t l 'axe longitudinal de l 'inclusion, et qui intdresse ~t la fois la couronne
de tubules p6riph6riques et le faisceau tubulaire accessoire (Fig. 15), permet d'6valuer
la longueur du corps central de ces inclusions/~ 2 # environ. Cette m4me image montre
que les tubules pdriph6riques paraissent constituer un fuseau qui v a s e rdsoudre pro-
gressivement dans le nucl4oplasme. En r6sum6, ces inclusions sont constitu4es par une association de filaments paral-
l~les ~ disposition plus r6guli6re ~t la p6ripMrie qu 'au centre, et d 'une couronne de
tubules pdriph4riques en une seule couche. Le corps central de l ' inclusion tubulo-
filamenteuse a un diamhtre plus important , mais une longueur plus faible que les
inclusions filamenteuses. Ces inclusions se distinguent encore des pr4c6dentes non seulement par la pr4sence
d 'une couronne pdriph6rique de tubules, mais encore par l 'existence d 'une coque granulaire et d 'un faisceau tubulaire accessoire (Fig. 14). Par ailleurs, comme nous
l 'avons dit, le corps granulaire accessoire est sph6rique, n 'est plus accol6 au corps
central de l ' inclusion et parai t plus important que dans le cas pr6c6dent.
C. Etude des inclusions cristalloMes (Figs. 16, 17, 18, 20)
La platine goniom6trique nous a permis d'expliquer 1'extreme diversit4 des images
d'inclusions ~ cristallo~des ~ qui apparait dans les conditions classiques d 'observation.
Fondamentalement , ces inclusions sont constitu6es, comme les pr6c6dentes, par des
tubules, des filaments et les m4mes structures associ6es (coque granulaire, faisceau
F~. 17a et b. Marne inclusion dont la forme polygonale r6sulte d'une incidence de coupe indiqu6e par le sch6ma 17c. (a) 0 °. (b) 45 °. La relation entre l'angle de bascule et l'angle apparent form6 par les deux couches de filaments permet d'6tablir avec certitude que les couches sont ddcal6es de 60 °. × 70 000. F~. 18. (b) 39 +. (c) 45 °. Deux coupes s6ri6es d'une m~me inclusion selon des plans ddfinis approxima- tivernent par la figure 18 a. Les tubules p6riph6riques forment une sorte de fuseau. Noter la pr6sence de lacunes dans le corps central. Noter aussi que la platine goniom6trique permet de corriger les variations de positions d'une m4me structure coup6e en s6rie sur le m4me treillis (l'aspect identique des deux images ayant 6t6 obtenu grace fi une 16g6re bascule), x 56 000.


464 SEITE, ESCAIG ET COUINEAU
SCFII~MA 1. Repr6sentation simplifi6e du mode d'organisation le plus frdquent d'une inclusion de type cristalloide. La partie centrale paralldl6pip6dique est constitude par des filaments dispos6s en 12 couches d6cal6es de 60 °. Les filaments sont dispos6s soit par paires, soit r6gulibrement 5 l'intdrieur de chaque couche. Les tubules p6riph6riques sont situ6s parall61ement aux deux grandes faces du parall616pip~de dans lequel s'inscrivent les filaments (angle de 30 ° environ par rapport au grand axe de l'inclusion). Sont 6galement repr6sent6s : le court faisceau tubulaire accessoire (f t) et le corps granulaire satellite (cgs). 11 est clair que selon l'incidence de coupes, ce mod61e peut foumir une in- finit6 d'aspects diffdrents. En r6alit6, les tubules ne sont pas strictement parall6les et forment peut- 6tre une sorte de fuseau, mais nous ignorons leur mode exact de terminaison dans le nucldoplasme. Le corps granulaire satellite peut 6tre soit du c6t6 oppos6 au faisceau de tubules associ6s, soit du m~me c6t6. I1 ne d6prime pas toujours les tubules p6riph6riques. La coque granulaire n'est pas repr6sent6e.
tubula i re p6riphdrique et corps granula i re satellite). Elles en diff6rent essentiel lement
pa r la d isposi t ion des f i laments qui se fai t en couches supetTosdes et pa r le mode de
r@ar t i t i on des tubules p6riph6riques.
a. Organisation des filaments en couches. On peut dis t inguer deux cat6gories de
couches de f i laments, l 'une dans laquelle les f i laments de 70 g 80 A sont r6guli6rement
espaces de 75 A, l ' au t re dans laquelle des f i laments de 100/t 110 ~ sont group6s pa r
paires de 200/t 250 A, chaque paire 6tant dis tante de 450 A environ de la suivante. Les
deux types de couches se superposent en a l te rnant : la d i rect ion des f i laments de l 'une
faisant un angle de 60 ° avec la direct ion des f i laments de la suivante (Fig, 17). La m~me
valeur avait 6t6 not6 entre les deux couches fibri l laires des treillis intranucldaires
(f ibri l lar lattice) d6crits pa r Chandle r et Willis (7) dans des neurones du Ra t et de la
Souris.
Pour les inclusions/~ 12 couches, l ' a l te rnance est r6guli~re sur 5 couches, la 6~me
est ident ique ~t la 56me (mais ses f i laments font toujours un angle de 60 ° avec ceux de

MICROFILAMENTS ET MICROTUBULES NUCLI~AIRES 465
Fro. 19. Mod61e destin6 ~t montrer les angles relatifs entre les directions des filaments et des tubules.
la 5~me couche), puis l'alternance redevient r6guli6re. Dans certains cas, la rupture de l'alternance r6guli6re ne se fait pas par apposition d'une 56me et d'une 6~me couche identiques, mais s'effectue au niveau d'une seule couche mixte qui comprend des fila- ments 6quidistants et des filaments groupds par paires (Fig. 16). Les sch6mas 1 et 2 rendent compte de cette organisation qui correspond aux inclusions les plus fr6quentes. Ces mod6les ~t 12 couches s'inscrivent dans un volume parall61@ip6dique dont la longueur, la largeur et la hauteur sont respectivement de 2 #, 0,8 # et 0,15/~ environ.
Deux remarques peuvent encore ~tre fakes : 1. Lorsque les filaments par paires ne sont pas parall~les au faisceau d'61ectrons, la
couche prend l'aspect de lignes continues de faible densit6 (Fig. 16) ou m~me devient tr6s impr6cise (Fig. 20/)); par contre, dans les m~mes conditions, les couches o/1 les filaments sont disposes r6guli6rement apparaissent sous la forme de lignes continues tr6s denses (Figs. 16 et 20/)) ce qui peut donner l'impression que le corps central est constitu6 par la superposition de feuillets (Figs. 4 et 20);
2. L'arrangement des filaments par rapport ~t ceux de la couche identique suivante (soit 2 couches apr6s) se fait selon une disposition hexagonale (Fig. 21).
b. R@artition des tubules pdriphdriques. Comme dans le cas pr6c6dent, le corps central filamenteux est entour6 par des tubules p6riph6riques qui, cette fois, sont
466 SEITE, ESCAIG ET COUINEAU
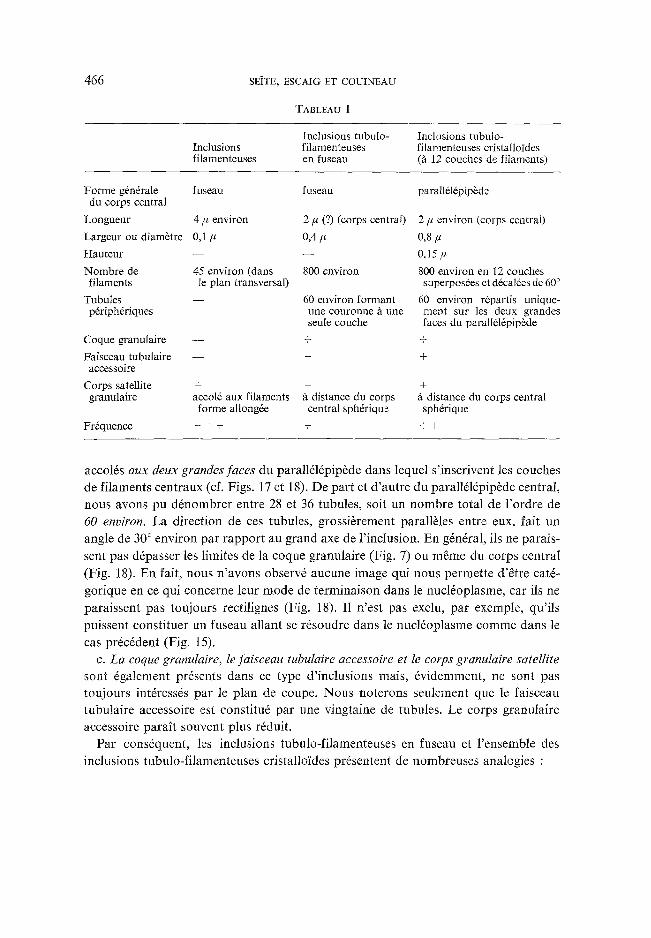
TABLEAU I
Inclusions tubulo- Inclusions filamenteuses filamenteuses en fuseau
Inclusions tubulo- filamenteuses cristalloides (fl 12 couches de filaments)
Forme g6n6rale du corps central
Longueur
Largeur ou diam~tre
Hauteur
Nombre de filaments
Tubules pdriphdriques
Coque granulaire
Faisceau tubulaire accessoire
Corps satellite granulaire
Fr6quence
fuseau fuseau parall616pipbde
4/~ environ 2 # (?) (corps central) 2 # environ (corps central)
0,1/~ 0,4/~ 0,8/,t
- - - - 0,15/~
45 environ (dans 800 environ 800 environ en 12 couches le plan transversal) superpos6es et d6cal6es de 60 °
- - 60 environ formant 60 environ r6partis unique- une couronne fl une ment sur les deux grandes seule couche faces du paralldl6pipbde
- - + +
- - + +
+ + + accol6 aux filaments /t distance du corps fi distance du corps central forme allongde central sph6rique sph6rique
+ + + + + +
accol6s aux deux grandes faces du parall616pip6de dans lequel s'inscrivent les couches de filaments centraux (cf. Figs. 17 et 18). De part et d 'autre du parall616pip~de central, nous avons pu ddnombrer entre 28 et 36 tubules, soit un hombre total de l 'ordre de 60 environ. La direction de ces tubules, grossi6rement parall61es entre eux, fait un angle de 30 ° environ par rapport au grand axe de l'inclusion. En g6ndral, ils ne parais- sent pas d6passer les limites de la coque granulaire (Fig. 7) ou m~me du corps central
(Fig. 18). En fait, nous n'avons observ6 aucune image qui nous permette d'atre catS- gorique en ce qui concerne leur mode de terminaison dans le nucl6oplasme, car ils ne paraissent pas toujours rectilignes (Fig. 18). I1 n'est pas exclu, par exemple, qu'ils puissent constituer un fuseau allant se r6soudre dans le nucldoplasme comme dans le cas pr6cddent (Fig. 15).
c. La coque granulaire, le faiseeau tubulaire accessoire et le corps granulaire satellite sont 4galement prdsents dans ce type d'inclusions mais, 4videmment, ne sont pas toujours int6ress4s par Je plan de coupe. Nous noterons seulement que le faisceau tubulaire accessoire est constitu6 par une vingtaine de tubules. Le corps granulaire accessoire parait souvent plus r4duit.
Par cons6quent, les inclusions tubulo-filamenteuses en fuseau et l 'ensemble des inclusions tubulo-filamenteuses cristalloides pr6sentent de nombreuses analogies :

MICROFILAMENTS ET MICROTUBULES NUCLEAIRES 467
b c
SCHEMA 2. Repr6sentation tr~s sch6matique (coupe transversale) des trois types fondamentaux d'inclusions nucl6aires. (a) Inclusion filamenteuse dont les filaments sont directement au contact du nucl6oplasme. Le corps granulaire satellite est accol6 aux filaments sur une partie de l'inclusion (cf. Fig. 12). (b) Inclusion tubulo-filamenteuse en fuseau, o/1 les filaments centraux sont entour6s par une couche unique de tubules p6riph6riques; le corps granulaire satellite paralt plus volumineux. (c) Inclusion tubulo-filamenteuse cristalloide/~ 12 couches de filaments; le corps granulaire satellite est moins r6gulier et parait ~tre plus r6duit que dans les deux cas pr6c6dents. Dans les deux derniers cas, sont repr6sent6s 6galement la coque granutaire et le faisceau tubulaire accessoire.
- - associa t ion de tubules et de f i laments;
- - pr6sence des 3 structures associ6es;
- - marne nombre approx imat i f de f i laments ;
- - marne nombre approx imat i f de tubules p6riph6riques.
La diff6rence essentielle r6side dans l ' o rgan i sa t ion des f i laments ~t l ' in t6r ieur du
corps central .
D. Remarques
a. L ' ensemble des r6sultats est r6sum6 dans le t ab leau I, pa r les sch6mas 1 et 2, et le
mod61e de la figure 19. I1 est impor t an t de rappe le r que les d imensions indiqu6es dans
ce tab leau ont 6t6 6tablies h pa r t i r de coupes don t l ' incidence est quelconque. On peut
avoi r une bonne pr6cision pour les diam~tres, pa r contre il est difficile d 'appr6c ier les
longuenrs r~elles h pa r t i r des longueurs observ6es.
Pour les inclusions f i lamenteuses, la valeur indiqu6e ( tableau I) co r respond ~t la
valeur m a x i m u m observ6e; toutes les valeurs inf6rieures sont tr~s v ra i semblab lement
dues ~ des or ienta t ions de coupes s'61oignant du p lan longi tudinal .
30 - 711825 g. Uttrastructure ResearcI~

468 SEITE, ESCAIG ET COUINEAU
Pour les inclusions tubulo-filamenteuses en fuseau, la valeur indiqu6e comprend la
couronne p6riph6rique de tubules, mais il n'est pas possible de pr6ciser la longueur des filaments.
Pour les inclusions tubulo-filamenteuses cristalloides, les valeurs observ6es sont
plus significatives dans la mesure o~ la forme g6om6trique des images correspond ~t
des plans mieux d6finis.
La not ion de fr6quence appelle 6galement un commentaire, car la probabilit6 de
rencontrer la coupe d 'une inclusion est d 'au tant plus grande que sa longueur est
61ev6e. C'est pourquoi il est possible que les inclusions filamenteuses donnent ] 'im-
pression d'etre plus nombreuses par rappor t aux autres qu'elles ne le sont en r6alit& Par contre les inclusions tubulo-filamenteuses en fuseau sont effectivement beaucoup
plus rares que les inclusions cristalloides. b. Certaines inclusions, rares ou m~me exceptionnelles, pr6sentent une structure qui
s'6carte des trois modules fondamentaux. Dans le cas des inclusions cristallo~des, les diff6rences portent essentiellement sur le
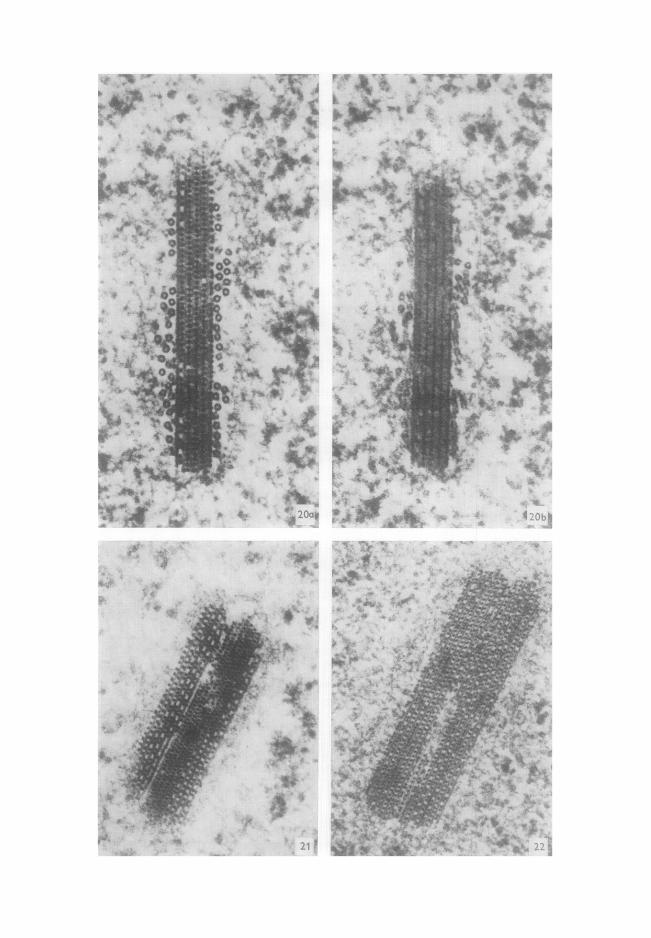
hombre de couches de filaments qui peut ~tre de 10 (Fig. 20) au lieu de 12 et atteindre
18, 28 (Fig. 21) et m~me 30 (Fig. 22). Dans ces inclusions exceptionnelles ~t 28 et 30
couches, nous n 'avons pas rencontr6 de tubules p6riph6riques.
Une autre 6ventualit~ rare, toujours pour ces inclusions cristallo~des, est repr6sent~e
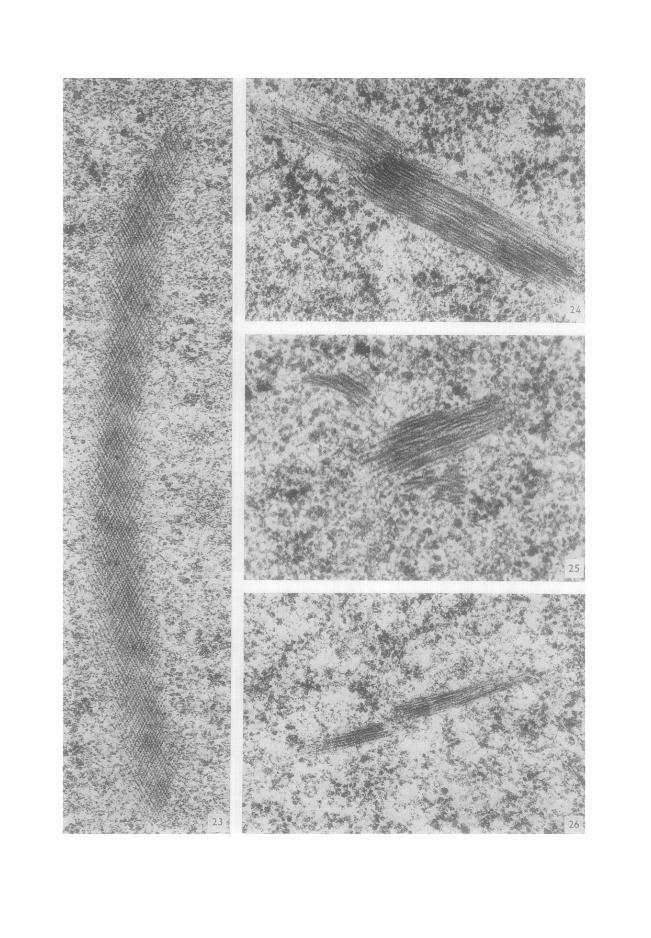
par une inclusion arqu6e, tr6s longue (2,3 #) (Fig. 23). Dans le cas des inclusions filamenteuses, nous avons pu observer exceptionnelle-
ment des inclusions off les filaments ne sont pas rectilignes (Fig. 24), des inclusions multiples (Fig. 25) et d 'autres enfin off le faisceau est nettement discontinu (Fig. 26).
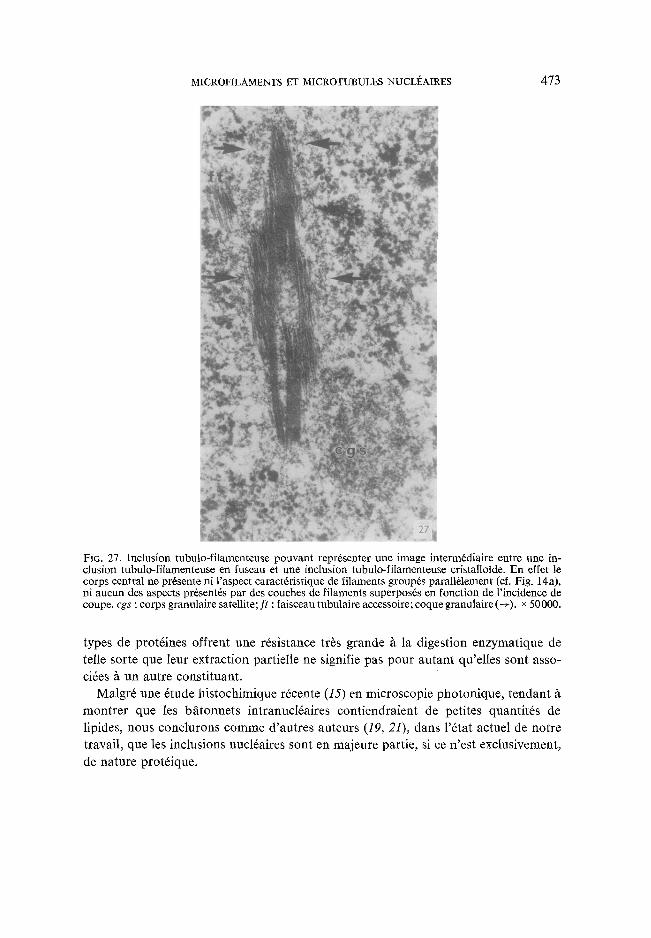
c. Une image rare (Fig. 27) a retenu notre attention, car elle pourrai t correspondre une inclusion tubulo-filamenteuse interm6diaire entre le type fuseau et le type cristal-
loide. d. Sur plus de 15 000 sections de noyaux, nous avons observ6 b, 6 reprises seulement
deux sections d'inclusions nettement distinctes dans la m~me coupe d 'un mEme noyau.
FI~. 20. Coupe transversale d'une inclusion h 10 couches. (a) Bascule + 21°. Les couches de filaments ~. disposition r6guli~re et celles off ils sont dispos6s par paires (apparaissant sous la forme d'une ligne continue de faible densit6) alternent r6guli~rement. La deuxi~me touche en partant de la gauche est form6e ~t la fois par des paires de filaments et par des filaments 6quidistants (couche mixte). Les tubules p6riph6riques sont tr~s nets. (b) Bascule -21°. La m~me inclusion paraR ~tre nniquement constitu6e par des lignes continues gt forte densit6 (filaments r6guli~rement r6partis de la Fig. 20a) ou ~ faible densit& L'ensemble peut donner l'impression de feuillets superpos6s, x 115 000. F~6. 21. Inclusion ~t 28 couches (6ventualit6 rare) paraissant constitu6e par 1'apposition de deux mod+le ~ 10 et 18 couches. Absence de tubules p6riph6riques. On peut noter sous cette incidence favorable, la disposition hexagonale des filaments. Comme dans la figure 20a, pr6sence de couches mixtes, x 75 000. Fx6. 22. Inclusion ~t 30 couches, x 80 000.

470 SEITE, ESCAIG ET COUINEAU
DISCUSSION
1. Microfilaments et microtubules nucldaires des neurones sympathiques
L'6tude en microscopic 61ectronique ordinaire de plus de 600 sections d'inclusions nucl6aires nous a fourni une tr6s grande vari6t6 d'images qui correspondent it la plupart de celles qui ont 6t6 observ6es dans d'autres territoires par d'autres auteurs.
Le lecteur trouvera une analyse bibliographique sur ce sujet dans notre travail pr6c6dent (34), ainsi que dans des travaux rdcemment parus (5, 6, 8, 10-12, 14, 17-23, 30, 31, 39, 41). Nous rapellerons seulement 5 cet 6gard que les images les plus fr6quem- ment d6crites jusqu'ici sont celles de faisceaux fibrillaires. Les images de ~ cristalloi- des ~, de ~ treillis fibriUaires ~ ou d'inclusions paracristallines, d'interpr6tation diffi- cile sinon impossible 5 l 'observation directe, sont plus rarement d6crites. Ce n'est que
de fagon tou t / t fair exceptionnelle que des tubules nucl6aires authentiques ont 6t6 signal6s dans des cellules normales.
L'6tude d 'un tr~s grand nombre de cas et l 'emploi d'une platine d'observation goniom6trique nous ont permis de d6montrer que les b~tonnets intranucl6aires sont constitu6s en r6alit6 par deux 616ments fondamentaux : des filaments de 70 ~ 110 A de diam&re et des tubules de 170/t 180 A de diam&re environ.
Ces inclusions nucl6aires ~ filaments e t / t tubules ont 6t6 observ6es dans tous les ganglions stellaires des 23 animaux 6tudi6s (pr6alablement vaccin6s). Ceci ne signifie pas que tousles noyaux d'une population de neurones contiennent au marne moment des constituants de ce type, mais cel/l signifie seulement, comme nous le verrons (35) que tousles noyaux sont susceptibles d'offrir de telles structures au cours de l'activit6
physiologique des neurones sympathiques, dont le niveau est tr+s variable, ~t chaque instant, selon les conditions physiologiques individuelles (rythme cardiaque, fr6- quence respiratoire, thermor6gulation, etc.).
C' est pourquoi nous sommes amends d considdrer ces inclusions nucldaires comme des constituants normaux du noyau des neurones sympathiques et ¢l distinguer leurs dldments eonstitutifs sous les termes de microfilaments et de microtubules.
Cette terminologie r6pond it un souci de clarification et permet, dans une perspec- tive de recherches ult6rieures, de souligner/t la fois des diff6rences 6videntes mais aussi des analogies entre les microfilaments et les microtubules nucl6aires d'une part et les neurofilaments et les microtubules cytoplasmiques d'autre part.
FIG. 23. Volumineuse inclusion cristalloide se distinguant par sa forme arqu6e et sa longueur sup6- rieure (2,3 #) aux valeurs les plus fr6quentes. × 60 000. FIG. 24. Les filaments paraissent former une boucle 5, leur extrdmit6. × 60 000. FIG. 25. Trois groupes de filaments. × 80 000. FIG. 26. Discontinuit6 des filaments, x 50 000.

472 SE'ITE, ESCAIG ET COUINEAU
Sur le plan purement ultrastructural, nous pouvons noter que le diambtre des microfilaments nucl6aires est de l 'ordre de 70 ?~ 80 A sauf dans le cas off ils sont r6unis par paires dans les inclusions cristallNdes (100-110 A). Ces valeurs s'inscrivent bien darts les limites que l 'on reconnalt au diam6tre des neurofilaments cytoplasmiques soit
80/t 110 X (26, 32, 33). Pour les microtubules nucl~aires, la difference est plus marquee, car leur diam6tre
est nettement inf~rieur (170 tt 180 A au lieu de 200 ~t 240 A) ~t celui des microtubules cytoplasmiques. Mais il faut souligner que ces derniers peuvent presenter de grandes variations de leur diamatre (28) de telle sorte qu 'on a pu en distinguer quatre cat6- gories (3).
En r6alit~, l 'analogie nous parait atre beaucoup plus fondamentale. La pr6sence de tubules ou de filaments nucl6aires t6moignerait de l'existence darts le noyau de pro- t6ines fibreuses dont il est 6videmment difficile de dire, en l'6tat actuel de nos travaux, si elles sont identiques ou seulement analogues par eertaines propri6t6s aux prot~ines fibreuses du cytoplasme. A ce sujet nous rappellerons que ces derni6res suscitent actuellement un grand int6r~t (cf. 32, 33). De nombreux travaux, dont nous ne citerons que les revues g~n&ales r~centes, se sont efforc~s en effet de d6terminer leurs parti- cularit6s ultrastructurales (27-29, 33), leur nature bioehimique (1, 4, 9, 13, 16, 40) et leur r61e qui parait ~tre d6terminant dans les ph6nom~nes de migration intra-axonale en particulier (cf. 32, 33).
Actuellement nous ignorons tout de la signification de la pr6senee de telles prot6ines dans le noyau de cellules qui ne sont pas impliqu6es dans les ph6nom6nes de division. I1 est ~galement impossible de proposer une hypoth6se fonctionnelle en se r~f&ant aux r61es qui leur sont attribuds dans le cytoplasme (architectural, maintenance du neu- rone, transports intracellulaires). Enfin nous n'avons observ6 aucune image pouvant sugg6rer une migration 6ventuelle de ces 616ments dans le cytoplasme comme cel/t a
~t6 avanc6 (21). Par contre l 'augmentation de la fr~quence des inclusions nucl6aires aprbs stimula-
tion 61ectrique 6tablit pour la premibre lois une corrdlation entre ces structures et le
niveau d'activit6 des neurones (35). Les 6tudes cytochimiques ne nous ont pas permis de d6finir nettement la nature de
ces inclusions. Le test de Silverman et Glick et les digestions enzymatiques nous ont montr6 qu'elles sont de nature prot6ique. Mais l'impossibilit6 de parvenir fi une extraction totale dolt faire retenir deux hypoth6ses. Nous ne pouvons pas dire, en effet, si les inclusions nucl6aires sont constitu6es exclusivement par une prot6ine ou si un autre constituant dont nous ignorons la nature (mais qui n'est sans doute pas de I 'ARN) est prdsent. Toutefois il est frappant de constater que dans un d61ai de 2 heures ou m~me de trois heures, sur certaines coupes, les microtubules cytoplasmiques de nature prot6ique sont encore fr6quemment ddcelables. I1 est possible que certains
MICROFILAMENTS ET MICROTUBULES NUCLt~AIRES 473
FIG. 27. Inclusion tubulo-filamenteuse pouvant repr6senter une image interm6diaire entre une in- clusion tubulo-filamenteuse en fuseau et une inclusion tubulo-filamenteuse cristaIloide. En effet le corps central ne pr6sente ni l'aspect caract6ristique de filaments group6s parall~lement (cf. Fig. 14a), ni aucun des aspects pr6sent6s par des couches de filaments superpos6s en fonction de l'incidence de coupe, cgs : corps granulaire satellite; ft : faisceau tubulaire accessoire; coque granulaire (4) . x 50000.
types de prot6ines offrent une r6sistance tr6s grande /t la d igest ion enzymat ique de
telle sorte que leur ext rac t ion par t ie l le ne signifie pas pour au tan t qu'el les sont asso-
ci6es ~ un autre const i tuant .
Malgr6 une 6tude h is tochimique r6cente (15) en microscopie pho ton ique , t e n d a n t / t
mon t r e r que les bf i tonnets int ranucl6aires cont iendra ien t de peti tes quanti t6s de
lipides, nous conclurons comme d 'au t res auteurs (19, 21), dans l '6tat actuel de not re
travail , que les inclusions nucl6aires sont en majeure pa t t ie , si ce n 'es t exclusivement,
de na ture prot6ique.

474 SEITE, ESCAIG ET COUINEAU
2. Organisation ultrastructurale des inclusions
I1 est difficile a priori de penser que les trois vari6t6s d'inclusions que nous avons raises en 6vidence puissent repr6senter des entit6s totalement ind6pendantes.
En effet, les deux vari6t6s d'inclusions tubulo-filamenteuses pr6sentent de fortes analogies (cf. Tableau I) :
- - l 'association de microfilaments et de microtubules; - - un m~me hombre approximatif (800 environ) de microfilaments; - - un m~me hombre approximatif de microtubules p6riph6riques (60 environ); - - des longueurs sans doute trbs voisines du corps central; - - la pr6sence des trois structures associ6es.
Les inclusions cristallo~des ne diffbrent des autres, en derni6re analyse, que par la r6partition en couches hautement organis6es et la r6partition pr6f6rentielle des micro-
tubules p6riph6riques. Dbs lots il est possible d'6mettre l 'hypoth6se que le passage de Fun/ t l 'autre type,
dans le sens inclusion tubulo4ilamenteuse en fuseau-inelusion tubulo-filamenteuse cristallo~de, est un ph6nombne dynamique aboutissant ~t une redistribution des 616- merits constitutifs par une sorte de glissement des microfilaments les uns sur les autres.
Pour appuyer cette hypoth6se, nous noterons que les inclusions tubulo-filarnen- teuses en fuseau sont rares, ce qui est un argument en faveur de leur caract6re transi- toire, et nous tiendrons compte de l 'image de la Fig. 27 que nous n'avons pas eu la possibilit6 d'observer ~t l'aide de la platine goniom6trique, mais qui para~t correspondre / tune inclusion mixte,
Quoiqu'il en soit, cette hypoth~se serait particuli6rement int6ressante dans la mesure off des recherches ult6rieures d6montreraient que les prot6ines fibreuses du
noyau, comme celle du cytoplasme, pr6sentent un certain nombre d'analogies avec
des prot6ines telle que l'actine (32, 33). Le probl6me des interrelations entre les inclusions filamenteuses et les pr6c6dentes
est encore plus difficile ~t aborder. Nous pouvons constater seulement que ces inclu- sions sont environ au minimum 2 fois plus longues que les inclusions tubulo-fila- menteuses. Leur 6volution vers le type tubulo-filamenteux en fuseau pourrait se faire alors par un raccourcissement avec une imbrication des microfilaments. Dans ce cas, il faudrait admettre qu'un certain nombre de microfilaments p6riph6riques se r6- organiseraient en microtubules. Par analogie cette hypoth6se pourrait s 'accorder avec la notion d6fendue par certains auteurs (26, 27, 43) de l'existence d'une sorte de ~< ba- lancement >> entre les neurofilaments et les neurotubules r6sultant d'une r6organisation dans l 'espace d'une subunit6 prot6ique commune aux deux structures. Toutefois cette notion para~t contredite par la r6cente raise en 6vidence (9, 13) de deux subunit6s prot6iques distinctes (tubuline et filarine).

MICROFILAMENTS ET MICROTUBULES NUCLEAIRES 475
Si cette conception uniciste nous parait la plus vraisemblable dans l'6tat actuel de nos recherches, il est 6vident qu'il reste h tenter de mettre ~tjour le d6terminisme de ces rapports dynamiques, question qui pose du marne coup celle de la signification fonc- tionnelle de ce type d'inclusion.
3. Les structures assocides
Parmi celles que nous avons d6crites une seule, ~ le corps granulaire satellite ~,
int6resse ~t la fois les trois vari6t6s d'inclusions. Ce corps granulaire pourrait cor- respondre au ~ corps granulo-fibrillaire ~) associ6 aux filaments intranucl6aires qui a 6t6 d6crit dans les neurones de l 'embryon de Poulet (21).
Darts notre mat6riel le corps granulaire satellite, vraisemblablement prot6ique, offre la particularit6 d'etre de forme allong6e et d'etre tr~s 6troitement accol6 aux filaments des inclusions filamenteuses sur une partie de leur trajet. Par contre il est grossi&e- ment spMrique, diminue de taille et s'61oigne du corps central darts le cas des inclu- sions tubulo-filamenteuses. Ces faits sugg&ent qu'il s'agit d'une structure dynamique dont l'6volution est vraisemblablement 6troitement li6e au degr6 de complexit6 des inclusions nucl6aires. D6s lots, ~t titre d'hypoth6se, on peut penser qu'elle pourrait intervenir darts le mode d 'arrangement r6ciproque des microfilaments et des micro- tubules nucl6aires ou encore qu'elle est plus ou moins directement impliqu6e dans la synth~se de ces 616ments, comme cel~t a 6t6 sugg6r6 par ailleurs (21).
Les autres structures associ6es, coque granulaire et ~ faisceau microtubulaire accessoire )) n 'avaient pas 6t6 observ6es jusqu'ici ~t notre connaissance. La coque granulaire, de nature vraisemblablement prot6ique, parait isoler le corps central des inclusions du reste du nucl6oplasme. Signe-t-elle un arr& au moins provisoire de leur 6volution; repr6sente-t-elle l ' image de subunit6s prot6iques en voie d'organisation? Nous n 'avons pas d 'arguments permettant de discuter de son r61e. De m~me, ~t l 'heure actuelle, nous sommes dans l'impossibilit6 d'envisager la signification du faisceau microtubulaire accessoire dont nous noterons toutefois qu'il a une orientation pr6cise par rapport aux microtubules p&iph6riques du corps central. Nous esp6rons que des recherches en cours, dont le but est d'6tudier l'6volution de ces structures en fonction du temps, nous permettrons d 'apporter un 616ment de r6ponse.
4. Un autre probl6me se pose : celui des relations que peuvent pr6senter ces micro- filaments et ces microtubules, quel que soit leur mode d'arrangement, avec les autres entit6s morphologiques du nucl6oplasme. Un des caract6res les plus nets de ces in- clusions est prdcis6ment de n 'avoir aucune localisation pr6f6rentielle pouvant sug- g6rer, comme Font fait certains auteurs (21), l'existence de rapports fonctionnels avec le nucl6ole ou l 'enveloppe nucl6aire.
Cette remarque ne signifie pas pour autant que de tels rapports n'existent pas mais nous noterons que ces probl6mes, qui touchent aux interrelations fonctionnelles entre

476 SEITE, ESCAIG ET COUINEAU
les diffSrents constituants nucl~aires et les b~tonnets nucl~aires, d~passent sans doute
les possibilit6s de la seule cytologie descriptive.
Les m~mes difficult5s concernent les relations dventuelles entre les ~< corps nucl6ai-
res >~ et les inclusions fi microfilaments et/~ mierotubules. Darts un travail pr6c6dent
sur les m6mes neurones, nous avions montr~ que ces corps nucl~aires sont tr~s fr&
quents et que leur nombre augmente au cours de l 'hyperactivit6 consecutive ~ l 'action de la r~serpine (34). La fr~quence et la structure de ces deux types d' inclusion sont
diffSrentes et il est possible d 'observer ~t la fois un ou plusieurs corps nucl~aires et un b~ttonnet intranuclSaire dans le m~me noyau, dans des aires nucl6oplasmiques tout ~t f a r distinctes. L'existence d 'une relation entre ces deux types d ' inclusion parait d~s lots peu vraisemblable, mais n'est dvidemment pas exclue.
Enfin la question se pose de savoir si les inclusions nucl~aires d~crites par d 'autres
auteurs et dans d 'autres territoires, y compris dans des cellules v6g~tales (42), peuvent
~tre consid~rSes comme de m~me nature que celles que nous venons de d6crire dans
les neurones sympathiques. Autrement dit de savoir si cette organisation ultrastruc-
turale tr6s complexe des microfilaments et des microtubules nucl~aires est ou n'est
pas un ph~nom6ne g6n~ral.
Dans l 'dtat actuel des recherches, il resterait /t d6montrer l'existence, dans ces
diverses categories cellulaires, de structures tubulaires intranucl~aires ~t l '6tat normal, voir s'il existe une association microfi laments-microtubules et, si tel est le cas,
savoir si leur mode d 'association est identique ou n o n / t celui que nous avons mis en ~vidence sur notre mat6riel. C'est donc un vaste champ d'investigation qui s 'ouvre
dans ce domaine encore tr~s peu connu, sans qu'il soit possible pour l ' instant d'6mettre
une opinion fond6e.
Quoiqu' i l en soit, nous soulignerons le fait que seule l 'observation syst6matique
d 'un grand nombre de noyaux ~ la platine goniom6trique nous a permis de fournir un
616merit de r6ponse aux probl6mes demeur6s non r6solus jusqu'ici de l 'organisat ion
ultrastructurale des b~ttonnets intranucl6aires 2 de la cellule nerveuse. En mont ran t que cette organisation repose, en derni~re analyse, sur la pr6sence de
microtubules et de microfilaments, cette 6tude nous parait susceptible non seulement
d 'ouvr i r de nouvelles perspectives de recherches sur l 'ultrastructure du noyau cellulaire, mais aussi de jeter les bases n6cessaires h des recherches destin6es ~ aborder la question de la signification biologique de ces inclusions. Cet aspect du problbme est envisag6 dans un autre travail qui a permis de mettre en 6vidence une augmentat ion de la fr6- quence des b~ttonnets intranucl6aires sous l ' influence de la stimulation 61ectrique (35).
II est bien 6vident qu'il s'agit d'un terme de microscopie photonique qu'il est difficile sinon im- possible de transposer dans tous les cas au domaine ultrastructural. Nous avons conserv6 cette terminologie non seulement pour sa valeur historique, mais aussi parce qu'elle permet, dans l'6tat actuel de nos connaissances, de distinguer imm6diatement une entit6 relativement d6finie parmi toutes celles qui entrent dans le cadre tr6s vague,/~ l'heure actuelle, des (( inclusions nucl6aires ~.

MICROFILAMENTS ET MICROTUBULES NUCLI~AIRES 477
S U M M A R Y
Nuclear inclusions of sympathetic neurons which can be correlated to << intranuclear rodlets >> of light microscopy are mainly, if not exclusively, of protein nature. They offer a wide variety of aspects when observed in usual conditions under electron microscope.
The electron microscope goniometer observation, done on several hundred inclusions, show that they are formed by two fundamental elements: microfilaments and microtubules which we consider as normal constituents of the nucleus. They built up filamentous and tubulofilamentous inclusions. The tubulofilamentous inclusions are either spindles formed by parallel microfilaments covered by one layer of microtubules or complex crystalloids. In this last case the microfilaments are in parallel layers (usually 12) forming a 60 ° angle with each other. We have observed a granular satellite body present in all inclusions. In the case of tubulofilamentous inclusions we have found two other associated structures: a granular shell and a short microtubular bundle.
This study is discussed with regard to the problems raised by the neuronal fibrous pro- teins. Hypothesis are proposed concerning the dynamics of the interrelation between the different varieties of inclusions.
Nous remercions le Dr. Luciani-Vuillet ainsi que Madame Bottini, Monsieur Cataldo, biologistes du C.N.R.S. et Madame Ferru pour l 'aide qu'ils nous ont apport6 au cours de ce travail.
BIBLIOGRAPHIE
1. ADELMAN, M. R., BoRISY, G. G., SHELANSKI, M. L., WEISENBERG, R. C. et TAYLOR, E. W., Fed. Proc. Fed. Soc. Exp. Biol. 27, 1186 (1968).
2. ANDERSON, W. A. et ANDRI~, J., .jr. Microseop. (Paris) 7, 343 (1968). 3. BEHNKE, O. et F(JRER, A., J. Cell Sci. 2, 169 (1967). 4. BORISY, G. G. et TAYLOR, E. W., dr. Cell Biol. 34, 535 (1967). 5. BOQUIST, L., d. Cell Biol. 43, 377 (1969). 6. BfDTTNER, D. W. et HORSTMANN, E., Exp. Cell Res. 49, 686 (1968). 7. CHANDLER, R. L. et WILLIS, R., J. Cell Sci. 1, 283 (1966). 8. DAHL, E., J. Anat. 106, 255 (1970). 9. DAVISON, P. F. et HUNEEUS, F. C., d. Mol. Biol, 52, 429 (1970).
10. DIXON, J. S., Anat. Rec. 168, 179 (1970). 11. GAMBETTI, P. et GONATAS, N. K., Riv. Patol. Nerv. Mental 88, 188 (1967). 12. GONATAS, N. K., d r. Neuropathol. 25, 177 (1966). 13. HUNEEUS, F. C. et DAVISON, P. F., dr. Mol. Biol. 52, 415 (1970). 14. KARLSSON, U., dr. Ultrastruct. Res. 16, 429 (1966). 15. KIM, S. U., MASUROVSKY, E. B., BENITEZ, U. H. et MURRAY, M. R., Histochimie 24, 33
(1970). 16. KIRK/'ATRICI¢, J. B., Fed. Proc. Fed. Soc. Exp. Biol. 27, 247 (1968). 17. KtBRNER, H. K. et FELDHEGE, A., Cytobiologie 1, 203 (1970). 18. KRALL, J. F., Z. Zellforsch. Mikrosk. Anat. 97, 45 (1969). 19. LANE, N. J., dr. Cell Biol. 40, 286 (1969). 20. MAGALHAES, M. M., Exp. Cell Res. 47, 628 (1967).

478 SEiTE, ESCAIG ET COUINEAU
21. MASUROVSKY, E. B., BENITEZ, H. H., KIM, S. U. et MURRAY, M. R., J. Cell Biol. 44, 172 (1970).
22. MEIER, C., Experientia 25, 294 (1969). 23. MISRABI, M., a r. Anat. 104, 588 (1968). 24. MONNERON, A., J. Microsc. (Paris) 5, 583 (1966). 25. MONNERON, A. et BERNrIARD, W., J. Microsc. (Paris) 5, 697 (1966). 26. PETERS, A., PA•AY, S. L., WEBSTER, H. DE F., The fine structure of nervous system.
Harper (Hoeber) New York, 1970. 27. PETERS, A. et VA~JOHN, J. E., ,L Cell Biol. 32, 113 (1967). 28. POCHON-MASSON, J., Ann. Biol. 6, 361 (1967). 29. PORTER, K. R., Principles Biomol. Organ. Ciba Found. Syrnp. 1965, p. 308 (1966). 30. QUATRINI, D., d. Microsc. (Paris) 5, 365 (1966). 31. RA~NE, C. S. et F~ELD, E. J., Brain Res. 10, 266 (1968). 32. SCHMITT, F. O., Neurosci. Res. Progr. Bull. 117 (1968). 33. - - dans BARONDES, S. H. (Ed.), Cellular dynamics of the neuron, p. 95. Academic
Press, New York, 1969. 34. SE~TE, R., J. Ultrastruct. Res. 30, 152 (1970). 35. SEiTE, R., MEI, N. et COUINEAU, S. (sous presse). 36. S~EGESMUND, K. A., DUTTA, C. R. et Fox, C. A., J. Anat. 98, 93 (1964). 37. SmVERMAN, L. et GLICK, D., 3". Cell Biol. 40, 761 (1969). 38. S~MARD, R., Int. Rev. Cytol. 28, 169 (1970). 39. SOTELO, C. et PALAY, S. L., J. Cell Biol. 36, 151 (1968). 40. STEPHENS, R. E. et LINCK, R. W., J. Mol. Biol. 40, 497 (1969). 41. VIWER, E., J. Microsc. (Paris) 4, 559 (1965). 42. WERG1N, W. P., GRURER, P. J. et NEWCOMB, E. H., d. Ultrastruct. Res. 30, 533 (1970). 43. WlNIEWSKI, H., SHELANSKI, M. L. et TERRY, R. D., J. Cell Biol. 38, 224 (1968).