Embed Size (px)
Citation preview

- 1 -
MUSÉUM NATIONAL D’HISTOIRE NATURELLE DE PARIS DÉPARTEMENT DE PRÉHISTOIRE
THÈSE
pour obtenir le grade de
DOCTEUR DU MUSÉUM NATIONAL D’HISTOIRE NATURELLE DE PARIS
Discipline : Préhistoire
présentée et soutenue publiquement dans l’amphithéâtre de l’Institut de Paléontologie Humaine
par
Anne BOUTEAUX
Le 27 octobre 2005
Paléontologie, paléoécologie et taphonomie des mammifères du Pléistocène moyen ancien du site à hominidés de Sangiran
(Java central, Indonésie)
Directeurs de thèse : François SÉMAH Anne-Marie MOIGNE
Rapporteurs : John DE VOS : Conservateur général au Museum Naturalis de Leiden, Pays Bas. Friedemann SCHRENK : Professeur à l’université Goethe de Francfort, Allemagne. Examinateurs : Jean-Philip BRUGAL : Directeur de recherches au C.N.R.S., Université Aix-Marseille Tony DJUBIANTONO : Directeur de recherches, directeur du centre national indonésien pour l’archéologie, Jakarta, Indonésie.I. Anne-Marie MOIGNE : Maître de Conférences au Muséum National d’Histoire Naturelle, Paris. François SÉMAH : Professeur au Muséum National d’Histoire Naturelle, Paris.

- 2 - 2

- 3 -
REMERCIEMENTS
Je tiens à remercier en premier lieu Monsieur le Professeur Henry de Lumley qui m’a accueilli au sein de l’Institut de Paléontologie Humaine depuis mon D.E.A.
Je remercie Monsieur le Professeur François Sémah d’avoir accepté de diriger ce
travail et de m’avoir permis de découvrir la préhistoire à Java. Je remercie également Madame Anne-Marie Moigne pour m’avoir fait découvrir les
espèces indonésiennes et pour le temps qu’elle m’a consacrée depuis le début de mes recherches afin de m’orienter et de me conseiller.
J’adresse mes remerciements à Monsieur le Professeur Teuku Jacob et Madame Etty
Indriati pour m’avoir accueilli dans leur laboratoire de l’université de Gajah Mada à Yogyakarta et m’avoir fait confiance pour étudier ces collections inédites ainsi que toute l’équipe du laboratoire pour son aide et sa gentillesse.
En Indonésie toujours, je remercie Messieurs Harry Widianto et Harry Truman Simanjuntak de m’avoir rencontrée et soutenue lors de chacune de mes visites. Je n’oublie pas non plus Mademoiselle Elfrida Anjarwati et toute l’équipe du Muséum de Sangiran pour avoir organisé mon excursion sur le dôme.
Je remercie l’équipe de Punung et plus particulièrement Kasman, Budiman et Stevanus ainsi que Pak Rokus pour ses conseils et sa bonne humeur.
Je remercie Monsieur John de Vos pour m’avoir reçue au Museum Naturalis de
Leiden et m’avoir permis d’étudier les collections Dubois. Je souhaite remercier également Madame Anne-Marie Sémah, Monsieur Florent
Détroit , Monsieur Christophe Falguères et Madame Brigitte Deniaux pour leur disponibilité et leurs conseils judicieux lors de leurs relectures. Et je remercie Monsieur Hervé Monchot pour ses conseils sur les statistiques et ses remarques judicieuses.
Je remercie tout particulièrement les membres du jury pour avoir accepté de
m’accorder du temps et de leurs compétences afin de juger ce travail. Mes remerciements vont également à tout le personnel technique et administratif de
l’Institut de Paléontologie Humaine et notamment à Marie-Thérèse Barbaud et Nicolas Claret.

- 4 - 4
Je remercie les étudiants Rury et Donan du laboratoire de Biologie de l’université de Gajah Mada pour m’avoir aidée dans mon travail au laboratoire et sur le terrain et m’avoir permis de découvrir la vie indonésienne. J’ai une pensée toute particulière pour tous mes amis indonésiens (Enno, Icuk, Daisy, Rohani, Darman, Jaya…) pour leur gentillesse qui a facilité mon intégration.
Un grand merci aussi : - à mes collègues de l’IPH : Vincent, Laurent, Sébastien, Emiliano, Caroline,
Guillaume, Marie, Erwan, Anne-Sophie, Matthieu, Xavier… - à mes collègues du C.E.R.P. de Tautavel : Pierre, Tony, Nicolas, Olivier, Arnaud… - à tous les amis qui m’ont toujours encouragée : Joffrey, Céline, Stéphanie, Caroline
et Jean-Michel, Edwige et Eric, Marie-Pierre et les autres… - et bien sûr à toute ma famille. Enfin, je remercie mes parents et ma sœur pour m’avoir toujours soutenu depuis mes
premiers ramassages de fossiles vers l’âge de cinq ans à Nanteuil. Et bien sûr je remercie Simon pour m’avoir accompagnée tout au long de ce périple et redonné du courage quand celui-ci me manquait.

- 5 - 5
A ma mère.

- 6 -

- 7 -
TABLE DES MATIERES Introduction ............................................................................................................................ 11 Premier chapitre..................................................................................................................... 15 Présentation du cadre de l’étude........................................................................................... 15
1. L’indonésie et l’île de Java.............................................................................................. 15 1.1 Géographie actuelle................................................................................................... 15 1.2 Contexte géotectonique ............................................................................................. 16 1.3 Eustatisme ................................................................................................................. 18 1.4 Géologie de l’île de Java ........................................................................................... 21
1.4.1. Les montagnes du sud ....................................................................................... 21 1.4.2. La grande dépression centrale ........................................................................... 21 1.4.3. La zone anticlinale de Bogor-Nord Seraju-Kendeng ........................................ 22 1.4.4. Les unités de la zone septentrionale.................................................................. 22
1.5. Climat ....................................................................................................................... 24 1.6. Végétation ................................................................................................................ 24 1.7. Faune ........................................................................................................................ 25 1.8. Migrations ................................................................................................................ 29 1.9. Historique des recherches paléontologiques à Java ................................................. 30
1.9.1. Les premières découvertes : 1888-1900............................................................ 31 1.9.2. L’expédition Selenka : 1906-1908 .................................................................... 33 1.9.3. L’entre deux guerres : von Koenigswald .......................................................... 34 1.9.4. De l’après guerre jusqu’à nos jours : le développement de la recherche indonésienne................................................................................................................. 35
1.10 Présentation du dôme de Sangiran .......................................................................... 37 1.10.1. Cadre géographique...................................................................................... 37
1.10.1.1. Actuel ....................................................................................................... 37 1.10.1.2. Paléogéographie ....................................................................................... 37
1.10.2. Cadre géologique et stratigraphique................................................................ 38 1.10.2.1. Origine du dôme....................................................................................... 38 1.10.2.2. Stratigraphie du dôme .............................................................................. 38
1.10.3. Palynologie...................................................................................................... 44 1.10.4. Datations.......................................................................................................... 46 1.10.5. Découvertes..................................................................................................... 48
1.10.5.1. Anthropologie........................................................................................... 48 1.10.5.2. Paléontologie............................................................................................ 49 1.10.5.3. Industrie lithique ...................................................................................... 49
1.10.6. Présentation des sites étudiés .......................................................................... 50 1.10.6.1. Tanjung..................................................................................................... 50 1.10.6.2. Sendang Busik.......................................................................................... 52 1.10.6.3. Ngrejeng Plupuh....................................................................................... 52 1.10.6.4. Grogol Plupuh .......................................................................................... 52 1.10.6.5. Bukuran .................................................................................................... 52 1.10.6.6. Dayu ......................................................................................................... 53
2. Cadre biostratigraphique de cette etude .......................................................................... 54 2.1. Récapitulatif des listes fauniques de Java au Pléistocène ........................................ 54 2.2. Biostratigraphie établie par von Koenigswald ......................................................... 59

- 8 - 8
2.3. Biostratigraphie établie par de Vos et Sondaar ........................................................ 62 2.4 Critiques .................................................................................................................... 68 2.5. Synthèse sur le cadre biostratigraphique de Java ..................................................... 72 2.6. Conclusion sur la colonisation de Java .................................................................... 73
Second chapitre ...................................................................................................................... 77 Etude paléontologique............................................................................................................ 77
1. Matériel osseux ............................................................................................................... 77 1.1. Travail préparatoire de laboratoire........................................................................... 77 1.2. Base de données ....................................................................................................... 78 1.3. Mesures et photographies......................................................................................... 78 1.4. Autres collections..................................................................................................... 79
2. Description de la faune des sites de Sangiran ................................................................. 80 2.1. Ordre Proboscidea Illiger, 1811 ............................................................................... 80
2.1.1. Famille Stegodontidae Young-Hopwood, 1935................................................ 81 2.1.2. Famille Elephantidae Gray, 1821...................................................................... 85 2.1.3. Proboscidiens indéterminés............................................................................... 89
2.2. Ordre Perissodactyla Owen, 1848............................................................................ 89 2.2.1. Famille Rhinocerotidae Gray, 1821 .................................................................. 89
2.3. Ordre Artiodactyla Owen, 1848 ............................................................................... 91 2.3.1. Famille Hippopotamidae Gray, 1821 ................................................................ 91 2.3.2. Famille Suidae Gray, 1821................................................................................ 95 2.3.3. Famille Bovidae Gray, 1821 ............................................................................. 97
2.3.3.1. Sous-famille Bovinae Gray, 1821 .............................................................. 97 2.3.4. Famille Cervidae Goldfuss, 1820.................................................................... 108
2.3.4.1. Sous-famille Cervinae Goldfuss, 1820..................................................... 109 2.3.5. Conclusion sur les herbivores ......................................................................... 124
2.4. Ordre Carnivora Bowdich, 1821 ............................................................................ 124 2.4.1. Famille Hyaenidae Gray, 1821........................................................................ 125
2.5. Ordre Primates........................................................................................................ 125 2.6. Reptiles et poissons ................................................................................................ 126
2.6.1. Ordre des Testudines....................................................................................... 127 2.6.2. Ordre des Crocodylia ...................................................................................... 128 2.6.3. Poissons........................................................................................................... 128 2.6.4. Gastéropodes ................................................................................................... 128
2.7. Comparaisons des collections issues des fouilles et de la surface ......................... 129 2.8. Collections fouillées............................................................................................... 133 2.9. Spectres fauniques.................................................................................................. 135 2.10. Âge des animaux .................................................................................................. 141 2.11. Richesse biologique.............................................................................................. 143 2.12. Biostratigraphie .................................................................................................... 144
2.12.1. Tanjung.......................................................................................................... 144 2.12.2. Sendang Busik............................................................................................... 145 2.12.3. Ngrejeng Plupuh (= Ngrejeng)...................................................................... 145 2.12.4. Grogol Plupuh (= Grogol) ............................................................................. 145 2.12.5. Bukuran ......................................................................................................... 146 2.12.6. Dayu ............................................................................................................. 146

- 9 - 9
Troisième chapitre................................................................................................................ 149 Etude des paléoenvironnements.......................................................................................... 149
1. Autoécologie ................................................................................................................. 149 1.1. Données sur les espèces actuelles .......................................................................... 150
1.1.1. Proboscidiens .................................................................................................. 151 1.1.2. Périssodactyles ................................................................................................ 152 1.1.3. Artiodactyles ................................................................................................... 153 1.1.4. Carnivores ....................................................................................................... 155 1.1.5. Reptiles............................................................................................................ 157 1.1.6. Conclusion....................................................................................................... 157
2. Synécologie ................................................................................................................... 158 2.1. Cénogramme .......................................................................................................... 158 2.2. Histogrammes écologiques .................................................................................... 159 2.3. Spectre d’association faunique............................................................................... 163 2.4. Conclusion.............................................................................................................. 165
3. Données climatiques et palynologiques ........................................................................ 166 3.1. Palynologie............................................................................................................. 166 3.2. Sédimentologie....................................................................................................... 166 3.3. Conclusion.............................................................................................................. 167
4. Endémisme et extinctions.............................................................................................. 167 4.1. Endémisme ............................................................................................................. 167 4.2. Extinctions.............................................................................................................. 168
Quatrième chapitre .............................................................................................................. 171 Etude taphonomique ............................................................................................................ 171
1. Répartition spatiale........................................................................................................ 171 2. Conservation.................................................................................................................. 173
2.1. Quantification des restes ........................................................................................ 173 2.2. Représentation squelettique.................................................................................... 175
2.2.1. Représentation squelettique générale .............................................................. 176 2.2.2. Représentation squelettique des bovidés et des cervidés ................................ 178 2.2.3. Représentation squelettique par classe de taille .............................................. 181 2.2.4. Comparaisons de Tanjung et Sendang Busik avec d’autres sites de plein air. 185 2.2.5. Représentations squelettiques des taxons minoritaires ................................... 190
2.3. Indices de conservation squelettique...................................................................... 192 2.4. Indices de représentation squelettique.................................................................... 193 2.5. Conclusion.............................................................................................................. 195
3. Fragmentation................................................................................................................ 196 3.1. Taille des fragments osseux ................................................................................... 196 3.2. Indice général d’intensité de la fragmentation : IGIF ............................................ 199 3.3. Indice de l’intensité de fragmentation des ossements : IFO................................... 200 3.4. Fragmentation des os longs .................................................................................... 201 3.5. Fragmentation des dents......................................................................................... 202 3.6. Forme des esquilles (NRI ou ND).......................................................................... 202
4. Agents taphonomiques .................................................................................................. 217 4.1. Agents climato-édaphiques .................................................................................... 217
4.1.1. Climat .............................................................................................................. 217 4.1.2. Formation du site fossilifère............................................................................ 217

- 10 - 10
4.1.3. Nature des dépôts ............................................................................................ 218 4.1.4. Description des actions de l’eau...................................................................... 219
4.1.4.1. Oxydes...................................................................................................... 220 4.1.4.2. Dissolution ............................................................................................... 220 4.1.4.3. Concrétions............................................................................................... 221 4.1.4.4. Couleurs ................................................................................................... 222
4.1.5. Fissuration ....................................................................................................... 222 4.1.6. Conclusion....................................................................................................... 224
4.2. Transport fluviatile................................................................................................. 224 4.2.1. Groupes de Voorhies....................................................................................... 225 4.2.2. Rapport dents/vertèbres................................................................................... 228 4.2.3. Groupes de Badgley ........................................................................................ 229 4.2.4. Connexions anatomiques................................................................................. 232
4.3. Agents biologiques non anthropiques .................................................................... 233 4.4. Action anthropique................................................................................................. 235
4.4.1. Formation de l’assemblage.............................................................................. 236 4.4.2. Composition de l’assemblage.......................................................................... 236 4.4.3. Modification de la surface des ossements ....................................................... 238
5. Discussion ..................................................................................................................... 239 Synthèse................................................................................................................................. 243
1. Tanjung.......................................................................................................................... 243 2. Sendang Busik............................................................................................................... 246 3. Ngrejeng Plupuh............................................................................................................ 247 4. Grogol Plupuh ............................................................................................................... 249 5. Bukuran ......................................................................................................................... 250 6. Dayu .............................................................................................................................. 251
Conclusion............................................................................................................................. 253 Références bibliographiques ............................................................................................... 259 Liste des figures .................................................................................................................... 297 Liste des tableaux ................................................................................................................. 301 Annexes ................................................................................................................................. 305 Liste des tableaux des annexes............................................................................................... 369

Introduction.
- 11 -
INTRODUCTION
Depuis très longtemps des fossiles de vertébrés ont été découverts à Java où ils étaient
considérés comme des restes des victimes de batailles mythiques (Sémah F. et al., 1993a). Les premières études paléontologiques ont débuté, sur cette île, dans la seconde moitié du XIXème siècle (Raden Saleh, 1867 ; Martin, 1887) en même temps que l’intérêt porté à l’étude des sites qui livraient les premiers fossiles humains dans le monde augmentait. C’est en effet à la suite de la théorie de l’évolution de Darwin (1859) que l’origine et l’évolution de l’homme sont devenues des sujets d’étude majeurs et que les recherches paléontologiques à Java se sont développées.
En effet, à la fin du XIXème siècle, les recherches sur le « chaînon manquant » de l’évolution humaine se multiplient, notamment en Indonésie où un jeune docteur néerlandais, Dubois, débute des recherches à Sumatra mais sans succès. Après la découverte anthropologique de Wajak sur l’île de Java (1889), il décide en 1890 de mener de nouvelles recherches dans cette zone. Il entreprend des fouilles sur le site de Trinil et découvre les premiers restes de Pithecanthropus erectus ou Homo erectus de Java (Dubois, 1894).
L’île de Java devient alors une zone privilégiée de découvertes et de nombreuses campagnes de fouilles et de prospections ont été menées principalement dans l’est et le centre de l’île et plus particulièrement à Sangiran, le site de notre étude. De nombreux restes d’Homo erectus y ont été découverts ainsi que de très nombreux restes fauniques et des outils lithiques.
Von Koenigswald, un paléontologue allemand (d’origine danoise) engagé et naturalisé par la suite par les néerlandais, fut le premier à mettre au jour de l’industrie lithique préhistorique et une mandibule d’Homo erectus sur le dôme de Sangiran (von Koenigswald, 1936, 1940). Depuis lors, les recherches n’ont pas cessé dans cette zone (Lizon-Sureau, 1979 ; Sémah F., 1984 ; Watanabe & Kadar, 1985 ; Sémah A.M., 1986 ; Sémah F. et al., 1993b, 2002). Depuis les années 70, le Centre National Indonésien de Recherches Archéologiques en collaboration avec le laboratoire de bio-paléoanthropologie de l’université de Gajah Mada (Yogyakarta) organisent un programme de recherches sur le dôme de Sangiran (Sartono, 1964a ; Jacob, 1964, 1975b ; Koeshardjono, 1988). De nombreuses campagnes de fouilles et de prospections sont menées et de nombreux restes anthropologiques et fauniques sont mis au jour notamment dans les localités de Tanjung, Sendang Busik, Ngrejeng Plupuh, Grogol Plupuh, Bukuran et Dayu. Plusieurs de ces collections de faune sont aujourd’hui conservées au laboratoire de bio-paléoanthropologie de l’université de Gajah Mada (Yogyakarta) où nous avons mené cette étude.

- 12 - 12
Actuellement, Sangiran est le site le plus riche de l’île de Java au niveau paléoanthropologique, 60 individus y ont été récoltés. Ces découvertes représentent 50 % des restes d’Homo erectus dans le monde (Simanjuntak, 2001) ce qui en fait un site d’un intérêt colossal pour l’évolution humaine. C’est ce qui lui a valu d’être classé au patrimoine mondial de l’humanité par l’U.N.E.S.C.O. dès 1996.
En parallèle, des recherches anthropologiques, de nombreux fossiles de mammifères et de reptiles du Pléistocène ont pu être mis au jour. En ce qui concerne la paléontologie, Sangiran présente une succession sédimentaire complète du Pliocène supérieur au Pléistocène moyen, ce qui en fait un site exceptionnel du point de vue de la chronologie, de la biostratigraphie et des paléoenvironnements de Java au Quaternaire. Il est également l’un des sites fossilifères les plus riches de l’île de Java pour le Pléistocène moyen.
Malheureusement, la majorité des découvertes sur le dôme ne possèdent pas un cadre stratigraphique précis et ont été faites en surface. Les nombreuses études menées sur les collections fouillées de Dubois et de von Koenigswald (von Koenigswald, 1933 et 1935 ; Brongersma, 1937 ; Hooijer 1946a, 1946b, 1947, 1950, 1955, 1958, 1960 et 1962 ; de Vos et al., 1982b, Sondaar, 1984, van den Bergh et al., 1996 et 2001) ont permis de déterminer les diverses espèces présentes à Java, leur association, leur succession et leur origine. Néanmoins, peu d’unités fauniques proviennent d’un cadre stratigraphique précis et seul un petit nombre d’entre elles ont pu être datées directement. Dans ce contexte, les fossiles provenant des campagnes de fouilles menées à Sangiran sont d’une importance toute particulière (Aimi & Aziz, 1985 ; Sémah F. et al., 1993b ; Moigne et al., 2004) notamment au niveau biostratigraphique. Les collections issues de ces fouilles (Aimi & Aziz, 1985) ont permis de dater les anciennes collections par comparaison des espèces présentes et des formations sédimentaires (Leinders et al., 1985). L’apport de collections paléontologiques, telles que celles qui font l’objet de la présente thèse, doit donc permettre de compléter et d’affiner le cadre biostratigraphique de Java afin de mieux appréhender la succession des migrations des mammifères au cours du Pléistocène.
Les données provenant de l’étude de ces six nouveaux sites de Sangiran vont
également compléter les données sur la paléoécologie du dôme, lors du Pléistocène moyen, obtenues à partir de l’étude du pollen (Sémah A.M., 1986) et des dépôts sédimentaires (Sémah F., 1984 ; Itihara et al., 1985 ; Larick et al., 2001). La nature des espèces fossiles va nous informer sur le paléoenvironnement d’Homo erectus au Pléistocène moyen.
Des analyses biostratigraphiques et paléoécologiques ont été menées classiquement
sur les collections paléontologiques javanaises mais très peu d’entre elles ont considéré l’importance des mammifères par rapport aux hominidés et quelle pouvait être leur interaction (Choi, 2003). À Sangiran, sur le site de Ngebung 2, un sol d’habitat a pu être mis en évidence dans le niveau A où des outils et des restes fauniques ont été retrouvés associés (Sémah et al.,

- 13 - 13
1993, 2001a, 2002). Il nous a donc paru indispensable et nécessaire de considérer les assemblages de notre étude dans une dimension archéozoologique pour tenter d’analyser les relations entre l’homme et les mammifères dans le passé. Pour ce faire, nous avons réalisé une analyse taphonomique des restes osseux pour définir l’origine des assemblages et essayer de faire ressortir un lien entre la faune présente et Homo erectus. Ceci a pour but de mieux connaître le comportement de subsistance de cet hominidé en milieu de type insulaire.
Dans un premier temps, nous présentons le contexte de notre étude. Nous débutons par
une présentation de l’île de Java puis des recherches paléontologiques qui s’y sont déroulées avant d’aborder plus particulièrement le dôme de Sangiran et les sites considérés dans notre étude. Nous faisons également le point sur les données biostratigraphiques préalable indispensable avant de présenter nos propres résultats.
Dans un second chapitre, nous effectuons l’étude paléontologique du matériel afin de définir précisément les taxons présents dans chacun des six gisements étudiés. Chaque espèce est traitée au niveau morphologique et ostéométrique afin de la comparer à celles des populations d’autres gisements de Sangiran ou de Java, pour la replacer dans un cadre biostratigraphique.
Dans le chapitre suivant, nous utilisons les déterminations paléontologiques pour tenter de reconstituer le paléoenvironnement des sites de Sangiran à partir des méthodes autoécologiques et synécologiques classiques et en faisant une revue critique de ces méthodes pour des sites de plein air en contexte fluviatile.
La dernière grande partie aborde l’étude de la taphonomie. Après avoir estimé la conservation et la fragmentation des restes osseux nous examinons chaque reste afin de mettre en évidence les stigmates de l’action des différents agents responsables de l’accumulation des ossements et/ou des modifications intervenues sur la composition initiale des assemblages. Ceci nous permet de proposer une interprétation pour l’origine des assemblages osseux des sites considérés. Notre recherche a donc pour objectifs d’affiner le cadre biostratigraphique de l’île de Java, de compléter les données sur l’environnement d’Homo erectus à Sangiran et de connaître l’origine des assemblages osseux. Toutes ces approches permettront de mieux comprendre les interactions de l’Homo erectus javanais avec son milieu au Pléistocène moyen et donc de permettre de compléter les données sur son comportement de subsistance.

- 14 -

- 15 -
PREMIER CHAPITRE PRESENTATION DU CADRE DE L’ETUDE
1. L’INDONESIE ET L’ILE DE JAVA
1.1 Géographie actuelle
L’île de Java se situe en Indonésie. L’archipel indonésien est composé d’environ treize mille îles qui se répartissent sur plus de 5 000 km d’est en ouest et 2 000 km du nord au sud (figure 1). Les îles principales sont Sumatra, Bali, Bornéo (Kalimantan), Célèbes (Sulawesi) et Java (Aarsse, 1993).
Figure 1 : Carte générale de l’Indonésie (archipel indonésien en orangé)
(d’après www.intercarto.com)
Java est limitée au Sud par l’Océan Indien, au nord par la Mer de Java, à l’est par le détroit de Bali et à l’ouest par le détroit de la Sonde (figures 1 et 2). Elle se situe entre 6° et 9° de latitude sud et 105 et 114° de longitude est. Cette île s’allonge de 1 000 km d’ouest en est et jusqu’à 180 km du nord au sud (Sevin, 1993).

Premier chapitre : Présentation du cadre de l’étude.
- 16 - 16
Figure 2 : Carte générale de l’île de Java
(d’après www.lonelyplanet.com/mapimages/south_east_asia/java/java.gif)
Cette île, d’une surface de 127 000 km², est la plus peuplée d’Indonésie (environ 120 millions d’habitants) et se divise en trois parties administratives : Java ouest, Java central et Java est. L’île totalise 33 volcans dont 17 en activité. La majorité se situe dans la chaîne montagneuse centrale et constitue la plus importante concentration de volcans actifs du monde (Sevin, 1993). Cette île, riche en ressources minières, a fait l’objet de nombreuses études et prospections géologiques hollandaises lors de la colonisation. Ces recherches sont à l’origine des nombreuses découvertes paléontologiques faites à partir des années trente (van Es, 1931, 1936).
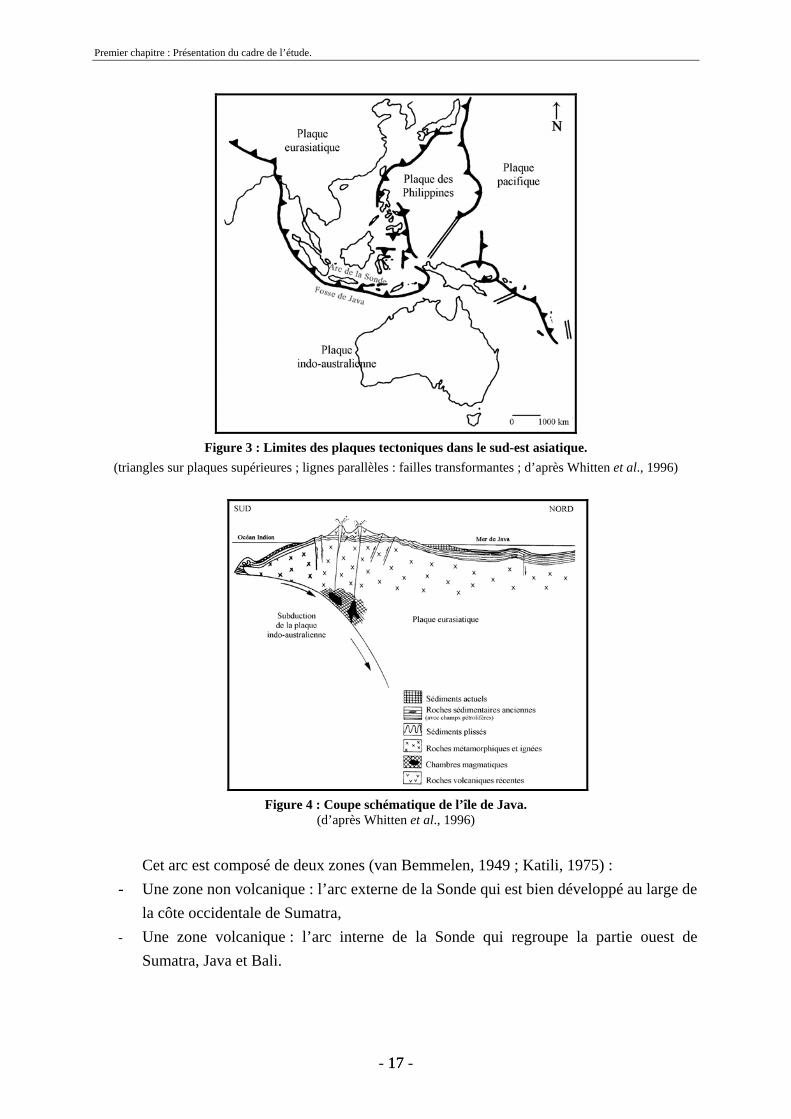
1.2 Contexte géotectonique L’archipel indonésien résulte du contact de quatre grandes plaques tectoniques : la plaque eurasiatique, la plaque indo-australienne, la plaque océanique pacifique et la plaque des Philippines (figures 3 et 4). La subduction de la plaque indo-australienne sous la plaque eurasiatique a formé l’arc insulaire de la Sonde et la fosse de Java en avant de celui-ci (Katili, 1975).
Premier chapitre : Présentation du cadre de l’étude.
- 17 - 17
Figure 3 : Limites des plaques tectoniques dans le sud-est asiatique.
(triangles sur plaques supérieures ; lignes parallèles : failles transformantes ; d’après Whitten et al., 1996)
Figure 4 : Coupe schématique de l’île de Java.
(d’après Whitten et al., 1996)
Cet arc est composé de deux zones (van Bemmelen, 1949 ; Katili, 1975) :
- Une zone non volcanique : l’arc externe de la Sonde qui est bien développé au large de la côte occidentale de Sumatra,
- Une zone volcanique : l’arc interne de la Sonde qui regroupe la partie ouest de Sumatra, Java et Bali.

Premier chapitre : Présentation du cadre de l’étude.
- 18 -
À l’Oligocène (30 Ma), l’arc insulaire de Sumatra à Flores prend son orientation actuelle. Le plateau de la Sonde, stable au niveau sismique, est bordé par l’arc insulaire. Il forme une vaste plate-forme pré-tertiaire à la pointe Sud du continent asiatique (Saint Marc et al.,1977). Il est exondé à Kalimantan et est recouvert par des mers peu profondes (Mer de Java, Mer de Flores) ne dépassant pas 100 mètres de profondeur dans les autres zones.
Au cours du Quaternaire, lors des alternances de périodes interglaciaires et glaciaires, ce plateau a donc été successivement submergé et émergé ce qui a permis la création de « ponts terrestres » (Sondaar, 1984) qui ont favorisé les migrations des faunes continentales vers les îles, et plus particulièrement celle des mammifères. Ceci a permis le développement d’espèces endémiques sur les différentes îles. Selon Saint Marc et al. (1977), la formation de l’île de Java résulte de la succession de trois phases orogéniques :
- Pendant l’Oligocène moyen, - Au Miocène moyen, - Et au Plio-Pléistocène.
1.3 Eustatisme
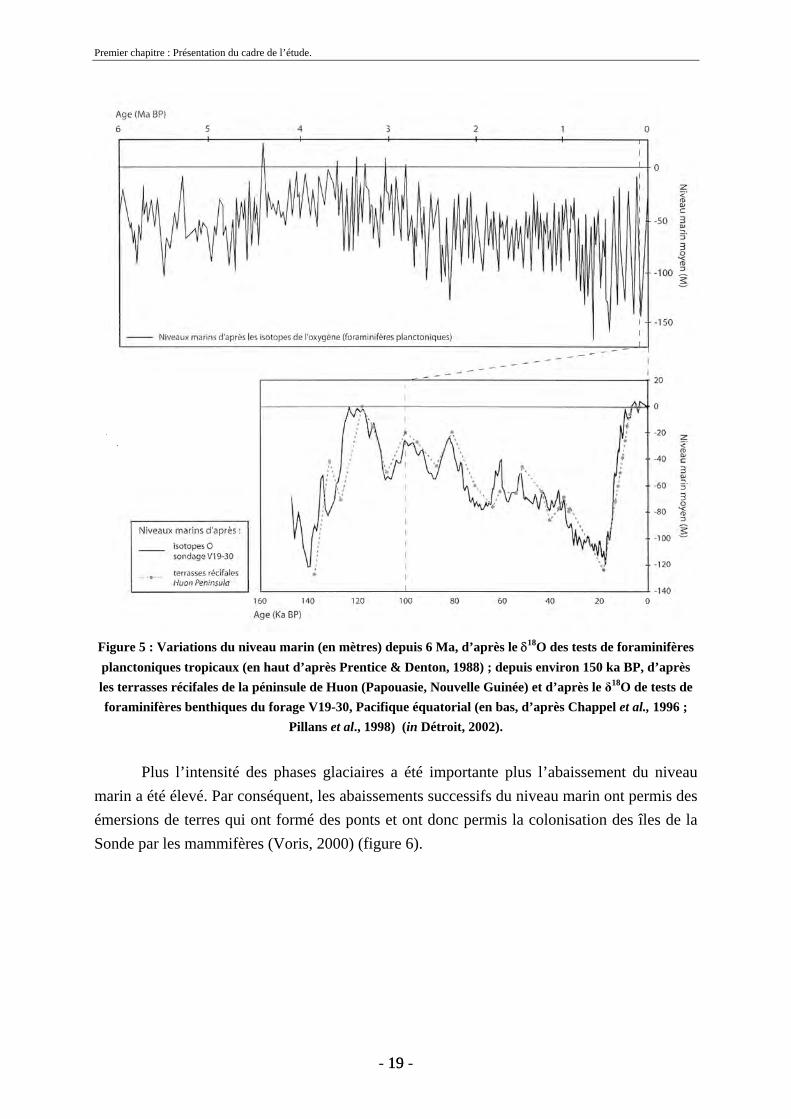
Dans cette zone du globe, les variations eustatiques, liées aux phases glaciaires et interglaciaires, ont permis l’émersion des terres dans des zones de mer peu profondes du Pliocène à l’Holocène. Ces variations n’ont pas été constantes au cours de cette période (figure 5) (Prentice & Denton, 1988 ; Pillans et al. 1998).
De 2,8 à 2,4 Ma, l’intensité des abaissements du niveau marin a augmenté. De 2,4 Ma à 0,8 Ma la variation moyenne du niveau marin est de -70 mètres avec des maximums à -120 mètres environ. A 0,8 Ma nous observons un abaissement maximum de -150 mètres environ avec une moyenne de 90 m (Prentice & Denton, 1988).
À partir du Pléistocène moyen, d’importantes variations eustatiques sont constatées. Les variations du niveau marin montrent de fortes amplitudes entre les périodes glaciaires et interglaciaires. Par exemple, entre 0,4 et 0,5 Ma, on observe une amplitude de 135 mètres (Prentice & Denton, 1988).
Premier chapitre : Présentation du cadre de l’étude.
- 19 - 19
Figure 5 : Variations du niveau marin (en mètres) depuis 6 Ma, d’après le δ18O des tests de foraminifères planctoniques tropicaux (en haut d’après Prentice & Denton, 1988) ; depuis environ 150 ka BP, d’après les terrasses récifales de la péninsule de Huon (Papouasie, Nouvelle Guinée) et d’après le δ18O de tests de foraminifères benthiques du forage V19-30, Pacifique équatorial (en bas, d’après Chappel et al., 1996 ;
Pillans et al., 1998) (in Détroit, 2002).
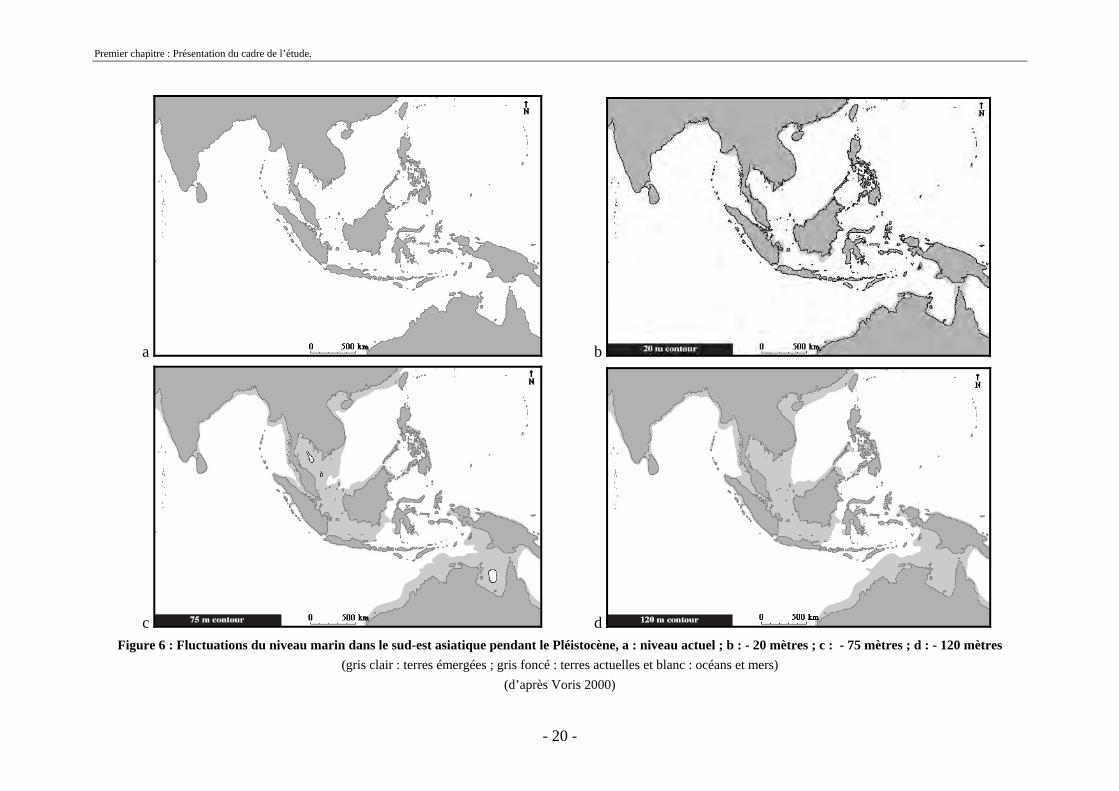
Plus l’intensité des phases glaciaires a été importante plus l’abaissement du niveau
marin a été élevé. Par conséquent, les abaissements successifs du niveau marin ont permis des émersions de terres qui ont formé des ponts et ont donc permis la colonisation des îles de la Sonde par les mammifères (Voris, 2000) (figure 6).
Premier chapitre : Présentation du cadre de l’étude.
- 20 -
a b
c d Figure 6 : Fluctuations du niveau marin dans le sud-est asiatique pendant le Pléistocène, a : niveau actuel ; b : - 20 mètres ; c : - 75 mètres ; d : - 120 mètres
(gris clair : terres émergées ; gris foncé : terres actuelles et blanc : océans et mers) (d’après Voris 2000)

Premier chapitre : Présentation du cadre de l’étude.
- 21 -
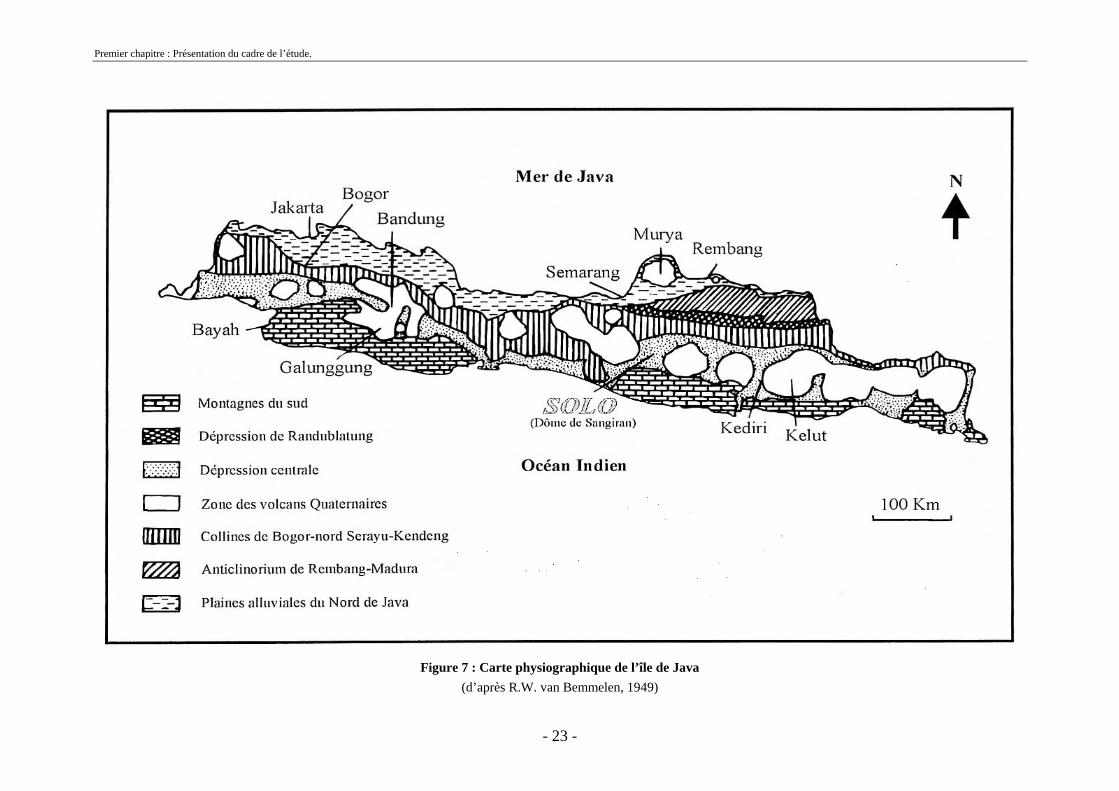
1.4 Géologie de l’île de Java
L’île de Java peut se découper en quatre grands ensembles géologiques (figure 7): les montagnes du sud, la grande dépression centrale, la zone anticlinale de Bogor-Nord Seraju-Kendeng et les unités de la zone septentrionale. Nous nous intéresserons tout particulièrement à la zone géologique où se situe le dôme de Sangiran (la grande dépression centrale) et aux zones qui lui sont contigües.
1.4.1. Les montagnes du sud
Ces montagnes se situent dans les zones ouest, centrale et est de Java, au sud du dôme de Sangiran, elles datent principalement du miocène (figure 7).
Elles sont constituées de dépôts volcaniques, les « old-andesites » où s’intercalent des calcaires à lépidocyclines du Miocène inférieur et moyen (van Bemmelen, 1949). Une régression a eu lieu après la formation des calcaires du Miocène supérieur. Au Pléistocène, cette zone s’est soulevée et inclinée vers le sud lors de la formation du grand anticlinal central. Cette surrection a permis le drainage des eaux à l’origine du système karstique actuel. Les montagnes du sud forment donc le flanc sud du grand anticlinal. Les produits de leur érosion se retrouvent dans la formation de Kabuh du Pléistocène moyen à Sangiran.
Le système karstique a isolé de petites collines calcaires de quelques dizaines de mètres de hauteur dans lesquelles existent des grottes. Certaines ont servi d’abri à l’homme au cours du Pléistocène supérieur et de l’Holocène et peuvent contenir du matériel archéologique+.
1.4.2. La grande dépression centrale
La grande dépression centrale de Java (figure 7) est comblée d’alluvions plio-
pléistocènes et de dépôts volcaniques. Elle peut être divisée en deux zones :
• La zone de Bandung : Elle se trouve dans la partie ouest de Java. Elle est constituée de plusieurs plaines
séparées par des complexes volcaniques (van Bemmelen, 1949 ; Sémah F., 1984 ; Djubiantono, 1992).
• La zone de Solo : Elle se trouve dans la partie est de Java. Le dôme de Sangiran se situe dans cette
dépression ainsi que le site de Trinil. + La grotte de Song Terus (Java Est) se situe dans la zone des Gunung Sewu (« les mille collines ») où une équipe franco-indonésienne : la Mission Quaternaire et Préhistoire en Indonésie (M.Q.P.I.), dirigée par Sémah et Simanjuntak, organise actuellement des fouilles et poursuit des recherches sur l’industrie lithique, la faune et les restes humains (Sémah F. et al., 2002 ; Détroit, 2002 ; Gallet, 2004 ; Hameau, 2004 ; Chacornac-Rault, 2004).

Premier chapitre : Présentation du cadre de l’étude.
- 22 - 22
Le long de sa partie axiale, de nombreux volcans « géants » quaternaires sont visibles et leurs dépôts recouvrent les formations tertiaires. En outre, de nombreux événements tectoniques locaux se sont produits qui ont conduit à la formation de grands grabens et de rides anticlinales affectant les sédiments (van Bemmelen, 1949). Ces dernières sont nombreuses à proximité du volcan Lawu où se situent les dômes de Sangiran, de Gemolong et de Bringinan (Sémah F., 1984 ; Djubiantono, 1992).
1.4.3. La zone anticlinale de Bogor-Nord Seraju-Kendeng
Cette zone se trouve au nord de la dépression centrale (figure 7). Elle est composée de grandes collines qui, à l’ouest, constituent la zone de Bogor, au centre la chaîne Nord Serayu et à l’est la zone des Kendeng (van Bemmelen, 1949 ; Djubiantono, 1992).
• La chaîne des Kendeng :
L’anticlinorium des Kendeng est un système de plis de direction est-ouest et est situé à l’est de Java au nord du dôme de Sangiran. Il montre des plis asymétriques.
Cette chaîne est moins élevée que la précédente. Elle s’étend de l’est du volcan Ungaran jusqu’au détroit de Madura. Son émergence date environ de la fin du Pliocène et cette phase tectonique s’est ensuite propagée à l’est, suivie par la régression de la mer (van Bemmelen, 1949 ; Sémah F., 1984).
Les produits de son érosion se retrouvent dans les dépôts sédimentaires du dôme de Sangiran dans les couches de Kabuh plus précisément.
1.4.4. Les unités de la zone septentrionale
Il existe trois principales unités dans la zone septentrionale (figure 7), que nous ne
détaillerons pas ici (van Bemmelen, 1949 ; Djubiantono, 1992):
• La dépression de Randublatung : Elle se trouve au nord des collines des Kendeng. Son remplissage est constitué de
dépôts marins et continentaux allant du Miocène au Pléistocène. • Les collines de Rembang-Madura :
Elles se trouvent au nord-est de Java. Ces collines sont constituées principalement par des calcaires du Miocène.
• Les plaines alluviales du Nord : Elles se situent tout au long de la côte Nord de Java. Elles sont composées d’alluvions
transportées par les rivières qui se jettent dans la mer de Java.
Premier chapitre : Présentation du cadre de l’étude.
- 23 -
Figure 7 : Carte physiographique de l’île de Java
(d’après R.W. van Bemmelen, 1949)

Premier chapitre : Présentation du cadre de l’étude.
- 24 -
1.5. Climat
Traversée par l’équateur, l’Indonésie connaît, selon les îles, un climat tropical ou équatorial. La zone équatoriale (Sumatra, Bornéo, Célèbes, Papua), au climat très humide, ne connaît qu’une très courte saison sèche. Dans la zone tropicale (Java, Bali, petites îles de la Sonde), la saison humide (de novembre à avril environ) alterne avec une longue saison sèche (de mai à octobre environ) (Whitten et al., 1996).
Les variations annuelles des températures indonésiennes sont faibles : de 24 à 26°C en moyenne, la température ne descendant jamais en dessous de 20°C, sauf en montagne. La pluviosité annuelle moyenne oscille entre 1 780 et 3 175 mm en plaine et peut atteindre 5 000 mm dans les régions montagneuses de Kalimantan, où tous les mois sont pluvieux (Whitten et al., 1996 ; encyclopédie Microsoft Encarta 2000).
Java connaît un climat tropical de moussons avec deux saisons, une humide et une sèche lié au déplacement de la Zone de Convergence Inter-Tropicale (Z.C.I.T.) (Koninck, 1994). La longueur de ces saisons varie suivant les régions de Java. En effet, l’île de Java se localise à cheval sur les zones orientale et occidentale de l’Indonésie et subit donc ces deux influences. La zone orientale plus sèche est soumise aux alizés (climat semi-aride) alors que la zone occidentale est très humide et voit se développer une forêt tropicale humide (climat tropical semi-humide) (Sevin, 1993 ; Whitten et al. 1996).
1.6. Végétation
La flore indonésienne, comme la faune, subi deux influences, l’une asiatique à l’ouest et l’autre australienne à l’est. L’isolement insulaire a favorisé le développement de l’endémisme (Groves, 1984).
L’ Indonésie est un pays forestier où il existe de forts contrastes de végétation d’une île à l’autre (Chacornac-Rault, 2004). Cet archipel présente une grande diversité de milieux : forêts tropicales humides, plaines alluviales, mangroves, volcans en activité, récifs coralliens jusqu’à des glaciers sur son plus haut sommet (5030 mètres Puncak Jaya, Papua) (Whitten et al., 1993).
Presque 65 % du territoire indonésien sont couverts de forêts équatoriales ou tropicales et de savanes arborées, surtout concentrées à Kalimantan, Sumatra et dans l’est de l’archipel. L’essence dominante dans ces forêts tropicales est Dipterocarpus (Backer & Bakhuisen Van Den Brinck, 1965).
L’Indonésie compte environ 2 500 espèces d’arbres et près de 5 500 espèces végétales à fleurs. L’île de Bornéo, notamment, détient l’un des écosystèmes les plus riches au monde (800 espèces d’arbres) (encyclopédie Microsoft Encarta 2000).

Premier chapitre : Présentation du cadre de l’étude.
- 25 -
À Java, le type de végétation dépend de nombreux paramètres : l’insularité, l’activité volcanique, la pluviométrie, l’altitude mais aussi et surtout de l’exploitation du milieu par l’homme.
Les grandes formations végétales présentes sur Java sont (Backer & Bakhuizen van den Brinck, 1965):
- La forêt tropicale humide, lorsque la pluviosité est supérieure à 2 000 millimètres, - La forêt de mousson, lorsque la pluviosité est inférieure à 2 000 millimètres pendant
plus de deux mois, - La forêt marécageuse, - Les formations de plages, - La forêt de mangrove.
La végétation actuelle de l’île de Java a été modifiée par l’homme. Beaucoup de zones de Java est présentent actuellement des formations végétales qui ne caractérisent pas le couvert originel (Sémah F. et al., 2002). La forme primaire de cette végétation n’existe quasiment plus, elle a été remplacée par les cultures (champs, rizières, plantations forestières) (Durand, 1994).
Il est possible de retrouver les influences des deux grandes tendances climatiques actuelles dans les zones refuges de cette végétation primaire :
- À l’ouest, une région humide où domine la forêt tropicale humide (Rain Forest) avec par exemple les espèces de type Dipterocarpus, Podocarpus et Altingia,
- À l’est, une région à saison sèche marquée où se développe une forêt plus ouverte, proche de la forêt de mousson (Sémah A.M., 1993) avec les taxons de type Mimosaceae, Poaceae et Mallotus, par exemple.
1.7. Faune
L’Indonésie se situe à un carrefour de diverses zones biogéographiques et donc possède une biodiversité unique. Elle appartient à deux des six provinces indomalaises décrites par Corbet et Hill (1992) :
- La province sondaïque qui inclut la Thaïlande du sud et la Malaisie, Sumatra, Bornéo, Palawan, Mentawai, et Java, et,
- La province de Wallace qui inclut Célèbes, les petites îles de la Sonde, et les Moluques.
Cependant suivant l’origine des espèces, trois grandes zones biogéographiques distinctes sont observables:
- La province sondaïque ou la plate forme de la Sonde dont les faunes montrent une forte influence continentale (figure 8),

Premier chapitre : Présentation du cadre de l’étude.
- 26 -
- La province de Wallace (Wallacea) qui se situe à l’est de la ligne de Wallace, où les faunes présentent un fort endémisme et peu d’influence continentale ou australienne (figure 8),
- La province australienne (Papouasie, Nouvelle Guinée, Australie) ou plate-forme de Sahul dont les faunes présentent de fortes influences australiennes (figure 8).
Ces trois provinces ont été limitées par des lignes zoogéographiques proposées par
divers auteurs (Wallace, 1860 ; Huxley, 1868 ; Weber, 1902 ; Pelseneer, 1904 ; Beaufort, 1913 ; Mayr, 1944) (figure 8) suivant la nature et la répartition des différentes espèces d’animaux.
De 1854 à 1862, Wallace, un naturaliste britannique sillonna l’Indonésie et collecta de nombreux spécimens d’animaux pour étudier la distribution des espèces (van den Bergh, 1999). À la suite de ses recherches, Wallace proposa une limite zoogéographique, « la ligne de Wallace », qui séparait la plate-forme de la Sonde de Célèbes à l’ouest et de Lombok à l’est (Wallace, 1860). Elle passe entre Bali et Lombok et entre Bornéo et Célèbes.
Figure 8 : Carte des provinces et des limites zoogéographiques de l’Asie du sud-est avec un abaissement
du niveau marin d’environ 120 mètres. (zones gris foncé : terres émergées actuellement ; zones gris clair : zones émergées lors des maximums
glaciaires avec le réseau hydrographique en noir ; zones blanches : mer ou océan ; d’après Mayr, 1944, Simpson, 1977 et van den Bergh, 1999)

Premier chapitre : Présentation du cadre de l’étude.
- 27 -
En 1868, Huxley proposa une extension de la ligne de Wallace en direction du nord. Cette ligne passe entre Bali et Lombok, et entre Bornéo et Célèbes mais elle tourne dans le Pacifique entre Mindanao et les îles Sanghir. Elle ne limite pas les régions indomalaise et australienne mais indique plutôt la limite de la plate-forme de la Sonde qui fut émergée pendant les phases glaciaires pléistocènes (figure 8).
Par la suite, Pelseneer (1904) proposa une ligne séparant les îles de l’ouest où les éléments indomalais étaient prédominants des îles de l’est sur lesquelles les éléments australo-papouasiens avaient une supériorité numérique. Cette ligne fut nommée ligne de Weber (figure 8).
Enfin, une ligne qui limitait la région australe à l’ouest a été nommée la ligne de Lydekker et fut citée par de Beaufort (1913). Elle passe entre l’Australie et Timor, les îles Aru et Kai, Missol et Seram et Waigeao et Halmahera (figure 8).
La multiplication des lignes zoogéographiques ne permettait pas d’avoir une vision claire car les différents chercheurs s’étaient basés sur l’étude de groupes d’animaux (chauve-souris, mammifères, oiseaux…) mais non sur l’ensemble de la faune (Simpson, 1977). La faculté de dispersion ou de franchissement des bras de mer n’est pas la même pour tous les groupes d’organismes et peut varier au sein d’un même d’un groupe. Nous retiendrons la proposition de Simpson (1977) qui définit deux ensembles : une région faunique à l’ouest comprenant le plateau de la Sonde et limitée par la ligne d’Huxley, et une région faunique australienne comprenant le plateau de Sahul et limitée par la ligne de Lydekker. Entre ces deux régions, la zone est considérée comme intermédiaire et est appelée « Wallacea » ou zone de Wallace (figure 8).
Alors que Wallace s’est basé sur l’étude de groupes vivants sans disposer de données sur la géologie, la théorie des plaques tectoniques et l’augmentation des données géologiques et géophysiques dans le sud-est asiatique ont apporté des informations plus complètes sur la formation des différentes îles (van den Bergh, 1999). Dans ce cadre, il ne semble pas incohérent de définir la zone entre les plateaux de la Sonde et de Sahul comme une zone de transition subissant les influences de ces deux grandes régions fauniques avec des dispersions locales. Les faunes de chaque île devrait être étudiée séparément car elles possèdent des compositions, des affinités, des histoires et des écologies différentes (van den Bergh, 1999). La province qui nous intéresse est la province sondaïque, mais nous devons tenir compte des interactions de toutes ces provinces.
La province sondaïque est redécoupée en six sous-provinces (Corbet & Hill, 1992) : • la sous-province malaise (Thaïlande du sud et Malaisie), • la sous-province de Sumatra, • la sous-province de Mentawai, • la sous-province de Java,

Premier chapitre : Présentation du cadre de l’étude.
- 28 -
• la sous-province de Bornéo et • la sous province de Palawan.
La faune de la sous-province de Java est quelque peu appauvrie comparée à celles de Sumatra et de Bornéo (Corbet & Hill, 1992). À Java, les ordres de petite taille sont majoritaires comme les chiroptères et les rongeurs (figure 9). Les herbivores ne représentent que 9 % du total des groupes.
5%
47%
4%10%
1%8%
21%1%
1%
1%1%
Pholidota
Insectivora
Scandentia
Dermoptera
Chiroptera
Primates
Carnivora
Perissodactyla
Artiodactyla
Rodentia
Lagomorpha
Figure 9 : Proportion des ordres de mammifères actuels de Java. (d’après Corbet & Hill, 1992)
Cette distribution est probablement due aux extinctions causées par la perte de
l’habitat avec ou sans exploitation directe par l’homme. En effet, Java a été plus fortement peuplée par les hommes et plus intensivement cultivée (champs, rizières, plantations forestières) que les autres îles depuis au moins 2 000 ans, ce qui a fait disparaître le milieu de vie de beaucoup d’espèces (Durand, 1994).
Les faunes des provinces sondaïque et indochinoise ont beaucoup plus d’affinités entre elles, car elles possèdent 47 espèces en commun, qu’avec la faune des Philippines, avec laquelle elles n’en partagent que 9 (Corbet & Hill, 1992 ; Tougard, 1998).
En outre, le taux d’endémisme varie suivant les îles. Les îles de la Sonde ont un taux d’endémisme assez bas : Java 23 %, Bornéo 32,5 %, par rapport à celui de Luzon ou Mindanao (79,5 % et 73,3 %) (Groves, 1984, 1985). Ceci confirme que les îles de la Sonde ont été en connexion, dans un passé récent, entre elles et avec le continent et que les Philippines ont été, au contraire, isolées. Cependant certaines espèces sont communes aux Philippines et aux îles de la Sonde, donc une migration a dû s’effectuer entre les Philippines et Célèbes (Groves, 1984, 1985).

Premier chapitre : Présentation du cadre de l’étude.
- 29 -
1.8. Migrations
Les connexions entre les îles de la Sonde et le continent se sont effectuées à la faveur des abaissements du niveau marin lors des périodes glaciaires. Ces abaissements ont permis la création de ponts terrestres qui ont favorisé les migrations d’un certain nombre d’espèces.
Néanmoins, les ponts terrestres ne se formaient pas entre chaque île à chaque glaciation et les migrations étaient possibles pour certains animaux ayant des facultés pour nager ou flotter et pour se déplacer en troupeau (de Vos et al., 1994 ; Dermitzakis et al., 1997). En effet, le système digestif des hippopotames, des éléphants et des cerfs produit de nombreux gaz, ce qui leur permet de bien flotter dans l’eau. La présence de ces espèces dans une faune fossile suggère la possibilité de migrations en milieu insulaire.
Les éléphants aiment se baigner et certains individus qui vivent sur des îles vont actuellement dans la mer ouverte loin des côtes du Sri Lanka et d’Inde et traversent les larges estuaires du Bangladesh (Sondaar, 1977 ; Johnson, 1980). Les cerfs fuient dans l’eau s’ils sont en danger et ont souvent été observés nageant.
La passage de certains bras de mer a également été possible pour des petits animaux embarqués sur des radeaux naturels (Simpson, 1965).
Les deux grands modèles proposés pour les migrations sur les îles de la Sonde sont (figure 10) :
- Une voie de migration continentale passant par la Malaisie provenant de l’Inde ou de la Chine,
- Une voie de migration passant par les Philippines. La deuxième voie de migration semble plus improbable, car la profondeur du bras de
mer séparant Taiwan et Luzon peut atteindre 1 000 mètres (Heaney, 1985). Cette valeur est bien supérieure au maximum d’abaissement du niveau marin constaté à 0,8 Ma (environ 150 mètres). S’il y a eu des migrations entre ces deux îles, elles n’ont pu s’effectuer que par des animaux nageurs ou des petits mammifères ayant traversé sur des radeaux naturels.
En revanche, la profondeur du bras de mer entre Palawan et Bornéo n’est que de 145 mètres au maximum, ce qui a sans doute permis des migrations entre ces deux îles. Ceci est confirmé par la composition de la faune actuelle de l’île de Palawan. En effet, 58 % des espèces de mammifères de cette île se retrouvent à Bornéo (Heaney, 1985).

Premier chapitre : Présentation du cadre de l’étude.
- 30 -
Figure 10 : Voies de migrations possibles lors de la formation de la plate-forme de la Sonde (= Sundaland)
au cours du Pléistocène moyen. (zone grises : terres émergées ; zones blanches : mer ou océan ; a : route indo-malaise du Pléistocène inférieur et
moyen ; b : route sino-malaise du Pléistocène supérieur ; fond de carte Voris, 2000)
La faune des Philippines, bien qu’isolée, présente plus d’affinités avec la faune de la
province sondaïque qu’avec celle de Chine du sud. Aucune forme provenant de Chine du sud ne se retrouve aux Philippines. Par conséquent, il semble que les Philippines aient été isolées de la zone continentale de l’Asie, excepté entre Bornéo et Palawan (Heaney, 1985 ; Groves, 1985).
Pour Java, il semble improbable qu’une migration de Chine du Sud soit passée par les Philippines puis par Bornéo ou Célèbes.
Nous pouvons donc penser qu’une succession de migrations a eu lieu sur l’île de Java
depuis 2,6 Ma (von Koenigswald, 1934a, 1949), époque des premiers dépôts sédimentaires continentaux, et nous allons nous efforcer d’en retrouver les traces à travers l’histoire de la découverte des fossiles de mammifères dans différentes zones de Java.
1.9. Historique des recherches paléontologiques à Java
Les sites fossilifères javanais se situent fréquemment dans des zones de culture et les ossements de vertébrés ont attiré depuis toujours l’attention des villageois. Les Javanais y

Premier chapitre : Présentation du cadre de l’étude.
- 31 -
voyaient les restes des victimes des grandes batailles mythiques de la tradition du Wayang (théâtre javanais) et récoltaient ces fossiles. De nombreux sites paléontologiques étaient donc connus et avaient suscité l’intérêt des javanais bien avant que les chercheurs européens n’arrivent (Raden Saleh, 1894) (figure 11).
Figure 11 : Carte des principaux sites paléontologiques de Java
(d’après Sémah A.M. et al., 1993)
Passionné par la culture javanaise ancienne, le célèbre peintre et naturaliste Raden
Saleh avait visité ces sites et écrit plusieurs articles à leur sujet dans les années 1860. Il avait même organisé des fouilles dans la région de Kedung Brubus (figure 11) (Sémah F. et al., 1993a).
1.9.1. Les premières découvertes : 1888-1900
En 1888, van Rietschoten découvrit le premier crâne humain fossile à Wajak (Java est)
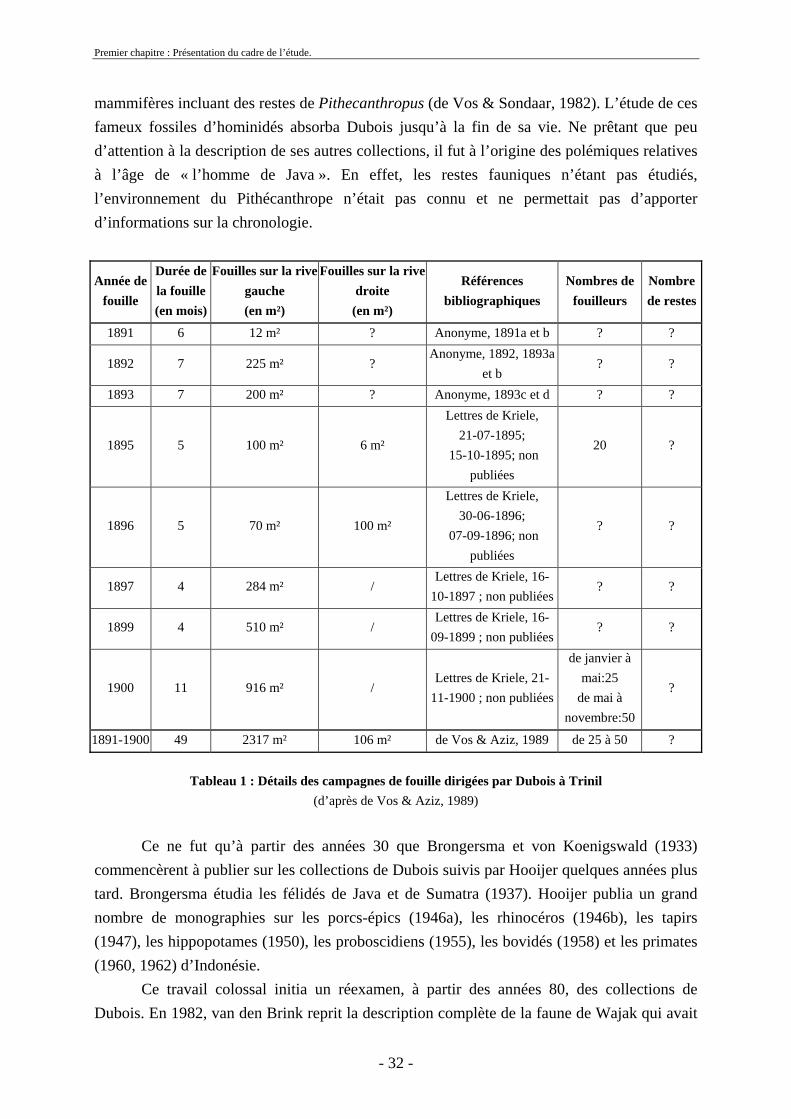
qui fut attribué plus tard à Homo sapiens. La même année, Dubois, jeune médecin néerlandais qui recherchait à l’ouest de Sumatra le « chaînon manquant » de l’évolution de l’homme en Indonésie, s’est intéressé à cette découverte. Il entreprit en 1890 des fouilles sur ce site où de nombreux restes de mammifères sont découverts ainsi qu’un deuxième crâne nommé Wajak II (Dubois, 1920 ; von Koenigswald, 1949 ; van den Bergh, 1999). Il alla également à Kedung Brubus (figure 11) pour collecter des fossiles et y découvrit en 1889 un fragment de mandibule humaine qu’il attribua plus tard à Pithecanthropus erectus (Dubois, 1924). En 1891, il débuta des fouilles à Trinil (figure 11 et tableau 1) qui se sont poursuivies jusqu’en 1900, sur les rives de la rivière Solo. Sur ce site, il mit au jour des restes de Pithécanthrope (une calotte crânienne, des dents, un fémur) et un très grand nombre de restes fauniques (Dubois, 1908).
Dès 1890, Dubois fut le premier à diriger des fouilles et à collecter systématiquement les fossiles à Java (tableau 1). En 1895, il laissa la direction des fouilles de Trinil à Kriele et De Winter jusqu’en 1900, et revint en Europe avec une importante collection de reptiles et de
Premier chapitre : Présentation du cadre de l’étude.
- 32 -
mammifères incluant des restes de Pithecanthropus (de Vos & Sondaar, 1982). L’étude de ces fameux fossiles d’hominidés absorba Dubois jusqu’à la fin de sa vie. Ne prêtant que peu d’attention à la description de ses autres collections, il fut à l’origine des polémiques relatives à l’âge de « l’homme de Java ». En effet, les restes fauniques n’étant pas étudiés, l’environnement du Pithécanthrope n’était pas connu et ne permettait pas d’apporter d’informations sur la chronologie.
Année de fouille
Durée de la fouille (en mois)
Fouilles sur la rive gauche (en m²)
Fouilles sur la rive droite
(en m²)
Références bibliographiques
Nombres de fouilleurs
Nombre de restes
1891 6 12 m² ? Anonyme, 1891a et b ? ?
1892 7 225 m² ? Anonyme, 1892, 1893a
et b ? ?
1893 7 200 m² ? Anonyme, 1893c et d ? ?
1895 5 100 m² 6 m²
Lettres de Kriele, 21-07-1895;
15-10-1895; non publiées
20 ?
1896 5 70 m² 100 m²
Lettres de Kriele, 30-06-1896;
07-09-1896; non publiées
? ?
1897 4 284 m² / Lettres de Kriele, 16-
10-1897 ; non publiées? ?
1899 4 510 m² / Lettres de Kriele, 16-
09-1899 ; non publiées? ?
1900 11 916 m² / Lettres de Kriele, 21-
11-1900 ; non publiées
de janvier à mai:25
de mai à novembre:50
?
1891-1900 49 2317 m² 106 m² de Vos & Aziz, 1989 de 25 à 50 ?
Tableau 1 : Détails des campagnes de fouille dirigées par Dubois à Trinil (d’après de Vos & Aziz, 1989)
Ce ne fut qu’à partir des années 30 que Brongersma et von Koenigswald (1933)
commencèrent à publier sur les collections de Dubois suivis par Hooijer quelques années plus tard. Brongersma étudia les félidés de Java et de Sumatra (1937). Hooijer publia un grand nombre de monographies sur les porcs-épics (1946a), les rhinocéros (1946b), les tapirs (1947), les hippopotames (1950), les proboscidiens (1955), les bovidés (1958) et les primates (1960, 1962) d’Indonésie.
Ce travail colossal initia un réexamen, à partir des années 80, des collections de Dubois. En 1982, van den Brink reprit la description complète de la faune de Wajak qui avait

Premier chapitre : Présentation du cadre de l’étude.
- 33 -
été partiellement effectuée par Brongersma (1937) et Hooijer (1947, 1962). De Vos (1983, 1984, 1985, 1989, 1993) et Sondaar (1981, 1984), seuls ou en collaboration (1982, 1983, 1994), ont travaillé principalement sur la biostratigraphie de Java et ont remis en cause celle établie par von Koenigswald en 1934. Hardjasasmita (1987) a étudié les suidés d’Indonésie et van den Bergh (1999) les éléphantidés d’Indonésie.
Le matériel paléontologique, issu de ces fouilles ou de la prospection, est actuellement
conservé dans de très bonnes conditions au nouveau Naturalis Museum de Leiden (Pays-Bas). Nous avons eu la chance de pouvoir observer et étudier certains de ces fossiles.
En ce qui concerne Sangiran, Dubois visita le site en 1893 mais il ne décida pas d’engager de fouilles dans cette zone (Bartstra, 1985).
1.9.2. L’expédition Selenka : 1906-1908
Après le départ de Dubois de l’île de Java, une expédition pluridisplinaire allemande
fut organisée à Trinil entre 1906 et 1908. En 1906, elle était dirigée par les époux Selenka puis, suite à la mort de son époux, par Eleonor Selenka avec l’aide de Blanckenhorn (tableau 2). Ces campagnes de fouilles n’ont pas permis de découvrir de nouveaux fossiles d’Homo erectus. Néanmoins, un très grand nombre de restes de la faune représentative des couches de Trinil sensu stricto a été mis au jour. Cette faune a été décrite par Stremme et Janensch (Selenka & Blankenhorn, 1911). Ces études permirent pour la première fois de donner une image claire de la faune contemporaine du Pithécanthrope.
Les restes issus de ces fouilles ont été conservés à Berlin et Munich, malheureusement une partie de ces fossiles a disparu durant la deuxième guerre mondiale. Actuellement, ce qui reste de cette collection est conservé au Museum für Naturkunde, à l’Université Humboldt de Berlin (Allemagne).
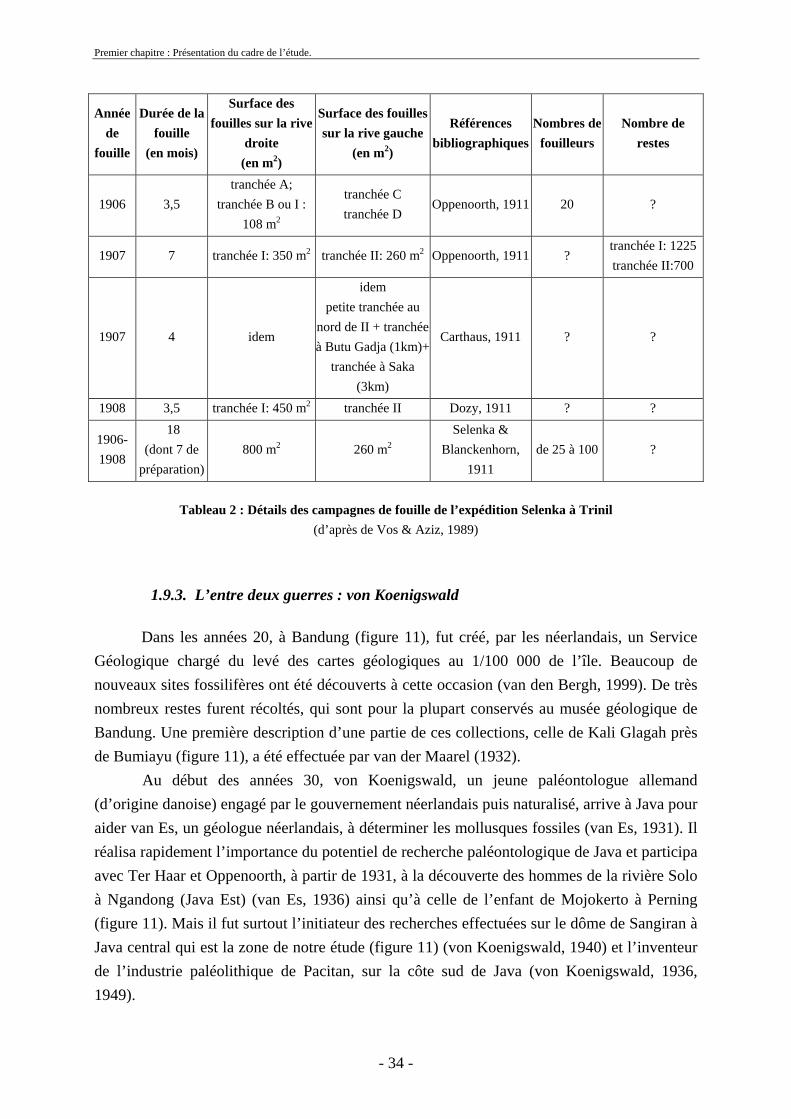
Premier chapitre : Présentation du cadre de l’étude.
- 34 -
Année de
fouille
Durée de la fouille
(en mois)
Surface des fouilles sur la rive
droite (en m2)
Surface des fouilles sur la rive gauche
(en m2)
Références bibliographiques
Nombres de fouilleurs
Nombre de restes
1906 3,5 tranchée A;
tranchée B ou I : 108 m2
tranchée C tranchée D
Oppenoorth, 1911 20 ?
1907 7 tranchée I: 350 m2 tranchée II: 260 m2 Oppenoorth, 1911 ? tranchée I: 1225 tranchée II:700
1907 4 idem
idem petite tranchée au
nord de II + tranchée à Butu Gadja (1km)+
tranchée à Saka (3km)
Carthaus, 1911 ? ?
1908 3,5 tranchée I: 450 m2 tranchée II Dozy, 1911 ? ?
1906-1908
18 (dont 7 de
préparation) 800 m2 260 m2
Selenka & Blanckenhorn,
1911 de 25 à 100 ?
Tableau 2 : Détails des campagnes de fouille de l’expédition Selenka à Trinil
(d’après de Vos & Aziz, 1989)
1.9.3. L’entre deux guerres : von Koenigswald
Dans les années 20, à Bandung (figure 11), fut créé, par les néerlandais, un Service
Géologique chargé du levé des cartes géologiques au 1/100 000 de l’île. Beaucoup de nouveaux sites fossilifères ont été découverts à cette occasion (van den Bergh, 1999). De très nombreux restes furent récoltés, qui sont pour la plupart conservés au musée géologique de Bandung. Une première description d’une partie de ces collections, celle de Kali Glagah près de Bumiayu (figure 11), a été effectuée par van der Maarel (1932).
Au début des années 30, von Koenigswald, un jeune paléontologue allemand (d’origine danoise) engagé par le gouvernement néerlandais puis naturalisé, arrive à Java pour aider van Es, un géologue néerlandais, à déterminer les mollusques fossiles (van Es, 1931). Il réalisa rapidement l’importance du potentiel de recherche paléontologique de Java et participa avec Ter Haar et Oppenoorth, à partir de 1931, à la découverte des hommes de la rivière Solo à Ngandong (Java Est) (van Es, 1936) ainsi qu’à celle de l’enfant de Mojokerto à Perning (figure 11). Mais il fut surtout l’initiateur des recherches effectuées sur le dôme de Sangiran à Java central qui est la zone de notre étude (figure 11) (von Koenigswald, 1940) et l’inventeur de l’industrie paléolithique de Pacitan, sur la côte sud de Java (von Koenigswald, 1936, 1949).

Premier chapitre : Présentation du cadre de l’étude.
- 35 -
En 1934, il découvrit des éclats de calcédoine et de jaspe à Sangiran sur le site de Ngebung (von Koenigswald & Gosh, 1973) ainsi que des restes fauniques (von Koenigswald, 1940). En 1935, voulant chercher l’origine de la matière première de ces éclats il part à Pacitan. Il y découvre un grand nombre d’outils et définit l’industrie paléolithique pacitanienne. À cette même période, aux environs du village de Punung à proximité de la ville de Pacitan (figure 11), de 1934 à 1937, il récolte de nombreuses dents de mammifères dans des fissures du karst. Par la suite, ces restes ont été étudiés par Badoux (1959).
Von Koenigswald, en 1934, établit la première biostratigraphie de Java (tableau 7) à partir de l’étude des fossiles des collections de Kali Glagah, de Ci Julang, de Jetis, de Trinil et de Ngandong (figure 11).
Les collections de fossiles récoltées par von Koenigswald sont conservées actuellement à l’Intitut Senckenberg de Francfort (Allemagne), au Quaternary Geology Laboratory de Bandung et au Geological Research and Development Center (GRDC) de Bandung (Indonésie).
Après 1934, le dôme de Sangiran devint le site à hominidés le plus riche de Java et l’un de ses plus importants gîtes fossilifères jusqu’à aujourd’hui.
Après la seconde guerre mondiale, von Koenigswald revînt à Java et il commença à
former les premiers chercheurs indonésiens. Dans les années 60, van Heekeren et Houbolt récoltèrent des fossiles sur le dôme de
Sangiran qui furent en partie décrits par Hooijer (1964).
1.9.4. De l’après guerre jusqu’à nos jours : le développement de la recherche indonésienne
Sartono (géologue et paléontologue), Soejono (préhistorien) et Jacob (anthropologue)
ont poursuivi l’œuvre de von Koenigswald et de van Heekeren, à partir des années 60 et ont mis en place des recherches systématiques : fouilles et prospections. Dans les années 70, le Centre National de Recherches Archéologiques Indonésien a développé un programme de recherches en collaboration avec le laboratoire de bio-paléoanthropologie de la faculté de médecine de l’université de Gajah Mada (Yogyakarta).
Les études se sont poursuivies principalement sur le dôme de Sangiran où des dizaines d’individus d’Homo erectus ont été découverts mais des fouilles ont aussi été menées sur les sites de Miri, Sambungmacan, Ngawi, Ngandong, Pati Ayam et Trinil (figure 11). La majorité des restes fauniques mis au jour depuis lors sont conservés à Java, au laboratoire de bio-paléoanthropologie de l’université de Gajah Mada de Yogyakarta, au Centre National de Recherches Archéologiques Indonésien de Yogyakarta, au Geological Research and Development Center (G.R.D.C.) de Bandung, au Centre National de Recherches Archéologiques Indonésien de Jakarta et au Musée de Sangiran.

Premier chapitre : Présentation du cadre de l’étude.
- 36 -
Les restes fauniques faisant l’objet de notre étude proviennent du laboratoire de bio-paléoanthropologie de l’université de Gajah Mada de Yogyakarta.
Une collaboration nippo-indonésienne entre l’Université de Tokyo et le Service Géologique indonésien, entre 1976 et 1979, a permis de mener une étude pluridisciplinaire sur la stratigraphie, la paléontologie et la chronologie de divers sites javanais dont Sangiran (Watanabe & Kadar, 1985). Aimi et Aziz ont étudié les restes fauniques découverts lors de sondages réalisés pendant cette période et ont proposé des listes fauniques pour les sites de Sambungmacan, Trinil, Mojokerto et Sangiran. À Sangiran, des tranchées ont été creusées sur le site de Bukuran dans les couches de Pucangan (cf. 1.10.2.2.) et des ramassages de surface ont été effectués à Tanjung, Sendang Busik, Bukuran et Dayu (figure 12 et annexe 1).
Ces fossiles sont conservés au Geological Research and Development Center (G.R.D.C.) de Bandung.
En 1989, une équipe franco-indonésienne du Muséum national d’Histoire naturelle de
Paris et du Centre National de Recherches Archéologiques Indonésien de Jakarta a entrepris une étude stratigraphique des dépôts du dôme de Sangiran (Widianto et al., 1994). A Bukuran, à la base de la formation de Pucangan a été découvert un squelette d’Hexaprotodon sivalensis koenigswaldi. Il est actuellement étudié par Moigne et Due Awe (M.Q.P.I.).
À partir des années 90, Simanjuntak, Djubiantono et Widianto, du Centre National de
Recherches Archéologiques Indonésien de Jakarta et de Yogyakarta, ont organisé plusieurs campagnes de fouilles sur le dôme de Sangiran. De 1990 à 1998, ces fouilles furent menées sur la zone de Ngebung 2 (figure 12 et annexe 1), à Sangiran, en collaboration avec une équipe du Muséum national d’Histoire naturelle de Paris dirigée par F. Sémah.
Lors de celles-ci trois niveaux ont pu être mis en évidence (A, B et U) dans la formation de Kabuh (cf. 1.10.2.2.). Dans le niveau A des restes fauniques ont été découverts en association avec des outils, ce qui est la preuve à Sangiran d’un niveau d’occupation humaine et suggère une possible interaction entre les hommes et les animaux. Une dent (NG91/G-10) a été mise au jour dans la couche archéologique de Ngebung et un fragment de fémur d’Homo erectus (Kresna 11) a également été trouvé sur l’affleurement du Grenzbank dans le périmètre de fouille (Sémah F. et al., 1993b, 2001a, 2002). Le matériel faunique a été étudié par Moigne et Due Awe (Mission Quaternaire et Préhistoire en Indonésie M.N.H.N./C.N.R.S./I.R.D.) (Moigne et al., 2004) et est conservé au Musée de Miri.
Nous utiliserons Ngebung 2 comme site de comparaison tout au long de notre étude.
Parallèlement, la région des Montagnes du sud a suscité un intérêt croissant. En 1953, Soejono et Basoeki prospectèrent la région (van Heekeren, 1972). Depuis 1992, Simanjuntak du Centre National de Recherches Archéologiques Indonésien de Jakarta organise régulièrement des fouilles et des prospections dans cette zone.

Premier chapitre : Présentation du cadre de l’étude.
- 37 -
Depuis 1994, des fouilles sont menées à proximité du village de Punung dans les grottes de Song Terus et de Tabuhan, par la Mission du Quaternaire et de la Préhistoire en Indonésie (Mission Quaternaire et Préhistoire en Indonésie M.N.H.N./C.N.R.S./I.R.D.). Un grand nombre d’outils lithiques et de fossiles ont été découverts lors de ces campagnes ainsi que des restes humains et une sépulture d’Homo sapiens à Song Terus (Détroit, 2002 ; Sémah F. et al., 2002). La majorité des découvertes archéologiques est conservée dans le village de Punung. La faune est en cours d’étude actuellement (M.Q.P.I.) et va permettre de compléter les données sur la biostratigraphie du Pléistocène moyen et supérieur à Java.
En conclusion, nous remarquons que l’île de Java est une zone privilégiée pour les
découvertes paléoanthropologiques et paléontologiques dans le sud-est asiatique. La majorité des restes paléoanthropologiques proviennent du site de Sangiran.
Peu de restes fauniques ont été trouvés lors de fouilles sur le dôme de Sangiran (Aimi & Aziz, 1985 ; Koeshardjono, 1988 ; Moigne et al., 2004 ; Sémah F. et al., 1993b, 2001a, 2002). C’est le faible nombre de ces restes qui justifie de prendre en compte l’ensemble des collections les plus complètes qui n’ont pas encore été étudiées à ce jour pour comprendre la succession des migrations à Sangiran et donc à Java au Plio-Pléistocène.
1.10 Présentation du dôme de Sangiran
1.10.1. Cadre géographique
1.10.1.1. Actuel
Sangiran est un des plus importants sites à Homo erectus dans le monde. Il est localisé dans la plaine de Kalioso-Surakarta à 15 km au nord de la ville de Solo dans le centre de l’île de Java (figure 11). Il s’étend sur 4 districts administratifs, respectivement : Gondangrejo, Kalijambe, Gemolong, et Plupuh. Le nom « Sangiran » est celui d’un hameau se situant au centre du dôme.
1.10.1.2. Paléogéographie
Une régression a débuté dans la dépression de Solo à la limite Gauss-Matuyama, vers 2,6 Ma. À Sangiran, l’environnement passe d’une mer ouverte à une lagune (Djubiantono & Sémah F., 1993). À la limite Plio-Pléistocène, à environ 1,8 Ma, une importante phase volcanique entraîne le dépôt de lahars dans cette zone. Ceux-ci ont en partie comblé le lagon et permis le dépôt d’argiles de marais littoral (Djubiantono & Sémah F., 1993). A la fin du Pléistocène inférieur, vers 0,8 Ma, l’influence marine disparaît peu à peu à Sangiran et se

Premier chapitre : Présentation du cadre de l’étude.
- 38 -
dépose donc un conglomérat où se mêlent des mollusques marins avec des restes fauniques continentaux et des restes humains. La dépression de Solo est limitée au sud par les Montagnes du Sud et au nord par les collines des Kendeng qui se sont formées au Pliocène supérieur. Les produits de l’érosion de ces montagnes se retrouvent, au Pléistocène moyen, dans la formation de Kabuh sous forme de calcédoine, de bois silicifié et de quartz (van Bemmelen, 1949 ; Sartono, 1964b). L’activité volcanique a joué un rôle important lors de la régression marine au niveau du comblement et de la formation des volcans dans cette zone, ce qui a permis, à terme, d’isoler complètement Sangiran du milieu marin (Djubiantono, 1992 ; Sémah F. et al., 2001a). Ces volcans étaient proches des grands volcans actuels tels que le Lawu, qui appartiennent à l’arc interne de la Sonde. Les anciens volcans étaient déjà présents il y a environ 2 Ma, à la fin du Pliocène et ont formé d’importants dépôts tels des lahars et des brèches (van Bemmelen, 1949).
À Sangiran, les produits volcaniques se retrouvent lors de deux grandes périodes, à la limite Plio-Pléistocène, (environ 1,8 Ma) à la base de la formation de Pucangan sous forme d’épais lahars, ainsi que dans la formation de Notopuro (environ 0,2 Ma) (Sémah F., 1984) sous forme de dépôts volcano-sédimentaires correspondants à l’émergence des grands volcans actuels.
1.10.2. Cadre géologique et stratigraphique
Le dôme de Sangiran se trouve dans la dépression de la rivière Solo (figures 7 et 11) et fait partie d’un ensemble de dômes érodés comme Gemolong, Onto et Bringinan. Ils ont pour origine des bombements magmatiques avortés ou des remontées diapiriques comme dans le cas de Sangiran (Sémah F., 1984 ; Sémah A.M. et al., 1993).
1.10.2.1. Origine du dôme
Du point de vue géologique, Sangiran est un anticlinal érodé (figure 12). Cet anticlinal
est approximativement orienté Nord-Sud (axe à N 30 E) et mesure 8 km sur 6 (Sémah F., 1984). Ce dôme se trouve en bordure des collines des Kendeng et est recoupé par la rivière Cemoro dans une direction est-ouest (Sémah F., 1984). Cette rivière serpente à environ 100 m d’altitude, avant de rejoindre la rivière Solo, ce qui confère une morphologie de « boutonnière » au dôme de Sangiran. Ceci forme donc un bassin entouré de collines pouvant atteindre l’altitude de 180 mètres au-dessus du niveau de la mer (van Es, 1931).
1.10.2.2. Stratigraphie du dôme
L’intérêt majeur de ce site, du point de vue stratigraphique, est de présenter une série
complète du Pliocène final au Pléistocène moyen (Wantanabe & Kadar, 1985 ; Sémah F.,
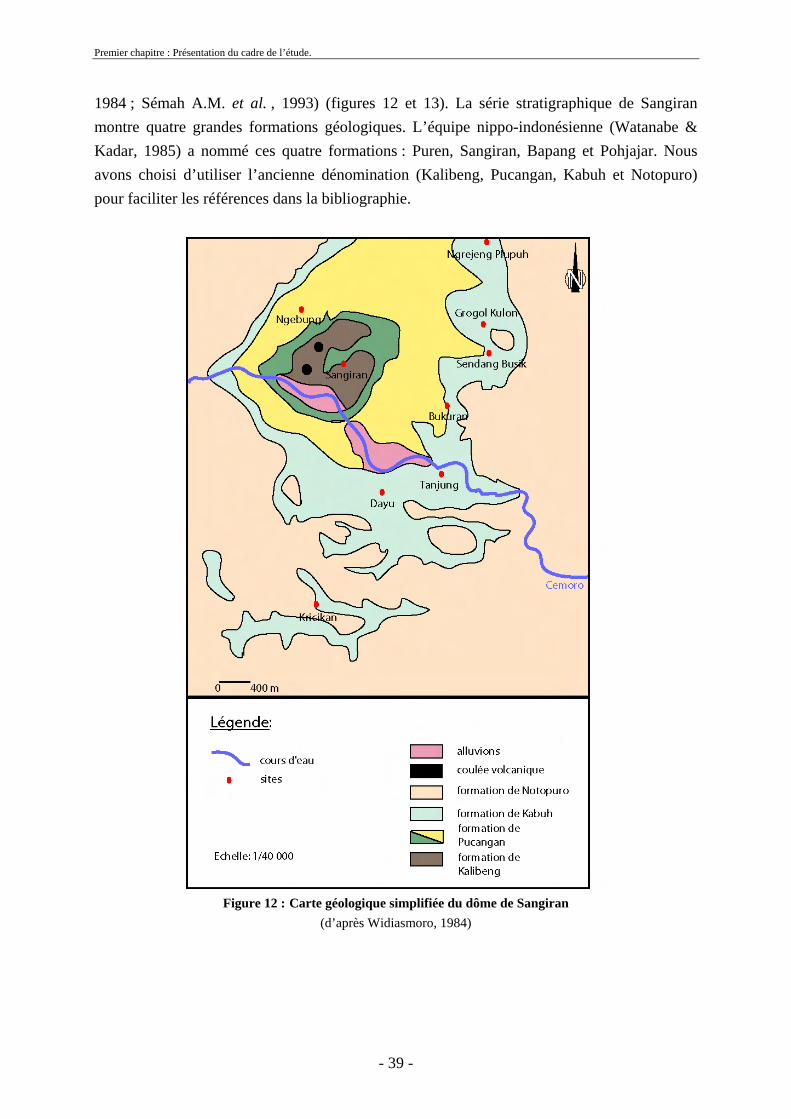
Premier chapitre : Présentation du cadre de l’étude.
- 39 -
1984 ; Sémah A.M. et al. , 1993) (figures 12 et 13). La série stratigraphique de Sangiran montre quatre grandes formations géologiques. L’équipe nippo-indonésienne (Watanabe & Kadar, 1985) a nommé ces quatre formations : Puren, Sangiran, Bapang et Pohjajar. Nous avons choisi d’utiliser l’ancienne dénomination (Kalibeng, Pucangan, Kabuh et Notopuro) pour faciliter les références dans la bibliographie.
Figure 12 : Carte géologique simplifiée du dôme de Sangiran
(d’après Widiasmoro, 1984)

Premier chapitre : Présentation du cadre de l’étude.
- 40 -
• La formation de Kalibeng (= Puren)
La série stratigraphique débute avec la formation de Kalibeng supérieur (Pliocène supérieur) car le Kalibeng inférieur n’est pas visible sur les coupes du dôme de Sangiran. Le faciès principal consiste en des argiles bleues (blue clays) (figures 13 et 14) et grises, à Turritella, souvent riches en matière organique qui contiennent à plusieurs reprises des passées plus sableuses très riches en foraminifères et mollusques (Sémah F., 1984).
Au sommet de la série, il existe un banc de calcaire côtier à Balanus. Celui-ci caractérise un faciès marin peu profond correspondant à une phase de régression (figure 13). Ce calcaire représente le niveau de transition avec la formation suivante. Il est suivi par un dépôt de grès pyroclastique jaunâtre à Corbicula exporrecta M. qui suggère les premiers niveaux d’eau douce (Lizon-Sureau, 1979).
Figure 13 : Colonne stratigraphique du dôme de Sangiran
(d’après Sémah F. et al., 1998)

Premier chapitre : Présentation du cadre de l’étude.
- 41 -
Figure 14 : Argiles bleues lagunaires de Kalibeng supérieur (vers –2 Ma)
(photo © Semenanjung)
• La formation de Pucangan (= Sangiran)
La formation de Pucangan (Pléistocène inférieur) (figures 12 et 13), débute par un
niveau de brèche volcanique et de lahars d’une épaisseur variable (figure 13) qui peut être séparé du niveau à Corbicula par quelques mètres de dépôts argileux (Sémah F., 1984 ; Sémah F. et al., 2001a). La présence de ces lahars paraît refléter un développement de l’activité volcanique dans la région à cette époque. Dans ces niveaux ont été retrouvés les plus anciens fossiles de vertébrés continentaux de Sangiran (dents de crocodiles) (van Es, 1931). La stratigraphie se poursuit par des argiles noires palustres et lacustres (black clays) (figure 15) parfois recoupées par des couches de cendres volcaniques ou de diatomites (Lizon-Sureau, 1979). L’épaisseur de cette formation varie entre 75 et 100 mètres. Les dépôts argileux sont riches en matière organique suggérant un milieu de dépôt plutôt proche d’un milieu palustre ou lacustre. Deux passées de diatomites ont été observées ce qui laisse à penser que deux transgressions marines au moins ont eu lieu à cette époque à Sangiran (Sémah F., 1984). Les déterminations effectuées dans la plus récente caractérisent un milieu littoral saumâtre (Lizon-Sureau, 1979).
Des fossiles de vertébrés ont été découverts à plusieurs reprises ainsi que des restes d’Homo erectus dans cette formation de Pucangan (von Koenigswald, 1940 ; Aimi & Aziz, 1985 ; Widianto et al., 1994).

Premier chapitre : Présentation du cadre de l’étude.
- 42 -
Figure 15 : Argiles noires de Pucangan à Bukuran.
(photo Bouteaux)
• La formation de Kabuh (= Bapang) La transition avec la formation précédente se fait au niveau d’un niveau nommé
« Grenzbank » (von Koenigswald, 1940). Il s'agit d'un conglomérat calcifié où se retrouvent mélangés des éléments marins et des graviers provenant de l’érosion des collines des Kendeng et des montagnes du sud (figure 7). Ce niveau n’est pas observable sur toutes les coupes du dôme de Sangiran, il est absent par exemple près de la coupe de Pagerejo (Sémah F., 1984). Ce faciès marque un changement drastique dans la sédimentation car il est caractéristique du comblement définitif de la lagune. Après la formation de Pucangan, l’influence marine disparaît définitivement et laisse place à des dépôts continentaux (Sémah F. et al., 2002).
Le Grenzbank, et également les sédiments se situant au-dessus, sont composés de dépôts synorogéniques provenant des collines des Kendeng et des montagnes du sud qui se soulèvent une nouvelle fois vers 0,9 Ma ainsi que de dépôts volcaniques (van Bemmelen, 1949; Sémah F., 1984 ).
La formation volcano-sédimentaire de Kabuh (Pléistocène moyen) (figures 12 et 13) est composée d’argiles, de graviers et de sables fluviatiles à stratification entrecroisée (figure 16). Dans la fraction grossière, des petits galets de calcédoine translucide qui ont servi de matière première aux éclats de Sangiran, ont été observés ainsi que des tectites et des ponces (Sémah F., 1984).
Cette formation a livré un grand nombre de restes fauniques ainsi que la majeure partie des hominidés fossiles de Sangiran.

Premier chapitre : Présentation du cadre de l’étude.
- 43 -
Figure 16 : Lits à stratifications entrecroisées de Kabuh à Bukuran.
(photo Bouteaux)
La formation de Kabuh a été plus précisément étudiée à Bapang (Larick et al., 2001). Cinq cycles ont pu être mis en évidence (C1 à C5).
Chaque cycle est composé de deux faciès sédimentaires (a et b) : - Les couches du faciès « a » sont composées de sables et de graviers à stratifications
planes ou entrecroisées. Les « clasts » sont composés d’andésite et de ponces dacitiques arrondies aussi bien que de quartz micro cristallin provenant de l’altération des roches carbonatées. Ces couches caractérisent un régime fluvio-lacustre.
- Les couches du faciès « b » sont composées de sables et de limons tufacés provenant de l’altération de verre volcanique mélangés à des cristaux angulaires à subangulaires et des « clasts » altérés. C1b, C2b et C4b ont été décrits respectivement auparavant comme « Lower, Middle and Upper Tuffs » (Suzuki et al., 1985). Ces couches caractérisent des niveaux de cendres altérées.
• La formation de Notopuro (= Pohjajar)
Les couches de Kabuh sont surmontées par une série discordante (Djubiantono, 1977)
de brèches volcaniques et de lahars dits de Notopuro (figure 17). Parfois, au-dessus de ceux-ci nous trouvons des dépôts alluviaux récents (figures 12 et 13).

Premier chapitre : Présentation du cadre de l’étude.
- 44 -
Figure 17 : Limite entre les couches de Kabuh et les lahars de Notopuro.
(photo Garrigues © Semenanjung)
De la fin du Pliocène jusqu'à l'Holocène, les formations sédimentaires, de Kalibeng, de Pucangan, de Kabuh et de Notopuro ont pu se déposer dans la région de Sangiran, et ce n'est qu'ensuite qu'a eu lieu le bombement du dôme et son érosion par la rivière Cemoro.
1.10.3. Palynologie
Dans le faciès des argiles bleues de la formation de Kalibeng, le pollen de
Rhizophoraceae, caractéristiques des mangroves (figure 18), prédomine dans la courbe pollinique de la mangrove. Les présences ou absences de ces grains de pollen permettent de suivre les légères oscillations de la ligne de rivage et de penser que Sangiran était, au Pliocène, une lagune entourée de mangroves (Sémah A.M., 1986).
Il y a 2,5 Ma environ, la mer qui couvrait la région de Solo commence à se retirer. Le contenu pollinique nous permet d’observer que le littoral est alors occupé par la forêt de mangrove. À cette période, les côtes présentent une forêt marécageuse et les reliefs une forêt tropicale humide.

Premier chapitre : Présentation du cadre de l’étude.
- 45 -
Figure 18 : Diagramme pollinique simplifié des séries de Kalibeng et de Pucangan de Sangiran.
(A : argiles bleues ; B : calcaire à Balanes ; C : argiles noires ; c1 : brèche volcanique ; c2 : diatomites ; c3 : tuf ; 1 : mangrove ; 2 : forêt de marais ; 3 : Cyperaceae ; 4 : forêt humide ; 5 : forêt ouverte ; 6 : Poaceae ; 7 : AP
(arboreal pollen) ; 8 : NAP (non arboreal pollen)) (in Sémah F. et al., 1998)
Ensuite dans la formation de Pucangan, le faciès des argiles « noires » renferme du pollen de taxons caractéristiques des mangroves et des plages, tels que les Rhizophoraceae et les Arecaceae (palmiers), dont le nombre diminue fortement. En parallèle, nous observons le développement de familles d’herbacées comme les Cyperaceae (figure 18) dans le diagramme pollinique, exception faite des niveaux à diatomites.
La composition de la forêt des reliefs varie selon les changements climatiques. Pendant les périodes à saison sèche marquée, la forêt est assez ouverte proche de celle actuelle à Java est. La forêt tropicale humide persiste et correspond probablement à des forêts galeries le long des zones plus humides comme les cours d’eau. Pendant les périodes plus humides, la forêt tropicale humide, telle que celle de Java ouest actuellement, domine le paysage.
À Sangiran, durant le Pléistocène inférieur, pendant lequel se dépose la formation de
Pucangan, la représentation de la mangrove dans les diagrammes polliniques régresse pour disparaître définitivement vers - 0,7 Ma et laisse la place à la forêt humide, avec la présence de pollens de Fagaceae, d’Altingia excelsa, d’Engelhardia et de Podocarpus.

Premier chapitre : Présentation du cadre de l’étude.
- 46 -
Dans les couches de Kabuh, la végétation, dont les caractères évoluent en fonction des oscillations climatiques, reflète des conditions globalement plus sèches. Le paysage est plus ouvert avec la présence de pollens de Poaceae, d’Asteraceae, de Fabaceae et de Mimosaceae (Sémah A.M., 1986 ; Sémah F. et al., 2002).
1.10.4. Datations
Beaucoup de datations ont été réalisées sur le dôme de Sangiran par différentes méthodes (tableau 3):
- Les méthodes K/Ar et 40Ar/39Ar (von Koenigswald, 1964 ; Jacob & Curtis, 1971 ; Jacob, 1975a ; Curtis, 1981; Nishimura et al., 1981 ; Orchiston & Siesser, 1982 ; Swisher et al., 1994 ; Saleki, 1997 ; Larick et al., 2001 ; Bettis et al., 2004),
- La méthode par traces de fission (TF) (Suzuki et al., 1982 ; Nishimura et al., 1978, 1981),
- Les calibrations paléomagnétiques (Sémah et al., 1980 ; Yokohama et al., 1980 ; Sémah F., 1982, 1986 ; Shimizu et al., 1985),
- La méthode U/Th et ESR (Saleki, 1997)
La plupart de ces méthodes sont basées sur des échantillons provenant de dépôts volcaniques et selon les méthodes les âges peuvent être très différents (tableau 3) parce que l’origine des produits volcaniques est difficile à établir à Sangiran et plus précisément dans les couches de Kabuh. Il existe donc toujours un risque que les échantillons, telles que les ponces, aient pu être remaniées à partir de structures volcaniques beaucoup plus anciennes (Sémah F. et al., 2001a ; Langbroek & Roebroeks, 2000).
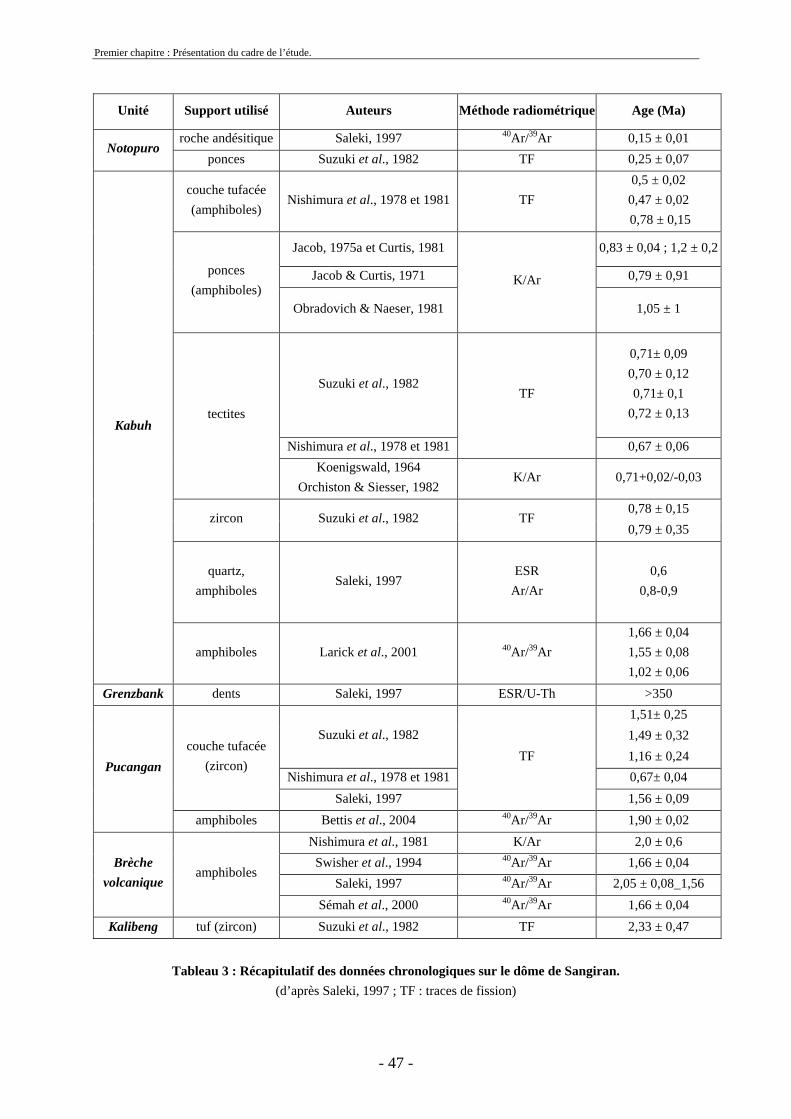
Premier chapitre : Présentation du cadre de l’étude.
- 47 -
Unité Support utilisé Auteurs Méthode radiométrique Age (Ma)
roche andésitique Saleki, 1997 40Ar/39Ar 0,15 ± 0,01 Notopuro
ponces Suzuki et al., 1982 TF 0,25 ± 0,07
couche tufacée (amphiboles)
Nishimura et al., 1978 et 1981 TF 0,5 ± 0,02
0,47 ± 0,02 0,78 ± 0,15
Jacob, 1975a et Curtis, 1981 0,83 ± 0,04 ; 1,2 ± 0,2
Jacob & Curtis, 1971 0,79 ± 0,91 ponces (amphiboles)
Obradovich & Naeser, 1981
K/Ar
1,05 ± 1
Suzuki et al., 1982
0,71± 0,09 0,70 ± 0,12 0,71± 0,1
0,72 ± 0,13
Nishimura et al., 1978 et 1981
TF
0,67 ± 0,06
tectites
Koenigswald, 1964 Orchiston & Siesser, 1982
K/Ar 0,71+0,02/-0,03
0,78 ± 0,15 zircon Suzuki et al., 1982 TF
0,79 ± 0,35
quartz, amphiboles
Saleki, 1997
ESR Ar/Ar
0,6
0,8-0,9
Kabuh
amphiboles Larick et al., 2001 40Ar/39Ar 1,66 ± 0,04 1,55 ± 0,08 1,02 ± 0,06
Grenzbank dents Saleki, 1997 ESR/U-Th >350 1,51± 0,25 1,49 ± 0,32 Suzuki et al., 1982 1,16 ± 0,24
Nishimura et al., 1978 et 1981 0,67± 0,04
couche tufacée (zircon)
Saleki, 1997
TF
1,56 ± 0,09
Pucangan
amphiboles Bettis et al., 2004 40Ar/39Ar 1,90 ± 0,02 Nishimura et al., 1981 K/Ar 2,0 ± 0,6 Swisher et al., 1994 40Ar/39Ar 1,66 ± 0,04
Saleki, 1997 40Ar/39Ar 2,05 ± 0,08_1,56 Brèche
volcanique amphiboles
Sémah et al., 2000 40Ar/39Ar 1,66 ± 0,04 Kalibeng tuf (zircon) Suzuki et al., 1982 TF 2,33 ± 0,47
Tableau 3 : Récapitulatif des données chronologiques sur le dôme de Sangiran.
(d’après Saleki, 1997 ; TF : traces de fission)

Premier chapitre : Présentation du cadre de l’étude.
- 48 -
Néanmoins, au niveau du paléomagnétisme, il existe des données plus fiables. Les lahars se situant entre le faciès marin et les argiles noires coïncident avec l’événement d’Olduvai. Ils peuvent être contemporains de cet événement ou s’être déposés juste après (Sémah F. 1986 ; Sémah F. et al., 2000).
Les enregistrements paléomagnétiques proches de la limite Brunhes/Matuyama entre la partie supérieure des argiles noires et des séries fluviatiles sont moins clairs. Différentes interprétations ont été données (Shimizu et al., 1985 ; Sémah F., 1986). Cette limite doit être placée juste avant ou juste après le début des dépôts de la série fluviatile (Sémah F. et al., 1998). La seconde de ces deux propositions est confirmée par les datations 40Ar/39Ar obtenues à Ngebung 2 (Saleki, 1997) au nord-ouest du dôme de Sangiran. La transition entre les formations de Pucangan et de Kabuh, le Grenzbank, pourrait être datée de la fin du Pléistocène inférieur entre l’événement de Jaramillo et la limite Brunhes/Matuyama (Sémah F. et al., 2001a).
La chronologie des hominidés fossiles javanais est difficile car elle repose sur un grand nombre de données radiométriques qui ne sont pas forcément en concordance les unes avec les autres. Ceci est le résultat d’un cadre géologique complexe et de l’absence d’informations solides sur la position stratigraphique de la plupart des fossiles et des échantillons datés (Theunissen et al., 1990 ; Langbroek & Roebroeks, 2000).
1.10.5. Découvertes
Le site de Sangiran est très riche du point de vue archéologique en restes d’hominidés et de faune.
1.10.5.1. Anthropologie
Environ une soixantaine d’individus d’Homo erectus ont été retrouvés dans cette zone
s’étendant sur une période de 1 million d’années. Ceci représente 65 % des hominidés fossiles d’Indonésie et 50 % de tous les Homo erectus découverts dans le monde (Simanjuntak, 2001). Ce site est donc d’un intérêt majeur, en particulier, pour la compréhension de la migration et de l’évolution des Homo erectus à Java et, en général, pour la compréhension de l’évolution humaine en général (annexe 2).
Deux des trois formes d’Homo erectus connues à Java sont présentes à Sangiran. Il est possible de distinguer trois formes d’Homo erectus à Java du point de vue
anatomique (Widianto, 2001): - Une forme archaïque, provenant de la formation de Pucangan de Sangiran (1,7 à 0,7
Ma), comme Sangiran 4 et 6,

Premier chapitre : Présentation du cadre de l’étude.
- 49 -
- Une forme classique, caractérisée par les fossiles de Trinil et de Sangiran, dans la formation de Kabuh (0,7 à 0,4 Ma),
- Une forme récente, comprenant les fossiles de Sambungmacan, Ngawi et Ngandong (après 0,3 Ma). En général, les découvertes anthropologiques de Sangiran sont très difficiles à replacer
dans un cadre stratigraphique précis. Ces fossiles ont été le plus souvent trouvés dans des champs par des paysans.
1.10.5.2. Paléontologie
Les restes fauniques sont les plus abondants et sont retrouvés principalement dans les couches du Pléistocène. Ces restes peuvent être des fossiles de mammifères, le plus souvent des herbivores, mais aussi de reptiles.
Des dents de crocodile, considérées comme les restes des plus anciens vertébrés continentaux de Sangiran, ont été retrouvées à la base de la formation de Pucangan (van Es, 1931). Dans les couches de Pucangan, le contenu fossilifère est moins riche que dans les couches de Kabuh car le milieu de sédimentation ne favorise pas une aussi bonne conservation des fossiles (Sémah A.M. et al.,1993 ; de Vos et al., 1982b ; Sondaar, 1984).
Dans le Grenzbank, mélangés à des éléments marins brisés et du matériel érodé des montagnes environnantes, de nombreux ossements de mammifères ont été découverts recouverts de concrétions caractéristiques. Dans les couches de Kabuh, le plus grand nombre de restes de mammifères fossiles a été retrouvé à Sangiran (Sémah F. et al., 2002).
1.10.5.3. Industrie lithique
En ce qui concerne les restes lithiques, ils ne se retrouvent que dans certaines localités
du dôme. Leur répartition est plus limitée, mais les recherches actuelles contribuent à en mettre au jour. Les premières découvertes d’industrie lithique ont été menées par G.H.R. von Koenigswald dès 1934 sur le site de Ngebung. Il a mis au jour des éclats de calcédoine et de jaspe. Von Koenigswald attribua ces éclats à la période du Pléistocène moyen. Cependant, l’ancienneté de ces éclats a été par la suite remise en cause (Movius, 1944 ; Bartstra, 1985).
Finalement, lors de la fouille de Ngebung 2, l’hypothèse de von Koenigswald a été confirmée et une riche industrie mise au jour dans un contexte stratigraphique précis (ensemble A). Cette industrie est composée de nombreux artéfacts: polyèdres et hâchereaux en andésite à grain fin, bolas, choppers, chopping tools et un galet faisant penser à un percuteur en quartz (Simanjuntak & Sémah, 1996).
Depuis 1993, le Centre National de Recherches Archéologiques de Yogyakarta a entrepris de nouvelles recherches sur l’industrie lithique dans cette zone (Widianto et al., 1996 ; Jatmiko, 2001). Les outils, et plus précisément, ceux de grande taille, sont rares. Les

Premier chapitre : Présentation du cadre de l’étude.
- 50 -
fouilles à Ngledok et Dayu (éclats) permettent de penser que les industries de Sangiran étaient plus anciennes que ce que G.H.R. von Koenigswald avait proposé (Widianto et al., 1996). L’Homo erectus a donc fabriqué des outils à Sangiran.
1.10.6. Présentation des sites étudiés Les collections des sites étudiés proviennent des fouilles menées dans les années 60, 70 et 80, sous la direction de Jacob, par le laboratoire de bio-paléoanthropologie de l’université de Gajah Mada où elles sont actuellement conservées. Les sites concernés se localisent principalement à l’est du dôme de Sangiran (figure 19 et annexe 1). Le choix de ces sites est souvent lié à une découverte paléoanthropologique faite préalablement par les villageois.
1.10.6.1. Tanjung
Tanjung est situé à environ 2 kilomètres au sud-est du centre du dôme à proximité de la rivière Cemoro (figure 19 et annexe 1). En juillet et août 1963, dans les environs du village de Tanjung à proximité d’un affluent de la rivière Cemoro sur le banc opposé à celui où fut découvert Sangiran 3 en 1937 (Jacob, 1964), des fragments crâniens appartenant à une même calotte crânienne de Pithécanthrope furent mis au jour. Ce crâne fut nommé Sangiran 10 ou Pithecanthropus VI (Sartono, 1964a).
Des campagnes de fouilles furent organisées en 1963 (août et septembre) et 1964 ainsi qu’en 1982 (du 16 au 30 juin 1982) (Koeshardjono, 1988).
Ce site fut choisi pour les raisons suivantes : - Il se trouvait proche du site de découverte du Pithécanthrope III (1937), - En juillet et août 1963, un os pariétal, un os zygomatique et d’autres fragments
crâniens y furent découverts par le même agriculteur, - Des fossiles de Bubalus sp. y furent mis au jour en juillet 1963, - Il se localise dans la partie supérieure fossilifère des couches de Kabuh (Jacob, 1964)
(tableau 4). Cette collection, environ 1109 restes, est la plus importante en nombre de restes :.
Les fouilles de 1963 (21 fossiles), 1964 (897 fossiles) et 1982 (191 fossiles) ne se situèrent pas au même niveau topographique, celle de 1982 est localisée au-dessus de celles de 1963 et 1964 (Koeshardjono, communication personnelle). L’industrie lithique associée au matériel faunique est composée d’une bola provenant des niveaux de 1982 et de rares éclats provenant des fouilles de 1963-64 qui furent perdus dans les années 60. Cependant, aucun outil n’a été découvert en association avec le crâne.
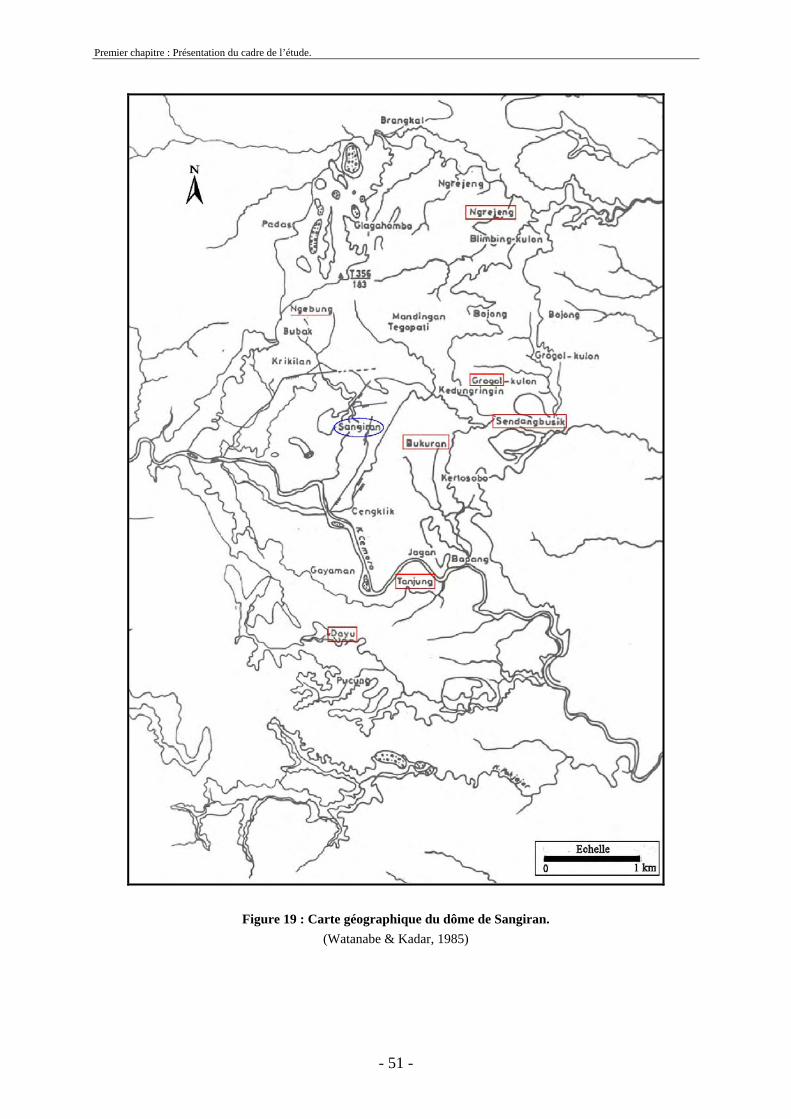
Premier chapitre : Présentation du cadre de l’étude.
- 51 -
Figure 19 : Carte géographique du dôme de Sangiran. (Watanabe & Kadar, 1985)

Premier chapitre : Présentation du cadre de l’étude.
- 52 -
1.10.6.2. Sendang Busik
Sendang Busik est situé à environ un kilomètre à l’est du centre du dôme de Sangiran (figure 19 et annexe 1). En avril 1979, un villageois découvrit deux fragments de crâne de Pithécanthrope sur ce site, ils furent nommés S37a et S37b (Baba & Aziz, 1991). Des fouilles furent entreprises à la suite de ces deux découvertes, en 1980 (Sendang Busik I), du 08 au 27 septembre 1980 et également en 1981 (Sendang Busik II), du 19 juin au 03 juillet 1981 (Koeshardjono, 1988). En 1982, un autre crâne de Pithécanthrope fut découvert dans cette même zone. Sendang Busik se situe, au niveau stratigraphique, dans les niveaux de Kabuh (tableau 4). Lors des fouilles de Sendang Busik I, 82 restes ont été mis au jour ainsi que 62 lors des ramassages de surface. Pour Sendang Busik II, 311 restes proviennent des fouilles et 15 de la surface. Aucune pièce lithique n’a été retrouvée en association avec le matériel faunique.
1.10.6.3. Ngrejeng Plupuh
Le site de Ngrejeng Plupuh est situé à environ 1,5 kilomètres au nord-est du centre du dôme (figure 19 et annexe 1). Sur ce site aucune découverte anthropologique n’a été faite. Une fouille fut entreprise sur le site du 07 au 26 avril 1985 (Koeshardjono, 1988). Les restes issus de cette fouille sont notés NKJ (Ngrejeng Kali Jambe, district de Karagancan) ou NP (Ngrejeng Plupuh district de Sragen), ces deux noms sont dus à un changement de district car le site était en limite des districts de Sragen et de Karagancan. Ngrejeng Plupuh est localisé dans les formations de Pucangan et de Kabuh (tableau 4). D’après des indications provenant des fiches accompagnant les ossements, 24 restes proviendraient de la formation de Pucangan (« Lempung »), ce qui est minoritaire dans l’assemblage. Lors de ces fouilles, 245 fossiles furent découverts et 203 fossiles en surface mais aucune pièce lithique ne fut trouvée.
1.10.6.4. Grogol Plupuh
Grogol Plupuh est localisé à environ 750 mètres à l’est du centre du dôme de Sangiran
(figure 19 et annexe 1). Sur ce site aucune découverte anthropologique n’a été faite. Des fouilles furent mises en place du 09 au 24 septembre 1983 à Grogol Plupuh (Koeshardjono, 1988). Grogol Plupuh se situe dans la formation de Kabuh (tableau 4). A partir des fouilles, 212 restes fauniques furent mis au jour, ainsi que 68 restes à partir de la surface mais aucune pièce lithique ne fut trouvée.
1.10.6.5. Bukuran
Bukuran se trouve à environ 500 mètres à l’est du centre du dôme (figure 19 et annexe 1). Sur ce site, von Koenigswald découvrit en 1936 une mandibule de Pithécanthrope

Premier chapitre : Présentation du cadre de l’étude.
- 53 -
nommée Sangiran 1b et en 1938-39 un maxillaire et une calotte dénommée Sangiran 4 (von Koenigswald, 1940).
Des fouilles furent conduites du 14 novembre au 08 décembre 1979 sur ce site (Koeshardjono, 1988). Et en 1980, une calotte crânienne de Pithécanthrope (Sangiran 38) fut découverte, de même, en 1989 une autre calotte crânienne (Hanoman 1) (Widianto & Grimaud-Hervé, 2000 ; Widianto et al., 1994) fut mise au jour, et en 1997, un crâne a été trouvé à proximité de ce site (Widianto, 2004 communication personnelle) ainsi qu’une incisive (Baba et al., 2000). C’est sur ce site également qu’un squelette d’Hexaprotodon sivalensis koenigswaldi a été découvert en 1989 (Widianto et al., 1994).
Bukuran se trouve dans les formations sédimentaires de Pucangan et de Kabuh (tableau 4). Le matériel fouillé provenant de ce site représente une collection moins importante en nombres de restes fauniques, 144 sont issus des fouilles néanmoins 346 restes sont issus des ramassages de surface.
1.10.6.6. Dayu Dayu se situe à environ un kilomètre au sud du centre du dôme de Sangiran (figure 19
et annexe 1). Du 17 au 31 octobre 1986 des fouilles furent menées à Dayu (Koeshardjono, 1988). Ce site est localisé dans la formation de Kabuh et plus précisément à sa base (Pasir conglomeratik). Cette collection est moins importante du point de vue du nombre de restes, seuls 33 fossiles proviennent des collections fouillées et 25 des ramassages de surface. Une pierre, peut-être un outil, fut découverte sur ce site.
En résumé, les restes fossiles des sites ne proviennent pas tous de la même formation
sédimentaire. La position stratigraphique des sites de Ngrejeng et Bukuran n’est pas très bien connue et donc peu précise (tableau 4). Néanmoins, nous pouvons penser que la majorité des restes issus des fouilles sur l’ensemble des sites proviennent de la formation de Kabuh.
KABUH Tanjung Sendang Busik
Grogol Plupuh
GRENZBANK
Ngrejeng Plupuh
Bukuran Dayu
PUCANGAN
Tableau 4 : Récapitulatif des positions stratigraphiques des sites étudiés.

Premier chapitre : Présentation du cadre de l’étude.
- 54 -
2. CADRE BIOSTRATIGRAPHIQUE DE CETTE ETUDE
Après avoir présenté la zone de l’étude, il nous a paru indispensable de faire le point sur les données paléontologiques et biostratigraphiques de l’île de Java au cours du Quaternaire avant de d’exposer nos propres résultats dans ce domaine.
Comme nous avons pu le voir précédemment la colonisation de l’île de Java par les
mammifères s’est faite au cours de plusieurs migrations lors des abaissements périodiques du niveau marin pendant le Quaternaire. Il était donc très important d’établir une chronologie des migrations à partir de l’apparition, de l’évolution et de la disparition des espèces à Java.
Les fossiles de mammifères sont connus à Java depuis longtemps mais aucune description scientifique n’a été publiée avant 1887. À cette date, Martin, un paléontologue néerlandais, décrit des restes de Stegodon envoyés au Naturalis Museum de Leiden. Il propose une relation avec les Siwaliks indiennes, laquelle sera confirmée par les recherches ultérieures (Colbert, 1943 ; von Koenigswald, 1934a ; Hooijer, 1952 ; Badoux, 1959 et de Vos, 1993). En 1932, van der Maarel suite à son étude de la faune de Bumiayu, considère cette dernière comme antérieure à la faune de Trinil et il date la faune de Bumiayu du Pliocène supérieur ou du Pléistocène inférieur. Cette proposition de cadre biostratigraphique pour les faunes de Java sera le début d’une longue série d’hypothèses dont nous présentons ici les arguments.
2.1. Récapitulatif des listes fauniques de Java au Pléistocène
Grâce à ces études sur le cadre biostratigraphique de Java nous avons pu effectuer un récapitulatif des listes fauniques établies par les différents auteurs, dans un premier temps, pour les localités datées du Pléistocène inférieur et moyen (tableau 5). Nous avons ajouté les données sur la faune de la grotte de Song Terus étudiée par A.M. Moigne et R. Due Awe (M.Q.P.I.).
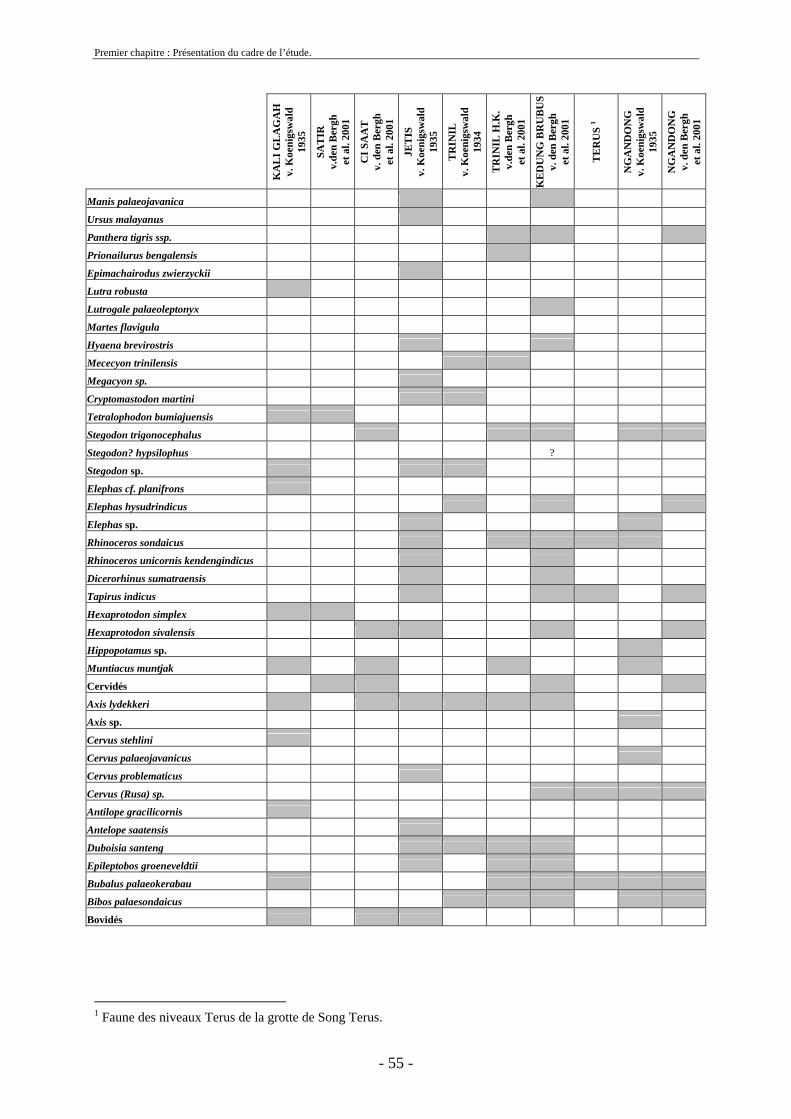
Premier chapitre : Présentation du cadre de l’étude.
- 55 -
KA
LI G
LA
GA
H
v. K
oeni
gsw
ald
19
35
SAT
IR
v.de
n B
ergh
et
al.
2001
CI S
AA
T
v. d
en B
ergh
et
al.
2001
JET
IS
v. K
oeni
gsw
ald
1935
TR
INIL
v.
Koe
nigs
wal
d
1934
TR
INIL
H.K
.
v.
den
Ber
gh
et a
l. 20
01
KE
DU
NG
BR
UB
US
v. d
en B
ergh
et a
l. 20
01
TE
RU
S 1
NG
AN
DO
NG
v. K
oeni
gsw
ald
1935
NG
AN
DO
NG
v. d
en B
ergh
et a
l. 20
01
Manis palaeojavanica
Ursus malayanus
Panthera tigris ssp.
Prionailurus bengalensis
Epimachairodus zwierzyckii
Lutra robusta
Lutrogale palaeoleptonyx
Martes flavigula
Hyaena brevirostris
Mececyon trinilensis
Megacyon sp.
Cryptomastodon martini
Tetralophodon bumiajuensis
Stegodon trigonocephalus
Stegodon? hypsilophus ?
Stegodon sp.
Elephas cf. planifrons
Elephas hysudrindicus
Elephas sp.
Rhinoceros sondaicus
Rhinoceros unicornis kendengindicus
Dicerorhinus sumatraensis
Tapirus indicus
Hexaprotodon simplex
Hexaprotodon sivalensis
Hippopotamus sp.
Muntiacus muntjak
Cervidés
Axis lydekkeri
Axis sp.
Cervus stehlini
Cervus palaeojavanicus
Cervus problematicus
Cervus (Rusa) sp.
Antilope gracilicornis
Antelope saatensis
Duboisia santeng
Epileptobos groeneveldtii
Bubalus palaeokerabau
Bibos palaesondaicus
Bovidés
1 Faune des niveaux Terus de la grotte de Song Terus.
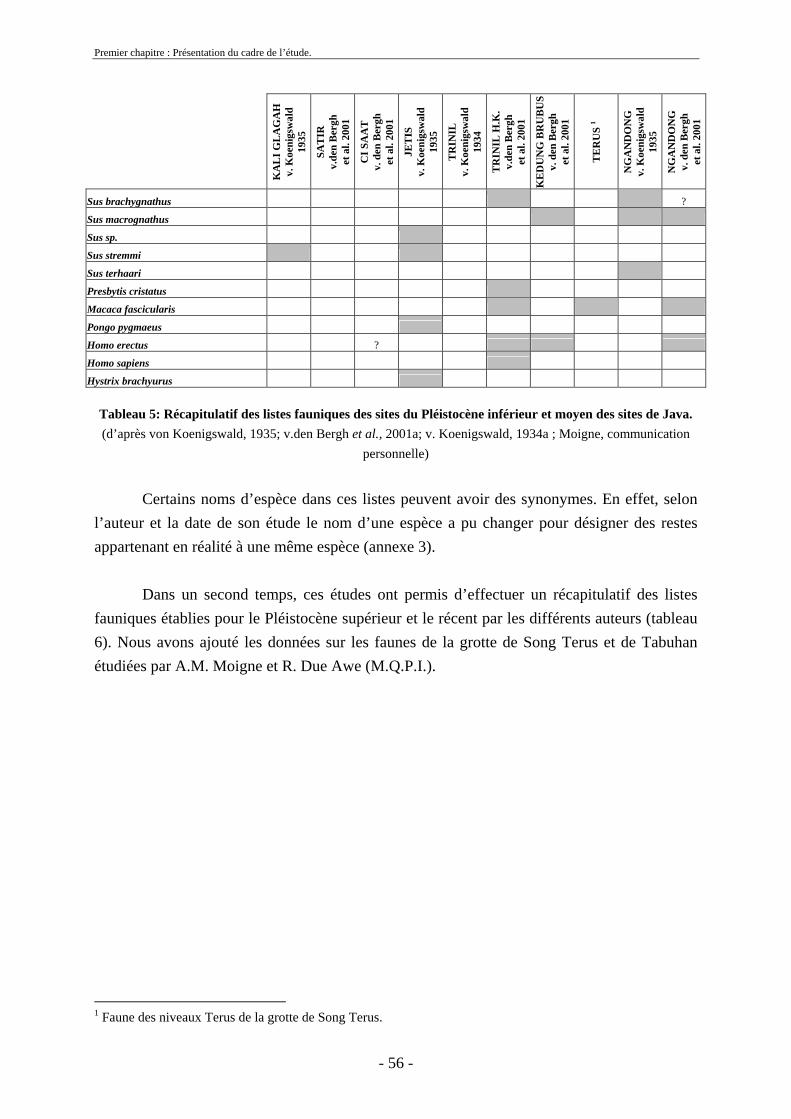
Premier chapitre : Présentation du cadre de l’étude.
- 56 -
KA
LI G
LA
GA
H
v. K
oeni
gsw
ald
19
35
SAT
IR
v.de
n B
ergh
et
al.
2001
CI S
AA
T
v. d
en B
ergh
et
al.
2001
JET
IS
v. K
oeni
gsw
ald
1935
TR
INIL
v.
Koe
nigs
wal
d
1934
TR
INIL
H.K
.
v.
den
Ber
gh
et a
l. 20
01
KE
DU
NG
BR
UB
US
v. d
en B
ergh
et a
l. 20
01
TE
RU
S 1
NG
AN
DO
NG
v. K
oeni
gsw
ald
1935
NG
AN
DO
NG
v. d
en B
ergh
et a
l. 20
01
Sus brachygnathus ?
Sus macrognathus
Sus sp.
Sus stremmi
Sus terhaari
Presbytis cristatus
Macaca fascicularis
Pongo pygmaeus
Homo erectus ?
Homo sapiens
Hystrix brachyurus
Tableau 5: Récapitulatif des listes fauniques des sites du Pléistocène inférieur et moyen des sites de Java. (d’après von Koenigswald, 1935; v.den Bergh et al., 2001a; v. Koenigswald, 1934a ; Moigne, communication
personnelle)
Certains noms d’espèce dans ces listes peuvent avoir des synonymes. En effet, selon
l’auteur et la date de son étude le nom d’une espèce a pu changer pour désigner des restes appartenant en réalité à une même espèce (annexe 3).
Dans un second temps, ces études ont permis d’effectuer un récapitulatif des listes fauniques établies pour le Pléistocène supérieur et le récent par les différents auteurs (tableau 6). Nous avons ajouté les données sur les faunes de la grotte de Song Terus et de Tabuhan étudiées par A.M. Moigne et R. Due Awe (M.Q.P.I.).
1 Faune des niveaux Terus de la grotte de Song Terus.
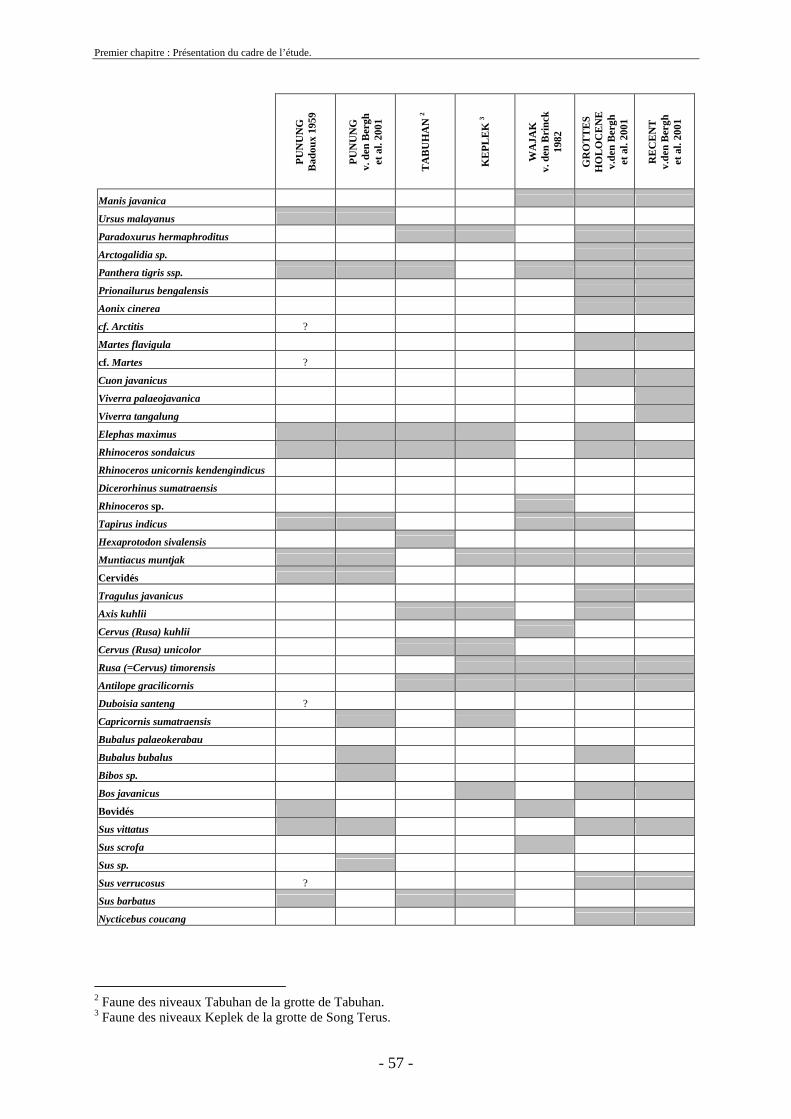
Premier chapitre : Présentation du cadre de l’étude.
- 57 -
PUN
UN
G
B
adou
x 19
59
PUN
UN
G
v.
den
Ber
gh
et
al.
2001
TA
BU
HA
N 2
KE
PLE
K 3
WA
JAK
v.
den
Bri
nck
1982
GR
OT
TE
S H
OL
OC
EN
E
v.
den
Ber
gh
et
al.
2001
RE
CE
NT
v.
den
Ber
gh
et a
l. 20
01
Manis javanica
Ursus malayanus
Paradoxurus hermaphroditus
Arctogalidia sp.
Panthera tigris ssp.
Prionailurus bengalensis
Aonix cinerea
cf. Arctitis ?
Martes flavigula
cf. Martes ?
Cuon javanicus
Viverra palaeojavanica
Viverra tangalung
Elephas maximus
Rhinoceros sondaicus
Rhinoceros unicornis kendengindicus
Dicerorhinus sumatraensis
Rhinoceros sp.
Tapirus indicus
Hexaprotodon sivalensis
Muntiacus muntjak
Cervidés
Tragulus javanicus
Axis kuhlii
Cervus (Rusa) kuhlii
Cervus (Rusa) unicolor
Rusa (=Cervus) timorensis
Antilope gracilicornis
Duboisia santeng ?
Capricornis sumatraensis
Bubalus palaeokerabau
Bubalus bubalus
Bibos sp.
Bos javanicus
Bovidés
Sus vittatus
Sus scrofa
Sus sp.
Sus verrucosus ?
Sus barbatus
Nycticebus coucang
2 Faune des niveaux Tabuhan de la grotte de Tabuhan. 3 Faune des niveaux Keplek de la grotte de Song Terus.
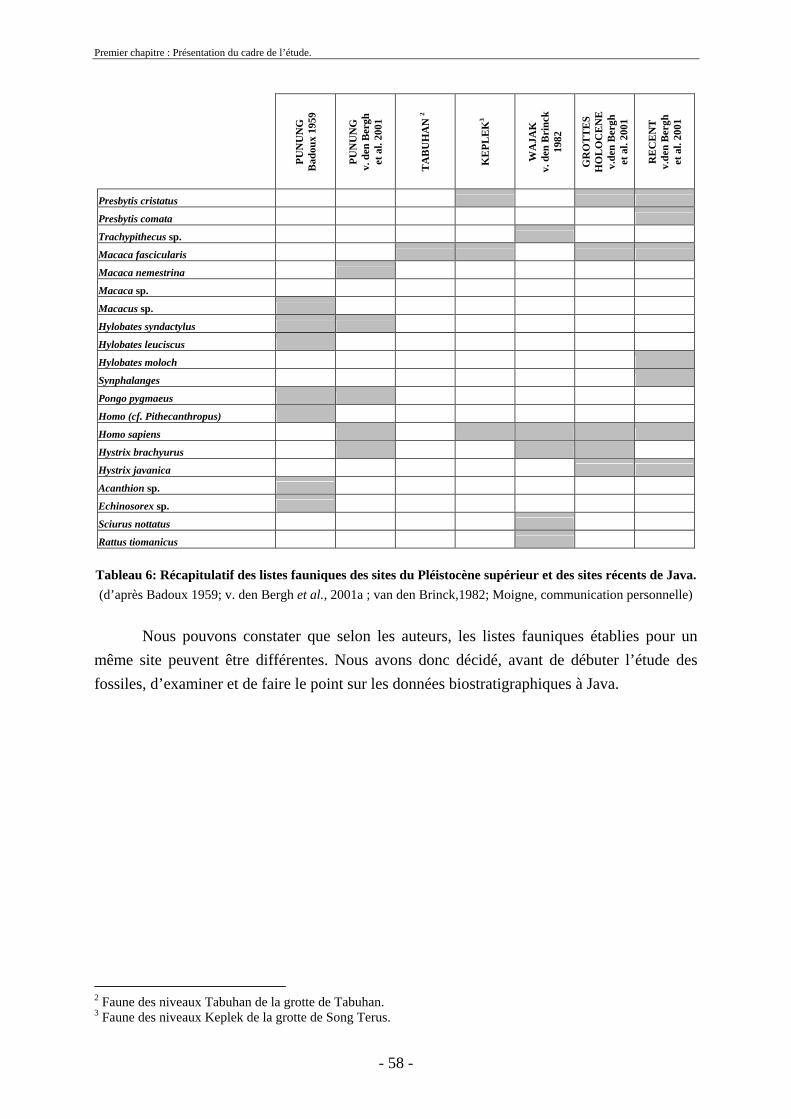
Premier chapitre : Présentation du cadre de l’étude.
- 58 -
PUN
UN
G
B
adou
x 19
59
PUN
UN
G
v.
den
Ber
gh
et
al.
2001
TA
BU
HA
N 2
KE
PLE
K3
WA
JAK
v.
den
Bri
nck
1982
GR
OT
TE
S H
OL
OC
EN
E
v.
den
Ber
gh
et
al.
2001
RE
CE
NT
v.
den
Ber
gh
et a
l. 20
01
Presbytis cristatus
Presbytis comata
Trachypithecus sp.
Macaca fascicularis
Macaca nemestrina
Macaca sp.
Macacus sp.
Hylobates syndactylus
Hylobates leuciscus
Hylobates moloch
Synphalanges
Pongo pygmaeus
Homo (cf. Pithecanthropus)
Homo sapiens
Hystrix brachyurus
Hystrix javanica
Acanthion sp.
Echinosorex sp.
Sciurus nottatus
Rattus tiomanicus
Tableau 6: Récapitulatif des listes fauniques des sites du Pléistocène supérieur et des sites récents de Java. (d’après Badoux 1959; v. den Bergh et al., 2001a ; van den Brinck,1982; Moigne, communication personnelle)
Nous pouvons constater que selon les auteurs, les listes fauniques établies pour un même site peuvent être différentes. Nous avons donc décidé, avant de débuter l’étude des fossiles, d’examiner et de faire le point sur les données biostratigraphiques à Java.
2 Faune des niveaux Tabuhan de la grotte de Tabuhan. 3 Faune des niveaux Keplek de la grotte de Song Terus.

Premier chapitre : Présentation du cadre de l’étude.
- 59 -
2.2. Biostratigraphie établie par von Koenigswald Von Koenigswald (1934a) a été le premier à proposer un cadre biostratigraphique pour
l’île de Java à partir des associations fauniques provenant de six localités différentes. Il a proposé l’âge de celles-ci en les comparant avec les biostratigraphies établies en Inde et en Chine (tableau 7).
Inde Java Chine Holocène / Sampung /
Grottes de Carnul Ngandong Grotte supérieure
(Choukoutien)
Narmada Boulder conglomerate
Couches de Notopuro (sans faune)
Trinil Choukoutien
Pléistocène supérieur Pléistocène moyen Pléistocène inférieur Pinjor Jetis Nihowan
Kali Glagah Pliocène supérieur Pliocène moyen
Tatrot Ci Julang
/
Tableau 7: Corrélation des faunes de mammifères en Asie.
(von Koenigswald, 1949)
La plus ancienne association faunique enregistrée est Ci Julang (tableau 7) près de Banjar, à la limite entre Java Ouest et Java Central (figure 20).
Figure 20 : Carte des sites paléontologiques de Java.
Pour von Koenigswald (1934a, 1949, 1956), cette faune contenant Elephas
(Archidiskodon) planifrons, Merycopotamus et Hippopotamus est clairement d’origine

Premier chapitre : Présentation du cadre de l’étude.
- 60 -
indienne, et peut être corrélée avec la faune de la zone du Tatrot des Siwaliks datée du Pliocène. L’arrivée de ces genres à Java marque une invasion Indo-Malaise au Pliocène moyen. D’un point de vue paléozoologique deux faits peuvent être mentionnés ici : la complète absence du cheval, qui est présent en Inde mais pas à Java ni en Chine du Sud, et l’abondance d’Hippopotamus à Java et en Inde mais pas en Chine.
La faune de Kali Glagah (tableau 7) n’est pas si bien définie, elle a été relevée dans un
seul site proche de Tegal à l’ouest de Java près de Bumiayu (figure 20). Le fossile principal est le trilophonte Mastodon bumiajuensis* (van der Maarel, 1932) qui a des défenses seulement sur le maxillaire. En outre, nous avons du Stegodon, de l’Elephas cf. planifrons, du Sus stremmi, du Muntiacus, du Cervus stehlini, du Bos et l’hexaprotodonte Hippopotamus simplex dont un squelette complet est conservé dans la collection de Bandung. Parmi les reptiles, des crocodiles et une tortue géante sont présents, cette dernière est identique ou très proche de Colossochelys atlas provenant des Siwaliks.
Vient ensuite l’association de la faune de Jetis (tableau 7 et figure 20) qui a une large distribution. Cet horizon est connu sur toute la limite nord de la dépression centrale avec les Kendeng, entre Solo et Mojokerto (figure 11). Cette faune est très riche. Les carnivores trouvés sont Felis, Epimachairodus, Lutra, Megacyon, Ursus, Hyaena* et beaucoup d’autres petites formes. Les artiodactyles sont communs : Hippopotamus (Hippopotamus koenigswaldi*), des suidés, des cerfs (le plus typique Cervus zwaani∗ et le grand Cervus problematicus), des antilopes (Antelope modjokertensis* et Antelope saatensis), du Bos et le primitif Leptobos, le Rhinoceros sondaicus*, le Rhinoceros kendengindicus. Tapirus* et Nestoritherium représentent les périssodactyles. De plus, le Stegodon, l’Elephas, le rare Cryptomastodon, un fourmilier géant (Manis palaeojavanica), des porcs-épics, des lagomorphes, des petits singes, des gibbons et un grand orang (Pongo) sont présents associés à des restes d’Homo erectus (von Koenigswald, 1933).
D’après von Koenigswald (1934a, 1949 et 1956), la faune de Jetis peut être corrélée avec la faune du Pinjor en Inde datée du Pléistocène inférieur (Colbert, 1943). Cependant la présence de Tapirus, de l’ours malais, des gibbons et de l’orang, qui n’ont jamais été trouvés en Inde mais sont typiques des dépôts de fissures de la Chine du Sud, montre que cette faune est marquée par des influences chinoises.
La faune de Trinil (tableau 7) provenant de Java est (figure 20), n’est pas très
différente de celle de Jetis mais Epimachairodus, Nestoritherium, Leptobos∗ et d’autres types plus anciens ont disparu. Les fossiles principaux sont un petit cerf Axis, Cervus (Axis) lydekkeri* et une petite antilope Duboisia kroesenii*. Le buffle, le banteng (Bibos banteng*), ∗ liste des synonymes, annexe 3.

Premier chapitre : Présentation du cadre de l’étude.
- 61 -
le Stegodon et un éléphant Elephas cf. namadicus ont été trouvés également. Le fameux Pithecanthropus erectus de Dubois provient de cet horizon.
La faune de Trinil peut être corrélée avec la faune du « Boulder Conglomerate » et avec les couches inférieures de la Narmada en Inde datées du Pléistocène moyen (von Koenigswald, 1934a, 1949, 1956).
La faune de Ngandong (tableau 7) est la plus moderne, connue seulement à partir
d’anciennes terrasses au Nord et à l’Ouest de Ngawi (figure 20). Les antilopes ont complètement disparu. Hippopotamus, Stegodon et Elephas sont hautement spécialisés. Le banteng moderne et Cervus hippelaphus∗ sont abondants. Les fossiles principaux sont un cerf Axis, Cervus palaeojavanicus et un grand suidé, Sus terhaari. Les restes d’homme fossile dans ces couches ont été attribués à la forme récente d’Homo erectus.
Pour von Koenigswald (1934a, 1949, 1956), les couches de Trinil occupent une
position centrale dans la séquence stratigraphique. L’âge peut être déterminé par la spécialisation des éléphants. Les éléphants de ces couches sont très proches ou identiques à Elephas namadicus d’Inde. Celui-ci en Inde n’apparaît qu’à partir de la formation du « Boulder Conglomerate » et des couches inférieures de la Narmada. En conséquence, les couches de Trinil ne peuvent pas représenter le Pléistocène inférieur. La faune des couches de Jetis, qui est un mélange de survivants du tertiaire avec des types plus avancés, serait une association typique du Pléistocène inférieur, la faune de Trinil une association du Pléistocène moyen et celle plus moderne de Ngandong du Pléistocène supérieur (tableau 7) (von Koenigswald, 1934a). Von Koenigswald (1935, 1940) a également tenté de corréler les associations fauniques qu’il avait définies avec les faunes retrouvées dans les horizons stratigraphiques du dôme de Sangiran lors de ses fouilles à Ngebung. Il a considéré que la faune de Ci Julang se retrouvait dans la formation de Kalibeng à Sangiran car les deux sites présentaient des restes d’un mastodon, la faune de Jetis dans la formation de Pucangan et la faune de Trinil dans la formation de Kabuh. Ce cadre biostratigraphique établi pour l’île de Java par von Koenigswald a été utilisé jusqu’en 1982.
∗ liste des synonymes, annexe 3.

Premier chapitre : Présentation du cadre de l’étude.
- 62 -
2.3. Biostratigraphie établie par de Vos et Sondaar Au début des années 80, de Vos et Sondaar (1982) décident de réviser la biostratigraphie établie par von Koenigswald (1934a). Pour ce faire, ils reprennent l’étude des faunes de mammifères du Pléistocène des collections de Dubois. Ces auteurs considéraient que von Koenigswald avait utilisé des fossiles provenant de plusieurs localités voisines ce qui lui donnait des assemblages fauniques hétérogènes et que les fossiles marqueurs utilisés pour définir ces unités biostratigraphiques représentaient des découvertes isolées d’espèces très rares qu’il était difficile de retrouver dans d’autres associations fauniques. Leur étude a tout d’abord porté sur les sites de Trinil et de Kedung Brubus. Ils n’ont considéré que les fossiles dont l’origine était certaine, c’est à dire provenant du site même de Trinil (Trinil H.K.) ou de Kedung Brubus et non de leurs environs. Trinil H.K. signifie : « Trinil Haupt-Knochen Schicht » c’est à dire qui est exclusivement basé sur les fossiles provenant de la principale couche fossilifère près de Trinil. Trinil H.K. est différente de la faune de Trinil considérée par von Koenigswald (1935). Ces résultats ont été ensuite comparés avec ceux de l’expédition Selenka à Trinil (Selenka & Blankenhorn, 1911), qui avaient permis de préciser le contexte stratigraphique de ce site.
De Vos et Sondaar (1982), ont donc pu mettre en évidence que la composition des faunes de Trinil H.K. et de Kedung Brubus était distincte et que ces faunes devaient être chronologiquement différentes. Cette différence n’était pas due au paléoenvironnement car Trinil H.K. et Ngandjar (localité de la faune de Kedung Brubus) sont proches géographiquement, mais elle était, plutôt, liée à l’apparition d’espèces plus « évoluées » dans la faune de Kedung Brubus telles qu’Epileptobos groeneveldtii∗ ou Elephas hysudrindicus*. D’autre part, les grandes ressemblances des faunes de Jetis et de Kedung Brubus montraient probablement le même âge, plus jeune que celui de Trinil H.K. (tableau 8), alors que pour von Koenigswald en 1935, Trinil et Kedung Brubus étaient du même âge et Jetis était d’un âge plus ancien (tableau 7).
∗ liste des synonymes, annexe 3.

Premier chapitre : Présentation du cadre de l’étude.
- 63 -
Von Koenigswald
(1934)
De Vos et al. (1982)
Listes fauniques simplifiées
Ngandong Ngandong Formes plus avancées, absence de Duboisia
Trinil Kedung
Brubus Nouvelles arrivées de Hyaena, Tapirus, Epileptobos et Elephas
Jetis Trinil H.K. Pauvre en espèces, principalement Axis et Duboisia, grands bovidés
Tableau 8: Comparaison des interprétations biostratigraphiques de l’île de Java en 1982.
(d’après von Koenigswald, 1934a et de Vos et al., 1982b)
En outre, Trinil H.K. est plus pauvre en espèces que Kedung Brubus, ce qui traduit
une position plus isolée de l’île de Java au moment du dépôt de la faune de Trinil H.K. En ce qui concerne la faune de Kedung Brubus, l’arrivée d’espèces différentes ou le renouvellement du stock d’espèces montrent un contact, à cette période, entre Java et le continent asiatique.
Par la suite, en 1983, de Vos a poursuivi la révision de la biostratigraphie de Java et
plus précisément de celle de la localité de Punung (Java Est) proche de la ville de Pacitan (figure 11).
La faune de Punung a été découverte par von Koenigswald entre 1934 et 1937 et a été étudiée par Badoux (1959). Cette collection de Punung est composée de restes provenant des localités de Punung I, redécouvertes récemment (de Vos et al., communication personnelle), à proximité de Mendolo Kidul, et de Punung II, à proximité de Tabuhan, mais de Vos a considéré la collection dans son ensemble. Il compare cette faune avec celles des grottes de Sumatra, Lida Ajer et Sibrambang, récoltées par Dubois entre 1887 et 1890. Pour cet auteur, il existe une ressemblance entre ces différents sites car leur taphonomie est proche. Les porcs-épics semblent avoir rongé la majorité des ossements ce qui a pour conséquence la prédominance des restes dentaires dans les assemblages. De plus, les associations fauniques sont très proches au niveau des espèces, le nombre d’éléments de chaque espèce et le paléoenvironnement sont comparables.
De Vos compare, ensuite, la faune de Punung à celle de Trinil, et conclut que cette dernière est antérieure à celle de Punung en accord avec Badoux. Dans un deuxième temps, il compare la faune de Punung à celle de Ngandong et propose l’antériorité de cette dernière par rapport à la faune de Punung en comparant la présence et le degré d’évolution des espèces. En effet, à Ngandong, les espèces archaïques telles qu’Hippopotamus, Cervus (Axis) lydekkeri, Stegodon trigonocephalus∗, Elephas hysudrindicus*, sont présentes alors qu’elles n’apparaissent pas dans la liste faunique de Punung. De plus, Pongo pygmaeus est très
∗ liste des synonymes, annexe 3.

Premier chapitre : Présentation du cadre de l’étude.
- 64 -
abondant à Punung et absent à Ngandong. Enfin, il compare la faune de Punung à celle de Wajak. Dans la faune de Wajak (van den Brink, 1982), Pongo sp., Elephas maximus, Ursus malayanus, Macaca et Hylobates sont absents ainsi que dans la faune actuelle de Java. Il en résulte que pour de Vos, la faune de Punung est plus ancienne que celle de Wajak et présente un environnement différent. La faune de Punung caractérise un milieu de forêt humide alors que la faune de Wajak et celle de Ngandong indiquent un milieu de forêt ouverte.
L’unité biostratigraphique de Punung se situe alors entre les unités de Ngandong et de Wajak (tableau 9).
Sumatra Java Environnement
Actuel Wajak Forêt ouverte
Grottes de Sumatra Punung Forêt humide Ngandong Forêt ouverte Kedung Brubus Forêt ouverte Trinil Forêt ouverte
Tableau 9: Récapitulatif des successions fauniques de l’île de Java et de leur environnement.
(de Vos, 1983)
Dernièrement, un nouveau site a été découvert à Punung : Gunung Dawu (Storm et
al., sous presse). Les espèces retrouvées sur ce site sont caractéristiques d’une forêt tropicale et confirment les résultats de de Vos.
Enfin, Sondaar, l’année suivante (1984), présente une récapitulation sur l’évolution de
la faune et la biostratigraphie des mammifères de l’île de Java. Tout d’abord, il met en relief les différentes unités biostratigraphiques (tableau 10).
Dans ce récapitulatif, la faune de la localité de Ci Julang n’est pas considérée comme une unité biostratigraphique utilisable, comme le pensait von Koenigswald (1934a, 1949) (tableau 7), car les restes fauniques ne proviennent pas d’un cadre stratigraphique précis.
La localité au nord-ouest de Bumiayu présente un mélange de faunes de différents niveaux stratigraphiques, récolté sur les rives de trois rivières : Kali Glagah, Kali Biuk et Ci Saat. Pour Sondaar les dépôts de Kali Glagah contiennent deux faunes différentes, Satir et Ci Saat, car il existe une très grande différence de niveaux stratigraphiques entre les fouilles dans la partie basse de Kali Glagah et celles du nord-ouest de Ci Saat. Satir possède peu d’espèces et celles-ci sont moins évoluées que celles de Ci Saat. En comparant la liste faunique de Ci Saat avec celle de Trinil cet auteur met en évidence des différences entre ces deux faunes et donc les individualise comme deux unités biostratigraphiques à part entière alors que von Koenigswald (1934a, 1949) n’en considérait qu’une seule : Kali Glagah.
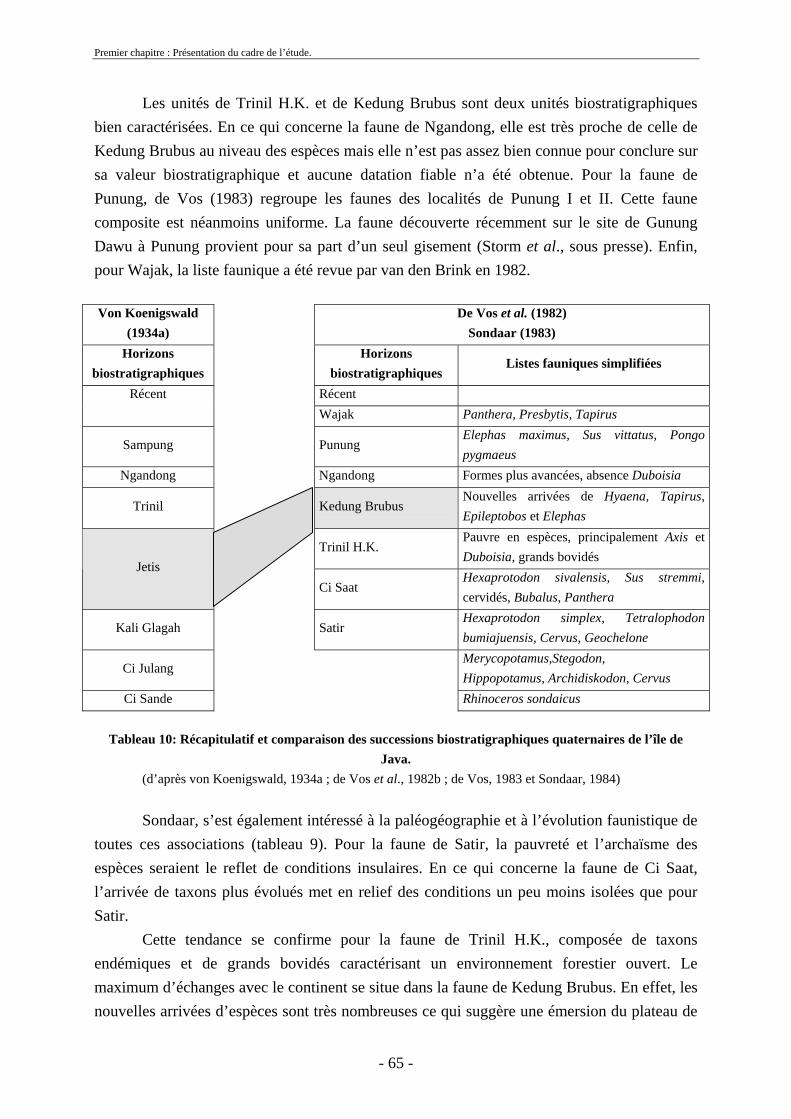
Premier chapitre : Présentation du cadre de l’étude.
- 65 -
Les unités de Trinil H.K. et de Kedung Brubus sont deux unités biostratigraphiques bien caractérisées. En ce qui concerne la faune de Ngandong, elle est très proche de celle de Kedung Brubus au niveau des espèces mais elle n’est pas assez bien connue pour conclure sur sa valeur biostratigraphique et aucune datation fiable n’a été obtenue. Pour la faune de Punung, de Vos (1983) regroupe les faunes des localités de Punung I et II. Cette faune composite est néanmoins uniforme. La faune découverte récemment sur le site de Gunung Dawu à Punung provient pour sa part d’un seul gisement (Storm et al., sous presse). Enfin, pour Wajak, la liste faunique a été revue par van den Brink en 1982.
Von Koenigswald
(1934a)
De Vos et al. (1982) Sondaar (1983)
Horizons biostratigraphiques
Horizons
biostratigraphiques Listes fauniques simplifiées
Récent Récent Wajak Panthera, Presbytis, Tapirus
Sampung Punung Elephas maximus, Sus vittatus, Pongo pygmaeus
Ngandong Ngandong Formes plus avancées, absence Duboisia
Trinil
Kedung Brubus Nouvelles arrivées de Hyaena, Tapirus, Epileptobos et Elephas
Trinil H.K. Pauvre en espèces, principalement Axis et Duboisia, grands bovidés
Jetis Ci Saat
Hexaprotodon sivalensis, Sus stremmi, cervidés, Bubalus, Panthera
Kali Glagah Satir Hexaprotodon simplex, Tetralophodon bumiajuensis, Cervus, Geochelone
Ci Julang Merycopotamus,Stegodon, Hippopotamus, Archidiskodon, Cervus
Ci Sande
Rhinoceros sondaicus
Tableau 10: Récapitulatif et comparaison des successions biostratigraphiques quaternaires de l’île de Java.
(d’après von Koenigswald, 1934a ; de Vos et al., 1982b ; de Vos, 1983 et Sondaar, 1984)
Sondaar, s’est également intéressé à la paléogéographie et à l’évolution faunistique de toutes ces associations (tableau 9). Pour la faune de Satir, la pauvreté et l’archaïsme des espèces seraient le reflet de conditions insulaires. En ce qui concerne la faune de Ci Saat, l’arrivée de taxons plus évolués met en relief des conditions un peu moins isolées que pour Satir.
Cette tendance se confirme pour la faune de Trinil H.K., composée de taxons endémiques et de grands bovidés caractérisant un environnement forestier ouvert. Le maximum d’échanges avec le continent se situe dans la faune de Kedung Brubus. En effet, les nouvelles arrivées d’espèces sont très nombreuses ce qui suggère une émersion du plateau de

Premier chapitre : Présentation du cadre de l’étude.
- 66 -
la Sonde à cette époque. La faune de Ngandong étant peu connue, elle est rapprochée, au niveau du paléoenvironnement, de celle de Kedung Brubus. La faune de Punung voit l’arrivée d’Elephas maximus et de Pongo pygmaeus, qui montrent encore un lien avec le continent, et donc une période glaciaire. Toutefois ce lien est moins important que dans la faune de Kedung Brubus. En outre, le paléoenvironnement de la faune de Punung est différent de celui des faunes précédentes, les espèces sont caractéristiques d’un milieu de forêt humide. Ceci est confirmé par les espèces découvertes sur le nouveau site de Gunung Dawu (Storm et al., sous presse). Enfin, la faune de Wajak est typique d’un milieu forestier ouvert pendant une période plus froide que l’actuel. Finalement, Sondaar, tente d’effectuer des corrélations biostratigraphiques entre différentes régions javanaises. La faune de Jetis (Java Est) serait alors la même que celle de Kedung Brubus. De la même façon, à Sangiran (Java central), il propose de replacer les unités biostratigraphiques dans les unités stratigraphiques existantes (Pucangan, et Kabuh).
Par la suite, Leinders et al. (1985) exposent de manière plus détaillée la biozonation standard du dôme de Sangiran. Les corrélations lithostratigraphiques détaillées et les fouilles systématiques effectuées par le programme de coopération et de recherche nippo-indonésien CTA-41 ont permis d’obtenir de nouvelles données stratigraphiques. À partir de ces données, les hypothèses suivantes ont pu être proposées (tableau 11) :
- La faune de la partie moyenne des argiles noires de la formation de Pucangan, principalement des cervidés et des hippopotamidés (Aimi & Aziz, 1985), est interprétée par Sondaar (1984) comme une faune insulaire et non stable très proche de la faune de Satir.
- Dans la partie supérieure des argiles noires de la formation de Pucangan, Bubalus, Bibos et Stegodon ont été retrouvés. Ces espèces rappellent celles de la faune de Ci Saat.
- À partir du conglomérat calcifié se situant à la base de la formation de Kabuh, le Grenzbank, Aimi et Aziz (1985) ont découvert une faune diversifiée avec des carnivores, des rhinocérotidés, des suidés et une antilope endémique Duboisia santeng. Cette faune peut-être corrélée à la faune de Trinil H.K.
- La première apparition du genre Elephas a été observée dans les couches de Kabuh au-dessus du Grenzbank. Cette présence permet de rapprocher la faune des couches de Kabuh de celle de la localité de Kedung Brubus.

Premier chapitre : Présentation du cadre de l’étude.
- 67 -
Biozones Dates Stratigraphie de Sangiran Faune de Kedung
Brubus 0,8 Ma Partie supérieure de la formation de Kabuh.
Faune de Trinil H.K.
1 Ma Dans la partie inférieure de la formation de Kabuh incluant le Grenzbank.
Faune de Ci Saat 1,2 Ma Partie supérieure des argiles noires de la formation de Pucangan sous le Grenzbank.
Faune de Satir 1,5 Ma Partie moyenne des argiles noires de la formation de Pucangan sous le tuff 9.
Tableau 11: Corrélations biostratigraphiques du dôme de Sangiran
(d’après Leinders et al., 1985)
Leinders et al. (1985) proposent une chronologie des faunes de Satir, Ci Saat, Trinil
H.K. et Kedung Brubus en se basant sur les dates des formations de Sangiran (tableau 11). Ces dates ont été obtenues par différentes méthodes : la méthode des traces de fission sur les zircons dans les tuffs (Suzuki et al., 1985), la méthode des traces de fission sur les tectites et la corrélation avec des couches de tectites découvertes dans des carottes sous-marines à la limite Brunhes-Matuyama (Ninkovitch & Burckle, 1978) et enfin avec la magnétostratigraphie (Sémah F., 1984). Tout ceci leur permet de conclure que la faune de Satir serait datée d’environ 1,5 million d’années, la faune de Ci Saat de 1,2 million d’années, la faune de Trinil H.K. de 1 million d’années et la faune de Kedung Brubus de 0,8 million d’années (tableau 11).
D’autre part, Matsu’ura (1982) a démontré que la composition en fluor varie dans des unités lithologiques de nature différente à Sangiran. À partir de ce concept, il a conclu que tous les fossiles d’hominidés de la zone de Sangiran provenaient du Grenzbank et des couches de Kabuh.
Par la suite, de Vos (1985) a intégré les découvertes anthropologiques, dont celles de
Sangiran, dans les différentes successions fauniques établies par Sondaar (1984) (tableau 12). Pour la localité de Trinil, neuf fossiles d’hominidés ont été découverts qui seraient
attribués à la forme classique et à la forme archaïque d’Homo erectus. À Kedung Brubus, deux fossiles d’hominidés ont été retrouvés. La mandibule serait attribuée à une forme d’Homo sp. indet. (Tobias, 1966) et la détermination du fémur est plus problématique. Après avoir été déterminé comme un fémur de Pithecanthropus (Dubois, 1937), il fut déterminé par les paléontologues comme un fémur de grand carnivore. En conclusion, de Vos (1985) considère la mandibule comme appartenant à un hominidé d’une forme plus progressive que celle de Trinil. Pour la localité de Sangiran, de nombreux restes d’hominidés ont été mis au jour. Parmi ces fossiles, certains ont été attribués à la forme archaïque d’Homo erectus et les autres à un Homo erectus de forme classique. À Ngandong, quatorze restes d’hominidés ont
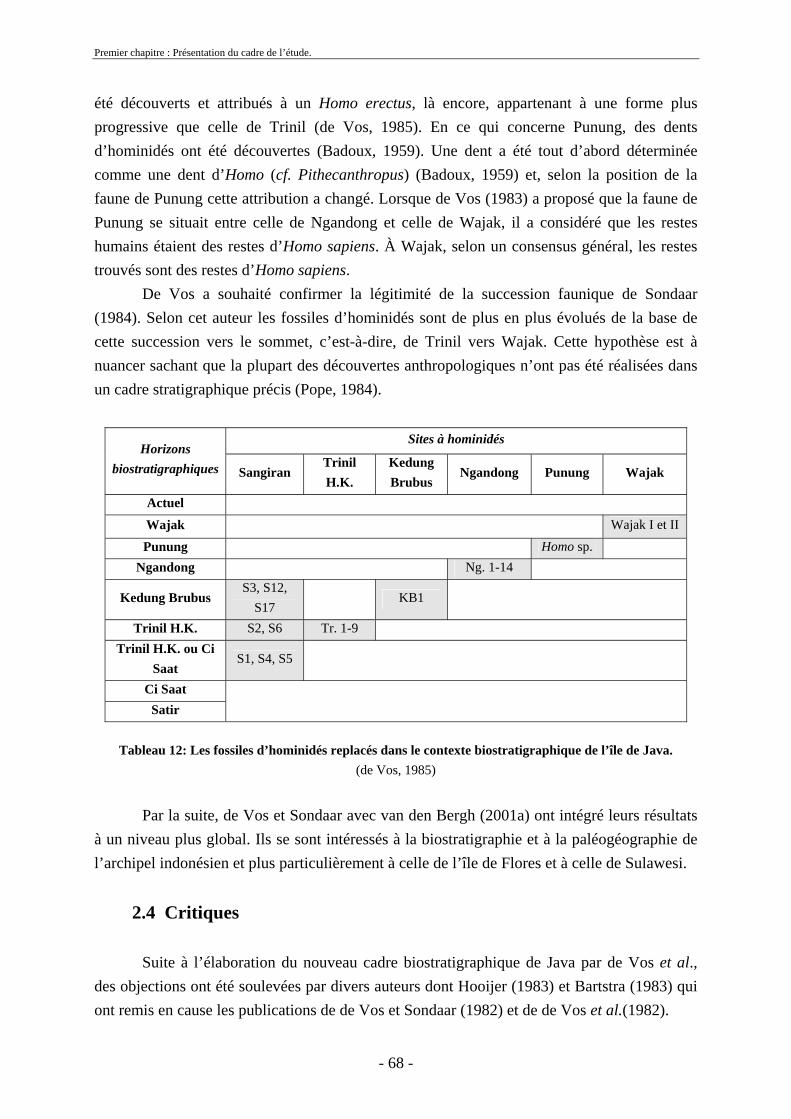
Premier chapitre : Présentation du cadre de l’étude.
- 68 -
été découverts et attribués à un Homo erectus, là encore, appartenant à une forme plus progressive que celle de Trinil (de Vos, 1985). En ce qui concerne Punung, des dents d’hominidés ont été découvertes (Badoux, 1959). Une dent a été tout d’abord déterminée comme une dent d’Homo (cf. Pithecanthropus) (Badoux, 1959) et, selon la position de la faune de Punung cette attribution a changé. Lorsque de Vos (1983) a proposé que la faune de Punung se situait entre celle de Ngandong et celle de Wajak, il a considéré que les restes humains étaient des restes d’Homo sapiens. À Wajak, selon un consensus général, les restes trouvés sont des restes d’Homo sapiens.
De Vos a souhaité confirmer la légitimité de la succession faunique de Sondaar (1984). Selon cet auteur les fossiles d’hominidés sont de plus en plus évolués de la base de cette succession vers le sommet, c’est-à-dire, de Trinil vers Wajak. Cette hypothèse est à nuancer sachant que la plupart des découvertes anthropologiques n’ont pas été réalisées dans un cadre stratigraphique précis (Pope, 1984).
Sites à hominidés Horizons
biostratigraphiques Sangiran Trinil H.K.
Kedung Brubus
Ngandong Punung Wajak
Actuel
Wajak Wajak I et II
Punung Homo sp. Ngandong Ng. 1-14
Kedung Brubus S3, S12,
S17 KB1
Trinil H.K. S2, S6 Tr. 1-9 Trinil H.K. ou Ci
Saat S1, S4, S5
Ci Saat Satir
Tableau 12: Les fossiles d’hominidés replacés dans le contexte biostratigraphique de l’île de Java.
(de Vos, 1985)
Par la suite, de Vos et Sondaar avec van den Bergh (2001a) ont intégré leurs résultats
à un niveau plus global. Ils se sont intéressés à la biostratigraphie et à la paléogéographie de l’archipel indonésien et plus particulièrement à celle de l’île de Flores et à celle de Sulawesi.
2.4 Critiques
Suite à l’élaboration du nouveau cadre biostratigraphique de Java par de Vos et al., des objections ont été soulevées par divers auteurs dont Hooijer (1983) et Bartstra (1983) qui ont remis en cause les publications de de Vos et Sondaar (1982) et de de Vos et al.(1982).

Premier chapitre : Présentation du cadre de l’étude.
- 69 -
Pour Hooijer, Kedung Brubus est une localité qui contient deux faunes : Jetis (Pléistocène inférieur) et Trinil (Pléistocène moyen). Les couches de Jetis se situent en dessous de celles de Trinil mais elles contiennent des éléments de la faune « sino-malaise » de von Koenigswald qui suggèrent un âge du Pléistocène moyen et non inférieur et donc une contemporanéité des deux faunes. En outre, pour cet auteur, l’unité de Kedung Brubus de de Vos et al. (1982) contient également des restes provenant de la localité de Gunung Butak.
Hooijer remet aussi en cause l’exclusivité des fossiles utilisés pour définir Trinil H.K., car selon cet auteur, une unité biostratigraphique ne peut pas être définie à partir d’un seul site. Un autre argument est que les fouilles de Dubois ont dû être conduites dans deux unités stratigraphiques différentes d’âge Pléistocène moyen et Pléistocène supérieur.
De Vos et al. (1982) suggéraient que la localité de Trinil ait été située sur une île d’après la nature endémique de la faune. Pour Hooijer, l’insularité de la localité de Trinil n’était pas possible compte tenu de la taille de la zone, inférieure à quelques kilomètres de diamètre, qui ne prenait pas en compte les sites environnants, comme Tawang où Epileptobos groeneveldtii a été trouvé et Ngancer où de l’Elephas hysudrindicus a été découvert. Les migrations de mammifères auraient été nombreuses pour une si petite île. Il a en fait mal interprété les propos de de Vos et al. (1982) qui ne considéraient pas exclusivement la zone de Trinil.
En outre, il constate l’omission d’une DM4 d’Elephas celebensis dans l’unité de Trinil H.K., ce qui remet en cause l’arrivée d’espèces évoluées, et notamment d’Elephas, à partir de l’unité de Kedung Brubus et donc la postériorité de celle-ci par rapport à Trinil H.K. Enfin pour Hooijer, comme pour Dubois, Epileptobos groeneveldtii était déjà présent à Trinil.
Pour sa part, Bartstra (Hooijer & Bartstra, 1983) remet en cause l’article de de Vos et
al. (1982) à cause des dates des deux unités. Pour cet auteur, l’association faunique de Kedung Brubus est antérieure à celle de Trinil H.K. En effet, il a retrouvé les localisations des découvertes de Dubois et Gunung Butak se situe juste au Nord de Kedung Brubus. Or d’après van Es (1931), la localité de Gunung Butak est plus ancienne, du point de vue stratigraphique, que celle de Kedung Brubus. Gunung Butak correspondrait à la formation du Pléistocène inférieur, qui est antérieure à celle du Pléistocène moyen qui contient la faune de Kedung Brubus. De plus, Dubois aurait enregistré les fossiles de Gunung Butak comme provenant de Kedung Brubus et il aurait aussi enregistré pour cette localité des os achetés dont le lieu précis de découverte n’était pas connu. Pour Bartstra, la faune de Kedung Brubus serait donc le résultat d’un mélange de deux faunes d’âge différent (Pléistocène inférieur et moyen) et ne peut pas être utilisée en tant qu’unité biostratigraphique.
En outre, au sujet de Trinil H.K., Bartstra a repris les arguments géologiques de van Es (1931) et Duyfjes (1936) pour conclure que les dépôts de la faune de Trinil sont des dépôts de terrasses mélangés et donc qu’elle correspond à un mélange de deux faunes d’âges

Premier chapitre : Présentation du cadre de l’étude.
- 70 -
différents (Pléistocène moyen et supérieur). Par conséquent, l’unité de Trinil H.K. ne peut pas non plus être utilisée pour une étude biostratigraphique et chronologique.
À la suite des critiques de Hooijer et Bartstra (1983) qui ont remis totalement en cause la théorie de de Vos et al. (1982b), Sondaar et al. (1983) ont proposé une réplique et ont argumenté leur position.
Les fossiles provenant de la localité de Gunung Butak auraient été nommés comme tels par Dubois et n’ont pas été incorporés dans la liste faunique de Kedung Brubus. Les fossiles de Kedung Brubus proviennent bien du site de Kedung Brubus, à partir d’une zone limitée et d’un seul niveau stratigraphique. En outre, la zone de Butak, où l’on peut retrouver les formations du Pléistocène inférieur et moyen, n’aurait pas livré de fossiles.
Pour Trinil H.K., la liste faunique est basée sur des fossiles provenant d’un seul niveau stratigraphique qui est le niveau inférieur de la rivière Solo en saison sèche et non les strates qui se situent 4 ou 5 mètres au-dessus de ce niveau. Ceci est confirmé par des analyses du fluor contenu dans plusieurs fossiles et qui montrent la même période de dépôt de ces fossiles (Bergman & Karsten, 1952). De plus, si cette faune était un mélange, le nombre de taxons différents serait plus important. En ce qui concerne la DM4 d’Elephas celebensis de Trinil, elle aurait été achetée et donc sa position stratigraphique précise ne serait pas connue. Pour le genre Epileptobos, il ne se trouve pas dans la localité de Trinil exactement mais dans celle de Tawang.
En outre, pour Sondaar, Duyfjes (1936) a donné une signification chronostratigraphique à la lithostratigraphie pour définir la formation du Pléistocène inférieur et sa limite avec celle du Pléistocène moyen.
Le débat s’est poursuivi avec Hooijer et Kurten (1984) et Hooijer (1984) qui réfutent à
nouveau l’argumentation de de Vos et Sondaar et justifient l’ancienneté de la faune de Kedung Brubus par rapport à celle de Trinil.
Hooijer se base sur les taxons et remet en cause plusieurs points de leur raisonnement dont les arguments essentiels sont repris ici. Pour cet auteur, Manis palaeojavanica, Elephas hysudrindicus, Rhinoceros kendengindicus, Epileptobos groeneveldtii sont des formes endémiques de l’île de Java et ne sont donc pas des formes provenant du continent asiatique. La faune de Kedung Brubus présente alors plus de formes endémiques que celle de Trinil ce qui indiquerait une ancienneté de la faune de Kedung Brubus par rapport à celle de Trinil. De plus, d’après les notes de von Koenigswald (1934a) et l’interposition de Hooijer (1952), les couches de Jetis de Kedung Brubus se situent sous les couches de Trinil et sont donc plus anciennes.
En outre, Day et Molleson (1973) ont mis en évidence les limites de l’analyse avec le fluor (Matsu’ura, 1982) sur le fémur de Trinil. Cet auteurs ont mené des analyses avec le fluor et d’autres analyses chimiques qui leur ont permis de conclure qu’il n’était pas possible de distinguer les os provenant de la faune de Jetis de ceux provenant de la faune de Trinil à
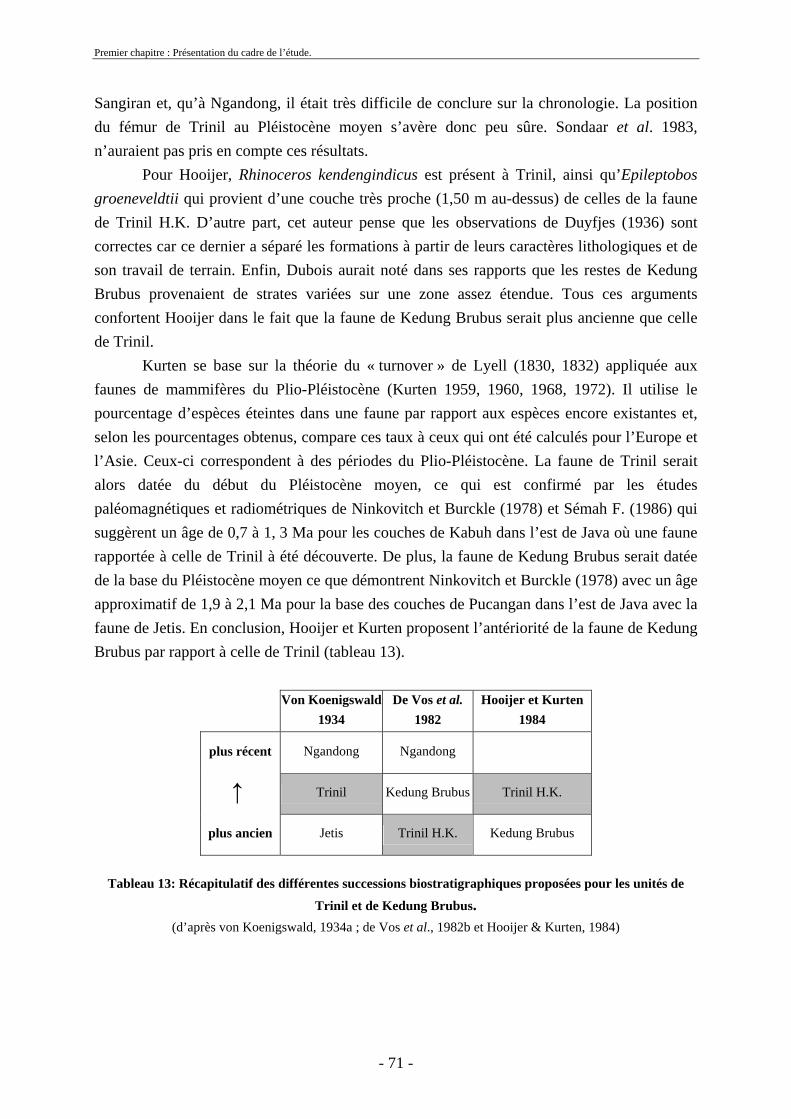
Premier chapitre : Présentation du cadre de l’étude.
- 71 -
Sangiran et, qu’à Ngandong, il était très difficile de conclure sur la chronologie. La position du fémur de Trinil au Pléistocène moyen s’avère donc peu sûre. Sondaar et al. 1983, n’auraient pas pris en compte ces résultats.
Pour Hooijer, Rhinoceros kendengindicus est présent à Trinil, ainsi qu’Epileptobos groeneveldtii qui provient d’une couche très proche (1,50 m au-dessus) de celles de la faune de Trinil H.K. D’autre part, cet auteur pense que les observations de Duyfjes (1936) sont correctes car ce dernier a séparé les formations à partir de leurs caractères lithologiques et de son travail de terrain. Enfin, Dubois aurait noté dans ses rapports que les restes de Kedung Brubus provenaient de strates variées sur une zone assez étendue. Tous ces arguments confortent Hooijer dans le fait que la faune de Kedung Brubus serait plus ancienne que celle de Trinil.
Kurten se base sur la théorie du « turnover » de Lyell (1830, 1832) appliquée aux faunes de mammifères du Plio-Pléistocène (Kurten 1959, 1960, 1968, 1972). Il utilise le pourcentage d’espèces éteintes dans une faune par rapport aux espèces encore existantes et, selon les pourcentages obtenus, compare ces taux à ceux qui ont été calculés pour l’Europe et l’Asie. Ceux-ci correspondent à des périodes du Plio-Pléistocène. La faune de Trinil serait alors datée du début du Pléistocène moyen, ce qui est confirmé par les études paléomagnétiques et radiométriques de Ninkovitch et Burckle (1978) et Sémah F. (1986) qui suggèrent un âge de 0,7 à 1, 3 Ma pour les couches de Kabuh dans l’est de Java où une faune rapportée à celle de Trinil à été découverte. De plus, la faune de Kedung Brubus serait datée de la base du Pléistocène moyen ce que démontrent Ninkovitch et Burckle (1978) avec un âge approximatif de 1,9 à 2,1 Ma pour la base des couches de Pucangan dans l’est de Java avec la faune de Jetis. En conclusion, Hooijer et Kurten proposent l’antériorité de la faune de Kedung Brubus par rapport à celle de Trinil (tableau 13).
Von Koenigswald
1934 De Vos et al.
1982 Hooijer et Kurten
1984
plus récent Ngandong Ngandong
↑ Trinil Kedung Brubus Trinil H.K.
plus ancien Jetis Trinil H.K. Kedung Brubus
Tableau 13: Récapitulatif des différentes successions biostratigraphiques proposées pour les unités de
Trinil et de Kedung Brubus. (d’après von Koenigswald, 1934a ; de Vos et al., 1982b et Hooijer & Kurten, 1984)

Premier chapitre : Présentation du cadre de l’étude.
- 72 -
2.5. Synthèse sur le cadre biostratigraphique de Java
En conclusion, le cadre biostratigraphique établi par von Koenigswald a été utilisé jusque dans les années 80, avant que de Vos et Sondaar ne le révisent.
Même si de Vos et Sondaar ont effectué un travail énorme sur les collections et ont répondu à la plupart des objections posées par Hooijer, Bartstra et Kurten, il reste des points peu clairs au sujet de la chronologie des faunes de Trinil H.K. et de Kedung Brubus. Les zones d’ombres persistent au niveau des espèces endémiques et de leur importance dans ces faunes, de l’insularité de la localité de Trinil dans l’émergence de l’île de Java, de la fiabilité de la méthode au fluor et de la présence d’Epileptobos groeneveldtii et d’Elephas hysudrindicus à Trinil.
Il existe, également, un manque crucial d’informations sur la faune de Ngandong que ce soit au niveau de sa nature ou de sa chronologie (de Vos et al., 1982b). Il semble judicieux de la placer avec beaucoup de prudence dans la succession faunique proposée (Sondaar, 1984) avant d’entreprendre ou d’étudier les restes de nouvelles fouilles de cette localité.
La faune de Punung est constituée essentiellement de restes dentaires qui ont été retrouvés dans des fissures du karst de deux localités différentes et qui par conséquent ne possèdent pas un cadre stratigraphique précis. Il nous paraît donc délicat de proposer une unité biostratigraphique basée sur les restes provenant de fissures karstiques même si ces premiers résultats sont confirmés par les récents travaux sur le site de Gunung Dawu (Storm et al., sous presse). Les recherches menées actuellement dans les grottes de Tabuhan et de Song Terus (M.Q.P.I.) permettront d’affiner le cadre biostratigraphique pour cette période car l’origine stratigraphique des découvertes fauniques dans ces grottes est bien connue.
Finalement, depuis 1984, la succession biostratigraphique quaternaire utilisée pour
Java est celle établie par de Vos et Sondaar (tableaux 5 et 6) mais elle reste un modèle obtenu à partir d’anciennes collections et doit être appliquée avec prudence. Comme nous avons pu le constater l’utilisation de collections anciennes peut être problématique pour connaître la source exacte des fossiles, la localisation et la nature exacte des niveaux stratigraphiques. Pour Sangiran, le cadre biostratigraphique n’est pas tout à fait clair et il est basé sur des ressemblances de quelques espèces avec les unités biostratigraphiques de Java. Il paraît donc indispensable de compléter ces données par de nouveaux résultats. Sangiran permet de donner un cadre chronologique aux autres unités biostratigraphiques de Java car il existe une succession stratigraphique du Pliocène supérieur au Pléistocène moyen. Néanmoins, bien que de bons résultats aient été obtenus sur les tufs (Saleki, 1997), le problème de l’origine des ponces datées dans les niveaux de Kabuh ne permet pas de donner une chronologie fiable pour les différentes faunes rapportées à ce niveau stratigraphique.

Premier chapitre : Présentation du cadre de l’étude.
- 73 -
Les collections fouillées actuellement dont l’origine stratigraphique est bien connue permettront d’affiner le cadre biostratigraphique de Java dans l’avenir et d’éclairer les zones d’ombres, notamment celles de la chronologie des collections de Trinil et de Kedung Brubus. Les fouilles menées sur le dôme de Sangiran devraient permettre de progresser vers l’établissement d’un cadre biostratigraphique pour cette aire géographique et de le comparer au cadre général proposé en 1984.
2.6. Conclusion sur la colonisation de Java
Divers auteurs ont proposé des corrélations entre les faunes plio-pléistocènes de Java, d’Inde et de Chine (tableau 14).
JAVA INDE* INDE** INDE*** CHINE
Narmada Trinil
Boulder Conglomerate Choukoutien
Jetis Pinjor Nihowan
Kali Glagah
Ci Julang
Tatrot
Dhok Pathan
Tableau 14: Corrélations des faunes javanaises, indiennes et chinoises plio-pléistocènes. (d’après Badoux, 1959: * Colbert, 1943 ; **von Koenigswlad, 1956 ; ***Hooijer, 1952 et von Koenigswald,
1949)
Badoux (1959) a également proposé une corrélation entre Java et l’Inde qui est proche
de celle de von Koenigswald où il y a intégré la faune de Punung. De Vos (1993) a réétudié les collections de von Koenigswald et a proposé un nouveau
modèle :
- L’Inde : En Inde, divers sites ont livré des restes paléontologiques qui ont permis de proposer
les corrélations entre les faunes indiennes et javanaises. Dans les couches de la Narmada Homo erectus a été retrouvé en association avec Bos namadicus, Bubalus palaeoindicus, Leptobos sp., Cervus duvauceli, Hexaprotodon namadicus, Stegodon ganesa, Stegodon insignis, Elephas namadicus et Gazella sp. (de Lumley & Sonakia, 1985).

Premier chapitre : Présentation du cadre de l’étude.
- 74 -
Les couches des Siwaliks supérieures regroupent les formations du Tatrot, du Pinjor et du Boulder Conglomerate. Dans ces formations de nombreuses espèces sont présentes comme Hexaprotodon sivalensis, Caprolagus sivalensis, Homotherium ultimum, Nestoritherium sivalense, Megantereon, Stegodon insignis, Elephas hysudricus, Boselaphini et Merycopotamus dissimilis qui sont reliées avec les espèces de Java.
- La Birmanie :
De même, en Birmanie, des restes paléontologiques provenant des couches d’Irrawaddy ont été étudiés par Colbert (1943). Il a décrit les espèces suivantes : Stegolophodon latidens, Stegodon insignis birmanicus, Elephas hysudricus, Rhinoceros sivalensis, Hexaprotodon cf. sivalensis, Merycopotamus dissimilis et un Boselaphinae. Colbert a pu mettre en évidence que la faune des couches d’Irrawaddy et celle de Trinil avaient des espèces en commun.
Quatre des espèces endémiques pléistocènes de Java sont proches d’espèces des
Siwaliks, de la Narmada et de la Birmanie : - Stegodon trigonocephalus avec Stegodon ganesa (= Stegodon insignis) - Elephas hysudrindicus avec Elephas hysudricus - Duboisia santeng avec Boselaphus namadicus - Epileptobos groeneveldtii avec Bos namadicus.
- La Chine :
Von Koenigswald a proposé une origine chinoise pour certains genres comme Pongo et Hylobates dans le Pléistocène inférieur et moyen (tableau 14). De Vos et Sondaar (1982) et Sondaar (1984) ont démontré que le concept de von Koenigswald des assemblages de Jetis et de Trinil du Pléistocène inférieur et moyen était erroné car il était basé sur des associations composées de fossiles provenant des fissures de Punung datés du Pléistocène supérieur et de couches plus anciennes. La faune de Punung est une faune de forêt tropicale qui montre des affinités avec les faunes du Pléistocène supérieur de Sumatra, de Bornéo, du Vietnam et des fissures de la Chine du Sud (Kwangsi et Yunnan) (de Vos, 1993 ; van den Bergh et al., 2001a). Ainsi, la voie sinomalaise n’a été probablement empruntée qu’à partir du Pléistocène supérieur lors du stade isotopique 6 (van den Bergh et al., 1996).
Les faunes les plus anciennes de Java présentent des espèces proches de celles des
Siwaliks ou du sud-est asiatique ou des espèces ayant une plus large distribution incluant la Chine. Et la faune de Punung datée du Pléistocène supérieur a des affinités avec les faunes chinoises. Il existe cependant une exception pour Tetralophodon bumiajuensis. Saegusa (1996) a démontré que cette espèce devait être en fait nommée Sinomastodon. Ce genre est connu du Miocène supérieur au Pléistocène inférieur en Chine et au Japon mais pas en Inde. Il

Premier chapitre : Présentation du cadre de l’étude.
- 75 -
a donc probablement migré vers le sud lors des premiers épisodes froids du Pliocène supérieur.
Ainsi deux voies principales de migration sont proposées pour la colonisation de l’île de Java par les mammifères : une voie indomalaise et une sinomalaise.
En ce qui concerne les migrations humaines, dans la faune de Ci Saat la présence humaine n’est pas certifiée (tableau 5). À partir de la faune de Trinil H.K., la présence d’Homo erectus est avérée jusqu’à la faune de Ngandong. À partir du Pléistocène supérieur, et de la faune de Punung, Homo sapiens apparaît (tableau 6).

- 76 -

- 77 -
SECOND CHAPITRE ETUDE PALEONTOLOGIQUE
Après avoir défini le cadre de notre étude, nous allons maintenant aborder la seconde partie de notre travail : l’étude paléontologique.
1. MATERIEL OSSEUX Le matériel étudié provient des six sites : Tanjung, Sendang Busik, Ngrejeng Plupuh, Grogol Plupuh, Bukuran et Dayu (Koeshardjono, 1988) (tableau 15 et annexe 1).
TANJUNG SENDANG BUSIK NGREJENG GROGOL BUKURAN DAYU
1963 1964 Années de
fouilles 1982
1980 (I) 1981 (II) 1985 1983 1979 1986
1109 dont 15 en 1963
906 en 1964 Nombre total de
restes (NRT) 191 en 1982
393 dont 82 en 1980
311 en 1981245 212 144 33
Jours de fouilles 14 en 1982 19 en 1980 14 en 1981 19 15 24 14
Nombre de restes (NR) des ramassages de
surface
175 62 en 1980 15 en 1981 203 68 346 25
Tableau 15 : Récapitulatif des données concernant les campagnes de fouilles et les ramassages de surface de Tanjung, Sendang Busik, Ngejeng, Grogol, Bukuran et Dayu.
Dans un premier temps, pour chaque site, nous avons étudié la collection fouillée et dans un second temps, la collection des ramassages de surface.
1.1. Travail préparatoire de laboratoire
Chaque reste a été sorti de son sac et recollé lorsqu’il était fragmenté de manière récente. L’étiquette contenue dans le sac a été conservée et une fiche d’identification a été ajoutée où le site, la zone, le numéro, le type, l’espèce et les mesures (longueur : L ; largeur : l et épaisseur : e) ont été notés (figure 21).

Second chapitre : Etude paléontologique.
- 78 -
Site/Zone: Numéro: Type: Espèce: Mesures: L/l/e
Figure 21 : Etiquette des collections de Sangiran
(L : longueur ; l : largeur ; e : épaisseur)
Pour une première détermination du matériel, nous nous sommes basés sur du matériel
de comparaison présent au laboratoire et provenant de divers sites javanais. Nous avons également consulté des ouvrages d’anatomie comparée (Hue, 1907 ; Schmid, 1972 ; Pales & Lambert, 1971 ; Pales & Garcia, 1981 ; Barone, 1999; Hillson, 1986).
Ensuite, pour améliorer les déterminations, nous avons utilisé les différentes diagnoses établies pour les différentes espèces de Java (Stremme, 1911 ; van der Maarel, 1932 ; von Koenigswald, 1933, 1935, 1940 ; Hooijer, 1946b, 1950, 1955, 1958 ; Badoux, 1959 ; Hardjasasmita, 1987 ; van den Bergh, 1999 ; Zaim et al., 2003 et Moigne et al., 2004).
1.2. Base de données Les informations reportées sur les étiquettes ont été compilées dans une base de données, dans un premier temps, sur papier puis informatisée. Elles ont été complétées par de nombreux critères archéologiques (numéro, carré, date, profondeur, association…) paléontologiques (classe, ordre famille, genre, espèce) et taphonomiques (traces de dissolution, de fissuration, d’oxydes, de carnivores ou anthropiques…). Les définitions de ces critères sont données dans le lexique des faunes établi au laboratoire de Préhistoire du Centre Européen de Recherches Préhistoriques de Tautavel (= C.E.R.P.T.) prévu pour l’exploitation de la base de données du matériel archéologique et préhistorique du Département de Préhistoire du M.N.H.N.
1.3. Mesures et photographies Lorsque cela était possible des mesures paléontologiques de l’ossement ont été prises afin de les comparer avec celles prises sur le matériel d’autres collections.
Pour les mesures concernant les proboscidiens, nous avons utilisé celles établies par Hooijer (1955), van den Bergh (1999) et Beden (1979) et pour les rhinocérotidés, celles proposées par Guérin (1980). En ce qui concerne les artiodactyles nous avons utilisé les mesures de Desse et al. (1986) et pour les suidés nous avons également utilisé celles établies par Hardjasasmita (1987). En outre, des photographies numériques de certains ossements ont été prises afin de poursuivre l’étude hors laboratoire et de comparer avec d’autres collections.

Second chapitre : Etude paléontologique.
- 79 -
Les mesures des diamètres mésio-distal (DMD) des dents ont été prises au niveau du collet des dents et celles des diamètres vestibulo-lingual sont les valeurs maximum observées.
Pour l’étude du matériel des ramassages de surface (895 restes au total) la méthodologie est restée la même. Parfois, le reste ne possédait pas de numéro et nous en avons attribué un nouveau en commençant à R 10 000 (R = remanié). Tous les restes d’un même site ont ensuite été rassemblés sur une ou plusieurs étagères proches afin de faciliter leur étude dans la salle des collections du laboratoire de bio-paléoanthropologie de l’université de Gajah Mada.
1.4. Autres collections Nous sommes allés également au Naturalis Museum de Leiden (Pays Bas) pour
étudier les collections ramenées par Dubois. Cette étude nous a permis d’effectuer une base de mesures et de photographies de références sur les cervidés, les bovidés, les rhinocérotidés et les proboscidiens pour comparer nos données.
Nous nous sommes enfin rendus au Muséum de Sangiran pour comparer et étudier les types de fossiles découverts en surface dans cette zone. À Sangiran, nous avons aussi visité les sites considérés dans cette étude afin de mieux comprendre leur stratigraphie et leur environnement. Il a parfois été difficile de retrouver le lieu exact des fouilles car elles ont été organisées pour la plupart il y a plus de vingt ans. Nous avons donc fait appel à d’anciens fouilleurs et à des habitants des villages alentours.
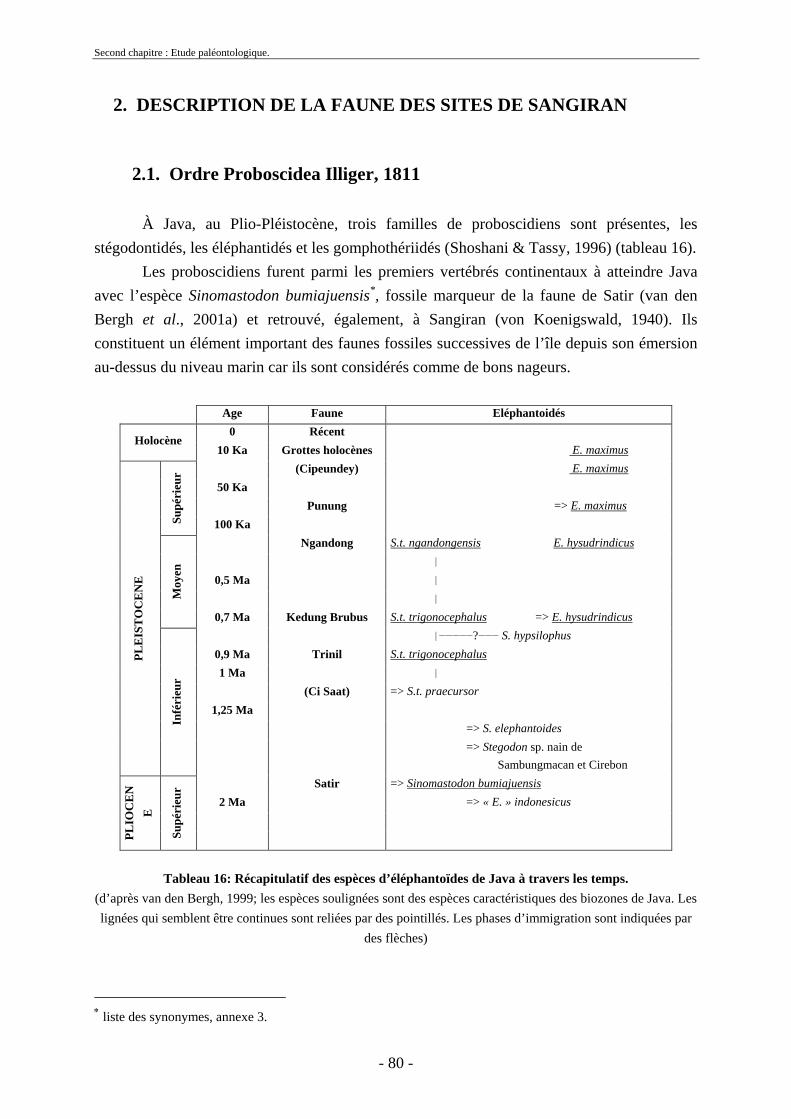
Second chapitre : Etude paléontologique.
- 80 -
2. DESCRIPTION DE LA FAUNE DES SITES DE SANGIRAN
2.1. Ordre Proboscidea Illiger, 1811
À Java, au Plio-Pléistocène, trois familles de proboscidiens sont présentes, les stégodontidés, les éléphantidés et les gomphothériidés (Shoshani & Tassy, 1996) (tableau 16).
Les proboscidiens furent parmi les premiers vertébrés continentaux à atteindre Java avec l’espèce Sinomastodon bumiajuensis*, fossile marqueur de la faune de Satir (van den Bergh et al., 2001a) et retrouvé, également, à Sangiran (von Koenigswald, 1940). Ils constituent un élément important des faunes fossiles successives de l’île depuis son émersion au-dessus du niveau marin car ils sont considérés comme de bons nageurs.
Age Faune Eléphantoidés 0 Récent
Holocène 10 Ka Grottes holocènes E. maximus
(Cipeundey) E. maximus 50 Ka
Punung => E. maximus
Supé
rieu
r
100 Ka Ngandong S.t. ngandongensis E. hysudrindicus
0,5 Ma M
oyen
0,7 Ma Kedung Brubus S.t. trigonocephalus => E. hysudrindicus −−−−−?−−− S. hypsilophus
0,9 Ma Trinil S.t. trigonocephalus 1 Ma
(Ci Saat) => S.t. praecursor 1,25 Ma
=> S. elephantoides
PLE
IST
OC
EN
E
Infé
rieu
r
=> Stegodon sp. nain de Sambungmacan et Cirebon
Satir => Sinomastodon bumiajuensis 2 Ma => « E. » indonesicus
PLIO
CE
NE
Supé
rieu
r
Tableau 16: Récapitulatif des espèces d’éléphantoïdes de Java à travers les temps.
(d’après van den Bergh, 1999; les espèces soulignées sont des espèces caractéristiques des biozones de Java. Les lignées qui semblent être continues sont reliées par des pointillés. Les phases d’immigration sont indiquées par
des flèches)
∗ liste des synonymes, annexe 3.

Second chapitre : Etude paléontologique.
- 81 -
Dès 1857, Falconer propose que « les stégodontes constituent un groupe intermédiaire de proboscidiens à partir duquel les autres espèces divergent par leurs caractères dentaires, avec 2 branches, les mastodontes et les éléphants ». Actuellement deux hypothèses principales sont soutenues pour l’origine de cette famille :
- Cette famille se rapporterait au groupe des mastodontes zygodontes des Mammuthidae (Maglio, 1973),
- Cette famille est attribuée au groupe comprenant les gomphothériidés et les éléphantidés (Tassy, 1982).
Cette deuxième proposition a été confirmée par les analyses cladistiques et les découvertes récentes de fossiles (Saegusa, 1996).
La forme des dents de ces deux familles (Stegodontidae et Elephantidae) diffère au
niveau de la forme des vallées séparant les lames (Maglio, 1973). Les lames adjacentes sont fermées ou en forme de Y à la base chez les stégodontidés et ouvertes ou en forme de U ou V chez les éléphantidés. Ceci est à nuancer car chez certains stégodontes primitifs les vallées peuvent être en forme de V.
2.1.1. Famille Stegodontidae Young-Hopwood, 1935
Cette famille est composée de deux genres Stegolophodon et Stegodon. Elle apparaît
tout d’abord en Asie pendant le Miocène inférieur et devient une des espèces dominantes de la faune asiatique durant le Pliocène et le Pléistocène. Elle est actuellement éteinte (Osborn, 1942 ; Hooijer, 1955 ; Saegusa, 1996).
Stegodon est un élément important des faunes de vertébrés terrestres fossiles en Indonésie. Des membres de ce groupe étaient présents partout en Asie pendant le Plio-Pléistocène et semblent avoir été parmi les premiers colonisateurs d’îles. Des stégodontes ont été identifiés des Phillippines jusqu’en Indonésie (Sulawesi, Sangihe, Flores, Sumba, Timor et Java) et dans les principales îles du Japon (van den Bergh, 1999).
D’après Hooijer (1955) une espèce principale est présente dans le Pléistocène de Java : Stegodon trigonocephalus* Martin, 1887, qui représente la majorité des restes. Il a déterminé également une espèce de petite taille subhypsodonte qui provient de la faune de Jetis (Pléistocène inférieur) : Stegodon hypsilophus* Hooijer, 1954.
Pour van den Bergh (1999), à partir du plus archaïque Stegodon elephantoides, trois sous-espèces successives de S. trigonocephalus peuvent maintenant être reconnues à Java. La détermination de l’espèce Stegodon elephantoides Clift, 1828, originaire de Birmanie (Saegusa, 1996), s’est faite récemment (1993) à partir d’une mandibule trouvée à Bukuran dans la formation de Pucangan (Sangiran, Java central) et datée entre 1,2 et 1,3 Ma (van den ∗ liste des synonymes, annexe 3.
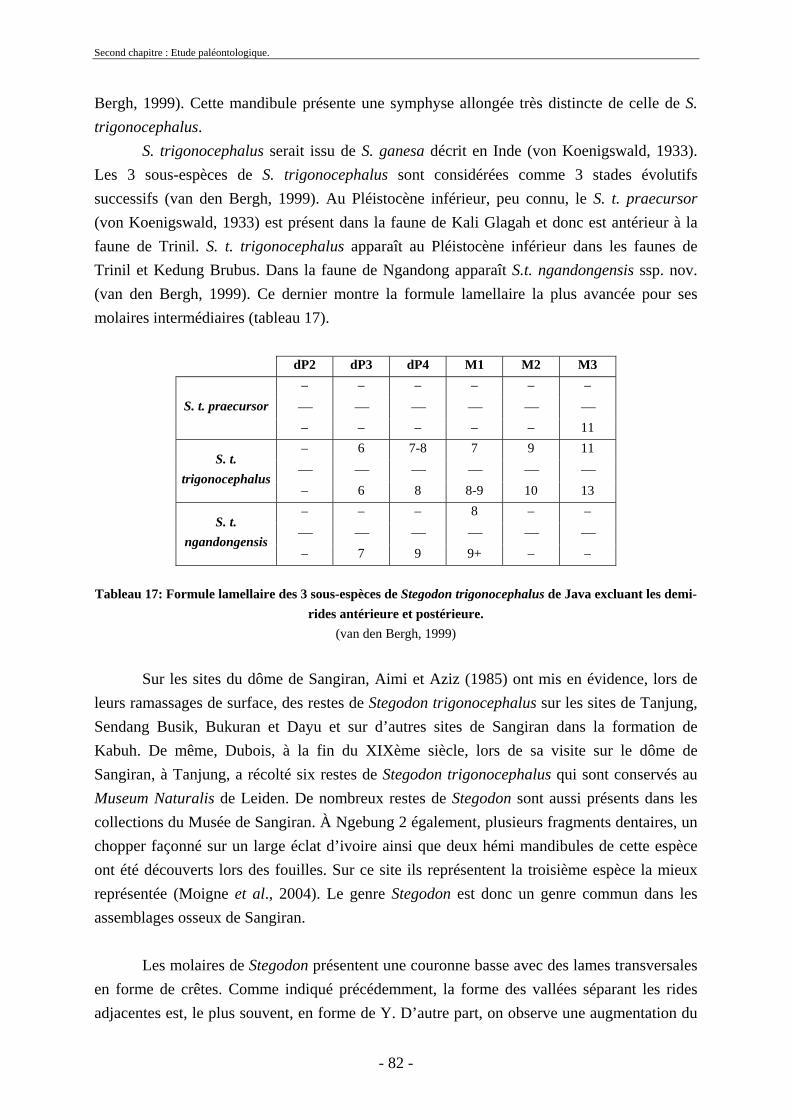
Second chapitre : Etude paléontologique.
- 82 -
Bergh, 1999). Cette mandibule présente une symphyse allongée très distincte de celle de S. trigonocephalus.
S. trigonocephalus serait issu de S. ganesa décrit en Inde (von Koenigswald, 1933). Les 3 sous-espèces de S. trigonocephalus sont considérées comme 3 stades évolutifs successifs (van den Bergh, 1999). Au Pléistocène inférieur, peu connu, le S. t. praecursor (von Koenigswald, 1933) est présent dans la faune de Kali Glagah et donc est antérieur à la faune de Trinil. S. t. trigonocephalus apparaît au Pléistocène inférieur dans les faunes de Trinil et Kedung Brubus. Dans la faune de Ngandong apparaît S.t. ngandongensis ssp. nov. (van den Bergh, 1999). Ce dernier montre la formule lamellaire la plus avancée pour ses molaires intermédiaires (tableau 17).
dP2 dP3 dP4 M1 M2 M3 – – – – – – ⎯ ⎯ ⎯ ⎯ ⎯ ⎯ S. t. praecursor – – – – – 11 – 6 7-8 7 9 11 ⎯ ⎯ ⎯ ⎯ ⎯ ⎯
S. t. trigonocephalus
– 6 8 8-9 10 13 – – – 8 – – ⎯ ⎯ ⎯ ⎯ ⎯ ⎯
S. t. ngandongensis
– 7 9 9+ – –
Tableau 17: Formule lamellaire des 3 sous-espèces de Stegodon trigonocephalus de Java excluant les demi-rides antérieure et postérieure.
(van den Bergh, 1999) Sur les sites du dôme de Sangiran, Aimi et Aziz (1985) ont mis en évidence, lors de
leurs ramassages de surface, des restes de Stegodon trigonocephalus sur les sites de Tanjung, Sendang Busik, Bukuran et Dayu et sur d’autres sites de Sangiran dans la formation de Kabuh. De même, Dubois, à la fin du XIXème siècle, lors de sa visite sur le dôme de Sangiran, à Tanjung, a récolté six restes de Stegodon trigonocephalus qui sont conservés au Museum Naturalis de Leiden. De nombreux restes de Stegodon sont aussi présents dans les collections du Musée de Sangiran. À Ngebung 2 également, plusieurs fragments dentaires, un chopper façonné sur un large éclat d’ivoire ainsi que deux hémi mandibules de cette espèce ont été découverts lors des fouilles. Sur ce site ils représentent la troisième espèce la mieux représentée (Moigne et al., 2004). Le genre Stegodon est donc un genre commun dans les assemblages osseux de Sangiran.
Les molaires de Stegodon présentent une couronne basse avec des lames transversales
en forme de crêtes. Comme indiqué précédemment, la forme des vallées séparant les rides adjacentes est, le plus souvent, en forme de Y. D’autre part, on observe une augmentation du

Second chapitre : Etude paléontologique.
- 83 -
nombre de crêtes en allant vers les troisièmes molaires mais aussi au cours de l’évolution des espèces. Les lames ou crêtes sont compressées et tectiformes. L’émail est composé de deux couches (une externe, une interne) de même épaisseur (figure 22) (van den Bergh, 1999). La table d’usure des molaires inférieures présente une forme concave et celle des molaires supérieures une forme convexe. Les fragments de molaire isolée sont difficiles à identifier.
Le genre Stegodon est représenté par un seul reste dans les collections fouillées. Le
fragment d’une lame de dent jugale (SBII/4/23) provient du site de Sendang Busik. Il faut cependant noter que des restes de Stegodon sp. et plus précisément des fragments de dents jugales ont été retrouvés dans le matériel récolté en surface dans tous les sites hormis à Dayu. Ces fragments sont les plus nombreux à Tanjung (18) et à Bukuran (16) (tableau 18). Tanjung Sendang Busik Ngrejeng Grogol Bukuran F S F S F S F S F S Stegodon sp. 0 18 1 4 0 5 0 2 0 16
Tableau 18 : Nombre de restes du genre Stegodon sur les sites du dôme de Sangiran.
(F : fouilles ; S : surface)
Parmi les 18 fragments dentaires retrouvés à Tanjung, cinq ont pu être plus précisément déterminés car ils possédaient un nombre suffisant de lames (tableau 19). Pour la nomenclature du nombre de lames, chez les Stegodon et les Elephas, nous avons utilisé la méthode classique (Maglio, 1973 ; Beden, 1979) modifiée par van den Bergh (1999) c’est à dire que les demi-lames postérieures et antérieures ne sont pas comptées et sont nommées « x ». Les autres fragments de dents ont été considérés comme des fragments de dents jugales de Stegodon sp. Dents supérieures :
- TDJ64/surface/R10235♦ présente une table d’usure convexe ce qui est caractéristique d’une molaire supérieure. Cette dent fragmentée est constituée de 3 lames à partir de la demi-lame postérieure. Ses dimensions rentrent dans le domaine de variation des M2 de Stegodon trigonocephalus (Hooijer, 19551 ; van den Bergh, 1999). Nous avons déterminé cette dent comme une M2 de Stegodon trigonocephalus (tableau 19).
- La table d’usure de TDJ64/surface/6♦ est convexe, comme pour TDJ64/surface/ R10235. Cette dent fragmentée est constituée de 4 lames à partir de la demi-lame postérieure. Sa fréquence laminaire et ses hauteurs rentrent dans le domaine de variation de celles des M3
♦ planche I, annexe 4. 1 mesures détaillées, tableaux 87, 88 et 89, annexe 5.

Second chapitre : Etude paléontologique.
- 84 -
de Stegodon trigonocephalus (Hooijer, 19551 ; van den Bergh, 1999). Nous avons déterminé cette dent comme une M3 de Stegodon trigonocephalus (tableau 19).
Dents inférieures :
- TDJ64/surface/R10219♦ et TDJ64/surface/ R10224♦ présentent des tables d’usure plutôt concaves. Sur TDJ64/surface/ R10224 la hauteur des lames augmente de l’arrière vers l’avant. Cette dent se courbe vers l’extérieur du côté vestibulaire. TDJ64/surface/R10219 est beaucoup plus usée que TDJ64/surface/ R10224. Ces deux dents possèdent un nombre de lames assez important et sont quasiment entières. Leur fréquence laminaire est élevée mais leur largeur et leur hauteur sont faibles. Ceci suggère une morphologie de déciduale. Et si l’on compare donc ses mesures nous constatons qu’elles rentrent dans le domaine de variation des dimensions des D4 de Stegodon trigonocephalus (Hooijer, 19551 ; van den Bergh, 1999). Nous avons donc déterminé ces deux dents comme des D4 gauches de Stegodon trigonocephalus (tableau 19).
- TDJ64/surface/ R10227♦ présente une table d’usure plutôt concave et elle se courbe vers l’extérieur. Sa dernière cuspide se situe sur la face linguale. Cette dent fragmentée est constituée de 4 lames à partir de la demi-lame postérieure. Ses largeurs et ses hauteurs sont proches de celles des M2 de Stegodon trigonocephalus (Hooijer, 19551 ; van den Bergh, 1999). Nous l’avons déterminée comme une M2 droite de Stegodon trigonocephalus (tableau 19).
Ne disposant que de dents fragmentées nous n’avons pas pu pousser la détermination
jusqu’à la sous-espèce. La position stratigraphique de ces dents en surface ne nous permet pas d’affiner le cadre biostratigraphique de ces découvertes et donc leur détermination.
En conclusion, le nombre de restes de cette espèce est faible dans les collections
fouillées mais assez important en surface car ils sont bien visibles par les collecteurs, ce qui confirme les résultats d’Aimi et Aziz (1985).
♦ planche I, annexe 4. 1 mesures détaillées, tableau 90, annexe 5.
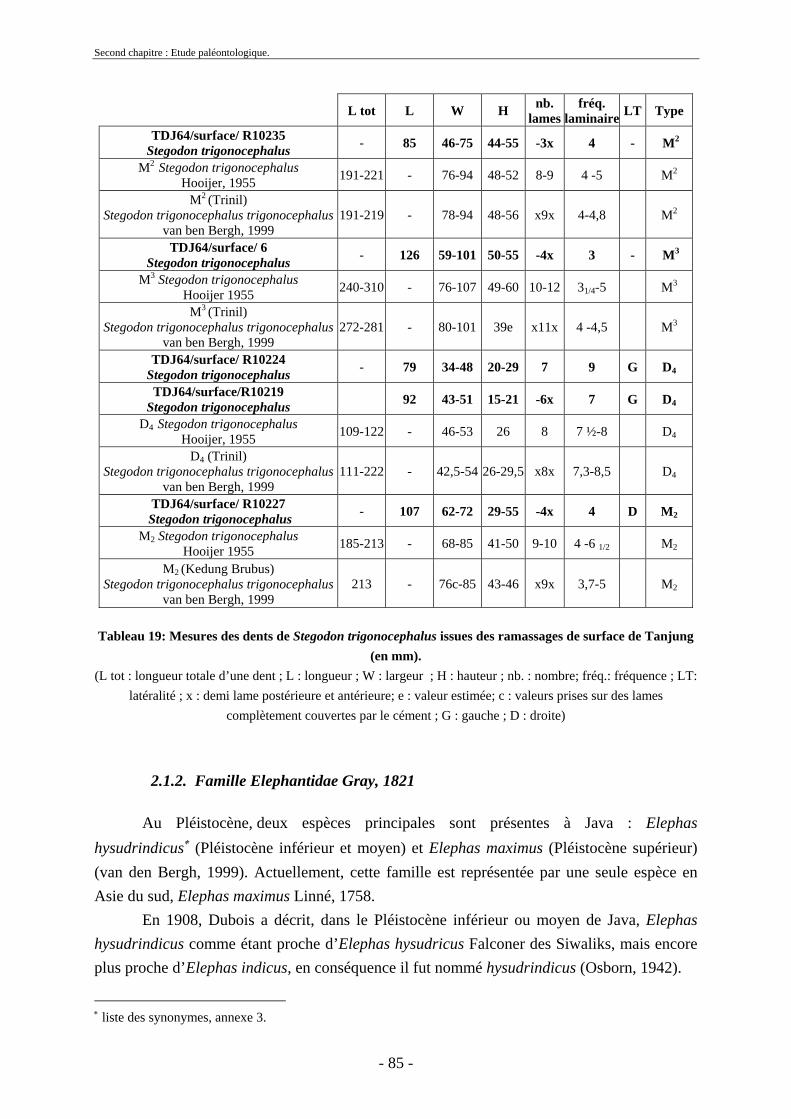
Second chapitre : Etude paléontologique.
- 85 -
L tot L W H nb. lames
fréq. laminaire LT Type
TDJ64/surface/ R10235 Stegodon trigonocephalus - 85 46-75 44-55 -3x 4 - M2
M2 Stegodon trigonocephalus Hooijer, 1955 191-221 - 76-94 48-52 8-9 4 -5 M2
M2 (Trinil) Stegodon trigonocephalus trigonocephalus
van ben Bergh, 1999 191-219 - 78-94 48-56 x9x 4-4,8 M2
TDJ64/surface/ 6 Stegodon trigonocephalus - 126 59-101 50-55 -4x 3 - M3
M3 Stegodon trigonocephalus Hooijer 1955 240-310 - 76-107 49-60 10-12 31/4-5 M3
M3 (Trinil) Stegodon trigonocephalus trigonocephalus
van ben Bergh, 1999 272-281 - 80-101 39e x11x 4 -4,5 M3
TDJ64/surface/ R10224 Stegodon trigonocephalus - 79 34-48 20-29 7 9 G D4
TDJ64/surface/R10219 Stegodon trigonocephalus 92 43-51 15-21 -6x 7 G D4
D4 Stegodon trigonocephalus Hooijer, 1955 109-122 - 46-53 26 8 7 ½-8 D4
D4 (Trinil) Stegodon trigonocephalus trigonocephalus
van ben Bergh, 1999 111-222 - 42,5-54 26-29,5 x8x 7,3-8,5 D4
TDJ64/surface/ R10227 Stegodon trigonocephalus - 107 62-72 29-55 -4x 4 D M2
M2 Stegodon trigonocephalus Hooijer 1955 185-213 - 68-85 41-50 9-10 4 -6 1/2 M2
M2 (Kedung Brubus)
Stegodon trigonocephalus trigonocephalus van ben Bergh, 1999
213 - 76c-85 43-46 x9x 3,7-5 M2
Tableau 19: Mesures des dents de Stegodon trigonocephalus issues des ramassages de surface de Tanjung
(en mm). (L tot : longueur totale d’une dent ; L : longueur ; W : largeur ; H : hauteur ; nb. : nombre; fréq.: fréquence ; LT:
latéralité ; x : demi lame postérieure et antérieure; e : valeur estimée; c : valeurs prises sur des lames complètement couvertes par le cément ; G : gauche ; D : droite)
2.1.2. Famille Elephantidae Gray, 1821
Au Pléistocène, deux espèces principales sont présentes à Java : Elephas
hysudrindicus∗ (Pléistocène inférieur et moyen) et Elephas maximus (Pléistocène supérieur) (van den Bergh, 1999). Actuellement, cette famille est représentée par une seule espèce en Asie du sud, Elephas maximus Linné, 1758.
En 1908, Dubois a décrit, dans le Pléistocène inférieur ou moyen de Java, Elephas hysudrindicus comme étant proche d’Elephas hysudricus Falconer des Siwaliks, mais encore plus proche d’Elephas indicus, en conséquence il fut nommé hysudrindicus (Osborn, 1942).
∗ liste des synonymes, annexe 3.

Second chapitre : Etude paléontologique.
- 86 -
Les molaires d’éléphant sont constituées d’un nombre variable de lames transversales rangées les unes derrières les autres et soudées entre elles par leur base. Ces molaires possèdent une couronne haute. La couche d’émail qui les recouvre et leur donnera leur forme définitive se développe progressivement de l’apex vers la base des lames. Les deux couches de cet émail sont d’une épaisseur différente, l’émail externe est plus fin que l’interne (figure 22) (Saegusa, 1996). Les lames d’une molaire supérieure sont presque toujours parallèles entre elles. Sur une molaire inférieure par contre, les axes des lames sont généralement convergents de la base vers le sommet et le profil des lames est en forme de « S ». La table d’usure, si elle est suffisamment développée, présente un profil longitudinal très caractéristique : convexe sur une molaire supérieure, concave sur une molaire inférieure (Beden, 1979).
À Tanjung, Dubois avait retrouvé une M3 fragmentée d’Elephas hysudrindicus qui est
conservée au Naturalis Museum de Leiden. De même, von Koenigswald a mis en évidence une espèce d’éléphantidé Elephas cf. namadicus à Sangiran (von Koenigswald, 1940). Aimi et Aziz (1985) ont aussi observé la présence d’Elephas sp., sur quelques sites du dôme situés dans la formation de Kabuh. Le genre Elephas est moins commun que le genre Stegodon dans les assemblages de Sangiran.
Un fragment de dent de proboscidien (TDJ82/I/26♦) du site de Tanjung a été déterminé comme un fragment de lame d’Elephas sp. En effet, ce reste présente un émail extérieur fin (2 mm) caractéristique de celui des dents des éléphants (figure 22).
Figure 22 : Comparaison de la structure de l’émail des Stegodontinae et des Elephantinae.
(inner enamel : émail interne ; outer enamel : émail externe ; cementum : cément ; Saegusa, 1996)
♦ planche II, annexe 4.

Second chapitre : Etude paléontologique.
- 87 -
Dans les collections de surface, 8 dents d’Elephas ont été retrouvées à Tanjung (tableau 20). Six ont pu être plus précisément déterminées mais les dents TDJ64/surface/R10221 et TDJ64/surface/R10234 n’ont pu l’être compte tenu de leur trop faible nombre de lames. Nous les avons donc considérées comme des fragments de molaires d’Elephas sp.
Dents supérieures :
- TDJ64/surface/R10223♦ présente une table d’usure convexe et des lames dont le profil est vertical. Cette dent ne montre pas de traces d’usure. Cependant, sur l’une de ces faces, on peut observer une altération qui ne permet pas de mesurer les largeurs des lames de cette dent. Cette dent est constituée de 4 lames depuis la demi-lame postérieure. Ses hauteurs sont dans le domaine inférieur des hauteurs de M2 d’Elephas hysudrindicus et Elephas maximus (Hooijer, 1955). Mais nous pouvons penser que celles-ci sont sous-estimées pour cette dent car les lames sont cassées à leur base. En outre, la fréquence laminaire est proche de celle des M2 d’Elephas hysudrindicus et d’Elephas maximus (Hooijer, 19551). Nous avons déterminé cette dent comme une M2 d’Elephas sp. (tableau 20).
- TDJ64/surface/R10222♦ est enchâssée dans l’os du maxillaire. Sa table d’usure est plutôt convexe. Les valeurs de sa largeur et de sa fréquence laminaire sont proches de celles des M3 d’ Elephas hysudrindicus et Elephas maximus (Hooijer, 19551). Nous avons déterminé cette dent comme une M3 droite d’Elephas sp. (tableau 20).
- TDJ64/surface/R10236♦ et TDJ64/surface/R10231♦ présentent des lames au profil droit et une table d’usure convexe caractéristiques des jugales supérieures. TDJ64/surface/R10236 est constituée de 3 lames médianes très altérées. Ses mesures sont donc difficiles à interpréter. De même, les lames de TDJ64/surface/R10231 sont fragmentées à leur base et peu nombreuses. Par conséquent, nous avons déterminé ces dents comme des fragments de molaires supérieures d’Elephas sp. (tableau 20). Dents inférieures :
- TDJ64/surface/R10226♦ et TDJ64/surface/R10220♦ présentent une table d’usure plutôt concave, contrairement aux dents précédentes. Leurs lames du côté postérieur montrent un profil en « S » caractéristique des molaires inférieures. TDJ64/surface/R10226 est composée de 6 lames à partir de la demi-lame postérieure. TDJ/surface/R10220 est composée de 4 lames à partir de la lame juste avant la demi-lame postérieure. En outre, leurs largeurs et leurs hauteurs rentrent dans le domaine de variation de celles des M3 d’Elephas hysudrindicus
1 mesures détaillées, tableaux 91, 92, 93, 94, 95 et 96, annexe 5. ♦ planche II, annexe 4.
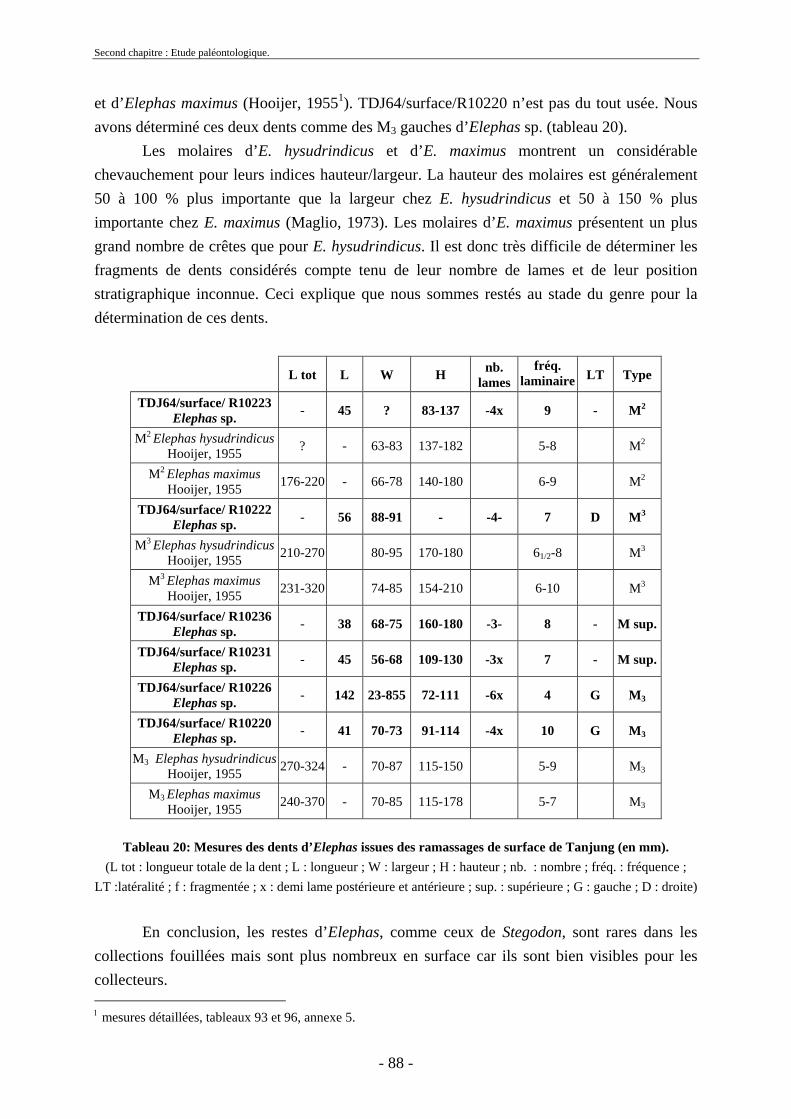
Second chapitre : Etude paléontologique.
- 88 -
et d’Elephas maximus (Hooijer, 19551). TDJ64/surface/R10220 n’est pas du tout usée. Nous avons déterminé ces deux dents comme des M3 gauches d’Elephas sp. (tableau 20).
Les molaires d’E. hysudrindicus et d’E. maximus montrent un considérable chevauchement pour leurs indices hauteur/largeur. La hauteur des molaires est généralement 50 à 100 % plus importante que la largeur chez E. hysudrindicus et 50 à 150 % plus importante chez E. maximus (Maglio, 1973). Les molaires d’E. maximus présentent un plus grand nombre de crêtes que pour E. hysudrindicus. Il est donc très difficile de déterminer les fragments de dents considérés compte tenu de leur nombre de lames et de leur position stratigraphique inconnue. Ceci explique que nous sommes restés au stade du genre pour la détermination de ces dents.
L tot L W H nb. lames
fréq. laminaire LT Type
TDJ64/surface/ R10223 Elephas sp. - 45 ? 83-137 -4x 9 - M2
M2 Elephas hysudrindicus Hooijer, 1955 ? - 63-83 137-182 5-8 M2
M2 Elephas maximus Hooijer, 1955 176-220 - 66-78 140-180 6-9 M2
TDJ64/surface/ R10222 Elephas sp. - 56 88-91 - -4- 7 D M3
M3 Elephas hysudrindicus Hooijer, 1955 210-270 80-95 170-180 61/2-8 M3
M3 Elephas maximus Hooijer, 1955 231-320 74-85 154-210 6-10 M3
TDJ64/surface/ R10236 Elephas sp. - 38 68-75 160-180 -3- 8 - M sup.
TDJ64/surface/ R10231 Elephas sp. - 45 56-68 109-130 -3x 7 - M sup.
TDJ64/surface/ R10226 Elephas sp. - 142 23-855 72-111 -6x 4 G M3
TDJ64/surface/ R10220 Elephas sp. - 41 70-73 91-114 -4x 10 G M3
M3 Elephas hysudrindicus Hooijer, 1955 270-324 - 70-87 115-150 5-9 M3
M3 Elephas maximus Hooijer, 1955 240-370 - 70-85 115-178 5-7 M3
Tableau 20: Mesures des dents d’Elephas issues des ramassages de surface de Tanjung (en mm).
(L tot : longueur totale de la dent ; L : longueur ; W : largeur ; H : hauteur ; nb. : nombre ; fréq. : fréquence ; LT :latéralité ; f : fragmentée ; x : demi lame postérieure et antérieure ; sup. : supérieure ; G : gauche ; D : droite)
En conclusion, les restes d’Elephas, comme ceux de Stegodon, sont rares dans les collections fouillées mais sont plus nombreux en surface car ils sont bien visibles pour les collecteurs. 1 mesures détaillées, tableaux 93 et 96, annexe 5.
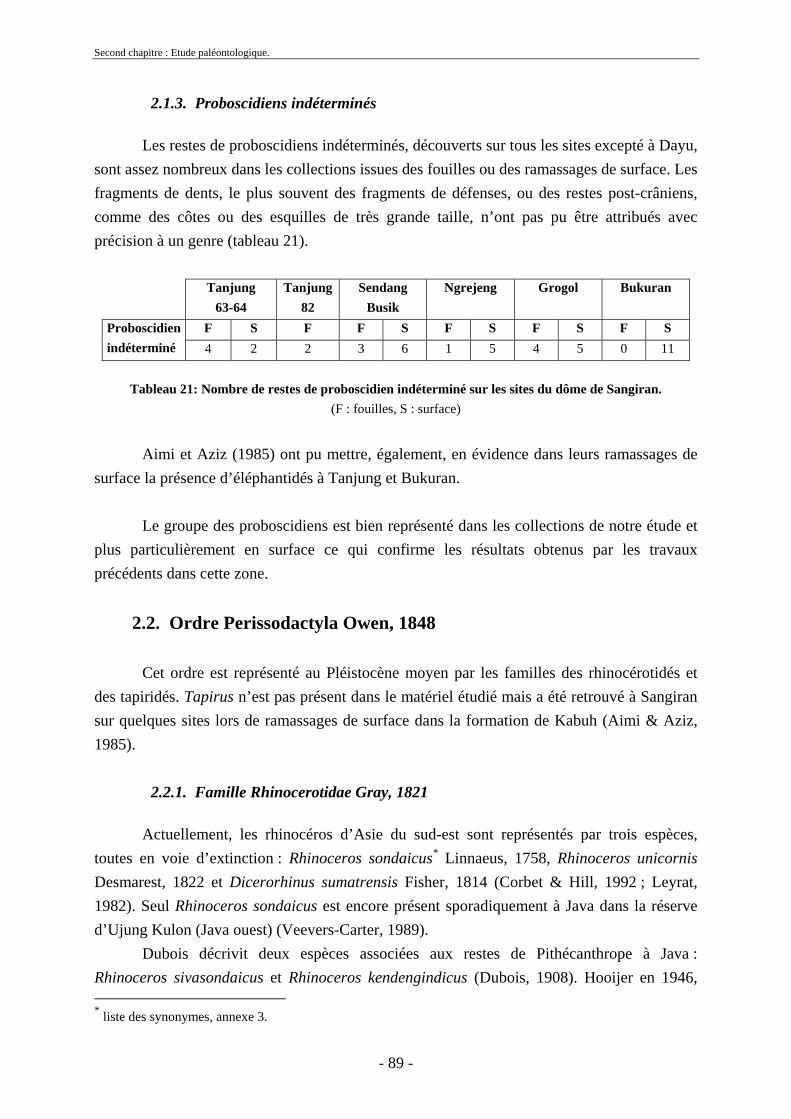
Second chapitre : Etude paléontologique.
- 89 -
2.1.3. Proboscidiens indéterminés
Les restes de proboscidiens indéterminés, découverts sur tous les sites excepté à Dayu, sont assez nombreux dans les collections issues des fouilles ou des ramassages de surface. Les fragments de dents, le plus souvent des fragments de défenses, ou des restes post-crâniens, comme des côtes ou des esquilles de très grande taille, n’ont pas pu être attribués avec précision à un genre (tableau 21).
Tanjung
63-64 Tanjung
82 Sendang
Busik Ngrejeng Grogol Bukuran
F S F F S F S F S F S Proboscidien indéterminé 4 2 2 3 6 1 5 4 5 0 11
Tableau 21: Nombre de restes de proboscidien indéterminé sur les sites du dôme de Sangiran.
(F : fouilles, S : surface)
Aimi et Aziz (1985) ont pu mettre, également, en évidence dans leurs ramassages de surface la présence d’éléphantidés à Tanjung et Bukuran.
Le groupe des proboscidiens est bien représenté dans les collections de notre étude et
plus particulièrement en surface ce qui confirme les résultats obtenus par les travaux précédents dans cette zone.
2.2. Ordre Perissodactyla Owen, 1848
Cet ordre est représenté au Pléistocène moyen par les familles des rhinocérotidés et des tapiridés. Tapirus n’est pas présent dans le matériel étudié mais a été retrouvé à Sangiran sur quelques sites lors de ramassages de surface dans la formation de Kabuh (Aimi & Aziz, 1985).
2.2.1. Famille Rhinocerotidae Gray, 1821
Actuellement, les rhinocéros d’Asie du sud-est sont représentés par trois espèces, toutes en voie d’extinction : Rhinoceros sondaicus* Linnaeus, 1758, Rhinoceros unicornis Desmarest, 1822 et Dicerorhinus sumatrensis Fisher, 1814 (Corbet & Hill, 1992 ; Leyrat, 1982). Seul Rhinoceros sondaicus est encore présent sporadiquement à Java dans la réserve d’Ujung Kulon (Java ouest) (Veevers-Carter, 1989).
Dubois décrivit deux espèces associées aux restes de Pithécanthrope à Java : Rhinoceros sivasondaicus et Rhinoceros kendengindicus (Dubois, 1908). Hooijer en 1946, * liste des synonymes, annexe 3.

Second chapitre : Etude paléontologique.
- 90 -
reprit l’étude de ces formes et fit de la première une sous-espèce de l’actuel R. sondaicus et observa que la deuxième était proche de R. unicornis. La révision de ce matériel par Guérin (1980) a confirmé que Rhinoceros sivasondaicus était une sous-espèce de Rhinoceros sondaicus et que R. kendengindicus était une sous-espèce de R. unicornis (Guérin & Faure, 2002). En résumé, R. sivasondaicus est le synonyme de R. sondaicus sivasondaicus et R. kendengindicus est un synonyme de R. unicornis kendengindicus.
Des restes de rhinocerotidés ont été décrit à Ngebung et à Tanjung et sur d’autres sites du dôme dans la formation de Kabuh (Aimi & Aziz, 1985) ainsi que des restes de Rhinoceros sondaicus à Ngebung 2 (Moigne et al., 2004).
Des restes post-crâniens de rhinocéros ont été mis au jour dans les collections fouillées
de Tanjung. Ces restes se composent d’un sésamoïde (TDJ82/IV/142♦) et d’une extrémité distale de fémur (TDJ64/H14/B18♦).
- L’épiphyse distale du fémur porte en face crâniale une trochlée plus haute que large
constituée d’une large gorge limitée par deux lèvres dont la médiale est à la fois beaucoup plus saillante et beaucoup plus haute que la latérale. En face caudale l’épiphyse distale porte deux gros condyles dont le médial est plus volumineux, séparés par une profonde fosse intercondylienne. Cette épiphyse est donc caractéristique d’un fémur droit (Guérin, 1980).
Le diamètre transversal (DT) de cette extrémité distale de fémur est de 118 mm, son diamètre antéro-postérieur (DAP) n’est malheureusement pas mesurable. Cette valeur est proche de celles des diamètres transversaux de Dicerorhinus sumatrensis actuel et d’un Rhinoceros sondaicus sivasondaicus de petite taille de Kedung Brubus (Guérin & Faure, 20021). Sans le DAP, nous ne pouvons pas conclure avec précision sur l’espèce et nous avons donc attribué ce fémur à Rhinoceros sp. L’épiphyse étant soudée, l’individu était donc un adulte.
- TDJ82/IV/142 présente un corps trapézoïdal avec sur sa face postérieure une surface
articulaire elliptique. Une légère crête sépare cette dernière en deux parties ce qui lui confère une double concavité (Lacombat, 1999). Nous avons donc attribué ce sésamoïde à Rhinoceros sp.
Nous avons, également, observé deux fragments de dents indéterminables de Rhinoceros sp. (NKJ85/surface/R10192 et SBI/D9/surface/R10009) dans les collections de surface de Ngrejeng Plupuh et Sendang Busik I.
Les restes de rhinocérotidés sont rares dans les assemblages osseux de notre étude.
♦ planche III, annexe 4. 1 mesures détaillées, tableau 97, annexe 5.

Second chapitre : Etude paléontologique.
- 91 -
2.3. Ordre Artiodactyla Owen, 1848 Cet ordre représente le plus grand nombre de restes dans les sites étudiés. Les animaux
appartenant à cet ordre sont des ongulés herbivores. A Java, au Pléistocène moyen, les artiodactyles étaient représentés par les familles des hippopotamidés, des suidés, des bovidés et des cervidés.
2.3.1. Famille Hippopotamidae Gray, 1821
Les hippopotames ont maintenant disparu d’Asie (Dennell, 1985), et ne se retrouvent qu’en Afrique avec les espèces : Hippopotamus amphibius Linné, 1758 et Choeropsis liberiensis Morton, 1844 (Faure, 1983).
Les hippopotamidés fossiles d’Asie, en général, et de Java, en particulier, possédaient
trois incisives par hémimandibule et sont donc qualifiés d’hexaprotodontes (Joleaud, 1920 et 1921 ; von Koenigswald, 1934b). Cette famille fut parmi les premiers colonisateurs de l’île de Java avec Hexaprotodon simplex présent dans la faune de Satir (van den Bergh et al., 2001a).
Les trois formes d’hippopotame de Java sont des sous-espèces d’Hexaprotodon sivalensis Falconer et Cautley, 1846. Ces trois sous-espèces, H. sivalensis sivajavanicus*, H. sivalensis koenigswaldi*, H. sivalensis soloensis*, diffèrent des formes continentales par leur taille plus petite. Ces trois sous-espèces javanaises se distinguent les unes des autres par la position de l’orbite, la forme de la mandibule et la position des incisives sur la mandibule. H. sivalensis sivajavanicus et H. sivalensis koenigswaldi sont caractéristiques du Pléistocène inférieur alors que H. sivalensis soloensis caractérise le Pléistocène moyen et supérieur (Hooijer, 1950). Cependant cette diagnose est actuellement peu utilisée car les restes d’Hexaprotodon sont le plus souvent des restes dentaires qui sont très difficiles à attribuer à une sous-espèce et la détermination reste le plus souvent au niveau de l’espèce (de Vos, communication personnelle).
Lors des ramassages de surface d’Aimi et Aziz à Sangiran (1985), la présence de l’hippopotame avait été constatée à Tanjung et Bukuran et sur divers sites du dôme situés dans la formation de Kabuh ainsi que lors de leur fouille à Bukuran dans les couches de Pucangan. Cette présence a aussi été confirmée sur le site de Ngebung 2 (Moigne et al., 2004). Von Koenigswald avait également noté la présence d’Hippopotamus antiquus* dans les couches de Pucangan (von Koenigswald, 1940).
À Bukuran, un squelette entier d’hippopotame a été mis au jour, en 1989, par une équipe franco-indonésienne du Muséum national d’Histoire naturelle de Paris et du Centre
* liste des synonymes, annexe 3.

Second chapitre : Etude paléontologique.
- 92 -
National de Recherches Archéologiques Indonésien de Jakarta (Widianto et al., 1994). Ce squelette a été découvert dans une tranchée creusée dans le cadre d’une étude stratigraphique des dépôts du dôme de Sangiran, au sommet de la formation de Pucangan. Il a pu être complètement reconstitué et correspond au squelette décrit par von Koenigswald conservé au Geological Research and Development Center de Bandung. Ce dernier a été attribué à H. sivalensis koenigswaldi par Hooijer (Moigne, communication personnelle).
L’hippopotame est donc une espèce commune du dôme de Sangiran au Pléistocène. L’hippopotame de Java ou Hexaprotodon sivalensis se retrouve dans le matériel issu des fouilles de Sendang Busik et de Bukuran. Dents inférieures :
- SBII/5/34♦ est caniniforme avec une carène tranchante bien marquée à l’avant ce
qui est caractéristique de la première déciduale (Faure, 1983). Nous n’avons pas pu comparer ses mesures avec celles d’autres D1. Son diamètre mésio-distal (=DMD) est de 22 mm et son diamètre vestibulo-lingual (=DVL) de 13 mm. Elle a donc été déterminée comme une D1 gauche d’Hexaprotodon sivalensis.
- La table d’usure de SBII/9/17♦ présente des dessins en forme de trèfle,
caractéristique des jugales d’hippopotamidé. Sa longueur est plus importante que sa largeur comme chez les jugales inférieures. Nous avons observé la présence d’un tubercule postérieur supplémentaire caractéristique des M3 (Faure, 1983). Cette dent présente donc la morphologie d’une M3 d’hippopotame. De plus, ses mesures rentrent dans le domaine de variation des M3
d’H. sivalensis sivajavanicus de la collection Dubois du Naturalis Museum de Leiden (Hooijer, 19501) (tableau 22). Néanmoins, SBII/9/17 a été déterminée comme une M3 gauche très usée d’Hexaprotodon sivalensis car il est difficile de déterminer des sous-espèces à partir de dents. Cet individu devait être un individu très âgé, sachant que la M3 est la dernière molaire à sortir lors de l’éruption dentaire.
Type DMD DVL M3 de H. s. sivajavanicus 48-55 30-36 M3 de H. s. koenigswaldi 57-60 31-35
M3 de H. s. soloensis 58 32 SBII/9/17 50 36
Tableau 22: Dimensions des M3
d’H. sivalensis sivajavanicus, d’H. sivalensis koenigswaldi et d’H. sivalensis soloensis de la collection Dubois du Naturalis Museum de Leiden (Hooijer, 1950) et de SBII/9/17 (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual )
♦ planche III, annexe 4. 1 mesures détaillées, tableau 99, annexe 5.

Second chapitre : Etude paléontologique.
- 93 -
- Le reste post-crânien issu du site de Bukuran est un fragment d’extrémité proximale de radius gauche (BK79/II/A1/13). La surface antérieure du radius est convexe d’un côté à l’autre, tandis que les surfaces médiales et postérieures sont aplaties (Hooijer, 19501). Collections de surface :
Dans le matériel issu des ramassages de surface, des restes d’Hexaprotodon ont été
trouvés à Bukuran, Ngrejeng et Sendang Busik. Ces restes sont principalement des dents BK79/surface/B1/R10228, NKJ85/surface/R10188, BK79/surface/B1/R10229 et SBI/surface/D9/R10010.
- BK79/surface/B1/R10229♦ présente un tubercule simple, crochu, muni d’une petite
carène postérieure typique des prémolaires d’hippopotamidé (Faure, 1983). Cette dent présente également un cingulum périphérique. Seul le paracône est visible car cette dent est assez usée. Cette morphologie suggère celle d’une P3 d’Hippopotamus amphibius (Hooijer, 1950). Son DMD est de 23 mm et son DVL de 15 mm. Nous n’avons pas pu comparer ces mesures avec celles de spécimens fossiles javanais mais ces dimensions sont très inférieures à celles des P3 d’H. amphibius (DMD : 29,5-43 mm et DVL : 22-34 mm) (Faure, 1983). Ceci confirme la plus petite taille des spécimens javanais par rapport aux formes continentales. Nous avons donc considéré cette dent comme une P3 droite d’Hexaprotodon sivalensis.
- SBI/surface/D9/R10010♦ présente un seul tubercule principal, comme
BK79/surface/B1/R10229 qui est caractéristique des prémolaires d’hippopotamidé. Ce tubercule est muni de quatre styles lui donnant un contour vaguement cruciforme. Ce tubercule présente à sa base un cingulum très épais qui tend à s’ouvrir vers l’arrière. Nous pouvons observer sur cette dent le protocône, le paracône et le métacône. Cette morphologie correspond à celle des P4 d’Hippopotamus amphibius (Hooijer, 19501). Son DMD est de 24 mm et son DVL de 19 mm. Nous n’avons pas pu comparer ces mesures avec celles de spécimens fossiles javanais mais ses dimensions sont très inférieures à celles des P4 d’H. amphibius (DMD : 21-36 mm et DVL : 26-38 mm) (Faure, 1983). Ceci confirme, encore une fois, la plus petite taille des spécimens javanais par rapport aux formes continentales. Cette dent a été considérée comme une P4 gauche d’Hexaprotodon sivalensis.
- BK79/surface/B1/R10228♦ présente quatre tubercules principaux. Sa section est
presque carrée et sa table d’usure possède un contour en forme de feuille de trèfle. En outre, un cingulum entoure la dent sur ses quatre faces. Toutes ces caractéristiques suggèrent une morphologie de molaire supérieure d’hippopotamidé. De plus, la paire de tubercules 1 mesures détaillées, tableau 100, annexe 5. ♦ planche III, annexe 4.
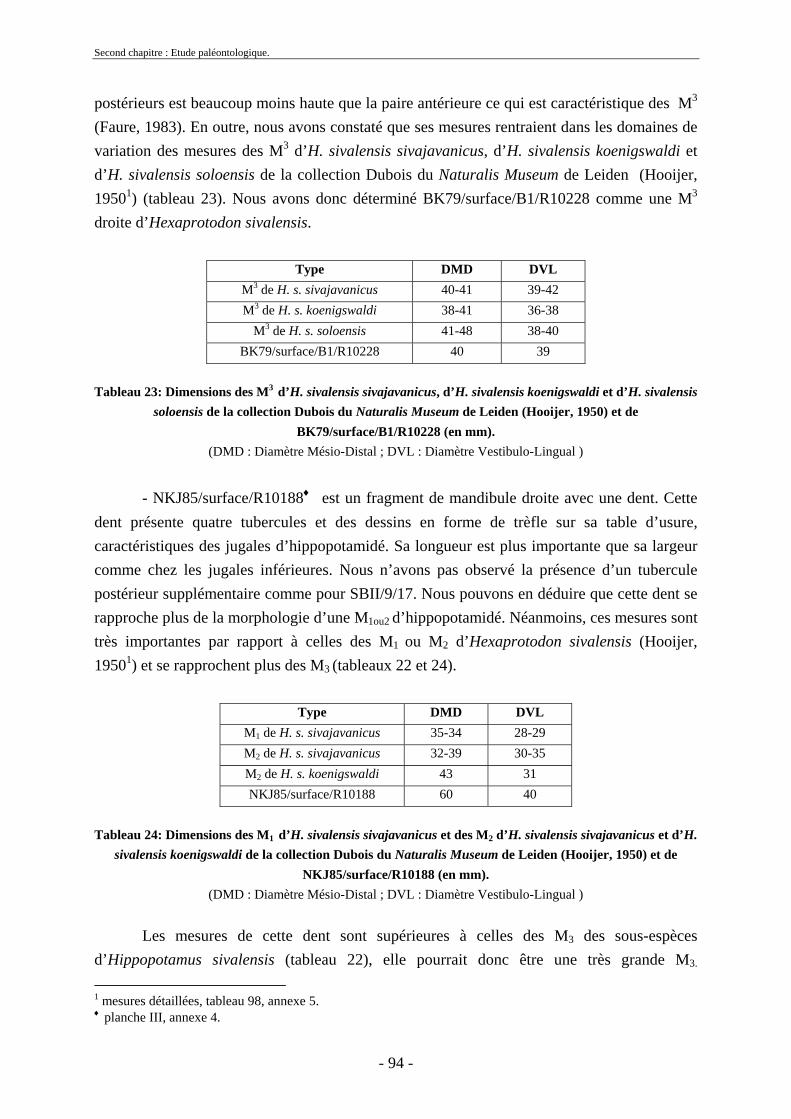
Second chapitre : Etude paléontologique.
- 94 -
postérieurs est beaucoup moins haute que la paire antérieure ce qui est caractéristique des M3
(Faure, 1983). En outre, nous avons constaté que ses mesures rentraient dans les domaines de variation des mesures des M3 d’H. sivalensis sivajavanicus, d’H. sivalensis koenigswaldi et d’H. sivalensis soloensis de la collection Dubois du Naturalis Museum de Leiden (Hooijer, 19501) (tableau 23). Nous avons donc déterminé BK79/surface/B1/R10228 comme une M3 droite d’Hexaprotodon sivalensis.
Type DMD DVL M3 de H. s. sivajavanicus 40-41 39-42 M3 de H. s. koenigswaldi 38-41 36-38
M3 de H. s. soloensis 41-48 38-40 BK79/surface/B1/R10228 40 39
Tableau 23: Dimensions des M3 d’H. sivalensis sivajavanicus, d’H. sivalensis koenigswaldi et d’H. sivalensis
soloensis de la collection Dubois du Naturalis Museum de Leiden (Hooijer, 1950) et de BK79/surface/B1/R10228 (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual )
- NKJ85/surface/R10188♦ est un fragment de mandibule droite avec une dent. Cette
dent présente quatre tubercules et des dessins en forme de trèfle sur sa table d’usure, caractéristiques des jugales d’hippopotamidé. Sa longueur est plus importante que sa largeur comme chez les jugales inférieures. Nous n’avons pas observé la présence d’un tubercule postérieur supplémentaire comme pour SBII/9/17. Nous pouvons en déduire que cette dent se rapproche plus de la morphologie d’une M1ou2 d’hippopotamidé. Néanmoins, ces mesures sont très importantes par rapport à celles des M1 ou M2 d’Hexaprotodon sivalensis (Hooijer, 19501) et se rapprochent plus des M3 (tableaux 22 et 24).
Type DMD DVL M1 de H. s. sivajavanicus 35-34 28-29 M2 de H. s. sivajavanicus 32-39 30-35 M2 de H. s. koenigswaldi 43 31 NKJ85/surface/R10188 60 40
Tableau 24: Dimensions des M1
d’H. sivalensis sivajavanicus et des M2 d’H. sivalensis sivajavanicus et d’H. sivalensis koenigswaldi de la collection Dubois du Naturalis Museum de Leiden (Hooijer, 1950) et de
NKJ85/surface/R10188 (en mm). (DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual )
Les mesures de cette dent sont supérieures à celles des M3 des sous-espèces d’Hippopotamus sivalensis (tableau 22), elle pourrait donc être une très grande M3. 1 mesures détaillées, tableau 98, annexe 5. ♦ planche III, annexe 4.

Second chapitre : Etude paléontologique.
- 95 -
Cependant, du côté postérieur de cette dent, nous pouvons observer la présence d’une alvéole dentaire pour accueillir la M3. Donc, d’après sa morphologie et la présence de cette alvéole postérieure sur la mandibule, nous considérons cette dent comme une M2 droite. Les mesures de NKJ85/surface/R10188 ne correspondent pas aux mesures prises sur les fossiles de Java, nous pouvons nous demander si elles sont exactes ou si nous sommes en présence d’un individu de très grande taille.
À partir de ce matériel, nous n’avons pas pu déterminer la sous-espèce d’Hexaprotodon sivalensis, ne possédant pas la partie incisive de la mandibule et la hauteur des branches horizontales.
2.3.2. Famille Suidae Gray, 1821
Cette famille est actuellement représentée à Java par trois espèces : Sus verrucosus Müller et Schlegel, 1843, Sus scrofa* Linné, 1758 et Sus domesticus, Linné 1758 (Corbet & Hill, 1992).
Pendant le Pléistocène, quatre espèces sont déterminées sur les sites de Java : Sus brachygnathus* Dubois, 1908 ; Sus macrognathus* Dubois, 1908 et Sus sangirensis von Koenigswald, 1963, Sus stremmi von Koenigswald, 1933 (Hardjasasmita, 1987). Sus stremmi est considéré par Hardjasasmita comme un nomen dubium car le nombre de restes lui paraît insuffisant pour permettre de décrire cette espèce.
Aimi et Aziz (1985) ont mis en évidence la présence de restes de Sus brachygnathus à Sendang Busik, Ngrejeng et Ngebung. Lors des fouilles du site de Ngebung 2, cette espèce a également été retrouvée (Moigne et al., 2004). À Tanjung, Dubois avait aussi découvert, lors de ses ramassages de surface, une M3 de Sus brachygnathus (collections Dubois du Naturalis Museum de Leiden).
- Une dent fragmentée de suidé a été découverte dans la collection fouillée de Bukuran (BK79/III/A1/2♦). Cette dent présente deux lobes et une partie du troisième. BK79/III/A1/2 présente une morphologie proche de celle d’une molaire inférieure droite de Sus avec ses trois cuspides. En outre, le diamètre vestibulo-ligual (DVL) de cette dent est de 14,7 mm ce qui correspond aux mesures des M3 de Sus brachygnathus où le DVL varie de 14,5 à 20 mm (tableau 25) (Hardjasasmita, 19871). BK79/III/A1/2 est donc une M3 droite fragmentée et usée attribuée à Sus brachygnathus.
* liste des synonymes, annexe 3. ♦ planche III, annexe 4. 1 mesures détaillées, tableau 101, annexe 5.

Second chapitre : Etude paléontologique.
- 96 -
- Une incisive supérieure a été découverte sur le site de Ngrejeng (NKJ85/II/13♦). Elle ne montre pas de collet et sa racine est courbée. Son DVL est de 9 mm et son DMD de 6 mm. Cette dent a été déterminée comme une incisive supérieure de Sus sp.
- De plus, un fragment distal de tibia gauche (TDJ82/I/29♦) a pu être attribué au genre
Sus sp. dans la collection fouillée de Tanjung. Les gorges de la cochlée distale sont peu profondes et le relief intermédiaire est peu saillant, ce qui est typique de la famille des suidés (Barone, 1999). La gorge oblique médiale est nettement plus petite que la gorge latérale. Son diamètre antéro-postérieur (DAP) est de 23,5 mm et son diamètre transverse (DT) de 25 mm.
Collections de surface : - Dans les ramassages de surface, à Sendang Busik I, nous avons trouvé une dent entière (SBI/surface/R10063♦). Cette dent possède trois lobes à deux cuspides et un quatrième avec une seule cuspide plus ou moins importante. Cette morphologie correspond à celle des M3 de suidé.
L’espèce Sus brachygnathus diffère de S. macrognathus au niveau de la M3. La M3 de S. macrognathus est relativement plus longue que celle de Sus brachygnathus. Dans la vallée médiane sur le bord buccal une cuspide accessoire est présente, celle-ci est plus large et plus complexe que le même élément de Sus brachygnathus.
Ici, nous n’avons pas observé la présence de ce cuspide accessoire et les mesures de cette dent sont proches de celles des M3 de Sus brachygnathus des sites de Trinil, Kebonduren et Kaligede (Hardjasamita, 19871) (tableau 25). Nous avons donc déterminé cette dent comme une M3 droite de Sus brachygnathus bien qu’elle paraisse un peu plus gracile au niveau du DVL que les M3 mesurées par Hardjasasmita (1987).
Type DMD DVL
M3 de Sus brachygnathus 29,1-37,8 14,5-20 SBI/surface/R10063 32 13
Tableau 25: Dimensions des M3 de Sus brachygnathus de Trinil, Kebonduren et Kaligede (Hardjasasmita,
1987) et de SBI/surface/R10063 (en mm). (DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual )
La famille des suidés est très peu représentée dans les collections fouillées et de
surface.
♦ planche III, annexe 4. 1 mesures détaillées, tableau 101, annexe 5.

Second chapitre : Etude paléontologique.
- 97 -
2.3.3. Famille Bovidae Gray, 1821 Actuellement, à Java, cette famille est représentée par Bos javanicus (ou banteng)
d’Alton, 1823, Bubalus bubalis (ou arnee ou kerbau) Linné, 1758, Capra hircus, Linné 1758 et Ovis aries, Linné 1758 (Corbet & Hill, 1992). Toutes ces espèces ont été domestiquées. Cette famille est très importante pour tous les sites car la majorité des restes fouillés appartiennent à celle-ci et plus particulièrement à la sous-famille des Bovinae.
2.3.3.1. Sous-famille Bovinae Gray, 1821
Au Pléistocène moyen, quatre espèces principales sont connues à Java (Hooijer,
1958) : Bubalus palaeokerabau∗ Dubois, 1908 ; Bibos palaesondaicus* Dubois, 1908 et Epileptobos groeneveldtii* Hooijer, 1956 qui sont de grande taille et un bovidé de petite taille Duboisia santeng* (Dubois, 1891). D’autres auteurs, ont déterminé une espèce proche de Duboisia santeng mais de plus grande taille : Duboisia saatensis* von Koenigswald, 1934. Cette détermination est basée sur quatre dents trouvées à Sangiran et à Bumiaju (Geraads, 1979a).
Bubalus palaeokerabau Dubois, 1908 Les ramassages de surface d’Aimi et Aziz (1985) à Glagahombo et les ramassages de
Dubois à Tanjung (collection du Naturalis Museum de Leiden) ont confirmé la présence de Bubalus palaeokerabau dans cette zone. En outre, cette espèce a été mise en évidence lors des fouilles de Ngebung 2 (Moigne et al., 2004).
Bubalus palaeokerabau est un bovidé de grande taille. Les chevilles osseuses de cette espèce sont subtriangulaires en section, à face supérieure est plate, et l'angle antéro-supérieur est défini (figure 23). Les cornes divergent en arrière avec un angle de presque 180° et se courbent de façon très régulière, vers l’extérieur et en arrière (Hooijer, 1958). L’envergure de celles-ci est très importante, elle peut varier entre 83 et 165 cm et est supérieure à celles des Bubalus actuels de Java (Hooijer, 1958).
∗ liste des synonymes, annexe 3.

Second chapitre : Etude paléontologique.
- 98 -
Figure 23: Section moyenne d’une cheville osseuse gauche de Bubalus palaeokerabau, au-dessus de la base, échelle 3/8.
(d’après Stremme, 1911)
- Des restes de Bubalus palaeokerabau ont été mis au jour lors des fouilles de
Tanjung, 2 fragments de chevilles osseuses (TDJ82/IV/128 et TDJ64/I9/58 ) et un crâne entier (Jacob, 1975 b) (figures 24 a et b). Ce crâne est conservé au musée du laboratoire de bio-paléoanthropologie de l’université Gadjah Mada de Yogyakarta.
a b
Figure 24: Crâne de Bubalus palaeokarabau découvert lors des fouilles de 1964 à Tanjung. (a Jacob, après 1971 et b Jacob, 1975b)
- TDJ82/IV/128 est un fragment médian de cheville osseuse. Sa hauteur est de 20 mm et son diamètre antéro-postérieur (DAP) est de 60 mm.
- TDJ64/I9/58 est un fragment médian de cheville osseuse. Sa hauteur est de 47 cm et don DAP de 63 mm.

Second chapitre : Etude paléontologique.
- 99 -
Collections de surface :
En surface, une dent a été découverte à Bukuran (BK79/surface/D2/R10215♦) et un fragment de cheville osseuse (NKJ85/surface/R10128) à Ngrejeng Plupuh.
- BK79/surface/D2/R10215 est une dent dont les mesures de longueur et largeur sont
très similaires avec des styles proéminents et fortement renflés. Nous avons calculé l’indice d’hypsodontie de cette dent selon la méthode d’Heintz (1970) :
Ih = (hauteur de la molaire au métacône x 100)/ longueur de la molaire au collet. Cet indice d’hypsodontie atteint très rarement la valeur 100 pour les molaires de
cervidés alors que pour les molaires de bovidés cet indice est toujours supérieur à 100 (Heintz, 1970). Nous n’avons pas utilisé la méthode de Janis (1984) par manque de deuxième et troisième molaires associées. L’indice d’hypsodontie, selon Heintz, de BK79/surface/D2/R10215 est égal à 190. Ces caractères suggèrent la morphologie d’une molaire supérieure de bovidé. De plus, cette dent possède un style distal très prononcé caractéristique des M3 (Hillson, 1986). Ses dimensions sont très supérieures à celles de Bibos javanicus et Epileptobos groeneveldtii (Hooijer, 1958) (tableau 26). BK79/surface/D2/R10215 a donc été déterminée comme une M3 gauche de Bubalus palaeokerabau.
Type DMD DVL
M3 de Bibos javanicus 27-29 18-20 M3 d’Epileptobos groeneveldtii 28-30 18-21
BK79/surface/D2/R10215 40 31
Tableau 26: Dimensions des M3 de Bibos javanicus et Epileptobos groeneveldtii (Hooijer, 1958) et de BK79/surface/D2/R10215 (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual )
- NKJ85/surface/R10128 est un fragment médian de cheville osseuse de 126 mm de
longueur. Son DAP est de 97 mm et sa hauteur de 64 mm.
Bibos palaesondaicus Dubois, 1908
Bibos palaesondaicus du Pléistocène est probablement l’ancêtre direct de la forme vivante du banteng de Java : Bibos javanicus* d'Alton, 1823. Le crâne de Bibos palaesondaicus est proportionnellement plus large que celui de Bibos javanicus (Hooijer, 1958). Les chevilles osseuses de cette espèce sont ovales en coupe à leur base et leur diamètre ♦ planche V, annexe 4. *liste des synonymes, annexe 3.

Second chapitre : Etude paléontologique.
- 100 -
horizontal dépasse le vertical d'un tiers. Les chevilles diminuent rapidement en diamètre pendant qu’elles s'incurvent doucement vers le haut. La surface des chevilles osseuses montre des rainures longitudinales, plus distinctes sur les faces inférieures et postérieures (Hooijer, 1958) (figure 25).
Figure 25: Section transversale proche de la pointe d’une cheville osseuse gauche de Bibos palaesondaicus, échelle 2/5.
(d’après Stremme, 1911)
Sur divers sites dans la formation de Kabuh, Aimi et Aziz (1985) ont découvert des
restes de Bibos palaesondaicus lors de leur ramassage de surface. A Ngebung 2, aucun reste de cette espèce n’a été découvert (Moigne et al., 2004).
Deux fragments de chevilles osseuses (TDJ64/H7/2B♦ et TDJ64/F4/89♦) et un crâne de Bibos palaesondaicus (Jacob, après 1971?) (figure 26) ont été observés dans la collection fouillée de Tanjung. En ce qui concerne le crâne, n’ayant pu y avoir accès, nous ne savons pas où il est entreposé, néanmoins, il est pris en compte pour la suite de cette étude.
Figure 26: Crâne de Bibos palaesondaicus découvert lors des fouilles de 1964 à Tanjung. (Jacob, après 1971)
♦ planche V, annexe 4.

Second chapitre : Etude paléontologique.
- 101 -
- TDJ64/H7/2B est un fragment médian de cheville osseuse fragmenté aux deux extrémités. Nous avons pu observer la présence d’une rainure profonde sur la face inférieure. Son diamètre antéro-postérieur (DAP) est de 82 mm et sa hauteur est de 55 mm.
- TDJ64/F4/89 est un fragment de frontal avec une partie de la cheville osseuse. Cette cheville s’incurve doucement vers le haut. Son DAP est de 130 mm au niveau du frontal et sa hauteur, au même niveau, est de 73 mm.
Dans les autres collections fouillées ou de surface nous n’avons pas pu mettre en évidence de restes attribuables à Bibos palaesondaicus.
Epileptobos groeneveldtii a été retrouvé parmi le matériel du site de Ngebung 2
(Moigne et al., 2004), en surface dans la formation de Kabuh sur le site de Bapang (Aimi & Aziz, 1985) et dans la formation de Pucangan (von Koenigswald, 1940). Sur les sites de notre étude nous n’avons pas mis en évidence sa présence.
Duboisia santeng (Dubois, 1891)
Duboisia santeng est un petit bovidé endémique à Java, aujourd’hui éteint, qui, par ses caractères dentaires et crâniens, selon Hooijer (1958) se rapproche plutôt des espèces actuelles Boselaphus tragocamelus et Tetraceros quadricornis.
Lors des ramassages de surface d’Aimi et Aziz (1985), des fossiles de Duboisia santeng ont été retrouvés entre autres à Tanjung et à Sendang Busik et également sur le site de Ngebung 2 (Moigne et al., 2004). Von Koenigswald avait également remarqué sa présence dans la formation de Kabuh (von Koenigswald, 1940). Cette espèce est très commune sur les sites javanais du Pléistocène.
Les chevilles osseuses de cette espèce de bovidé sont courtes et sub-triangulaires à la
base, mais plutôt compressées d'en haut vers le bas. Duboisia présente des cornes chez les deux sexes. La morphologie de ses dents est proche de celles des petits cervidés javanais mais sa dentition est très hypsodonte et on note le plus souvent la présence d’un pilier basilaire. De plus, la surface de l’émail est lisse (Hooijer, 1958).
Dans le matériel, huit dents ont été mises au jour dans les collections fouillées de
Tanjung, Ngrejeng et Bukuran. TDJ82/IV/46, TDJ82/IV/58, TDJ82/IV/138, NKJ85/III/10005, NKJ85/III/1, NKJ85/III/16, NKJ85/III/15 et BK79/II/A1/10004.

Second chapitre : Etude paléontologique.
- 102 -
Dents supérieures : - TDJ82/IV/46♦ et NKJ85/III/10005♦ présentent une forme carrée. Ces dents sont bilobées à quatre cuspides. Les cônes sont bien soudés et nous pouvons remarquer la présence d’un pilier basilaire. L’émail de ces dents est lisse. Nous avons calculé ensuite l’indice d’hypsodontie (Ih) (Heintz, 1970) de ces dents défini précédemment pour les collections de surface:
Les indices d’hypsodontie des deux dents considérées sont : - Ih TDJ82/IV/46 = 193, - Ih NKJ85/III/10005 = 106.
Tous ces caractères semblent correspondre à la morphologie des molaires supérieures de bovidé. Afin de corroborer cela, nous avons calculé l’indice i (Heintz, 1970).
i = (largeur * 100)/ longueur Sachant que les dents des cervidés sont proportionnellement plus larges que celles des
bovidés, cet indice est supérieur à 100 pour les molaires supérieures de cervidés et inférieur à 100 pour les molaires supérieures de bovidés. Cet indice peut également être utilisé sur des dents usées ou très usées car la largeur n’est pas modifiée au cours de l’usure, et donc il ne croît que très légèrement en fonction de l’usure.
Pour les deux dents considérées les valeurs de cet indice sont : - i (TDJ82/IV/46) = 96, - i (NKJ85/III/10005) = 93, ce qui démontre leur attribution aux bovidés.
Enfin, ceci est confirmé par la comparaison de leurs dimensions avec celles des M1 et M2 de Duboisia santeng des collections Dubois du Naturalis Museum de Leiden, de la collection de Punung (Badoux, 19591), de la collection de Trinil (Stremme, 19111) et des M1 et M2 d’Epileptobos groeneveldtii de Leiden. Les dimensions de TDJ81/IV/46 et NKJ85/III/10005 rentrent dans le domaine de variation des M1 et M2 de Duboisia santeng et semblent plus proches des dimensions des M2 (figure 27).
♦ planche V, annexe 4. 1 mesures détaillées, tableaux 104, 106 et 107, annexe 5.
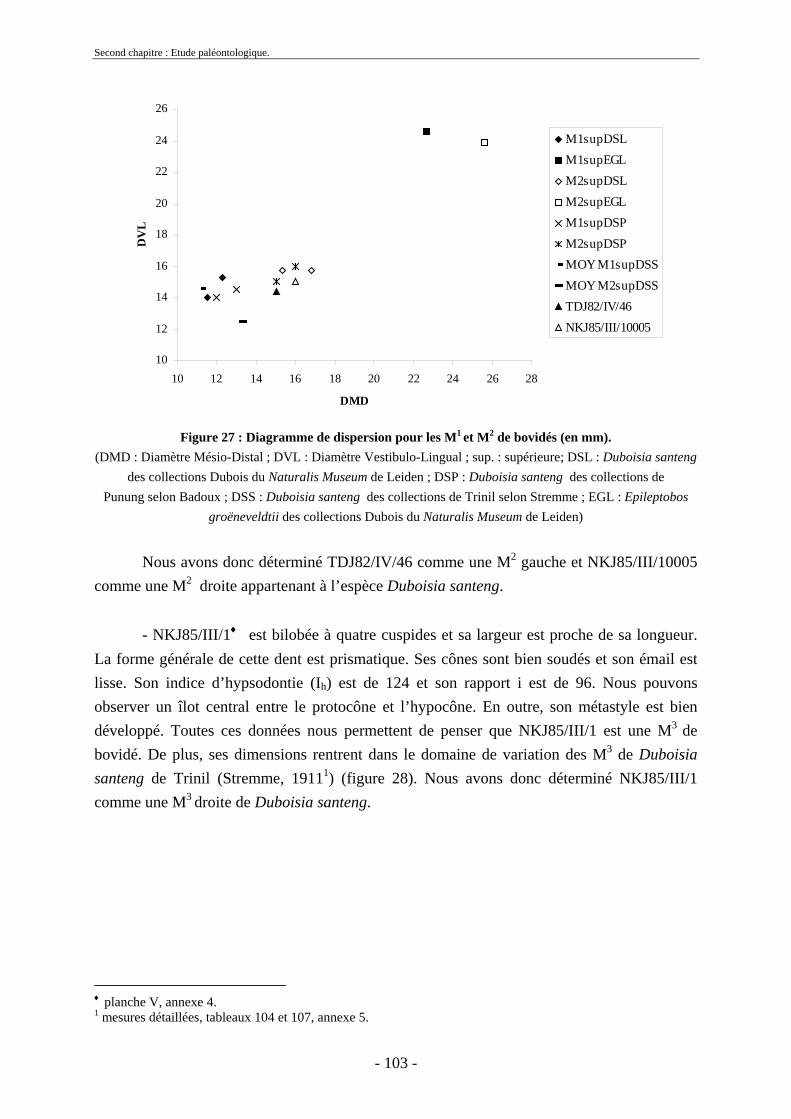
Second chapitre : Etude paléontologique.
- 103 -
10
12
14
16
18
20
22
24
26
10 12 14 16 18 20 22 24 26 28
DMD
DV
LM1supDSLM1supEGLM2supDSLM2supEGLM1supDSPM2supDSPMOY M1supDSSMOY M2supDSSTDJ82/IV/46NKJ85/III/10005
Figure 27 : Diagramme de dispersion pour les M1 et M2 de bovidés (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual ; sup. : supérieure; DSL : Duboisia santeng des collections Dubois du Naturalis Museum de Leiden ; DSP : Duboisia santeng des collections de
Punung selon Badoux ; DSS : Duboisia santeng des collections de Trinil selon Stremme ; EGL : Epileptobos groëneveldtii des collections Dubois du Naturalis Museum de Leiden)
Nous avons donc déterminé TDJ82/IV/46 comme une M2 gauche et NKJ85/III/10005
comme une M2 droite appartenant à l’espèce Duboisia santeng.
- NKJ85/III/1♦ est bilobée à quatre cuspides et sa largeur est proche de sa longueur. La forme générale de cette dent est prismatique. Ses cônes sont bien soudés et son émail est lisse. Son indice d’hypsodontie (Ih) est de 124 et son rapport i est de 96. Nous pouvons observer un îlot central entre le protocône et l’hypocône. En outre, son métastyle est bien développé. Toutes ces données nous permettent de penser que NKJ85/III/1 est une M3 de bovidé. De plus, ses dimensions rentrent dans le domaine de variation des M3 de Duboisia santeng de Trinil (Stremme, 19111) (figure 28). Nous avons donc déterminé NKJ85/III/1 comme une M3 droite de Duboisia santeng.
♦ planche V, annexe 4. 1 mesures détaillées, tableaux 104 et 107, annexe 5.
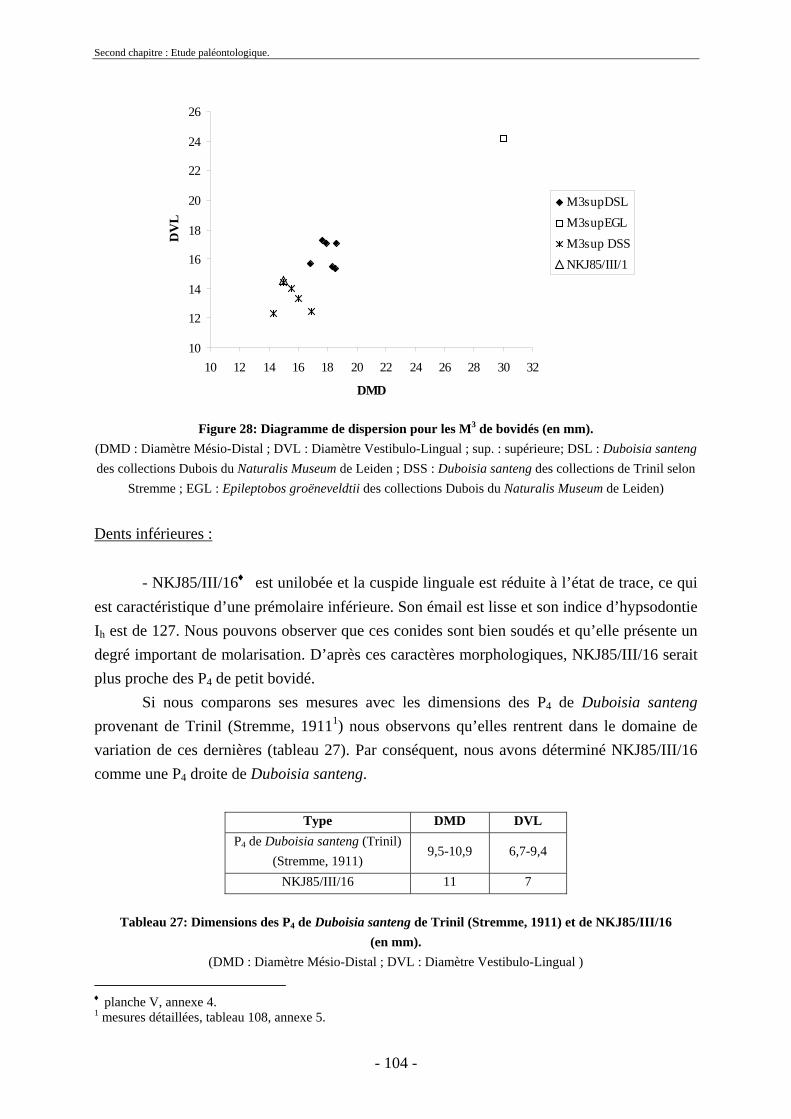
Second chapitre : Etude paléontologique.
- 104 -
10
12
14
16
18
20
22
24
26
10 12 14 16 18 20 22 24 26 28 30 32
DMD
DV
L
M3supDSLM3supEGLM3sup DSSNKJ85/III/1
Figure 28: Diagramme de dispersion pour les M3 de bovidés (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual ; sup. : supérieure; DSL : Duboisia santeng des collections Dubois du Naturalis Museum de Leiden ; DSS : Duboisia santeng des collections de Trinil selon
Stremme ; EGL : Epileptobos groëneveldtii des collections Dubois du Naturalis Museum de Leiden) Dents inférieures :
- NKJ85/III/16♦ est unilobée et la cuspide linguale est réduite à l’état de trace, ce qui est caractéristique d’une prémolaire inférieure. Son émail est lisse et son indice d’hypsodontie Ih est de 127. Nous pouvons observer que ces conides sont bien soudés et qu’elle présente un degré important de molarisation. D’après ces caractères morphologiques, NKJ85/III/16 serait plus proche des P4 de petit bovidé.
Si nous comparons ses mesures avec les dimensions des P4 de Duboisia santeng provenant de Trinil (Stremme, 19111) nous observons qu’elles rentrent dans le domaine de variation de ces dernières (tableau 27). Par conséquent, nous avons déterminé NKJ85/III/16 comme une P4 droite de Duboisia santeng.
Type DMD DVL P4 de Duboisia santeng (Trinil)
(Stremme, 1911) 9,5-10,9 6,7-9,4
NKJ85/III/16 11 7
Tableau 27: Dimensions des P4 de Duboisia santeng de Trinil (Stremme, 1911) et de NKJ85/III/16 (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual ) ♦ planche V, annexe 4. 1 mesures détaillées, tableau 108, annexe 5.
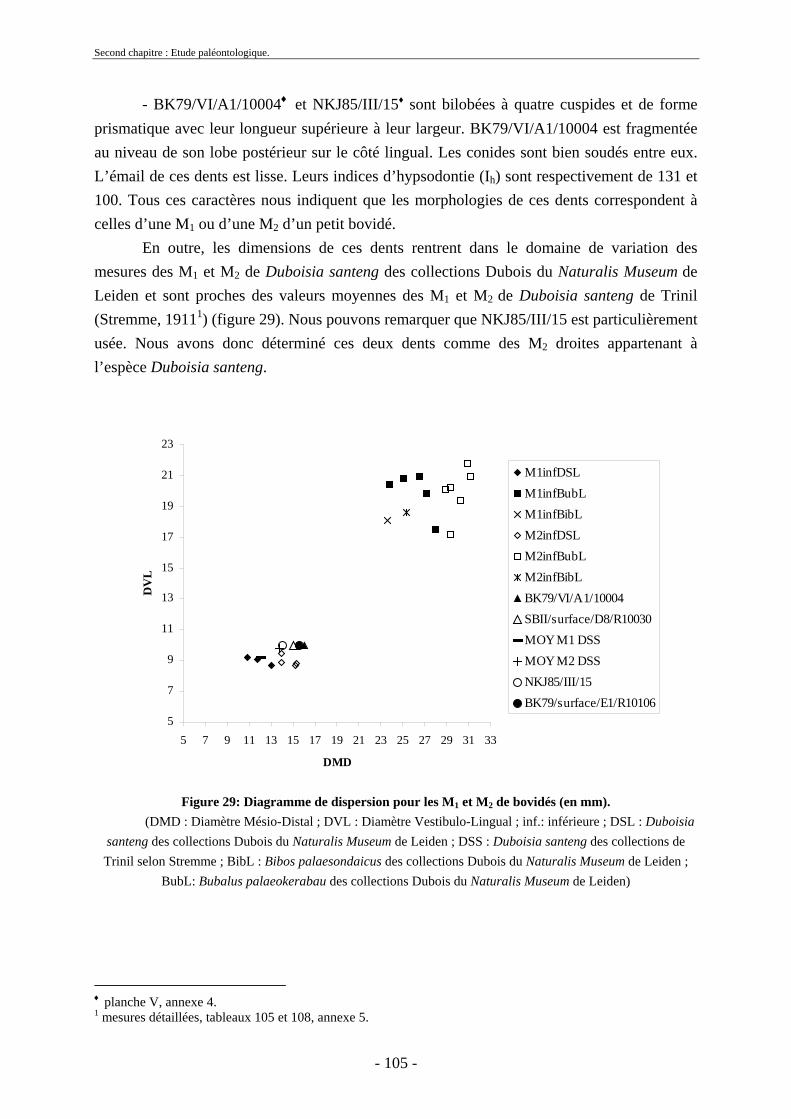
Second chapitre : Etude paléontologique.
- 105 -
- BK79/VI/A1/10004♦ et NKJ85/III/15♦ sont bilobées à quatre cuspides et de forme prismatique avec leur longueur supérieure à leur largeur. BK79/VI/A1/10004 est fragmentée au niveau de son lobe postérieur sur le côté lingual. Les conides sont bien soudés entre eux. L’émail de ces dents est lisse. Leurs indices d’hypsodontie (Ih) sont respectivement de 131 et 100. Tous ces caractères nous indiquent que les morphologies de ces dents correspondent à celles d’une M1 ou d’une M2 d’un petit bovidé.
En outre, les dimensions de ces dents rentrent dans le domaine de variation des mesures des M1 et M2 de Duboisia santeng des collections Dubois du Naturalis Museum de Leiden et sont proches des valeurs moyennes des M1 et M2 de Duboisia santeng de Trinil (Stremme, 19111) (figure 29). Nous pouvons remarquer que NKJ85/III/15 est particulièrement usée. Nous avons donc déterminé ces deux dents comme des M2 droites appartenant à l’espèce Duboisia santeng.
5
7
9
11
13
15
17
19
21
23
5 7 9 11 13 15 17 19 21 23 25 27 29 31 33
DMD
DV
L
M1infDSLM1infBubLM1infBibLM2infDSLM2infBubLM2infBibLBK79/VI/A1/10004SBII/surface/D8/R10030MOY M1 DSSMOY M2 DSSNKJ85/III/15BK79/surface/E1/R10106
Figure 29: Diagramme de dispersion pour les M1 et M2 de bovidés (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual ; inf.: inférieure ; DSL : Duboisia santeng des collections Dubois du Naturalis Museum de Leiden ; DSS : Duboisia santeng des collections de Trinil selon Stremme ; BibL : Bibos palaesondaicus des collections Dubois du Naturalis Museum de Leiden ;
BubL: Bubalus palaeokerabau des collections Dubois du Naturalis Museum de Leiden)
♦ planche V, annexe 4. 1 mesures détaillées, tableaux 105 et 108, annexe 5.

Second chapitre : Etude paléontologique.
- 106 -
- TDJ82/IV/58♦ et TDJ82/IV/138♦ sont des molaires fragmentées. Leurs conides sont bien soudés et leur émail est lisse. La valeur de leurs indices d’hypsodontie Ih est la même, elle est de 142. Sur TDJ82/IV/138 nous notons la présence du lobe distal très réduit et de forme cylindrique caractéristique des M3. Pour TDJ82/IV/58 nous ne pouvons pas déterminer avec certitude la nature de cette dent jugale inférieure gauche car elle est bilobée et le deuxième lobe est fragmenté.
En outre, étant donné leur fragmentation, nous n’avons pas pu comparer leurs mesures avec des dents trouvées sur d’autres sites mais, d’après leur morphologie et leur proportion, nous les considérons comme appartenant à Duboisia santeng. Leur taille est proportionnelle à celle des molaires de Duboisia santeng des collections Dubois du Naturalis Museum de Leiden1. TDJ82/IV/138 a donc été déterminée comme une M3 gauche fragmentée de Duboisia santeng et TDJ82/IV/58 comme un fragment de dent jugale inférieure gauche de Duboisia santeng. Collections de surface :
Au niveau des collections de surface, des dents de Duboisia santeng ont été observées dans les collections de Sendang Busik (SBII/surface/D8/R10030 et SBII/surface/D8/R10031) et de Bukuran (BK79/surface/E1/R10106 et BK79/surface/C1/R10350).
- SBII/surface/D8/R10030♦ et BK79/surface/E1/R10106♦ ont été déterminées comme
des M2 droites de Duboisia santeng. En effet, leurs indices d’hypsodontie Ih sont respectivement de 140 et 116 et elles présentent la même morphologie que BK79/II/A1/10004 et NKJ85/III/15. Elles rentrent également dans le domaine de variation des mesures des M1 et M2 de Duboisia santeng des collections Dubois du Naturalis Museum de Leiden et des valeurs moyennes des M1 et M2 de Duboisia santeng de Trinil (Stremme, 19111) (figure 29). Leurs dimensions sont plus proches de celles des M2.
- SBII/surface/D8/R10031 est fragmentée et nous n’avons donc pas pu comparer ses
mesures, mais sa morphologie et ses proportions sont très proches des M1 et M2 de Duboisia santeng des collections de Dubois du Naturalis Museum de Leiden1. Nous avons donc déterminé cette dent comme un fragment de M1ou2 droite de Duboisia santeng.
BK79/surface/C1/R10350♦ est un fragment de dent jugale inférieure de Duboisia santeng.
♦ planche V, annexe 4. 1 mesures détaillées, tableaux 105 et 108, annexe 5.
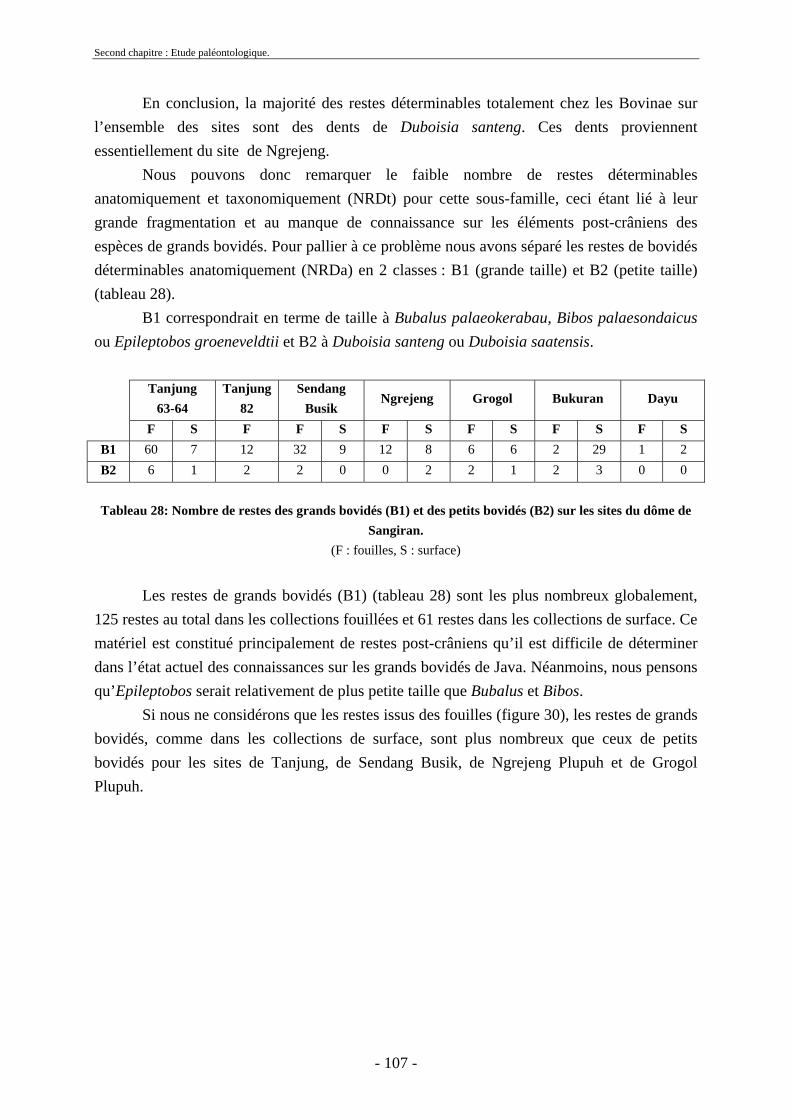
Second chapitre : Etude paléontologique.
- 107 -
En conclusion, la majorité des restes déterminables totalement chez les Bovinae sur l’ensemble des sites sont des dents de Duboisia santeng. Ces dents proviennent essentiellement du site de Ngrejeng.
Nous pouvons donc remarquer le faible nombre de restes déterminables anatomiquement et taxonomiquement (NRDt) pour cette sous-famille, ceci étant lié à leur grande fragmentation et au manque de connaissance sur les éléments post-crâniens des espèces de grands bovidés. Pour pallier à ce problème nous avons séparé les restes de bovidés déterminables anatomiquement (NRDa) en 2 classes : B1 (grande taille) et B2 (petite taille) (tableau 28).
B1 correspondrait en terme de taille à Bubalus palaeokerabau, Bibos palaesondaicus ou Epileptobos groeneveldtii et B2 à Duboisia santeng ou Duboisia saatensis. Tanjung
63-64 Tanjung
82 Sendang
Busik Ngrejeng Grogol Bukuran Dayu
F S F F S F S F S F S F S B1 60 7 12 32 9 12 8 6 6 2 29 1 2 B2 6 1 2 2 0 0 2 2 1 2 3 0 0
Tableau 28: Nombre de restes des grands bovidés (B1) et des petits bovidés (B2) sur les sites du dôme de
Sangiran. (F : fouilles, S : surface)
Les restes de grands bovidés (B1) (tableau 28) sont les plus nombreux globalement,
125 restes au total dans les collections fouillées et 61 restes dans les collections de surface. Ce matériel est constitué principalement de restes post-crâniens qu’il est difficile de déterminer dans l’état actuel des connaissances sur les grands bovidés de Java. Néanmoins, nous pensons qu’Epileptobos serait relativement de plus petite taille que Bubalus et Bibos.
Si nous ne considérons que les restes issus des fouilles (figure 30), les restes de grands bovidés, comme dans les collections de surface, sont plus nombreux que ceux de petits bovidés pour les sites de Tanjung, de Sendang Busik, de Ngrejeng Plupuh et de Grogol Plupuh.
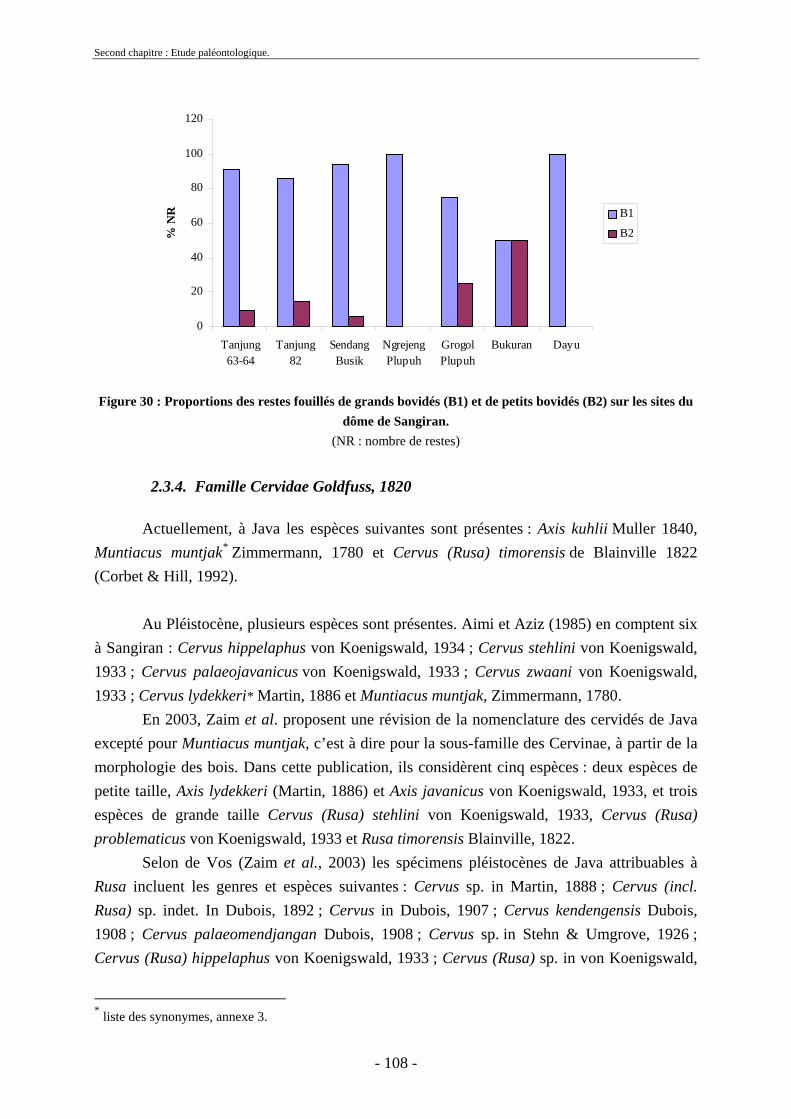
Second chapitre : Etude paléontologique.
- 108 -
0
20
40
60
80
100
120
Tanjung63-64
Tanjung82
SendangBusik
NgrejengPlupuh
GrogolPlupuh
Bukuran Dayu
% N
R B1
B2
Figure 30 : Proportions des restes fouillés de grands bovidés (B1) et de petits bovidés (B2) sur les sites du
dôme de Sangiran. (NR : nombre de restes)
2.3.4. Famille Cervidae Goldfuss, 1820
Actuellement, à Java les espèces suivantes sont présentes : Axis kuhlii Muller 1840,
Muntiacus muntjak* Zimmermann, 1780 et Cervus (Rusa) timorensis de Blainville 1822 (Corbet & Hill, 1992). Au Pléistocène, plusieurs espèces sont présentes. Aimi et Aziz (1985) en comptent six à Sangiran : Cervus hippelaphus von Koenigswald, 1934 ; Cervus stehlini von Koenigswald, 1933 ; Cervus palaeojavanicus von Koenigswald, 1933 ; Cervus zwaani von Koenigswald, 1933 ; Cervus lydekkeri* Martin, 1886 et Muntiacus muntjak, Zimmermann, 1780.
En 2003, Zaim et al. proposent une révision de la nomenclature des cervidés de Java excepté pour Muntiacus muntjak, c’est à dire pour la sous-famille des Cervinae, à partir de la morphologie des bois. Dans cette publication, ils considèrent cinq espèces : deux espèces de petite taille, Axis lydekkeri (Martin, 1886) et Axis javanicus von Koenigswald, 1933, et trois espèces de grande taille Cervus (Rusa) stehlini von Koenigswald, 1933, Cervus (Rusa) problematicus von Koenigswald, 1933 et Rusa timorensis Blainville, 1822.
Selon de Vos (Zaim et al., 2003) les spécimens pléistocènes de Java attribuables à Rusa incluent les genres et espèces suivantes : Cervus sp. in Martin, 1888 ; Cervus (incl. Rusa) sp. indet. In Dubois, 1892 ; Cervus in Dubois, 1907 ; Cervus kendengensis Dubois, 1908 ; Cervus palaeomendjangan Dubois, 1908 ; Cervus sp. in Stehn & Umgrove, 1926 ; Cervus (Rusa) hippelaphus von Koenigswald, 1933 ; Cervus (Rusa) sp. in von Koenigswald,
* liste des synonymes, annexe 3.

Second chapitre : Etude paléontologique.
- 109 -
1933 ; Cervus sp. in von Koenigswald, 1933 ; Cervus (Rusa) oppenoorthi von Koenigswald, 1933 et Cervus hippelaphus von Koenigswald, 1934.
Cette famille est très importante pour tous les sites du dôme de Sangiran car une
grande partie des restes fouillés appartiennent à ce groupe et plus particulièrement à la sous-famille des Cervinae.
2.3.4.1. Sous-famille Cervinae Goldfuss, 1820
Aimi et Aziz (1985), lors de leur étude sur le dôme de Sangiran, ont mis en évidence
de nombreuses espèces de cervidés. À Bukuran, dans les tranchées effectuées dans les couches de Pucangan, ils ont découvert des restes de Cervus hippelaphus, de Cervus zwaani, de Cervus lydekkeri et de Cervus sp., ces mêmes espèces ont été retrouvées lors des ramassages de surface sur ce même site. À Tanjung, Sendang Busik et Dayu, entre autres, ils ont pu également mettre en évidence la présence de Cervus lydekkeri, et celle de Cervus hippelaphus à Sendang Busik. Nous retiendrons que ce qu’ils qualifient comme l’espèce Cervus lydekkeri est maintenant considérée comme Axis lydekkeri, de même l’espèce qu’ils nomment Cervus hippelaphus, est aujourd’hui nommée Cervus (Rusa) sp. Von Koenigswald (1940) avait noté la présence de Cervus zwaani et C. problematicus dans la formation de Pucangan et la présence d’Axis lydekkeri dans la formation de Kabuh (Grenzbank).
Sur le site de Ngebung 2, la population de cervidés décrite dans le niveau principal (niveau A) est assez complète et homogène pour décrire un stade évolutif de l’espèce Axis lydekkeri : Axis lydekkeri ngebungensis. D’autre part, des restes de Cervus (Rusa) sp. sont présents dans les niveaux supérieurs (B et U) (Moigne et al., 2004).
Dans notre matériel, nous avons pu mettre en évidence la présence de deux espèces de
Cervinae : Axis lydekkeri* (Martin, 1886) et Cervus (Rusa) sp.* in von Koenigswald, 1933. La diagnose des cervidés est basée principalement sur l’étude des bois, lorsqu’ils
étaient absents nous avons utilisé les dents. Pour les diagnoses des dents de cervidés nous avons choisi la nomenclature de Heintz (1970). Cette nomenclature dentaire est celle de la théorie trituberculaire de Cope (1874) et d’Osborn (1888 et 1904), modifiée et complétée par les travaux de Scott (1892), Gregory (1916), Simpson (1925) et Schwartz (1937).
* liste des synonymes, annexe 3.

Second chapitre : Etude paléontologique.
- 110 -
Axis lydekkeri (Martin, 1886) et Axis sp. Des fossiles du petit cervidé Axis sp. ont été découverts lors des fouilles sur tous les
sites excepté à Dayu. Ces restes sont principalement des dents et des bois. À Tanjung, 7 bois, 3 dents et 2 scapulas ont été mis au jour, à Sendang Busik, 2 bois, à Bukuran, un bois et une scapula, à Ngrejeng Plupuh, une scapula et enfin à Grogol Plupuh un bois et trois dents.
Les bois d’Axis lydekkeri ont une forme de lyre. Le bois entier est composé d’une base
se poursuivant par un premier andouiller qui fait un angle d’environ 90° avec le merrain et ensuite un andouiller central, court, rentrant vers l’intérieur et faisant un angle avec le merrain d’également environ 90°. La deuxième partie du merrain est longue. La surface du bois d’Axis est assez lisse (Zaim et al., 2003) Néanmoins, sur la plupart des restes, nous avons pu observer une légère ornementation caractérisée par de fines rainures. Ici, nous avons illustré nos propos par un dessin provenant de l’ouvrage de von Koenigswald (1933). Ces bois sont ceux d’un Axis axis indien actuel qui permettent de donner une idée de la forme des bois d’un Axis lydekkeri javanais (figure 31).
Figure 31 : Bois d’Axis axis indien (1/5 nat.)
(von Koenigswald, 1933) Bois :
Parmi les bois, trois ont pu être attribués à l’espèce Axis lydekkeri : TDJ64/G11F11/13B10B♦, TDJ64/I4/930♦ et BK79/III/B1/1♦.
TDJ64/G11F11/13B10B est un bois de chute gauche où l’andouiller basilaire est fragmenté et où l’épois est fragmenté également. TDJ64/I4/930 correspondrait à un bois de chute où la rose est fragmentée et où la partie supérieure est absente c’est à dire l’andouiller central et l’épois. BK79/III/B1/1 est une partie supérieure de bois, c’est à dire l’andouiller central et l’épois. Les dimensions de ces bois rentrent dans le domaine de variation des bois
♦ planche VI, annexe 4.

Second chapitre : Etude paléontologique.
- 111 -
d’Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden et d’Axis sp. du Muséum de Sangiran (tableau 29).
Pour les autres bois de nos collections, nous n’avons pas pu déterminer l’espèce (Axis lydekkeri ou Axis javanicus) et nous sommes restés au niveau du genre Axis, compte tenu de la fragmentation et des mesures comparées à celles des collections de Dubois du Naturalis Museum de Leiden (tableau 29) et du Musée de Sangiran.
Cinq bois ont été retrouvés à Tanjung, deux à Sendang Busik et un à Grogol Plupuh : - TDJ64/G1/575 : bois de chute, partie basale (rose et base) avec la première
partie du merrain au-dessus de l’andouiller basilaire, - TDJ64/C2/1043 et TDJ64/D12/286 : bois de chute, partie basale (rose et
base) avec l’andouiller basilaire, - TDJ64/F6/228 et TDJ64/C8/1076 : fragment de frontal avec la base du bois
(pivot, rose et base), - SBII/6/14 : partie basale du bois (rose et base) avec l’andouiller basilaire
cassé, - SBII/9/20 : fragment de frontal avec la base du bois (pivot, rose et base), - GP83/II/A3/4 : partie basale du bois (rose et base) avec l’andouiller basilaire
cassé.
Nous remarquons que les dimensions de TDJ64/G1/575 sont inférieures à celles des autres bois (tableau 29). Cette différence est-elle due à une différence d’âge ou à une spéciation différente ? Ceci est difficile à savoir compte tenu des multiples variations des bois. Des variations peuvent exister à l’intérieur d’une même espèce, en fonction de l’âge, en fonction des saisons, en fonction du milieu ou en fonction d’anomalies (Heintz, 1970).
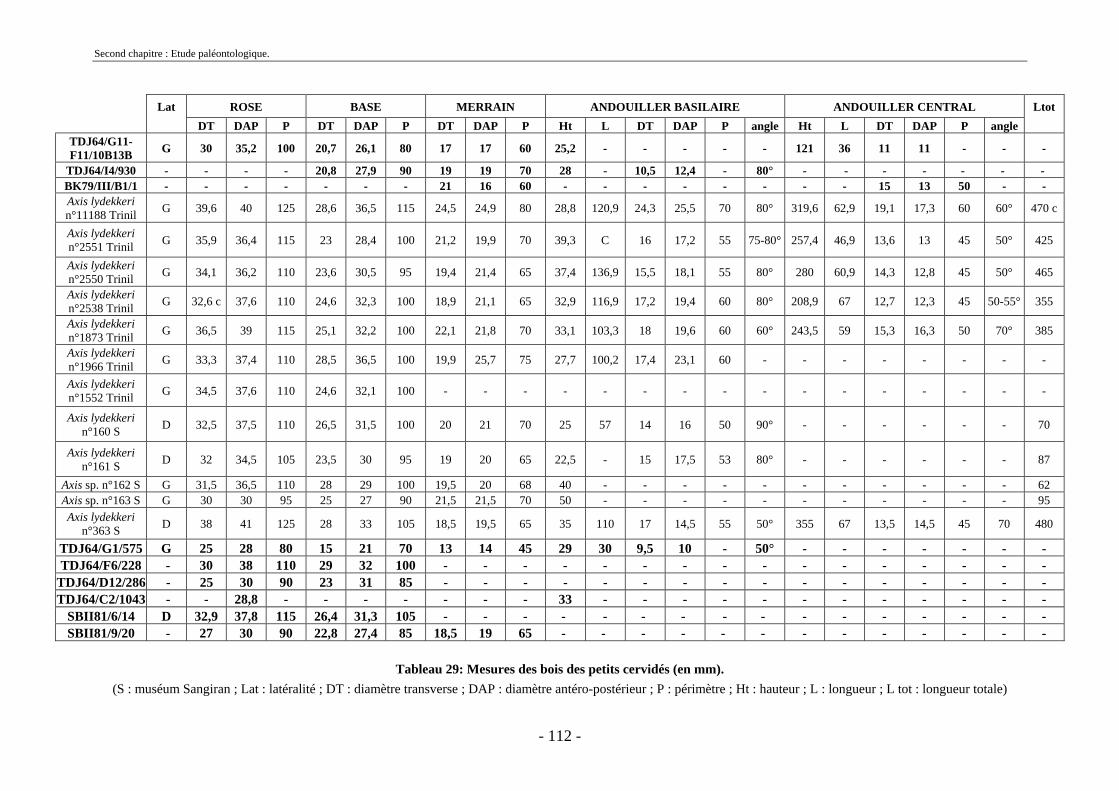
Second chapitre : Etude paléontologique.
- 112 -
ROSE BASE MERRAIN ANDOUILLER BASILAIRE ANDOUILLER CENTRAL
Lat DT DAP P DT DAP P DT DAP P Ht L DT DAP P angle Ht L DT DAP P angle
Ltot
TDJ64/G11-F11/10B13B G 30 35,2 100 20,7 26,1 80 17 17 60 25,2 - - - - - 121 36 11 11 - - -
TDJ64/I4/930 - - - - 20,8 27,9 90 19 19 70 28 - 10,5 12,4 - 80° - - - - - - - BK79/III/B1/1 - - - - - - - 21 16 60 - - - - - - - - 15 13 50 - - Axis lydekkeri n°11188 Trinil G 39,6 40 125 28,6 36,5 115 24,5 24,9 80 28,8 120,9 24,3 25,5 70 80° 319,6 62,9 19,1 17,3 60 60° 470 c
Axis lydekkeri n°2551 Trinil G 35,9 36,4 115 23 28,4 100 21,2 19,9 70 39,3 C 16 17,2 55 75-80° 257,4 46,9 13,6 13 45 50° 425
Axis lydekkeri n°2550 Trinil G 34,1 36,2 110 23,6 30,5 95 19,4 21,4 65 37,4 136,9 15,5 18,1 55 80° 280 60,9 14,3 12,8 45 50° 465
Axis lydekkeri n°2538 Trinil G 32,6 c 37,6 110 24,6 32,3 100 18,9 21,1 65 32,9 116,9 17,2 19,4 60 80° 208,9 67 12,7 12,3 45 50-55° 355
Axis lydekkeri n°1873 Trinil G 36,5 39 115 25,1 32,2 100 22,1 21,8 70 33,1 103,3 18 19,6 60 60° 243,5 59 15,3 16,3 50 70° 385
Axis lydekkeri n°1966 Trinil G 33,3 37,4 110 28,5 36,5 100 19,9 25,7 75 27,7 100,2 17,4 23,1 60 - - - - - - - -
Axis lydekkeri n°1552 Trinil G 34,5 37,6 110 24,6 32,1 100 - - - - - - - - - - - - - - - -
Axis lydekkeri n°160 S D 32,5 37,5 110 26,5 31,5 100 20 21 70 25 57 14 16 50 90° - - - - - - 70
Axis lydekkeri n°161 S D 32 34,5 105 23,5 30 95 19 20 65 22,5 - 15 17,5 53 80° - - - - - - 87
Axis sp. n°162 S G 31,5 36,5 110 28 29 100 19,5 20 68 40 - - - - - - - - - - - 62 Axis sp. n°163 S G 30 30 95 25 27 90 21,5 21,5 70 50 - - - - - - - - - - - 95 Axis lydekkeri
n°363 S D 38 41 125 28 33 105 18,5 19,5 65 35 110 17 14,5 55 50° 355 67 13,5 14,5 45 70 480
TDJ64/G1/575 G 25 28 80 15 21 70 13 14 45 29 30 9,5 10 - 50° - - - - - - - TDJ64/F6/228 - 30 38 110 29 32 100 - - - - - - - - - - - - - - - -
TDJ64/D12/286 - 25 30 90 23 31 85 - - - - - - - - - - - - - - - - TDJ64/C2/1043 - - 28,8 - - - - - - - 33 - - - - - - - - - - - -
SBII81/6/14 D 32,9 37,8 115 26,4 31,3 105 - - - - - - - - - - - - - - - - SBII81/9/20 - 27 30 90 22,8 27,4 85 18,5 19 65 - - - - - - - - - - - - -
Tableau 29: Mesures des bois des petits cervidés (en mm).
(S : muséum Sangiran ; Lat : latéralité ; DT : diamètre transverse ; DAP : diamètre antéro-postérieur ; P : périmètre ; Ht : hauteur ; L : longueur ; L tot : longueur totale)

Second chapitre : Etude paléontologique.
- 113 -
Dents supérieures :
- GP83/II/b2/74♦ est bilobée à quatre cuspides et sa longueur est proche de sa largeur. Ses cônes sont indépendants et nous avons observé le pli protoconal caractéristique des molaires supérieures de cervidé. Son émail est ridé. Cette dent présente des styles développés notamment le parastyle et le mésostyle. Son indice d’hypsodontie (Heintz, 1970) est inférieur à 100 ce qui confirme son appartenance à la famille des cervidés :
- Ih GP83/II/b2/74 = 92. Les deux cuspides de cette dent ne présentent pas la même forme, la cuspide
antérieure présente une forme plus carrée que la postérieure ce qui est caractéristique des M1. De plus, cette dent rentre dans le domaine de variation des M1 et M2 d’Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden et d’Axis nssp. de Ngebung 2 (figure 31) et est un peu plus éloignée des moyennes des mesures des M1 et M2 de Trinil (Stremme, 19111). Ces différences sont peut-être dues à la méthode de prise de mesures. Ses dimensions sont plus proches de celles des M1 d’Axis lydekkeri. GP83/II/b2/74 a donc été déterminée comme une M1 droite. - TDJ82/IV/91♦ et GP83/II/A1/11♦ sont bilobées à quatre cuspides. Leur longueur est proche de leur largeur. Les cônes de ces deux dents sont peu soudés. L’indice i (Heintz, 1970) de ces dents sont de 119 et de 113, ce qui est caractéristique des molaires supérieures de cervidés car ils sont supérieurs à 100. Leur émail est ridé. Toutes ces données nous indiquent que ces dents sont des M1ou2 de cervidé. Les deux cuspides de ces dents sont de même forme et de même taille ce qui confirme une morphologie de M2.
Nous avons comparé les dimensions de ces dents avec celles des M1ou2 d’Axis lydekkeri et de Cervus kendengensis des collections de Dubois du Naturalis Museum de Leiden et celles d’Axis nssp. de Ngebung 2 1 (figure 32). Nous observons que leurs dimensions sont plus proches de celles des M2 d’Axis lydekkeri.
Nous avons donc considéré TDJ82/IV/91 comme une M2 gauche et GP83/II/A1/11 comme une M2 gauche appartenant au genre Axis.
♦ planche VII, annexe 4. 1 mesures détaillées, tableaux 109, 111, 113 et 116, annexe 5.
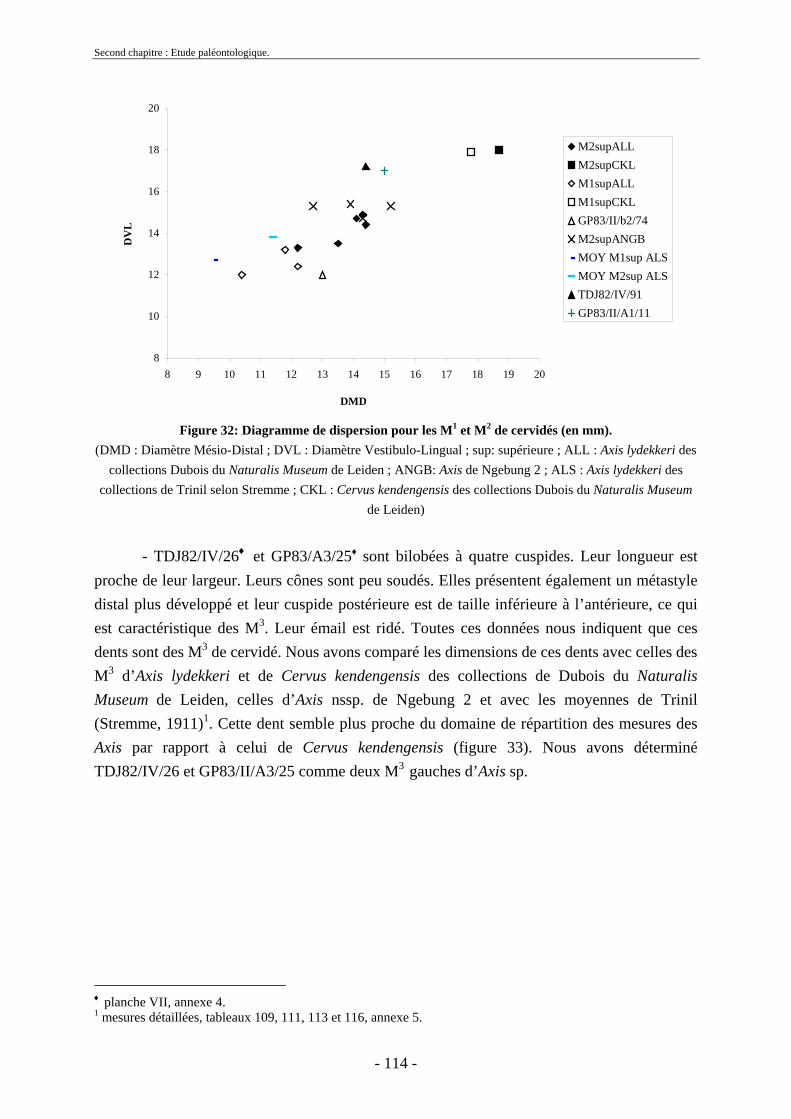
Second chapitre : Etude paléontologique.
- 114 -
8
10
12
14
16
18
20
8 9 10 11 12 13 14 15 16 17 18 19 20
DMD
DV
L
M2supALLM2supCKLM1supALLM1supCKLGP83/II/b2/74M2supANGBMOY M1sup ALSMOY M2sup ALSTDJ82/IV/91GP83/II/A1/11
Figure 32: Diagramme de dispersion pour les M1 et M2 de cervidés (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual ; sup: supérieure ; ALL : Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden ; ANGB: Axis de Ngebung 2 ; ALS : Axis lydekkeri des
collections de Trinil selon Stremme ; CKL : Cervus kendengensis des collections Dubois du Naturalis Museum de Leiden)
- TDJ82/IV/26♦ et GP83/A3/25♦ sont bilobées à quatre cuspides. Leur longueur est
proche de leur largeur. Leurs cônes sont peu soudés. Elles présentent également un métastyle distal plus développé et leur cuspide postérieure est de taille inférieure à l’antérieure, ce qui est caractéristique des M3. Leur émail est ridé. Toutes ces données nous indiquent que ces dents sont des M3 de cervidé. Nous avons comparé les dimensions de ces dents avec celles des M3 d’Axis lydekkeri et de Cervus kendengensis des collections de Dubois du Naturalis Museum de Leiden, celles d’Axis nssp. de Ngebung 2 et avec les moyennes de Trinil (Stremme, 1911)1. Cette dent semble plus proche du domaine de répartition des mesures des Axis par rapport à celui de Cervus kendengensis (figure 33). Nous avons déterminé TDJ82/IV/26 et GP83/II/A3/25 comme deux M3 gauches d’Axis sp.
♦ planche VII, annexe 4. 1 mesures détaillées, tableaux 109, 111, 113 et 116, annexe 5.
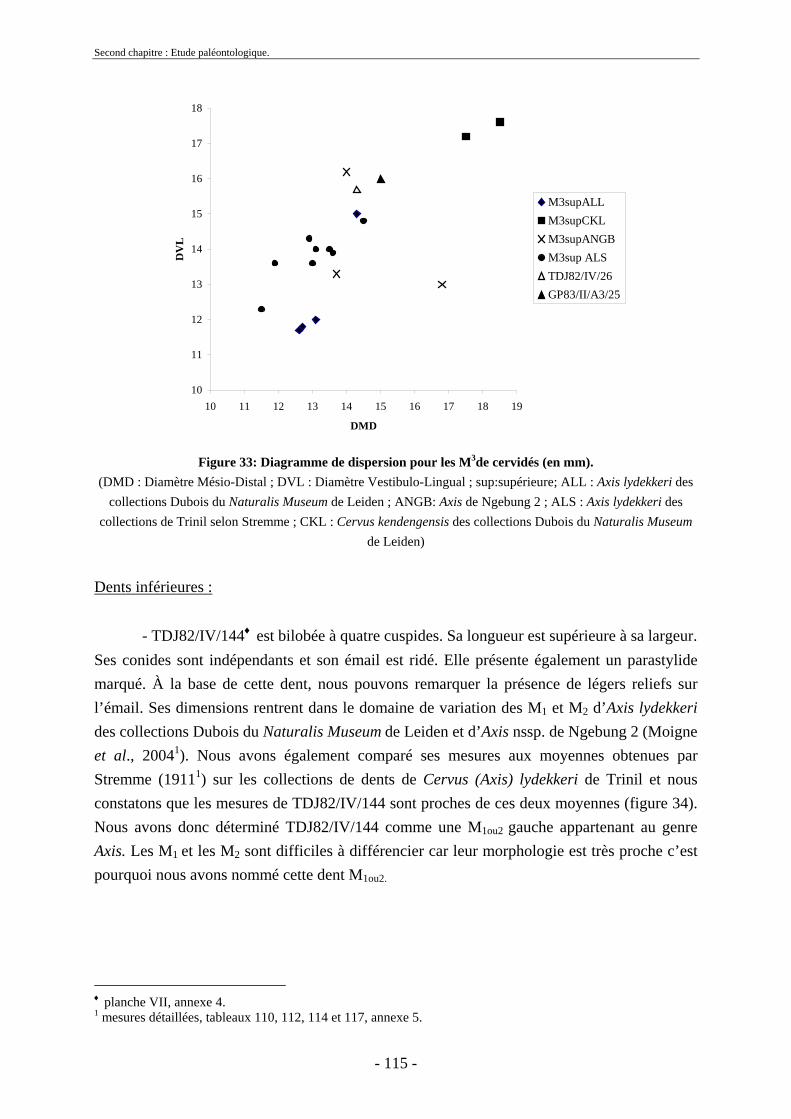
Second chapitre : Etude paléontologique.
- 115 -
10
11
12
13
14
15
16
17
18
10 11 12 13 14 15 16 17 18 19
DMD
DV
L
M3supALLM3supCKLM3supANGBM3sup ALSTDJ82/IV/26GP83/II/A3/25
Figure 33: Diagramme de dispersion pour les M3de cervidés (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual ; sup:supérieure; ALL : Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden ; ANGB: Axis de Ngebung 2 ; ALS : Axis lydekkeri des
collections de Trinil selon Stremme ; CKL : Cervus kendengensis des collections Dubois du Naturalis Museum de Leiden)
Dents inférieures :
- TDJ82/IV/144♦ est bilobée à quatre cuspides. Sa longueur est supérieure à sa largeur. Ses conides sont indépendants et son émail est ridé. Elle présente également un parastylide marqué. À la base de cette dent, nous pouvons remarquer la présence de légers reliefs sur l’émail. Ses dimensions rentrent dans le domaine de variation des M1 et M2 d’Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden et d’Axis nssp. de Ngebung 2 (Moigne et al., 20041). Nous avons également comparé ses mesures aux moyennes obtenues par Stremme (19111) sur les collections de dents de Cervus (Axis) lydekkeri de Trinil et nous constatons que les mesures de TDJ82/IV/144 sont proches de ces deux moyennes (figure 34). Nous avons donc déterminé TDJ82/IV/144 comme une M1ou2 gauche appartenant au genre Axis. Les M1 et les M2 sont difficiles à différencier car leur morphologie est très proche c’est pourquoi nous avons nommé cette dent M1ou2.
♦ planche VII, annexe 4. 1 mesures détaillées, tableaux 110, 112, 114 et 117, annexe 5.
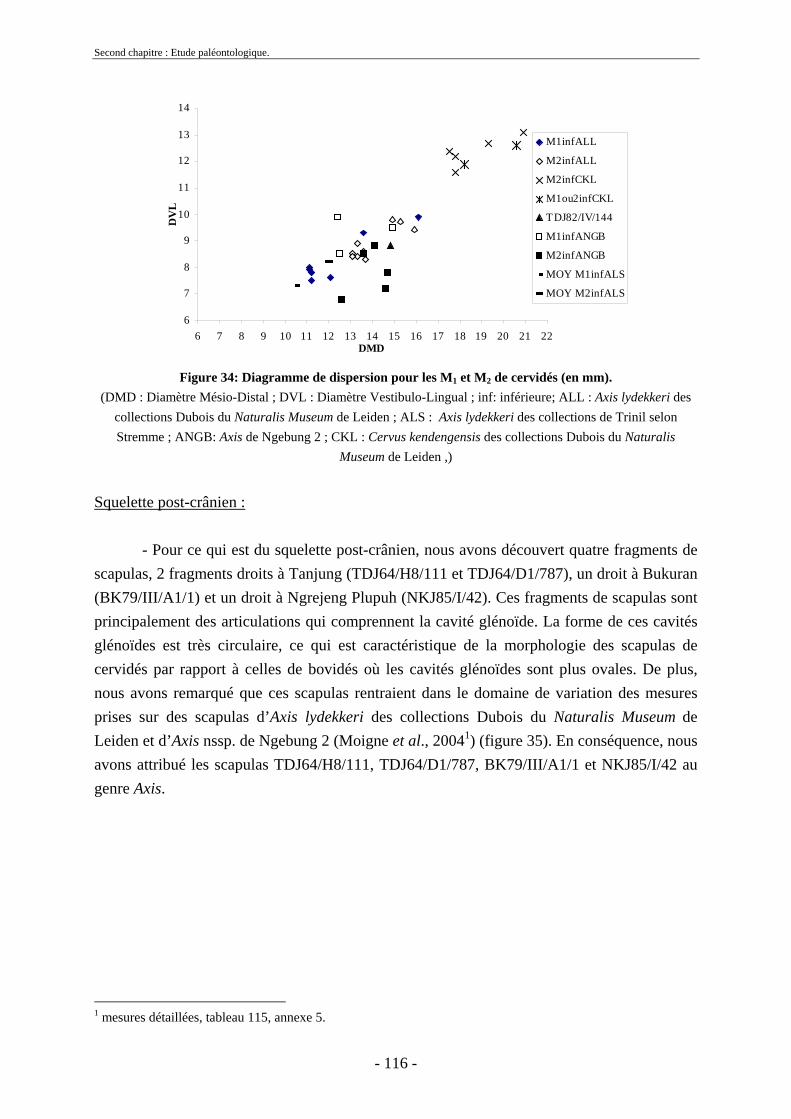
Second chapitre : Etude paléontologique.
- 116 -
6
7
8
9
10
11
12
13
14
6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22DMD
DV
L
M1infALL
M2infALL
M2infCKL
M1ou2infCKL
TDJ82/IV/144
M1infANGB
M2infANGB
MOY M1infALS
MOY M2infALS
Figure 34: Diagramme de dispersion pour les M1 et M2 de cervidés (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual ; inf: inférieure; ALL : Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden ; ALS : Axis lydekkeri des collections de Trinil selon Stremme ; ANGB: Axis de Ngebung 2 ; CKL : Cervus kendengensis des collections Dubois du Naturalis
Museum de Leiden ,)
Squelette post-crânien :
- Pour ce qui est du squelette post-crânien, nous avons découvert quatre fragments de scapulas, 2 fragments droits à Tanjung (TDJ64/H8/111 et TDJ64/D1/787), un droit à Bukuran (BK79/III/A1/1) et un droit à Ngrejeng Plupuh (NKJ85/I/42). Ces fragments de scapulas sont principalement des articulations qui comprennent la cavité glénoïde. La forme de ces cavités glénoïdes est très circulaire, ce qui est caractéristique de la morphologie des scapulas de cervidés par rapport à celles de bovidés où les cavités glénoïdes sont plus ovales. De plus, nous avons remarqué que ces scapulas rentraient dans le domaine de variation des mesures prises sur des scapulas d’Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden et d’Axis nssp. de Ngebung 2 (Moigne et al., 20041) (figure 35). En conséquence, nous avons attribué les scapulas TDJ64/H8/111, TDJ64/D1/787, BK79/III/A1/1 et NKJ85/I/42 au genre Axis.
1 mesures détaillées, tableau 115, annexe 5.
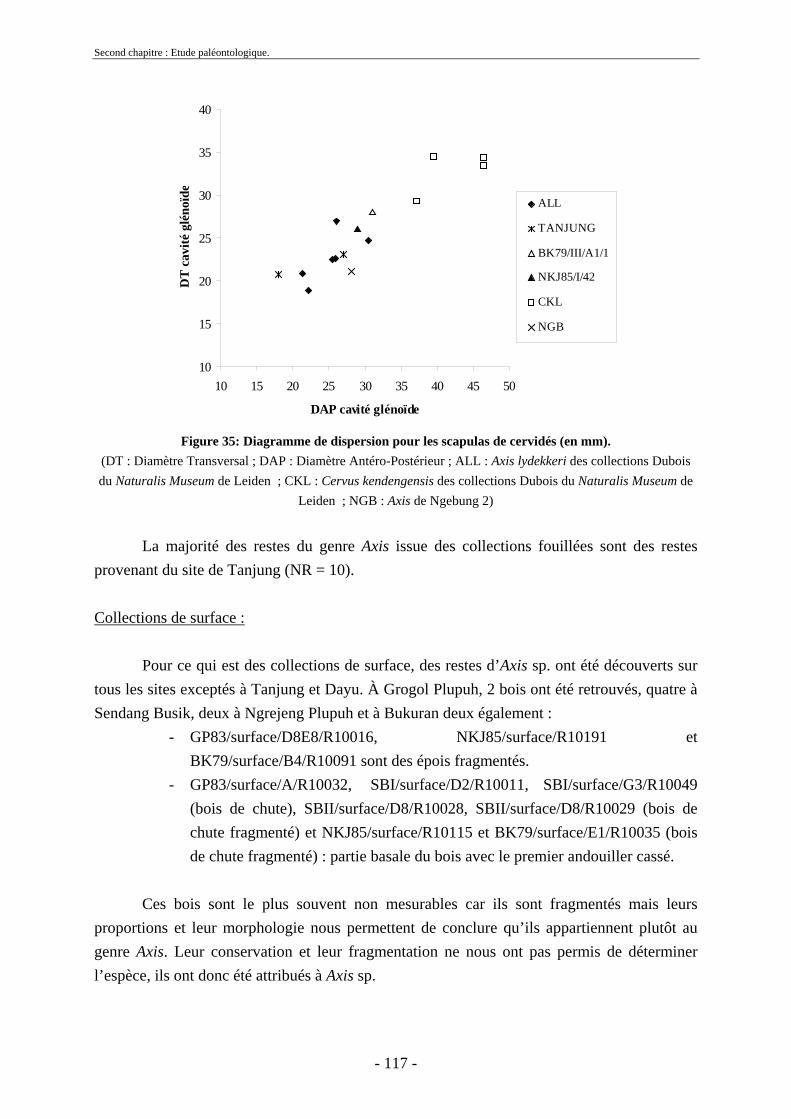
Second chapitre : Etude paléontologique.
- 117 -
10
15
20
25
30
35
40
10 15 20 25 30 35 40 45 50
DAP cavité glénoïde
DT
cav
ité g
léno
ïde
ALL
TANJUNG
BK79/III/A1/1
NKJ85/I/42
CKL
NGB
Figure 35: Diagramme de dispersion pour les scapulas de cervidés (en mm).
(DT : Diamètre Transversal ; DAP : Diamètre Antéro-Postérieur ; ALL : Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden ; CKL : Cervus kendengensis des collections Dubois du Naturalis Museum de
Leiden ; NGB : Axis de Ngebung 2)
La majorité des restes du genre Axis issue des collections fouillées sont des restes
provenant du site de Tanjung (NR = 10). Collections de surface :
Pour ce qui est des collections de surface, des restes d’Axis sp. ont été découverts sur tous les sites exceptés à Tanjung et Dayu. À Grogol Plupuh, 2 bois ont été retrouvés, quatre à Sendang Busik, deux à Ngrejeng Plupuh et à Bukuran deux également :
- GP83/surface/D8E8/R10016, NKJ85/surface/R10191 et BK79/surface/B4/R10091 sont des épois fragmentés.
- GP83/surface/A/R10032, SBI/surface/D2/R10011, SBI/surface/G3/R10049 (bois de chute), SBII/surface/D8/R10028, SBII/surface/D8/R10029 (bois de chute fragmenté) et NKJ85/surface/R10115 et BK79/surface/E1/R10035 (bois de chute fragmenté) : partie basale du bois avec le premier andouiller cassé.
Ces bois sont le plus souvent non mesurables car ils sont fragmentés mais leurs proportions et leur morphologie nous permettent de conclure qu’ils appartiennent plutôt au genre Axis. Leur conservation et leur fragmentation ne nous ont pas permis de déterminer l’espèce, ils ont donc été attribués à Axis sp.
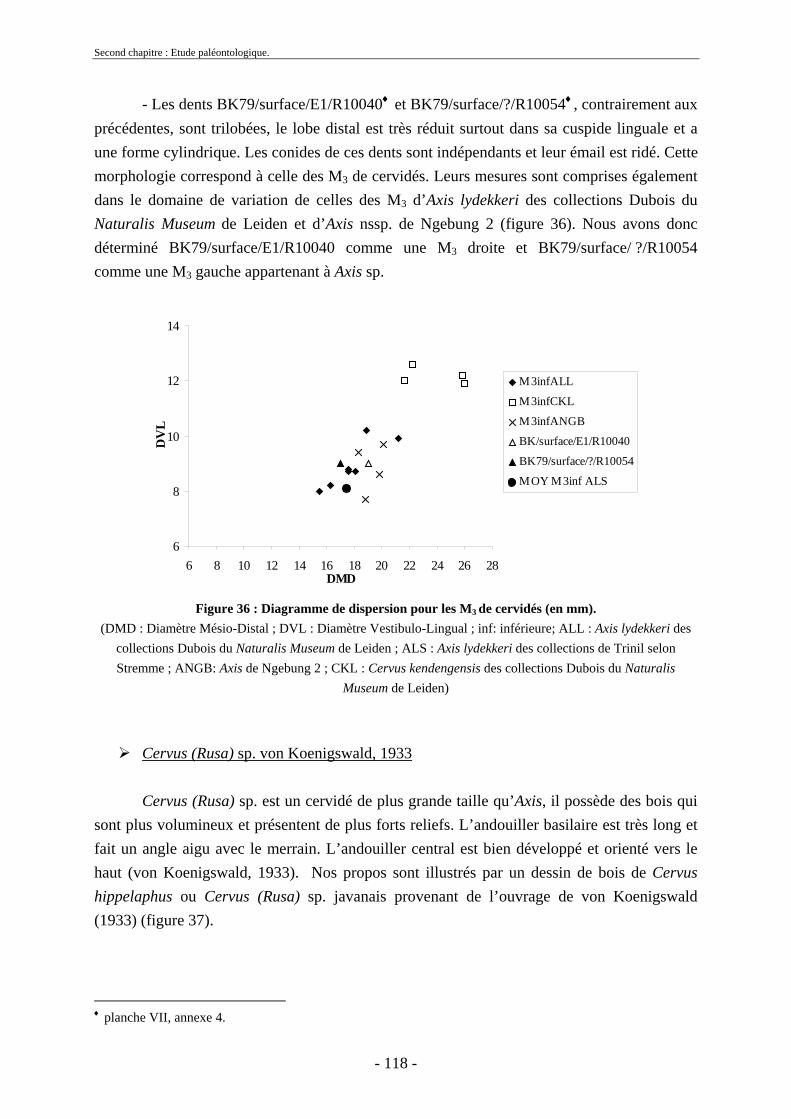
Second chapitre : Etude paléontologique.
- 118 -
- Les dents BK79/surface/E1/R10040♦ et BK79/surface/?/R10054♦, contrairement aux précédentes, sont trilobées, le lobe distal est très réduit surtout dans sa cuspide linguale et a une forme cylindrique. Les conides de ces dents sont indépendants et leur émail est ridé. Cette morphologie correspond à celle des M3 de cervidés. Leurs mesures sont comprises également dans le domaine de variation de celles des M3 d’Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden et d’Axis nssp. de Ngebung 2 (figure 36). Nous avons donc déterminé BK79/surface/E1/R10040 comme une M3 droite et BK79/surface/ ?/R10054 comme une M3 gauche appartenant à Axis sp.
6
8
10
12
14
6 8 10 12 14 16 18 20 22 24 26 28DMD
DV
L
M3infALL
M3infCKL
M3infANGB
BK/surface/E1/R10040
BK79/surface/?/R10054
MOY M3inf ALS
Figure 36 : Diagramme de dispersion pour les M3 de cervidés (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual ; inf: inférieure; ALL : Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden ; ALS : Axis lydekkeri des collections de Trinil selon Stremme ; ANGB: Axis de Ngebung 2 ; CKL : Cervus kendengensis des collections Dubois du Naturalis
Museum de Leiden)
Cervus (Rusa) sp. von Koenigswald, 1933
Cervus (Rusa) sp. est un cervidé de plus grande taille qu’Axis, il possède des bois qui sont plus volumineux et présentent de plus forts reliefs. L’andouiller basilaire est très long et fait un angle aigu avec le merrain. L’andouiller central est bien développé et orienté vers le haut (von Koenigswald, 1933). Nos propos sont illustrés par un dessin de bois de Cervus hippelaphus ou Cervus (Rusa) sp. javanais provenant de l’ouvrage de von Koenigswald (1933) (figure 37).
♦ planche VII, annexe 4.

Second chapitre : Etude paléontologique.
- 119 -
Figure 37 : Bois d’un Cervus hippelaphus javanais. (1/10 nat.)
(von Koenigswald, 1933) Des restes de Cervus (Rusa) sp., espèce de grande taille, ont été découverts dans les
collections fouillées de Ngrejeng et Sendang Busik. Ce grand cerf est représenté par sept dents NG85/III/18, NKJ85/I/43, NKJ85/I/11, NKJ85/II/12, NKJ85/II/18, NKJ85/III/11 et SBII/11/47.
Dents supérieures :
- NG85/III/18♦ présente un seul cône lingual et deux renflements sur le côté labial. Ce schéma est proche de celui des molaires mais semble avoir subi une compression antéro-postérieure, morphologie caractéristique des prémolaires. D’autre part, nous pouvons observer que le style antéro-labial ou parastyle est très prononcé sur cette dent. Ce type de morphologie est typique des P4 où l’importance du lobe antérieur prédomine. Les cônes linguaux et vestibulaires paraissent peu soudés et l’émail est ridé. Tous ces caractères permettent de rapprocher la morphologie de cette dent de celle d’une P4 de cervidé.
De plus, son DMD est de 9 mm et son DVL de 12 mm, ces dimensions sont supérieures aux moyennes calculées à partir des mesures de Stremme (19111) pour Axis lydekkeri (DMD moyen = 6,7 mm et DVL moyen = 10 mm). Cette dent semble donc d’une taille nettement supérieure à celle des P4 des Axis lydekkeri des collections de Selenka, nous l’avons donc considérée comme une P4 droite de Cervus (Rusa) sp.
- NKJ85/I/43♦ est de forme carrée, elle est bilobée et possède quatre cuspides. Son
indice i est de 112 et son Ih de 69. Ses cônes sont peu soudés et son émail est ridé. Au niveau de l’hypocône, on peut remarquer la présence d’un éperon. Les styles sont bien marqués sur la face vestibulaire. Toutes ces données permettent de rapprocher la morphologie de cette dent
♦ planche VIII, annexe 4. 1 mesures détaillées, tableaux 109 et 116, annexe 5.
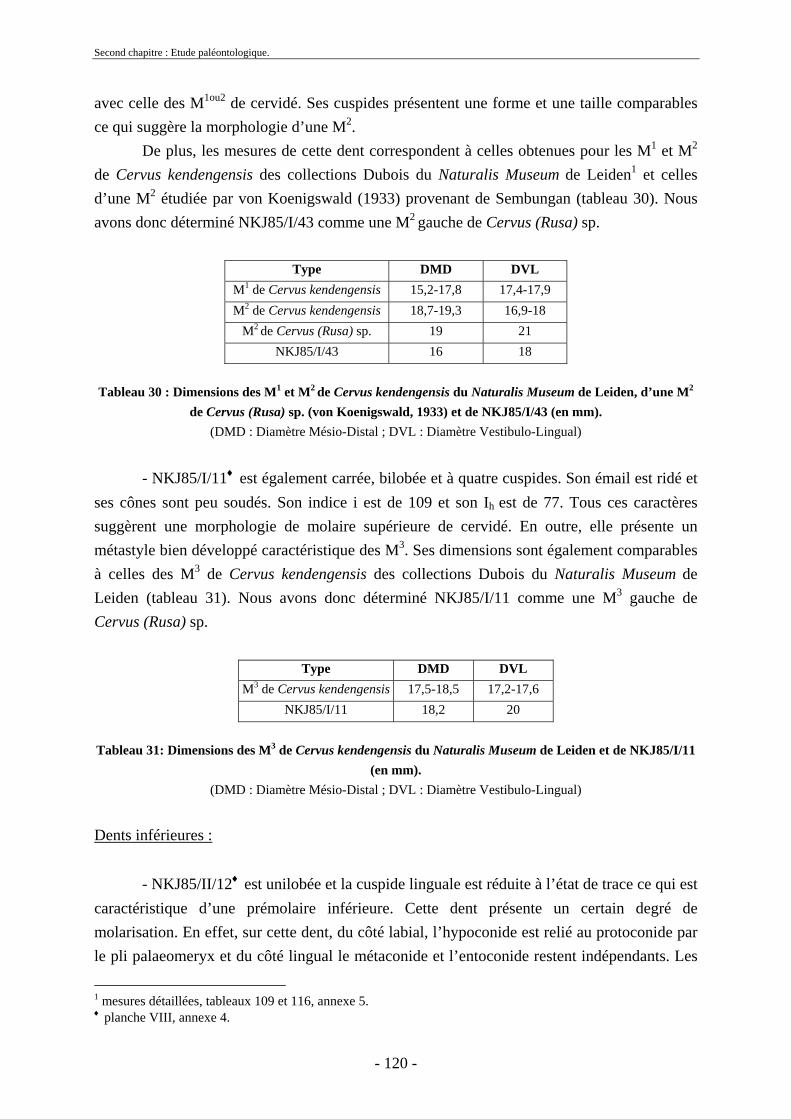
Second chapitre : Etude paléontologique.
- 120 -
avec celle des M1ou2 de cervidé. Ses cuspides présentent une forme et une taille comparables ce qui suggère la morphologie d’une M2.
De plus, les mesures de cette dent correspondent à celles obtenues pour les M1 et M2 de Cervus kendengensis des collections Dubois du Naturalis Museum de Leiden1 et celles d’une M2 étudiée par von Koenigswald (1933) provenant de Sembungan (tableau 30). Nous avons donc déterminé NKJ85/I/43 comme une M2 gauche de Cervus (Rusa) sp.
Type DMD DVL
M1 de Cervus kendengensis 15,2-17,8 17,4-17,9 M2 de Cervus kendengensis 18,7-19,3 16,9-18
M2 de Cervus (Rusa) sp. 19 21 NKJ85/I/43 16 18
Tableau 30 : Dimensions des M1 et M2 de Cervus kendengensis du Naturalis Museum de Leiden, d’une M2
de Cervus (Rusa) sp. (von Koenigswald, 1933) et de NKJ85/I/43 (en mm). (DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
- NKJ85/I/11♦ est également carrée, bilobée et à quatre cuspides. Son émail est ridé et
ses cônes sont peu soudés. Son indice i est de 109 et son Ih est de 77. Tous ces caractères suggèrent une morphologie de molaire supérieure de cervidé. En outre, elle présente un métastyle bien développé caractéristique des M3. Ses dimensions sont également comparables à celles des M3 de Cervus kendengensis des collections Dubois du Naturalis Museum de Leiden (tableau 31). Nous avons donc déterminé NKJ85/I/11 comme une M3 gauche de Cervus (Rusa) sp.
Type DMD DVL M3 de Cervus kendengensis 17,5-18,5 17,2-17,6
NKJ85/I/11 18,2 20
Tableau 31: Dimensions des M3 de Cervus kendengensis du Naturalis Museum de Leiden et de NKJ85/I/11 (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
Dents inférieures :
- NKJ85/II/12♦ est unilobée et la cuspide linguale est réduite à l’état de trace ce qui est caractéristique d’une prémolaire inférieure. Cette dent présente un certain degré de molarisation. En effet, sur cette dent, du côté labial, l’hypoconide est relié au protoconide par le pli palaeomeryx et du côté lingual le métaconide et l’entoconide restent indépendants. Les
1 mesures détaillées, tableaux 109 et 116, annexe 5. ♦ planche VIII, annexe 4.
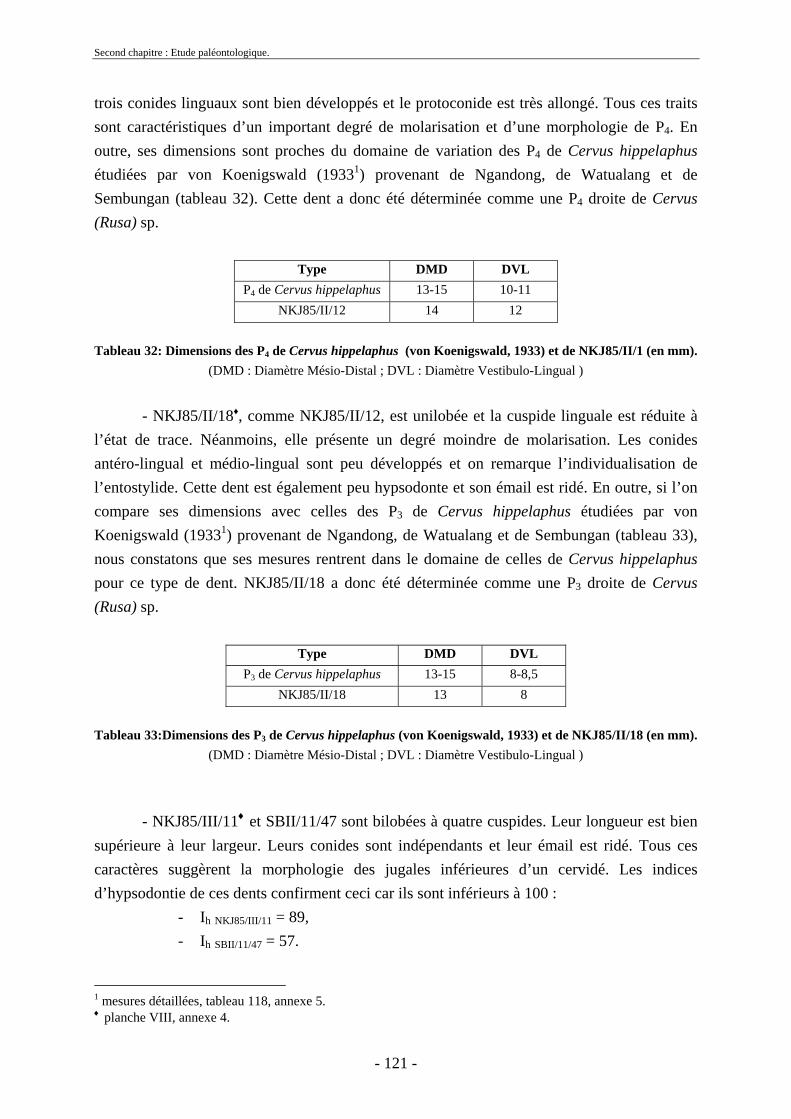
Second chapitre : Etude paléontologique.
- 121 -
trois conides linguaux sont bien développés et le protoconide est très allongé. Tous ces traits sont caractéristiques d’un important degré de molarisation et d’une morphologie de P4. En outre, ses dimensions sont proches du domaine de variation des P4 de Cervus hippelaphus étudiées par von Koenigswald (19331) provenant de Ngandong, de Watualang et de Sembungan (tableau 32). Cette dent a donc été déterminée comme une P4 droite de Cervus (Rusa) sp.
Type DMD DVL
P4 de Cervus hippelaphus 13-15 10-11 NKJ85/II/12 14 12
Tableau 32: Dimensions des P4 de Cervus hippelaphus (von Koenigswald, 1933) et de NKJ85/II/1 (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual )
- NKJ85/II/18♦, comme NKJ85/II/12, est unilobée et la cuspide linguale est réduite à
l’état de trace. Néanmoins, elle présente un degré moindre de molarisation. Les conides antéro-lingual et médio-lingual sont peu développés et on remarque l’individualisation de l’entostylide. Cette dent est également peu hypsodonte et son émail est ridé. En outre, si l’on compare ses dimensions avec celles des P3 de Cervus hippelaphus étudiées par von Koenigswald (19331) provenant de Ngandong, de Watualang et de Sembungan (tableau 33), nous constatons que ses mesures rentrent dans le domaine de celles de Cervus hippelaphus pour ce type de dent. NKJ85/II/18 a donc été déterminée comme une P3 droite de Cervus (Rusa) sp.
Type DMD DVL P3 de Cervus hippelaphus 13-15 8-8,5
NKJ85/II/18 13 8
Tableau 33:Dimensions des P3 de Cervus hippelaphus (von Koenigswald, 1933) et de NKJ85/II/18 (en mm). (DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual )
- NKJ85/III/11♦ et SBII/11/47 sont bilobées à quatre cuspides. Leur longueur est bien supérieure à leur largeur. Leurs conides sont indépendants et leur émail est ridé. Tous ces caractères suggèrent la morphologie des jugales inférieures d’un cervidé. Les indices d’hypsodontie de ces dents confirment ceci car ils sont inférieurs à 100 :
- Ih NKJ85/III/11 = 89, - Ih SBII/11/47 = 57.
1 mesures détaillées, tableau 118, annexe 5. ♦ planche VIII, annexe 4.
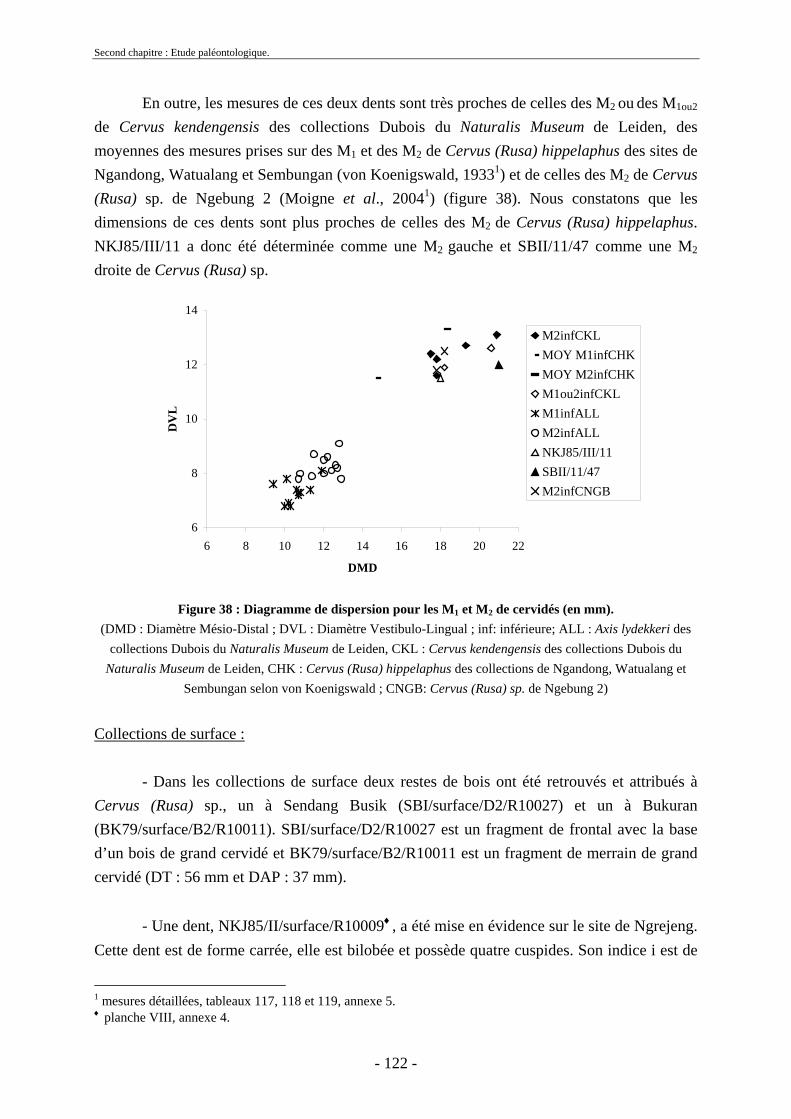
Second chapitre : Etude paléontologique.
- 122 -
En outre, les mesures de ces deux dents sont très proches de celles des M2 ou des M1ou2 de Cervus kendengensis des collections Dubois du Naturalis Museum de Leiden, des moyennes des mesures prises sur des M1 et des M2 de Cervus (Rusa) hippelaphus des sites de Ngandong, Watualang et Sembungan (von Koenigswald, 19331) et de celles des M2 de Cervus (Rusa) sp. de Ngebung 2 (Moigne et al., 20041) (figure 38). Nous constatons que les dimensions de ces dents sont plus proches de celles des M2 de Cervus (Rusa) hippelaphus. NKJ85/III/11 a donc été déterminée comme une M2 gauche et SBII/11/47 comme une M2
droite de Cervus (Rusa) sp.
6
8
10
12
14
6 8 10 12 14 16 18 20 22
DMD
DV
L
M2infCKLMOY M1infCHKMOY M2infCHKM1ou2infCKLM1infALLM2infALLNKJ85/III/11SBII/11/47M2infCNGB
Figure 38 : Diagramme de dispersion pour les M1 et M2 de cervidés (en mm).
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual ; inf: inférieure; ALL : Axis lydekkeri des collections Dubois du Naturalis Museum de Leiden, CKL : Cervus kendengensis des collections Dubois du
Naturalis Museum de Leiden, CHK : Cervus (Rusa) hippelaphus des collections de Ngandong, Watualang et Sembungan selon von Koenigswald ; CNGB: Cervus (Rusa) sp. de Ngebung 2)
Collections de surface : - Dans les collections de surface deux restes de bois ont été retrouvés et attribués à Cervus (Rusa) sp., un à Sendang Busik (SBI/surface/D2/R10027) et un à Bukuran (BK79/surface/B2/R10011). SBI/surface/D2/R10027 est un fragment de frontal avec la base d’un bois de grand cervidé et BK79/surface/B2/R10011 est un fragment de merrain de grand cervidé (DT : 56 mm et DAP : 37 mm). - Une dent, NKJ85/II/surface/R10009♦, a été mise en évidence sur le site de Ngrejeng. Cette dent est de forme carrée, elle est bilobée et possède quatre cuspides. Son indice i est de
1 mesures détaillées, tableaux 117, 118 et 119, annexe 5. ♦ planche VIII, annexe 4.
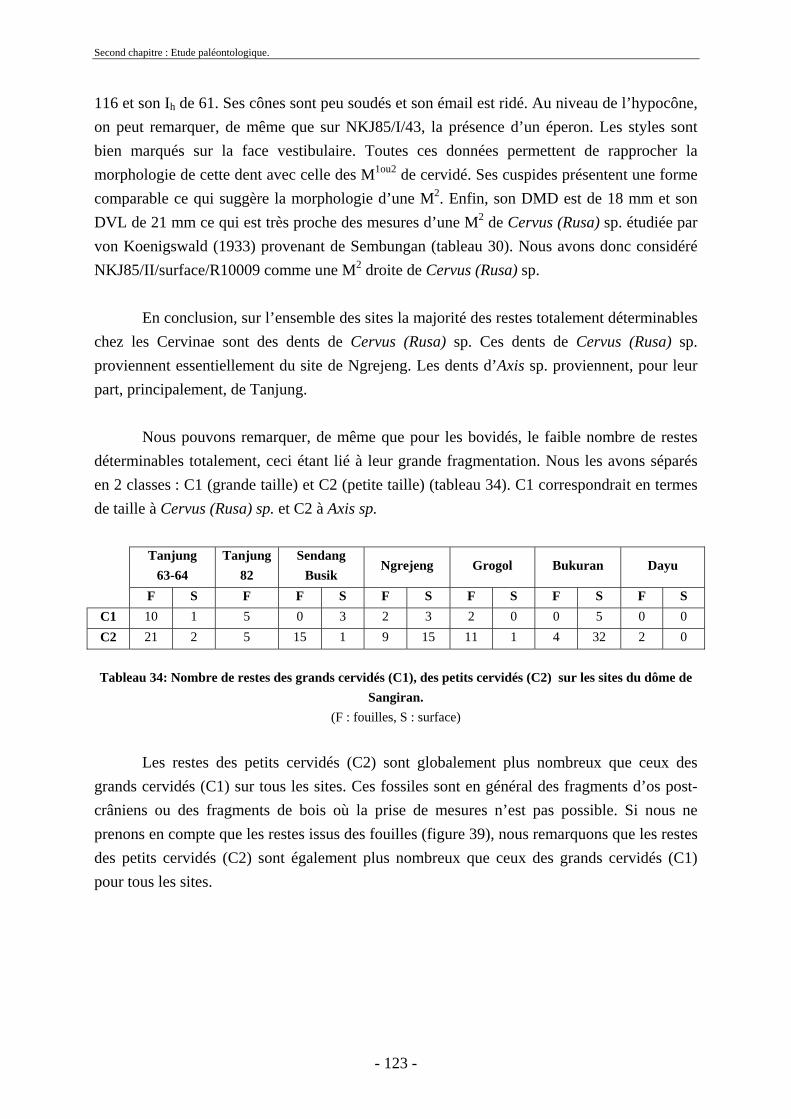
Second chapitre : Etude paléontologique.
- 123 -
116 et son Ih de 61. Ses cônes sont peu soudés et son émail est ridé. Au niveau de l’hypocône, on peut remarquer, de même que sur NKJ85/I/43, la présence d’un éperon. Les styles sont bien marqués sur la face vestibulaire. Toutes ces données permettent de rapprocher la morphologie de cette dent avec celle des M1ou2 de cervidé. Ses cuspides présentent une forme comparable ce qui suggère la morphologie d’une M2. Enfin, son DMD est de 18 mm et son DVL de 21 mm ce qui est très proche des mesures d’une M2 de Cervus (Rusa) sp. étudiée par von Koenigswald (1933) provenant de Sembungan (tableau 30). Nous avons donc considéré NKJ85/II/surface/R10009 comme une M2 droite de Cervus (Rusa) sp.
En conclusion, sur l’ensemble des sites la majorité des restes totalement déterminables
chez les Cervinae sont des dents de Cervus (Rusa) sp. Ces dents de Cervus (Rusa) sp. proviennent essentiellement du site de Ngrejeng. Les dents d’Axis sp. proviennent, pour leur part, principalement, de Tanjung.
Nous pouvons remarquer, de même que pour les bovidés, le faible nombre de restes déterminables totalement, ceci étant lié à leur grande fragmentation. Nous les avons séparés en 2 classes : C1 (grande taille) et C2 (petite taille) (tableau 34). C1 correspondrait en termes de taille à Cervus (Rusa) sp. et C2 à Axis sp.
Tanjung
63-64 Tanjung
82 Sendang
Busik Ngrejeng Grogol Bukuran Dayu
F S F F S F S F S F S F S C1 10 1 5 0 3 2 3 2 0 0 5 0 0 C2 21 2 5 15 1 9 15 11 1 4 32 2 0
Tableau 34: Nombre de restes des grands cervidés (C1), des petits cervidés (C2) sur les sites du dôme de
Sangiran. (F : fouilles, S : surface)
Les restes des petits cervidés (C2) sont globalement plus nombreux que ceux des grands cervidés (C1) sur tous les sites. Ces fossiles sont en général des fragments d’os post-crâniens ou des fragments de bois où la prise de mesures n’est pas possible. Si nous ne prenons en compte que les restes issus des fouilles (figure 39), nous remarquons que les restes des petits cervidés (C2) sont également plus nombreux que ceux des grands cervidés (C1) pour tous les sites.
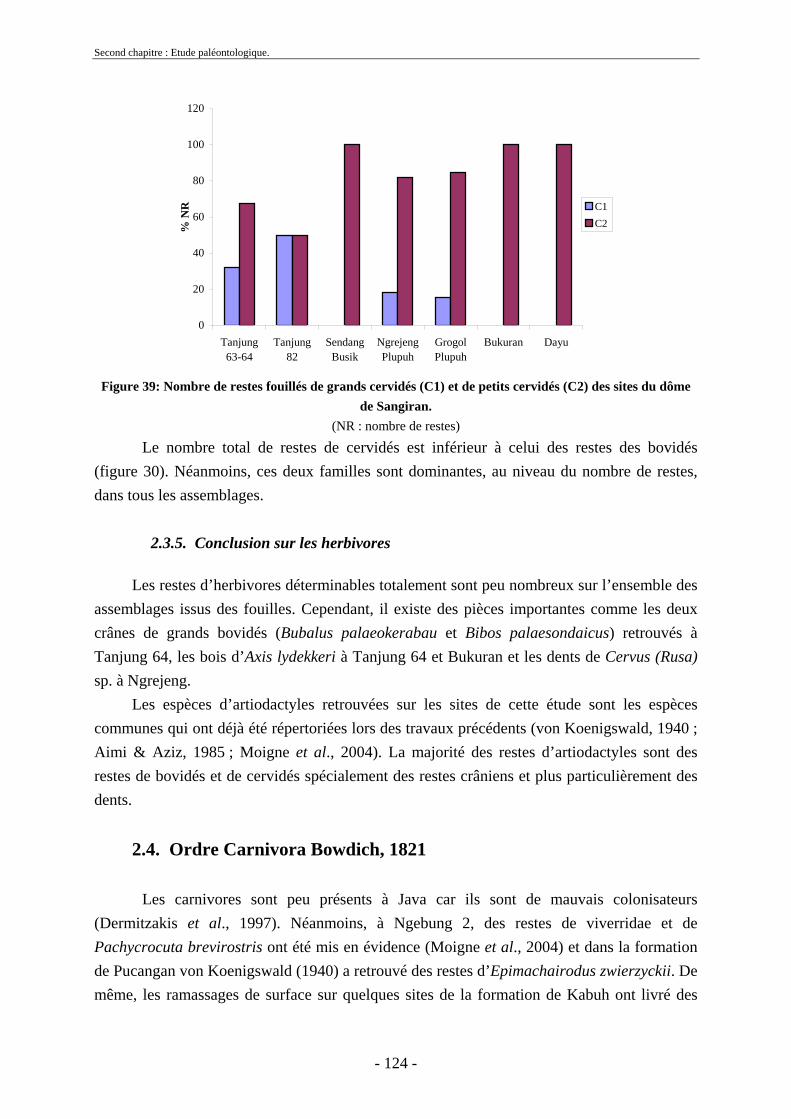
Second chapitre : Etude paléontologique.
- 124 -
0
20
40
60
80
100
120
Tanjung63-64
Tanjung82
SendangBusik
NgrejengPlupuh
GrogolPlupuh
Bukuran Dayu
% N
R C1C2
Figure 39: Nombre de restes fouillés de grands cervidés (C1) et de petits cervidés (C2) des sites du dôme
de Sangiran. (NR : nombre de restes)
Le nombre total de restes de cervidés est inférieur à celui des restes des bovidés (figure 30). Néanmoins, ces deux familles sont dominantes, au niveau du nombre de restes, dans tous les assemblages.
2.3.5. Conclusion sur les herbivores
Les restes d’herbivores déterminables totalement sont peu nombreux sur l’ensemble des assemblages issus des fouilles. Cependant, il existe des pièces importantes comme les deux crânes de grands bovidés (Bubalus palaeokerabau et Bibos palaesondaicus) retrouvés à Tanjung 64, les bois d’Axis lydekkeri à Tanjung 64 et Bukuran et les dents de Cervus (Rusa) sp. à Ngrejeng.
Les espèces d’artiodactyles retrouvées sur les sites de cette étude sont les espèces communes qui ont déjà été répertoriées lors des travaux précédents (von Koenigswald, 1940 ; Aimi & Aziz, 1985 ; Moigne et al., 2004). La majorité des restes d’artiodactyles sont des restes de bovidés et de cervidés spécialement des restes crâniens et plus particulièrement des dents.
2.4. Ordre Carnivora Bowdich, 1821
Les carnivores sont peu présents à Java car ils sont de mauvais colonisateurs (Dermitzakis et al., 1997). Néanmoins, à Ngebung 2, des restes de viverridae et de Pachycrocuta brevirostris ont été mis en évidence (Moigne et al., 2004) et dans la formation de Pucangan von Koenigswald (1940) a retrouvé des restes d’Epimachairodus zwierzyckii. De même, les ramassages de surface sur quelques sites de la formation de Kabuh ont livré des

Second chapitre : Etude paléontologique.
- 125 -
restes de Cuon javanicus trinilensis, Crocuta crocuta sinensis, Panthera tigris, P. pardus et d’ Homotherium zwierzyckii (Aimi & Aziz, 1985).
Très peu de restes de carnivores ont été découverts sur les sites de notre étude parmi
les restes déterminables.
2.4.1. Famille Hyaenidae Gray, 1821
Actuellement, cette famille a disparu de Java et quasiment de toute l’Asie, sauf en Inde (Pautret-Homerville, 2001). Au Pléistocène moyen, l’espèce présente à Java était Pachycrocuta brevirostris* Aymard, 1856 (Geraads, 1979b). Cette hyène était robuste et approchait la taille d’un lion actuel (Turner & Anton, 1996).
Dans le matériel fouillé un fragment de dent a été découvert à Bukuran
(BK79/VI/A1/9). Ce fragment de dent est un protoconide peu large de prémolaire inférieure droit attribué à Pachycrocuta brevirostris (annexe 4, planche IX).
2.5. Ordre Primates
Trachypithecus sp. a été retrouvé à Ngebung 2 (Moigne et al., 2004) mais nous n’avons pas constaté sa présence sur les sites de notre étude. Des restes de macaque ont pu être mis en évidence sur de rares sites dans la formation de Kabuh, en surface (Aimi & Aziz, 1985).
Sur tous les sites de cette étude, exceptés à Ngrejeng Grogol et Dayu, des restes d’Homo erectus ou Pithecanthropus ont été découverts par des villageois avant les fouilles. Ces restes sont des fragments de crâne :
- Tanjung (1963) : S6, une calotte crânienne composée des deux pariétaux, du frontal, de l’occipital, des deux temporaux, du sphénoïde et de l’os zygomatique gauche (Jacob 1964 ; Sartono, 1964a),
- Sendang Busik (1979): S37a, un fragment crânien composé d’une partie postérieure du pariétal gauche, du mastoïde et des os de l’occipital proche de l’astérion gauche ; S37b, un fragment crânien composé du pariétal gauche, de la moitié droite de l’occipital supérieur et d’une portion du mastoïde. Ces deux fragments n’ont pas été retrouvés en contact mais ont été considérés comme appartenant au même crâne d’après leur couleur et leur texture (Baba & Aziz, 1991),
- Bukuran : S1b (1936), une mandibule droite et S4 (1938-39) un fragment crânien avec un fragment de maxillaire (von Koenigswald, 1940).
* liste des synonymes, annexe 3.

Second chapitre : Etude paléontologique.
- 126 -
De même à Ngebung, dans les différentes zones fouillées, de nombreux restes de Pithécanthrope ont été retrouvés (von Koenigswald, 1940 ; Sémah et al., 1993 ; Widianto, 2001) (annexe 2).
2.6. Reptiles et poissons
Lors des fouilles d’Aimi et Aziz (1985), la présence des tortues, des crocodiles et des poissons a été observée à Bukuran et dans la plupart de leurs ramassages de surface. A Ngebung 2 (Moigne et al., 2004), des restes de ces animaux ont été mis au jour. Dubois, lors de ces ramassages de surface à Tanjung, avait découvert un humérus fragmenté de crocodile qui est conservé au Naturalis Museum de Leiden.
Ce groupe est bien représenté sur l’ensemble du dôme de Sangiran et sur les sites de notre étude (tableau 35).
Tanjung
63-64 Tanjung
82 Sendang
Busik Ngrejeng Grogol Bukuran Dayu
F S F F S F S F S F S F S Tortues 14 10 8 5 7 3 3 1 3 3 7 1 2
Crocodiles 2 0 4 6 0 2 0 0 0 9 2 0 0 Poissons 0 0 0 0 0 3 1 0 0 1 0 0 0
Gastéropodes 0 0 0 0 0 0 0 0 0 2 0 0 0
Tableau 35: Nombre de restes des reptiles et des poissons sur les sites du dôme de Sangiran. (F : fouilles, S : surface)
D’autre part, si l’on ne considère que les restes fouillés, nous pouvons constater que les fossiles de tortues et de crocodiles sont beaucoup plus nombreux (en pourcentage de nombre de restes) que ceux de poissons et de gastéropodes et que la tortue est majoritaire à Tanjung (dans les deux niveaux), tandis que le crocodile l’est à Bukuran (figure 40).
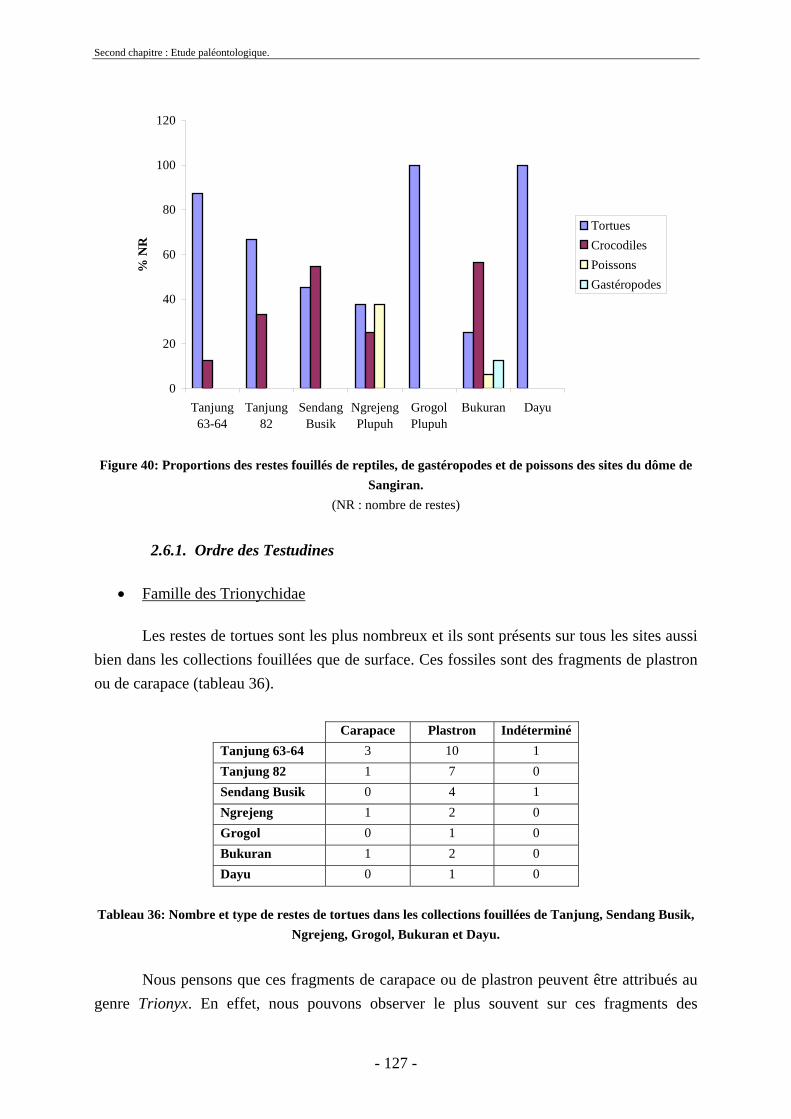
Second chapitre : Etude paléontologique.
- 127 -
0
20
40
60
80
100
120
Tanjung63-64
Tanjung82
SendangBusik
NgrejengPlupuh
GrogolPlupuh
Bukuran Dayu
% N
R
TortuesCrocodilesPoissonsGastéropodes
Figure 40: Proportions des restes fouillés de reptiles, de gastéropodes et de poissons des sites du dôme de
Sangiran. (NR : nombre de restes)
2.6.1. Ordre des Testudines
• Famille des Trionychidae
Les restes de tortues sont les plus nombreux et ils sont présents sur tous les sites aussi
bien dans les collections fouillées que de surface. Ces fossiles sont des fragments de plastron ou de carapace (tableau 36).
Carapace Plastron Indéterminé Tanjung 63-64 3 10 1 Tanjung 82 1 7 0 Sendang Busik 0 4 1 Ngrejeng 1 2 0 Grogol 0 1 0 Bukuran 1 2 0 Dayu 0 1 0
Tableau 36: Nombre et type de restes de tortues dans les collections fouillées de Tanjung, Sendang Busik,
Ngrejeng, Grogol, Bukuran et Dayu.
Nous pensons que ces fragments de carapace ou de plastron peuvent être attribués au
genre Trionyx. En effet, nous pouvons observer le plus souvent sur ces fragments des

Second chapitre : Etude paléontologique.
- 128 -
empreintes creuses, similaires à celles d’une balle de golf ♦, caractéristiques du genre Trionyx (Sobolik & Steele, 1996).
2.6.2. Ordre des Crocodylia
• Famille des Crocodylidae Le crocodile, pour sa part, est présent sur tous les sites hormis à Grogol Plupuh et
Dayu. Les restes attribués au crocodile sont, principalement, des dents isolées♦ sur tous les sites. En outre, un fragment de mandibule a été trouvé sur le site de Ngrejeng (NP85/IV/2♦). Ces restes rendent difficile la détermination générique.
Lors de l’étude des restes de crocodiles de Trinil (Janensch, 1911) deux espèces ont été déterminées : Gavialis bengawanicus et Crocodilus ossifragus.
2.6.3. Poissons
Sur deux sites, à Ngrejeng Plupuh et à Bukuran, nous avons observé des restes de
poissons. Ces restes sont trois dents (NKJ85/II/11♦, NKJ85/II/35 et NKJ85/II/61) pouvant être attribuées à des Chondrichthyens (poisson à squelette cartilagineux) de la sous classe des Eslamobranches et peut-être de l’ordre des squaliformes et de la famille des squalidés.En outre, une vertèbre (BK79/IV/A1/7♦) a été retrouvée à Bukuran, elle serait plutôt attribuée à un Ostéichtyen (poisson osseux).
Des dents de poissons de Trinil (Hennig, 1911) ont été attribuées au genre Prionodon.
2.6.4. Gastéropodes
En outre, à Bukuran, deux opercules de gastéropodes sont présents (BK79/IV/A1/4♦ et BK79/IV/A1/10000). Ces éléments ou plaques permettent de fermer la coquille des gastéropodes pour se protéger des prédateurs (Limondin-Lozouet, 2002). BK79/IV/A1/10000 est un petit fragment d’opercule (L :11 mm, l : 10 mm et e : 1 mm) alors que BK79/IV/A1/4 est quasi complet et présente une forme semi-circulaire avec une apophyse. La longueur de ce dernier est de 19 mm, sa largeur de 11 mm et son épaisseur de 1 mm.
♦ planche IX, annexe 4.

Second chapitre : Etude paléontologique.
- 129 -
2.7. Comparaisons des collections issues des fouilles et de la surface
Nous remarquons des similitudes entre les listes fauniques établies à partir des restes issus des différentes campagnes de fouilles et celles établies à partir des ramassages de surface qui ont eu lieu en même temps que ces fouilles. Ce nombre de restes issus des ramassages de surface n’est pas le même, proportionnellement, suivant les collections (tableau 37). À Tanjung 82, il n’y a pas eu de ramassage de surface.
Pour comparer la diversité taxonomique entre les assemblages issus des fouilles et des ramassages de surface, nous avons calculé un coefficient de similarité. Nous avons choisi d’utiliser le coefficient de communauté de Jaccard (S), qui est un coefficient binaire excluant les zéros et qui donne un poids égal aux différents termes utilisés (Daget, 1979 ; Legendre & Legendre, 1984).
S = a/a+b+c - a : le nombre de descripteurs pour lesquels les deux objets sont codés 1 c’est à dire le
nombre d’espèces en commun, - b et c : le nombre de descripteurs pour lesquels les deux objets sont codés
différemment (0 et 1).
NR fouilles NR surface S Tanjung 63-64 921 85 / Sendang Busik 393 78 0,4 Ngrejeng 245 203 0,4 Grogol 212 68 0,5 Bukuran 144 346 0,43 Dayu 33 26 /
Tableau 37: Nombres de restes (NR) issus des fouilles et des ramassages de surface et coefficient de similarité des collections de Tanjung 63-64, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.
Le calcul du coefficient de similarité pour les collections de Dayu n’a pas été possible
car nous n’avons pas pu déterminer d’espèce et pour Tanjung 63-64, la collection fouillée et de surface ne possédaient pas d’espèces en commun. Pour les autres sites, la valeur du coefficient de similarité est environ la même entre 0,4 et 0,5 ce qui n’est pas très élevé, pour des restes provenant des mêmes sites.
La liste faunique obtenue à Tanjung 63-64 à partir des fossiles trouvés en surface est proche des listes établies (ramassages liés à la fouille, ramassages d’Aimi et Aziz et de Dubois), mises à part les espèces Stegodon trigonocephalus et Hexaprotodon sivalensis qui n’ont pas été déterminées dans la collection issue des fouilles (tableau 38).

Second chapitre : Etude paléontologique.
- 130 -
En ce qui concerne le site de Sendang Busik, la diversité de la liste faunique est très proche de celle des listes issue des ramassages de surface, exceptée pour l’espèce Sus sp. qui n’a été déterminée qu’en surface (tableau 38).
À Ngrejeng, la liste faunique établie à partir des restes récoltés en surface paraît plus variée pour les mammifères. En effet, les espèces Stegodon sp., Rhinoceros sp. et Hexaprotodon sivalensis ne se retrouvent pas dans la liste faunique des restes fouillés. Néanmoins, des fossiles de crocodiles et de poisson n’ont pas été observés dans les collections de surface (tableau 38).
À Grogol, les collections issues des fouilles et de la surface, récoltées en 1983, sont quasiment identiques à une espèce près : Stegodon sp. Ces listes ne sont pas très diversifiées.
À Bukuran, les deux listes établies à partir des collections fouillées et de surface de 1979 sont très proches. L’espèce Stegodon sp. est seulement présente en surface alors que Sus sp. n’est présente que dans la collection fouillée. Par contre, les listes issues de la fouille et des ramassages d’Aimi et Aziz sont plus riche en espèces (tableau 38).
Enfin à Dayu, les listes fauniques sont toutes identiques à une espèce près, Stegodon trigonocephalus, qui a été retrouvée seulement dans les ramassages d’Aimi et Aziz. Ces trois listes sont pauvres en espèces (tableau 38).
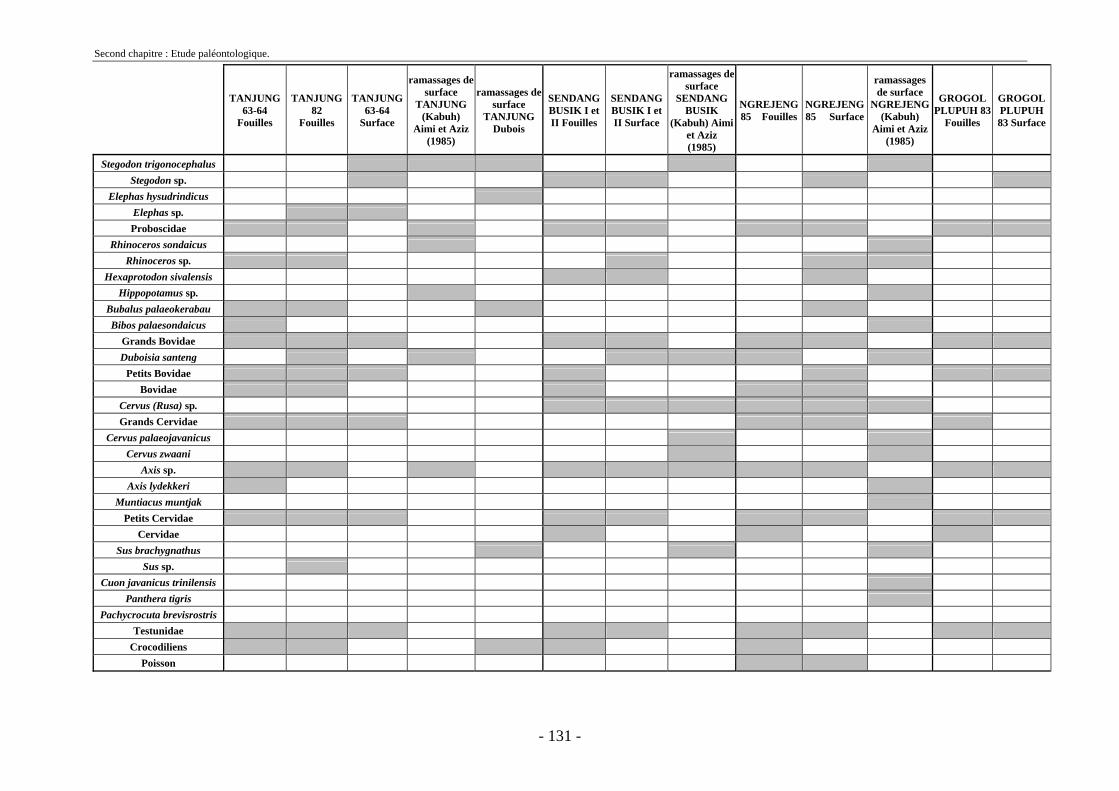
Second chapitre : Etude paléontologique.
- 131 -
TANJUNG
63-64 Fouilles
TANJUNG 82
Fouilles
TANJUNG 63-64
Surface
ramassages de surface
TANJUNG (Kabuh)
Aimi et Aziz (1985)
ramassages de surface
TANJUNG Dubois
SENDANG BUSIK I et II Fouilles
SENDANG BUSIK I et II Surface
ramassages de surface
SENDANG BUSIK
(Kabuh) Aimi et Aziz (1985)
NGREJENG 85 Fouilles
NGREJENG 85 Surface
ramassages de surface
NGREJENG (Kabuh)
Aimi et Aziz (1985)
GROGOL PLUPUH 83
Fouilles
GROGOL PLUPUH 83 Surface
Stegodon trigonocephalus Stegodon sp.
Elephas hysudrindicus Elephas sp. Proboscidae
Rhinoceros sondaicus Rhinoceros sp.
Hexaprotodon sivalensis Hippopotamus sp.
Bubalus palaeokerabau Bibos palaesondaicus
Grands Bovidae Duboisia santeng
Petits Bovidae Bovidae
Cervus (Rusa) sp. Grands Cervidae
Cervus palaeojavanicus Cervus zwaani
Axis sp. Axis lydekkeri
Muntiacus muntjak Petits Cervidae
Cervidae Sus brachygnathus
Sus sp. Cuon javanicus trinilensis
Panthera tigris Pachycrocuta brevisrostris
Testunidae Crocodiliens
Poisson
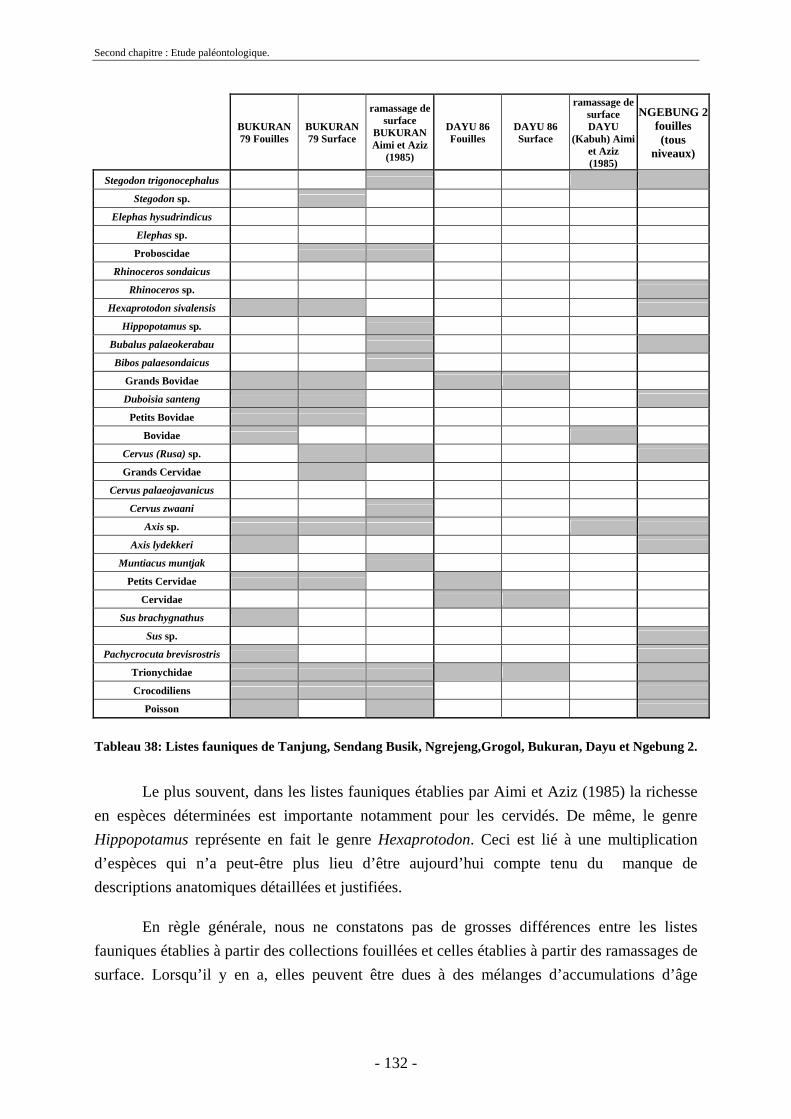
Second chapitre : Etude paléontologique.
- 132 -
BUKURAN 79 Fouilles
BUKURAN 79 Surface
ramassage de surface
BUKURAN Aimi et Aziz
(1985)
DAYU 86 Fouilles
DAYU 86 Surface
ramassage de surface DAYU
(Kabuh) Aimi et Aziz (1985)
NGEBUNG 2 fouilles (tous
niveaux)
Stegodon trigonocephalus Stegodon sp.
Elephas hysudrindicus Elephas sp. Proboscidae
Rhinoceros sondaicus Rhinoceros sp.
Hexaprotodon sivalensis Hippopotamus sp.
Bubalus palaeokerabau Bibos palaesondaicus
Grands Bovidae Duboisia santeng
Petits Bovidae Bovidae
Cervus (Rusa) sp. Grands Cervidae
Cervus palaeojavanicus Cervus zwaani
Axis sp. Axis lydekkeri
Muntiacus muntjak Petits Cervidae
Cervidae Sus brachygnathus
Sus sp. Pachycrocuta brevisrostris
Trionychidae Crocodiliens
Poisson
Tableau 38: Listes fauniques de Tanjung, Sendang Busik, Ngrejeng,Grogol, Bukuran, Dayu et Ngebung 2.
Le plus souvent, dans les listes fauniques établies par Aimi et Aziz (1985) la richesse
en espèces déterminées est importante notamment pour les cervidés. De même, le genre Hippopotamus représente en fait le genre Hexaprotodon. Ceci est lié à une multiplication d’espèces qui n’a peut-être plus lieu d’être aujourd’hui compte tenu du manque de descriptions anatomiques détaillées et justifiées.
En règle générale, nous ne constatons pas de grosses différences entre les listes fauniques établies à partir des collections fouillées et celles établies à partir des ramassages de surface. Lorsqu’il y en a, elles peuvent être dues à des mélanges d’accumulations d’âge
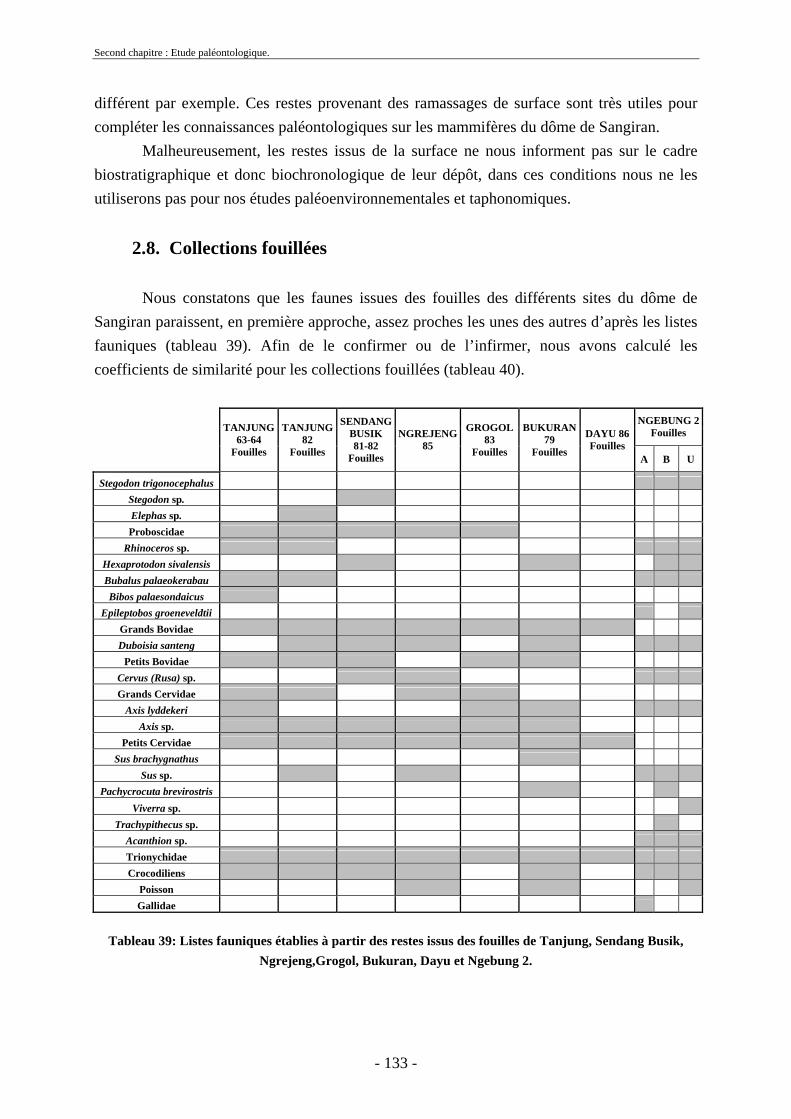
Second chapitre : Etude paléontologique.
- 133 -
différent par exemple. Ces restes provenant des ramassages de surface sont très utiles pour compléter les connaissances paléontologiques sur les mammifères du dôme de Sangiran.
Malheureusement, les restes issus de la surface ne nous informent pas sur le cadre biostratigraphique et donc biochronologique de leur dépôt, dans ces conditions nous ne les utiliserons pas pour nos études paléoenvironnementales et taphonomiques.
2.8. Collections fouillées Nous constatons que les faunes issues des fouilles des différents sites du dôme de Sangiran paraissent, en première approche, assez proches les unes des autres d’après les listes fauniques (tableau 39). Afin de le confirmer ou de l’infirmer, nous avons calculé les coefficients de similarité pour les collections fouillées (tableau 40).
NGEBUNG 2
Fouilles
TANJUNG 63-64
Fouilles
TANJUNG 82
Fouilles
SENDANG BUSIK 81-82
Fouilles
NGREJENG 85
GROGOL 83
Fouilles
BUKURAN 79
Fouilles
DAYU 86 Fouilles
A B U
Stegodon trigonocephalus Stegodon sp. Elephas sp. Proboscidae
Rhinoceros sp. Hexaprotodon sivalensis Bubalus palaeokerabau Bibos palaesondaicus
Epileptobos groeneveldtii Grands Bovidae Duboisia santeng
Petits Bovidae Cervus (Rusa) sp. Grands Cervidae
Axis lyddekeri Axis sp.
Petits Cervidae Sus brachygnathus
Sus sp. Pachycrocuta brevirostris
Viverra sp. Trachypithecus sp.
Acanthion sp. Trionychidae Crocodiliens
Poisson Gallidae
Tableau 39: Listes fauniques établies à partir des restes issus des fouilles de Tanjung, Sendang Busik,
Ngrejeng,Grogol, Bukuran, Dayu et Ngebung 2.
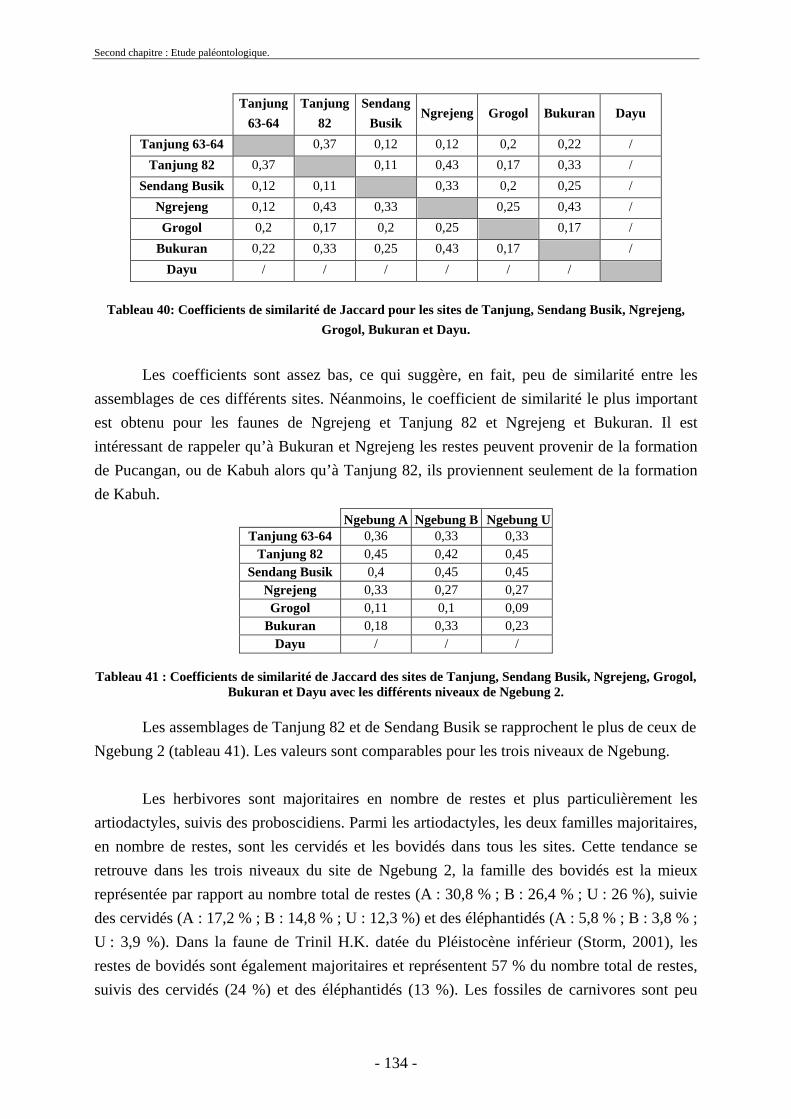
Second chapitre : Etude paléontologique.
- 134 -
Tanjung 63-64
Tanjung 82
Sendang Busik
Ngrejeng Grogol Bukuran Dayu
Tanjung 63-64 0,37 0,12 0,12 0,2 0,22 / Tanjung 82 0,37 0,11 0,43 0,17 0,33 /
Sendang Busik 0,12 0,11 0,33 0,2 0,25 / Ngrejeng 0,12 0,43 0,33 0,25 0,43 / Grogol 0,2 0,17 0,2 0,25 0,17 /
Bukuran 0,22 0,33 0,25 0,43 0,17 / Dayu / / / / / /
Tableau 40: Coefficients de similarité de Jaccard pour les sites de Tanjung, Sendang Busik, Ngrejeng,
Grogol, Bukuran et Dayu.
Les coefficients sont assez bas, ce qui suggère, en fait, peu de similarité entre les assemblages de ces différents sites. Néanmoins, le coefficient de similarité le plus important est obtenu pour les faunes de Ngrejeng et Tanjung 82 et Ngrejeng et Bukuran. Il est intéressant de rappeler qu’à Bukuran et Ngrejeng les restes peuvent provenir de la formation de Pucangan, ou de Kabuh alors qu’à Tanjung 82, ils proviennent seulement de la formation de Kabuh.
Ngebung A Ngebung B Ngebung UTanjung 63-64 0,36 0,33 0,33
Tanjung 82 0,45 0,42 0,45 Sendang Busik 0,4 0,45 0,45
Ngrejeng 0,33 0,27 0,27 Grogol 0,11 0,1 0,09
Bukuran 0,18 0,33 0,23 Dayu / / /
Tableau 41 : Coefficients de similarité de Jaccard des sites de Tanjung, Sendang Busik, Ngrejeng, Grogol,
Bukuran et Dayu avec les différents niveaux de Ngebung 2. Les assemblages de Tanjung 82 et de Sendang Busik se rapprochent le plus de ceux de Ngebung 2 (tableau 41). Les valeurs sont comparables pour les trois niveaux de Ngebung.
Les herbivores sont majoritaires en nombre de restes et plus particulièrement les artiodactyles, suivis des proboscidiens. Parmi les artiodactyles, les deux familles majoritaires, en nombre de restes, sont les cervidés et les bovidés dans tous les sites. Cette tendance se retrouve dans les trois niveaux du site de Ngebung 2, la famille des bovidés est la mieux représentée par rapport au nombre total de restes (A : 30,8 % ; B : 26,4 % ; U : 26 %), suivie des cervidés (A : 17,2 % ; B : 14,8 % ; U : 12,3 %) et des éléphantidés (A : 5,8 % ; B : 3,8 % ; U : 3,9 %). Dans la faune de Trinil H.K. datée du Pléistocène inférieur (Storm, 2001), les restes de bovidés sont également majoritaires et représentent 57 % du nombre total de restes, suivis des cervidés (24 %) et des éléphantidés (13 %). Les fossiles de carnivores sont peu

Second chapitre : Etude paléontologique.
- 135 -
nombreux à Trinil H.K. comme à Ngebung 2. Ces représentations sont donc typiques de Java et confirment que les carnivores ont peu migré vers cette île contrairement aux herbivores.
Pour tous les sites de notre étude, nous remarquons aussi que les restes déterminables anatomiquement et taxonomiquement (=NRDt) sont peu nombreux dans les collections fouillées. Sur l’ensemble des assemblages, les restes déterminables anatomiquement et taxonomiquement sont le plus souvent des dents isolées.
Afin de quantifier la détermination des restes à Sangiran nous avons calculé le taux de
détermination des différents assemblages fouillés (Brugal & Patou-Mathis, 1993) (tableau 42) où le Nombre de Restes Déterminables anatomiquement et totalement (NRD) est divisé par le Nombre de Restes Total (NRT):
Taux de détermination : (NRD/NRT)*100.
Tanjung 63-64
Tanjung 82
Sendang Busik
Ngrejeng Grogol Bukuran Dayu
Taux de détermination
36,5 % 47,9 % 46 % 52 % 42 % 51 % 33 %
Tableau 42: Taux de détermination des sites de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et
Dayu.
Les taux de détermination ne sont pas les mêmes suivant les sites. Ce taux est le plus élevé à Ngrejeng et à Bukuran, aux environs de 50 %. À Tanjung 63-64 et Dayu ce taux est plus faible aux environs de 35 %. Ces différences sont liées à la nature des restes présents sur les sites, à la conservation et la fracturation des assemblages.
2.9. Spectres fauniques Nous avons ensuite calculé le nombre minimum d’individus (NMI) de chaque espèce de mammifère. Pour ce faire, nous avons choisi d’utiliser le NMI de fréquence (NMIf) basé sur le nombre le plus élevé d’éléments latéralisés (Poplin, 1976) (tableaux 42 à 47). Ces valeurs sont très délicates à exploiter car chaque espèce est représentée par très peu de restes dans chaque assemblage. En effet dans ce cas, le NMI surestime la fréquence relative d’un taxon représenté par peu de restes. De plus, l’utilisation de celui-ci implique une équivalence entre un reste et un animal entier ce qui n’est pas toujours le cas car certaines parties anatomiques de l’animal peuvent avoir été transportées séparément dans le site (Brugal et al., 1994). En outre, nous avons indiqué le nombre minimum d’éléments ou NME (=MNE Minimum Number of Elements). Il s’agit du nombre d’éléments d’une partie anatomique, quel que soit l’âge, le sexe et le côté, c’est donc une estimation du nombre minimal pour une catégorie anatomique désignée (Binford, 1984). Nous remarquons que le NME est très peu différent du NR.
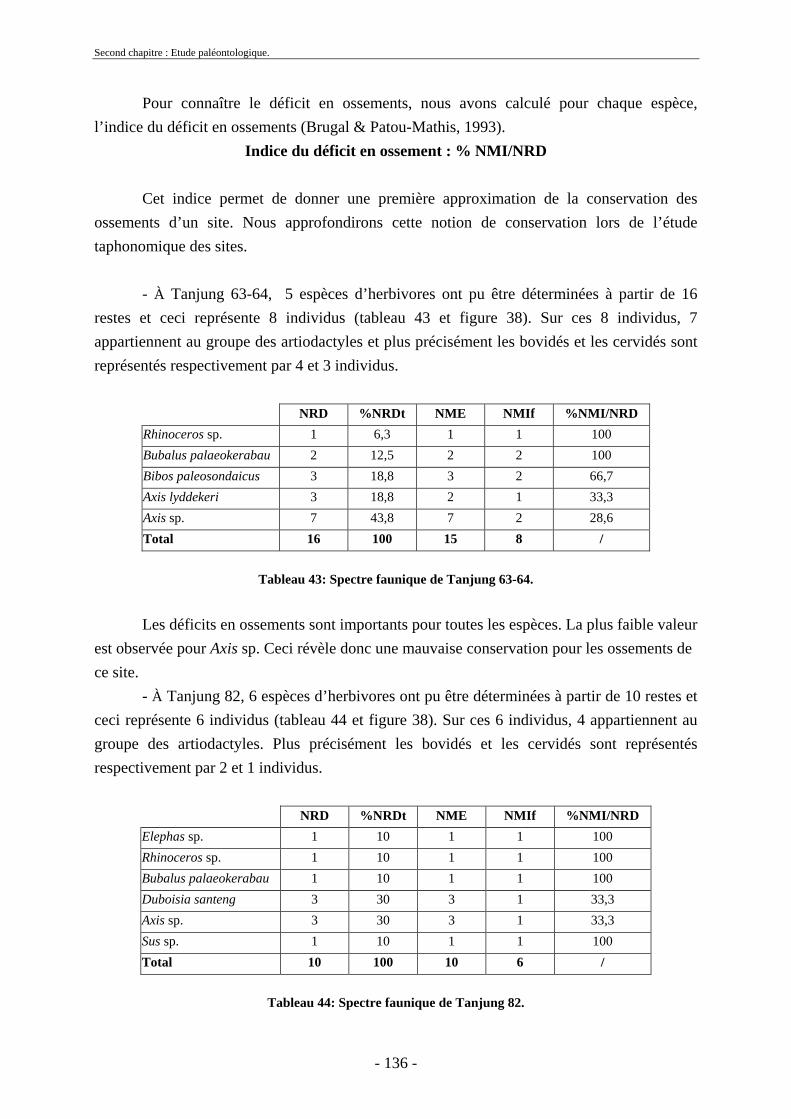
Second chapitre : Etude paléontologique.
- 136 -
Pour connaître le déficit en ossements, nous avons calculé pour chaque espèce, l’indice du déficit en ossements (Brugal & Patou-Mathis, 1993).
Indice du déficit en ossement : % NMI/NRD
Cet indice permet de donner une première approximation de la conservation des ossements d’un site. Nous approfondirons cette notion de conservation lors de l’étude taphonomique des sites. - À Tanjung 63-64, 5 espèces d’herbivores ont pu être déterminées à partir de 16 restes et ceci représente 8 individus (tableau 43 et figure 38). Sur ces 8 individus, 7 appartiennent au groupe des artiodactyles et plus précisément les bovidés et les cervidés sont représentés respectivement par 4 et 3 individus.
NRD %NRDt NME NMIf %NMI/NRD Rhinoceros sp. 1 6,3 1 1 100 Bubalus palaeokerabau 2 12,5 2 2 100 Bibos paleosondaicus 3 18,8 3 2 66,7 Axis lyddekeri 3 18,8 2 1 33,3 Axis sp. 7 43,8 7 2 28,6 Total 16 100 15 8 /
Tableau 43: Spectre faunique de Tanjung 63-64.
Les déficits en ossements sont importants pour toutes les espèces. La plus faible valeur
est observée pour Axis sp. Ceci révèle donc une mauvaise conservation pour les ossements de ce site. - À Tanjung 82, 6 espèces d’herbivores ont pu être déterminées à partir de 10 restes et ceci représente 6 individus (tableau 44 et figure 38). Sur ces 6 individus, 4 appartiennent au groupe des artiodactyles. Plus précisément les bovidés et les cervidés sont représentés respectivement par 2 et 1 individus.
NRD %NRDt NME NMIf %NMI/NRD Elephas sp. 1 10 1 1 100 Rhinoceros sp. 1 10 1 1 100 Bubalus palaeokerabau 1 10 1 1 100 Duboisia santeng 3 30 3 1 33,3 Axis sp. 3 30 3 1 33,3 Sus sp. 1 10 1 1 100 Total 10 100 10 6 /
Tableau 44: Spectre faunique de Tanjung 82.

Second chapitre : Etude paléontologique.
- 137 -
Le déficit en ossements est très important pour toutes les espèces excepté pour Duboisia santeng et Axis sp. - À Sendang Busik, 4 espèces d’herbivores ont pu être déterminées à partir de seulement 6 restes et ces restes représentent 5 individus. Sur ces 5 individus, 4 appartiennent au groupe des artiodactyles et parmi ces 4, 2 sont des cervidés (tableau 45 et figure 38).
NRD %NRDt NME NMIf %NMI/NRD Stegodon sp. 1 16,7 1 1 100 Hexaprotodon sivalensis 2 33,3 2 2 100 Cervus (Rusa) sp. 1 16,7 1 1 100 Axis sp. 2 33,3 2 1 50 Total 6 100 6 5 /
Tableau 45: Spectre faunique de Sendang Busik.
Les indices de déficit en ossements sont très élevés, ce qui révèle une conservation
encore moins bonne qu’à Tanjung.
- À Ngrejeng, 4 espèces ont été déterminées à partir de 12 restes et cela représente 5 individus. Nous pouvons remarquer la présence de deux individus appartenant aux cervidés et un individu aux bovidés (tableau 46 et figure 38).
NRD %NRDt NME NMIf %NMI/NRD Duboisia santeng 4 33,3 4 2 50 Cervus (Rusa) sp. 6 50,0 6 1 16,7 Axis sp. 1 8,3 1 1 100 Sus sp. 1 8,3 1 1 100 Total 12 100 12 5 /
Tableau 46: Spectre faunique de Ngrejeng.
Les indices de déficit en ossements, de même qu’à Sendang Busik, sont très élevés, ce
qui révèle une conservation encore moins bonne qu’à Tanjung. Pour Cervus (Rusa) sp., ce déficit est moindre que pour les autres espèces de Ngrejeng mais reste encore assez élevé.
- À Grogol, seulement une espèce de cervidé a été déterminée à partir de 4 restes : Axis sp. ; et ces restes représentent un individu (tableau 47 et figure 38).
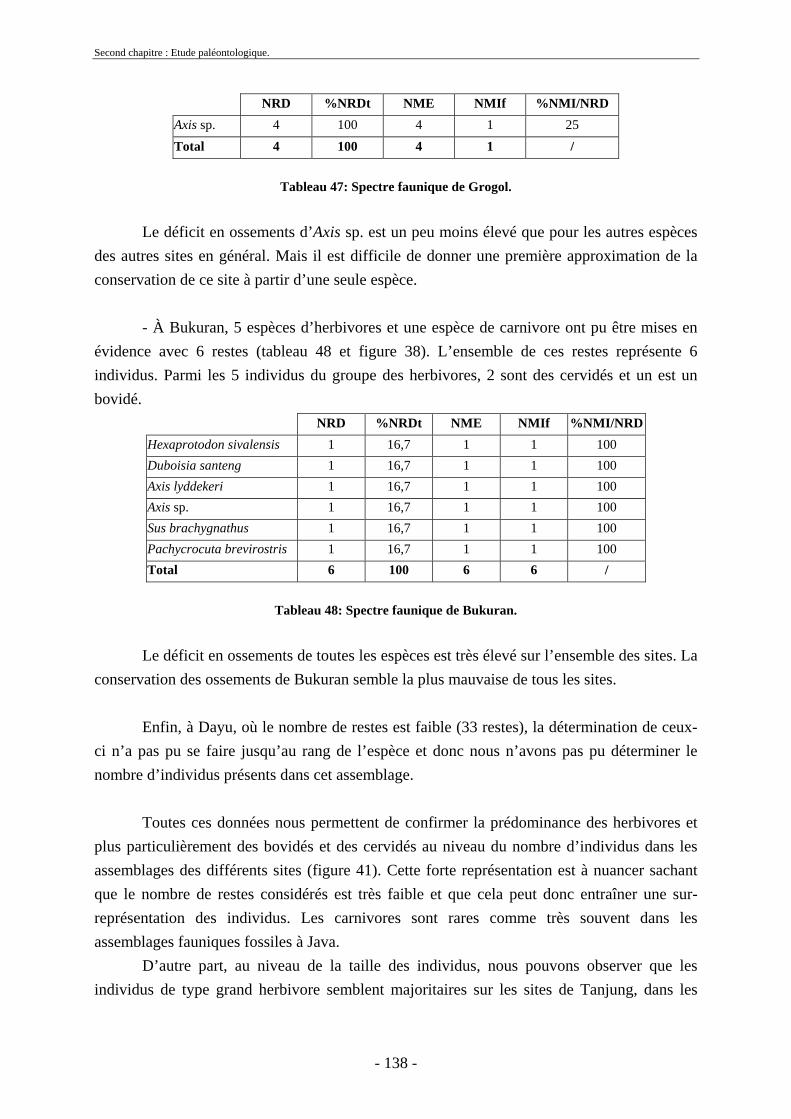
Second chapitre : Etude paléontologique.
- 138 -
NRD %NRDt NME NMIf %NMI/NRD Axis sp. 4 100 4 1 25 Total 4 100 4 1 /
Tableau 47: Spectre faunique de Grogol.
Le déficit en ossements d’Axis sp. est un peu moins élevé que pour les autres espèces
des autres sites en général. Mais il est difficile de donner une première approximation de la conservation de ce site à partir d’une seule espèce.
- À Bukuran, 5 espèces d’herbivores et une espèce de carnivore ont pu être mises en évidence avec 6 restes (tableau 48 et figure 38). L’ensemble de ces restes représente 6 individus. Parmi les 5 individus du groupe des herbivores, 2 sont des cervidés et un est un bovidé.
NRD %NRDt NME NMIf %NMI/NRD Hexaprotodon sivalensis 1 16,7 1 1 100 Duboisia santeng 1 16,7 1 1 100 Axis lyddekeri 1 16,7 1 1 100 Axis sp. 1 16,7 1 1 100 Sus brachygnathus 1 16,7 1 1 100 Pachycrocuta brevirostris 1 16,7 1 1 100 Total 6 100 6 6 /
Tableau 48: Spectre faunique de Bukuran.
Le déficit en ossements de toutes les espèces est très élevé sur l’ensemble des sites. La conservation des ossements de Bukuran semble la plus mauvaise de tous les sites.
Enfin, à Dayu, où le nombre de restes est faible (33 restes), la détermination de ceux-ci n’a pas pu se faire jusqu’au rang de l’espèce et donc nous n’avons pas pu déterminer le nombre d’individus présents dans cet assemblage. Toutes ces données nous permettent de confirmer la prédominance des herbivores et plus particulièrement des bovidés et des cervidés au niveau du nombre d’individus dans les assemblages des différents sites (figure 41). Cette forte représentation est à nuancer sachant que le nombre de restes considérés est très faible et que cela peut donc entraîner une sur-représentation des individus. Les carnivores sont rares comme très souvent dans les assemblages fauniques fossiles à Java.
D’autre part, au niveau de la taille des individus, nous pouvons observer que les individus de type grand herbivore semblent majoritaires sur les sites de Tanjung, dans les

Second chapitre : Etude paléontologique.
- 139 -
deux niveaux, et de Sendang Busik, alors que sur les autres sites les petits bovidés et les cervidés sont majoritaires.
Au niveau du nombre de restes, les restes de grands bovidés et de petits cervidés sont majoritaires sur tous les sites.
Ces données nous donnent une image de la faune contemporaine du Pithécanthrope et donc une première idée des espèces pouvant interagir avec les hominidés.
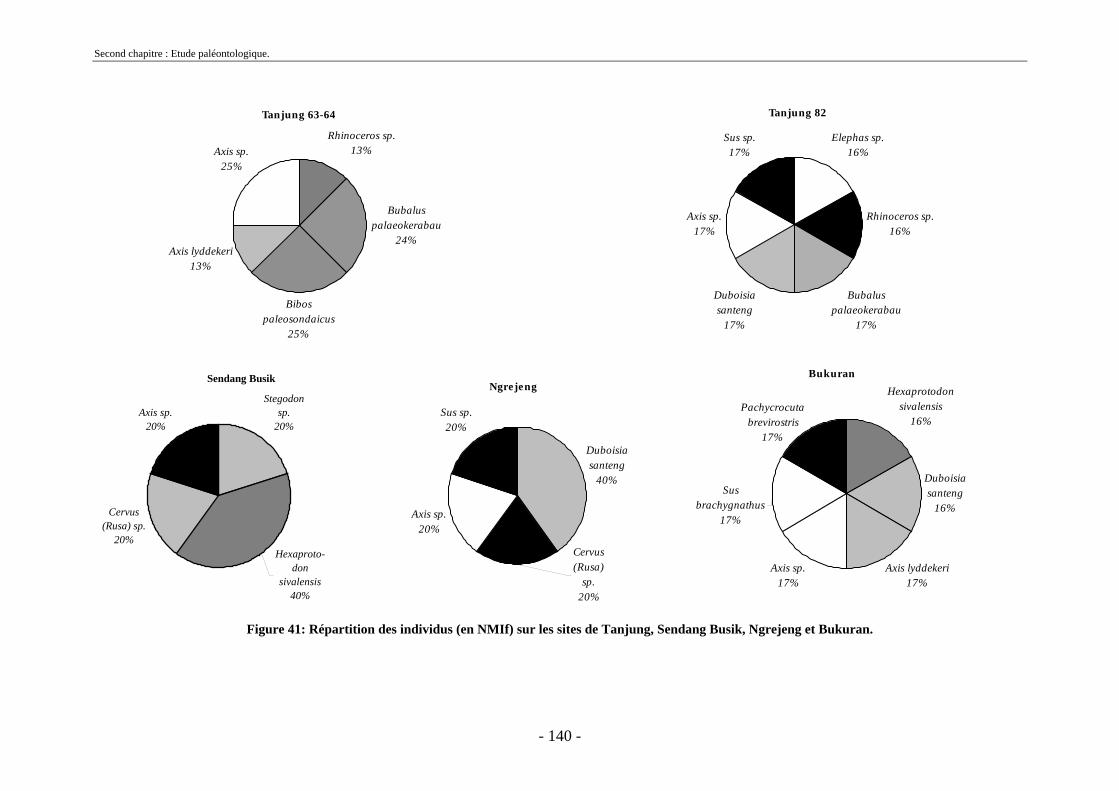
Second chapitre : Etude paléontologique.
- 140 -
Tanjung 63-64
Rhinoceros sp.13%
Bibos paleosondaicus
25%
Axis lyddekeri13%
Axis sp.25%
Bubalus palaeokerabau
24%
Tanjung 82
Elephas sp.16%
Rhinoceros sp.16%
Bubalus palaeokerabau
17%
Duboisia santeng
17%
Axis sp.17%
Sus sp.17%
Sendang Busik
Stegodon sp.
20%
Cervus (Rusa) sp.
20%
Axis sp.20%
Hexaproto-don
sivalensis40%
Ngrejeng
Duboisia santeng
40%
Axis sp.20%
Sus sp.20%
Cervus (Rusa)
sp.20%
Bukuran
Duboisia santeng
16%
Axis lyddekeri17%
Axis sp.17%
Sus brachygnathus
17%
Pachycrocuta brevirostris
17%
Hexaprotodon sivalensis
16%
Figure 41: Répartition des individus (en NMIf) sur les sites de Tanjung, Sendang Busik, Ngrejeng et Bukuran.

Second chapitre : Etude paléontologique.
- 141 -
2.10. Âge des animaux
Pour déterminer l’âge des animaux, la méthode basée sur les stades de croissance et le degré d’abrasion des dents est la plus fréquemment utilisée (Klein et al., 1981 ; Davis, 1983 ; Morrison & Whitridge, 1997) mais elle présente également des désavantages (Quimby & Gaab, 1957 ; Stallibrass, 1982). Il peut exister des différences entre des individus de deux régions différentes, entre deux individus de la même population, entre deux hémi-mandibules du même animal, entre les incisives et les jugales et entre les dents de la même hémi-mandibule. Dans notre zone d’étude aucune étude de l’âge des animaux n’a encore été tentée hormis pour les proboscidiens (van den Bergh, 1999).
D’après les restes dentaires il a été difficile de proposer de nombreuses classes d’âges car nous n’avions pas de mandibules pour comparer les stades d’usure des dents isolées.
Les hippopotamidés:
Pour l’ensemble des restes des sites considérés, seul l’âge relatif de deux individus de l’espèce Hexaprotodon sivalensis a pu être connu à Sendang Busik. SBII/5/34 est une première déciduale inférieure appartenant donc à un individu juvénile et SBII/9/17 est une M3
très usée provenant certainement d’un individu très âgé car cette dent est la dernière jugale qui apparaît lors de l’éruption dentaire (tableau 50). Les suidés:
Le degré d’abrasion de la M3 de Sus brachygnathus de Bukuran (BK79/III/A1/2) indique un individu âgé car elle est usée (tableau 53). Les bovidés:
À Ngrejeng, d’après l’usure de la M2 de Duboisia santeng (NKJ85/III/15) nous pouvons penser que cette dent provient d’un individu âgé (tableau 52).
Pour le reste des individus il est difficile de donner des âges plus précis qu’adulte.
Lorsque nous avions des restes autres que dentaires nous n’avons pas proposé d’âge (= points d’interrogation dans les tableaux 49 à 54) hormis pour certains os longs épiphysés comme le fémur de Rhinoceros sp. à Tanjung.
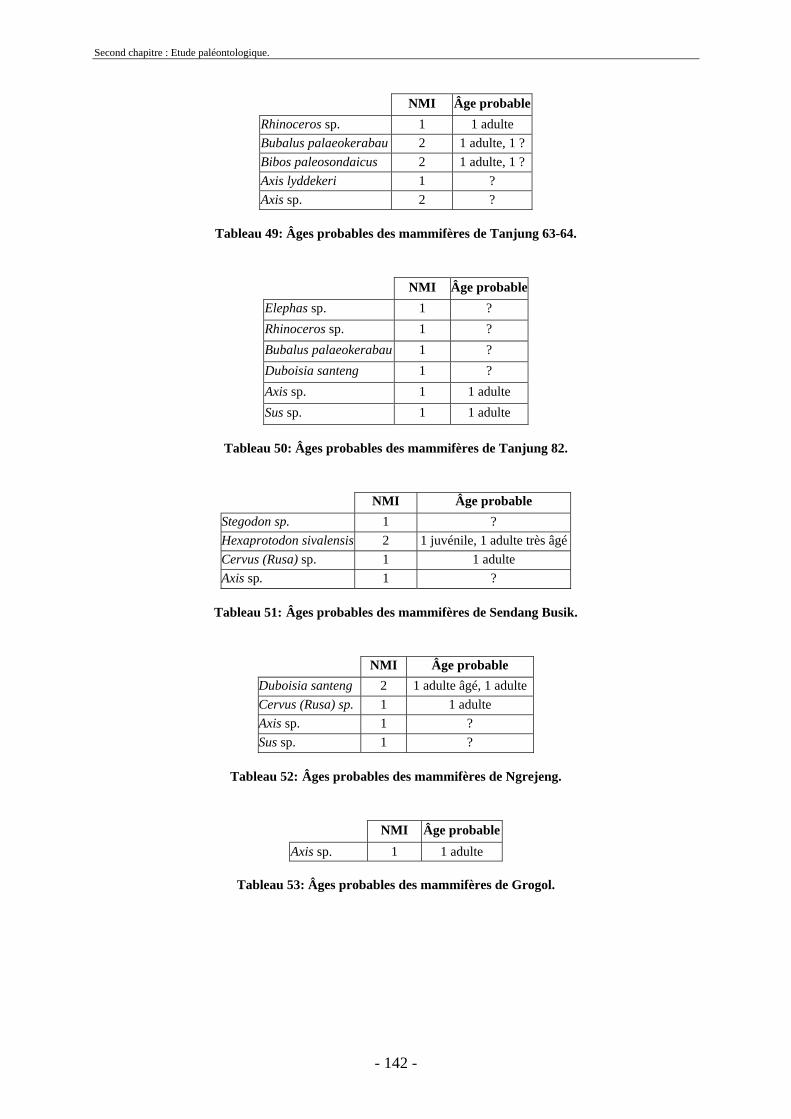
Second chapitre : Etude paléontologique.
- 142 -
NMI Âge probableRhinoceros sp. 1 1 adulte Bubalus palaeokerabau 2 1 adulte, 1 ?Bibos paleosondaicus 2 1 adulte, 1 ?Axis lyddekeri 1 ? Axis sp. 2 ?
Tableau 49: Âges probables des mammifères de Tanjung 63-64.
NMI Âge probableElephas sp. 1 ? Rhinoceros sp. 1 ? Bubalus palaeokerabau 1 ? Duboisia santeng 1 ? Axis sp. 1 1 adulte Sus sp. 1 1 adulte
Tableau 50: Âges probables des mammifères de Tanjung 82.
NMI Âge probable Stegodon sp. 1 ? Hexaprotodon sivalensis 2 1 juvénile, 1 adulte très âgé Cervus (Rusa) sp. 1 1 adulte Axis sp. 1 ?
Tableau 51: Âges probables des mammifères de Sendang Busik.
NMI Âge probable Duboisia santeng 2 1 adulte âgé, 1 adulteCervus (Rusa) sp. 1 1 adulte Axis sp. 1 ? Sus sp. 1 ?
Tableau 52: Âges probables des mammifères de Ngrejeng.
NMI Âge probableAxis sp. 1 1 adulte
Tableau 53: Âges probables des mammifères de Grogol.

Second chapitre : Etude paléontologique.
- 143 -
NMI Âge probableHexaprotodon sivalensis 1 1 adulte Duboisia santeng 1 1 adulte Axis lyddekeri 1 ? Axis sp. 1 ? Sus brachygnathus 1 1 adulte âgéPachycrocuta brevirostris 1 ?
Tableau 54: Âges probables des mammifères de Bukuran.
2.11. Richesse biologique La richesse biologique d’un gisement peut dépendre de nombreux facteurs : l’intervention humaine ou des carnivores, la richesse en espèces vivantes de la zone considérée, des événements catastrophiques… Afin d’appréhender cette richesse biologique nous avons calculé un indice. Cet indice (D) a été établi par Margalef (Daget, 1979 ; Legendre & Legendre, 1984) (tableau 55):
D = n-1/ln N (n : nombre d’espèces, ln : logarithme néperien, N : nombre total d’individus)
Indice de richesse
(D) Tanjung 63-64 1,92
Tanjung 82 2,79 Sendang Busik 1,86
Ngrejeng 1,86 Bukuran 2,79
Tableau 55: Indice de richesse biologique des sites de Tanjung, Sendang Busik, Ngrejeng et Bukuran.
Nous n’avons pas pu calculer cet indice à Grogol car il n’existe qu’une espèce déterminable et à Dayu car aucune espèce n’a été déterminée. La richesse biologique est plus importante dans les assemblages de Tanjung 82 et de Bukuran. Pour Bukuran, cette hypothèse est à nuancer, sachant que les restes de ce site proviennent de plusieurs formations ce qui a pu contribuer à la richesse biologique de même qu’à Ngrejeng. Á titre de comparaison, ce même indice varie entre 1,913 et 3,395 dans les couches C et D (Monchot, 1998) du site de plein air naturel de Tourville situé dans la basse vallée de la Seine et daté du Pléistocène moyen (Descombes, 1980). De même sur le site de plein air archéologique du Mont Dol (Ille-et-Vilaine) daté du Paléolithique moyen (Simonet & Monnier, 1991) cet indice est de 3,508 (Monchot, 1998). Ceci ne nous permet pas de conclure sur la nature des sites considérés.
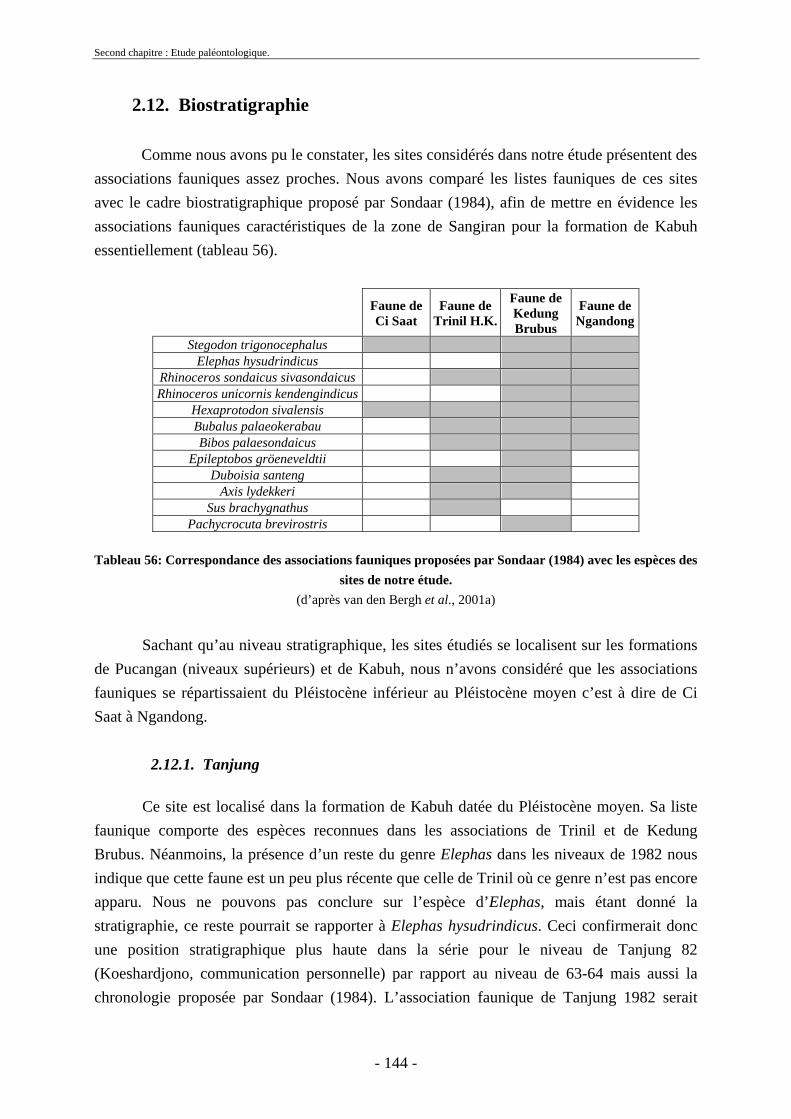
Second chapitre : Etude paléontologique.
- 144 -
2.12. Biostratigraphie
Comme nous avons pu le constater, les sites considérés dans notre étude présentent des associations fauniques assez proches. Nous avons comparé les listes fauniques de ces sites avec le cadre biostratigraphique proposé par Sondaar (1984), afin de mettre en évidence les associations fauniques caractéristiques de la zone de Sangiran pour la formation de Kabuh essentiellement (tableau 56).
Faune de Ci Saat
Faune de Trinil H.K.
Faune de Kedung Brubus
Faune de Ngandong
Stegodon trigonocephalus Elephas hysudrindicus
Rhinoceros sondaicus sivasondaicus Rhinoceros unicornis kendengindicus
Hexaprotodon sivalensis Bubalus palaeokerabau Bibos palaesondaicus
Epileptobos gröeneveldtii Duboisia santeng
Axis lydekkeri Sus brachygnathus
Pachycrocuta brevirostris Tableau 56: Correspondance des associations fauniques proposées par Sondaar (1984) avec les espèces des
sites de notre étude. (d’après van den Bergh et al., 2001a)
Sachant qu’au niveau stratigraphique, les sites étudiés se localisent sur les formations de Pucangan (niveaux supérieurs) et de Kabuh, nous n’avons considéré que les associations fauniques se répartissaient du Pléistocène inférieur au Pléistocène moyen c’est à dire de Ci Saat à Ngandong.
2.12.1. Tanjung Ce site est localisé dans la formation de Kabuh datée du Pléistocène moyen. Sa liste faunique comporte des espèces reconnues dans les associations de Trinil et de Kedung Brubus. Néanmoins, la présence d’un reste du genre Elephas dans les niveaux de 1982 nous indique que cette faune est un peu plus récente que celle de Trinil où ce genre n’est pas encore apparu. Nous ne pouvons pas conclure sur l’espèce d’Elephas, mais étant donné la stratigraphie, ce reste pourrait se rapporter à Elephas hysudrindicus. Ceci confirmerait donc une position stratigraphique plus haute dans la série pour le niveau de Tanjung 82 (Koeshardjono, communication personnelle) par rapport au niveau de 63-64 mais aussi la chronologie proposée par Sondaar (1984). L’association faunique de Tanjung 1982 serait

Second chapitre : Etude paléontologique.
- 145 -
assez proche de celle de Kedung Brubus alors que celle de Tanjung 63-64 serait assez proche de celle de Trinil. Le niveau où le crâne Sangiran 10 a été découvert, a lui-même été daté d’environ 0,83 Ma (Jacob, 1975a). Ce niveau correspond à celui de Tanjung 63-64. La présence d’Elephas à Sangiran est donc postérieure à cette période et coïncide avec la date donnée pour l’association de Kedung Brubus (0,8 Ma, Leinders et al., 1985) (tableau 56).
2.12.2. Sendang Busik Ce site se localise dans la formation de Kabuh. Les espèces présentes sur ce site le sont également dans l’ensemble de la succession des quatre associations de Sondaar (tableau 49). Seules les espèces Duboisia santeng et Axis lydekkeri nous permettent d’affiner la biostratigraphie et de proposer une association plus proche des faunes de Trinil H.K. et de Kedung Brubus (tableau 57).
2.12.3. Ngrejeng Plupuh (= Ngrejeng) Ce site se trouve sur les formations de Pucangan et de Kabuh. Le cadre biostratigraphique est donc ici plus difficile à définir. Cervus (Rusa) sp. apparaît au niveau de l’association de Kedung Brubus, les premiers suidés au niveau de Trinil H.K. et les premiers Axis à partir de l’association de Ci Saat. Cet assemblage faunique doit être l’image d’un mélange de plusieurs associations entre Ci Saat, Trinil H.K et Kedung Brubus. D’après des indications sédimentaires, 24 restes proviendraient d’une passée argileuse sableuse (Lempung Pasir). Nous pouvons nous interroger sur ces indications, serait-ce la formation de Pucangan ou non ?
- Si oui, une dent de Duboisia santeng est présente parmi ces 24 restes or cette espèce n’apparaît que dans l’unité de Trinil H.K. dans le modèle biostratigraphique de Sondaar (van den Bergh et al., 2001a). Vue sa position stratigraphique, cette espèce ne peut pas appartenir à l’unité de Trinil H.K. Dans ce cas un bovidé doit être ajouté dans les niveaux plus anciens que cette dernière association (tableau 57) et alors le cadre biostratigraphique est complété.
- Si non, nous nous trouvons dans la formation de Kabuh, peut-être à sa base et la position de la dent de Duboisia santeng est alors en concordance avec les résultats obtenus par Sondaar (1984) et serait donc caractéristique de Trinil H.K.
Nous ne pouvons donc pas conclure en raison des incertitudes sur l’origine des fossiles.
2.12.4. Grogol Plupuh (= Grogol) Ce site se trouve également dans la formation de Kabuh. Sa liste faunique est moins variée que celles des deux sites précédents. L’espèce Axis lydekkeri, seule espèce présente

Second chapitre : Etude paléontologique.
- 146 -
dans la liste, nous indique que cette association est probablement assez proche de celles de Trinil H.K. ou de Kedung Brubus (tableau 57).
2.12.5. Bukuran Ce site, comme celui de Ngrejeng Plupuh, se localise à la fois sur les formations de Pucangan et de Kabuh. Il est encore délicat de positionner cet assemblage car il provient de deux formations sédimentaires. Les espèces Axis et Pachycrocuta proviennent d’un niveau de sable conglomératique (Pasir Conglomeratik). Ce niveau ne peut pas être considéré comme le Grenzbank car il n’est pas nommé comme tel et à Bukuran celui-ci n’apparaît pas dans la stratigraphie (Widianto et al., 1994). Cependant dans la colonne stratigraphique obtenue suite à l’étude de diverses tranchées, la base de la formation est constituée d’un sable avec des graviers (Widianto et al., 1994). Dans ce contexte, nous pouvons penser que les espèces Axis et Pachycrocuta se situent dans la partie inférieure de la formation de Kabuh. Dans le cadre biostratigraphique proposé par Sondaar, la hyène apparaît dans l’association de Kedung Brubus c’est à dire dans la partie supérieure de la formation de Kabuh selon Leinders et al. (1985). La hyène semble donc présente à Sangiran dès l’association de Trinil H.K.
En outre, Sus brachygnathus apparaît au niveau de l’association de Trinil H.K. et il se situe dans la formation de Kabuh au-dessus du Grenzbank d’après les indications sédimentaires (Pasir Halus). L’espèce Duboisia santeng est caractéristique des associations de Trinil H.K. et de Kedung Brubus et Axis lydekkeri de celle de Ci Saat, Trinil H.K. et de Kedung Brubus (tableau 57). Cet assemblage de Bukuran semble être un mélange de différentes associations non contemporaines qui peuvent aller de l’association de Ci Saat à celle de Kedung Brubus.
2.12.6. Dayu Ce site se localise à la base de la formation de Kabuh, dans le Grenzbank. Le nombre de restes provenant de ce site ne nous permet pas de donner des informations sur son cadre biostratigraphique.
Nos résultats bien que fragmentaires sur la nature des espèces nous ont permis de compléter le cadre biostratigraphique proposé par Sondaar (1984) et Leinders et al. (1985) notamment pour la hyène. Ces premières conclusions sont prometteuses pour de nouvelles recherches dans cette zone (tableau 57). De nouvelles fouilles à Sangiran, bien calées stratigraphiquement, permettraient de progresser dans les connaissances sur la succession des migrations de mammifères à Java.
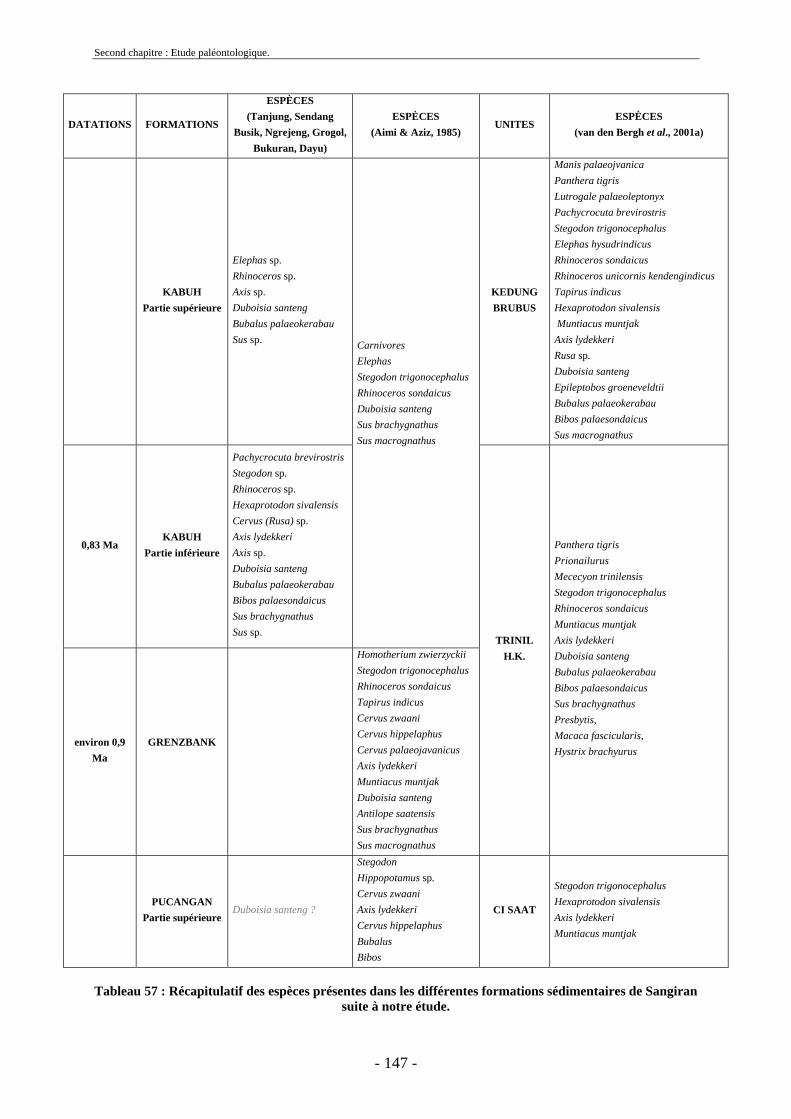
Second chapitre : Etude paléontologique.
- 147 -
DATATIONS FORMATIONS
ESPÈCES (Tanjung, Sendang
Busik, Ngrejeng, Grogol, Bukuran, Dayu)
ESPÈCES (Aimi & Aziz, 1985)
UNITES ESPÈCES
(van den Bergh et al., 2001a)
KABUH Partie supérieure
Elephas sp. Rhinoceros sp. Axis sp. Duboisia santeng Bubalus palaeokerabau Sus sp.
KEDUNG BRUBUS
Manis palaeojvanica Panthera tigris Lutrogale palaeoleptonyx Pachycrocuta brevirostris Stegodon trigonocephalus Elephas hysudrindicus Rhinoceros sondaicus Rhinoceros unicornis kendengindicus Tapirus indicus Hexaprotodon sivalensis Muntiacus muntjak Axis lydekkeri Rusa sp. Duboisia santeng Epileptobos groeneveldtii Bubalus palaeokerabau Bibos palaesondaicus Sus macrognathus
0,83 Ma KABUH
Partie inférieure
Pachycrocuta brevirostris Stegodon sp. Rhinoceros sp. Hexaprotodon sivalensis Cervus (Rusa) sp. Axis lydekkeri Axis sp. Duboisia santeng Bubalus palaeokerabau Bibos palaesondaicus Sus brachygnathus Sus sp.
Carnivores Elephas Stegodon trigonocephalus Rhinoceros sondaicus Duboisia santeng Sus brachygnathus Sus macrognathus
environ 0,9 Ma
GRENZBANK
Homotherium zwierzyckii Stegodon trigonocephalus Rhinoceros sondaicus Tapirus indicus Cervus zwaani Cervus hippelaphus Cervus palaeojavanicus Axis lydekkeri Muntiacus muntjak Duboisia santeng Antilope saatensis Sus brachygnathus Sus macrognathus
TRINIL H.K.
Panthera tigris Prionailurus Mececyon trinilensis Stegodon trigonocephalus Rhinoceros sondaicus Muntiacus muntjak Axis lydekkeri Duboisia santeng Bubalus palaeokerabau Bibos palaesondaicus Sus brachygnathus Presbytis, Macaca fascicularis, Hystrix brachyurus
PUCANGAN Partie supérieure
Duboisia santeng ?
Stegodon Hippopotamus sp. Cervus zwaani Axis lydekkeri Cervus hippelaphus Bubalus Bibos
CI SAAT
Stegodon trigonocephalus Hexaprotodon sivalensis Axis lydekkeri Muntiacus muntjak
Tableau 57 : Récapitulatif des espèces présentes dans les différentes formations sédimentaires de Sangiran
suite à notre étude.

- 148 -

- 149 -
TROISIEME CHAPITRE ETUDE DES PALEOENVIRONNEMENTS
Comme nous l’avons vu précédemment, à partir des unités biostratigraphiques
proposées par de Vos et al. (1982), des reconstitutions paléoenvironnementales ont été également proposées. Pour les unités de Trinil, de Kedung Brubus et Ngandong les grands bovidés prédominent en nombre de restes ce qui révèle un milieu forestier plutôt ouvert. Pour l’unité de Satir, le paléoenvironnement de type mangrove a été déduit des études palynologiques obtenues à Sangiran dans la formation de Pucangan (Sémah A.M., 1986). Enfin, à Punung, l’arrivée de Pongo et Hylobates reflète un environnement de type forestier humide.
À Sangiran, des reconstitutions paléoenvironnementales ont été réalisées à partir de la palynologie et de la sédimentologie. Nous nous proposons donc à partir des méthodes utilisées couramment pour les mammifères sur des sites européens de reconstituer le paléoenvironnement des espèces fossiles du Pléistocène moyen du dôme de Sangiran.
1. AUTOECOLOGIE
L’autoécologie étudie les relations entre un type d’organisme et les facteurs de son environnement.
Un niveau fossilifère dans un gisement correspond à un ensemble de niches écologiques imbriquées. Or chaque espèce occupe une niche écologique bien précise, définie par de nombreux paramètres dont les valeurs extrêmes constituent autant de facteurs limites : température et humidité du biotope, nature du paysage végétal, orographie, régime alimentaire, adaptations locomotrices, taille des individus, caractéristiques éthologiques (Delpech & Guérin, 1996). Les niches écologiques des espèces existant encore à l’heure actuelle sont présumées n’avoir pas changé. On peut avoir une idée de celles des espèces disparues en prenant en compte les éléments suivants :
- Analogies avec les formes actuelles les plus proches, - Données de l’anatomie fonctionnelle : hypsodontie ou brachyodontie des dents
jugales, port de tête haut ou bas, rapports des segments de membres, robustesse des os des extrémités, etc,

Troisième chapitre : Etude des paléoenvironnements.
- 150 -
150
- Données fournies par les faunes et les flores habituellement associées aux espèces considérées (Delpech & Guérin, 1996).
À Java, la plupart des espèces du Pléistocène moyen ont disparu et il est donc plus
difficile de tenter de reconstituer leur milieu de vie. Néanmoins, si nous essayons de rapprocher ces espèces fossiles des espèces actuelles sauvages et du milieu de vie de ces dernières, nous pouvons tenter une première approche des paléoenvironnements en suivant une démarche actualiste.
1.1. Données sur les espèces actuelles
Les données sur l’écologie actuelle en Indonésie peuvent donc apporter des informations sur le milieu de vie des grandes familles du Pléistocène moyen à Java. Sur nos sites, les grands ordres de mammifères présents sont les proboscidiens, les périssodactyles, les artiodactyles, les carnivores et les primates.
Les informations sur les espèces actuelles ont été tirées principalement des ouvrages de Veevers (1989) et de Corbet et Hill (1992). Nous avons vu précédemment que le nombre d’espèces de petits mammifères (masse < 10 kilogrammes) est beaucoup plus important que celui des grands mammifères. Malheureusement, sur les sites du dôme de Sangiran étudiés les restes de petits mammifères ne se sont pas conservés ou n’ont pas été repérés lors des fouilles.
Dans ces conditions, nous avons tenté de comparer la diversité actuelle de Java et la diversité fossile de Sangiran (au Pléistocène moyen) au niveau des ordres de grands mammifères. Nous avons comparé le nombre de genres présents dans chaque ordre en commun (figure 42). Nous constatons que les artiodactyles et les périssodactyles (pour Tanjung) représentaient à Sangiran une proportion plus importante qu’actuellement à Java alors que cette tendance s’inverse pour les carnivores et les primates.

Troisième chapitre : Etude des paléoenvironnements.
- 151 -
151
Actuel
21%
45%
31%
3%
Primates
Carnivora
Perissodactyla
Artiodactyla
Tanjung14%
0%14%
72%
Sendang Busik20% 0%
0%
80% Figure 42: Proportions relatives des ordres actuels et fossiles des mammifères de Java en nombre
d’espèces. (d’après Corbet & Hill, 1992)
Nous pouvons penser que ces différences entre l’actuel et le fossile sont dues essentiellement à l’augmentation de la pression anthropique qui favorise le développement d’espèces de taille moins importante et d’espèces ne faisant pas l’objet d’une prédation.
Notre but n’est pas de dresser un inventaire exhaustif de toutes les espèces actuelles de Java. En effet, nous avons choisi de regrouper les informations écologiques utiles pour l’association définie au Pléistocène moyen sur Sangiran.
Dans un premier temps, nous nous sommes intéressés aux herbivores actuels. Les herbivores en général sont plus sensibles aux variations de l’environnement puisque la végétation qu’ils consomment dépend directement du substrat et du climat.
1.1.1. Proboscidiens
Pour les proboscidiens, seule l’espèce Elephas maximus Linné, 1758 (= éléphant d’Asie) persiste en Indonésie aujourd’hui. Cette espèce se retrouve à Sumatra et Kalimantan. La taille au garrot d’Elephas maximus peut atteindre 3 mètres et son poids 5 tonnes. Il se nourrit de feuilles ou de jeunes pousses de divers palmiers, de plantes grimpantes ou d’herbe. Il vit en bordure de forêt à proximité de l’eau. Il a besoin de se baigner régulièrement.
Les Stegodon ont, eux, complètement disparu. Ils étaient confinés à des régions au climat tropical, subtropical ou tempéré chaud (van den Bergh, 1999). Ils étaient des brouteurs

Troisième chapitre : Etude des paléoenvironnements.
- 152 -
152
forestiers qui se sont développés dans une région forestière ou semi-forestière orientale comme l’Inde (Burma), la Chine, le Japon et plus au sud Java, Bornéo, et les Philippines (Osborn, 1942).
Les proboscidiens d’Indonésie semblent donc caractériser un milieu forestier ou en limite à proximité d’un cours d’eau.
1.1.2. Périssodactyles
L’ordre des périssodactyles, et plus particulièrement la famille des rhinocerotidés, est
représenté, actuellement, dans l’archipel indonésien et en Inde par les deux espèces suivantes : Rhinoceros sondaicus (= rhinocéros de la Sonde) et Rhinoceros unicornis (= rhinocéros de l’Inde).
Rhinoceros sondaicus Desmarest, 1822
Depuis 1930, il n’est présent qu’à Java Ouest (Ujung Kulon). Sa taille est inférieure à celle de Rhinoceros unicornis, sa taille au garrot est de 1,75 mètre et son poids d’environ 2 tonnes. Le rhinocéros unicorne javanais broute des buissons, des feuilles, de jeunes pousses et aussi des épineux. Il vit dans la jungle dense à proximité de la mer (Hoogerwerf, 1970). Il a besoin d’eau, de boue et d’ombre (figure 43).
Rhinoceros sondaicus sivasondaicus présent au Pléistocène moyen à Java serait une sous-espèce de ce Rhinoceros sondaicus. Les dents des espèces fossiles semblent plus volumineuses que celles des espèces actuelles, de même les métapodes et le radius sont de plus grande taille alors que les humérus, les fémurs et les tibias sont de plus petite taille que ceux des espèces actuelles (Guérin, 1980).
Rhinoceros unicornis Linné, 1758 Il est aussi appelé le rhinocéros unicorne de l’Inde. C’est le plus grand des rhinocéros
actuels, sa taille au garrot est de 2 mètres et son poids varie de 3 à 4 tonnes. Il se nourrit d’herbes, de bourgeons et de plantes tendres. Aujourd’hui, il vit dans des réserves du Nord de l’Inde et au Népal dans la forêt dense ou dans les zones marécageuses à, proximité d’un point d’eau (figure 43).
Au Pléistocène moyen, Rhinoceros unicornis kendengindicus est présent sur l’île de Java. Cependant, l’espèce Rhinoceros unicornis a disparu d’Indonésie mais reste présente sporadiquement en Inde et au Népal. La forme fossile retrouvée à Java : Rhinoceros unicornis kendengindicus, est identique au niveau du crâne à l’actuelle mais les jugales sont plus hypsodontes et de forme différente (Guérin, 1980).
De même, que les proboscidiens, les rhinocérotidés indonésiens ou indiens caractérisent un milieu forestier à proximité d’un point d’eau.

Troisième chapitre : Etude des paléoenvironnements.
- 153 -
153
1.1.3. Artiodactyles
Aujourd’hui, à Java, les artiodactyles sont représentés par les familles des bovidés, des cervidés, des suidés et des tragulidés. La famille des bovidés est représentée, pour sa part, par Bos javanicus (= banteng), Bubalus bubalis (= arnee), Capra hircus (= chèvre) et Ovis aries (= mouton). Ces deux dernières espèces sont domestiques et ont été amenées sur l’île par les hommes. Les deux autres espèces à l’état sauvage, vivent dans des biotopes particuliers.
Bos javanicus d'Alton, 1823 Le banteng se trouve actuellement à Java, Bali et Kalimantan. Sa hauteur au garrot
varie entre 1,30 et 1,70 mètre et son poids entre 500 et 900 kilogrammes. C’est un animal brouteur qui se retrouve dans les parcs et les forêts. Il vit souvent dans les forêts de bambous, où il mange les jeunes pousses et les jeunes feuilles, ou dans des forêts denses avec des trouées. Cette espèce a été domestiquée.
Bos javanicus descendrait de Bibos palaesondaicus du Pléistocène (Hooijer, 1958).
Bubalus bubalis Linné, 1758 Très répandu dans le monde. Sa hauteur au garrot est d’environ 1,80 mètre et son
poids varie entre 500 et 1000 kilogrammes. Il vit près ou dans les grandes rivières des jungles ou dans les marécages (figure 43). Cette espèce a également été domestiquée. La taille de cette espèce est comparable à celle de Bubalus palaeokerabau mais ces chevilles osseuses sont moins imposantes que celles de Bubalus palaeokerabau qui peuvent atteindre 275 centimètres d’envergure (Hooijer, 1958). Néanmoins, Bubalus bubalis est d’une taille supérieure au buffle dosmestique de Java.
La taille de Duboisia santeng correspondrait plus ou moins à celle de l’anoa actuel (van den Bergh, 1988). L’anoa ou Bubalus depressicornis Smith, 1827 se retrouve actuellement aux Sulawesi et à Buru, mais pas à Java. Sa hauteur au garrot est d’environ 1 mètre et son poids de 200 kilogrammes. Du point de vue phylogénétique, Duboisia santeng est plus proche de Boselaphus tragocamelus (= nilgai). Du point de vue morphologique, Duboisia santeng est proche de Tetraceros quadricornis (= antilope à quatre cornes) et ceci serait lié à la nature identique de leur biotope : les forêts galeries le long des rivières dans des zones ouvertes (van den Bergh, 1988).
Boselaphus tragocamelus Pallas, 1766
Il vit actuellement en Amérique du nord et en Asie du sud. Sa hauteur au garrot varie de 119 à 150 centimètres et son poids de 109 à 288 kilogrammes. Il mange principalement de

Troisième chapitre : Etude des paléoenvironnements.
- 154 -
154
l’herbe. Il vit dans des milieux ouverts et secs. En Inde, il se retrouve au pied des montagnes de l’Himalaya.
Tetraceros quadricornis de Blainville, 1816 Il vit en Inde et au Népal. Sa hauteur au garrot varie de 55 à 65 centimètres et son poids de 15 à 25 kilogrammes. Il est brouteur et se nourrit d’herbe, de pousses et de fruits. Il vit dans la forêt ouverte à proximité d’un point d’eau (figure 43).
Le banteng et l’antilope à quatre cornes caractérisent donc un milieu forestier alors que l’arnee est caractéristique d’un milieu plus ouvert où l’eau est omniprésente.
Pour ce qui est de la famille des cervidés, les espèces javanaises répertoriées sont les suivantes : Cervus (Rusa) timorensis (= rusa ou sambar de la Sonde) de Blainville 1822, Axis kuhlii Müller et Schlegel, 1844 et Muntiacus muntjak Zimmermann, 1780 (= muntjak). Ces espèces vivent dans des biotopes caractéristiques et ne sont pas réparties de la même façon dans tout l’archipel indonésien. En ce qui concerne la sous-famille des cervinae, seuls Cervus (Rusa) timorensis et Axis kuhlii sont encore présents à Java.
Cervus (Rusa) timorensis de Blainville 1822 Il est retrouvé à Java, Bali, Flores, Timor, Sulawesi et Maluku. Sa taille au garrot varie
de 83 à 110 centimètres et son poids de 70 à 160 kilogrammes. Il se nourrit d’herbe et de feuilles. Le cerf Rusa vit dans des espaces plus secs et plus ouverts que les autres espèces de cervidés. Il vit dans les plaines herbeuses, les collines parsemées d’arbres et de buissons ou dans la forêt ouverte (figure 43).
Axis kuhlii Müller et Schlegel 1844 Il se trouve actuellement uniquement sur l’île de Bawean. Sa hauteur au garrot varie
de 65 à 70 centimètres et son poids de 35 à 60 kilogrammes. Le cerf axis consomme presque uniquement de l’herbe. Il vit sur des collines boisées ou occasionnellement dans des plantations de tecks (figure 43). Ces données sur les cervidés actuels d’Indonésie permettent de mettre en évidence leur milieu de vie. Le Rusa semble caractériser un milieu forestier plus ouvert et plus sec qu’Axis.
En ce qui concerne la famille des suidés, les espèces Sus scrofa vittatus (= sanglier), Sus domesticus (= cochon) et Sus verrucosus sont présentes actuellement en Indonésie. Sus domesticus Linné, 1758 est l’espèce domestiquée par l’homme et ne caractérise donc pas un milieu particulier.

Troisième chapitre : Etude des paléoenvironnements.
- 155 -
155
Sus scrofa vittatus Linné, 1758 Il est présent partout en Malaisie et en Indonésie. Sa hauteur au garrot est de 1 mètre et
son poids varie de 100 à 150 kilogrammes. Ces animaux qui mangent toutes sortes de nourriture et se reproduisent en grand nombre s’adaptent à tous les milieux (figure 43).
Sus verrucosus Müller, 1840
Il se retrouve à Java, à Bawean et à Madura. Sa hauteur au garrot varie de 70 à 90 centimètres et son poids de 44 à 108 kilogrammes. Il est omnivore. Il vit dans la forêt secondaire à une altitude inférieure à 800 m d’altitude. C’est une espèce menacée. Par conséquent, les suidés ont une faculté d’adaptation importante, mais ils sont présents plus souvent dans les milieux forestiers. Enfin, la famille des hippopotamidés n’est plus représentée en Asie, de nos jours, bien qu’elle ait été une famille importante durant tout le Pléistocène (Dennell, 2005). Actuellement, seul Hippopotamus amphibius Linné, 1758 (hippopotame de grande taille) et Choeropsis liberiensis Morton, 1844 (hippopotame nain de la forêt équatoriale) sont présents en Afrique. Hippopotamus amphibius vit dans des prairies à herbe rase la nuit et dans des bourbiers, des rivières, des lacs le jour (Faure, 1983). Les hippopotames sont très liés à la présence d’eau. Leur physiologie leur impose de rester immergés plusieurs heures par jour. Ils caractérisent un milieu aquatique : fleuves, lacs, marais et constituent donc un bon indicateur d’environnement. Les jugales sont brachyodontes bunodontes. Leur émail est très épais et permet donc la consommation des graminées en dépit de la brachyodontie (Guérin & Patou-Mathis, 1996).
1.1.4. Carnivores
Les carnivores sont assez rares dans nos assemblages. Les carnivores ne sont eux pas directement liés à la végétation car ils se situent en bout de chaîne trophique et dépendent donc du type de proies qu’ils consomment. La hyène a disparu de l’Asie du sud-est et n’est présente actuellement qu’en Afrique et en Inde. La hyène n’est pas caractéristique d’un milieu de vie, elle est ubiquiste.
Les espèces carnivores actuelles présentes à Java sont : Panthera tigris, Panthera pardus, Prionailurus bengalensis, Herpestes javanicus, Arctictis binturong, Paradoxurus hermaphroditus, Prionodon linsang, Viverricula indica, Aonyx cinerea, Lutrogale perspicillata, Mydaus javanensis, Martes flavigula, Mustela lutreolina et Cuon alpinus. Ces espèces se retrouvent le plus souvent dans un milieu forestier.
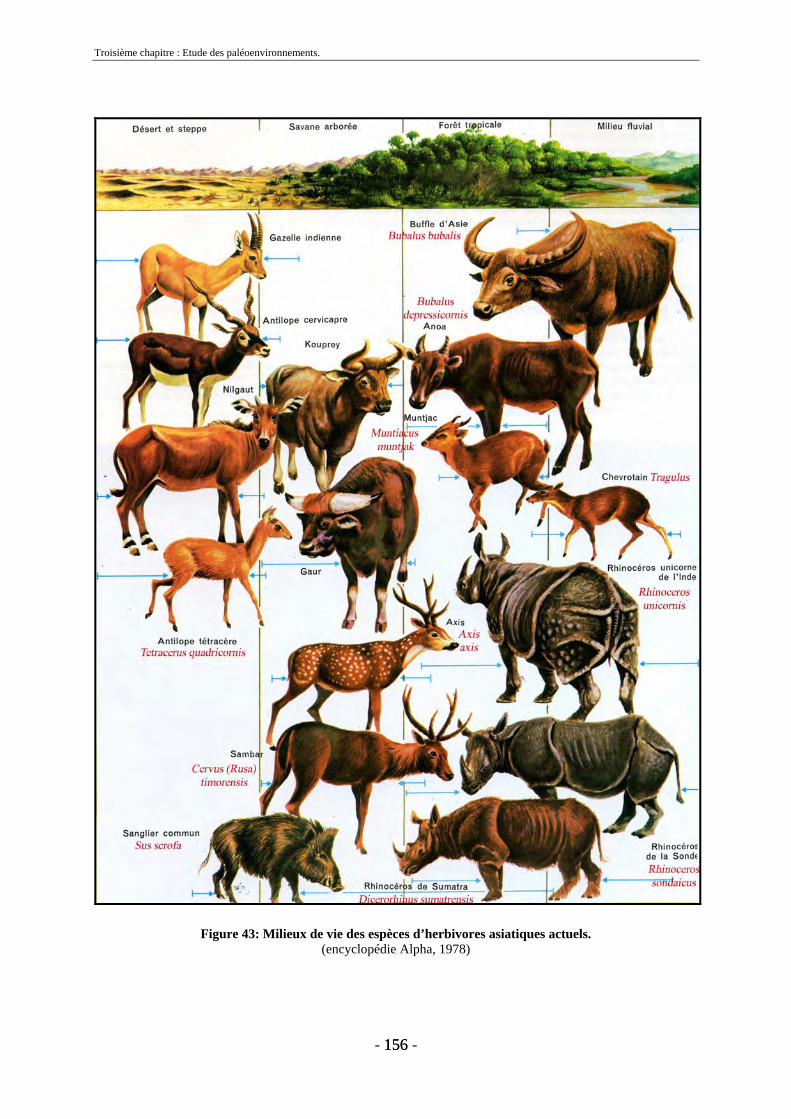
Troisième chapitre : Etude des paléoenvironnements.
- 156 -
156
Figure 43: Milieux de vie des espèces d’herbivores asiatiques actuels. (encyclopédie Alpha, 1978)

Troisième chapitre : Etude des paléoenvironnements.
- 157 -
157
1.1.5. Reptiles
Hormis les mammifères, les reptiles sont de bons indicateurs de milieu. En effet, ce sont des vertébrés ectothermes c’est-à-dire que la température interne de leur corps est déterminée par la température de l’environnement (Bailon & Rage, 1992). Ils peuvent contribuer de façon importante à la reconstitution des paléoenvironnements dans lesquels vivait Homo erectus. La présence des crocodiles et des tortues indique donc la proximité d’un cours d’eau ou d’un lac. De même, la présence des poissons et des gastéropodes confirme celle de l’eau.
1.1.6. Conclusion
En conclusion, la majorité des espèces actuelles de mammifères de Java qui nous
intéressent, exceptée la hyène, caractérisent un milieu forestier plus ou moins ouvert. De plus, les éléphantidés, les hippopotamidés et le buffle sont de bons indicateurs de la présence d’un cours d’eau, d’un lac ou d’un marais à proximité.
Avec ces résultats, nous pouvons proposer des paléoenvironnements pour les
différents sites considérés. À Tanjung 63-64, les deux espèces de bovidés suggèrent un milieu forestier ouvert
(Bibos et Bubalus) de même que l’Elephas. Seules les espèces Axis et Rhinoceros caractérisent un milieu forestier plus dense. Les espèces présentes sur le site de Tanjung 63-64 semblent caractériser un milieu forestier assez ouvert et proche d’un cours d’eau car nous notons la présence du Bubalus, du Rhinoceros, de l’Elephas, qui sont des espèces aquaphiles, ainsi que la présence de reptiles.
À Tanjung 82, les espèces Rhinoceros, Axis et Sus évoquent un milieu forestier alors que les espèces Bubalus et Duboisia reflètent un milieu plus ouvert. Les espèces des niveaux de 82 caractérisent un milieu peu différent de celui du niveau 63-64.
À Sendang Busik, Stegodon, Duboisia et Cervus suggèrent plutôt un milieu forestier ouvert alors qu’Axis caractérise un milieu forestier assez dense. L’espèce Hexaprotodon sivalensis caractérise un milieu aquatique et des restes de tortue et de crocodiles ont également été observés. Le paléoenvironnement de Sendang Busik semble assez proche de celui de Tanjung.
À Ngrejeng, nous avons quatre espèces déterminables : deux espèces de milieu forestier dense (Axis et Sus) et deux de milieu forestier ouvert (Cervus et Duboisia). Nous pouvons donc proposer un paléoenvironnement de type mosaïque. Cependant l’origine stratigraphique des restes étant peu précise (formations de Kabuh et de Pucangan) il est difficile de tenter une reconstitution du paléoenvironnement sur des dépôts recouvrant une

Troisième chapitre : Etude des paléoenvironnements.
- 158 -
158
période de temps si importante et sachant que les environnements de ces deux formations sont très différents.
À Grogol et à Dayu, nous n’avons pas pu proposer une reconstitution de paléoenvironnements par manque de données.
À Bukuran, nous observons la présence de deux espèces typiques du milieu forestier dense (Axis et Sus), d’une espèce typique d’un milieu forestier ouvert (Duboisia), d’une espèce ubiquiste (Pachycrocuta) et d’une aquatique (Hexaprotodon). L’interprétation est assez délicate et nous pouvons penser que le paléoenvironnement du site de Bukuran était assez proche d’un milieu forestier avec des zones plus ou moins ouvertes. Néanmoins, comme à Ngrejeng l’origine stratigraphique des restes est peu précise, il est donc difficile de tenter une reconstitution du paléoenvironnement.
L’application de l’autoécologie est délicate. En effet, le climat de cette zone a changé
depuis le Pléistocène et l’environnement a changé car la pression anthropique est très importante actuellement. La plupart des espèces sauvages actuelles d’Indonésie sont en voie d’extinction notamment du fait de la pression anthropique. L’augmentation de la population de l’île de Java a réduit très considérablement la taille et le nombre des niches écologiques disponibles pour les espèces sauvages. Certaines espèces se cantonnent dans des zones refuges afin de se protéger. Le muntjak, par exemple, vit actuellement en altitude à partir de 2500 mètres à Java alors qu’il se retrouve dans toutes les îles de la Sonde à toutes les altitudes (Corbet & Hill, 1992). Par conséquent, la majorité de ces espèces se situent maintenant dans des réserves naturelles afin d’être protégées de l’influence anthropique.
2. SYNECOLOGIE
La synécologie étudie l’écologie des communautés et des interrelations entre une communauté et son environnement. Pour tenter une reconstitution du paléoenvironnement, il existe également des méthodes synécologiques : le cénogramme, le diagramme écologique et le spectre d’association faunique.
2.1. Cénogramme
Cette méthode a été créée par Valverde (1964) puis modifiée dans les années 80 (Legendre & Sudre, 1984 ; Legendre, 1986 et 1989). Le cénogramme est utilisé pour préciser l’environnement d’une couche archéologique. Le principe de cette méthode est de classer les mammifères par ordre de poids décroissant sans tenir compte du groupe trophique. Le calcul

Troisième chapitre : Etude des paléoenvironnements.
- 159 -
159
du poids des espèces se fait à l’aide d’une relation allométrique étroite existante entre la surface de la première molaire inférieure et le poids corporel de l’animal.
Cette méthode n’est pas applicable dans notre étude car nous ne possédons pas tous les types d’espèces. En effet, nous n’avons pas retrouvé de restes de micromammifères dans notre matériel ce qui biaise les classes de poids. En outre, du fait de l’insularité de Java, les espèces qui y sont présentes évoluent d’une manière différente des espèces continentales. En effet, le plus souvent nous assistons à une diminution en taille des espèces continentales dans un milieu insulaire. Ce phénomène sera étudié en détail plus loin.
2.2. Histogrammes écologiques Cette méthode a été proposée en premier lieu pour étudier des populations de mammifères actuels (Fleming, 1973). Ensuite, elle fut appliquée à des fossiles (Andrews et al., 1979) puis modifiée (Guérin & Faure, 2002). Le principe est de caractériser les sites par un ensemble de quatre histogrammes prenant en considération différents paramètres (tableaux 59, 60 et 61):
-un histogramme taxinomique (figures 44, 45 et 46) comportant huit classes (R : Rongeurs, I : Insectivores, Pri : Primates, Ar : Artiodactyles, C : Carnivores, Per : Périssodactyles, Pro : Proboscidiens, A : Autres ordres), -un histogramme des masses (figures 44, 45 et 46) comportant sept classes (AB : moins de 1 kilogramme, C : de 1 à 10 kilogrammes, D : de 10 à 45 kilogrammes, E : de 45 à 100 kilogrammes, F : de 100 à 200 kilogrammes, G : de 200 à 1000 kilogrammes et H : plus de 1000 kilogrammes), -un histogramme des adaptations alimentaires (figures 44, 45 et 46) comportant six classes (I : mangeurs d’insectes, F : frugivores et granivores, HB : herbivores brachyodontes, HH : herbivores hypsodontes, C : carnassiers et O : omnivores), -un histogramme des adaptations locomotrices (figures 44, 45 et 46) comportant six classes (GTf : grands mammifères terrestres de type forestier, Gtu : grands mammifères terrestres de type ubiquiste, GTc : grands mammifères terrestres de type coureur, PT : petits mammifères, Gr-Ar : grimpeurs et arboricoles, Aq : aquatiques, Ae : aériens, Fo : fouisseurs).
La méthode des histogrammes écologiques peut s’appliquer partiellement sur nos sites. Nous n’avons pris en compte pour cette méthode que les sites de Tanjung et Sendang Busik. En effet, ce sont les deux seuls sites dont la provenance stratigraphique est unique
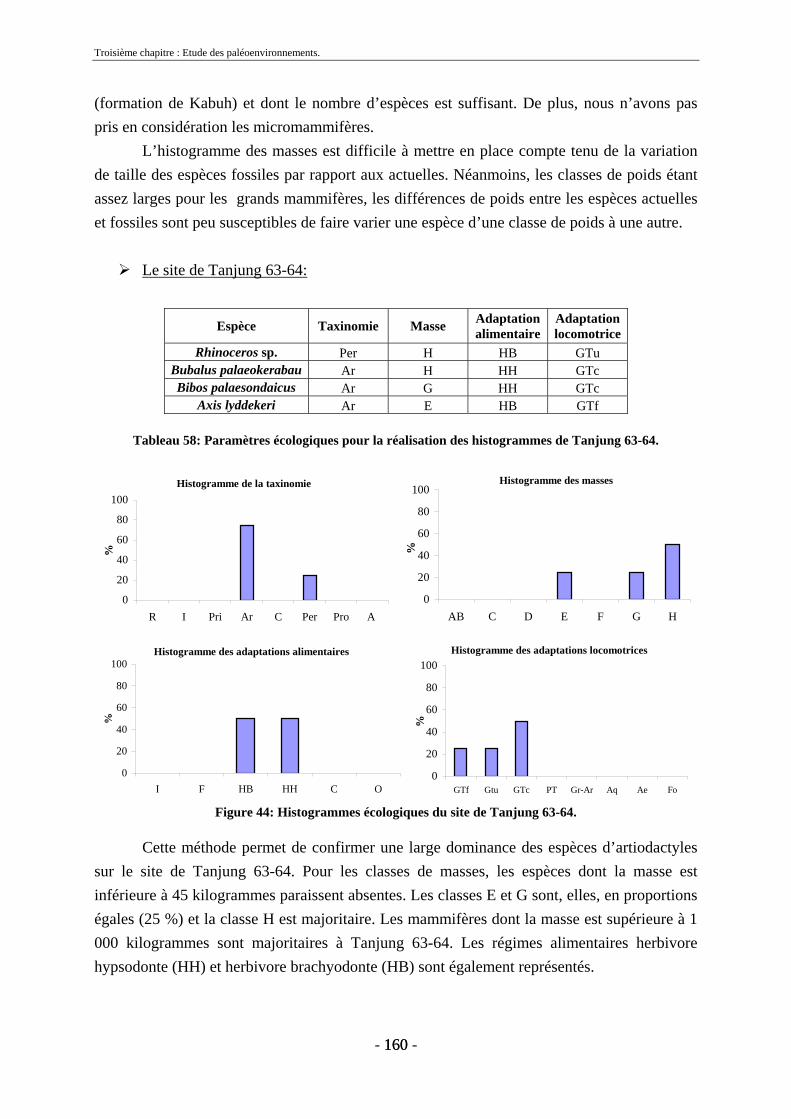
Troisième chapitre : Etude des paléoenvironnements.
- 160 -
160
(formation de Kabuh) et dont le nombre d’espèces est suffisant. De plus, nous n’avons pas pris en considération les micromammifères.
L’histogramme des masses est difficile à mettre en place compte tenu de la variation de taille des espèces fossiles par rapport aux actuelles. Néanmoins, les classes de poids étant assez larges pour les grands mammifères, les différences de poids entre les espèces actuelles et fossiles sont peu susceptibles de faire varier une espèce d’une classe de poids à une autre.
Le site de Tanjung 63-64:
Espèce Taxinomie Masse Adaptation alimentaire
Adaptation locomotrice
Rhinoceros sp. Per H HB GTu Bubalus palaeokerabau Ar H HH GTc Bibos palaesondaicus Ar G HH GTc
Axis lyddekeri Ar E HB GTf
Tableau 58: Paramètres écologiques pour la réalisation des histogrammes de Tanjung 63-64.
Histogramme de la taxinomie
0
20
40
60
80
100
R I Pri Ar C Per Pro A
%
Histogramme des masses
0
20
40
60
80
100
AB C D E F G H
%
Histogramme des adaptations alimentaires
0
20
40
60
80
100
I F HB HH C O
%
Histogramme des adaptations locomotrices
0
20
40
60
80
100
GTf Gtu GTc PT Gr-Ar Aq Ae Fo
%
Figure 44: Histogrammes écologiques du site de Tanjung 63-64.
Cette méthode permet de confirmer une large dominance des espèces d’artiodactyles
sur le site de Tanjung 63-64. Pour les classes de masses, les espèces dont la masse est inférieure à 45 kilogrammes paraissent absentes. Les classes E et G sont, elles, en proportions égales (25 %) et la classe H est majoritaire. Les mammifères dont la masse est supérieure à 1 000 kilogrammes sont majoritaires à Tanjung 63-64. Les régimes alimentaires herbivore hypsodonte (HH) et herbivore brachyodonte (HB) sont également représentés.

Troisième chapitre : Etude des paléoenvironnements.
- 161 -
161
Tout ceci nous donne des informations sur le paléoenvironnement de ces espèces au moment de leur mort. Ce milieu devait être forestier mais assez ouvert du fait de l’importance des espèces herbivores hypsodontes, des espèces de grande taille et de type coureur.
Le site de Tanjung 82:
Espèces Taxinomie Masse Adaptation alimentaire
Adaptation locomotrice
Elephas sp. Pro H HH GTu Rhinoceros sp. Per H HB GTu
Bubalus palaeokerabau Ar H HH GTc Duboisia santeng Ar F HH GTc Axis lydekkeri* Ar E HB GTf
Sus sp. Ar E O GTf
Tableau 59: : Paramètres écologiques pour la réalisation des histogrammes de Tanjung 82.
Histogramme de la taxinomie
0
20
40
60
80
100
R I Pri Ar C Per Pro A
%
Histogramme des masses
0
20
40
60
80
100
AB C D E F G H
%
Histogramme des adaptations alimentaires
0
20
40
60
80
100
I F HB HH C O
%
Histogramme des adaptations locomotrices
0
20
40
60
80
100
GTf Gtu GTc PT Gr-Ar Aq Ae Fo
%
Figure 45: Histogrammes écologiques du site de Tanjung 82.
À Tanjung 82, les artiodactyles sont dominants par rapport aux périssodactyles et aux
proboscidiens. Pour les classes de masses, les espèces dont la masse est inférieure à 45 kilogrammes paraissent absentes et la classe H est majoritaire comme dans les niveaux de 63-64. Les mammifères dont le poids est supérieur à 1 000 kilogrammes sont nombreux à Tanjung 82. Le régime alimentaire herbivore hypsodonte (HH) est mieux représenté que le régime herbivore brachyodonte (HB). Les grands mammifères de type forestier, coureur et ubiquiste sont représentés en proportions égales. Le milieu de vie de ces espèces devait être
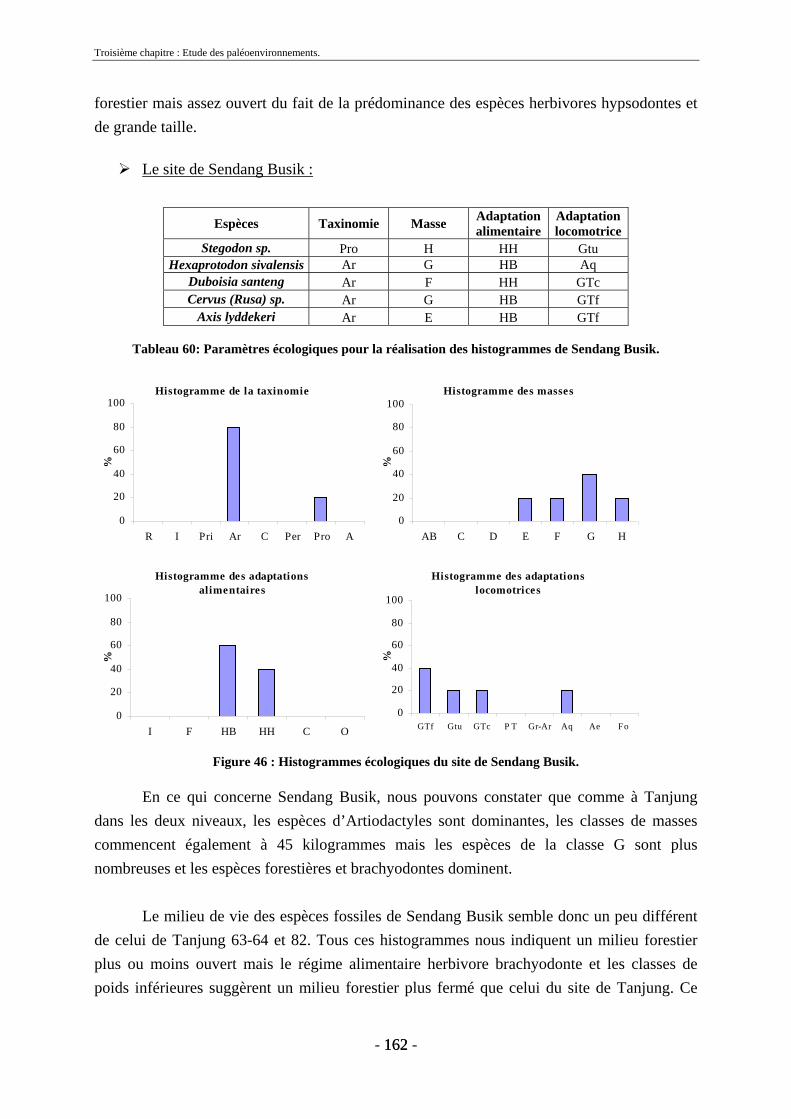
Troisième chapitre : Etude des paléoenvironnements.
- 162 -
162
forestier mais assez ouvert du fait de la prédominance des espèces herbivores hypsodontes et de grande taille.
Le site de Sendang Busik :
Espèces Taxinomie Masse Adaptation alimentaire
Adaptation locomotrice
Stegodon sp. Pro H HH Gtu Hexaprotodon sivalensis Ar G HB Aq
Duboisia santeng Ar F HH GTc Cervus (Rusa) sp. Ar G HB GTf
Axis lyddekeri Ar E HB GTf
Tableau 60: Paramètres écologiques pour la réalisation des histogrammes de Sendang Busik.
Histogramme de la taxinomie
0
20
40
60
80
100
R I Pri Ar C Per Pro A
%
Histogramme des masses
0
20
40
60
80
100
AB C D E F G H
%
Histogramme des adaptations alimentaires
0
20
40
60
80
100
I F HB HH C O
%
Histogramme des adaptations locomotrices
0
20
40
60
80
100
GTf Gtu GTc P T Gr-Ar Aq Ae Fo
%
Figure 46 : Histogrammes écologiques du site de Sendang Busik.
En ce qui concerne Sendang Busik, nous pouvons constater que comme à Tanjung dans les deux niveaux, les espèces d’Artiodactyles sont dominantes, les classes de masses commencent également à 45 kilogrammes mais les espèces de la classe G sont plus nombreuses et les espèces forestières et brachyodontes dominent. Le milieu de vie des espèces fossiles de Sendang Busik semble donc un peu différent de celui de Tanjung 63-64 et 82. Tous ces histogrammes nous indiquent un milieu forestier plus ou moins ouvert mais le régime alimentaire herbivore brachyodonte et les classes de poids inférieures suggèrent un milieu forestier plus fermé que celui du site de Tanjung. Ce

Troisième chapitre : Etude des paléoenvironnements.
- 163 -
163
milieu était peut-être plus proche d’une forêt type galerie au bord d’un cours d’eau avec la présence de l’hippopotame.
2.3. Spectre d’association faunique
La méthode du spectre d’association faunique a été utilisée jusqu’à maintenant pour tenter de reconstituer des environnements préhistoriques européens en proposant des regroupements d’herbivores et de carnivores à partir de données actualistes mais aussi spécifiques du Pléistocène (Delpech, 1983 ; Auguste, 1995). Elle peut être adaptée à de nouveaux milieux car nous nous localisons dans une zone tropicale où les climats sont et ont été très différents de ceux d’Europe.
Afin d’établir les spectres d’associations fauniques pour certains sites du dôme de Sangiran, nous avons proposé quatre types de milieu à partir des espèces présentes sur les sites (figure 47):
Le milieu de forêt tropicale humide : GMFT
Ce milieu est composé d’une forêt dense et humide (rain forest) constituée d’essences
comme Dipterocarpus, Podocarpus, Castanopsis, Altingia, Quercus (Sémah A.M., 1986). Ce milieu est fermé, il ne peut donc pas abriter d’espèces de mammifères de grande taille ou d’espèces avec des appendices frontaux très développés. Les espèces qui vivaient dans ce type de milieu sont considérées comme des espèces du groupe de milieu de forêt tropicale (GMFT). Sur les sites de notre étude nous avons classé les espèces Axis lydekkeri et Rhinoceros sp. dans ce groupe.
Le milieu de forêt décidue ou de savane arborée : GMFS
Lorsque la saison sèche dure plus de quatre mois la forêt tropicale humide fait place à
la forêt décidue ou forêt de mousson (monsoon forest) et si la saison sèche continue plus longtemps cette forêt passe à une savane arborée (Sémah A.M., 1986). La forêt de mousson est composée d’espèces décidues qui perdent périodiquement leur feuillage pour s’adapter aux conditions climatiques. La savane arborée est constituée de zones herbeuses et de zones plus boisées. L’importance des herbacées dans ce milieu est plus forte, les arbres et les fougères sont beaucoup moins présents et les taxons tels que les Mimosaceae, Dodanaea, Poaceae, Fabaceae se développent ainsi que les arbres de forêt claire comme Mallotus (Sémah A.M., 1986). Ce milieu est plus ouvert et abrite des espèces de mammifères de plus grande taille ou possédant des appendices frontaux développés. Les espèces qui vivaient dans ce type de milieu ouvert sont considérées comme appartenant au groupe de milieu de forêt décidue ou de

Troisième chapitre : Etude des paléoenvironnements.
- 164 -
164
savane arborée (GMFS). Nous avons répertorié les espèces Cervus (Rusa) sp., Bibos palaesondaicus et Duboisia santeng dans cette catégorie.
Le milieu de forêt galerie : GMFG
Ce milieu est composé d’une forêt se situant au bord des cours d’eau ou ripisylve. Elle
est composée d’espèces vivant dans des zones humides, tels que les Meliaceae, le Callophyllum et le Ficus (Sémah A.M., 1986). Ce milieu se développe le long des cours d’eau et abritent des espèces de mammifères qui se baignent quotidiennement. Les espèces qui vivaient dans ce type de milieu ouvert sont considérées comme appartenant au groupe de milieu de forêt galerie (GMFG). Sur les sites de notre étude nous avons classé les espèces Hexaprotodon sivalensis, Stegodon trigonocephalus, Elephas hysudrindicus et Bubalus palaeokerabau dans ce groupe.
Le milieu ubiquiste : GMU Les espèces de mammifères vivant dans ce milieu ont une grande faculté d’adaptation
et ne sont pas inféodés à un biotope précis. Nous avons classé l’espèce Pachycrocuta brevirostris dans ce groupe.
Tanjung 63-64
25%
50%
25%
Tanjung 82
17%
50%
33%
Sendang Busik
20%
20%60%
GMFS
GMFT
GMFG
Figure 47: Spectres d’association faunique de Tanjung 63-64, Tanjung 82 et de Sendang Busik.
(en NMI)

Troisième chapitre : Etude des paléoenvironnements.
- 165 -
165
Les espèces du milieu forestier tropical représentent une proportion plus importante par rapport aux espèces des autres milieux en nombre d’individus dans les deux niveaux du site de Tanjung. Les deux spectres d’association faunique de Tanjung sont très proches. A Sendang Busik, les espèces du milieu de forêt galerie sont plus nombreuses que celles des milieux de forêt tropicale et de savane arborée en nombre d’individus.
Le milieu de vie des espèces fossiles de Tanjung paraît avoir été plus forestier que celui de Sendang Busik. Ce résultat est différent de celui des histogrammes écologiques basés sur le nombre d’espèces et non sur le nombre d’individus. Néanmoins, la méthode des spectres fauniques nous confirme que le site de Sendang Busik semblait plus proche de la rivière que les niveaux de Tanjung.
2.4. Conclusion
Les méthodes des histogrammes écologiques et du spectre faunique nous permettent de penser que le milieu de vie des espèces fossiles des deux niveaux du site de Tanjung (dans les deux niveaux) était proche d’un milieu forestier assez ouvert à proximité d’un cours d’eau car le nombre de grandes espèces est important ainsi que celui des espèces hypsodontes.
Ces mêmes méthodes à Sendang Busik suggèrent un milieu forestier ouvert mais moins qu’à Tanjung car les espèces de très grande taille ne sont pas majoritaires alors que les espèces brachyodontes le sont. En outre, les espèces vivant à proximité d’un milieu aquatique sont nombreuses. Nous pouvons donc penser que le milieu de vie des espèces fossiles de Sendang Busik était proche d’un milieu forestier type forêt galerie bordant un cours d’eau.
Les méthodes synécologiques classiques utilisées sur des sites préhistoriques
européens sont difficilement adaptables aux sites du dôme de Sangiran. En effet, les climats, les milieux et les espèces sont très différents. Nous avons essayé de modifier ces méthodes afin qu’elles soient mieux adaptées aux sites étudiés. Néanmoins, ces méthodes sont assez approximatives et particulièrement dans les sites de plein air en milieu fluviatile où le transport a pu agir sur les ossements. Par conséquent, le milieu de dépôt des ossements peut-être assez éloigné du milieu de vie des espèces ce qui peut biaiser les résultats. En outre, les espèces déterminables sont peu nombreuses ce qui biaise également les résultats sur les paléoenvironnements.
Afin de confirmer ou d’infirmer nos résultats paléoenvironnementaux obtenus à partir de l’étude des mammifères, nous les avons comparés avec ceux obtenus pour la taphonomie et notamment avec ceux sur le transport des ossements (cf. chapitre suivant) ainsi qu’avec ceux d’autres disciplines.

Troisième chapitre : Etude des paléoenvironnements.
- 166 -
166
3. DONNEES CLIMATIQUES ET PALYNOLOGIQUES
3.1. Palynologie Comme nous l’avons vu précédemment dans les couches de Kabuh, la végétation, dont les caractères évoluent en fonction des oscillations climatiques, reflète des conditions globalement plus sèches et donc un milieu plus ouvert que dans les niveaux de Pucangan. Les espèces de la strate herbacée (= NAP : non arboreal pollen) sont donc plus nombreuses que celles du couvert arboréen (= AP : arboreal pollen) (Sémah A.M., 1986). Les données palynologiques confirment les reconstitutions des paléoenvironnements obtenues à Tanjung et Sendang Busik avec les espèces de mammifères.
En effet, d’après les espèces de mammifères le paléoenvironnement de Sangiran devait être assez proche d’un milieu forestier avec des zones plus ouvertes et le passage d’un cours d’eau.
3.2. Sédimentologie La sédimentologie nous apporte des informations sur l’environnement au moment du dépôt des sédiments. Comme nous l’avons vu précédemment, les différentes couches du dôme de Sangiran nous informent sur le paléoenvironnement de cette zone au cours du temps. Les argiles bleues de Kalibeng supérieur sont caractéristiques d’un milieu de mer peu profonde. Les couches de Pucangan évoquent un milieu palustre ou lacustre. Le Grenzbank montre le début des dépôts continentaux qui se poursuivent dans les couches volcano-sédimentaires de Kabuh où la sédimentation fluviatile prédomine (Lizon-Sureau, 1979 ; Sémah F., 1984 ; Larick et al., 2001). Les restes fossiles des sites que nous avons considérés (Tanjung et Sendang Busik) proviennent des couches de Kabuh. Ces fossiles se sont donc déposés dans un environnement fluviatile. Lizon-Sureau (1979) a pu mettre en évidence des variations du régime fluviatile dans ces couches. À la base, elle constate un courant modéré et régulier et même un court épisode lagunaire. Dans la partie moyenne de Kabuh, une reprise du régime fluviatile est notée suivi de conditions lacustres. Enfin, dans la partie supérieure, le milieu de sédimentation est variable : un régime fluviatile, puis torrentiel et à nouveau fluviatile. Ces variations peuvent être dues en partie au climat mais pas seulement. En effet, la sédimentation lacustre peut-être due aussi à la divagation des lits des cours d’eau ou à la subsidence.
Ces données sédimentologiques nous permettent de confirmer les résultats obtenus avec les espèces de mammifères à Tanjung et Sendang Busik. En effet, sur ces deux sites, les

Troisième chapitre : Etude des paléoenvironnements.
- 167 -
167
espèces de milieu forêt galerie sont en proportions non négligeables (Tanjung) ou même majoritaires (Sendang Busik).
En outre, sur ces deux sites, nous avons découvert des restes de tortue et de crocodile, espèces aquatiques révélant la présence d’un point d’eau à proximité du milieu de dépôt.
Pour ce qui est des sites de Ngrejeng et de Bukuran où les fossiles proviennent des formations de Kabuh et de Pucangan, le milieu était plus ou moins forestier avec toujours un point d’eau à proximité, qui dans la formation de Pucangan, a pu être de type lac ou marais plutôt que rivière comme dans Kabuh.
3.3. Conclusion
La palynologie et la sédimentologie permettent de confirmer les résultats paléoenvironnementaux obtenus à partir de l’étude des espèces de mammifères sur les sites de Tanjung et de Sendang Busik.
À Tanjung, nous pouvons penser que les espèces de mammifères et que donc Homo erectus vivaient dans un milieu mosaïque constitué de zones forestières et de zones plus ouvertes, le tout à proximité d’un cours d’eau type rivière.
À Sendang Busik, de même que pour Tanjung, nous pouvons penser que les espèces de mammifères et Homo erectus évoluaient dans un milieu mosaïque cependant plus proche du cours d’eau qu’à Tanjung. Nous verrons ultérieurement si cette hypothèse peut être confirmée lors de l’étude taphonomique.
4. ENDEMISME ET EXTINCTIONS
4.1. Endémisme Les espèces insulaires voient leur taille varier par rapport aux formes continentales
dont elles descendent (Dermitzakis et al., 1997). Dans ce cadre, Hooijer (1949) a comparé des éléments des faunes de Sumatra et de Java du Plio-Pléistocène avec les faunes actuelles de cette même zone. Les espèces choisies étaient : Dicerorhinus sumatrensis, Rhinoceros sondaicus, Tapirus indicus, Acanthion brachyurus, Panthera tigris et Pongo pygmaeus. Il a constaté que depuis leur immigration sur la plate forme de la Sonde de nombreuses espèces ont diminué en taille ce qui s’est traduit dans certains cas par un changement dans la structure de leurs membres et de leurs pattes. De plus, il a remarqué que cette diminution en taille pendant le Quaternaire avait été plus importante à Java qu’à Sumatra et a conclu que les espèces pionnières de Sumatra devaient être d’une taille plus importante que celles de Java.

Troisième chapitre : Etude des paléoenvironnements.
- 168 -
168
Cependant, van den Bergh (1988, 1999) a remarqué une augmentation de la taille de Duboisia et des Stegodon du Pléistocène inférieur au Pléistocène moyen à Java, de même Hardjasasmita (1987) a constaté cette tendance chez les suidés et Hooijer (1950) chez les hippopotamidés. Ceci s’explique probablement par le fait que le paléoenvironnement a changé du Pléistocène inférieur au Pléistocène moyen et que l’espace s’est ouvert au détriment de la mangrove et ceci a permis une augmentation de la taille de certaines espèces.
4.2. Extinctions Des extinctions catastrophiques sont communes à ces îles, elles peuvent être de
différents types (Dermitzakis, 1997) : - Un changement de faune : une faune continentale avec de grands prédateurs envahit
l’île et provoque l’extinction des nains, - Un appauvrissement très ancien d’une faune endémique insulaire qui est suivi d’une
autre faune distincte mais appauvrie, - À un niveau plus local, des éruptions volcaniques.
Les causes peuvent donc être multiples : catastrophes naturelles, changements climatiques ou arrivées de grands prédateurs ou une combinaison de ces facteurs.
Sur l’île de Java, au Plio-Pléistocène, des renouvellements d’espèces sont observables
au cours de la succession biostratigraphique (Sondaar, 1984 ; van den Bergh et al., 1996, 2001a).
La faune de Satir est considérée comme une faune de type insulaire (de Vos, 1985 ; de Vos et al., 1994 ; Dermitzakis et al., 1997) composée entre autres par l’association caractéristique : Tetralophodon bumiajuensis, Geochelone atlas et Hexaprotodon simplex. L’unité faunique qui lui succède est très différente. Elle est non insulaire. Dans la faune de Ci Saat, le Tetralophodon est remplacé par le Stegodon et l’Hexaprotodon simplex est remplacé par l’H. sivalensis. La tortue, quant à elle, disparaît à l’unité faunique suivante, Trinil H.K. Nous assistons à un renouvellement des espèces de la faune de Satir. Celle-ci est liée à l’arrivée d’espèces et de groupes continentaux comme le tigre, les bovidés, les rhinocerotidés et les suidés. De la faune de Trinil à Kedung Brubus, les espèces ne s’éteignent pas. Mais la faune de Kedung Brubus est beaucoup plus riche que celle de Trinil H.K., témoignant d’une plus longue période de connexion de l’île avec le continent. La faune de Ngandong est peu connue mais semble assez proche, du point de vue de l’adaptation paléoenvironnementale, de celle de Kedung Brubus.
Un deuxième grand renouvellement ou « turnover » a lieu après cette période avec l’arrivée de la faune de Punung. Les faunes caractérisées par l’association Stegodon-Homo erectus observées depuis l’unité de Ci Saat sont remplacées par une faune de forêt tropicale humide caractérisée par l’association Pongo pygmaeus-Homo sapiens.

Troisième chapitre : Etude des paléoenvironnements.
- 169 -
169
Par conséquent, deux grands renouvellements ou « turnover » sont donc notés au Plio-Pléistocène sur l’île de Java (Dermitzakis et al., 1997).
Nous nous sommes intéressés plus précisément à deux espèces disparues aujourd’hui
de l’île de Java et présentes au Pléistocène moyen : Pachycrocuta brevirostris et Rhinoceros unicornis.
Les hyènes n’ont pas été retrouvées dans les dépôts postérieurs à ceux de la faune de Kedung Brubus. Dans une région plus au nord en Thaïlande, Tougard (1998) a remarqué la présence de Crocuta crocuta ultima qui en Chine a remplacé Pachycrocuta brevirostris sinensis. Crocuta crocuta ultima se retrouve également dans des dépôts plus récents en Chine du sud, au Laos et au Cambodge. D’après Kurten (1956), ce remplacement serait écologique, Crocuta crocuta ultima aurait occupé la niche écologique laissée vacante par Pachycrocuta brevirostris suite au refroidissement de 0,44-0,48 Ma. A Java, Pachycrocuta brevirostris n’a pas été remplacée par une autre espèce d’hyène, mais son extinction est peut-être également liée à un refroidissement à cette période.
Rhinoceros unicornis n’est actuellement retrouvé que dans des réserves du nord de l’Inde et au Népal alors qu’il a été observé à Kedung Brubus au Pléistocène moyen (van den Bergh et al., 2001a) et dans les sites de l’Holocène en Thaïlande et en Asie (Tougard, 1998) ce qui montre qu’il était présent dans une grande partie de l’Asie du sud-est. Cette extinction semble donc plus liée à une prédation excessive et à une destruction de son habitat par l’homme qu’à une cause climatique.
Après l’étude des fossiles et de leur paléoenvironnement, il nous a paru essentiel de
considérer ces assemblages du point de vue taphonomique afin de mieux comprendre les liens entre Homo erectus et les mammifères et donc de tenter une approche de son comportement de subsistance.

- 170 -

- 171 -
QUATRIEME CHAPITRE ETUDE TAPHONOMIQUE
La taphonomie a été définie par Efremov (1940) comme la science qui étudie les lois de l’enfouissement des restes animaux et végétaux (Olson, 1980 ; Fosse, 1992). Lyman (1987) a proposé, plus tard, une définition assez claire pour les ossements : « la science qui traite de toutes les histoires post-mortem, pré- et post-enfouissement des restes osseux ».
Actuellement, la taphonomie est divisée en deux parties (Noe-Nygaard, 1977) : - La biostratinomie étudie les processus subis par les restes organiques des animaux et des végétaux de leur mort jusqu’à leur enfouissement dans les sédiments, - L’étude diagénétique étudie les transformations chimiques et physiques affectant les restes après leur enfouissement. À Sangiran, un très grand nombre de fossiles fut découvert en surface ou lors de
fouilles. À ce jour, peu d’études taphonomiques, excepté à Ngebung 2, ont été menées sur des assemblages osseux de ce dôme. Il est alors intéressant de connaître l’origine et l’évolution de ces accumulations qui peuvent avoir subi différentes actions taphonomiques anté ou post-dépositionnelles expliquant la conservation et la fragmentation des ossements.
Dans un premier temps, nous allons aborder la répartition, puis la conservation et la fragmentation des différents assemblages osseux issus des fouilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu afin de comprendre l’origine et l’histoire de ceux-ci en contexte de plein air et en milieu fluviatile.
1. REPARTITION SPATIALE
À Tanjung 63, seul un secteur (II) est indiqué pour l’ensemble des pièces. À Tanjung 64, les restes se répartissent sur 225 carrés représentant une surface de 900 m² si l’on considère que les carrés de fouille sont de 2 mètres sur 2. Nous nous ne possédons pas de données sur l’épaisseur de la fouille. Les restes sont les plus nombreux dans la zone de C à F et de 1 à 5. Les carrés les plus riches sont D1 (41 restes) et E1 (33 restes) qui présentent une fréquence de plus de 30 restes (annexe 6). Toute la partie supérieure de la zone fouillée semble plus riche en restes. Nous n’avons pas de données sur les profondeurs où les fossiles ont été découverts. Au niveau statistique, nous avons testé la répartition des ossements de Tanjung 64 afin de savoir si elle était aléatoire ou pas. Pour ce faire nous avons utilisé la méthode de Dacey (1973) (annexe 7). D’après les résultats obtenus la répartition des

Quatrième chapitre : Etude taphonomique.
- 172 -
ossements sur le site de Tanjung 64 ne serait pas aléatoire. Ceci peut-être lié à différents facteurs : le type de fouille, l’action des carnivores ou de l’homme dans une zone du site ou l’action du transport fluviatile (barre de méandre…). En outre, nous avons testé l’association des restes déterminables et des esquilles sur ce même site et ces deux types de restes sont associés au niveau spatial d’après le test de Dacey (1973) (annexe 7).
À Tanjung 82, les restes se répartissent sur 5 carrés (I à V) et sur une surface de 20 m² si l’on considère que les carrés de fouille sont de 2 mètres sur 2 (annexe 6) et sur des profondeurs allant de 39 à 402 centimètres. Nous observons la même surface à Ngrejeng, Grogol et Dayu (annexe 6). Á Tanjung 82, le carré le plus riche en restes est le carré IV où 153 fossiles ont été découverts. Dans le carré II, seule une bola a été découverte (annexe 11). C’est le seul outil retrouvé sur l’ensemble des sites.
À Sendang Busik I, les restes se répartissent sur 5 carrés (de 4 à 8) mais à Sendang Busik II, ils se répartissent sur 11 carrés (de 1 à 11) et donc sur une surface plus importante de 44 m² si l’on considère des carrés de 2 mètres sur 2 et sur des profondeurs (z) allant de 8 à 557 centimètres. Á Sendang Busik I, les carrés les plus riches sont les carrés 7 et 8 et à Sendang Busik II, les carrés les plus riches sont les carrés 5 et 6 et 8 à 11. Dans tous ces carrés, le nombre de restes est supérieur à 30 restes (annexe 6). Les deux fouilles ont été considérées comme une seule dans la suite de notre étude.
À Ngrejeng, les 5 carrés (I à V) sont riches en restes, seul le carré IV présente 20 à 30 restes alors que les autres ont plus de 30 restes (annexe 6). Les profondeurs des découvertes varient de 36 à 480 centimètres.
À Grogol , les restes se répartissent sur 5 carrés (A1 à A3, et, B1 et B2), les carrés A3 (51 restes) et B2 (111 restes) sont les plus riches en restes (annexe 6). Les découvertes se distribuent sur des profondeurs variant de 22 à 410 centimètres.
À Bukuran, les restes se répartissent sur une surface plus importante d’environ 36 m² si l’on considère des carrés de 2 mètres de côté et sur des profondeurs allant de 73 à 486 centimètres. Des fossiles ont été trouvés sur 12 des 18 carrés au total. Le carré le plus riche est B1-VI avec 47 restes (annexe 6).
Enfin, à Dayu, parmi les cinq carrés, le plus riche ne présente que 20 à 30 restes (annexe 6). Les découvertes se répartissent sur des profondeurs variant de 66 à 541 centimètres.
La répartition spatiale nous montre que suivant les sites, la surface et la richesse des carrés diffèrent. La plupart des sites n’ont été fouillés que sur environ 5 carrés, ceci représente des petites surfaces de fouilles d’environ 20 m². Il est donc plus difficile d’obtenir des informations sur la répartition des ossements sur de telles surfaces. Cette répartition spatiale nous donne une première image de la distribution des fossiles suivant les zones du dôme et sur les méthodes de fouilles de l’époque. Mis à part à Tanjung 64, les surfaces de fouilles sont assez réduites et les durées également (entre 14 et 24 jours).

Quatrième chapitre : Etude taphonomique.
- 173 -
Après avoir analysé la répartition spatiale des restes osseux nous nous sommes intéressés à leur conservation.
2. CONSERVATION
L’étude taxinomique et la répartition spatiale nous ont donné une première image de la nature et de la distribution des restes osseux déterminables sur les différents sites de Sangiran. Nous allons maintenant considérer l’ensemble des restes pour chaque assemblage au niveau qualitatif et quantitatif.
2.1. Quantification des restes
Pour mieux saisir et comprendre l’origine et l’histoire d’un assemblage osseux, il est nécessaire d’effectuer de bons dénombrements. Différentes méthodes de décompte ont ainsi été mises au point pour faciliter le traitement des données (Poplin, 1976 ; Klein & Cruz-Uribe, 1984 ; Grayson, 1979 ; Badgley, 1986a ; Brugal et al. 1994 ; Lyman, 1994).
Le nombre total des restes (NRT ou TNF : Total Number of Fragments) pour chaque collection peut être séparé en plusieurs catégories de restes :
• Le NRDt représente le nombre de restes déterminés taxonomiquement et anatomiquement (= NISP : Number of Identified Specimen) ;
• Le NRDa correspond au nombre de restes déterminés anatomiquement ;
• le NRI (= ND, non déterminable, flake bones) correspond au nombre de restes indéterminés ou esquilles.
Par conséquent, le NRT est égal à la somme de ces trois catégories :
NRT = NRDt + NRDa + NRI
Dans notre étude, le NRDt varie environ entre 15 et 43 % du NRT, le NRDa est lui
compris entre environ 5 et 20 % et le NRI entre 50 et 67 % (tableau 61). Pour Tanjung, les restes sont issus de deux campagnes de fouilles distinctes menées à
18 ans d’intervalle. La situation de la fouille de 1982, aurait été un peu différente de celle de 1963-64, elle serait plus haute au niveau topographique (Koeshardjono, communication personnelle). Nous pouvons nous demander si ces deux assemblages de Tanjung ont eu la même histoire taphonomique. Nous les avons donc considérés comme deux assemblages distincts.
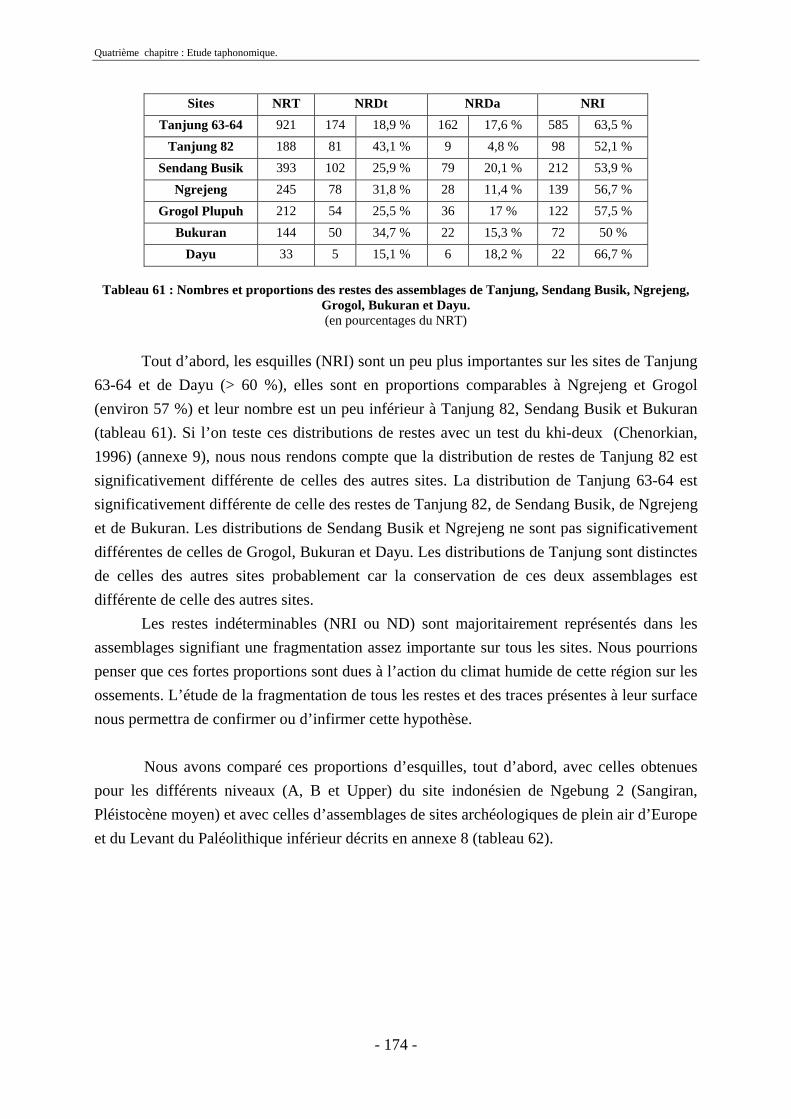
Quatrième chapitre : Etude taphonomique.
- 174 -
Sites NRT NRDt NRDa NRI Tanjung 63-64 921 174 18,9 % 162 17,6 % 585 63,5 %
Tanjung 82 188 81 43,1 % 9 4,8 % 98 52,1 % Sendang Busik 393 102 25,9 % 79 20,1 % 212 53,9 %
Ngrejeng 245 78 31,8 % 28 11,4 % 139 56,7 % Grogol Plupuh 212 54 25,5 % 36 17 % 122 57,5 %
Bukuran 144 50 34,7 % 22 15,3 % 72 50 % Dayu 33 5 15,1 % 6 18,2 % 22 66,7 %
Tableau 61 : Nombres et proportions des restes des assemblages de Tanjung, Sendang Busik, Ngrejeng,
Grogol, Bukuran et Dayu. (en pourcentages du NRT)
Tout d’abord, les esquilles (NRI) sont un peu plus importantes sur les sites de Tanjung
63-64 et de Dayu (> 60 %), elles sont en proportions comparables à Ngrejeng et Grogol (environ 57 %) et leur nombre est un peu inférieur à Tanjung 82, Sendang Busik et Bukuran (tableau 61). Si l’on teste ces distributions de restes avec un test du khi-deux (Chenorkian, 1996) (annexe 9), nous nous rendons compte que la distribution de restes de Tanjung 82 est significativement différente de celles des autres sites. La distribution de Tanjung 63-64 est significativement différente de celle des restes de Tanjung 82, de Sendang Busik, de Ngrejeng et de Bukuran. Les distributions de Sendang Busik et Ngrejeng ne sont pas significativement différentes de celles de Grogol, Bukuran et Dayu. Les distributions de Tanjung sont distinctes de celles des autres sites probablement car la conservation de ces deux assemblages est différente de celle des autres sites.
Les restes indéterminables (NRI ou ND) sont majoritairement représentés dans les assemblages signifiant une fragmentation assez importante sur tous les sites. Nous pourrions penser que ces fortes proportions sont dues à l’action du climat humide de cette région sur les ossements. L’étude de la fragmentation de tous les restes et des traces présentes à leur surface nous permettra de confirmer ou d’infirmer cette hypothèse.
Nous avons comparé ces proportions d’esquilles, tout d’abord, avec celles obtenues
pour les différents niveaux (A, B et Upper) du site indonésien de Ngebung 2 (Sangiran, Pléistocène moyen) et avec celles d’assemblages de sites archéologiques de plein air d’Europe et du Levant du Paléolithique inférieur décrits en annexe 8 (tableau 62).

Quatrième chapitre : Etude taphonomique.
- 175 -
Ngebung A
Ngebung B
Ngebung Upper
Terra Amata (France)
Soleilhac (France)
Notarchirico* (Italie)
Holon (Israël)
NRT 729 358 324 33176 3000 975 1568 NRI 321 168 163 30497 2278 588 996 % 44 46,9 50,3 78,9 75,9 60,3 63,5
Tableau 62 : Proportions des esquilles (NRI) sur les sites de plein air de Ngebung 2, de Terra Amata (Serre, 1987), de Soleilhac (Fosse & Bonifay, 1994), de Notarchirico (Cassoli et al., 1999) et d’Holon
(Monchot & Horwitz, 2002). (A : niveau A ; B : niveau B ; Upper : niveau supérieur ; * niveau Alpha, en pourcentages du NRT)
Les proportions des esquilles du site de Ngebung 2 sont un peu inférieures à celles
obtenues sur tous les sites de notre étude. Le niveau qui se rapproche le plus des sites de notre étude serait le niveau U de Ngebung 2 qui présente une majorité d’esquilles.
Le nombre d’esquilles est toujours très important et majoritaire dans les sites archéologiques de plein air quelles que soient les régions géographiques ou les climats. Le site de Notarchirico est le plus proche des sites de Sangiran au niveau de la proportion des esquilles, les autres présentant un nombre d’esquilles un peu plus important. Néanmoins, au niveau statistique+ (test du khi-deux), les proportions d’esquilles de tous les sites de comparaison de plein air choisis sont très significativement différentes de celles des sites de notre étude excepté pour Dayu et Terra Amata, mais vu le faible nombre de restes à Dayu, ce résultat est peu significatif. Les proportions d’esquilles des sites de Sangiran entre eux ne sont pas significativement différentes+.
Nous pouvons nous demander si ces fortes proportions reflètent des conditions de conservation différentes selon la localisation des sites ou si cela est dû à l’importance de l’échantillonnage. Ceci nous a conduit à approfondir l’étude de la conservation des restes déterminables notamment avec les représentations squelettiques des assemblages.
2.2. Représentation squelettique Les modalités de la conservation des restes déterminables sont liées à la répartition squelettique des assemblages, c’est-à-dire à la répartition du nombre de restes déterminables suivant les parties anatomiques. Dans un site archéologique, les os n’ont pas la même probabilité de conservation. En effet, les ossements de taxons différents ne se conservent pas de la même manière dans un assemblage (Behrensmeyer & Dechant Boaz, 1980). De la même façon, dans un même squelette les éléments anatomiques différents ne présentent pas la même probabilité de conservation (Shipman, 1981a ; Kreutzer, 1992 ; Auguste 1994 b; Lyman, 1994). Nous avons choisi d’utiliser plusieurs types de représentations squelettiques (générale, des bovidés et des cervidés et par classe de taille) qui peuvent paraître redondantes mais sont nécessaires pour appréhender la conservation à différents niveaux d’analyse.
+ tableaux 121 et 122, annexe 9.
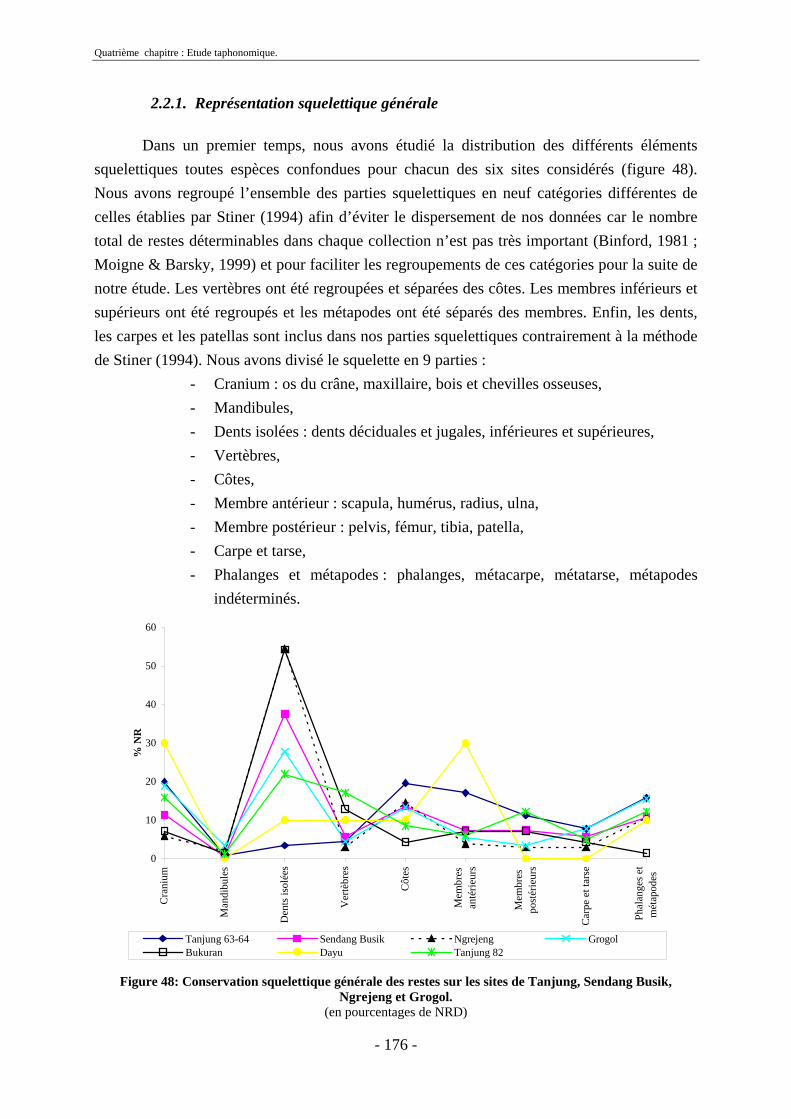
Quatrième chapitre : Etude taphonomique.
- 176 -
2.2.1. Représentation squelettique générale Dans un premier temps, nous avons étudié la distribution des différents éléments squelettiques toutes espèces confondues pour chacun des six sites considérés (figure 48). Nous avons regroupé l’ensemble des parties squelettiques en neuf catégories différentes de celles établies par Stiner (1994) afin d’éviter le dispersement de nos données car le nombre total de restes déterminables dans chaque collection n’est pas très important (Binford, 1981 ; Moigne & Barsky, 1999) et pour faciliter les regroupements de ces catégories pour la suite de notre étude. Les vertèbres ont été regroupées et séparées des côtes. Les membres inférieurs et supérieurs ont été regroupés et les métapodes ont été séparés des membres. Enfin, les dents, les carpes et les patellas sont inclus dans nos parties squelettiques contrairement à la méthode de Stiner (1994). Nous avons divisé le squelette en 9 parties :
- Cranium : os du crâne, maxillaire, bois et chevilles osseuses, - Mandibules, - Dents isolées : dents déciduales et jugales, inférieures et supérieures, - Vertèbres, - Côtes, - Membre antérieur : scapula, humérus, radius, ulna, - Membre postérieur : pelvis, fémur, tibia, patella, - Carpe et tarse, - Phalanges et métapodes : phalanges, métacarpe, métatarse, métapodes
indéterminés.
0
10
20
30
40
50
60
Cra
nium
Man
dibu
les
Den
ts is
olée
s
Ver
tèbr
es
Côt
es
Mem
bres
anté
rieur
s
Mem
bres
post
érie
urs
Car
pe e
t tar
se
Phal
ange
s et
mét
apod
es
% N
R
Tanjung 63-64 Sendang Busik Ngrejeng GrogolBukuran Dayu Tanjung 82
Figure 48: Conservation squelettique générale des restes sur les sites de Tanjung, Sendang Busik,
Ngrejeng et Grogol. (en pourcentages de NRD)
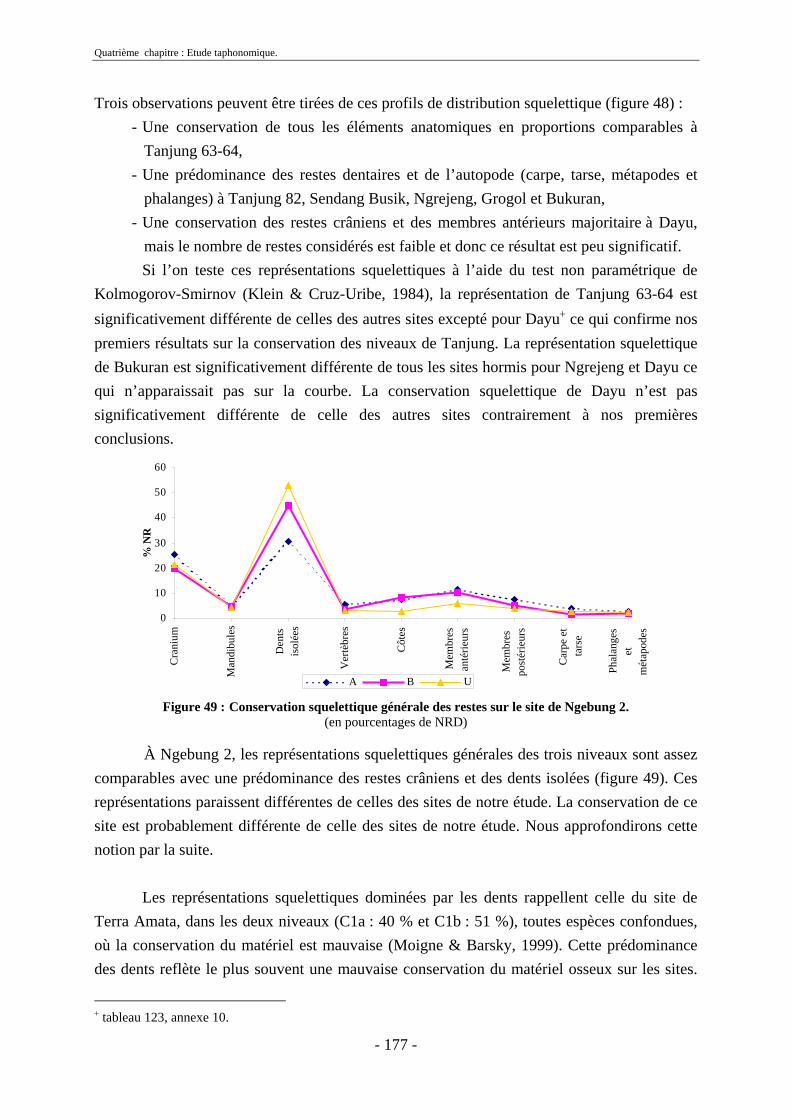
Quatrième chapitre : Etude taphonomique.
- 177 -
Trois observations peuvent être tirées de ces profils de distribution squelettique (figure 48) : - Une conservation de tous les éléments anatomiques en proportions comparables à
Tanjung 63-64, - Une prédominance des restes dentaires et de l’autopode (carpe, tarse, métapodes et
phalanges) à Tanjung 82, Sendang Busik, Ngrejeng, Grogol et Bukuran, - Une conservation des restes crâniens et des membres antérieurs majoritaire à Dayu,
mais le nombre de restes considérés est faible et donc ce résultat est peu significatif. Si l’on teste ces représentations squelettiques à l’aide du test non paramétrique de Kolmogorov-Smirnov (Klein & Cruz-Uribe, 1984), la représentation de Tanjung 63-64 est significativement différente de celles des autres sites excepté pour Dayu+ ce qui confirme nos premiers résultats sur la conservation des niveaux de Tanjung. La représentation squelettique de Bukuran est significativement différente de tous les sites hormis pour Ngrejeng et Dayu ce qui n’apparaissait pas sur la courbe. La conservation squelettique de Dayu n’est pas significativement différente de celle des autres sites contrairement à nos premières conclusions.
0
10
20
30
40
50
60
Cra
nium
Man
dibu
les
Den
tsis
olée
s
Ver
tèbr
es
Côt
es
Mem
bres
anté
rieur
s
Mem
bres
post
érie
urs
Car
pe e
tta
rse
Phal
ange
set
mét
apod
es
% N
R
A B U Figure 49 : Conservation squelettique générale des restes sur le site de Ngebung 2.
(en pourcentages de NRD)
À Ngebung 2, les représentations squelettiques générales des trois niveaux sont assez comparables avec une prédominance des restes crâniens et des dents isolées (figure 49). Ces représentations paraissent différentes de celles des sites de notre étude. La conservation de ce site est probablement différente de celle des sites de notre étude. Nous approfondirons cette notion par la suite. Les représentations squelettiques dominées par les dents rappellent celle du site de Terra Amata, dans les deux niveaux (C1a : 40 % et C1b : 51 %), toutes espèces confondues, où la conservation du matériel est mauvaise (Moigne & Barsky, 1999). Cette prédominance des dents reflète le plus souvent une mauvaise conservation du matériel osseux sur les sites.
+ tableau 123, annexe 10.

Quatrième chapitre : Etude taphonomique.
- 178 -
La tendance différente observée à Tanjung 63-64 peut-elle être expliquée par une conservation due à des agents différents ou par un nombre de restes plus important. Pour tenter de répondre à cette interrogation nous proposons tout d’abord d’étudier plus en détails la conservation squelettique des taxons majoritaires.
2.2.2. Représentation squelettique des bovidés et des cervidés Nous nous sommes intéressés en premier lieu à la représentation squelettique des deux grandes familles dominantes sur tous les sites, les bovidés et les cervidés (figures 50 et 51). Le site de Dayu n’a pas été figuré car les bovidés ne sont représentés que par un seul métacarpien et les cervidés par deux bois et un fragment de crâne.
Tanjung 63-64 et Tanjung 82
0102030405060708090
100
Cra
nium
Man
dibu
les
Den
tsis
olée
s
Ver
tèbr
es
Côt
es
Mem
bre
anté
rieur
Mem
bre
post
érie
ur
Car
pe e
tta
rse
Phal
ange
set
mét
apod
es
% N
R
Bovidés Tanjung 63-64 Bovidés Tanjung 82Cervidés Tanjung 63-64 Cervidés Tanjung 82
Figure 50 : Représentations squelettiques des bovidés et des cervidés du site de Tanjung pour les niveaux
de 1963-64 et 1982. (en pourcentages de nombres de restes (NR))
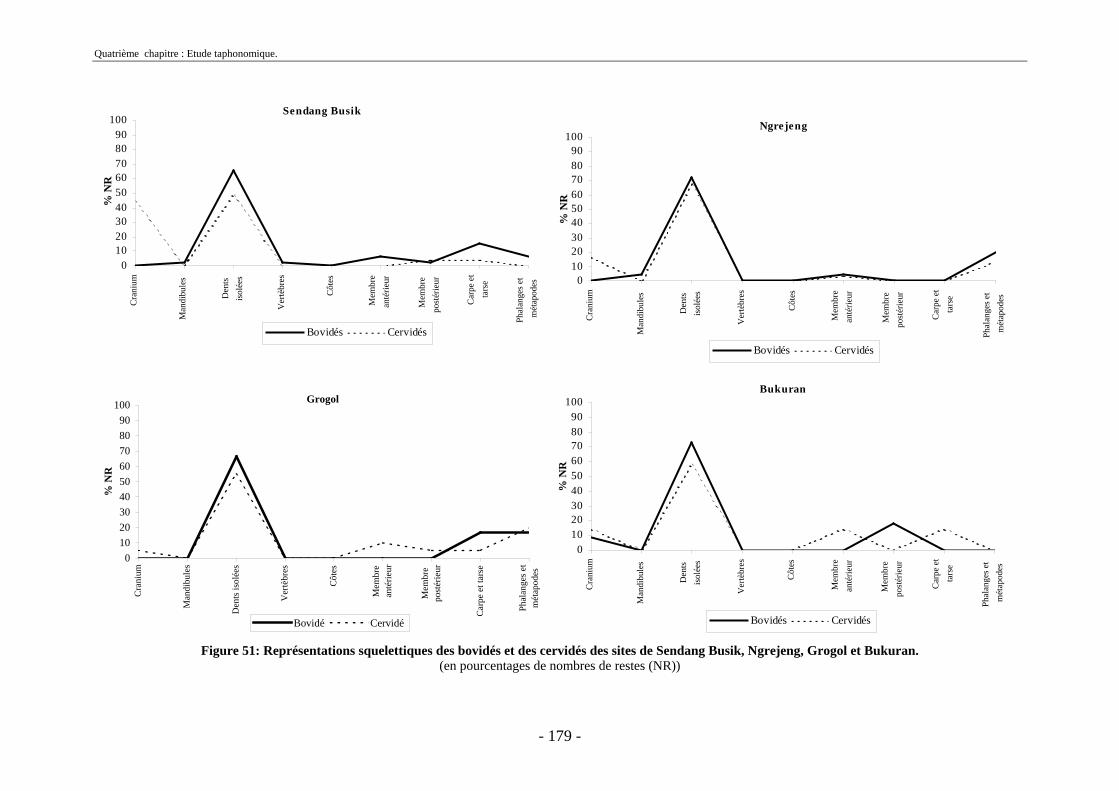
Quatrième chapitre : Etude taphonomique.
- 179 -
Sendang Busik
0102030405060708090
100C
rani
um
Man
dibu
les
Den
tsis
olée
s
Ver
tèbr
es
Côt
es
Mem
bre
anté
rieur
Mem
bre
post
érie
ur
Car
pe e
tta
rse
Phal
ange
s et
mét
apod
es
% N
R
Bovidés Cervidés
Ngrejeng
0102030405060708090
100
Cra
nium
Man
dibu
les
Den
tsis
olée
s
Ver
tèbr
es
Côt
es
Mem
bre
anté
rieur
Mem
bre
post
érie
ur
Car
pe e
tta
rse
Phal
ange
s et
mét
apod
es
% N
R
Bovidés Cervidés
Grogol
0102030405060708090
100
Cra
nium
Man
dibu
les
Den
ts is
olée
s
Ver
tèbr
es
Côt
es
Mem
bre
anté
rieur
Mem
bre
post
érie
ur
Car
pe e
t tar
se
Phal
ange
s et
mét
apod
es
% N
R
Bovidé Cervidé
Bukuran
0102030405060708090
100
Cra
nium
Man
dibu
les
Den
tsis
olée
s
Ver
tèbr
es
Côt
es
Mem
bre
anté
rieur
Mem
bre
post
érie
ur
Car
pe e
tta
rse
Phal
ange
s et
mét
apod
es
% N
R
Bovidés Cervidés
Figure 51: Représentations squelettiques des bovidés et des cervidés des sites de Sendang Busik, Ngrejeng, Grogol et Bukuran. (en pourcentages de nombres de restes (NR))

Quatrième chapitre : Etude taphonomique.
- 180 -
- Les bovidés (figures 50 et 51): Le squelette axial et cingulaire ainsi que les membres antérieurs et postérieurs sont très faiblement représentés ou totalement absents sur la plupart des sites. Dans la collection de Tanjung, pour les deux niveaux, l’ensemble des parties squelettiques est représenté. À Sendang Busik, Ngrejeng, Grogol et Bukuran, les restes majoritaires sont les restes crâniens et plus précisément les dents isolées. Les restes les plus nombreux après les restes crâniens sont les os de l’autopode ou du membre postérieur. À Holon (annexe 8), l’auroch présente une conservation squelettique comparable à ces sites avec une prédominance des dents et des restes de l’autopode (Monchot & Horwitz, 2002). À Ngebung, pour les grands bovidés, dans les 3 niveaux, les restes dentaires sont majoritaires suivis des éléments des membres antérieurs. Cependant, à Tanjung, dans les deux niveaux, les restes les plus nombreux sont les os de l’autopode et la proportion de restes des membres antérieurs est supérieure à celle des dents. Nous observons quasiment le même cas de figure à Notarchirico (annexe 8), dans le niveau Alpha, pour le Bos qui est représenté par 40 % de restes de l’autopode et 11 % pour les membres antérieurs (Cassoli et al., 1999). Les dents sont les éléments fossiles les plus minéralisés, donc les plus résistants à l’altération et ceux qui se conservent le mieux. Pour ce qui est des os de l’autopode, ce sont en général des os de petite taille et assez denses qui résistent mieux à l’altération que les os des membres. Leur prédominance dans les assemblages confirme donc une mauvaise conservation générale des restes de bovidés pour l’ensemble des sites considérés car seuls les éléments les plus résistants à l’altération ont été conservés. - Les cervidés (figures 50 et 51): L’ensemble des parties squelettiques est représenté dans les deux niveaux de Tanjung. À Sendang Busik, Ngrejeng, Grogol et Bukuran, les os du squelette axial ne sont pas présents. À Sendang Busik et Ngrejeng, les os des membres antérieurs sont également absents. Enfin à Ngrejeng et Bukuran, les os des membres postérieurs ne sont pas représentés. Sur tous les sites, les restes crâniens (crâne, mandibules et dents isolées) sont les plus abondants. Le grand nombre d’os du crâne chez les cervidés par rapport aux bovidés s’explique par la présence de nombreux fragments de bois dans les assemblages. Les bois sur le site de Tanjung 63-64 représentent 100 % des restes crâniens, à Tanjung 82, 94 %, à Sendang Busik 91 %, à Ngrejeng 10 %, à Grogol 93 %, à Bukuran 100 % et à Dayu 67 %. À Tanjung 63-64 et à Sendang Busik, les os du crâne sont plus nombreux que les dents alors que c’est l’inverse à Tanjung 82, Ngrejeng, Grogol et Bukuran.

Quatrième chapitre : Etude taphonomique.
- 181 -
Les représentations de Tanjung 82 et de Tanjung 63-64 pour les cervidés sont différentes de celles des autres sites où les dents dominent, comme pour les bovidés. À Tanjung 63-64, les dents sont très peu représentées alors que les os de l’autopode sont majoritaires. De même pour Dama à Notarchirico, les dents sont minoritaires (environ 5%) et les restes de l’autopode sont très majoritaires (environ 64%) (Cassoli et al., 1999 ; annexe 8). À Tanjung 82, les dents et les vertèbres sont prédominantes, ce qui est différent de la représentation des bovidés pour ce même site.
Les os de l’autopode chez les cervidés sont moins bien représentés que chez les bovidés. Néanmoins, les tendances des répartitions squelettiques chez les bovidés et les cervidés sont assez comparables si l’on exclut les bois des restes crâniens. Les dents et les os de l’autopode sont prédominants ce qui est caractéristique d’une mauvaise conservation des fossiles de ces deux familles. Tanjung montre une conservation différente des autres sites, pour les bovidés et les cervidés, ce qui confirme une conservation des ossements différente sur ce site. En considérant ces deux grands groupes majoritaires (bovidés et cervidés), nous excluons la notion de différence de taille au sein de ces groupes. En effet, dans ces deux familles, il existe des espèces de taille très différente qui peuvent avoir eu une histoire taphonomique différente selon la taille des ossements. Duboisia santeng est, par exemple, une espèce de bovidé d’une taille très inférieure à celle de Bibos palaesondaicus ou Bubalus palaeokerabau (Hooijer, 1958). De même, chez les cervidés, l’espèce Axis lydekkeri est d’une taille inférieure à celle de Cervus (Rusa) sp. (Zaim et al., 2003). Nous avons donc choisi d’établir la représentation squelettique par classe de taille.
2.2.3. Représentation squelettique par classe de taille Nous avons choisi de séparer l’ensemble des restes de bovidés et de cervidés, en classes de taille, sans inclure les fragments de bois (hors fragments de frontal) dans la suite de notre étude afin d’éviter une sur-représentation de ceux-ci car leur perte n’est pas liée nécessairement à la mort de l’animal :
- B1 : bovidés de grande taille, Bubalus palaeokerabau, Bibos palaesondaicus, Epileptobos groeneveldtii (masse ≥ 500 kg),
- B2C1 : bovidés de petite taille, Duboisia santeng et cervidé de grande taille, Cervus (Rusa) sp. (100 kg ≤ masse ≤ 500 kg),
- C2 : cervidés de petite taille, Axis sp., Axis lydekkeri (masse ≤ 100 kg). En utilisant ces classes de taille, nous avons choisi de regrouper les parties anatomiques en grands ensembles (figure 52) pour mettre en relief les parties squelettiques riches en viande (tronc et partie supérieure des membres : B, C et D) par rapport aux éléments
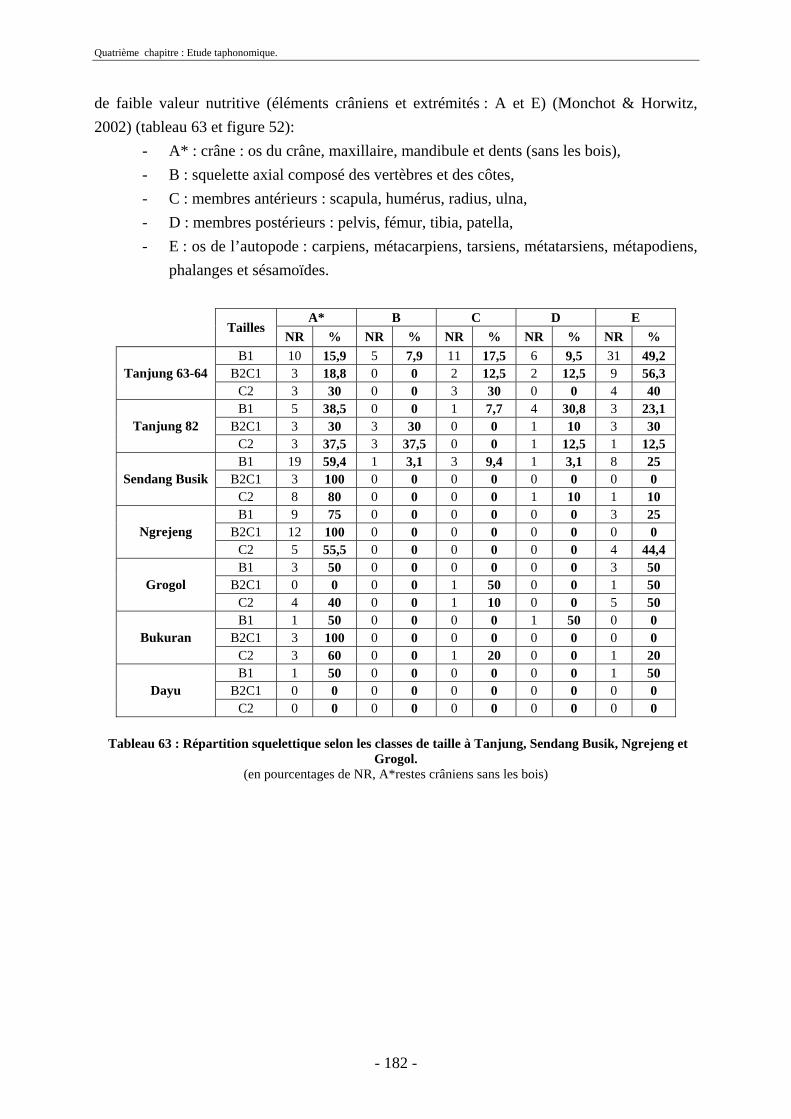
Quatrième chapitre : Etude taphonomique.
- 182 -
de faible valeur nutritive (éléments crâniens et extrémités : A et E) (Monchot & Horwitz, 2002) (tableau 63 et figure 52):
- A* : crâne : os du crâne, maxillaire, mandibule et dents (sans les bois), - B : squelette axial composé des vertèbres et des côtes, - C : membres antérieurs : scapula, humérus, radius, ulna, - D : membres postérieurs : pelvis, fémur, tibia, patella, - E : os de l’autopode : carpiens, métacarpiens, tarsiens, métatarsiens, métapodiens,
phalanges et sésamoïdes.
A* B C D E
Tailles NR % NR % NR % NR % NR %
B1 10 15,9 5 7,9 11 17,5 6 9,5 31 49,2 B2C1 3 18,8 0 0 2 12,5 2 12,5 9 56,3 Tanjung 63-64
C2 3 30 0 0 3 30 0 0 4 40 B1 5 38,5 0 0 1 7,7 4 30,8 3 23,1
B2C1 3 30 3 30 0 0 1 10 3 30 Tanjung 82 C2 3 37,5 3 37,5 0 0 1 12,5 1 12,5 B1 19 59,4 1 3,1 3 9,4 1 3,1 8 25
B2C1 3 100 0 0 0 0 0 0 0 0 Sendang Busik C2 8 80 0 0 0 0 1 10 1 10 B1 9 75 0 0 0 0 0 0 3 25
B2C1 12 100 0 0 0 0 0 0 0 0 Ngrejeng C2 5 55,5 0 0 0 0 0 0 4 44,4 B1 3 50 0 0 0 0 0 0 3 50
B2C1 0 0 0 0 1 50 0 0 1 50 Grogol C2 4 40 0 0 1 10 0 0 5 50 B1 1 50 0 0 0 0 1 50 0 0
B2C1 3 100 0 0 0 0 0 0 0 0 Bukuran C2 3 60 0 0 1 20 0 0 1 20 B1 1 50 0 0 0 0 0 0 1 50
B2C1 0 0 0 0 0 0 0 0 0 0 Dayu C2 0 0 0 0 0 0 0 0 0 0
Tableau 63 : Répartition squelettique selon les classes de taille à Tanjung, Sendang Busik, Ngrejeng et
Grogol. (en pourcentages de NR, A*restes crâniens sans les bois)
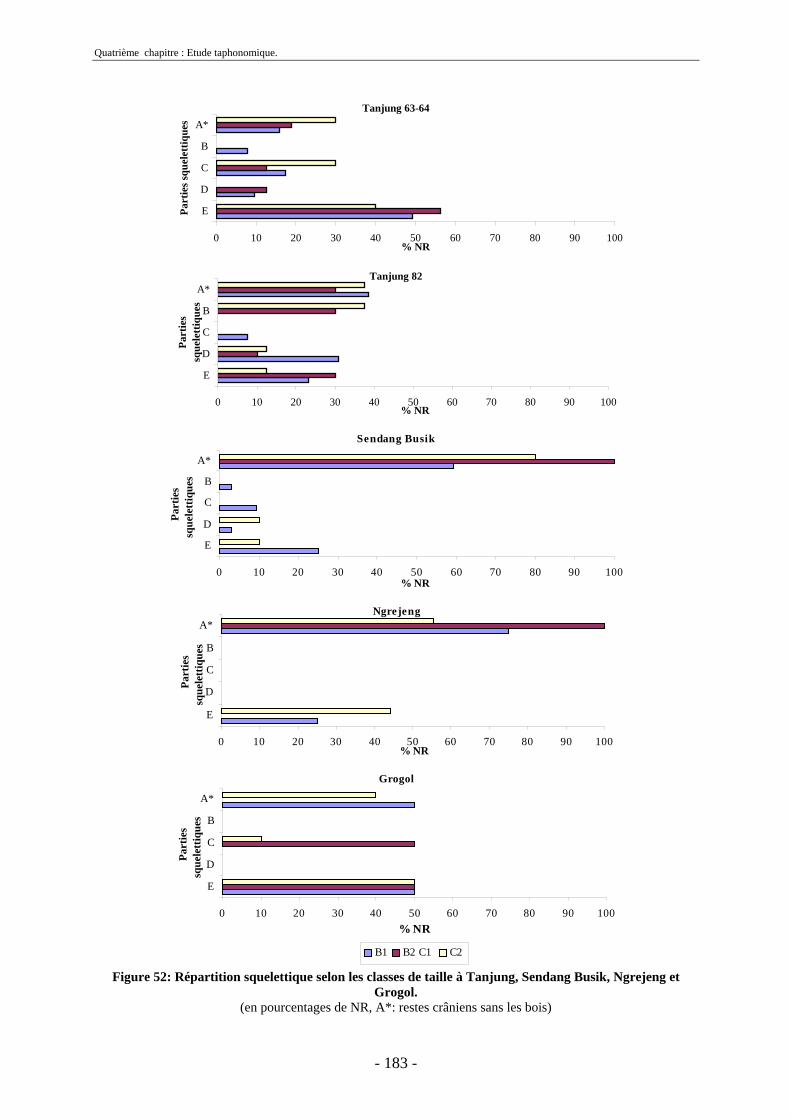
Quatrième chapitre : Etude taphonomique.
- 183 -
Tanjung 63-64
0 10 20 30 40 50 60 70 80 90 100
E
D
C
B
A*
Part
ies s
quel
ettiq
ues
% NR
Tanjung 82
0 10 20 30 40 50 60 70 80 90 100
E
D
C
B
A*
Part
ies
sque
lett
ique
s
% NR
Sendang Busik
0 10 20 30 40 50 60 70 80 90 100
E
D
C
B
A*
Part
ies
sque
lett
ique
s
% NR
Ngrejeng
0 10 20 30 40 50 60 70 80 90 100
E
D
C
B
A*
Part
ies
sque
lett
ique
s
% NR
Grogol
0 10 20 30 40 50 60 70 80 90 100
E
D
C
B
A*
Part
ies
sque
lett
ique
s
% NR
B1 B2 C1 C2
Figure 52: Répartition squelettique selon les classes de taille à Tanjung, Sendang Busik, Ngrejeng et Grogol.
(en pourcentages de NR, A*: restes crâniens sans les bois)

Quatrième chapitre : Etude taphonomique.
- 184 -
Tanjung 63-64 et 82 (figure 52):
Les répartitions squelettiques de Tanjung 82 et 63-64 sont proches car presque toutes les parties anatomiques sont représentées pour la plupart des classes de taille. Les restes des parties riches en viande sont mieux représentés à Tanjung, les deux niveaux confondus, que sur les autres sites. À Tanjung 82, les restes des éléments riches en viande sont proportionnellement plus nombreux qu’à Tanjung 63-64 notamment pour les classes C2 et B2C1. À Tanjung 63-64, les restes des extrémités sont prédominants pour les trois classes de taille alors qu’à Tanjung 82, ce sont les restes crâniens et du squelette axial qui sont prédominants. Sendang Busik et Ngrejeng (figure 52): Pour les 3 classes de taille, la majorité des restes appartiennent aux os du crâne. Les os des extrémités des classes B1 et C2 sont moins nombreux qu’à Tanjung. La classe B2C1 sur ces deux sites est sur-représentée par les restes crâniens. À Sendang Busik et Ngrejeng, toutes les parties squelettiques ne sont pas représentées pour toutes les classes de taille et la majorité des restes observés provient d’éléments pauvres en viande. Grogol (figure 52): Pour les classes B1 et C2, les restes provenant du crâne (A*) et des extrémités (E) sont majoritairement représentés. Pour la classe B1, les restes du squelette axial (B), les restes des membres antérieurs (C) et postérieurs (D) sont absents. La classe B2C1 présente le même nombre de restes provenant des membres antérieurs et des extrémités. Ces valeurs sont peu significatives compte tenu du nombre de restes considérés. Les restes des éléments crâniens, du squelette axial et des membres postérieurs ne sont pas représentés. La classe C2, hormis les restes de A* et E, présente un reste issu des membres antérieurs. À Grogol, comme à Sendang Busik et à Ngrejeng, toutes les parties squelettiques ne sont pas représentées pour toutes les classes de taille et la majorité des restes représentés sont des restes provenant d’éléments pauvres en viande. De plus, les restes du squelette axial et des membres postérieurs sont absents pour toutes les classes de taille. Nous avons appliqué ensuite le test non paramétrique de Kolmogorov-Smirnov car l’effectif total des différents sites pouvait être inférieur à 40 et que certaines valeurs pouvaient être inférieures à 5. Les représentations simplifiées des différentes classes de taille pour chaque site ne sont pas significativement différentes+. De même, si l’on compare les
+ tableau 124, annexe 10.

Quatrième chapitre : Etude taphonomique.
- 185 -
représentations squelettiques simplifiées d’une classe de taille pour les différents sites, le résultat du test est très majoritairement non significatif excepté pour Tanjung 63-64 avec Sendang Busik et Ngrejeng pour la classe B1 et pour la classe B2C1 à Ngrejeng avec les deux niveaux de Tanjung+ . En conclusion, les deux niveaux de Tanjung semble présenter une tendance différente pour leur représentation squelettique simplifiée par rapport aux autres sites mais ceci n’est pas confirmé statistiquement.
2.2.4. Comparaisons de Tanjung et Sendang Busik avec d’autres sites de plein air
Nous avons comparé les données obtenues à Sangiran avec celles obtenues pour un site de plein air de la même région (Ngebung 2). Néanmoins, les données taphonomiques sur des sites de plein air en Asie du sud-est étant très rares, nous avons élargi notre comparaison à des sites de plein air d’Europe, d’Amérique du Nord et du Moyen Orient datés du Miocène, du Paléolithique inférieur, moyen et de l’Actuel. Nous avons choisi des sites, naturels ou archéologiques, où les données sur les représentations squelettiques des bovidés et des cervidés étaient suffisantes et accessibles. Puis nous nous sommes basés sur la méthode utilisée par Monchot et Horwitz (2002) pour le site d’Holon (tableau 64). Pour ce qui est des données sur Sangiran, seules les données provenant des classes de taille B1 à Tanjung 63-64 et de la classe de taille B1 à Sendang Busik ont été exploitées. Ce choix a été fait pour éviter les absences c’est-à-dire que les sites où certains éléments anatomiques étaient absents n’ont pas été considérés dans cette comparaison.
Les sites de comparaisons se répartissent en plusieurs groupes : Les sites riches en restes crâniens (A > 45%): Holon B (6), Soleilhac (8),
Ngebung A (10), Ngebung B (11), Wallertheim B1 (12) et Sendang Busik (26), Les sites riches en éléments du squelette axial (B > 45%) : Cagny l’Epinette
(5), Mont Sainte Hélène 1 et 3 (21 et 22), Les sites où les éléments riches en viande sont abondants (C ou D > 20%):
Terra Amata (2), Mutzig (15), Mont Dol (18), Les sites riches en extrémités des membres (E > 40%): Notarchirico (3 et 4),
Soucy 1 (9), Mutzig (15), Mont Dol (17 et 18), Tourville D (19 et 20), Tanjung (24 et 25).
Certains de ces sites se situent très près d’un point d’eau (rivière, lac, bord de mer) :
Isernia la Pineta, Terra Amata, Notarchirico, Ngebung 2, Tanjung, Sendang Busik, ce qui a pu perturber leur distribution squelettique. De même, d’autres sites ont subi les actions
+ tableaux 125 à 128, annexe 10.
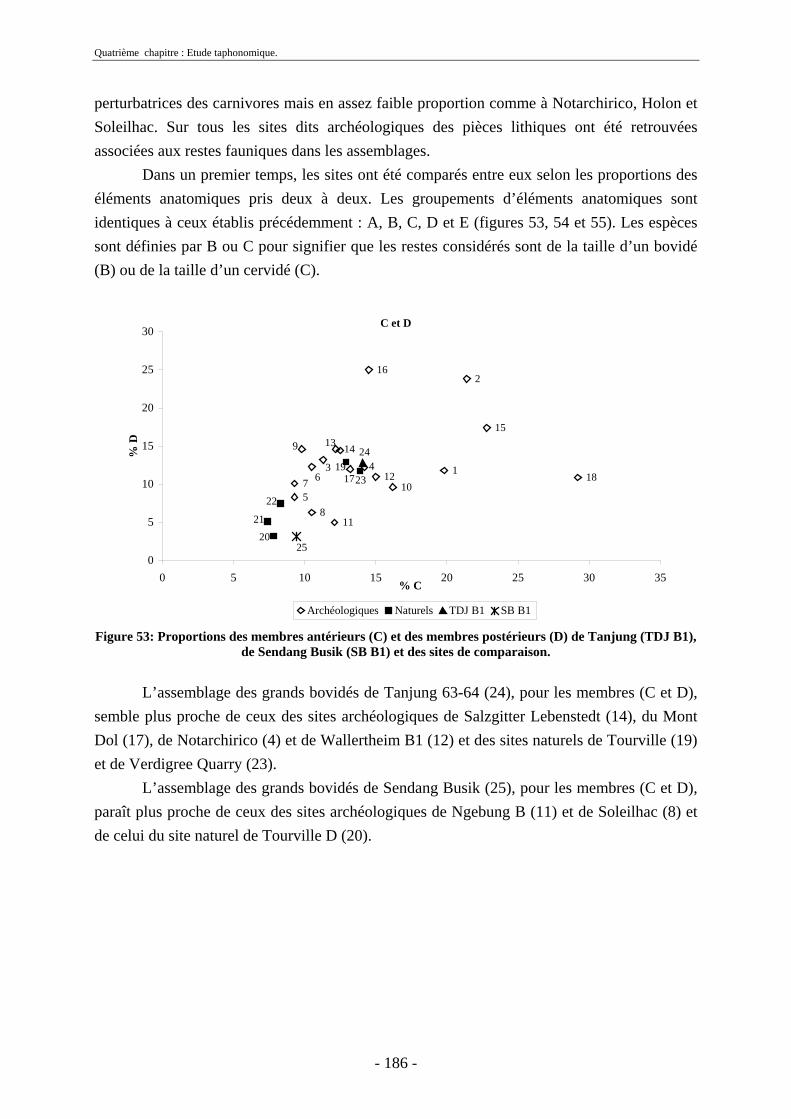
Quatrième chapitre : Etude taphonomique.
- 186 -
perturbatrices des carnivores mais en assez faible proportion comme à Notarchirico, Holon et Soleilhac. Sur tous les sites dits archéologiques des pièces lithiques ont été retrouvées associées aux restes fauniques dans les assemblages.
Dans un premier temps, les sites ont été comparés entre eux selon les proportions des éléments anatomiques pris deux à deux. Les groupements d’éléments anatomiques sont identiques à ceux établis précédemment : A, B, C, D et E (figures 53, 54 et 55). Les espèces sont définies par B ou C pour signifier que les restes considérés sont de la taille d’un bovidé (B) ou de la taille d’un cervidé (C).
C et D
417
367
58
11
14139
101 18
15
162
122319
22
20
21
24
250
5
10
15
20
25
30
0 5 10 15 20 25 30 35% C
% D
Archéologiques Naturels TDJ B1 SB B1
Figure 53: Proportions des membres antérieurs (C) et des membres postérieurs (D) de Tanjung (TDJ B1), de Sendang Busik (SB B1) et des sites de comparaison.
L’assemblage des grands bovidés de Tanjung 63-64 (24), pour les membres (C et D),
semble plus proche de ceux des sites archéologiques de Salzgitter Lebenstedt (14), du Mont Dol (17), de Notarchirico (4) et de Wallertheim B1 (12) et des sites naturels de Tourville (19) et de Verdigree Quarry (23).
L’assemblage des grands bovidés de Sendang Busik (25), pour les membres (C et D), paraît plus proche de ceux des sites archéologiques de Ngebung B (11) et de Soleilhac (8) et de celui du site naturel de Tourville D (20).
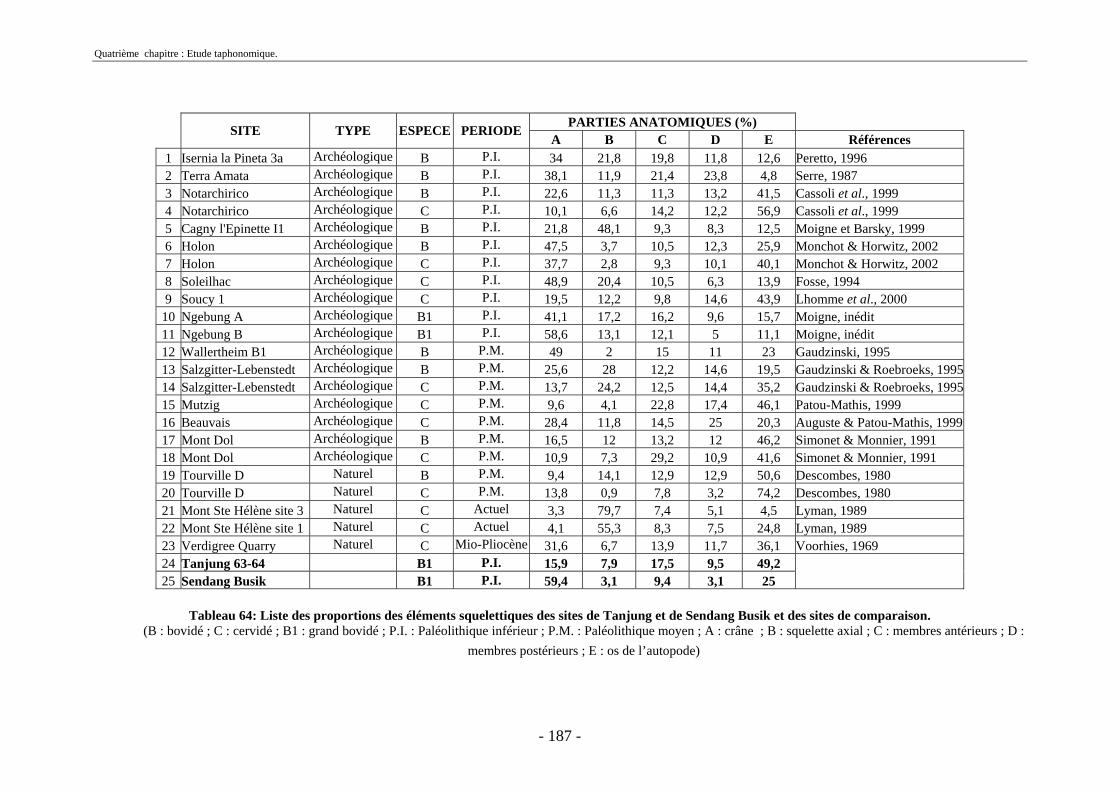
Quatrième chapitre : Etude taphonomique.
- 187 -
PARTIES ANATOMIQUES (%)
SITE TYPE ESPECE PERIODE A B C D E Références
1 Isernia la Pineta 3a Archéologique B P.I. 34 21,8 19,8 11,8 12,6 Peretto, 1996 2 Terra Amata Archéologique B P.I. 38,1 11,9 21,4 23,8 4,8 Serre, 1987 3 Notarchirico Archéologique B P.I. 22,6 11,3 11,3 13,2 41,5 Cassoli et al., 1999 4 Notarchirico Archéologique C P.I. 10,1 6,6 14,2 12,2 56,9 Cassoli et al., 1999 5 Cagny l'Epinette I1 Archéologique B P.I. 21,8 48,1 9,3 8,3 12,5 Moigne et Barsky, 1999 6 Holon Archéologique B P.I. 47,5 3,7 10,5 12,3 25,9 Monchot & Horwitz, 2002 7 Holon Archéologique C P.I. 37,7 2,8 9,3 10,1 40,1 Monchot & Horwitz, 2002 8 Soleilhac Archéologique C P.I. 48,9 20,4 10,5 6,3 13,9 Fosse, 1994 9 Soucy 1 Archéologique C P.I. 19,5 12,2 9,8 14,6 43,9 Lhomme et al., 2000
10 Ngebung A Archéologique B1 P.I. 41,1 17,2 16,2 9,6 15,7 Moigne, inédit 11 Ngebung B Archéologique B1 P.I. 58,6 13,1 12,1 5 11,1 Moigne, inédit 12 Wallertheim B1 Archéologique B P.M. 49 2 15 11 23 Gaudzinski, 1995 13 Salzgitter-Lebenstedt Archéologique B P.M. 25,6 28 12,2 14,6 19,5 Gaudzinski & Roebroeks, 199514 Salzgitter-Lebenstedt Archéologique C P.M. 13,7 24,2 12,5 14,4 35,2 Gaudzinski & Roebroeks, 199515 Mutzig Archéologique C P.M. 9,6 4,1 22,8 17,4 46,1 Patou-Mathis, 1999 16 Beauvais Archéologique C P.M. 28,4 11,8 14,5 25 20,3 Auguste & Patou-Mathis, 199917 Mont Dol Archéologique B P.M. 16,5 12 13,2 12 46,2 Simonet & Monnier, 1991 18 Mont Dol Archéologique C P.M. 10,9 7,3 29,2 10,9 41,6 Simonet & Monnier, 1991 19 Tourville D Naturel B P.M. 9,4 14,1 12,9 12,9 50,6 Descombes, 1980 20 Tourville D Naturel C P.M. 13,8 0,9 7,8 3,2 74,2 Descombes, 1980 21 Mont Ste Hélène site 3 Naturel C Actuel 3,3 79,7 7,4 5,1 4,5 Lyman, 1989 22 Mont Ste Hélène site 1 Naturel C Actuel 4,1 55,3 8,3 7,5 24,8 Lyman, 1989 23 Verdigree Quarry Naturel C Mio-Pliocène 31,6 6,7 13,9 11,7 36,1 Voorhies, 1969 24 Tanjung 63-64 B1 P.I. 15,9 7,9 17,5 9,5 49,2 25 Sendang Busik B1 P.I. 59,4 3,1 9,4 3,1 25
Tableau 64: Liste des proportions des éléments squelettiques des sites de Tanjung et de Sendang Busik et des sites de comparaison.
(B : bovidé ; C : cervidé ; B1 : grand bovidé ; P.I. : Paléolithique inférieur ; P.M. : Paléolithique moyen ; A : crâne ; B : squelette axial ; C : membres antérieurs ; D : membres postérieurs ; E : os de l’autopode)
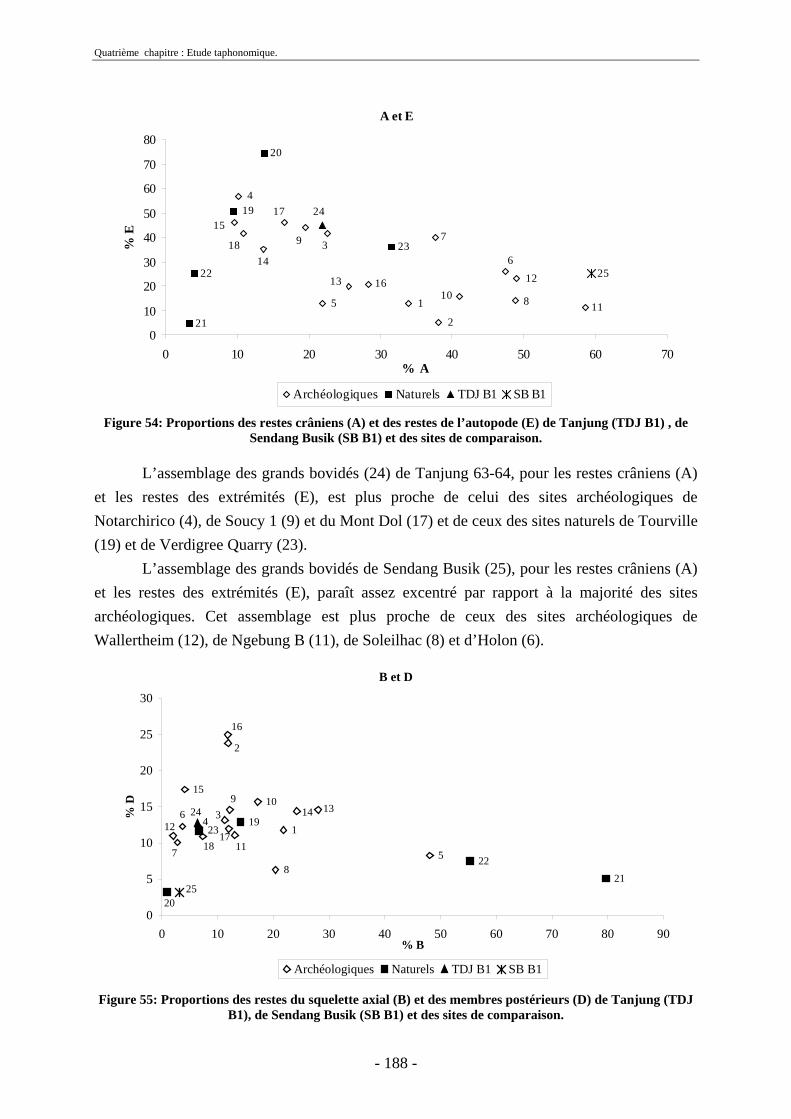
Quatrième chapitre : Etude taphonomique.
- 188 -
A et E
1110 8
126
2
15
13 16
7
1418
15
417
9 3 23
19
20
21
22
24
25
0
10
20
30
40
50
60
70
80
0 10 20 30 40 50 60 70% A
% E
Archéologiques Naturels TDJ B1 SB B1
Figure 54: Proportions des restes crâniens (A) et des restes de l’autopode (E) de Tanjung (TDJ B1) , de Sendang Busik (SB B1) et des sites de comparaison.
L’assemblage des grands bovidés (24) de Tanjung 63-64, pour les restes crâniens (A)
et les restes des extrémités (E), est plus proche de celui des sites archéologiques de Notarchirico (4), de Soucy 1 (9) et du Mont Dol (17) et de ceux des sites naturels de Tourville (19) et de Verdigree Quarry (23).
L’assemblage des grands bovidés de Sendang Busik (25), pour les restes crâniens (A) et les restes des extrémités (E), paraît assez excentré par rapport à la majorité des sites archéologiques. Cet assemblage est plus proche de ceux des sites archéologiques de Wallertheim (12), de Ngebung B (11), de Soleilhac (8) et d’Holon (6).
B et D
17
323
18
9
2
16
15
612
7
1314
1
10
8
115
2419
20
2122
4
25
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70 80 90% B
% D
Archéologiques Naturels TDJ B1 SB B1
Figure 55: Proportions des restes du squelette axial (B) et des membres postérieurs (D) de Tanjung (TDJ B1), de Sendang Busik (SB B1) et des sites de comparaison.

Quatrième chapitre : Etude taphonomique.
- 189 -
L’assemblage des grands bovidés de Tanjung 63-64 (24), pour le squelette axial et les membres postérieurs (B et D), est le plus proche de ceux des sites archéologiques de Notarchirico (3 et 4), du Mont Dol (17 et 18), d’Holon (6), et de celui du site naturel de Verdigree Quarry (23).
L’assemblage des grands bovidés de Sendang Busik (25), pour le squelette axial et les membres postérieurs (B et D), est plus proche de celui du site archéologique de Soleilhac (8) et de celui du site naturel de Tourville D (20).
Nous pouvons constater que certains sites reviennent plusieurs fois au cours de ces
comparaisons des proportions des éléments squelettiques car leur représentation se rapproche de celles de nos sites. L’origine des représentations squelettiques de ces sites peut nous donner des indices sur l’origine de celles des grands bovidés de Tanjung 63-64 et de Sendang Busik. La présentation de ces sites se situe en annexe 8. Tanjung 63 –64:
L’assemblage de Tanjung 63-64 pour les grands bovidés (24) se rapproche des assemblages des sites archéologiques de Notarchirico (3 ou 4) et du Mont Dol (17 ou 18) pour les représentations des éléments squelettiques pris deux à deux. Ces deux sites sont des sites archéologiques de périodes différentes où l’action anthropique a été clairement démontrée (annexe 8).
En ce qui concerne les sites naturels, l’assemblage du site de Tanjung (24) se rapproche de celui de Verdigree Quarry (annexe 8) pour les trois comparaisons. En outre, le site de Tanjung (24) se rapproche de Tourville (19) pour deux des trois comparaisons (annexe 8).
Il semble qu’il ne soit pas facile de rapprocher l’assemblage de Tanjung (24) d’un type de site archéologique ou naturel. Nous pouvons penser que ce résultat confirme une multiplicité des agents à l’origine de l’assemblage osseux des grands bovidés de Tanjung.
Sendang Busik:
L’assemblage du site de Sendang Busik pour les grands bovidés (25) paraît très différent des assemblages de Tanjung. Pour les sites archéologiques, cette accumulation de Sendang Busik se rapproche de l’assemblage de Soleilhac (8) (annexe 8) pour toutes les proportions d’éléments squelettiques et de celui de Ngebung 2 (niveau B) (annexe 8) pour deux des trois représentations des éléments squelettiques pris deux à deux.
Au niveau des sites naturels, l’assemblage de Sendang Busik (25) se rapproche de
celui de Tourville D (20) (annexe 8) pour deux des trois comparaisons.

Quatrième chapitre : Etude taphonomique.
- 190 -
L’accumulation (25) du site de Sendang Busik paraît différente de celles de Tanjung (24). La composition de l’assemblage de Sendang Busik semble se rapprocher de celle du site archéologique voisin de Ngebung B (11) et de Soleilhac (8) où l’origine de l’assemblage est en partie anthropique. L’assemblage de Tanjung (24) est assez proche de ceux des sites archéologiques européens de Notarchirico (4) et du Mont Dol (17 et 18) où l’action anthropique a été mise en évidence et est une des actions à l’origine des assemblages.
Il n’est pas facile de rapprocher les assemblages des grands bovidés de Tanjung 63-64 et de Sendang Busik de ceux de sites archéologiques ou naturels dont l’origine des accumulations osseuses est connue. Ceci révèle une origine multiple et non unique de ces assemblages de grands bovidés de Tanjung et de Sendang Busik. Nous pouvons penser que le transport fluviatile a joué un certain rôle dans ces accumulations car la plupart des sites archéologiques ou naturels dont les proportions des éléments squelettiques sont les plus proches de celles de Tanjung et de Sendang Busik, sont des sites en milieu fluviatile où il y a eu plus ou moins de transport des ossements.
Nous avons appliqué le test de Kolmogorov-Smirnov+ à ces représentations
squelettiques simplifiées pour comparer les assemblages des grands bovidés de Tanjung 63-64 et de Sendang Busik avec les sites de comparaisons. À partir de ce test, nous pouvons conclure que l’assemblage de Tanjung 63-64 pour les grands bovidés n’est pas significativement différent des assemblages de Notarchirico (cervidés et bovidés), de Soucy 1, de Salzgitter Lebenstedt (cervidés), de Mutzig, du Mont Dol (cervidés et bovidés), de Tourville (cervidés) et de Verdigree Quarry. Ceci confirme nos précédents résultats pour Notarchirico, le Mont Dol, Tourville et Verdigree Quarry. L’assemblage des grands bovidés de Sendang Busik n’est pas significativement différent de ceux d’Holon (cervidés et bovidés), de Soleilhac, de Ngebung (niveaux A et B) et de Wallertheim B1. De même que pour Tanjung ces analyses statistiques pour l’assemblage de Sendang Busik permettent de compléter nos premières conclusions.
Après l’étude des représentations squelettiques, nous pouvons penser que le site de
Tanjung a eu une histoire taphonomique différente des autres sites considérés. Pour confirmer cette hypothèse nous avons étudié la représentation squelettique pour les taxons minoritaires.
2.2.5. Représentations squelettiques des taxons minoritaires
Les proboscidiens sont représentés en majorité par des fragments de dents isolées et plus précisément des fragments de défenses sur les sites de Tanjung, Sendang Busik, Ngrejeng et Grogol (chapitre II). Ceci est assez fréquent dans les sites de plein air du
+ tableaux 128 et 129, annexe 10.

Quatrième chapitre : Etude taphonomique.
- 191 -
Paléolithique inférieur par exemple en France à Terra-Amata (Serre, 1987, 1991 ; annexe 8) et Soleilhac (Fosse & Bonifay, 1991), en Europe à Notarchirico (Cassoli et al., 1999 ; annexe 8) et Ambrona (Villa et al., 2005) et au Proche-Orient à Holon (Monchot & Horwitz, sous presse ; annexe 8). Les os longs de proboscidiens sont souvent très spongieux et il est donc difficile d’extraire la moelle sans fragmenter d’une manière intensive l’os ce qui rend difficile, en général, la détermination d’os longs de proboscidiens (Monchot & Horwitz, sous presse). Les rhinocérotidés sont représentés par une épiphyse distale de fémur (Tanjung 63-64) et un grand sésamoïde (Tanjung 82) issus du site de Tanjung (chapitre II). Les hippopotamidés sont représentés par des dents inférieures isolées provenant du site de Sendang Busik (chapitre II). Les suidés sont représentés par un fragment de dent inférieure isolée à Bukuran, une incisive à Ngrejeng et par une épiphyse distale de tibia à Tanjung 82 (chapitre II). Les hyénidés sont représentés un fragment de dent inférieure à Bukuran (chapitre II). En ce qui concerne les reptiles de tous les sites, les crocodiles ne sont représentés que par des dents isolées et les tortues par des fragments de carapace et de plastron. Les poissons sont représentés par des dents isolées et une vertèbre et les gastéropodes par deux opercules (chapitre II).
Les représentations squelettiques de tous les taxons minoritaires (en nombre de restes) sont assez similaires, en effet, les dents sont majoritaires et plus précisément les dents inférieures. Les restes dentaires des taxons minoritaires proviennent des sites de Sendang Busik, Ngrejeng et Bukuran alors que les restes post-crâniens proviennent du site de Tanjung.
Le même type de conservation est observable pour les bovidés et les cervidés et pour les taxons minoritaires, les dents sont majoritaires sur tous les sites exceptés à Tanjung. Les représentations squelettiques des taxons minoritaires confirment donc celles des taxons majoritaires de ces mêmes sites. Il est peu aisé de comparer ces données avec d’autres sites car le plus souvent les représentations squelettiques des taxons minoritaires ne sont pas décrites.
Nous avons poursuivi l’étude de la conservation des assemblages osseux en utilisant
des indices de conservation et des indices squelettiques.
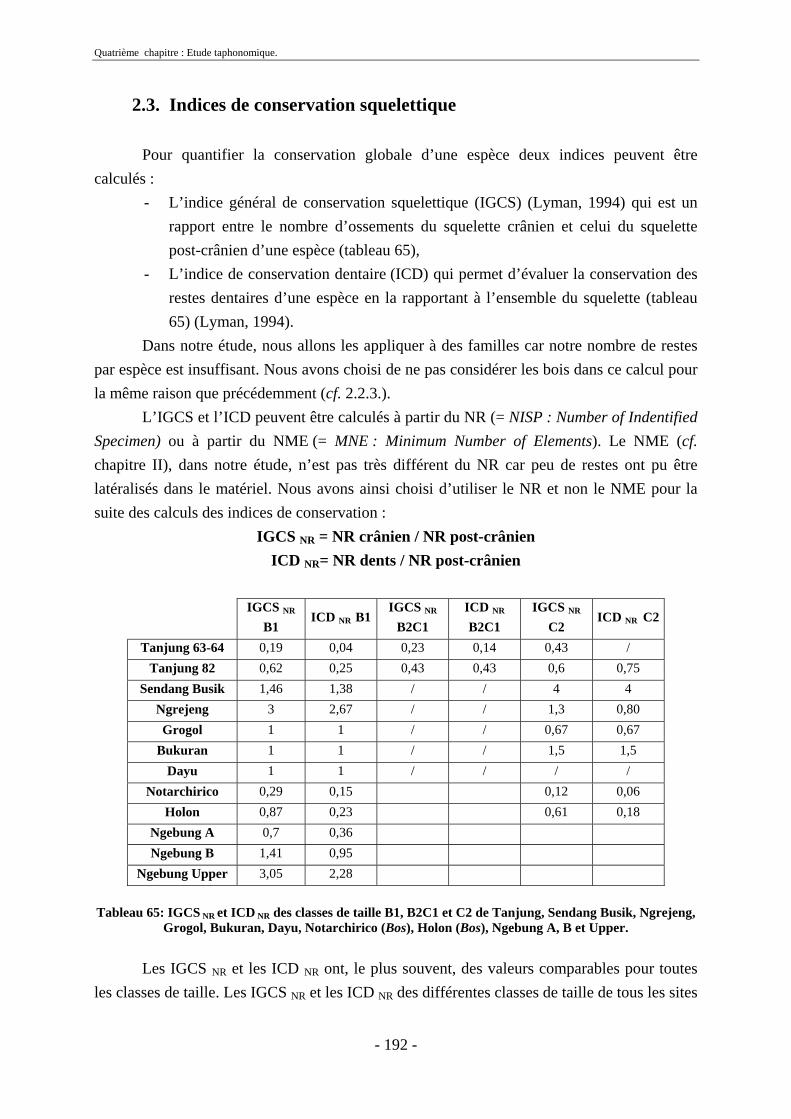
Quatrième chapitre : Etude taphonomique.
- 192 -
2.3. Indices de conservation squelettique Pour quantifier la conservation globale d’une espèce deux indices peuvent être calculés :
- L’indice général de conservation squelettique (IGCS) (Lyman, 1994) qui est un rapport entre le nombre d’ossements du squelette crânien et celui du squelette post-crânien d’une espèce (tableau 65),
- L’indice de conservation dentaire (ICD) qui permet d’évaluer la conservation des restes dentaires d’une espèce en la rapportant à l’ensemble du squelette (tableau 65) (Lyman, 1994).
Dans notre étude, nous allons les appliquer à des familles car notre nombre de restes par espèce est insuffisant. Nous avons choisi de ne pas considérer les bois dans ce calcul pour la même raison que précédemment (cf. 2.2.3.). L’IGCS et l’ICD peuvent être calculés à partir du NR (= NISP : Number of Indentified Specimen) ou à partir du NME (= MNE : Minimum Number of Elements). Le NME (cf. chapitre II), dans notre étude, n’est pas très différent du NR car peu de restes ont pu être latéralisés dans le matériel. Nous avons ainsi choisi d’utiliser le NR et non le NME pour la suite des calculs des indices de conservation :
IGCS NR = NR crânien / NR post-crânien ICD NR= NR dents / NR post-crânien
IGCS NR
B1 ICD NR B1
IGCS NR B2C1
ICD NR B2C1
IGCS NR C2
ICD NR C2
Tanjung 63-64 0,19 0,04 0,23 0,14 0,43 / Tanjung 82 0,62 0,25 0,43 0,43 0,6 0,75
Sendang Busik 1,46 1,38 / / 4 4 Ngrejeng 3 2,67 / / 1,3 0,80 Grogol 1 1 / / 0,67 0,67
Bukuran 1 1 / / 1,5 1,5 Dayu 1 1 / / / /
Notarchirico 0,29 0,15 0,12 0,06 Holon 0,87 0,23 0,61 0,18
Ngebung A 0,7 0,36 Ngebung B 1,41 0,95
Ngebung Upper 3,05 2,28
Tableau 65: IGCS NR et ICD NR des classes de taille B1, B2C1 et C2 de Tanjung, Sendang Busik, Ngrejeng,
Grogol, Bukuran, Dayu, Notarchirico (Bos), Holon (Bos), Ngebung A, B et Upper. Les IGCS NR et les ICD NR ont, le plus souvent, des valeurs comparables pour toutes les classes de taille. Les IGCS NR et les ICD NR des différentes classes de taille de tous les sites

Quatrième chapitre : Etude taphonomique.
- 193 -
sont souvent supérieurs à 1 ce qui indique que la proportion des restes crâniens et plus précisément des dents est nettement supérieure à celle des restes post-crâniens, exceptés à Tanjung et Grogol. À Sendang Busik, Ngrejeng et Bukuran, l’IGSC NR de la classe B2C1 n’a pas pu être calculé car la présence de restes post-crâniens n’a pas été constatée dans cette classe et à Grogol, au contraire, les restes crâniens sont absents. Les valeurs des indices obtenues à Tanjung pour les deux niveaux sont assez proches des valeurs de Notarchirico, Holon et Ngebung A (annexe 8). Les indices généraux de conservation squelettique supérieurs à 1 (à Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu) se rapprochent des valeurs obtenues à Ngebung dans les niveaux B et Upper pour les grands bovidés. À Dayu, nous n’avons pas pu déterminer de restes dans les classes B2C1 et C2. Pour les sites de Dayu et Bukuran, le nombre de restes considérés dans ces trois classes est très faible ou nul, ce qui rend la valeur de ces IGCS et ICD peu significative. Ces indices nous permettent de confirmer que les restes crâniens et particulièrement les restes dentaires sont en majorité dans la plupart des assemblages considérés excepté à Tanjung et Grogol. Cependant à Grogol, Bukuran et Dayu les résultats sont peu significatifs car le nombre de restes considérés est faible.
2.4. Indices de représentation squelettique Par la suite, afin de mettre en évidence plus clairement l’importance des différentes
parties squelettiques nous avons calculé différents indices prenant en compte les restes crâniens, le squelette axial, le squelette appendiculaire et les membres (tableau 66) (Potts, 1983 ; Blumenschine, 1986 ; Monchot & Horwitz, 2002).
Les trois premiers indices (crânial/axial ; crânial/squelette appendiculaire et crânial/membre) permettent de mettre en évidence l’importance des restes crâniens par rapport aux différentes parties du squelette post-crânien. Les autres indices considèrent les différentes parties du squelette post-crânien (antérieur/postérieur ; axial/squelette appendicualire et axial/membre).
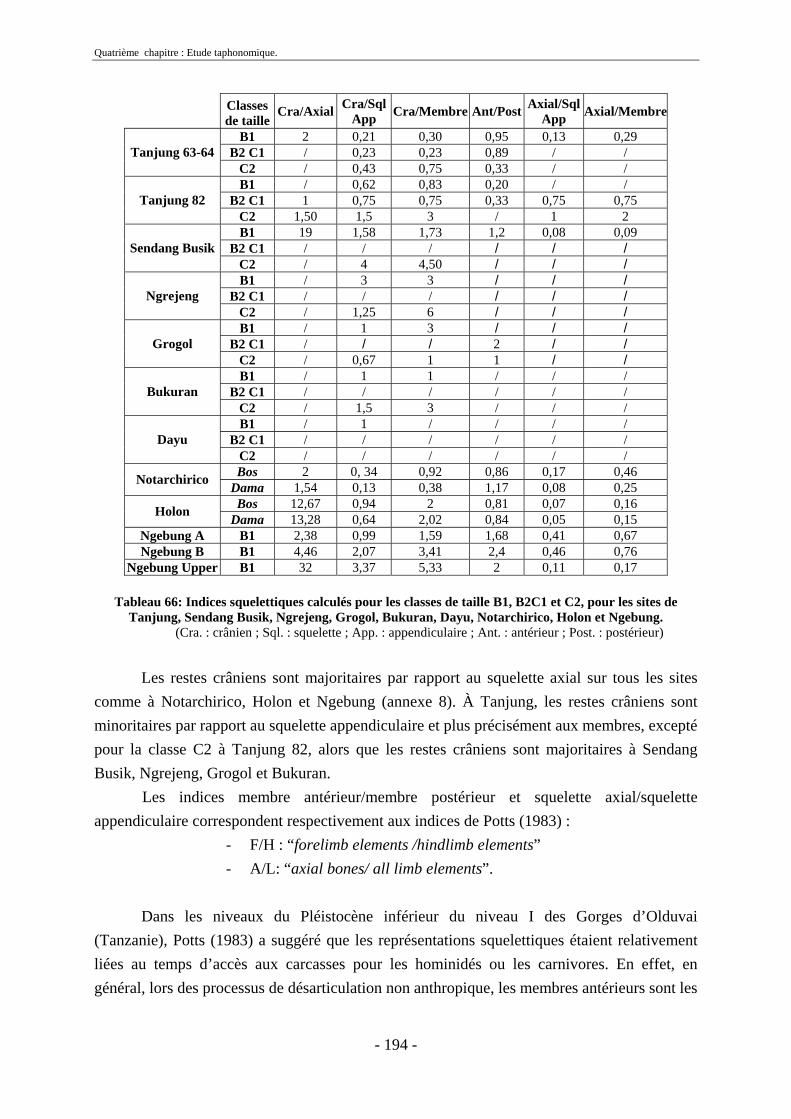
Quatrième chapitre : Etude taphonomique.
- 194 -
Classes de taille
Cra/Axial Cra/Sql App Cra/Membre Ant/Post Axial/Sql
App Axial/Membre
B1 2 0,21 0,30 0,95 0,13 0,29 B2 C1 / 0,23 0,23 0,89 / / Tanjung 63-64
C2 / 0,43 0,75 0,33 / / B1 / 0,62 0,83 0,20 / /
B2 C1 1 0,75 0,75 0,33 0,75 0,75 Tanjung 82 C2 1,50 1,5 3 / 1 2 B1 19 1,58 1,73 1,2 0,08 0,09
B2 C1 / / / / / / Sendang Busik C2 / 4 4,50 / / / B1 / 3 3 / / /
B2 C1 / / / / / / Ngrejeng C2 / 1,25 6 / / / B1 / 1 3 / / /
B2 C1 / / / 2 / / Grogol C2 / 0,67 1 1 / / B1 / 1 1 / / /
B2 C1 / / / / / / Bukuran C2 / 1,5 3 / / / B1 / 1 / / / /
B2 C1 / / / / / / Dayu C2 / / / / / / Bos 2 0, 34 0,92 0,86 0,17 0,46 Notarchirico Dama 1,54 0,13 0,38 1,17 0,08 0,25 Bos 12,67 0,94 2 0,81 0,07 0,16 Holon Dama 13,28 0,64 2,02 0,84 0,05 0,15
Ngebung A B1 2,38 0,99 1,59 1,68 0,41 0,67 Ngebung B B1 4,46 2,07 3,41 2,4 0,46 0,76
Ngebung Upper B1 32 3,37 5,33 2 0,11 0,17
Tableau 66: Indices squelettiques calculés pour les classes de taille B1, B2C1 et C2, pour les sites de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran, Dayu, Notarchirico, Holon et Ngebung.
(Cra. : crânien ; Sql. : squelette ; App. : appendiculaire ; Ant. : antérieur ; Post. : postérieur)
Les restes crâniens sont majoritaires par rapport au squelette axial sur tous les sites comme à Notarchirico, Holon et Ngebung (annexe 8). À Tanjung, les restes crâniens sont minoritaires par rapport au squelette appendiculaire et plus précisément aux membres, excepté pour la classe C2 à Tanjung 82, alors que les restes crâniens sont majoritaires à Sendang Busik, Ngrejeng, Grogol et Bukuran. Les indices membre antérieur/membre postérieur et squelette axial/squelette appendiculaire correspondent respectivement aux indices de Potts (1983) :
- F/H : “forelimb elements /hindlimb elements” - A/L: “axial bones/ all limb elements”.
Dans les niveaux du Pléistocène inférieur du niveau I des Gorges d’Olduvai
(Tanzanie), Potts (1983) a suggéré que les représentations squelettiques étaient relativement liées au temps d’accès aux carcasses pour les hominidés ou les carnivores. En effet, en général, lors des processus de désarticulation non anthropique, les membres antérieurs sont les

Quatrième chapitre : Etude taphonomique.
- 195 -
premiers séparés du squelette axial, ensuite les membres postérieurs sont désarticulés et enfin les éléments du squelette axial restent le plus souvent sur le site de mort de l’animal.
Pour une carcasse complète de bovidé, F/H est égal à 1 et A/L est égal à 1,6. Dans le cas d’un animal chassé le rapport A/L serait inférieur à 1,6 et le rapport F/H supérieur à 1 et dans le cas d’un animal charogné F/H serait inférieur à 1 et A/L supérieur à 1,6.
À Tanjung 63-64, pour les classes B1 et B2C1, et à Grogol, pour la classe C2, les
proportions des restes des membres antérieurs et postérieurs sont proches. À Sendang Busik, pour la classe B1, et à Grogol, pour la classe B2C1, les restes des membres antérieurs sont un peu plus nombreux comme à Holon. À Tanjung 63-64, pour la classe C2 et à Tanjung 82 pour les classes B1 et B2C1, le nombre de restes des membres postérieurs est légèrement supérieur comme sur les sites de Ngebung et de Notarchirico (Dama). Pour tous les autres cas le calcul n’a pas été possible. Nous ne constatons pas une nette différence entre les proportions des membres antérieurs et postérieurs.
En ce qui concerne le squelette axial, il est sous-représenté à Tanjung 63-64 pour toutes les classes de taille, à Tanjung 82 pour la classe B2C1 et à Sendang Busik pour la classe B1 par rapport au squelette appendiculaire et plus particulièrement par rapport aux membres. Dans les autres cas, les éléments du squelette axial sont absents ou les restes du squelette appendiculaire sont absents. D’une manière générale, les restes du squelette axial sont sous-représentés sur tous les sites et pour toutes les classes d’âge et à Notarchirico, Holon et Ngebung également.
Cette sous-représentation en éléments du squelette axial peut être due à divers phénomènes comme l’intensité de la diagenèse, l’importance du transport fluviatile comme à Notarchirico (Cassoli et al., 1999 ; annexe 8) ou les activités de charognage des hommes et/ou des carnivores.
2.5. Conclusion Ces différentes méthodes de dénombrement nous permettent de conclure que la conservation générale des sites du dôme de Sangiran considérés est mauvaise pour tous les taxons et plus particulièrement pour les bovidés et les cervidés qui sont les taxons majoritaires de ces assemblages. Le site de Tanjung confirme une tendance différente de celle des autres sites. En effet, sur ces derniers, seuls les éléments les plus résistants à l’altération, telles que les dents, sont représentés en majorité dans les assemblages osseux alors qu’à Tanjung tous les éléments sont plus ou moins représentés. Après avoir étudié la conservation des restes de ces sites, nous nous sommes intéressés à la fragmentation, sachant que la majorité des restes sont des fragments osseux, dans le but de mieux comprendre les actions étant à l’origine ou ayant modifié ces ossements.

Quatrième chapitre : Etude taphonomique.
- 196 -
3. FRAGMENTATION
Si nous considérons l’ensemble des assemblages osseux de tous les sites, nous remarquons l’absence d’os longs entiers (humérus, radio-ulna, fémur, tibia et métapodes). Cette absence d’os longs entiers souligne une grande fragmentation des os longs sur tous les sites. Ainsi, il nous paraît indispensable d’étudier la fragmentation des ossements afin de mieux comprendre l’origine et compléter l’histoire taphonomique des différents sites.
3.1. Taille des fragments osseux
Dans un premier temps, nous nous sommes intéressés à la longueur des restes déterminables et des esquilles composant les différents assemblages. Pour ce faire, nous avons divisé les restes en quatre classes de taille proposées par Brugal et Patou-Mathis (1993) (figure 56):
- 0 < classe I ≤ 20 millimètres, - 20 < classe II ≤ 50 millimètres, - 50 < classe III ≤ 100 millimètres, - classe IV > 100 millimètres.
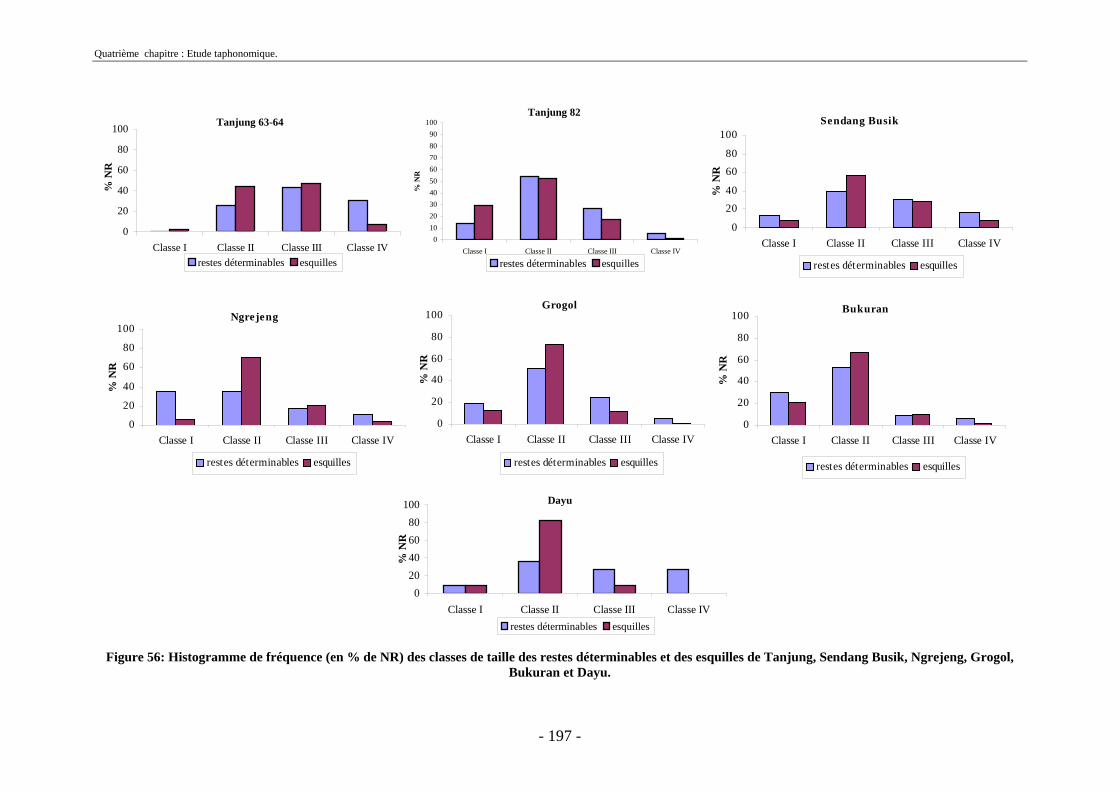
Quatrième chapitre : Etude taphonomique.
- 197 -
Tanjung 63-64
0
20
40
60
80
100
Classe I Classe II Classe III Classe IV
% N
R
restes déterminables esquilles
Tanjung 82
0102030405060708090
100
Classe I Classe II Classe III Classe IV
% N
R
restes déterminables esquilles
Sendang Busik
0
2040
6080
100
Classe I Classe II Classe III Classe IV
% N
R
restes déterminables esquilles
Ngrejeng
0
20
40
60
80
100
Classe I Classe II Classe III Classe IV
% N
R
restes déterminables esquilles
Grogol
0
20
40
60
80
100
Classe I Classe II Classe III Classe IV%
NR
restes déterminables esquilles
Bukuran
0
20
40
60
80
100
Classe I Classe II Classe III Classe IV
% N
R
restes déterminables esquilles
Dayu
020406080
100
Classe I Classe II Classe III Classe IV
% N
R
restes déterminables esquilles
Figure 56: Histogramme de fréquence (en % de NR) des classes de taille des restes déterminables et des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.

Quatrième chapitre : Etude taphonomique.
- 198 -
La majorité des restes présente une longueur supérieure à 20 millimètres et elle est le plus souvent comprise entre 20 et 50 millimètres. Cette absence de restes de très petite taille (< 10 millimètres) peut s’expliquer par un problème de reconnaissance du fragment osseux lors de la fouille ou par une absence de tamisage.
Ces valeurs, se retrouvent dans divers types de sites où l’activité humaine a été mise en évidence : site de plein air, grotte et abri. Sur le site anthropique mésolithique de plein air de Noyen (Bridault, 1994), les fragments d’os longs déterminés et indéterminés sont également compris entre 20 et 60 millimètres et sur le site de plein air d’Holon (Monchot & Horwitz, sous presse) la majorité des longueurs des esquilles est comprise entre 15 et 50 millimètres. Dans les différents niveaux des grottes de Pomongwe en Afrique du sud (Brain, 1981), la majorité des esquilles ont une longueur comprise entre 25 et 50 millimètres. De même, dans les niveaux L.S.A. (Lower Stone Age) et M.S.A. (Middle Stone Age) de l’abri de Bushman en Afrique du sud (Brain, 1981), la majorité des esquilles ont une longueur comprise entre 25 et 50 millimètres. Á Notarchirico, 44 % des restes ont une longueur inférieure à 50 millimètres et 34 % ont une longueur entre 50 et 100 millimètres (Cassoli et al., 1999).
La distribution des longueurs des esquilles du site de Tanjung 63-64 est significativement différente de celles des autres sites de notre étude lors de l’utilisation du test de Kolmogorov-Smirnov +. De même, la distribution des longueurs des esquilles de Sendang Busik est significativement différente de celles de Tanjung (pour les deux niveaux), de Grogol et de Bukuran. Tanjung 63-64 et Sendang Busik ne semblent pas avoir la même distribution de la longueur de leurs esquilles par rapport aux autres sites.
Pour les restes déterminables, les représentations semblent différentes de celles des esquilles. Ils sont plus nombreux que les esquilles dans les classes de taille III et IV, excepté à Tanjung 82 et Ngrejeng où les dents isolées facilement déterminables sont plus nombreuses et augmentent la classe de taille I. Les différences observées au niveau du nombre de restes déterminables de la classe de taille IV pour tous les sites s’expliquent aisément car il est plus facile, en général, de déterminer des fragments de grande taille qui sont susceptibles d’être plus proches de la forme de la pièce originale que des fragments de petite taille.
Au niveau statistique, avec un test de Kolmogorov-Smirnov, seules les distributions des esquilles, à Tanjung 63-64 et à Ngrejeng, sont significativement différentes de celles des restes déterminables+. Pour les distributions des restes déterminables entre elles, à Tanjung 63-64, la distribution est significativement différente de celles des autres sites sauf pour Sendang Busik et Dayu et à Sendang Busik, elle est significativement différente de celle de Ngrejeng et de Bukuran+.
+ tableaux 130 à 132, annexe 10.

Quatrième chapitre : Etude taphonomique.
- 199 -
199
Au sein des sites nous pouvons observer trois grandes tendances : 1) Les esquilles et les restes déterminables sont d’une taille entre 20 et 100 mm.
Les esquilles et les restes déterminables montrent des proportions comparables dans les classes II et III : Tanjung 63-64,
2) Les esquilles et les restes déterminables présentent une distribution de leur longueur comparable : Sendang Busik,
3) Les esquilles et les restes déterminables sont les mieux représentés dans la classe II entre 20 et 50 mm. La classe II est prédominante : Tanjung 82, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.
En résumé, les longueurs des restes déterminables se répartissent environ de la même
façon que celles des esquilles excepté pour la classe IV pour les deux tendances observées. Encore une fois, le site de Tanjung semble être différent des autres. La taille des esquilles et des restes déterminables sur ce site est différente de celle des autres sites, comme la conservation et la fragmentation.
Pour continuer l’étude de la fragmentation, nous avons calculé différents indices et nous avons observé la fragmentation de divers types de restes.
3.2. Indice général d’intensité de la fragmentation : IGIF
Pour poursuivre l’étude de la fragmentation nous avons choisi de calculer l’indice général d’intensité de la fragmentation (IGIF) (Brugal & Patou-Mathis, 1993). Cet indice permet de quantifier la fragmentation générale du matériel pour un site en faisant le rapport entre le nombre de restes déterminables totalement et le nombre de restes total :
IGIF = % NRDt/ NRT.
Nous observons que l’IGIF est de 27,8 % en moyenne, ce qui n’est pas très élevé. Cet indice est de 18,9 % à Tanjung 63-64 et de 15,1 % à Dayu, ce qui est peu important. À Sendang Busik (25,9 %), à Grogol (25,5 %), à Ngrejeng (31,8 %) et à Bukuran (34,7 %) l’indice général d’intensité de la fragmentation est compris entre 25 et 35 %. Il est le plus important à Tanjung 82 avec 43,1 %.
À Holon, l’indice général d’intensité de la fragmentation est de 36 %, à Notarchirico il est de 39 %, à Ngebung A il est de 56 %, à Ngebung B de 53 % et Ngebung Upper de 46 %. Ces valeurs sont un peu supérieures à celles observées sur les sites de notre étude. Tout ceci confirme que les restes déterminables totalement sont minoritaires dans les assemblages et que donc le matériel a subi une grande fragmentation.
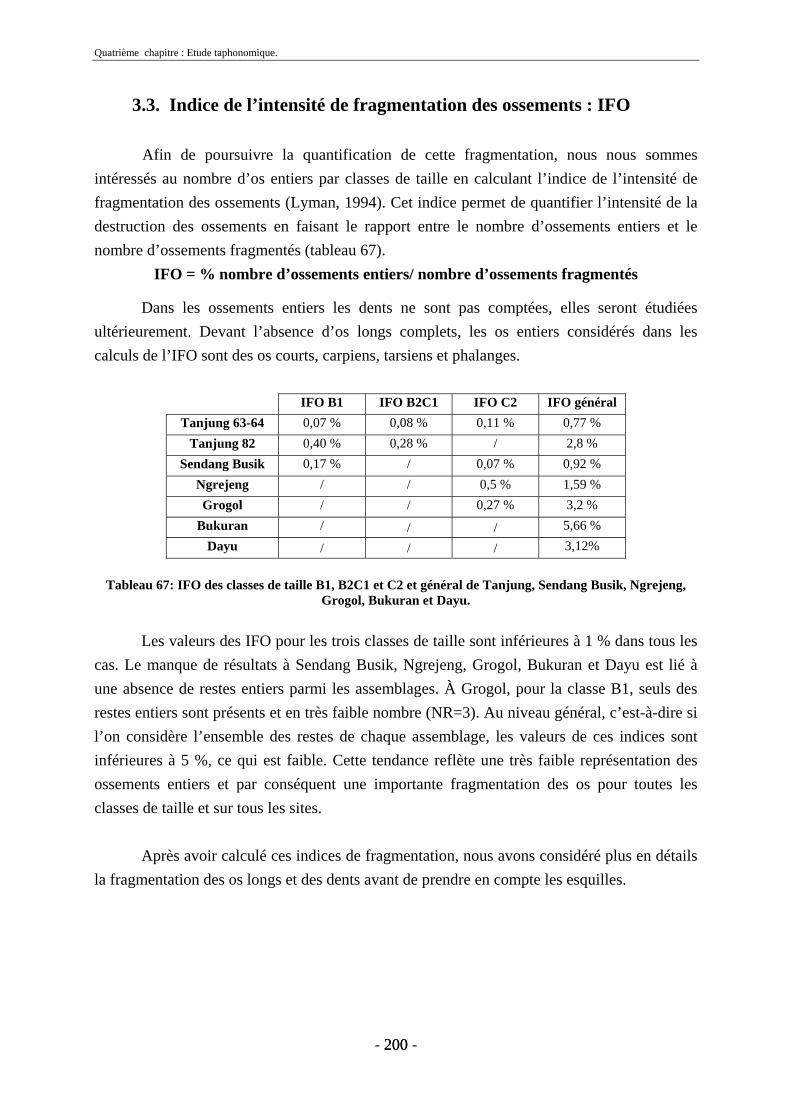
Quatrième chapitre : Etude taphonomique.
- 200 -
200
3.3. Indice de l’intensité de fragmentation des ossements : IFO
Afin de poursuivre la quantification de cette fragmentation, nous nous sommes intéressés au nombre d’os entiers par classes de taille en calculant l’indice de l’intensité de fragmentation des ossements (Lyman, 1994). Cet indice permet de quantifier l’intensité de la destruction des ossements en faisant le rapport entre le nombre d’ossements entiers et le nombre d’ossements fragmentés (tableau 67).
IFO = % nombre d’ossements entiers/ nombre d’ossements fragmentés
Dans les ossements entiers les dents ne sont pas comptées, elles seront étudiées ultérieurement. Devant l’absence d’os longs complets, les os entiers considérés dans les calculs de l’IFO sont des os courts, carpiens, tarsiens et phalanges.
IFO B1 IFO B2C1 IFO C2 IFO général Tanjung 63-64 0,07 % 0,08 % 0,11 % 0,77 %
Tanjung 82 0,40 % 0,28 % / 2,8 % Sendang Busik 0,17 % / 0,07 % 0,92 %
Ngrejeng / / 0,5 % 1,59 % Grogol / / 0,27 % 3,2 %
Bukuran / / / 5,66 % Dayu / / / 3,12%
Tableau 67: IFO des classes de taille B1, B2C1 et C2 et général de Tanjung, Sendang Busik, Ngrejeng,
Grogol, Bukuran et Dayu.
Les valeurs des IFO pour les trois classes de taille sont inférieures à 1 % dans tous les cas. Le manque de résultats à Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu est lié à une absence de restes entiers parmi les assemblages. À Grogol, pour la classe B1, seuls des restes entiers sont présents et en très faible nombre (NR=3). Au niveau général, c’est-à-dire si l’on considère l’ensemble des restes de chaque assemblage, les valeurs de ces indices sont inférieures à 5 %, ce qui est faible. Cette tendance reflète une très faible représentation des ossements entiers et par conséquent une importante fragmentation des os pour toutes les classes de taille et sur tous les sites.
Après avoir calculé ces indices de fragmentation, nous avons considéré plus en détails
la fragmentation des os longs et des dents avant de prendre en compte les esquilles.
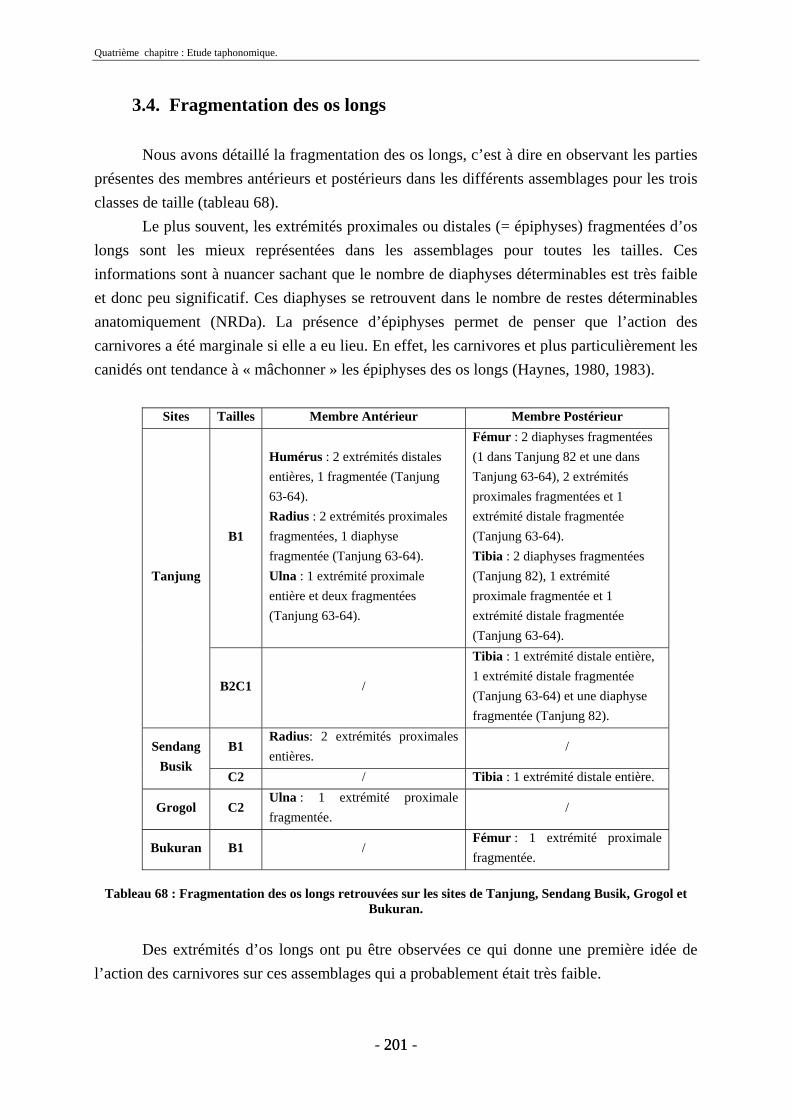
Quatrième chapitre : Etude taphonomique.
- 201 -
201
3.4. Fragmentation des os longs
Nous avons détaillé la fragmentation des os longs, c’est à dire en observant les parties présentes des membres antérieurs et postérieurs dans les différents assemblages pour les trois classes de taille (tableau 68).
Le plus souvent, les extrémités proximales ou distales (= épiphyses) fragmentées d’os longs sont les mieux représentées dans les assemblages pour toutes les tailles. Ces informations sont à nuancer sachant que le nombre de diaphyses déterminables est très faible et donc peu significatif. Ces diaphyses se retrouvent dans le nombre de restes déterminables anatomiquement (NRDa). La présence d’épiphyses permet de penser que l’action des carnivores a été marginale si elle a eu lieu. En effet, les carnivores et plus particulièrement les canidés ont tendance à « mâchonner » les épiphyses des os longs (Haynes, 1980, 1983).
Sites Tailles Membre Antérieur Membre Postérieur
B1
Humérus : 2 extrémités distales entières, 1 fragmentée (Tanjung 63-64). Radius : 2 extrémités proximales fragmentées, 1 diaphyse fragmentée (Tanjung 63-64). Ulna : 1 extrémité proximale entière et deux fragmentées (Tanjung 63-64).
Fémur : 2 diaphyses fragmentées (1 dans Tanjung 82 et une dans Tanjung 63-64), 2 extrémités proximales fragmentées et 1 extrémité distale fragmentée (Tanjung 63-64). Tibia : 2 diaphyses fragmentées (Tanjung 82), 1 extrémité proximale fragmentée et 1 extrémité distale fragmentée (Tanjung 63-64).
Tanjung
B2C1 /
Tibia : 1 extrémité distale entière, 1 extrémité distale fragmentée (Tanjung 63-64) et une diaphyse fragmentée (Tanjung 82).
B1 Radius: 2 extrémités proximales entières.
/ Sendang Busik
C2 / Tibia : 1 extrémité distale entière.
Grogol C2 Ulna : 1 extrémité proximale fragmentée.
/
Bukuran B1 / Fémur : 1 extrémité proximale fragmentée.
Tableau 68 : Fragmentation des os longs retrouvées sur les sites de Tanjung, Sendang Busik, Grogol et
Bukuran.
Des extrémités d’os longs ont pu être observées ce qui donne une première idée de l’action des carnivores sur ces assemblages qui a probablement était très faible.

Quatrième chapitre : Etude taphonomique.
- 202 -
202
Afin de compléter cette étude sur la fragmentation des ossements nous avons pris en compte les dents.
3.5. Fragmentation des dents
Dans la plupart des assemblages, les dents sont en proportion non négligeable et il nous a paru logique de les séparer des ossements et d’étudier plus précisément leur fragmentation (tableau 69).
Dents entières Dents fragmentées NR % NR % Tanjung 63-64 9 50 9 50
Tanjung 82 4 37 7 63 Sendang Busik 20 30,3 46 69,7
Ngrejeng 15 26,8 41 73,2 Grogol 4 16 21 84
Bukuran 9 23,7 29 76,3 Dayu 0 0 1 100
Tableau 69 : Nombre et proportions des dents entières et fragmentées dans les assemblages de Tanjung,
Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. Dans tous les assemblages, la majorité des dents sont fragmentées avec une moyenne d’environ 71 % du total des dents (tableau 68). Ceci révèle donc une mauvaise conservation générale sur tous les sites alors que les dents sont pourtant les éléments les plus minéralisés et les plus résistants à l’altération. Sachant qu’elles ne peuvent pas être utilisées comme élément nutritif, cette fragmentation est liée à l’importance de l’action édaphique ou diagénétique sur ces assemblages que nous quantifierons ultérieurement.
3.6. Forme des esquilles (NRI ou ND)
Pour prolonger l’étude de la fragmentation, nous nous sommes intéressés plus particulièrement aux fragments indéterminés ou esquilles. Nous avons effectué sur celles-ci un certain nombre de mesures : longueur, largeur et épaisseur.
Dans un premier temps, nous avons établi les histogrammes de fréquences de chaque dimension afin de mettre en évidence leur répartition pour les quatre sites considérés (tableau 70). Pour ce faire, afin de faciliter les comparaisons entre les sites nous avons volontairement écarté des esquilles hors norme : une esquille dont l’épaisseur était de 62 millimètres à Tanjung 63-64 (TDJ64/ ?/1108), quatre grosses esquilles dont la longueur était supérieure à 200 millimètres à Sendang Busik (SBI/D1/K8/23 ; SBI/D1/7/36 ; SBII/11/48 ; SBII/8/27) et deux esquilles de longueur 240 et 93 millimètres à Bukuran (BK79/IV/A1/1 ; BK79/I/C1/2). Ces esquilles sont des fragments d’ossements d’herbivores de très grande taille.
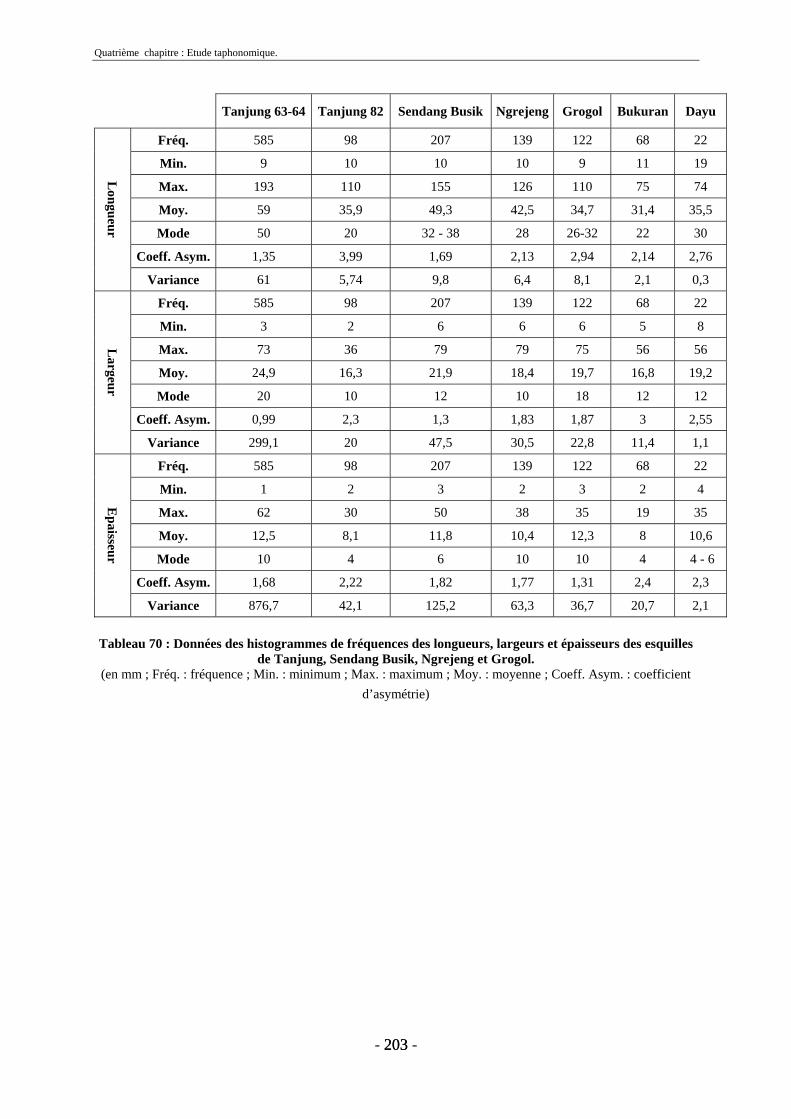
Quatrième chapitre : Etude taphonomique.
- 203 -
203
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran Dayu
Fréq. 585 98 207 139 122 68 22
Min. 9 10 10 10 9 11 19
Max. 193 110 155 126 110 75 74
Moy. 59 35,9 49,3 42,5 34,7 31,4 35,5
Mode 50 20 32 - 38 28 26-32 22 30
Coeff. Asym. 1,35 3,99 1,69 2,13 2,94 2,14 2,76
Longueur
Variance 61 5,74 9,8 6,4 8,1 2,1 0,3
Fréq. 585 98 207 139 122 68 22
Min. 3 2 6 6 6 5 8
Max. 73 36 79 79 75 56 56
Moy. 24,9 16,3 21,9 18,4 19,7 16,8 19,2
Mode 20 10 12 10 18 12 12
Coeff. Asym. 0,99 2,3 1,3 1,83 1,87 3 2,55
Largeur
Variance 299,1 20 47,5 30,5 22,8 11,4 1,1
Fréq. 585 98 207 139 122 68 22
Min. 1 2 3 2 3 2 4
Max. 62 30 50 38 35 19 35
Moy. 12,5 8,1 11,8 10,4 12,3 8 10,6
Mode 10 4 6 10 10 4 4 - 6
Coeff. Asym. 1,68 2,22 1,82 1,77 1,31 2,4 2,3
Epaisseur
Variance 876,7 42,1 125,2 63,3 36,7 20,7 2,1
Tableau 70 : Données des histogrammes de fréquences des longueurs, largeurs et épaisseurs des esquilles
de Tanjung, Sendang Busik, Ngrejeng et Grogol. (en mm ; Fréq. : fréquence ; Min. : minimum ; Max. : maximum ; Moy. : moyenne ; Coeff. Asym. : coefficient
d’asymétrie)

Quatrième chapitre : Etude taphonomique.
- 204 -
204
- Interprétation de la dimension des esquilles selon leur longueur (figure 57 et tableau 70): Nous observons que les répartitions sont différentes suivant les sites. À Tanjung 63-64, le mode des longueurs est à 50 millimètres et à Sendang Busik il est entre 32 et 38 millimètres, alors qu’il est plus faible sur tous les autres sites. Ceci confirme nos premières données sur la longueur plus importante des esquilles de Tanjung 63-64 (figure 55) et sur les résultats statistiques obtenus pour les distributions des longueurs de Tanjung 63-64 et Sendang Busik. La moyenne et la valeur maximale sont, également, plus importantes à Tanjung 63-64+ et à Sendang Busik que sur les autres sites. Néanmoins, les valeurs minimales sont assez comparables suivant les sites, aux alentours de 10 millimètres, excepté à Dayu où la longueur minimum est de 19 millimètres. Les esquilles de Tanjung 63-64 et Sendang Busik sont donc proportionnellement plus longues que sur les autres sites. Ceci est sans doute lié à la nature des espèces présentes dans les assemblages, en effet à Tanjung 63-64 et Sendang Busik la majorité des espèces sont des grands herbivores (chapitre II). La fracturation des os de grands herbivores produit donc des esquilles de plus grande taille. La distribution à Tanjung 63-64 est beaucoup plus étalée que sur les autres sites c’est à dire que les longueurs des esquilles sont plus mal classées.
+ tableaux 130 à 132, annexe 10.
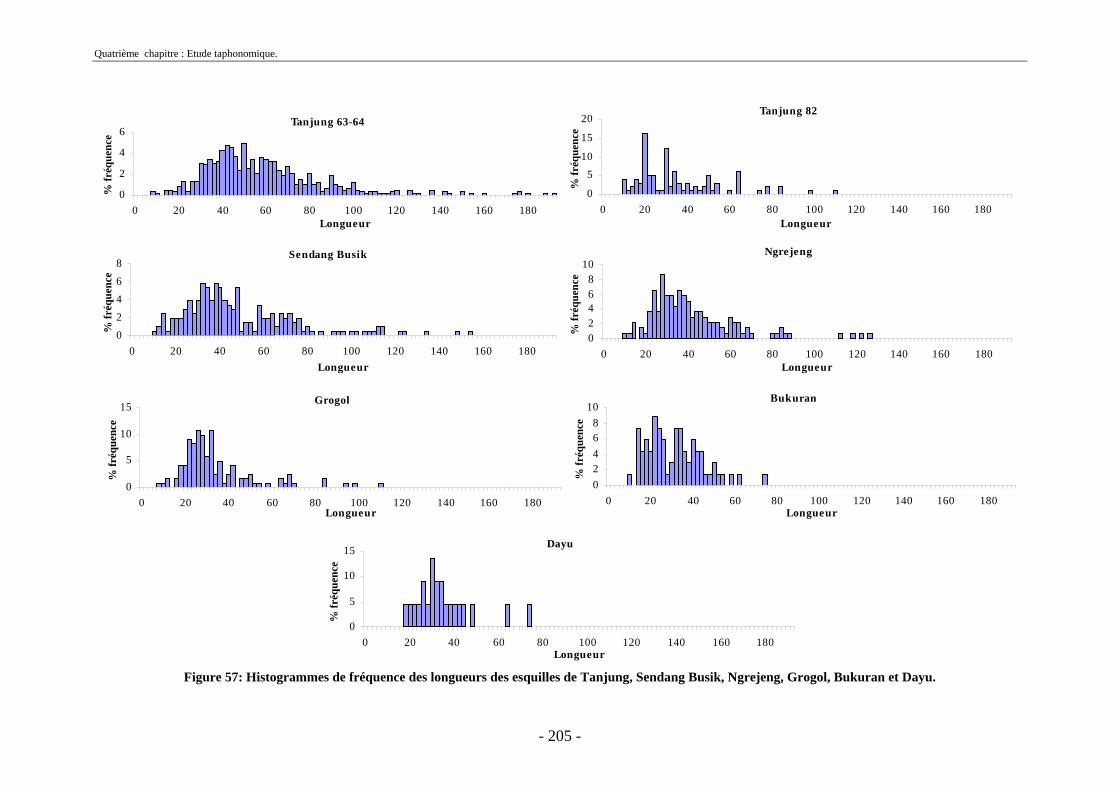
Quatrième chapitre : Etude taphonomique.
- 205 -
Tanjung 63-64
0
2
4
6
0 20 40 60 80 100 120 140 160 180Longueur
% fr
éque
nce
Tanjung 82
0
510
15
20
0 20 40 60 80 100 120 140 160 180Longueur
% fr
éque
nce
Sendang Busik
02468
0 20 40 60 80 100 120 140 160 180Longueur
% fr
éque
nce
Ngrejeng
02468
10
0 20 40 60 80 100 120 140 160 180Longueur
% fr
éque
nce
Grogol
0
5
10
15
0 20 40 60 80 100 120 140 160 180Longueur
% fr
éque
nce
Bukuran
02468
10
0 20 40 60 80 100 120 140 160 180Longueur
% fr
éque
nce
Dayu
0
5
10
15
0 20 40 60 80 100 120 140 160 180Longueur
% fr
éque
nce
Figure 57: Histogrammes de fréquence des longueurs des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.

Quatrième chapitre : Etude taphonomique.
- 206 -
- Interprétation de la dimension des esquilles selon leur largeur (figure 58 et tableau 70):
Les modes sont assez comparables pour Tanjung 63-64 et Grogol aux environs de 20 millimètres alors qu’à Sendang Busik, Ngrejeng, Bukuran et Dayu il n’est qu’à 10-12 millimètres. Les moyennes des largeurs des esquilles de Tanjung 63-64 et Sendang Busik sont assez proches et sont supérieures à celles des autres sites. La moyenne la plus faible est à Tanjung 82.
À Sendang Busik et à Ngrejeng, les valeurs maximums sont les plus importantes (79 millimètres). La largeur minimum à Tanjung 63-64 et 82 est inférieure aux valeurs minimums des autres sites. Les esquilles de Tanjung 63-64 et Sendang Busik paraissent donc plus larges que celles des autres sites. Ceci est sûrement dû à la nature des espèces présentes sur les sites comme pour les longueurs des esquilles.
De même que pour les longueurs la distribution des largeurs de Tanjung 63-64 est beaucoup plus étalée que pour les autres sites.
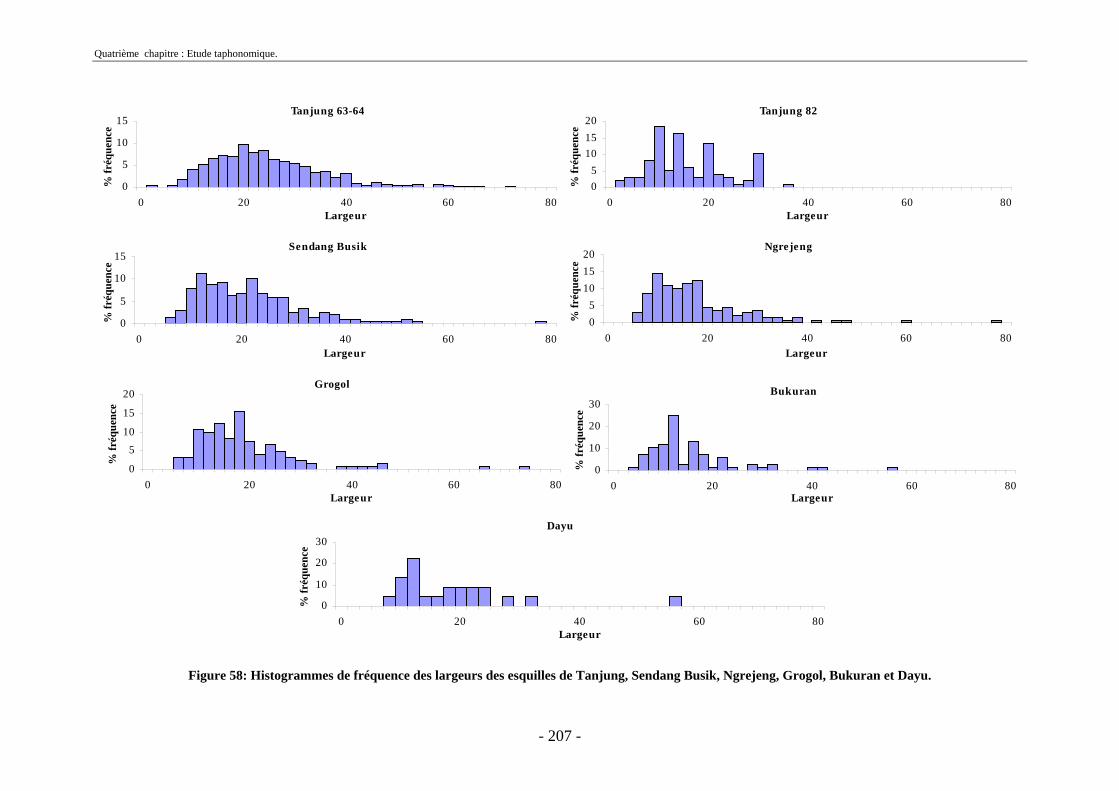
Quatrième chapitre : Etude taphonomique.
- 207 -
Tanjung 63-64
0
5
10
15
0 20 40 60 80Largeur
% fr
éque
nce
Tanjung 82
05
101520
0 20 40 60 80Largeur
% fr
éque
nce
Sendang Busik
0
5
10
15
0 20 40 60 80Largeur
% fr
éque
nce
Ngrejeng
05
101520
0 20 40 60 80Largeur
% fr
éque
nce
Grogol
0
510
15
20
0 20 40 60 80Largeur
% fr
éque
nce
Bukuran
0
10
20
30
0 20 40 60 80Largeur
% fr
éque
nce
Dayu
0
10
20
30
0 20 40 60 80Largeur
% fr
éque
nce
Figure 58: Histogrammes de fréquence des largeurs des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.

Quatrième chapitre : Etude taphonomique.
- 208 -
- Interprétation de la dimension des esquilles selon leur épaisseur (figure 59 et tableau 70):
Les valeurs des épaisseurs semblent plus homogènes que celles des longueurs et des largeurs. Ceci est logique car la fragmentation modifie, principalement, les ossements au niveau de la longueur et de la largeur. Les modes des épaisseurs sont identiques pour les sites de Tanjung 63-64, Ngrejeng et Grogol (10 millimètres). À Tanjung 82, Sendang Busik, Bukuran et Dayu, le mode des épaisseurs est un peu plus faible (4-6 millimètres). Les moyennes des épaisseurs sont comprises entre 10 et 12 millimètres environ, hormis à Bukuran où elle est de 8 millimètres. Les valeurs maximales des épaisseurs sont, également, les plus importantes à Tanjung 63-64 et Sendang Busik. Les valeurs minimales, quant à elles, sont assez proches sur l’ensemble des sites et comprises entre 1 et 4 millimètres. Il est donc plus difficile d’interpréter les résultats obtenus à partir des épaisseurs des esquilles car les épaisseurs des os sont liées à la nature de l’os, à la partie considérée de l’os et à l’espèce. En effet, l’épaisseur des esquilles peut être liée à la nature même des os, à l’origine des esquilles, à l’altération ou à la taille des animaux dont proviennent ces esquilles. Un ossement, suivant sa nature, peut être un os long, un os plat ou un os spongieux. De même, les esquilles obtenues à partir d’un même ossement, peuvent avoir une épaisseur différente suivant la partie dont elles sont extraites (diaphyse ou épiphyse). La taille de l’animal joue également un rôle, des os de grands animaux présenteront des épaisseurs plus importantes. Et l’altération peut produire également, si elle est suffisamment importante, une abrasion de la surface de l’os et donc réduire l’épaisseur de celui-ci.
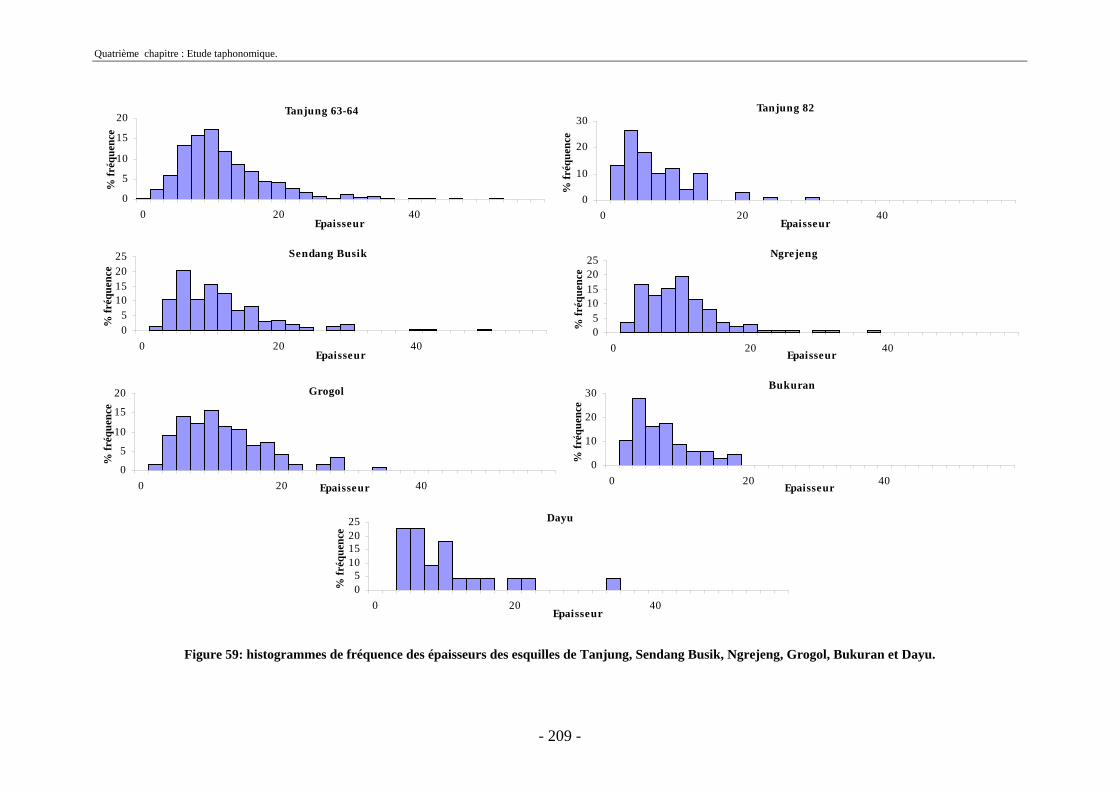
Quatrième chapitre : Etude taphonomique.
- 209 -
Tanjung 63-64
0
5
10
15
20
0 20 40Epaisseur
% fr
éque
nce
Tanjung 82
0
10
20
30
0 20 40Epaisseur
% fr
éque
nce
Sendang Busik
05
10152025
0 20 40Epaisseur
% fr
éque
nce
Ngrejeng
05
10152025
0 20 40Epaisseur
% fr
éque
nce
Grogol
0
5
10
15
20
0 20 40Epaisseur
% fr
éque
nce
Bukuran
0
10
20
30
0 20 40Epaisseur
% fr
éque
nce
Dayu
05
10152025
0 20 40Epaisseur
% fr
éque
nce
Figure 59: histogrammes de fréquence des épaisseurs des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.

Quatrième chapitre : Etude taphonomique.
- 210 -
À l’aide de ces trois variables (L, l et e) 3 types de sites semblent se dégager: - Les sites où les esquilles ont une longueur moyenne supérieure à 45 millimètres, une
largeur moyenne supérieure à 20 millimètres et une épaisseur moyenne supérieure à 11 millimètres : Tanjung 63-64 et Sendang Busik,
- Les sites intermédiaires : Ngrejeng, Grogol et Dayu, - Les sites où les esquilles ont des longueurs moyennes d’environ 35 millimètres, des
largeurs moyennes aux alentours de 16 millimètres et des épaisseurs moyennes d’environ 8 millimètres : Tanjung 82 et Bukuran. Pour intégrer ces trois variables, longueur (L), largeur (l) et épaisseur (e), dans un
même diagramme nous proposons de construire un diagramme triangulaire de forme et de sphéricité (tableau 71 et figures 60 à 66) (Sneed & Folk, 1958). Ce diagramme permet, en géologie, de placer les galets dans dix classes de formes différentes suivant leur degré de compaction, leur épaisseur et leur allongement (Sneed & Folk, 1958). Cette méthode a déjà donné des résultats en archéozoologie sur le site de la Baume de Goulon (Provence) daté du Paléolithique supérieur (Monchot, 1998) et sur celui d’Holon (Monchot & Horwitz, sous presse). Ces diagrammes permettent de mettre en évidence la forme des esquilles ce qui peut nous informer sur les phénomènes à l’origine de la fragmentation.
Les dix classes de formes sont définies comme suit: - C : compacte (compact), - CP : compacte et plate (compact and platy), - CB : compacte et lamellaire (compact and bladed), - CE : compacte et allongée (compact and elongated), - P : plate (platy), - B : lamellaire (bladed), - E : allongée (elongated), - VP : très plate (very platy), - VB : très lamellaire (very bladed), - VE : très allongée (very elongated).
Les côtés du triangle sont définis par les valeurs des rapports (e/L), (L-l/L-e) et le degré de sphéricité de l’os : ³√( e²/Ll).
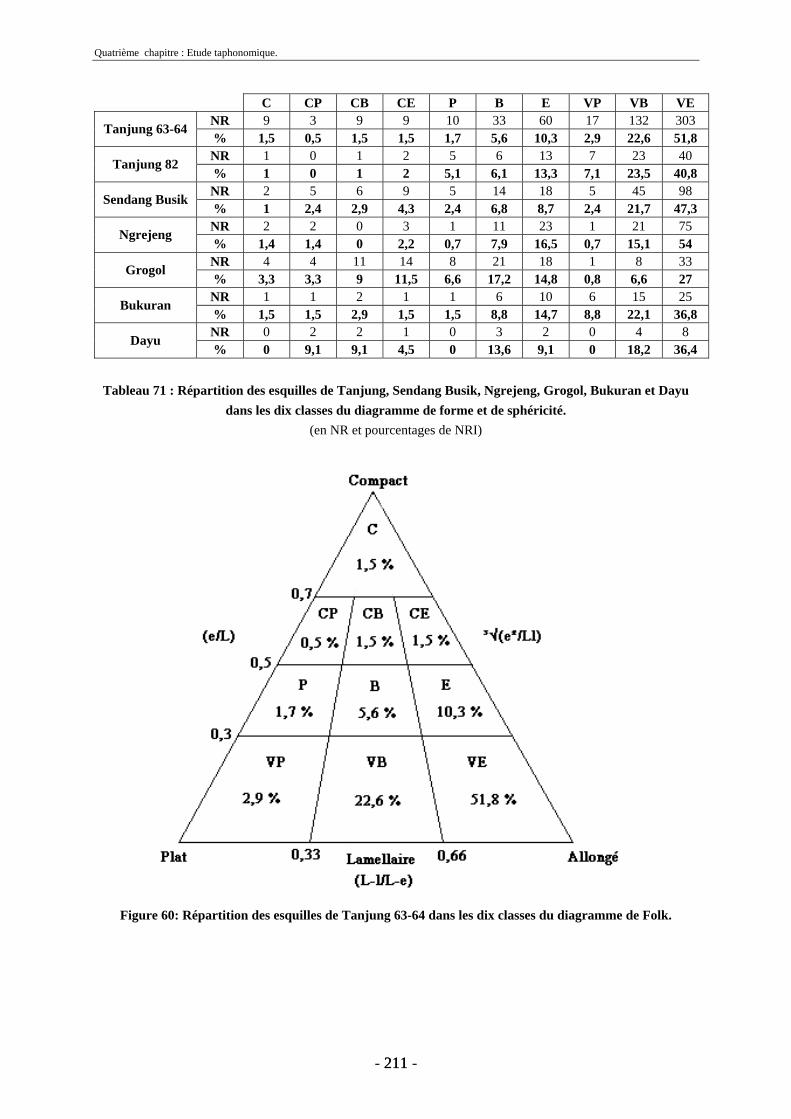
Quatrième chapitre : Etude taphonomique.
- 211 - 211
C CP CB CE P B E VP VB VE NR 9 3 9 9 10 33 60 17 132 303 Tanjung 63-64 % 1,5 0,5 1,5 1,5 1,7 5,6 10,3 2,9 22,6 51,8 NR 1 0 1 2 5 6 13 7 23 40 Tanjung 82 % 1 0 1 2 5,1 6,1 13,3 7,1 23,5 40,8 NR 2 5 6 9 5 14 18 5 45 98 Sendang Busik % 1 2,4 2,9 4,3 2,4 6,8 8,7 2,4 21,7 47,3 NR 2 2 0 3 1 11 23 1 21 75 Ngrejeng % 1,4 1,4 0 2,2 0,7 7,9 16,5 0,7 15,1 54 NR 4 4 11 14 8 21 18 1 8 33 Grogol % 3,3 3,3 9 11,5 6,6 17,2 14,8 0,8 6,6 27 NR 1 1 2 1 1 6 10 6 15 25 Bukuran % 1,5 1,5 2,9 1,5 1,5 8,8 14,7 8,8 22,1 36,8 NR 0 2 2 1 0 3 2 0 4 8 Dayu % 0 9,1 9,1 4,5 0 13,6 9,1 0 18,2 36,4
Tableau 71 : Répartition des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu
dans les dix classes du diagramme de forme et de sphéricité. (en NR et pourcentages de NRI)
Figure 60: Répartition des esquilles de Tanjung 63-64 dans les dix classes du diagramme de Folk.

Quatrième chapitre : Etude taphonomique.
- 212 - 212
Figure 61: Répartition des esquilles de Tanjung 82 dans les dix classes du diagramme de Folk.
Figure 62 : Répartition des esquilles de Sendang Busik dans les dix classes du diagramme de Folk.

Quatrième chapitre : Etude taphonomique.
- 213 - 213
Figure 63: Répartition des esquilles de Ngrejeng dans les dix classes du diagramme de Folk.
Figure 64 : Répartition des esquilles de Grogol dans les dix classes du diagramme de Folk.

Quatrième chapitre : Etude taphonomique.
- 214 - 214
Figure 65: Répartition des esquilles de Bukuran dans les dix classes du diagramme de Folk.
Figure 66: Répartition des esquilles de Dayu dans les dix classes du diagramme de Folk.

Quatrième chapitre : Etude taphonomique.
- 215 - 215
Sur tous les sites, excepté à Grogol, la classe de forme la mieux représentée est la classe VE suivie de la classe VB (figures 60 à 66). Les éléments très allongés et très lamellaires sont les mieux représentés. Le site de Grogol montre une distribution différente, la classe VE est toujours majoritaire mais en plus faible proportion que sur les autres sites et la classe suivante la mieux représentée est la classe B. Les éléments lamellaires (B) et allongés (E) sont mieux représentés à Grogol que sur les autres sites. Au niveau statistique cette tendance se retrouve : en effet, la distribution des classes d’esquilles à Grogol est significativement différente de celles des autres sites sauf pour Dayu (test de Kolmogorov-Smirnov, annexe 10). Les distributions des formes des esquilles à Tanjung ne sont pas significativement différentes de celles des autres sites sauf avec celle de Grogol. Tanjung ne s’individualise pas au niveau de la forme de ses esquilles par rapport aux autres sites de l’étude.
Dans les six sites considérés les esquilles présentant une forme très allongée (VE) sont les plus nombreuses. Cette forme des esquilles peut être liée à une fissuration lors de l’exposition à l’air libre des ossements (« weathering »), l’os est alors desséché et présente un bord de fracture rugueux, ou à une fracturation volontaire des ossements par l’homme ou les carnivores, dans ce cas sur os frais et le bord de fracture est lisse (Behrensmeyer, 1978 ; Villa & Mahieu, 1991 ; Bridault, 1994).
Ces fortes proportions des éléments très allongés ont également été observées sur le site de la Baume de Goulon (Salernes, Var) où 64,4 % des esquilles sont de type très allongées (Monchot, 1998 ; annexe 8) et sur le site d’Holon où 43,3 % des esquilles sont de type très allongées (Monchot & Horwitz, sous presse ; annexe 8). Nous avons testé ces proportions à l’aide d’un test de Kolmogorov-Smirnov+. Il en ressort que la distribution d’Holon est différente significativement de celles de Tanjung 63-64, de Grogol et de Ngrejeng. La distribution des esquilles pour la Baume de Goulon est significativement différente de celle de Grogol et de Bukuran. Il est donc difficile de conclure sur l’origine de cette fracturation. La fracturation intentionnelle sur os frais est donc peut-être absente ou beaucoup moins présente sur les sites de notre étude qu’à Holon et à la Baume de Goulon.
Cette classification n’étant pas très facile à utiliser dans notre analyse, nous avons choisi de réduire les 10 classes proposées. Nous avons regroupé toutes les classes d’éléments compacts, toutes les classes d’éléments plats, toutes les classes d’éléments lamellaires et enfin toutes les classes d’éléments allongés (tableau 72).
+ tableau 133, annexe 10.

Quatrième chapitre : Etude taphonomique.
- 216 - 216
C P B E Tanjung 63-64 5,1 % 5 % 28,2 % 62,1 %
Tanjung 82 4,1 % 12 % 29,6 % 54,1 % Sendang Busik 10,6 % 4,8 % 28,5 % 56 %
Ngrejeng 5 % 1,4 % 23 % 70,5 % Grogol 27 % 7,4 % 23,8 % 41,8 %
Bukuran 7,4 % 10,3 % 30,9 % 51,5 % Dayu 22,7 % 0 % 31,8 % 45,5 %
Tableau 72 : Répartition des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu
dans les classes de formes. (en pourcentages de NRI)
La majorité des esquilles de tous les sites sont de type allongé (E). Nous pouvons mettre en évidence deux grandes tendances dans les formes d’esquille. À Grogol et à Dayu, les classes des éléments compacts et lamellaires sont en proportions assez importantes et comparables alors que la classe des éléments plats est faible ou absente. Sur les autres sites, les classes des éléments compacts et plats sont en proportions beaucoup plus faibles que les classes des éléments lamellaires et allongés. Nous avons observé le nombre d’individus de grande taille présents dans ces assemblages. À Tanjung 63-64, sur 8 individus déterminés, 5 sont des grands herbivores. À Tanjung 82, sur 6 individus, 3 sont des grands herbivores. À Sendang Busik, sur 5 individus déterminés, 3 sont des grands herbivores. À Ngrejeng et à Grogol, aucun individu de grande taille n’a été déterminé. Enfin, à Bukuran, sur 6 individus, un seul a été attribué à un grand herbivore. Il ne semble pas y avoir de lien entre les sites où les grands herbivores sont majoritaires et la forme des esquilles. En effet, à Bukuran et Ngrejeng nous observons les mêmes tendances que pour Tanjung et Sendang Busik alors que les grands herbivores paraissent moins présents dans les assemblages. De même, pour les épiphyses, nous avons compté le nombre d’épiphyses entières d’os longs (métapodes compris) tous taxons confondus sur tous les sites. À Tanjung 63-64, neuf épiphyses ont été comptées, à Tanjung 82 deux, à Sendang Busik trois, à Ngrejeng deux et à Grogol une. À Bukuran et Dayu, nous n’en avons pas observé. À Grogol et Dayu, où le nombre d’esquilles de type compact est le plus important, nous observons un nombre d’épiphyses entières très faible ou nul et une absence d’espèces de grands herbivores. La classe compacte est donc peut-être liée à une plus grande fragmentation des épiphyses qui produit des esquilles plus compactes alors que les esquilles allongées sont liées à une plus grande fragmentation des diaphyses des os longs.
Après avoir observé la répartition spatiale, une mauvaise conservation générale des assemblages et une grande fragmentation des restes, nous pouvons nous demander quels sont les agents à l’origine de ces phénomènes et quels sont ceux qui peuvent expliquer les différences observées entre Tanjung et les autres sites.

Quatrième chapitre : Etude taphonomique.
- 217 - 217
4. AGENTS TAPHONOMIQUES
Nous verrons, parmi les agents taphonomiques, tout d’abord, les agents climato-édaphiques et le transport fluviatile, puis, les agents biologiques non-anthropiques et enfin l’action anthropique.
4.1. Agents climato-édaphiques
4.1.1. Climat Les facteurs climatiques interviennent avant et après l’enfouissement des os. Pour les
zones tropicales, la conservation est généralement mauvaise. Le processus de dessication – hydratation et l’activité biologique des racines et des micro-organismes peuvent provoquer la fragmentation des ossements avant l’enfouissement (Auguste, 1994a). Mais la conservation est possible si l’enfouissement est rapide.
L’action du climat peut aussi intervenir après le dépôt et donc agir sur la percolation et sur la végétation, par exemple.
Sur tous les sites, nous pouvons penser que l’action du climat avant l’enfouissement n’a pas été très importante car les os ont été conservés et que l’enfouissement a donc été assez rapide. Nous préciserons ceci lors de l’étude de la fissuration.
4.1.2. Formation du site fossilifère
La nature du site influe largement sur la concentration des ossements. Les sites de notre étude sont des sites de plein air. Si le contexte géomorphologique et sédimentaire est favorable, l’enfouissement peut être rapide permettant une conservation des ossements. Par contre, si l’évolution géologique est importante, les os peuvent être détériorés et disparaître (Behrensmeyer, 1978 ; Auguste, 1994b).
Les dépôts sédimentaires (formation de Kabuh) des sites de notre étude suggèrent des
dépôts fluviatiles et donc la proximité d’un cours d’eau par rapport au milieu de dépôt. Les accumulations osseuses peuvent être alors le produit d’un transport fluviatile et d’un dépôt en un lieu privilégié des cadavres, de manière régulière ou lors d’une crue (Badgley, 1986a ; Behrensmeyer, 1988 ; Auguste, 1994b). La conservation des ossements sur un site de plein air dépend de conditions locales variables. Plus un os se retrouvera éloigné du lit majeur d’une rivière plus il aura des chances de se conserver car il subira moins les effets du transport. Nos différences de conservation sur le site de Tanjung par rapport aux autres sites

Quatrième chapitre : Etude taphonomique.
- 218 - 218
peut s’expliquer probablement en partie par la position éloignée du site par rapport au lit d’une paléorivière.
4.1.3. Nature des dépôts
En ce qui concerne le sol, il joue le rôle de milieu de conservation pour les restes
fauniques et donc sa nature va influencer leur conservation et leur fossilisation (Auguste, 1994b). Un sédiment à éléments grossiers est un mauvais milieu de conservation et de fossilisation pour les ossements car lors de son dépôt il peut avoir un effet mécanique sur eux et il peut y avoir une certaine porosité qui sera propice à l’infiltration des eaux. Au contraire, les sédiments fins sont peu poreux ce qui ne favorise pas l’écoulement des eaux ni la destruction des ossements.
Les dépôts volcano-sédimentaires de la formation de Kabuh sont principalement constitués par des sables (Lizon-Sureau, 1979). Ce type de dépôt n’est normalement pas favorable à une bonne conservation des ossements car la taille de ses éléments est assez importante. Cependant, dans la région du dôme de Sangiran, et plus précisément dans la formation de Kabuh de nombreux fossiles ont été découverts malgré la nature des dépôts sédimentaires.
Dans la région du rift est africain, des environnements possédant de hautes concentrations en calcium, comme ceux où le type de volcanisme est fortement calcique, semblent favoriser la minéralisation des os et leur plus grande résistance aux destructions de surface (Behrensmeyer, 1973).
À titre d’exemple, la formation volcano-sédimentaire de Kabuh à Bapang a été divisée en 5 cycles (C1 à C5) (Larick et al., 2001). Chaque cycle présente deux faciès différents, un faciès sédimentaire (a) et un faciès volcanique (b). Les fossiles sont présents dans différents cycles mais toujours dans le faciès « a ». À Tanjung les fossiles auraient été découverts dans le cycle C3a et à Sendang Busik dans le cycle C2a (Larick et al., 2001, figure 68). Les fragments de ponces associés au faciès « a » montrent une composition chimique composée principalement de minéraux sodi-calciques (phénocristaux de plagioclase) ou calciques (phénocristaux de hornblende) et d’apatite (Ca5(PO4)3). Ce dernier minéral rentre dans l’épigénie des débris d’os et des dents c’est à dire qu’il remplace progressivement les minéraux constituant l’os ou la dent, au cours de la fossilisation (Foucault & Raoult, 1995). Les abondances de ces minéraux sont proches de celles des fragments de ponces du volcan Lawu situé à 30 km à l’est (Larick et al., 2001). Par conséquent, après l’enfouissement, la percolation a pu transporter ces minéraux provenant de l’altération de ces fragments de ponces qui ont favorisé la fossilisation et donc la conservation des ossements.
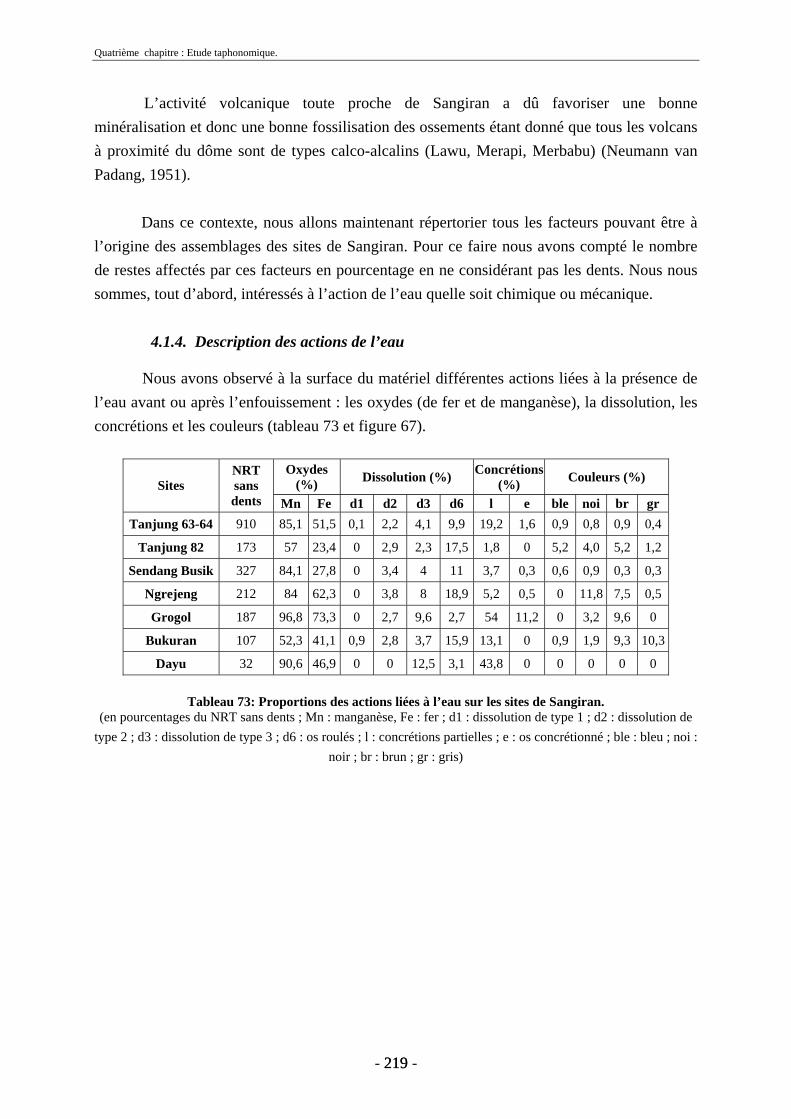
Quatrième chapitre : Etude taphonomique.
- 219 - 219
L’activité volcanique toute proche de Sangiran a dû favoriser une bonne minéralisation et donc une bonne fossilisation des ossements étant donné que tous les volcans à proximité du dôme sont de types calco-alcalins (Lawu, Merapi, Merbabu) (Neumann van Padang, 1951).
Dans ce contexte, nous allons maintenant répertorier tous les facteurs pouvant être à
l’origine des assemblages des sites de Sangiran. Pour ce faire nous avons compté le nombre de restes affectés par ces facteurs en pourcentage en ne considérant pas les dents. Nous nous sommes, tout d’abord, intéressés à l’action de l’eau quelle soit chimique ou mécanique.
4.1.4. Description des actions de l’eau
Nous avons observé à la surface du matériel différentes actions liées à la présence de
l’eau avant ou après l’enfouissement : les oxydes (de fer et de manganèse), la dissolution, les concrétions et les couleurs (tableau 73 et figure 67).
Oxydes (%) Dissolution (%) Concrétions
(%) Couleurs (%) Sites NRT sans dents Mn Fe d1 d2 d3 d6 l e ble noi br gr
Tanjung 63-64 910 85,1 51,5 0,1 2,2 4,1 9,9 19,2 1,6 0,9 0,8 0,9 0,4
Tanjung 82 173 57 23,4 0 2,9 2,3 17,5 1,8 0 5,2 4,0 5,2 1,2
Sendang Busik 327 84,1 27,8 0 3,4 4 11 3,7 0,3 0,6 0,9 0,3 0,3
Ngrejeng 212 84 62,3 0 3,8 8 18,9 5,2 0,5 0 11,8 7,5 0,5
Grogol 187 96,8 73,3 0 2,7 9,6 2,7 54 11,2 0 3,2 9,6 0
Bukuran 107 52,3 41,1 0,9 2,8 3,7 15,9 13,1 0 0,9 1,9 9,3 10,3
Dayu 32 90,6 46,9 0 0 12,5 3,1 43,8 0 0 0 0 0
Tableau 73: Proportions des actions liées à l’eau sur les sites de Sangiran.
(en pourcentages du NRT sans dents ; Mn : manganèse, Fe : fer ; d1 : dissolution de type 1 ; d2 : dissolution de type 2 ; d3 : dissolution de type 3 ; d6 : os roulés ; l : concrétions partielles ; e : os concrétionné ; ble : bleu ; noi :
noir ; br : brun ; gr : gris)
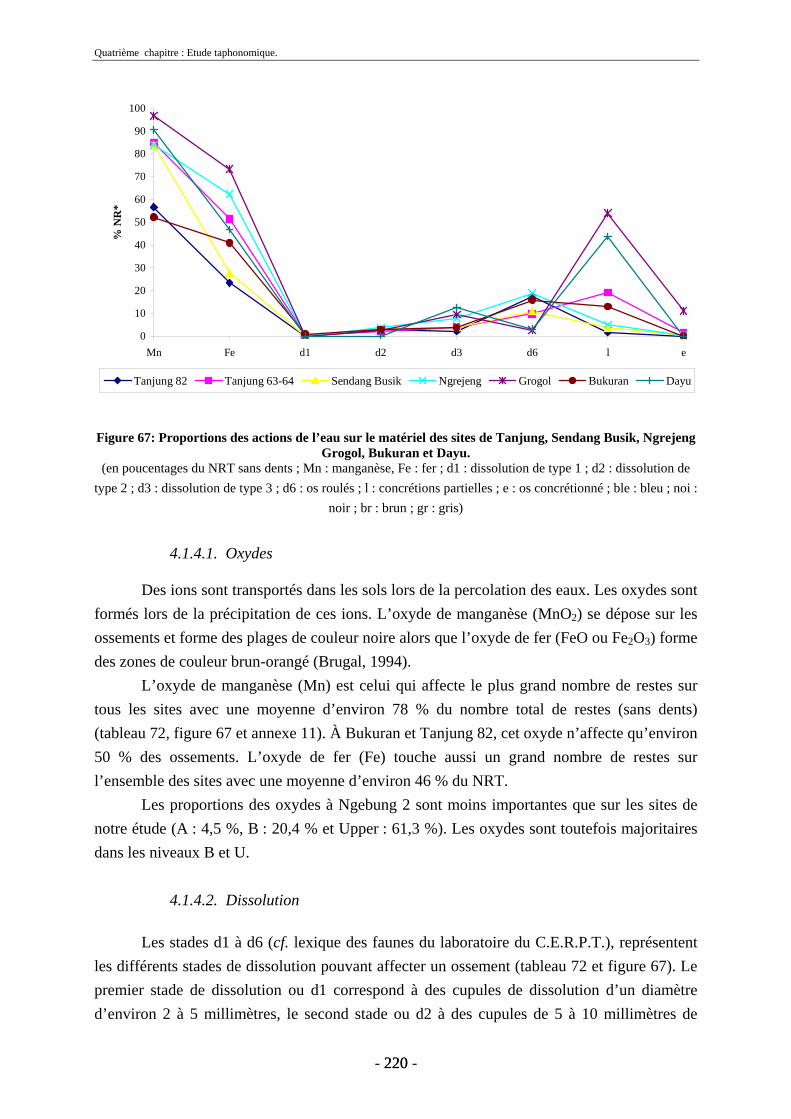
Quatrième chapitre : Etude taphonomique.
- 220 - 220
0
10
20
30
40
50
60
70
80
90
100
Mn Fe d1 d2 d3 d6 l e
% N
R*
Tanjung 82 Tanjung 63-64 Sendang Busik Ngrejeng Grogol Bukuran Dayu
Figure 67: Proportions des actions de l’eau sur le matériel des sites de Tanjung, Sendang Busik, Ngrejeng
Grogol, Bukuran et Dayu. (en poucentages du NRT sans dents ; Mn : manganèse, Fe : fer ; d1 : dissolution de type 1 ; d2 : dissolution de
type 2 ; d3 : dissolution de type 3 ; d6 : os roulés ; l : concrétions partielles ; e : os concrétionné ; ble : bleu ; noi : noir ; br : brun ; gr : gris)
4.1.4.1. Oxydes
Des ions sont transportés dans les sols lors de la percolation des eaux. Les oxydes sont
formés lors de la précipitation de ces ions. L’oxyde de manganèse (MnO2) se dépose sur les ossements et forme des plages de couleur noire alors que l’oxyde de fer (FeO ou Fe2O3) forme des zones de couleur brun-orangé (Brugal, 1994).
L’oxyde de manganèse (Mn) est celui qui affecte le plus grand nombre de restes sur tous les sites avec une moyenne d’environ 78 % du nombre total de restes (sans dents) (tableau 72, figure 67 et annexe 11). À Bukuran et Tanjung 82, cet oxyde n’affecte qu’environ 50 % des ossements. L’oxyde de fer (Fe) touche aussi un grand nombre de restes sur l’ensemble des sites avec une moyenne d’environ 46 % du NRT.
Les proportions des oxydes à Ngebung 2 sont moins importantes que sur les sites de notre étude (A : 4,5 %, B : 20,4 % et Upper : 61,3 %). Les oxydes sont toutefois majoritaires dans les niveaux B et U.
4.1.4.2. Dissolution
Les stades d1 à d6 (cf. lexique des faunes du laboratoire du C.E.R.P.T.), représentent
les différents stades de dissolution pouvant affecter un ossement (tableau 72 et figure 67). Le premier stade de dissolution ou d1 correspond à des cupules de dissolution d’un diamètre d’environ 2 à 5 millimètres, le second stade ou d2 à des cupules de 5 à 10 millimètres de

Quatrième chapitre : Etude taphonomique.
- 221 - 221
diamètre, le troisième stade ou d3 à des zones de desquamation de plusieurs centimètres sur l’ensemble de l’os et le dernier stade ou d6 à un aspect roulé de l’os lié au transport de l’os.
L’importance de la dissolution de type 1 est minoritaire sur tous les assemblages quand elle est observable. La d3 affecte un plus grand nombre de restes que la d2 sur tous les sites mais cette action est mineure, elle touche 6,3 % des restes en moyenne dans un assemblage. La d6, à Tanjung (pour les deux niveaux), Sendang Busik, Bukuran et Ngrejeng est majoritaire, ce qui signifie qu’environ 14,6 % en moyenne des restes de ces assemblages ont subi un transport. Alors qu’à Grogol et Dayu l’importance de la d6 est nettement plus faible, environ 3 % du NRT.
Le stade de dissolution affectant le plus de restes est d6 suggèrant qu’une partie du matériel a subi un transport mais cette action reste mineure sur l’ensemble des assemblages. Ces restes roulés sont principalement des esquilles.
Les traces de dissolutions de type d1 et d2 sont plus nombreuses à Ngebung 2 que sur les sites considérés dans notre étude et le phénomène s’inverse pour les dissolutions de type 3 et 6 (tableau 74).
Dissolution (%)
Niveaux NRT sans dents d1 d2 d3 d6
A 729 3,2 8,9 2,3 5,0 B 358 3,5 8,5 1,4 4,2
Upper 324 2,8 8,5 2,4 7,3
Tableau 74 : Proportions des restes affectés par les différents types de dissolution dans les trois niveaux de Ngebung 2.
(en pourcentages du NRT sans dents de chaque niveau)
4.1.4.3. Concrétions
Les concrétions de type « l » sont des concrétions présentes sur une partie de l’os et celles de type « e » recouvrent tout l’os (cf. lexique des faunes du laboratoire du C.E.R.P.T.) (tableau 72 et figure 67).
Les concrétions de type « l » affectent un grand nombre de restes notamment dans les assemblages de Grogol et de Dayu avec 54 % et 43,8 % respectivement des ossements présentant des concrétions pour seulement 19,2 % à Tanjung 63-64. Les concrétions de type « e » sont moins présentes sur les restes que celles de type « l ». À Bukuran et Dayu elles sont absentes et à Tanjung, Sendang Busik et Ngrejeng elles sont mineures. Néanmoins, à Grogol, les concrétions de type « e » affectent 11,2 % des restes.
La présence de ces concrétions confirme des infiltrations d’eau chargée en minéraux dans tous les sites et plus particulièrement à Grogol et Dayu.

Quatrième chapitre : Etude taphonomique.
- 222 - 222
4.1.4.4. Couleurs
Nous avons distingué 4 couleurs différentes : bleu (ble), noir (noi), brun (br) et gris (gr) sur tous les restes (dents comprises) sur l’ensemble des sites hormis à Dayu (tableau 72 et figure 67). Un reste coloré présente une teinte bleue, noire, grise ou brune sur toute sa surface. Cette coloration n’est pas due à l’action du feu mais à des éléments transportés par l’eau comme peuvent l’être le fer ou le manganèse.
À Tanjung 63-64 et à Sendang Busik cette action est mineure, peu de restes sont colorés. Le plus grand nombre d’ossements colorés a été observé à Ngrejeng et à Bukuran et nous savons que les restes de ces deux sites proviennent de la formation de Kabuh et de Pucangan. La coloration des os paraît plus accentuée sur les sites dont certains restes peuvent provenir de la formation de Pucangan. Ceci pourrait alors s’expliquer par la forte présence de matière organique dans cette formation qui a pu colorer les os d’une manière caractéristique.
Après l’étude des traces liées aux actions de l’eau, nous nous sommes intéressés aux
manifestations de l’action du climat sur les ossements.
4.1.5. Fissuration
En outre, des traces laissées par le « weathering » sont observables sur les fossiles des sites de Sangiran (tableau 75 et annexe 11).
Le « weathering » a été défini comme « la décomposition naturelle des os et des dents par des processus physico-chimiques opérant sur ou dans le sol » (Behrensmeyer, 1990). De nombreux facteurs peuvent influencer le weathering : l’humidité, la température, le pH du sol, la taille et l’âge des animaux, le type d’os, la durée de l’exposition au soleil et aux intempéries et la végétation (Behrensmeyer, 1978). Ce processus va être progressif de la mort de l’animal à la disparition des ossements ou bien à leur enfouissement. Six étapes de conservation du reste osseux ont donc pu être distinguées au cours du temps après la mort de l’animal sur des sites de l’est africain (Behrensmeyer, 1978):
- stade 0 : aucun signe de détérioration de l’os ⇒ 0 à 1 an, - stade 1 : apparition de craquelures parallèles à la structure fibreuse (cracking) ⇒ 0
à 3 ans, - stade 2 : apparition d’écaillements à la surface de l’os (flacking) ⇒ 2 à 6 ans, - stade 3 : la surface des os devient rugueuse et fibreuse ⇒ 4 à 15 ans, - stade 4 : ouverture des craquelures, des esquilles se forment (splitting) ⇒ 6 à 15
ans et plus, - stade 5 : détachement de grosses esquilles ⇒ 6 à 15 ans et plus.
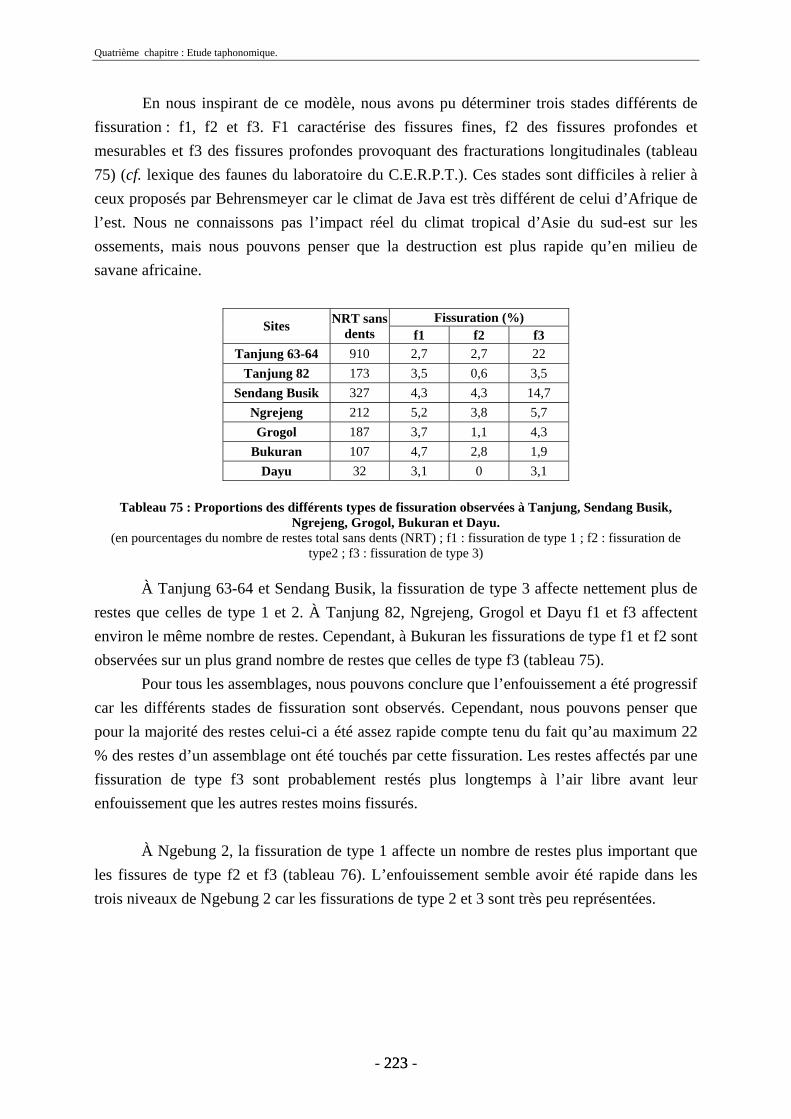
Quatrième chapitre : Etude taphonomique.
- 223 - 223
En nous inspirant de ce modèle, nous avons pu déterminer trois stades différents de fissuration : f1, f2 et f3. F1 caractérise des fissures fines, f2 des fissures profondes et mesurables et f3 des fissures profondes provoquant des fracturations longitudinales (tableau 75) (cf. lexique des faunes du laboratoire du C.E.R.P.T.). Ces stades sont difficiles à relier à ceux proposés par Behrensmeyer car le climat de Java est très différent de celui d’Afrique de l’est. Nous ne connaissons pas l’impact réel du climat tropical d’Asie du sud-est sur les ossements, mais nous pouvons penser que la destruction est plus rapide qu’en milieu de savane africaine.
Fissuration (%) Sites NRT sans
dents f1 f2 f3 Tanjung 63-64 910 2,7 2,7 22
Tanjung 82 173 3,5 0,6 3,5 Sendang Busik 327 4,3 4,3 14,7
Ngrejeng 212 5,2 3,8 5,7 Grogol 187 3,7 1,1 4,3
Bukuran 107 4,7 2,8 1,9 Dayu 32 3,1 0 3,1
Tableau 75 : Proportions des différents types de fissuration observées à Tanjung, Sendang Busik,
Ngrejeng, Grogol, Bukuran et Dayu. (en pourcentages du nombre de restes total sans dents (NRT) ; f1 : fissuration de type 1 ; f2 : fissuration de
type2 ; f3 : fissuration de type 3)
À Tanjung 63-64 et Sendang Busik, la fissuration de type 3 affecte nettement plus de restes que celles de type 1 et 2. À Tanjung 82, Ngrejeng, Grogol et Dayu f1 et f3 affectent environ le même nombre de restes. Cependant, à Bukuran les fissurations de type f1 et f2 sont observées sur un plus grand nombre de restes que celles de type f3 (tableau 75).
Pour tous les assemblages, nous pouvons conclure que l’enfouissement a été progressif car les différents stades de fissuration sont observés. Cependant, nous pouvons penser que pour la majorité des restes celui-ci a été assez rapide compte tenu du fait qu’au maximum 22 % des restes d’un assemblage ont été touchés par cette fissuration. Les restes affectés par une fissuration de type f3 sont probablement restés plus longtemps à l’air libre avant leur enfouissement que les autres restes moins fissurés.
À Ngebung 2, la fissuration de type 1 affecte un nombre de restes plus important que
les fissures de type f2 et f3 (tableau 76). L’enfouissement semble avoir été rapide dans les trois niveaux de Ngebung 2 car les fissurations de type 2 et 3 sont très peu représentées.

Quatrième chapitre : Etude taphonomique.
- 224 - 224
Fissuration (%) Niveaux
NRT sans dents f1 f2 f3
A 729 6,3 1,0 0,3 B 358 10,2 0,7 0,7
Upper 324 8,1 1,6 1,2
Tableau 76 : Proportions des restes affectés par les différents types de fissuration dans les trois niveaux de Ngebung 2.
(en pourcentages du NRT sans dents de chaque niveau ; f1 : fissuration de type 1 ; f2 : fissuration de type2 ; f3 : fissuration de type 3)
4.1.6. Conclusion L’action chimique de l’eau sur ces assemblages est prédominante (tableau 72 et figure
67). En effet, des traces d’oxydes de manganèse et de fer ont été observées sur une grande partie des restes. De plus, des ossements présentent des concrétions (l et e) et des traces de dissolution (d1, d2 et d3). Enfin, nous avons pu observer une coloration sur divers os. Tous ces stigmates suggèrent un milieu humide avant et pendant l’enfouissement. Ceci pourrait être probablement lié au climat (pluies) mais aussi à la proximité d’un cours d’eau (transport des ossements ou crues). Ceci est confirmé par le fait qu’une partie des restes semble avoir été « roulée » (d6) synonyme de transport par l’eau. L’eau a donc eu également une action mécanique, certes moins importante que son action chimique mais non négligeable, sur les ossements des différents assemblages. Il paraît donc indispensable d’analyser l’action du transport fluviatile sur ces assemblages. Enfin, les proportions des différents types de fissuration indiquent un enfouissement progressif des ossements, mais cette action n’est pas majeure sur les assemblages.
Les ossements des niveaux de Ngebung 2, paraissent avoir été enfouis assez rapidement car la fissuration de type f3 est négligeable. Ces restes ont subi l’action chimique de l’eau mais d’une manière moins intense que les restes des sites de notre étude. En ce qui concerne l’action mécanique de l’eau, elle semble négligeable au vu des proportions des restes roulés (d6). Les assemblages des niveaux de Ngebung 2 ont moins subi l’influence des agents climato-édaphiques que les assemblages des sites considérés ici. Le niveau U paraît le plus proche au niveau taphonomique des assemblages des sites de notre étude.
4.2. Transport fluviatile
Les fossiles des sites de Sangiran ont été découverts totalement ou en partie dans la formation de Kabuh qui est une formation volcano-sédimentaire en contexte fluviatile. Il est important de considérer l’allochtonie ou l’autochtonie de ces restes osseux par rapport à leur milieu de dépôt initial. Nous avons choisi d’utiliser différentes méthodes d’étude du transport fluviatile pour recouper et confirmer nos résultats.
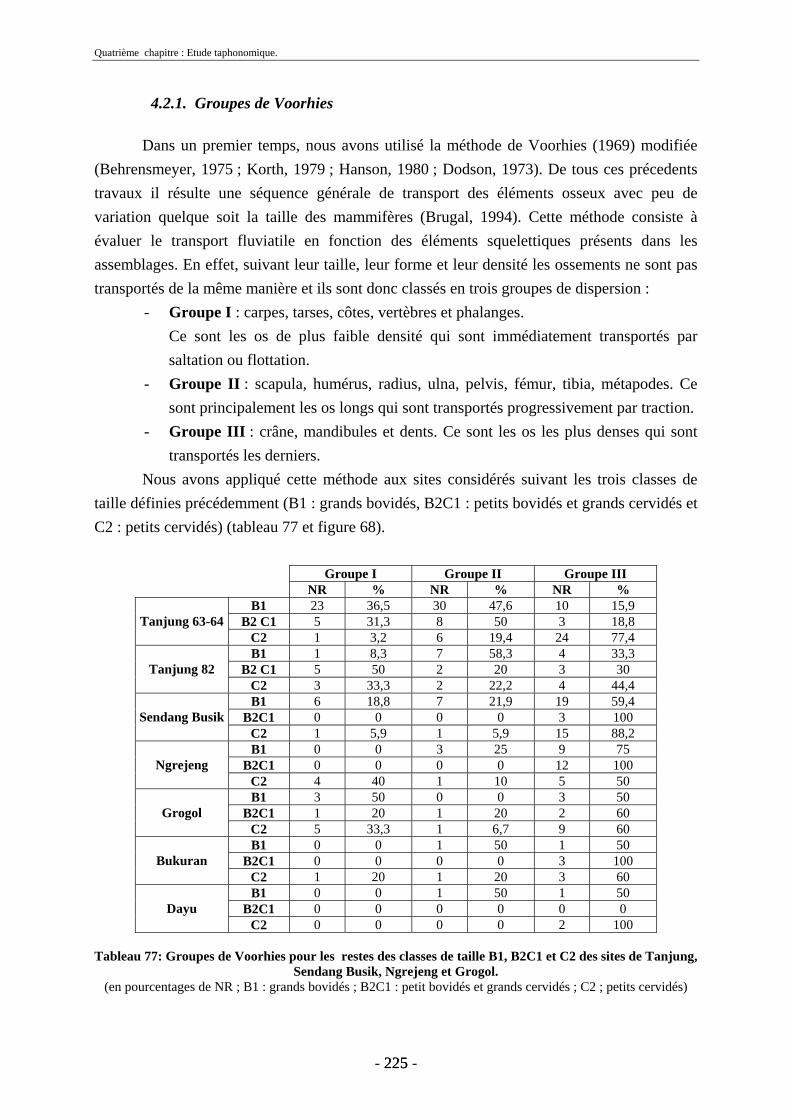
Quatrième chapitre : Etude taphonomique.
- 225 - 225
4.2.1. Groupes de Voorhies
Dans un premier temps, nous avons utilisé la méthode de Voorhies (1969) modifiée (Behrensmeyer, 1975 ; Korth, 1979 ; Hanson, 1980 ; Dodson, 1973). De tous ces précedents travaux il résulte une séquence générale de transport des éléments osseux avec peu de variation quelque soit la taille des mammifères (Brugal, 1994). Cette méthode consiste à évaluer le transport fluviatile en fonction des éléments squelettiques présents dans les assemblages. En effet, suivant leur taille, leur forme et leur densité les ossements ne sont pas transportés de la même manière et ils sont donc classés en trois groupes de dispersion :
- Groupe I : carpes, tarses, côtes, vertèbres et phalanges. Ce sont les os de plus faible densité qui sont immédiatement transportés par saltation ou flottation.
- Groupe II : scapula, humérus, radius, ulna, pelvis, fémur, tibia, métapodes. Ce sont principalement les os longs qui sont transportés progressivement par traction.
- Groupe III : crâne, mandibules et dents. Ce sont les os les plus denses qui sont transportés les derniers.
Nous avons appliqué cette méthode aux sites considérés suivant les trois classes de taille définies précédemment (B1 : grands bovidés, B2C1 : petits bovidés et grands cervidés et C2 : petits cervidés) (tableau 77 et figure 68).
Groupe I Groupe II Groupe III NR % NR % NR %
B1 23 36,5 30 47,6 10 15,9 B2 C1 5 31,3 8 50 3 18,8 Tanjung 63-64
C2 1 3,2 6 19,4 24 77,4 B1 1 8,3 7 58,3 4 33,3
B2 C1 5 50 2 20 3 30 Tanjung 82 C2 3 33,3 2 22,2 4 44,4 B1 6 18,8 7 21,9 19 59,4
B2C1 0 0 0 0 3 100 Sendang Busik C2 1 5,9 1 5,9 15 88,2 B1 0 0 3 25 9 75
B2C1 0 0 0 0 12 100 Ngrejeng C2 4 40 1 10 5 50 B1 3 50 0 0 3 50
B2C1 1 20 1 20 2 60 Grogol C2 5 33,3 1 6,7 9 60 B1 0 0 1 50 1 50
B2C1 0 0 0 0 3 100 Bukuran C2 1 20 1 20 3 60 B1 0 0 1 50 1 50
B2C1 0 0 0 0 0 0 Dayu C2 0 0 0 0 2 100
Tableau 77: Groupes de Voorhies pour les restes des classes de taille B1, B2C1 et C2 des sites de Tanjung,
Sendang Busik, Ngrejeng et Grogol. (en pourcentages de NR ; B1 : grands bovidés ; B2C1 : petit bovidés et grands cervidés ; C2 ; petits cervidés)
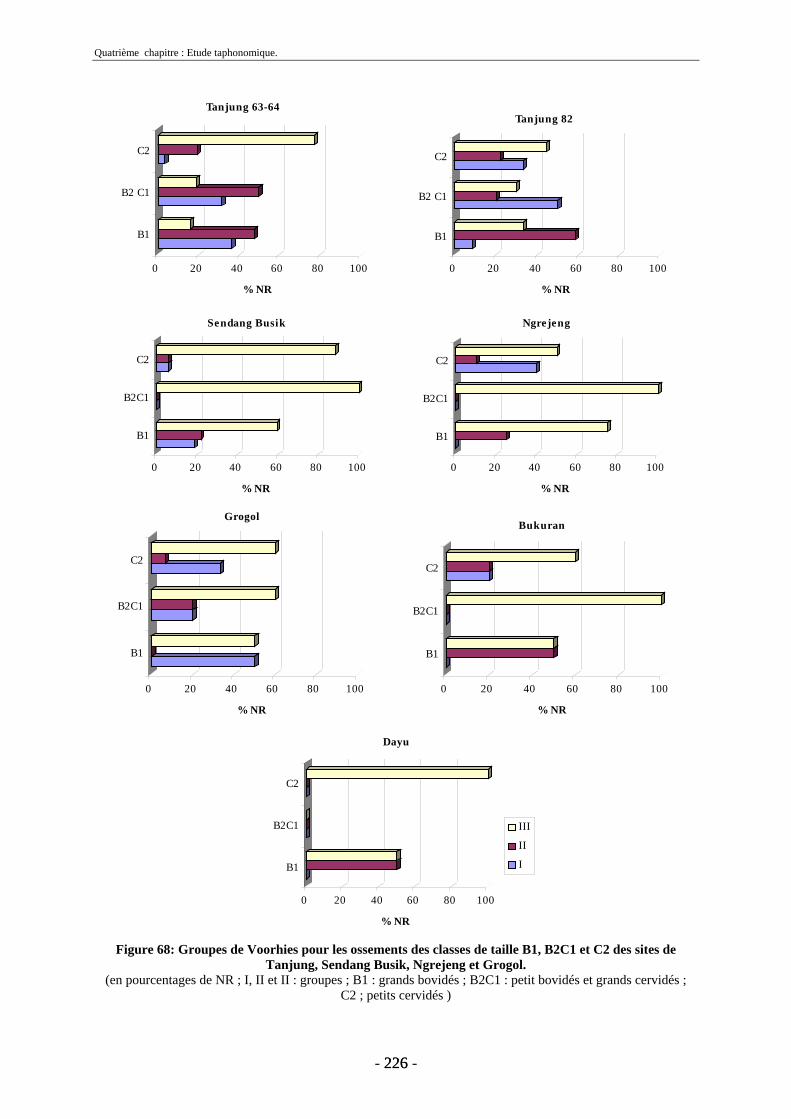
Quatrième chapitre : Etude taphonomique.
- 226 - 226
0 20 40 60 80 100
% NR
B1
B2 C1
C2
Tanjung 63-64
0 20 40 60 80 100
% NR
B1
B2 C1
C2
Tanjung 82
0 20 40 60 80 100
% NR
B1
B2C1
C2
Sendang Busik
0 20 40 60 80 100
% NR
B1
B2C1
C2
Ngrejeng
0 20 40 60 80 100
% NR
B1
B2C1
C2
Grogol
0 20 40 60 80 100
% NR
B1
B2C1
C2
Bukuran
0 20 40 60 80 100
% NR
B1
B2C1
C2
Dayu
III
II
I
Figure 68: Groupes de Voorhies pour les ossements des classes de taille B1, B2C1 et C2 des sites de
Tanjung, Sendang Busik, Ngrejeng et Grogol. (en pourcentages de NR ; I, II et II : groupes ; B1 : grands bovidés ; B2C1 : petit bovidés et grands cervidés ;
C2 ; petits cervidés )

Quatrième chapitre : Etude taphonomique.
- 227 - 227
À Tanjung, pour les deux niveaux, tous les groupes d’ossements sont représentés pour toutes les classes de taille dans les deux assemblages (tableau 76 et figure 68). Néanmoins, la classe des petits cervidés à Tanjung 63-64 présente une grande prédominance des restes du groupe III et très peu du groupe I.
Ces représentations plaident en faveur d’une absence de transport fluviatile pour les deux niveaux de Tanjung excepté peut-être pour la classe des petits cervidés à Tanjung 63-64. La taille inférieure des ossements de la classe des petits cervidés a sûrement permis un transport plus facile des éléments. Au niveau statistique, les résultats du test de Kolmogorov-Smirnov permettent de conclure que la distribution des petits cervidés de Tanjung 63-64 pour les groupes de Voorhies est significativement différente de celle des grands bovidés (B1) ou des petits bovidés et grands cervidés de ce même site+.
À Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu, les groupes de Voorhies sont
moins bien représentés pour toutes les classes de taille par rapport à Tanjung 63-64 et 82. En effet, sur ces sites les trois classes de taille présentent le plus souvent une prédominance des éléments les plus denses c’est à dire des éléments du groupe III. Les éléments moins denses (groupe I et II) semblent avoir été transportés en partie hors des sites. Néanmoins, à Sendang Busik les classes B1 et C2 et à Grogol, les classes B2C1 et C2 présentent les trois groupes.
La classe C2 à Ngrejeng et Grogol et la classe B1 à Grogol, paraissent avoir subi un transport moins important de ces éléments les moins denses. Les restes sont donc probablement plus autochtones par rapport à leur milieu de dépôt. À Ngrejeng, Grogol, Bukuran et Dayu le nombre de restes considéré dans les différentes classes de taille n’est pas très important et donc les résultats observés sont moins significatifs que sur les sites précédemment étudiés. D’un point de vue statistique+, la majorité des distributions des classes de taille pour les groupes de Voorhies des différents sites de notre étude ne sont pas significativement différentes hormis à Tanjung 63-64 où la classe des grands bovidés est significativement différente de celles de Sendang Busik et Ngrejeng+.
Par conséquent, l’influence du transport fluviatile sur les assemblages osseux paraît
plus importante sur les sites de Sendang Busik, Ngrejeng, Bukuran et Dayu pour toutes les classes de taille et à Tanjung pour la classe C2 (figure 68) mais ceci n’est pas vérifié au niveau des statistiques. À Grogol, cette influence fluviatile semble moins prononcée qu’à Sendang Busik et Ngrejeng. Cette hypothèse reste délicate car le nombre de restes considéré sur ce site est très faible pour toutes les classes de taille. À Tanjung, dans les deux niveaux, l’influence du transport fluviatile semble moins importante qu’à Sendang Busik et Ngrejeng pour les classes de taille B1 et B2C1.
+ tableaux 134 à 137, annexe 10.

Quatrième chapitre : Etude taphonomique.
- 228 - 228
Ces résultats sont à interpréter avec prudence car le nombre de restes considérés pour utiliser cette méthode est bien inférieur au nombre total d’ossements sur les différents sites. En ajoutant les esquilles, les effectifs des groupes I et II seraient augmentés. Cette méthode n’est pas très bien adaptée aux sites de Sangiran car il faudrait pouvoir trier correctement toutes les esquilles des éléments classés dans les groupes I et II et réaliser des expériences de transport.
Pour confirmer ces premiers résultats, nous avons ensuite analysé les rapports dents/vertèbres puis utilisé la méthode de Badgley.
4.2.2. Rapport dents/vertèbres
Afin de confirmer ces premiers résultats nous avons utilisé un autre rapport, le rapport dents/vertèbres (Behrensmeyer, 1975) (tableau 78). Ce rapport a été proposé comme un indice du transport fluviatile. En effet, dans un squelette de mammifère vivant le rapport du nombre de dents par rapport au nombre de vertèbres est d’environ 1. Les dents sont les éléments les plus denses alors que les vertèbres sont les moins denses et figurent parmi les premiers éléments transportés par l’eau.
Dans les assemblages considérés nous avons calculé ce rapport pour les trois classes de taille (B1, B2C1 et C2) des sept sites. Ce calcul n’a été possible que pour les classes des grands bovidés (B1) de Tanjung 63-64 et de Sendang Busik et celle des petits bovidés et des grands cervidés (B2C1) et des petits cervidés (C2) de Tanjung 82.
Dents/Vertèbres B1 B2C1 C2 Tanjung 63-64 0,5 / /
Tanjung 82 / 1 3 Sendang Busik 18 / /
Tableau 78: Rapport dents/vertèbres des classes de taille B1, B2C1 et C2 des sites de Tanjung et Sendang
Busik. À Tanjung 63-64, ce rapport est peu élevé pour la classe B1. À Tanjung 82, pour la
classe B2C1, ce rapport est très proche de celui d’un squelette de mammifère vivant. Ces deux valeurs sont très proches des valeurs obtenues dans le bassin actuel d’Amboseli au Kenya, dans les dépôts d’environnement deltaïque de la formation de Koobi Fora au Kenya (Behrensmeyer & Dechant-Boaz, 1980), dans les dépôts du Miocène supérieur d’une plaine d’inondation ou d’un bord de chenal des Siwaliks (Badgley, 1986a). Ces deux rapports sont donc caractéristiques d’une absence de transport fluviatile.
À Sendang Busik, pour la classe B1 et à Tanjung 82, pour la classe C2, ce rapport est supérieur à 1 et témoigne d’une grande perte des vertèbres dans cet assemblage osseux. Pour
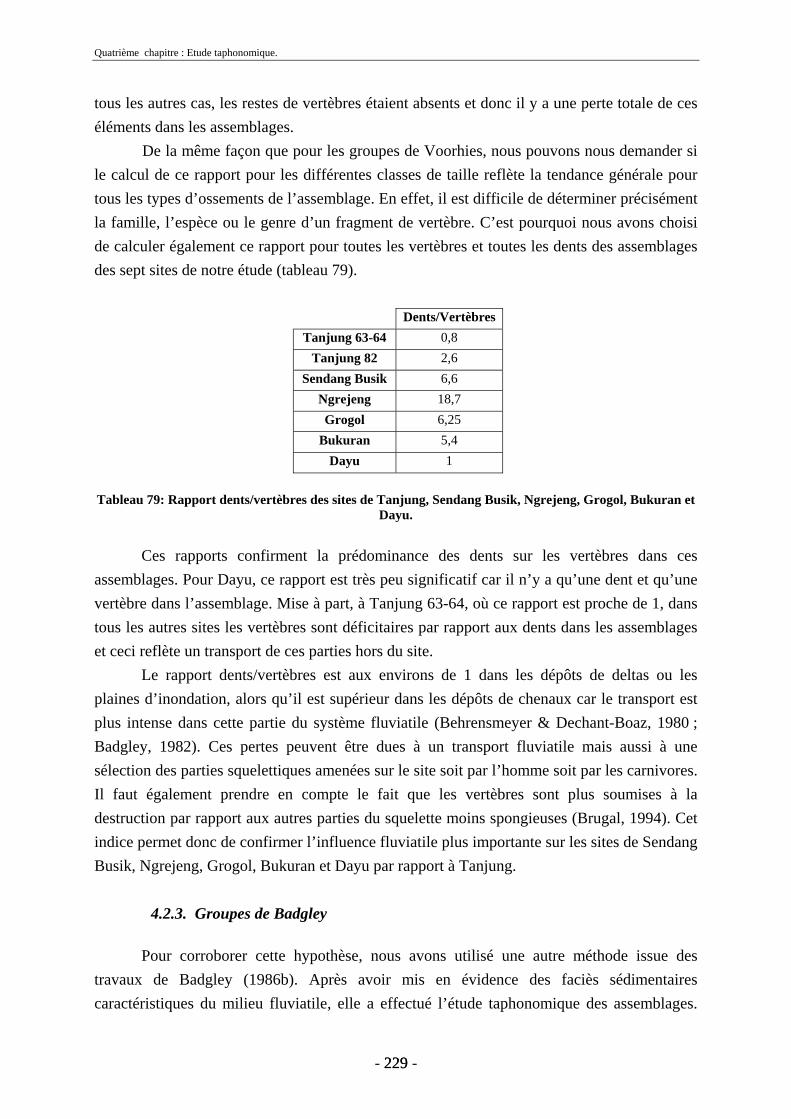
Quatrième chapitre : Etude taphonomique.
- 229 - 229
tous les autres cas, les restes de vertèbres étaient absents et donc il y a une perte totale de ces éléments dans les assemblages.
De la même façon que pour les groupes de Voorhies, nous pouvons nous demander si le calcul de ce rapport pour les différentes classes de taille reflète la tendance générale pour tous les types d’ossements de l’assemblage. En effet, il est difficile de déterminer précisément la famille, l’espèce ou le genre d’un fragment de vertèbre. C’est pourquoi nous avons choisi de calculer également ce rapport pour toutes les vertèbres et toutes les dents des assemblages des sept sites de notre étude (tableau 79).
Dents/Vertèbres Tanjung 63-64 0,8
Tanjung 82 2,6 Sendang Busik 6,6
Ngrejeng 18,7 Grogol 6,25
Bukuran 5,4 Dayu 1
Tableau 79: Rapport dents/vertèbres des sites de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et
Dayu. Ces rapports confirment la prédominance des dents sur les vertèbres dans ces
assemblages. Pour Dayu, ce rapport est très peu significatif car il n’y a qu’une dent et qu’une vertèbre dans l’assemblage. Mise à part, à Tanjung 63-64, où ce rapport est proche de 1, dans tous les autres sites les vertèbres sont déficitaires par rapport aux dents dans les assemblages et ceci reflète un transport de ces parties hors du site.
Le rapport dents/vertèbres est aux environs de 1 dans les dépôts de deltas ou les plaines d’inondation, alors qu’il est supérieur dans les dépôts de chenaux car le transport est plus intense dans cette partie du système fluviatile (Behrensmeyer & Dechant-Boaz, 1980 ; Badgley, 1982). Ces pertes peuvent être dues à un transport fluviatile mais aussi à une sélection des parties squelettiques amenées sur le site soit par l’homme soit par les carnivores. Il faut également prendre en compte le fait que les vertèbres sont plus soumises à la destruction par rapport aux autres parties du squelette moins spongieuses (Brugal, 1994). Cet indice permet donc de confirmer l’influence fluviatile plus importante sur les sites de Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu par rapport à Tanjung.
4.2.3. Groupes de Badgley
Pour corroborer cette hypothèse, nous avons utilisé une autre méthode issue des travaux de Badgley (1986b). Après avoir mis en évidence des faciès sédimentaires caractéristiques du milieu fluviatile, elle a effectué l’étude taphonomique des assemblages.
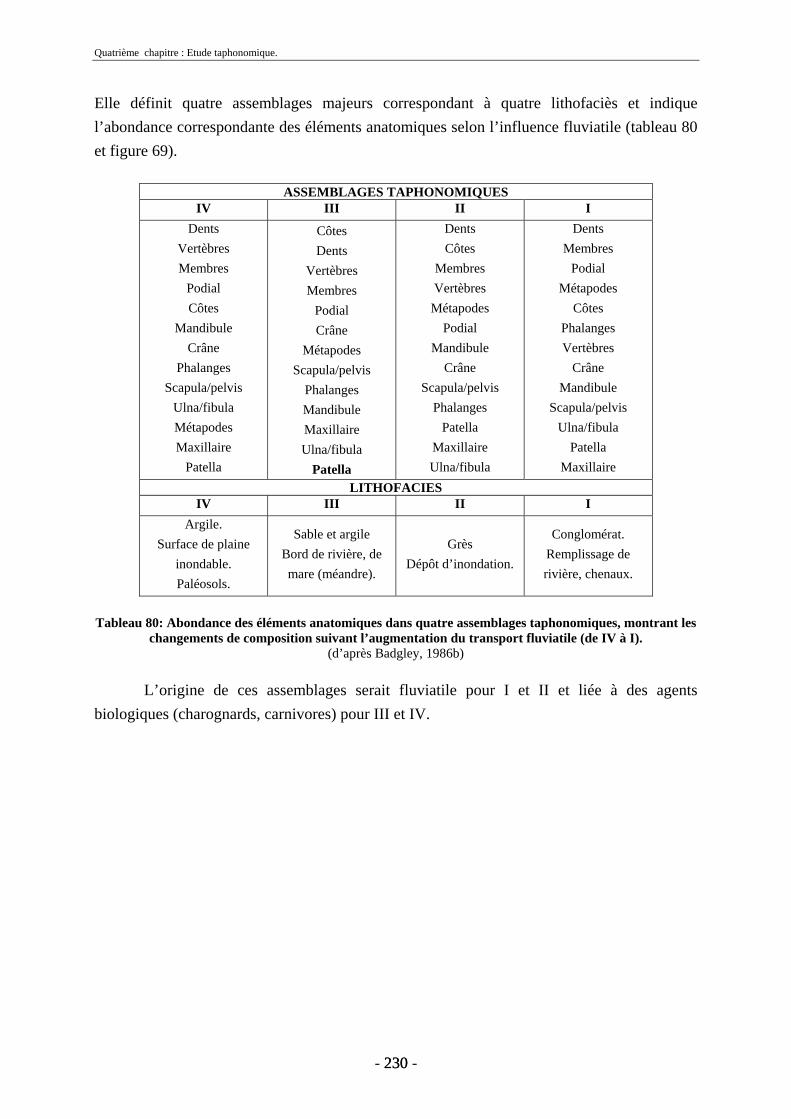
Quatrième chapitre : Etude taphonomique.
- 230 - 230
Elle définit quatre assemblages majeurs correspondant à quatre lithofaciès et indique l’abondance correspondante des éléments anatomiques selon l’influence fluviatile (tableau 80 et figure 69).
ASSEMBLAGES TAPHONOMIQUES IV III II I
Dents Vertèbres Membres
Podial Côtes
Mandibule Crâne
Phalanges Scapula/pelvis
Ulna/fibula Métapodes Maxillaire
Patella
Côtes Dents
Vertèbres Membres
Podial Crâne
Métapodes Scapula/pelvis
Phalanges Mandibule Maxillaire Ulna/fibula
Patella
Dents Côtes
Membres Vertèbres
Métapodes Podial
Mandibule Crâne
Scapula/pelvis Phalanges
Patella Maxillaire Ulna/fibula
Dents Membres
Podial Métapodes
Côtes Phalanges Vertèbres
Crâne Mandibule
Scapula/pelvis Ulna/fibula
Patella Maxillaire
LITHOFACIES IV III II I
Argile. Surface de plaine
inondable. Paléosols.
Sable et argile Bord de rivière, de mare (méandre).
Grès Dépôt d’inondation.
Conglomérat. Remplissage de rivière, chenaux.
Tableau 80: Abondance des éléments anatomiques dans quatre assemblages taphonomiques, montrant les
changements de composition suivant l’augmentation du transport fluviatile (de IV à I). (d’après Badgley, 1986b)
L’origine de ces assemblages serait fluviatile pour I et II et liée à des agents biologiques (charognards, carnivores) pour III et IV.
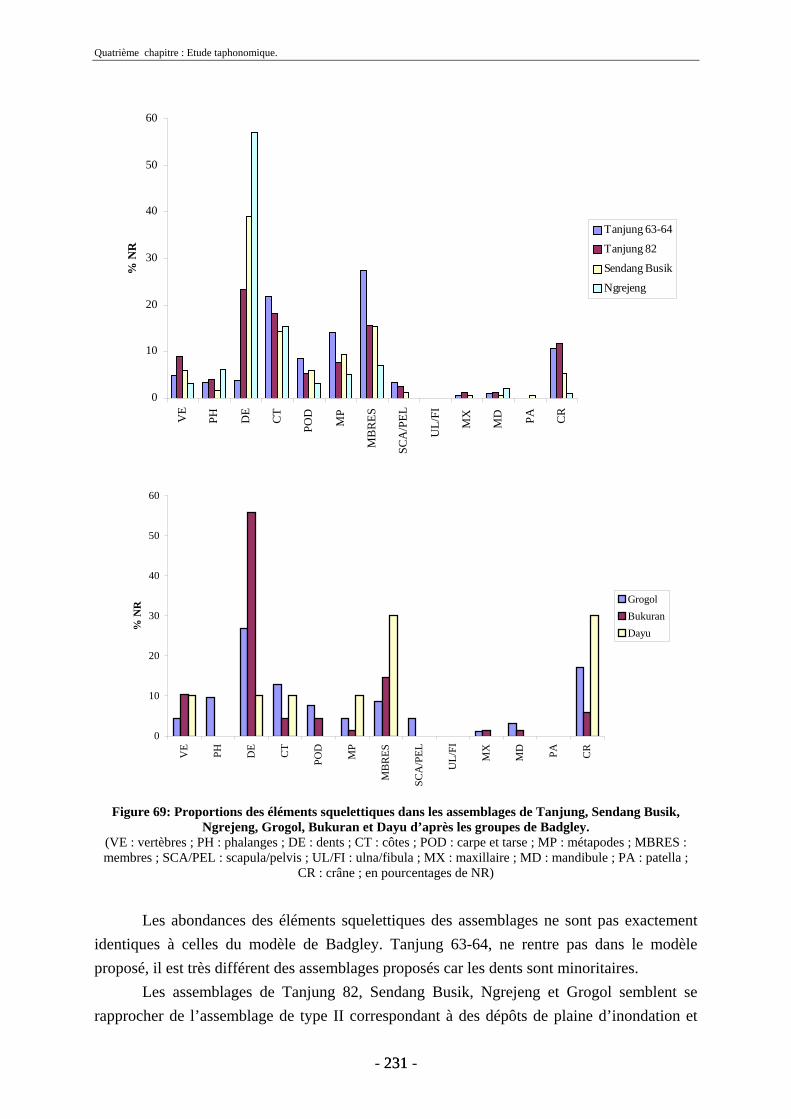
Quatrième chapitre : Etude taphonomique.
- 231 - 231
0
10
20
30
40
50
60
VE
PH DE
CT
POD
MP
MB
RES
SCA
/PEL
UL/
FI
MX
MD PA CR
% N
R
Tanjung 63-64
Tanjung 82
Sendang Busik
Ngrejeng
0
10
20
30
40
50
60
VE
PH DE
CT
POD
MP
MB
RES
SCA
/PEL
UL/
FI
MX
MD PA CR
% N
R GrogolBukuranDayu
Figure 69: Proportions des éléments squelettiques dans les assemblages de Tanjung, Sendang Busik,
Ngrejeng, Grogol, Bukuran et Dayu d’après les groupes de Badgley. (VE : vertèbres ; PH : phalanges ; DE : dents ; CT : côtes ; POD : carpe et tarse ; MP : métapodes ; MBRES : membres ; SCA/PEL : scapula/pelvis ; UL/FI : ulna/fibula ; MX : maxillaire ; MD : mandibule ; PA : patella ;
CR : crâne ; en pourcentages de NR)
Les abondances des éléments squelettiques des assemblages ne sont pas exactement identiques à celles du modèle de Badgley. Tanjung 63-64, ne rentre pas dans le modèle proposé, il est très différent des assemblages proposés car les dents sont minoritaires. Les assemblages de Tanjung 82, Sendang Busik, Ngrejeng et Grogol semblent se rapprocher de l’assemblage de type II correspondant à des dépôts de plaine d’inondation et

Quatrième chapitre : Etude taphonomique.
- 232 - 232
donc à un transport moins intense. Pour Bukuran, l’assemblage paraît plus proche de l’assemblage I correspondant à un dépôt de chenal et donc à un transport plus intense que pour l’assemblage II. Pour Dayu, nous n’avons pas considéré les résultats qui sont peu significatifs car le nombre de dents est très faible. Nous notons que cette méthode établie à partir de sites indiens (Siwaliks, Pakistan), nous permet de quantifier l’influence fluviatile sur les assemblages de notre étude mais que nous ne retrouvons pas exactement les mêmes types d’assemblages et que donc nous devons rester prudents sur ces données.
4.2.4. Connexions anatomiques Les connexions anatomiques peuvent donner des informations sur la dispersion des
ossements. Si de nombreux restes sont en connexion anatomique ils auront subi peu de transport et seront plus proches du lieu de mort de l’animal (Behrensmeyer, 1988 ; Brugal, 1994). Sur l’ensemble des sites, aucune connexion anatomique n’a été observée dans les assemblages ceci renforce donc l’idée d’un transport d’une partie des assemblages.
Néanmoins, nous avons constaté des associations de restes à Tanjung, Sendang Busik et Ngrejeng qui peuvent être liées à des fractures avant enfouissement si les deux restes proviennent de deux carrés différents ou à des fractures post-dépositionnelles si les deux restes proviennent du même carré:
- TDJ82/I/2 et TDJ82/I/3, ce sont deux fragments de diaphyse de tibia d’un grand bovidé (B1).
- TDJ64/G11/13B et TDJ64/F11/10B, ce sont deux fragments d’un bois d’Axis lydekkeri.
- SBII/11/48 et SBII/8/27, ce sont deux fragments d’un os plat, probablement un coxal, de grand herbivore (GH),
- NP85/V/49 et NP85/V/50, ce sont deux fragments de côtes de grand herbivore (GH).
À Ngebung 2, les restes de grands bovidés (B1) des trois niveaux ont peu subi
l’influence fluviatile car pour la méthode de Voorhies, les trois groupes d’éléments sont représentés. Cependant, le groupe III est le plus abondant (tableau 81).
Niveaux Groupe I Groupe II Groupe III Rapport dents/vertèbres
A 25,38 % 33,50 % 41,12 % 2,3 B 17,17 % 24,24 % 58,59 % 7,8
Upper 7,06 % 17,65 % 75,29 % 24
Tableau 81 : Groupes de Voorhies pour la classe des grands bovidés du site de Ngebung 2. (en pourcentages de NR)

Quatrième chapitre : Etude taphonomique.
- 233 - 233
Le rapport dents/vertèbres pour les grands bovidés (B1) sur ce site est très élevé pour les trois niveaux (A : 2,3 ; B : 7,8 ; Upper : 24). Le niveau Upper semble avoir subi un transport plus important que les niveaux A et B ceci confirme en partie les données obtenues sur les proportions d’os roulés.
À Notarchirico, l’étude des groupes de Badgley dans le niveau Alpha, l’absence d’éléments en connexion, la rareté des restes d’espèces de petite taille et d’individus juvéniles ont permis de mettre en évidence que l’accumulation a subi un courant d’intensité moyenne à forte (Tagliacozzo et al., 1999 ; annexe 8).
À Holon, pour le Bos et le Dama, les éléments du groupes I de Voorhies sont moins bien représentés que les éléments des groupes II et III. D’autre part, les rapports dents/vertèbres pour ces deux espèces sont élevés (Dama : 22,7 et Bos : 9,16). En conséquence, les restes de ces deux espèces semblent avoir subi un transport fluviatile mais celui-ci n’est pas le principal facteur à l’origine de leur représentation squelettique (Monchot & Horwitz, sous presse ; annexe 8).
L’action du transport fluviatile semble avoir été moins importante à Tanjung que sur
les autres sites ce qui pourrait confirmer une position plus éloignée de ce site par rapport au lit d’une paléorivière.
Après avoir passé en revue les traces laissées par les agents climato-édaphiques et les effets du transport fluviatile, nous avons considéré les traces laissées par les agents biologiques non anthropiques.
4.3. Agents biologiques non anthropiques
Les agents biologiques non anthropiques peuvent agir sur les ossements avant ou après leur enfouissement. Ils peuvent être de différentes natures : les microbes, les plantes, les insectes, les reptiles, les oiseaux, les rongeurs, les artiodactyles et les carnivores (Miller, 1994).
Les racines des plantes peuvent provoquer des réactions chimiques à la surface des os,
ce qui produit des figures de corrosion à leur surface et donc des vermiculations caractéristiques (Andrews, 1990). Sur les sites de Sangiran, nous n’avons pas mis en évidence de traces liées à l’action des plantes dans notre matériel.
Les reptiles et plus particulièrement les crocodiles sont des prédateurs très efficaces qui peuvent attaquer ou « charogner » des petits et des grands vertébrés (Shoop & Ruckdeschel, 1990) et qui peuvent digérer les os. Les os durant la digestion sont déminéralisés et le résidu organique restant est rapidement détruit après la défécation (Fisher, 1981). En conséquence, seuls les ossements qui ne sont pas totalement digérés, qui sont trop encombrants, qui sont trop grands ou qui sont négligés par le crocodile pourront subsister et

Quatrième chapitre : Etude taphonomique.
- 234 - 234
parvenir jusqu’à nous (Behrensmeyer, 1973 ; Fougeirol, communication personnelle). Bien que des dents de crocodiles aient été retrouvées sur les sites de Tanjung, Sendang Busik, Ngrejeng et Bukuran, aucune trace explicite de leur action n’a pu être mise en évidence sur les ossements. Nous pouvons penser qu’ils ont eu une certaine action sur les assemblages même si aucune trace lisible n’a été observée à la surface des ossements.
Les porcs-épics sont connus pour leur action sur les os dans les sites archéologiques (Shipman, 1981b). Ils rongent les os secs et les racines des dents dans le but d’user leurs incisives qui sont en croissance continue et de pallier leurs déficiences en phosphate et calcium de leur alimentation car ces deux éléments sont importants pour la croissance de leurs épines (Tougard, 1998 ; de Vos, 1983 ; Zeitoun et al., 2005). Aucune trace de l’action des porcs-épics n’a pu être mise en évidence au sein de nos différents assemblages osseux. En règle générale, les carnivores modifient les assemblages osseux d’origine en apportant leur proie ou en agissant directement sur les ossements. Selon les taxons, les carnivores ont des actions spécifiques sur les ossements (Miller, 1994). Les canidés sucent les os ce qui donne des formes typiques en « fourchette » et laisse des traces de crocs (Haynes, 1980, 1983). Les félidés déchiquètent la viande ce qui provoque des stries et laisse des traces de crocs (Blumenschine, 1986 ; Brain, 1981). Les hyènes sont des briseuses et mangeuses d’os, elles les avalent et parfois les régurgitent, ceci produit une grande fracturation des os (Sutcliff, 1970 ; Hill, 1983, 1989a ; Blumenschine, 1986, 1988 ; Horwitz, 1990 ; Cruz-Uribe, 1991 ; Marean & Spencer, 1991) et à leur surface des traces de dissolution caractéristiques de l’action des sucs gastriques (Andrews, 1990 ; Guadelli, 1989).
À Tanjung, Sendang Busik, Grogol et Dayu nous n’avons pas observé la présence de restes ou de traces laissées par des carnivores. À Ngrejeng, deux pièces semblent présenter des traces de « mâchonnage » NKJ85/III/14 (fragment de côte d’un herbivore de taille moyenne) et NKJ85/II/37 (esquille) (annexe 11). Et à Bukuran, un ossement BK79/VI/B1/30 (esquille) présente également des traces de carnivore (annexe 11). Cette dernière observation corrobore les résultats de la liste faunique de ce site car un reste de Pachycrocuta brevirostris a été mis en évidence.
L’action des carnivores sur nos assemblages semble marginale lorsqu’elle est observable. En outre, la présence de cylindres, ou de diaphyses sans épiphyses, est peu importante dans les accumulations alors qu’elle est généralement élevée dans des repaires d’hyènes par exemple (Binford, 1981 ; Brain, 1981). Les éléments déterminables des membres sont assez rares dans nos assemblages alors qu’ils sont bien représentés et particulièrement par rapport aux restes du squelette axial dans les repaires de carnivores (Hill, 1989a ; Blumenschine & Marean, 1993 ; Fosse et al., 1998 ; Guadelli, 1989). Enfin, dans les repaires il est fréquent de retrouver des restes de carnivores notamment des individus juvéniles (Fosse, 1994), or dans notre cas seul un reste de carnivore a été déterminé sur

Quatrième chapitre : Etude taphonomique.
- 235 - 235
l’ensemble des sites de notre étude. Nous pouvons penser que l’action des carnivores a été très marginale sur l’ensemble des assemblages de notre étude.
À Ngebung 2, l'action des carnivores (3 restes soit 0,1 % du NRT) et plus précisément
celle des canidés a pu être observée, ce sont principalement des stries et des traces de crocs (Moigne, 2004, communication personnelle) (tableau 82).
Traces de carnivore Niveaux NR % NRT*
A 2 0,3 B 0 0
Upper 1 0,4
Tableau 82 :Action des carnivores dans les niveaux de Ngebung 2. (en NR et pourcentages du NRTsans dents de chaque niveau)
Les carnivores ont également eu une action mineure sur les assemblages des différents
niveaux du site de Ngebung 2. De la même façon, à Holon, les traces de carnivores sont peu nombreuses et sont observables sur seulement 2,1 % des restes (Monchot & Horwitz, sous presse ; annexe 8). À Notarchirico, dans le niveau Alpha, l’influence des carnivores paraît également mineure, seulement deux restes présentent des traces pouvant être attribuées à des carnivores (Tagliacozzo et al., 1999 ; annexe 8).
Les agents biologiques non anthropiques n’ont donc eu qu’une action mineure sur les
assemblages de notre étude. Pour finir, nous avons considéré l’action anthropique sur les assemblages afin de
mieux connaître le lien homme-animal au Pléistocène moyen à Sangiran.
4.4. Action anthropique
L’homme préhistorique intervient à trois niveaux différents dans l’histoire des assemblages osseux (Patou-Mathis, 1994):
- sur la formation de l’assemblage, - sur la composition de l’assemblage, - sur les ossements eux-mêmes (modification des formes et des surfaces).
De nombreuses recherches ont été menées sur l’action anthropique pouvant être à l’origine des assemblages osseux dans des sites archéologiques (Foley, 1983 ; Turner, 1984 ; Klein & Cruz-Uribe, 1984 ; Stiner, 1990) ou dans des sites sub-actuels ou actuels (Binford, 1978, 1983 ; Yellen, 1977).

Quatrième chapitre : Etude taphonomique.
- 236 - 236
4.4.1. Formation de l’assemblage
Dans un gisement archéologique, la présence, la nature et l’état des ossements témoignent de la fonction du site. Les sites peuvent donc avoir différentes fonctions : site d’habitat, site d’abattage, site de « charognage » ou site de boucherie.
Les sites d’habitat peuvent être de trois types (Patou-Mathis, 1994): - le camp de base : habitat de longue durée où les restes fauniques sont très
abondants, - le camp temporaire : temps d’occupation court (une saison) où les restes fauniques
sont abondants, - la halte de chasse : très courte durée (quelques jours) pour la préparation du gibier
(Otte & Patou-Mathis, 1992). Le site d’abattage est le lieu de chasse où l’industrie lithique est rare. Il y a une
absence d’os longs et une abondance des côtes et des vertèbres. Les esquilles sont rares également (Girard &David, 1982 ; Jaubert & Brugal, 1990 ; Buehler , 1997 ; Fisher & Roll, 1999 ; Todd & Rapson, 1999).
Le site de charognage est le lieu de prélèvement de viande sur des animaux morts naturellement ou tués par un prédateur autre que l’homme. Le matériel lithique et osseux y est peu abondant (Shipman, 1983 ; Villa, 1990 ; Villa et al., 2005).
Le site de boucherie est une zone de préparation des animaux. Les outils sont peu nombreux, les déchets osseux sont assez fréquents (certains présentant des marques de boucherie) et les os riches en viande sont rares (Binford, 1978 ; Girard & David, 1982 ; Jaubert & Brugal, 1990).
Suivant la fonction du site, le matériel osseux voit son abondance varier, sa nature changer et est plus ou moins associé à de l’industrie lithique.
D’après ces premières descriptions, les sites de notre étude ne semblent pas être des sites d’habitat car les outils lithiques sont très rares et les restes ne sont pas très nombreux.
4.4.2. Composition de l’assemblage
La détermination et la quantification d’un assemblage osseux peut informer sur les
techniques d’acquisition (chasse et charognage) et de traitement du gibier. Pour quantifier un assemblage il existe de nombreux indices que nous avons utilisés précédemment (Poplin, 1976 ; Klein & Cruz-Uribe, 1984 ; Brugal & Patou-Mathis, 1993 ; Brugal et al., 1994).
L’étude de l’âge et du sexe des individus peut également apporter des informations sur l’acquisition du gibier. Des chasses spécialisées peuvent apparaître sur une espèce, une classe d’âge, un sexe, une taille ou une saison. Des abattages massifs, des chasses opportunistes ou organisées et des pratiques de charognage peuvent être mises en évidence.
L’analyse des courbes de mortalité des populations présentes sur un site archéologique peut permettre de proposer des hypothèses sur l’origine des accumulations osseuses car leur

Quatrième chapitre : Etude taphonomique.
- 237 - 237
profil peut être corrélé avec la configuration des vestiges (Lyman, 1994). Dans le cas d’une mortalité catastrophique (noyade, éruption volcanique…) de nombreux animaux meurent en même temps dans un même endroit. La courbe de mortalité décroît régulièrement des plus jeunes aux plus âgés. Pour une mortalité naturelle ou attritionnelle, les animaux meurent de façon régulière et continue dans le temps, la répartition des ossements est donc plus diffuse et l’enfouissement est progressif (Shipman, 1983). Les individus très âgés et les jeunes sont majoritaires dans ce type d’assemblage. Exceptés ces deux modèles, les animaux peuvent être piégés à tout âge par les sédiments meubles le long des cours d’eau et être transportés par l’eau avant de s’échouer sur les berges (Fosse, 1994). Enfin, s’il existe une chasse sélective anthropique la mortalité est qualifiée de cynégétique. La courbe met alors en évidence des adultes jeunes et des adultes.
Cependant, une récente étude sur le site fluviatile d’Untermassfeld daté du Pléistocène inférieur (Kahlke & Gaudzinski, 2005), a mis en évidence que les causes de mortalité établies à partir des assemblages fauniques d’un site archéologique pouvaient être très complexes. Sur ce site fluviatile, des crues ont agit avec un impact sélectif sur une communauté animale et ont produit différents types de mortalités. Les différences de courbes de mortalité selon les espèces s’expliqueraient en partie par l’éthologie et les capacités physiologiques des animaux lors d’un événement comme une crue. En effet, les capacités pour nager ne sont pas les mêmes suivant les espèces. Les proboscidiens et les hippopotames sont de bons nageurs alors que les rhinocéros le sont beaucoup moins ce qui expliquent que de nombreux individus adultes aient été retrouvés dans le matériel d’ Untermassfeld. Pour les bons nageurs, les individus jeunes et âgés sont majoritairement représentés car ils sont plus vulnérables.
Dans les assemblages de notre étude, les âges des individus ont été difficiles à connaître. À Tanjung 63-64, pour une surface de 900 m² nous avons déterminé 8 individus, une mort catastrophique paraît peu probable de même qu’une mort naturelle vu l’âge relatif déduit pour certains individus (3 adultes). Il est risqué de proposer un modèle cynégétique compte tenu des données en notre possession mais il n’est pas à exclure car les hypothèses précédentes (mort naturelle ou catastrophique) ne sont pas satisfaisantes. Le même cas de figure s’observe à Tanjung 82 où la surface de fouille est beaucoup plus réduite qu’à Tanjung 63-64. Le nombre d’individus y est proportionnellement plus important mais il n’y a pas d’espèce dominante représentée ce qui exclut le modèle catastrophique. À Sendang Busik, les hippopotames sont peut-être morts de manière naturelle vu leur âge probable où se sont noyés lors d’une crue mais pour les autres individus l’interprétation est plus délicate. À Ngrejeng et Bukuran, il est ardu d’émettre des hypothèses compte tenu du contexte stratigraphique de ces deux sites.
Nos données parcellaires sur l’âge d’une partie des individus ne nous permettent pas de choisir un modèle de mortalité pour chaque site.
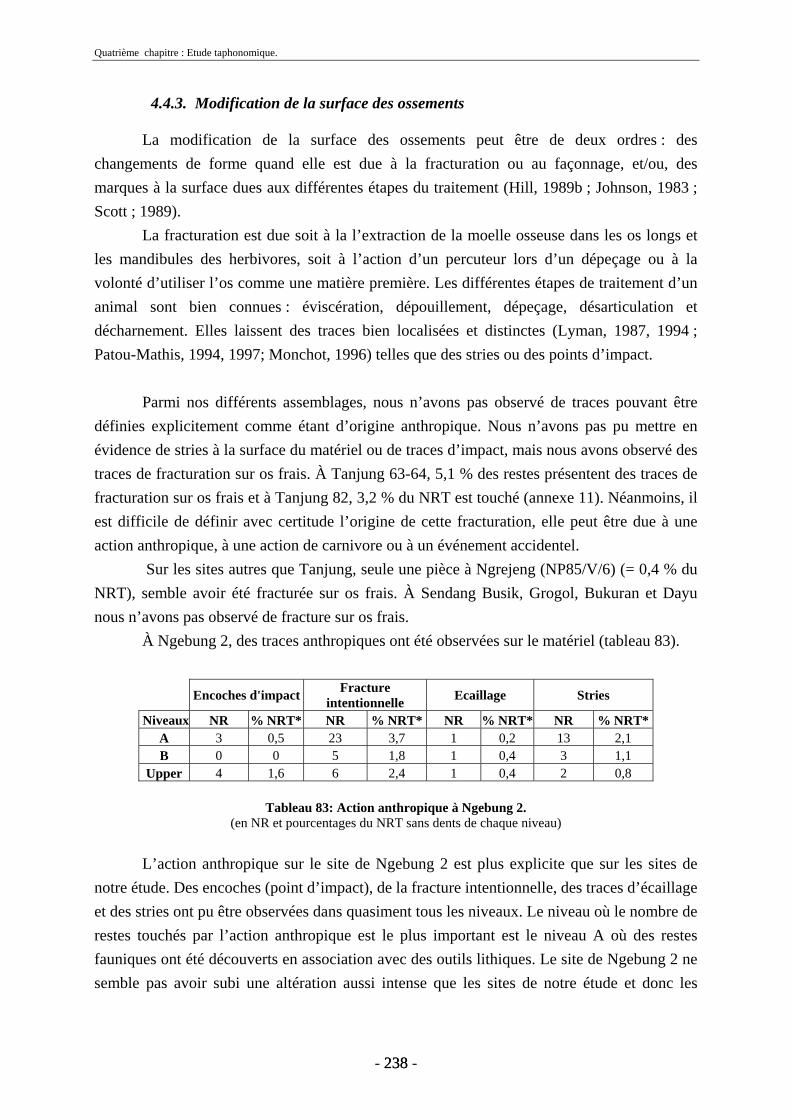
Quatrième chapitre : Etude taphonomique.
- 238 - 238
4.4.3. Modification de la surface des ossements
La modification de la surface des ossements peut être de deux ordres : des changements de forme quand elle est due à la fracturation ou au façonnage, et/ou, des marques à la surface dues aux différentes étapes du traitement (Hill, 1989b ; Johnson, 1983 ; Scott ; 1989).
La fracturation est due soit à la l’extraction de la moelle osseuse dans les os longs et les mandibules des herbivores, soit à l’action d’un percuteur lors d’un dépeçage ou à la volonté d’utiliser l’os comme une matière première. Les différentes étapes de traitement d’un animal sont bien connues : éviscération, dépouillement, dépeçage, désarticulation et décharnement. Elles laissent des traces bien localisées et distinctes (Lyman, 1987, 1994 ; Patou-Mathis, 1994, 1997; Monchot, 1996) telles que des stries ou des points d’impact. Parmi nos différents assemblages, nous n’avons pas observé de traces pouvant être définies explicitement comme étant d’origine anthropique. Nous n’avons pas pu mettre en évidence de stries à la surface du matériel ou de traces d’impact, mais nous avons observé des traces de fracturation sur os frais. À Tanjung 63-64, 5,1 % des restes présentent des traces de fracturation sur os frais et à Tanjung 82, 3,2 % du NRT est touché (annexe 11). Néanmoins, il est difficile de définir avec certitude l’origine de cette fracturation, elle peut être due à une action anthropique, à une action de carnivore ou à un événement accidentel.
Sur les sites autres que Tanjung, seule une pièce à Ngrejeng (NP85/V/6) (= 0,4 % du NRT), semble avoir été fracturée sur os frais. À Sendang Busik, Grogol, Bukuran et Dayu nous n’avons pas observé de fracture sur os frais.
À Ngebung 2, des traces anthropiques ont été observées sur le matériel (tableau 83).
Encoches d'impact Fracture intentionnelle Ecaillage Stries
Niveaux NR % NRT* NR % NRT* NR % NRT* NR % NRT* A 3 0,5 23 3,7 1 0,2 13 2,1 B 0 0 5 1,8 1 0,4 3 1,1
Upper 4 1,6 6 2,4 1 0,4 2 0,8
Tableau 83: Action anthropique à Ngebung 2. (en NR et pourcentages du NRT sans dents de chaque niveau)
L’action anthropique sur le site de Ngebung 2 est plus explicite que sur les sites de
notre étude. Des encoches (point d’impact), de la fracture intentionnelle, des traces d’écaillage et des stries ont pu être observées dans quasiment tous les niveaux. Le niveau où le nombre de restes touchés par l’action anthropique est le plus important est le niveau A où des restes fauniques ont été découverts en association avec des outils lithiques. Le site de Ngebung 2 ne semble pas avoir subi une altération aussi intense que les sites de notre étude et donc les

Quatrième chapitre : Etude taphonomique.
- 239 - 239
traces ont pu être observées plus aisément. L’homme et les carnivores semblent avoir eu une action plus importante à Ngebung 2 que sur les sites de notre étude.
À Holon, 3,3 % des restes présentent des modifications de surface de type anthropique. En effet, des stries, des points d’impact ainsi que des éclats d’os ont pu être observés (Horwitz & Monchot, 2002). À Notarchirico, quelques restes présentent des points d’impact et des éclats de percussion ont également été retrouvés qui seraient le résultat d’une fracturation anthropique intentionnelle sur os frais pour extraire la moelle (Cassoli et al., 1999). Mais ces restes sont en très faible proportion parmi l’assemblage du niveau Alpha. L’action anthropique sur ces deux sites est plus importante que sur les sites de notre étude, néanmoins elle reste mineure dans l’origine des accumulations osseuses.
5. DISCUSSION
Les sites de Sangiran ne présentent pas tous les mêmes répartitions spatiales et donc pas les mêmes conditions de conservation.
En ce qui concerne la conservation des différentes parties anatomiques, nous constatons que Tanjung montre une conservation différente où tous les types d’éléments squelettiques sont représentés alors que sur les autres sites, les restes crâniens et de l’autopode sont principalement représentés. Ceci est dû en partie à un nombre plus élevé de restes considérés pour Tanjung 63-64. Les restes crâniens de tous les sites sont en majorité des dents isolées. Sur tous les sites et pour toutes les classes de taille B1, B2C1 et C2, les restes les plus nombreux sont des restes d’éléments pauvres en viande. Nous pouvons donc constater que la conservation générale de ces sites est mauvaise.
En outre, la fracturation de ces ossements est importante car très peu d’os entiers sont présents dans les assemblages et aucun os long entier n’a été observé sur l’ensemble des sites.
Les esquilles sont les restes majoritaires sur tous les sites. Ces dernières ont, le plus souvent, une forme allongée avec une proportion très majoritaire de ces formes à Tanjung 63-64, Sendang Busik et Ngrejeng. La majorité de ces esquilles ont une taille comprise entre 20 et 50 millimètres à Ngrejeng, Grogol, Bukuran et Dayu. Les esquilles sont d’une taille plus grande sur les sites de Tanjung 63-64 et de Sendang Busik. Nous pouvons donc observer une grande fragmentation des ossements composant les assemblages ce qui réduit les chances de retrouver des restes entiers ou des traces d’agents taphonomiques.
Après avoir dressé ce constat, il était important de mettre en évidence les facteurs étant à l’origine de cette conservation.
Tous ces sites de plein air se trouvent principalement ou en partie dans une formation
volcano-sédimentaire en contexte fluviatile (formation de Kabuh). Les sols ont évolué au

Quatrième chapitre : Etude taphonomique.
- 240 - 240
cours du temps, et plus précisément sous des climats chauds, le drainage du sol et le pH ont provoqué une altération géochimique post-dépositionnelle qui a modifié la composition chimique de l’os.
L’eau a donc été un agent taphonomique important pour tous les sites que ce soit au niveau chimique ou mécanique. L’oxydation, et plus particulièrement l’oxydation du manganèse, est l’action qui touche le plus de restes sur tous les sites suivie de la dissolution et des concrétions. L’action chimique post-dépositionnelle de l’eau est donc prépondérante dans la formation de tous ces assemblages. Son action mécanique ou le transport fluviatile paraissent ne pas avoir eu la même intensité sur tous les sites. Ce transport paraît moins important à Tanjung que sur les autres sites exceptés à Bukuran ou Dayu où les données sont trop fragmentaires. Mais nous l’avons vu, les méthodes utilisées pour mettre en évidence cette action ne sont pas très bien adaptées à ce type d’assemblage.
L’action des agents biologiques est très minoritaire, nous n’avons observé des traces d’action de carnivores que sur un nombre très faible de restes sur les sites de Ngrejeng et Bukuran. Pour ce qui est de l’action des crocodiles, nous pouvons supposer qu’ils en ont eu une mais nous n’avons pas mis en évidence de traces explicites.
Enfin, l’action anthropique n’a pas pu être mise en évidence clairement. Nous n’avons pas observé de stries ou de traces d’impacts de percussions sur l’ensemble des sites. À Tanjung et Ngrejeng des traces de fracturation sur os frais ont pu être constatées, mais leur origine ne peut pas être précisée. Cependant à Tanjung, une bola (dans le niveau de 82) et des éclats (dans les niveaux 63-64) ont été retrouvés.
La grande fragmentation des assemblages semble liée en partie au weathering, en partie au transport fluviatile et à Tanjung peut-être en partie à l’action anthropique. En conclusion, l’origine de ces accumulations osseuses ne semble pas unique mais plutôt multiple. L’eau paraît néanmoins l’agent principal de ces accumulations. Différents agents ont plus ou moins agi sur les assemblages de ces différents sites. Ceci a été confirmé par la comparaison de la représentation squelettique des grands bovidés de Tanjung 63-64 et de Sendang Busik avec d’autres sites de plein air.
À la lumière de ces premiers résultats nous avons décidé de nous pencher plus particulièrement sur les sites de Tanjung et de Sendang Busik et sur leur présumée position stratigraphique dans la formation de Kabuh.
La formation de Kabuh est composée d’alternances de faciès (figure 70). Ces deux faciès (a et b) indiquent un régime fluviatile avec des dépôts de courant de chenaux d’énergie basique surmontés par des dépôts d’un courant de très basse énergie et des bancs sédimentaires. Les séries sédimentaires deviennent plus fines de la base au sommet des faciès. A la base des cycles C2a, C3a et C5a, la reprise de l’action fluviatile a lessivé les surfaces du « Lower Tuff », « Middle Tuff » et « Upper Tuff ». Dans le faciès « a », les fossiles se situent

Quatrième chapitre : Etude taphonomique.
- 241 - 241
dans les lentilles de sédiment grossier et plus spécialement dans les unités de la base (Larick et al., 2001).
Selon Larick et Zaim, Tanjung 63-64 se situerait à la base du cycle C3a et d’après eux à cette position, il y a une reprise de l’action fluviatile et un lessivage. D’après nos résultats sur le transport fluviatile, les ossements de Tanjung 63-64 n’auraient été que très peu transportés. Nous pouvons donc penser que la position de cet assemblage dans la colonne stratigraphique n’est pas exactement à la base du cycle C3a. Les datations de Larick et al. (2001) ne coïncident pas également avec les premières obtenues à Tanjung (Jacob, 1975a) à environ 800 000 ans et qui paraissent plus plausibles par rapport aux résultats paléomagnétiques (Sémah F., 1984).
En ce qui concerne les assemblages de Tanjung, nous ne pouvons pas les considérer comme un seul et même assemblage et nous pouvons penser que Tanjung 82 ne se trouve pas à la même position que Tanjung 63-64 dans la colonne stratigraphique. D’après le nombre de restes roulés et les rapports dents/vertèbres, l’assemblage de 82 paraît avoir subi un transport fluviatile plus important que le niveau de 63-64. Néanmoins, ce transport paraît moins puissant que sur les autres sites de Sangiran car les représentations squelettiques sont assez complètes. Nous n’avons pas observé de différences très significatives entre ces deux assemblages, les différences observées peuvent être dues à une distance plus importante de la rive ou à une position plus éloignée par rapport à la base d’un cycle sédimentaire pour l’assemblage de 1963-64 par rapport à l’assemblage de 1982 qui aurait plus subi les effets d’un transport fluviatile, mais d’une intensité moins importante que pour les autres sites.
Figure 70 : Téphrostratigraphie de la formation de Kabuh.
(Cycles sédimentaires : orange, jaune, faciès « a » ; vert, faciès « b ») (Larick et al., 2001)

Quatrième chapitre : Etude taphonomique.
- 242 - 242
La position stratigraphique de l’assemblage de Sendang Busik est moins contestable, car il est situé dans le cycle C2a mais non à sa base, donc l’action du transport fluviatile a pu jouer un rôle mais être d’une intensité moins importante qu’à la base.

- 243 -
SYNTHESE Suite aux résultats paléontologiques, biostratigraphiques, paléoenvironnementaux et taphonomiques obtenus sur les différents sites du dôme de Sangiran, nous pouvons proposer une synthèse de ces résultats site par site.
1. TANJUNG
La collection de Tanjung est constituée de deux niveaux différents (63-64 et 82) fouillés à environ 20 ans d’intervalle. Nous avons distingué ces deux niveaux et nous les avons considérés individuellement.
À Tanjung 63-64 sur une surface totale d’environ 900 m², 921 restes ont été mis au jour. Cette distribution, du point de vue statistique, n’est pas aléatoire. À Tanjung 82 sur une surface d’environ 20 m², 188 restes ont été découverts. Ces deux niveaux se situent dans la formation de Kabuh au-dessus du niveau du Grenzbank. En outre, une collection de surface a été récoltée à Tanjung 63-64 ( 85 restes).
Ces fossiles sont principalement, dans les deux niveaux, des restes d’artiodactyles. Huit individus ont pu être dénombrés pour cinq espèces à Tanjung 63-64. Parmi ces huit individus, cinq ont une masse supérieure à 500 kilogrammes.:
- Rhinoceros sp. : 1 individu, - Bubalus palaeokerabau : 2 individus, - Bibos palaesondaicus : 2 individus, - Axis lydekkeri : 1 individu, - Axis sp. : 2 individus.
Six individus ont été déterminés pour six espèces dans le niveau de 82 dont trois ont une masse supérieure à 500 kilogrammes :
- Elephas sp.: 1 individu, - Rhinoceros sp. : 1 individu, - Bubalus palaeokerabau : 1 individu, - Duboisia santeng : 1 individu, - Axis sp. : 1 individu, - Sus sp. : 1 individu. Les bovidés et les cervidés sont dominants dans ces deux assemblages et les
carnivores sont absents. L’âge des individus dans ces deux niveaux n’a pas pu être précisé. La

Synthèse.
- 244 -
richesse biologique est plus importante dans le niveau de 82. Toutefois, parmi tous les sites étudiés, la liste faunique de Tanjung 63-64 présente le plus de similarités avec celle de Tanjung 82, qui elle-même se rapproche le plus de celle de Ngrejeng.
La position stratigraphique du niveau de 82 semble plus haute dans la série que celle du niveau de Tanjung 63-64 (Koeshardjono, communication personnelle). Ceci se confirme au niveau biostratigraphique, l’association de Tanjung 82 paraît plus récente que celle de Tanjung 63-64. La présence de l’éléphant rapproche l’association faunique de Tanjung 82 de celle de Kedung Brubus plutôt que de Trinil H.K. Tanjung 63-64 serait un dépôt plus ancien que celui de 82. Il pourrait se situer à la base de l’unité de Kedung Brubus ou à la limite supérieure de Trinil H.K. et constituer une zone charnière entre deux horizons. Ceci est confirmé par les datations dans le niveau de Tanjung 64 qui ont donné un âge de 0,83 Ma (Jacob, 1975a). D’après Leinders et al. (1985) l’unité de Trinil H.K. est datée d’environ 1 à 0,9 Ma alors que celle de Kedung Brubus serait datée de 0,8 à 0,7 Ma. Tout ceci confirmerait donc la biozonation établie par Sondaar (1984).
Au niveau paléoenvironnemental, la présence de grands bovidés et de mammifères de grande taille (rhinocéros et éléphant) dans les deux niveaux ainsi que d’Axis suggèrent un milieu forestier ouvert. Celui-ci était proche d’un cours d’eau vue la présence d’espèces aquaphiles et de reptiles. Nous n’avons pas constaté de différences de paléomilieux entre les deux niveaux.
La conservation des restes est un peu différente entre Tanjung 63-64 et 82. Les restes majoritaires dans les deux assemblages sont les esquilles (Tanjung 63-64 : 63,5 % et Tanjung 82 : 52,1 %). Dans le niveau de 63-64, les os de l’autopode sont prédominants parmi les éléments squelettiques. À Tanjung 82, les dents sont plus nombreuses et les restes de l’autopode (carpe, tarse, métapodes et phalanges) sont aussi bien représentés. Les restes majoritaires dans ces deux assemblages sont des restes provenant d’éléments pauvres en viande, même si leurs proportions sont inférieures aux autres sites de Sangiran. La représentation squelettique de l’assemblage des grands bovidés de Tanjung 63-64 est assez proche de celles des sites archéologiques du Mont Dol et de Notarchirico et du site naturel de Verdigree Quarry.
Les esquilles de Tanjung 82 sont moins nombreuses que dans les niveaux de Tanjung 63-64 et elles ont une taille inférieure. Leur forme est le plus souvent allongée ou lamellaire dans les deux niveaux. Le matériel semble moins fragmenté dans les niveaux de 82, comme le confirment l’indice général d’intensité de la fragmentation (IGIF), l’indice de fragmentation des ossements (IFO) et le nombre des esquilles. Les dents sont par contre plus fragmentées dans les niveaux de 82 que de 63-64. Ces deux types de conservation diffèrent de celle des autres sites de Sangiran.
Les agents climato-édaphiques n’ont pas agi avec la même intensité sur les deux niveaux. Dans les niveaux de 82, l’action chimique de l’eau paraît moins intense que dans Tanjung 63-64. Les oxydes, la dissolution, la fissuration et le concrétionnement sont moins

Synthèse.
- 245 -
présents. Par contre, la proportion d’os roulés est plus importante dans le niveau de 82 et révèle un transport fluviatile d’une partie des ossements, peut-être dans une plaine d’inondation, ce qui est confirmé par les différentes méthodes de Voorhies et de Badgley. Ce transport fluviatile semble très faible dans le niveau de Tanjung 63-64 ce qui est confirmé par la présence de deux crânes entiers de grands bovidés. Ce dépôt devait se localiser à une certaine distance du lit d’une paléorivière. Aucune trace d’agents biologiques n’a été mise en évidence dans ces deux niveaux, mais des restes de crocodiles ont été observés. Des fractures sur os frais ont été observées dans les deux niveaux cependant l’origine de cette fracturation n’est pas facile à définir clairement. Une bola a été découverte dans le carré III de Tanjung 82 et des éclats dans le niveau 63-64.
L’origine des assemblages du site de Tanjung semble donc multiple. Dans le niveau de 63-64, le weathering, avant l’enfouissement, a joué un rôle non négligeable et l’action chimique de l’eau a été prépondérante après l’enfouissement. La présence du crâne (Sangiran 10), la découverte d’éclats, les traces de fracturation sur os frais et la répartition spatiale dans ce niveau suggèrent une interaction entre les mammifères et Homo erectus. S’il y a eu action anthropique sur cet assemblage, elle a été mineure. Ce site semble assez proche d’une zone d’occupation de très courte durée.
Dans ces deux niveaux des concentrations primaires d’ossements ont subi une
évolution post-dépositionnelle qui a masqué les traces d’actions anté-dépositionnelles. Nous pouvons proposer un scénario de formation et d’évolution des dépôts de
Tanjung: - Tanjung 63-64 : 1) Une accumulation de cadavres s’est produite (dépôt primaire) liée peut-être à
une prédation opportuniste de l’homme ou des crocodiles ou à un piège dans les sédiments meubles au bord d’une rivière. Cette accumulation ne peut pas être liée à une mortalité catastrophique car il n’y a pas d’espèce dominante et l’enfouissement a été progressif, ni à une accumulation naturelle car la majorité des individus sont des adultes.
2) L’Homme a pu faire un court passage sur les carcasses de même que les crocodiles,
3) L’enfouissement a été progressif, il y a eu l’action du weathering sur une partie du matériel (fragmentation),
4) L’évolution géochimique post-dépositionnelle a altéré les surfaces des ossements.
D’après la très faible intensité du transport fluviatile, la présence d’os roulés dans cet assemblage pose un problème. Il peut s’agir d’un apport initial sur le site ou bien d’un apport entre la phase 1 et 2 lié à une grande crue ou à une coulée boueuse (dépôt secondaire).

Synthèse.
- 246 -
- Tanjung 82 : 1) Une accumulation de cadavres s’est produite (dépôt primaire). L’origine de
celle-ci est comparable à celle de Tanjung 63-64, 2) L’Homme a pu faire un court passage sur les carcasses de même que les
crocodiles, 3) Une crue dans une plaine d’inondation a pu transporter une partie de
l’assemblage et a apporté des os roulés (dépôt secondaire), 4) L’enfouissement a été plus rapide que pour Tanjung 63-64, 5) L’évolution géochimique post-dépositionnelle a altéré les surfaces des
ossements.
2. SENDANG BUSIK À Sendang Busik, deux fouilles ont été menées à un an d’intervalle dans la formation
de Kabuh : Sendang Busik I (82 restes sur une surface de 20 m²) et Sendang Busik II (311 restes sur une surface de 44 m²). Nous avons choisi de considérer ces deux assemblages comme provenant d’une même zone fouillée. De même qu’à Tanjung 63-64, des collections de surface ont été constituées en parallèle des fouilles (78 restes).
Cinq individus ont pu être déterminés à Sendang Busik dont trois ont une masse supérieure à 500 kilogrammes :
- Stegodon sp.: 1 individu, - Hexaprotodon sivalensis : 2 individus, - Cervus (Rusa) sp., 1 individu, - Axis sp., 1 individu. Cette liste faunique est moins riche que celle du site de Tanjung pour les deux niveaux
néanmoins les cervidés sont bien représentés. Les carnivores sont absents. Un âge a pu être attribué aux deux Hexaprotodon : un individu juvénile et un très âgé. Pour les autres individus cela n’a pas été possible.
Au niveau biostratigraphique, cette association est proche de l’association de Trinil H.K. et donc se situe environ entre 1 et 0,8 Ma. Les reptiles ajoutés aux espèces décrites caractérisent un milieu de vie assez proche de celui de Tanjung : un milieu forestier ouvert en bordure d’un cours d’eau.
Les dents sont les éléments squelettiques majoritaires chez les bovidés et les cervidés quelle que soit la classe de taille et elles sont le plus souvent fragmentées (69,7 %). Les éléments

Synthèse.
- 247 -
squelettiques prédominants sont pauvres en viande. La distribution squelettique des grands bovidés se rapproche de celles des sites archéologiques de Soleilhac et de Ngebung 2 dans le niveau B et du site naturel de Tourville D. Les esquilles sont majoritaires dans l’assemblage mais en proportion inférieure à Tanjung 63-64 et la plupart sont de forme allongée. Les ossements entiers sont très rares ce qui est confirmé par l’indice de fragmentation des ossements (IFO).
Les agents climato-édaphiques ont touché cet assemblage dans des proportions comparables à celles de Tanjung 63-64. Cependant, le transport fluviatile paraît plus important qu’à Tanjung 82 (indices de Voorhies et proportions dents/vertèbres) bien que la proportion d’os roulés (d6) soit inférieure.
Sur ce site l’action biologique et l’action anthropique n’ont pas été mises en évidence et aucun outil lithique n’a été retrouvé. Cependant, des restes de crocodiles ont été déterminés ce qui implique qu’ils ont pu avoir une action sur l’assemblage. L’hypothèse d’une action anthropique sur cet assemblage semble peu probable.
L’action mécanique de l’eau semble à l’origine de cette accumulation. Le scénario de formation possible de cette accumulation pourrait être le suivant :
1) Une accumulation de cadavres s’est produite (dépôt primaire), les hippopotames sont peut-être morts naturellement ou se sont noyés mais pour les autres individus l’interprétation est plus délicate. Ils ont peut-être été piégés dans les sédiments meubles le long de la rivière ou ont subi la prédation des crocodiles.
2) Les crocodiles ont pu faire un court passage sur les carcasses, 3) Une crue a pu transporter une partie de l’assemblage et a apporté des os
roulés (dépôt secondaire), 4) L’enfouissement a été progressif, il y a eu l’action du weathering sur une
partie du matériel, 5) L’évolution géochimique post-dépositionnelle a altéré les surfaces des
ossements.
3. NGREJENG PLUPUH La fouille a été menée en 1985 dans les formations de Pucangan et de Kabuh. Lors de celle-ci, 245 restes osseux ont été mis au jour sur une surface de 20 m². En outre, une collection de surface a été constituée en parallèle (203 restes).
Cinq individus ont pu être déterminés dont la masse est inférieure à 500 kilogrammes : - Duboisia santeng : 2 individus, - Cervus (Rusa) sp. : 1 individu,

Quatrième chapitre : Etude taphonomique.
- 248 - 248
- Axis sp. : 1 individu, - Sus sp. : 1 individu. L’un des deux individus de Duboisia santeng a pu être déterminé comme un adulte
âgé. Les cervidés et les bovidés prédominent dans cet assemblage. Les carnivores sont absents. La richesse biologique de cette liste faunique est comparable à celle du site de Sendang Busik. Cette liste se rapproche le plus de celles établies pour Tanjung 82 et Bukuran. Le taux de détermination sur ce site est le plus important parmi tous les sites étudiés mais il faut garder à l’esprit que ces restes proviennent de deux formations sédimentaires. La position biostratigraphique de cette association est difficile à définir compte tenu du manque de précision stratigraphique sur les dépôts fossilifères. Cette association faunique peut être l’image d’un mélange de plusieurs unités biostratigraphiques entre Ci Saat, Trinil H.K et Kedung Brubus. Le paléoenvironnement de ces espèces est un milieu forestier ouvert proche d’un cours d’eau ou d’un lac compte tenu de la nature argileuse de la formation de Pucangan. Cette hypothèse est délicate à formuler compte tenu du cadre stratigraphique.
La conservation squelettique à Ngrejeng est comparable à celle de Sendang Busik. En effet, les restes dentaires sont majoritaires et les restes du squelette axial sont déficitaires. Les esquilles sont majoritaires dans l’assemblage, leur forme est en général allongée et leur taille est comprise entre 20 et 50 millimètres. Aucun ossement entier n’a été mis en évidence. Les dents sont également très fragmentées.
Parmi les agents climato-édaphiques, les oxydes touchent une grande partie des restes de Ngrejeng et la proportion d’os roulés est la plus importante de tous les sites considérés. Les traces de fissuration sont peu importantes. Le transport fluviatile semble avoir été un agent non négligeable dans la genèse de cet assemblage au vu des résultats des méthodes de Voorhies et Badgley et du pourcentage d’os roulés. Le milieu de dépôt de cet assemblage serait proche d’un dépôt de plaine d’inondation.
Des traces de carnivores ont été mises en évidence sur deux restes de ce site et des restes de crocodiles ont été découverts également. Une pièce présente une fracture sur os frais. L’action biologique a été mineure sur cette accumulation. L’action majeure à l’origine de cet assemblage semble avoir été l’action mécanique de l’eau. Aucune trace d’action anthropique n’a pu être observée sur cet assemblage.
Le scénario de formation des différents assemblages est délicat à proposer sachant qu’il y a eu plusieurs dépôts à différentes époques car les restes proviennent des formations de Pucangan et de Kabuh :
1) Des cadavres se sont déposés (dépôt primaire) à plusieurs époques. Un premier dépôt a dû avoir lieu dans la formation de Pucangan avec notamment les restes de poissons. Et ensuite divers dépôts se sont succédés. Ces morts peuvent être liées à un piégeage dans les sédiments meubles au bord d’une

Synthèse.
- 249 -
rivière notamment pour les restes provenant de la formation de Kabuh ou à une prédation par les crocodiles ou les carnivores.
2) Les carnivores et les crocodiles ont pu faire un court passage sur les carcasses, 3) Une crue a pu transporter une partie de l’assemblage et a apporté des os
roulés (dépôt secondaire), 4) L’enfouissement a été rapide, 5) L’évolution géochimique post-dépositionnelle a altéré les surfaces des
ossements.
4. GROGOL PLUPUH À Grogol Plupuh, des fouilles en 1983 sur une surface de 20 m² ont permis de mettre au jour 212 restes dans la formation de Kabuh. Environ la moitié de ces restes semblent provenir du niveau du Grenzbank (Kalsirudit). Une collection de surface a été établie en parallèle des fouilles (68 restes). Un seul individu (Axis sp.), dont l’âge n’a pas pu être précisé, a pu être déterminé mais des restes de bovidés et de cervidés indéterminés ont également été observés. Les carnivores sont absents. La pauvreté de cette liste ne nous permet pas de définir un cadre biostratigraphique pour cette association. Le paléomilieu est lui aussi difficile à définir avec une seule espèce qui est forestière en milieu fermé et très peu de restes de reptiles. Au niveau taphonomique, les dents et les restes de l’autopode sont majoritaires chez les cervidés et les bovidés. Les résultats sont à nuancer car le nombre de restes considéré est souvent faible et donc peu significatif. La majorité des restes de cet assemblage sont des esquilles dont la longueur varie entre 20 et 50 millimètres. La forme de ces esquilles est le plus souvent allongée mais le nombre d’esquilles lamellaires et plates n’est pas négligeable. Cet assemblage semble donc très fragmenté ce qui est confirmé par l’indice général d’intensité de la fragmentation (IGIF) et l’indice de fragmentation des ossements (IFO) ainsi que par la proportion des dents fragmentées. Ces restes montrent surtout des traces d’oxydes et de concrétionnements typiques du Grenzbank. La fissuration, la dissolution et le nombre d’os roulés sont peu importants. Cependant, d’après les méthodes de Voorhies et de Badgley, cet assemblage a subi un transport fluviatile mais moins intense qu’à Ngrejeng et Sendang Busik et peut-être dans une plaine d’inondation.
Aucune trace biologique ou anthropique n’a été mise en évidence. Aucun reste de crocodile n’a été observé. L’agent principal de formation de cette accumulation semble encore être l’eau.

Synthèse.
- 250 -
Le scénario de formation de cet assemblage peut être le suivant : 1) Quelques cadavres se sont déposés (dépôt primaire). La mort peut être
accidentelle, là aussi peut-être liée à un piégeage dans les sédiments meubles. 2) Une crue a pu transporter une partie de l’assemblage et a apporté des os
roulés (dépôt secondaire), 3) L’enfouissement a été rapide, 4) L’évolution géochimique post-dépositionnelle a été importante et a altéré les
surfaces des ossements avec de nombreuses concrétions.
5. BUKURAN Des fouilles ont été conduites à Bukuran en 1979 sur une surface de 72 m² (144 restes) dans les formations de Pucangan et de Kabuh. Néanmoins, la majorité des restes se situe dans la partie inférieure de la formation de Kabuh, dans le Grenzbank (109 restes). Une collection de surface importante a été récoltée pendant la période des fouilles (346 restes). Six individus ont pu être déterminés dans l’assemblage de Bukuran dont un a une masse supérieure à 500 kilogrammes:
- Hexaprotodon sivalensis : 1 individu, - Duboisia santeng : 1 individu, - Axis lydekkeri : 1 individu, - Axis sp. : 1 individu, - Sus brachygnathus : 1 individu, - Pachycrocuta brevirostris : 1 individu. Parmi ces six individus nous trouvons deux cervidés et un bovidé. Pachycrocuta
brevirostris est le seul carnivore déterminé sur tous les sites considérés. Le suidé serait un adulte âgé. Cette liste se rapproche le plus de celle de Ngrejeng. La richesse biologique de cet assemblage est comparable à celle de Tanjung 82 mais ici les restes proviennent de deux formations. Au niveau biostratigraphique, il est encore délicat de positionner cet assemblage car il provient de deux formations sédimentaires. Cette association faunique peut être l’image d’un mélange de plusieurs unités biostratigraphiques entre Ci Saat, Trinil H.K et Kedung Brubus. Les espèces Axis et Pachycrocuta proviennent du niveau de la base de la formation de Kabuh (Pasir Conglomeratik). Dans le cadre biostratigraphique proposé initialement (Sondaar, 1984), la hyène apparaît dans l’association de Kedung Brubus. La présence du fossile dans la partie inférieure de Kabuh, tend à indiquer que sur le dôme de Sangiran la hyène se retrouve dans l’unité biostratigraphique de Trinil H.K. Le paléoenvironnement des espèces de Bukuran est un milieu forestier ouvert proche d’une rivière ou d’un lac d’après la nature argileuse de la formation de Pucangan. Cette

Synthèse.
- 251 -
hypothèse est délicate à formuler compte tenu du cadre stratigraphique. Au niveau taphonomique, les dents sont les éléments majoritaires chez les cervidés et les bovidés. La représentation squelettique pour les différentes classes de taille n’est pas pertinente en raison du peu de restes. Les restes de Bukuran sont très fragmentés et les esquilles sont ici aussi majoritaires. La plupart des longueurs des esquilles sont comprises entre 20 et 50 millimètres. Aucun os entier n’a été retrouvé et les dents sont très majoritairement fragmentées.
La proportion des ossements oxydés est moins importante que sur les autres sites de notre étude. La fissuration et la dissolution sont peu importantes. Par contre, les os roulés sont assez nombreux et les concrétionnements sont très importants. Les restes colorés sont aussi bien représentés.
Le transport fluviatile semble le plus important sur ce site et selon la méthode de Badgley le milieu de dépôt de cet assemblage serait proche d’un dépôt de chenal.
Des traces de carnivores ont été observées sur un reste et des dents de crocodiles ont été retrouvées également. Aucune trace d’activité anthropique n’a été mise en évidence. L’agent principal de formation de cette accumulation semble donc être l’eau.
Le scénario de formation de ces assemblages peut être le suivant, sachant qu’il y a eu différents dépôts à différentes époques :
1) Des cadavres se sont déposés à plusieurs époques (dépôt primaire). Un premier dépôt a dû avoir lieu dans la formation de Pucangan avec notamment les restes de poissons. Et ensuite divers dépôts se sont succédés. Ces morts peuvent être liées à un piégeage dans les sédiments meubles au bord d’une rivière notamment pour les restes provenant de la formation de Kabuh ou à une prédation par les crocodiles ou les carnivores.
2) Les carnivores et les crocodiles ont pu faire un court passage sur les carcasses,
3) Le transport fluviatile a été assez important et a apporté des os roulés et une partie de l’assemblage a pu être transportée et les restes se seraient déposés dans une zone du lit de la rivière (dépôt secondaire),
4) L’enfouissement a été rapide, 5) L’évolution géochimique post-dépositionnelle a été importante et a altéré
les surfaces des ossements avec de nombreuses concrétions.
6. DAYU Ce site a été fouillé en 1983 sur une surface de 20 m² dans la partie inférieure de la formation de Kabuh et plus particulièrement à sa base (Pasir Conglomeratik). Le nombre

Synthèse.
- 252 -
faible de restes (33) ne nous permet pas d’obtenir beaucoup de résultats. Une collection de surface a été établie lors des fouilles (26 restes). Nous n’avons pas pu déterminer d’individus sur ce site. L’étude biostratigraphique est donc impossible. Si l’on se base sur le contexte lithostratigraphique et sur les données bibliographiques en paléontologie, cet assemblage serait proche de l’association de Trinil H.K. car il est situé dans le Grenzbank. L’étude du paléoenvironnement est également impossible.
Les esquilles sont les plus nombreuses dans l’assemblage. Les éléments les mieux représentés sont les éléments crâniens et les membres antérieurs, mais ceci est à nuancer car le nombre de restes considéré est souvent faible et donc peu significatif.
Les esquilles très majoritaires ont leur longueur comprise entre 20 et 50 millimètres. Cette grande fracturation est confirmée par l’indice général d’intensité de la fragmentation (IGIF), l’indice de l’intensité de fragmentation des ossements (IFO) et le pourcentage de dents fragmentées.
Les restes sont très fortement affectés par les traces d’oxydes, les zones de desquamation étendues (d3) et les concrétions (l), peu par la fissuration. Ces concrétions sont caractéristiques du niveau du Grenzbank comme à Bukuran. Le transport fluviatile est difficile à mettre en évidence car le nombre de restes est faible.
L’origine de cette accumulation semble difficile à définir du fait du manque de données au niveau du nombre de restes et car l’évolution post-dépositionnelle qui altére les ossements a été très intense.
Le scénario de formation de cet assemblage a pu être le suivant : 1) Quelques cadavres se sont déposés (dépôt primaire). Ces morts peuvent être
liées à un piégeage dans les sédiments meubles au bord d’une rivière ou à une prédation par les crocodiles.
2) Nous ne pouvons pas proposer de scénario pour cette phase vu le manque de données,
3) L’enfouissement a été rapide, 4) L’évolution géochimique post-dépositionnelle a été importante et a altéré les
surfaces des ossements avec de nombreuses concrétions.

- 253 -
CONCLUSION
Les déterminations des espèces obtenues lors de notre étude confirment celles des travaux précédents (Aimi & Aziz, 1985 ; van den Bergh et al., 2001a). La présence de certaines espèces nous a permis de caler biostratigraphiquement ces associations fauniques (tableau 83). La zone du dôme de Sangiran permet ou permettra d’affiner le cadre biostratigraphique de l’île de Java notamment pour l’espèce Pachycrocuta brevirostris sur le site de Bukuran.
FORMATIONS ESPECES
Kabuh Partie supérieure
Elephas sp., Rhinoceros sp., Axis sp., Duboisia santeng, Bubalus palaeokerabau, Sus sp.
Kabuh Partie inférieure
Pachycrocuta brevirostris Stegodon sp., Rhinoceros sp., Hexaprotodon sivalensis, Cervus (Rusa) sp., Axis lydekkeri, Axis sp., Duboisia santeng, Bubalus palaeokerabau, Bibos palaesondaicus, Sus brachygnathus, Sus sp.,
Tableau 84 : Récapitulatif des espèces présentes dans les différentes formations sédimentaires de Sangiran
suite à notre étude.
La nature des espèces nous informe sur les paléoenvironnements du dôme de Sangiran au Pléistocène moyen ancien. Les espèces indiquent des paléomilieux assez homogènes suivant les sites. Les espèces présentes dans la formation de Kabuh vivaient dans un milieu proche d’une forêt ouverte de type forêt décidue ou savane arborée proche d’un point d’eau (rivière). Ces résultats nous permettent de confirmer les données obtenues par la palynologie, qui reflètent une ouverture du paysage due à une aridité relative du climat à cette époque (Sémah A.M., 1986) et l’opinion de Belwood (1997) qui place les hominidés de Sangiran dans un milieu mosaïque. Ce changement majeur du climat a permis les migrations de mammifères sur l’île de Java et plus précisément à Sangiran. Là, l’espace était assez ouvert

Conclusion.
- 254 -
et présentait des cours d’eau ce qui était favorable à l’installation des mammifères et de l’Homo erectus. Ces conclusions vont contre celles proposées par Pope (1984) qui considère que le milieu de vie d’Homo erectus était la forêt tropicale.
Au niveau taphonomique, la plupart des sites hormis Tanjung, montrent une représentation squelettique dominée par les restes dentaires et les restes de l’autopode. L’intensité de la fragmentation est importante pour tous les sites. Cette fragmentation est le résultat de plusieurs actions : le weathering et le transport fluviatile principalement. Comme nous l’avons démontré, l’action de l’eau est prédominante sur tous ces assemblages, quelle soit chimique ou mécanique. Nous avons tenté de quantifier le transport fluviatile mais il faudrait compléter ces premiers résultats par une étude de l’orientation des ossements et par une étude sur les paléocourants. À la suite de ceci, nous proposons de positionner les différents sites de Sangiran par rapport au lit d’une rivière en prenant en compte les résultats des groupes de Voorhies, de Badgley et des rapports dents/vertèbres (figure 71). Le milieu de dépôt du site de Bukuran devait être proche d’un dépôt de chenal alors que les milieux de dépôt des sites de Tanjung 82, Sendang Busik, Ngrejeng et Grogol étaient des milieux de dépôt plus calmes de type plaine d’inondation. Tanjung 63-64 devait être assez éloigné du lit de la rivière même si 10 % de ses restes sont roulés. L’étude du transport fluviatile permet de compléter les données sur la datation et donc d’être prudent sur l’interprétation des dates obtenues à partir des ponces qui ont pu être transportées à partir d’un dépôt primaire plus ancien (Langbroek & Roebroeks, 2000). En effet, les datations obtenues à partir de tels produits volcaniques peuvent être beaucoup plus anciennes que les niveaux où ces ponces sont situées actuellement.
Figure 71 : Hypothèse des positions des sites par rapport à un cours d’eau d’après les assemblages de Badgley.
Au Pléistocène moyen, certains individus sont morts au bord d’une rivière. Cette mort a pu être accidentelle, naturelle ou liée à une prédation. Ces carcasses ont peut-être été

Conclusion.
- 255 - 255
charognées par les carnivores ou Homo erectus, puis en partie transportée par l’eau, les éléments les plus denses restant sur place.
L’enfouissement des restes a dû être assez rapide car les ossements n’ont pas subi une très grande fissuration. Les ossements ont pu être ensevelis par des coulées de débris volcano-sédimentaires à la suite de fortes pluies ou lors d’éruptions volcaniques. Ensuite, l’action géochimique a eu un effet important sur la surface de ces ossements que ce soit des traces d’oxydes ou de concrétions liées à la nature des minéraux présents ou transportés dans le sol.
Si Homo erectus a eu une influence sur les assemblages, celle-ci n’est pas évidente. Le transport fluviatile et la forte action chimique sur les ossements ont pu en partie dissimuler des stigmates d’actions anté-dépositionnelles.
En outre, les outils sont rares sur l’ensemble des sites hormis des éclats à Tanjung 63-64 et une bola à Tanjung 82. Des outils ont pourtant été découverts sur le dôme de Sangiran tout d’abord à Ngebung par von Koenigswald (1936), dans les années 70 Soejono a également retrouvé un chopping tool sur ce site (Soejono, 1982) puis dans les années 90, à Ngebung 2, l’équipe franco-indonésienne de la Mission Quaternaire et Préhistoire en Indonésie (M.Q.P.I.) a mis au jour des outils associés à des restes fauniques (Sémah F. et al., 2002). Depuis 1995, de nouvelles recherches sur l’industrie lithique ont été entreprises dans cette zone par Widianto (Widianto et al.,1996 ; Jatmiko, 2001). Il a pu mettre en évidence la présence d’outils dès la base de la formation de Kabuh. Par conséquent, des outils sont présents à Sangiran au Pléistocène moyen ancien, alors pourquoi sont-ils si rares dans les assemblages de notre étude ? Nous pouvons proposer différentes hypothèses :
- Les assemblages sont éloignés des zones d’activités où les hommes ont pu laisser leurs outils car ils ont été transportés,
- Les hommes ne laissaient pas leurs outils sur des sites de charognage, - Les outils n’ont pas été transportés de la même manière que les ossements, ils étaient
plus denses et n’ont pas ou très peu bougé de leur lieu de dépôt et c’est le cas contraire pour les éclats, étant légers ils ont été transportés plus loin. Les outils peuvent être en andésite et en calcédoine qui sont des matériaux résistants à l’altération,
- Le nombre d’outils à cette époque était faible (Pope, 1989, 1995), - La matière première n’était pas disponible dans la zone de Sangiran. Ceci est
confirmé par les premières recherches de von Koenigswald (1940) sur l’origine de la matière première des éclats de Ngebung dans les montagnes du sud (Simanjuntak & Sémah F., 1996).
- Le site de Ngebung 2 a peu subi le transport fluviatile et donc se rapproche d’un sol d’habitat et donc d’un dépôt primaire ce qui explique la présence des outils.
Si les outils sont présents dans cette zone, alors de la même façon pourquoi les traces
anthropiques liées à leur utilisation sont-elles très rares ? Le transport fluviatile et l’évolution

Conclusion.
- 256 - 256
géochimique post-dépositionnelle sont probablement à l’origine de la sous-estimation du nombre de ces traces. Cette rareté des traces anthropiques a également été remarquée par Choi (2003), qui n’a trouvé que cinq restes présentant des stries sur un total de 30 000 ossements étudiés provenant de Java. Ces cinq restes proviennent néanmoins de ramassages de surface effectués à Sangiran. Des stigmates anthropiques ont été observés sur des fossiles de Ngebung 2 mais en faible quantité. À partir de ces résultats, il est difficile de conclure que le comportement de subsistance d’Homo erectus était basé sur la consommation de viande comme le propose Choi (2003).
De nombreux restes humains ont été découverts sur le dôme de Sangiran et plus particulièrement sur une partie des sites de notre étude comme Tanjung, Sendang Busik, Bukuran et Dayu dans les couches de Kabuh. La présence de l’homme est donc attestée à cette époque. Ces restes crâniens sont associés aux restes fauniques et sont donc contemporains de ceux-ci. Ces fragments de crânes ne présentent pas un aspect roulé et ont donc peu subi le transport fluviatile ce qui s’explique également par leur densité (groupe III de Voorhies). Ils proviennent probablement du même stock que les ossements de mammifères non roulés.
Homo erectus a dû choisir le dôme de Sangiran, pour sa situation proche des cours d’eau et l’abondance des espèces de mammifères. Le nombre d’espèces présentes semble faible par rapport à la biodiversité d’un écosystème tropical actuel. Nous pouvons nous interroger sur ceci. Est-ce que cela est dû à une chasse sélective ou à la conservation, à la fragmentation et à une altération géochimique trop intenses ou tout simplement à une diversification des sources alimentaires ? L’hypothèse d’une chasse paraît peu probable mais une activité de charognage a très bien pu avoir lieu. Huffman (2001) défend la thèse de la diversité des sources alimentaires. Pour cet auteur, la diversité actuelle des milieux géographiques (volcans, côtes, plaines…) reflète la diversité des habitats et des sources d’approvisionnement pour Homo erectus.
L’action anthropique au Pléistocène semble bien avoir existé mais il n’est pas aisé de la mettre en évidence sur le dôme de Sangiran compte tenu du transport fluviatile et de l’action diagénétique importante.
Cette rareté des traces nous conduit à nous poser diverses interrogations sur la stratégie de subsistance d’Homo erectus :
- Son comportement de subsistance à Java est-il différent de celui des mêmes hominidés dans d’autres zones du globe ? L’insularité a-t-elle provoqué une adaptation de ces hominidés à des conditions particulières ?
Selon Huffman (2001), le comportement de subsistance d’Homo erectus à Java ne serait pas différent de celui des Homo erectus d’Afrique, mais le cadre géographique et l’écologie sont eux différents. Par conséquent, à Java vu la diversité des milieux (volcans, côtes, plaines, mangroves…) et donc des habitats, les sources de nourriture varient et sont très diversifiées (fruits, tubercules, noix, champignons, animaux aquatiques, oiseaux, invertébrés, petits

Conclusion.
- 257 - 257
mammifères…) ce qui ne demande pas un comportement spécialisé sur une espèce ou sur une activité (chasse, charognage, cueillette…). La rareté des outils s’expliquerait alors par la disponibilité de la matière première selon les zones.
En outre, si l’on compare ces données avec celles obtenues pour des sites à Homo erectus de la même époque en Afrique de l’est (Bunn, 1983 ; Bunn & Kroll, 1986) ou en Europe tel que les sites d’Ambrona (Villa et al., 2005) et de Torralba (Villa, 1990) nous constatons que la fréquence des traces anthropiques est également très faible.
- Le faible nombre d’outils retrouvés implique-t-il l’utilisation d’une autre matière tel que le bois (bambou) ?
Selon Pope (1989 ; 1995), le bambou a été très utilisé par les hominidés en Asie du Sud-Est au détriment de la matière première lithique ce qui expliquerait la rareté des outils lithiques sur les sites et celle des stigmates sur les ossements. Mais il est difficile d’émettre cette hypothèse sachant la difficulté de retrouver des traces de cette activité et que les expérimentations menées avec le bambou n’ont pas été très concluantes (Choi, 2003).
- Le régime alimentaire d’Homo erectus à Java était-il essentiellement végétarien ?
Selon Jacob (2001), la morphologie crânienne de cet hominidé reflète une intense mastication d’aliments végétaux mais il pense également que ce dernier devait pratiquer la chasse. On peut penser que sa nourriture était diversifiée et que la consommation de viande était aléatoire suivant les opportunités. Ceci a dû favoriser le charognage. Il n’est donc pas aisé de proposer une hypothèse de comportement de subsistance pour Homo erectus à Java. Néanmoins, nos résultats sont plus en accord avec l’hypothèse d’Huffman.
À partir de collections, certes parfois restreintes en nombre de restes, nous avons pu extraire de nombreux résultats cohérents en utilisant et croisant différentes méthodes et notamment au cours de l’étude taphonomique. Ces méthodes peuvent parfois sembler redondantes mais elles sont nécessaires pour valider et recouper nos résultats. Nous avons rencontré à diverses reprises des problèmes d’applications de méthodes généralement utilisées pour l’étude des sites archéologiques et qui en milieu tropical ou sur des échantillons de petite taille doivent être modifiées. Il serait donc important de développer de nouvelles méthodes notamment pour exploiter au maximum les esquilles.
Ces premiers résultats taphonomiques sont très importants pour comprendre l’origine des accumulations osseuses dans cette région. La systématisation de l’étude taphonomique des assemblages sur l’île de Java permettrait de compléter les données parcellaires de la taphonomie en milieu tropical et de mieux comprendre le comportement de subsistance d’Homo erectus à Java au Pléistocène afin de le comparer avec des comportements mieux connus en Europe, en Afrique ou en Asie.

Conclusion.
- 258 - 258
Toutes ces données obtenues à partir des collections fouillées de ces sites sont très intéressantes et démontrent l’intérêt d’entreprendre de nouvelles campagnes de fouilles dans cette région pour tenter de trouver des sites en place qui n’auront que très peu subi le transport fluviatile. Ces fouilles devront être organisées dans un cadre stratigraphique bien précis pour apporter un maximum d’informations et permettre ainsi de compléter ce travail.

- 259 -
REFERENCES BIBLIOGRAPHIQUES AARSSE R., 1993, L’Indonésie, coll. Méridiens, Peuples et Pays du Monde, Karthala ed.,
Paris, 184 pp. AIMI M. & AZIZ F., 1985, Vertebrate fossils from the Sangiran dome, Mojokerto, Trinil
and Sambungmacan areas, in : Quaternary geology of the hominid fossil bearing formations in Java, Geological Research and Development Centre ed., Bandung, 155-197.
ANDREWS P., 1990, Owls, caves and fossils : predation, preservation and accumulation of
small mammal bones in caves with an analysis of the Pleistocene cave faunas from Westbury-sub-Mendip, Somerset, UK, University of Chicago Press ed., Chicago, 231 pp.
ANDREWS P., LORD J.M. & NESBIT-EVANS E.M., 1979, Patterns of ecological
diversity in fossil and mammalian faunas, Biological Journal of the Linnean Society, London, 11, 177-205.
ANONYME, 1891 a, Palaeontologische onderzoekingen op Java, Verslag v.h. Mijnwezen
over het 3de kwartaal 1891, 12-14. ANONYME, 1891 b, Palaeontologische onderzoekingen op Java, Verslag v.h. Mijnwezen
over het 4de kwartaal 1891, 12-15. ANONYME, 1892, Palaeontologische onderzoekingen op Java, Verslag v.h. Mijnwezen over
het 2de kwartaal 1892, 14-17. ANONYME, 1893 a, Palaeontologische onderzoekingen op Java, Verslag v.h. Mijnwezen
over het 3de kwartaal 1892, 10-14. ANONYME, 1893 b, Palaeontologische onderzoekingen op Java, Verslag v.h. Mijnwezen
over het 4de kwartaal 1892, 11-12. ANONYME, 1893 c, Palaeontologische onderzoekingen op Java, Verslag v.h. Mijnwezen
over het 2de kwartaal 1893, 12-13.

Références bibliographiques
- 260 -
ANONYME, 1893 d, Palaeontologische onderzoekingen op Java, Verslag v.h. Mijnwezen over het 3de kwartaal 1893, 15-17.
ANONYME, 1894, Palaeontologische onderzoekingen op Java, Verslag v.h. Mijnwezen over
het 4de kwartaal 1893, 14-15. AUGUSTE P., 1994 a, Thème I : Actions climatiques et édaphiques - Synthèse générale, in :
Patou-Mathis M. (ed), Taphonomie/bone modification - Outillage peu élaboré en os et bois de Cervidés IV, Artefacts, 9, 17-27.
AUGUSTE P., 1994 b, Introduction générale - La fossilisation, in : Patou-Mathis M. (ed),
Taphonomie/bone modification - Outillage peu élaboré en os et bois de Cervidés IV, Artefacts, 9, 11-14.
AUGUSTE P., 1995, Cadres biostratigraphiques et paléoécologiques du peuplement dans la
France septentrionale durant le Pléistocène. Apports de l’étude paléontologique des grands mammifères du gisement de Biache Saint Vaast (Pas de Calais), Thèse de doctorat, M.N.H.N., I.P.H., Paris.
AUGUSTE P. & PATOU-MATHIS M., 1999, Beauvais (Oise) : un nouveau gisement du
Paléolithique moyen dans le Bassin parisien, in : The Role of Early Humans in the Accumulation of European Lower and Middle Palaeolithic Bone Assemblages, Römisch-Germanisches Zentralmuseum, 42, 343-365.
AZIZ F. & DE VOS J., 1999, The fossil faunas from the Citarum area, West Java, Indonesia,
D.E.I.N.S.E.A., 7, 21-32. BABA H. & AZIZ F., 1991, Hominid skull fragments found in 1979 from Sangiran, Central
Java, Bulletin of the National Science Museum, Tokyo, serie D, 17, 1-8. BABA H., AZIZ F., NARASAKI S., SUDIJONO, KAIFU Y., SUPRIJO A., HYODO M.,
SUSANTO E.E. & JACOB T., 2000, A new hominid incisor from Sangiran, Central Java, Journal of Human Evolution, 38, 855-862.
BACKER C.A. & BAKHUISEN VAN DEN BRINK R.C., 1965, Flora of Java, 2, N.V.P.,
Noordhoff ed., Groningen, 641 pp. BADGLEY C., 1982, Community reconstruction of a Siwalik mammalian assemblage, PhD,
Yale University.

Références bibliographiques
- 261 -
BADGLEY C., 1986a, Counting Individuals in Mammalian Fossil Assemblages from Fluvial Environments, PALAIOS, vol. 1, 328-338.
BADGLEY C., 1986b, Taphonomy of mammalian fossil remains from Siwalik rocks of
Pakistan, Paleobiology, 12 (2), 119-142. BADOUX D.M., 1959, Fossils mammals from two fissure deposits at Punung (Java),
Drukkerij en uitgeversmij v/h Kemink en Zoon N.V. Domplein 2, 153 pp. BAILON S. & RAGE J.-C., 1992, Amphibiens et reptiles du Quaternaire, Relations avec
l’homme, Mém. Soc. Géol. France, (160), 95-100. BARONE R., 1999, Anatomie comparée des mammifères domestiques - Ostéologie, Vigot
ed., Paris, tome 1, 761 pp. BARTSTRA G.-J., 1983, The fauna from Trinil, type locality of Homo erectus : a
reinterpretation, Comment I : The vertebrate bearing-deposits of Kedung Brubus and Trinil, Java, Indonesia, Geologie en Mijnbouw, 16 (7746), 329-336.
BARTSTRA G.-J., 1985, Sangiran, the stone implements of Ngebung, and the paleolithic of
Java, Modern Quaternary Research in South-East Asia, 9, 99-113. BEAUFORT L.F. de, 1913, Fishes of the eastern part of the Indo-Australian Archipelago,
with remarks on its zoogeography, Bijdr. Tot de Dierk., 19, 95-164. BEDEN M., 1979, Les éléphants (Loxodonta et Elephas) d'Afrique Orientale: systématique,
phylogénie, intérêt biochronologique, Thèse de doctorat, université de Poitiers, 567 pp. BEHRENSMEYER A.K., 1973, The taphonomy and paleoecology of Plio-Pleistocene
vertebrate assemblages, East of Lake Rudolf, Kenya, PhD, Harvard University, 188 pp. BEHRENSMEYER A.K., 1975, The taphonomy and paleoecology of Plio-Pleistocene
vertebrate assemblages of Lake Rudolf , Kenya, Bulletin of the Museum of Comparative Zoology , 146, 473-578.
BEHRENSMEYER A. K., 1978, Taphonomic and ecologic information from bone
weathering, Paleobiology, 4, 150-162.

Références bibliographiques
- 262 -
BEHRENSMEYER A.K., 1988, Vertebrate preservation in fluvial channels, Palaeogeography, palaeoclimatology, palaeoecology, 63, 183-199.
BEHRENSMEYER A. K., 1990, Experimental taphonomy workshop, congrès ICAZ, 23 mai
1990, 25pp. BEHRENSMEYER A.K. & DECHANT-BOAZ D.E., 1980, The recent bones of Amboseli
Park, Kenya, East African paleoecology, in : Behrensmeyer A.K. & Hill A.P. (eds.), Fossils in the making - Vertebrate taphonomy and paleoecology, University of Chicago Press ed., 72-92.
BELLWOOD P., 1997, Prehistory of the Indo-Malayan Archipelago, revisited edition,
University of Hawai’i Press ed., Honolulu, 384 pp. BEMMELEN VAN R.W., 1949, The Geology of Indonesia and Adjacent Archipelagos,
Martinus Nijhoff ed., Den Haag, 1, 732 pp. BERGH VAN DEN G.D., 1988, The endemic pleistocene bovid from Java (Indonesia), 56
pp. BERGH VAN DEN G.D., 1999, The Late Neogene elephantoid-bearing faunas of Indonesia
and their palaeozoogeographic implications. A study of the terrestrial faunal succession of Sulawesi, Flores, and Java, including evidence for early hominid dispersal east of Wallace’s Line, Scripta Geologica, 117, 419 pp.
BERGH VAN DEN G.D., DE VOS J., SONDAAR P.Y. & AZIZ F., 1996, Pleistocene
zoogeographic evolution of Java (Indonesia) and glacio-eustatic sea level fluctuations : a background for the presence of Homo, Bulletin Indo-Pacific Prehistory Association, 1, (14), 7-21.
BERGH VAN DEN G.D., DE VOS J. & SONDAAR P.Y., 2001 a, The Late Quaternary
palaeogeography of mammal evolution in the Indonesian Archipelago, PALAEO, (171), 385-408.
BERGH VAN DEN G.D., DE VOS J., AZIZ F. & MORWOOD M.J., 2001 b,
Elephantoidea in the Indonesian region : new Stegodon findings from Flores, The World of Elephants, International Congress, Rome 2001, 623-627.

Références bibliographiques
- 263 -
BERGMANN C., 1847, Uber die Verhaltnisse der Warmekonomie der Thiere zu ihrer Grosse, Gottingen studien, 3, 595-708.
BERGMAN R.A.M. & KARSTEN P., 1952, The fluorine content of Pithecanthropus and of
other specimens from the Trinil fauna, Verhandelingen der Koninklijke Nederlandse akademie van wetenschappen, B, LV, 150-152.
BETTIS E.A., ZAIM Y., LARICK R.R., CIOCHON R.L., SUMINTO, RIZAL Y.,
REAGAN M. & HEIZLER M., 2004, Landscape development preceding Homo erectus immigration into Central Java, Indonesia: the Sangiran Formation Lower Lahar, PALAEO, 206, 115-131.
BIBERSON B. & AGUIRRE E., 1965, Expériences de taille d'outils préhistoriques dans des
os d'éléphant, Quaternaria, VIII, 165-183. BINFORD L.R., 1978, Nunamiut Ethnoarchaeology, Academic Press ed., New York, 283
pp. BINFORD L.R., 1981, Bones - Ancient man and modern myths, Academic Press ed., New
York, 320 pp. BINFORD L.R., 1983, In pursuit of the past, Thames and Hudson ed., New York, 256 pp. BINFORD L.R., 1984, Faunal Remains from Klasies River mouth, Academic Press ed., New
York, 283 pp. BLUMENSCHINE J., 1986, Early hominid scavenging opportunities : implications of
carcass availibility in the Serengeti and Ngorongoro ecosystems, BAR International Series, 283.
BLUMENSCHINE J., 1988, An experimental model of the timing of hominid and carnivore
influence on archaeological bones assemblages, Journal of Archaeological Science, 15, 483-502.
BLUMENSCHINE J. & MAREAN C.W., 1993, A carnivore’s view of archaeological bone
assemblages, in : Hudson J. (ed.), From bones to behavior : ethnoarchaeological and experimental contributions to the interpretation of faunal remains, Center for Archaeological Investigations, occasional paper, University of Southern Illinois Press, (21), 273-300.

Références bibliographiques
- 264 -
BOITANI L. & BARTOLI S., 1983, Les Mammifères, Nathan ed., Paris, 512 pp. BRAIN C.K., 1981, The hunters or the hunted ? - An introduction to African cave
taphonomy, University of Chicago Press ed., Chicago, 376 pp. BRIDAULT A., 1994, La fragmentation osseuse : modèle d’analyse pour les séries
mésolithiques, in : Patou-Mathis M. (ed), Taphonomie/bone modification - Outillage peu élaboré en os et bois de Cervidés IV, Artefacts, 9, 155-166.
BRINK VAN DEN L.M., 1982, On the mammal fauna of the Wajak Cave, Java (Indonesia),
Modern Quaternary Research in South-East Asia, 7, 177-193. BRONGERSMA L.D., 1937, Notes on some recent and fossil cats, chiefly from the Malay
Archipelago, Brill ed., Leiden, 89 pp. BRUGAL J.-P., 1994, Introduction générale, Action de l’eau sur les ossements et les
assemblages fossiles, in : Patou-Mathis M. (ed), Taphonomie/bone modification - Outillage peu élaboré en os et bois de Cervidés IV, Artefacts, 9, 121-129.
BRUGAL J.-P. & JAUBERT J., 1991, Les gisements paléontologiques pléistocènes à
indices de fréquentation humaine : un nouveau type de comportement de prédation ?, Paléo, (3), 15-41.
BRUGAL J.-P. & PATOU-MATHIS M., 1993, L’assemblage osseux de l’abri des
Canalettes : présentation générale, in : Meignen L. (ed.), L’abri des Canalettes, un habitat moustérien sur les grands Causses (Nant, Aveyron): fouilles 1980-1986, C.N.R.S. ed., 77-87.
BRUGAL J.-P., DAVID F. & FARIZY C., 1994, Quantification d’un assemblage osseux :
paramètres et tableaux, in : Patou-Mathis M. (ed.), Taphonomie/bone modification - Outillage peu élaboré en os et bois de Cervidés IV, Artefacts, 9, 143-153.
BUEHLER K., 1997, Where's the Cliff ? Late Archaic Bison Kills in the Southern Plains,
Plains Anthropologist Memoir, 29, (159). BUNN H.T., 1983, Evidence on the diet and subsitence pattern of Plio-Pleisotocene hominids
at Koobi For a, Kenya and Olduvai Gorge, Tanzania, in : Clutton-Brock J. & Grigson C. (eds.), Animals and Archaeology: 1.Hunters and their prey, BAR International Series, 163, 21-30.

Références bibliographiques.
- 265 - 265
BUNN H.T. & KROLL E., 1986, Systematic butchery by Plio-Pleisotocene hominids at Olduvai Gorge, Tanzania, Current anthropology, 27, (5), 431-452.
CARTHAUS E., 1911, Arbeitsbericht über die Ausgrabungen, II, Teil, Die Arbeiten von
August bis November 1907, in : Selenka & Blanckenhorn (eds.), Die Pithecanthropus- Schichten auf Java, Geologische und Paläontologische Ergebnisse der Trinil-Expedition (1907 und 1908), Leipzig, 38-39.
CASSOLI P.F., FABBO G., FIORE I., PIPERNO M. & TAGLIACOZZO A., 1999,
Taphonomic analysis of some levels of the Lower Palaeolithic site of Notarchirico (Venosa, Basilicata, Italy), in : The Role of Early Humans in the Accumulation of European Lower and Middle Palaeolithic Bone Assemblages, Römisch-Germanisches Zentralmuseum, 42, 53-172.
CHACORNAC-RAULT M., 2004, Etude palynologique et paléoenvironnementale de
sondages holocènes dans les Gunung Sewu (Java, Indonésie) - Reconstitution de l’environnement, impacts climatiques et anthropiques - Mise en évidence de la néolithisation, Thèse de Doctorat, M.N.H.N., I.P.H., Paris, 365 pp.
CHAPPEL J., OMURA A., ESAT T., MC CULLOCH M., PANDOLFI J, OTA Y. &
PILLANS B., 1996, Reconciliation of Late Quaternary sea levels derived from coral terraces at Huon peninsula with deep sea oxygen isotope records, Earth and Planetary Science Letters, 141, 227-236.
CHAZAN M., MONCHOT H., PORAT N., LISTER A., DAVIES P. & HORWITZ L.,
2001, Le site acheuléen de plein air d’Holon (Israël) : premiers résultats, Compte-Rendus de l’Académie des Sciences, série Sciences de la Terre et de l’Univers, Paris, 332, 201-207.
CHAZAN M., MONCHOT H. & HORWITZ L.K., (sous presse), Spatial analysis of lithics
artefacts and fauna from Holon, in : M. Chazan & L.K. Horwitz (eds.), The Lower Paleolithic Site of Holon, Israel, Peabody Museum, Harvard University.
CHENORKIAN R., 1996, Pratique archéologique statistique et graphique, Errance ed., Paris, 162 pp.
CHOI K., 2003, Subsistence and tool use behavior of Homo erectus in Java: an experimental
and taphonomic approach, PhD, University of Wisconsin-Madison, 302 pp.

Références bibliographiques.
- 266 - 266
COLBERT E.H., 1943, Pleistocene vertebrates collected in Burma by the American Southeast Asiatic expedition, Transactions of the American Philosophical Society, part III, 395-429.
COLBERT E.H. & HOOIJER D.A., 1953, Pleistocene mammals from the limestone
fissures of Szechwan, China, Bulletin of the American Museum of Natural history, 102, 1-134.
COPE E.D., 1874, On the homologies and origin of the types of molar teeth of mammalia
educabilia, Journal of the Academy of Naural Scences of Philadelphia, 2ème série, 8, art. III, 71-91.
CORBET G.B. & HILL J.E., 1992, The mammals of the Indomalayan Region : a systematic
review, Natural History Museum publications, 488 pp. CRUZ-URIBE K, 1991, Distinguishing hyena from hominid bone accumulations, Journal of
Field Archaeology, 18, 467-486. CURTIS G.H., 1981, Man’s immediate forerunners - Establishing a relevant time scale in
anthropological and archaeological research, Philosophical Transactions of the Royal Society of London, series B, 292, 7-20.
DACEY M.F., 1973, Statistical tests of spatial association in the locations of tool types,
American antiquity, 38, (3), 320-328. DAGET J., 1979, Les modèles mathématiques en écologie, Masson ed., Paris, 172 pp. DARWIN C., 1859, Sur l’origine des espèces, Flammarion ed., Paris, 608 pp. DAVIS S.J.M., 1983, The age profiles of Gazelles predated by ancien Man in Israel: possible
evidence seasonality to sedentism in the Natufian, Paléorient, 9, (1), 55-62. DAY M.H. & MOLLESON T.I., 1973, The Trinil femora, Human Evolution, 11, 127-154. DELPECH F., 1983, Les faunes du Paléolithique supérieur dans le Sud-Ouest de la France,
coll. Cahiers du Quaternaire, C.N.R.S. ed., 453 pp.

Références bibliographiques.
- 267 - 267
DELPECH F. & GUERIN C., 1996, Paléoenvironnement : l’animal, le milieu, le climat, in : Guérin C. & Patou-Mathis M. (eds.), Les grands mammifères plio-pléistocènes d’Europe, Masson ed., Paris, 243-253.
DENNELL R.W., 2005, Early Pleistocene hippopotamid extinctions, monsoonal climates,
and river system histories in South and South West Asia : comment on Jablonski (2004) (“The hippo’s tale: how the anatomy and physiology of Late Neogene Hexaprotodon shed light on Late Neogene environmental change” Quaternary International, 117, 119-123), Quaternary International, 126-128, 283-287.
DERMITZAKIS M., SONDAAR P.Y., DE VOS J. & VAN DEN BERGH G.D., 1997,
Colonisation of Islands by man with special reference to Crete, Anthropologia, 4, 5-26. DESCOMBES J.C., 1980, La première faune rissienne de la basse vallée de la basse vallée
de la Seine, Implications biostratigraphiques et paléoécologiques, Thèse de doctorat, Université de Poitiers, 214 pp.
DESSE J., CHAIX L. & DESSE-BERSET N., 1986, Ostéo : base-réseau de données
ostéométriques pour l’archéozoologie, coll. Notes et monographies techniques, (20), C.N.R.S. éd., 161 pp.
DETROIT F., 2002, Origine et évolution des Homo sapiens en Asie du Sud-Est :
Descriptions et analyses morphométriques de nouveaux fossiles, Thèse de doctorat, M.N.H.N., I.P.H., Paris, 444 pp.
DODSON P., 1973, The significance of small bones in paleoecological interpretation,
Univ.Wyoming Contrib.Geol., Laramie, 12, (1), 15-19. DOZY C.M., 1911, Arbeitsbericht über die Ausgrabungen, III, Teil, Die Arbeiten im Jahre
1908, in : Selenka & Blanckenhorn (eds.), Die Pithecanthropus- Schichten auf Java, Geologische und Paläontologische Ergebnisse der Trinil-Expedition (1907 und 1908), Leipzig, 40-42.
DJUBIANTONO T., 1977, Geologi daerah Kertosobo, Sangiran, Skripsi Sarjana Muda pada
Akademi Geologi dan Pertambangan Bandung, 98 pp. DJUBIANTONO T., 1992, Les derniers dépôts marins de la dépression de Solo (Java
Central, Indonésie), Chronostratigraphie et paléogéographie, Thèse de doctorat, M.N.H.N., I.P.H., Paris, 206 pp.

Références bibliographiques.
- 268 - 268
DJUBIANTONO T. & SEMAH F., 1992, L’évolution de la région de Solo au Quaternaire, Les Dossiers d’Archéologie, 184, 46-48.
DUBOIS E., 1894, Pithecanthropus erectus : eine menschenaehnliche Uebergangsform aus
Java, Landesdruckerei ed., Batavia, 39 pp. DUBOIS E., 1896, Pithecanthropus erectus, eine menschenaehnliche Uebergangsform,
Compte rendu des séances du 3ème Congrès international de Zoologie, 251-271. DUBOIS E., 1908, Das geologische alter der Kendeng-oder Trinil fauna, Tijdschrift van het
Koninklijk Nederlansch Aardrijkskundig Genootschap, 25, 1235-1270. DUBOIS E., 1920, De proto-Australische fossiele mensch van Wadjak (Java), I-II,
Proceedings Koninklijke Akademie van Wetenschappen te Amsterdam, 19, 88-105, 866-887.
DUBOIS E., 1924, On the principal characters of the cranium and the teeth of
Pithecanthropus erectus, Proceedings Koninklijke Nederlandse Akademie van Wetenschappen, 27, 265-278.
DUBOIS E., 1937, On the Fossil Human Skulls Recently Discovered in Java and
Pithecanthropus erectus, MAN, January 1937, 1-7. DUIJFJES J., 1936, Zur Geologie und Stratigraphie des Kendenggebietes zwischen Trinil
und Soerabaja (Java), De Ingenieur in Nederlandsch Indië, 8, 136-149. DURAND F., 1994, Les forêts en Asie du Sud-Est – Recul et exploitation, Le cas de
l’Indonésie, L’Harmattan ed., Paris, 396 pp. EFREMOV J.A., 1940, Taphonomy : new branch of Paleontology, Pan-American geologist,
74, (2), 81-93. ES VAN L.J.C., 1931, The Age of Pithecanthropus, Martinus Nijhoff ed., Den Haag, 142 pp. ES VAN L.J.C., 1936, L’Homme fossile aux Indes néerlandaises, Report of XVI Session,
United States of America 2, International Geological Congress, Washington, D.C., 1231-1237.

Références bibliographiques.
- 269 - 269
FAE M., 1996, Lithobiostratigraphy and fossil hominids of the Sangiran-Krikilan area, Java (Yogyakarta, Indonesia), Memoire di Scienze Geologiche, 48, 143-153.
FALCONER H., 1857, On the species of mastodon and elephant occurring in the fossil state
in Great Britain, Part I : Mastodon, Quarterly Journal of the Geological Society of London, 13, p 307- 360.
FALCONER H. & CAUTLEY P.T., 1846, Fauna antiqua sivalensis being the fossil zoology
of the Siwalik hills in the North of India, S.Elder ed., London, 92 pp. FAURE M., 1983, Les hippopotamidae (mammalia, artiodactyla) d’Europe occidentale,
Thèse de doctorat, Université C.Bernard, Lyon. FISHER D.C., 1981, Crocodilian scatology, microvertebrate concentrations, and enamel-less
teeth, Paleobiology, 7, (2), 262-275. FISHER J.W. & ROLL T.E., 1999, Prehistoric human exploitation of bison in the great
plains of Montana (USA) during the last 3 000 years, in : Brugal J.P., David F., Enloe J.G. & Jaubert J. (eds.), Le bison : gibier et moyen de subsistance des hommes du Paléolithique aux Paléoindiens des Grandes Plaines, Actes du colloque international Toulouse 6-10 juin 1995, A.P.D.C.A. ed., 417-436.
FLEMING T.H., 1973, Numbers of mammal species in north and central America forest
communities, Ecology, Durham, 54 (3), 555-563. FOLEY R., 1983, Modelling hunting strategies and inferring predators behaviors from prey
attributes, in : Clutton-Brock J. & Grigson C. (eds.), Animals and Archaeology : 1.Hunters and their prey, BAR International Series, 163, 63-75.
FOSSE P., 1992, Taphonomie des grands mammifères pléistocènes : essai de définition,
Préhistoire et Anthropologie Méditerranéennes, 1, 169-177 FOSSE P., 1994, Taphonomie paléolithique : les grands mammifères de Soleilhac (Haute
Loire) et de Lunel-Viel 1 (Hérault), Thèse de doctorat, Université de Provence Aix-Marseille 1, 205 pp.
FOSSE P. & BONIFAY M.F., 1991, Les vestiges osseux de Soleilhac : approche
taphonomique, in : Bonifay E. & Vandermeersch B. (eds.), Actes du 114ème Congrès National des Sociétés Savantes (Paris, 3-9 avril, 1989), C.T.H.S. ed., Paris, 115-133.

Références bibliographiques.
- 270 - 270
FOSSE P., BRUGAL J.P., GUADELLI J.L., MICHEL P. & TOURNEPICHE J.F., 1998, Les repaires d’hyènes des cavernes en Europe occidentale : présentation et comparaison de quelques assemblages osseux, in : Economie préhistorique : les comportements de subsistance au Paléolithique, XVIIIèmes rencontres internationales d’Antibes, 1997, APDCA ed., 43-61.
FOUCAULT A. & RAOULT J.-F., 1995, Dictionnaire de géologie, Masson ed., Paris, 324
pp. GALLET X., 2004, Dynamique de la sédimentation dans les grottes du karst de Punung
(Pacitan, Java) - Relations avec les occupations paléolithiques, Thèse de doctorat, M.N.H.N., I.P.H., Paris, 387 pp.
GAUDZINSKI S., 1995, Wallertheim revisited : a re-analysis of the fauna from the Middle
Palaeolithic site of Wallertheim (Rheinhessen, Germany), Journal of Archaeological Science, 22, 51-66.
GAUDZINSKI S. & ROEBROEKS W., 2000, Adults only, reindeer hunting at the Middle
Palaeolithic site Salzgitter Lebenstedt, Northern Germany, Journal of Archaeological Science, 38, 497-521.
GENET-VARCIN E., 1979, Les Hommes Fossiles, coll. L’Homme et ses origines, Boubée
ed., Paris, 79-89. GERAADS D., 1979, Un nouveau bovidé (Artiodactyla, Mammalia) du Plio-Pléistocène de
Java : Duboisia (?) sartonoi n.sp., Bulletin du Muséum National d’Histoire Naturelle, Paris, 4ème série, 1, section G, (4), 371-376.
GERAADS D., 1979, Nouvelles données sur Hyaena brevirostris bathygnatha Dubois,
(Carnivora, Mammalia) du Pléistocène de Java, Compte Rendu sommaire de la Société géologique de France, 2, 80-82.
GIRARD C. & DAVID F., 1982, Á propos de la chasse spécialisée du Paléolithique moyen :
l’exemple de Mauran (Haute Garonne), Bulletin de la Société préhistorique de France, 79, (1), 11-12.
GRAYSON D.K., 1979, On the quantification of vertebrate archaeofaunas, Advances in
archaeological method theory, New York, 2, 199-237.

Références bibliographiques.
- 271 - 271
GREGORY W.K., 1916, Studies of the evolution of the Primates, Bulletin of the American Museum of Natural History, New York, 35, art. 19, 239-355.
GROVES C.P., 1984, Mammal faunas and the palaeogeography of the Indo-Australian
region, Courier Forshungs Institut Senckenberg, 69, 267-273. GROVES C.P., 1985, Plio-Pleistocene mammals in island southeast Asia, Modern
Quaternary Research in South-East Asia, 9, 43-54. GUADELLI J.-L., 1989, Etude taphonomique du repaire d’hyènes de Camiac (Gironde,
France), Eléments de comparaison entre un site naturel et un gisement préhistorique, Bull. de l’A.F.E.Q., 2, 91-100.
GUERIN C., 1980, Les rhinocéros (Mammalia, Perissodactyla) du Miocène terminal au
Pléistocène supérieur en europe occidentale - Comparaison avec les espèces actuelles, Documents des laboratoires de géologie, Lyon, (79), 1185 pp.
GUERIN C. & PATOU-MATHIS M., 1996, Les grands mammifères Plio-Pléistocènes
d’Europe, Masson ed., Paris, 291 pp. GUERIN C. & FAURE M., 2002, Les grands mammifères, in : Miskovsky J.C. (ed.)
Géologie de la Préhistoire : méthodes, techniques, applications, Maison de la géologie éd., Paris, 859-887.
HAMEAU S., 2004, Datation des sites paléolithiques du Pléistocène inférieur et supérieur de
Punung (Pacitan, Java) - Application des méthodes ESR et U/Th aux grottes de Song Terus et Goa Tabuhan, Thèse de doctorat, M.N.H.N, I.P.H., Paris, 200 pp.
HANSON C.B., 1980, Fluvial taphonomic processes : models and experiments, in :
Behrensmeyer A.K. & Hill A.P. (eds.), Fossils in the making - Vertebrate taphonomy and paleoecology, University of Chicago Press ed., 156-181.
HARDJASASMITA H.S., 1987, Taxonomy and phylogeny of the Suidae (Mammalia) in
Indonesia, Scripta geologica, 85, 1-69. HAYNES G., 1980, Prey bones and predators : potential ecologic information from analysis
of bones sites, OSSA, 7, 75-97.

Références bibliographiques.
- 272 - 272
HAYNES G., 1983, A guide for differentiating mammalian carnivore taxa responsible for gnaw damage to herbivore limb bones, Paleobiology, 14, (1), 81-90.
HEANEY L.R., 1985, Zoogeographic evidence for middle and late Pleistocene land bridges
to the Philippine islands, Modern Quaternary Research in South-East Asia, 9, 127-143. HEEKEREN VAN H.R., 1955, New investigations on the lower Palaeolithic Pajitan culture
in Java, Bulletin of the Archaeological Service of the Republic of Indonesia, 1, 28 pp. HEEKEREN VAN H.R., 1972, The stone Age of Indonesia, Verhandelingen v.h. Koninklijk
Inst. Voor Taal-, Land- en Volkenkunde, Martin Nijhoff ed., Den Haag, (61), 247 pp. HEINTZ E., 1970, Les cervidés villafranchiens de France et d’Espagne, Mémoires du
M.N.H.N., Paris, série C, tome XXII. HENNIG E., 1911, Die Fischreste, in : Selenka L. & Blankenhorn M. (eds.), Die
Pithecanthropus-Schichten auf Java-geologische und paläontologische, ergebnisse der Trinil-Expedition, Verlag von Wilhem Engelmann ed., Leipzig, 54-60.
HILL A., 1983, Hyenas and early hominids, in : Clutton-Brock J. & Grigson C. (eds.),
Animals and Archaeology, 1: Hunters and their Prey, BAR International Series, 163, 87-92.
HILL A., 1989 a, Bone modification by modern spotted hyenas, in : Bonnichsen R. & Sorg
M.H. (eds.), Bone Modification, Center for the Study of the First Americans, University of Maine, Orono, 169-178.
HILL A.P., 1989 b, Problems and prospects of interpreting modified bones from the
archaeological record, in : Bonnichsen R. & Sorg M.H. (eds.), Bone Modification, Center for the Study of the First Americans, University of Maine, Orono.
HILLSON S., 1986, Teeth, collection: Cambridge Manuals in Archaeology, Cambridge
University Press, London, 376 pp. HOOGERWERF A., 1970, Udjung Kulon, the land of the last javan rhinoceros, E.J.Brill ed.,
Leiden, 512 pp. HOOIJER D.A., 1946 a, Some remarks on recent, prehistoric and fossil porcupines from the
Malay Archipelago, Zoologische mededelingen, Leiden, 26, 251-267.

Références bibliographiques.
- 273 - 273
HOOIJER D.A., 1946 b, Prehistoric and fossil rhinoceroses from the Malay archipelago and India, Zoologische mededelingen, Leiden, 26, 1-138.
HOOIJER D.A., 1947, On fossil and prehistoric remains of Tapirus from Java, Sumatra and
China, Zoologische mededelingen, Leiden, 27, 253-299. HOOIJER D.A., 1948 a, Prehistoric teeth of man and of the orang-utan from Central
Sumatra, with notes on the fossil orang-utan from Java and Southern China, Zoologische mededelingen, Leiden, 29, 175-301.
HOOIJER D.A., 1948 b, Pleistocene vertebrates from Celebes II, Testudo margae nov. spec.,
Proceedings koninklijke nederlandse akademie van wetenschappen, Amsterdam, B, 51, (9), 1169-1182.
HOOIJER D.A., 1949, Mammalian evolution in the quaternary of southern and eastern Asia,
Evolution, 3, (2), 125-128. HOOIJER D.A., 1950, The fossil hippopotamidae of Asia, with notes on the recent species,
Zoologische Verhandelingen, Leiden, 8, 149 pp. HOOIJER D.A., 1952, Fossil mammals and the Plio-Pleistocene boundary in Java,
Proceedings koninklijke nederlandse akademie van wetenschappen, Amsterdam, B, 55, (4), 436-443.
HOOIJER D.A., 1954 a, Pleistocene vertebrates from Celebes. X, Testudinata, Proceedings
koninklijke nederlandse akademie van wetenschappen, Amsterdam, B, 57, (4), 486-489. HOOIJER D.A., 1954 b, A pygmy Stegodon from the middle Pleistocene of eastern Java,
Zoologische mededelingen, Leiden, 33, (14), 91-102. HOOIJER D.A., 1954 c, Crocodilian remains from the Pleistocene of Celebes, Copeia, (4),
263-266. HOOIJER D.A., 1955, Fossil Proboscidea from the Malay archipelago and the Punjab,
Zoologische Verhandelingen, Leiden, 28, 163 pp. HOOIJER D.A., 1956 a, Epileptobos gen. Nov. for Leptobos groeneveldtii Dubois from the
middle Pleistocene of Java, Zoologische mededelingen, Leiden, 34, (17), 239-241.

Références bibliographiques.
- 274 - 274
HOOIJER D.A., 1956 b, The valid name of the Banteng: Bibos javanicus, Zoologische mededelingen, Leiden, 34, (14), 223-226.
HOOIJER D.A., 1958, Fossil Bovidae from the Malay archipelago and the Punjab,
Zoologische Verhandelingen, Leiden, 38, 125 pp. HOOIJER D.A., 1960, Quaternary gibbons from the Malay archipelago, Zoologische
Verhandelingen, Leiden, 46, 44 pp. HOOIJER D.A., 1962, Quaternary langurs and macaques from the malay archipelago,
Zoologische Verhandelingen, Leiden, 55, 64 pp. HOOIJER D.A., 1964, New records of mammals from the Middle Pleistocene of Sangiran,
Central Java, Zoologische mededelingen, Leiden, 40, (10), 74-88. HOOIJER D.A., 1969, The stegodon from Timor, Proc. Kon. Akad. Wetensch., Amsterdam,
B, 72, (3), 203-210. HOOIJER D.A., 1972, Stegodon trigonocephalus florensis Hooijer and Stegodon timorensis
Sartono from the Pleistocene of Flores and Timor, Proceedings koninklijke nederlandse akademie van wetenschappen, Amsterdam, B, 75, (1), 12-33.
HOOIJER D.A., 1974, Elephas celebensis (Hooijer) from the Pleistocene of Java,
Zoologische mededelingen, Leiden, 48, (11), 85-93. HOOIJER D.A., 1975, Quaternary mammals west and east of Wallace's line, Modern
Quaternary Research in South-East Asia, 1, 37-46. HOOIJER D.A., 1982, The extinct giant land tortoise and the pygmy stegodont of Indonesia,
Modern Quaternary Research in South-East Asia, 7, 171-176. HOOIJER D.A., 1983, The fauna from Trinil, type locality of Homo erectus : a
reinterpretation, Comment II: Remarks upon the Dubois collection of fossil mammals from Trinil and Kedung Brubus in Java, Geologie en Mijnbouw, 16, (7746), 337-338.
HOOIJER D.A., 1984, The mammalian faunas of Trinil and Kedungbrubus in Java once
more, Modern Quaternary Research in South-East Asia, 8, 95-102.

Références bibliographiques.
- 275 - 275
HOOIJER D.A. & COLBERT E.H., 1951, A note on the Plio-Pleistocene boundary in the Siwalik series of India and in Java, American Journal of Science, 249, 533-538.
HOOIJER D.A. & KURTEN B., 1984, Trinil and Kedung Brubus: the Pithecanthropus-
bearing fossil faunas of Java and their relative age, Annales zoologici Fennici, 21, 135-141.
HORWITZ L.K., 1990, The origin of partially digested bones recovered from archaeological
contexts in Israel, Paleorientologica, 16, (1), 97-106. HORWITZ L.K. & MONCHOT H., 2002, Choice cuts : hominid butchery activities at the
lower paleolithic site of Holon, in : Buitenhuis H., Choyke A.M., Mashkour M. & Al-Shiyab A.H. (eds.), Proceedings of the Vth international symposium on the archaeology of southwestern Asia and adjacent areas-ICAZ, Archaeology of the Near East V, 48-61.
HUFFMAN O. F., 2001, Plio-Pleistocene environmental variety in eastern Java and early
Homo erectus paleoecology - A geological perspective, in : T. Simanjuntak, B. Prasetyo et R. Handini (eds.), Proceedings of the International Colloquium on Sangiran – Sangiran : Man, Culture, and Environment in Pleistocene Times, 231-256.
HUE E., 1907, Musée ostéologique - Etude de la faune quaternaire - Ostéologie des
mammifères, Paris, 8, (1-2), 50 pp. HUXLEY T.H., 1868, On the classification and distribution of the Alectoromorphae and
Heteromorphae, Proceedings of the Zoological Society of London, 294-319. ITIHARA M., SUDIJONO, KADAR D., SHIBASAKI T., KUMAI H., YOSHIKAWA S.,
AZIZ F., SOERADI T. WIKARNO, KADAR A.P., HASIBUAN F. & KAGEMORI Y., 1985, Geology and stratigraphy of the Sangiran area, in : Quaternary geology of the hominid fossil bearing formations in Java, Geological Research and Development Centre, Bandung, 12-43.
JACOB T., 1964, A new hominid skull cap from Pleistocene Sangiran, Anthropologica, 6,
(1), 97-104. JACOB T., 1966, The sixth skullcap of Pithecanthropus erectus, American Journal of
Physical Anthropology, 25, (3), 243-250.

Références bibliographiques.
- 276 - 276
JACOB T., 1967, Recent Pithecanthropus finds in Indonesia, Current Anthropology, 8, (5), 501-504.
JACOB T., après 71, Prehistoric riches from Indonesia, ?, 25-29. JACOB T., 1975 a, Indonesia, in : K.P. Oakley, B.G. Campbell & T.I. Molleson (eds.),
Catalogue of fossil hominids, Part III : Americas, Asia, Australasia, Bristish Museum ed., London, 104-116.
JACOB T., 1975 b, L’Homme de Java, La Recherche, (62), 1027-1032. JACOB T., 1982, Pengembangan ilmu tentang lingkungan dalam penelitian arkeologi, B.
Bioanthrop. Indon., 2, (3), 139-144. JACOB T., 1984, The fossil skull cap from Sambungmachan and its implication to human
evolution, Berkala Bioanthropologi Indonesia, V, 1, 19-27. JACOB T., 2001, Biological aspects of Homo erectus through the time-space continuum, in :
T. Simanjuntak, B. Prasetyo et R. Handini (eds.), Proceedings of the International Colloquium on Sangiran - Sangiran: Man, Culture, and Environment in Pleistocene Times, 19-23.
JACOB T. & CURTIS G.H., 1971, Preliminary potassium/argon of early man in Java,
Contrib. Univ. of Calif. Arch. Res. Facilit., 12, 50. JANENSCH W., 1911, Die Reptilienreste-Crocodilia, in : Selenka L. & Blankenhorn M.
(eds.), Die Pithecanthropus-Schichten auf Java-geologische und paläontologische, ergebnisse der Trinil-Expedition, Verlag von Wilhem Engelmann ed., Leipzig, 61-74.
JANIS C.M., 1984, The significance of fossil ungulate communities as indicators of
vegetation structure and climate, in : Brenchley P.J. (ed.), Fossils and Climate, John Wiley and Sons ed., New York, 85-104.
JATMIKO, 2001, New discoveries of paleolithic tools in some paleolithic sites in Indonesia,
in : T. Simanjuntak, B. Prasetyo & R. Handini (eds.), Proceedings of the International Colloquium on Sangiran, Sangiran : Man, Culture, and Environment in Pleistocene Times, 171-184.

Références bibliographiques.
- 277 - 277
JAUBERT J. & BRUGAL J.P., 1990, Contribution à l’étude du mode de vie au Paléolithique moyen : les chasseurs d’Aurochs de la Borde, in : Jaubert J., Lorblanchet M., Laville H., Slott-Moller R., Turq A. & Brugal J.P., Les chasseurs d’Aurochs de la Borde : un site du Paléolithique moyen (Livernon, Lot), DAF, (27), 127-145.
JOLEAUD L., 1920, Contribution à l'étude des hippopotames fossiles, Bull. Soc. Zool. de
France, 4, (20), 13-26. JOLEAUD L., 1921, Considérations sur le système dentaire des hippopotames, Bull. Soc.
Zool. de France, 46, 18-22. JOHNSON D.L., 1980, Problems in the land vertebrate zoogeography of certain islands and
the swimming powers of elephants, Journal of Biogeography, 7, 383-398. JOHNSON E., 1983, A framework for interpretation in bone technology, in : Carnivores,
human scavengers and predators, Arch. Ass. Dept. of Arch. ed., Univ. of Calgary, 55-93.
KAHLKE H.D., 1986, Biostratigraphical Correlations (Mammals) of Quaternary Continental
Deposits of Europe and the Far East, Quartärpaläontologie, 6, 83-86. KAHLKE R.D. & GAUDZINSKI S., 2005, The blessing of a great flood : differentiation of
mortality patterns in the large mammal record of the Lower Pleistocene fluvial site of Untermassfeld (Germany) and its relevance for the interpretation of faunal assemblages from archaeological sites, Journal of Archaeological Science, (32), 1202-1222.
KATILI J.A., 1975, Volcanism and plate tectonics in the indonesian island arcs,
Tectonophysics, 26, 165-188. KLEIN R.G., WOLF C., FREEMAN L.G. & ALLWARDEN K., 1981, The use of dental
crown heights for constructing age profiles of Red Deer and similar species in archaeological samples, Journal of Archaeological Science, 8, 1-31.
KLEIN R.G. & CRUZ-URIBE K., 1984, The analysis of animal bones from archaeological
sites, University of Chicago Press ed., 266 pp. KOENIGSWALD VON G.H.R., 1933, Beitrag zur Kenntnis der fossilen Wirbeltiere Javas-
I. Teil., Wetenschappelijke Mededeelingen, dienst van den mijnbouw in Nederlansch-Indië, 23, 1-127.

Références bibliographiques.
- 278 - 278
KOENIGSWALD VON G.H.R., 1934 a, Zur Stratigraphie des javanischen Pleistocän, De Ingenieur in Nederlandsch Indië, 1 (4), 185-201.
KOENIGSWALD VON G.H.R., 1934 b, Die Spezialisation des Incisivengebisses bei den
javanischen Hippopotamidae, Proceedings koninklijke nederlandse akademie van wetenschappen, B, 37, (9), 653-659.
KOENIGSWALD VON G.H.R., 1935, Die fossilen Säugetierfaunen Javas, Proceedings
koninklijke nederlandse akademie van wetenschappen, B, 38, 188-198. KOENIGSWALD VON G.H.R., 1936, Early Palaeolithic stone implements from Java,
Bulletin of the Raffles Museum Singapore, 1, 52-60. KOENIGSWALD VON G.H.R., 1939, Das Pleistocän Javas, Quartär, 28-53. KOENIGSWALD VON G.H.R., 1940, Neue Pithecanthropus-funde 1936-1938 - Ein
Beitrag zur Kenntnis der Praehominiden, Wetenschappelijke Mededeelingen, dienst van den mijnbouw in Nederlansch-Indië, 28, 235 pp.
KOENIGSWALD VON G.H.R., 1949, Vertebrate stratigraphy of the Neogene and
Quaternary, in : Bemmelen van, The Geology of Indonesia and Adjacent Archipelagos, Martinus Nijhoff ed., Den Haag, 1, 91-94.
KOENIGSWALD VON G.H.R., 1956, Remarks on the correlation of mammalian faunas of
Java and India and the Plio-Pleistocene boundary, Proceedings koninklijke nederlandse akademie van wetenschappen, Amsterdam, B, 59, (3), 204-210.
KOENIGSWALD VON G.H.R., 1964, Potassium-Argon dates and early man : Trinil,
Report of VI the I.N.Q.U.A. Congress, Warsaw, 325-327. KOENIGSWALD VON G.H.R. & GOSH A.K., 1973, Stone implements from the Trinil
beds of Sangiran, Central Java, Kon. Ned. Akad. Wetensch., Proceedings koninklijke nederlandse akademie van wetenschappen, series B, 76, (1), 1-34.
KOESHARDJONO, 1988, Kehidupan ekonomi masa lampau berdasakan data arkeologi,
Jilid II, Analisis hasil penelitian arkeologi II, Departemen pendidikan dan kedudayaan, Jakarta, 51-60, non publié.
KONINCK de R., 1994, L’Asie du sud-est, Masson ed., Paris, 317 pp.

Références bibliographiques.
- 279 - 279
KORTH W.W., 1979, Taphonomy of microvertebrate fossil assemblages, Annals of the Carnegie Museum, Pittsburgh, 48, 235-285.
KREUTZER L.A., 1992, Bison and deer bone mineral densities : comparisons and
implications for interpretation of archaeological faunas, Journal of Archaeological Science, 19, 271-294.
KURTEN B., 1956, The status and affinities of Hyaena sinensis Owen and Hyaena ultima
Matsumoto, American Museum Novitates, 1764, 1-48. KURTEN B., 1959, On the longevity of mammalian species in the Tertiary, Comment. Biol.
Soc. Sci. Fennica, 21, (4), 1-14. KURTEN B., 1960, An attempted parallellization of the quaternary mammalian faunas of
China and Europe, Commentationes Biologicae, societas scientarum Fennica, Helsinki, 23, (8), 1-12.
KURTEN B., 1968, Pleistocene mammals of Europe, Weidenfeld & Nicolson ed., London, i-
viii, 1-317. KURTEN B., 1972, The “half-life” concept in evolution illustrated from various mammalian
groups, in : Bishop, W.W. & Miller J.A. (eds.), Calibration of hominoid evolution, Scottish Academic Press, Edinburgh, 187-194.
LACOMBAT F., 1999, Le rhinocéros du Pléistocène moyen d’Isernia la Pineta (Molise,
Italie) - Systématique, Taphonomie, Palethnographie, Mémoire de DEA, M.N.H.N., I.P.H., Paris, 118 pp.
LANGBROEK M. & ROEBROEKS W., 2000, Extraterrestrial evidence on the age of the
hominids from Java, Journal of Human Evolution, 38, 595-600. LARICK R., CIOCHON R.L., ZAIM Y., SUDIJONO, SUMINTO, RIZAL Y., AIZ F.,
REAGAN M. & HEIZLER M., 2001, Early Pleistocene 40Ar/39Ar ages for Bapang formation hominins, Central Jawa, Indonesia, P.N.A.S., 98, (9), 4866-4871.

Références bibliographiques.
- 280 - 280
LEFEVRE D., RAYNAL J.-P., VERNET G., PILEYRE T., PIPERNO M., SANZELLE S., FAIN J., MIALLER D. & MONTRET M., 1994, Sédimentation, volcanisme et présence humaine dans le basin de Venosa (Basilicata, Italie) au Pléistocène Moyen : exemple du site de Notarchirico, Bulletin de la Société Préhistorique de France, 91, 103-111.
LEGENDRE L. & LEGENDRE P., 1984, Ecologie numérique - La structure des données
écologiques, Masson ed., Paris, 335 pp. LEGENDRE S., 1986, Analysis of mammalian communities from the late Eocene and
Oligocene of Southern France, Palaeovertabrata, Montpellier, 16, (4), 191-212. LEGENDRE S., 1989, Les communautés de mammifères du Paléogène (Eocène supérieur et
oligocène) d’Europe occidentale : structures, milieux et évolution, Münchner Geowiss. Abh., (A), 16, 1-110.
LEGENDRE S. & SUDRE J., 1984, La communauté de Mammifères de Robiac (Eocène
supérieur), 10e R.A.S.T.-Bordeaux, Société Géologique de France ed., Paris, 352 pp. LEINDERS J.J.M., SONDAAR P.Y. & DE VOS J., 1985, The age of hominid-bearing
deposits of Java : state of the art, Geologie en Minjbouw, 64, 167-173. LEYRAT J.C., 1982, Contribution à l'étude des rhinocéros asiatiques, Thèse, Ecole nationale
vétérinaire d'Alfort, 91 pp. LHOMME V., CONNET N., BEMILLI C. & CHAUSSE C., 2000, Essai d’interprétation
du site paléolithique inférieur de Soucy 1 (Yonne), Gallia Préhistoire, 42, p1-44. LIMONDIN-LOZOUET N., 2002, Les mollusques continentaux, in : Miskovsky J.C. (ed.)
Géologie de la Préhistoire : méthodes, techniques, applications, Maison de la géologie éd., Paris, 773-784.
LIZON-SUREAU B., 1979, Essai de reconstitution du paléoenvironnement du
pithécanthrope de Java, du Pléistocène moyen, dans la région de Sangiran (Java central), par les méthodes d'analyse en sédimentologie classique et moderne, Thèse de doctorat, Université Paris VI, 109 pp.

Références bibliographiques.
- 281 - 281
LUMLEY H. de & SONAKIA A., 1985, Contexte stratigraphique et archéologique de l’Homme de la Narmada, Hathnora, Madhya Pradesh, Inde, L’Anthropologie, Paris, 89, (1), 3-12.
LYELL C., 1830, Principles of geology I, John Murray ed., London. LYELL C., 1832, Principles of geology II, John Murray ed., London. LYMAN R.L., 1987, Zooarchaeology and taphonomy: a general consideration, J. of
Ethnobiology, 7, (1), 93-117. LYMAN R.L., 1989, Taphonomy of cervids killed by the May 18, 1980, volcanic eruption of
Mount St. Helens, Washington, USA, in : Bonnichsen R. & Sorg H. (eds.), Bone modification, University of Maine, Center for the study of the first Americans, 149-167.
LYMAN R.L., 1994, Vertebrate taphonomy, Cambridge University Press. MAC NAB B.K., 1971, On the ecological significance of the Bergmann’s rule, Ecology, 52
(5), 845-854. MAAREL VAN DER F.H., 1932, Contribution of the knowledge of the fossil mammalian
fauna of Java, Wetenschappelijke Mededeelingen, 15, 230 pp. MAGLIO V.J., 1973, Origin and evolution of elephantidae, Transactions of the American
Philosophical Society, 63, (3), 1-149. MAREAN C.W. & SPENCER L.M., 1991, Impact of carnivore ravaging on
zooarchaeological measures of element abundance, American Antiquity, 56, (4), 645-658.
MATSU’URA S., 1982, A chronological framing for the Sangiran hominids, Bulletin of the
National Science Museum, Tokyo, 8, 53 pp. MAYR E., 1944, Wallace's line in the light of recent zoogeographic studies, Quaterly Review
of Biology, 19, 1-14. MAZZA P., 1991, Interrelations between Pleistocene hippopotami of Europe and Africa,
Bolletino della Societa Paleontologica Italiana, 30, (2), 153-186.

Références bibliographiques.
- 282 - 282
MILLER S.J., 1994, Biological agents of bone modification, in : Patou-Mathis M. (ed), Taphonomie/bone modification - Outillage peu élaboré en os et bois de Cervidés IV, Artefacts, 9, 67-75.
MOIGNE A.-M. & BARSKY D.R., 1999, Large mammal assemblages from Lower
Palaeolithic sites in France : la Caune de l’Arago, Terra Amata, Orgnac 3 et Cagny l’Epinette, in : The Role of Early Humans in the Accumulation of European Lower and Middle Palaeolithic Bone Assemblages, Römisch-Germanisches Zentralmuseum, 42, 219-235.
MOIGNE A.M., DUE AWE R., SEMAH F. & SEMAH A.M., 2004, The cervid from
Ngebung site (“Kabuh series”, Sangiran dome, Central Java) and their biostratigraphical significance, Modern Quaternary Research in South-East Asia, 18, 31-44.
MONCHOT H., 1996, Les Petits Bovidés (genre Ovis, Hemitragus, Capra et Rupicapra) de
la Caune de l’Arago (Tautavel, Pyrénées-Orientales), Thèse de doctorat, Université Aix-Marseille II.
MONCHOT H., 1998, Acquisition et exploitation du gibier à la fin du Paléolithique
supérieur à la Baume de Goulon (Salernes, Var), Géologie Méditerranéenne, XXV, (2), 57-73.
MONCHOT H. & HORWITZ L.K., 2002, Représentation squelettique au Paléolithique
inférieur, le site d’Holon (Israël), Paléorient, 28, (2), 71-86. MONCHOT H. & HORWITZ L.K., (sous presse), Faunal taphonomie, in : M. Chazan &
L.K. Horwitz (eds.), The Lower Paleolithic Site of Holon, Israel, Peabody Museum, Harvard University.
MORRISON D. & WHITRIDGE P., 1997, Estimating the age and sexe of Caribou from
mandibular measurements, Journal of Archaeological Science, 24, 1093-1106. MOVIUS H.L., 1944, Early Man and Pleistocene Stratigraphy in Southern and Eastern Asia,
Papers of the Peabody Museum of American archaeology and ethnology, Harvard University, 19, (3), 125 pp.
NEUMANN VAN PADANG M., 1951, Indonesia, XIII, Vulcani ed., 271 pp.

Références bibliographiques.
- 283 - 283
NINKOVICH D. & BURCKLE L.H., 1978, Absolute age of the base of the hominid-bearing beds in Eastern Java, Nature, 275, 306-308.
NISHIMURA S., SASAJIMA S., THIO K.H. & HEHUWAT F., 1978, The second report
of fission-track dating in the Sunda arc, Proc. 2nd Working Group meeting (Biostratigraphic datum-planes of the Pacific Neogene), 163-164.
NISHIMURA S., IKEDA T. & YOKOYAMA T., 1981, A fundamental investigation on the
fission-track method, Report for the Grant-in-Aid for Scientific Research, 5-11. NOE-NYGAARD N., 1977, Butchering and marrow fracturing as a taphonomic factor in
archaeological deposits, Paleobiology, 3, (2), 218-237. OBRADOVICH J.D. & NAESER C.W., 1981, Indonesia, Age of Homo erectus from Java,
Geological Survey Research, Geol. Survey Prof. Paper, 1275, 285-286. OLSON E.C., 1980, Taphonomy : its history and role in community evolution, in :
Behrensmeyer A.K. & Hill A.P. (eds.), Fossils in the making - Vertebrate taphonomy and paleoecology, University of Chicago Press ed., 5-19.
OPPENOORTH F., 1911, Arbeitsbericht über die Ausgrabungen, I, Teil, Die Arbeiten des
Jahres 1907 bis August, in : Selenka & Blanckenhorn (eds.), Die Pithecanthropus- Schichten auf Java, Geologische und Paläontologische Ergebnisse der Trinil-Expedition (1907 und 1908), Leipzig, 26-38.
ORCHISTON D.W. & SIESSER W.G., 1982, Chronostratigraphy of the Plio-Pleistocene
fossil hominids of Java, Modern Quaternary Research in South-East Asia, 7, 131-149. OSBORN F.H., 1888, The nomenclature of the mammalian molar cups, American
Naturalist., 22, (262), 926-928. OSBORN F.H., 1904, Paleontological evidence for the original tritubercular theorie, The
American Journal of Science, série 4, 17, (100), 321-323. OSBORN F.H., 1942, Proboscidea, The American Museum Press ed., 2, 805-1599. OTTE M. & PATOU-MATHIS M., 1992, Comportements de subsistance au Paléolithique
moyen en Europe, Paléo, (4), 29-34.

Références bibliographiques.
- 284 - 284
PALES L. & LAMBERT C., 1971, Atlas ostéologique des mammifères du Quaternaire - Les membres : herbivores, carnivores, C.N.R.S. ed., 1.
PALES L. & GARCIA M.A., 1981, Atlas ostéologique des mammifères du Quaternaire -
Tête, rachis, ceintures scapulaire et pelvienne : herbivores, carnivores, homme, C.N.R.S. ed., 2.
PATOU-MATHIS M., 1994, Actions anthropiques sur les assemblages osseux et sur les
ossements du Paléolithique : état des recherches, in : Patou-Mathis M. (ed), Taphonomie/bone modification - Outillage peu élaboré en os et bois de Cervidés IV, Artefacts, 9, 133-142.
PATOU-MATHIS M., 1997, Etude de la fracturation des os longs de renne - Méthode et
résultats : le niveau magdalénien « archaïque » de la grotte de Tournal à Bize (Aude), in : Hannus L.A., Rossum L. & Winham P. (eds.), Proceedings of the 1993 bone modification conference, Hot Springs, South Dakota, Archaeology laboratory Augustana College ed., 1, 195-219.
PATOU-MATHIS M., 1999, A new Middle Palaeolithic site in Alsace : Mutzig 1 (Bas
Rhin), Subsistence behaviour, in : The Role of Early Humans in the Accumulation of European Lower and Middle Palaeolithic Bone Assemblages, Römisch-Germanisches Zentralmuseum, 42, 325-341.
PAUTRET-HOMERVILLE C., 2001, Etude de l’hyène des cavernes (Crocuta crocuta
spelaea) en Ardèche - Origine, évolution et extinction d’un carnivore pléistocène, Mémoire de DEA, M.N.H.N., I.P.H., Paris, 90 pp.
PELSENEER P., 1904, La « Ligne de Weber », limite zoologique de l’Asie et de l’Australie,
Bull. Classe Sci. Acad. Roy. Belgique, 1904, 1001-1022. PERETTO C., 1996, I reperti paleontologici de giacimento paleolitico di Isernia la Pineta.
Isernia : Cosmo Lannone ed., 493 pp. PILLANS B., CHAPPELL J. & NAISH T.R., 1998, A review of the Milankovitch climatic
beat : template for Plio-Pleistocene sea-level changes and sequence stratigraphy, Sedimentary Geology, 122, 5-21.

Références bibliographiques.
- 285 - 285
POPE G.G., 1984, The Antiquity and Paleoenvironment of the asian hominidae, in : R.O.Whyte (ed.), The evolution of the east asian environment, University of Hong Kong, 2, 822-847.
POPE G.G., 1989, Bamboo and human evolution, Natural History, 98, 48-57. POPLIN F., 1976, Remarques théoriques et pratiques sur les unités utilisées dans les études
d’ostéologie quantitative, particulièrement en archéologie préhistorique, Thèmes spécialisés, Actes du IXème congrès de l’U.I.S.P.P., Nice 13-18 septembre 1976, 124-141.
POTTS R., 1983, Foraging for faunal resources by early hominids at Olduvai Gorge,
Tanzania, in : Clutton-Brock J. & Grigson C. (eds.), Animals and Archaeology: 1.Hunters and their prey, BAR International Series, 163, 51-62.
PRENTICE M.L. & DENTON G.H., 1988, Deep-sea oxygen isotope record, the global ice
sheet system, and hominid evolution, in : Grine F. (ed.), The evolutionary history of the robust Australopithecines, de Gruyter ed., New York, 383-403.
QUIMBY D.C. & GAAB J.E., 1957, Mandibular dentition as an age indicator in Rocky
Mountain Elk, The Journal of Wildlife Management, 21, (4), 435-451. RADEN SALEH, 1867, Over fossiele beenderen van den Pandan, Natuurwetenschappelijk
Tijdschrift voor Nederlandsch Indie, 29, 422-423. REIS K.R. & GARONG A.M., 2001, Late Quaternary terrestrial vertebrates from Palawan
Island, Philippines, PALAEO, 171, 409-421. RIGHTMIRE G.P., 1987, L'évolution des premiers hominidés en Asie du Sud-Est,
L'Anthropologie, 91, n°2, p455-466. SAEGUSA H., 1996, Stegodontidae : evolutionary relationships, in : Shoshani J. et Tassy P.
(eds.), The Proboscidea. Evolution and Palaeoecology of Elephants and their relatives, Oxford University Press ed., Oxford, 178-190.
SAINT-MARC P., PALTRINIERI F. & SITUMORANG B., 1977, Le Cénozoïque
d’Indonésie Occidentale, Bulletin de la Société Géologique de France, 7ème série, XIX, 1, 125-133.

Références bibliographiques.
- 286 - 286
SALEKI H., 1997, Apport d’une intercomparaison de méthodes nucléaires (230Th/234U, ESR et 40Ar/39Ar) à la datation de couches fossilifères pléistocènes dans le dôme de Sangiran (Java, Indonésie), Thèse de doctorat, M.N.H.N., I.P.H., Paris.
SARTONO S., 1964 a, On a new find of another Pithecanthropus skull: an announcement,
Bulletin of the Geological Survey of Indonesia, 1, (1), 1-5. SARTONO S., 1964 b, Stratigraphy and sedimentation of the Easternmost part of Gunung
Sewu (East Jawa), Direktorat Geologi Bandung, Publik. Tek., Geologi, (1). SARTONO S., 1971, Observations on a new skull of Pithecanthropus erectus
(Pithecanthrope VIII) from Sangiran, Central Java, Proc. Acad. Sci. AMST, B, 74, 185-194.
SARTONO S., 1991, Homo (Pithecanthropus) erectus : le débat sans fin, L'Anthropologie,
95, (1), 123-136. SCHEPARTZ L.A., STOUTAMIRE S. & BEKKEN D.A., 2005, Stegodon orientalis from
Panxian Dadong, a Middle Pleistocene archaeological site in Guizhou, South China: taphonomy, population structure and evidence for human interactions, Quaternary International, 126-128, 271-282.
SCHMID E., 1972, Atlas of animal bones for Prehistorians, Archaeologists, and Quaternary
Geologists, Elsevier Publishing Company, 152 pp. SCHWARZ E., 1937, Die fossilen Antilopen von Oldoway (Wissenschaftliche Ergebnisse
der Oldoway- Expedition, 1913, n.F., H.4), Borntraeger ed., Leipzig, 1-90. SCOTT W.B., 1892, Evolution of the premolar teeth in the mammals, Proc. Acad. Nat. Sci.,
44, 406-444. SCOTT K., 1989, Mammoth bones modified by man : evidence from La Cotte de St Brelade,
Jersey, Channel Islands, in : Bonnichsen R. & Sorg H. (eds.), Bone modification, University of Maine, Center for the study of the first Americans, 335-346.
SELENKA L. & BLANKENHORN M., 1911, Die Pithecanthropus-Schichten auf Java-
geologische und paläontologische, ergebnisse der Trinil-Expedition, Verlag von Wilhem Engelmann ed., Leipzig, 225 pp.

Références bibliographiques.
- 287 - 287
SEMAH A.M., 1986, Le milieu naturel lors du premier peuplement de Java, Thèse de doctorat d’Etat, Université de Provence Aix-Marseille I.
SEMAH A.M., 1993, Le Pithécanthrope et la forêt, Les Dossiers d’Archéologie, 184, 50-55. SEMAH A.M., SEMAH F. & DJUBIANTONO T., 1993, Les grands sites fossilifères de
Java, Les Dossiers d’Archéologie, 184, 20-25. SEMAH F., 1982, Pliocene and Pleistocene geomagnetic reversals recorded in the Gemolong
and Sangiran domes (Central Java), Modern Quaternary Research in South-East Asia, 7, 151-164.
SEMAH F., 1984, Stratigraphie et paléomagnétisme du Pliocène supérieur et du Pléistocène
de l’île de Java - Indonésie, Thèse de doctorat, Université de Provence Aix-Marseille I. SEMAH F., 1986, Le peuplement ancien de Java; ébauche d'un cadre chronologique,
l'Anthropologie, 90, (3), 359-400. SEMAH F., SARTONO S., ZAIM Y & DJUBIANTONO T., 1980, Premiers résultats
concernant l’étude paléomagnétique de la partie ouest du dôme de Sangiran (Java, Indonésie), Compte-Rendus de l’Académie des Sciences, Paris, B, 290, 477-480.
SEMAH F., SEMAH A.M., SARTONO S., ZAIM Y. & DJUBIANTONO T., 1981-1982,
L'âge et l'environnement des Homo erectus de Java : nouveaux résultats paléomagnétiques et palynologiques, L'Anthropologie, 85-86, (3), 509-516.
SEMAH F., PURWASITO A. & DJUBIANTONO T., 1993 a, Un fascinant chaînon
manquant, Les Dossiers d’Archéologie, 184, 4-11. SEMAH F., SEMAH A.M., DJUBIANTONO T. & DRIWANTORO D., 1993 b, La
fouille du site de Ngebung, Les Dossiers d’Archéologie, 184, 68-74. SEMAH F., SALEKI H., FALGUERES C., FERAUD G. & DJUBIANTONO T., 2000,
Did Early Man reach during the Late Pliocene ?, Journal of Archaeological Science, 27, 763-769.

Références bibliographiques.
- 288 - 288
SEMAH F., SEMAH A.M. & DJUBIANTONO T., 2001 a, From the shoreline to the slopes of the volcanoes : the long Pithecanthropus trek, in : T. Simanjuntak, B. Prasetyo et R. Handini (eds.), Proceedings of the International Colloquium on Sangiran – Sangiran : Man, Culture, and Environment in Pleistocene Times, 195-218.
SEMAH F., FALGUERES C., GRIMAUD-HERVE D. & SEMAH A.M., 2001 b, Origine
des peuplements et chronologie des cultures paléolithiques dans le sud-est asiatique, Colloque International de la Fondation Singer Polignac, Paris, 3-5 juin 1998.
SEMAH F., SEMAH A.M. & SIMANJUNTAK T., 2002, More Than a Million Years of
Human Occupation in Insular Southeast Asia - The Early Archaeology of Eastern and Central Java, in : Julio Mercader (ed.), Under the Canopy - The Archaeology of Tropical Rain Forests, 161-190.
SERRE F., 1987, Contribution à l’étude des grands mammifères du site pléistocène moyen
de Terra Amata (Nice, Alpes-Maritimes) - Paléontologie, taphonomie et paléœthnologie, Mémoire de D.E.A., M.N.H.N., I.P.H., Paris, 237 pp.
SEVIN O., 1993, L’Indonésie, coll. Que sais-je, P.U.F., (801), 128 pp. SHIMIZU Y., MUBRATO B., SIAGIAN H. & UNTUNG M., 1985, A paleomagnetic
study in the Sangiran area, in : Watanabe N. et Kadar D. (eds.), Quaternary Geology of the Hominid Fossil Bearing Formations in Java, Special publication of Geological Research and Development Centre, Bandung, (4), 274-307.
SHIPMAN P., 1981 a, Life history of a fossil - An introduction to Taphonomy and
Paleoecology, Harvard University Press ed., 222 pp. SHIPMAN P., 1981 b, Applications of scanning electron microscopy to tahonomic problems,
in : Cantwell A.M., Griffin J.B. & Rotschid N.A. (eds.), The Research Potential of Anthropological Museum Collections, New York Academy of Sciences, 357-385.
SHIPMAN P., 1983, Early Hominid lifestyle : hunting and gathering, or foraging and
scavenging ?, in : Clutton-Brock J. & Grigson C. (eds.), Animals and Archaeology: 1.Hunters and their prey, BAR International Series, 163, 31-49.
SHOOP C.R. & RUCKDESCHEL C.A., 1990, Alligators as Predators on Terrestrial
Mammals, The American Midland Naturalist, 124, 407-412.

Références bibliographiques.
- 289 - 289
SHOSHANI J. & TASSY P., 1996, The Proboscidea, Evolution and Palaeoecology of Elephants and Their Relatives, Oxford Science Publications, 472 pp.
SIMANJUNTAK H.T., 2001, Sangiran site: problems and the balance of research, in : T.
Simanjuntak, B. Prasetyo et R. Handini (eds.), Proceedings of the International Colloquium on Sangiran - Sangiran: Man, Culture, and Environment in Pleistocene Times, 3-15.
SIMANJUNTAK H.T. & SEMAH F., 1996, A new insight into the Sangiran flake industry,
Bulletin of the Indo-Pacific Prehistory Association, 14, 22-26. SIMONET P. & MONNIER J.-L., 1991, Approche paléo-écologique et taphonomique de la
grande faune du gisement moustérien du Mont-Dol (Ille et Vilaine, France), Quaternaire, 2, 5-15.
SIMPSON G.G., 1925, Mesozoic Mammalia III: Preliminary comparison of Jurassic
Mammals except Multituberculates, The American Journal of Science, série 5, 10, (60), 559-569.
SIMPSON G.G., 1965, The Geography of Evolution, Chilton Books ed., New York, 249 pp. SIMPSON G.G., 1977, Too many lines : the limits of the oriental and Australian
zoogeographic regions, Proceedings of the American Philosophical Society, 121, n° 2, p107-120.
SOBOLIK K.D. & STEELE D.G., 1996, A turtle atlas to facilitate archaeological
identifications, Mammoth Site of Hot Springs (ed.), U.S.A., 117 pp. SOEJONO R.P., 1982, New data on the paleolithic industry in Indonesia, 1er congrès
international de paléontologie humaine, Nice, colloque l’Homo erectus et la place de l’homme de Tautavel parmi les hominidés fossiles, II, 578-592.
SONDAAR P.Y., 1977, Insularity and its effect on mammal evolution, in : M.K. Hecht, P.C.
Goody & B.M. Hecht (eds.), Major patterns in vertebrate evolution, Plenum Press ed., New York, 671-707.
SONDAAR P.Y., 1981, The Geochelone faunas of the indonesian archipelago and their
palaeogeographical and biostratigraphical significance, Modern Quaternary Research in South-East Asia, 6, 111-120.

Références bibliographiques.
- 290 - 290
SONDAAR P.Y., 1984, Faunal evolution and the mammalian biostratigraphy of Java, Courier Forshungs Institut Senckenberg., 69, 219-235.
SONDAAR P.Y., DE VOS J. & LEINDERS J.J.M., 1983, The fauna from Trinil, type
locality of Homo erectus : a reinterpretation, Reply facts and fiction around the fossil mammals of Java, Geologie en Mijnbouw, 16, (7746), 339-343.
SONDAAR P.Y., VAN DEN BERGH G.D., MUBROTO B., AZIZ F., DE VOS J. &
BATU U.L., 1994, Middle Pleistocene faunal turnover and colonization of Flores (Indonesia) by Homo erectus, Compte-Rendus de l’Académie des Sciences, série II, (319), 1255-1262.
SNEED E.D. & FOLK R.L., 1958, Pebbles in the lower Colorado River, Texas, A study in
particle morphogenesis, Journal of Geology, 66, 114-150. STALLIBRASS S., 1982, The use of cement layers for absolute ageing of mammalian teeth:
a selective review of the literature, with suggestions for further studies and alternative applications, in : Wilson B., Grigson C. & Paynes S. (eds.), Ageing and sexing animal bones from archaeological sites, BAR Series, 109, 109-126.
STEHN C.E. & UMBGROVE J.H.F., 1929, Bijdrage tot de geologie der vlakte van
Bandoeng, Tijdschr. kon. Ned. aardr. Gen., 2nd serie, XLVI, 301-314. STINER M.C., 1990, The use of mortality patterns in archaeological studies of Hominid
predatory adaptation, Journal of Anthropological Archaeology, 9, 305-351. STINER M.C., 1994, Honor among thieves - A zooarchaeological study of Neandertal
ecology, Princeton University Press ed., Princeton, 447 pp. STORM P., 2001, Life and death of Homo erectus in Australasia: an environmental approach
to the fate of a paleospecies, in : Sémah F., Falguères C., Grimaud-Hervé D. & Sémah A.M. (eds.), Origine des peuplements et chronologie des cultures paléolithiques dans le sud-est asiatique, Colloque International de la Fondation Singer Polignac, 3-5 juin 1998, 279-298.
STORM P., AZIZ F., DE VOS J., DIKDIK KOSASIH, SINUNG BASKORO,
NGALIMAN, VAN DEN HOEK OSTENDE L.W., sous presse, Late Pleistocene Homo sapiens in a tropical rainforest fauna in East Java, Journal of Human Evolution, 10 pp.

Références bibliographiques.
- 291 - 291
STREMME H., 1911, Die Saügetiere mit Ausnahme der Proboscidier, in : Selenka L. & Blankenhorn M. (eds), Die Pithecanthropus-Schichten auf Java-geologische und paläontologische, ergebnisse der Trinil-Expedition, Verlag von Wilhem Engelmann ed., Leipzig, 82-150.
SUTCLIFF A.J., 1970, Spotted hyaena : crusher, gnawer, digestor and collector of bones,
Nature, 227, 1110-1113. SUZUKI M., WIKARNO, BUDIDANTOSO, SAEFUDIN I. & ITIHARA M., 1985,
Fission track ages of pumice, tuff layers and javites of hominid fossils bearing formation in Sangiran area, Central Java, in : Watanabe N. & Kadar D. (eds.), Quaternary Geology of the Hominid Fossil Bearing Formations in Java, Special publication of Geological Research and Development Centre, Bandung, (4), 309-357.
SWISHER C.C., CURTIS G.H., JACOB T., GETTY A.G., SUPRIJO A. &
WIDIASMORO, 1994, Age of the earliest known hominids in Java, Indonesia, Science, 263, 1118-1121.
TAGLIACOZZO A., CASSOLI P.F., CURCI A. & FIORE I., 1999, Taphonomical
analysis of the bone remains of level Alpha, in : Piperno M. (ed.), Notarchirico, Un sito del Pleistocene medioiniziale nel lacino di Venosa, Osanna, Venosa, 517-520.
TASSY P., 1982, Les principales dichotomies dans l’histoire des proboscidea (mammalia) :
une approche phylogénétique, Géobios, Mem. Spec. 6, 225-245. TER HAAR I.C., 1929, Geological guide to the locality of fossil vertebrates in the Kali
Glagah-Boemiajoe district, IVth Pacific Science Congress, 1-15. TER HAAR I.C., 1934, Homo-Soloënsis, De Ingenieur in Nederlansch-Indië, 4, 52-60. TERRA H. de & PATERSON T.T., 1939, Studies on the ice-age in India and associated
human cultures, Carnegie Magazine, (493), 1-354. THEUNISSEN B., DE VOS J., SONDAAR P.Y. & AZIZ F., 1990, The establishment of a
chronological framework for the hominid-bearing deposits of Java, a historical survey, in : Laporte (ed.), Establishment of a Geologic Framework for Palaeoanthropology, Geological Society of America Special Paper, Boulder, Colorado, 242, 39-53.

Références bibliographiques.
- 292 - 292
TOBIAS P., 1966, A re-examination of the Kedung Brubus mandible, Zoologische Mededelingen, Leiden, 41, 307-320.
TODD L.C. & RAPSON D.J., 1999, Formational analysis of bison bonebeds and
interpretation of Paleoindian subsistence, in : Brugal J.P., David F., Enloe J.G. & Jaubert J. (eds.), Le bison: gibier et moyen de subsistance des homes du Paléolithique aux Paléoindiens des Grandes Plaines, Actes du colloque international Toulouse 6-10 juin 1995, A.P.D.C.A. ed., 479-499.
TONG H., 2000, Les Rhinocéros des sites à fossiles humains de Chine, L'Anthropologie,
104, 523-529. TOUGARD C., 1998, Les faunes de grands mammifères du Pléistocène moyen terminal de
Thaïlande dans leur cadre phylogénétique, paléoécologique et biochronologique, Thèse de doctorat, Université Montpellier II, 175 pp.
TURNER A., 1984, Identifying bone-accumulating agents, in : Hall Mandal (ed.), Frontiers :
Southern African Archaeology today, BAR, 207, 334-339. TURNER A. & ANTON M., 1996, The Giant Hyaena, Pachycrocuta brevirostris
(Mammalia, Carnivora, Hyaenidae), GEOBIOS, Villeurbanne, 29, (4), 455-468. VALLI A., 2001, Le Gisement Villafranchien moyen de St Vallier (Drôme) : nouvelles
données paléontologiques (cervidae, bovinae) et taphonomiques, Documents des laboratoires de géologie, Lyon, (153), 274 pp.
VALVERDE J.A., 1964, Remarques sur la structure et l’évolution des communautés de
Vertébrés terrestres, I : Structure d’une communauté, II : Rapports entre prédateurs et proies, La Terre et la vie, Paris, 111, 121-153.
VEEVERS-CARTER W., 1989, Land mammals of Indonesia, Pt Intermasa ed., Indonesia,
116 pp. VILLA P., 1990, Torralba and Aridos : elephant exploitation in middle Plesitocene Spain,
Journal of Human Evolution, 19, 299-309. VILLA P. & MAHIEU E., 1991, Breakage patterns of human long bones, Journal of Human
Evolution, 21, 27-48.

Références bibliographiques.
- 293 - 293
VILLA P., SOTO E., SANTONJA M., PEREZ-GONZALEZ A., MORA R., PARCERISAS J. & SESE C., 2005, New data from Ambrona : closing the hunting versus scavenging debate, Quaternay International, 126-128, 223-250.
VOORHIES M.R., 1969, Taphonomy and population dynamics of an early Pliocene
vertebrate fauna, Knox County, Nebraska - Contributions to geology, University of Wyoming, Special Paper, 1, 69 pp.
VORIS H.K., 2000, Maps of Pleistocene Sea Levels in South East Asia: Shorelines, River
Systems, Time Durations, Journal of Biogeography, 27, 1153-1168. VOS J. de, 1983, The Pongo faunas from Java and Sumatra and their significance for
biostratigraphical and paleo-ecological interpretations, Proceedings koninklijke nederlandse akademie van wetenschappen, 4, (86), 417-425.
VOS J. de, 1984, Reconsideration of Pleistocene cave faunas from South China and their
relation to the faunas from Java, Courier Forshungs Institut Senckenberg, 69, 259-266. VOS J. de, 1985, Faunal Stratigraphy and Correlation of the Indonesian Hominid Sites, in :
Delson E. (ed.), Ancestors, The Hard Evidence, Liss ed., New York, 215-220. VOS J. de, 1989, The environment of Homo erectus from Trinil H.K., Proceedings of the
second International Congress of Human Palaeontology, 225-229. VOS J. de, 1993, The migration of Homo erectus and Homo sapiens in South-East Asia and
the Indonesian archipelago, in : Bower J.R.F. et Sartono S. (ed.), Evolution and ecology of Homo erectus, Pithecantropus, Centennial 1893-1993 Congress, 1, 238-259.
VOS J. de & SONDAAR P.Y., 1982, The importance of the « Dubois collection »
reconsidered, Modern Quaternary Research in South-East Asia, 7, 35-63. VOS J. de, SARTONO S., HARDJASASMITA S. & SONDAAR P.Y., 1982, The fauna
from Trinil, type locality of Homo erectus - A reinterpretation , Geologie en Minjouw, 16, (7746), 207-216.
VOS J. de & AZIZ F., 1989, The excavations by Dubois (1891-1900), Selenka (1906-1908),
and the Geological Survey by the Indonesian-Japanese Team (1976-1977) at Trinil (Java, Indonésie), Journal of the anthropological society of Nippon, 97, (3), 407-420.

Références bibliographiques.
- 294 - 294
VOS J. de, SONDAAR P.Y., VAN DEN BERGH G.D. & AZIZ F., 1994, The Homo bearing deposits of Java and its ecological context, Courier Forshungs Institut Senckenberg, 171, 129-140.
WALLACE A.R., 1860, On the zoological geography of the Malay Archipelago, Zooogical
Journalof the Linnaean Society of London, 4, 172-184. WANPO H., CIOCHON R., YUMIN G., LARICK R., QIREN F., SCHWAREZ H.,
YONGE C., DE VOS J. & RINK W., 1995, Early Homo and associated artefacts from Asia, Nature, 378, 275-278.
WATANABE N. & KADAR D., 1985, Quaternary geology of the hominid bearing
formations in Java, Special publication of Geological Research and Development Centre, (4), 378 pp.
WEBER M., 1902, Der Indo-australische Archipel und die Geschichte seiner Tierwelt, Jena,
46 pp. WHITTEN T., WHITTEN J. & CUBITT G., 1993, Indonésie sauvage, nature et paysage
de l’archipel indonésien, Soline ed., Paris, 207 pp. WHITTEN T., SOERIAATMADAJA R.E. & AFFIF S.A., 1996, The Ecology of Java and
Bali, Périplus ed., Hong Kong, 2, 1028 pp. WIDIANTO H., 2001, The perspective on the evolution of Javanese Homo erectus based on
morphological and stratigraphic characteristics, in : T. Simanjuntak, B. Prasetyo et R. Handini (eds.), Proceedings of the International Colloquium on Sangiran - Sangiran: Man, Culture, and Environment in Pleistocene Times, 24-45.
WIDIANTO H., SEMAH A.M., DJUBIANTONO T. & SEMAH F., 1994, A tentative
reconstruction of the cranial human remains of Hanoman 1 from Bukuran, Sangiran (Central Java), Courier Forschungs-Institut Senkenberg, 171, 47-59.
WIDIANTO H. , BUDIANTO H. & SIMANJUNTAK T. 1996, Laporan penelitian
Sangiran jilid 1, Berita Penelitian Arkeologi, Pusat Penelitian Arkeologi Nasional, Jakarta, 46.

Références bibliographiques.
- 295 - 295
WIDIANTO H. & GRIMAUD-HERVE D., 2000, Un nouveau crâne humain fossile dans le dôme de Sangiran (Java, Indonésie), Compte-Rendus de l’Académie des Sciences, série sciences de la Terre et de l’Univers, Paris, 330, 883-888.
WIDIASMORO, 1984, Geologi daerah Tanjung dan petrogenesa Formasi Kabuh di daerah
Sangiran Tengah, Jawa Tengah, Ditujakan kepada Proyek Penelitian Paleoanthropoloagi Nasional.
YELLEN J.E., 1977, Cultural patterning in faunal remains : evidence from the Kung
Bushmen, in : Ingersoll D., Yellen J.E. & Mc Donald W. (ed), Experimental Archaeology, Columbia University Press ed., 271-331.
YOKOHAMA T., HADIWISASTRA S., HAYASHIDA A. & HAWTORO W., 1980,
Preliminary report on paleomagnetism of the Plio-Pleistocene series in Sangiran and Trinil areas, Central Java, Indonesia, in : S. Nishimura (ed.), Physical Geology of Indonesian Island Arcs, Kyoto University, 88-96.
ZAIM Y., DE VOS J., HUFFMAN O.F., AZIZ F., KAPPELMAN J. & RIZAL Y., 2003,
A new antler specimen from the 1936 Perning hominid site, East Jawa, Indonesia, attributable to Axis lydekkeri (Martin, 1886), Journal of Mineral Technology, Bandung, 10, (2), 1-9.
ZEITOUN V., SEVEAU A., FORESTIER H., THOMAS H., LENOBLE A., LAUDET
F., ANTOINE P.-O., DEBRUYNE R., GINSBURG L., MEIN P., WINAYALAI C., CHUMDEE N., DOYASA T., KIJNGAM A. & NAKBUNLUNG S., 2005, Découverte d’un assemblage faunique à Stegodon-Ailuropoda dans une grotte du Nord de la Thaïlande (Ban Fa Suai, Chiang Dao), Compte-Rendus Palevol, 4, 255-264.
Autres : Encyclopédie Alpha, 1978, L’asie tropicale, Alpha ed., Paris, 7, 300 pp. Encyclopédie Microsoft Encarta 2000, CD-Rom. Liste des sites internet : - www.intercarto.com - www.lonelyplanet.com/mapimages/south_east_asia/java/java.gif

- 296 -

- 297 -
LISTE DES FIGURES Figure 1 : Carte générale de l’Indonésie (archipel indonésien en orangé)............................... 15 Figure 2 : Carte générale de l’île de Java (Indonésie).............................................................. 16 Figure 3 : Limites des plaques tectoniques dans le sud-est asiatique....................................... 17 Figure 4 : Coupe schématique de l’île de Java......................................................................... 17 Figure 5 : Variations du niveau marin (en mètres) depuis 6 Ma, d’après le δ18O des tests de
foraminifères planctoniques tropicaux (en haut d’après Prentice & Denton, 1988) ; depuis environ 150 ka BP, d’après les terrasses récifales de la péninsule de Huon (Papouasie, Nouvelle Guinée) et d’après le δ18O de tests de foraminifères benthiques du forage V19-30, Pacifique équatorial (en bas, d’après Chappel et al., 1996 ; Pillans et al., 1998) (in Détroit, 2002). ................................................................................................. 19
Figure 6 : Fluctuations du niveau marin dans le sud-est asiatique pendant le Pléistocène, a : niveau actuel ; b : - 20 mètres ; c : - 75 mètres ; d : - 120 mètres ................................... 20
Figure 7 : Carte physiographique de l’île de Java.................................................................... 23 Figure 8 : Carte des provinces et des limites zoogéographiques de l’Asie du sud-est avec un
abaissement du niveau marin d’environ 120 mètres. ....................................................... 26 Figure 9 : Proportion des ordres de mammifères actuels de Java. ........................................... 28 Figure 10 : Voies de migrations possibles lors de la formation de la plate-forme de la Sonde
(= Sundaland) au cours du Pléistocène moyen. ............................................................... 30 Figure 11 : Carte des principaux sites paléontologiques de Java............................................. 31 Figure 12 : Carte géologique simplifiée du dôme de Sangiran ................................................ 39 Figure 13 : Colonne stratigraphique du dôme de Sangiran...................................................... 40 Figure 14 : Argiles bleues lagunaires de Kalibeng supérieur (vers –2 Ma)............................. 41 Figure 15 : Argiles noires de Pucangan à Bukuran.................................................................. 42 Figure 16 : Lits à stratifications entrecroisées de Kabuh à Bukuran........................................ 43 Figure 17 : Limite entre les couches de Kabuh et les lahars de Notopuro. .............................. 44 Figure 18 : Diagramme pollinique simplifié des séries de Kalibeng et de Pucangan de
Sangiran............................................................................................................................ 45 Figure 19 : Carte géographique du dôme de Sangiran. ............................................................ 51 Figure 20 : Carte des sites paléontologiques de Java. .............................................................. 59 Figure 21 : Etiquette des collections de Sangiran .................................................................... 78 Figure 22 : Comparaison de la structure de l’émail des Stegodontinae et des Elephantinae. .. 86 Figure 23: Section moyenne d’une cheville osseuse gauche de Bubalus palaeokerabau, au-
dessus de la base, échelle 3/8. .......................................................................................... 98 Figure 24: Crâne de Bubalus palaeokarabau découvert lors des fouilles de 1964 à Tanjung. 98 Figure 25: Section transversale proche de la pointe d’une cheville osseuse gauche de Bibos
palaesondaicus, échelle 2/5............................................................................................ 100 Figure 26: Crâne de Bibos palaesondaicus découvert lors des fouilles de 1964 à Tanjung. . 100 Figure 27 : Diagramme de dispersion pour les M1 et M2 de bovidés (en mm). ..................... 103 Figure 28: Diagramme de dispersion pour les M3 de bovidés (en mm)................................. 104 Figure 29: Diagramme de dispersion pour les M1 et M2 de bovidés (en mm). ...................... 105 Figure 30 : Proportions des restes fouillés de grands bovidés (B1) et de petits bovidés (B2) sur
les sites du dôme de Sangiran. ....................................................................................... 108 Figure 31 : Bois d’Axis axis indien (1/5 nat.)......................................................................... 110 Figure 32: Diagramme de dispersion pour les M1 et M2 de cervidés (en mm). ..................... 114 Figure 33: Diagramme de dispersion pour les M3de cervidés (en mm)................................. 115 Figure 34: Diagramme de dispersion pour les M1 et M2 de cervidés (en mm). ..................... 116

Liste des figures.
- 298 - 298
Figure 35: Diagramme de dispersion pour les scapulas de cervidés (en mm). ...................... 117 Figure 36 : Diagramme de dispersion pour les M3 de cervidés (en mm). .............................. 118 Figure 37 : Bois d’un Cervus hippelaphus javanais. (1/10 nat.) ............................................ 119 Figure 38 : Diagramme de dispersion pour les M1 et M2 de cervidés (en mm). .................... 122 Figure 39: Nombre de restes fouillés de grands cervidés (C1) et de petits cervidés (C2) des
sites du dôme de Sangiran. ............................................................................................. 124 Figure 40: Proportions des restes fouillés de reptiles, de gastéropodes et de poissons des sites
du dôme de Sangiran. ..................................................................................................... 127 Figure 41: Répartition des individus (en NMIf) sur les sites de Tanjung, Sendang Busik,
Ngrejeng et Bukuran. ..................................................................................................... 140 Figure 42: Proportions relatives des ordres actuels et fossiles des mammifères de Java en
nombre d’espèces. .......................................................................................................... 151 Figure 43: Milieux de vie des espèces d’herbivores asiatiques actuels. ................................ 156 Figure 44: Histogrammes écologiques du site de Tanjung 63-64. ......................................... 160 Figure 45: Histogrammes écologiques du site de Tanjung 82. .............................................. 161 Figure 46 : Histogrammes écologiques du site de Sendang Busik. ....................................... 162 Figure 47: Spectres d’association faunique de Tanjung 63-64, Tanjung 82 et de Sendang
Busik............................................................................................................................... 164 Figure 48: Conservation squelettique générale des restes sur les sites de Tanjung, Sendang
Busik, Ngrejeng et Grogol. ............................................................................................ 176 Figure 49 : Conservation squelettique générale des restes sur le site de Ngebung 2. ............ 177 Figure 50 : Représentations squelettiques des bovidés et des cervidés du site de Tanjung pour
les niveaux de 1963-64 et 1982...................................................................................... 178 Figure 51: Représentations squelettiques des bovidés et des cervidés des sites de Sendang
Busik, Ngrejeng, Grogol et Bukuran.............................................................................. 179 Figure 52: Répartition squelettique selon les classes de taille à Tanjung, Sendang Busik,
Ngrejeng et Grogol......................................................................................................... 183 Figure 53: Proportions des membres antérieurs (C) et des membres postérieurs (D) de
Tanjung (TDJ B1), de Sendang Busik (SB B1) et des sites de comparaison................. 186 Figure 54: Proportions des restes crâniens (A) et des restes de l’autopode (E) de Tanjung
(TDJ B1) , de Sendang Busik (SB B1) et des sites de comparaison. ............................. 188 Figure 55: Proportions des restes du squelette axial (B) et des membres postérieurs (D) de
Tanjung (TDJ B1), de Sendang Busik (SB B1) et des sites de comparaison................. 188 Figure 56: Histogramme de fréquence (en % de NR) des classes de taille des restes
déterminables et des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. .............................................................................................................................. 197
Figure 57: Histogrammes de fréquence des longueurs des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. .................................................................. 205
Figure 58: Histogrammes de fréquence des largeurs des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. ............................................................................. 207
Figure 59: histogrammes de fréquence des épaisseurs des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. .................................................................. 209
Figure 60: Répartition des esquilles de Tanjung 63-64 dans les dix classes du diagramme de Folk................................................................................................................................. 211
Figure 61: Répartition des esquilles de Tanjung 82 dans les dix classes du diagramme de Folk......................................................................................................................................... 212
Figure 62 : Répartition des esquilles de Sendang Busik dans les dix classes du diagramme de Folk................................................................................................................................. 212
Figure 63: Répartition des esquilles de Ngrejeng dans les dix classes du diagramme de Folk......................................................................................................................................... 213

Liste des figures.
- 299 - 299
Figure 64 : Répartition des esquilles de Grogol dans les dix classes du diagramme de Folk.213 Figure 65: Répartition des esquilles de Bukuran dans les dix classes du diagramme de Folk.
........................................................................................................................................ 214 Figure 66: Répartition des esquilles de Dayu dans les dix classes du diagramme de Folk. .. 214 Figure 67: Proportions des actions de l’eau sur le matériel des sites de Tanjung, Sendang
Busik, Ngrejeng Grogol, Bukuran et Dayu. ................................................................... 220 Figure 68: Groupes de Voorhies pour les ossements des classes de taille B1, B2C1 et C2 des
sites de Tanjung, Sendang Busik, Ngrejeng et Grogol. ................................................. 226 Figure 69: Proportions des éléments squelettiques dans les assemblages de Tanjung, Sendang
Busik, Ngrejeng, Grogol, Bukuran et Dayu d’après les groupes de Badgley. ............... 231 Figure 70 : Téphrostratigraphie de la formation de Kabuh. ................................................... 241 Figure 71 : Hypothèse des positions des sites par rapport à un cours d’eau d’après les
assemblages de Badgley................................................................................................. 254

- 300 -

- 301 -
LISTE DES TABLEAUX Tableau 1 : Détails des campagnes de fouille dirigées par Dubois à Trinil ............................. 32 Tableau 2 : Détails des campagnes de fouille de l’expédition Selenka à Trinil....................... 34 Tableau 3 : Récapitulatif des données chronologiques sur le dôme de Sangiran. ................... 47 Tableau 4 : Récapitulatif des positions stratigraphiques des sites étudiés. .............................. 53 Tableau 5: Récapitulatif des listes fauniques des sites du Pléistocène inférieur et moyen des
sites de Java. ..................................................................................................................... 56 Tableau 6: Récapitulatif des listes fauniques des sites du Pléistocène supérieur et des sites
récents de Java.................................................................................................................. 58 Tableau 7: Corrélation des faunes de mammifères en Asie. .................................................... 59 Tableau 8: Comparaison des interprétations biostratigraphiques de l’île de Java en 1982...... 63 Tableau 9: Récapitulatif des successions fauniques de l’île de Java et de leur environnement.
.......................................................................................................................................... 64 Tableau 10: Récapitulatif et comparaison des successions biostratigraphiques quaternaires de
l’île de Java....................................................................................................................... 65 Tableau 11: Corrélations biostratigraphiques du dôme de Sangiran........................................ 67 Tableau 12: Les fossiles d’hominidés replacés dans le contexte biostratigraphique de l’île de
Java................................................................................................................................... 68 Tableau 13: Récapitulatif des différentes successions biostratigraphiques proposées pour les
unités de Trinil et de Kedung Brubus............................................................................... 71 Tableau 14: Corrélations des faunes javanaises, indiennes et chinoises plio-pléistocènes...... 73 Tableau 15 : Récapitulatif des données concernant les campagnes de fouilles et les ramassages
de surface de Tanjung, Sendang Busik, Ngejeng, Grogol, Bukuran et Dayu. ................ 77 Tableau 16: Récapitulatif des espèces d’éléphantoïdes de Java à travers les temps................ 80 Tableau 17: Formule lamellaire des 3 sous-espèces de Stegodon trigonocephalus de Java
excluant les demi-rides antérieure et postérieure. ............................................................ 82 Tableau 18 : Nombre de restes du genre Stegodon sur les sites du dôme de Sangiran............ 83 Tableau 19: Mesures des dents de Stegodon trigonocephalus issues des ramassages de surface
de Tanjung (en mm). ........................................................................................................ 85 Tableau 20: Mesures des dents d’Elephas issues des ramassages de surface de Tanjung (en
mm). ................................................................................................................................. 88 Tableau 21: Nombre de restes de proboscidien indéterminé sur les sites du dôme de Sangiran.
.......................................................................................................................................... 89 Tableau 22: Dimensions des M3
d’H. sivalensis sivajavanicus, d’H. sivalensis koenigswaldi et d’H. sivalensis soloensis de la collection Dubois du Naturalis Museum de Leiden (Hooijer, 1950) et de SBII/9/17 (en mm). ........................................................................ 92
Tableau 23: Dimensions des M3 d’H. sivalensis sivajavanicus, d’H. sivalensis koenigswaldi et d’H. sivalensis soloensis de la collection Dubois du Naturalis Museum de Leiden (Hooijer, 1950) et de BK79/surface/B1/R10228 (en mm). .............................................. 94
Tableau 24: Dimensions des M1 d’H. sivalensis sivajavanicus et des M2 d’H. sivalensis
sivajavanicus et d’H. sivalensis koenigswaldi de la collection Dubois du Naturalis Museum de Leiden (Hooijer, 1950) et de NKJ85/surface/R10188 (en mm). .................. 94
Tableau 25: Dimensions des M3 de Sus brachygnathus de Trinil, Kebonduren et Kaligede (Hardjasasmita, 1987) et de SBI/surface/R10063 (en mm). ............................................ 96
Tableau 26: Dimensions des M3 de Bibos javanicus et Epileptobos groeneveldtii (Hooijer, 1958) et de BK79/surface/D2/R10215 (en mm). ............................................................. 99

Liste des tableaux.
- 302 - 302
Tableau 27: Dimensions des P4 de Duboisia santeng de Trinil (Stremme, 1911) et de NKJ85/III/16 .................................................................................................................. 104
Tableau 28: Nombre de restes des grands bovidés (B1) et des petits bovidés (B2) sur les sites du dôme de Sangiran. ..................................................................................................... 107
Tableau 29: Mesures des bois des petits cervidés (en mm). .................................................. 112 Tableau 30 : Dimensions des M1 et M2 de Cervus kendengensis du Naturalis Museum de
Leiden, d’une M2 de Cervus (Rusa) sp. (von Koenigswald, 1933) et de NKJ85/I/43 (en mm). ............................................................................................................................... 120
Tableau 31: Dimensions des M3 de Cervus kendengensis du Naturalis Museum de Leiden et de NKJ85/I/11 (en mm).................................................................................................. 120
Tableau 32: Dimensions des P4 de Cervus hippelaphus (von Koenigswald, 1933) et de NKJ85/II/1 (en mm). ...................................................................................................... 121
Tableau 33:Dimensions des P3 de Cervus hippelaphus (von Koenigswald, 1933) et de NKJ85/II/18 (en mm). .................................................................................................... 121
Tableau 34: Nombre de restes des grands cervidés (C1), des petits cervidés (C2) sur les sites du dôme de Sangiran. ..................................................................................................... 123
Tableau 35: Nombre de restes des reptiles et des poissons sur les sites du dôme de Sangiran......................................................................................................................................... 126
Tableau 36: Nombre et type de restes de tortues dans les collections fouillées de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. ................................................... 127
Tableau 37: Nombres de restes (NR) issus des fouilles et des ramassages de surface et coefficient de similarité des collections de Tanjung 63-64, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. .............................................................................................. 129
Tableau 38: Listes fauniques de Tanjung, Sendang Busik, Ngrejeng,Grogol, Bukuran, Dayu......................................................................................................................................... 132
Tableau 39: Listes fauniques établies à partir des restes issus des fouilles de Tanjung, Sendang Busik, Ngrejeng,Grogol, Bukuran, Dayu et Ngebung 2.................................. 133
Tableau 40: Coefficients de similarité de Jaccard pour les sites de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. ............................................................................. 134
Tableau 41 : Coefficients de similarité de Jaccard des sites de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu avec les différents niveaux de Ngebung 2. .......... 134
Tableau 42: Taux de détermination des sites de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu............................................................................................................. 135
Tableau 43: Spectre faunique de Tanjung 63-64. .................................................................. 136 Tableau 44: Spectre faunique de Tanjung 82......................................................................... 136 Tableau 45: Spectre faunique de Sendang Busik. .................................................................. 137 Tableau 46: Spectre faunique de Ngrejeng. ........................................................................... 137 Tableau 47: Spectre faunique de Grogol................................................................................ 138 Tableau 48: Spectre faunique de Bukuran. ............................................................................ 138 Tableau 49: Âges probables des mammifères de Tanjung 63-64. ......................................... 142 Tableau 50: Âges probables des mammifères de Tanjung 82................................................ 142 Tableau 51: Âges probables des mammifères de Sendang Busik. ......................................... 142 Tableau 52: Âges probables des mammifères de Ngrejeng. .................................................. 142 Tableau 53: Âges probables des mammifères de Grogol....................................................... 142 Tableau 54: Âges probables des mammifères de Bukuran. ................................................... 143 Tableau 55: Indice de richesse biologique des sites de Tanjung, Sendang Busik, Ngrejeng et
Bukuran. ......................................................................................................................... 143 Tableau 56: Correspondance des associations fauniques proposées par Sondaar (1984) avec
les espèces des sites de notre étude. ............................................................................... 144

Liste des tableaux.
- 303 - 303
Tableau 57 : Récapitulatif des espèces présentes dans les différentes formations sédimentaires de Sangiran suite à notre étude....................................................................................... 147
Tableau 58: Paramètres écologiques pour la réalisation des histogrammes de Tanjung 63-64......................................................................................................................................... 160
Tableau 59: : Paramètres écologiques pour la réalisation des histogrammes de Tanjung 82.161 Tableau 60: Paramètres écologiques pour la réalisation des histogrammes de Sendang Busik.
........................................................................................................................................ 162 Tableau 61 : Nombres et proportions des restes des assemblages de Tanjung, Sendang Busik,
Ngrejeng, Grogol, Bukuran et Dayu. ............................................................................. 174 Tableau 62 : Proportions des esquilles (NRI) sur les sites de plein air du Pléistocène moyen de
Ngebung 2 et du Paléolithique inférieur de Terra Amata (Serre, 1987), Soleilhac (Fosse & Bonifay, 1994), Notarchirico (Cassoli et al., 1999) et Holon (Monchot & Horwitz, 2002)............................................................................................................................... 175
Tableau 63 : Répartition squelettique selon les classes de taille à Tanjung, Sendang Busik, Ngrejeng et Grogol......................................................................................................... 182
Tableau 64: Liste des proportions des éléments squelettiques des sites de Tanjung et de Sendang Busik et des sites de comparaison. .................................................................. 187
Tableau 65: IGCS NR et ICD NR des classes de taille B1, B2C1 et C2 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran, Dayu, Notarchirico (Bos), Holon (Bos), Ngebung A, B et Upper. ..................................................................................................................... 192
Tableau 66: Indices squelettiques calculés pour les classes de taille B1, B2C1 et C2, pour les sites de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran, Dayu, Notarchirico, Holon et Ngebung. ......................................................................................................... 194
Tableau 67: IFO des classes de taille B1, B2C1 et C2 et général de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. ............................................................................. 200
Tableau 68 : Fragmentation des os longs retrouvées sur les sites de Tanjung, Sendang Busik, Grogol et Bukuran.......................................................................................................... 201
Tableau 69 : Nombre et proportions des dents entières et fragmentées dans les assemblages de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. .................................... 202
Tableau 70 : Données des histogrammes de fréquences des longueurs, largeurs et épaisseurs des esquilles de Tanjung, Sendang Busik, Ngrejeng et Grogol. .................................... 203
Tableau 71 : Répartition des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu dans les dix classes du diagramme de forme et de sphéricité. ........... 211
Tableau 72 : Répartition des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu dans les classes de formes.................................................................. 216
Tableau 73: Proportions des actions liées à l’eau sur les sites de Sangiran. .......................... 219 Tableau 74 : Proportions des restes affectés par les différents types de dissolution dans les
trois niveaux de Ngebung 2............................................................................................ 221 Tableau 75 : Proportions des différents types de fissuration observées à Tanjung, Sendang
Busik, Ngrejeng, Grogol, Bukuran et Dayu. .................................................................. 223 Tableau 76 : Proportions des restes affectés par les différents types de fissuration dans les
trois niveaux de Ngebung 2............................................................................................ 224 Tableau 77: Groupes de Voorhies pour les restes des classes de taille B1, B2C1 et C2 des
sites de Tanjung, Sendang Busik, Ngrejeng et Grogol. ................................................. 225 Tableau 78: Rapport dents/vertèbres des classes de taille B1, B2C1 et C2 des sites de Tanjung
et Sendang Busik. ........................................................................................................... 228 Tableau 79: Rapport dents/vertèbres des sites de Tanjung, Sendang Busik, Ngrejeng, Grogol,
Bukuran et Dayu............................................................................................................. 229

Liste des tableaux.
- 304 - 304
Tableau 80: Abondance des éléments anatomiques dans quatre assemblages taphonomiques, montrant les changements de composition suivant l’augmentation du transport fluviatile (de IV à I). ...................................................................................................................... 230
Tableau 81 : Groupes de Voorhies pour la classe des grands bovidés du site de Ngebung 2......................................................................................................................................... 232
Tableau 82 :Action des carnivores dans les niveaux de Ngebung 2. ..................................... 235 Tableau 83: Action anthropique à Ngebung 2. ...................................................................... 238 Tableau 84 : Récapitulatif des espèces présentes dans les différentes formations sédimentaires
de Sangiran suite à notre étude....................................................................................... 253

- 305 -
ANNEXES
- Annexe 1 : Carte géographique au 1/25000 du dôme de Sangiran. - Annexe 2 : Liste des principaux restes d’hominidés retrouvés sur l’île de Java et sur le
dôme de Sangiran. - Annexe 3 : Liste de synonymes d’espèces de Java. - Annexe 4 : Planches photographiques des restes fauniques déterminables. - Annexe 5 : Tableaux de mesures. - Annexe 6 : Répartition spatiale. - Annexe 7 : Tests statistiques sur la répartition spatiale. - Annexe 8 : Présentation des sites de comparaison. - Annexe 9 : Méthode du khi-deux ou khi-carré. - Annexe 10 : Méthode du test statistique de Kolmogorov-Smirnov. - Annexe 11 : Planche photographique des traces taphonomiques.
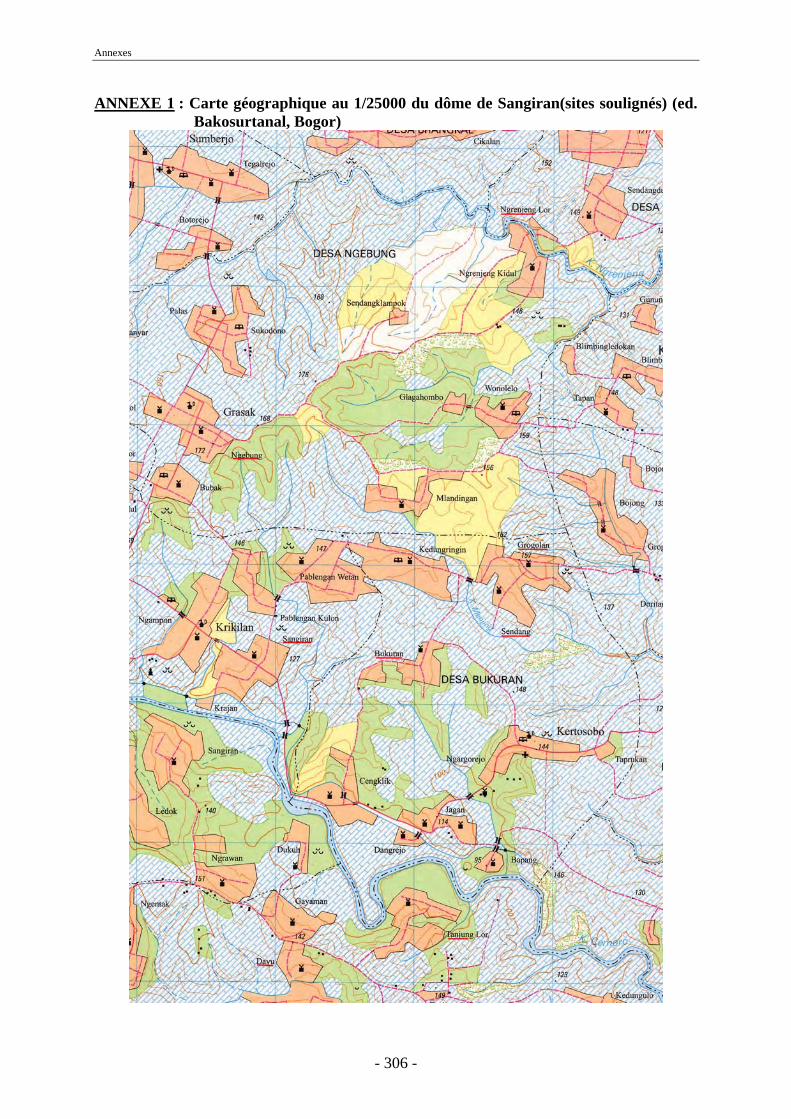
Annexes
- 306 -
ANNEXE 1 : Carte géographique au 1/25000 du dôme de Sangiran(sites soulignés) (ed. Bakosurtanal, Bogor)
Annexes
- 307 -
Légende :
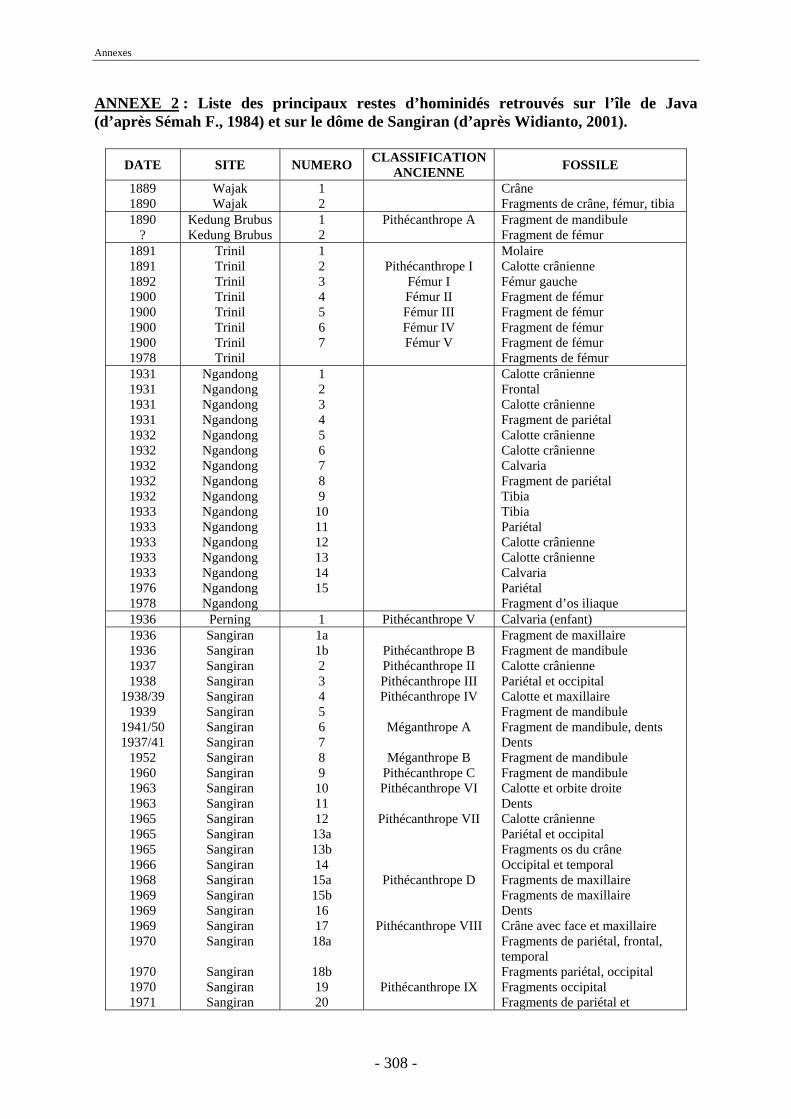
Annexes
- 308 -
ANNEXE 2 : Liste des principaux restes d’hominidés retrouvés sur l’île de Java (d’après Sémah F., 1984) et sur le dôme de Sangiran (d’après Widianto, 2001).
DATE SITE NUMERO CLASSIFICATION ANCIENNE FOSSILE
1889 1890
Wajak Wajak
1 2
Crâne Fragments de crâne, fémur, tibia
1890 ?
Kedung Brubus Kedung Brubus
1 2
Pithécanthrope A Fragment de mandibule Fragment de fémur
1891 1891 1892 1900 1900 1900 1900 1978
Trinil Trinil Trinil Trinil Trinil Trinil Trinil Trinil
1 2 3 4 5 6 7
Pithécanthrope I
Fémur I Fémur II Fémur III Fémur IV Fémur V
Molaire Calotte crânienne Fémur gauche Fragment de fémur Fragment de fémur Fragment de fémur Fragment de fémur Fragments de fémur
1931 1931 1931 1931 1932 1932 1932 1932 1932 1933 1933 1933 1933 1933 1976 1978
Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong Ngandong
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15
Calotte crânienne Frontal Calotte crânienne Fragment de pariétal Calotte crânienne Calotte crânienne Calvaria Fragment de pariétal Tibia Tibia Pariétal Calotte crânienne Calotte crânienne Calvaria Pariétal Fragment d’os iliaque
1936 Perning 1 Pithécanthrope V Calvaria (enfant) 1936 1936 1937 1938
1938/39 1939
1941/50 1937/41
1952 1960 1963 1963 1965 1965 1965 1966 1968 1969 1969 1969 1970
1970 1970 1971
Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran
Sangiran Sangiran Sangiran
1a 1b 2 3 4 5 6 7 8 9
10 11 12 13a 13b 14 15a 15b 16 17 18a
18b 19 20
Pithécanthrope B Pithécanthrope II Pithécanthrope III Pithécanthrope IV
Méganthrope A
Méganthrope B
Pithécanthrope C Pithécanthrope VI
Pithécanthrope VII
Pithécanthrope D
Pithécanthrope VIII
Pithécanthrope IX
Fragment de maxillaire Fragment de mandibule Calotte crânienne Pariétal et occipital Calotte et maxillaire Fragment de mandibule Fragment de mandibule, dents Dents Fragment de mandibule Fragment de mandibule Calotte et orbite droite Dents Calotte crânienne Pariétal et occipital Fragments os du crâne Occipital et temporal Fragments de maxillaire Fragments de maxillaire Dents Crâne avec face et maxillaire Fragments de pariétal, frontal, temporal Fragments pariétal, occipital Fragments occipital Fragments de pariétal et
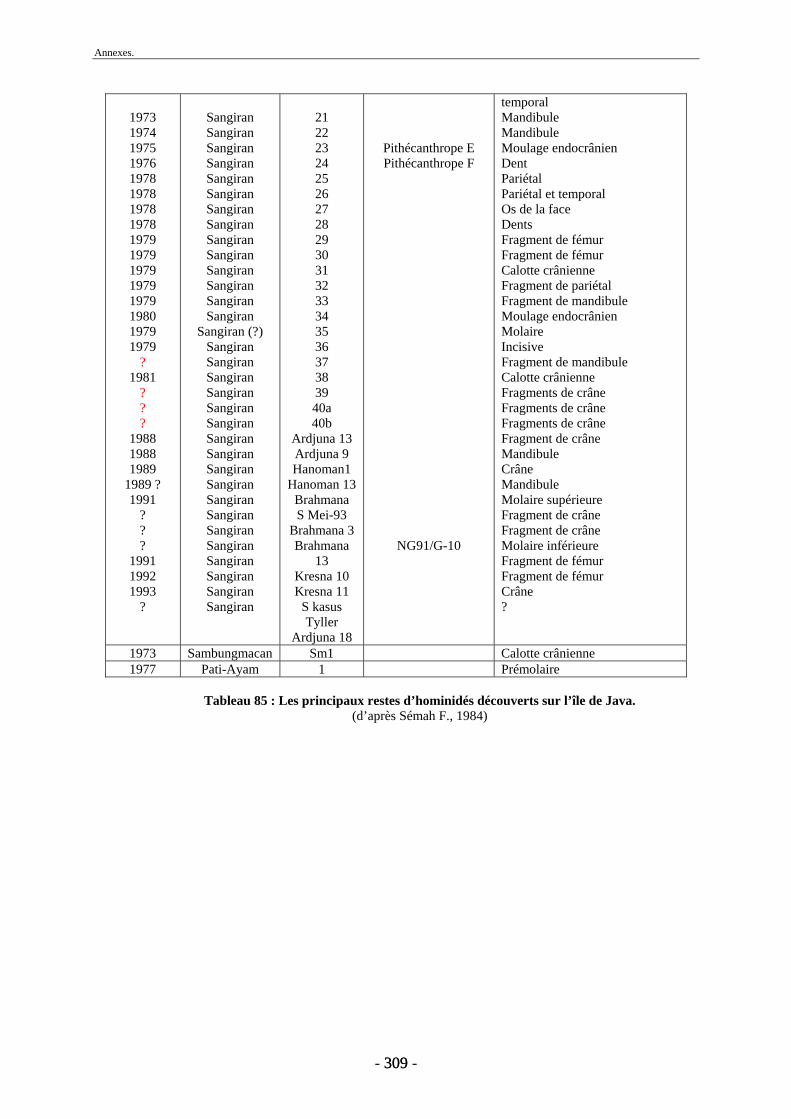
Annexes.
- 309 - 309
1973 1974 1975 1976 1978 1978 1978 1978 1979 1979 1979 1979 1979 1980 1979 1979
? 1981
? ? ?
1988 1988 1989
1989 ? 1991
? ? ?
1991 1992 1993
?
Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran
Sangiran (?) Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran Sangiran
21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40a 40b
Ardjuna 13 Ardjuna 9 Hanoman1
Hanoman 13 Brahmana S Mei-93
Brahmana 3 Brahmana
13 Kresna 10 Kresna 11
S kasus Tyller
Ardjuna 18
Pithécanthrope E Pithécanthrope F
NG91/G-10
temporal Mandibule Mandibule Moulage endocrânien Dent Pariétal Pariétal et temporal Os de la face Dents Fragment de fémur Fragment de fémur Calotte crânienne Fragment de pariétal Fragment de mandibule Moulage endocrânien Molaire Incisive Fragment de mandibule Calotte crânienne Fragments de crâne Fragments de crâne Fragments de crâne Fragment de crâne Mandibule Crâne Mandibule Molaire supérieure Fragment de crâne Fragment de crâne Molaire inférieure Fragment de fémur Fragment de fémur Crâne ?
1973 Sambungmacan Sm1 Calotte crânienne 1977 Pati-Ayam 1 Prémolaire
Tableau 85 : Les principaux restes d’hominidés découverts sur l’île de Java.
(d’après Sémah F., 1984)
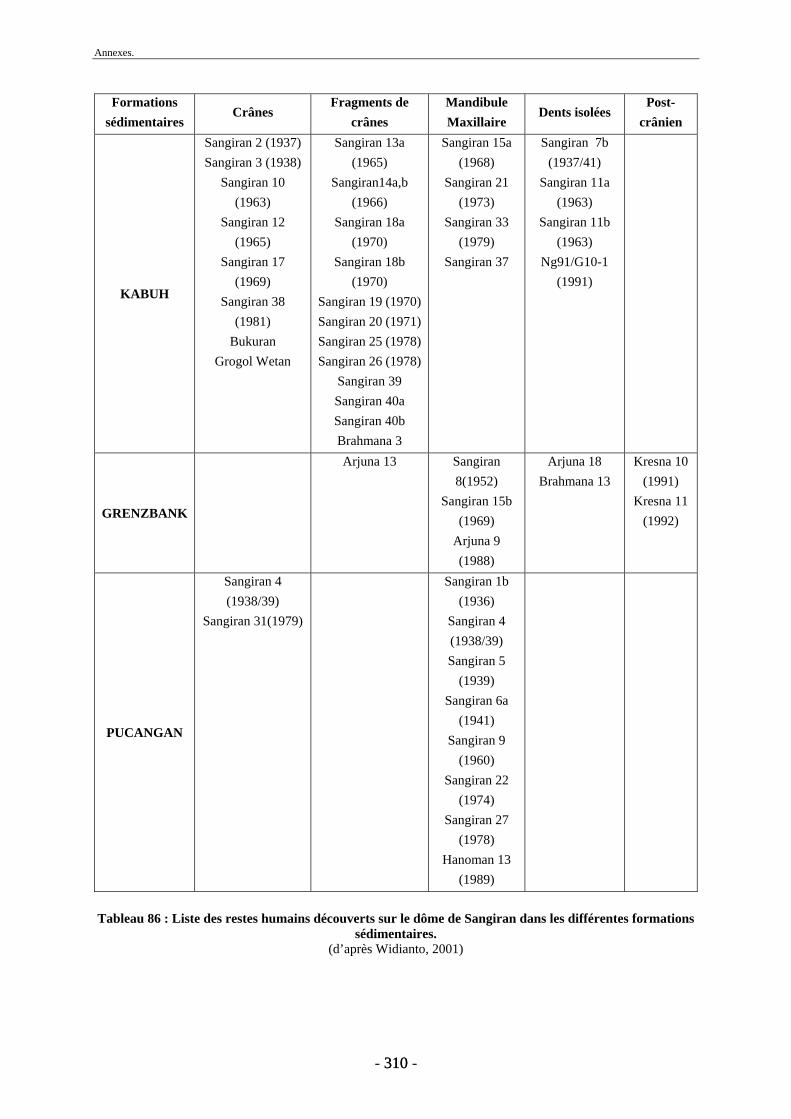
Annexes.
- 310 - 310
Formations sédimentaires
Crânes Fragments de
crânes Mandibule Maxillaire
Dents isolées Post-
crânien
KABUH
Sangiran 2 (1937) Sangiran 3 (1938)
Sangiran 10 (1963)
Sangiran 12 (1965)
Sangiran 17 (1969)
Sangiran 38 (1981)
Bukuran Grogol Wetan
Sangiran 13a (1965)
Sangiran14a,b (1966)
Sangiran 18a (1970)
Sangiran 18b (1970)
Sangiran 19 (1970) Sangiran 20 (1971) Sangiran 25 (1978) Sangiran 26 (1978)
Sangiran 39 Sangiran 40a Sangiran 40b Brahmana 3
Sangiran 15a (1968)
Sangiran 21 (1973)
Sangiran 33 (1979)
Sangiran 37
Sangiran 7b (1937/41)
Sangiran 11a (1963)
Sangiran 11b (1963)
Ng91/G10-1 (1991)
GRENZBANK
Arjuna 13 Sangiran 8(1952)
Sangiran 15b (1969)
Arjuna 9 (1988)
Arjuna 18 Brahmana 13
Kresna 10 (1991)
Kresna 11 (1992)
PUCANGAN
Sangiran 4 (1938/39)
Sangiran 31(1979)
Sangiran 1b (1936)
Sangiran 4 (1938/39) Sangiran 5
(1939) Sangiran 6a
(1941) Sangiran 9
(1960) Sangiran 22
(1974) Sangiran 27
(1978) Hanoman 13
(1989)
Tableau 86 : Liste des restes humains découverts sur le dôme de Sangiran dans les différentes formations
sédimentaires. (d’après Widianto, 2001)

Annexes.
- 311 - 311
ANNEXE 3 : Liste de synonymes d’espèces de Java. PROBOSCIDIENS: Sinomastodon bumiajuensis (van der Maarel, 1932) : in Saegusa, 1995 Mastodon sp.: in van der Maarel, 1932 ? Mastodon sp.: in van der Maarel, 1932 Mastodon (Trilophodon) bumiajuensis (van der Maarel, 1932): in von Koenigswald, 1933 Stegodon trigonocephalus Martin, 1887 Mastodon elephantoides in Junghuhn, 1857 Stegodon aff. bombifrons Falconer & Cautley, 1846: in Martin, 1887 Stegodon cliftii Falconer & Cautley, 1846: in Martin, 1887 Stegodon airawana : in Martin, 1890 Stegodon javanoganesa Dubois, 1908 Stegodon ganesa var. javanicus Dubois, 1908 Stegodon bondolensis : in van der Maarel, 1932 Stegodon trigonocephalus trigonocephalus Martin, 1887; in von Koenigswald, 1933 Stegodon trigonocephalus praecursor von Koenigswald, 1935 Stegodon hypsilophus Hooijer, 1954 Elephas celebensis : in Hooijer,1974 Elephas hysudrindicus Dubois, 1908 Euelephas namadicus : in Martin, 1887 Euelephas hysudricus : in Martin, 1887 Elephas indicus : in Dubois, 1891 Palaeoloxodon hysudrindicus (Dubois, 1908) : in Osborn, 1942 Elephas ? maximus L. fossilis in van der Maarel, 1932 PERISSODACTYLES: Rhinoceros sondaicus Desmarest, 1822 Rhinoceros javanicus Geoffroy & Cuvier, 1824 Rhinoceros javanus Cuvier, 1829 Rhinoceros nasalis Gray, 1868 Rhinoceros sivasondaicus Dubois, 1908 Aceratherium boschi Koenigswald, 1933 Tapirus indicus Desmarest, 1819 Tapirus pandanicus Dubois, 1908

Annexes.
- 312 - 312
ARTIODACTYLES: Hippopotamidés : Hexaprotodon sivalensis sivajanicus Dubois, 1908 Hippopotamus : in Dubois, 1891 Hexaprotodon sivajavanicus Dubois, 1908 Hippopotamus sivajavanicus : in Martin, 1919 Hippopotamus (Hexaprotodon) simplex von Koenigswald, 1933 Hippopotamus simplex: in von Koenigswald, 1934 Hexaprotodon simplex : in von Koenigswald, 1939 Hexaprotodon sivalensis koenigswaldi (Hooijer, 1948) Hippopotamus sp. in Cosijn, 1931 Hippopotamus (Hexaprotodon) antiquus : in von Koenigswald, 1934 Hippopotamus antiquus Desmarest, 1822 : in von Koenigswald, 1934 Hippopotamus koenigswaldi Hooijer, 1948 Hippopotamus sivalensis soloensis Hooijer, 1950 Hippopotamus sp. : in Stremme, 1911 Hippopotamus (Hexaprotodon) sivajavanicus : in von Koenigswald, 1933 Hippopotamus (Hexaprotodon) namadicus : in von Koenigswald, 1934 Hippopotamus namadicus : in von Koenigswald, 1934 Bovidés: Epileptobos groeneveldtii (Dubois, 1908): in Hooijer, 1958 Leptobos groeneveldtii Dubois, 1908 Leptobos cosijni von Koenigswald, 1934 Bibos cosijni : in von Koenigswald, 1934 Duboisia santeng (Dubois, 1891) : in Brongersma, 1936 Anoa santeng Dubois, 1891 Tetraceros kroesenii Dubois, 1907 Tetracerus kroesenii : in Dubois, 1908 Duboisia kroesenii : in Stremme, 1911 Antilope modjokertensis von Koenigswald, 1934 Duboisia säatensis von Koenigswald, 1934 Duboisia (?) sartonoi n. sp. Geraads, 1979 Bos javanicus d'Alton, 1823 Bos leucoprymnus Quoy & Gaimard, 1830 Bos sondaicus Temminck, 1839 Bos banteng Wagner, 1844 Bos bantiger Schlegel et Müller, 1845

Annexes.
- 313 - 313
Bibos palaesondaicus Dubois, 1908 Bison sivalensis : in Martin, 1887 Bibos banteng Dubois, 1891 Bibos protocavifrons Dubois, 1908 Bibos palaeosondaicus Stremme, 1911 Leptobos dependicornus Dubois, 1908 Bibos sondaicus fossilis in van der Maarel, 1932 Bos (Bibos) banteng palaeosondaicus in von Koenigswald, 1933 Leptobos dependicornis : in Pilgrim, 1939 Bubalus palaeokerabau Dubois, 1908 Bubalus buffelus : in Dubois, 1891 Buffelus palaeokerabau : in Stremme, 1911 ? Bos sp. : in Stehn & Umbgrove, 1929 Buffelus bubalus? var. sondaicus fossilis : in van der Maarel, 1932 Bos (Bubalus) bubalis palaeokerabau : in von Koenigswald, 1933 Bos (Bubalus) cf. bubalis : in von Koenigswald, 1934 Cervidés: Axis lydekkeri (Martin, 1886) Cervus lydekkeri Martin, 1886 Cervus liriocerus Dubois, 1907 Cervus (Axis) lydekkeri : in Stremme, 1911 Cervus zwaani von Koenigswald, 1933 Muntiacus muntjak (Zimmermann, 1780) Cervus muntjak Zimmermann, 1780 Cervus (Rusa) sp. : in von Koenigswald, 1933 Cervus sp. : in Martin, 1888 Cervus (incl. Rusa) sp. indet. : in Dubois, 1892 Cervus : in Dubois, 1907 Cervus kendengensis Dubois, 1908 Cervus palaeomendjangan Dubois, 1908 Cervus sp.: in Stehn & Umgrove, 1926, Cervus (Rusa) hippelaphus : in von Koenigswald, 1933 Cervus sp.: in von Koenigswald, 1933 Cervus (Rusa) oppenoorthi von Koenigswald, 1933 Cervus hippelaphus : in von Koenigswald, 1934 Suidés: Sus brachygnathus Dubois, 1908 Sus hysudricus : in Martin, 1890 Sus celebensis Müller & Schlegel, 1843 Sus vatualangensis von Koenigswald, 1933 Sus barbatus : in Badoux, 1959

Annexes.
- 314 - 314
Sus macrognathus Dubois, 1908 Sus verrucosus Hodgson, 1847 : in Dubois, 1891 Sus coerti von Koenigswald, 1934 Sus verrucosus Müller & Schlegel, 1842 : in Badoux, 1959 Sus scrofa Linné, 1758 Sus milleri Jentink, 1905 CARNIVORA : Panthera tigris Linné, 1758 Felis groeneveldtii Dubois, 1907 Felis oxygnatha Dubois, 1908 Felis trinilensis Dubois, 1908 Feliopsis palaeojavanica Stremme, 1911 Prionailurus bengalensis Kerr, 1792 Felis javanensis Desmarest, 1816 ? Felis undata : in Desmarest, 1816 Felis minuta Temminck, 1824 Felis microgale Dubois, 1908 Prionailurus bengalensis microgale : in Hemmer, 1971 Pachycrocuta brevirostris Aymard, 1856 Hyaena brevirostris bathygnatha Dubois, 1908 Crocuta (?) bathygnatha : in Brongersma, 1935 Cuon javanicus Pallas, 1811 Cuon alpinus Desmarest, 1820 PRIMATES: Macaca fascicularis Raffles, 1821 Macaca mordax Thomas & Wroughton, 1909 Macaca resima Thomas & Wroughton, 1909 Pithecus baweanus Elliot, 1910 Pithecus cupidus Elliot, 1910 RONGEURS : Hystrix Linné, 1758 Acanthion Cuvier, 1823 Thecurus Lyon, 1907

Annexes.
- 315 - 315
ANNEXE 4 : Planches photographiques des restes fauniques déterminables.

Annexes.
- 316 - 316
PLANCHE I - Ordre des proboscidiens : Famille des Stegodontidae : Collections de surface :
I a- M2 de Stegodon trigonocephalus,TDJ64/surface/R10235, vue occlusale. I b- M3 de Stegodon trigonocephalus,TDJ64/surface/6, vue occlusale. I c- D4 gauche de Stegodon trigonocephalus, TDJ64/surface/R10219, vue occlusale. I d- D4 gauche de Stegodon trigonocephalus,TDJ64/surface/R10224, vue occlusale. I e- M2 droite de Stegodon trigonocephalus,TDJ64/surface/R10227, vue occlusale.

Annexes.
- 317 - 317
PLANCHE I
a b
c d e

Annexes.
- 318 - 318
PLANCHE II - Ordre des proboscidiens : Famille des Elephantidae: Collections fouillées: II a - Fragment de dent d’Elephas sp., TDJ82/I/26, vue latérale.
II b - Fragment de dent d’Elephas sp., TDJ82/I/26, vue antérieure. Collections de surface :
II c- M2 d’Elephas sp., TDJ64/surface/R10223, vue latérale. II d- M3 droite d’Elephas sp., TDJ64/surface/R10222, vue occlusale.
II e- Molaire supérieure d’Elephas sp., TDJ64/surface/R10236, vue latérale. II f- Molaire supérieure d’Elephas sp., TDJ64/surface/R10231, vue vestibulaire.
II g- M3 gauche d’Elephas sp., TDJ64/surface/R10226, vue occlusale II h- M3
gauche d’Elephas sp., TDJ64/surface/R10220, vue linguale.

Annexes.
- 319 - 319
PLANCHE II
a b
c d e
f g h

Annexes.
- 320 - 320
PLANCHE III - Ordre des Périssodactyles: Famille des Rhinocerotidae : Collections fouillées: III a- Sésamoïde de Rhinoceros sp., TDJ82/IV/142, vue basale. III b- Fémur de Rhinoceros sp., TDJ64/H14/B18, épiphyse distale, vue basale. - Ordre des Artiodactyles : Famille des Hippopotamidae : Collections fouillées : III c- D1 gauche d’Hexaprotodon sivalensis, SBII/5/34, vue vestibulaire. III d- M3 gauche d’Hexaprotodon sivalensis, SBII/9/17, vue occlusale. Collections de surface : III e- P3 droite d’Hexaprotodon sivalensis, BK79/surface/B1/R10229, vue occlusale. III f- P4 gauche d’Hexaprotodon sivalensis, SBI/surface/D9/R10010, vue occlusale. III g- Mandibule droite d’Hexaprotodon sivalensis, NKJ85/surface/R10188, vue occlusale. III h- M3 droite d’Hexaprotodon sivalensis, TDJ64/surface/R10228, vue occlusale.

Annexes.
- 321 - 321
PLANCHE III
a b
c d
e f
g h

Annexes.
- 322 - 322
PLANCHE IV - Ordre des Artiodactyles : suite
Famille des Suidae : Collections fouillées : IV a- M3 droite fragmentée de Sus brachygnathus, BK79/SIII/A1/2, vue occlusale. IV b- Incisive supérieure de Sus sp, NKJ85/II/13, vue latérale. IV c- Tibia gauche de Sus sp., TDJ82/I/29, épiphyse distale, vue médiale. Collections de surface : IV d- M3 droite de Sus brachygnathus, SBI80/surface/R10063, vue occlusale.
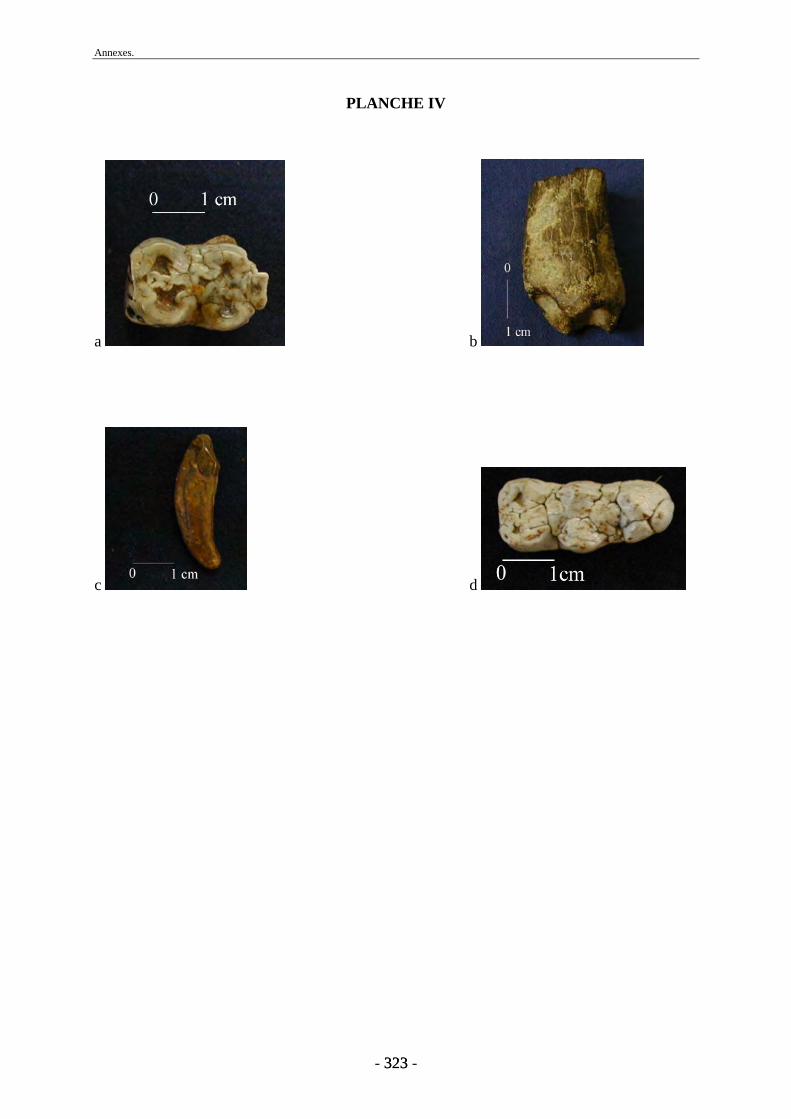
Annexes.
- 323 - 323
PLANCHE IV
a b
c d

Annexes.
- 324 - 324
PLANCHE V - Ordre des Artiodactyles : suite Famille des Bovidae : Collections fouillées : V a- Cheville osseuse de Bibos palaesondaicus, TDJ64/F4/89, vue antérieure. V b- Cheville osseuse de Bibos palaesondaicus, TDJ64/H7/2B, vue supérieure. V c- M2 gauche de Duboisia santeng, TDJ82/IV/46, vue occlusale. V d- M2 droite de Duboisia santeng, NKJ85/III/10005, vue occlusale. V e- M3 droite de Duboisia santeng, NKJ85/III/1, vue occlusale. V f- P4 droite de Duboisia santeng, NKJ85/III/16, vue occlusale. V g- M2 droite fragmentée de Duboisia santeng, BK79/VI/A1/10004, vue occlusale. V h- M2 droite de Duboisia santeng, NKJ85/III/15, vue occlusale. V i- M3 gauche fragmentée de Duboisia santeng, TDJ82/IV/138, vue occlusale. V j- Molaire inférieure gauche fragmentée de Duboisia santeng, TDJ82/IV/58, vue occlusale. Collections de surface : V k- M3 gauche de Bubalus palaeokerabau, BK79/surface/D2/R10215, vue occlusale. V l- M2 droite de Duboisia santeng, SBII/surface/D8/R10030, vue occlusale. V m- M2 droite de Duboisia santeng, BK79/surface/E1/R10106, vue vestibulaire. V n- Molaire inférieure fragmentée de Duboisia santeng, BK79/surface/C1/R10350, vue linguale.
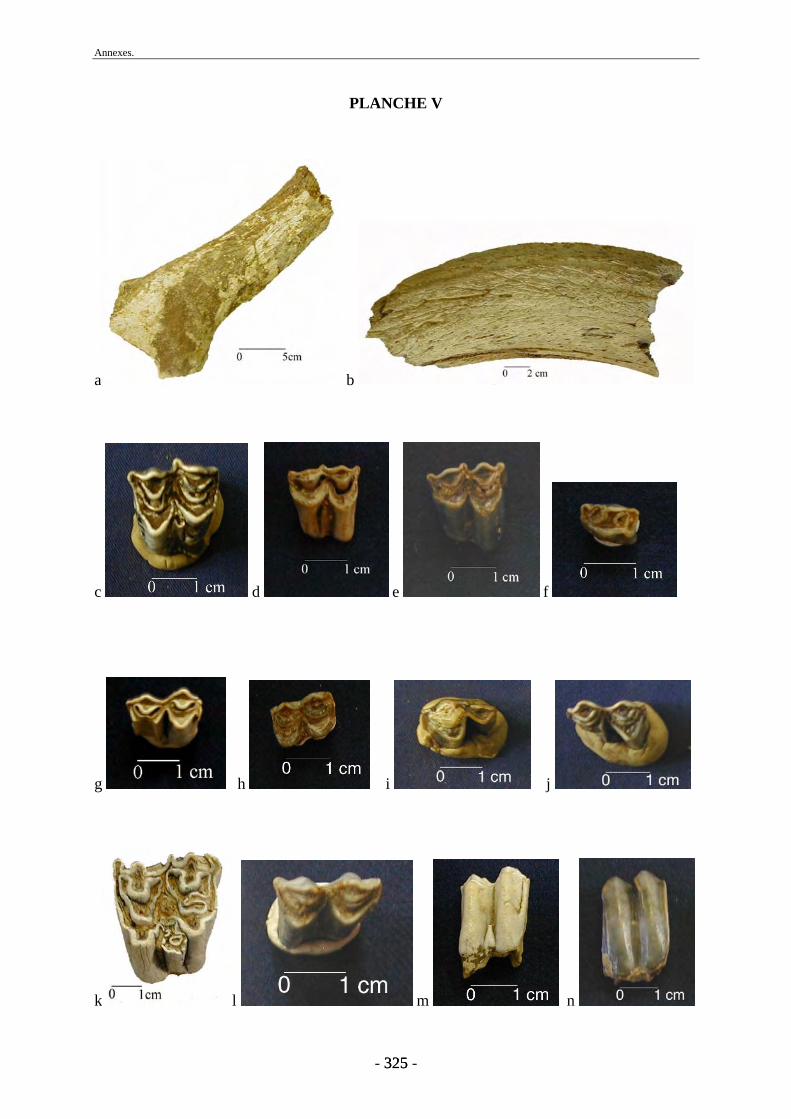
Annexes.
- 325 - 325
PLANCHE V
a b
c d e f
g h i j
k l m n

Annexes.
- 326 - 326
PLANCHE VI - Ordre des Artiodactyles : suite Famille des Cervidae : Collections fouillées : VI a- Bois de chute gauche d’Axis lydekkeri, TDJ64/G11F11/13B10B, vue latérale. VI b- Bois de chute d’Axis lydekkeri, TDJ64/I4/930, vue latérale. VI c- Bois d’Axis lydekkeri, BK79/III/B1/1, partie supérieure, vue mésiale.

Annexes.
- 327 - 327
PLANCHE VI
a b
c

Annexes.
- 328 - 328
PLANCHE VII - Ordre des Artiodactyles : suite Famille des Cervidae : Collections fouillées : VII a- M1 droite d’Axis sp., GP83/II/b2/74, vue occlusale. VII b- M2 gauche d’Axis sp., TDJ82/IV/91, vue occlusale. VII c- M2 gauche d’Axis sp., GP83/II/A1/11, vue occlusale. VII d- M3 gauche d’Axis sp., TDJ82/IV/26, vue occlusale VII e- M3 gauche d’Axis sp., GP83/II/A3/25, vue occlusale. VII f- M1ou2 gauche d’Axis sp., TDJ82/IV/144, vue occlusale. Collections de surface : VII g– M3 droite d’Axis sp., BK79/surface/E1/R10040, vue occlusale. VII h- M3 gauche d’Axis sp., BK79/surface/R10054, vue occlusale.

Annexes.
- 329 - 329
PLANCHE VII
a b c
d e
f
g h

Annexes.
- 330 - 330
PLANCHE VIII Collections fouillées: VIII a- P4 droite de Cervus (Rusa) sp., NKJ85/III/18, vue occlusale. VIII b- M2 gauche de Cervus (Rusa) sp., NKJ85/I/43, vue occlusale. VIII c- M3 gauche de Cervus (Rusa) sp., NKJ85/I/11, vue occlusale. VIII d- P4 droite de Cervus (Rusa) sp., NKJ85/II/12, vue occlusale. VIII e- P3 droite de Cervus (Rusa) sp., NKJ85/II/18, vue occlusale. VIII f- M2 gauche de Cervus (Rusa) sp., NKJ85/III/11, vue occlusale. Collections de surface: VIII g- M2 droite de Cervus (Rusa) sp., NKJ85/II/surface/R10009, vue occlusale.

Annexes.
- 331 - 331
PLANCHE VIII
a b c
d e f
g

Annexes.
- 332 - 332
PLANCHE IX Carnivores : IX a- Fragment de prémolaire inférieure droite de Pachycrocuta brevirostris, BK79/VI/A1/9, vue occlusale. Reptiles et poissons: IX b- Hypoplastron de Trionyx, TDJ64/10155, vue ventrale. IX c- Mandibule de crocodile, NP85/IV/2, vue occlusale. IX d- Dent de crocodile, BK79/SIV/A1/10, vue vestibulaire. IX e- Vertèbre de poisson, BK79/SIV/A1/10, vue caudale. IX f- Dent de squalidé, NKJ85/II/11, vue latérale. IX g- Opercule de gastéropode, BK79/SIV/A1/4, vue occlusale.
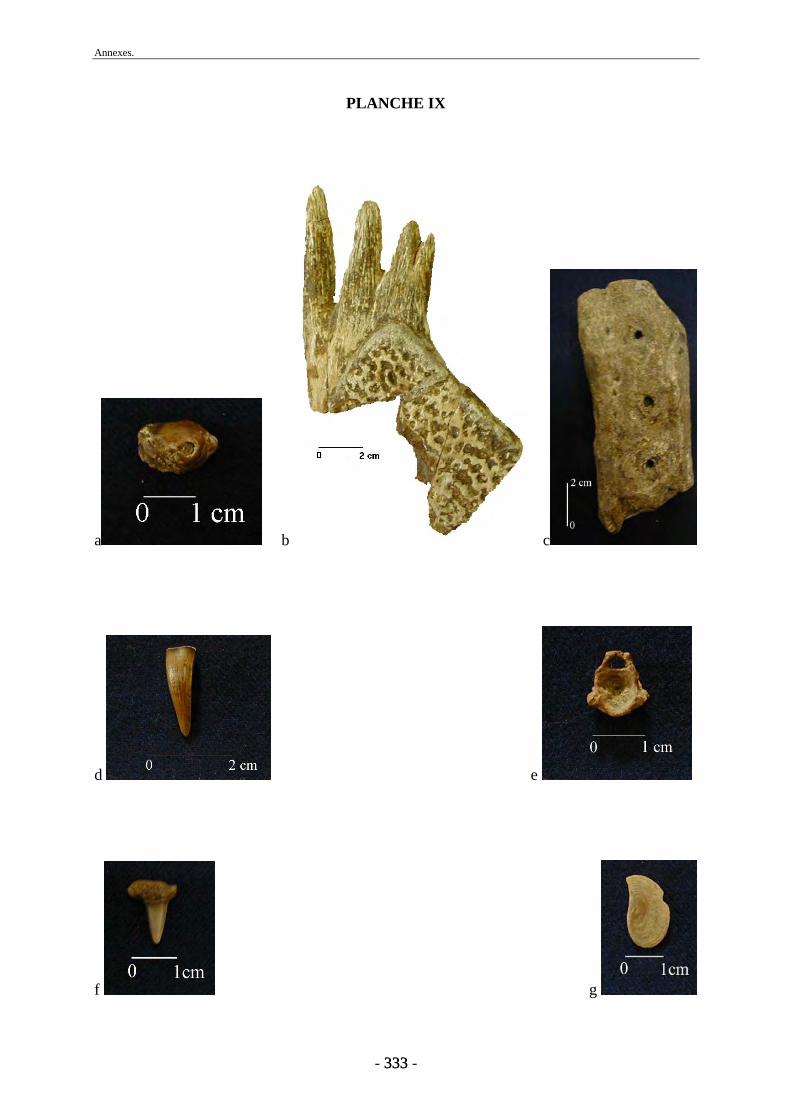
Annexes.
- 333 - 333
PLANCHE IX
a b c
d e
f g
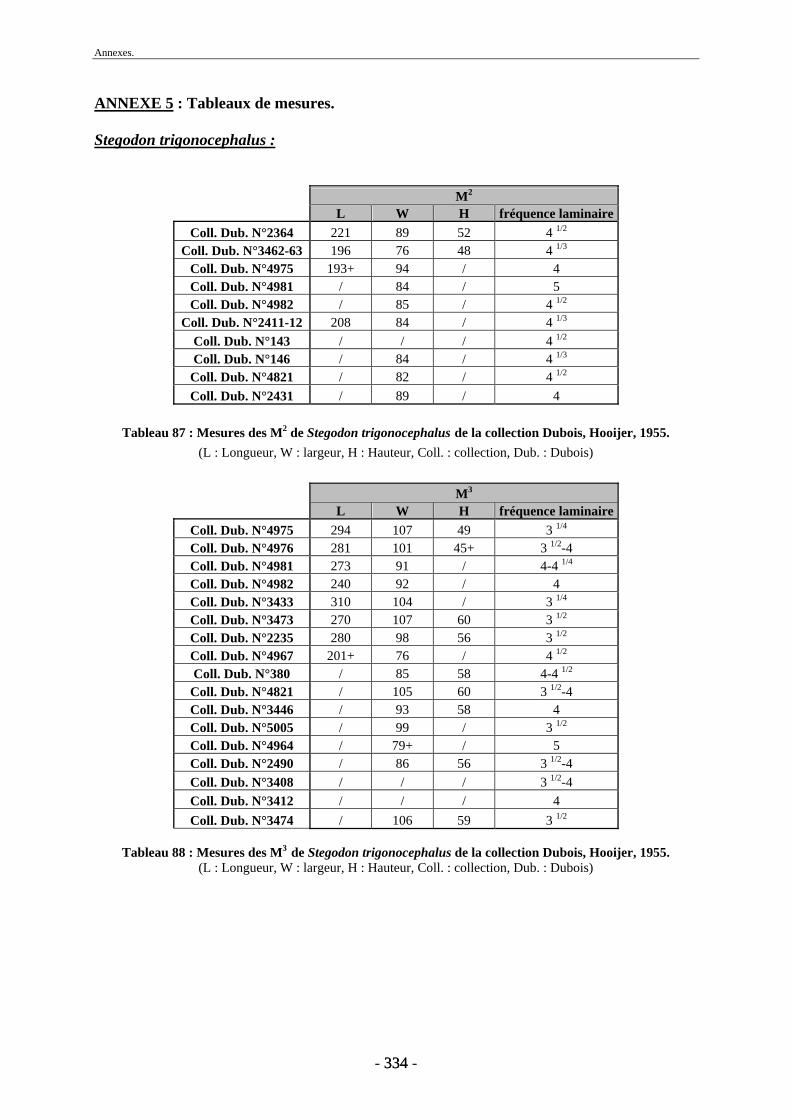
Annexes.
- 334 - 334
ANNEXE 5 : Tableaux de mesures. Stegodon trigonocephalus :
M2 L W H fréquence laminaire
Coll. Dub. N°2364 221 89 52 4 1/2 Coll. Dub. N°3462-63 196 76 48 4 1/3
Coll. Dub. N°4975 193+ 94 / 4 Coll. Dub. N°4981 / 84 / 5 Coll. Dub. N°4982 / 85 / 4 1/2
Coll. Dub. N°2411-12 208 84 / 4 1/3 Coll. Dub. N°143 / / / 4 1/2 Coll. Dub. N°146 / 84 / 4 1/3
Coll. Dub. N°4821 / 82 / 4 1/2 Coll. Dub. N°2431 / 89 / 4
Tableau 87 : Mesures des M2 de Stegodon trigonocephalus de la collection Dubois, Hooijer, 1955.
(L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Dub. : Dubois)
M3 L W H fréquence laminaire
Coll. Dub. N°4975 294 107 49 3 1/4 Coll. Dub. N°4976 281 101 45+ 3 1/2-4 Coll. Dub. N°4981 273 91 / 4-4 1/4 Coll. Dub. N°4982 240 92 / 4 Coll. Dub. N°3433 310 104 / 3 1/4 Coll. Dub. N°3473 270 107 60 3 1/2 Coll. Dub. N°2235 280 98 56 3 1/2 Coll. Dub. N°4967 201+ 76 / 4 1/2 Coll. Dub. N°380 / 85 58 4-4 1/2
Coll. Dub. N°4821 / 105 60 3 1/2-4 Coll. Dub. N°3446 / 93 58 4 Coll. Dub. N°5005 / 99 / 3 1/2 Coll. Dub. N°4964 / 79+ / 5 Coll. Dub. N°2490 / 86 56 3 1/2-4 Coll. Dub. N°3408 / / / 3 1/2-4 Coll. Dub. N°3412 / / / 4 Coll. Dub. N°3474 / 106 59 3 1/2
Tableau 88 : Mesures des M3
de Stegodon trigonocephalus de la collection Dubois, Hooijer, 1955. (L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Dub. : Dubois)
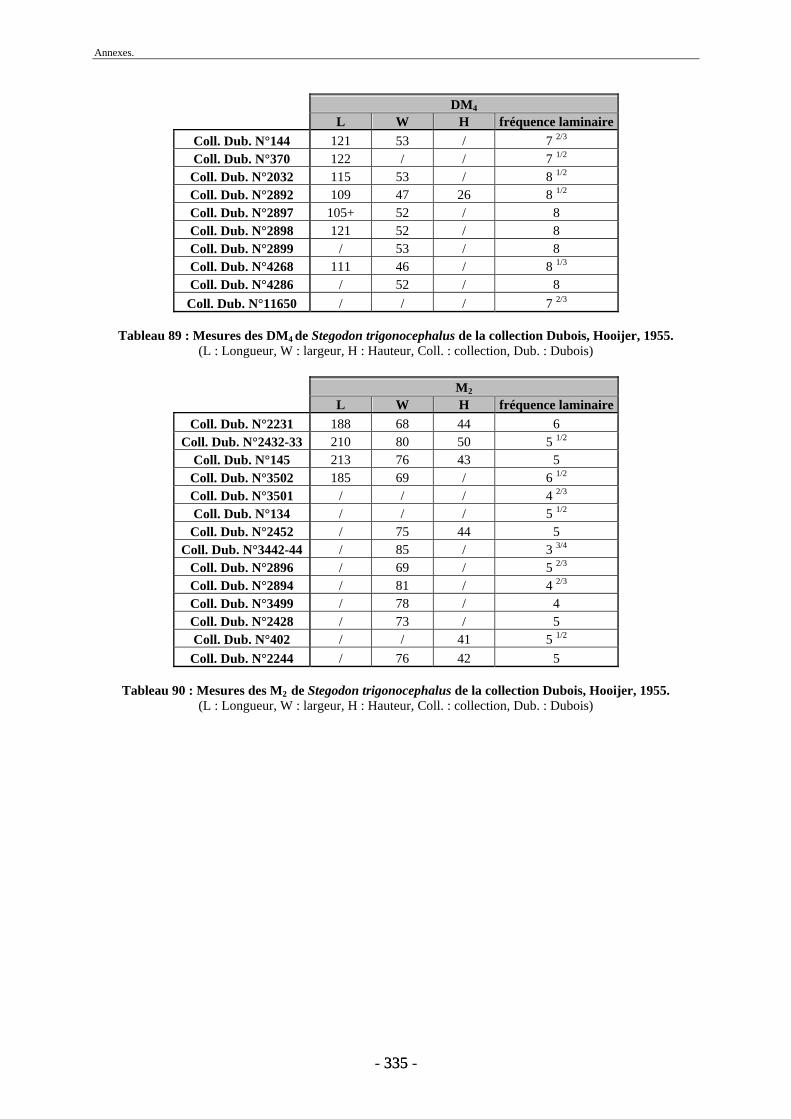
Annexes.
- 335 - 335
DM4 L W H fréquence laminaire
Coll. Dub. N°144 121 53 / 7 2/3 Coll. Dub. N°370 122 / / 7 1/2
Coll. Dub. N°2032 115 53 / 8 1/2 Coll. Dub. N°2892 109 47 26 8 1/2 Coll. Dub. N°2897 105+ 52 / 8 Coll. Dub. N°2898 121 52 / 8 Coll. Dub. N°2899 / 53 / 8 Coll. Dub. N°4268 111 46 / 8 1/3 Coll. Dub. N°4286 / 52 / 8
Coll. Dub. N°11650 / / / 7 2/3
Tableau 89 : Mesures des DM4 de Stegodon trigonocephalus de la collection Dubois, Hooijer, 1955. (L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Dub. : Dubois)
M2 L W H fréquence laminaire
Coll. Dub. N°2231 188 68 44 6 Coll. Dub. N°2432-33 210 80 50 5 1/2
Coll. Dub. N°145 213 76 43 5 Coll. Dub. N°3502 185 69 / 6 1/2 Coll. Dub. N°3501 / / / 4 2/3 Coll. Dub. N°134 / / / 5 1/2
Coll. Dub. N°2452 / 75 44 5 Coll. Dub. N°3442-44 / 85 / 3 3/4
Coll. Dub. N°2896 / 69 / 5 2/3 Coll. Dub. N°2894 / 81 / 4 2/3 Coll. Dub. N°3499 / 78 / 4 Coll. Dub. N°2428 / 73 / 5 Coll. Dub. N°402 / / 41 5 1/2
Coll. Dub. N°2244 / 76 42 5
Tableau 90 : Mesures des M2 de Stegodon trigonocephalus de la collection Dubois, Hooijer, 1955. (L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Dub. : Dubois)
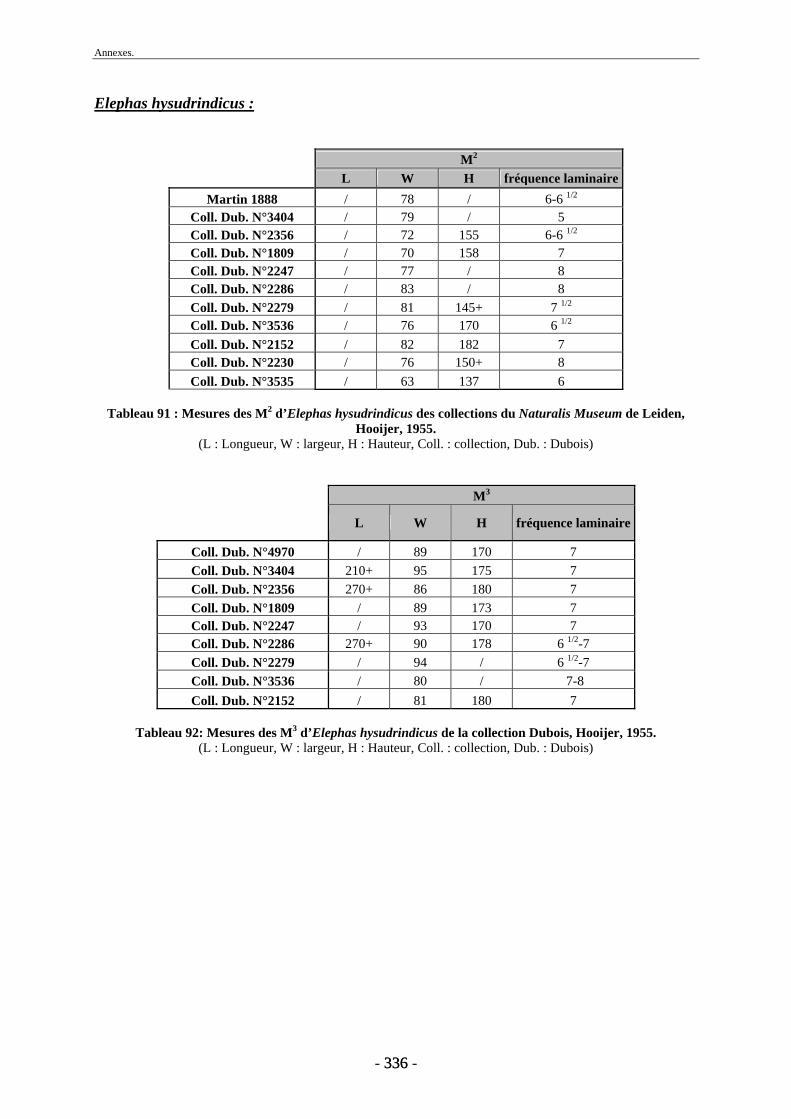
Annexes.
- 336 - 336
Elephas hysudrindicus :
M2 L W H fréquence laminaire
Martin 1888 / 78 / 6-6 1/2 Coll. Dub. N°3404 / 79 / 5 Coll. Dub. N°2356 / 72 155 6-6 1/2 Coll. Dub. N°1809 / 70 158 7 Coll. Dub. N°2247 / 77 / 8 Coll. Dub. N°2286 / 83 / 8 Coll. Dub. N°2279 / 81 145+ 7 1/2 Coll. Dub. N°3536 / 76 170 6 1/2 Coll. Dub. N°2152 / 82 182 7 Coll. Dub. N°2230 / 76 150+ 8 Coll. Dub. N°3535 / 63 137 6
Tableau 91 : Mesures des M2 d’Elephas hysudrindicus des collections du Naturalis Museum de Leiden,
Hooijer, 1955. (L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Dub. : Dubois)
M3
L W H fréquence laminaire
Coll. Dub. N°4970 / 89 170 7 Coll. Dub. N°3404 210+ 95 175 7 Coll. Dub. N°2356 270+ 86 180 7 Coll. Dub. N°1809 / 89 173 7 Coll. Dub. N°2247 / 93 170 7 Coll. Dub. N°2286 270+ 90 178 6 1/2-7 Coll. Dub. N°2279 / 94 / 6 1/2-7 Coll. Dub. N°3536 / 80 / 7-8 Coll. Dub. N°2152 / 81 180 7
Tableau 92: Mesures des M3 d’Elephas hysudrindicus de la collection Dubois, Hooijer, 1955.
(L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Dub. : Dubois)
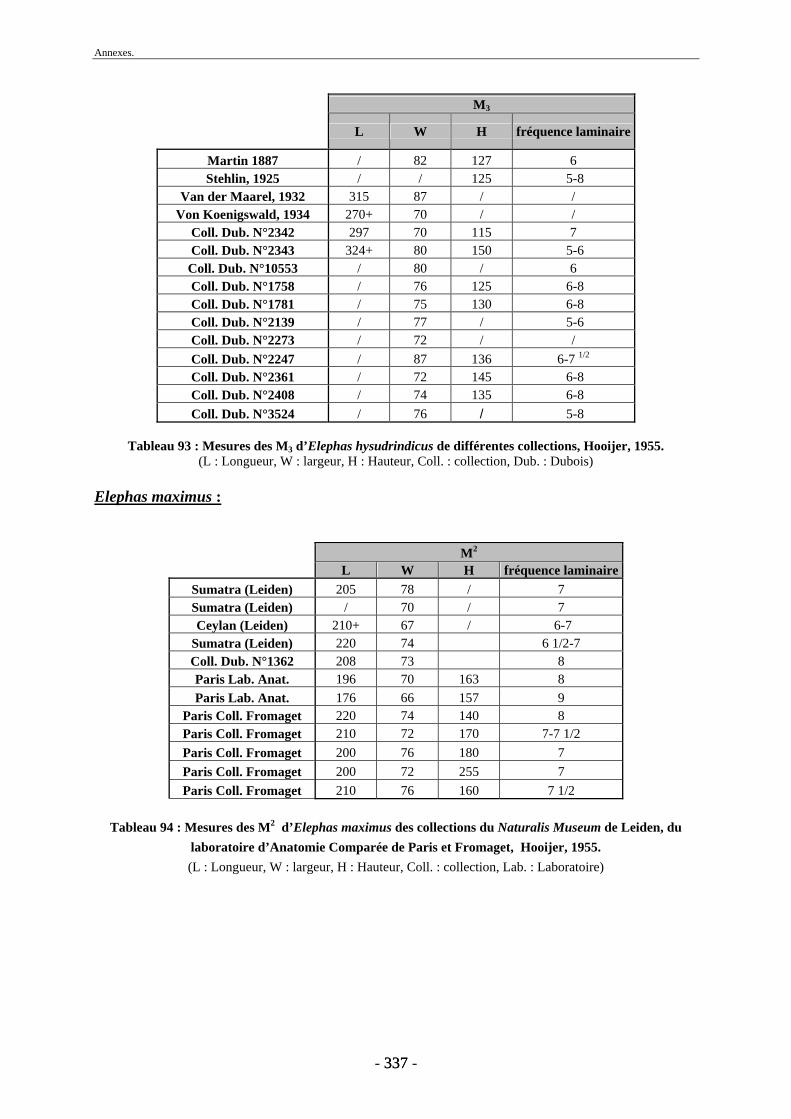
Annexes.
- 337 - 337
M3
L W H fréquence laminaire
Martin 1887 / 82 127 6 Stehlin, 1925 / / 125 5-8
Van der Maarel, 1932 315 87 / / Von Koenigswald, 1934 270+ 70 / /
Coll. Dub. N°2342 297 70 115 7 Coll. Dub. N°2343 324+ 80 150 5-6
Coll. Dub. N°10553 / 80 / 6 Coll. Dub. N°1758 / 76 125 6-8 Coll. Dub. N°1781 / 75 130 6-8 Coll. Dub. N°2139 / 77 / 5-6 Coll. Dub. N°2273 / 72 / / Coll. Dub. N°2247 / 87 136 6-7 1/2 Coll. Dub. N°2361 / 72 145 6-8 Coll. Dub. N°2408 / 74 135 6-8 Coll. Dub. N°3524 / 76 / 5-8
Tableau 93 : Mesures des M3 d’Elephas hysudrindicus de différentes collections, Hooijer, 1955.
(L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Dub. : Dubois) Elephas maximus :
M2 L W H fréquence laminaire
Sumatra (Leiden) 205 78 / 7 Sumatra (Leiden) / 70 / 7 Ceylan (Leiden) 210+ 67 / 6-7
Sumatra (Leiden) 220 74 6 1/2-7 Coll. Dub. N°1362 208 73 8 Paris Lab. Anat. 196 70 163 8 Paris Lab. Anat. 176 66 157 9
Paris Coll. Fromaget 220 74 140 8 Paris Coll. Fromaget 210 72 170 7-7 1/2 Paris Coll. Fromaget 200 76 180 7 Paris Coll. Fromaget 200 72 255 7 Paris Coll. Fromaget 210 76 160 7 1/2
Tableau 94 : Mesures des M2 d’Elephas maximus des collections du Naturalis Museum de Leiden, du
laboratoire d’Anatomie Comparée de Paris et Fromaget, Hooijer, 1955. (L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Lab. : Laboratoire)
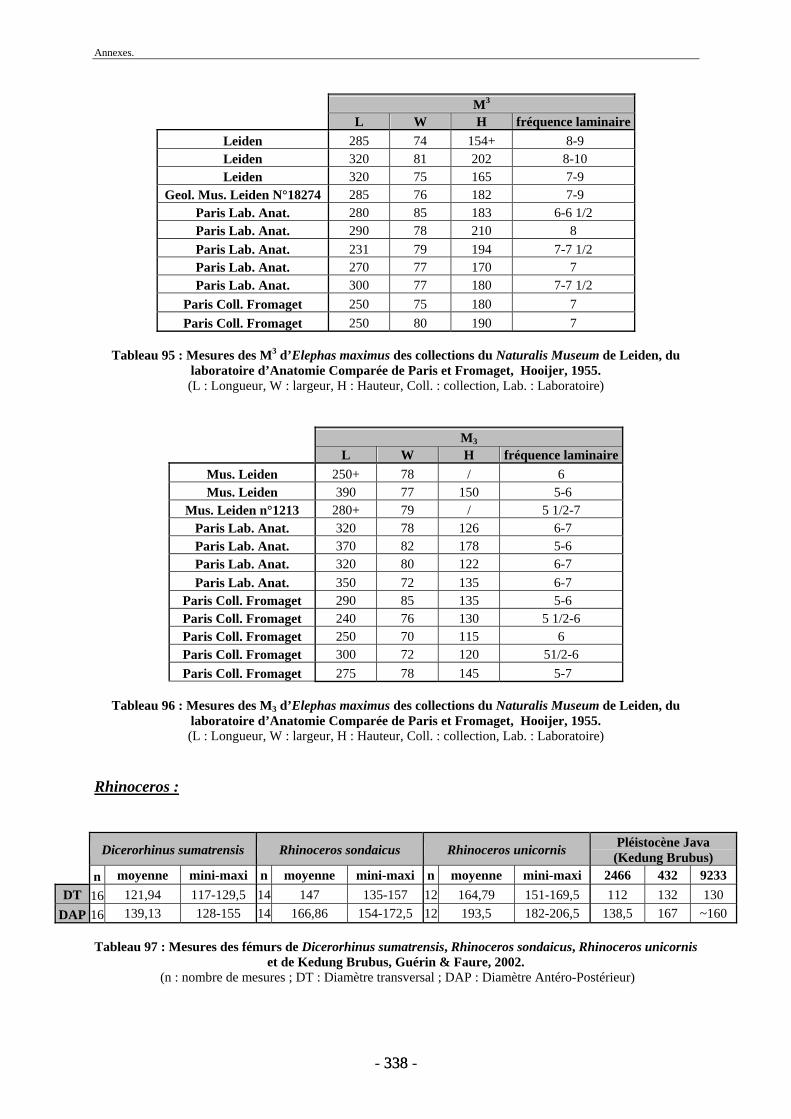
Annexes.
- 338 - 338
M3 L W H fréquence laminaire
Leiden 285 74 154+ 8-9 Leiden 320 81 202 8-10 Leiden 320 75 165 7-9
Geol. Mus. Leiden N°18274 285 76 182 7-9 Paris Lab. Anat. 280 85 183 6-6 1/2 Paris Lab. Anat. 290 78 210 8 Paris Lab. Anat. 231 79 194 7-7 1/2 Paris Lab. Anat. 270 77 170 7 Paris Lab. Anat. 300 77 180 7-7 1/2
Paris Coll. Fromaget 250 75 180 7 Paris Coll. Fromaget 250 80 190 7
Tableau 95 : Mesures des M3 d’Elephas maximus des collections du Naturalis Museum de Leiden, du
laboratoire d’Anatomie Comparée de Paris et Fromaget, Hooijer, 1955. (L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Lab. : Laboratoire)
M3 L W H fréquence laminaire
Mus. Leiden 250+ 78 / 6 Mus. Leiden 390 77 150 5-6
Mus. Leiden n°1213 280+ 79 / 5 1/2-7 Paris Lab. Anat. 320 78 126 6-7 Paris Lab. Anat. 370 82 178 5-6 Paris Lab. Anat. 320 80 122 6-7 Paris Lab. Anat. 350 72 135 6-7
Paris Coll. Fromaget 290 85 135 5-6 Paris Coll. Fromaget 240 76 130 5 1/2-6 Paris Coll. Fromaget 250 70 115 6 Paris Coll. Fromaget 300 72 120 51/2-6 Paris Coll. Fromaget 275 78 145 5-7
Tableau 96 : Mesures des M3 d’Elephas maximus des collections du Naturalis Museum de Leiden, du
laboratoire d’Anatomie Comparée de Paris et Fromaget, Hooijer, 1955. (L : Longueur, W : largeur, H : Hauteur, Coll. : collection, Lab. : Laboratoire)
Rhinoceros :
Dicerorhinus sumatrensis Rhinoceros sondaicus Rhinoceros unicornis Pléistocène Java (Kedung Brubus)
n moyenne mini-maxi n moyenne mini-maxi n moyenne mini-maxi 2466 432 9233 DT 16 121,94 117-129,5 14 147 135-157 12 164,79 151-169,5 112 132 130
DAP 16 139,13 128-155 14 166,86 154-172,5 12 193,5 182-206,5 138,5 167 ~160
Tableau 97 : Mesures des fémurs de Dicerorhinus sumatrensis, Rhinoceros sondaicus, Rhinoceros unicornis et de Kedung Brubus, Guérin & Faure, 2002.
(n : nombre de mesures ; DT : Diamètre transversal ; DAP : Diamètre Antéro-Postérieur)
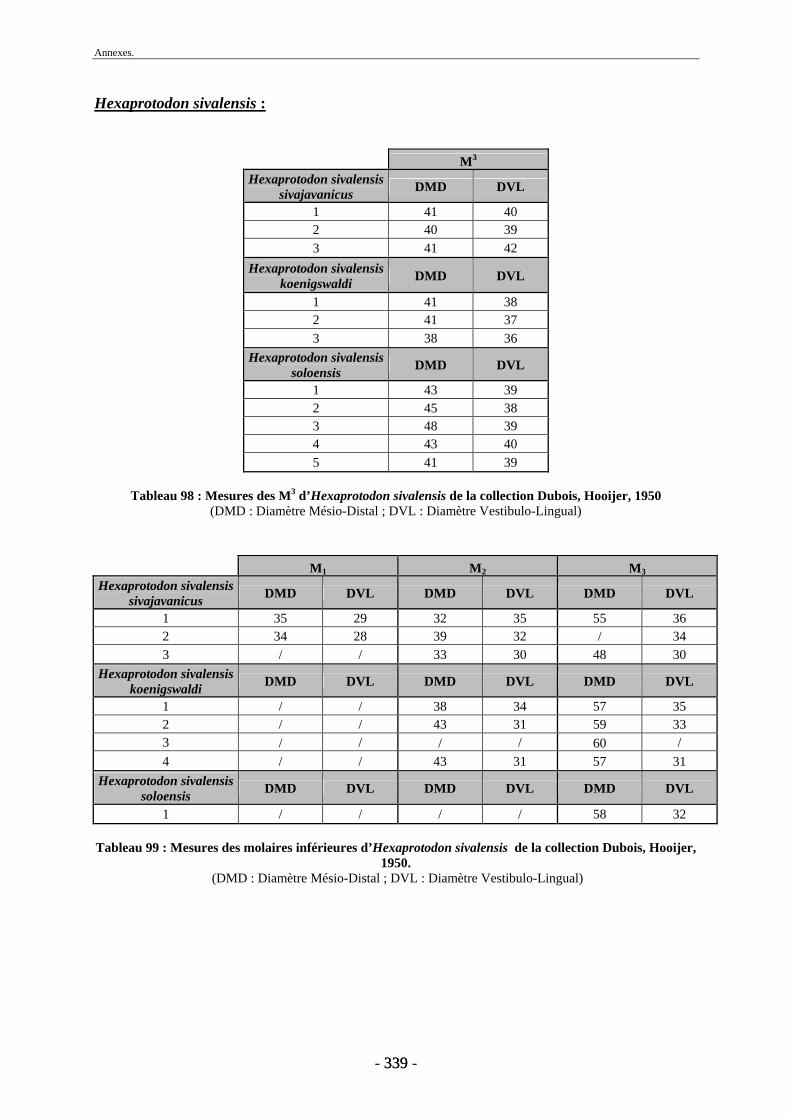
Annexes.
- 339 - 339
Hexaprotodon sivalensis :
M3 Hexaprotodon sivalensis
sivajavanicus DMD DVL
1 41 40 2 40 39 3 41 42
Hexaprotodon sivalensis koenigswaldi DMD DVL
1 41 38 2 41 37 3 38 36
Hexaprotodon sivalensis soloensis DMD DVL
1 43 39 2 45 38 3 48 39 4 43 40 5 41 39
Tableau 98 : Mesures des M3 d’Hexaprotodon sivalensis de la collection Dubois, Hooijer, 1950
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual) M1 M2 M3 Hexaprotodon sivalensis
sivajavanicus DMD DVL DMD DVL DMD DVL
1 35 29 32 35 55 36 2 34 28 39 32 / 34 3 / / 33 30 48 30
Hexaprotodon sivalensis koenigswaldi DMD DVL DMD DVL DMD DVL
1 / / 38 34 57 35 2 / / 43 31 59 33 3 / / / / 60 / 4 / / 43 31 57 31
Hexaprotodon sivalensis soloensis DMD DVL DMD DVL DMD DVL
1 / / / / 58 32
Tableau 99 : Mesures des molaires inférieures d’Hexaprotodon sivalensis de la collection Dubois, Hooijer, 1950.
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
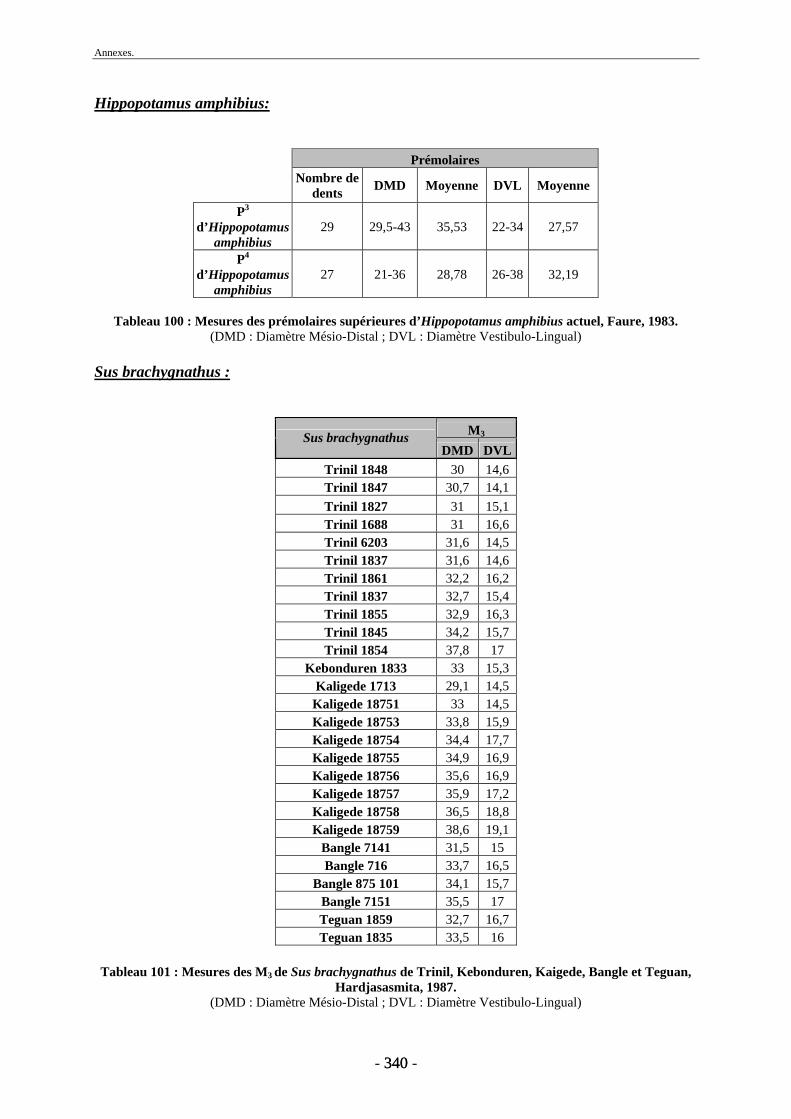
Annexes.
- 340 - 340
Hippopotamus amphibius:
Prémolaires
Nombre de dents DMD Moyenne DVL Moyenne
P3
d’Hippopotamus amphibius
29 29,5-43 35,53 22-34 27,57
P4
d’Hippopotamus amphibius
27 21-36 28,78 26-38 32,19
Tableau 100 : Mesures des prémolaires supérieures d’Hippopotamus amphibius actuel, Faure, 1983.
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual) Sus brachygnathus :
M3 Sus brachygnathus DMD DVL
Trinil 1848 30 14,6 Trinil 1847 30,7 14,1 Trinil 1827 31 15,1 Trinil 1688 31 16,6 Trinil 6203 31,6 14,5 Trinil 1837 31,6 14,6 Trinil 1861 32,2 16,2 Trinil 1837 32,7 15,4 Trinil 1855 32,9 16,3 Trinil 1845 34,2 15,7 Trinil 1854 37,8 17
Kebonduren 1833 33 15,3 Kaligede 1713 29,1 14,5
Kaligede 18751 33 14,5 Kaligede 18753 33,8 15,9 Kaligede 18754 34,4 17,7 Kaligede 18755 34,9 16,9 Kaligede 18756 35,6 16,9 Kaligede 18757 35,9 17,2 Kaligede 18758 36,5 18,8 Kaligede 18759 38,6 19,1
Bangle 7141 31,5 15 Bangle 716 33,7 16,5
Bangle 875 101 34,1 15,7 Bangle 7151 35,5 17 Teguan 1859 32,7 16,7 Teguan 1835 33,5 16
Tableau 101 : Mesures des M3 de Sus brachygnathus de Trinil, Kebonduren, Kaigede, Bangle et Teguan,
Hardjasasmita, 1987. (DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
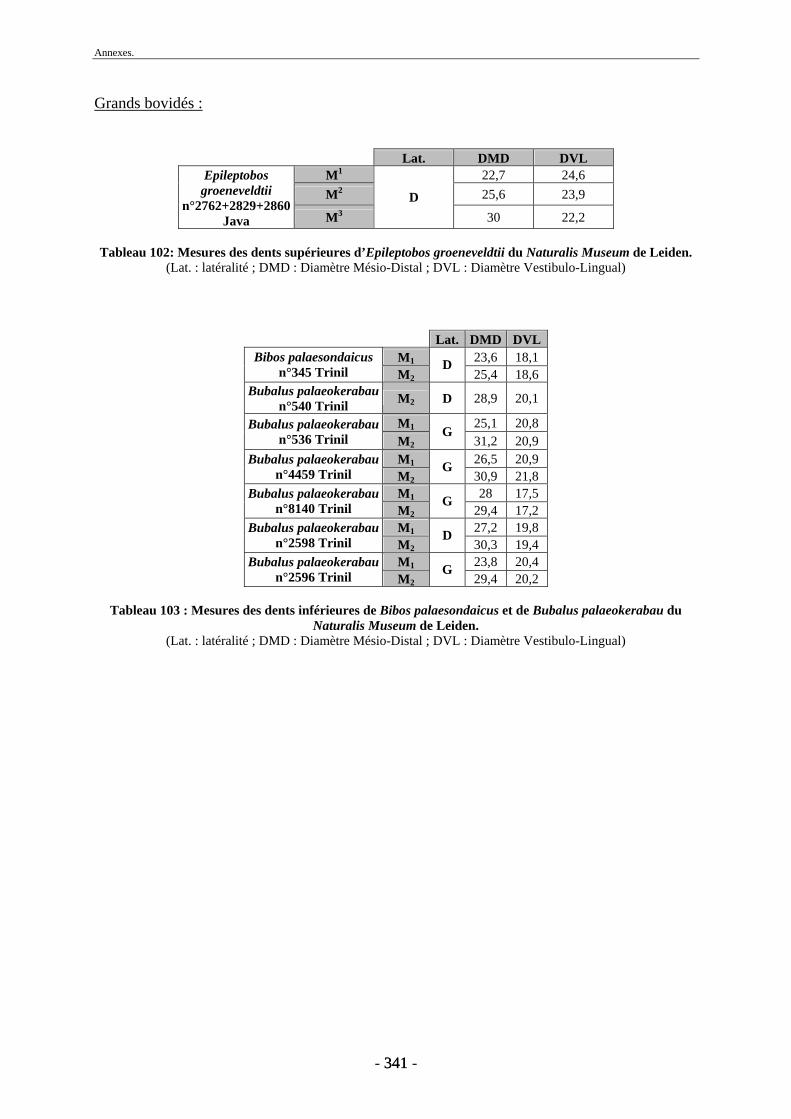
Annexes.
- 341 - 341
Grands bovidés :
Lat. DMD DVL M1 22,7 24,6 M2 25,6 23,9
Epileptobos groeneveldtii
n°2762+2829+2860 Java M3
D 30 22,2
Tableau 102: Mesures des dents supérieures d’Epileptobos groeneveldtii du Naturalis Museum de Leiden.
(Lat. : latéralité ; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
Lat. DMD DVLM1 23,6 18,1 Bibos palaesondaicus
n°345 Trinil M2 D
25,4 18,6 Bubalus palaeokerabau
n°540 Trinil M2 D 28,9 20,1
M1 25,1 20,8 Bubalus palaeokerabau n°536 Trinil M2
G 31,2 20,9
M1 26,5 20,9 Bubalus palaeokerabau n°4459 Trinil M2
G 30,9 21,8
M1 28 17,5 Bubalus palaeokerabau n°8140 Trinil M2
G 29,4 17,2
M1 27,2 19,8 Bubalus palaeokerabau n°2598 Trinil M2
D 30,3 19,4
M1 23,8 20,4 Bubalus palaeokerabau n°2596 Trinil M2
G 29,4 20,2
Tableau 103 : Mesures des dents inférieures de Bibos palaesondaicus et de Bubalus palaeokerabau du
Naturalis Museum de Leiden. (Lat. : latéralité ; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
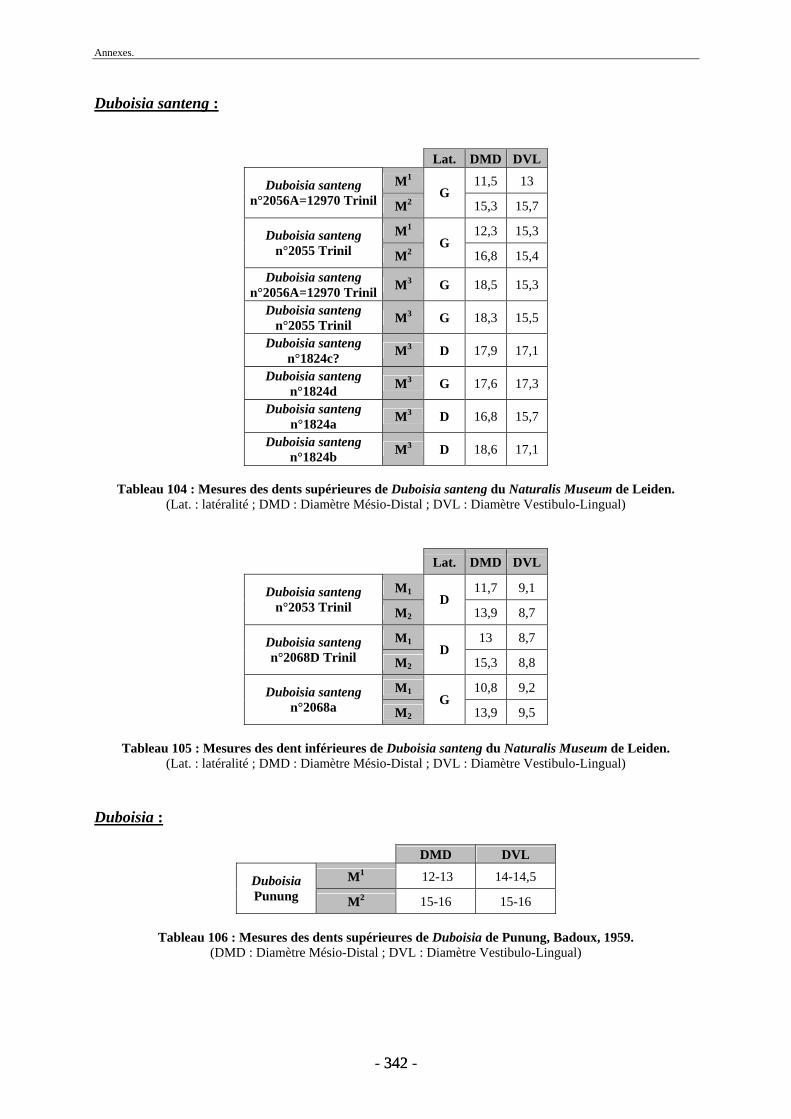
Annexes.
- 342 - 342
Duboisia santeng :
Lat. DMD DVL
M1 11,5 13 Duboisia santeng n°2056A=12970 Trinil M2
G 15,3 15,7
M1 12,3 15,3 Duboisia santeng n°2055 Trinil M2
G 16,8 15,4
Duboisia santeng n°2056A=12970 Trinil M3 G 18,5 15,3
Duboisia santeng n°2055 Trinil M3 G 18,3 15,5
Duboisia santeng n°1824c? M3 D 17,9 17,1
Duboisia santeng n°1824d M3 G 17,6 17,3
Duboisia santeng n°1824a M3 D 16,8 15,7
Duboisia santeng n°1824b M3 D 18,6 17,1
Tableau 104 : Mesures des dents supérieures de Duboisia santeng du Naturalis Museum de Leiden.
(Lat. : latéralité ; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
Lat. DMD DVL
M1 11,7 9,1 Duboisia santeng n°2053 Trinil M2
D 13,9 8,7
M1 13 8,7 Duboisia santeng n°2068D Trinil M2
D 15,3 8,8
M1 10,8 9,2 Duboisia santeng n°2068a M2
G 13,9 9,5
Tableau 105 : Mesures des dent inférieures de Duboisia santeng du Naturalis Museum de Leiden.
(Lat. : latéralité ; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual) Duboisia :
DMD DVL
M1 12-13 14-14,5 Duboisia Punung M2 15-16 15-16
Tableau 106 : Mesures des dents supérieures de Duboisia de Punung, Badoux, 1959.
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
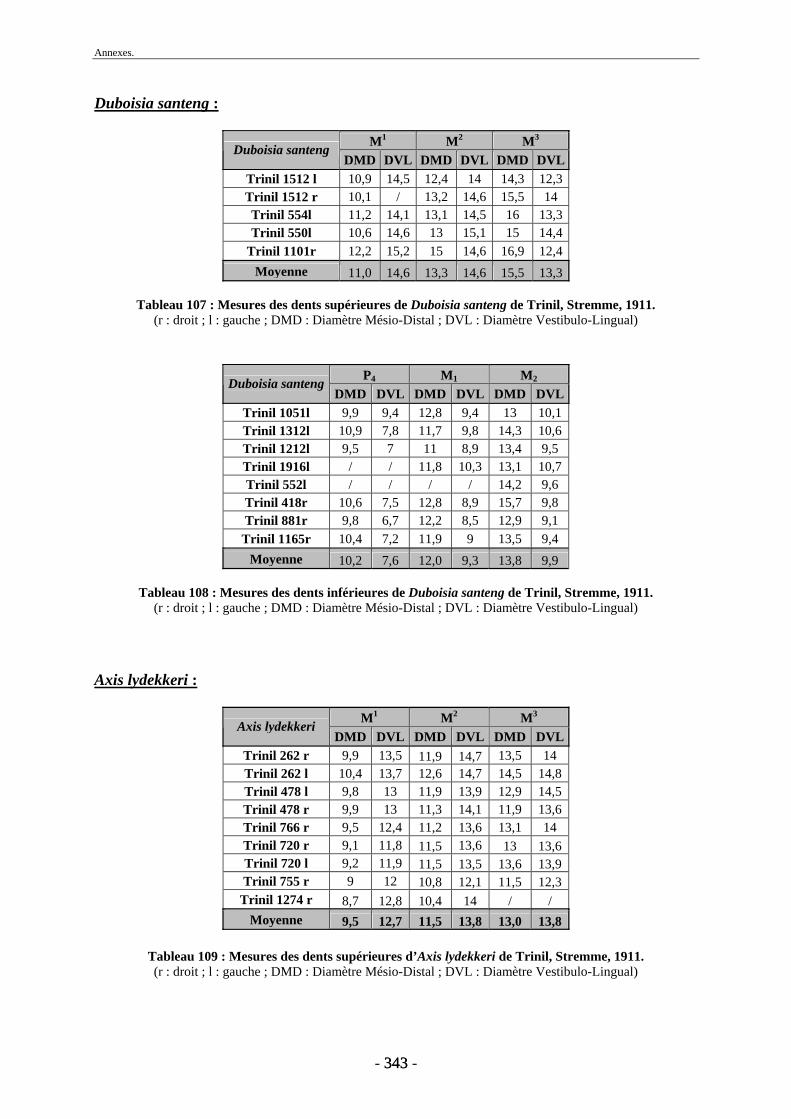
Annexes.
- 343 - 343
Duboisia santeng :
M1 M2 M3 Duboisia santeng DMD DVL DMD DVL DMD DVL
Trinil 1512 l 10,9 14,5 12,4 14 14,3 12,3 Trinil 1512 r 10,1 / 13,2 14,6 15,5 14 Trinil 554l 11,2 14,1 13,1 14,5 16 13,3 Trinil 550l 10,6 14,6 13 15,1 15 14,4
Trinil 1101r 12,2 15,2 15 14,6 16,9 12,4 Moyenne 11,0 14,6 13,3 14,6 15,5 13,3
Tableau 107 : Mesures des dents supérieures de Duboisia santeng de Trinil, Stremme, 1911.
(r : droit ; l : gauche ; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
P4 M1 M2 Duboisia santeng DMD DVL DMD DVL DMD DVL
Trinil 1051l 9,9 9,4 12,8 9,4 13 10,1 Trinil 1312l 10,9 7,8 11,7 9,8 14,3 10,6 Trinil 1212l 9,5 7 11 8,9 13,4 9,5 Trinil 1916l / / 11,8 10,3 13,1 10,7 Trinil 552l / / / / 14,2 9,6 Trinil 418r 10,6 7,5 12,8 8,9 15,7 9,8 Trinil 881r 9,8 6,7 12,2 8,5 12,9 9,1
Trinil 1165r 10,4 7,2 11,9 9 13,5 9,4 Moyenne 10,2 7,6 12,0 9,3 13,8 9,9
Tableau 108 : Mesures des dents inférieures de Duboisia santeng de Trinil, Stremme, 1911.
(r : droit ; l : gauche ; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual) Axis lydekkeri :
M1 M2 M3 Axis lydekkeri DMD DVL DMD DVL DMD DVL
Trinil 262 r 9,9 13,5 11,9 14,7 13,5 14 Trinil 262 l 10,4 13,7 12,6 14,7 14,5 14,8 Trinil 478 l 9,8 13 11,9 13,9 12,9 14,5 Trinil 478 r 9,9 13 11,3 14,1 11,9 13,6 Trinil 766 r 9,5 12,4 11,2 13,6 13,1 14 Trinil 720 r 9,1 11,8 11,5 13,6 13 13,6 Trinil 720 l 9,2 11,9 11,5 13,5 13,6 13,9 Trinil 755 r 9 12 10,8 12,1 11,5 12,3
Trinil 1274 r 8,7 12,8 10,4 14 / / Moyenne 9,5 12,7 11,5 13,8 13,0 13,8
Tableau 109 : Mesures des dents supérieures d’Axis lydekkeri de Trinil, Stremme, 1911. (r : droit ; l : gauche ; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
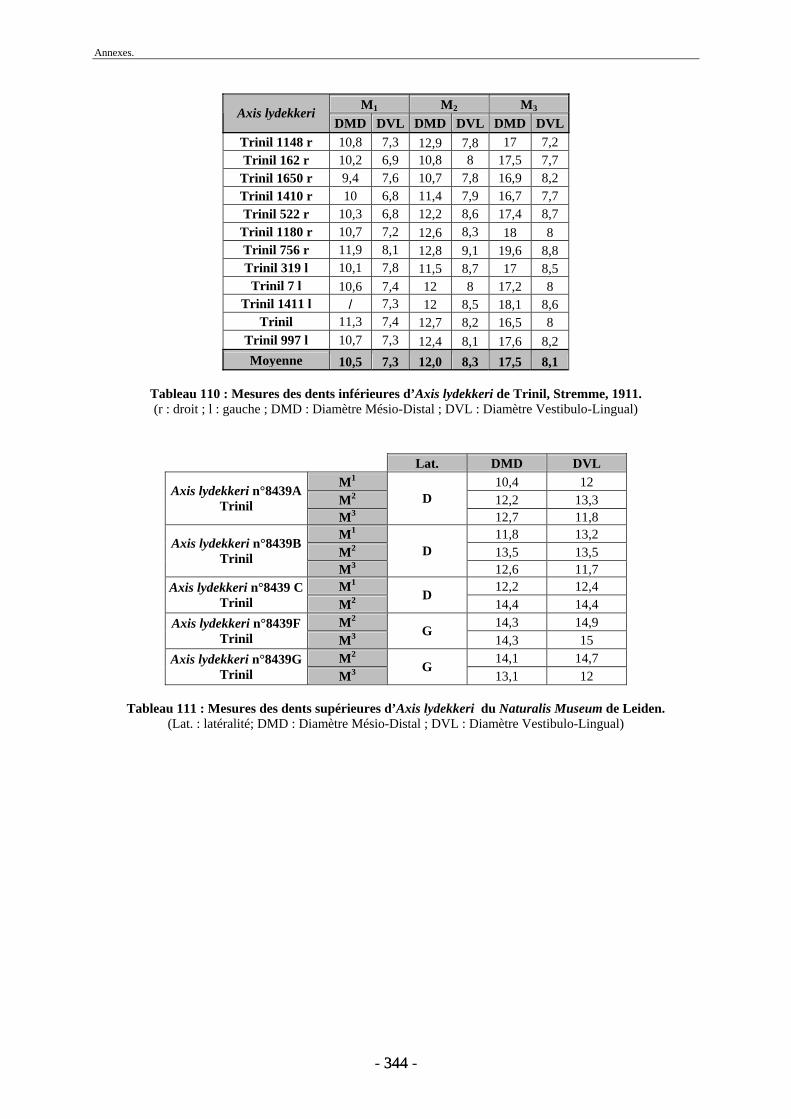
Annexes.
- 344 - 344
M1 M2 M3 Axis lydekkeri DMD DVL DMD DVL DMD DVL
Trinil 1148 r 10,8 7,3 12,9 7,8 17 7,2 Trinil 162 r 10,2 6,9 10,8 8 17,5 7,7
Trinil 1650 r 9,4 7,6 10,7 7,8 16,9 8,2 Trinil 1410 r 10 6,8 11,4 7,9 16,7 7,7 Trinil 522 r 10,3 6,8 12,2 8,6 17,4 8,7
Trinil 1180 r 10,7 7,2 12,6 8,3 18 8 Trinil 756 r 11,9 8,1 12,8 9,1 19,6 8,8 Trinil 319 l 10,1 7,8 11,5 8,7 17 8,5
Trinil 7 l 10,6 7,4 12 8 17,2 8 Trinil 1411 l / 7,3 12 8,5 18,1 8,6
Trinil 11,3 7,4 12,7 8,2 16,5 8 Trinil 997 l 10,7 7,3 12,4 8,1 17,6 8,2 Moyenne 10,5 7,3 12,0 8,3 17,5 8,1
Tableau 110 : Mesures des dents inférieures d’Axis lydekkeri de Trinil, Stremme, 1911. (r : droit ; l : gauche ; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
Lat. DMD DVL M1 10,4 12 M2 12,2 13,3
Axis lydekkeri n°8439A Trinil
M3 D
12,7 11,8 M1 11,8 13,2 M2 13,5 13,5
Axis lydekkeri n°8439B Trinil
M3 D
12,6 11,7 M1 12,2 12,4 Axis lydekkeri n°8439 C
Trinil M2 D
14,4 14,4 M2 14,3 14,9 Axis lydekkeri n°8439F
Trinil M3 G
14,3 15 M2 14,1 14,7 Axis lydekkeri n°8439G
Trinil M3 G
13,1 12
Tableau 111 : Mesures des dents supérieures d’Axis lydekkeri du Naturalis Museum de Leiden. (Lat. : latéralité; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
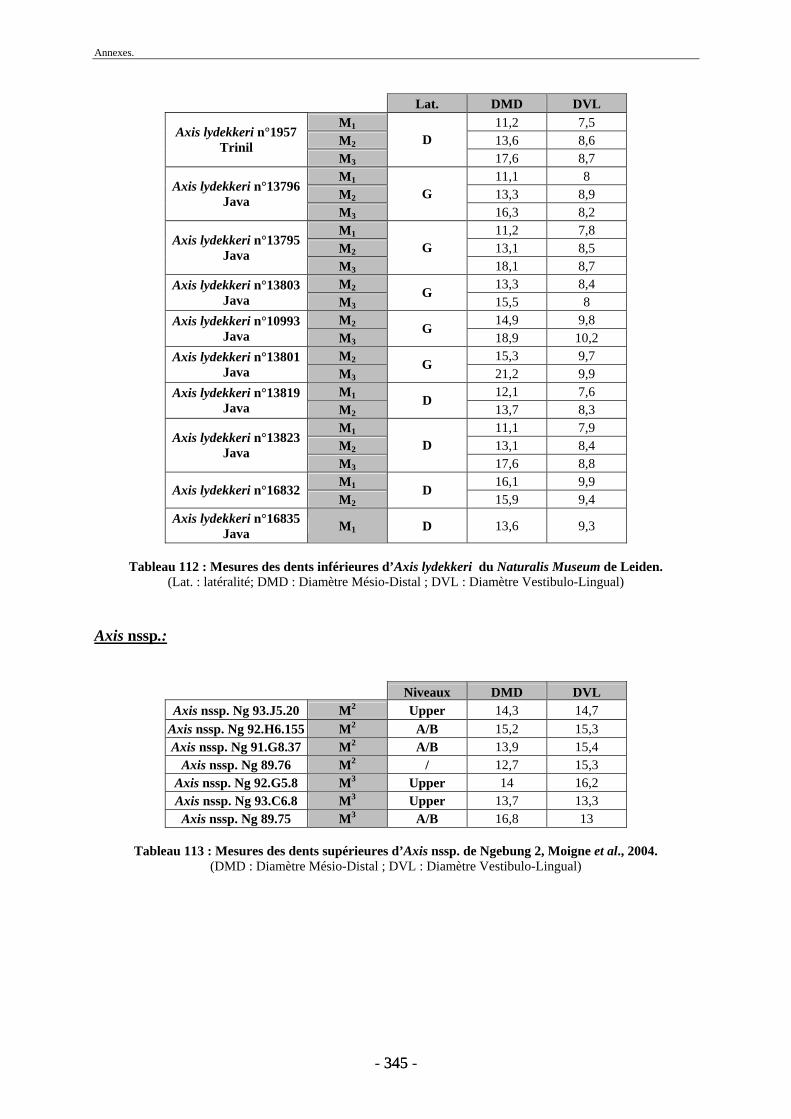
Annexes.
- 345 - 345
Lat. DMD DVL M1 11,2 7,5 M2 13,6 8,6 Axis lydekkeri n°1957
Trinil M3
D 17,6 8,7
M1 11,1 8 M2 13,3 8,9 Axis lydekkeri n°13796
Java M3
G 16,3 8,2
M1 11,2 7,8 M2 13,1 8,5 Axis lydekkeri n°13795
Java M3
G 18,1 8,7
M2 13,3 8,4 Axis lydekkeri n°13803 Java M3
G 15,5 8
M2 14,9 9,8 Axis lydekkeri n°10993 Java M3
G 18,9 10,2
M2 15,3 9,7 Axis lydekkeri n°13801 Java M3
G 21,2 9,9
M1 12,1 7,6 Axis lydekkeri n°13819 Java M2
D 13,7 8,3
M1 11,1 7,9 M2 13,1 8,4 Axis lydekkeri n°13823
Java M3
D 17,6 8,8
M1 16,1 9,9 Axis lydekkeri n°16832 M2
D 15,9 9,4
Axis lydekkeri n°16835 Java M1 D 13,6 9,3
Tableau 112 : Mesures des dents inférieures d’Axis lydekkeri du Naturalis Museum de Leiden.
(Lat. : latéralité; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual) Axis nssp.:
Niveaux DMD DVL Axis nssp. Ng 93.J5.20 M2 Upper 14,3 14,7
Axis nssp. Ng 92.H6.155 M2 A/B 15,2 15,3 Axis nssp. Ng 91.G8.37 M2 A/B 13,9 15,4
Axis nssp. Ng 89.76 M2 / 12,7 15,3 Axis nssp. Ng 92.G5.8 M3 Upper 14 16,2 Axis nssp. Ng 93.C6.8 M3 Upper 13,7 13,3 Axis nssp. Ng 89.75 M3 A/B 16,8 13
Tableau 113 : Mesures des dents supérieures d’Axis nssp. de Ngebung 2, Moigne et al., 2004.
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
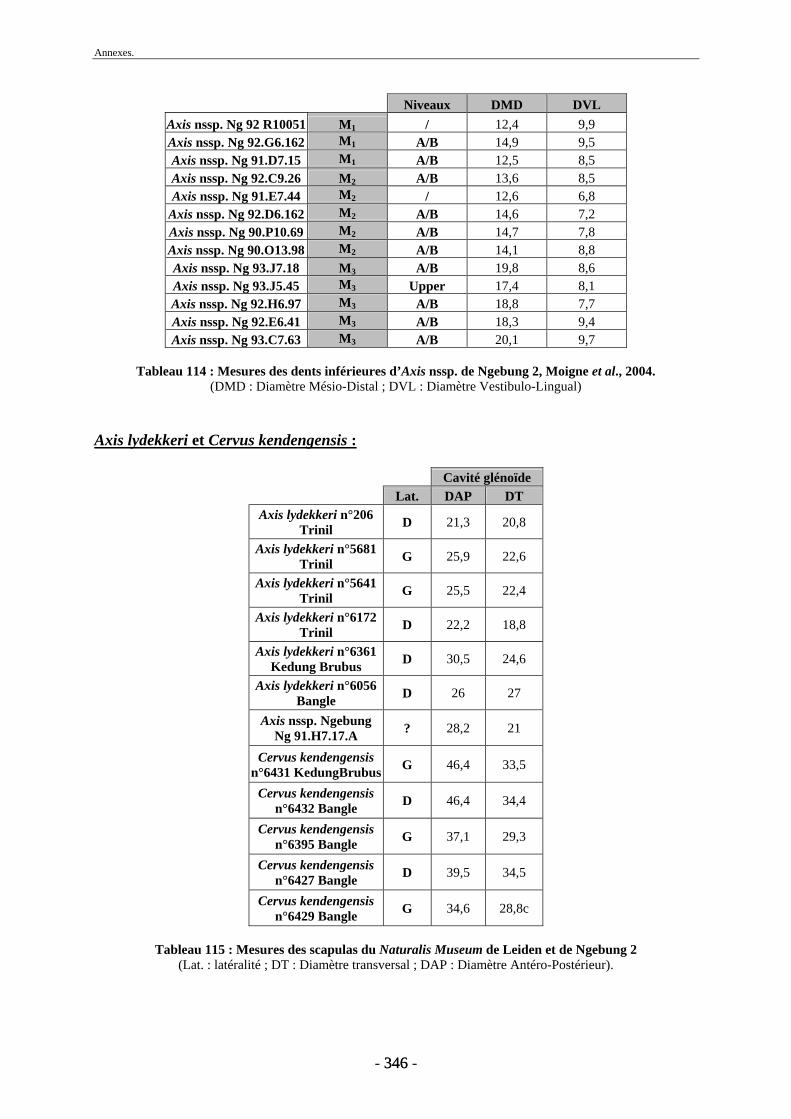
Annexes.
- 346 - 346
Niveaux DMD DVL Axis nssp. Ng 92 R10051 M1 / 12,4 9,9 Axis nssp. Ng 92.G6.162 M1 A/B 14,9 9,5 Axis nssp. Ng 91.D7.15 M1 A/B 12,5 8,5 Axis nssp. Ng 92.C9.26 M2 A/B 13,6 8,5 Axis nssp. Ng 91.E7.44 M2 / 12,6 6,8
Axis nssp. Ng 92.D6.162 M2 A/B 14,6 7,2 Axis nssp. Ng 90.P10.69 M2 A/B 14,7 7,8 Axis nssp. Ng 90.O13.98 M2 A/B 14,1 8,8 Axis nssp. Ng 93.J7.18 M3 A/B 19,8 8,6 Axis nssp. Ng 93.J5.45 M3 Upper 17,4 8,1 Axis nssp. Ng 92.H6.97 M3 A/B 18,8 7,7 Axis nssp. Ng 92.E6.41 M3 A/B 18,3 9,4 Axis nssp. Ng 93.C7.63 M3 A/B 20,1 9,7
Tableau 114 : Mesures des dents inférieures d’Axis nssp. de Ngebung 2, Moigne et al., 2004.
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
Axis lydekkeri et Cervus kendengensis :
Cavité glénoïde Lat. DAP DT
Axis lydekkeri n°206 Trinil D 21,3 20,8
Axis lydekkeri n°5681 Trinil G 25,9 22,6
Axis lydekkeri n°5641 Trinil G 25,5 22,4
Axis lydekkeri n°6172 Trinil D 22,2 18,8
Axis lydekkeri n°6361 Kedung Brubus D 30,5 24,6
Axis lydekkeri n°6056 Bangle D 26 27
Axis nssp. Ngebung Ng 91.H7.17.A ? 28,2 21
Cervus kendengensis n°6431 KedungBrubus G 46,4 33,5
Cervus kendengensis n°6432 Bangle D 46,4 34,4
Cervus kendengensis n°6395 Bangle G 37,1 29,3
Cervus kendengensis n°6427 Bangle D 39,5 34,5
Cervus kendengensis n°6429 Bangle G 34,6 28,8c
Tableau 115 : Mesures des scapulas du Naturalis Museum de Leiden et de Ngebung 2
(Lat. : latéralité ; DT : Diamètre transversal ; DAP : Diamètre Antéro-Postérieur).
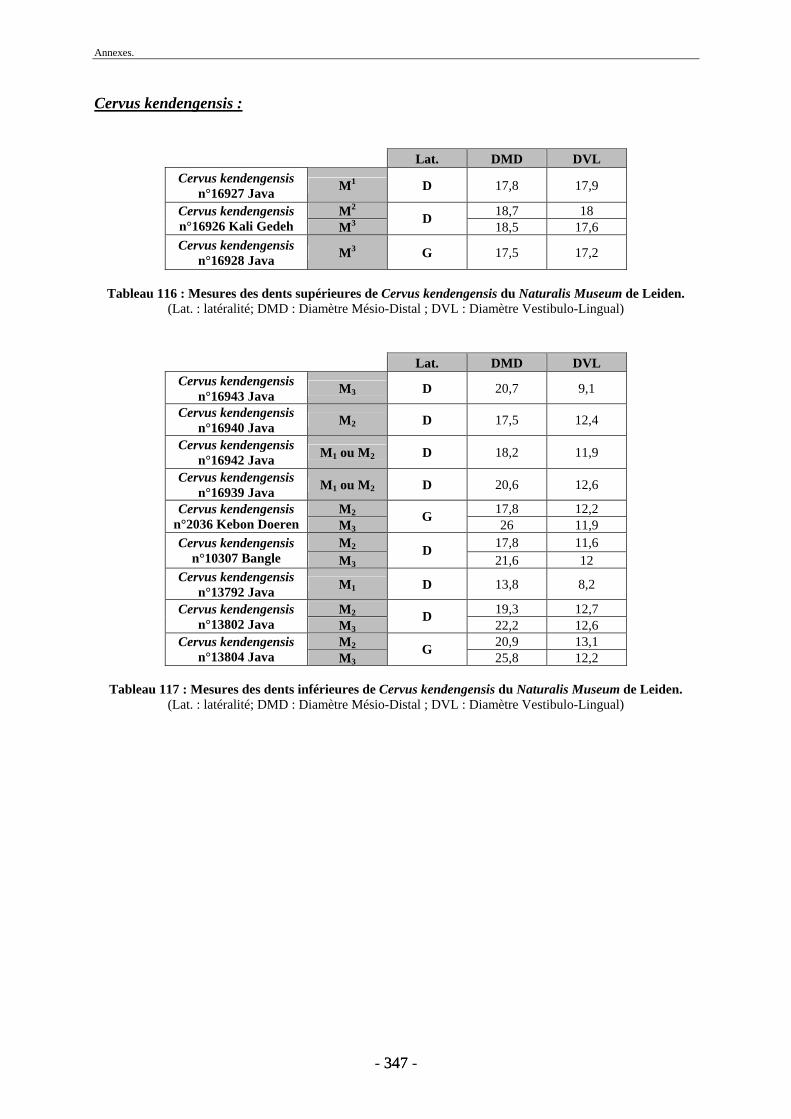
Annexes.
- 347 - 347
Cervus kendengensis :
Lat. DMD DVL Cervus kendengensis
n°16927 Java M1 D 17,8 17,9
M2 18,7 18 Cervus kendengensis n°16926 Kali Gedeh M3 D 18,5 17,6 Cervus kendengensis
n°16928 Java M3 G 17,5 17,2
Tableau 116 : Mesures des dents supérieures de Cervus kendengensis du Naturalis Museum de Leiden.
(Lat. : latéralité; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
Lat. DMD DVL Cervus kendengensis
n°16943 Java M3 D 20,7 9,1
Cervus kendengensis n°16940 Java M2 D 17,5 12,4
Cervus kendengensis n°16942 Java M1 ou M2 D 18,2 11,9
Cervus kendengensis n°16939 Java M1 ou M2 D 20,6 12,6
M2 17,8 12,2 Cervus kendengensis n°2036 Kebon Doeren M3
G 26 11,9 M2 17,8 11,6 Cervus kendengensis
n°10307 Bangle M3 D
21,6 12 Cervus kendengensis
n°13792 Java M1 D 13,8 8,2
M2 19,3 12,7 Cervus kendengensis n°13802 Java M3
D 22,2 12,6
M2 20,9 13,1 Cervus kendengensis n°13804 Java M3
G 25,8 12,2
Tableau 117 : Mesures des dents inférieures de Cervus kendengensis du Naturalis Museum de Leiden. (Lat. : latéralité; DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
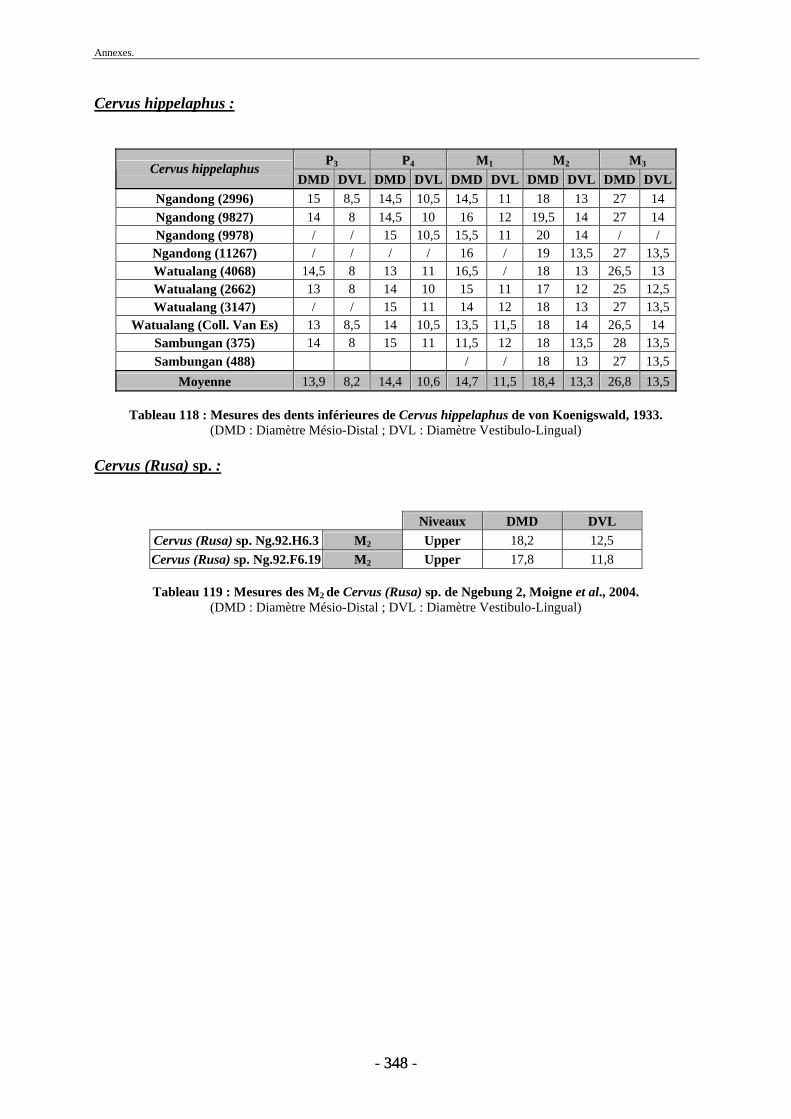
Annexes.
- 348 - 348
Cervus hippelaphus :
P3 P4 M1 M2 M3 Cervus hippelaphus DMD DVL DMD DVL DMD DVL DMD DVL DMD DVL
Ngandong (2996) 15 8,5 14,5 10,5 14,5 11 18 13 27 14 Ngandong (9827) 14 8 14,5 10 16 12 19,5 14 27 14 Ngandong (9978) / / 15 10,5 15,5 11 20 14 / /
Ngandong (11267) / / / / 16 / 19 13,5 27 13,5Watualang (4068) 14,5 8 13 11 16,5 / 18 13 26,5 13 Watualang (2662) 13 8 14 10 15 11 17 12 25 12,5Watualang (3147) / / 15 11 14 12 18 13 27 13,5
Watualang (Coll. Van Es) 13 8,5 14 10,5 13,5 11,5 18 14 26,5 14 Sambungan (375) 14 8 15 11 11,5 12 18 13,5 28 13,5Sambungan (488) / / 18 13 27 13,5
Moyenne 13,9 8,2 14,4 10,6 14,7 11,5 18,4 13,3 26,8 13,5
Tableau 118 : Mesures des dents inférieures de Cervus hippelaphus de von Koenigswald, 1933. (DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
Cervus (Rusa) sp. :
Niveaux DMD DVL Cervus (Rusa) sp. Ng.92.H6.3 M2 Upper 18,2 12,5 Cervus (Rusa) sp. Ng.92.F6.19 M2 Upper 17,8 11,8
Tableau 119 : Mesures des M2 de Cervus (Rusa) sp. de Ngebung 2, Moigne et al., 2004.
(DMD : Diamètre Mésio-Distal ; DVL : Diamètre Vestibulo-Lingual)
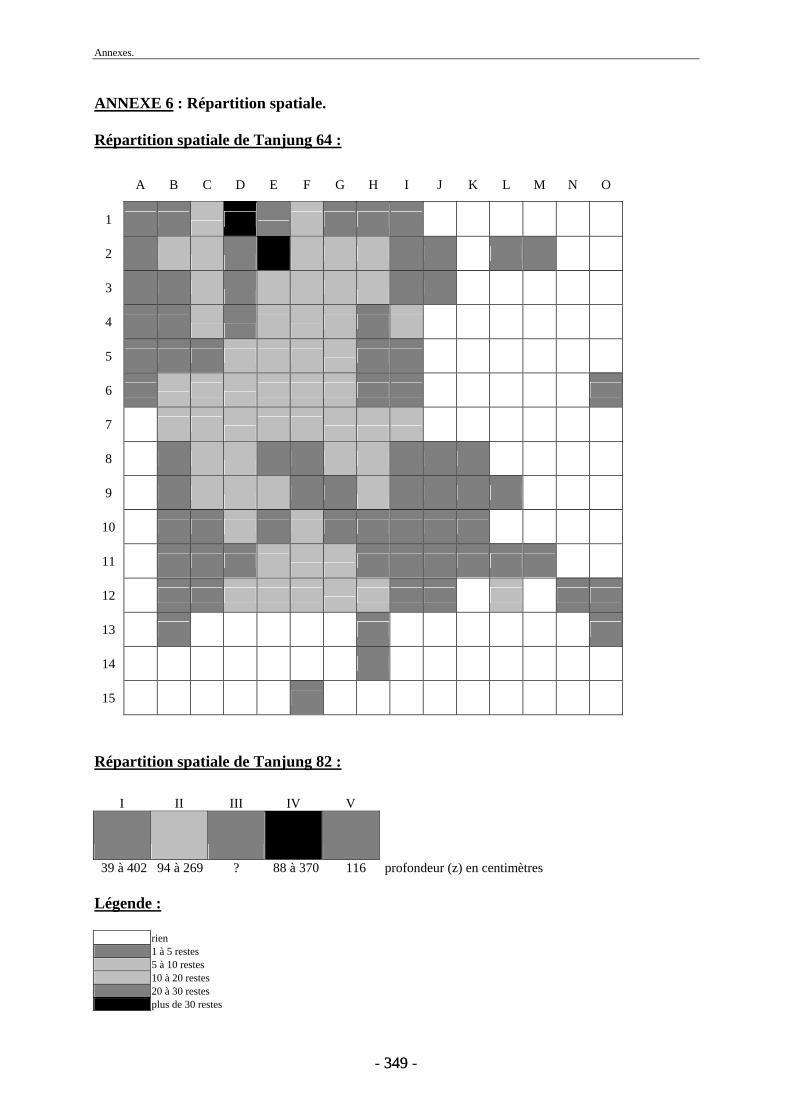
Annexes.
- 349 - 349
ANNEXE 6 : Répartition spatiale. Répartition spatiale de Tanjung 64 :
A B C D E F G H I J K L M N O
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
Répartition spatiale de Tanjung 82 :
I II III IV V
39 à 402 94 à 269 ? 88 à 370 116 profondeur (z) en centimètres Légende : rien 1 à 5 restes 5 à 10 restes 10 à 20 restes 20 à 30 restes plus de 30 restes
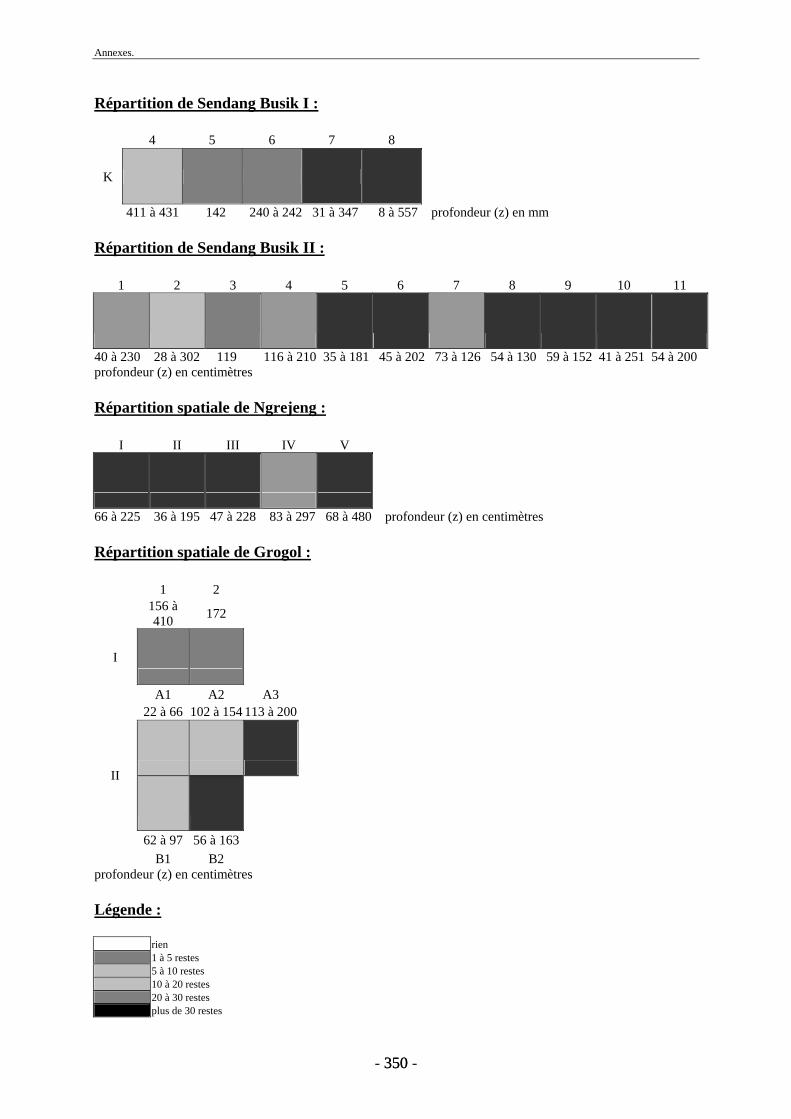
Annexes.
- 350 - 350
Répartition de Sendang Busik I : 4 5 6 7 8
K
411 à 431 142 240 à 242 31 à 347 8 à 557 profondeur (z) en mm Répartition de Sendang Busik II :
1 2 3 4 5 6 7 8 9 10 11
40 à 230 28 à 302 119 116 à 210 35 à 181 45 à 202 73 à 126 54 à 130 59 à 152 41 à 251 54 à 200 profondeur (z) en centimètres Répartition spatiale de Ngrejeng :
I II III IV V
66 à 225 36 à 195 47 à 228 83 à 297 68 à 480 profondeur (z) en centimètres Répartition spatiale de Grogol : 1 2
156 à 410 172
I
A1 A2 A3 22 à 66 102 à 154 113 à 200
II
62 à 97 56 à 163 B1 B2 profondeur (z) en centimètres Légende : rien 1 à 5 restes 5 à 10 restes 10 à 20 restes 20 à 30 restes plus de 30 restes

Annexes.
- 351 - 351
Répartition spatiale de Bukuran :
A1 B1 C1
I
385 à 486 207 à 369 73 à 253
II
? 267 à 269
III
277 à 382 226 à 325 77 à 156
IV
204 à 345
V
267 à 280
VI
105 à 257 271 à 395 profondeur (z) en centimètres Répartition spatiale de Dayu :
I II III IV V
102 à 294 85 à 131 248 à 541 155 66 à 252 profondeur (z) en centimètres Légende : rien 1 à 5 restes 5 à 10 restes 10 à 20 restes 20 à 30 restes plus de 30 restes

Annexes
- 352 -
ANNEXE 7 : Tests statistiques sur la répartition spatiale. Test de la distribution spatiale des vestiges archéologiques (Dacey, 1973) : Ho : la distribution spatiale des vestiges est aléatoire. La formule de ce test est : khi-deux = nv/m N : nombre de carrés. V : variance de la moyenne. M : moyenne du nombre de vestiges par carré. - Si le khi-deux est supérieur à la valeur seuil obtenue dans les tables pour un degré de liberté de (n-1) alors l’hypothèse Ho est rejetée et donc la répartition spatiale des vestiges n’est pas aléatoire.
- Si le khi-deux est inférieur à la valeur seuil obtenue dans les tables pour un degré de liberté de (n-1) alors l’hypothèse Ho est conservée et la répartition spatiale des vestiges est aléatoire.
Dans notre cas, la variance de notre échantillon est de 41,74 et la moyenne est de 3,87
vestiges par carré à Tanjung 64. Le khi-deux a une valeur de 2428,9 ce qui est très supérieur à la valeur seuil pour un degré de liberté de 224 à 0,05. En conséquence la répartition spatiale des ossements de Tanjung 64 n’est pas aléatoire.
Test de l’association spatiale des vestiges (Dacey, 1973) : Ho : il n’existe pas d’association de vestiges.
Si l’on considère dans chaque carré l’absence et la présence de deux types de restes (0 : absence ; 1 : présence), on obtient des couples de données et leur fréquence. Ensuite ces fréquences sont testées à partir d’un test de khi-deux classique (cf. annexe 9) (Chenorkian, 1996).
- Si la valeur du khi-deux est supérieure à celle de la valeur seuil des tables à 0,05 pour un degré de liberté (ddl) donné, alors Ho sera rejetée et donc les deux types de restes sont associés sur le site.
- Si la valeur du khi-deux est inférieure à celle de la valeur seuil des tables à 0,05 pour un degré de liberté (ddl) donné, alors Ho sera conservée et donc les deux types de restes ne sont pas associés sur le site. Dans notre cas, la valeur du khi-deux est égale à 88,02 ce qui est très supérieur à la valeur seuil (3,84) pour un ddl de 1 à 0,05. En conséquence nous pouvons penser qu’il existe une corrélation entre la distribution des esquilles et des restes indéterminés.

Annexes
- 353 -
ANNEXE 8 : Présentation des principaux sites de comparaison
- Ngebung 2 est un site de plein air indonésien (Sangiran) en milieu fluviatile daté du Pléistocène moyen (Saleki, 1997 ; Sémah F. et al., 1993b, 2001a et 2002). Le niveau B est un niveau archéologique remanié ayant subi une altération plus importante que le niveau A où des pièces lithiques ont été trouvées en association avec les restes fauniques. Sur ces derniers des traces de fracturation intentionnelle et des stries ont pu être mises en évidence. Des traces de fracturation intentionnelle et des stries ont également été observées sur quelques restes dans le niveau B.
- Terra Amata (Alpes-Maritimes, France) est un site d’habitat de plein air en bord de la mer Méditerranée où de l’industrie lithique et de la faune ont été découvertes (Serre, 1987, 1991) datées du Pléistocène moyen (environ 380 000 ans). Les espèces d’herbivores dominantes dans l’assemblage sont l’éléphant antique (Palaeoloxodon antiquus) et le cerf élaphe (Cervus elaphus) présentes dans les deux ensembles C1a (plage) et C1b (dune).
- Soleilhac (Haute-Loire, France) est une halte de chasse située sur une presqu’île de
petite taille où ont été retrouvés des vestiges lithiques et des restes fauniques datés d’environ 900 000 ans (Fosse & Bonifay, 1991 ; Fosse, 1994). Ces restes fauniques sont principalement constitués de cervidés (Praemegaceros solihacus et Cervus) et d’éléphantidés (Palaeoloxodon antiquus). L’Homme semble avoir été l’auteur principal de l’accumulation osseuse, bien que l’activité des carnivores ait été attestée mais reste mineure. 446 pièces lithiques en basalte, quartz, silex et granite y ont été découvertes associées aux restes fauniques (Fosse & Bonifay, 1991 ; Fosse, 1994).
- Notarchirico (Basilicata, Italie) est un site de plein air italien en contexte fluviatile
où de l’industrie et des restes fauniques ont été découverts (Cassoli et al., 1999). Le niveau Alpha de ce site serait daté à 230 000 +/- 50 000 ans (Lefèvre et al., 1994). Les espèces dominantes dans ce niveau sont le daim (Dama clactoniana) et l’éléphant (Elephas antiquus). Des traces anthropiques de fracturation ont pu être mises en évidence sur certains ossements. Divers facteurs sont à l’origine de l’assemblage du niveau Alpha. Cette accumulation serait un mélange de restes introduits par le transport fluviatile et d’un second dépôt provenant de l’érosion des rives composé de restes de carcasses d’animaux morts naturellement ou qui ont été chassés par les carnivores ou l’homme, dans l’environnement proche du cours d’eau. La très forte proportion des restes fauniques et leur association avec des outils lithiques dans ce niveau impliquent une origine anthropique, il est possible qu’elle soit caractérisée par un palimpseste de diverses accumulations de restes (Cassoli et al., 1999).

Annexes.
- 354 - 354
- Holon (Israël) est un site de plein air de la côte israélienne daté d’environ 200 000 ans localisé à proximité d’un ancien bras de rivière (Chazan et al., 2001 ; Monchot & Horwitz, 2002). Sur ce site, des vestiges lithiques et fauniques ont été mis au jour. Les espèces dominantes sont le daim de Mésopotamie (Dama mesopotamica), l’auroch (Bos primigenius) et l’éléphant antique (Palaeoloxodon antiquus). Holon est considéré comme un site d’abattage et/ou de dépeçage.
- Le site du Mont Dol est un site de plein air français (Ille-et-Vilaine) daté du
Paléolithique moyen (âge Melisey II environ 90 000 ans) (Simonet & Monnier, 1991). Ce site se localise au pied d’une butte granitique, le Mont Dol. Les restes fauniques et lithiques proviennent, principalement, des couches 6 à 9 constituées par plusieurs coulées de solifluxion. L’action anthropique a pu être mise en évidence à travers la fracturation systématique des os longs et crâniens pour la récupération de la moelle et de la cervelle et par l’observation de quelques stries laissées sur les os.
- Verdigree Quarry est un site de plein air américain (Knox County, Nebraska)
couvrant une période allant du Miocène au Pliocène inférieur (Voorhies, 1969). Ce site a subi une forte influence fluviatile, en effet, les sédiments où se situent les fossiles semblent s’être déposés très rapidement pendant une crue.
- Tourville D est un site naturel de plein air français (basse vallée de la Seine) daté du
Pléistocène moyen (stades isotopiques 6 et 7) (Descombes, 1980). Ce niveau D se situe dans un dépôt de chenal. Le matériel de ce niveau semble avoir été très transporté sur un courte période de temps et ne présente pas de traces anthropiques ou de carnivores. Une telle accumulation paraît être un enfouissement dans le chenal de cadavres apportés par le courant (Descombes, 1980).
- Le site de la Baume de Goulon est une grotte du Paléolithique supérieur (environ 10
000 ans) où de la faune, principalement des restes de Cervus elaphus et de Bos primigenius, et de l’industrie lithique ont été retrouvés. Sur ces deux sites, ces proportions s’expliquent par une fracturation intentionnelle sur os frais (Monchot, 1998).
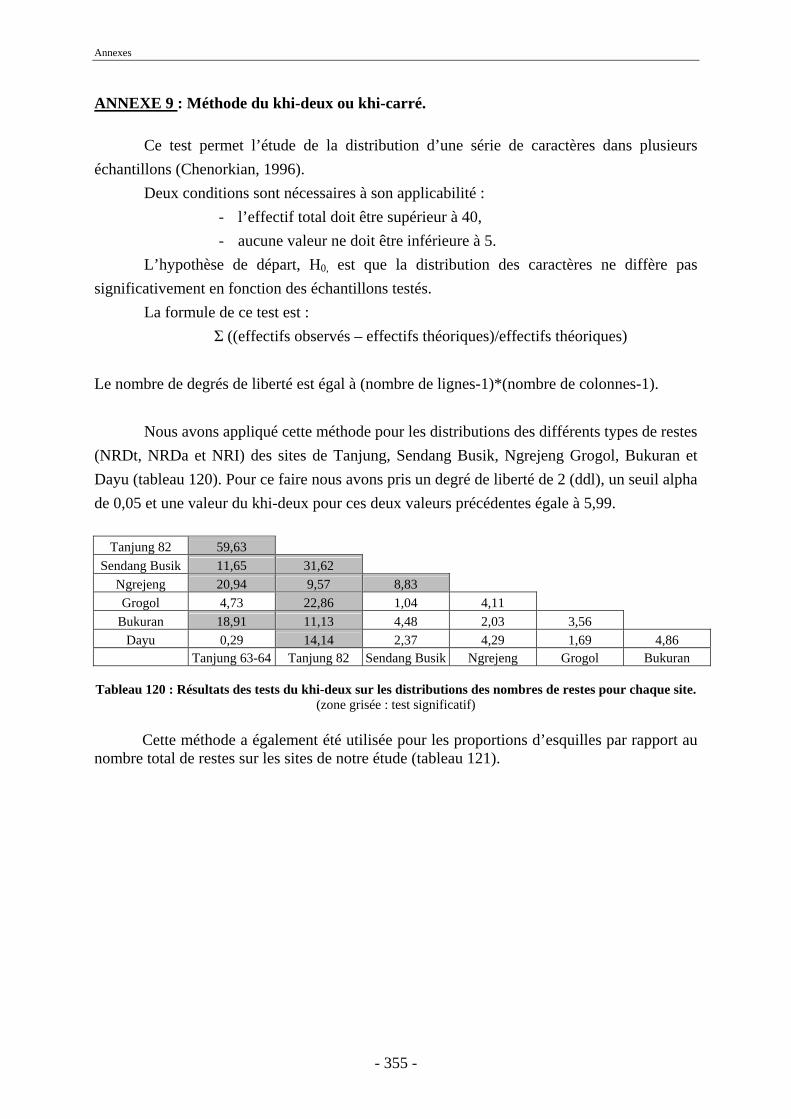
Annexes
- 355 -
ANNEXE 9 : Méthode du khi-deux ou khi-carré.
Ce test permet l’étude de la distribution d’une série de caractères dans plusieurs échantillons (Chenorkian, 1996).
Deux conditions sont nécessaires à son applicabilité : - l’effectif total doit être supérieur à 40, - aucune valeur ne doit être inférieure à 5.
L’hypothèse de départ, H0, est que la distribution des caractères ne diffère pas significativement en fonction des échantillons testés.
La formule de ce test est : Σ ((effectifs observés – effectifs théoriques)/effectifs théoriques)
Le nombre de degrés de liberté est égal à (nombre de lignes-1)*(nombre de colonnes-1).
Nous avons appliqué cette méthode pour les distributions des différents types de restes
(NRDt, NRDa et NRI) des sites de Tanjung, Sendang Busik, Ngrejeng Grogol, Bukuran et Dayu (tableau 120). Pour ce faire nous avons pris un degré de liberté de 2 (ddl), un seuil alpha de 0,05 et une valeur du khi-deux pour ces deux valeurs précédentes égale à 5,99.
Tanjung 82 59,63 Sendang Busik 11,65 31,62
Ngrejeng 20,94 9,57 8,83 Grogol 4,73 22,86 1,04 4,11
Bukuran 18,91 11,13 4,48 2,03 3,56 Dayu 0,29 14,14 2,37 4,29 1,69 4,86
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 120 : Résultats des tests du khi-deux sur les distributions des nombres de restes pour chaque site. (zone grisée : test significatif)
Cette méthode a également été utilisée pour les proportions d’esquilles par rapport au
nombre total de restes sur les sites de notre étude (tableau 121).
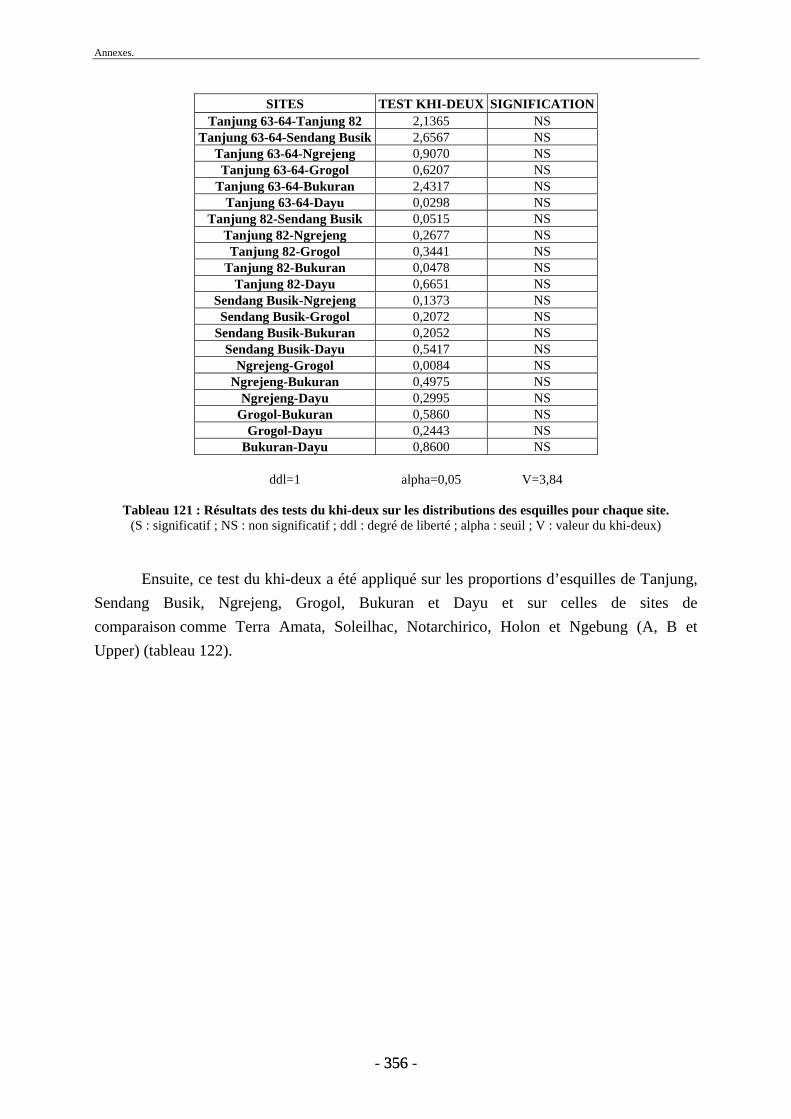
Annexes.
- 356 - 356
SITES TEST KHI-DEUX SIGNIFICATION Tanjung 63-64-Tanjung 82 2,1365 NS
Tanjung 63-64-Sendang Busik 2,6567 NS Tanjung 63-64-Ngrejeng 0,9070 NS Tanjung 63-64-Grogol 0,6207 NS
Tanjung 63-64-Bukuran 2,4317 NS Tanjung 63-64-Dayu 0,0298 NS
Tanjung 82-Sendang Busik 0,0515 NS Tanjung 82-Ngrejeng 0,2677 NS Tanjung 82-Grogol 0,3441 NS
Tanjung 82-Bukuran 0,0478 NS Tanjung 82-Dayu 0,6651 NS
Sendang Busik-Ngrejeng 0,1373 NS Sendang Busik-Grogol 0,2072 NS
Sendang Busik-Bukuran 0,2052 NS Sendang Busik-Dayu 0,5417 NS
Ngrejeng-Grogol 0,0084 NS Ngrejeng-Bukuran 0,4975 NS
Ngrejeng-Dayu 0,2995 NS Grogol-Bukuran 0,5860 NS
Grogol-Dayu 0,2443 NS Bukuran-Dayu 0,8600 NS
ddl=1 alpha=0,05 V=3,84
Tableau 121 : Résultats des tests du khi-deux sur les distributions des esquilles pour chaque site.
(S : significatif ; NS : non significatif ; ddl : degré de liberté ; alpha : seuil ; V : valeur du khi-deux)
Ensuite, ce test du khi-deux a été appliqué sur les proportions d’esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu et sur celles de sites de comparaison comme Terra Amata, Soleilhac, Notarchirico, Holon et Ngebung (A, B et Upper) (tableau 122).
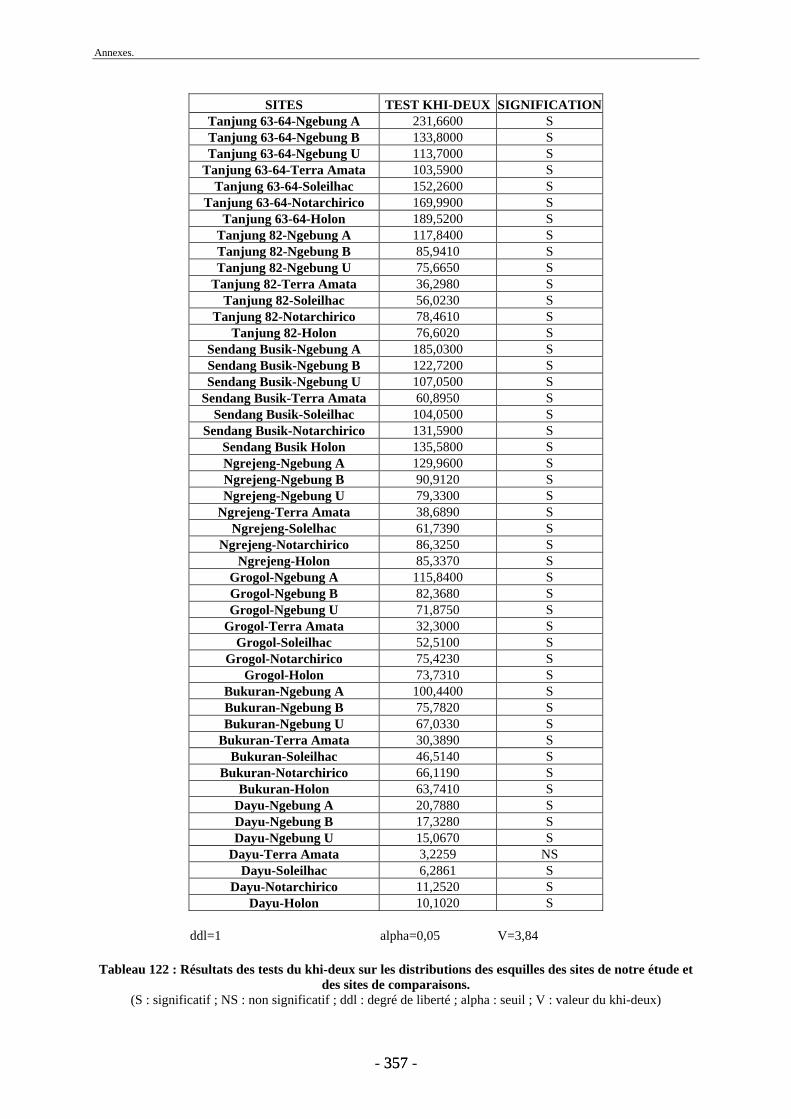
Annexes.
- 357 - 357
SITES TEST KHI-DEUX SIGNIFICATION Tanjung 63-64-Ngebung A 231,6600 S Tanjung 63-64-Ngebung B 133,8000 S Tanjung 63-64-Ngebung U 113,7000 S
Tanjung 63-64-Terra Amata 103,5900 S Tanjung 63-64-Soleilhac 152,2600 S
Tanjung 63-64-Notarchirico 169,9900 S Tanjung 63-64-Holon 189,5200 S
Tanjung 82-Ngebung A 117,8400 S Tanjung 82-Ngebung B 85,9410 S Tanjung 82-Ngebung U 75,6650 S
Tanjung 82-Terra Amata 36,2980 S Tanjung 82-Soleilhac 56,0230 S
Tanjung 82-Notarchirico 78,4610 S Tanjung 82-Holon 76,6020 S
Sendang Busik-Ngebung A 185,0300 S Sendang Busik-Ngebung B 122,7200 S Sendang Busik-Ngebung U 107,0500 S
Sendang Busik-Terra Amata 60,8950 S Sendang Busik-Soleilhac 104,0500 S
Sendang Busik-Notarchirico 131,5900 S Sendang Busik Holon 135,5800 S Ngrejeng-Ngebung A 129,9600 S Ngrejeng-Ngebung B 90,9120 S Ngrejeng-Ngebung U 79,3300 S
Ngrejeng-Terra Amata 38,6890 S Ngrejeng-Solelhac 61,7390 S
Ngrejeng-Notarchirico 86,3250 S Ngrejeng-Holon 85,3370 S
Grogol-Ngebung A 115,8400 S Grogol-Ngebung B 82,3680 S Grogol-Ngebung U 71,8750 S
Grogol-Terra Amata 32,3000 S Grogol-Soleilhac 52,5100 S
Grogol-Notarchirico 75,4230 S Grogol-Holon 73,7310 S
Bukuran-Ngebung A 100,4400 S Bukuran-Ngebung B 75,7820 S Bukuran-Ngebung U 67,0330 S
Bukuran-Terra Amata 30,3890 S Bukuran-Soleilhac 46,5140 S
Bukuran-Notarchirico 66,1190 S Bukuran-Holon 63,7410 S
Dayu-Ngebung A 20,7880 S Dayu-Ngebung B 17,3280 S Dayu-Ngebung U 15,0670 S
Dayu-Terra Amata 3,2259 NS Dayu-Soleilhac 6,2861 S
Dayu-Notarchirico 11,2520 S Dayu-Holon 10,1020 S
ddl=1 alpha=0,05 V=3,84
Tableau 122 : Résultats des tests du khi-deux sur les distributions des esquilles des sites de notre étude et
des sites de comparaisons. (S : significatif ; NS : non significatif ; ddl : degré de liberté ; alpha : seuil ; V : valeur du khi-deux)
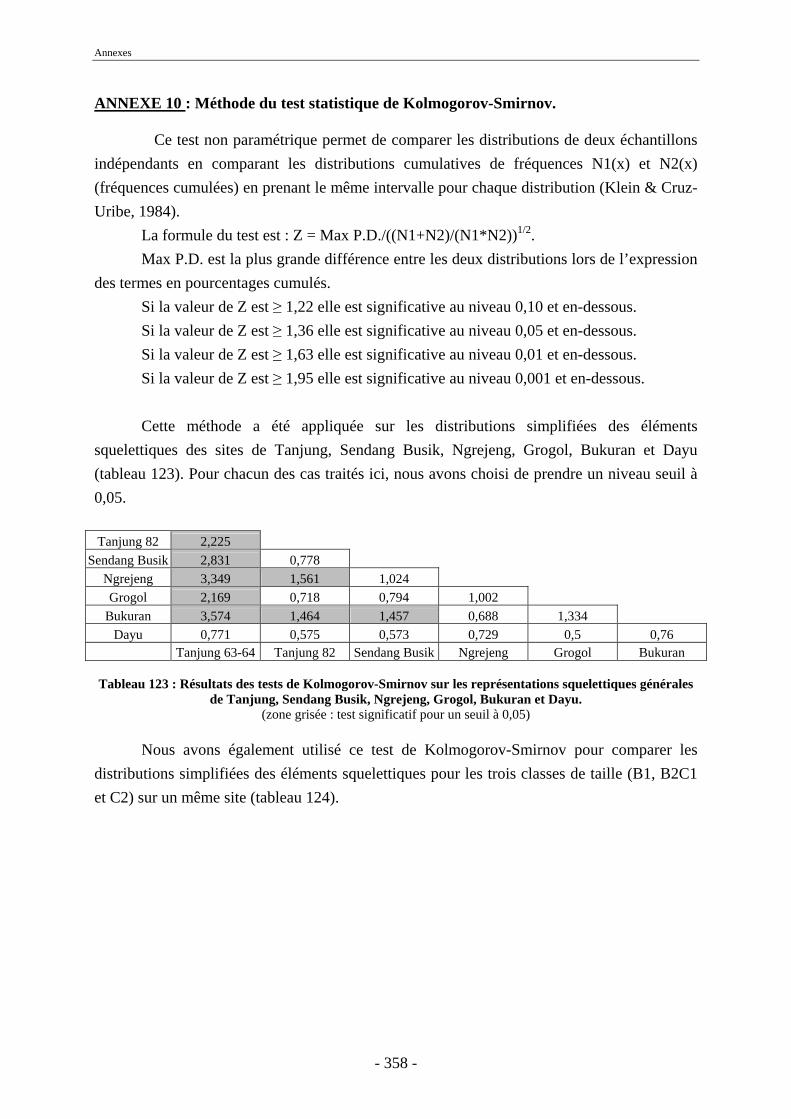
Annexes
- 358 -
ANNEXE 10 : Méthode du test statistique de Kolmogorov-Smirnov.
Ce test non paramétrique permet de comparer les distributions de deux échantillons indépendants en comparant les distributions cumulatives de fréquences N1(x) et N2(x) (fréquences cumulées) en prenant le même intervalle pour chaque distribution (Klein & Cruz-Uribe, 1984).
La formule du test est : Z = Max P.D./((N1+N2)/(N1*N2))1/2. Max P.D. est la plus grande différence entre les deux distributions lors de l’expression
des termes en pourcentages cumulés. Si la valeur de Z est ≥ 1,22 elle est significative au niveau 0,10 et en-dessous. Si la valeur de Z est ≥ 1,36 elle est significative au niveau 0,05 et en-dessous. Si la valeur de Z est ≥ 1,63 elle est significative au niveau 0,01 et en-dessous. Si la valeur de Z est ≥ 1,95 elle est significative au niveau 0,001 et en-dessous. Cette méthode a été appliquée sur les distributions simplifiées des éléments
squelettiques des sites de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu (tableau 123). Pour chacun des cas traités ici, nous avons choisi de prendre un niveau seuil à 0,05. Tanjung 82 2,225
Sendang Busik 2,831 0,778 Ngrejeng 3,349 1,561 1,024 Grogol 2,169 0,718 0,794 1,002
Bukuran 3,574 1,464 1,457 0,688 1,334 Dayu 0,771 0,575 0,573 0,729 0,5 0,76
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 123 : Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques générales de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.
(zone grisée : test significatif pour un seuil à 0,05)
Nous avons également utilisé ce test de Kolmogorov-Smirnov pour comparer les distributions simplifiées des éléments squelettiques pour les trois classes de taille (B1, B2C1 et C2) sur un même site (tableau 124).
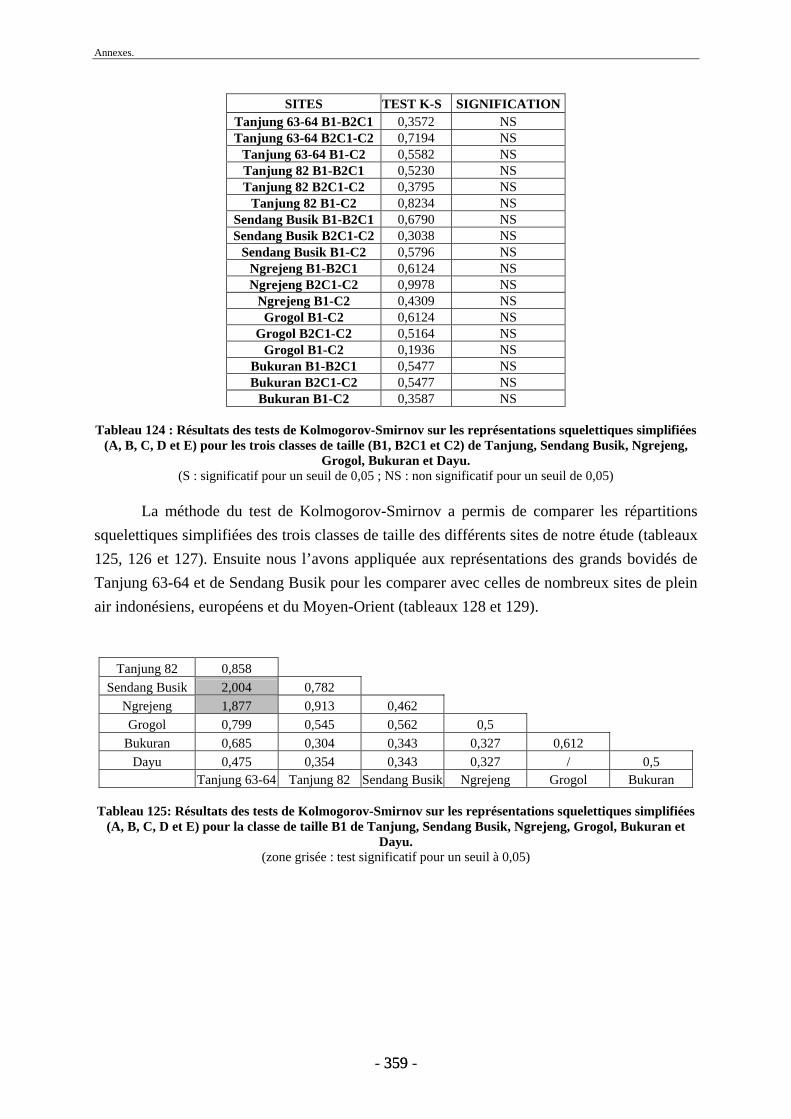
Annexes.
- 359 - 359
SITES TEST K-S SIGNIFICATION Tanjung 63-64 B1-B2C1 0,3572 NS Tanjung 63-64 B2C1-C2 0,7194 NS
Tanjung 63-64 B1-C2 0,5582 NS Tanjung 82 B1-B2C1 0,5230 NS Tanjung 82 B2C1-C2 0,3795 NS
Tanjung 82 B1-C2 0,8234 NS Sendang Busik B1-B2C1 0,6790 NS Sendang Busik B2C1-C2 0,3038 NS
Sendang Busik B1-C2 0,5796 NS Ngrejeng B1-B2C1 0,6124 NS Ngrejeng B2C1-C2 0,9978 NS
Ngrejeng B1-C2 0,4309 NS Grogol B1-C2 0,6124 NS
Grogol B2C1-C2 0,5164 NS Grogol B1-C2 0,1936 NS
Bukuran B1-B2C1 0,5477 NS Bukuran B2C1-C2 0,5477 NS
Bukuran B1-C2 0,3587 NS
Tableau 124 : Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour les trois classes de taille (B1, B2C1 et C2) de Tanjung, Sendang Busik, Ngrejeng,
Grogol, Bukuran et Dayu. (S : significatif pour un seuil de 0,05 ; NS : non significatif pour un seuil de 0,05)
La méthode du test de Kolmogorov-Smirnov a permis de comparer les répartitions
squelettiques simplifiées des trois classes de taille des différents sites de notre étude (tableaux 125, 126 et 127). Ensuite nous l’avons appliquée aux représentations des grands bovidés de Tanjung 63-64 et de Sendang Busik pour les comparer avec celles de nombreux sites de plein air indonésiens, européens et du Moyen-Orient (tableaux 128 et 129).
Tanjung 82 0,858 Sendang Busik 2,004 0,782
Ngrejeng 1,877 0,913 0,462 Grogol 0,799 0,545 0,562 0,5
Bukuran 0,685 0,304 0,343 0,327 0,612 Dayu 0,475 0,354 0,343 0,327 / 0,5
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 125: Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour la classe de taille B1 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et
Dayu. (zone grisée : test significatif pour un seuil à 0,05)
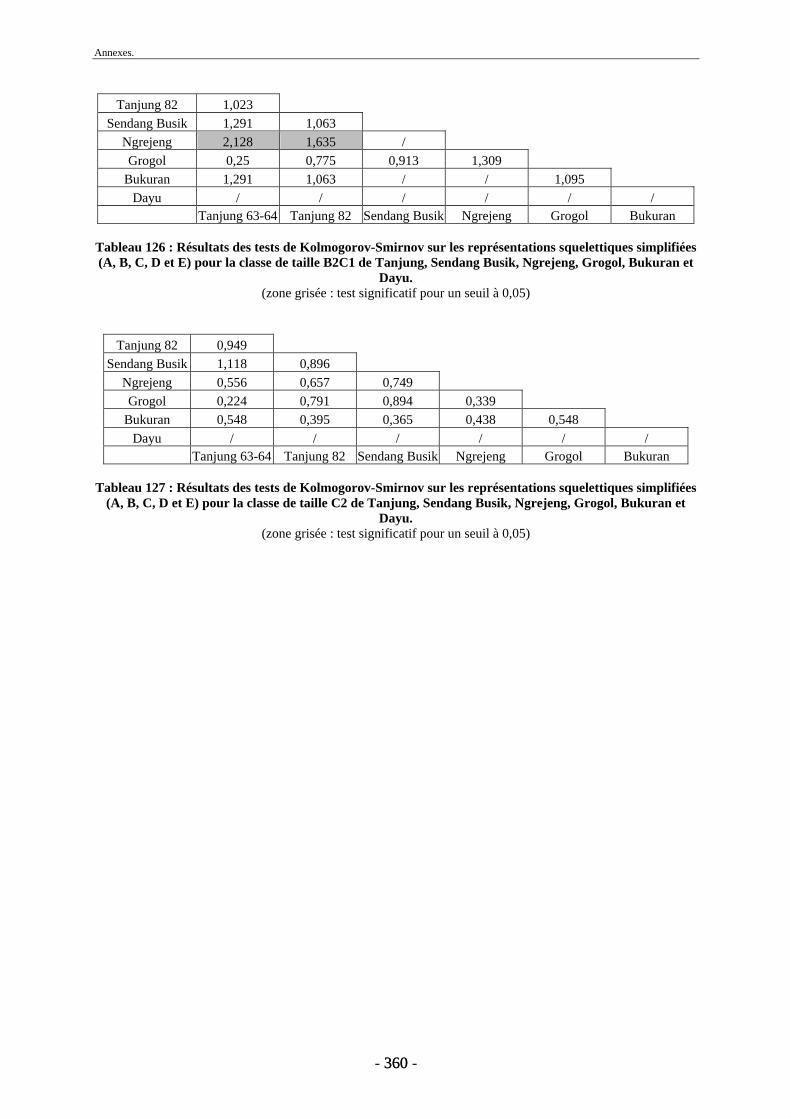
Annexes.
- 360 - 360
Tanjung 82 1,023 Sendang Busik 1,291 1,063
Ngrejeng 2,128 1,635 / Grogol 0,25 0,775 0,913 1,309
Bukuran 1,291 1,063 / / 1,095 Dayu / / / / / /
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 126 : Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour la classe de taille B2C1 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et
Dayu. (zone grisée : test significatif pour un seuil à 0,05)
Tanjung 82 0,949 Sendang Busik 1,118 0,896
Ngrejeng 0,556 0,657 0,749 Grogol 0,224 0,791 0,894 0,339
Bukuran 0,548 0,395 0,365 0,438 0,548 Dayu / / / / / /
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 127 : Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour la classe de taille C2 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et
Dayu. (zone grisée : test significatif pour un seuil à 0,05)
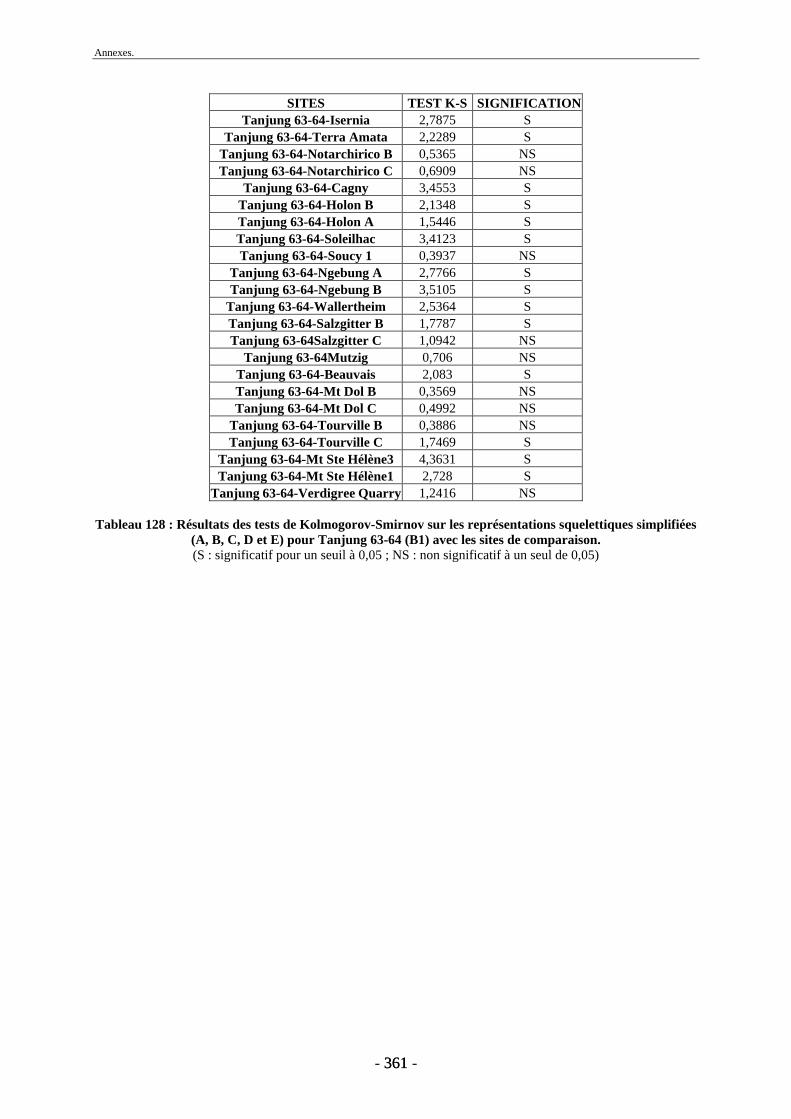
Annexes.
- 361 - 361
SITES TEST K-S SIGNIFICATION Tanjung 63-64-Isernia 2,7875 S
Tanjung 63-64-Terra Amata 2,2289 S Tanjung 63-64-Notarchirico B 0,5365 NS Tanjung 63-64-Notarchirico C 0,6909 NS
Tanjung 63-64-Cagny 3,4553 S Tanjung 63-64-Holon B 2,1348 S Tanjung 63-64-Holon A 1,5446 S Tanjung 63-64-Soleilhac 3,4123 S Tanjung 63-64-Soucy 1 0,3937 NS
Tanjung 63-64-Ngebung A 2,7766 S Tanjung 63-64-Ngebung B 3,5105 S
Tanjung 63-64-Wallertheim 2,5364 S Tanjung 63-64-Salzgitter B 1,7787 S Tanjung 63-64Salzgitter C 1,0942 NS
Tanjung 63-64Mutzig 0,706 NS Tanjung 63-64-Beauvais 2,083 S Tanjung 63-64-Mt Dol B 0,3569 NS Tanjung 63-64-Mt Dol C 0,4992 NS
Tanjung 63-64-Tourville B 0,3886 NS Tanjung 63-64-Tourville C 1,7469 S
Tanjung 63-64-Mt Ste Hélène3 4,3631 S Tanjung 63-64-Mt Ste Hélène1 2,728 S
Tanjung 63-64-Verdigree Quarry 1,2416 NS
Tableau 128 : Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour Tanjung 63-64 (B1) avec les sites de comparaison. (S : significatif pour un seuil à 0,05 ; NS : non significatif à un seul de 0,05)
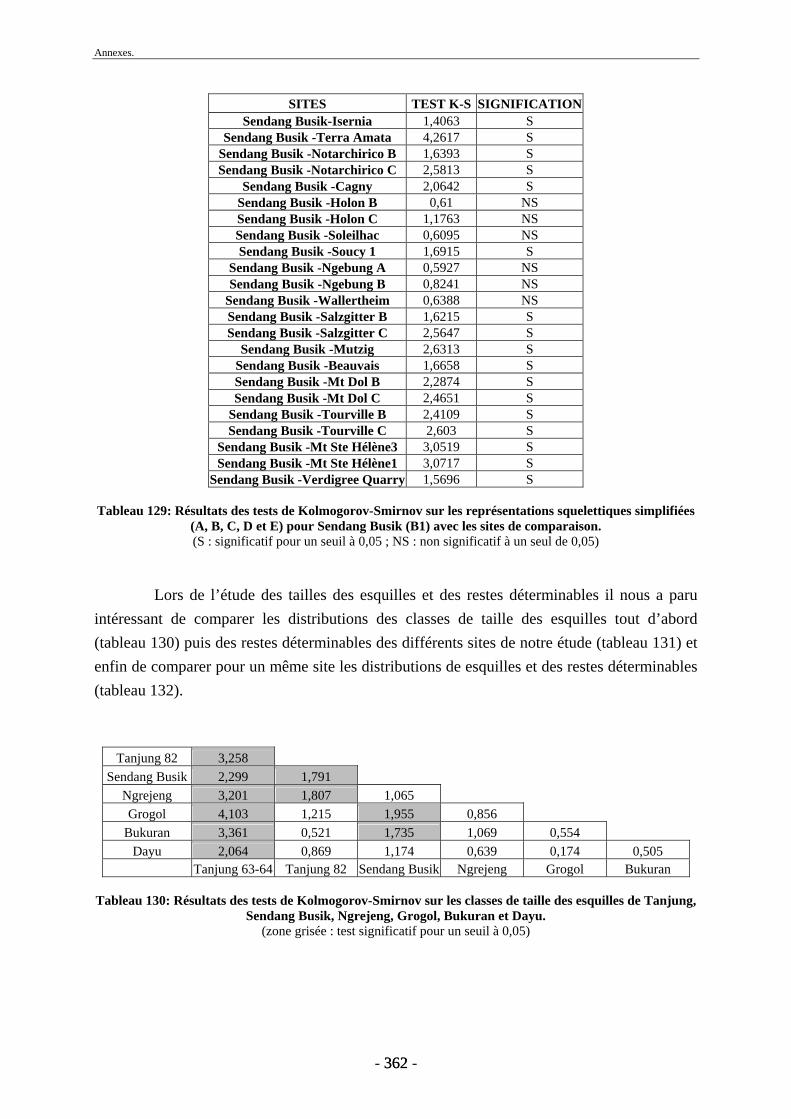
Annexes.
- 362 - 362
SITES TEST K-S SIGNIFICATION Sendang Busik-Isernia 1,4063 S
Sendang Busik -Terra Amata 4,2617 S Sendang Busik -Notarchirico B 1,6393 S Sendang Busik -Notarchirico C 2,5813 S
Sendang Busik -Cagny 2,0642 S Sendang Busik -Holon B 0,61 NS Sendang Busik -Holon C 1,1763 NS Sendang Busik -Soleilhac 0,6095 NS Sendang Busik -Soucy 1 1,6915 S
Sendang Busik -Ngebung A 0,5927 NS Sendang Busik -Ngebung B 0,8241 NS
Sendang Busik -Wallertheim 0,6388 NS Sendang Busik -Salzgitter B 1,6215 S Sendang Busik -Salzgitter C 2,5647 S
Sendang Busik -Mutzig 2,6313 S Sendang Busik -Beauvais 1,6658 S Sendang Busik -Mt Dol B 2,2874 S Sendang Busik -Mt Dol C 2,4651 S
Sendang Busik -Tourville B 2,4109 S Sendang Busik -Tourville C 2,603 S
Sendang Busik -Mt Ste Hélène3 3,0519 S Sendang Busik -Mt Ste Hélène1 3,0717 S
Sendang Busik -Verdigree Quarry 1,5696 S
Tableau 129: Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour Sendang Busik (B1) avec les sites de comparaison. (S : significatif pour un seuil à 0,05 ; NS : non significatif à un seul de 0,05)
Lors de l’étude des tailles des esquilles et des restes déterminables il nous a paru intéressant de comparer les distributions des classes de taille des esquilles tout d’abord (tableau 130) puis des restes déterminables des différents sites de notre étude (tableau 131) et enfin de comparer pour un même site les distributions de esquilles et des restes déterminables (tableau 132).
Tanjung 82 3,258 Sendang Busik 2,299 1,791
Ngrejeng 3,201 1,807 1,065 Grogol 4,103 1,215 1,955 0,856
Bukuran 3,361 0,521 1,735 1,069 0,554 Dayu 2,064 0,869 1,174 0,639 0,174 0,505
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 130: Résultats des tests de Kolmogorov-Smirnov sur les classes de taille des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.
(zone grisée : test significatif pour un seuil à 0,05)
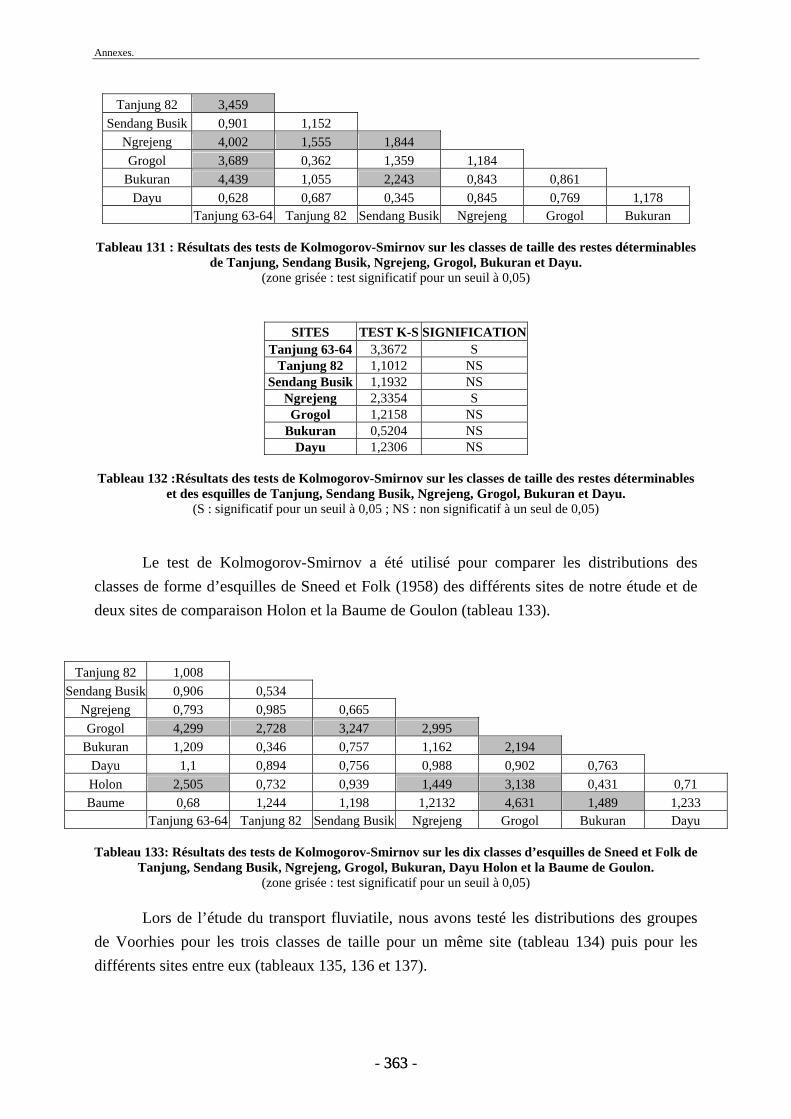
Annexes.
- 363 - 363
Tanjung 82 3,459 Sendang Busik 0,901 1,152
Ngrejeng 4,002 1,555 1,844 Grogol 3,689 0,362 1,359 1,184
Bukuran 4,439 1,055 2,243 0,843 0,861 Dayu 0,628 0,687 0,345 0,845 0,769 1,178
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 131 : Résultats des tests de Kolmogorov-Smirnov sur les classes de taille des restes déterminables de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.
(zone grisée : test significatif pour un seuil à 0,05)
SITES TEST K-S SIGNIFICATIONTanjung 63-64 3,3672 S
Tanjung 82 1,1012 NS Sendang Busik 1,1932 NS
Ngrejeng 2,3354 S Grogol 1,2158 NS
Bukuran 0,5204 NS Dayu 1,2306 NS
Tableau 132 :Résultats des tests de Kolmogorov-Smirnov sur les classes de taille des restes déterminables
et des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. (S : significatif pour un seuil à 0,05 ; NS : non significatif à un seul de 0,05)
Le test de Kolmogorov-Smirnov a été utilisé pour comparer les distributions des classes de forme d’esquilles de Sneed et Folk (1958) des différents sites de notre étude et de deux sites de comparaison Holon et la Baume de Goulon (tableau 133).
Tanjung 82 1,008 Sendang Busik 0,906 0,534
Ngrejeng 0,793 0,985 0,665 Grogol 4,299 2,728 3,247 2,995
Bukuran 1,209 0,346 0,757 1,162 2,194 Dayu 1,1 0,894 0,756 0,988 0,902 0,763 Holon 2,505 0,732 0,939 1,449 3,138 0,431 0,71 Baume 0,68 1,244 1,198 1,2132 4,631 1,489 1,233
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran Dayu
Tableau 133: Résultats des tests de Kolmogorov-Smirnov sur les dix classes d’esquilles de Sneed et Folk de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran, Dayu Holon et la Baume de Goulon.
(zone grisée : test significatif pour un seuil à 0,05)
Lors de l’étude du transport fluviatile, nous avons testé les distributions des groupes de Voorhies pour les trois classes de taille pour un même site (tableau 134) puis pour les différents sites entre eux (tableaux 135, 136 et 137).
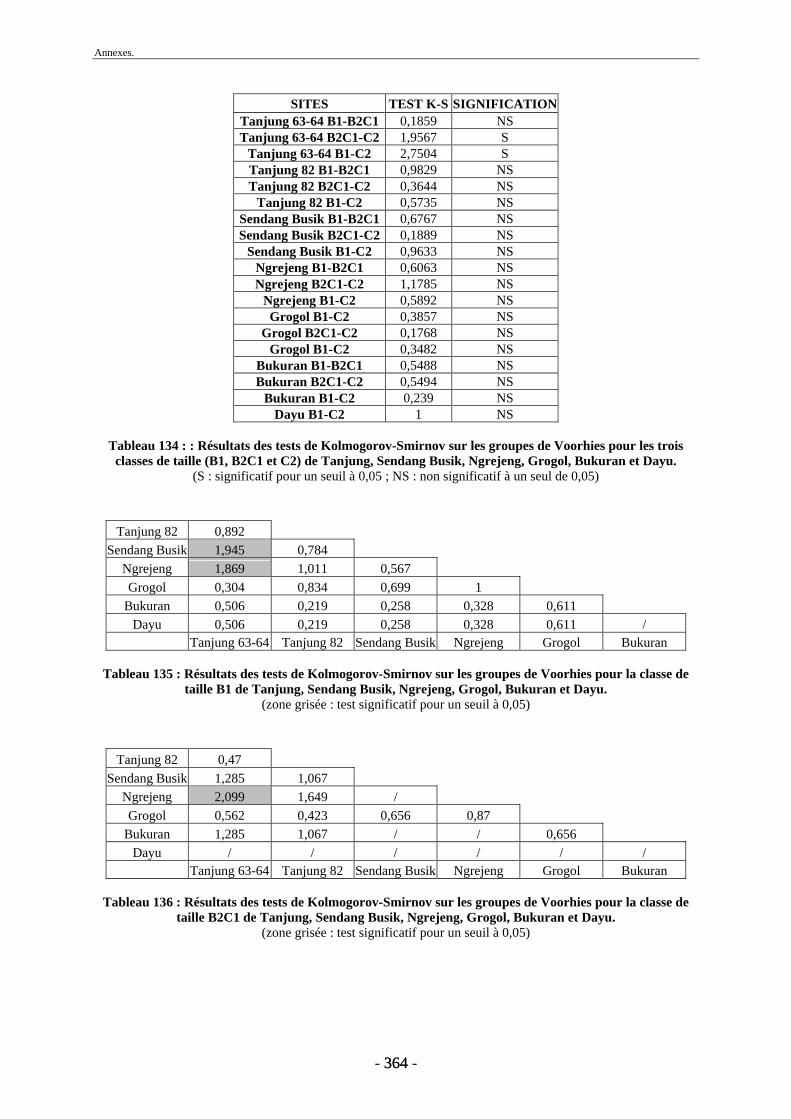
Annexes.
- 364 - 364
SITES TEST K-S SIGNIFICATIONTanjung 63-64 B1-B2C1 0,1859 NS Tanjung 63-64 B2C1-C2 1,9567 S
Tanjung 63-64 B1-C2 2,7504 S Tanjung 82 B1-B2C1 0,9829 NS Tanjung 82 B2C1-C2 0,3644 NS
Tanjung 82 B1-C2 0,5735 NS Sendang Busik B1-B2C1 0,6767 NS Sendang Busik B2C1-C2 0,1889 NS
Sendang Busik B1-C2 0,9633 NS Ngrejeng B1-B2C1 0,6063 NS Ngrejeng B2C1-C2 1,1785 NS
Ngrejeng B1-C2 0,5892 NS Grogol B1-C2 0,3857 NS
Grogol B2C1-C2 0,1768 NS Grogol B1-C2 0,3482 NS
Bukuran B1-B2C1 0,5488 NS Bukuran B2C1-C2 0,5494 NS
Bukuran B1-C2 0,239 NS Dayu B1-C2 1 NS
Tableau 134 : : Résultats des tests de Kolmogorov-Smirnov sur les groupes de Voorhies pour les trois
classes de taille (B1, B2C1 et C2) de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. (S : significatif pour un seuil à 0,05 ; NS : non significatif à un seul de 0,05)
Tanjung 82 0,892 Sendang Busik 1,945 0,784
Ngrejeng 1,869 1,011 0,567 Grogol 0,304 0,834 0,699 1
Bukuran 0,506 0,219 0,258 0,328 0,611 Dayu 0,506 0,219 0,258 0,328 0,611 /
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 135 : Résultats des tests de Kolmogorov-Smirnov sur les groupes de Voorhies pour la classe de taille B1 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.
(zone grisée : test significatif pour un seuil à 0,05)
Tanjung 82 0,47 Sendang Busik 1,285 1,067
Ngrejeng 2,099 1,649 / Grogol 0,562 0,423 0,656 0,87
Bukuran 1,285 1,067 / / 0,656 Dayu / / / / / /
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 136 : Résultats des tests de Kolmogorov-Smirnov sur les groupes de Voorhies pour la classe de taille B2C1 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.
(zone grisée : test significatif pour un seuil à 0,05)
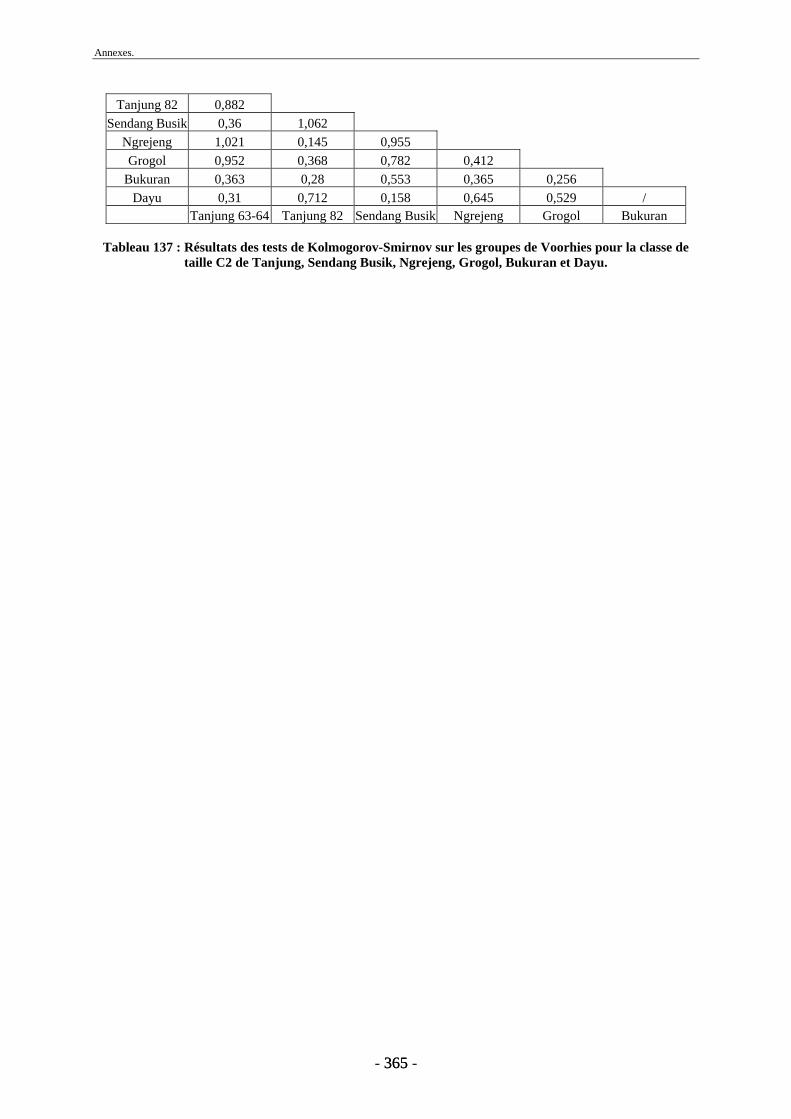
Annexes.
- 365 - 365
Tanjung 82 0,882 Sendang Busik 0,36 1,062
Ngrejeng 1,021 0,145 0,955 Grogol 0,952 0,368 0,782 0,412
Bukuran 0,363 0,28 0,553 0,365 0,256 Dayu 0,31 0,712 0,158 0,645 0,529 /
Tanjung 63-64 Tanjung 82 Sendang Busik Ngrejeng Grogol Bukuran
Tableau 137 : Résultats des tests de Kolmogorov-Smirnov sur les groupes de Voorhies pour la classe de taille C2 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu.

Annexes.
- 366 - 366
ANNEXE 11 : Planche photographique des traces taphonomiques.
a- TDJ64/F3/241 : esquille avec traces d’oxyde de manganèse, vue externe. b- TDJ64/F6/269 : esquille avec traces d’oxyde de fer, vue externe. c- TDJ64/surface/ R10028 : esquille avec traces de fissure type f2, vue externe. d- TDJ64/G5/86 : esquille avec traces de fissure type f3, vue interne. e- NKJ85/III/14: côte avec traces de mâchonnage de carnivore, vue externe. f- NKJ85/II/37: esquille avec traces de mâchonnage de carnivore, vue externe. g- BK79/VI/B1/30: esquille avec traces de mâchonnage de carnivore, vue externe. h- TDJ64/C4/10181 : esquille avec fracture sur os frais, vue externe. i- TDJ82/III/5 : bola.

Annexes.
- 367 - 367
a b c d
e f g h

Annexes.
- 368 - 368
i

- 369 -
LISTE DES TABLEAUX DES ANNEXES Tableau 85 : Les principaux restes d’hominidés découverts sur l’île de Java. ...................... 309 Tableau 86 : Liste des restes humains découverts sur le dôme de Sangiran dans les différentes
formations sédimentaires................................................................................................ 310 Tableau 87 : Mesures des M2 de Stegodon trigonocephalus de la collection Dubois, Hooijer,
1955................................................................................................................................ 334 Tableau 88 : Mesures des M3
de Stegodon trigonocephalus de la collection Dubois, Hooijer, 1955................................................................................................................................ 334
Tableau 89 : Mesures des DM4 de Stegodon trigonocephalus de la collection Dubois, Hooijer, 1955................................................................................................................................ 335
Tableau 90 : Mesures des M2 de Stegodon trigonocephalus de la collection Dubois, Hooijer, 1955................................................................................................................................ 335
Tableau 91 : Mesures des M2 d’Elephas hysudrindicus des collections du Naturalis Museum de Leiden, Hooijer, 1955................................................................................................ 336
Tableau 92: Mesures des M3 d’Elephas hysudrindicus de la collection Dubois, Hooijer, 1955......................................................................................................................................... 336
Tableau 93 : Mesures des M3 d’Elephas hysudrindicus de différentes collections, Hooijer, 1955................................................................................................................................ 337
Tableau 94 : Mesures des M2 d’Elephas maximus des collections du Naturalis Museum de Leiden, du laboratoire d’Anatomie Comparée de Paris et Fromaget, Hooijer, 1955.... 337
Tableau 95 : Mesures des M3 d’Elephas maximus des collections du Naturalis Museum de Leiden, du laboratoire d’Anatomie Comparée de Paris et Fromaget, Hooijer, 1955.... 338
Tableau 96 : Mesures des M3 d’Elephas maximus des collections du Naturalis Museum de Leiden, du laboratoire d’Anatomie Comparée de Paris et Fromaget, Hooijer, 1955.... 338
Tableau 97 : Mesures des fémurs de Dicerorhinus sumatrensis, Rhinoceros sondaicus, Rhinoceros unicornis et de Kedung Brubus, Guérin & Faure, 2002. ............................ 338
Tableau 98 : Mesures des M3 d’Hexaprotodon sivalensis de la collection Dubois, Hooijer, 1950................................................................................................................................ 339
Tableau 99 : Mesures des molaires inférieures d’Hexaprotodon sivalensis de la collection Dubois, Hooijer, 1950. ................................................................................................... 339
Tableau 100 : Mesures des prémolaires supérieures d’Hippopotamus amphibius actuel, Faure, 1983................................................................................................................................ 340
Tableau 101 : Mesures des M3 de Sus brachygnathus de Trinil, Kebonduren, Kaigede, Bangle et Teguan, Hardjasasmita, 1987. ................................................................................... 340
Tableau 102: Mesures des dents supérieures d’Epileptobos groeneveldtii du Naturalis Museum de Leiden. ........................................................................................................ 341
Tableau 103 : Mesures des dents inférieures de Bibos palaesondaicus et de Bubalus palaeokerabau du Naturalis Museum de Leiden. .......................................................... 341
Tableau 104 : Mesures des dents supérieures de Duboisia santeng du Naturalis Museum de Leiden............................................................................................................................. 342
Tableau 105 : Mesures des dent inférieures de Duboisia santeng du Naturalis Museum de Leiden............................................................................................................................. 342
Tableau 106 : Mesures des dents supérieures de Duboisia de Punung, Badoux, 1959. ........ 342 Tableau 107 : Mesures des dents supérieures de Duboisia santeng de Trinil, Stremme, 1911.
........................................................................................................................................ 343 Tableau 108 : Mesures des dents inférieures de Duboisia santeng de Trinil, Stremme, 1911.
........................................................................................................................................ 343

Annexes.
- 370 - 370
Tableau 109 : Mesures des dents supérieures d’Axis lydekkeri de Trinil, Stremme, 1911. ... 343 Tableau 110 : Mesures des dents inférieures d’Axis lydekkeri de Trinil, Stremme, 1911. .... 344 Tableau 111 : Mesures des dents supérieures d’Axis lydekkeri du Naturalis Museum de
Leiden............................................................................................................................. 344 Tableau 112 : Mesures des dents inférieures d’Axis lydekkeri du Naturalis Museum de
Leiden............................................................................................................................. 345 Tableau 113 : Mesures des dents supérieures d’Axis nssp. de Ngebung 2, Moigne et al., 2004.
........................................................................................................................................ 345 Tableau 114 : Mesures des dents inférieures d’Axis nssp. de Ngebung 2, Moigne et al., 2004.
........................................................................................................................................ 346 Tableau 115 : Mesures des scapulas du Naturalis Museum de Leiden et de Ngebung 2....... 346 Tableau 116 : Mesures des dents supérieures de Cervus kendengensis du Naturalis Museum
de Leiden. ....................................................................................................................... 347 Tableau 117 : Mesures des dents inférieures de Cervus kendengensis du Naturalis Museum de
Leiden............................................................................................................................. 347 Tableau 118 : Mesures des dents inférieures de Cervus hippelaphus de von Koenigswald,
1933................................................................................................................................ 348 Tableau 119 : Mesures des M2 de Cervus (Rusa) sp. de Ngebung 2, Moigne et al., 2004. ... 348 Tableau 120 : Résultats des tests du khi-deux sur les distributions des nombres de restes pour
chaque site. (zone grisée : test significatif) .................................................................... 355 Tableau 121 : Résultats des tests du khi-deux sur les distributions des esquilles pour chaque
site. ................................................................................................................................. 356 Tableau 122 : Résultats des tests du khi-deux sur les distributions des esquilles des sites de
notre étude et des sites de comparaisons. ....................................................................... 357 Tableau 123 : Résultats des tests de Kolmogorov-Smirnov sur les représentations
squelettiques générales de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu......................................................................................................................................... 358
Tableau 124 : Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour les trois classes de taille (B1, B2C1 et C2) de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. ........................ 359
Tableau 125: Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour la classe de taille B1 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. ............................................................................. 359
Tableau 126 : Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour la classe de taille B2C1 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. ................................................... 360
Tableau 127 : Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour la classe de taille C2 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. ................................................... 360
Tableau 128 : Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour Tanjung 63-64 (B1) avec les sites de comparaison. .................................................................................................................. 361
Tableau 129: Résultats des tests de Kolmogorov-Smirnov sur les représentations squelettiques simplifiées (A, B, C, D et E) pour Sendang Busik (B1) avec les sites de comparaison.362
Tableau 130: Résultats des tests de Kolmogorov-Smirnov sur les classes de taille des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. ............................... 362
Tableau 131 : Résultats des tests de Kolmogorov-Smirnov sur les classes de taille des restes déterminables de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu......... 363

Annexes.
- 371 - 371
Tableau 132 :Résultats des tests de Kolmogorov-Smirnov sur les classes de taille des restes déterminables et des esquilles de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. .............................................................................................................................. 363
Tableau 133: Résultats des tests de Kolmogorov-Smirnov sur les dix classes d’esquilles de Sneed et Folk de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran, Dayu Holon et la Baume de Goulon........................................................................................................... 363
Tableau 134 : : Résultats des tests de Kolmogorov-Smirnov sur les groupes de Voorhies pour les trois classes de taille (B1, B2C1 et C2) de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. .............................................................................................. 364
Tableau 135 : Résultats des tests de Kolmogorov-Smirnov sur les groupes de Voorhies pour la classe de taille B1 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. . 364
Tableau 136 : Résultats des tests de Kolmogorov-Smirnov sur les groupes de Voorhies pour la classe de taille B2C1 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu......................................................................................................................................... 364
Tableau 137 : Résultats des tests de Kolmogorov-Smirnov sur les groupes de Voorhies pour la classe de taille C2 de Tanjung, Sendang Busik, Ngrejeng, Grogol, Bukuran et Dayu. 365





























