Embed Size (px)
Citation preview

Molecular Phylogenetics and Evolution 38 (2006) 794–807www.elsevier.com/locate/ympev
Phylogeny of the ectomycorrhizal mushroom genus Alnicola (Basidiomycota, Cortinariaceae) based on rDNA sequences with special
emphasis on host speciWcity and morphological characters
Pierre-Arthur Moreau a,¤, Ursula Peintner b, Monique Gardes c
a Laboratoire de Botanique, Faculté des Sciences Pharmaceutiques et Biologiques, 3 rue du Professeur Laguesse, BP 83, F-59006 LILLE Cédex, Franceb Institute of Microbiology, University Innsbruck, Technikerstr. 25, A-6020 Innsbruck, Austria
c Laboratoire Evolution et Diversité Biologique (UMR 5174), Bât 4R3, Université Paul Sabatier—Toulouse III, 118 route de Narbonne, F-31062 Toulouse Cédex, France
Received 1 June 2005; revised 7 September 2005; accepted 10 October 2005Available online 28 November 2005
Abstract
Alnicola ( D Naucoria, pro parte) is a mushroom genus of strictly temperate, obligately ectomycorrhizal species, traditionally includedin the family Cortinariaceae. Most Alnicola spp. are primarily host speciWc on Alnus, although a few are mycobionts of Salix or otherhosts. The diVerent species of Alnicola exhibit unique morphological (cystidia, pileipellis) and cytological (dikaryotic or monokaryotichyphae) characters. This makes the genus Alnicola of particular interest for studying the evolution of host speciWcity and morphologicalcharacters in ectomycorrhizal basidiomycetes. We used a combination of classical morphological and phylogenetic methods (rDNA ITSand LSU sequences) to address the following questions: (i) Is Alnicola monophyletic? And (ii) Are characters like host speciWcity ormicroscopical structures synapomorphic for certain clades? The study included nearly all currently known European Alnicola sp. Ourresults demonstrated that, on one hand, the genus Alnicola is polyphyletic, with sistergroup relationships to Hebeloma, Anamika or theclades /Hymenogaster I and /Hymenogaster II. On the other hand, Alnicola splits into three well-supported clades corresponding to thesections Alnicola, Submelinoides, and Salicicolae. The strict host-speciWcity to Alnus is a derived character and has occurred at least twice.The following morphological characters are synapomorphic for deWned clades: the spindle-shaped hymenial cystidia for sect. Alnicola, thehymeniform pileipellis for sect. Submelinoides, and monocaryotic/clampless hyphae for sect. Salicicolae and its sistergroup /Hymenogas-ter II. As a taxonomical consequence, polyphyly of Alnicola implies that the sects. Submelinoides and Salicicolae need to be segregatedfrom Alnicola. 2005 Elsevier Inc. All rights reserved.
Keywords: Alnus ectomycorrhiza; Anamika; Hebeloma; Hymenogaster; ITS and LSU rDNA sequences
1. Introduction
Alnicola Kühner ( D Naucoria (Fr.: Fr.) P. Kumm. pp.) isa genus of strictly temperate, obligately ectomycorrhizalfungi (Kühner, 1926), traditionally included in the familyCortinariaceae (Singer, 1986), tribe Hebelomeae (Kühner,1980). The name Naucoria (in the sense of Moser, 1983) has
* Corresponding author. Fax: +33 3 20 96 43 64.E-mail address: [email protected] (P.-A. Moreau).
1055-7903/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.ympev.2005.10.008
been used as an alternative to Alnicola Kühner, but fornomenclatural and practical reasons Alnicola is used herefollowing Redhead (1984) and Kühner (1987), with A. lute-oloWbrillosa Kühner proposed as the lectotype (Moreau,2005). Most Alnicola spp. are strictly host-speciWc withalders (Alnus spp.), while a small number of species aremycobionts of willows (Salix spp.) or other hosts. The host-speciWcity exhibited by Alnicola is in marked contrast tothat shown in the sister taxa Hebeloma (Aanen et al., 2000b;Bruchet, 1974) and presumably Hymenogaster (accordingto weak evidence for host associations reported in the

P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807 795
literature; Pegler and Young, 1987; Smith, 1966; Söhner,1962). The host-speciWcity suggests a close evolutionaryrelationship between Alnicola and their host plants.
Alnicola is a comparatively small genus with approxi-mately 60 species mentioned in the literature (reviewed byMoreau, 2005). Based on the current classiWcation (Küh-ner, 1981; Singer, 1986), two subgenera are recognized:subgenus Alnicola with spindle-shaped cheilocystidia(about 30 European species) and subgenus Salicicolae(J.E. Lange) Kühner (Lange, 1939; Kühner, 1981) withclub-shaped cheilocystidia (about eight European spe-cies). The latter subgenus is divided into two sections: sect.Salicicolae without clamp connections and with a pseudo-parenchymatous pileipellis, and sect. SubmelinoideaeSinger, with clamp connections and with a hymeniformpileipellis (Singer, 1939; Moreau, 2005). Based on mor-phological and ecological characters, Romagnesi (1942),Kühner (1980), and Singer (1986) postulated the system-atic position of the genus Alnicola in the family Cortinari-aceae, tribe Hebelomeae. This was conWrmed bymolecular analyses focusing on other Cortinariaceae(Aanen et al., 2000a). Peintner et al. (2001) also conWrmedthe phylogenetic aYnities of some species of Hymenogas-ter, a secotioid genus, with Alnicola and Hebeloma, andwith the currently monospeciWc genus Anamika (also theWrst tropical genus reported so far in this tribe, Thomaset al., 2002). The Cortinariaceae are one of the most spe-cies-rich families amongst the Basidiomycota, and manytaxonomic and phylogenetic studies have been devoted tothis family (Aanen et al., 2000a,b; Gulden et al., 2001;Matheny, 2004; Peintner et al., 2001, 2002, 2004; Thomaset al., 2002). However, Alnicola remains the most poorlyknown genus in this family. The rather uniform and unat-tractive macromorphology (Fig. 1) and the diYculty withproperly identifying species are clearly the main reasonfor this neglect. This is further exacerbated by often con-tradictory monographs and/or of primarily local treat-ments (e.g., Bon, 1992; Orton, 1960, 1984; Reid, 1984).
Alnicola comprises a relatively small number of speciescompared to other genera of Cortinariaceae (e.g., Cortina-rius and Hebeloma), but it shows the greatest variety ofmicroscopical structures in the family. Therefore, thisgroup of ectomycorrhizal fungi is particularly attractive for
a combination of classical morphological and phylogeneticstudies. Moreover, the strict host-speciWcity makes thisgenus of particular interest for the study of coevolutionaryprocesses in plant–fungus interactions. Therefore, Alnicolawas chosen as the subject of more extensive research by oneof us (P.-A. M.), including nomenclatural (Horak andMoreau, 2004; Moreau, 2005) and taxonomic revisions(Moreau, 2004; Moreau and Deïana, 2004; Moreau andGarcia, 2005). In this paper, we present phylogenetic analy-ses of a wide range of Alnicola spp. based on rDNAsequences. Due to the small number of taxa analyzed inprevious publications (Aanen et al., 2000a,b; Peintner et al.,2001), the following two main questions remained unsolvedand are addressed in this study: (i) Do Alnicola spp. have amonophyletic origin, and what is the position of Alnicolaspp. in the tribe Hebelomeae (or the /hebelomatoid clade,Peintner et al., 2001; Thomas et al., 2002)? (ii) What mor-phological and ecological (host-speciWcity) characters aresynapomorphic, and what characters are useful in a system-atic reconstruction of the genus Alnicola?
2. Materials and methods
2.1. Taxonomic sampling
Sampling included collections of Alnicola representingall subgenera, sections, and most species as deWned by Küh-ner (1980). Sequences of Hymenogaster spp. and of mostHebeloma spp. included were taken from GenBank. Somespecies in the genus Agrocybe (A. erebia, A. praecox, and A.semiorbicularis) were chosen as the outgroup, as earlierstudies (Moncalvo et al., 2002; Thomas et al., 2002) andpreliminary multiple gene data (including ITS, LSU,EF1alpha, and ATP6) focusing on the relationship ofbrown-spored genera (Peintner, unpublished) have shownAgrocybe to be the best possible sister group. Taxon sam-pling, voucher specimens, geographical locations, and Gen-Bank accession numbers are listed in Table 1. Speciesinterpretations and nomenclature follow Orton (1960),Singer (1977), and Moreau (2005). To distinguish betweenclade names and formal taxonomic epithets, clade namesare not italicized and are preceded with the symbol “/”(Moncalvo et al., 2002).
Fig. 1. Fruitbodies of Alnicola spp. (A) A. umbrina (sect. Alnicola). (B) A. macrospora (sect. Salicicolae). (C) A. inculta (sect. Submelinoides). Photos P.-A.Moreau.
796 P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807
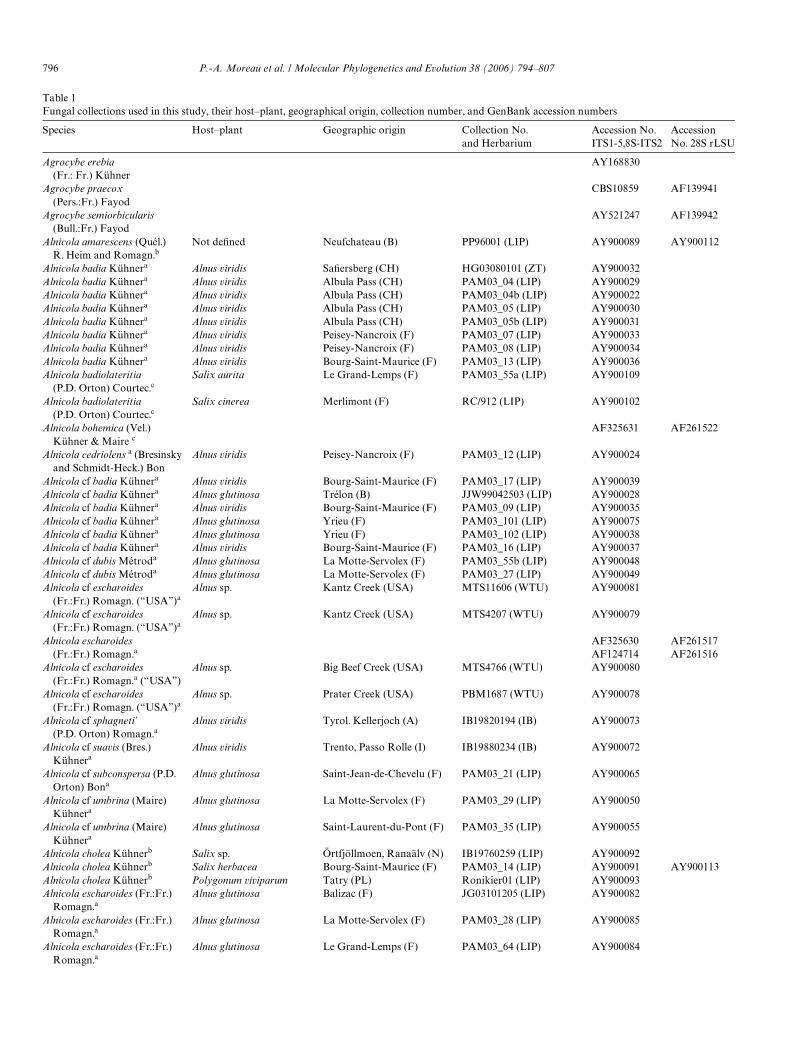
Table 1Fungal collections used in this study, their host–plant, geographical origin, collection number, and GenBank accession numbers
Species Host–plant Geographic origin Collection No. and Herbarium
Accession No. ITS1-5,8S-ITS2
Accession No. 28S rLSU
Agrocybe erebia (Fr.: Fr.) Kühner
AY168830
Agrocybe praecox(Pers.:Fr.) Fayod
CBS10859 AF139941
Agrocybe semiorbicularis (Bull.:Fr.) Fayod
AY521247 AF139942
Alnicola amarescens (Quél.)R. Heim and Romagn.b
Not deWned Neufchateau (B) PP96001 (LIP) AY900089 AY900112
Alnicola badia Kühnera Alnus viridis SaWersberg (CH) HG03080101 (ZT) AY900032Alnicola badia Kühnera Alnus viridis Albula Pass (CH) PAM03_04 (LIP) AY900029Alnicola badia Kühnera Alnus viridis Albula Pass (CH) PAM03_04b (LIP) AY900022Alnicola badia Kühnera Alnus viridis Albula Pass (CH) PAM03_05 (LIP) AY900030Alnicola badia Kühnera Alnus viridis Albula Pass (CH) PAM03_05b (LIP) AY900031Alnicola badia Kühnera Alnus viridis Peisey-Nancroix (F) PAM03_07 (LIP) AY900033Alnicola badia Kühnera Alnus viridis Peisey-Nancroix (F) PAM03_08 (LIP) AY900034Alnicola badia Kühnera Alnus viridis Bourg-Saint-Maurice (F) PAM03_13 (LIP) AY900036Alnicola badiolateritia
(P.D. Orton) Courtec.cSalix aurita Le Grand-Lemps (F) PAM03_55a (LIP) AY900109
Alnicola badiolateritia(P.D. Orton) Courtec.c
Salix cinerea Merlimont (F) RC/912 (LIP) AY900102
Alnicola bohemica (Vel.) Kühner & Maire c
AF325631 AF261522
Alnicola cedriolens a (Bresinsky and Schmidt-Heck.) Bon
Alnus viridis Peisey-Nancroix (F) PAM03_12 (LIP) AY900024
Alnicola cf badia Kühnera Alnus viridis Bourg-Saint-Maurice (F) PAM03_17 (LIP) AY900039Alnicola cf badia Kühnera Alnus glutinosa Trélon (B) JJW99042503 (LIP) AY900028Alnicola cf badia Kühnera Alnus viridis Bourg-Saint-Maurice (F) PAM03_09 (LIP) AY900035Alnicola cf badia Kühnera Alnus glutinosa Yrieu (F) PAM03_101 (LIP) AY900075Alnicola cf badia Kühnera Alnus glutinosa Yrieu (F) PAM03_102 (LIP) AY900038Alnicola cf badia Kühnera Alnus viridis Bourg-Saint-Maurice (F) PAM03_16 (LIP) AY900037Alnicola cf dubis Métroda Alnus glutinosa La Motte-Servolex (F) PAM03_55b (LIP) AY900048Alnicola cf dubis Métroda Alnus glutinosa La Motte-Servolex (F) PAM03_27 (LIP) AY900049Alnicola cf escharoides
(Fr.:Fr.) Romagn. (“USA”)aAlnus sp. Kantz Creek (USA) MTS11606 (WTU) AY900081
Alnicola cf escharoides (Fr.:Fr.) Romagn. (“USA”)a
Alnus sp. Kantz Creek (USA) MTS4207 (WTU) AY900079
Alnicola escharoides AF325630 AF261517(Fr.:Fr.) Romagn.a AF124714 AF261516
Alnicola cf escharoides(Fr.:Fr.) Romagn.a (“USA”)
Alnus sp. Big Beef Creek (USA) MTS4766 (WTU) AY900080
Alnicola cf escharoides (Fr.:Fr.) Romagn. (“USA”)a
Alnus sp. Prater Creek (USA) PBM1687 (WTU) AY900078
Alnicola cf sphagneti’ (P.D. Orton) Romagn.a
Alnus viridis Tyrol. Kellerjoch (A) IB19820194 (IB) AY900073
Alnicola cf suavis (Bres.) Kühnera
Alnus viridis Trento, Passo Rolle (I) IB19880234 (IB) AY900072
Alnicola cf subconspersa (P.D. Orton) Bona
Alnus glutinosa Saint-Jean-de-Chevelu (F) PAM03_21 (LIP) AY900065
Alnicola cf umbrina (Maire) Kühnera
Alnus glutinosa La Motte-Servolex (F) PAM03_29 (LIP) AY900050
Alnicola cf umbrina (Maire) Kühnera
Alnus glutinosa Saint-Laurent-du-Pont (F) PAM03_35 (LIP) AY900055
Alnicola cholea Kühnerb Salix sp. Örtfjöllmoen, Ranaälv (N) IB19760259 (LIP) AY900092Alnicola cholea Kühnerb Salix herbacea Bourg-Saint-Maurice (F) PAM03_14 (LIP) AY900091 AY900113Alnicola cholea Kühnerb Polygonum viviparum Tatry (PL) Ronikier01 (LIP) AY900093Alnicola escharoides (Fr.:Fr.)
Romagn.aAlnus glutinosa Balizac (F) JG03101205 (LIP) AY900082
Alnicola escharoides (Fr.:Fr.) Romagn.a
Alnus glutinosa La Motte-Servolex (F) PAM03_28 (LIP) AY900085
Alnicola escharoides (Fr.:Fr.) Romagn.a
Alnus glutinosa Le Grand-Lemps (F) PAM03_64 (LIP) AY900084
P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807 797
(continued on next page)
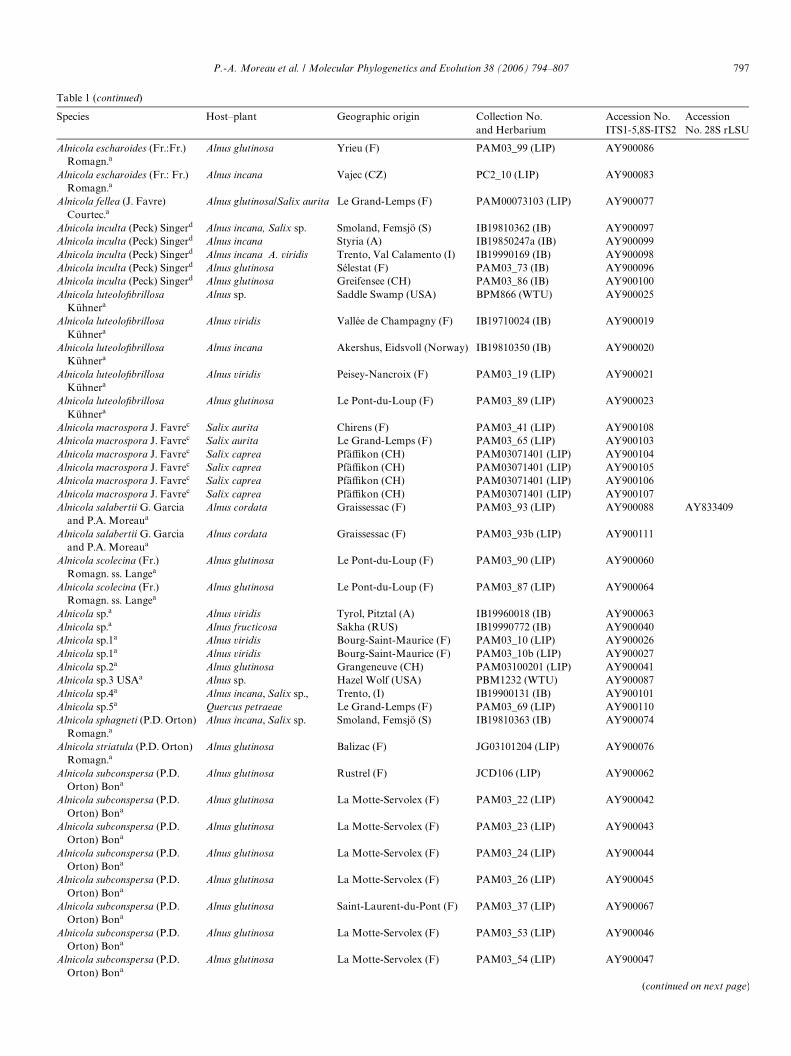
Table 1 (continued)
Species Host–plant Geographic origin Collection No. and Herbarium
Accession No. ITS1-5,8S-ITS2
Accession No. 28S rLSU
Alnicola escharoides (Fr.:Fr.) Romagn.a
Alnus glutinosa Yrieu (F) PAM03_99 (LIP) AY900086
Alnicola escharoides (Fr.: Fr.) Romagn.a
Alnus incana Vajec (CZ) PC2_10 (LIP) AY900083
Alnicola fellea (J. Favre) Courtec.a
Alnus glutinosa/Salix aurita Le Grand-Lemps (F) PAM00073103 (LIP) AY900077
Alnicola inculta (Peck) Singerd Alnus incana, Salix sp. Smoland, Femsjö (S) IB19810362 (IB) AY900097Alnicola inculta (Peck) Singerd Alnus incana Styria (A) IB19850247a (IB) AY900099Alnicola inculta (Peck) Singerd Alnus incana A. viridis Trento, Val Calamento (I) IB19990169 (IB) AY900098Alnicola inculta (Peck) Singerd Alnus glutinosa Sélestat (F) PAM03_73 (IB) AY900096Alnicola inculta (Peck) Singerd Alnus glutinosa Greifensee (CH) PAM03_86 (IB) AY900100Alnicola luteoloWbrillosa
KühneraAlnus sp. Saddle Swamp (USA) BPM866 (WTU) AY900025
Alnicola luteoloWbrillosa Kühnera
Alnus viridis Vallée de Champagny (F) IB19710024 (IB) AY900019
Alnicola luteoloWbrillosa Kühnera
Alnus incana Akershus, Eidsvoll (Norway) IB19810350 (IB) AY900020
Alnicola luteoloWbrillosa Kühnera
Alnus viridis Peisey-Nancroix (F) PAM03_19 (LIP) AY900021
Alnicola luteoloWbrillosa Kühnera
Alnus glutinosa Le Pont-du-Loup (F) PAM03_89 (LIP) AY900023
Alnicola macrospora J. Favrec Salix aurita Chirens (F) PAM03_41 (LIP) AY900108Alnicola macrospora J. Favrec Salix aurita Le Grand-Lemps (F) PAM03_65 (LIP) AY900103Alnicola macrospora J. Favrec Salix caprea PfäYkon (CH) PAM03071401 (LIP) AY900104Alnicola macrospora J. Favrec Salix caprea PfäYkon (CH) PAM03071401 (LIP) AY900105Alnicola macrospora J. Favrec Salix caprea PfäYkon (CH) PAM03071401 (LIP) AY900106Alnicola macrospora J. Favrec Salix caprea PfäYkon (CH) PAM03071401 (LIP) AY900107Alnicola salabertii G. Garcia
and P.A. MoreauaAlnus cordata Graissessac (F) PAM03_93 (LIP) AY900088 AY833409
Alnicola salabertii G. Garcia and P.A. Moreaua
Alnus cordata Graissessac (F) PAM03_93b (LIP) AY900111
Alnicola scolecina (Fr.) Romagn. ss. Langea
Alnus glutinosa Le Pont-du-Loup (F) PAM03_90 (LIP) AY900060
Alnicola scolecina (Fr.) Romagn. ss. Langea
Alnus glutinosa Le Pont-du-Loup (F) PAM03_87 (LIP) AY900064
Alnicola sp.a Alnus viridis Tyrol, Pitztal (A) IB19960018 (IB) AY900063Alnicola sp.a Alnus fructicosa Sakha (RUS) IB19990772 (IB) AY900040Alnicola sp.1a Alnus viridis Bourg-Saint-Maurice (F) PAM03_10 (LIP) AY900026Alnicola sp.1a Alnus viridis Bourg-Saint-Maurice (F) PAM03_10b (LIP) AY900027Alnicola sp.2a Alnus glutinosa Grangeneuve (CH) PAM03100201 (LIP) AY900041Alnicola sp.3 USAa Alnus sp. Hazel Wolf (USA) PBM1232 (WTU) AY900087Alnicola sp.4a Alnus incana, Salix sp., Trento, (I) IB19900131 (IB) AY900101Alnicola sp.5a Quercus petraeae Le Grand-Lemps (F) PAM03_69 (LIP) AY900110Alnicola sphagneti (P.D. Orton)
Romagn.aAlnus incana, Salix sp. Smoland, Femsjö (S) IB19810363 (IB) AY900074
Alnicola striatula (P.D. Orton) Romagn.a
Alnus glutinosa Balizac (F) JG03101204 (LIP) AY900076
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa Rustrel (F) JCD106 (LIP) AY900062
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa La Motte-Servolex (F) PAM03_22 (LIP) AY900042
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa La Motte-Servolex (F) PAM03_23 (LIP) AY900043
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa La Motte-Servolex (F) PAM03_24 (LIP) AY900044
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa La Motte-Servolex (F) PAM03_26 (LIP) AY900045
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa Saint-Laurent-du-Pont (F) PAM03_37 (LIP) AY900067
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa La Motte-Servolex (F) PAM03_53 (LIP) AY900046
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa La Motte-Servolex (F) PAM03_54 (LIP) AY900047
798 P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807
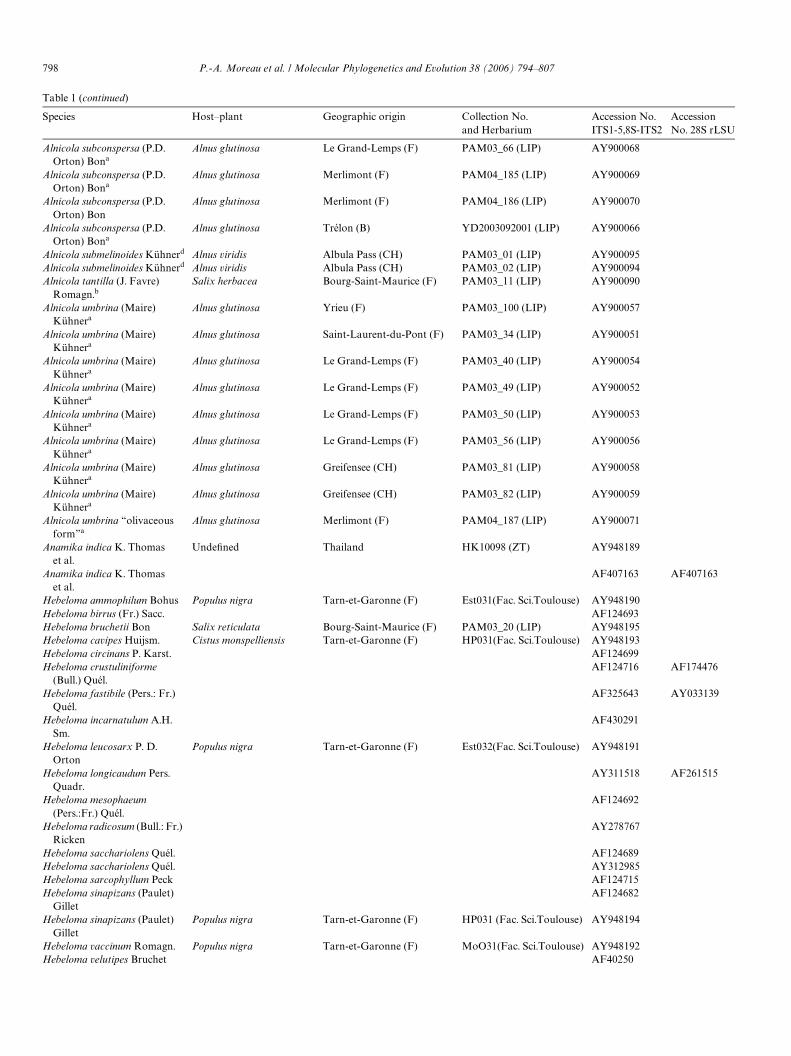
Table 1 (continued)
Species Host–plant Geographic origin Collection No. and Herbarium
Accession No. ITS1-5,8S-ITS2
Accession No. 28S rLSU
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa Le Grand-Lemps (F) PAM03_66 (LIP) AY900068
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa Merlimont (F) PAM04_185 (LIP) AY900069
Alnicola subconspersa (P.D. Orton) Bon
Alnus glutinosa Merlimont (F) PAM04_186 (LIP) AY900070
Alnicola subconspersa (P.D. Orton) Bona
Alnus glutinosa Trélon (B) YD2003092001 (LIP) AY900066
Alnicola submelinoides Kühnerd Alnus viridis Albula Pass (CH) PAM03_01 (LIP) AY900095Alnicola submelinoides Kühnerd Alnus viridis Albula Pass (CH) PAM03_02 (LIP) AY900094Alnicola tantilla (J. Favre)
Romagn.bSalix herbacea Bourg-Saint-Maurice (F) PAM03_11 (LIP) AY900090
Alnicola umbrina (Maire) Kühnera
Alnus glutinosa Yrieu (F) PAM03_100 (LIP) AY900057
Alnicola umbrina (Maire) Kühnera
Alnus glutinosa Saint-Laurent-du-Pont (F) PAM03_34 (LIP) AY900051
Alnicola umbrina (Maire) Kühnera
Alnus glutinosa Le Grand-Lemps (F) PAM03_40 (LIP) AY900054
Alnicola umbrina (Maire) Kühnera
Alnus glutinosa Le Grand-Lemps (F) PAM03_49 (LIP) AY900052
Alnicola umbrina (Maire) Kühnera
Alnus glutinosa Le Grand-Lemps (F) PAM03_50 (LIP) AY900053
Alnicola umbrina (Maire) Kühnera
Alnus glutinosa Le Grand-Lemps (F) PAM03_56 (LIP) AY900056
Alnicola umbrina (Maire) Kühnera
Alnus glutinosa Greifensee (CH) PAM03_81 (LIP) AY900058
Alnicola umbrina (Maire) Kühnera
Alnus glutinosa Greifensee (CH) PAM03_82 (LIP) AY900059
Alnicola umbrina “olivaceous form”a
Alnus glutinosa Merlimont (F) PAM04_187 (LIP) AY900071
Anamika indica K. Thomas et al.
UndeWned Thailand HK10098 (ZT) AY948189
Anamika indica K. Thomas et al.
AF407163 AF407163
Hebeloma ammophilum Bohus Populus nigra Tarn-et-Garonne (F) Est031(Fac. Sci.Toulouse) AY948190Hebeloma birrus (Fr.) Sacc. AF124693Hebeloma bruchetii Bon Salix reticulata Bourg-Saint-Maurice (F) PAM03_20 (LIP) AY948195Hebeloma cavipes Huijsm. Cistus monspelliensis Tarn-et-Garonne (F) HP031(Fac. Sci.Toulouse) AY948193Hebeloma circinans P. Karst. AF124699Hebeloma crustuliniforme
(Bull.) Quél.AF124716 AF174476
Hebeloma fastibile (Pers.: Fr.) Quél.
AF325643 AY033139
Hebeloma incarnatulum A.H. Sm.
AF430291
Hebeloma leucosarx P. D. Orton
Populus nigra Tarn-et-Garonne (F) Est032(Fac. Sci.Toulouse) AY948191
Hebeloma longicaudum Pers. Quadr.
AY311518 AF261515
Hebeloma mesophaeum (Pers.:Fr.) Quél.
AF124692
Hebeloma radicosum (Bull.: Fr.) Ricken
AY278767
Hebeloma sacchariolens Quél. AF124689Hebeloma sacchariolens Quél. AY312985Hebeloma sarcophyllum Peck AF124715Hebeloma sinapizans (Paulet)
GilletAF124682
Hebeloma sinapizans (Paulet) Gillet
Populus nigra Tarn-et-Garonne (F) HP031 (Fac. Sci.Toulouse) AY948194
Hebeloma vaccinum Romagn. Populus nigra Tarn-et-Garonne (F) MoO31(Fac. Sci.Toulouse) AY948192Hebeloma velutipes Bruchet AF40250

P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807 799
2.2. Morphological observations
The following characters were investigated anddescribed qualitatively from fresh or dry fruitbodies revivedin 5% KOH: (i) macromorphology, including shape (agari-coid/gastroid), colors, gill insertion, development of the veil,striations on pileus, organoleptic properties; (ii) micromor-phology, including spore shape and size, ornamentation,color in 5% KOH and Melzer’s reagent, shape of cheilocy-stidia, pileal structure, presence of clamp connections; (iii)cytology, including number of nuclei per hypha in tramaand subhymenium (according to Clémençon, 1985).
2.3. Molecular techniques
DNA isolation and sequencing protocols followed thoseof previous studies by Gardes and Bruns (1993) and Peint-ner et al. (2001). Primers used for PCR ampliWcation andsequencing of the ITS region were ITS1-F and ITS4-B(Gardes and Bruns, 1993). For the ampliWcation of therDNA LSU, the primer pair LROR and LR5 was used, andsequencing was performed with the primers LR3R andLR16. AmpliWcation protocols followed thermal cyclingconditions described by Vilgalys and Hester (1990) andGardes and Bruns (1993). Sequencing was performed by anindependent laboratory (Genome Express, Grenoble, F).
2.4. Phylogenetic analyses
Sequences were aligned manually and regions of ambig-uous alignment were excluded from the analyses. If not oth-erwise stated, gaps were treated as missing data. Maximumparsimony (MP) analyses were carried out with the com-puter program PAUP* 4.0b10 (SwoVord, 2003). Heuristicsearches were conducted with 100 random sequence addi-tion replicates, holding a single tree at each step, MaxTreesset to 2000, and TBR branch-swapping. Branch supportwas assessed by nonparametric bootstrapping with 100
pseudoreplicates, simple taxon addition in each replicate,with Wve trees held at each step, and NNI branch-swapping(Salamin et al., 2003).
Bayesian analysis of the datasets was conducted by usingMrBayes 3.0B2 (Huelsenbeck and Ronquist, 2001). Each ofthe four Markov chains, three heated and one cold, wasstarted from a random tree, and all four chains ran simulta-neously for 3 £ 106 generations, with trees being sampledevery 100 generations for a total of 30,000 trees. After thelikelihoods of the trees in each chain converged, we dis-carded the Wrst 600–700 trees as burn in. Branch lengths ofthe trees were saved and a Bayesian consensus phylogrambased on mean branch lengths was calculated for each runbased on the remaining trees. Branch lengths for the Wguresare maximum-likelihood estimates and are scaled in termsof expected numbers of nucleotide substitutions per site.These trees did not diVer signiWcantly between runs, there-fore the consensus phylogram resulting from the thirdBayesian analysis was randomly chosen to represent thephylogeny of Alnicola (Fig. 2). Posterior probabilities(BPP) were made by using the computer program PAUP*:a majority-rule consensus tree was calculated based ontrees remaining after the burn from all three Bayesian runs.
A Bayesian analysis to test for the monophyly of Alni-cola was conducted by adding an input Wle containing a treeresolved only at a node leading to all Alnicola spp., andconducting a constrained Bayesian analysis as describedabove. Consensus phylograms obtained in the constrainedand unconstrained analyses were compared in PAUP usingthe Shimodaira–Hasegawa (SH) test (Shimodaira andHasegawa, 1999) and RELL approximation with 1000bootstrap replicates (Goldman and Whelan, 2000).
Clades with BPP higher than 95% and a few clades withBPP higher than 70% considered as systematically signiW-cant (supported by strong morphological characters), werenamed, as far as possible, according to the traditional clas-siWcations. No formal epithets are applied to clade names,as this will be done in a separate publication.
Table 1 (continued)
a Alnicola (subgen. Alnicola) sect. Alnicola.b Alnicola (subgen. Alnicola) sect. Amarescens (Kühner, 1981).c Alnicola (subgen. Salicicolae) sect. Salicicolae (Lange, 1939; Kühner, 1981).d Alnicola (subgen. Salicicolae) sect. Submelinoideae (Singer, 1939).
Species Host–plant Geographic origin Collection No. and Herbarium
Accession No. ITS1-5,8S-ITS2
Accession No. 28S rLSU
Hymenogaster alnicola A. H. Sm.
AF325632
Hymenogaster alnicola A. H. Sm.
AF325632
Hymenogaster bulliardii Vitt. AF325641Hymenogaster griseus Vitt. AF325636 DQ133941Hymenogaster olivaceus Vitt. AF325642Hymenogaster parksii Zeller &
DodgeAF325638
Hymenogaster subalpinus A.H. Sm.
AF325640 AF325640
Hymenogaster tener Berk. & Broome
AF325633 AF325633

800 P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807
A second analysis was carried out with a restricted set of65 ITS sequences focusing on /Alnicola. Alnicola incultawas used as the outgroup. Analyses were carried out asabove, except the sequences were more easily aligned andtherefore more characters could be included in the analysis.Gaps were treated as a 5th character in the MP search.
To infer basal relationships among subclades of the/hebelomoid, a combined alignment of 17 rDNA ITS andLSU sequences was generated. Most LSU sequences weregenerated using the same DNA extract as for the ITSsequence, the rest was downloaded from GenBank. ITS andLSU sequences were analyzed together as described above.
3. Results
3.1. Sequence alignments and phylogenetic analyses
The alignment of the 112 rDNA ITS sequences consistedof 859 characters of which 595 were included, 308 were con-stant, and 234 were parsimony informative. The MP analy-sis of the whole dataset yielded 2000 equally parsimonioustrees (tree score D 814 steps, CI D 0.5319, homoplasy index(HI) D 0.4681, RI D 0.8306, rescaled consistency index(RC) D 0.4418). The MP trees are not shown, as they do notdiVer signiWcantly from trees resulting from the Bayesiananalysis.
The Bayesian consensus phylogram shown (Fig. 2) wascalculated using 9300 trees from the single randomly cho-sen Bayesian analysis and BPP were computed with theoverall 23,000 trees remaining after burn in.
Shimodaira–Hasegawa tests rejected monophyly of thegenus Alnicola, but monophyly of subgenus Alnicola (corre-sponding to /Alnicola) was not rejected (Table 2). Mono-phyly of subgenus Salicicolae (corresponding to the twoclades /Bohemica and /Submelinoides) was rejected.
The following further hypotheses were also tested usingconstrained analyses: monophyly of Alnicola spp. with thesame type of cystidia (subgenus Alnicola and subgenus Sal-icicolae, respectively); monophyly of species withoutclamps (/Hymenogaster II and /Bohemica), monophyleticgroups with host speciWcity (Alnicola spp. strictly associatedwith Alnus, and Alnicola spp. associated with Salix or withother hosts), monophyly of the two clades of Hymenogas-ter. Monophyletic origins for all of these clades wererejected (Table 2).
The alignment of the smaller, restricted 65 rDNA ITSdataset consisted of 859 characters of which 595 wereincluded, 431 were constant, and 124 were parsimony infor-mative. The MP analysis yielded 2000 equally parsimonioustrees (tree score D 266 steps, CI D 0.7556, HI D 0.2444,RI D 0.8997, RC D 0.6798) (trees not shown). The Bayesianconsensus phylogram was calculated using 9500 trees fromthe third Bayesian analysis. This phylogram is not shown asit only diVers from the larger one shown in Fig. 2 by highersupport values (bootstrap/BPP) for the clades /Alnicola(100/100), /Alnicola ss. str. (72/), and by the moderately sup-ported sistergroup relationship of /Alnicola s.s. to the cladecontaining A. “sphagneti” (84/86).
The combined dataset of ITS and LSU sequences includeda total 17 taxa; Agrocybe praecox and A. semiorbicularis were
Fig. 2. Consensus phylogram of 9300 trees as calculated with the program MrBayes. BPP and BS values >70 are given besides branches as BPP/BS. Well-supported branches (BPP D 100 or BS > 90%) are in boldface. Gray branches have BPP and BS < 50. Species names in larger letters represent severalsequences of the same Alnicola morphospecies (refer to Table 1 for collection numbers).
100/100
100/97
/100
97/61
100/62
55/
Alnicola luteolofibrillosa
Alnicola escharoides
Alnicola escharoides
Alnicola salabertii
Alnicola amarescens
Alnicola cholea
Hymenogaster griseus
Hy. subalpinus
Alnicola submelinoides
Alnicola bohemica
Hymenogaster tener
Anamika indica
Hebeloma fastibile
Hebeloma crustuliniforme
Hebeloma longicaudum
Agrocybe praecox
Agrocybe semiorbicularis
/heb
elom
eae
clad
e
/ALNICOLA
/HYMENOGASTER I
/HYMENOGASTER II
/BOHEMICA
/SUBMELINOIDES
/ANAMIKA
/HEBELOMA
/AGROCYBE
/AL
NIC
OL
A S.S.
0.01 substitutions/site
66/62

P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807 801
used as the outgroup. The alignment of these 17 sequencesconsisted of 1609 included characters: 1268 were constant,178 were parsimony informative. The MP analysis of thiscombined dataset yielded 21 equally parsimonious trees(tree score D 603 steps, CI D 0.6998, HI D 0.3002, RI D0.5690, RCD0.3982). These trees were topologically identicalto the Bayesian consensus phylograms. Bayesian analyseswere run as described above (trees not shown). The Bayes-ian consensus phylogram was calculated using 9500trees from the third Bayesian analysis, and BPP were calcu-lated with a total of 29,253 trees left after the burnin(Fig. 3).
3.2. The clades of Alnicola
All Anamika, Hebeloma, Hymenogaster, and Alnicolaspecies included in this study fall into a well-supported (BS100, BPP 100) clade (Fig. 2), which is consistent with the /
hebelomatoid identiWed by Moncalvo et al. (2002). Thegenus Alnicola, as currently deWned, is polyphyletic: treetopologies consistently resolve three clades of Alnicola spp.(Fig. 2 from top down): subclade /Alnicola (consisting ofspecies of subgenus Alnicola, BPP 78, but see below aboutFig. 3), subclade /Submelinoides (sect. Submelinoides, BPP99), and subclade /Bohemica (sect. Salicicolae ss. str., BPP100). Other, moderately supported subclades of /hebelomatoidare /Anamika, /Hebeloma, /Hymenogaster II, and /Hyme-nogaster I. Relationships between the major clades of/hebelomatoid are unresolved.
The clade /Alnicola (Fig. 2) is characterized by shortbranches, which reXect the low divergence found in the ribo-somal ITS gene identiWed in the second analysis. Due to theinclusion of more characters in the second analysis (Fig. 3),/Alnicola is well supported with BPP 100 and BS 100. Twosubclades surrounding A. cholea and A. amarescens–A. tan-tilla form a monophyletic sister group to /Alnicola ss. str.
Table 2Shimodaira–Hasegawa tests (one-tailed RELL bootstrap with 1000 replicates) for constrained vs. unconstrained tree topologies (consensus phylogramsresulting from the Bayesian analysis)
¤ P < 0.05 are signiWcantly diVerent and written in bold letters.
Constraints to monophyly -lnL DiV P
none (2nd chain) 5 454.5880 0 bestAlnicola, Hebeloma and Hymenogaster 5 750.9754 252.6215 0.000¤
Alnicola 5 708.3903 253.8023 0.000¤
Hymenogaster 5 715.1425 260.5545 0.000¤
Subgenus Alnicola 5 496.5513 41.96332 0.296Alnicola spp. with the same type of cystidia
(subgenus Alnicola and subgenus Submelinoides)5 802.6675 304.3134 0.000¤
Species without clamps (/Bohemica and /Hymenogaster II) 5 570.0225 115.4345 0.007¤
Host-speciWc monophyletic clades(either strict Alnus-speciWc or with Salix and other hosts)
5 612.8746 158.2865 0.000¤
Fig. 3. Consensus phylogram of 9500 trees as calculated with the program MrBayes for ITS and LSU sequences. BPP and BS values >70 are given besidesbranches as BPP/BS. Well-supported branches (BPP > 95) are boldface (refer to Table 1 for collections number).

802 P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807
Other distinct clades are formed by species classiWed as A.luteoloWbrillosa, A. badia, “A. escharoides USA,” A. escharo-ides, and A. salabertii. Due to their low divergence, the fol-lowing taxa cannot easily be distinguished based on ITSsequences: A. dubis/A. scolecina/A. subconspersa/A. umbrina.
Analysis of combined ITS and LSU data (Fig. 3)revealed the same clades as in the ITS phylogeny. Relation-ships between the major clades cannot be resolved, but inthis phylogeny both Alnicola and Hymenogaster arepolyphyletic: /Alnicola (agaricoid) has close relationshipsto /Hymenogaster I (gasteroid), while /Bohemica (agari-coid) has a weakly supported sistergroup relationship to/Hymenogaster II (gasteroid).
3.3. Morphology of studied collections
All morphological features reported by recent authorsincluding macromorphology, cystidia shape, spore size, andornamentation were conWrmed (Bon, 1992; Kühner, 1931;Lange, 1939; Moreau, 2004; Orton, 1960; Reid, 1984;Romagnesi, 1942). Presence of clamp connections and pilei-
pellis structure were often insuYciently documented by theabovementioned authors and have been checked for all col-lections. We found the following diVerent types of pileipel-lis structures (Fig. 4) to be characteristic for genera,subgenera or sections supported by the rDNA data (termi-nology based on Clémençon, 2004):
• Tomentocutis with repent incrusted hyphae, on a pseu-doparenchymatous subpellis: A. bohemica, A. cholea,A. rubriceps, Anamika indica.
• Tomentocutis with smooth +/¡ erected hyphae, on apseudoparenchymatous subpellis: A. amarescens, A. fel-lea, A. geraniolens, A. salabertii, A. tantilla.
• Ixotomentocutis, usually on a pseudoparenchymatoussubpellis: Hebeloma spp.
• Irregular epidermioid cutis (pseudoparenchymatous toepithelioid), without or with incomplete suprapellis:A. badiolateritia, A. macrospora.
• Hymeniderm: A. inculta, A. submelinoides.• Irregular trichoderm (with transitions to epithelium),
with or without superWcial velar hyphae: A. badia,
Fig. 4. Synthetic phylogenetic reconstruction of tribe Hebelomeae based on phylogram (Fig. 3), in relation with signiWcant macromorphological, micro-morphological, and cytological characteristics of fruitbodies. Legends for pileal structures (A–E) in the text (3.3.).

P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807 803
A. dubis, A. escharoides, A. luteoloWbrillosa, A. scolecina,A. striatula, A. suavis, A. subconspersa, A. umbrina, A.spp (all unidentiWed collections).
Clamp connections were found in all species of Hebelo-meae except Alnicola sect. Salicicolae (/Bohemica) and someHymenogaster (/Hymenogaster II). Kühner (1942) reportedAlnicola bohemica as the Wrst case amongst Cortinariaceae ofa fruitbody consisting of monokaryotic hyphae only. One ofus (P.-A. M.) investigated the cytology of all clampless spe-cies according to the methods described by Kühner (1942)and found them all to be monokaryotic (in the subhyme-nium). This cytological anomaly deWnes two sister clades: oneagaricoid (/BohemicaDAlnicola sect. Salicicolae) and onegasteroid (/Hymenogaster II DHymenogaster pp.).
4. Discussion
4.1. Phylogeny of Alnicola
Ribosomal sequence data demonstrate that all species ofthis large /hebelomeae clade, although morphologicallyquite variable, are comparatively conservative within theirrDNA ITS region compared to, e.g., Galerina (Gulden et al.,2001) or Leccinum (Den Bakker et al., 2004). We were ableto align most nucleotide positions of all ingroup taxa.Other studies found similar low levels of variation in ITSsequences in the ectomycorrhizal genera Hebeloma andCortinarius (Aanen et al., 2000a,b; Peintner et al., 2004).
However, despite this low level of variation, rDNAsequence data are very useful for addressing the systematicsand evolution of the genus Alnicola. It is clear that speciescurrently classiWed in the genus Alnicola are polyphyletic inorigin. The subgenus Alnicola forms a distinct, and well-supported clade. In addition, the support for this cladeincreases with increased character sampling (Fig. 3). Theanalysis focusing on /Alnicola (tree not shown, see Fig. 2),with the A. inculta clade as an outgroup, also clearly dem-onstrates that /Alnicola is a distinct clade. The distinctclade including A. amarescens is the sister-group of /Alni-cola s.s (here including clade A. “sphagneti”) and Alnicolacholea appears to be a sister group to the two former clades.In contrast, species of Alnicola subgenus Submelinoidesconsistently form two well-supported clades (BPP > 95%)with possible sister-group relationships to Hebeloma/Ana-mika or to /Hymenogaster II.
4.2. Ecology and host speciWcity
The strict and characteristic ectomycorrhizal associationbetween Alnicola and Alnus is observed exclusively in twoclades (Fig. 4): /Submelinoides and /Alnicola, but the latterwith the exclusion of the A. cholea and the A. amarescensgroups ( D /Alnicola sensu stricto). In these two last groupsthe species are associated with Salicaceae and other hostplants, e.g., Polygonum viviparum. Also species of /Bohe-mica are not associated with Alnus, but mainly with Salica-
ceae. Thus, the present study strongly suggests that thestrict association of Alnicola spp. with Alnus is a derivedcharacter, which has evolved independently at least twice:in /Alnicola sensu stricto and in /Submelinoides.
When comparing host speciWcity of Alnicola spp. withHebeloma spp., it is striking that no Hebeloma species isknown to occur under Alnus, even occasionally; but a num-ber of Hebeloma spp. are strictly or preferentially associ-ated with Salicaceae (Bon, 2002; Bruchet, 1974; Favre,1948; Vesterholt, 1989). In the genus Hymenogaster, as faras the information is available to us, only one species(Hy. alnicola) is strictly associated with Alnus (Smith, 1966)but more information on the ecology of Hymenogaster speciesis required for a more precise interpretation of these data.
Our phylogenetic reconstruction based on rDNAsequences implies that Alnus host specialists evolved fromgeneralist ancestors with a wider host range. This process isprobably still going on in /Alnicola sensu stricto: AlnicolaluteoloWbrillosa is a generalist symbiont associated with var-ious species of Alnus and may have diverged early in theevolution of this lineage. Other Alnicola species are associ-ated with only one species of Alnus (e.g., A. salabertii withAlnus cordata, or A. umbrina and A. subconspersa withAlnus glutinosa). Another example of a narrow host range isfound in /Submelinoides: A. inculta is so far only reportedunder Alnus sect. Alnus (A. glutinosa and A. incana inEurope), while Alnicola submelinoides is, with very fewexceptions, found under Alnus viridis. This pattern suggeststhat these Alnicola mutualists specialized to their alderhosts, but this hypothesis needs to be conWrmed by furtheranalyses including more extra-European taxa and usingcomparative methods with a fully resolved Alnus phylogeny(Navarro et al., 2003; Chen and Li, 2004).
Our phylogenetic analyses suggest that the associationof Alnicola spp. with Alnus results from a very strong, irre-versible adaptation of the fungus to the host. If it occurs,this coevolutionary process might also explain the low spe-cies diversity amongst Alnus-associated ectomycorrhizalfungi (dominated by Alnicola) compared to other host-trees(Brunner and Horak, 1990; Bujakiewicz, 1989; Dorninger,1993; Molina, 1981).
More generally, the putative coevolution of Betulaceae–Salicaceae and Hebelomeae may have resulted in cospecia-tion of both mycobionts and their hosts, driven by hostswitching or restricting of host speciWcity. The more or lessXexible symbiotic association of Betulaceae-Salicaceae withHebelomeae is a plesiomorphic character. Similar observa-tions were made also for other groups of ectomycorrhizalfungi, e.g., for Rhizopogon spp. (Molina and Trappe, 1994)or Leccinum spp. (Den Bakker et al., 2004). No species ofthese genera are reported in association with Alnus.
4.3. General variability of morphological characters in Hebelomeae
Comparing tribe Hebelomeae to the closely related tribeCortinarieae (including Cortinarius, Cuphocybe, Rozites,

804 P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807
Thaxterogaster, etc.; corresponding to the /cortinarioidclade of Peintner et al., 2001, 2002), the overall variabilityof characters such as spore ornamentation and shape, pilealstructure, presence of a veil, colors and organoleptic char-acters is similar, or even greater, in Hebelomeae (Kühner,1980; Peintner et al., 2001; Singer, 1986). In both tribes,sequestrate species, species without clamp connections, andspecies with peculiar pileal structures (e.g., a hymenoderm)occur. However, more species with two-spored basidia(exceptional in Cortinarieae) and proportionally manymore host-speciWc mycorrhizal species (with Betulaceaeand Salicaceae) belong to the Hebelomeae, unless Corti-narieae is found to be more species-rich (500–2500 speciesin Europe; Courtecuisse and Duhem, 1994; Kirk et al.,2001).
4.4. Pileal structure and cystidia
The pileal diVerentiation in the Hebelomeae is the mostimportant amongst all the Cortinariaceae (Kühner, 1980).Members of the genus Hebeloma usually have a thick, pseu-doparenchymatous subpellis, and a more or less developedixotomentocutis (Bon, 2002; Bruchet, 1970). Similar but notgelatinized pileipellis structures are found in /Anamika(Thomas et al., 2002) and /Bohemica pp. (A. bohemica, A.rubriceps; Kühner, 1981; Orton, 1984). In contrast, all taxaof Submelinoides have a typical hymeniform pileipellis; thischaracter can be regarded as a synapomorphy for /Submeli-noides.
Pileipellis structures of species of /Alnicola (includingboth the A. cholea and the A. amarescens groups) are simi-lar to those in Hebeloma, /Anamika, and /Bohemica. Allspecies of this clade have a dry tomentocutis with pseudo-parenchymatous subpellis, but without a gelatinized supra-pellis (Kühner, 1981; Fig. 4).
Cheilo- and caulocystidia are lanceolate (Urtica-like) inAlnicola, with thickened walls in /A. amarescens and /A.cholea. In contrast, in /Submelinoides and /Bohemica,cheilo- and caulocystidia are cylindro-claviform (Singer,1986; Fig. 4).
4.5. Clamp connections, cytology, and sexuality
All species belonging to the Cortinariaceae are reportedto have dikaryotic hyphae (Kühner, 1980), except for Alni-cola bohemica, which has been reported to have monokary-otic hyphae (Kühner, 1942; Moreau and Deïana, 2004).Thus monokaryons appear to be characteristic and possi-bly a synapomorphy of the whole /Bohemica clade and alsoof clade /Hymenogaster II, further conWrming that thesetwo clades are sister-groups and that an early modiWcationof sexual behavior occurred before stabilization of fruit-body shape (secotioid/agaricoid).
Unfortunately, only Smith (1966) and Pegler and Young(1987) consider the occurrence or lack of clamp connec-tions in their descriptions of Hymenogaster spp. In orderto ascertain the conservation of the monokaryotic state in
/Hymenogaster I (Hy. gardneri, Hy. parksii, Hy. subalpi-nus), more collections need to be examined for clamp con-nections and nuclear status. Whether or not the monokaryonoriginates from parthenogenesis (haploid), as assumed byKühner (1942), or from early fusion of nuclei in the dikar-yons (diploid), as observed in other fungal genera such asArmillaria (Grillo et al., 2000), should be investigated.
4.6. Evolution and radiation of agaricoid and sequestrate ( D secotioid or gastroid) groups
This study points out that sequestrate forms have animportant but often neglected role in the recent evolution-ary history of Hebelomeae, and Agaricales in general. Forbetter taxonomic and systematic treatments of the Agarica-les, characters of such groups should be more closely inves-tigated in comparison with their agaricoid sister groups. Inour study, species with gastroid fruitbodies fall into twodistinct clades in Hebelomeae and are not monophyletic.Moreover, each of the two gastroid clades /Hymenogaster Iand /Hymenogaster II share some very unusual characterswith their respective agaricoid sister-groups: spindle-shaped cystidia occur in species of /Hymenogaster I(H. subalpinus; Smith, 1966) and in /Alnicola, and mono-karyons occur in Hymenogaster II and in /Bohemica. Thesecharacters do not occur elsewhere in the family. Sharedcharacters of agaricoid and gastroid clades suggest that anepisode of extensive speciation (perhaps though adaptiveradiation) occurred at the origin of the whole tribe. Thisexplosive origin is marked by frequent transitions fromagaricoid to gastroid fruitbodies. Transitional forms (illus-trated by secotioid species in other clades or genera, e.g.,Cortinarius) are not observed amongst agaricoid subcladesof Hebelomeae and both sequestrate subclades are exclu-sively gastroid. This suggests that subclades of Hebelomeaehave arrived at more stable morphologies and ecologiesafter the initial radiation processes.
Recent phylogenetic analyses (e.g., Hibbett et al., 1997;Peintner et al., 2001) have clearly rejected Singer’s (1951)hypothesis of secotioid species as a transitional stepbetween gasteromycetes and agaricomycetes. Sequestrategroups are polyphyletic and they have close relationshipswith many diVerent groups of agaricoid fungi (Hibbettet al., 1997). Heim (1971) suggested that the “secotioid syn-drome” could be a characteristic of recently evolvedgroups. Sequestrate fungi probably result from a rapid andactive period of radiation, possibly corresponding to fastcolonization of new ecological niches (e.g., host nicheexpansion or edaphic niche diVerentiation). There is strongevidence that secotioid forms occur mostly in agaricoidgroups where an active radiation is still going on, e.g., in thespecies groups Arcangeliella borziana/Lactarius aurantiacus(Peter et al., 2001), Descolea/Setchelliogaster (Peintneret al., 2001), or Hydnangium/Laccaria (Mueller and Pine,1995). Therefore, occurrence of sequestrate forms could beused as markers for periods of adaptive radiation inAgaricales.

P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807 805
4.7. Synapomorphic characters
Our phylogenetic reconstruction shows how moleculartools can help to interpret morphological and ecologicalcharacters in Basidiomycetes and to assess their taxonomicvalue. In the monophyletic tribe Hebelomeae, most groups(genera and/or sections) described in the literature wereconWrmed as monophyletic using evidence from moleculardata, but our reconstruction results in a re-evaluation of theanatomical (cystidia, pileipellis) and ecological (host-speci-Wcity) characters originally used to delimit these groups.
Important synapomorphic characters of selected clades(Fig. 4) are the following: (i) the hymeniform pileipellis (B)is a unique, synapomorphic character for /Submelinoides;(ii) the lanceolate cystidia are a synapomorphy for /Alni-cola; (iii) the lack of clamp connections and monokaryotichyphae are synapomorphic characters for /Bohemica and/Hymenogaster II; (iv) the strict ectomycorrhizal associa-tion with Alnus spp. is an independently derived characterof both, /Alnicola sensu stricto, and /Submelinoides; and (v)the gastroid morphology is characteristic of both /Hyme-nogaster I and /Hymenogaster II.
Thus, each clade of Hebelomeae exhibits a diagnosticcombination of characters: in species belonging to /Alni-cola, the pileipellis is a tomentocutis with pseudoparenchy-matous subpellis (Fig. 4A), cheilo- and caulocystidia arelanceolate, and clamp connections are present. Moreover,species of the subclade /Alnicola sensu stricto are character-ized by strict ectomycorrhizal associations with Alnus spp.,while species of both subclades /A. amarescens and /A. cho-lea are characterized by ectomycorrhizal associations withSalicaceae. In species belonging to /Submelinoides, thepileipellis is hymeniform (Fig. 4B), cheilo- and caulocysti-dia are cylindro-claviform, clamp connections are present,and they are strict ectomycorrhizal symbionts of Alnus spp.In species belonging to /Bohemica, the pileipellis is a tom-entocutis with pseudoparenchymatous subpellis (Fig. 4E),cheilo- and caulocystidia are cylindro-claviform, clampconnections are lacking, hyphae are monokaryotic,and they are ectomycorrhizal symbionts of the Salicaceae.The /Hebeloma and /Anamika clades are characterized by awell-diVerentiated and stratiWed suprapellis consisting ofslender hyphae with pseudoparenchymatous subpellis, andthe suprapellis can be gelatinized (Hebeloma, Fig. 4D) ornot (Anamika, Fig. 4C).
4.8. Taxonomic conclusions and perspectives
If all subclades of the /Hebelomeae clade are to be con-sidered on the same taxonomic level, the polyphyleticnature of Alnicola has two possible taxonomic implications:(i) all species of Hebelomeae are assigned to the samegenus, with subgeneric distinction delimited by its subc-lades (this was already suggested by Kühner, 1980; Hyme-nogaster not being considered). (ii) All subclades arerecognized at the generic level. In the latter case, /Submeli-noides and /Bohemica need to be segregated from Alnicola
and /Hymenogaster II would also need to be segregatedfrom the genus Hymenogaster. A detailed systematic andnomenclatural discussion of Alnicola will be topic of a sepa-rate paper.
The present study elucidates the origin and evolution ofectomycorrhizal fungi in the genus Alnicola and relatedgroups. Phylogenetic analyses reject a single origin of spe-cies currently classiWed in Alnicola and clearly show theadvantages and limitations of selected anatomical and eco-logical characters for the systematic treatments of thisgenus. The rDNA sequences used in this study are suitablefor addressing questions at the genus and sectional levels,but in some groups of /Alnicola there was insuYcient phy-logenetic resolution to address species concepts andspeciation processes. A more robust phylogeny of Alnus-associated Alnicola species will require evidence fromseveral other independent loci. In conclusion, the evolution-ary history of these mutualistic fungi has resulted in com-plex and sometimes also very strict associations with plantgenera of the families Betulaceae and Salicaceae (Aanenet al., 2000b). These intriguing patterns should be exploredwith more data including sampling extra-European isolatesand by parallel comparisons with host tree phylogenies.
Acknowledgments
We thank Edith Lang (ETH, Zürich, CH), Sophie Millotand Nicolas Paoli (Université Paul Sabatier, Toulouse, FR)for their technical assistance, and Dr. Anne-Marie Fiore(Genève, CH), Dr. Dirk Redecker (Basel, CH) Dr. HannesGamper, Dr. Egon Horak, Dr. Adrian Leuchtmann, Dr.Jim Mant and Dr. Alex Widmer (ETH, Zürich, CH) fortheir helpful advices. We are also grateful to all colleagueswho provided us with specimens used for sequencing: Dr.Joe F. Ammirati (University of Washington, Seattle, USA),André Bidaud (Meyzieu, FR), Dr. Régis Courtecuisse (Uni-versité Lille 2, FR), Jean-Claude Déiana (Annemasse, FR),Yves Deneyer (Wasmes, B), Maurice Durand (La Motte-Servolex, F), Jacques Guinberteau (INRA Bordeaux, FR),Dr. Patrick B. Matheny (Clark University, Worcester,USA), Paul Pirot (Neuchâteau, B), Roberto FernándezSasia (Biskaia, SP) and Jean-Jacques Wuilbaut (Mons, B).The Wrst author thanks especially Dr. Peter Edwards (Geo-botanisches Institut, ETH Zürich, CH) for having allocatedcredits for the realization of this study in the context of hispost-doctoral period. We Wnally thank Dr. Andy Taylor(SU) and Bryn Dentinger (USA) for critically reading theWnal manuscript, for helpful suggestions, and English veriW-cation, and two anonymous reviewers for their accuratecomments and valuable suggestions on our manuscript.
References
Aanen, D.K., Kuyper, T.W., Hoekstra, R.F., 2000a. Phylogenetic relation-ships in the genus Hebeloma based on ITS 1 and 2 sequences, with spe-cial emphasis on the Hebeloma crustuliniforme complex. Mycologia 92,269–281.

806 P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807
Aanen, D.K., Kuyper, T.W., Hoekstra, R.F., 2000a. Phylogenetic relation-ships in the genus Hebeloma based on ITS 1 and 2 sequences, with spe-cial emphasis on the Hebeloma crustuliniforme complex. Mycologia 92,269–281.
Aanen, D.K., Kuyper, T.W., Mes, T.H.M., Hoekstra, R.F., 2000b. The evo-lution of reproductive isolation in the ectomycorrhizal Hebeloma crust-uliniforme aggregate (Basidiomycetes) in northwestern Europe: aphylogenetic approach. Evolution 54 (4), 1192–1206.
Bon, M., 1992. Clé monographique des espèces galéro-naucorioïdes. Doc.mycol. 21 (84), 1–89.
Bon, M., 2002. Clé de détermination du genre Hebeloma (Fr.) Kumm.(Agaricomycetidae—Cortinariales). Doc. mycol. 30 (123), 3–39.
Bruchet, G., 1970. Contribution à l’étude du genre Hebeloma (Fr.) Kum-mer; partie spéciale. Bull. mens. Soc. Linn. Lyon 39 (6 suppl.), 3–132.
Bruchet, G., 1974. Recherches sur l’écologie des Hebeloma arctico-alpins(Basidiomycètes—Agaricales). Étude spéciale de l’aptitude ectomy-corrhizogène des espèces. In: Travaux mycologiques dédiés à R. Küh-ner. Bull. mens. Soc. Linn. Lyon 43, 85–96.
Brunner, I., Horak, E., 1990. Mycoecological analysis of Alnus-associatedmacrofungi in the region of the Swiss National Park as recorded by J.Favre (1960). Mycol. Helv. 4, 111–139.
Bujakiewicz, A.M., 1989. Macrofungi in the alder and alluvial forests invarious parts of Europe and North America. Op. Bot. 100, 29–41.Chen, Z., Li, J., 2004. Pylogenetics and Biogeography of Alnus (Betula-ceae) Inferred from Sequences of Nuclear Ribosomal DNA ITSRegion. Int. J. Pl. Sci. 165 (2), 325–335.
Chen, Z., Li, J., 2004. Phylogenetics and biogeography of Alnus (Betula-ceae) inferred from sequences of nuclear ribosomal DNA ITS region.Int. J. Plant Sci. 165 (2), 325–335.
Clémençon, H., 1985. Schwärzende Lyophyllum-Arten Europas. Z. Mykol.52, 61–84.
Clémençon, H., 2004. Cytology and plectology of the Hymenomycetes.Bibliotheca mycological 199. J. Cramer, Berlin, Germany.
Courtecuisse, R., Duhem, B., 1994. Guide des champignons de France etd’Europe. Delachaux & Niestlé, Lausanne, Switzerland.
Den Bakker, H.C., Zuccarello, G.C., Kuyper, T.W., Noordeloos, M.E.,2004. Evolution and host speciWcity in the ectomycorrhizal genus Lecc-inum. New Phytol. 163 (1), 201–215.
Dorninger, A., 1993. Pilzsoziologie von Grau- und Grünerlebestanden.Univ. Innsbruck, Doktorarbeitung diss, Austria.
Favre, J., 1948. Les associations fongiques des hauts-marais jurassienset de quelques régions voisines. Beitr. KryptogamenX. Schweiz 10 (3),1–228. 1–6.
Gardes, M., Bruns, T.D., 1993. ITS primers with enhanced speciWcity forbasidiomycetes—application to the identiWcation of mycorrhizae andrusts. Mol. Ecol. 2, 113–118.
Goldman, N., Whelan, S., 2000. Statistical tests of gamma-distributed rateheterogeneity in models of sequence evolution in phylogenetics. Mol.Biol. Evol. 17, 975–978.
Grillo, R., Korhonen, K., Hantula, J., Hietala, A.M., 2000. Genetic evi-dence for somatic haploidization in developing fruit bodies of Armil-laria tabescens. Fungal Genet. Biol. 30 (2), 135–145.
Gulden, G., Dunham, S., Stockman, J., 2001. DNA studies in the Galerinamarginata complex. Mycol. Res. 105, 432–440.
Heim, R., 1971. The interrelationships between the Agaricales and gastero-mycetes. In: Petersen, R.H. (Ed.), Evolution in the higher Basidiomyce-tes. University of Tennessee, Knoxville, TN, USA, pp. 505–534.
Hibbett, D.S., Pine, E.M., Langer, E., Langer, G., Donoghue, M.J., 1997.Evolution of gilled mushrooms and puVballs inferred from ribosomalDNA sequences. Proc. Natl. Acad. Sci. Unit. St. Amer. 94, 12002–12006.
Horak, E., Moreau, P.-A., 2004 (2005). Les Naucoria nouveaux de la FloreAnalytique. Bull. trim. Soc. mycol. Fr. 120 (1–4), 215–237.
Huelsenbeck, J.P., Ronquist, F.R., 2001. MrBayes: Bayesian inference ofphylogeny. Biometrics 17, 754–755.
Kirk, P.M., Cannon, P.F., David, J.C., Stalpers, J.A., 2001. Ainsworth &Bisby’s dictionary of the fungi, ninth ed. CAB International,Egham, UK.
Kühner, R., 1926. Contribution à l’étude des Hyménomycètes et spéciale-ment des Agaricales. Botaniste 17 (1–4), 1–224.
Kühner, R., 1931. Description de quelques espèces nouvelles de Naucoriadu groupe Alnicola. Bull. trim. Soc. mycol. Fr. 47, 237–243.
Kühner, R., 1942. Observations taxinomiques et cytologiques sur quelquesNaucoria du groupe Alnicola. Ann. Univ. (Lyon), 3e sér Sci. nat., 1–15.
Kühner, R., 1980. Les Hyménomycètes agaricoïdes (Agaricales, Tricholo-matales, Plutéales, Russulales). Étude générale et classiWcation. Bull.mens Soc. Linn. Lyon numéro spécial 49, 1–927.
Kühner, R., 1981. Agaricales de la zone alpine. Genre Alnicola Kühner.Trav. sci. Parc natl Vanoise 11, 119–127.
Kühner, R., 1987. Quelques problèmes de nomenclature posés par le taxonNaucoria Fries (Agaricales). Bull. trim. Soc. mycol. Fr. 103 (3), 227–238.
Lange, J.E., 1939. Flora agaricina danica 4. Copenhagen, 1–119 (tab.)121–160.
Matheny, P.B., 2004. Improving phylogenetic inference of mushroomswith RPB1 and RPB2 nucleotide sequences (Inocybe; Agaricales). Mol.Phylogenet. Evol. 35, 1–20.
Molina, R., 1981. Ectomycorrhizal speciWcity in the genus Alnus. Can. J.Bot. 59, 325–334.
Molina, R., Trappe, J.M., 1994. Biology of the ectomycorrhizal genusRhizopogon. I. Host associations, host-speciWcity and pure culture syn-theses. New Phytol. 126, 653–675.
Moncalvo, J.-M., Vilgalys, R., Redhead, S.A., Johnson, J.E., James, T.Y.,Aime, M.C., Hofstetter, V., Verduin, S.J.W., Larsson, E., Baroni, T.J.,Thorn, R.G., Jacobsson, S., Clemencon, H., Miller, O.K., 2002. Onehundred and seventeen clades of euagarics. Mol. Phylogenet. Evol. 23,357–400.
Moreau, P.-A., 2004. Qu’est-ce que Tubaria umbrina? Bull. sem. Féd.Assoc. mycol. médit. 25 (1), 3–18.
Moreau, P.-A., 2005. A nomenclatural revision of the genus Alnicola(Cortinariaceae). Fungal Div. 21, 121–155.
Moreau, P.-A., Deïana, J.-C., 2004. Un Alnicola des saules: Alnicola saliciset sa variabilité. Bull. mycol. bot. Dauphiné-Savoie 73, 41–50.
Moreau, P.-A., Garcia, G., 2005. Alnicola salabertii, sp. nov., mycorhiziqued’ Alnus cordata, et deux autres Alnicola à petites spores. Bull. trim.Soc. mycol. Fr. 120 (1–4), 273–292.
Moser, M., 1983. Die Röhrlinge und Blätterpilze. In: Gams, H. (Ed.), KleineKryptogamenXora 2, second ed. G. Fischer, Stuttgart, pp. 1–534.
Mueller, G.M., Pine, E.M., 1995. Evidence for monophyly of Hydnangium,Laccaria, and Podohydnangium based on rDNA sequence data. Inocu-lum, Newsletter mycol. Soc. Amer. 46 (3), 30.
Navarro, E., Bousquet, J., Moiroud, A., Munive, A., Piou, D., Normand, P.,2003. Molecular phylogeny of Alnus (Betulaceae), inferred from ribo-somal DNA ITS sequences. Pl. Soil 254, 207–217.
Orton, P.D., 1960. New check-list of British Agarics and Boleti. Part III.Notes on genera and species in the list. Trans. Brit. mycol. Soc. 43 (2),159–439.
Orton, P.D., 1984. Notes on British Agarics: viii. Notes Roy. bot. Gard.Edinburgh 41 (3), 565–624.
Pegler, D.N., Young, T.W.K., 1987. A reassessment of the British species ofHymenogaster (Basidiomata: Cortinariales). Notes Roy. bot. Gard.Edinburgh 44 (3), 437–485.
Peintner, U., Moncalvo, J.M., Vilgalys, R., 2004. Toward a better under-standing of the infrageneric relationships in Cortinarius (Agaricales,Basidiomycota). Mycologia 96, 1042–1058.
Peintner, U., Moser, M., Horak, E., Vilgalys, R., 2002. Phylogeny of Roz-ites, Cuphocybe and Rapacea inferred from ITS and LSU rDNAsequences. Mycologia 94 (4), 620–629.
Peintner, U., Bougher, N.L., Castellano, M.A., Moncalvo, J.M., Moser,M.M., Trappe, J.M., Vilgalys, R., 2001. Multiple origins of sequestratefungi related to Cortinarius (Cortinariaceae). Am. J. Bot. 88, 2168–2179.
Peter, M., Buchler, U., Ayer, F., Egli, S., 2001. Ectomycorrhizas and molec-ular phylogeny of the hypogeous russuloid fungus Arcangeliella borzi-ana. Mycol. Res. 105, 1231–1238.
Redhead, S.A., 1984. Mycological observations, 4–12: on Kuehneromyces,Stropharia, Marasmius, Mycena, Geopetalum, Omphalopsis, Phaeomar-asmius, Naucoria and Prunulus. Sydowia 37, 246–270.

P.-A. Moreau et al. / Molecular Phylogenetics and Evolution 38 (2006) 794–807 807
Reid, D.A., 1984. A revision of the British species of Naucoria sensu lato.Trans. Brit. mycol. Soc. 82 (2), 197–237.
Romagnesi, H., 1942. Description de quelques espèces d’Agarics och-rosporés. Bull. trim. Soc. mycol. Fr. 58 (4), 121–169.
Salamin, N., Chase, M.W., Hodkinson, T.R., Savolainen, V., 2003. Assess-ing internal support with large phylogenetic DNA matrices. Mol.Phylogenet. Evol. 51, 112–126.
Shimodaira, H., Hasegawa, A., 1999. Multiple comparisons of log-likelihoodswith applications to phylogenetic inference. Mol. Biol. Evol. 16, 1114–1116.
Singer, R., 1939. Notes sur quelques Basidiomycètes. Ve série Rev. Mycol.Paris 4 (1–2), 64–72.
Singer, R., 1951. Thaxterogaster—a new link between Gasteromycetes andAgaricales. Mycologia 43, 215–288.
Singer, R., 1977 (1978). Keys for identiWcation of the species of AgaricalesI. Sydowia 30 (1–6), 192–279.
Singer, R., 1986. The Agaricales in modern taxonomy, fourth ed. Koeltz,Königstein, Germany.
Smith, A.H., 1966. Notes on Dendrogaster, Gymnoglossum, Protoglossumand species of Hymenogaster. Mycologia 58, 104–125.
SwoVord, D.L., 2003. PAUP*. Phylogenetic analysis using parsimony(* and other methods). Version 4. Sinauer Associates, Sunderland, MA,USA.
Söhner, E., 1962. Die Gattung Hymenogaster Vitt. Eine monographischeStudie mit besonderer Berücksichtigung der bayerischen Arten. Beih.Nova Hedwigia 2, 1–113.
Thomas, K.A., Peintner, U., Moser, M., Manimohan, P., 2002. Anamika, anew mycorrhizal genus of Cortinariaceae from India and its phyloge-netic position based on ITS and LSU sequences. Mycol. Res. 108, 245–251.
Vesterholt, J., 1989. A revision of Hebeloma sect. Indusiata in the Nordiccountries. Nord. J. Bot. 9 (3), 289–319.
Vilgalys, R., Hester, M., 1990. Rapid genetic identiWcation and mapping ofenzymatically ampliWed ribosomal DNA from several Cryptococcusspecies. J. Bacteriol. 172, 4238–4246







![[Psilocybin FR]Guide de Culture des Champignons Magiques pour champotes-The Magic Mushroom Growers Guide-[hallucinogene psilocybe mexique entheogene mycologie psychoactif psychédélique]](https://img.pdfslide.fr/doc/110x75/557213f6497959fc0b936c70/psilocybin-frguide-de-culture-des-champignons-magiques-pour-champotes-the.jpg)




![Presentation F. Chateauraynaud.ppt [Mode de compatibilité] · individuals and events sometimes make a difference. Like mushroom spores, they may germinate in unexpected places, reshaping](https://img.pdfslide.fr/doc/110x75/5ec75bfa1d81065d2965f699/presentation-f-mode-de-compatibilit-individuals-and-events-sometimes-make-a.jpg)
![Presentation F. Chateauraynaud.ppt [Mode de compatibilité]...• Anna Tsing, The Mushroom at the End of the World. On the Possibility of Life in Capitalist Ruins (2015) Anna Tsing,](https://img.pdfslide.fr/doc/110x75/5f3cc572680ccd291b406c0e/presentation-f-mode-de-compatibilit-a-anna-tsing-the-mushroom-at-the.jpg)















