Embed Size (px)
Citation preview

Chapitre 46
Culture embryonnaire prolongée et éclosion assistée
Y. Ménézo, A. Hazout, A. Veiga, E.J. Servy, P. Cohen-Bacrie, J. Chouteau, A.M. Junca et M. Dumont
Culture embryonnaire prolongée
Introduction
L a culture prolongée correspondait au départ à une nécessité : les biotechnologies chez les animaux domestiques à intérêt zootechnique
nécessitent, pour le transfert embryonnaire, un transfert au stade blastocyste. En eff et, le transfert aux stades précoces, après fécondation in vitro et culture de courte durée, n’est tout simplement pas possible : les embryons sont expulsés de l’utérus. Seul le transfert de morula tardives (5 jours) ou de blastocyste est opérant. Les problèmes de culture de l’embryon du stade 1 au blastocyste se heurtent, dans de nombreux cas, à un blocage au moment du cycle cellulaire correspondant à la transition du contrôle maternel au contrôle embryonnaire (MZT ou mater-nal to zygotic transition) : souris : 2-4 C, homme : 4-8 C, vache, lapin : 8-16 C. Ce cycle cellulaire dure en moyenne 24 heures pendant lesquelles démarrent les premières transcriptions embryonnaires ; l’em-bryon prend son autonomie par rapport aux réserves d’ARN et de protéines stockées pendant la croissance de l’ovocyte, qui se sont épuisées pendant les premiè-res divisions. Il est par ailleurs toujours très diffi cile de reproduire effi cacement les conditions in vivo pour les reproduire en culture in vitro. Les rendements dépassent très rarement 50 %. La coculture, i.e. la culture de l’embryon avec des cellules somatiques, a alors été une étape évidente.La coculture des embryons de bovins avec des vési-cules trophoblastiques, impliquant un système embryotrophique autocrine, a été la première tech-nologie permettant l’obtention de veaux vivants par culture embryonnaire d’embryons du stade 1-cellule au blastocyste puis transfert embryon-naire (1, 2). Puis, naturellement, les cellules épi-théliales du tractus génital femelle (utilisation d’un eff et paracrine) ont été utilisées (3). Les pre-miers essais chez les primates concernaient les cel-lules tubaires humaines (4) et utérines (singe) (5). Ensuite, la démonstration que la culture embryon-naire in vitro n’était pas hormonodépendante (6) a permis le passage à des lignées établies d’origine
non génitale, notamment les cellules Vero, cellules épithéliales de rein de singe, utilisées en pharmacie industrielle pour la fabrication des vaccins (7-10), largement employées en FIV humaine, et les cellu-les BRL (buff alo rat liver) utilisées chez les bovins. Si l’utilisation de la coculture perdure pour les utilisations biotechnologiques vétérinaires, cette technique a été plutôt abandonnée pour la FIV humaine, surtout en ce qui concerne l’utilisation de lignées établies.Depuis, deux types de milieux défi nis de culture sont actuellement utilisés : 1) les milieux dits « séquen-tiels » : un premier milieu est utilisé jusqu’à l’activation génomique, puis un autre est employé à partir du stade j3 ; 2) les milieux utilisés de la fécondation à l’obtention du blastocyste sans modifi cation de formule.
Diff érentes techniques
Le développement embryonnaire préimplantatoire est biphasique. Les premières divisions se réalisent à partir des réserves maternelles, stockées durant la croissance ovocytaire. Cette première phase est nécessairement plus fragile. Ainsi, toute anoma-lie du stockage impliquant la division cellulaire et/ ou la protection contre des agressions exté-rieures (ROS notamment) ou les régulations (répa-ration de l’ADN : « DNA repair ») peuvent amener soit à un arrêt de développement, soit au déclen-chement de l’apoptose. Dans un deuxième temps, entre les stades 4 et 8 cellules, les transcriptions embryonnaires démarrent, l’embryon devient plus autonome et est alors capable de gérer plus effi cacement les variations d’environnement. Le cycle de l’activation génomique est le plus long, il dure 24 heures, pour la plupart des mammifères, et c’est là que les arrêts de développement sont les plus fréquents. Cependant, cette vision est quand même un peu schématique, dans la mesure où des transcrits maternels importants perdurent aussi jusqu’au stade blastocyste où ils s’expriment.
Coculture
Elle peut être défi nie comme la culture simultanée de cellules somatiques avec l’embryon. Les premiers
C. Poncelet, et al., Physiologie, pathologie et thérapie de la reproduction chez l’humain© Springer-Verlag France, Paris 2011

522 Physiologie, pathologie et thérapie de la reproduction chez l’humain
aminés soufrés entrent très fortement dans la protection antiradicalaire. Certains de ces milieux de première phase médiocres peuvent également augmenter le risque de jumeaux monozygotiques par absence d’acides aminés soufrés et excès de glucose, générateurs de radicaux libres via l’acti-vité mitochondriale. Le risque mentionné de la formation excessive d’ammoniaque ne tient pas plus, dans la mesure où le carbonate d’ammonium formé dans les milieux de culture est très insta-ble et libère l’ammoniac qui est éliminé par les fl ux gazeux. Il faut noter à ce propos que l’ammoniac libéré dans la coculture est recyclé par les cellu-les du support cellulaire. La teneur en glucose est réduite dans ces milieux de première phase afi n de diminuer les risques d’apoptose induite par les radicaux libres.Le milieu de deuxième phase est généralement plus riche et complet.Il est à noter que la culture du stade zygote au stade blastocyste dans un seul milieu (global) est réalisable pour l’embryon humain qui a une cer-taine plasticité. Elle est quasi impossible pour les espèces domestiques à intérêt zootechnique.
Diff érences coculture/milieux simples
Il est assez communément admis que la coculture aboutit à de meilleurs résultats que les milieux dits « simplifi és », qu’ils soient séquentiels ou non (10, 22, 23). L’éclosion et la synthèse d’hCG sont notamment plus précoces. Dans notre expérience, la diff érence en faveur de la coculture a été très importante, surtout concernant la viabilité des blastocystes congelés. Il est plus que certain que les facteurs de croissance sont responsables de cette diff érence (24). Ils conjuguent eff ets trophiques et antiapoptotiques (GMCSF, IGF1). De fait, ces fac-teurs de croissance agissent dans un ensemble har-monieux : certains sont inhibiteurs d’autres activa-teurs. Ceci est spécialement vrai pour la régulation, dans le jeune blastocyste, du ratio masse cellulaire interne/trophoblaste (fi g. 1).Comme certaines cytokines (IL-6 mais surtout le LIF), ils améliorent la formation de blastocystes et l’éclosion. Le PAF semble également effi cace. À l’heure actuelle, seul le GMCSF semble devoir avoir une application commerciale. Il faut noter que la plupart de ces facteurs de croissance souvent étudiés individuellement sont présents simulta-nément dans les systèmes de coculture : ceci est spécialement vrai pour le LIF et l’IL-6. Il faut noter que GP130, récepteur d’IL-6 et du LIF et le récep-teur bêta du LIF sont présents sous forme de RNA, dans l’ovocyte et le jeune embryon. Le LIF inter-vient via la voie JAKs/STAT et freine l’apoptose (Ménézo et Tosti, communication personnelle). Ainsi, le même eff ecteur peut, de plus, utiliser
essais datent des débuts des années 1980 et ont été liés à l’essor fantastique du transfert embryonnaire bovin, quand a été possible la FIV bovine.La possibilité que la production des facteurs tro-phiques embryonnaires ne soit pas sous dépen-dance hormonale a été montrée d’abord chez la souris et les bovins par l’utilisation de cellules d’animaux prépubères (6), puis de cellules désyn-chronisées (11). Il est à remarquer que les systè-mes de cocultures avec des cellules endométria-les humaines ne plaident pas plus en faveur de l’hormono dépendance de la culture jusqu’au stade blastocyste, tout en ayant le désavantage de l’ac-cessibilité et de la quantité de matériel cellulaire disponible. Il faut également prendre en consi-dération le fait que les fi broblastes seuls sont de bien médiocres eff ecteurs du développement embryonnaire. De fait, cette effi cacité trophique semble bien liée aux épithéliums transporteurs. C’est pourquoi les cellules Vero, cellules pérenni-sées, faciles à cultiver et très contrôlées, ont eu un succès important du fait de leur facilité d’utilisa-tion. De fait, la majorité des informations quant aux régulations de la formation du blastocyste – eff et paternels et maternels (8, 12, 13), sélection cytogénétique (14) ; jusqu’à la congélation (14, 15) et la biopsie (16) des blastocystes – ont été obtenues avec les cellules Vero. Cependant, il faut également noter que des techniques de cocultures autologues ont été mises au point à partir soit de biopsies d’endomètres (17, 18), soit de cellules du cumulus (19). Cependant, le problème des cellules non pérennisées réside dans un possible vieillisse-ment qui peut altérer les capacités trophiques et dans le fait que ces cellules supportent assez mal le « repiquage ».
Milieux séquentiels
Ils sont basés sur l’aspect biphasique du développe-ment embryonnaire préimplantatoire. Le premier milieu, de la fécondation au stade 8 cellules, doit être plus « protecteur ». Si nous prenons l’exemple des radicaux libres, inducteurs incontournables de l’apoptose embryonnaire, l’embryon possède, avant l’activation génomique, une capacité limitée à les neutraliser. In vivo, il est entouré de systèmes de protection redondants dans le tractus génital femelle. Par ailleurs, dans les conditions in vitro, il y a une génération spontanée de radicaux libres dans certains milieux trop « simples » (20). Il a été soutenu que certains acides aminés (AA), dont les AA essentiels soufrés, sont toxiques lors de la pre-mière phase de culture (G séries) (21). Ce concept est incorrect et par ailleurs dangereux : la dépriva-tion des acides aminés soufrés est non seulement inductrice d’apoptose mais elle peut provoquer des anomalies d’empreinte. Par ailleurs, les acides

Culture embryonnaire prolongée et éclosion assistée 523
deux récepteurs diff érents. D’une façon globale, la coculture donne de meilleurs résultats que tout facteur de croissance ajouté individuellement. C’est bien l’objet du débat quant à l’addition d’un facteur de croissance spécifi que dans les milieux de culture. Encore une fois, c’est probablement l’équi-libre entre eff ets activateurs et eff ets inhibiteurs qui permet, comme in vivo dans le tractus génital, une croissance harmonieuse. Les cellules épithé-liales sécrètent aussi des anti-radicaux libres, pro-tecteurs de l’embryon (hypotaurine, glutathion, etc.) (25). Une des conséquences immédiatement observable est l’absence de jumeaux monozygotes en coculture, au contraire des milieux séquentiels, parfois mal conçus (26). De toute façon, la for-mation de radicaux libres est spontanée dans les milieux de culture, incubés in vitro en présence d’oxygène (20).
Pourquoi la culture prolongée ?
De fait, tout transfert embryonnaire avant l’acti-vation génomique est un geste « aveugle », même si certains critères cinétiques et morphologiques ont été respectés. Comme le stade morula com-pacte est un stade « fragile », impliquant un remo-delage cellulaire total, le transfert au stade blasto-cyste devient une évidence. La controverse quant à l’aspect de la sélection des meilleurs embryons a perduré. D’un point de vue cytogénétique, le nombre d’anomalies chromosomiques est réduit au stade blastocyste, ce qui peut permettre une
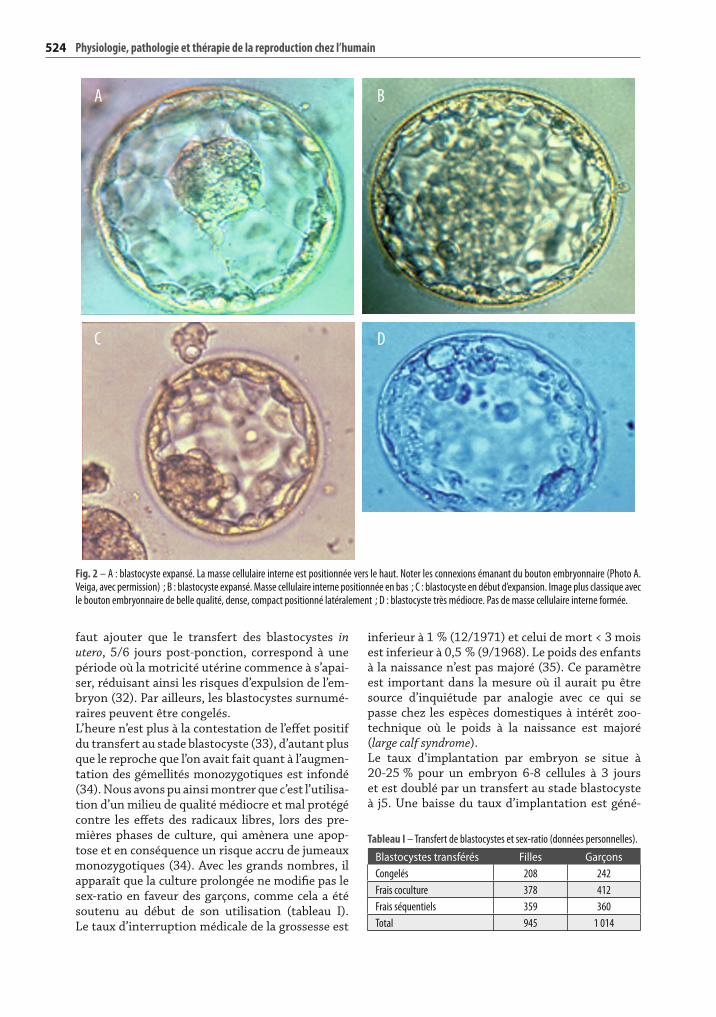
sélection pour les porteurs de translocation (27). Cette sélection n’est cependant pas parfaite, pas plus d’ailleurs qu’elle ne l’est in vivo (voir triso-mies). Par ailleurs, il peut se produire des régula-tions chromo somiques, dans le sens d’une amélio-ration, entre les stades de segmentation précoces et le blastocyste. Des embryons diagnostiqués « déséquilibrés » aux stades précoces par diagno-stic préimplantatoire se retrouvent « normaux » au stade blastocyste (28). Certaines cellules anor-males peuvent ainsi être exclues et ne pas partici-per à la compaction. Ces observations, associées à l’effi cacité toute relative du diagnostic préimplan-tatoire, des aneuploïdies et autres déséquilibres chromosomiques, ont amené à une contestation vigoureuse de cette technique (29). Certaines équipes, notamment australiennes, privilégient d’ailleurs le diagnostic génétique des embryons au stade blastocyste (30, 31).Les conséquences de cet aspect sélectif de la culture prolongée amènent à choisir le stade blastocyste toutes les fois que l’on veut privilégier le transfert d’un seul embryon : utérus malformé, volonté déli-bérée de la patiente de ne pas avoir de jumeaux, etc. En présence d’échecs répétés de transfert aux stades précoces, il est important de déterminer les capacités réelles de développement des embryons conçus. Seule la culture prolongée pourra apporter un élément de réponse. Par ailleurs, il faut ajou-ter que l’analyse de la qualité des blasto cystes sur les critères morphologiques est aisée. La qualité et la densité de la masse cellulaire interne (bou-ton embryonnaire), le début d’expansion sont des éléments favorables faciles à observer (fi g. 2). Il
Fig. 1 – Facteurs de croissance et développement in vitro de l’embryon préimplantatoire.
524 Physiologie, pathologie et thérapie de la reproduction chez l’humain
Fig. 2 – A : blastocyste expansé. La masse cellulaire interne est positionnée vers le haut. Noter les connexions émanant du bouton embryonnaire (Photo A. Veiga, avec permission) ; B : blastocyste expansé. Masse cellulaire interne positionnée en bas ; C : blastocyste en début d’expansion. Image plus classique avec le bouton embryonnaire de belle qualité, dense, compact positionné latéralement ; D : blastocyste très médiocre. Pas de masse cellulaire interne formée.
A
C
B
D
faut ajouter que le transfert des blastocystes in utero, 5/6 jours post-ponction, correspond à une période où la motricité utérine commence à s’apai-ser, réduisant ainsi les risques d’expulsion de l’em-bryon (32). Par ailleurs, les blastocystes surnumé-raires peuvent être congelés.L’heure n’est plus à la contestation de l’eff et positif du transfert au stade blastocyste (33), d’autant plus que le reproche que l’on avait fait quant à l’augmen-tation des gémellités monozygotiques est infondé (34). Nous avons pu ainsi montrer que c’est l’utilisa-tion d’un milieu de qualité médiocre et mal protégé contre les eff ets des radicaux libres, lors des pre-mières phases de culture, qui amènera une apop-tose et en conséquence un risque accru de jumeaux monozygotiques (34). Avec les grands nombres, il apparaît que la culture prolongée ne modifi e pas le sex-ratio en faveur des garçons, comme cela a été soutenu au début de son utilisation (tableau I). Le taux d’interruption médicale de la grossesse est
inferieur à 1 % (12/1971) et celui de mort < 3 mois est inferieur à 0,5 % (9/1968). Le poids des enfants à la naissance n’est pas majoré (35). Ce paramètre est important dans la mesure où il aurait pu être source d’inquiétude par analogie avec ce qui se passe chez les espèces domestiques à intérêt zoo-technique où le poids à la naissance est majoré (large calf syndrome).Le taux d’implantation par embryon se situe à 20-25 % pour un embryon 6-8 cellules à 3 jours et est doublé par un transfert au stade blastocyste à j5. Une baisse du taux d’implantation est géné-
Tableau I – Transfert de blastocystes et sex-ratio (données personnelles).
Blastocystes transférés Filles GarçonsCongelés 208 242
Frais coculture 378 412
Frais séquentiels 359 360
Total 945 1 014
Culture embryonnaire prolongée et éclosion assistée 525
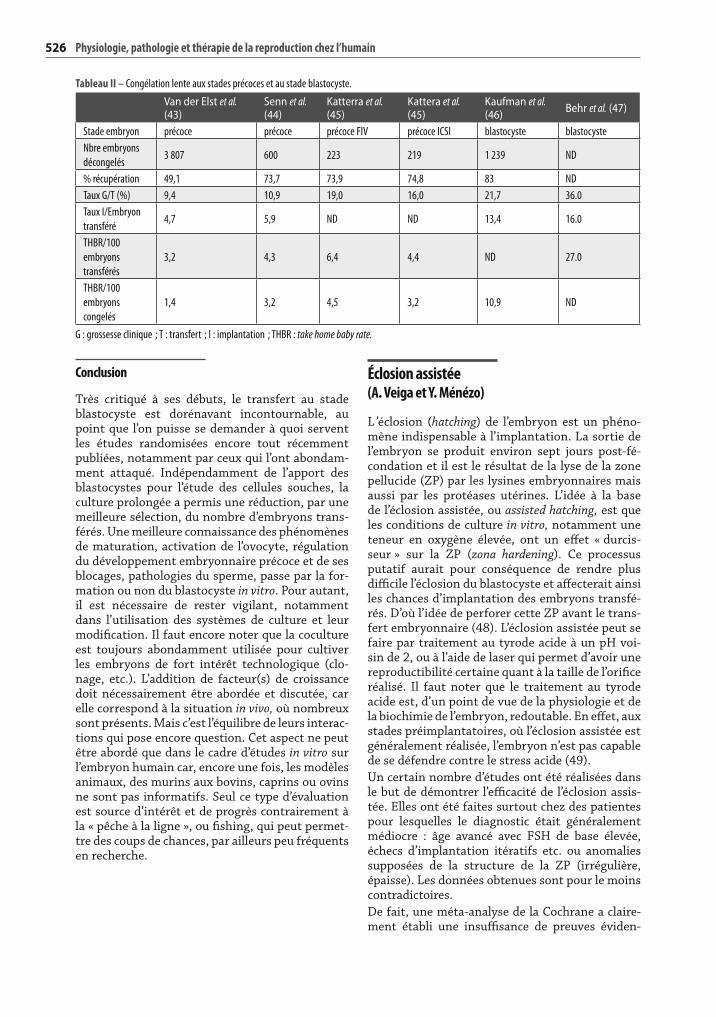
diminue le nombre d’embryons susceptibles d’être congelés : encore une fois la sélection des meilleurs n’est pas prise en compte pour les embryons pré coces ; de plus, rarement plus d’un tiers des embryons surnuméraires sont congelés sur la base des critères morphologiques. Les critères étudiés à retenir et disponibles dans la littérature quant à la congélation des blastocystes sont le pourcentage des couples ayant une congélation, le pourcentage de récupération des embryons après décongéla-tion, et le « take home baby rate » par embryon congelé et par embryon transféré (tableau II). Pour le transfert, nous avons toujours privilégié une récupération d’une nuit. Ceci permet une bien meilleure évaluation.
En ce qui concerne la vitrifi cation, il existe un point favorable indéniable : la durée de la manipulation technique 10-15 minutes pour la vitrifi cation contre près de 3 heures pour la congélation lente programmée. L’aspect « sécurité sanitaire » avec une augmentation potentielle de contamination d’une paillette ouverte à l’autre semble sur le point d’être réglé (40). Il apparaît pourtant immédiatement un élément que l’on peut qualifi er de défavorable : la très forte concentration en cryoprotecteurs toxi-ques contrairement au glycérol utilisé pour la congé-lation lente qui, lui, est un composé physiologique non toxique. Par ailleurs, il faut noter que l’éthylène glycol est un composé potentiellement toxique et mutagène (41). Pour la congélation au stade blasto-cyste, il n’y pas encore réellement d’évidence qu’une technique surpasse l’autre. Ce n’est sans doute pas à ce stade que la vitrifi cation se révélera la plus intéressante (42). Il faut simplement noter qu’en zootechnie la congélation lente demeure toujours la technique la plus employée.
ralement observée quand les blastocystes appa-raissent plus tardivement et sont transférés à j6. Quand ils sont obtenus à j7, il faut passer par la congélation car les transferts de blastocystes frais ne donnent pas de grossesse, alors que les blasto-cystes décongelés, transférés dans des utérus plus jeunes d’un point de vue hormonal, aboutissent à des naissances, confi rmant ainsi l’importance du respect de la fenêtre embryonnaire dans l’effi cacité de l’implantation chez la femme.
La congélation au stade blastocyste
D’un point de vue assez simpliste, deux théo-ries s’opposent. Doit-on éviter de prolonger la culture en suggérant que la structure embryon-naire sera altérée et rendue moins apte à la congélation ? Ainsi, faut-il congeler aux stades embryonnaires précoces, en sachant par ailleurs que la congélation entre 4 et 8 cellules à j2 ou j3 sera encore une fois un geste aveugle, ou doit-on congeler aux stades blastocystes ? D’autant plus que la sélection des meilleurs embryons à j3 est très aléatoire (36), ce que confirment les obser-vations d’Iwarson et al. (37), montrant un taux très élevé d’anomalies chromosomiques pour les embryons congelés aux stades précoces de leur développement. Là encore, la technique de culture jusqu’au stade blastocyste est intéres-sante, et c’est la protection contre les radicaux libres qui fera la différence car une peroxydation excessive des lipides des membranes les déforme et les rend beaucoup plus rigides donc plus fragi-les et « cassantes » (38).Cependant, il est extrêmement important d’ajou-ter deux paramètres :– le premier est purement cryobiologique. Une cel-
lule se congèle d’autant mieux qu’elle possède un rapport nucléocytoplasmique élevé i.e. qu’elle ne contient pas beaucoup d’eau : c’est le cas des cel-lules du blastocyste, pauvres en eau et de taille réduite, contrairement aux cellules des embryons plus précoces ;
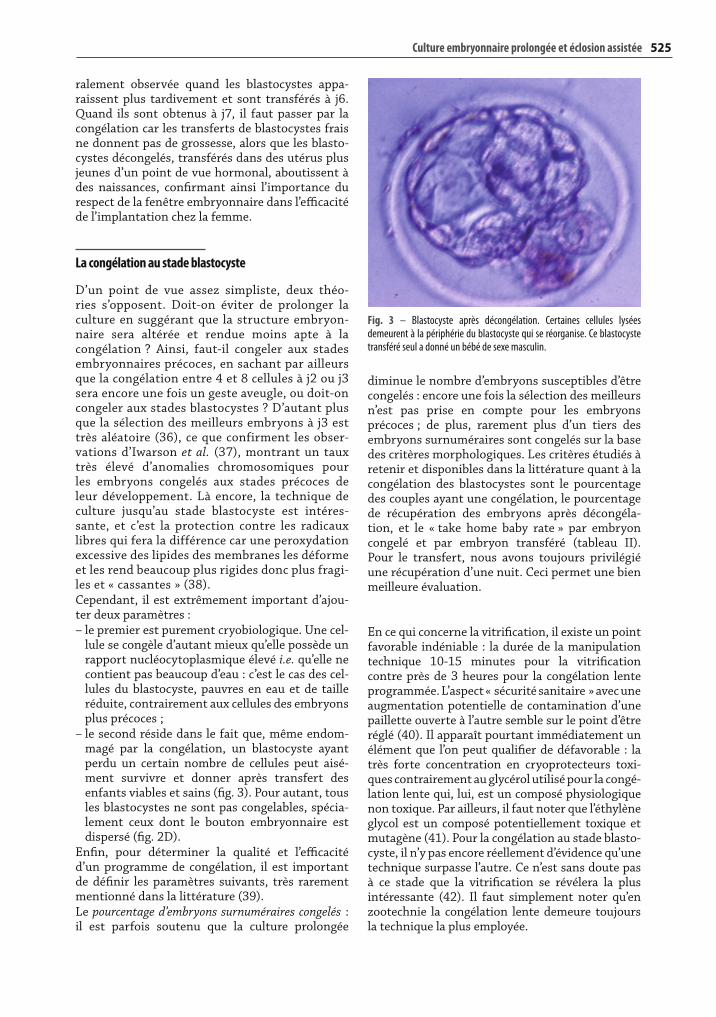
– le second réside dans le fait que, même endom-magé par la congélation, un blastocyste ayant perdu un certain nombre de cellules peut aisé-ment survivre et donner après transfert des enfants viables et sains (fi g. 3). Pour autant, tous les blastocystes ne sont pas congelables, spécia-lement ceux dont le bouton embryonnaire est dispersé (fi g. 2D).
Enfi n, pour déterminer la qualité et l’effi cacité d’un programme de congélation, il est important de défi nir les paramètres suivants, très rarement mentionné dans la littérature (39).Le pourcentage d’embryons surnuméraires congelés : il est parfois soutenu que la culture prolongée
Fig. 3 – Blastocyste après décongélation. Certaines cellules lysées demeurent à la périphérie du blastocyste qui se réorganise. Ce blastocyste transféré seul a donné un bébé de sexe masculin.
526 Physiologie, pathologie et thérapie de la reproduction chez l’humain
Éclosion assistée(A. Veiga et Y. Ménézo)
L ’éclosion (hatching) de l’embryon est un phéno-mène indispensable à l’implantation. La sortie de l’embryon se produit environ sept jours post-fé-condation et il est le résultat de la lyse de la zone pellucide (ZP) par les lysines embryonnaires mais aussi par les protéases utérines. L’idée à la base de l’éclosion assistée, ou assisted hatching, est que les conditions de culture in vitro, notamment une teneur en oxygène élevée, ont un eff et « durcis-seur » sur la ZP (zona hardening). Ce processus putatif aurait pour conséquence de rendre plus diffi cile l’éclosion du blastocyste et aff ecterait ainsi les chances d’implantation des embryons transfé-rés. D’où l’idée de perforer cette ZP avant le trans-fert embryonnaire (48). L’éclosion assistée peut se faire par traitement au tyrode acide à un pH voi-sin de 2, ou à l’aide de laser qui permet d’avoir une reproductibilité certaine quant à la taille de l’orifi ce réalisé. Il faut noter que le traitement au tyrode acide est, d’un point de vue de la physiologie et de la biochimie de l’embryon, redoutable. En eff et, aux stades préimplantatoires, où l’éclosion assistée est généralement réalisée, l’embryon n’est pas capable de se défendre contre le stress acide (49).Un certain nombre d’études ont été réalisées dans le but de démontrer l’effi cacité de l’éclosion assis-tée. Elles ont été faites surtout chez des patientes pour lesquelles le diagnostic était généralement médiocre : âge avancé avec FSH de base élevée, échecs d’implantation itératifs etc. ou anomalies supposées de la structure de la ZP (irrégulière, épaisse). Les données obtenues sont pour le moins contradictoires.De fait, une méta-analyse de la Cochrane a claire-ment établi une insuffi sance de preuves éviden-
Conclusion
Très critiqué à ses débuts, le transfert au stade blastocyste est dorénavant incontournable, au point que l’on puisse se demander à quoi servent les études randomisées encore tout récemment publiées, notamment par ceux qui l’ont abondam-ment attaqué. Indépendamment de l’apport des blastocystes pour l’étude des cellules souches, la culture prolongée a permis une réduction, par une meilleure sélection, du nombre d’embryons trans-férés. Une meilleure connaissance des phénomènes de matu ration, activation de l’ovocyte, régulation du développement embryonnaire précoce et de ses blocages, pathologies du sperme, passe par la for-mation ou non du blastocyste in vitro. Pour autant, il est nécessaire de rester vigilant, notamment dans l’utilisation des systèmes de culture et leur modifi cation. Il faut encore noter que la coculture est toujours abondamment utilisée pour cultiver les embryons de fort intérêt technologique (clo-nage, etc.). L’addition de facteur(s) de croissance doit nécessairement être abordée et discutée, car elle correspond à la situation in vivo, où nombreux sont présents. Mais c’est l’équilibre de leurs interac-tions qui pose encore question. Cet aspect ne peut être abordé que dans le cadre d’études in vitro sur l’embryon humain car, encore une fois, les modèles animaux, des murins aux bovins, caprins ou ovins ne sont pas informatifs. Seul ce type d’évaluation est source d’intérêt et de progrès contrairement à la « pêche à la ligne », ou fi shing, qui peut permet-tre des coups de chances, par ailleurs peu fréquents en recherche.
Tableau II – Congélation lente aux stades précoces et au stade blastocyste.
Van der Elst et al. (43)
Senn et al. (44)
Katterra et al. (45)
Kattera et al. (45)
Kaufman et al. (46) Behr et al. (47)
Stade embryon précoce précoce précoce FIV précoce ICSI blastocyste blastocyste
Nbre embryons
décongelés3 807 600 223 219 1 239 ND
% récupération 49,1 73,7 73,9 74,8 83 ND
Taux G/T (%) 9,4 10,9 19,0 16,0 21,7 36.0
Taux I/Embryon
transféré4,7 5,9 ND ND 13,4 16.0
THBR/100
embryons
transférés
3,2 4,3 6,4 4,4 ND 27.0
THBR/100
embryons
congelés
1,4 3,2 4,5 3,2 10,9 ND
G : grossesse clinique ; T : transfert ; I : implantation ; THBR : take home baby rate.

Culture embryonnaire prolongée et éclosion assistée 527
Menezo Y, Dale B (1995) Paternal contribution to success-13. ful embryogenesis. Hum Reprod 10: 1326-8Menezo Y, Veiga A (1997) Cryopreservation of blasto-14. cysts. In: Gomel V, Leung PCK, eds. In Vitro Fertilization and Assisted Reproduction, 10th World Congress of in Vitro Fertilization and Assisted Reproduction. Vancouver, Monduzzi: 49-53Menezo Y, Nicollet B, Herbaut N 15. et al. (1992) Freezing co-cultured human blastocysts. Fertil. Steril 58: 977-80Veiga A, Sandalinas M, Benkhalifa M 16. et al. (1997) Laser blastocyst biopsy for preimplantation diagnosis in the human. Zygote 5: 351-4Jayot S, Parneix I, Verdaguer S 17. et al. (1995) Coculture of embryos on homologous endometrial cells in patients with repeated failures of implantation. Fertil Steril 63: 109-14 Simon C, Mercader A, Garcia-Velasco J 18. et al. (1999) Cocul-ture of human embryos with autologous human endome-trial epithelial cells in patients with implantation failure. J Clin Endocrinol Metab 84: 2638-46Quinn P, Margalit R (1996) 19. Benefi cial eff ects of coculture with cumulus cells on blastocyst formation in a prospec-tive trial with supernumerary human embryos. J Assist Reprod Genet 13: 9-14Martín-Romero FJ, Miguel-Lasobras EM, Domínguez-20. Arroyo J et al. (2008) Contribution of culture media to oxidative stress and its eff ect on human oocytes. Reprod Biomed Online 17: 652-61Lane M, Hooper K, Gardner DK (2001) Eff ect of essential 21. amino acids on mouse embryo viability and ammonium production. J Assist Reprod Genet 18: 519-25Mercader A, Garcia-Velasco JA, Escudero E 22. et al. (2003) Clinical experience and perinatal outcome of blastocyst transfer after coculture of human embryos with human endometrial epithelial cells: a 5-year follow-up study. Fer-til Steril 80: 1162-8Dominguez F, Gadea B, Mercader A 23. et al. (2010) Embryo-logic outcome and secretome profi le of implanted blas-tocysts obtained after coculture in human endometrial epithelial cells versus the sequential system. Fertil Steril 93: 774-82 Richter KS (2008) 24. Th e importance of growth factors for preimplantation embryo development and in-vitro cul-ture.Curr Opin Obstet Gynecol 20: 292-304Ouhibi N, Hamidi J, Guillaud J, Menezo Y (1990) 25. Co-cul-ture of 1-cell mouse embryos on diff erent cell supports. Hum Reprod 5: 737-43Menezo Y, Sakkas D (2002) 26. Monozygotic twinning: is it related to apoptosis in the embryo? Hum Reprod 17: 247-8 Menezo Y, Chouteau J, Veiga A (2001) 27. In vitro fertilization and blastocyst transfer for carriers of chromosomal trans-location. Eur J Obstet Gynecol Reprod Biol 96: 193-5Veiga A, Gil Y, Boada M 28. et al. (1999) Confi rmation of diag-nosis in preimplantation genetic diagnosis (PGD) through blastocyst culture: preliminary experience. Prenat Diagn 19: 1242-7Fauser BC (2008) 29. Preimplantation genetic screening: the end of an aff air? Hum Reprod 23: 2622-5Boada M, Carrera M, De La Iglesia C 30. et al. (1998) Success-ful use of a laser for human embryo biopsy in preimplanta-tion genetic diagnosis: report of two cases. J Assist Reprod Genet 15: 302-7Mcarthur SJ, Leigh D, Marshall JT 31. et al. (2005) Pregnan-cies and live births after trophectoderm biopsy and pre-implantation genetic testing of human blastocysts. Fertil Steril 84: 1628-36Fanchin R, Ayoubi JM, Olivennes F 32. et al. (2000) Hormo-nal infl uence on the uterine contractility during ovarian stimulation. Hum Reprod 15 Suppl 1: 90-100
tes (50) et les conséquences à tirer sont les suivan-tes :– l’éclosion assistée n’augmente pas le taux d’im-
plantation et de grossesse en FIV, pour les pre-mières tentatives ;
– elle augmenterait les taux d’implantation chez des femmes ayant eu plusieurs échecs d’implan-tation ;
– il n’y a pas l’évidence d’un bénéfi ce pour les patien-tes d’âge avancé, pour les patientes ayant des embryons avec ZP épaisse, ou pour les embryons congelés ;
– en conséquence, il n’y a pas assez d’évidences, quant à un éventuel bénéfi ce, pour recommander l’éclosion assistée en routine de FIV/ICSI.
De fait, il est possible que l’on soit ici en présence d’un ratio risque/bénéfi ce assez confus : l’apport bénéfi que possible apporté par la perforation étant contrecarrée par l’eff et délétère lié à la technologie nécessaire pour réaliser cette perforation : pH for-tement acide ou ionisation liée au faisceau laser.
Références
Camous S, Heyman Y, Meziou W, Menezo Y (1984) Cleav-1. age beyond the block stage and survival after transfer of early bovine embryos cultured with trophoblastic vesicles. J Reprod Fert 72: 479-85Heyman Y, Menezo Y, Chesne P 2. et al. (1987) In vitro cleav-age of bovine and ovine early embryos: improved develop-ment using coculture with trophoblastic vesicles. Th eriog-enology 27: 59-68Gandolfi F, Moor RM (1987) 3. Stimulation of early embry-onic development in the sheep by co-culture with oviduct epithelial cells. J Reprod Fertil 81: 23-8Bongso A, Soon-Chye N, Sathananthan H 4. et al. (1989) Improved quality of human embryos when co-cultured with human ampullary cells. Hum Reprod 4: 706-13Goodeaux LL, Voelkel SA, Anzalone CA 5. et al. (1989) Th e eff ect of rhesus epithelial cell monolayers on in vitro growth of rhesus embryos. Th eriogenology 39: 197Menezo Y, Hamidi J, Khatchadourian C 6. et al. (1989) The murine prepuberal oviduct supports early embryo development in vitro. Developp Growth Differ 31: 551-5 Menezo Y, Guerin JF, Czyba JC (1990) Improvement of 7. human early embryo development in vitro by coculture on monolayers of Vero cells. Biol Reprod 42: 301-6 Menezo Y, Hazout A, Dumont M et al. (1992) 8. Coculture of embryos on Vero cells and transfer of blastocysts in humans. Hum Reprod 7 (Suppl 1): 101-6 Menezo Y, Janny L (1996) Is there a rationale for tubal 9. transfer in human ART? Hum Reprod 11: 1818-20Turner K, Lenton EA (1996) Th e infl uence of Vero cell 10. culture on human embryo development and chorionic gonadotrophin production in vitro. Hum Reprod 11: 1966-74Th ibodeaux JK, Menezo Y, Roussel DJ 11. et al. (1992) Co-culture of in vitro fertilized bovine embryos with oviduc-tal epithelial cells originating from diff erent stages of the estrous cycle J. Dairy Sci 75: 1448-55Janny L, Menezo Y (1996) Maternal age eff ect on early 12. human embryonic development and blastocyst formation. Mol Reprod Develop 45: 31-7

528 Physiologie, pathologie et thérapie de la reproduction chez l’humain
Blake DA, Farquhar CM, Johnson N 33. et al. (2007) Cleav-age stage versus blastocyst stage embryo transfer in assisted conception. Cochrane Database Syst Rev 17: CD002118Cassuto G, Chavrier M, Menezo Y (2003) 34. Culture condi-tions and not prolonged culture time are responsible for monozygotic twinning in human in vitro fertilization. Fer-til Steril 80: 462-3Menezo Y, Veiga A, Pouly JL (2000) 35. Assisted reproductive technology (ART) in humans: facts and uncertainties. Th e-riogenology 53: 599-610Milki AA, Hinckley MD, Gebhardt J 36. et al. (2002) Accuracy of day 3 criteria for selecting the best embryos. Fertil Steril 77: 1191-5Iwarsson E, Lundqvist M, Inzunza J 37. et al. (1999) A high degree of aneuploidy in frozen-thawed human preimplan-tation embryos. Hum Genet 104: 376-82Pryor WA, Stanley JP, Blair E (1976) 38. Autoxidation of poly-unsaturated fatty acids: II. A suggested mechanism for the formation of TBA-reactive materials from prostaglandin-like endoperoxides. Lipids 11: 370-9Menezo Y (2004) Blastocyst freezing. Eur J Obstet Gyne-39. col Reprod Biol 115 (Suppl 1): S12-5Vanderzwalmen P, Ectors F Grobet L 40. et al. (2009) Asep-tic vitrifi cation of blastocysts from infertile patients, egg donors and after IVM. Reprod Biomed Online 19: 700-7Klug S, Merker Hj, Jäckh R (2001) 41. Eff ects of ethylene gly-col and metabolites on in vitro development of rat embryos during organogenesis. Toxicol In Vitro 15: 635-42
Youssry M, Ozmen B, Zohni K 42. et al. (2008) Current aspects of blastocyst cryopreservation. Reprod Biomed Online 16: 311-20Van der Elst J, 43. Van den Abbeel E, Vitrier S et al. (1997) Selective transfer of cryopreserved human embryos with further cleavage after thawing increases delivery and implantation rates. Hum Reprod 12: 1513-21Senn 44. A, Vozzi C, Chanson A, De Grandi P, Germond M (2000) Prospective randomized study of two cryopreser-vation policies avoiding embryo selection: the pronucleate stage leads to a higher cumulative delivery rate than the early cleavage stage. Fertil Steril 74: 946-52Kattera 45. S, Shrivastav P, Craft I (1999) Comparison of pregnancy outcome of pronuclear- and multicellular-stage frozen-thawed embryo transfers. J Assist Reprod Genet 16: 358-62Kaufman RA, Menezo Y, Hazout A 46. et al. (1995) Cocultured blastocyst cryopreservation: experience of more than 500 transfer cycles. Fertil Steril 64: 1125-9Behr 47. B, Gebhardt J, Lyon J, Milki AA (2002) Factors relat-ing to a successful cryopreserved blastocyst transfer pro-gram. Fertil Steril 77: 697-9Cohen J (1991) 48. Assisted hatching of human embryos. J In-Vitro Fert Embryo Transf 8: 179-90Dale B, Menezo Y, Cohen J 49. et al. (1998) Intracellular pH regulation in the human oocyte. Hum Reprod 13: 964-70Seif MM, Edi-Osagie EC, Farquhar C 50. et al. (2005) Assisted hatching on assisted conception (IVF & ICSI). Cochrane Database Syst Rev 1: CD001894