Embed Size (px)
Citation preview

THÈSE
Pour l'obtention du grade deDOCTEUR DE L'UNIVERSITÉ DE POITIERS
UFR des sciences fondamentales et appliquéesLaboratoire Signalisation et transports ioniques membranaires - STIM (Poitiers)
(Diplôme National - Arrêté du 25 mai 2016)
École doctorale : Biologie-santé - Bio-santé (Limoges)Secteur de recherche : Aspects moléculaire et cellulaire de la biologie
Présentée par :Oualid Ayad
Caractérisation fonctionnelle des cellules souchescardiaques humaines dans un but thérapeutique
Directeur(s) de Thèse :Patrick Bois, Aurélien Chatelier
Soutenue le 12 décembre 2017 devant le jury
Jury :
Président Valérie Coronas Professeur des Universités, Université de Poitiers
Rapporteur Emmanuel Deval Chargé de recherche CNRS, Université de Nice-Sophia Antipolis
Rapporteur Christophe Vandier Professeur des Universités, Université de Tours
Membre Patrick Bois Professeur des Universités, Université de Poitiers
Membre Aurélien Chatelier Maître de conférences, Université de Poitiers
Membre Anne Aries Chercheur, Institut de recherche en hématologie et transplantion,Mulhouse
Pour citer cette thèse :Oualid Ayad. Caractérisation fonctionnelle des cellules souches cardiaques humaines dans un but thérapeutique[En ligne]. Thèse Aspects moléculaire et cellulaire de la biologie. Poitiers : Université de Poitiers, 2017. Disponiblesur Internet <http://theses.univ-poitiers.fr>

THESE
Pou l’o te tio du G ade de
DOCTEUR DE L’UNIVER“ITE DE POITIER“
(Faculté des Sciences Fondamentales et Appliquées) (Diplôme National - Arrêté du 25 mai 2016)
Ecole Doctorale : Biologie-santé
Secteur de Recherche : Aspects moléculaire et cellulaire de la biologie
Présentée par :
Oualid AYAD
************************
Caractérisation fonctionnelle des cellules souches cardiaques humaines dans un but thérapeutique
************************
Directeurs de Thèse : Pr. Patrick Bois & Dr. Aurélien Chatelier
************************
Soutenue le 12 décembre 2017
De a t la Co issio d’E a e
************************
JURY C. VANDIER Professeur, Université de Tours Rapporteur E. DEVAL CR CNRS, IPMC, Nice Rapporteur A. ARIES Chercheur, IRHT, Mulhouse Examinateur V. CORONAS Professeur, Université de Poitiers Examinateur A. CHATELIER MCU, Université de Poitiers Examinateur P. BOIS Professeur, Université de Poitiers Examinateur

Sommaire
1
SOMMAIRE
TABLE DES ILLUSTRATIONS ...................................................................................................................... 8
ABREVIATIONS ....................................................................................................................................... 12
INTRODUCTION ..................................................................................................................................... 16
Chapitre I : Cellules souches et régénération tissulaire ........................................................................ 16
1 Historique ...................................................................................................................................... 16
2 Propriétés des cellules souches ..................................................................................................... 17
2.1 L’auto-renouvellement .......................................................................................................... 17
2.2 Pouvoir de différenciation en plusieurs lignages cellulaires ................................................. 18
3 Types et potentiels de différenciation des cellules souches humaines : ...................................... 19
3.1 Les cellules souches embryonnaires (CSEs) : ......................................................................... 20
3.1.1 Les cellules souches totipotentes : ................................................................................ 20
3.1.2 Les cellules souches pluripotentes : .............................................................................. 21
3.2 Les cellules souches adultes: ................................................................................................. 21
3.2.1 Les cellules souches hématopoïétiques (CSHs) ............................................................. 22
3.2.2 Les cellules souches mésenchymateuses (CSMs) .......................................................... 24
3.2.3 Les cellules souches du tissu adipeux ............................................................................ 27
3.2.4 Les cellules progénitrices endothéliales ........................................................................ 27
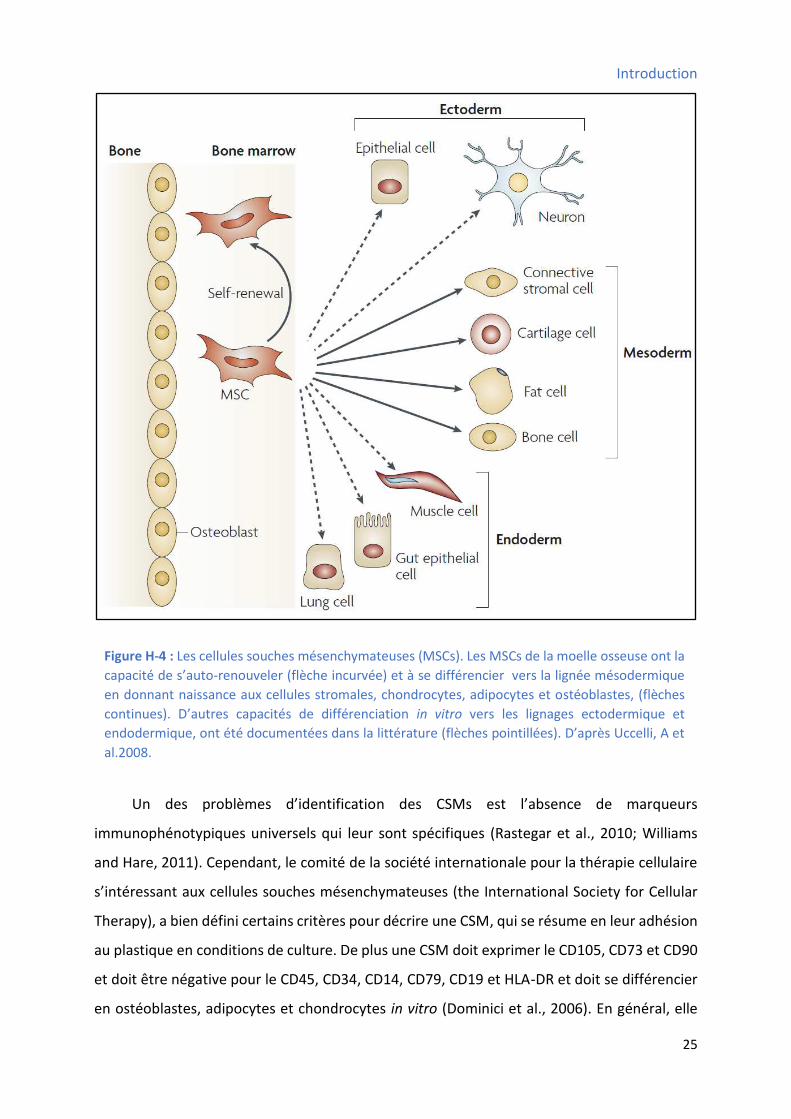
3.2.5 Les ellules sou hes de l’ pith liu i testi al : ............................................................ 29
3.2.6 Les ellules sou hes de l’ pide e ............................................................................... 31
3.2.7 Les ellules sou hes de l’ pith liu pul o ai e .......................................................... 32
3.2.8 Les cellules souches de la cornée et de la rétine .......................................................... 33
3.2.9 Les cellules souches/progénitrices hépatiques ............................................................. 35
3.2.10 Les cellules souches neurales (CSNs) du système nerveux central ............................... 36
3.2.11 Les cellules souches musculaires squelettiques ............................................................ 39
3.2.12 Les cellules souches cardiaques .................................................................................... 41
Chapitre II : Cellules souches et progéniteurs cardiaques .................................................................... 43
1 Nature des cellules souches cardiaques ........................................................................................ 43
2 Types des cellules souches cardiaques.......................................................................................... 45
2.1 Les CSCs c-kit positives (CSC c-kit+) ........................................................................................ 45
2.2 Les CSCs Sca1 positives (CSC Sca1+) ....................................................................................... 48
2.3 Les CSCs Isl-1 positives (CSCs Isl-1+)....................................................................................... 50
2.4 Les CSCs BCRP+ positives side-population (BCRP+ SP) ........................................................... 51

Sommaire
2
2.5 Les cellules dérivées des cardiosphères (CDCs) .................................................................... 53
2.6 Les CSCs W8B2 positives (CSC W8B2+) .................................................................................. 54
3 Modulateurs de la différenciation in vitro des CSCs adultes en cardiomyocytes ......................... 56
3.1 La 5-azacytidine ..................................................................................................................... 57
3.2 Le TGF-b: ................................................................................................................................ 58
3.3 L’a ide as o i ue : ............................................................................................................... 59
3.4 L’a ide ti oï ue : ................................................................................................................ 60
3.5 La dexaméthasone :............................................................................................................... 61
3.6 L’o to i e: ........................................................................................................................... 61
Chapitre III : Physiologie des cellules souches /progéniteurs cardiaques ............................................. 62
1 Le calcium : .................................................................................................................................... 62
1.1 Le calcium dans les cellules souches: .................................................................................... 62
1.1.1 L’a ti it al i ue spo ta e da s les ellules sou hes: .............................................. 63
1.1.2 Canaux VGCC et signalisation calciques ........................................................................ 64
1.1.3 Les stocks calciques intracellulaires dans les cellules souches: .................................... 67
1.2 Le calcium dans les cellules souches/progéniteurs cardiaques: ........................................... 67
2 Les canaux ioniques : ..................................................................................................................... 70
2.1 Les canaux ioniques dans les CSEs: ....................................................................................... 70
2.2 Les canaux ioniques dans les CSMs: ...................................................................................... 71
2.3 Les canaux ioniques dans les cellules souches/progéniteurs neurales: ................................ 72
2.4 Les canaux ioniques dans les CSCs: ....................................................................................... 73
3 Rôles des canaux ioniques dans la régulation de la prolifération et/ou la différenciation des
cellules souches: .................................................................................................................................... 73
3.1 Prolifération ........................................................................................................................... 73
3.2 Différenciation : ..................................................................................................................... 75
POSITION DU PROBLEME ...................................................................................................................... 76
MATERIELS ET METHODES .................................................................................................................... 78
I- Isolement et purification des CSCs W8B2+ à pa ti d’ ha tillo s hu ai s d’o eillettes d oites 78
1 Isolement des CSCs W8B2+ avec la méthode des explants ........................................................... 78
2 Purification des CSCs W8B2+ ......................................................................................................... 79
2.1 Enrichissement en CSCs W8B2+ par le système du tri cellulaire magnétique positif: ........... 79
2.1.1 Principe : ........................................................................................................................ 79
2.1.2 Protocole expérimental : ............................................................................................... 81
2.2 Purification des CSCs W8B2+ par le système de tri cellulaire par cytométrie en flux: .......... 82
2.2.1 Principe: ......................................................................................................................... 82
2.2.2 Protocole expérimental: ................................................................................................ 87

Sommaire
3
II- Immunophénotypage des CSCs W8B2+ ..................................................................................... 87
III- Conditions de culture et entretien des CSCs W8B2+ ................................................................. 88
IV- Congélation/décongélation des CSCs W8B2+ ............................................................................ 89
V- Test de formation de colonies (CFU-Fs). ....................................................................................... 89
1 Principe : ........................................................................................................................................ 89
2 Protocole expérimental: ................................................................................................................ 89
VI- Test de prolifération par comptage cellulaire ........................................................................... 90
1 Principe : ........................................................................................................................................ 90
2 Protocole expérimental: ................................................................................................................ 90
VII- Test de blessure (wound healing assay) .................................................................................... 90
1 Principe : ........................................................................................................................................ 90
2 Protocole expérimental : ............................................................................................................... 91
VIII- Test de viabilité cellulaire .......................................................................................................... 91
1 Principe : ........................................................................................................................................ 91
2 Protocole expérimental: ................................................................................................................ 92
IX- A al se du le ellulai e a e l’iodu e de p opidiu ............................................................ 92
1 Principe : ........................................................................................................................................ 92
2 Protocole expérimental : ............................................................................................................... 92
X- Immunocytochimie indirecte .................................................................................................... 93
1 Principe : ........................................................................................................................................ 93
2 Protocole expérimental : ............................................................................................................... 93
XI- Différenciation cardiaque in vitro des CSCs W8B2+................................................................... 94
1 Principe : ........................................................................................................................................ 94
2 Protocole expérimental : ............................................................................................................... 94
XII- Imagerie calcique ...................................................................................................................... 95
1 Principe : ........................................................................................................................................ 95
2 Protocol expérimental : ................................................................................................................. 96
XIII- RT-PCR « Reverse Transcription-Polymerase Chain Reaction » ................................................ 98
1 Principe : ........................................................................................................................................ 98
2 Protocole expérimental : ............................................................................................................... 98
2.1 Extraction des ARNs totaux ................................................................................................... 99
2.1.1 Extraction des ARNs totaux à partir des CSCs W8B2+ adhérentes en culture : ............. 99
2.1.2 Extraction des ARNs totaux à partir des EODHs : .......................................................... 99
2.2 Rétro-transcription ou RT (Reverse Transcription): ............................................................ 100
2.3 Amplification par PCR (Polymerase Chain Reaction) .......................................................... 100
XIV- RT-qPCR « Reverse Transcription- quantitative Polymerase Chain Reaction » en SybrGreen 101

Sommaire
4
1 Principe : ...................................................................................................................................... 101
2 Protocole expérimental : ............................................................................................................. 101
2.1 qPCR avec la technologie SybrGreen .................................................................................. 102
2.2 RT-qPCR à haut rendement ou TLDA (TaqMan Low Density Custom Array) ...................... 102
2.2.1 Principe: ....................................................................................................................... 102
2.2.2 Extraction des ARNs totaux ......................................................................................... 102
2.2.3 Transcription inverse ou RT (Reverse Transcription): ................................................. 103
2.2.4 qPCR avec la technologie TaqMan .............................................................................. 103
XV- Western blot ............................................................................................................................ 106
1 Principe: ....................................................................................................................................... 106
2 Protocole expérimental: .............................................................................................................. 107
XVI- Patch clamp ............................................................................................................................. 108
1 Principe : ...................................................................................................................................... 108
2 Protocole expérimental : ............................................................................................................. 108
XVII- Tests statistiques ..................................................................................................................... 109
RESULTATS ........................................................................................................................................... 110
Chapitre I : Isolement et caractérisation des CSCs W8B2+ .................................................................. 110
I- Isolement et caractérisation phénotypique des CSCs W8B2+ ..................................................... 111
1 Isolement des CSCs W8B2+ .......................................................................................................... 111
2 Capa it s d’auto-renouvellement et de prolifération des CSCs W8B2+ ..................................... 111
3 Immunophénotype des CSCs W8B2+ ........................................................................................... 114
II-Caractérisation génique des CSCs W8B2+ ........................................................................................ 115
1 P ofil d’e p essio des a ueu s a dia ues ........................................................................... 115
1.1 P ofil d’e p essio g i ue des facteurs de transcription cardiaques ............................... 115
1.2 P ofil d’e p essio g i ue des st u tu es o t a tiles a dia ues ................................... 117
1.3 P ofil d’e p essio g i ue des o e i es a dia ues. .................................................... 118
1.4 P ofil d’e p essio g i ue des peptides at iu ti ues.................................................... 119
2 P ofil d’e p essio g i ue des a au io i ues da s les C“Cs W B + ..................................... 119
2.1 P ofil d’e p essio g i ue des a au sodi ues da s les CSCs W8B2+ ............................ 120
2.2 P ofil d’e p essio g i ue des a au potassi ues da s les C“Cs W B + ........................ 121
2.3 P ofil d’e p essio g i ue des a au HCN da s les C“Cs W B + .................................... 127
2.4 P ofil d’e p essio g i ue des a au al i ues da s les C“Cs W B + ............................ 128
2.5 P ofil d’e p essio g i ue des t a spo teu s io i ues da s les C“Cs W B + .................. 129
3 P ofil d’e p essio g i ue des a teu s de l’ho ostasie al i ue da s les C“Cs W B + ......... 130
III-Caractérisation fonctionnelle des CSCs W8B2+ ............................................................................... 131
1 Différenciation cardiaque in vitro des CSCs W8B2+..................................................................... 131

Sommaire
5
1.1 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des marqueurs cardiaques ..................................................................................................................... 132
1.1.1 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des facteurs de transcription cardiaques .................................................................................... 132
1.1.2 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des structures contractiles cardiaques ........................................................................................ 134
1.1.3 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des connexines cardiaques .......................................................................................................... 136
1.2 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des canaux ioniques ............................................................................................................................... 137
1.2.1 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des canaux sodiques .................................................................................................................... 137
1.2.2 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des canaux potassiques ............................................................................................................... 138
1.2.3 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des canaux HCN ........................................................................................................................... 142
1.2.4 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des canaux calciques ................................................................................................................... 143
1.2.5 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio g i ue des t a spo teu s io i ues et/ou des a teu s de l’ho ostasie al i ue ................................. 145
2 Etude de l’a ti it al i ue au ou s de la diff e iatio a dia ue in vitro des CSCs W8B2+ . 151
2.1 “ig atu e de l’a ti it spo ta e al i ue au ou s de la diff e iatio a dia ue in vitro
des CSCs W8B2+ ............................................................................................................................... 151
2.2 Evolution des paramètres calciques au cours de la différenciation cardiaque in vitro des
CSCs W8B2+ ..................................................................................................................................... 153
2.2.1 Evolution de la fréquence des oscillations calciques au cours de la différenciation
cardiaque in vitro des CSCs W8B2+ .............................................................................................. 153
2.2.2 E olutio de l’a plitude des os illatio s al i ues au ou s de la diff e iatio cardiaque in vitro des CSCs W8B2+ .............................................................................................. 155
2.2.3 Evolution de la durée des oscillations calciques au cours de la différenciation
cardiaque in vitro des CSCs W8B2+ .............................................................................................. 155
2.2.4 E olutio de l’ai e des os illatio s al i ues au ou s de la diff e iatio a dia ue in
vitro des CSCs W8B2+ .................................................................................................................. 155
2.2.5 Evolution du TTP (time to peak) des oscillations calciques au cours de la différenciation
cardiaque in vitro des CSCs W8B2+ .............................................................................................. 157
2.2.6 Evolution du TTR (time to recovery) des oscillations calciques au cours de la
différenciation cardiaque in vitro des CSCs W8B2+ ..................................................................... 157
2.2.7 Evolution des pentes 1 et 2 des oscillations calciques au cours de la différenciation
cardiaque in vitro des CSCs W8B2+ .............................................................................................. 158
2.2.8 E olutio du pou e tage du te ps d’a ti it al i ue au ou s de la diff e iatio cardiaque in vitro des CSCs W8B2+ .............................................................................................. 160

Sommaire
6
2.3 Identification des acteurs impliqués dans la génération des oscillations calciques dans les
CSCs W8B2+ différenciées ............................................................................................................... 161
2.3.1 Effet de l’i hi itio de l’ ha geu NCX su les os illatio s al i ues da s les C“Cs W8B2+ différenciées pendant 28 jours........................................................................................ 161
2.3.2 Effet de l’i hi itio de la “ERCA su les os illatio s al i ues da s les C“Cs W B +
différenciées pendant 28 jours ................................................................................................... 162
2.3.3 Effet de l’i hi itio des a au CaV de type L sur les oscillations calciques dans les CSCs
W8B2+ différenciées pendant 28 jours........................................................................................ 163
2.3.4 Effet de l’i hi itio des epteu s à l’IP su les os illatio s al i ues da s les C“Cs W8B2+ différenciées pendant 28 jours........................................................................................ 164
2.3.5 Effet de l’i hi itio des epteu s à la a odi e su les os illatio s al i ues da s les CSCs W8B2+ différenciées pendant 28 jours ............................................................................... 170
Chapitre II : Rôle du canal BKCa da s la gulatio de la p olif atio et de l’auto-renouvellement des
CSCs W8B2+ ......................................................................................................................................... 172
1 Cad e de l’ tude .......................................................................................................................... 172
2 Résumé des résultats .................................................................................................................. 173
3 Article scientifique: « FUNCTIONAL BKCa CHANNEL IN HUMAN RESIDENT CARDIAC STEM CELLS
EXPRESSING W8B2». ........................................................................................................................... 174
4 BKCa et oscillations calciques dans les CSCs W8B2+ différenciées ............................................... 192
Chapitre III : Effets de la sphingosine 1-phosphate sur les propriétés des CSCs W8B2+ ..................... 196
1 Contexte bibliographique ............................................................................................................ 196
1.1 S1P et signalisation .............................................................................................................. 196
1.2 S1P et tissu cardiaque ......................................................................................................... 197
1.3 S1P et cellules souches ........................................................................................................ 198
2 Co te te de l’ tude ..................................................................................................................... 198
3 Résultats ...................................................................................................................................... 199
3.1 P ofil d’e p essio des i epteu s à la “ P da s les C“Cs W B + ............................... 199
3.2 Effet de la S1P sur les capacités prolifératives des CSCs W8B2+ ......................................... 201
3.3 I pli atio des “ PR da s l’effet a tip olif atif de la “ P o se da s les C“Cs W B + 204
3.4 Effet de la “ P su les p op i t s d’auto-renouvellement des CSCs W8B2+ ....................... 205
3.5 Effet de la S1P sur les capacités migratoires des CSCs W8B2+ ............................................ 206
DISCUSSION ......................................................................................................................................... 207
I-Isolement et caractérisation des CSCs W8B2+ .................................................................................. 207
II-P ofil d’e p essio g i ue des anaux ioniques dans les CSCs W8B2+ .......................................... 208
III-Rôle des canaux ioniques dans les CSCs W8B2+ .............................................................................. 209
IV-P ofil d’e p essio g i ue des a teu s al i ues da s les C“Cs W B + ....................................... 211
V-Différenciation cardiaque in vitro des CSCs W8B2+ ......................................................................... 213
1 Profil des marqueurs cardiaques ................................................................................................. 213

Sommaire
7
2 Profil des transcrits codant pour les canaux ioniques ................................................................. 215
3 Oscillations calciques au cours de la différenciation cardiaque in vitro des CSCs W8B2+ .......... 219
4 La sphingosine 1-phosphate (S1P) joue un rôle important dans les CSCs W8B2+ ...................... 223
CONCLUSION GENERALE ET PERSPECTIVES ........................................................................................ 227
REFERENCES BIBLIOGRAPHIQUES ....................................................................................................... 230
RESUME ............................................................................................................................................... 266
ANNEXES .............................................................................................................................................. 268

Table des illustrations
8
TABLE DES ILLUSTRATIONS
Liste des figures
Figure H-1 : Hiérarchie de la différenciation des cellules souches. ................................................... 19
Figure H-2 : La hiérarchie des cellules souches. .................................................................................... 20
Figure H-3 : Les cellules souches hématopoïétiques ...................................................................... 23 Figure H-4 : Les cellules souches mésenchymateuses .................................................................... 25
Figure H-5 : Les cellules progénitrices endothéliales ...................................................................... 28 Figure H-6 : Les cellules souches intestinales ................................................................................ 30 Figure H-7 : Les cellules souches de l'épiderme interfolliculaire ..................................................... 32 Figure H-8 : Les ellules sou hes de l’ pith liu pul o ai e .......................................................... 33 Figure H-9 : Les cellules souches du limbe scléro-cornéen .............................................................. 34 Figure H-10 : Les cellules progénitrices hépatiques ....................................................................... 36 Figure H-11 : Les cellules souches neuronales ............................................................................... 38 Figure H-12 : Les cellules satellites du muscle strié squelettique .................................................... 40 Figure H-13 : Concept des cellules souches cardiaques ................................................................... 42 Figure H-14 : Hiérarchie de la croissance et de la différenciation des cellules souches cardiaques..... 44 Figure H-15 : Illustration schématique des cellules souches cardiaques exprimant c-kit et de leur
pouvoir de différenciation ............................................................................................................ 46 Figure H-16 : Illustration schématique des cellules souches cardiaques exprimant Sca-1 et de leur
pouvoir de différenciation ............................................................................................................ 50 Figure H-17 : Illustration schématique des cellules souches cardiaques exprimant BCRP et de leur
pouvoir de différenciation ............................................................................................................ 53 Figure H-18 : Illust atio s h ati ue de l’e p essio fo tio elle des a au et epteu s calciques au cours de la différenciation des CSEs et des CSMs (du tissu adipeux et de la moelle
osseuse) ..................................................................................................................................... 66 Figure H-19 : Voies de signalisations majeures impliquées dans le maintien de la pluripotence ou de
la favorisation de la différenciation des cellules souches cardiaques .............................................. 69
Figure H-20 : Structure chimique de la sphingosine 1-phosphate ………………………………………………….196
Figure MM-1 : “ h a du p oto ole d’isole e t et de pu ifi ation des CSCs W8B2+ à partir des
échantillons auriculaires humains ................................................................................................. 78 Figure MM-2 : Schéma du principe de la technique du tri cellulaire magnétique positif ................... 80 Figure MM-3 : Schéma simplifié du principe de la technique du tri cellulaire par FACS ..................... 83 Figure MM-4 : “ h a si plifi du p i ipe du fo tio e e t d’u to t e de flu .................. 84 Figure MM-5 : Schéma simplifié des paramètres clés mesurés par le cytomètre de flux ................... 85 Figure MM-6 : Schéma simplifié du protocole de différenciation cardiaque in vitro utilisé pour
différencier les CSCs W8B2+ .......................................................................................................... 95 Figure MM-7 : “ h a du M a is e d’a tio de la GCaMP ......................................................... 95 Figure MM-8 : E e ple d’i age de fluo es e e o te ue lo s des os illatio s al i ues ap s traitement avec le logiciel Image J................................................................................................. 97 Figure MM-9 : T a d’u e os illation calcique obtenue au 28ème jour de différenciation montrant les
différents paramètres calculés avec le logiciel IDL .......................................................................... 98 Figure R-1 : Proportions des CSCs W8B2+ durant les différentes étapes de purification .................. 112 Figure R-2 : Morphologie des CSCs W8B2+ isolées et des colonies formées ................................... 113 Figure R-3 : Capa it d’auto-renouvellement et de prolifération des CSCs W8B2+ ......................... 113

Table des illustrations
9
Figure R-4 : Pourcentage des CSCs W8B2+ positives pour les marqueurs de surface indiqués
déterminé par cytométrie en flux................................................................................................ 114 Figure R-5 : P ofil d’e p essio des fa teu s de t a s iptio a dia ues da s les C“Cs W B + ....... 116 Figure R-6 : P ofil d’e p essio g i ue des st u tu es o t a tiles a dia ues da s les C“Cs W B +
................................................................................................................................................. 117 Figure R-7 : P ofil d’e p essio g i ue des o e i es a diaques dans les CSCs W8B2+ .............. 118 Figure R-8 : P ofil d’e p essio g i ue des peptides at iu ti ues de t pe A et de t pe B da s les CSCs W8B2+ déterminé par RT-qPCR ........................................................................................... 119 Figure R-9 : P ofil d’e p essio g i ue des a au sodi ues oltage-dépendant dans les CSCs W8B2+
déterminé par RT-qPCR .............................................................................................................. 120 Figure R-10 : P ofil d’e p essio g i ue des sous-unités alpha des canaux potassiques voltage-
dépendant dans les CSCs W8B2+ déterminé par RT-qPCR ............................................................. 122 Figure R-11 : Ca a t isatio de l’e pression des ARNm codant pour KV4.2 et KV4.3 dans les CSCs
W8B2+ par RT-PCR ..................................................................................................................... 122 Figure R-12 : P ofil d’e p essio g i ue des sous-unités bêta des canaux potassiques voltage-
dépendant dans les CSCs W8B2+ déterminé par RT-qPCR ............................................................. 123 Figure R-13 : Ca a t isatio de l’e p essio des ARN oda t pou KCa . , KCa . et KCa . da s les CSCs W8B2+ par RT-PCR ......................................................................................................... 124 Figure R-14 : P ofil d’e p essio g i ue des a au potassi ues TWIK et TA“K da s les C“Cs W B +
déterminé par RT-qPCR .............................................................................................................. 125 Figure R-15 : Profil d’e p essio g i ue des a au potassi ues e tifia ts e t a ts a e leu s sous-
unités régulatrices) et le courant sensible au baryum enregistré dans les CSCs W8B2+ .................. 126
Figure R-16 : P ofil d’e p essio g i ue des a au HCN da s les C“Cs W B + déterminé par RT-
qPCR ........................................................................................................................................ 127 Figure R-17 : P ofil d’e p essio g i ue des a au al i ues da s les C“Cs W B + déterminé par
RT-qPCR ................................................................................................................................... 129 Figure R-18 : P ofil d’e p essio g i ue des ha geu s Na+/K+, SERCA, PMCA et NCX1 dans les CSCs
W8B2+ déterminé par RT-qPCR .................................................................................................. 130 Figure R-19 : P ofil d’e p essio g i ue des a teu s i pli u s da s l’ho ostasie al i ue da s les CSCs W8B2+ déterminé par RT-qPCR ........................................................................................... 131 Figure R-20 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g ique des
facteurs de transcription cardiaques .......................................................................................... 133
Figure R-21 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des structures contractiles cardiaques .............................................................................................. 135
Figure R-22 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des peptides natriurétiques de type A et de type B déterminé par RT-qPCR ........................................ 136 Figure R-23 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des connexines déterminé par RT-qPCR ............................................................................................ 137
Figure R-24 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux sodiques voltage-dépendant déterminé par RT-qPCR ....................................................... 138 Figure R-25 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux potassiques voltage-dépendant déterminé par RT-qPCR .................................................. 139 Figure R-26 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des sous-
unités bêta des canaux potassiques voltage-dépendant déterminé par RT-qPCR ........................... 140 Figure R-27 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux potassiques TWIK et TASK déterminé par RT-qPCR .......................................................... 141 Figure R-28 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des sous-
unités régulatrices et des canaux potassiques rectifiants entrants déterminé par RT-qPCR ............ 142

Table des illustrations
10
Figure R-29 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux HCN déterminé par RT-qPCR .......................................................................................... 143 Figure R-30 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux calciques déterminé par RT-qPCR ................................................................................... 144 Figure R-31 : Courant dynamique enregistré sur les CSCs W8B2+ différenciées en configuration
cellule-entière ........................................................................................................................... 145 Figure R-32 : Effet de la différenciatio a dia ue des C“Cs W B + su l’e p essio g i ue des échangeurs Na+/K+, SERCA, PMCA et NCX1 déterminé par RT-qPCR ............................................ 146
Figure R-33 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des a teu s i pli u s da s l’ho ostasie calcique déterminé par RT-qPCR ....................................... 147 Figure R-34 : Regroupement hiérarchique des gènes dans les CSCs W8B2+, les CSCs W8B2+ après
différenciation cardiaque (Diff) et les CSCs W8B2+ après différenciation cardiaque et stimulation
optogénétique du CHR2 ............................................................................................................. 150 Figure R-35 : T pes d’a ti it s al i ues spo ta es e egist es au ème jour et au 28ème jour de
différenciation des CSCs W8B2+ .................................................................................................. 152 Figure R-36 : E olutio de la f ue e et de l’a plitude de l’a ti it al i ue spo ta e au ou s des 4 semaines de différenciation cardiaque des CSCs W8B2+ ...................................................... 154 Figure R-37 : E olutio de la du e et de l’ai e de l’a ti it al i ue spo ta e au ou s des semaines de différenciation cardiaque des CSCs W8B2+ ............................................................... 156 Figure R-38 : E olutio de TTP et de TTR de l’a ti it al ique spontanée au cours des 4 semaines de
différenciation cardiaque des CSCs W8B2+................................................................................... 158 Figure R-39 : E olutio de la pe te et de la pe te de l’a ti it al i ue spo ta e au ou s des semaines de différenciation cardiaque des CSCs W8B2+ ............................................................... 159 Figure R-40 : E olutio du te ps d’a ti it e p i e pou e tage de l’a ti it al i ue spo ta e au cours des 4 semaines de différenciation cardiaque des CSCs W8B2+ ......................................... 160 Figure R-41 : Effet de SEA- su l’a ti it al i ue spo ta e e egist e su les C“Cs W8B2+
après 28 jours de différenciation ................................................................................................ 162 Figure R-42 : Effet de la thapsiga gi e su l’a ti it al i ue spo ta e e egist e su les C“Cs W8B2+ après 28 jours de différenciation ..................................................................................... 163 Figure R-43 : Effet de la if dipi e su l’a ti it al i ue spo ta e e egist e su les C“Cs W B +
après 28 jours de différenciation ................................................................................................ 164 Figure R-44 : Effet de la stospo gi e C su les l’a plitude, la f ue e, l’ai e et la du e des oscillations calciques spontanées enregistrées sur les CSCs W8B2+ après 28 jours de différenciation
................................................................................................................................................. 165 Figure R-45 : Effet de la xéstospongine C sur le TTP , le TTR , la pente1 , la pente 2 et le pourcentage
du te ps d’a ti it des os illatio s al i ues spo ta es e egist es sur les CSCs W8B2+ après 28
jours de différenciation .............................................................................................................. 166 Figure R-46 : Effet du 2-APB su l’a plitude, la f ue e, l’ai e et la du e des os illatio s al i ues spontanées enregistrées sur les CSCs W8B2+ après 28 jours de différenciation ............................ 168 Figure R-47 : Effet du 2-APB sur le TTP , le TTR , la pente1 , la pente 2 et le pourcentage du temps
d’a ti it des os illatio s al i ues spo ta es e egist es su les C“Cs W B + après 28 jours de
différenciation ........................................................................................................................... 169 Figure R-48 : Effet de la a odi e su l’a plitude, la f ue e, l’ai e et la du e des os illatio s calciques spontanées enregistrées sur les CSCs W8B2+ après 28 jours de différenciation .............. 170 Figure R-49 : Effet de la ryanodine sur le TTP , le TTR , la pente 1 , la pente 2 et le pourcentage du
te ps d’a ti it des os illatio s al i ues spo ta es e egist es su les C“Cs W B + après 28 jours
de différenciation ...................................................................................................................... 171

Table des illustrations
11
Figure R-50 : Effet de la pa illi e su l’a ti it al i ue spo ta e e egist e su les C“Cs W B +
après 28 jours de différenciation ................................................................................................ 192 Figure R-51 : Effet de la pa illi e su l’a plitude, la f ue e, l’ai e et la du e des os illatio s calciques spontanées enregistrées sur les CSCs W8B2+ après 28 jours de différenciation ............... 194 Figure R-52 : Effet de la paxilline sur le TTP , le TTR , la pente 1 , la pente 2 et le pourcentage du
te ps d’a ti it des os illatio s al i ues spo ta es e egist es su les C“Cs W B + après 28 jours
de différenciation ...................................................................................................................... 195 Figure R-53 : Ca a t isatio de l’e p ession des ARNm codant pour les récepteurs à la sphingosine 1
phosphate de type 1 (S1PR1) et de type 2 (S1PR2) dans les CSCs W8B2+ par RT-PCR ..................... 199 Figure R-54 : Ca a t isatio de l’e p essio des ARN oda t pou les epteu s à la sphi gosi e phosphate de type S1PR3, de type S1PR4 et de type S1PR5 dans les CSCs W8B2+ par RT-PCR ........ 200 Figure R-55 : P ofil d’e p essio g i ue des i epteu s à la “ P “ PR , “ PR , “ PR “ PR et S1PR5) dans les CSCs W8B2+ déterminé par RT-qPCR ................................................................... 201 Figure R-56 : Test de prolifération cellulaire des CSCs W8B2+ par comptage cellulaire en présence de
la S1P ou en situation contrôle .................................................................................................. 202 Figure R-57 : Test de viabilité cellulaire des CSCs W8B2+ par cytométrie en flux en présence de S1P
ou en situation contrôle ............................................................................................................ 202 Figure R-58 : Analyse du cycle cellulaire par cytométrie de flux en présence de la S1P ou en situation
contrôle .................................................................................................................................... 203 Figure R-59 : Test de prolifération cellulaire des CSCs W8B2+ en présence de la S1P, S1P + JTE013 ,
S1P + W146, S1P + BML241 ou en situation contrôle .................................................................. 204 Figure R-60 : Test de formation de colonies CFU-Fs formées par les CSCs W8B2+ en présence de la
S1P , S1P+JTE013, S1P+W146 ou en situation contrôle ............................................................... 205 Figure R-61 : Test de migration cellulaire après 7 heures en présence de la S1P, S1P+JTE013,
S1P+W146 ou en situation contrôle ........................................................................................... 206 Figure D-1 : Mod le h poth ti ue d’u pote tiel d’a tio g pa u e ellule W B +diff e i e ................................................................................................................................................. 218 Figure C-1 : “h a apitulatif des p i ipau g es do t l’e p essio a ie ap s diff e iatio cardiaque in vitro des CSCs W8B2+ ............................................................................................ 229
Liste des tableaux
Tableau H-1 : Principaux marqueurs des cellules souches cardiaques (CSCs) ................................... 45 Tableau H-2 : Protocoles de différenciation cardiaques utilisés pour différents types de CSCs ......... 57 Tableau MM-1 : Liste des a ti o ps utilis s pou l’i u oph ot page pa to t ie e flu ...... 88 Tableau MM-2 : Liste des a ti o ps utilis s pou l’i u o to hi ie i di e te ............................. 94 Tableau MM-3 : Liste des amorces sens et anti-sens utilisées pour la PCR .................................... 101 Tableau MM-4 : Liste des sondes et des 96 gènes analysés par TLDA ........................................... 104 Tableau MM-5 : Liste des anticorps utilisés pour le Western Blot ................................................ 107 Tableau R-1 : Etat d’e p essio des p i ipau g es do t l’e p essio a ie a a t et ap s différenciation cardiaque des CSCs W8B2+ .................................................................................. 148

Abréviations
12
ABREVIATIONS
A
AA : acide ascorbique ADN: acide désoxyribonucléique ADNc: acide désoxyribonucléique complémentaire ADSCs: adipose-derived stem cells ALP: alkaline phosphatase ANG: Angiopoietin ANKB: ankyrine B ANP: atrial natriuretic factor ARN: acide ribonucléique ATP: adénosine triphosphate ATF2: activating transcription factor 2 AT2: alveolar type 2 B
BCRP: breast cancer resistance protein BKCa : Large conductance calcium-activated potassium channel BMSCs: bone marrow stromal cells BNP: brain natriuretic peptide BrdUrd: bromodésoxyuridine C
Ca2+: calcium ion CaM: calmoduline CaMK-II: Ca2+/calmodulin-dependent protein kinase II CD: Cluster of differentiation CDCs : cellules dérivées des cardiosphères CFU-Fs: colony-forming unit-fibroblasts ChR2: channelrhodopsins 2 Cl- : chloride ion CLASP2: cytoplasmic Linker Associated Protein 2 CPEs: cellules progénitrices endothéliales CPHs: cellules progénitrices hépatiques CPS : prosurfactant protein C CREB: C-AMP response element-binding protein CSBAs : cellules souches broncho-alvéolaires CSCs: cellules souches cardiaques CSECs: ellules sou hes de l’ pith liu o e CSEs: Cellules souches embryonnaires CSHs: cellules souches hématopoïétiques CSIs: cellules souches intestinales CSKs: cellules souches des kératinocytes CSMs: cellules souches mésenchymateuses CSNs: cellules souches neurales CSEps: ellules sou hes de l’ pide e CSRs: cellules souches rétiniennes CT: cycle threshold
CX: connexine [Ca2+]i: concentration de calcium intracellulaire D
DG: dental gyrus DMSO: diméthylsulfoxyde

Abréviations
13
DNMT: DNA Methyltransferases dNTP: deoxynucleotide E
eCSCNs : cellules souches épidermiques issues de la crête neurale EDTA: ethylenediaminetetraacetic acid EGF: epidermal growth factor EGFP: enhanced green fluorescent protein EGF-R: epidermal growth factor receptor EGM-2: Endothelial Cell Growth Medium 2 EODHs: ha tillo s hu ai s d’o eillettes d oites ER : endoplasmic reticulum
F
FACS: fluorescence activated cell sorting FGF: Fibroblast growth factors FSC: forward scatter channel G
GFP: green fluorescent protein G-CSF: granulocyte colony-stimulating factor GPCs: glial progenitor cells GPI: glycerophosphatidylinositol H
hEag1 human Ether à Go-Go 1 HEPES 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid HCN Hyperpolarization-activated cyclic nucleotide–gated HLA: human leukocyte antigen HGF : Hepatocyte growth factor I
IA : A-type potassium current ICa.L : L-type calcium current ICa.T : T-type calcium current ICl : chloride current ICl.vol : Volume-sensitive chloride channels IDL: interactive data language IGF : insulin-like growth factor Ih : hyperpolarization-activated cation current IKir : cardiac inward rectifier potassium current IKCa : intermediate-conductance Ca2+-dependent potassium channels IKDR : delayed rectifier potassium current IMDM: Iscove's Modified Dulbecco's Medium INa : sodium current InsP3 : inositol trisphosphate IP: iodure de propidium iPSCs : induced pluripotent stem cells
IP3 : inositol trisphosphate IP3R: inositol trisphosphate receptor ISL1: ISL LIM Homeobox 1 Ito : cardiac transient outward potassium current
J
JNK: c-Jun N-terminal kinases

Abréviations
14
K
K+: potassium ion KCa: calcium-activated potassium channels Kv: voltage-gated potassium channels K2P: two-pore domain potassium channels
L
LIN- : lignage négatif L-VOCC: L type Voltage-operated calcium channels M
MACS: magnetic-activated cell sorting MAPK: mitogen-activated protein kinases MEF2C: myocyte enhancer factor 2C M-MLV: moloney murine leukemia virus MPC: myocyte progenitor cells MSCA-1: mesenchymal stem cell antigen 1 MYH: myosin heavy chain MYL: myosin light chain M199: medium 199 N
Na+: sodium ion Nav: voltage-gated sodium channels NCX: sodium-calcium exchanger Na+/K+ ATPase: sodium-potassium pump NFAT: nuclear factor of activated T-cells NF-KB: nuclear factor-kappa B NPCs: neural progenitor cells N-VOCC: N type Voltage-operated calcium channels O
OB: olfactory bulb OV-6: ovalbumine-6 P
PBS: phosphate-buffered saline PGF: placental growth factor PKC: protein kinase C PMCA : plasma membrane Ca2+ ATPase PMT : photomultiplicateur P/Q-VOCC : P/Q type Voltage-operated calcium channels pRb : retinoblastoma protein P2X P2Y: purinergic receptors Q
QSP : quantité suffisante pour R
RA: retinoic acid RAR: retinoic acid receptor RMS: rostro-migratory stream RPL13A: Ribosomal Protein L13a RT-PCR: reverse transcription polymerase chain reaction RYR: ryanodine receptor R-VOCC : R type Voltage-operated calcium channels S
SCA-1: stem cell antigen 1 SCF: stem cell factor

Abréviations
15
SCs: satellite cells SDF-1: stromal cell-derived factor-1 SERCA: sarco/endoplasmic reticulum Ca2+-ATPase SHH: Sonic hedgehog siRNA: Small interfering RNA SK: small conductance calcium-activated potassium channels SOC: store-operated channel SP: side population SSC: side scatter channel SUR : sous-unité régulatrice SVF: s u de eau fœtal SVZ : subventricular zone S1P: sphingosine 1-phosphate S1PR: sphingosine 1-phosphate receptor T
TA: transit amplifying TAK: tat-associated kinase THY-1: thymus cell antigen 1 TTP: time to peak TTR: time to recovery TRP: transient receptor potential TTX: tetrodotoxin TRPC1: transient receptor potential cation channel subfamily C member 1 TLDA: taqMan low density arrays TNNI: troponin I TNNT: troponin T TNAP: tissue nonspecific alkaline phosphatase TGFβ: transforming growth factor beta T-VOCC: T type Voltage-operated calcium channels U
UA: unité arbitraire V
VEGF: Vascular endothelial growth facto VPC: vascular progenitor cells VGCC HVA: voltage-dependent calcium channels high-voltage-activated VGCC LVA: voltage-dependent calcium channels low-voltage-activated 2-APB: 2-aminoethoxydiphenyl borate W
Wnt 11: wingless-type MMTV integration site family member 11

Introduction
16
INTRODUCTION
Chapitre I : Cellules souches et régénération tissulaire
1 Historique
La biologie des cellules souches a vu sa renaissance à la fin du siècle passé grâce à John
Gurdon (prix Nobel en 2012 avec Shinya Yamanaka), qui est parvenu (dans les années 1960)
à e pla e a e su s le o au d’u e ellule i atu e d’u ovocyte de grenouille avec le
o au d'u e ellule atu e d’o igi e i testi ale. Cet o o te s’est e suite d elopp
normalement.
En 1997, Wilmut et son groupe ont appliqué le résultat de John Gurdon pour démontrer
que le noyau des cellules somatiques avait un potentiel génétique complet en donnant
naissance à la brebis Dolly. Un an après, Thomson et al. (1998) ont développé une méthode
d’isole e t et de ultu e pou ai te i les ellules sou hes embryonnaires (CSEs) humaines
in vitro. Ces travaux ont ouvert les portes pour de nouvelles perspectives de recherche sur les
CSEs (Thomson et al., 1998).
Pou e ui est des ellules sou hes adultes, l’histoi e o e e avec Ernest McCulloch
et James Till (dans les années 1960) ui s’i t essaie t au effets des adiatio s d'une
explosion atomique) sur la capacité de la moelle osseuse à produire des cellules sanguines.
McCulloch a o se des g u eau su la ate d’u e sou is a a t eçu des i je tio s de
moelle. Ces deux chercheurs ont pensé que ces nodules étranges provenaient d'une seule
cellule, probablement de la moelle osseuse.
En 1966, Friedenstein et ses collègues ont caractérisé les p op i t s d’u e petite
population mixte de cellules de la moelle osseuse capable d’adh er au plastique in vitro.
Cette populatio i te tait o stitu e de ellules sou hes h atopoï ti ues et d’u e aut e
populatio ellulai e, i o ue à l’ po ue, apa le de do e aissa e à des ellules «
réticulaires » et à des ostéoblastes. Cette population inconnue correspond à e ue l’o
appelle maintenant les cellules souches mésenchymateuses (CSMs). Cette équipe a également
o t l’i po ta e de l’i te a tio des C“Ms avec le microenvironnement de la moelle
osseuse (ou niche hématopoïétique) mais aussi leur capacité à se différencier en tissu

Introduction
17
d’o igi e sode i ue. Da s e tai es o ditio s, les CSMs étaient capables de se
différencier en ostéoblastes, chondrocytes et adipocytes (Caplan, 1986; Piersma et al., 1985).
Dans les années 1990-2000, le pouvoir de différenciation myogénique des CSMs a été
démontré (Wakitani et al., 1995), notamment leur différenciation en cardiomyocytes in vivo
(Toma et al., 2002; Vacanti et al., 2005). Durant la même période une autre étude a pu mettre
en évidence un pouvoir immuno-modulateur des CSMs par surpression de la prolifération des
lymphocytes T (Di Nicola et al., 2002). Cette tude a atti pa ti uli e e t l’atte tio du
ilieu s ie tifi ue su l’appli atio th apeuti ue des CSMs lors de transplantations
allogéniques.
Ces études ont révélé la plasticité et le pouvoir multipotent des cellules souches adultes
et ont ouvert les possibilités de thérapies cellulaires.
Avant de faire une description plus exhaustive des différents types de cellules souches, il
est important de rappeler les propriétés intrinsèques qui les définissent.
2 Propriétés des cellules souches
2.1 L’auto-renouvellement
Les cellules souches sont définies comme étant des cellules indifférenciées dotées de
apa it d’auto-renouvellement et de diff e iatio e d’aut es t pes ellulai es. L’auto-
renouvellement est un processus par lequel une cellule souche se divise asymétriquement ou
symétriquement pour générer respectivement une ou deux cellules filles qui présentent un
potentiel de développement similaire à celui de la cellule mère. Les divisions symétriques
suppose t ue les ellules filles o se e t les a a t isti ues p op es d’u e ellule sou he
(Ulloa-Montoya et al., 2005). La apa it d’auto-renouvellement est essentielle pour les
cellules souches pour accroitre leur nombre durant le développement, être maintenues au
sein des tissus adultes et rétablir le stock des cellules souches après blessure. L’auto-
renouvellement se différencie de la prolifération même si les deux processus sont liés à la
division cellulaire. La prolifération est un terme plus générique qui regroupe tous les types de
di isio s des ellules sou hes et p og it i es. L’auto- e ou elle e t i pli ue u’au oi s
une des cellules filles possède le même potentiel de développement que la cellule mère.
Pour la plupart des cellules souches de mammifères, telles que les cellules souches
hématopoïétiques (CSHs) et les cellules souches neurales (CSNs), l'auto-renouvellement est

Introduction
18
une division avec maintien de la multipotence. Pour les cellules souches qui forment un seul
type de cellule fille, par exemple les cellules souches spermatogoniales, l'auto-renouvellement
est la division avec la maintenance de l'état indifférencié. La plupart des mécanismes
impliqués dans l'auto-renouvellement des cellules souches régulent ceux impliqués dans la
prolifération de nombreux types cellulaires (Molofsky et al., 2003; Nishino et al., 2008).
Les ellules sou hes o se e t pe da t de t s lo gues p iodes leu apa it d’auto-
e ou elle e t. Il est ie ta li u’à ha ue dupli atio du g o e, des e eu s de le tu e
ou des do ages de l’ADN peu e t su e i . Les ellules souches possèdent des mécanismes
de surveillance qui détectent les erreurs de réplication et éliminent les cellules dans lesquelles
apparaissent ces erreurs. Il y aurait intervention de la protéine p53 bien connue pour son rôle
de facteur anti-tumoral et pro-apoptotique, et de la protéine du rétinoblastome (pRb), elle
aussi facteur anti-tumoral. Ce mécanisme préviendrait ainsi la formation de tumeur par
utatio d’u e ellule sou he et p olif atio de ette ellule ut e MORRI“ON et
SPRADLING, 2008).
2.2 Pouvoir de différenciation en plusieurs lignages cellulaires
La différenciation est définie comme un changement dans le phénotype cellulaire en raison
de l’e p essio de g es sp ifi ues de la fo tio ellulai e sui a t le lig age au uel elle
appartient (Gargett, C.E. . Co e ous l’a o s d jà e tio , les cellules souches
e t e t da s la oie de diff e iatio pa des di isio s dites as t i ues i pli ua t u’u e
seule des deux cellules filles conserve les caractéristiques propres des cellules souches, alors
ue l’aut e ellule fille o ti ue da s la oie de la diff e iatio (Ulloa-Montoya et al., 2005).
Il semblerait que l’as t ie soit li e à u e as t ie ol ulai e au sei de la ellule
souche en division. Il y aurait une ségrégation asymétrique des protéines, des ARNs, des
o ga elles et de l’ADN. Il se le ait gale e t ue le fuseau essai e à la di isio ellulaire
soit lui aussi asymétrique (Morrison and Spradling, 2008). E fi l’e i o ement
i te ie d ait aussi da s l’as t ie des di isio s. La ellule fille o te ue e t a t da s la oie
de diff e iatio est appel e ellule d’a plifi atio t a sitoi e t a sit a plif i g p oge ito
ou cellule TA) (Gargett, 2007). Ces cellules TA, aussi appelées, suivant leur stade, cellules
progénitrices puis cellules précurseurs, possèdent des propriétés intermédiaires entre les
cellules souches adultes et les cellules différenciées : elles ont un potentiel de division limité,

Introduction
19
u e a se e de apa it à l’auto-renouvellement et donnent naissance progressivement aux
cellules différenciées (Figure H-1).
La division asymétrique jouerait aussi un rôle dans la apa it u’o t les ellules sou hes
à se ai te i lo gte ps : les utatio s de l’ADN se aie t sp ifi ue e t e o es da s la
ellule fille pou sui a t la diff e iatio alo s ue l’ADN o igi al, i tact, serait conservé dans
la cellule souche fille respo sa le de l’auto- e ou elle e t. Ce ph o e ’a epe da t
pas été décrit dans tous les systèmes (Morrison and Spradling, 2008).
3 Types et potentiels de différenciation des cellules souches humaines :
Le pouvoir de différenciation des cellules souches est différent selon le type de cellules
souches qui peuvent être totipotentes, pluripotentes ou multipotentes (Laurie, 2004;
Takahashi et al., 2007; Thomson et al., 1998) (Figure H-2).
Figure H-1 : Hiérarchie de la différenciation des cellules souches. Les cellules souches subissent des divisions cellulaires as t i ues, e ui leu pe et de s’auto-renouveler ou de se différencier pour donner naissance à des progéniteurs engagés. Ces derniers prolifèrent et donnent lieu à des « cellules amplificatrices de transit » ou (transit amplifying (TA) cells) plus différenciées, qui prolifèrent rapidement et se différencient finalement pour produire de nombreuses cellules fo tio elles diff e i es sa s apa it de p olif atio . D’ap s Ga gett, C.E et al., .
Introduction
20
3.1 Les cellules souches embryonnaires (CSEs) :
3.1.1 Les cellules souches totipotentes :
Les cellules souches ont la capacité de se différencier en tous types cellulaires
spécialisés. Le nombre de type de cellules spécialisées différents que peut engendrer une
cellule souche définit sa potentialité.
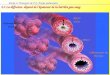
Figure H-2 : La hi a hie des ellules sou hes. Le z gote totipote t fo pa la fusio de l’o ule et le spermatozoïde se divise pour former la masse cellulaire interne (ICM ou inner cell mass) et le tissu extra-embryonnaire (EE ou extra-embryonic) du blastocyste. Lorsqu'elles sont isolées du blastocyste, les cellules de l'ICM peuvent être maintenues in vitro en culture sous la forme de lignées de cellules souches embryonnaires (CSEs) pluripotentes. Au cours du développement de l'embryon, les cellules souches pluripotentes de l'ICM sont de plus en plus restreintes dans leur potentiel de lignage et génèrent des cellules souches multipotentes tissu-spécifiques. Celles-ci incluent des cellules souches épidermiques (cellules souches du follicule pileux) qui forment la peau et les cheveux, des cellules souches hématopoïétiques dans la moelle osseuse qui donnent naissance à toutes les cellules hématopoïétiques, des cellules souches neurales dans la zone subventriculaire du cerveau, des cellules souches gastro-intestinales situées dans la crypte de l'intestin grêle, des cellules ovales qui donnent naissance au foie (non montrées) et des cellules souches mésenchymateuses qui résident dans la moelle osseuse et peuvent former des cellules osseuses, stromales et des adipocytes (non représentées). D’ap s E kfeldt, C.E et al. .

Introduction
21
Les ellules sou hes totipote tes o t la apa it de do e aissa e à ’i po te uel
t pe ellulai e o stitua t l’o ga is e hu ai ompris les tissus extra-embryonnaires
comme le placenta (Thomson et al., 1998). L’e iste e de es ellules sou hes totipote tes
est courte, et dure entre la fertilisation et la formation du blastocyste, lorsque le zygote ou
œuf f o d o e e à se di ise jus u’à a oi huit ellules ide ti ues (Figure H-2).
3.1.2 Les cellules souches pluripotentes :
Au uat i e jou du d eloppe e t e o ai e, l’a as ellulai e fo e le
lasto ste . Le lasto ste est o stitu de deu ou hes ; une couche externe qui se
transforme en tissus extra-embryonnaires et la couche interne qui est un ensemble de 30
ellules e i o ui fo e o t pa la suite l’o ga is e (Eckfeldt et al., 2005) (Figure H-2).
Ces cellules sont pluripotentes et ont le potentiel de se différencier en 200 types
ellulai es à l’esse e du o ps hu ai (Laurie, 2004). Ainsi, les cellules souches pluripotentes
de la couche interne du blastocyste représente une source pour créer des lignées de CSEs
humaines (Laurie, 2004; Thomson et al., 1998). A la différence des cellules souches
totipotentes, les ellules sou hes plu ipote tes e peu e t pas t e à l’o igi e d’u t e
hu ai puis u’elles sont incapables de constituer les tissus extra-embryonnaires essentiels
au développement embryonnaire. Les cellules souches pluripotentes peuvent donc former les
trois feuillets germinaux (endoderme, mésoderme et ectoderme) et se spécialiser ensuite en
d’aut es t pes de ellules sou hes ue so t les ellules sou hes ultipote tes (Enmon et al.,
2002).
3.2 Les cellules souches adultes:
On retrouve les cellules souches adultes multipotentes dans plusieurs tissus et organes
comme la moelle osseuse, le tissu adipeux, le sang périphérique, le muscle squelettique,
l’ pide e, le œu , le foie et le e eau. Ces ellules so t « o ga es-spécifiques », se forment
pe da t le d eloppe e t fœtal et pe du e t da s l’o ga is e adulte (figure H-2).
Elles sont indiffére i es a e u e apa it d’auto-renouvellement et une forte
prolifération et se différencient en cellules spécialisées avec des fonctions spécifiques
(Pittenger et al., 1999). Les cellules souches multipotentes comme par exemple les cellules
souches hématopoïétiques, mésenchymateuses et épidermiques, peuvent êtres différenciées
in vitro et utilisées en médecine régénérative (Ma, 2010; Macrin et al., 2017). Malgré leur

Introduction
22
pouvoir limité de différenciation, les cellules souches adultes multipotentes peuvent être
orientées vers plusieurs types cellulaires et facilement manipulées in vitro contrairement aux
CSEs do t l’utilisatio est sou e t t s li it e pou des uestio s thi ues. Les ellules
souches multipotentes adultes se présentent donc comme de bons candidats pour le
traitement de multiples pathologies dans le domaine de la médecine régénérative et de la
thérapie cellulaire. Cependant, il est nécessaire et indispensable de bien caractériser et
comprendre in vitro leu s a is es d’auto-renouvellement, de prolifération et de
différenciation.
Avant de développer plus spécifiquement les cellules progénitrices cardiaques il est
important de faire une description plus généraliste (synthétique) des différents types de
cellules souches adultes
3.2.1 Les cellules souches hématopoïétiques (CSHs)
Elles font partie des cellules souches adultes de la moelle osseuse les plus connues et
les plus étudiées. Les cellules souches hématopoïétiques sont des cellules multipotentes et
donnent naissance aux deux lignées myéloïde et lymphoïde des cellules sanguines. Ces cellules
ont été utilisées avec succès dans la thérapie cellulaire (Kuznetsov et al., 2001) et peuvent être
aussi trouvées dans le sang périphérique ou dans le sang du cordon ombilical des nouveaux-
nés. Les CSHs o t t à l’o igi e du d ut des e he hes ode es su les ellules sou hes
adultes grâce aux découvertes faites sur leur très forte capacité d’auto-renouvellement et de
différenciation en tous les types cellulaires du lignage hématopoïétique (figure H-3).

Introduction
23
Les CSHs peuvent être isolées par prélèvement direct de la moelle osseuse hématopoïétique
mais aussi prélevées dans le sang périphérique après stimulation par des cytokines (Sackstein,
2004). Les CSHs de la moelle osseuse expriment à leur surface des marqueurs qui permettent
de les identifier comme le CD34, KDR, CD45, AC133. Elles expriment peu les marqueurs THY-
1, KIT et SCA-1. Les CSHs ont la capacité de se différencier en toutes les lignées cellulaires qui
composent le sang. Leur pouvoir de différenciation in vivo en cardiomyocytes a été analysé
mais les résultats obtenus in vitro sont controversés ; des études ont montré que les CSHs
isolées de la moelle osseuse pouvaient exprimer plusieurs facteurs de transcription cardiaques
intégrant les protéines sarcomériques (Eisenberg et al., 2003).
Plusieurs études ont suggéré que des cellules isolées de moelle osseuse et donc
enrichies en CSHs pou aie t g e le o a de da s des od les a i au d’i fa tus
(Agbulut et al., 2003; Jackson et al., 2001; Kajstura et al., 2005; Orlic et al., 2001a). Cependant,
Figure H-3 : Les cellules souches hématopoïétiques (CSHs). Les CSHs sont dotées d'une capacité
d'auto-renouvellement (flèche incurvée) qui maintient leur stock et de potentiel de différenciation
multipotent qui permet de produire toutes les cellules du système hématopoïétique mature. À
esu e ue les C“Hs se diff e ie t, elles pe de t leu apa it d’auto-renouvellement et leur
pouvoir multiple de différenciation devient de plus en plus restreint. Les CSHs donnent naissance
à des progéniteurs (CLP, CMP, MEP et GMP) qui s’e gage t da s u p o essus de diff e iatio pour donner à la fin des cellules spécialisées (lymphocyte B, lymphpcyte T, cellules NK etc ...
CLP (common lymphoid progenitor); CMP (common myeloid progenitor); GMP, (granulocyte
monocyte progenitor); MEP (megakaryocyte erythrocyte progenitor); NK (natural killer). D’ap s Eckfeldt, C.E et al.2005.

Introduction
24
des publications ont montré que la même population de cellules adoptait uniquement un
phénotype hématopoïétique après transplantation (Balsam et al., 2004; Murry et al., 2004).
D’aut es st at gies o e la o ilisatio de ellules p u seu s h atopoï ti ues pa
du Stem Cell Factor et du Granulocyte-Colony Stimulating Factor ou G-CSF pendant le
d oule e t d’u i fa tus o t gale e t do des sultats o t adi toi es (Deten et al.,
2005; Orlic et al., 2001b).
3.2.2 Les cellules souches mésenchymateuses (CSMs)
Les cellules souches mésenchymateuses appelées également cellules souches stromales
ont été également retrouvées dans la moelle osseuse. Co e ous l’a o s d jà e tio
les CSMs de la moelle osseuse ont été décrites pour la première fois par Ernest McCulloch et
James Till dans les années 1960. D’aut es he heu s du a t les a es -80 (Friedenstein
(1970), Owen (1988), Tavassoli et Crosby (1970), qualifiaient ces cellules de « mechanocytes
dérivées de la moelle osseuse » ou encore « des fibroblastes stromaux ». Ce ’est u’à pa ti
de 1991 que Caplan nomma ces cellules ; «cellules souches mésenchymateuses ». Les CSMs
so t fusifo es et o t la p op i t d’adh e au plastique in vitro. A l’i e se, la plupa t des
autres types de cellules souches de la moelle osseuse comme les CSHs ’o t pas ette
p op i t d’adh e e au plasti ue (Friedenstein, 1995). Les CSMs peu e t s’auto-renouveler
et se différencier en lignage mésodermique (adipocyte, chondrocytes, ostéoblastes et cellules
stromales) (figure H-4).
Des études ont montré la capacité des CSMs à se transdifférencier in vitro e d’aut es
lignages (ectoderme et endoderme) mais cette transdifférenciation est controversée in vivo
(figure H-4). Elles interviennent in vivo lors des processus de réparations des tissus
e do ag s o e l’os, le a tilage, le us le, le liga e t, le te do et le st o a (Pittenger
et al., 1999; Psaltis et al., 2008). Les CSMs ont pu être isolées à partir de plusieurs tissus comme
le sang périphérique (Kuznetsov et al., 1997), le sang du cordon ombilical (Lee et al., 2004), le
sang foetal (Noort et al., 2002), la pulpe dentaire (Gronthos et al., 2000), le liquide amniotique
I ’t A ke et al., , le poumon (Fan et al., 2005), le foie (Campagnoli et al., 2001), le tissu
adipeux (Zuk et al., 2002), l’i testi (Bjerknes and Cheng, 2002) et le follicule pileux (Amoh et
al., 2005).
Introduction
25
U des p o l es d’ide tifi ation des CSMs est l’a se e de a ueu s
immunophénotypiques universels qui leur sont spécifiques (Rastegar et al., 2010; Williams
and Hare, 2011). Cependant, le comité de la société internationale pour la thérapie cellulaire
s’i t essa t au cellules souches mésenchymateuses (the International Society for Cellular
Therapy), a bien défini certains critères pour décrire une CSM, qui se résume en leur adhésion
au plastique en conditions de culture. De plus une CSM doit exprimer le CD105, CD73 et CD90
et doit être négative pour le CD45, CD34, CD14, CD79, CD19 et HLA-DR et doit se différencier
en ostéoblastes, adipocytes et chondrocytes in vitro (Dominici et al., 2006). En général, elle
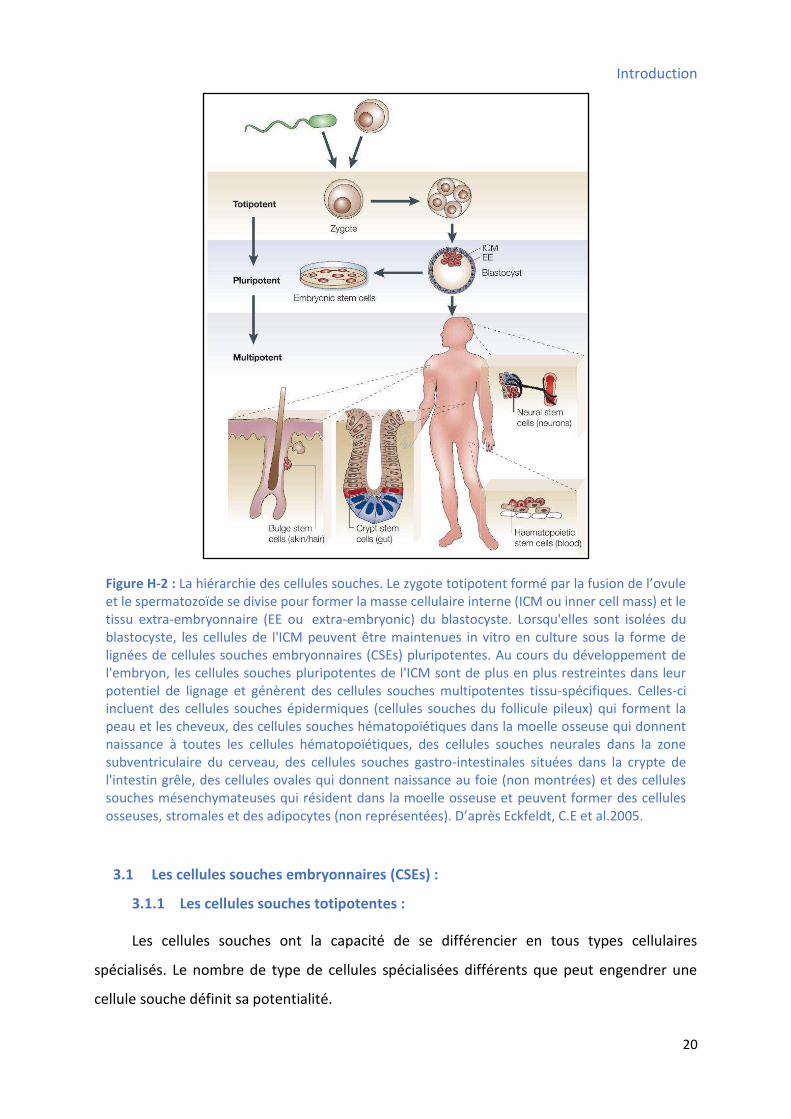
Figure H-4 : Les cellules souches mésenchymateuses (MSCs). Les MSCs de la moelle osseuse ont la
apa it de s’auto-renouveler (flèche incurvée) et à se différencier vers la lignée mésodermique
en donnant naissance aux cellules stromales, chondrocytes, adipocytes et ostéoblastes, (flèches
o ti ues . D’aut es apa it s de diff e iatio in vitro vers les lignages ectodermique et
e dode i ue, o t t do u e t es da s la litt atu e fl hes poi till es . D’ap s U elli, A et al.2008.

Introduction
26
’e p i e pas le CD , le CD , le CD et le CD (Majumdar et al., 2003). D’aut e pa t, les
CSMs sont connues pour être positives pour les peptides de surface SH2, SH3, SH4, le CD35
(trans-membrane protein), le CD73, le CD90 (Thy1), le CD123, le CD117, CD271 et le Stro-3-
positive mesenchymal precursor cell (Kuçi et al., 2010). Ces ellules sou hes doi e t d’aut e
pa t e p i e des a ueu s e o ai es o e l’O t , le Na og et le ““EA-4 (Gang et
al., 2007).
N a oi s, d’aut es t pes ellulai es, o e les CSHs, expriment également certains
des marqueurs déjà cités rendant la caractérisation des cellules souches mésenchymateuses
plus complexe (Kim and Ahn, 2012). Il en résulte que pour bien distinguer les CSMs des CSHs,
une sélection basée sur l'absence des marqueurs hématopoïétiques CD34, CD45 et CD14
serait suffisante (Majumdar et al., 2003).
Les CSM peuvent se différencier en plusieurs types cellulaires. La première
différenciation a été réalisée par Friedenstein et ses collaborateurs (1970) qui ont pu isoler et
différencier les CSMs en tissus osseux in vitro. Différents groupes ont montré par la suite que
les CSMs peuvent se différencier en cellules pancréatiques (Lee et al., 2004) , neuronales
(Woodbury et al., 2000), en os (Haynesworth et al., 1992), en cartilage (Yoo et al., 1998), en
muscle (Wakitani et al., 1995), stroma de la moelle (Majumdar et al., 1998), tendon et
ligament (Young et al., 1998), tissu adipeux (Dennis et al., 1999) et en différents tissus
connectifs (Studeny et al., 2004). Les CSMs de la moelle osseuse sont également capables de
freiner et « réparer » les altérations pulmonaires (Curley et al., 2012), le diabète (Si et al.,
2012), les désordres neurologiques (Edalatmanesh et al., 2011), les maladies rénales (Alfarano
et al., 2012) et les blessures hépatiques (Zhao et al., 2012). En plus de leur pouvoir de
différenciation en différents lignages mésenchymateux, les CSMs peuvent aussi donner
aissa e sous e tai es o ditio s de ultu e à d’aut es ellules sp ialisées comme les
cardiomyocytes.
Ces t a au o t pe is de ett e e ide e des ha ge e ts d’e p essio de
certains gènes cardiaques comme la chaîne lourde de myosine, la troponine T cardiaque, des
facteurs de transcription précoces cardiaques comme NKx2.5 et GATA4 (Reik, 2007). GATA4
est u gulateu ajeu de l’e p essio des g es a dia ues du a t la diff e iatio
(Heineke et al., 2007). P se t da s le œu adulte il joue u ôle de gulateu
transcriptionnel de nombreux gènes cardiaques comme le facteur natriuretic atrial, le peptide

Introduction
27
natriuretic de type B, la chaîne lourde de myosine et la troponine T cardiaque qui sont deux
protéines contractiles cruciales dans la fonction musculaire squelettique et cardiaque.
3.2.3 Les cellules souches du tissu adipeux
Des études ont montré que le tissu adipeux contient des cellules souches nommées
cellules mésenchymateuses dérivées du tissu adipeux (ADSCs ou Adipose tissue-derived stem
cells) (Nakagami et al., 2005). Ce tissu est a o da t, fa ile d’a s et ep se te u e
potentielle source de cellules pour des transplantations autologues. Les ADSCs sont
multipotentes et sont localisées au niveau du stroma vasculaire du tissu adipeux sous cutané.
Ces cellules ont beaucoup de similitude avec les CSMs de la moelle osseuse. Elles seraient
positives pour les marqueurs SCA-1 et CD44 et négatives pour les marqueurs KIT, CD11b,
CD31, CD34 ou CD45 (Nakagami et al., 2005). Elles présentent un phénotype proche de celui
des fibroblastes et peuvent se différencier in vitro en adipocytes, ostéoblastes et
chondroblastes. Les ADSCs secrètent de o euses toki es HGF, VEGF, PGF, TGFβ, FGF,
ANG-1 et ANG) mais aussi des facteurs angiogéniques comme le VEGF et le HGF. Les ADSC
partagent beaucoup de gènes avec les CSMs de la moelle osseuse et expriment des marqueurs
hématopoéitiques (CD34, CD133 et ABCG2) (Planat-Benard et al., 2004) suggérant une double
populatio d’AD“Cs : elles de t pe h atopoï ti ue et elles de t pe se h ateu .
3.2.4 Les cellules progénitrices endothéliales
Les cellules progénitrices endothéliales (CPEs) sont des cellules circulantes de la moelle
osseuse ui side t au sei de l’e doth liu et o t i ue t à la as uloge se et
l’ho ostasie as ulai e (Khakoo and Finkel, 2005) (figure H-5). Elles ont été identifiées pour
la première fois en 1997 (Asahara et al., 1997) et ont été isolées à partir du sang périphérique
comme étant des cellules exprimant le marqueur CD34. Ces cellules seraient natives de la
moelle osseuse.
.

Introduction
28
Les h a gio lastes p se ts pe da t l’e oge se se aie t à l’o igi e des ellules
précurseurs des endothéliums (Kubo and Alitalo, 2003; Risau and Flamme, 1995) mais
pourraient également provenir directement des CSHs (Kubo and Alitalo, 2003). En effet, les
CPEs partagent des marqueurs communs avec les CSHs comme le CD133 et CD34 mais elles
expriment en plus le marqueur endothélial FLK-1 (Liao et al., 2007). Les CPEs se présenteraient
comme une sous population de CSHs a a t la apa it d’a u i u ph ot pe e doth lial
(Liao et al., 2007). Les CPEs sont très peu nombreuses dans le sang périphérique et dans la
moelle, limitant leur utilisation (Nakagami et al., 2005). Ces cellules ont été utilisées en
thérapie cellulaire pour différentes pathologies comme certaines maladies cardiovasculaires
(Kawamoto and Losordo, 2008) et le diabète (Fadini et al., 2010).
Figure H-5 : Les cellules progénitrices endothéliales (CPEs). Les CPEs peuvent dériver de cellules
embryonnaires (hémangioblastes) et de plusieurs tissus adultes (cellules CD14+/ CD34+/CD133+ du
sang périphérique, moelle osseuse, paroi vasculaire et myocarde). Les CPEs sont impliquées
esse tielle e t da s la ee dothelisatio des aisseau l s s et da s l’a gioge se da s des zo es ischémiques. D’ap s Le i, A., a d Kajstu a, J. .

Introduction
29
3.2.5 Les ellules sou hes de l’ pith liu i testi al :
Les ellules sou hes de l’ pith liu i testi al fo t pa tie des ellules sou hes
pith liales des tissus à e ou elle e t apide et e pos s au ilieu e t ieu . L’i testi a u
ôle p ote teu ajeu pa la fo atio d'u e a i e a e le ilieu e t ieu d’où la
nécessité de se renouveler en permanence. Ce renouvellement (tous les deux à sept jours) est
assuré par la présence de cellules souches qui en se différenciant, donnent différents types
cellulaires avec différentes fonctions. Les cellules souches intestinales (CSIs) résident dans les
cryptes et expriment à leur surface les marqueurs (MSI-1), CD-24 et KIT (Reya and Clevers,
2005). Elles sont multipotentes et se différencient en quatre types cellulaires : les entérocytes
ôle d’a so ptio des ali e ts , les ellules u ipa es ali ifo es p ote tio de la su fa e
de l'épithélium par la sécrétion du mucus), les cellules entéroendocrines (sécrétion des
hormones peptidiques) et les cellules de Paneth (sécrétion de lysozymes et rôle immunitaire)
(Goodell et al., 2015) (figure H-6).
Les entérocytes, les cellules mucipares caliciformes et les cellules entéroendocrines se
diff e ie t e ig a t e s l’e t it des ptes et e haut des illosit s, es ellules
su isse t l’apoptose et des ua e t da s la lu i e i testinale. Les cellules de Paneth se
différencient en migrant vers la base de la crypte. Ces cellules ont une durée de vie plus longue
ue les aut es t pes ellulai es a a t d’ t e phago t es pa les ellules e i o a tes.
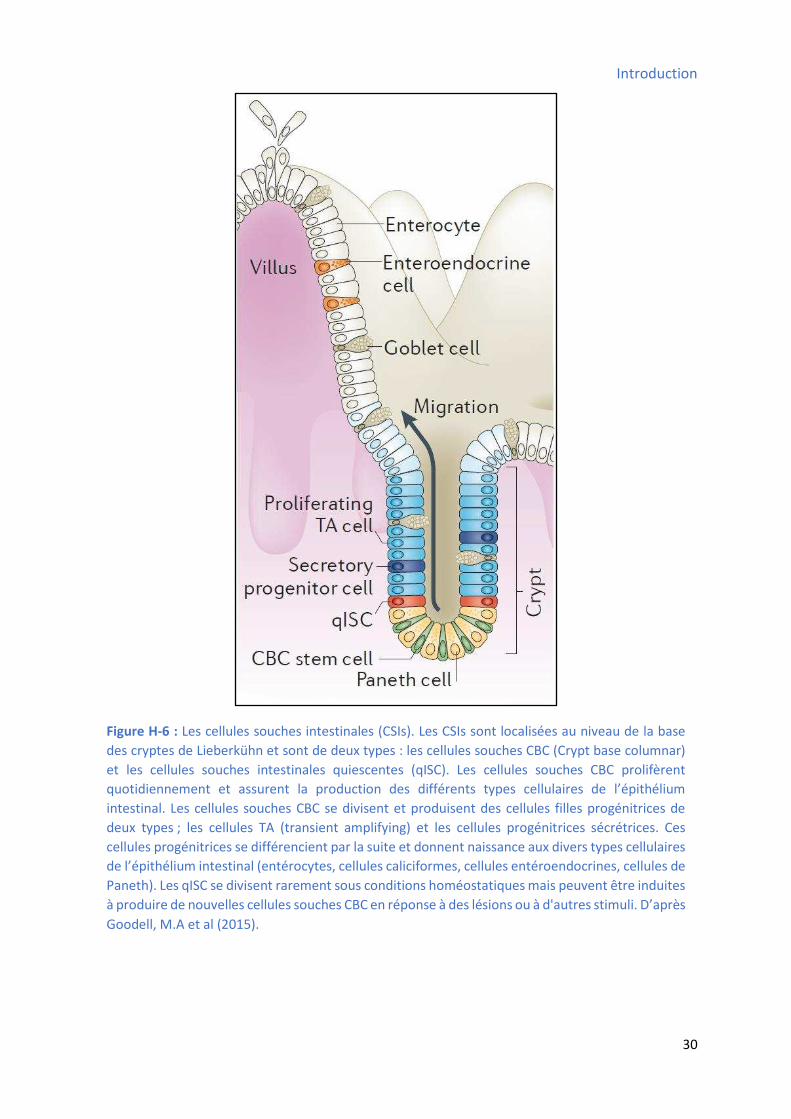
Introduction
30
Figure H-6 : Les cellules souches intestinales (CSIs). Les CSIs sont localisées au niveau de la base
des cryptes de Lieberkühn et sont de deux types : les cellules souches CBC (Crypt base columnar)
et les cellules souches intestinales quiescentes (qISC). Les cellules souches CBC prolifèrent
quotidienne e t et assu e t la p odu tio des diff e ts t pes ellulai es de l’ pith liu intestinal. Les cellules souches CBC se divisent et produisent des cellules filles progénitrices de
deux types ; les cellules TA (transient amplifying) et les cellules progénitrices sécrétrices. Ces
cellules progénitrices se différencient par la suite et donnent naissance aux divers types cellulaires
de l’ pith liu i testi al e t o tes, ellules ali ifo es, ellules e t oe do i es, ellules de Paneth). Les qISC se divisent rarement sous conditions homéostatiques mais peuvent être induites
à produire de nouvelles cellules souches CBC en réponse à des lésions ou à d'autres stimuli. D’ap s Goodell, M.A et al (2015).

Introduction
31
3.2.6 Les ellules sou hes de l’ pide e
L’ pide e est u tissu qui se renouvelle constamment par le phénomène de
des ua atio . Cette de i e o espo d à l’ li i atio des ellules de la ou he o e ui
sont replacées rapidement par les kératinocytes. Il existe ainsi une population de cellules
souches multipotentes apa les d’auto e ou elle e t et ui e p i e t des a ueu s
sp ifi ues o e la K , des ol ules d’adh sio o e des β -intégrines, des E-
adh i es et des β-caténines. Ces cellules souches portent le nom de cellules souches des
kératinocytes (CSKs) (Watt, 2002). Ces CSKs donnent rapidement des cellules intermédiaires
qui se détachent de la membrane basale et se différencient (Solanas and Benitah, 2013)
(figure H-7). On a ainsi des CSKs au niveau de la couche basale associée à des cellules
intermédiaires, des cellules intermédiaires dans la couche épineuse, des cellules différenciées
dans la couche granuleuse et des cellules mortes dans la couche cornée.
Les CSKs so t issues d’aut es ellules sou hes ultipote tes appel es ellules sou hes
de l’ pide e C“Eps). Les CSEps e p i e t les a ueu s CD , le K et la α -intégrine
(Mimeault and Batra, 2006). Les CSEps so t à l’o igi e des cellules progénitrices de la matrice
du follicule pileux, des cellules progénitrices des glandes sébacées et des cellules de la papille
de i ue. A la diff e e des C“Ks ui e do e t aissa e u’au cellules épidermiques,
les CSEps se différencient à la fois e ellules de l’ pide e et e ellules p og it i es de la
matrice du follicule pileux. En plus des CSKs et des CSEps, la peau contient également des
cellules souches pluripotentes épidermiques issues de la crête neurale (eCSCNs). Les eCSCNs
ont été découvertes par LI et al. en 2003 chez la souris comme des cellules exprimant la
nestine (marqueur des cellules de la crête neurale) et retrouvées en partie au niveau du bulbe
du follicule pileux. Les eCSCNs expriment aussi le CD34 et THY-1 et peuvent se différencier en
plusieurs lignages cellulaires incluant des neurones, des mélanocytes, des cellules de Schwann
et des cellules de Merkel (Mimeault and Batra, 2006) et (Sakaue and Sieber-Blum, 2015). Les
eNCSCs semblent être localisées au niveau du bulbe du follicule pileux mais leur localisation
change suivant le cycle folliculaire (Li et al., 2003).
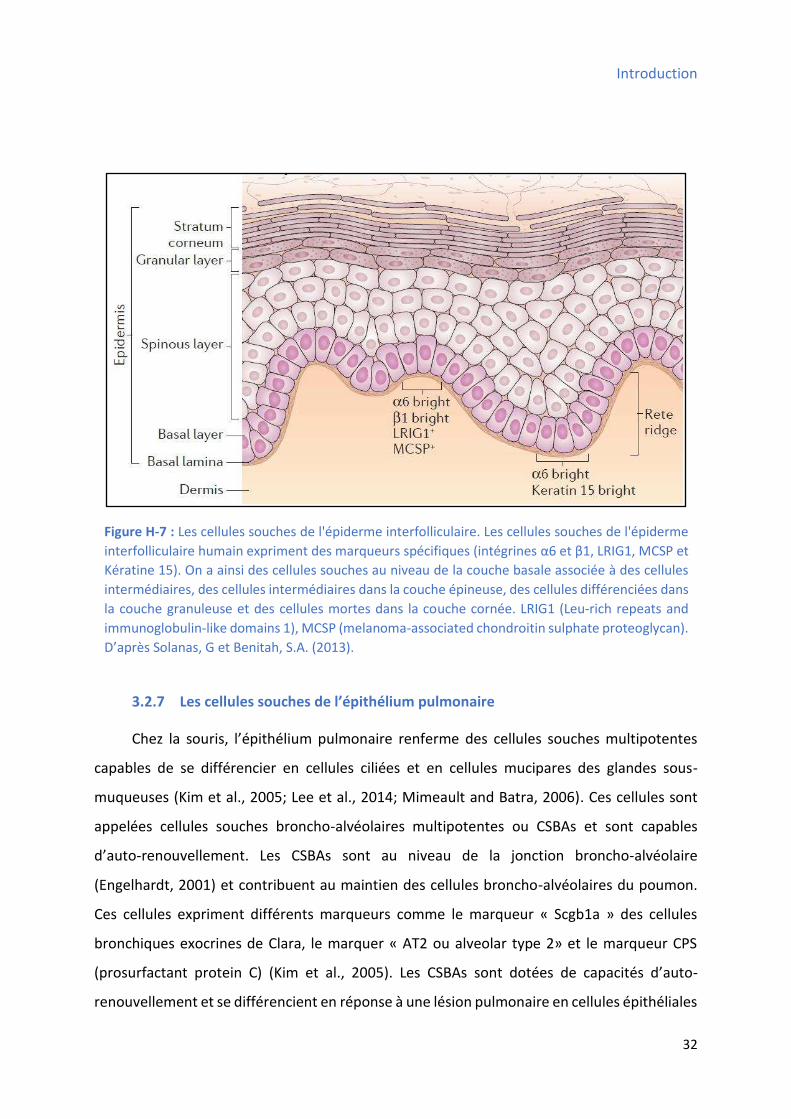
Introduction
32
3.2.7 Les ellules sou hes de l’ pith liu pul o ai e
Chez la sou is, l’ pith lium pulmonaire renferme des cellules souches multipotentes
capables de se différencier en cellules ciliées et en cellules mucipares des glandes sous-
muqueuses (Kim et al., 2005; Lee et al., 2014; Mimeault and Batra, 2006). Ces cellules sont
appelées cellules souches broncho-alvéolaires multipotentes ou CSBAs et sont capables
d’auto-renouvellement. Les CSBAs sont au niveau de la jonction broncho-alvéolaire
(Engelhardt, 2001) et contribuent au maintien des cellules broncho-alvéolaires du poumon.
Ces cellules expriment différents marqueurs comme le marqueur « Scgb1a » des cellules
bronchiques exocrines de Clara, le marquer « AT2 ou alveolar type 2» et le marqueur CPS
(prosurfactant protein C) (Kim et al., 2005). Les CSBAs so t dot es de apa it s d’auto-
renouvellement et se différencient en réponse à une lésion pulmonaire en cellules épithéliales
Figure H-7 : Les cellules souches de l'épiderme interfolliculaire. Les cellules souches de l'épiderme
interfolliculaire humain expriment des marqueurs spécifiques (intégrines α6 et β1, LRIG1, MCSP et
Kératine 15). On a ainsi des cellules souches au niveau de la couche basale associée à des cellules
intermédiaires, des cellules intermédiaires dans la couche épineuse, des cellules différenciées dans
la couche granuleuse et des cellules mortes dans la couche cornée. LRIG1 (Leu-rich repeats and
immunoglobulin-like domains 1), MCSP (melanoma-associated chondroitin sulphate proteoglycan). D’ap s “ola as, G et Benitah, S.A. (2013).
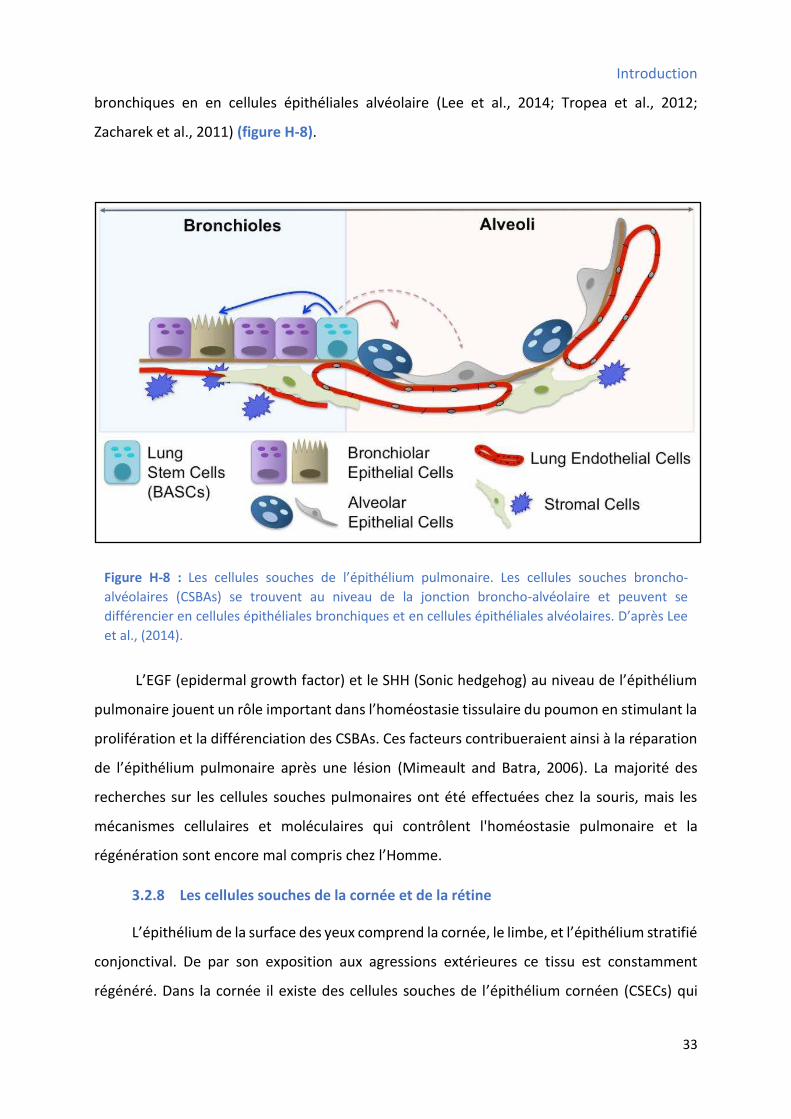
Introduction
33
bronchiques en en cellules épithéliales alvéolaire (Lee et al., 2014; Tropea et al., 2012;
Zacharek et al., 2011) (figure H-8).
L’EGF epide al g o th fa to et le “HH “o i hedgehog au i eau de l’ pith liu
pul o ai e joue t u ôle i po ta t da s l’ho ostasie tissulai e du pou o e sti ula t la
prolifération et la différenciation des CSBAs. Ces facteurs contribueraient ainsi à la réparation
de l’ pith liu pul o ai e ap s u e l sio (Mimeault and Batra, 2006). La majorité des
recherches sur les cellules souches pulmonaires ont été effectuées chez la souris, mais les
mécanismes cellulaires et moléculaires qui contrôlent l'homéostasie pulmonaire et la
g atio so t e o e al o p is hez l’Ho e.
3.2.8 Les cellules souches de la cornée et de la rétine
L’ pith liu de la su fa e des eu o p e d la o e, le li e, et l’ pith liu st atifi
conjonctival. De par son exposition aux agressions extérieures ce tissu est constamment
régénéré. Dans la cornée il existe des cellules souches de l’ pith liu o e C“ECs qui
Figure H-8 : Les ellules sou hes de l’ pith liu pul o ai e. Les ellules souches broncho-
alvéolaires (CSBAs) se trouvent au niveau de la jonction broncho-alvéolaire et peuvent se
différencier en cellules épithéliales bronchiques et en cellules épithéliales alvéolai es. D’ap s Lee et al., (2014).
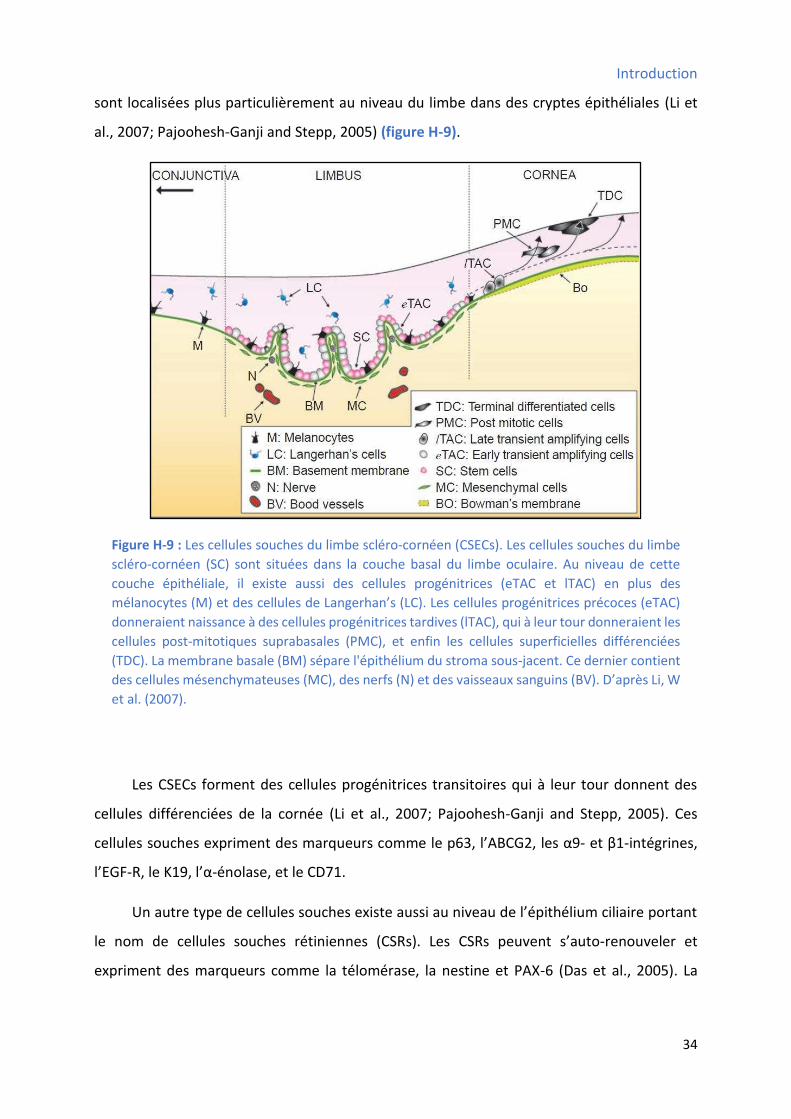
Introduction
34
sont localisées plus particulièrement au niveau du limbe dans des cryptes épithéliales (Li et
al., 2007; Pajoohesh-Ganji and Stepp, 2005) (figure H-9).
Les CSECs forment des cellules progénitrices transitoires qui à leur tour donnent des
cellules différenciées de la cornée (Li et al., 2007; Pajoohesh-Ganji and Stepp, 2005). Ces
cellules souches expriment des marqueurs co e le p , l’ABCG , les α - et β -intégrines,
l’EGF-R, le K , l’α-énolase, et le CD71.
U aut e t pe de ellules sou hes e iste aussi au i eau de l’ pith liu iliai e po ta t
le nom de cellules souches rétiniennes (CSRs). Les CSRs peu e t s’auto-renouveler et
expriment des marqueurs comme la télomérase, la nestine et PAX-6 (Das et al., 2005). La
Figure H-9 : Les cellules souches du limbe scléro-cornéen (CSECs). Les cellules souches du limbe
scléro-cornéen (SC) sont situées dans la couche basal du limbe oculaire. Au niveau de cette
couche épithéliale, il existe aussi des cellules progénitrices (eTAC et lTAC) en plus des
mélanocytes (M) et des ellules de La ge ha ’s LC . Les ellules p og it i es p o es eTAC donneraient naissance à des cellules progénitrices tardives (lTAC), qui à leur tour donneraient les
cellules post-mitotiques suprabasales (PMC), et enfin les cellules superficielles différenciées
(TDC). La membrane basale (BM) sépare l'épithélium du stroma sous-jacent. Ce dernier contient
des cellules mésenchymateuses (MC), des nerfs (N) et des aisseau sa gui s BV . D’ap s Li, W et al. (2007).

Introduction
35
prolifération et la différenciation des CSRs sont régulées par de nombreux facteurs et voies de
signalisation (Das et al., 2005).
3.2.9 Les cellules souches/progénitrices hépatiques
Dans des conditions physiopathologiques, par exemple une infection ou un
traumatisme, le foie peut initier un processus de régénération. Le renouvellement hépatique
est assuré par les hépatocytes et les cholangiocytes ou cellules des canaux biliaires qui sont
des cellules inter-mitotiques pouvant entrer en division après stimulation. Ces deux types
cellulaires assurent seulement une régénération à petite échelle lors de lésions mineures.
Des recherches ont révélé la présence de niches pour des cellules souches au sein des
canaux de Hering (canaux biliaires dans la région péri-portale). Ces cellules progénitrices
hépatiques (CPHs) présentent un phénotype intermédiaire entre les hépatocytes péri-portaux
et les cellules des canaux biliaires (Tarnowski and Sieron, 2006). Les CPHs assurent une
régénération à grande échelle et se différencient en hépatocytes et en cellules des canaux
biliaires (Van Haele and Roskams, 2017) (figure H-10). L’o igi e des CPHs serait soit les
hépatoblastes (eux-mêmes issus de cellules précurseurs endodermiques) ; soit des cellules
précurseurs endodermiques (Walkup and Gerber, 2006). Les CPHs expriment de manière
spécifique le OV-6 (ovalbumine-6) (Tarnowski and Sieron, 2006) mais expriment aussi des
marqueurs communs avec les hépatocytes immatures et les cellules des canaux biliaires
o e le CK , l’α-f top ot i e αFP , la -glutamyl-t a sf ase, l’al u i e ALB , le CK ,
et l’OC . Les CPHs sont aussi positives pour des marqueurs hématopoïétiques comme CD34,
THY-1, FLT-3 et KIT. Plusieurs facteurs (HGF, EGF, TGF-α, VEGF, SCF, TGF-β, et “DF-1) et voies
de sig alisatio s WNT/β-caténine, Notch) sont impliqués dans la régulation de la prolifération
et de la différenciation de ces cellules pendant la régénération hépatique. Des études ont
suggérés une source extra-hépatique des CPHs que serait la moelle osseuse (Tarnowski and
Sieron, 2006).

Introduction
36
3.2.10 Les cellules souches neurales (CSNs) du système nerveux central
Depuis très longtemps, le cerveau était considéré comme un organe dépourvu de
régénération avec un stock de neurones qui diminue tout au long de la vie. Mais des études
sur les rongeurs dans les années 196 o t l u’il e istait u ph o e de eu oge se
dans certaines zones du cerveau adulte de mammifères (Altman, 1963, 1969; Altman and Das,
1965). Plus tard entre 1970 et 1980, cette neurogenèse a été confirmée chez des rongeurs
Figure H-10 : Les cellules progénitrices hépatiques (CPHs). Les CPHs quiescentes (K19, K7, NCAM,
CD133+) prolifèrent après activation et donnent des CPHs (K19, K7, NCAM, CD133+, EpCAM). Ces
dernières peuvent se différencier à la fois en hépatocytes et en cholangiocytes, lors de
l'embryogenèse et suite à une lésion hépatique. Après une blessure, la différenciation des CPHs
dépend du site de la lésion. Après une perte d'hépatocytes, les CPHs a ti es s’e gage t da s u lignage hépatocytaire et donnent naissance à des CPHs (K19-, K7+, NCAM-, CD133-, EpCAM+) suite à
l’i hi itio de la oie de sig alisatio Not h. Ces dernières se différencient en hépatocytes matures
(K19-, K7-, NCAM-, CD133-, EpCAM-). Après la perte des voies biliaires, les CPHs a ti es s’e gage t dans un lignage cholangiocytaire et donnent des CPHs (K19+, K7+, NCAM+, CD133-, EpCAM+) via
l'activation de la voie Notch. Ces CPHs engagées se différencient après en cholangiocytes matures
(K19+, K7+, NCAM-, CD133-, EpCAM+). X: inhibition, ++: activation, numb: inhibiteur de la voie Notch.
D’ap s Va Haele, M., et Roska s, T. .

Introduction
37
da s l’hippo a pe et da s la zo e sous-ventriculaire près du ventricule latéral (Bayer, 1982;
Kaplan and Hinds, 1977).
Il a fallu attendre les années 1990 pour démontrer que la neurogenèse se produisait
gale e t da s le e eau des p i ates et de l’Ho e (Kuhn et al., 1996; Luskin, 1993; Seki
and Arai, 1993). La neurogenèse commence précisément dans deux zones du cerveau qui sont
le g us de t de tal g us ou DG de l’hippo a pe et la zo e sous-ventriculaire
(subventricular zone ou SVZ) (Taupin, 2006). Dans le DG, les nouvelles cellules neuronales sont
obtenues au niveau de la zone sous-granulaire (subgranular zone ou SGZ) puis elles migrent
da s la ou he des ellules g a ulai es d’où elles se p ojette t da s l’ai e CA de la o e
d’A o (Bond et al., 2015) (figure H-11 A).
Da s la “V), les ou elles ellules eu o ales o te ues ig e t jus u’au ul e olfa tif
(olfactory bulb ou OB), à travers la voie de migration rostrale (rostro-migratory stream ou
RMS), où elles se différencient en interneurones. Dans le cerveau adulte, la majorité de la
neurogenèse se fait dans la SVZ et la SGZ (LI et XIE, 2005; TARNOWSKI et SIERON, 2006). Une
eu oge se plus « dis te » a t appo t e da s d’aut es zo es du e eau o e le o ps
st i et le e e t i ule et l’ai e CA , hez les o geu s, ai si ue dans le néocortex chez
les primates non humains (Taupin, 2006).
Les CSNs so t dot es de apa it d’auto-renouvellement et elles sont considérées
comme des cellules souches multipotentes donnant naissance aux neurones, aux astrocytes
et aux oligodendrocytes (Taupin, 2006) (figure H-11 B). Les CSNs sont aussi les précurseurs
des cellules progénitrices neurales (neural progenitor cells ou NPCs) et gliales (glial progenitor
cells ou GPCs). Ces deux types de cellules progénitrices sont des cellules indifférenciées et
comparées aux CSNs, elles possèdent une capacité de prolifération limitée et ne peuvent pas
s’auto-renouveler (Taupin, 2006).
L’isole e t et la a a t isatio in vitro des CSNs ont commencé en 1992 avec les
travaux de REYNOLD et WEISS sur la souris. Ces travaux ont montré que les CSNs isolées à
partir de la SVZ pouvaient se différencier en neurones, en astrocytes et en oligodendrocytes.
Ces cellules avaient la particularité de se développer en neurosphères in vitro en présence de
l’EGF.
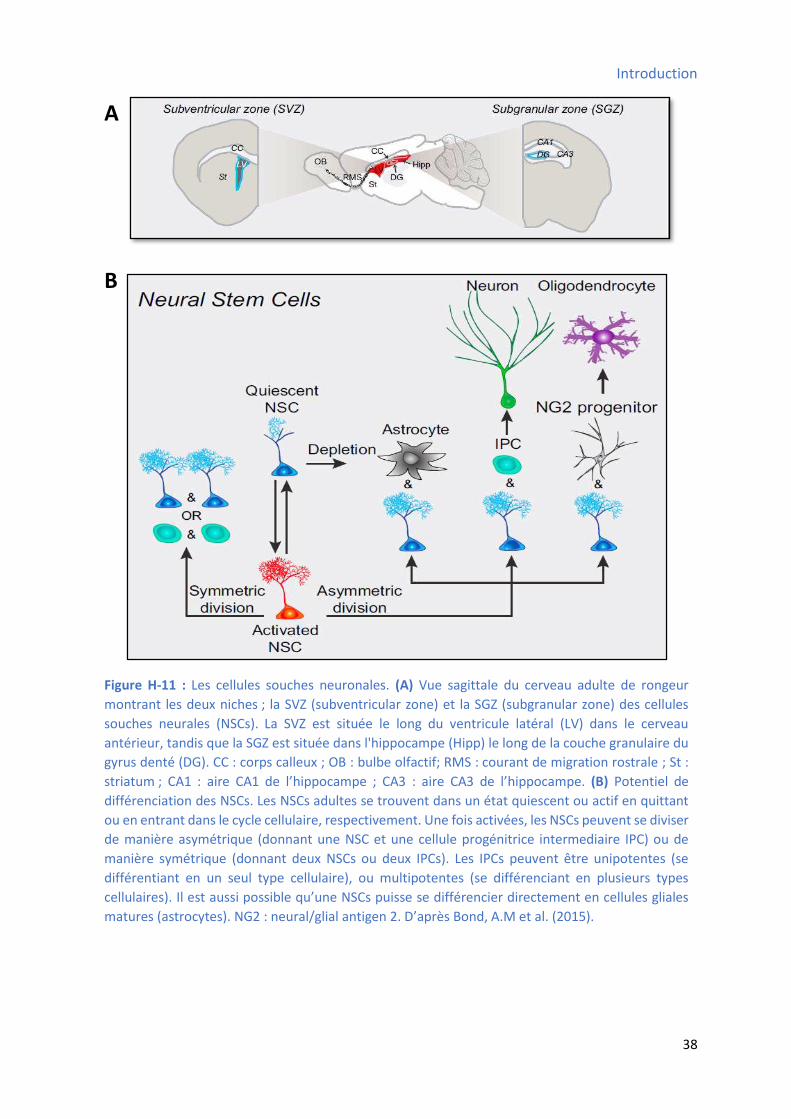
Introduction
38
A
B
Figure H-11 : Les cellules souches neuronales. (A) Vue sagittale du cerveau adulte de rongeur
montrant les deux niches ; la SVZ (subventricular zone) et la SGZ (subgranular zone) des cellules
souches neurales (NSCs). La SVZ est située le long du ventricule latéral (LV) dans le cerveau
antérieur, tandis que la SGZ est située dans l'hippocampe (Hipp) le long de la couche granulaire du
gyrus denté (DG). CC : corps calleux ; OB : bulbe olfactif; RMS : courant de migration rostrale ; St :
striatum ; CA1 : ai e CA de l’hippo a pe ; CA : ai e CA de l’hippo a pe. (B) Potentiel de
différenciation des NSCs. Les NSCs adultes se trouvent dans un état quiescent ou actif en quittant
ou en entrant dans le cycle cellulaire, respectivement. Une fois activées, les NSCs peuvent se diviser
de manière asymétrique (donnant une NSC et une cellule progénitrice intermediaire IPC) ou de
manière symétrique (donnant deux NSCs ou deux IPCs). Les IPCs peuvent être unipotentes (se
différentiant en un seul type cellulaire), ou multipotentes (se différenciant en plusieurs types
cellulaires . Il est aussi possi le u’u e N“Cs puisse se diff e ie di e te e t e ellules gliales matures (astrocytes). NG2 : eu al/glial a tige . D’ap s Bo d, A.M et al. (2015).

Introduction
39
La nestine marque les cellules souches neuro-épithéliales et les cellules du système nerveux
central durant le développement embryonnaire. En plus de la nestine, les cellules souches
fo a t es eu osph es e p i e t aussi d’aut es a ueu s o e LEX/““EA-1, AC133 et
NG2 (Tarnowski and Sieron, 2006). D’aut es uipes o t pu isole des NSCs à partir du cerveau
adulte et de la oelle pi i e hez de o euses esp es o p is l’Ho e (Li and Xie,
2005; Taupin, 2006). La neurogenèse du cerveau adulte est modulée par de nombreux stimuli
environnementaux et selon certaines conditions physiopathologiques (Taupin, 2006). Chez le
o geu , l’e e i e aug e te ait la eu oge se da s l’hippo a pe alo s ue l’a a e e t
da s l’âge la di i ue ait. L’isole e t so ial, l’al oolis e, le st ess et le a ue de so eil
diminuent la neurogenèse. La neurogenèse adulte survient dans les processus physiologiques
o e la oi e et d’app e tissage ais aussi da s des conditions pathologiques, comme
les pathologies neurologiques et les traumatismes du cerveau (Taupin, 2006).
3.2.11 Les cellules souches musculaires squelettiques
Le muscle strié squelettique renferme des cellules souches bien caractérisées qui
assurent son renouvellement et sa réparation lors des lésions. Les cellules souches
myogéniques, nommées cellules satellites (satellite cells ou SCs), se localisent entre deux
couches de lame basale et expriment des marqueurs comme N-CAM, CD56, la M-cadhérine,
PAX-7, MYO-D et MYF- . L’e p essio e es a ueu s a ie selo le stade de différenciation
(McCullagh and Perlingeiro, 2015; Zammit et al., 2006) (figure H-12). Les SCs se trouvent dans
u tat uies e t ais suite à leu a ti atio pa u sti ulus lessu e, t au atis e, …) ces
cellules prolifèrent, fusionnent et se différencient en myofibres (Christov et al., 2007; Hawke
and Garry, 2001). La différenciation des SCs en myofibres s’a o pag e de ha ge e ts
toplas i ues i po ta ts o e la du tio de l’h t o h o ati e, aug e tatio du
atio toplas e/ o au et aug e tatio du o e d’o ga elles i t a ellulai es (Hawke and
Garry, 2001; Tarnowski and Sieron, 2006).
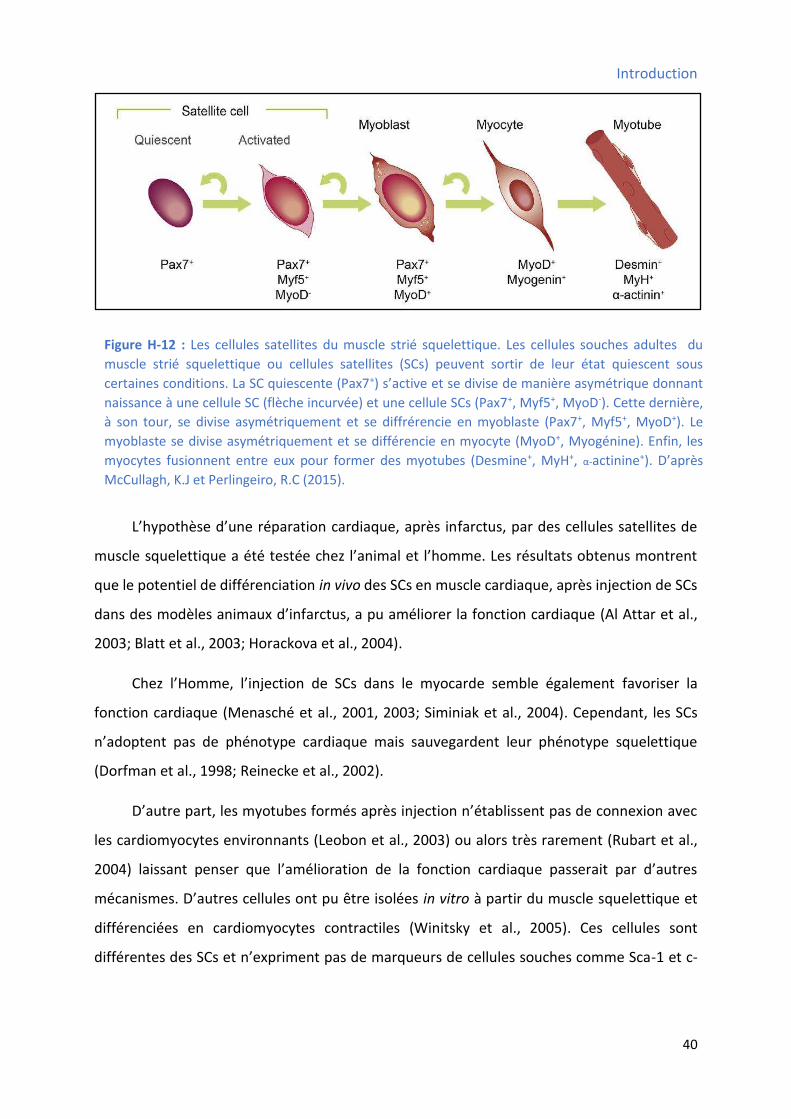
Introduction
40
L’h poth se d’u e pa atio a dia ue, ap s i farctus, par des cellules satellites de
us le s ueletti ue a t test e hez l’a i al et l’ho e. Les sultats o te us o t e t
que le potentiel de différenciation in vivo des SCs en muscle cardiaque, après injection de SCs
da s des od les a i au d’i farctus, a pu améliorer la fonction cardiaque (Al Attar et al.,
2003; Blatt et al., 2003; Horackova et al., 2004).
Chez l’Ho e, l’i je tio de “Cs da s le o a de se le gale e t favoriser la
fonction cardiaque (Menasché et al., 2001, 2003; Siminiak et al., 2004). Cependant, les SCs
’adopte t pas de ph ot pe ardiaque mais sauvegardent leur phénotype squelettique
(Dorfman et al., 1998; Reinecke et al., 2002).
D’aut e pa t, les otu es fo s ap s i je tio ’ ta lisse t pas de o e io a e
les cardiomyocytes environnants (Leobon et al., 2003) ou alors très rarement (Rubart et al.,
2004) laissa t pe se ue l’a lio atio de la fo tio a dia ue passe ait pa d’aut es
a is es. D’aut es ellules o t pu t e isol es in vitro à partir du muscle squelettique et
différenciées en cardiomyocytes contractiles (Winitsky et al., 2005). Ces cellules sont
différentes des SCs et ’e p i e t pas de a ueu s de ellules sou hes o e “ a-1 et c-
Figure H-12 : Les cellules satellites du muscle strié squelettique. Les cellules souches adultes du
muscle strié squelettique ou cellules satellites (SCs) peuvent sortir de leur état quiescent sous
certaines conditions. La SC quiescente (Pax7+ s’a ti e et se di ise de manière asymétrique donnant
naissance à une cellule SC (flèche incurvée) et une cellule SCs (Pax7+, Myf5+, MyoD-). Cette dernière,
à son tour, se divise asymétriquement et se diffrérencie en myoblaste (Pax7+, Myf5+, MyoD+). Le
myoblaste se divise asymétriquement et se différencie en myocyte (MyoD+, Myogénine). Enfin, les
myocytes fusionnent entre eux pour former des myotubes (Desmine+, MyH+, α-actinine+ . D’ap s McCullagh, K.J et Perlingeiro, R.C (2015).

Introduction
41
kit. Cette nouvelle population de cellules souche contractile pourrait, selon les auteurs,
représenter une alternative intéressante.
3.2.12 Les cellules souches cardiaques
Depuis lo gte ps, le œur des mammifères a été considéré comme un organe post-
mitotique dépourvu de tout pouvoir de régénération. Ce dogme a ainsi limité la recherche
fondamentale et clinique en cardiologie depuis plusieurs décennies. On pensait alors que suite
aux lésions cardiaques, les cardiomyocytes subissaient des hypertrophies cellulaires et ne
pouvaient pas être remplacés. Une autre hypothèse propose que pendant la vie
embryonnaire, les CSEs a dia ues s’e gage t da s u ph ot pe a dia ue et pe de t
définitivement leurs p op i t s de ellules sou hes da s le œu adulte (Anversa et al., 2006).
Il est vrai que, suite à un infarctus du myocarde, les cardiomyocytes nécrosés sont remplacés
par un tissu cicatriciel fibrotique. Cependant, quelques recherches ont pu montrer que les
a dio o tes peu e t se di ise de a i e peu f ue te da s le œur adulte (Beltrami
et al., 2001; Kajstura et al., 1998; Quaini et al., 2002; Reiss et al., 1994). Cette découverte de
divisions mitotiques cardiomyocytaires a cassé le dogme selon lequel le nombre de
cardiomyocytes était défini à la naissance et a éveillé la curiosité des chercheurs sur le fait que
ces nouveaux cardiomyocytes dériveraient de cellules souches cardiaques (figure H-13). En
effet, plusieu s uipes o t is e ide e l’e iste e de ellules sou hes a dia ues
(Cardiac Stem Cells ou CSCs) capables de se différencier en myocytes, cellules musculaires
lisses et en cellules endothéliales (Beltrami et al., 2003; Linke et al., 2005; Oh et al., 2003;
Urbanek et al., 2005). Cette découverte importante a donné de nouvelles perspectives
o e a t la iologie du œu et les mécanismes de son homéostasie tissulaire et de sa
régénération. Grâce à ces travaux, le concept de CSCs est né et le tissu cardiaque est désormais
d fi i o e u tissu apa le d’auto-renouvellement et les myocytes sont régénérés durant
toute la vie de l’o ga is e (Bergmann et al., 2015; Lázár et al., 2017). De même, il a été
montré que les C“Cs e s’e gage t pas i e si le e t e se diff e ia t du a t la ie
embryonnaire mais elles restent après la naissance apa les d’auto-renouvellement.
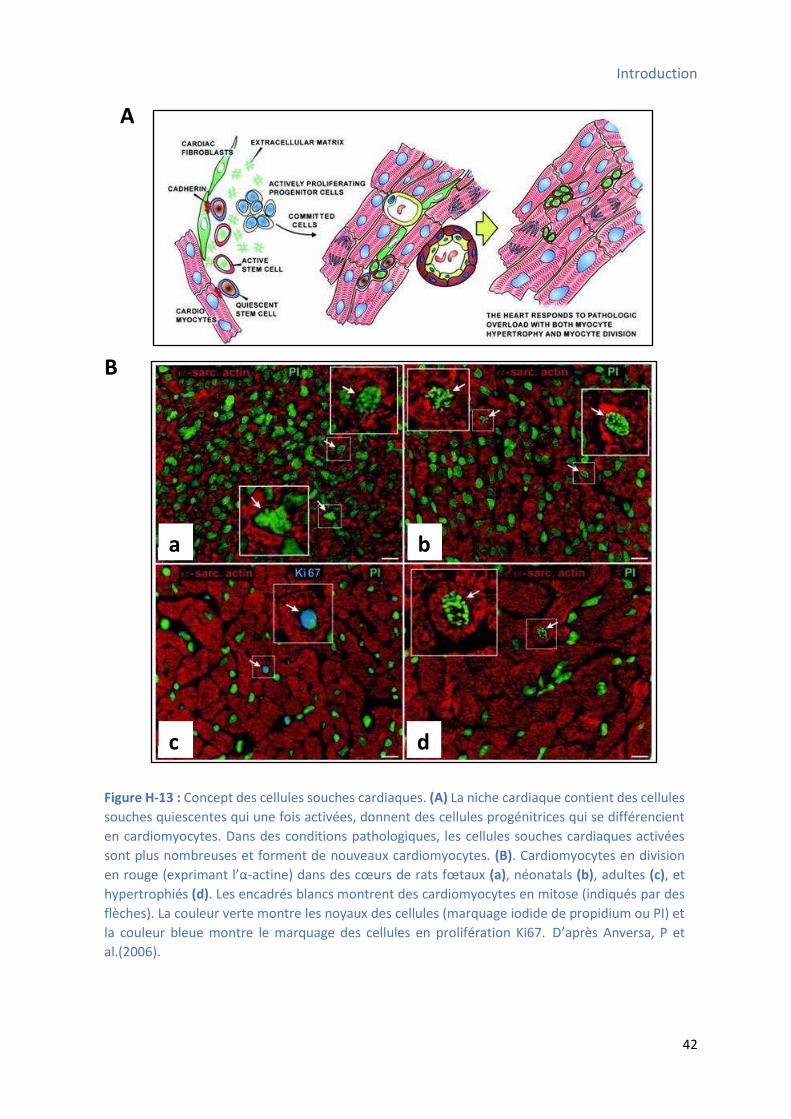
Introduction
42
B
b
d
a
c
Figure H-13 : Concept des cellules souches cardiaques. (A) La niche cardiaque contient des cellules
souches quiescentes qui une fois activées, donnent des cellules progénitrices qui se différencient
en cardiomyocytes. Dans des conditions pathologiques, les cellules souches cardiaques activées
sont plus nombreuses et forment de nouveaux cardiomyocytes. (B). Cardiomyocytes en division
e ouge e p i a t l’α-a ti e da s des œu s de ats fœtau (a), néonatals (b), adultes (c), et
hypertrophiés (d). Les encadrés blancs montrent des cardiomyocytes en mitose (indiqués par des
flèches). La couleur verte montre les noyaux des cellules (marquage iodide de propidium ou PI) et
la couleur bleue montre le marquage des cellules en prolifération Ki67. D’ap s A e sa, P et al.(2006).
A

Introduction
43
Suite à cette nouvelle découverte, plusieurs groupes de chercheurs ont pu isoler des
C“Cs à pa ti de œu s adultes e se asa t su l’e p essio de e tai s arqueurs de surface
déjà utilisés pou isole des ellules sou hes à pa ti d’aut es o ga es.
Chapitre II : Cellules souches et progéniteurs cardiaques
1 Nature des cellules souches cardiaques
Le ph ot pe des C“Cs ’est pas e o e ie d fi i. La o e latu e est i p ise et
plusieurs types de cellules cardiaques adultes ont été caractérisés et proposés comme des
cellules souches cardiaques (Iancu et al., 2015). Le nombre des CSCs chez différentes espèces,
o p is l’Ho e, est estimé à environ 1 CSC pour 8000 à 20000 myocytes (Anversa et al.,
2006). Un modèle de classement par hiérarchie de différenciation des CSCs a été proposé
(figure H-14). Selon ce modèle, une CSC originelle se divise de manière asymétrique pour
donner naissance à une cellule souche fille et une autre cellule fille nommée cellule
progénitrice (CPg). Cette CPg à son tour donne une variété de cellules progénitrices plus
sp ialis es ; les p u seu s de o tes MPg , u e ellule p og it i e d’e doth liu EPg
et enfin, une cellule progénitrice de muscle lisse (SMPg). Ces cellules donnent ensuite un
précurseur ; la MPg ui est à l’o igi e d’u e ellule p u seu de o tes MP , la EPg u e
ellule p u seu d’e doth liu EP et la “MPg u e ellule p u seu de us le lisse “MP .
Enfin, les précurseurs deviennent des « cellules amplificatrices transitoires » (transient
amplifying cells) et se différencient en myocytes matures des cellules endothéliales ou des
cellules du muscle lisse des vaisseaux sanguins. Les CSC dans ce modèle sont des cellules
lignage-négatives qui expriment uniquement c-kit, MDR1 ou Sca-1. Les progéniteurs
expriment des antigènes de cellules souches et des facteurs de transcription de cellules
cardiaques mais ne présentent pas de protéines cytoplasmiques spécifiques.
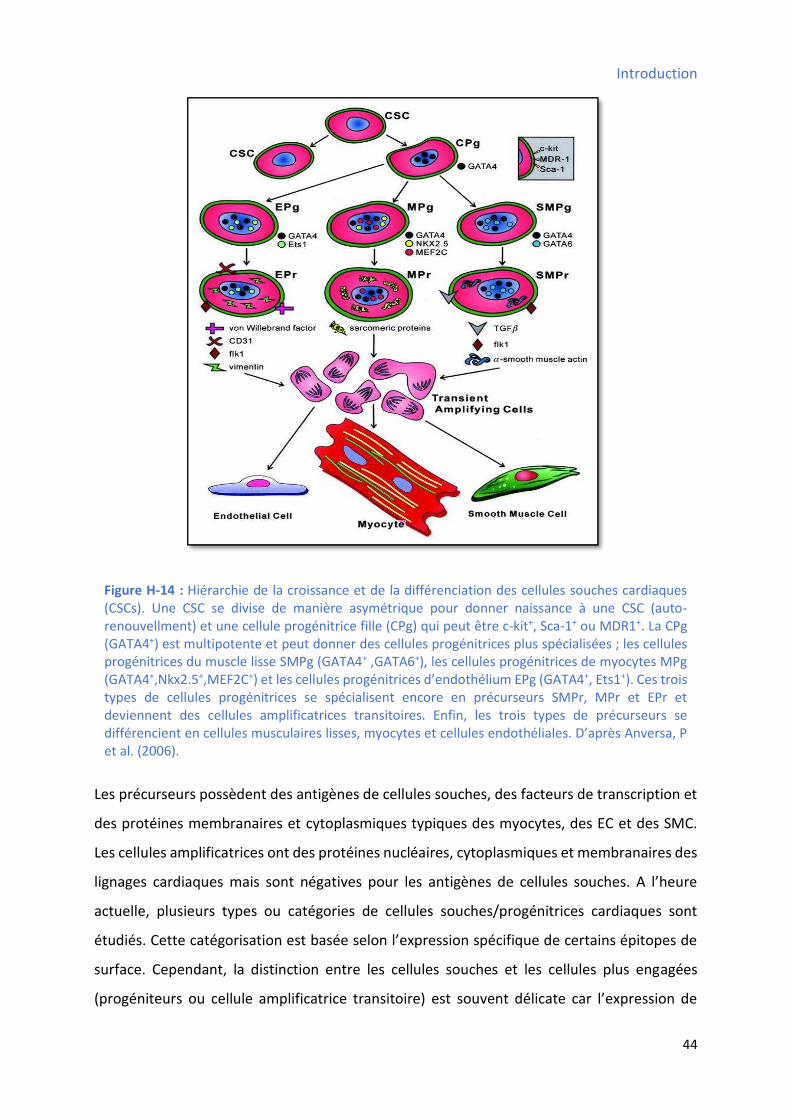
Introduction
44
Les précurseurs possèdent des antigènes de cellules souches, des facteurs de transcription et
des protéines membranaires et cytoplasmiques typiques des myocytes, des EC et des SMC.
Les cellules amplificatrices ont des protéines nucléaires, cytoplasmiques et membranaires des
lig ages a dia ues ais so t gati es pou les a tig es de ellules sou hes. A l’heu e
actuelle, plusieurs types ou catégories de cellules souches/progénitrices cardiaques sont
tudi s. Cette at go isatio est as e selo l’e p essio sp ifi ue de e tai s pitopes de
surface. Cependant, la distinction entre les cellules souches et les cellules plus engagées
p og iteu s ou ellule a plifi at i e t a sitoi e est sou e t d li ate a l’e p essio de
Figure H-14 : Hiérarchie de la croissance et de la différenciation des cellules souches cardiaques (CSCs). Une CSC se divise de manière asymétrique pour donner naissance à une CSC (auto-renouvellment) et une cellule progénitrice fille (CPg) qui peut être c-kit+, Sca-1+ ou MDR1+. La CPg (GATA4+) est multipotente et peut donner des cellules progénitrices plus spécialisées ; les cellules progénitrices du muscle lisse SMPg (GATA4+ ,GATA6+), les cellules progénitrices de myocytes MPg (GATA4+,Nkx2.5+,MEF2C+ et les ellules p og it i es d’e doth liu EPg GATA +, Ets1+). Ces trois types de cellules progénitrices se spécialisent encore en précurseurs SMPr, MPr et EPr et deviennent des cellules amplificatrices transitoires. Enfin, les trois types de précurseurs se diff e ie t e ellules us ulai es lisses, o tes et ellules e doth liales. D’ap s A e sa, P et al. (2006).
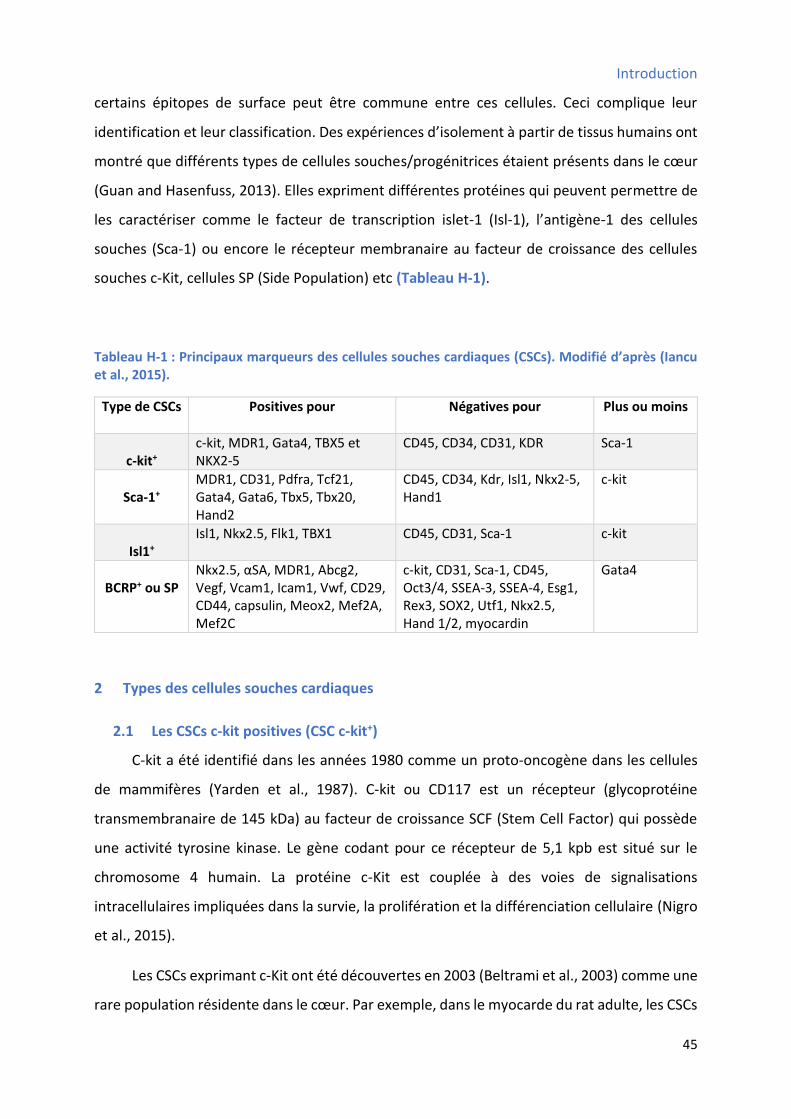
Introduction
45
certains épitopes de surface peut être commune entre ces cellules. Ceci complique leur
ide tifi atio et leu lassifi atio . Des e p ie es d’isole e t à partir de tissus humains ont
montré que différents types de cellules souches/progénitrices étaient présents da s le œu
(Guan and Hasenfuss, 2013). Elles expriment différentes protéines qui peuvent permettre de
les caractériser comme le facteur de transcription islet-1 (Isl- , l’a tig e-1 des cellules
souches (Sca-1) ou encore le récepteur membranaire au facteur de croissance des cellules
souches c-Kit, cellules SP (Side Population) etc (Tableau H-1).
Tableau H-1 : P i ipau a ueu s des ellules sou hes a dia ues CSCs . Modifi d’ap s Ia u et al., 2015).
Type de CSCs
Positives pour Négatives pour Plus ou moins
c-kit+
c-kit, MDR1, Gata4, TBX5 et NKX2-5
CD45, CD34, CD31, KDR Sca-1
Sca-1+
MDR1, CD31, Pdfra, Tcf21, Gata4, Gata6, Tbx5, Tbx20, Hand2
CD45, CD34, Kdr, Isl1, Nkx2-5, Hand1
c-kit
Isl1+
Isl1, Nkx2.5, Flk1, TBX1 CD45, CD31, Sca-1 c-kit
BCRP+ ou SP
Nk . , α“A, MDR , A g , Vegf, Vcam1, Icam1, Vwf, CD29, CD44, capsulin, Meox2, Mef2A, Mef2C
c-kit, CD31, Sca-1, CD45, Oct3/4, SSEA-3, SSEA-4, Esg1, Rex3, SOX2, Utf1, Nkx2.5, Hand 1/2, myocardin
Gata4
2 Types des cellules souches cardiaques
2.1 Les CSCs c-kit positives (CSC c-kit+)
C-kit a été identifié dans les années 1980 comme un proto-oncogène dans les cellules
de mammifères (Yarden et al., 1987). C-kit ou CD117 est un récepteur (glycoprotéine
transmembranaire de 145 kDa) au facteur de croissance SCF (Stem Cell Factor) qui possède
une activité tyrosine kinase. Le gène codant pour ce récepteur de 5,1 kpb est situé sur le
chromosome 4 humain. La protéine c-Kit est couplée à des voies de signalisations
intracellulaires impliquées dans la survie, la prolifération et la différenciation cellulaire (Nigro
et al., 2015).
Les CSCs exprimant c-Kit ont été découvertes en 2003 (Beltrami et al., 2003) comme une
a e populatio side te da s le œu . Pa e e ple, da s le o a de du at adulte, les C“Cs
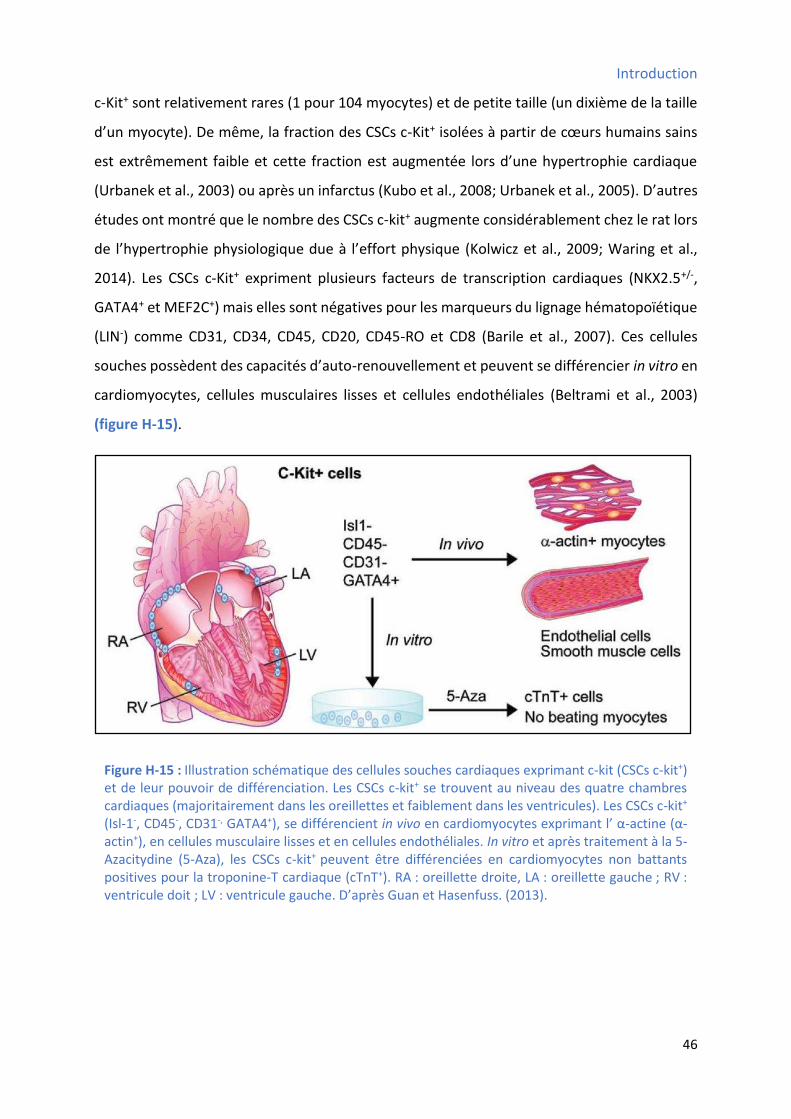
Introduction
46
c-Kit+ sont relativement rares (1 pour 104 myocytes) et de petite taille (un dixième de la taille
d’u o te . De e, la f a tio des C“Cs -Kit+ isol es à pa ti de œu s hu ai s sai s
est e t e e t fai le et ette f a tio est aug e t e lo s d’u e h pe t ophie a dia ue
(Urbanek et al., 2003) ou après un infarctus (Kubo et al., 2008; Urbanek et al., 2005). D’aut es
études ont montré que le nombre des CSCs c-kit+ augmente considérablement chez le rat lors
de l’h pe t ophie ph siologi ue due à l’effo t ph si ue (Kolwicz et al., 2009; Waring et al.,
2014). Les CSCs c-Kit+ expriment plusieurs facteurs de transcription cardiaques (NKX2.5+/-,
GATA4+ et MEF2C+) mais elles sont négatives pour les marqueurs du lignage hématopoïétique
(LIN-) comme CD31, CD34, CD45, CD20, CD45-RO et CD8 (Barile et al., 2007). Ces cellules
sou hes poss de t des apa it s d’auto-renouvellement et peuvent se différencier in vitro en
cardiomyocytes, cellules musculaires lisses et cellules endothéliales (Beltrami et al., 2003)
(figure H-15).
Figure H-15 : Illustration schématique des cellules souches cardiaques exprimant c-kit (CSCs c-kit+) et de leur pouvoir de différenciation. Les CSCs c-kit+ se trouvent au niveau des quatre chambres cardiaques (majoritairement dans les oreillettes et faiblement dans les ventricules). Les CSCs c-kit+ (Isl-1-, CD45-, CD31-, GATA4+), se différencient in vivo e a dio o tes e p i a t l’ α-a ti e α-actin+), en cellules musculaire lisses et en cellules endothéliales. In vitro et après traitement à la 5-Azacitydine (5-Aza), les CSCs c-kit+ peuvent être différenciées en cardiomyocytes non battants positives pour la troponine-T cardiaque (cTnT+). RA : oreillette droite, LA : oreillette gauche ; RV : ventricule doit ; LV : e t i ule gau he. D’ap s Gua et Hase fuss. .

Introduction
47
Les cellules CSCs c-kit+/Lin- ep se te t l’u e des p i ipales populatio s de ellules sou hes
t ou es da s le œu (Bollini et al., 2011). Ces cellules se localisent préférentiellement dans
l’o eillette d oite du œu hez l’adulte et hez l’e fa t (Itzhaki-Alfia et al., 2009; Mishra et al.,
2011). Cependant, l’e p essio de -kit ’est pas se seule e t au C“Cs a ette
protéine est aussi exprimée par les mastocytes (Kubo et al., 2008), les cellules endothéliales
(Castaldo et al., 2008), des cellules souches de la moelle osseuse (Chong et al., 2011), les
cellules dendritiques et mélanocytes (Fang et al., 2012), les cardiomyocytes postnatals (Li et
al., 2008) etc. A cause de cette expression non spécifique, les CSCs c-Kit+ représentent une
population hétérogène. E effet, l’a al se t a s ipto i ue au ou s de la diff e iatio
cardiaque des cellules c-Kit+ a l l’e iste e de plusieu s lasses de C“C -kit+; les cellules
c-Kit+KDR+ ou VPC (vascular progenitor cells) , les cellules c-Kit+KDR- ou MPC (myocyte
progenitor cells) et les c-kit+KDR+CD31+ (se différenciant en cellules endothéliales) (Bearzi et
al., 2009; Fang et al., 2012; Tallini et al., 2009). Une autre étude a montré que les CSC c-kit+
expriment des marqueurs mésenchymateux comme le CD105 et le CD29 (Gambini et al., 2011)
et peuvent se différencier en adipocytes et en ostéocytes (Itzhaki-Alfia et al., 2009). Il est
probable donc que les CSCs c-kit+ isolées in vitro soient contaminées par d'autres types
cellulaires (fibroblastes cardiaques, les mastocytes ou les cellules de lignée hématopoïétique).
Les données sur l'expression des marqueurs de lignage sont encore plus conflictuelles. Par
exemple, des études ont montré que les CSC c-kit+ n'expriment pas tous les marqueurs de
lignage cardiaques tels que GATA4, NKX2.5, MEF2C (Bearzi et al., 2007; Boni et al., 2008;
D’A a io et al., . De plus, da s les œu s des ou eau-nés et des jeunes enfants, le
pourcentage de cellules c-kit+ variait de 5% à 9% et seulement 1% à 3% des cellules c-kit+
étaient positives pour Nkx2.5+ (Mishra et al., 2011). De toute évidence, de nombreux aspects
de ces cellules restent à comprendre.
In vitro, les CSC c-kit+ ont des propriétés typiques de cellules souches et certaines études
in vivo ont montré que la transplantation de ces cellules améliore la fonction cardiaque chez
des od les a i au d’i fa tus du o a de (Ellison et al., 2013). Chez l’Ho e, l’i je tio
intra-coronarienne de CSCs c-Kit+ p e ie essai li i ue su l’Ho e de phase I a a lio
la fo tio s stoli ue du e t i ule gau he et a duit la taille de l’i farctus chez les patients
souffra t d’i suffisa e a dia ue (Bolli et al., 2011; Chugh et al., 2012). Une étude a
également montré que la perfusion intra-coronarienne de 20 millions de CSCs c-kit+ dans des

Introduction
48
œu s des po s e p o o ue au u do age al, h pati ue ou o a dia ue (Keith et
al., 2015). Néanmoins, la rétention de ces cellules dans le myocarde reste faible malgré le
nombre élevé injecté.
2.2 Les CSCs Sca1 positives (CSC Sca1+)
L'antigène de cellules souches-1 (Sca-1 ou Ly6A) est une protéine à ancre GPI
(glycerophosphatidylinositol) de 18 KDa qui appartient à la famille des gènes Ly6 (lymphocyte
activation protein-6). La famille des gènes Ly6 contient au moins 18 gènes liés étroitement sur
le h o oso e et ui pa tage t plus de % d’ho ologie de s ue e a e “ a-1 (LeClair
et al., 1986; Patterson et al., 2000; van de Rijn et al., 1989). Co e d’aut es p ot i es à a e
GPI, les protéines Ly6 ont une position membranaire idéale pour réguler ou co-activer des
voies de signalisation via des liaisons ligand- epteu ou d’aut es i te actions protéine-
protéine. Cependant le mécanisme moléculaire exact par lequel ces protéines agissent reste
non élucidé. Sca-1 a été initialement identifié comme marqueur de cellules souches
hématopoïétiques murines (Chong et al., 2014; Han et al., 1995) mais son expression est
également retrouvée da s d’aut es t pes ellulai es. En effet, plusieurs groupes de chercheurs
ont identifié la protéine Sca-1 chez la souris dans plusieurs tissus comme le système
squelettique (Short et al., 2001), la glande mammaire (Welm et al., 2002), la prostate (Burger
et al., 2005; Xin et al., 2005), le derme (Fernandes et al., 2004; Toma et al., 2001), le muscle
squelettique (Gussoni et al., 1999; Lee et al., 2000), le foie (Petersen et al., 2003; Wulf et al.,
2003), la moelle osseuse (Gojo et al., 2003) et le œu (Oh et al., 2003; Rosenblatt-Velin et al.,
2005). Toutefois, il ’a pas e o e t o t si es populatio s “ a-1 positives sont vraiment
des cellules souches tissu-spécifiques. Plusieurs sous-populations de cellules Sca-1+ ont été
a a t is es da s des od les u i s e se asa t su l’e p essio de a ueu s de su fa e
lignage-spécifique (Valente et al., 2014). Ces sous-populatio s ’e p i e t pas les a ueu s
endothéliaux (CD31) et hématopoïétiques (CD45) nommées Lignage négatif (ou Lin-) et
montrent un profil mésenchymateux (CD34-, CD29+, CD90+, CD105+ et CD44+). Ces cellules
Sca-1+ murines, ont été décrites comme pouvant se différencier en cardiomyocytes, cellules
musculaires lisses et cellules endothéliales. De plus, les CSCs Sca-1+ murines expriment
faiblement les gènes codant pour les facteurs de transcription cardiaques précoces GATA-4 et
NKX2.5 et les protéines sarcomériques (Matsuura et al., 2004).

Introduction
49
Il a été montré que l'injection intra-myocardique du facteur de croissance hépatique (HGF) et
du facteur de croissance lié à l'insuline (IGF) chez des modèles animaux facilite la migration et
la prolifération des cellules Sca-1+ c-kit+, qui semblent se différencier en petits myocytes
fonctionnels (Linke et al., 2005; Urbanek et al., 2005).
De plus, après injection intraveineuse des CSCs Sca-1+ da s des od les d’i fa tus
murins, il a été mis en évidence dans le myocarde atteint, la présence de cellules différenciées
en cardiomyocytes e p i a t l’α-actine sarcomérique et la connexine 43 (Oh et al., 2003).
Bien que l'expression de Sca-1 soit limitée aux souris, une protéine de type Sca-1 a été
d te t e pa i os opie o fo ale et pa Weste lot da s le œu du hie et de l’Ho e
(Leri et al., 2005). Comme Sca-1, cette protéine appartient probablement à la famille des
gènes Ly6. Des cellules semblables à Sca-1+ (Sca-1+-like o t t isol es à pa ti du œu
humain adulte en utilisant des anticorps Sca-1 anti-souris. Ces cellules Sca-1+-like humaines
sont négatives pour le CD45, CD34, CD133 et CD14, et positives pour le CD105, GATA4 et
Nkx2.5 (figure H-16). Les CSCs Sca-1+-like hu ai e fœtales et adultes, so t ultipote tes et
peuvent se différencier en cardiomyocytes fonctionnels, en cellules endothéliales et en
cellules musculaires lisses (van Vliet et al., 2008, 2010). Cepe da t, au u essai li i ue ’a
été rapporté avec cette population cellulaire.
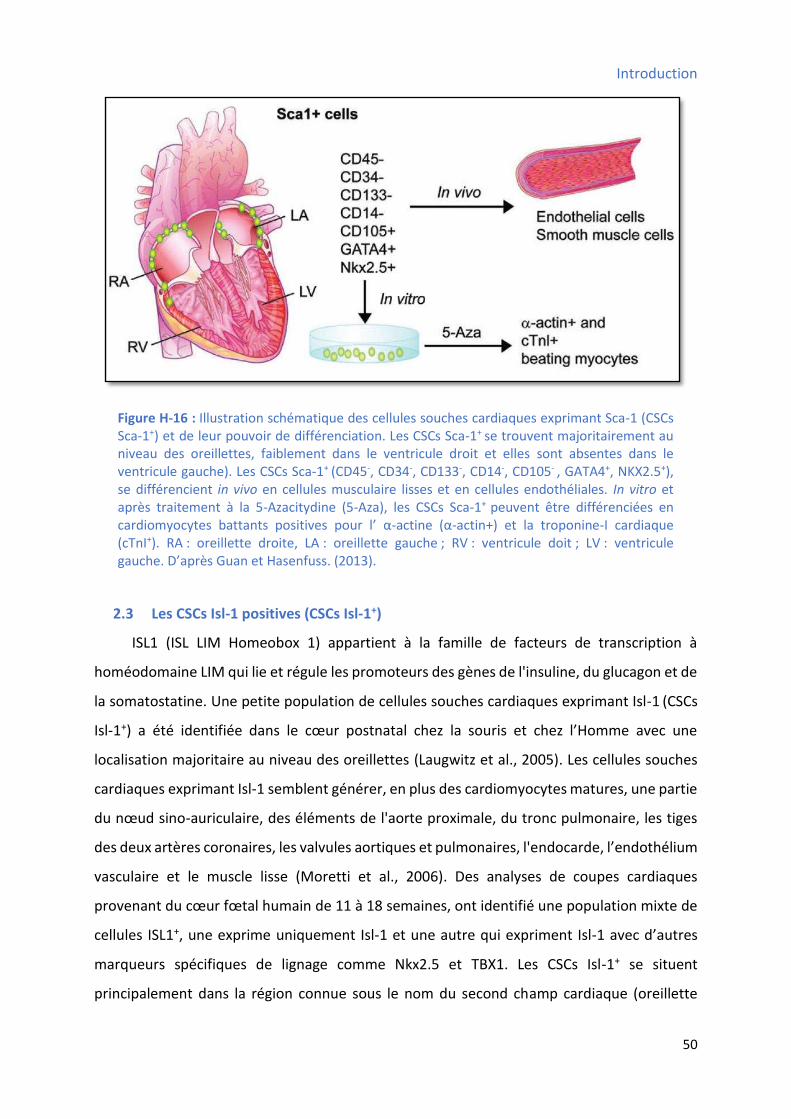
Introduction
50
2.3 Les CSCs Isl-1 positives (CSCs Isl-1+)
ISL1 (ISL LIM Homeobox 1) appartient à la famille de facteurs de transcription à
homéodomaine LIM qui lie et régule les promoteurs des gènes de l'insuline, du glucagon et de
la somatostatine. Une petite population de cellules souches cardiaques exprimant Isl-1 (CSCs
Isl-1+ a t ide tifi e da s le œu post atal hez la sou is et hez l’Ho e a e u e
localisation majoritaire au niveau des oreillettes (Laugwitz et al., 2005). Les cellules souches
cardiaques exprimant Isl-1 semblent générer, en plus des cardiomyocytes matures, une partie
du œud si o-auriculaire, des éléments de l'aorte proximale, du tronc pulmonaire, les tiges
des deux artères coronaires, les valvules aortiques et pulmonaires, l'endocarde, l’e doth liu
vasculaire et le muscle lisse (Moretti et al., 2006). Des analyses de coupes cardiaques
provenant du œu fœtal hu ai de 11 à 18 semaines, ont identifié une population mixte de
cellules ISL1+, une exprime uniquement Isl-1 et une autre qui expriment Isl- a e d’aut es
marqueurs spécifiques de lignage comme Nkx2.5 et TBX1. Les CSCs Isl-1+ se situent
principalement dans la région connue sous le nom du second champ cardiaque (oreillette
Figure H-16 : Illustration schématique des cellules souches cardiaques exprimant Sca-1 (CSCs Sca-1+) et de leur pouvoir de différenciation. Les CSCs Sca-1+ se trouvent majoritairement au niveau des oreillettes, faiblement dans le ventricule droit et elles sont absentes dans le ventricule gauche). Les CSCs Sca-1+ (CD45-, CD34-, CD133-, CD14-, CD105- , GATA4+, NKX2.5+), se différencient in vivo en cellules musculaire lisses et en cellules endothéliales. In vitro et après traitement à la 5-Azacitydine (5-Aza), les CSCs Sca-1+ peuvent être différenciées en a dio o tes atta ts positi es pou l’ α-a ti e α-actin+) et la troponine-I cardiaque
(cTnI+). RA : oreillette droite, LA : oreillette gauche ; RV : ventricule doit ; LV : ventricule gau he. D’ap s Gua et Hasenfuss. (2013).

Introduction
51
droite et le tractus d'éjection) (Bu et al., 2009). Juste après la naissance, des CSCs Isl-1+ ont
été identifiées chez la souris, le rat et l'Homme. In vitro, ces cellules sont capables de se
différencier en cardiomyocytes (Laugwitz et al., 2005). Durant le développement cardiaque,
les CSCs Isl1+ jouent un rôle primordial dans la création du ventricule droit, des oreillettes et
du t a tus d’ je tio (Cai et al., 2003). Ces cellules sont multipotentes et capables de se
différencier en tous les lignages cardiaques (Bu et al., 2009). Cependant, l'expression d'Isl1
dans les progéniteurs cardiovasculaires endogènes est étroitement corrélée avec l'âge. Les
tissus o atau et fœtau a dia ues o t des i eau d’e p ession élevés en progéniteurs
Isl1+, ais a e l’âge, l’e p essio de Isl est duit à des i eau fai les oi e i e ista ts
(Simpson et al., 2012). En outre, les marqueurs de surface des cellules Isl1 adultes n'ont pas
été bien définies (Serradifalco et al., 2011) car la plupart des travaux identifiant les
progéniteurs Isl1 reposent sur des études de traçage des lignages pendant le développement.
Malgré le pouvoir de différenciation multiple des CSCs Isl-1+, leu ua tit li it e da s le œu
adulte est insuffisante pour pouvoir envisager des thérapies cellulaires après infarctus de
myocarde.
2.4 Les CSCs BCRP+ positives side-population (BCRP+ SP)
Les cellules souches cardiaques BCRP+ “P o t t ide tifi es da s le œu de sou is
(Hierlihy et al., 2002) et da s le œu hu ai (Emmert et al., 2013). Les CSCs BCRP+ SP ou
“ide Populatio o t t o es ai si a elles o t la p op i t d’e lu e a ti e e t le
marqueu d’ADN Hoe hst de leu toplas e, e ui pe et de les s parer des autres
cellules par cytométrie de flux. Ces CSCs SP+ sont nommées BCRP+ SP car la protéine BCRP
(Breast Cancer Resistance Protein) est responsable du phénotype SP. La protéine BCRP a été
identifiée en tant que membre des transporteurs membranaires à ATP binding cassette (ABC).
C'est un peptide de 655 acides aminés avec une capacité à extruder une grande variété de
composés chimiques à partir des cellules (Hierlihy et al., 2002). Les CSCs BCRP+ SP+ de souris
peuvent se différencier in vitro e olo ies de ellules o t a tiles lo s u’elles so t ulti es
avec des cardiomyocytes primaires de rat (Hierlihy et al., 2002). Des études ont montré que
des cellules cardiaques SP+ de souris expriment un ensemble de marqueurs qui les distinguent
des autres types de cellules souches déjà connus. Ces cellules sont positives pour Sca-1, CD31
et CD38, expriment une télomerase active et des facteurs de transcription cardiaques comme
MEF2C, GATA-4 et TEF-1 (Oh et al., 2003, 2004). In vitro, les CSCs SP+ de souris se différencient

Introduction
52
en cardiomyocytes qui expriment Nkx2.5 et des protéines sarcomériques. De plus, après
i je tio da s u od le d’i fa tus hez la sou is, es ellules so t et ou es da s le
myocarde et se différencient à la périphérie de la zone infarcie (Oh et al., 2004). Dans la
population de CSCs SP+ murines, il existe une sous-population positive pour Sca-1 et négative
pour CD31 (Pfister et al., 2005). Cette population Sca-1+CD31- qui exprime déjà des marqueurs
de cellules musculaires et des cellules endothéliales présentent le plus grand potentiel de
différenciation vers le lignage cardiomyocytaire. Différencier les cellules SP cardiaques des
CSCs Sca-1+ reste une approche expérimentale difficile et non clairement définie. L'analyse
par FACS a suggéré que les cellules SP sont en fait une sous-population des CSCs Sca-1+ (Oh et
al., 2003). Par conséquent, il est probable que seules les cellules SP Sca-1+/CD31- soient
responsables du potentiel cardiomyogénique observé (Pfister et al., 2005). Une autre étude a
montré que les cellules Sca-1+/CD31- de souris isolées par tri cellulaire magnétique se
différencient en lignées endothéliales et cardiomyogéniques (Wang et al., 2006). In vitro, la
stimulation des cellules Sca-1+/CD31- avec le VEGF entraine une forte expression des
marqueurs endothéliaux. La stimulation par la 5-azacytidine et plusieurs facteurs de
croissance induit une différenciation cardiomyocytaire. Comme pour les cellules SP Sca-
1+/CD31- décrites (Pfister et al., 2005), la différenciation des cellules progénitrices Sca-1+/
CD31- est favorisée en co-culture avec des cardiomyocytes de rat. Environ 93% des CSCs SP+
sont Sca-1+/CD45-/c-kit- (Oh et al., 2003) mais les CSCs Sca-1+ ne sont pas toutes des cellules
SP+ (Noseda et al., 2015). Les CSCs SP+ sont des cellules quiescentes retrouvées dans les tissus
adultes, qui sont a a t is es pa l’e lusio spo ta e du olo a t Hoe hst ,
l’e p essio de la esti e et leu capacité à former des sphères in vitro (Tomita et al., 2005).
Fonctionnellement, les cellules CSCs SP+ cardiaques peuvent se différencier en multiples
lignages cellulaires (figure H-17). L'analyse transcriptomique réalisée sur les cellules SP de
souris a montré la présence de certains marqueurs endothéliaux (par exemple, VEGF, Vcam1,
Icam1, Vwf) mais une absence de CD31 (Martin et al., 2004). Tomita et ses collègues, ont
montré l'expression de CD29, CD44 à la surface des cellules SP et l'absence de marqueurs de
la lignée hématopoïétique (Tomita et al., 2005).
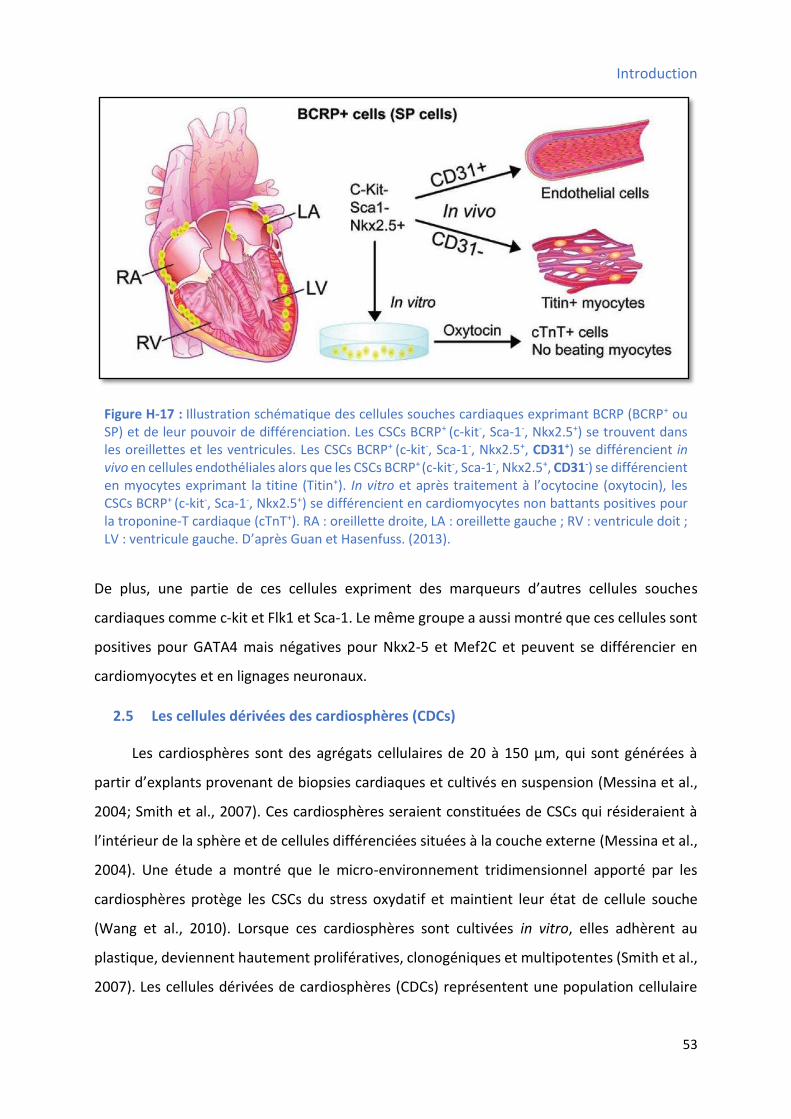
Introduction
53
De plus, u e pa tie de es ellules e p i e t des a ueu s d’aut es ellules sou hes
cardiaques comme c-kit et Flk1 et Sca-1. Le même groupe a aussi montré que ces cellules sont
positives pour GATA4 mais négatives pour Nkx2-5 et Mef2C et peuvent se différencier en
cardiomyocytes et en lignages neuronaux.
2.5 Les cellules dérivées des cardiosphères (CDCs)
Les a diosph es so t des ag gats ellulai es de à μ , ui so t g es à
pa ti d’e pla ts p o e a t de biopsies cardiaques et cultivés en suspension (Messina et al.,
2004; Smith et al., 2007). Ces cardiosphères seraient constituées de CSCs qui résideraient à
l’i t ieu de la sphère et de cellules différenciées situées à la couche externe (Messina et al.,
2004). Une étude a montré que le micro-environnement tridimensionnel apporté par les
cardiosphères protège les CSCs du stress oxydatif et maintient leur état de cellule souche
(Wang et al., 2010). Lorsque ces cardiosphères sont cultivées in vitro, elles adhèrent au
plastique, deviennent hautement prolifératives, clonogéniques et multipotentes (Smith et al.,
2007). Les cellules dérivées de cardiosphères (CDCs) représentent une population cellulaire
Figure H-17 : Illustration schématique des cellules souches cardiaques exprimant BCRP (BCRP+ ou SP) et de leur pouvoir de différenciation. Les CSCs BCRP+ (c-kit-, Sca-1-, Nkx2.5+) se trouvent dans les oreillettes et les ventricules. Les CSCs BCRP+ (c-kit-, Sca-1-, Nkx2.5+, CD31+) se différencient in
vivo en cellules endothéliales alors que les CSCs BCRP+ (c-kit-, Sca-1-, Nkx2.5+, CD31-) se différencient en myocytes exprimant la titine (Titin+). In vitro et ap s t aite e t à l’o to i e o to i , les CSCs BCRP+ (c-kit-, Sca-1-, Nkx2.5+) se différencient en cardiomyocytes non battants positives pour la troponine-T cardiaque (cTnT+). RA : oreillette droite, LA : oreillette gauche ; RV : ventricule doit ; LV : e t i ule gau he. D’ap s Gua et Hase fuss. .

Introduction
54
hétérogène composée de cellules indifférenciées et engagées, exprimant c-kit, Sca-1, CD105,
CD90 et CD29 (Messina et al., 2004). Les CDCs peuvent se différencier en cardiomyocytes
battants, en cellules musculaires lisses vasculaires et en cellules endothéliales (Tang et al.,
2009).
Des études in vivo sur des modèles animaux infarctus, ont montré ue l’ad inistration
des CDCs duit la taille de l'i fa tus, a lio e la f a tio d’ je tio du e t i ule gau he et
l'hémodynamique cardiaque (Carr et al., 2011; Chimenti et al., 2010; Johnston et al., 2009;
Smith et al., 2007). Chez l’Ho e, u essai li i ue de phase , onnu sous le nom de
« CADUCEUS », a o t ue l’i je tio i t a-coronarienne de CDCs réduit significativement
la masse de la cicatrice après infarctus (8,4 g au cours des six premiers mois et 12,9 g après un
a , ais au u e diff e e ’a t o se e o e a t la f a tio d’ je tio du e t i ule
gauche (Makkar et al., 2012).
2.6 Les CSCs W8B2 positives (CSC W8B2+)
W8B2 ou MSCA-1 (Mesenchymal Stem Cell Antigen-1) est une phosphatase alcaline non
spécifique (TNAP pour tissue nonspecific alkaline phosphatase) (Sobiesiak et al., 2010). Cette
enzyme (ALP pour alkaline phosphatase) est présente dans de nombreux tissus humains et
possède trois isoformes spécifiques du placenta, des cellules germinales et des intestins et
une isoforme tissulaire non spécifique (Devito et al., 2014).
L’e p essio du marqueur W8B2 chez la souris a été retrouvée dans plusieurs
populations de cellules progénitrices dans les zones neuronales embryonnaires, postnatales
et adultes (Langer et al., 2007). Chez l’Ho e, W B est utilis o e a ueu sp ifi ue
des CSEs Ade u i et al., ; O’Co o et al., , des cellules progénitrices du muscle
squelettique (Dellavalle et al., 2007) et aussi comme marqueur spécifique des cellules souches
mésenchymateuses de la moelle osseuse à haut potentiel prolifératif (Bühring et al., 2007).
L’e p essio de W8B2 a également été retrouvée dans les cellules souches
se h ateuses d’aut es tissus adultes hu ai s o e le p ioste de la â hoi e
(Alexander et al., 2013) et la gelée de Wharton (Devito et al., 2014).
Deux équipes ont rapporté pour la première fois la présence de cellules souches
cardiaques exprimant le marqueur W8B2 avec une provenance mésenchymateuse (Aguiar et

Introduction
55
al., 2011). Depuis, d’aut es he heu s o t pu isole et a a t ise e t pe de ellules sou hes
cardiaques exprimant le marqueur W8B2 et confirmer leur origine mésenchymateuse (Reus
et al., 2016; Zhang et al., 2015a). En effet, les cellules souches cardiaques exprimant W8B2
(CSCs W8B2+) sont positives pour les marqueurs mésenchymateux (CD29+ ,CD73+ ,CD90+
,CD105+ et CD106+), négatives pour les marqueurs hématopoïétiques endothéliaux (CD31-
,CD34- ,CD45- ,CD11b- ,CD19- et CD133-), pour le marqueur des fibroblastes cardiaques DDR2
et les marqueurs de CSCs (Aguiar et al., 2011; Reus et al., 2016; Rossini et al., 2011; Zhang et
al., 2015a).
Bien que les CSCs W8B2+ aient une origine mésenchymateuse, l'analyse génomique a
montré que ces cellules mésenchymateuses cardiaques diffèrent par leur expression génique
des cellules mésenchymateuses de la moelle osseuse ou encore les CSCs c-kit+ (Rossini et al.,
2011; Zhang et al., 2015a).
De plus, les CSCs W8B2+ expriment des facteurs de transcription spécifiques cardiaques
comme GATA4 et peuvent être différenciées en cellules endothéliales, cellules musculaires
lisses et en cellules ayant un phénotype cardiaque immature (Zhang et al., 2015a). Ces cellules
cardiaques immatures (cardiac-like cells), obtenues après différenciation in vitro des CSCs
W8B2+, ne possèdent pas de structures contractiles matures comme celles retrouvées dans
un cardiomyocyte mais répondent aux stimulations électriques (Zhang et al., 2015a).
Rossini et ses collègues ont aussi montré que les cellules souches cardiaques
mésenchymateuses, comparativement aux cellules souches mésenchymateuses de la moelle
osseuse, se différenciaient plus facilement vers le phénotype cardiovasculaire que vers les
phénotypes adipogénique et ostéogénique.
Les CSCs W8B2+ possèdent des capacités réparatrices cardiaques importantes
lo s u’elles so t i je t es da s des od les u i s d’i fa tus du o a de. E effet,
l’i je tio i t a- o a di ue de es ellules à des ats souff a t d’i fa tus h o i ue du
o a de a o t u’elles taie t apa les de ig e e s la i at i e fi euse et de se
différencier en cardiomyocytes (Rossini et al., 2011). Ces résultats ont été confirmés par les
t a au de )ha g et ses oll gues ui o t o t ue l’i je tio i t a-myocardique des CSCs
W8B2+ da s des od les d’i fa tus de o a de h o i ue de rats, améliorait la fraction

Introduction
56
d’ je tio du e t i ule gau he et aug e tait la as ula isatio de la i at i e fi euse ap s
trois semaines de transplantation (Zhang et al., 2015a).
Parallèlement à leurs capacités réparatrices, les CSCs W8B2+ possèdent un large
sécrétome composé de cytokines et de facteurs de croissance intervenant notamment dans
l'a gioge se, la su ie ellulai e, le hi iota tis e, la po se i u itai e, l’i fla atio
et le remodelage extracellulaire (Reus et al., 2016; Zhang et al., 2015a).
Il a été également montré que le milieu conditionné issu de cellules souches cardiaques
se h ateuse fa o isait l’a ti it a giog i ue des ellules e doth liales hu ai es
(microvasculaires et celles de la veine ombilicale), la survie et la prolifération des
cardiomyocytes de rats et des cellules souches mésenchymateuse humaines du tissu adipeux
et favorisait la différenciation cardiomyocytaire de la lignée myoblastique H9c2 (Reus et al.,
2016; Zhang et al., 2015a).
3 Modulateurs de la différenciation in vitro des CSCs adultes en cardiomyocytes
Plusieurs approches expérimentales ont été appliquées pour induire la différenciation
des CSCs en cardiomyocytes in vitro. Plusieurs protocoles de différenciation cardiaque existent
comme l'utilisation de milieux de culture spéciaux, la co-culture avec des cardiomyocytes
néonataux, la stimulation électrique, la stimulation mécanique, des approches
pharmacologiques utilisant par exemple l'ocytocine ou la 5-azacytidin, la stimulation par le
FGF- 2 ou encore le TGF-β (Pasha et al., 2008; Rosenblatt-Velin et al., 2005; Tran et al., 2014).
Divers groupes ont isolé et différencié des CSCs adultes en utilisant des protocoles de
différenciation différents (Chamuleau et al., 2009) (Tableau H-2).

Introduction
57
Tableau H-2 : Protocoles de différenciation cardiaques utilisés pour différents types de CSCs. Modifié
d’ap s Cha uleau et al., .
Type de CSCs
Protocole de différenciation Gènes cardiaques exprimés après
différenciation
c-kit+
-Milieu MEM + 10% SVF+dexamethasone -Co-culture avec cardiomyocytes de rats néonatals
Actine a-sarcomérique, myosine cardiaque, desmine, connexine 43
Sca-1+
-5-azacytidine -Oxytocine -5-aza + TGF-b
Troponine I, a-sarcomerique actine, myosine cardiaque, MHC (a et b), desmine, connexine 43
BCRP+ ou SP
Co-culture avec cellules cardiaque dans un milieu cardiaque spécifique
Troponine, a-actinine cardiaque
Isl1+
Co-culture avec cardiomyocytes néonatals
Troponine T, a-actinine cardiaque
Cardiosphères
-Co-culture avec cardiomyocytes de rats néonatals -Champs magnétique à très faible fréquence
Troponine, a-MHC
3.1 La 5-azacytidine
La 5-azacytidine est l'un des agents de déméthylation d'ADN le plus important et le
mieux caractérisé (Jüttermann et al., 1994). Elle s’i o po e da s l'ADN et fo e u o ple e
covalent et irréversible avec l'ADN méthyltransférase (DNMT) empêchant l'enzyme de
méthyler la position 5 des cytosines qui sont regroupées dans les îlots CpG régulateurs (Cheng
et al., 2003). Cette modification épigénétique par la 5-azacytidine conduit au démasquage des
g es ui e so t pas e p i s e aiso de l’h pe th latio de leu s p o oteu s (Keating,
2009). Bie u’il ait t précédemment montré que la 5-Azacytidine induit une différenciation
en cardiomyocytes de cellules souches mésenchymateuses in vitro, son effet sur les CSCs reste
peu clair. Concernant les CSMs issues du cordon ombilical, cette différenciation passerait par
l’a ti atio des ki ases e t a ellulai es (Qian et al., 2012).
D’aut es t a au o trent que suite au traitement avec la 5-azacytidine, les CSMs de la
oelle osseuse so t apa les d’e p i e des a ueu s sp ifi ues a dia ues et p se tent
des battements spontanés et un potentiel d'action mesurable, compatible avec un lignage

Introduction
58
myocytaire (Antonitsis et al., 2007; Naeem et al., 2013; Toma et al., 2002; Xu et al., 2004; Ye
et al., 2006).
3.2 Le TGF-b:
Le TGF-β est u fa teu de oissa e ultifo tio el t s i po ta t i pli u da s la
différenciation cardiaque des CSEs (Behfar et al., 2002; Slager et al., 1993). Les membres des
facteurs de croissance de la famille TGF-β o t t appo t s pou gule la ardiogenèse en
induisant la formation du premier champ cardiaque et en régulant les processus
morphologiques au cours du développement cardiovasculaire (Azhar et al., 2003; Brand and
Schneider, 1995).
Chez les mammifères, il existe trois isoformes de TGF-β1-3 (TGF-β , -β et -β (Schiller
et al., 2004). Durant le développement, ces trois isoformes sont présentes mais ont un profil
d’e p essio spatio-temporel différent (Pelton et al., 1991). En effet, les trois isoformes de
TGF ont montré des profils d'expression spécifiques à différents stades de développement
avec une localisation spécifique de TGF-β da s les fi es de l'œil e tode e , de TGF-β
dans le cortex de la glande surrénale (mésoderme) et de TGF-β da s l' pith liu o hl ai e
de l'oreille interne (endoderme).
De plus, l’e p essio des t ois isofo es au sei d’u e o ga e a ie en fonction du
stade e o ai e. Pa e e ple, da s le œu de sou is, le i eau d’e p essio de TGF-β
est élevé dans les coussinets atrioventriculaires et faible dans les ventricules au jour 11.5 de
développement. Au jour de développement 12,5, le niveau d’e p essio de TGF-β da s le
œu est le da s les e t i ules et les o eillettes et fai le da s les oussi ets at io-
ventriculaires.
Les protéines de la famille TGF-β se e t de sig au i du tifs pou la fo atio et la
différenciation des muscles cardiaques. Le traitement avec du TGF-β e og e a lio e la
formation du muscle cardiaque dans des CSEs de souris et des lignées cellulaires de carcinome
embryonnaire (Singla et al., 2005; Slager et al., 1993; Zeineddine et al., 2006). Dans le
myocarde infarci la famille TGF-β se ait su a ti e (Bujak and Frangogiannis, 2007; Deten et
al., 2001). L'implantation de CSMs de la moelle osseuse préprogrammée par le TGF-β da s le
o a de l s s’est l e effi a e, elle p o o ue u e g ation cardiaque accompagnée
d’u e i du tio de l’e p essio de gènes de différenciation myocardiques (Li et al., 2005).

Introduction
59
Réciproquement, la perturbation de la voie de signalisation TGF-β via un mutant dominant
négatif du récepteur au TGF-β e p he la diff e iatio des a dio o tes (Behfar et al.,
2002). L’i hi itio de l'activité du TGF-β duit l'a t du le ellulai e ai si ue l'e p essio
de l'inhibiteur des CDK : p21WAF1 et freine l'induction du facteur de transcription cardiaque
Nkx2.5. La voie de signalisation TGF-β joue u ôle i po ta t da s le maintien des capacités
dites « souches » des cellules souches adultes mais aussi celles des cellules ES (Kitisin et al.,
2007; Pucéat, 2007). Les effets observés par le TGF-β so t t s d pe da ts du te ps et de la
dose de traitement et seraient différents selon la source et le type de cellules souches (Pucéat,
2007). Il agit positivement sur la différenciation des myoblastes embryonnaires (Slager et al.,
1993) alo s u’il a u effet gatif su le p o essus de diff e iation myogénique des
myoblastes de lignée C2C12 (Gardner et al., 2011; Massagué et al., 1986).
E su , l’e se le des tudes e es su la diff e iatio des C“Cs adultes
humaines montrent clairement que la stimulation par TGF-β i duit et/ou a lio e la
différenciation en cardiomyocytes.
3.3 L’a ide as o i ue :
L'acide L-ascorbique (AA), souvent appelé vitamine C, est essentiel à la vie chez l'homme,
chez lequel ses propriétés antioxydantes protègent les cellules contre le stress oxydatif
(Padayatty et al., 2004). L’AA pou ait t e o sid o e u a teur important dans la
p olif atio et la diff e iatio ellulai e. E effet, Il a t o t ue l’AA a ait u effet
a tip olif atif asso i à l’i hi itio des g es essai es da s la p og essio du le
cellulaire (Belin et al., 2009). De plus, l’AA i hi e la diff e iatio des p adipo tes adultes
en adipocytes matures (Rahman et al., 2014). Plusieu s tudes e tes o t o t ue l’AA
est capable de reprogrammer des cellules somatiques murines et humaines en cellules
souches adultes (Esteban et al., 2010; Stadtfeld et al., 2012; Wang et al., 2011). Cette
reprogrammation est médiée par la déméthylation de l'ADN sur des loci spécifiques (Chen et
al., 2013).
L'AA favorise la différenciation cardiaque des CSEs (Chan et al., 2009; Sato et al., 2006)
et améliore la différenciation cardiaque des cellules souches pluripotentes murines par
stimulation de la voie de signalisation Wnt/β-catenine (Ivanyuk et al., 2015). La
différenciation des CSEs vers le lignage myogénique serait engagée via son récepteur

Introduction
60
intracellulaire SVCT2 (Rahman et al., 2017). La diff e iatio pa l’AA i pli ue la oie p
MAPK / CREB, probablement stimulée par l'AMPc via l'adénylate cyclase. De e fait, l’AA est
utilisé dans plusieurs protocoles de différenciation cardiaque des ESC et des cellules souches
pluripotentes induites ou iPSCs (Burridge et al., 2011; Kattman et al., 2011; Yang et al., 2008).
3.4 L’a ide ti oï ue :
L'acide rétinoïque (retinoic acid ou RA) est le dérivé actif de la vitamine A. Le RA régule,
par stimulation de ses récepteurs nucléaires (RAR ou retinoic acid receptor), des voies de
signalisation qui sont essentielles à la croissance des organes pendant le développement
embryonnaire (Duester, 2008; Niederreither and Dollé, 2008). La génération locale de RA
implique la rétinaldéhyde déshydrogénase 2 (Raldh2) catalyseur de la deuxième étape
d'oxydation dans la biosynthèse du RA. En effet, les embryons de souris déficients pour la
Raldh2 (Raldh2 -/-) présentent une carence sévère en AR et des défauts de développement
cardiaques (Niederreither et al., 1999, 2003). L’i hi itio de la sig alisatio pa le RA i duit
également des défauts de développement précoce cardiaques (Gruber et al., 1996; Hochgreb
et al., 2003). En excès, l’a ide ti oï ue p o o ue u la gisse e t du o pa ti e t si o-
auriculaire au détriment des tissus ventriculaires chez les embryons de poulets (Osmond et
al., 1991; Yutzey et al., 1994) et chez le poisson zèbre (Stainier and Fishman, 1992). Toutes ces
études indiquent que le RA est nécessaire en tant que morphogène pour un développement
cardiovasculaire normal, mais entraîne des défauts myocardiques si il est présent à des
concentrations anormales.
Dans les CSEs, le traitement avec le RA accélère la différenciation cardiaque et améliore
le développement des cardiomyocytes ventriculaires (Wobus et al., 1997). Par analogie à la
vitamine C, l’effet du RA su la diff e iatio des E“C (soit en cellules pacemaker soit en
cellules atriales) est dépendant de la dose et du te ps d’ad i ist atio (Gassanov et al.,
2008). Des effets de l’AR e o i aiso a e d’aut es ol ules su la diff e iatio des
cellules souches progénitrices in vitro, ont été observés mais les mécanismes moléculaires
restent complexes. Récemment, il a été montré que le RA en combinaison avec la 5-
azacytidine et le DMSO, favorise la différenciation cardiaque des CSMs humaines dérivées du
foie fœtal (Deng et al., 2016). D’aut e pa t, da s les CSMs hu ai es d i es du œu , le RA
(en combinaison avec la 5-azacytidine et le phényl butyrate) induit une différenciation de ces
cellules vers un phénotype cardiaque immature (Zhang et al., 2015a).

Introduction
61
3.5 La dexaméthasone :
La dexaméthasone est un glucocorticoïde synthétique, couramment administré à des
nourrissons prématurés, afin de réduire l'incidence et la gravité du syndrome de détresse
respiratoire (Liggins and Howie, 1972). Cependant, l'exposition aux glucocorticoïdes
synthétiques peut être nocive pour d'autres tissus et organes (Davis et al., 2011; Kelly et al.,
2012). Une étude récente indique que le traitement des rats nouveau-nés avec la
dexaméthasone pendant une période critique du développement cardiaque, diminue la
prolifération et le o e total des a dio o tes da s le œu via des modifications
épigénétiques (Gay et al., 2015). Cette étude montre également que la dexaméthasone
stimule une différenciation terminale cardiomyocytaire prématurée par la répression
épigénétique du gène cycline D2.
Ces résultats ont fourni de nouvelles pistes concernant la régulation de la maturation
des cardiomyocytes par les glucocorticoïdes. Cependant, les mécanismes moléculaires
impliqués durant la différenciation des cellules souches cardiaques en cardiomyocytes en
utilisant la dexaméthasone restent non étudiés. Ainsi, les différentes propriétés de la
dexaméthasone évoquées ci-dessus conduisent à son utilisation in vitro dans plusieurs
protocoles de différenciation de CSCs en cardiomyocytes (Matsuura et al., 2004; Zhang et al.,
2015a).
3.6 L’o to i e:
L'ocytocine ou oxytocine, est un neuropeptide composé de neuf acides aminés sécrétés
principalement par les noyaux hypothalamiques. L'ocytocine est également produite dans des
tissus p iph i ues o e l’ut us, le pla e ta, l’a ios, le o ps jau e o pus luteu , le
testi ule et le œu (Gimpl and Fahrenholz, 2001). Elle a longtemps été reconnue comme étant
principalement impliquée dans la régulation des fonctions reproductives, telles que
l'induction des contractions utérines, le réflexe d'éjection du lait et l'ovulation (Gimpl and
Fahrenholz, 2001). D’aut es ôles de l’o to i e o t t tudi s ota e t da s la gulatio
de différentes fonctions et comportements (comportement sexuel et maternel, la mémoire,
la cognition, la tolérance, les fonctions cardiovasculaires etc.) (Bussolati and Cassoni, 2001).
L'ocytocine est également considérée comme régulateur positif et négatif de la prolifération
cellulaire dans différents tissus (Cassoni et al., 2001). Cette hormone administrée en excès
hez le fœtus pe tu e le d eloppe e t a dia ue hez le at (Schriefer et al., 1982). Elle

Introduction
62
régulerait également la différenciation. En effet, le nombre de récepteu s à l’o to i e
augmente à partir du 7ème jour de gestation, le jour où la différenciation cardiaque
o e e hez l’e o de sou is (Mukaddam-Daher et al., 2002). Chez l’adulte, da s
l'i fa tus du o a de i duit e p i e tale e t hez les ats, l’appo t o ti u d’o to i e
améliore la guérison cardiaque, le travail cardiaque, réduit l'inflammation et stimule
l'angiogenèse (Gutkowska et al., 2014).
En résumé, l'ocytocine est décrite comme étant un puissant inducteur de différenciation
cardiomyocytaire dans les CSEs et les CSCs adultes de souris (Matsuura et al., 2004; Paquin et
al., 2002).
Chapitre III : Physiologie des cellules souches /progéniteurs cardiaques
1 Le calcium :
1.1 Le calcium dans les cellules souches:
Dans toutes les cellules et particulièrement dans les cellules somatiques de mammifères
cet ion divalent joue un rôle fondamental dans les processus de prolifération et la
différenciation. Cependant le rôle du calcium da s les ellules sou hes ’a t tudi ue
depuis quelques années (Heubach et al., 2004; Kawano et al., 2002). Cette recherche a suscité
un intérêt important dans le domaine car les données suggèrent que la signalisation calcique
est liée à la prolifération et à la différenciation cellulaire, deux fonctions fondamentales des
cellules souches.
Le calcium (Ca2+) est un cation ubiquitaire qui contrôle de nombreux processus
cellulaires tels que la prolifération, la différenciation cellulaire, et l'apoptose. Le mécanisme
général de la signalisation calcique est relativement simple et repose sur les variations
spatiotemporelles de cet ion (calcium libre). Dans les cellules eucaryotes, la concentration de
calcium intracellulaire [Ca2+]i au repos est comprise entre 50 et 100 nM et peut atteindre entre
1 et 10µM suite à une stimulation. Les composants moléculaires du réseau de signalisation
Ca2+ peuvent être divisés en plusieurs groupes selon leur fonction : il existe des canaux
calciques de la membrane plasmique qui permettent l'entrée de Ca2+ dans la cellule (comme
les canaux calciques indépendants et dépendants du voltage), des canaux de libération
calcique localisés aux niveaux des stocks intracellulaires (comme les canaux sensibles à
l’i ositol t iphosphate IP et à la ryanodine), des pompes et des échangeurs Ca2+ (comme la

Introduction
63
pompe PMCA (pour plasma membrane Ca2+ ATPase) et SERCA (sarco/endoplasmic reticulum
Ca2+-ATPase), l’ ha geu Na+/Ca2+) qui réduisent la concentration du calcium cytosolique
et/ou favorisent sa recapture dans les stocks intracellulaires; et enfin de nombreux senseurs
cytoplasmiques (comme la calséquestrine, la calmoduline, le phospholamban ou encore la
calcineurine A) qui traduisent les signaux calciques en activité cellulaire.
Plusieurs paramètres caractérisent les signaux calciques: leur origine, leur localisation,
leur fréquence, leur amplitude et leur cinétique. Ainsi, en fonction de la nature de ces signaux,
la ellule pa l’i te diai e de p ot i es lia t le al iu a la o e u e po se
appropriée (Berridge et al., 2000).
1.1.1 L’a tivit al i ue spo ta e da s les ellules sou hes:
Les oscillations calciques spontanées ont été observées à la fois dans les cellules
excitables et non-excitables et elles sont importantes pour le développement embryonnaire.
Les oscillations calciques contribuent à la régulation de plusieurs processus tels que la
fertilisation, la prolifération cellulaire, la sécrétion, la différenciation neuronale, la croissance
axonale, la prolifération et la migration neuronale etc. (Clapham, 2007; Montoro and Yuste,
2004).
Bie u’elles soie t des ellules dites « non-excitables », les cellules souches adultes
expriment certains canaux ioniques activés par le voltage (Heubach et al., 2004; Zahanich et
al., 2005). Une étude réalisée sur les CSEs et les CSMs a montré que ces deux types cellulaires
e po daie t pas au sti ulatio s pa le gluta ate, le GABA, l’o to i e ou la
dépolarisation confirmant leur non-excitabilité (Forostyak et al., 2013).
Cependant, la majorité des cellules souches ont généré des [Ca2+]i transitoires après
sti ulatio pa l’ATP ; % des CSEs, 90% des CSMs du tissu adipeux et 62% des CSMs de la
moelle osseuse (Kawano et al., 2006). La majorité des CSMs ont des récepteurs purinergiques
fonctionnels (Faroni et al., 2013; Kotova et al., 2014) de type P2X pour les CSMs du tissu
adipeux et de type P2X et P2Y pour les CSMs de la moelle osseuse (Coppi et al., 2007).
Les transitoires [Ca2+] spontanés se produisent principalement au cours des premiers
stades de la différenciation des précurseurs neuronaux et régulent la croissance des neurites
et l'apparition du phénotype GABAergique (Ciccolini et al., 2003). L'activité [Ca2+] spontanée
a été observée dans 31% des précurseurs neuronaux dérivés des CSEs humaines et elle est

Introduction
64
médiée par l'influx Ca2+ via les VGCC HVA (pour High-voltage-activated Voltage-gated calcium
channel) (Forostyak et al., 2013). L’i hi itio des t a sitoi es [Ca2+] spontanés (médiés par les
VGCC de type L et des canaux de type TRPC1) entraine une réduction significative de la
prolifération dans les neurones post-mitotiques dérivés des CSEs humaines (Weick et al.,
2009). L'activité [Ca2+] spontanée et synchronisée enregistrée dans les progéniteurs
neuronaux dérivés des CSNs fœtales hu ai es se ait d pe da te des jo tio s gap de t pe
connexine 43 et des VGCC (Jäderstad et al., 2010). En effet dans ces cellules neuronales,
l'inhibition des jonctions gap annule l'activité Ca2+ spontanée et la prolifération cellulaire
(Malmersjö et al., 2013).
Les CSMs de la moelle osseuse possèdent également des oscillations [Ca2+] spontanées
et des transitoires calciques (Kawano et al., 2003). Dans les CSMs de la moelle osseuse
humaines, les transitoires [Ca2+] spontanés résultent de la libération calcique des stocks du
réticulum endoplasmique (ER) médiée par IP3 (Kawano et al., 2003, 2006).
En résumé, les oscillations calciques spontanées se produisent dans plusieurs types
cellulaires; dans les CSEs, elles dépendent de l'influx Ca2+ via la membrane plasmique, alors
que dans les cellules souches adultes, elles sont déclenchées principalement par la libération
du Ca2+ depuis le RE. Les recherches dans ce domaine et plus particulièrement sur les cellules
sou hes adultes so t peu o euses. Les d ou e tes da s l’a e i pou aient aider à la
compréhension des mécanismes et du rôle de ces oscillations dans la prolifération et la
différenciation.
1.1.2 Canaux VGCC et signalisation calciques
Les canaux Ca2+ voltage-dépendant (Voltage-Gated Ca2+ Channels ou VGCC)
représentent la principale voie d'entrée de Ca2+ dans les cellules excitables. Les VGCC ont été
lass s e deu g oupes p i ipau : les a au à haut seuil d’a ti atio High Voltage-
Activated channels ou HVA) qui comprennent les canaux de type L, N-, P / Q et R et les canaux
à bas seuil d’a ti atio Lo Voltage-Activated channels ou LVA) ou canaux de type T (Catterall
and Swanson, 2015; Wang et al., 1999b). L'expression du VGCC de type L et T a été identifiée
dans de nombreux types de cellule souches.
En particulier, ils ont été détectés au niveau protéique dans les CSMs du tissu adipeux
(Bai et al., 2007; Jang et al., 2010) et dans les CSMs de la moelle osseuse (Heubach et al., 2004;

Introduction
65
Kawano et al., 2002; Zahanich et al., 2005). Certains travaux montrent que la modulation des
canaux de type T qui se produit pendant la progression dans le cycle cellulaire pourrait
contribuer à la maintenance de la capacité d'auto-renouvellement des CSEs de souris
(Rodríguez-Gómez et al., 2012). L'activation des VGCC LVA serait à l’o igi e des os illatio s
calciques dans les progéniteurs neuronales dérivés des CSEs suggérant leur intervention dans
le processus de prolifération cellulaire (Malmersjö et al., 2013). Les stocks calciques impliqués
dans ces oscillations induites dans les CSEs et les CSMs avant différenciation font intervenir
principalement les stocks calciques sensibles aux IP3 couplés aux vecteurs calciques
membranaires P2x,y et VGCC. Les VGCC favorisent la différenciation des CSEs ainsi que les
CSMs adultes de différentes origines (Forostyak et al., 2016) (figure H-18). En effet, au cours
de la différenciation neuronales des CSEs, ces dernières commencent à exprimer les canaux
calciques (L-VOCC, N-VOCC, P/Q-VOCC et R-VOCC). Les CSMs (du tissu adipeux et de la moelle
osseuse) quand elles sont différenciées en plusieurs lignages (neuronal, adipogénique et
ostéogénique) expriment les canaux calciques L-VOCC et T-VOCC (pour les CSMs du tissu
adipeux) et les canaux calciques P/Q-VOCC (pour les CSMs de la moelle osseuse). En
particulier, il a été démontré que la différenciation neuronale des CSEs dépend de la
coopération entre les canaux calciques VGCC de type L et les canaux de libération calcique
intracellulaire de type RyR2 (Yu et al., 2008) (figure H-18).
Le signal calcique spontané ou induit observé pendant la différenciation jouerait un rôle
fo da e tal da s l’e p essio de e tai s g es ota e t ceux impliqués dans le cycle
e du al iu . L’appa itio des epteu s à la a odi e et de diff e ts a au al i ues
t pes N, P/Q, R, T, L e t e aut es se ait o te po ai e à l’ olutio des p op i t s du sig al
calcique.
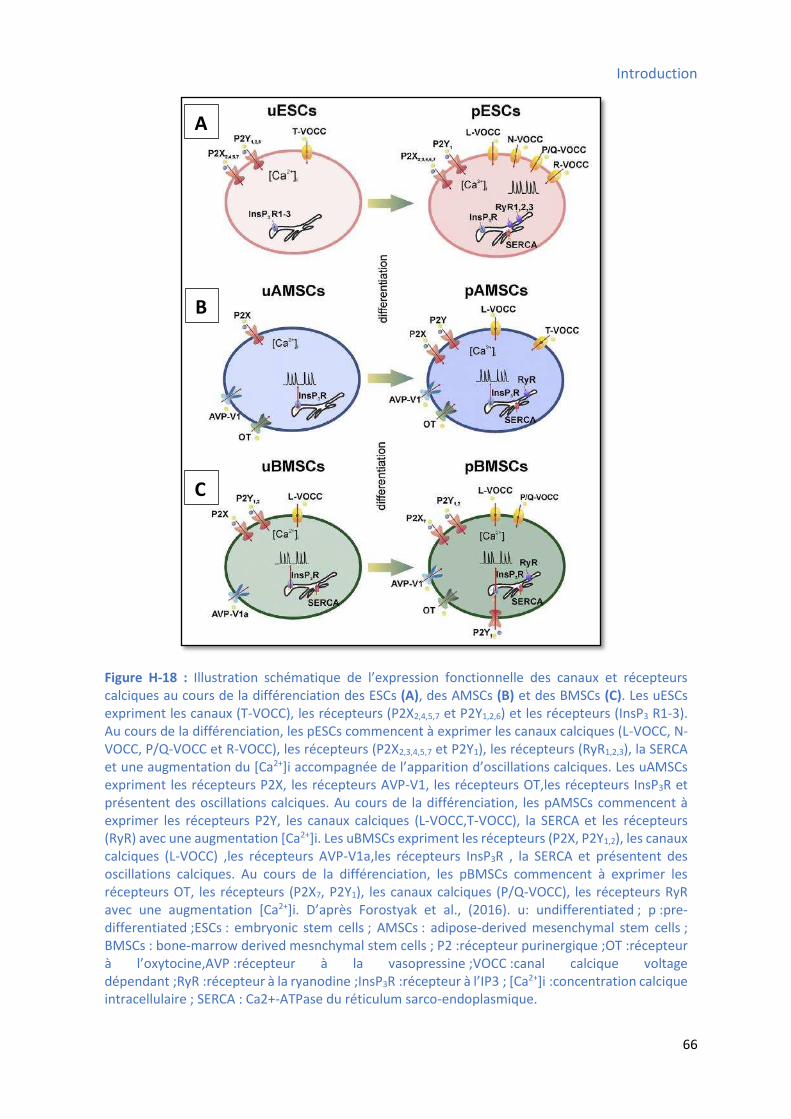
Introduction
66
Figure H-18 : Illust atio s h ati ue de l’e p essio fo tio elle des a au et epteu s calciques au cours de la différenciation des ESCs (A), des AMSCs (B) et des BMSCs (C). Les uESCs expriment les canaux (T-VOCC), les récepteurs (P2X2,4,5,7 et P2Y1,2,6) et les récepteurs (InsP3 R1-3). Au cours de la différenciation, les pESCs commencent à exprimer les canaux calciques (L-VOCC, N-VOCC, P/Q-VOCC et R-VOCC), les récepteurs (P2X2,3,4,5,7 et P2Y1), les récepteurs (RyR1,2,3), la SERCA et une augmentation du [Ca2+]i a o pag e de l’appa itio d’os illatio s al i ues. Les uAM“Cs expriment les récepteurs P2X, les récepteurs AVP-V1, les récepteurs OT,les récepteurs InsP3R et présentent des oscillations calciques. Au cours de la différenciation, les pAMSCs commencent à exprimer les récepteurs P2Y, les canaux calciques (L-VOCC,T-VOCC), la SERCA et les récepteurs (RyR) avec une augmentation [Ca2+]i. Les uBMSCs expriment les récepteurs (P2X, P2Y1,2), les canaux calciques (L-VOCC) ,les récepteurs AVP-V1a,les récepteurs InsP3R , la SERCA et présentent des oscillations calciques. Au cours de la différenciation, les pBMSCs commencent à exprimer les récepteurs OT, les récepteurs (P2X7, P2Y1), les canaux calciques (P/Q-VOCC), les récepteurs RyR avec une augmentation [Ca2+]i. D’ap s Fo ost ak et al., (2016). u: undifferentiated ; p :pre-differentiated ;ESCs : embryonic stem cells ; AMSCs : adipose-derived mesenchymal stem cells ; BMSCs : bone-marrow derived mesnchymal stem cells ; P2 :récepteur purinergique ;OT :récepteur à l’o to i e,AVP :récepteur à la vasopressine ;VOCC :canal calcique voltage dépendant ;RyR :récepteur à la ryanodine ;InsP3R : epteu à l’IP ; [Ca2+]i :concentration calcique intracellulaire ; SERCA : Ca2+-ATPase du réticulum sarco-endoplasmique.
A
C
B

Introduction
67
1.1.3 Les stocks calciques intracellulaires dans les cellules souches:
Le réticulum endoplasmique (ER) est le principal organe de stockage de Ca2+.
Parallèlement, deux mécanismes majeurs de libération de Ca2+ réticulaire existent via les
récepteurs à la ryanodine (RyR) et les récepteurs au 1,4,5-triphosphate d'inositol (InsP3). Le
remplissage des stocks calciques « vidés » est assu pa l’e iste e d’u e po pe Ca2+-ATPase
(SERCA).
La cascade de signalisation métabotropique PLC/InsP3 a été démontrée comme la voie
principale induisant la libération de Ca2+ de l'ER dans différents types de cellules souches. Des
tudes o t o t ue seuls les epteu s à l’I sP , et o les R R, so t fo tio els da s
les ESC de souris (Yanagida et al., 2004); dans les CSMs humaines du tissu adipeux (Kotova et
al., 2014; Tran et al., 2014) et dans les CSMs humaines de la moelle osseuse (Kawano et al.,
2002). La oie de li atio al i ue PLC/I sP este fo tio elle uelle ue soit l’o igi e ou
le stade de différenciation, ce qui laisse à penser que cette voie est conservée dans toutes les
cellules souches. Par exemple, les CSES et les MSC différenciés en cardiomyocytes, en
adipo tes ou e eu o es, poss de t toutes des epteu s fo tio els à l’I sP (Kawano
et al., 2006; Resende et al., 2010; Sedan et al., 2008; Tran et al., 2015). Cependant, les
récepteurs à la ryanodine sont peu caractérisés dans les cellules souches, même si ils
apparaissent lors de la différenciation. Par exemple, les cellules neuronales et cardiaques
dérivées des CSEs ou des CSMs, expriment des RyR fonctionnels (Forostyak et al., 2013;
Resende et al., 2010). Des tudes o t o t u’il y a un lien fonctionnel entre les RyR type2
et les VGCC de type L et que cela est important dans la différenciation neuronale (Yu et al.,
2008).
1.2 Le calcium dans les cellules souches/progéniteurs cardiaques:
Le al iu joue u ôle i po ta t da s le œu e a ti a t les p o essus de oissa e
(Berridge et al., 2000; Frey et al., 2000) et en modulant le comportement mécanique des
cardiomyocytes (Bers, 2002; Houser and Molkentin, 2008).
A l’heu e a tuelle, les tudes su l’a ti it et la sig alisatio al i ue da s les C“Cs so t
rares. Ferreira-Martins et ses collègues ont montré pour la première fois que les CSC c-kit+
humaines possèdent des oscillations calciques spontanées et que ces dernières jouent un rôle
essentiel dans la régulation de la prolifération cellulaire (Ferreira-Martins et al., 2009). Ces

Introduction
68
oscillations intrinsèques se produisent indépendamment du couplage avec des
cardiomyocytes ou de la présence de Ca2+ extracellulaire. Ces résultats ont été confirmés dans
les œu s de sou is t a sg i ues da s les uelles l’EGFP (pour enhanced green fluorescent
protein) était sous le contrôle du promoteur c-kit. Ces oscillations calciques ont été régulées
par la libération calcique depuis l'ER par l'activation des IP3R et la réabsorption du Ca2+ par la
SERCA. Les IP3R et les SERCA ont été fortement exprimés dans ces cellules alors que les RyR
n'ont pas été détectés.
D’aut e pa t, les C“Cs -kit+ exprimaient des canaux SOC (Store-Operated Channel ou
canaux calciques activés consécutivement à la déplétion des stocks de Ca2+ du réticulum
endoplasmique) et la pompe Ca2+ de la membrane plasmique. Bien que ces canaux et pompes
soient fonctionnels, ils n'ont aucun effet direct sur les oscillations de Ca2+. Néanmoins, la
fréquence des oscillations calciques peut être significativement augmentée par l'ATP et
l'histamine via les epteu s pu i gi ues et les epteu s à l’hista i e de t pe .
D’aut e pa t, il appa ait ue es oscillations calciques sont couplées avec l'entrée des
cellules dans le cycle cellulaire et l'incorporation de BrdUrd. L'induction des oscillations
calciques dans les CSCs c-kit+ a a t leu i je tio i t a o a di ue da s des œu s i fa is de
souris, amélio e ait la g effe et l’e pa sio de es ellules (Ferreira-Martins et al., 2009). Lors
de la différenciation des CSCs en cardiomyocytes, la famille des facteurs de croissance FGF et
TGFβ joue u ôle ajeu dans lequel intervient le signal calcique (Tonelli et al., 2012) (figure
H-19).
L'activation des récepteurs TGF-ß sti ule l’a ti atio de la tat-associated kinase (TAK),
une protéine qui appartient à la voie MAPK. Le TAK agit sur les facteurs de transcription ATF2
et CREB via la voie MAPK sensible au Ca2+ (Park et al., 2009). L'ATF2 peut également se lier aux
Smads pour favoriser l'expression des facteurs de transcription responsables du
développement du phénotype cardiaque.
Wnt11 participe également au processus de différenciation cardiaque, en agissant sur
des voies de signalisation dépendantes du Ca2+ médiées par la Protéine kinase
Ca2+/calmoduline-dépendante (CaMK-II), la proteine kinase C (PKC) et les c-Jun N-terminal
kinases (JNK) (Biteau et al., 2008).
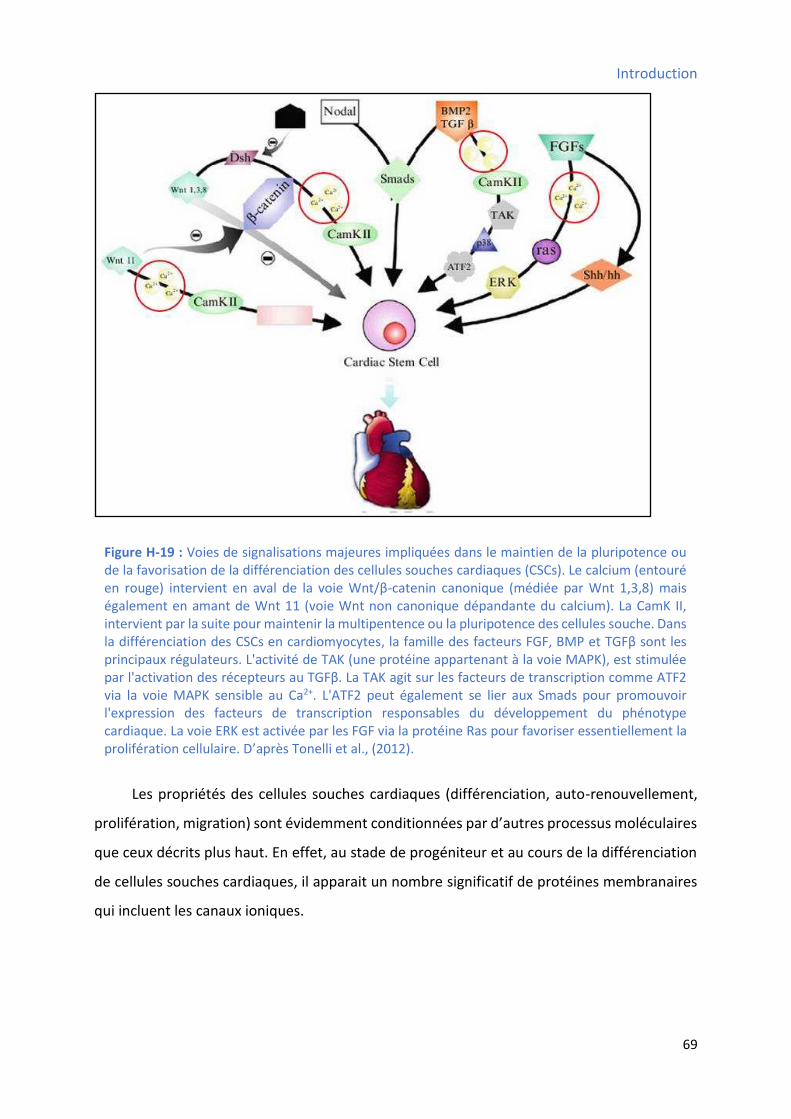
Introduction
69
Les propriétés des cellules souches cardiaques (différenciation, auto-renouvellement,
p olif atio , ig atio so t ide e t o ditio es pa d’aut es p o essus ol ulai es
que ceux décrits plus haut. En effet, au stade de progéniteur et au cours de la différenciation
de cellules souches cardiaques, il apparait un nombre significatif de protéines membranaires
qui incluent les canaux ioniques.
Figure H-19 : Voies de signalisations majeures impliquées dans le maintien de la pluripotence ou de la favorisation de la différenciation des cellules souches cardiaques (CSCs). Le calcium (entouré e ouge i te ie t e a al de la oie W t/β-catenin canonique (médiée par Wnt 1,3,8) mais également en amant de Wnt 11 (voie Wnt non canonique dépandante du calcium). La CamK II, intervient par la suite pour maintenir la multipentence ou la pluripotence des cellules souche. Dans la différenciation des CSCs en cardiomyocytes, la fa ille des fa teu s FGF, BMP et TGFβ so t les principaux régulateurs. L'activité de TAK (une protéine appartenant à la voie MAPK), est stimulée pa l'a ti atio des epteu s au TGFβ. La TAK agit su les fa teu s de t a s iptio o e ATF via la voie MAPK sensible au Ca2+. L'ATF2 peut également se lier aux Smads pour promouvoir l'expression des facteurs de transcription responsables du développement du phénotype cardiaque. La voie ERK est activée par les FGF via la protéine Ras pour favoriser essentiellement la prolifération cellulaire. D’ap s To elli et al., .

Introduction
70
2 Les canaux ioniques :
Comme nous l’a o s a o d su i te e t da s le hapit e p da t, les signaux
bioélectriques générés par les canaux ioniques jouent un rôle crucial dans la genèse de
l’e itatio le t i ue et da s la o du tio des sti ulatio s da s les ellules e ita les. Da s
les cellules non-excitables, les canaux ioniques interviennent dans différents processus
ellulai es o e la p olif atio , la ig atio et l’apoptose (Berridge et al., 1998, 2000;
Orrenius et al., 2003). E effet, il a t o t u’u e sti ulatio électrique conditionnante
(de 1 Hz pendant 7 à 14 jours) sur des progéniteurs cardiaques humains, fa o ise l’e p essio
de protéines membranaires et structurales (ex : Troponine I, SERCA2 Cx43, alpha-actine) ;
favorisant leur potentiel phénotypique cardiovasculaire (Llucià-Valldeperas et al., 2014).
Plusieurs études ont montré que les cellules souches expriment différents canaux
ioniques. Ces derniers sont exprimés de manière hétérogène et dépendent du type de cellule
souche. Le rôle de ces canaux dans les cellules souches reste peu étudié mais les données
dispo i les o t e t u’ils so t fo da e tau da s la gulatio de la p olif atio , la
différenciation et la migration.
De nombreux courants ioniques ont été caractérisés dans les cellules souches; ils
comprennent des courant potassiques rectifiants retardés voltage-dépendant ou IKDR (codé
par différents gènes KV), des courants K+ activés par le Ca2+ intracellulaire ou KCa (BKCa, KCa de
grande conductance, IKCa; KCa de conductance intermédiaire et SK4, KCa à petite conductance
comme le SK1,2,3), le courant transitoire sortant K+ (Ito) , le courant K+ rectifiant entrant (IKir),
le ou a t a ti pa l’h pe pola isatio Ih a au HCN et modulé par les nucléotides
cycliques intracellulaires et non sélectif aux cations, différents courants chlorures (ICl), le
courant sodium dépendant du voltage (INa), les courant calciques de types T et L (ICaT ,ICa.L)
et les courants TRP (transient receptor potential ) perméables aux cations (Li and Deng, 2011).
Tous ces courants étaient présents de manière hétérogène dans les cellules ES, les CSMs
de différentes origines (moelle osseuse, tissu adipeux et sang du cordon ombilical humain),
les cellules progénitrices neurales ou les cellules progénitrices cardiaques.
2.1 Les canaux ioniques dans les CSEs:
Il a été rapporté que les CSEs qui co-expriment les courants IKDR et BKCa sont retrouvés
dans 52% des cellules CSEs de souris et 100% des cellules CSEs humaines (Wang et al., 2005).

Introduction
71
Les IKDR des cellules CSEs de souris sont codés par les gènes KV1.1, KV1.2, KV1.3 et KV1.6 dans
des cellules CSEs alors que les IKDR des cellules ES humaines sont codés par les gènes KV7.2 et
Kv9.3. De plus, un courant Ih (codé par le gène HCN3) est présent dans 23% des cellules CSEs
de souris mais absent dans les cellules CSEs humaines. De même, les courants K rectifiant
entrant (IK1) responsables de la stabilisation du potentiel de membrane de repos n'étaient pas
présents dans les CSEs de souris (Wang et al., 2005).
2.2 Les canaux ioniques dans les CSMs:
La plupart des CSMs humaines de la moelle osseuse possèdent des courant KCa et IKDR
(Heubach et al., 2004; Li et al., 2005). Le courant KCa dans les CSMs de la moelle osseuse est
codé par le gène MaxiK (KCa1.1 ou Slo), comme le montrent plusieurs études (Heubach et al.,
2004; Kawano et al., 2003). De plus, des courants sodiques tetrodotoxine (TTX) sensibles
(codés par NaV1.7), des courants Ito (codés par KV1.4 et KV4.2) et des courants ICa.L (codés par
CaV1.2) sont présents dans une petite population (29%, 8% et 15% respectivement) de CSMs
humaines (Li et al., 2005).
Des courants IKCa (codé par KCa3.1), des courants Cl- sensibles au volume (codé par Clcn3)
et des courants IKir (codé par Kir2.1), sont présents dans les CSMs de la moelle osseuse de
souris (Tao et al., 2007). Le p ofil d’e p essio des a au io i ues da s les CSMs de souris
est différent de celui des cellules ES de souris, ce qui suggère que l'expression du canal ionique
peut dépendre de l'origine de la cellule souche. Il a été constaté que dans les CSMs de rat,
plusieurs courants ont été caractérisés; des courants INa TTX sensibles (16% des cellules) ; Ito
(10% des cellules) ; ICa.L (8% des cellules), IKDR (91% des cellules) et des BKCa coexprimés avec
des courants IKCa (33% des cellules) (Li et al., 2006). Dans les CSMs de la moelle osseuse de
lapin, il existe des courants IKDR (codés par KV1.2 et KV2.1) présents dans 78% des cellules, des
courants BKCa et IKCa coexprimés avec IKDR dans 29% des cellules, tandis que IKir (codé par
Kir1.1) est présent dans 28% des cellules (Deng et al., 2006). Les résultats obtenus sur les CSMs
de la oelle osseuse de at, de lapi ou de l’Ho e ette t e ide e des p ofils et des
ph ot pes d’e p essio de a au io iques différents, suggérant une expression
dépendante de l'espèce.
D’aut es tudes o t o t la présence de plusieurs courants dans les CSMs de la veine
du cordon ombilical humain. Ces cellules possèdent des courant ; BKCa (codé par KCa1.1) dans

Introduction
72
92% des cellules, Ito (codé par KV1.4 et KV4.2) dans 50%, INa TTX sensibles (codé par hNE-Na)
dans 30% des cellules et des courants IKir (codé par TWIK et Kir2.1) dans 5% des cellules.
Cependant, aucun courant IKDR typique n'a été enregistré, bien que les gènes KV1.1 et KV10.1
soient détectés dans ces cellules (Park et al., 2007). Dans les CSMs humaines du tissu adipeux,
des courants INa et Ito ont pu être enregistrés dans une petite population de cellules (8% et
19%). Les courants IKDR (73% des cellules), BKCa (codé par KCa1.1), IKCa (codé par KCa3.1) et
SKCa (codé par KCa2.3) sont présents dans ces cellules (Bai et al., 2007). Ces études suggèrent
que le profil d'expression des canaux ioniques dans les CSMs peut être tissu-spécifique.
2.3 Les canaux ioniques dans les cellules souches/progéniteurs neurales:
Une étude publiée en 1997, a montré que les cellules progénitrices oligodendrocytaires
et les oligodendrocytes différenciés provenant de rats néonatals, possèdent des courants IKDR
(codés par KV1.2, KV1.5 et KV1.6) et des courants potassiques IA (codés par KV1.4) (Attali et al.,
1997). Des études récentes ont montré également que les courants IKir (codé par Kir4.1 et
Kir5.1) et le IKDR (codé par KV3.1) sont présents dans les progéniteurs neuronaux de souris
issus de sphères (Yasuda and Adams, 2010; Yasuda et al., 2008).
De multiples canaux ioniques sont exprimés de manière hétérogène dans des cellules
souches neurales embryonnaires de rat. Ces cellules ont des courants potassqiues IA et IKDR
dans plus de 80% des cellules, des courants INa (TTX sensibles et TTX non sensibles) dans 22
% des cellules et et des courants ICa,L dans 19% des cellules souches neurales (Cai et al., 2004).
Des courants IKDR (codé par KV2.1) et IA (codé par KV4.3) ont été aussi retrouvés chez les
progéniteurs neuronaux embryonnaires de rat (Smith et al., 2008). Il a été signalé que les
cellules progénitrices du cerveau antérieur de rat néonatal co-expriment des courants INa
TTX sensibles, des courants IKDR et des courants IA (Stewart et al., 1999) alors que, seuls les
courants IKDR sont présents dans les cellules progénitrices de rat adulte (Liebau et al., 2006).
Concernant les cellules souches neurales embryonnaires humaines, des études ont
montré la présence de courants IA (codé par KV4.2) et de courants IKDR dans les CSEs neurales
hu ai es d i es du tissu e eu de fœtus a o t (Schaarschmidt et al., 2009). Quatre
types de courants ioniques (IA, IKDR, IKir et INa TTX sensibles) ont été également décrits dans
les ellules sou hes eu ales hu ai es du o te de fœtus a o t (Lim et al., 2008).

Introduction
73
2.4 Les canaux ioniques dans les CSCs:
Peu d’i fo atio s su l'e p essio des a au io i ues da s les ellules
souches/progénitrices cardiaques humaines sont disponibles. Une étude sur les cellules
progénitrices c-kit cardiaques de souris adultes a montré que ces cellules expriment des
courants IKDR (codé par KV1.1, KV1.2 et KV1.6), des courants ICl (codé par Clcn3) et des
courants IKir (codé par Kir1.1, Kir2.1 et Kir2 .2) (Han et al., 2010).
En 2014, Zhang et ses collègues ont caractérisé des courants ioniques dans les cellules
progénitrices cardiaques humaines c-kit+. Plusieurs courants ioniques étaient présents dans
ces cellules ; des courants BKCa codés par KCa.1.1 (dans 86% des cellules), des courants IKir
codés par Kir2.1 ( dans 84% des cellules), des courants Ito codés par KV4.2 et KV4.3 (dans 47%
des cellules) et des courants Na+ TTX sensibles codés par SCN3A et SCN8A (dans 61% des
cellules) (Zhang et al., 2014a). Ces résultats ont révélé pour la première fois que quatre types
de courants ioniques (BKCa, Ito, IKir et INa TTX sensibles) sont présents de manière hétérogène
dans les cellules cardiaques humaines c-kit+.
Une autre étude publiée en 2016 par Che et ses collègues, a identifié la présence de
canaux TRPV (transient receptor potential vanilloid) de type 2 et de type 4 sur les CSC c-kit+
(Che et al., 2016).
3 Rôles des canaux ioniques dans la régulation de la prolifération et/ou la différenciation
des cellules souches:
3.1 Prolifération
Les canaux ioniques jouent un rôle important dans le contrôle de la prolifération
cellulaire (MacFarlane and Sontheimer, 2000; Pardo, 2004). Dans les CSMs de la moelle
osseuse de rat, il a été montré que durant la transition G1/S du cycle cellulaire, le courant
IKDR diminue tandis que le courant IKCa augmente dans les MSC dérivés de la moelle osseuse
de rat (Deng et al., 2007).
De plus, l’e ti tio des a au IKDR ou les a au IKCa avec des siRNA ciblant KV1.2 et
KV2.1 (pour IKDR) ou KCa3.1 (pour IKCa) inhibe la prolifération cellulaire et provoque
l’a u ulatio des ellules e phase G / G du le, e ui sugg e ue es deux canaux
sont nécessaires à la régulation de la prolifération cellulaire de ces cellules (Deng et al., 2007;

Introduction
74
Wang et al., 2008). Le blocage du canal responsable du courant IKDR réduit considérablement
la prolifération des cellules CSEs murines et humaines (Wang et al., 2005), et celle des CSMs
humaines du tissu adipeux (Bai et al., 2007) mais pas la prolifération des CSC c-kit+ murines
(Han et al., 2010). Cependant, l'inhibition de IKDR favorise la prolifération de cellules
progénitrices neurales adultes de rat (Liebau et al., 2006; Yasuda and Adams, 2010; Yasuda et
al., 2008) et celle des progéniteurs neuronaux humains (Schaarschmidt et al., 2009).
De même, le blocage de IKCa réduit également la prolifération cellulaire dans les CSMs
murines de la moelle osseuse avec une accumulation des cellules dans la phase G0 / G1 du
cycle (Tao et al., 2008). Cependant, dans les CSMs humaines du tissu adipeu l'i hi itio d’IKCa
n'a pas d'effet inhibiteur sur la prolifération cellulaire (Bai et al., 2007).
L'effet régulateur de BKCa sur la prolifération a été reporté dans plusieurs types
cellulaires. L'inhibition de BKCa réduit la prolifération cellulaire des pré-adipocytes humains
(Hu et al., 2009) et celle des cellules ES murins (Wang et al., 2005) mais pas celle des CSMs
humaines du tissu adipeux (Bai et al., 2007). Cependant, l'inhibition de BKCa réduit la
p olif atio ellulai e et p o o ue l’a t du le e phase G /G da s les CSMs humaines
de la moelle osseuse (Zhang et al., 2014b). De la e a i e, l’i hi itio de BKCa réduit la
prolifération en accumulant les cellules en phase G0/G1 dans les CSCs c-kit+ humaines (Zhang
et al., 2015b). Le canal Cl- sensible au volume a été impliqué dans la prolifération cellulaire et
l'apoptose de plusieurs types cellulaires (Blackiston et al., 2009; Lang et al., 2006; Liu et al.,
2010). L’i hi itio de ICl. ol duit ota le e t la p olif atio ellulai e des CSMs murines
(par accumulation des cellules en phase G0/G1) en inhibant la cycline D et la cycline E (Tao et
al., 2008). De même, le blocage du canal ICl.vol diminue également la prolifération cellulaire
des CSC c-kit+ murines (Han et al., 2010). L’h pe pola isatio de la e a e est i pli u e
da s l’i hi itio de la p olif atio ellulai e (Hu et al., 2009; Kuhlmann et al., 2005).
Dans les cellules progénitrices neuronales adultes, la dépolarisation membranaire par
l’i hi itio de IKi ou par l'augmentation de la concentration de K+ extracellulaire favorise la
prolifération cellulaire (Yasuda et al., 2008). Cepe da t l’i hi ition du canal Kir2.1 dans les
CSCs c-kit+ hu ai es ’affe te pas la p olif atio ellulai e (Zhang et al., 2015b). Concernant
le rôle du courant Ito dans la prolifération, des études ont montré que l'activation du courant
IA (KV4.2) est une condition requise pour la prolifération dans les cellules progénitrices
neuronales embryonnaires humaines (Schaarschmidt et al., 2009) et ue l’i hi itio du a al

Introduction
75
KV4.2 réduit la prolifération cellulaire dans les préadipocytes humains (Hu et al., 2009). Une
tude e te a o t ue l’i hi itio des a au TRPV et TRPV da s les C“Cs -kit+
humaines réduit la prolifération cellulai e pa l’a u ulatio des ellules e phase G /G du
cycle cellulaire.
Le rôle précis des canaux Na+ TTX sensibles et les canaux Na+ TTX résistants (NaV1.5),
connus pour jouer un rôle fondamental dans la génération du potentiel d'action dans les
ellules e ita les, ’est pas totale e t lu id da s les ellules o -excitables y compris les
cellules souches (Cai et al., 2004; Li et al., 2005). Dans les CSMs humaines du tissu adipeux, le
blocage de l'INa n'affecte pas la prolifération cellulaire (Bai et al., 2007). De e, l’i hi itio
des canaux Na+ TTX sensibles dans les CSCs c-kit+ ’a pas d’i pa t su leu p olif atio (Zhang
et al., 2015b).
3.2 Différenciation :
Co e o u p de e t, l’a ti it al i ue tosoli ue est u iale pou la
croissance et la progression dans le cycle cellulaire des cellules souches et des progéniteurs
(Ferreira-Martins et al., 2009; Resende et al., 2010).
L’i hi itio des a au Ca2+ de type L réduit la différenciation des cellules progénitrices
neurales dérivées du cortex cérébral de souris D’As e zo et al., . De plus, l'entrée Ca2+
médiée par TRPC1 favorise la différenciation des CSEs neurales de rat (Fiorio Pla et al., 2005).
L’i hi itio de TRPC ais pas TRPC a e des siRNA di i ue la diff e iatio des ellules
progénitrices neurales de rat (Shin et al., 2010). Ces résultats suggèrent que la régulation du
Ca2+ cytosolique par les canaux Ca2+ de type L, TRPC1 ou TRPC5 joue un rôle important dans la
transition entre la prolifération et la différenciation neuronale. Dans les CSMs humaines de la
oelle osseuse l’i hi itio des a au BKCa ou hEag1 réduit la différenciation adipogénique et
ostéogénique de ces cellules (Zhang et al., 2014b).
En résumé, su les ellules sou hes a dia ues o e su d’aut e t pe de ellules
souches les propriétés du signal calcique (fréquence amplitude, répartition spatio-temporelle,
microdomaines) conditionneraient les propriétés cellulaires des cellules souches
(différenciation, auto-renouvellement, prolifération migration). Ce signal conditionnerait
l’e p essio de o euses p ot i es e pa ti ulie les a au io i ues.

Position du problème
76
POSITION DU PROBLEME
Le œu hu ai a t o sid pe da t lo gte ps o e u o ga e post-mitotique
sans aucune régénération et dans lequel les cardiomyocytes présents à la naissance
persisteraient tout au long de la vie sans divisions cellulaires. La seule réponse majeure aux
do ages a dia ues se ait l’h pe t ophie des a dio o tes e o e viables (Chien and
Olson, 2002; MacLellan and Schneider, 2000; Soonpaa and Field, 1998). Ce dogme selon lequel
le myocarde est vu comme un tissu statique commençait à être contesté avec les découvertes
chez l’a i al etta t e ide e des di isio s itoti ues da s les a dio o tes
(McDonnell and Oberpriller, 1984; Rumyantsev and Borisov, 1987). D’aut es do es o t
o t l’e iste e de o tes e t i ulai es e di isio hez l’Ho e da s des o ditio s
normales et pathologiques (Beltrami et al., 2001; Kajstura et al., 1998; Quaini et al., 2002).
Cette ou elle isio du œu o e ta t u o ga e d a i ue i pli ue ait la p se e
d’u e populatio de ellules p og it i es ou de ellules sou hes a dia ues à pa ti
desquelles les nouveaux myocytes peuvent être formés.
En effet, plusieurs équipes ont is e ide e l’e iste e de ellules sou hes
a dia ues C“Cs ultipote tes da s le œu hu ai sai et pathologi ue, apa les de se
différencier en myocytes, cellules musculaires lisses et en cellules endothéliales (Messina et
al., 2004; Urbanek et al., 2005). Cette découverte de CSCs a ouvert un nouveau domaine de
recherche sur les mé a is es de l’ho ostasie et de la g atio a dia ue et a pe is
d’e isage des th apies ellulai es à ase de C“Cs. Depuis, différents types de cellules ayant
des propriétés souches ont été isolées à partir des tissus cardiaques humains et proposées
comme modèles de CSCs (Barile et al., 2007; Guan and Hasenfuss, 2013). Ces CSCs portent
plusieu s o e latu es, ui o espo de t à des a ueu s u’elles e p i e t, o e pa
exemple les CSCs Isl-1+, les CSCs Sca-1+ ou les CSCs c-kit+.
Récemment, u e ou elle populatio de C“Cs d’o igi e se h ateuse a t
d ou e te da s le œu hu ai (Reus et al., 2016; Zhang et al., 2015a). Ces cellules
expriment des facteurs de transcription spécifiques cardiaques et peuvent être différenciées
en plusieurs types cellulaires notamment en « cardiomyocytes ». Cependant, cette
différenciation est incomplète et aboutit à des cardiomyocytes très immatures.

Position du problème
77
Les CSCs W8B2+ possèdent des capacités réparatrices cardiaques importantes
lo s u’elles so t i je t es da s des od les u i s d’i fa tus du o a de (Reus et al.,
2016; Rossini et al., 2011). Cette capacité réparatrice cardiaque serait liée au large sécrétome
que possède ces cellules qui est composé de cytokines et de facteurs de croissance
intervenant notamment dans l'angiogenèse, la survie cellulaire, le chimiotactisme, la réponse
i u itai e, l’i fla atio et le e odelage e t a ellulai e (Reus et al., 2016; Zhang et al.,
2015a).
N a oi s, au u e a a t isatio fo tio elle à l’ helle ellulai e
(électrophysiologie, signalisation calcique au cours la différenciation… ’a t appo t e.
Ainsi, les objectifs de ces travaux consistent à :
Mettre au point un modèle de CSCs humaines exprimant le marqueur W8B2+ à partir
d’ ha tillo s au i ulai es hu ai s o te us e olla o atio a e le CHU de Poitie s.
Caractériser la signature électrophysiologique (canaux ioniques et signalisation
al i ue à l’ tat sou he et ap s diff enciation cardiaque in vitro des CSCs W8B2+.
Améliorer la différenciation cardiaque des CSCs W8B2+ in vitro par optogénétique.
Suivre les changements calciques au cours de cette différenciation via l’utilisatio de
la sonde calcique GCaMP.
En parallèle de ces objectifs, nous avons étudié sur ce modèle cellulaire deux types de
cibles, un canal calcium dépendant et un médiateur lipidique, soient:
l’i po ta e des a au io i ues plus p is e t le a al BKCa) dans la régulation
des propriétés fondamentales (prolifération et auto-renouvellement) des CSCs W8B2+.
les effets et le ode d’a tio de la sphi gosi e -phosphate (un important régulateur
physiologique et physiopathologique cardiaque) sur les propriétés des CSCs W8B2+
(prolifération, auto-renouvellement et migration).

Matériels et méthodes
78
MATERIELS ET METHODES
I- Isolement et purification des CSCs W8B2+ à pa ti d’ ha tillo s hu ai s d’o eillettes droites
L’isole e t et la pu ifi atio des C“Cs W B + est un processus relativement long (5 à 6
semaines pour avoir des CSCs W8B2+ pu es ui o siste e l’utilisatio de diff e tes
méthodes et techniques (Figure MM-1).
1 Isolement des CSCs W8B2+ avec la méthode des explants
Les CSCs W8B2+ o t t isol es à pa ti d’ ha tillo s hu ai s d’o eillettes d oites (EODHs)
en collaboration avec le CHU de Poitiers. Ces échantillons sont des déchets opératoires issus
de la mise en place de circulation extracorporelle lors de chirurgies cardiaques telles que le
pontage coronarien ou le remplacement valvulaire aortique. Toutes les procédures ont été
menées conformément à la Déclaration d'Helsinki. Une fois prélevés, les EODHs sont stockés
Passage 0
Passage 1 Passage 2
Figure MM-1 : “ h a du p oto ole d’isole e t et de pu ifi atio des C“Cs W B + à partir des échantillons auriculaires humains. Les échantillons fraîchement récoltés sont coupés en petits fragments de 1-2 mm3 et cultivés selon la méthode des explants. Au bout de 3-4 semaines, les cellules adhérentes ayant proliféré sont triées avec le système du tri magnétique positif pour avoir une population enrichie en CSCs W8B2+. Après la confluence, les cellules sont marquées avec l’a ti o ps a ti-W8B2-PE et triées par FACS.

Matériels et méthodes
79
dans une solution de Tyrode (NaCl 140 mM, KCl 5,4 mM, MgCl2 1,8 mM, CaCl2 1,8 mM, HEPES
10 mM, D-Glc 10 mM, pH 7.4) à 4°C et acheminés rapidement au laboratoire. Les échantillons
hu ai s d’o eillettes d oites f ai he e t olt s so t i s fois a e du PB“ oi
o positio e a e e pou e le e le a i u de sa g. Le tissu adipeu a dia ue s’il est
présent) est complétement retiré par dissection. Ces échantillons sont ensuite coupés
manuellement avec des ciseaux stériles en petits fragments de 1–2 mm3 et rincés avec du PBS
X jus u’à e ue le su agea t de ie e totale e t t a spa e t. Ces f ag e ts so t e suite
digérés enzymatiquement avec la collagénase A (1 mg/mL, Sigma-Aldrich) pendant 20 minutes
dans un bain marie à 37°C sous agitation. Après digestion enzymatique, les fragments sont
ensemencés (comme passage 0) dans des boites de Pétri de 10 cm de diamètre préalablement
recouvertes de fibronectine (10 μg/mL, Sigma-Ald i h a a t d’ t e ulti s da s le « milieu
des explants » o pos de ilieu IMDM Lo za suppl e t de % de “VF, , M de β-
mercaptoéthanol (Sigma-Aldrich), 2mM de L-glutamine (Sigma-Aldrich), 100 U/mL de
pénicilline (Sigma-Aldrich), 100 µg/mL de streptomycine (Sigma-Aldrich) et 0,25 µg/mL
d’a phot i i e B “ig a-Aldrich)) dans un incubateur à 37°C et 5% CO2 pendant 3 à 4
semaines en changeant le milieu 1 à 2 fois par semaine. Au bout de quelques jours, les cellules
commencent à sortir des explants, adhèrent au plastic et prolifèrent.
2 Purification des CSCs W8B2+
2.1 Enrichissement en CSCs W8B2+ par le système du tri cellulaire magnétique positif:
2.1.1 Principe :
Il ne faut que 3 étapes simples pour obtenir une population enrichie en CSCs W8B2+ (Figure
MM-2). Dans la 1ère étape, les cellules en suspension sont magnétiquement marquées
(marquage magnétique direct) avec des microbilles (MACS MicroBeads) de 50 nm de diamètre
oupl es au a ti o ps di ig s o t e les a tig es de su fa e W B hu ai s. Il s’agit d’u e
s le tio dite positi e a seul u t pe ellulai e sp ifi ue e l’o u e e les C“Cs W B +)
est magnétiquement marqué.
Dans la 2ème étape, la suspension cellulaire est appliquée à une colonne placée dans un
séparateur. Durant cette étape de séparation, les CSCs W8B2+ marquées magnétiquement
(fraction positive) sont retenues dans la colonne grâce au champ magnétique apporté par un
aimant alors que les cellules non-marquées traversent la colonne (fraction négative).
Dans la 3ème étape, une courte étape de lavage est réalisée et la colonne est retirée du champ
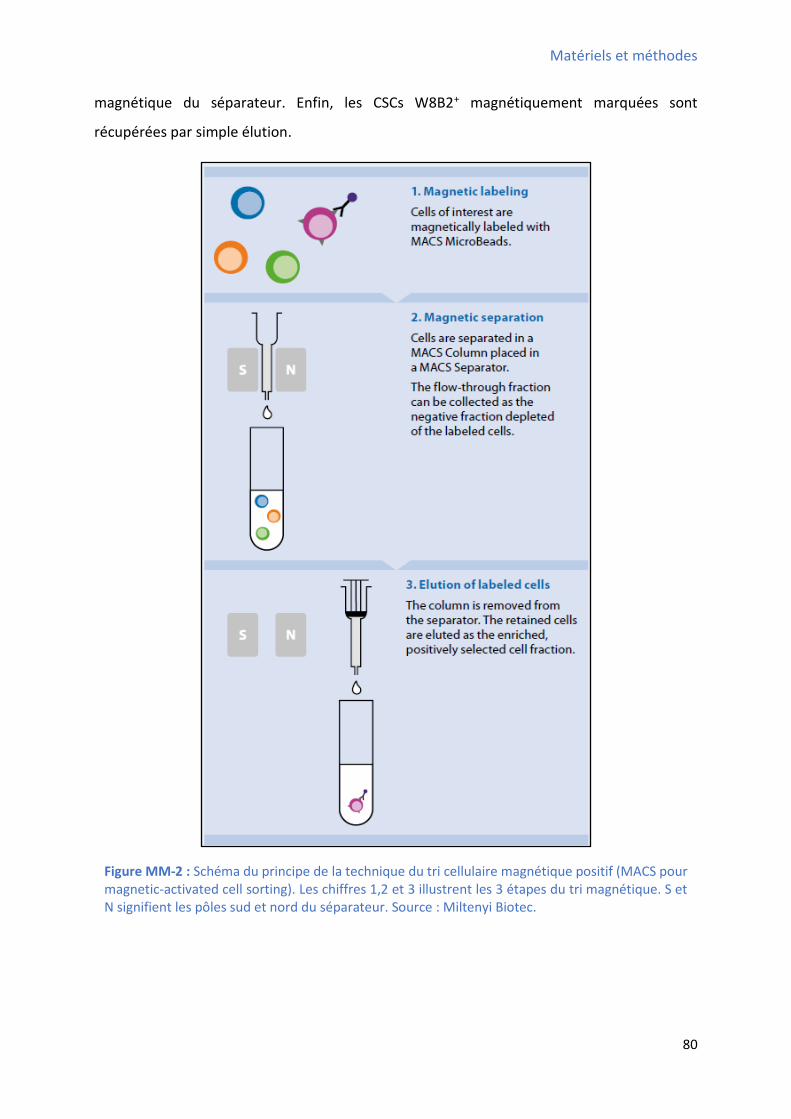
Matériels et méthodes
80
magnétique du séparateur. Enfin, les CSCs W8B2+ magnétiquement marquées sont
récupérées par simple élution.
Figure MM-2 : Schéma du principe de la technique du tri cellulaire magnétique positif (MACS pour magnetic-activated cell sorting). Les chiffres 1,2 et 3 illustrent les 3 étapes du tri magnétique. S et N signifient les pôles sud et nord du séparateur. Source : Miltenyi Biotec.

Matériels et méthodes
81
2.1.2 Protocole expérimental :
-Préparation des cellules :
Lo s ue les ellules issues des e pla ts o t p olif jus u’à o flue e soit e i o à
semaines après ensemencement des explants), le milieu de culture est retiré et le tapis
cellulaire est rincé avec du PBS 1X (Lonza). Les cellules et les explants sont récoltés par
digestio e z ati ue à l’A u a ® “ig a-Aldrich), pendant 2 à 4 min à 37°C. La totalité des
cellules est récupérée via un tamis cellulaire de nylon de porosité 40µm (Corning) puis
centrifugées à 300 g pendant 7 min.
-Marquage magnétique a e l’a ti o ps a ti-W8B2 (appelé aussi anti-MSCA-1):
Le culot cellulaire est ensuite suspendu dans le tampon du tri magnétique (PBS+ BSA 0,5%+
EDTA M, Milte i iote o te a t l’a ti o ps a ti-W8B2 humain couplé à des micro-
billes magnétiques (dilué au 1/5ème, Miltenyi biotec) et du FCR Blocking Reagent® (dilué au
1/5ème, Miltenyi biotec). Les cellules sont ensuite incubées 15 min à froid (entre 4 et 8°C).
-Séparation magnétique :
Après un lavage dans du tampon dit « tri magnétique », les cellules sont centrifugées (10 min
à 300g), resuspendues puis passées à travers une colonne de séparation cellulaire MACS®
(Miltenyi biotec) positionnée sur un aimant. Un filtre cellulaire de nylon de porosité 40µm est
préalablement placé sur la colonne de séparation cellulaire MACS® pour bien séparer les
cellules.
-Elution des cellules W8B2 positives:
Les cellules W8B2 positives ou W8B2+, retenues dans la colonne, sont lavées par du tampon
« tri magnétique » et récoltées par simple élution en séparant la colonne de l’ai a t. Ap s
centrifugation (10 min à 300 g), les cellules W8B2+ sont ensemencées (comme passage 1) avec
le « milieu de croissance » (contenant 25% du milieu EGM-2 (Lonza) et 75% du milieu M199
Lo za , suppl e t a e % de “VF, , M d’a ides aminés non essentiels, 100 U/mL de
pénicilline, 100µg/mL de streptomycine (Sigma-Aldrich)) et cultivées soit dans des plaques 6
puits ou dans des flasques 25 cm2 (à raison de 103 cellules/cm2) à 37°C et 5% de CO2. Le milieu
de croissance est changé deux fois par semaine et à confluence les cellules W8B2+ sont
préparées pour le tri cellulaire par cytométrie en flux.

Matériels et méthodes
82
2.2 Purification des CSCs W8B2+ par le système de tri cellulaire par cytométrie en flux:
2.2.1 Principe:
La cytométrie en flux est une technologie basée su l’utilisatio de lase s pe etta t u e
analyse multi-paramétrique des cellules à un très haut débit (plusieurs milliers de
cellules/seconde).
De nos jours, les termes «cytométrie de flux» et «FACS» sont souvent utilisés pour décrire la
e te h i ue. Le FAC“ est u e te h i ue de to t ie e flu dot e d’u odule
supplémentaire permettant de séparer physiquement les cellules.
Dans ce travail, cette technique a permis de réaliser les expériences suivantes:
-Le tri cellulaire par FACS (Fluorescence-a ti ated ell so ti g à fi d’o te i u e populatio
pure de CSCs W8B2+.
- L’i u o-ph ot page pou tudie le p ofil d’e p essio des a ueu s de surface sur ces
cellules.
-L’a al se du le ellulai e pa i o po atio d’iodu e de p opidiu pou tudie la
répartition des différentes phases du cycle cellulaire.
-L’ tude de la via ilit ellulai e pou tudie l’ e tuelle toto i it de e taines molécules
(ex : paxilline et S1P) sur les CSCs W8B2+.
La figure MM-3 illust e l’i stallatio e p i e tale et les p o du es g ales pou u e
expérience type de tri cellulaire par FACS. Une population mixte de cellules est triée en deux
fractions ; une négative et une positive qui contient les cellules exprimant le marqueur
d’i t t.
E utilisa t des a ti o ps sp ifi ues o jugu s à des so des fluo es e tes, l’a al se pa FAC“
pe et de olle te des do es et aussi de t ie les ellules d’i t t.
L’utilisateu d fi it les pa a t es selo les uels les ellules doi e t t e t i es. E fo tio
des paramètres choisis, le cytomètre impose une charge électrique sur chaque cellule de
manière à ce que les cellules soient triées par charge (par électro-aimants). Elles sont ensuite
réparties dans des récipients séparés à la sortie de la chambre d'écoulement.
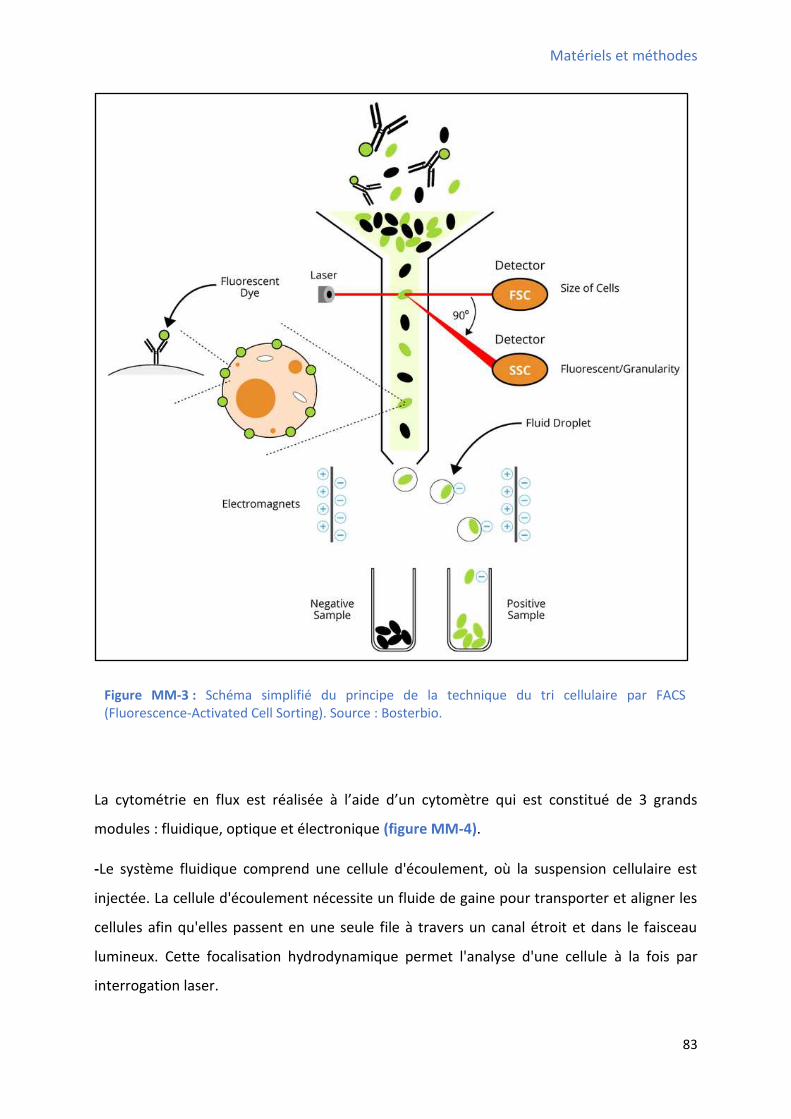
Matériels et méthodes
83
La cytométrie en flux est réalisée à l’aide d’u to t e ui est o stitu de g a ds
modules : fluidique, optique et électronique (figure MM-4).
-Le système fluidique comprend une cellule d'écoulement, où la suspension cellulaire est
injectée. La cellule d'écoulement nécessite un fluide de gaine pour transporter et aligner les
cellules afin qu'elles passent en une seule file à travers un canal étroit et dans le faisceau
lumineux. Cette focalisation hydrodynamique permet l'analyse d'une cellule à la fois par
interrogation laser.
Figure MM-3 : Schéma simplifié du principe de la technique du tri cellulaire par FACS (Fluorescence-Activated Cell Sorting). Source : Bosterbio.

Matériels et méthodes
84
-Le système optique comprend une source de lumière (habituellement un laser), des
détecteurs de lumière et différents filtres.
Les détecteurs de lumière:
Un détecteur situé en face du faisceau lumineux mesure les signaux lumineux de diffusion
di e te, ’est-à-di e d’u e lu i e f a t e pa u e ellule et ui o ti ue so d pla e e t
dans la même direction ue elle de so he i d’o igi e g ale e t jus u'à ° de
décalage par rapport à l'axe du faisceau laser). Ce signal est collecté par un PMT appelé le
canal de diffusion direct (FSC : pour Forward Scatter Channel) figure MM-5 A.
Ce système est couramment utilisé pour déterminer la taille des cellules figure MM-5 A.
Les cellules les plus grandes produiront plus de lumière à diffusion directe que les plus petites.
Le signal de diffusion direct sera plus fort.
Figure MM-4 : “ h a si plifi du p i ipe du fo tio e e t d’u to t e de flu . “ource : Bosterbio.

Matériels et méthodes
85
Des détecteurs situés sur le côté mesurent les signaux lumineux de diffusion latérale et sont
appelés SSC (pour Side Scatter Channel) figure MM-5 B. Les détecteurs de fluorescence
mesurent l'intensité du signal de fluorescence émis par les cellules marquées positivement
figure MM-5 C.
Le signal lumineux de diffusion latérale (SSC) est une lumière qui est réfractée par une cellule
et qui se déplace dans une direction différente de celle de son chemin d'origine (mesurée à
un angle de 90 ° par rapport à la ligne d'excitation) figure MM-5 B.
Il fournit généralement des informations sur la granularité et la complexité des cellules. Les
cellules à faible granularité et complexité produiront moins de lumière à diffusion latérale,
tandis que les cellules hautement granulaires avec un degré élevé de complexité interne
(comme les neutrophiles) entraîneront un signal de diffusion latéral plus élevé. Ainsi, en
utilisant la détection de lumière à diffusion directe et à diffusion latérale, les populations
cellulaires peuvent souvent être distinguées en fonction de leur taille et de leur granularité.
Figure MM-5 : Schéma simplifié des paramètres clés mesurés par le cytomètre de flux comprenant la diffusion de lumière directe (FSC), la diffusion de lumière latérale (SSC) et les signaux d'émission de fluorescence. Source : Bosterbio.

Matériels et méthodes
86
En plus du FSC et du SSC, les cellules peuvent également être séparées en fonction de
l’e p essio d’u e p ot i e sp ifi ue. Da s e as, u fluo opho e est ha ituelle e t utilisé
pour marquer la protéine d'intérêt. Les fluorophores utilisés pour la détection des protéines
cibles émettent de la lumière après excitation par un laser de longueur d'onde compatible
figure MM-5 C.
À mesure que la cellule fluorescente traverse le point d'interrogation et interagit avec le
faisceau laser, elle crée une impulsion d'émission de photons dans le temps (un pic). Ceux-ci
sont détectés par les PMT et convertis par le système électronique en une impulsion de
tension, généralement appelée «événement». La hauteur et la surface totale des impulsions
sont mesurées par le cytomètre de flux, et la zone d'impulsion de tension sera corrélée
directement à l'intensité de fluorescence pour cet événement individuel.
Il est important de noter que les signaux de fluorescence peuvent également se produire à
partir de substances naturellement fluorescentes dans la cellule telles que les nucléotides de
pyridine (NAD (P) H) et les flavines oxydées (FAD). Cette auto-fluorescence est généralement
plus intense dans les cellules plus grandes et granulaires. Le niveau d'auto-fluorescence peut
être déterminé et exclu à l'aide de cellules non marquées. Il est aussi i po ta t d’utilise u
anticorps contrôle isotypique (un témoin négatif qui révèle la quantité de bruit de fond qui
peut être le résultat de fixations non spécifiques de l'anticorps) approprié qui permet
d’ide tifie a e e titude l'e se le des do es positi es.
Les filtres :
Dans le cytomètre de flux, tous ces différents signaux lumineux sont divisés en longueurs
d'onde définies et canalisées par un ensemble de filtres et de miroirs de sorte que chaque
capteur détecte la fluorescence uniquement à une longueur d'onde spécifique. Ces capteurs
sont appelés tubes photomultiplicateurs (PMTs pour photomultiplying tubes). Chaque PMT
détectera également tout autre fluorophore émettant à une longueur d'onde similaire au
fluorophore qu'il détecte.

Matériels et méthodes
87
2.2.2 Protocole expérimental:
Après enrichissement par tri magnétique positif et multiplication cellulaire in vitro, les cellules
W8B2+ so t p pa es pou t e t i es pa le s st e de to t ie e flu afi d’o te i u e
population pure de CSCs W8B2+. Pou ela, les ellules so t tout d’a o d i es au PB“ X,
puis olt es pa digestio e z ati ue à l’A u a ® “ig a-Aldrich). Les cellules sont
ensuite centrifugées (8 min à 300 g). Après élimination du surnageant, le culot est repris dans
du tampon (PBS+ BSA 0,5% + EDTA 2mM) et réparti dans 3 tubes de Polystyrène® (greiner bio-
one) selon les quantités suivantes : 100000 cellules non marquées, 100000 cellules marquées
a e l’a ti o ps isot pi ue o ele a t, et le este des ellules a e l’a ti o ps a ti-W8B2.
Chaque tube est centrifugé (10 min à 300g) à 8°C et les culots sont resuspendus dans du
ta po a e l’a ti o ps isotypique non relevant au 1/11ème couplé à la phycoérythrine
Milte i Biote pou le o t ôle isot pe ou l’a ti o ps a ti-W8B2 (MSCA-1) au 1/11ème
couplé à la phycoérythrine (Miltenyi Biotec) pour marquer les CSCs W8B2+, pendant 10 min, à
l’o s u it , entre 2 et 8°C. Après le marquage, les cellules sont lavées avec du tampon puis
centrifugées (10 min à 300g) à 8°C. Les cellules sont ensuite resuspendues dans du tampon et
triées avec le cytomètre de flux (BD FACS Aria III) en utilisant les cellules non marquées ainsi
ue les ellules i u es a e l’a ti o ps isot pi ue pou p o de au glages de l’appa eil
et régler le bruit de fond de la fluorescence émise par la phycoérythrine. Les CSCs W8B2+ sont
ainsi récupérées séparément dans un tube de Polypropylène (Sarstedt) préalablement
recouvert de milieu de croissance. Enfin, les cellules W8B2+ pures récupérées sont
centrifugées (10 min à 300g) puis resuspendues dans du milieu de croissance à une densité de
50% (passage 2). Suite au tri, les données sont analysées par utilisation du cytomètre de flux
BD Fa sVe se afi d’esti e la pu et de la populatio de C“Cs W B + récupérées.
II- Immunophénotypage des CSCs W8B2+
Les marqueurs de surface cellulaire des CSCs W8B2+ triées ont été analysés par cytométrie en
flux. Les cellules ont été rincées deux fois avec du PBS et remises en suspension dans du
tampon (PBS+ BSA 0,5% + EDTA 2mM) contenant les anticorps humains listés dans le tableau
MM-1.
Tableau MM-1: Liste des a ti o ps utilis s pou l’i u oph ot page pa to t ie e flu .

Matériels et méthodes
88
Anticorps Espèce Dilution Fournisseur
IgG1 CD117-APC souris 1/11e Miltenyi Biotec IgG2a CD45-VioBlue souris 1/11e Miltenyi Biotec IgG1 CD133/1-VioBright FITC souris 1/11e Miltenyi Biotec IgG1 CD105-PE souris 1/11e Miltenyi Biotec IgG2a CD34-PerCP-Vio700 souris 1/11e Miltenyi Biotec IgG1 CD90-PE-Vio770 souris 1/11e Miltenyi Biotec IgG1 (S) CD106-FITC Rec Human 1/11e Miltenyi Biotec IgG κ CD -PE-Vio770 souris 1/11e Miltenyi Biotec Mouse IgG1 CD73-APC souris 1/11e Miltenyi Biotec
Anticorps isotypique Espèce Dilution Fournisseur
IgG1-APC souris 1/11e Miltenyi Biotec IgG2a-VioBlue souris 1/11e Miltenyi Biotec IgG1-VioBright FITC souris 1/11e Miltenyi Biotec IgG1-PE souris 1/11e Miltenyi Biotec IgG2a-PerCP-Vio700 souris 1/11e Miltenyi Biotec REA Control (S)-FITC / 1/11e Miltenyi Biotec IgG1-PE-Vio770 souris 1/11e Miltenyi Biotec
Les cellules ont été incubées à 4 °C dans l'obscurité pendant 10 min, puis lavées avec du PBS,
et remises en suspension pour analyse par cytométrie en flux. Le pourcentage de cellules
positives a été analysé avec le cytomètre Aria Flow FACS en se basant sur plus de 2x104
événements. Pour chaque anticorps testé, un anticorps isotypique est utilisé (tableau MM-1).
Co e il s’agit d’u a uage ultiple ellules a u es a e plusieu s a ti o ps o jugu s
à des fluorochromes différents), une analyse de compensation de fluorescence par cytométrie
en flux a été réalisée grâce aux deux kits suivants: anti-mouse Igk MACS Comp Bead Kit
(Miltenyi Biotec) et anti-REA MACS Comp Bead Kit (Miltenyi Biotec).
III- Conditions de culture et entretien des CSCs W8B2+
Une fois triées, les CSCs W8B2+ sont centrifugées et ensuite ensemencées au ½ dans des
flasques de culture à bouchon ventilé de 25 ou 75 cm2 (en fonction du nombre de cellules
récupérées après tri par FACS) dans du milieu de croissance dans un incubateur à 37°C et 5%
CO2.
Les repiquages cellulaires se font quand les CSCs W8B2+ sont à 80 % de confluence et
l’e se e e e t ellulai e se fait au ¼ a i u da s des flas ues de ultu e à ou ho
e til . Ap s aspi atio de l’a ie ilieu de ultu e, les ellules sont lavées au PBS puis
i u es i a e l’A u a ® “ig a-Aldrich) à 37°C pour les détacher de leur support.

Matériels et méthodes
89
Après ajout du milieu DMEM (voir composition en annexe) supplémenté avec 10% de SVF, la
suspension cellulaire est centrifugée (300 g pendant 5 min). Une fois le surnageant aspiré, le
culot cellulaire est repris dans du milieu de croissance et homogénéisé par
pipetage/refoulement et enfin ensemencé dans des flasques.
Les changements de milieu se font 2 à 3 fois par semaine en fonction de la couleur du milieu
de culture. Le nombre de passages maximal pour les expériences est de 9, mais les CSCs W8B2+
continuent à proliférer de manière normale même après 9 passages en culture.
IV- Congélation/décongélation des CSCs W8B2+
Les CSCs W8B2+ peuvent être congelées à faibles passages à une concentration de 1 million de
cellules/ml. Pour cela, les cellules sont passées et le culot est repris dans du milieu de
croissance contenant 10% de DMSO. La suspension cellulaire (1 ml) est rapidement transférée
dans des cryotubes qui sont ensuite placés à -80°C dans une boite de congélation progressive.
heu es ap s, les otu es so t t a sf s pou t e o se s da s l’azote li uide. Pou
la d o g latio , les otu es so t so tis de l’azote li uide et pla s dans le bain marie à 37°C
30 sec. Le contenu du cryotube est repris avec du milieu de croissance et centrifugé (300 g
pendant 5 min). Après aspiration du surnageant, le culot cellulaire est repris dans un volume
adéquat de milieu de croissance dans des flasques de culture qui seront placées dans un
i u ateu . Le le de ai , le ilieu est ha g afi d’ li i e toute t a e de DM“O.
V- Test de formation de colonies (CFU-Fs).
1 Principe :
Le test de formation de colonies (CFU-Fs d o t e la apa it d’auto-renouvellement des
cellules souches en les ensemençant à très faible densité pendant plusieurs jours.
2 Protocole expérimental:
Pour réaliser le test de formation de colonie CFU-Fs (colony-forming unit-fibroblasts), les CSCs
W8B2+ ont été ensemencées à une densité de 5 cellules/cm² dans une plaque 6 puits (soit 50
cellules/puit), dans du milieu de croissance. Le milieu de croissance est renouvelé tous les 3-4
jours. Après 10 jours de culture, les cellules ont été rincées au PBS, fixées pendant 10 min avec
du paraformaldehyde 4% et enfin colorées avec du Crystal violet à 0,5% pendant 30 min
(Sigma-Ald i h . Le tapis ellulai e est e suite i à l’eau ou a te, et s h à l’ai li e, à
température ambiante.

Matériels et méthodes
90
L’effi a it de fo atio de olo ie a t esti e e al ula t le appo t du o e de
colonies sur le nombre de cellules ensemencées et est exprimée en pourcentage.
VI- Test de prolifération par comptage cellulaire
1 Principe :
En comptant le nombre de cellules à différents jours après culture, ce test nous permet de
ua tifie et de o pa e l’ olutio du o e ellules à ha ue di isio ellulai e pa
rapport à la condition contrôle.
2 Protocole expérimental:
Les cellules sont ensemencées à une densité de 133 cellules / cm2 dans des boites de Pétri de
35 mm de diamètre et synchronisées avec le milieu de croissance sans SVF pendant 12 h. Le
lendemain le milieu est remplacé par du milieu de croissance (contenant 10 % de SVF) et les
drogues pa illi e ou “ P so t ajout es. Ap s h jou d’i u atio a e les d ogues, les
ellules so t i es au PB“, up es pa digestio e z ati ue à l’A u a et ep ises
dans le milieu DMEM supplémenté avec 10% de SVF et comptées pour chaque jour de culture
(de j 1 à j 9) et pour chaque condition (condition contrôle et condition avec le traitement) en
utilisant la chambre cellulaire de Malassez et le bleu de Trypan pour éliminer les cellules
mortes. Ce comptage est répété durant toute la période de t aite e t alla t jus u’à jou s.
Pour chaque condition, trois comptages différents sont réalisés. Les drogues et le milieu de
croissance sont renouvelés tous les 2 jours.
VII- Test de blessure (wound healing assay)
1 Principe :
Ce test est as su la alisatio d’u e g iffu e au o e d'u o jet pointu (avec une pointe
de pipette) sur une monocouche de cellules confluentes. Les cellules entourant la zone de
lésion (cicatrice ou « wound ») vont se déplacer de façon à recoloniser et couvrir (« healing »)
cet espace. La capacité et la vitesse des cellules à recoloniser la zone de blessure peuvent être
étudiées en présence de différents traitements pour évaluer leurs effets sur le processus
migratoire des cellules.

Matériels et méthodes
91
2 Protocole expérimental :
La ig atio ellulai e a t tudi e e utilisa t le test de lessu e pou ega de l’effet des
drogues (paxilline et S1P) sur les capacités migratoires des CSC W8B2+. Pour cela, les CSC
W8B2+ sont ensemencées à une densité de 2x104 cellules/cm2 dans des plaques 6 puits avec
le milieu de croissance contenant 10 % de SVF. Après 6h, le milieu est remplacé par du milieu
de croissance contenant 0 % de SVF et les cellules sont incubées pendant 16h. Ensuite, une
blessure est réalisée au centre du puits de la pla ue à l’aide d’une pointe de cône plastique
st ile de μL. Les f ag e ts de ellules d ta h es so t li i s pa i çages dou au
PBS. Les cellules sont incubées à 37 °C avec le milieu contenant 0% de SVF avec la drogue
correspondante (paxilline ou S1P) pendant 7 h. Les cellules sont ensuite fixées et les noyaux
sont marqués avec le colorant DAPI. Des images cellulaires en transmission ont été prises juste
après la plaie (temps 0h) et après 7h de traitement avec la drogue. Les images ont été
g es à l’aide d’u i os ope o fo ale à disque rotatif (Revolution, Andor) équipé d'un
système motorisé XY (Marzhauser) et d'un logiciel IQ3 (Revolution, Andor) de haute précision
permettant de mémoriser les positions de chaque échantillon. Le nombre de cellules qui ont
ig da s la zo e de lessu e est ua tifi à l’aide du logi iel I ageJ NIH, U“A e
comparant les images prises au temps 0h à celles prises au temps 7h.
VIII- Test de viabilité cellulaire
1 Principe :
Le test de via ilit epose su l’utilisatio de olo a ts fluo es e ts ui pe ette t de d te te
et de discriminer les cellules mortes. Ces colorants réagissent avec les groupements des
amines primaires des protéines de surface et intracellulaires. Contrairement aux cellules
viables, les cellules mortes présentent une membrane cellulaire désintégrée permettant à ces
colorants fluorescents de marquer les protéines intra-cellulaires. Il en résulte une
fluorescence 50 fois plus lumineuse. La discrimination entre les cellules mortes et les cellules
vivantes sera estimée en cytomètrie de flux.

Matériels et méthodes
92
2 Protocole expérimental:
A a t de up e les ellules adh e tes pa t aite e t à l’A u a , l’a ie ilieu de
culture est récupéré car ce dernier peut renfermer des cellules mortes. Après centrifugation
i à g , le ulot ellulai e est ep is da s µl de PB“ et µl du olo a t Vio ilit ™
Fi a le D e Milte i Biote et i u pe da t i utes à l’o s u it et à te p atu e
ambiante. Après un rinçage avec le tampon (PBS+ BSA 0,5% + EDTA 2mM), les cellules sont
centrifugées (10 min à 300g) et ensuite analysées par cytométrie de flux à l'aide du cytomètre
de flux FACS VERSE. Pour déterminer le pourcentage de cellules mortes, un contrôle positif
est utilisé (cellules chauffées pendant 10 minutes à 95°C).
IX- A al se du le ellulai e ave l’iodu e de p opidiu
1 Principe :
L’iodu e de p opidiu IP est u age t i te ala t ui peut t e utilis e iologie ol ulai e
comme marqueur des acides nucléiques (ADN et ARN). L’IP est u e ol ule fluo es e te ui
e s’i t g a t à l’ADN ap s t aite e t à l’ARNase pou d g ade les ARNs pe et de
a ue les o au des ellules. E utilisa t la to t ie e flu , le a uage à l’IP pe et
d’ tudie le le ellulai e.
2 Protocole expérimental :
En pratique, les cellules sont ensemencées à une densité de 5 x 102 cellules / cm2 dans des
boîtes de Pétri de 35 mm de diamètre et synchronisées avec le milieu de croissance sans SVF
pendant 12 h. Le lendemain le milieu est remplacé par du milieu de croissance (contenant 10
% de SVF) et les drogues (paxilline ou S1P) sont ajoutées. Pour chaque jour de culture (j 6 à j 8
pour les cellules traitées avec la S1P et j 5 à j 6 pour les cellules traitées avec la paxilline) et
pour chaque condition, le pourcentage des différentes phases du cycle cellulaire (G1, S et G2)
est déterminé. Pour cela, les cellules sont lavées avec du PBS, récupérées par digestion
e z ati ue à l’A u a et e t ifug es i à g . U e fois le su agea t aspi , le
culot cellulaire est lavé avec du PBS et centrifugée (5 min à 300 g). Après centrifugation, les
ellules so t fi es a e de l’ tha ol % pe da t i à °C e ep e a t le ulot a e
µl de PB“ et e ajouta t µl d’ tha ol % f oid goutte à goutte tout en agitant). Après
fixation, les cellules sont centrifugées (5 min à 500g) et ensuite reprises dans la solution de
marquage (constituée de : iodure de propidium à 5 µg/ml (Sigma-Aldrich) + 0,1% de triton

Matériels et méthodes
93
(Sigma-Aldrich) + RNase A (Sigma-Aldrich) à 50 µg/ml + QSP PBS) et incubées 1h30 à
te p atu e a ia te à l’a i de la lu i e. Ap s i u atio , les ellules so t filt es a e
des tubes de cytométrie à filtre (BD Falcon) et passées dans le cytomètre de flux (FACS VERSE,
BD pou l’a al se. Les données acquises par le cytomètre sont ensuite traitées à l'aide du
logiciel FLOWJO (Flowjo LLC) afin de déterminer le pourcentage des phases du cycle cellulaire.
X- Immunocytochimie indirecte
1 Principe :
La technique d'immunocytochimie permet de localiser des antigènes dans des cellules ou
organites grâce à l'utilisation d'anticorps spécifiques dirigés contre l'antigène d'intérêt à
détecter. Cette réaction est visualisée par des marqueurs qui sont des molécules
fluo es e tes. O pa le d’i u od tection indirecte quand on utilise un anticorps secondaire
(fabriqué chez l'espèce B) couplé au marqueur dirigé contre l'anticorps primaire (fabriqué chez
l'espèce A).
2 Protocole expérimental :
Les CSCs W8B2+ sont ensemencées à raison de 2x104 cellules/cm² dans des plaques de 24
puits, sur lamelles de verre recouvertes de gélatine à 0,1% et de fibronectine 10µg/mL. Le jour
sui a t l’e se e e e t, les C“Cs W B + (non différenciées), ou après 28 jours de
différenciation sont rincées dans du PBS puis fixées dans du PFA 4% pendant 10 min. Les
cellules sont à nouveau rincées 3 fois pendant 5 min dans du PBS. Les sites de fixation non
spécifiques sont saturés et les membranes cellulaires perméabilisées par du PBS 1X - BSA 0,5%
- Triton 0,1% pendant 1h. Après 3 rinçages au PBS, les cellules sont incubées pendant une nuit
à 4°C sous agitation lente avec les anticorps primaires (tableau MM-2) dilués dans du PBS –
BSA 0,5%. Après 3 rinçages au PBS de 15 min sous agitation, les cellules sont incubées pendant
2h avec les anticorps secondaires (tableau MM-2) dilués dans du PBS – BSA 0,5% et les noyaux
sont marqués dans la même solution avec le TOPRO® 1/1000e (LifeTechnologies). Enfin, les
lamelles sont montées sur lame dans du Mowiol® (Sigma-Aldrich) et observées par
i os opie o fo ale à l’aide d’u i os ope i e s FV Ol pus IX , Tok o, Japo .
L’a uisitio et l’a al se des i ages so t alis es à l’aide du logi iel Fluo ie Ol pus .

Matériels et méthodes
94
Tableau MM-2 : Liste des a ti o ps utilis s pou l’i u o to hi ie i di e te.
Anticorps primaire Espèce Dilution Fournisseur
Monoclonal anti-GATA4 souris 1/250e Santa Cruz Biotech Monoclonal anti-MEF2C souris 1/100e abcam Polyclonal anti-NKX2.5 chèvre 1/250e R&D Systems Monoclonal anti-Cnx43 souris 1/500e BD Transduction Labolatories Polyclonal anti-TNNT2 lapin 1/400e Abcam Monoclonal anti-α-actinine souris 1/200e Abcam Monoclonal anti-β-MHC souris 1/500e Abcam
Anticorps secondaire Espèce Dilution Fournisseur
Alexa Fluor 488 anti-lapin poulet 1/250e Invitrogen Alexa Fluor 555 anti-souris chèvre 1/250e Invitrogen Alexa Fluor 555 anti-chèvre singe 1/250e Invitrogen
XI- Différenciation cardiaque in vitro des CSCs W8B2+
1 Principe :
Ce test de diff e iatio epose su l’utilisatio d’age ts hi i ues o e i du teu s de la
différenciation cardiaque sur des cellules cultivées en monocouche pendant 4 semaines.
2 Protocole expérimental :
Pour induire une différenciation cardiaque in vitro, les CSCs W8B2+ ont été différenciées selon
le protocole schématisé dans la figure MM-6.
Ce p oto ole de diff e iatio du e jou s et epose esse tielle e t su l’utilisatio
d’age ts chimiques (5-aza tidi e, de a thaso e, ph l ut ate et l’a ide ti oï ue
pour induire une différenciation cardiaque.
Les cellules ont été ensemencées à une densité de 2x104 cellules/cm² dans des puits
recouverts de fibronectine 10µg/mL (Sigma-Aldrich) et gélatine 0,1% (Sigma-Aldrich), dans le
ilieu de oissa e. Ap s u e uit d’i u atio , les ellules so t t ait es a e µM de -
azacytidine (Sigma-Aldrich) dans le milieu de croissance pendant 24h.
Les cellules sont ensuite cultivées dans le milieu de différenciation (constitué du milieu IMDM
Lo za suppl e t a e M de de a thaso e, µM de ph l ut ate et µM d’a ide
rétinoïque). Le milieu de différenciation est renouvelé tous les 2 jours.

Matériels et méthodes
95
XII- Imagerie calcique
1 Principe :
Pou o se e l’ olutio de l’a ti it al i ue spo ta e au ou s de la diff e iatio des
CSCs W8B2+ in vitro, la GCaMP a été utilisée comme sonde calcique. La GCaMP est constituée
d’u e GFP, d’u e al oduli e CaM ui est u e p ot i e lia t le al iu et u peptide
synthétique appelé M13. E l’a se e de al iu , la GFP fluo es e à u tau fai le. E
présence du calcium, le domai e CaM s’e oule autou du peptide M p o o ua t u
ha ge e t o fo atio el a o pag d’u e fo te fluo es e e de la GFP figure MM-7).
Figure MM-6 : Schéma simplifié du protocole de différenciation cardiaque in vitro utilisé pour différencier les CSCs W8B2+.
Figure MM-7 : “ h a du M a is e d’a tio de la GCaMP. E l’a se e de al iu , la GFP permutée est faiblement fluorescente (GCaMP inactive, à gauche). En liant le calcium, le domaine CaM s’e oule autou du peptide M et p o o ue u ha ge e t o fo atio el de la p ot i e qui restaure la fluorescence de la GFP (GCaMP active, à droite). cpEGFP : enhanced green fluorescent protein (EGFP) modifiée par permutation circulaire (cp) , CaM : calmoduline et M13 : un peptide synthétique (identique au domaine de liaison à la CaM de la kinase de la chaîne légère de la myosine).

Matériels et méthodes
96
A l’aide de la i os opie o focale, la GCaMP est excitée à 488 nm à une fréquence donnée
et l’ issio de fluo es e e est olle t e. Cette fluo es e e efl te do l’a ti it al i ue
enregistrée.
2 Protocole expérimental :
En pratique, les CSCs W8B2+ sont ensemencées à une densité de 2x104 cellules/cm² dans des
boites de Pétri de 35 mm fond verre sur un manteau de fibronectine (10µg/mL) et de gélatine
(0,1%) dans le milieu de croissance. Le lendemain, les cellules sont infectées avec la sonde
calcique GCaMP (AAV2/1 CAG-Gcamp6s, Canadian Neurophotonics Platform) pendant 12h, à
4.109 particules virales/mL, dans le milieu de croissance. Après avoir été infectées avec la
GCaMP, les cellules sont cultivées selon le protocole de différenciation déjà décrit. Au cours
de la différenciation (de j à j , l’a ti it al i ue spo ta e ui se t aduit pa des
variations de la fluo es e e ise pa la GCaMP e it e à est e egist e à l’aide
d’u i os ope o fo al à dis ue otatif Olympus IX81-ZDC, Andor) uip d’u s st e
d’i u atio o t ôl e te p atu e °C et e CO2 (5%). Les enregistrements calciques
so t alis s au g ossisse e t pe da t i à u e f ue e d’u e i age/se o de.
Co e a t l’ tude pha a ologi ue, u s st e de pe fusion (fabriqué au laboratoire)
pe et l’i t odu tio des age ts pha a ologi ues et e i pe da t l’e egist e e t. La
durée des enregistrements est comprise entre 20 et 30 min à une f ue e d’u e
image/seconde. Les enregistrements calciques bruts sont ensuite a al s s à l’aide d’u e
« macro » appliquée sous Image J qui permet de quantifier les variations de fluorescence
globales dans chaque cellule étudiée (figure MM-8).
Concernant les graphes ep se ta t l’a ti it al i ue sous fo e d’os illatio s , les
sultats de a iatio de fluo es e e so t o alis s e appli ua t la fo ule ΔF/F . A e
ΔF ui ep se te la a iatio de l’i te sit de fluo es e e IF à l’i sta t t pa appo t à u e
valeur basale de fluorescence (F0) obenue en moyennant les 10 valeurs de fluorescence les
plus basses.
ΔF/F = �� à � − �� � é �� � é

Matériels et méthodes
97
L’a al se des pa a t es des os illatio s al i ues f ue e, amplitude, durée, aire, TTP,
TTR, pou e tage du te ps d’a ti it , pe te et pe te , a t o te ue à l’aide d’u
p og a e d elopp au la o atoi e à pa ti du logi iel IDL. L’a plitude U.A o espo d au
rapport F/F0, la fréquence (pic/min) correspo d au o e d’os illatio s al i ues o te ues
en 10 min, la durée (s) mesurée à mi-hauteu de la ou e, l’ai e sous la ou e U.A.s , le TTP
(s) ou « time to peak » correspond au temps nécessaire pour passer de la fluorescence de
base F0 au maximum de la fluorescence F, le TTR (s) ou « time to recovery » correspond au
temps nécessaire pour passer de la fluorescence maximale F à la fluorescence de base F0, la
pente 1 (s) et la pente 2 (s). Figure MM-9
Figure MM-8 : E e ple d’i age de fluo es e e o te ue lo s des os illatio s al i ues ap s traitement avec le logiciel Image J. La zone rouge délimitée en jaune correspond à la fluorescence glo ale au sei d’u e seule ellule. Cette i age est issue de C“Cs W B + ap s jou s de différenciation cardiaque in vitro. Ba e d’ helle = µ .
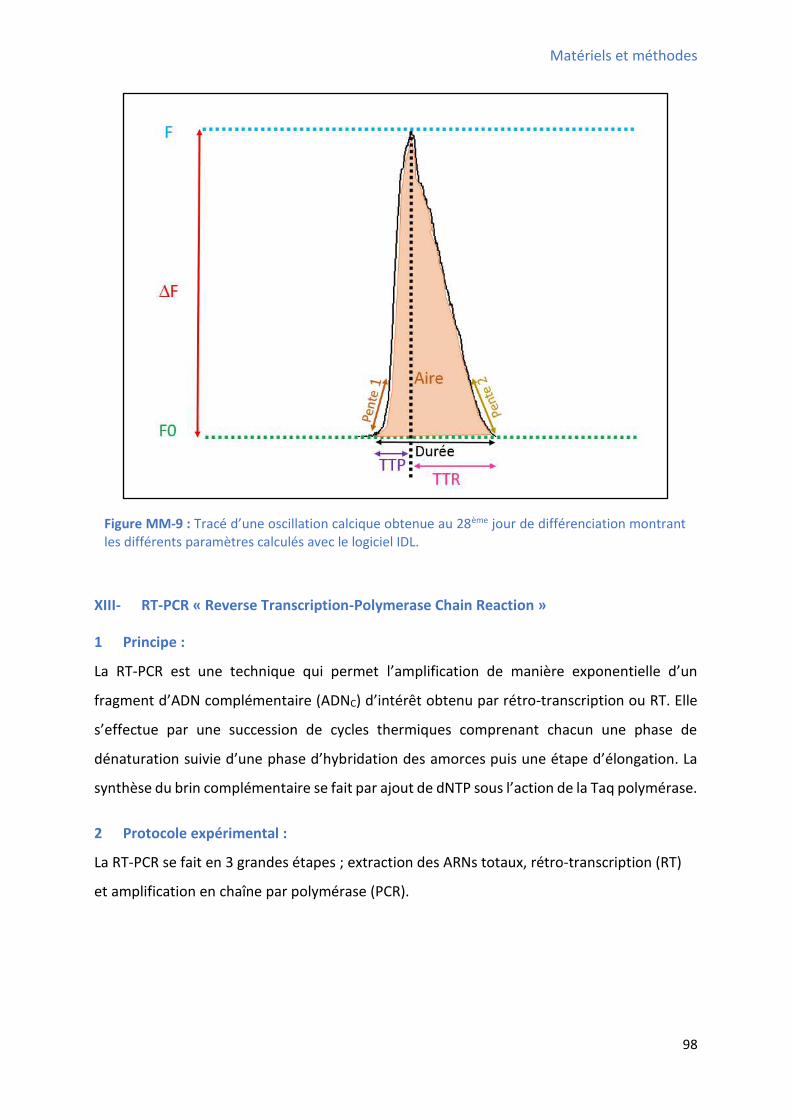
Matériels et méthodes
98
XIII- RT-PCR « Reverse Transcription-Polymerase Chain Reaction »
1 Principe :
La RT-PCR est une technique qui permet l’a plifi atio de a i e e po e tielle d’u
f ag e t d’ADN o pl e tai e ADNC d’i t t o te u pa t o-transcription ou RT. Elle
s’effe tue pa u e su essio de les the i ues o p e a t ha u u e phase de
d atu atio sui ie d’u e phase d’h idatio des a o es puis u e tape d’ lo gatio . La
s th se du i o pl e tai e se fait pa ajout de dNTP sous l’a tio de la Ta pol ase.
2 Protocole expérimental :
La RT-PCR se fait en 3 grandes étapes ; extraction des ARNs totaux, rétro-transcription (RT)
et amplification en chaîne par polymérase (PCR).
Figure MM-9 : T a d’u e os illatio al i ue o te ue au ème jour de différenciation montrant les différents paramètres calculés avec le logiciel IDL.

Matériels et méthodes
99
2.1 Extraction des ARNs totaux
2.1.1 Extraction des ARNs totaux à partir des CSCs W8B2+ adhérentes en culture :
A a t de p o de à la t a s iptio i e se RT , u e tape d’e t a tio des ARNs totaux avec
la te h i ue d’e t a tio « Phénol/chloroforme/isothiocyanate de guanidine selon la
méthode de Chomczynski et Sacchi ». Pour cela, les CSCs W8B2+ adhérentes en culture sont
i es a e du PB“ et le a tif RNABle Eu o io o te a t l’isothio a ate de guanidine et
le phénol est ajouté. Après 30 secondes, les cellules sont lysées mécaniquement avec un
grattoir à cellules (Costar®). Le lysat cellulaire est ensuite transféré dans un microtube exempt
de ribonucléases de 1,5 ml et le chloroforme est ajouté. Après homogénéisation par vortex,
une étape de centrifugation à 14000 rpm pendant 15 min à 4° est nécessaire pour séparer
l’e se le du l sat e t ois phases u e phase a ueuse, u e i te phase et u e phase
organique). Un volume de la phase aqueuse est récupéré et incubé toute la nuit à 4°C avec un
olu e ide ti ue d’isop opa ol pou p ipite les ARNs. Le le de ai , la solutio est
centrifugée pendant 15 min à 14 p à °C et l’isop opa ol est eti . Le ulot o te a t
les ARNs est la a e de l’ tha ol 70% pour éliminer les sels et de nouveau centrifugé
pendant 15 min à 14 p à °C. Ap s a oi eti l’ tha ol, le ulot d’ARNs est s h à
°C à l’aide d’u ai à se . E fi , les ARNs totau so t suspe dus da s de l’eau d sio is e
exempte de ribonucléases et chauffés 5 min à 65°C. La concentration en ARN de chaque
échantillon est mesurée en ng/ml par spectrophotométrie en utilisant le NanoDrop
Spectrophotometer ND-1000 (NanoDrop Technologies Inc, USA).
2.1.2 Extraction des ARNs totaux à partir des EODHs :
Les EODHs fraichement récoltés sont rincés 3 fois avec du PBS, dégraissés et ensuite coupés
manuellement avec des ciseaux stériles en petits fragments de 1–2 mm3. Ces fragments sont
transférés dans des tubes de 4,5 ml à fond rond (Greiner Bio-One) contenant du réactif RNABle
Eu o io . Les f ag e ts so t e suite o s Pol t o PT E, Fishe “ ie tifi jus u’à
o te tio d’u o at ho og e.
Le reste du protocole expérimental est le même que pour les cellules adhérentes en culture à
l’e eptio de l’ tape de p ipitatio des ARNs totau ui se fait à te p atu e a ia te
pendant 15 min.

Matériels et méthodes
100
2.2 Rétro-transcription ou RT (Reverse Transcription):
Les ARNs totaux précédemment extraits sont rétrotranscrits en ADN complémentaire (ADNc)
en utilisant les he a es u l otidi ues al atoi es ui se fi e t al atoi e e t su l’ARN afi
d’a o e la s th se du i d’ADN et l’e z e la t a s iptase i e se M-MLV) qui
atal se la a tio de s th se de l’ADN e utilisa t l’ARN o e at i e.
Après dissolution des ARNs totau , μL de p i so t ajout s au μL d’ARNs totau
µg). Le prémix (ou mélange réactionnel) est constitué de : 5 µl de tampon 5X (composé de 100
mM Tris–HCl pH 8.3, 150 mM KCl, 6.25 mM MgCl2, Invitrogen), 2,5 µl de DTT (20 mM,
Invitrogen), 1 µl de dNTPs (2 mM, I it oge et , µl de Ra do P i e Pd N . μg ,
I it oge et µl d’eau.
Ap s i u atio des ARNs à °C pe da t i , µl de d’i hi iteu de RNAse U,
RNaseOUT-Invitrogen) et 2 µl de transcriptase inverse M-MLV (400 U, Invitrogen) sont ajoutés
pou a oi u olu e fi al de μL.
L'ADNc est synthétisé à 37°C pendant 1 heure puis 25 µL d'eau stérile sont ajoutés. Les
enzymes restantes ont été désactivées par la chaleur (100°C, 2 min). Les ADNc peuvent être
conservés à -20°C.
2.3 Amplification par PCR (Polymerase Chain Reaction)
Ap s la p o du e de RT, , µl d’ADN ≈ g so t ajout s aux 22,5 µl de mélange
réactionnel de PCR. Ce mélange est composé de 2,5 µl de tampon 10X (composé de : 250 mM
de Tris-HCl pH8.4, 62,5 mM KCl, Invitrogen), 1 µl de MgCl2 (2,5 mM, Invitrogen), 0,25 µl de
dNTPs (277, µM, I it oge , µl d’a o es se s et a ti-sens (12 pM, Invitrogen), 0,25 µl de
Taq polymérase (1, U, I it oge et , µl d’eau. De l’huile io is e µl est ajoutée dans
ha ue tu e pou ite l’ apo atio à haute te p atu e.
La réaction de PCR est effectuée avec un thermocycleur (dénaturation à 95 ° C pendant 5
minutes, suivie de 38 cycles à 57°C pendant 30 s, 72°C pendant 30 s et 95°C Pendant 30 s.
Après le dernier cycle, les échantillons sont incubés à 57°C pendant 2 min et à 72°C pendant
10 minutes pour assurer une extension complète du produit. Toutes les amorces sont décrites
dans le tableau MM-3. La GAPDH est utilisée comme gène de ménage. Les produits amplifiés
sont séparés par électrophorèse sur gel d'agarose à 2% (contenant 0,01% de bromure
d'éthidium) dans du tampon Tris-Acétate-EDTA et visualisés en utilisant un Transilluminateur
UV.

Matériels et méthodes
101
Tableau MM-3 : Liste des amorces sens et anti-sens utilisées pour la PCR.
Gène Amorce sens Amorce anti-sens
GAPDH AAGGTCATCCCTGAGCTGAA TGACAAAGTGGTCGTTGAGG
MEF2C AAGAAGGAGATTTGTTTGGAGG CTTTGTAAATGTCACCTGTCTG
GATA4 CCCAATCTCGATATGTTTGAC CGTTCATCTTGTGGTAGAGG
TNNT2 CTGCTGTTCTGAGGGAGAGC CACCAAGTTGGGATGAACG
ACTC1 CCTTCATTGGTATGGAATCTG AATCTTCATGGTGCTTAGGAG
β-MHC GAGCTACGGGTTCCCCTCT GGAAGCTCTGCTACCCCTTTT
NKX2.5 AAAGAAAGGGTGTGAACTGC CGCACAGTAATGGTAAGGGA
KCa1.1 CCTGCAGCGAAGTATCATCA ACTCGAAGTGAAGCTGCCAT
Connexin43 GGGCGTTAAGGATCGGGTTAAGG GTTGGTGAGGAGCAGCCATTGAA
KCa3.1 GCCTCAGAGCAAAAGTCCCA AGGTAGAGCGCCCACGAG
KCa2.1 GAAGTTCCTCCAAGCTATCCA TCTTTCCGTTCCCTGGTCT
KV4.2 ACATGCAGAGCAAACGGAA GGACTGTGACTTGAAGGACGA
KV4.3 GCCTCCGAACTAGGCTTTCT CCCTGCGTTTATCAGCTCTC
Kir2.1 ACTTCCACTCCATGTCCC CTTTACTCTTCCCGTTCC
S1PR1 var 1 GACTCGAGCTGCGGTTTCC TCCTTGTCCGCGCTGATATT
S1PR1 var 2 GAGCGAGGCTGCGGTT CCGCGCTGATATTCAGCTTT
S1PR2 CGGCCTAGCCAGTTCTGAAA AATGGCGCAACAGAGGATGA
S1PR3 GGCCGCTGGAAAAGTTTAGTC GTTGAAAAAGGGCTCCTCCGT
S1PR4 TTGCAGTCTTGCGTGTGGAT GACCATGGGAAGCCCATTTG
S1PR5 var 1 GGAGCAGCGGGAACCTATCT CCCCAAGGCTGTGAACTGA
S1PR5 var 2 AGGGCGCAGACCTTGG GCCCCGACAGTAGGATGTTG
TRPM4 TTGGCATACTGGGAGACGCA GGCCCAAGATCGTCATCGT
TRPM7 TCACAGAACTTCCATTCCTGTTCA CTGTGACAGGCTCCCCTCTT
XIV- RT-qPCR « Reverse Transcription- quantitative Polymerase Chain Reaction » en
SybrGreen
1 Principe :
Cette te h ologie est as e su la d te tio et la ua tifi atio d’u epo te fluo es e t
le « Sybr Green » do t l’ issio est di e te e t p opo tio elle à la ua tit
d’a pli o s g s pendant la réaction de PCR. Le « Sybr Green » est un intercalant de
l’ADN.
2 Protocole expérimental :
La RT-qPCR comme la RT-PCR, s’effe tue e g a des tapes : extraction des ARNs totaux,
reverse transcription (RT) et amplification par PCR quantitative (qPCR ou real-time PCR).
-Les p oto oles d’e t a tio d’ARNs totau et de RT sont les mêmes que pour la RT-PCR.

Matériels et méthodes
102
2.1 qPCR avec la technologie SybrGreen
Après la procédure de RT, la qPCR (avec la technologie SybrGreen) est réalisée sur plaque de
96 puits, avec les amorces sens et anti-sens de chaque gène étudié (tableau MM-3). Le volume
final du mélange réactionnel est de 15 µl par puits, o te a t µl de Mi et µl d’ADN
g fi al/puits . Le Mi o tie t , µl de ouple d’a o es se s et a ti-sens) à 900 nM, 7,5
µl de réactif Power SYBR® Green PCR Master Mix (Thermo Fisher Scientific) et 1, µl d’eau.
Le Power SYBR® Green PCR Master Mix contient la Taq Gold® DNA Polymerase (enzyme de la
réaction), les dNTPs et les sels nécessaires pour la réaction. Chaque échantillon a été
pol is e t iple afi d’esti e au ieu le le seuil le th eshold d’appa itio de la
fluo es e e. L’eau est hoisie o e o t ôle gatif. La a tio d’a plifi atio o e e
pa u e d atu atio i itiale de l’ADN pe da t i à °C, pe etta t l’a ti atio de
l’ADN pol ase, et est sui ie de les su essifs o pos s d’u e tape de d atu atio
pe da t se à °C, puis d’u e tape d’h idatio des a o es et d’ lo gatio pe da t
1 min à 60°C. La quantification de produits amplifiés à chaque cycle se fait par détection de
fluo es e e g â e à l’appa eil Fast Real-Time PCR System (Applied Biosystems USA).
2.2 RT-qPCR à haut rendement ou TLDA (TaqMan Low Density Custom Array)
2.2.1 Principe:
La technologie TLDA (ou TaqMan Low Density Array) est une qPCR à haut rendement car elle
permet de déterminer rapidement le profil d'expression d'un grand nombre de gènes et sur
plusieu s ha tillo s tout e utilisa t u e fai le ua tit d’ADN . Co t ai e e t à la PCR
en plaque 96 puits, la TLDA est réalisée sur cartes micro-fluidiques composées de 384 puits.
Le p i ipe de la TLDA epose su l’utilisatio de so des fluo es e tes « sondes TaqMan ».
Comme pour la qPCR en « SybrGreen », le signal fluorescent résultant permet une mesure
quantitative de l'accumulation exponentielle du produit au cours des différents cycles de PCR.
2.2.2 Extraction des ARNs totaux
L’e t a tio des ARNs totau à pa ti des C“Cs W B + et des CSCs W8B2+ différenciées est
alis e a e u Kit d’e t a tio d’ARN Nu leo“pi ® Ma he e -Nagel). Les cellules sont
collectées après trypsinisation et centrifugation (8 min à 300 g) et lysées avec le tampon RA1
et le β-mercaptoethanol. Le lysat est ensuite filtré avec des colonnes (NucleoSpin® Filter) et

Matériels et méthodes
103
e t ifug s i à g . Ap s filt atio , l’ tha ol % est ajout au l sat et e de ie
est déposé sur des colonnes (NucleoSpin® RNA Column). Le lysat est ensuite centrifugé (30 s
à 11000 g). Après ajout du MDB (Membrane Desalting Buffer), le lysat est centrifugé (1 min à
g pou li i e les sels. L’ADN est e suite dig a e u e solutio dite de
« digestion » (DNase reaction mixture) pendant 15 min à température ambiante. Les ARNs
sont ensuite rincés avec les tampons RAW2 et RA3 (1er rinçage avec le tampon RAW2 et le 2ème
et 3ème i çage a e le ta po RA . E fi , les ARNs so t lu s da s µl d’eau RNase-free
water) et centrifugés (1 min à 11000 g). La concentration en ARN de chaque échantillon est
mesurée en ng/ml par spectrophotométrie en utilisant le NanoDrop Spectrophotometer ND-
1000 (NanoDrop Technologies Inc, USA). Les ARNs sont gardés à -8 °C jus u’à la p o hai e
étape.
2.2.3 Transcription inverse ou RT (Reverse Transcription):
Les ARNs totaux précédemment extraits sont rétrotranscrits en ADN complémentaires (ADNc)
en utilisant un Kit de transcription inverse (Super Script IV VILO, Invitrogen). Pour cela, 16 µl
d’ARNs totau o te a t µg d’ARNs totau o pl t s a e de l’eau so t ajout s aux 4 µl
de superscript IV VILO Master mix du kit (contenant tout ce qui est nécessaire pour la réaction
de RT). Après centrifugation rapide, le mélange est incubé 10 min à 25°C puis 20 min à 50°C
et enfin 5 min à 85°C. Après une rapide centrifugation, les ADNc obtenus sont conservés à -20
°C.
2.2.4 qPCR avec la technologie TaqMan
Cette expérimentation a été réalisée en collaboration avec le Docteur Nathalie Gaborit de
l'institut du thorax, équipe INSERM UMR 1087 / CNRS UMR 6291 à Nantes.
Le mélange réactionnel TaqMan® master mix (incluant tous les agents nécessaires pour la
réaction de PCR) est ajouté aux 384 puits de la carte (TaqMan® Array Micro Fluidic Card, AB)
préchargés ave des so des d’h d ol se fluo escentes TaqMan® et des amorces spécifiques
du gène à amplifier. La réaction de PCR est effectuée sur TLDA avec le système de détection
de s ue es ABI P is HT Applied Bios ste s . L’a plifi atio se fait e ua a te
cycles comprenant trois étapes : une étape de dénaturation de 30 secondes à 97°C et une
tape d’h idatio et d’ lo gatio d’u e i ute à . °C. Les do es so t e ueillies à
l’aide du logi iel Applied Bios ste s “D“ . et a al s es a e la thode de ua tifi atio
relative du cycle seuil (Ct) (Livak & Schmittgen, 2001).

Matériels et méthodes
104
Les 96 gènes humains sélectionnés pour leur expression génique (tableau MM-4) codent pour
des canaux potassiques, calciques, sodiques et HCN. Mais également pour des protéines
i pli u es da s la sig alisatio et l’ho ostasie al i ue, des o e i es, des fa teu s de
transcription et des protéines structurales cardiaques, des NP (peptide natriurétique) et enfin
4 gènes références pour la normalisation. Nous avons sélectionné le gène RPL13A (Ribosomal
Protein L13a) pour la normalisation des données, en tant que gène le plus uniformément
distribué. L'expression relative de chaque gène par rapport à RPL13A a été calculée pour
ha ue ha tillo ΔCt i di ue des do es o alis es . Le p ofil d’e p essio de ha ue
gène dans les CSCs W8B2+ est représenté avec la formule : 2-ΔCt vs RPL13A (x104). La variation
d'expression de chaque gène dans les CSCs W8B2+ différenciées versus les CSCs W8B2+ non
différenciées est représentée par le facteur –ΔΔCt.
Tableau MM-4: Liste des sondes et des 96 gènes analysés par TLDA
Référence de la sonde Nom du gène Nom de la protéine
Hs01093761_m1
Hs00245832_m1
Hs00958036_m1
Hs00426868_g1
Hs00544877_m1
Hs00193090_m1
Hs00608066_m1
Hs00167681_m1
Hs00167753_m1
Hs00367969_m1
Hs01103527_m1
Hs00984856_m1
Hs01021049_m1
Hs01100744_m1
Hs00300085_s1
Hs00968732_g1
Hs00154286_m1
Hs00748445_s1
Hs00270952_m1
Hs00271416_s1
Hs00987388_s1
Hs01085412_m1
Hs00606903_m1
Hs00380018_m1
ABCC8
ABCC9
ATP1A3
ATP1B1
ATP2A2
ATP2A3
ATP2B4
CACNA1C
CACNA1D
CACNA1G
CACNA1H
CACNA2D1
CACNA2D2
CACNB2
CALM1
CALM3
CASQ2
GJA1
GJA5
GJA7
GJD3
HCN1
HCN2
HCN3
SUR1
SUR2
Na/K-ATPase a3
Na/K-ATPase b1
SERCA2
SERCA3
Ca ATPase4
Cav1.2
Cav1.3
Cav3.1
Cav3.2
Cava2d1
Cava2d2
Cavb2
CALM1
CALM3
CASQ2
Cx43
Cx40
Cx45
Cx30.2
HCN1
HCN2
HCN3
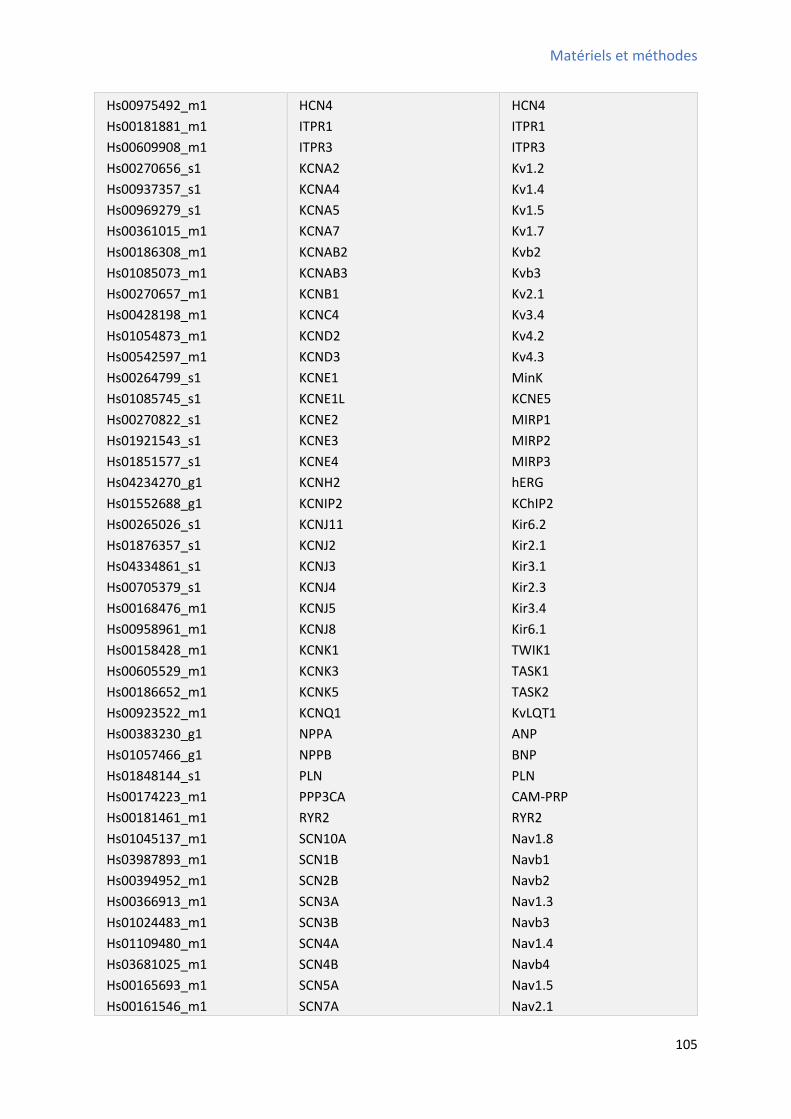
Matériels et méthodes
105
Hs00975492_m1
Hs00181881_m1
Hs00609908_m1
Hs00270656_s1
Hs00937357_s1
Hs00969279_s1
Hs00361015_m1
Hs00186308_m1
Hs01085073_m1
Hs00270657_m1
Hs00428198_m1
Hs01054873_m1
Hs00542597_m1
Hs00264799_s1
Hs01085745_s1
Hs00270822_s1
Hs01921543_s1
Hs01851577_s1
Hs04234270_g1
Hs01552688_g1
Hs00265026_s1
Hs01876357_s1
Hs04334861_s1
Hs00705379_s1
Hs00168476_m1
Hs00958961_m1
Hs00158428_m1
Hs00605529_m1
Hs00186652_m1
Hs00923522_m1
Hs00383230_g1
Hs01057466_g1
Hs01848144_s1
Hs00174223_m1
Hs00181461_m1
Hs01045137_m1
Hs03987893_m1
Hs00394952_m1
Hs00366913_m1
Hs01024483_m1
Hs01109480_m1
Hs03681025_m1
Hs00165693_m1
Hs00161546_m1
HCN4
ITPR1
ITPR3
KCNA2
KCNA4
KCNA5
KCNA7
KCNAB2
KCNAB3
KCNB1
KCNC4
KCND2
KCND3
KCNE1
KCNE1L
KCNE2
KCNE3
KCNE4
KCNH2
KCNIP2
KCNJ11
KCNJ2
KCNJ3
KCNJ4
KCNJ5
KCNJ8
KCNK1
KCNK3
KCNK5
KCNQ1
NPPA
NPPB
PLN
PPP3CA
RYR2
SCN10A
SCN1B
SCN2B
SCN3A
SCN3B
SCN4A
SCN4B
SCN5A
SCN7A
HCN4
ITPR1
ITPR3
Kv1.2
Kv1.4
Kv1.5
Kv1.7
Kvb2
Kvb3
Kv2.1
Kv3.4
Kv4.2
Kv4.3
MinK
KCNE5
MIRP1
MIRP2
MIRP3
hERG
KChIP2
Kir6.2
Kir2.1
Kir3.1
Kir2.3
Kir3.4
Kir6.1
TWIK1
TASK1
TASK2
KvLQT1
ANP
BNP
PLN
CAM-PRP
RYR2
Nav1.8
Navb1
Navb2
Nav1.3
Navb3
Nav1.4
Navb4
Nav1.5
Nav2.1

Matériels et méthodes
106
Hs00161567_m1
Hs01062258_m1
Hs00153998_m1
Hs99999903_m1
Hs04194366_g1
Hs00231122_m1
Hs00171403_m1
Hs00388359_m1
Hs00232018_m1
Hs00232622_m1
Hs01124217_g1
Hs00212560_m1
Hs04334749_m1
Hs00361155_m1
Hs00195612_m1
Hs00911929_m1
Hs00231763_m1
Hs00165957_m1
Hs00943911_m1
Hs01101425_m1
Hs01110632_m1
Hs00188163_m1
Hs00380556_m1
Hs00380518_m1
Hs00187842_m1
Hs01085598_g1
Hs00166405_m1
Hs99999901_s1
SCN9A
SLC8A1
ANK2
ACTB
RPL13A
GATA3
GATA4
GATA5
GATA6
HEY2
IRX3
IRX4
IRX5
TBX5
TBX3
TBX2
NKX2.5
TroponineI, cardiac
TroponineT,cardiac
MYH6
MYH7
RRAD
CLASP2
GPD1L
B2M
MYL7
MYL2
18S
Nav1.7
NCX1
ANKB
ACTB
RPL13A
GATA3
GATA4
GATA5
GATA6
HEY2
IRX3
IRX4
IRX5
TBX5
TBX3
TBX2
NKX2-5
TNNI3
TNNT2
MYH6
MYH7
RAD
CLASP2
GPD1L
B2M
MLV2A
MLC2V
18S
XV- Western blot
1 Principe:
Le Weste lot est u e te h i ue d’a al se pou ett e e ide e l’e p essio des
p ot i es d’i t t se t ou a t da s u la ge p ot i ue. Les p ot i es totales e t aites
sont séparées par électrophorèse en conditions dénaturantes selon leur poids moléculaire,
puis transférées sur une membrane (électrotransfert ou blot). La protéine d'intérêt est ensuite
d te t e pa i u od te tio i di e te pa u a ti o ps p i ai e sp ifi ue. L’a ti o ps
secondaire est couplé à un groupement enzymatique permettant une révélation par chimie-
luminescence.

Matériels et méthodes
107
2 Protocole expérimental:
Pour déterminer l'expression protéique de KCa1.1, NCX1 ,NCX3 et S1PR3, les cellules W8B2+
ou le tissu atrial humain ont été lysées dans le tampon de radioimmunoprécipitation
(radioimmunoprecipitation assay buffer) contenant en mM: Tris-HCl 50 pH: 8, NaCl 150, EDTA
5, 0,05% d'Igepal, 1% d'acide désoxycholique, 1% de Triton X-100, 0,1% de SDS) et
supplémenté avec des inhibiteurs des protéases et des phosphatases (Protease Inhibitor
Cocktail, Sigma-Aldrich - PhosSTOP Phosphatase Inhibitor Cocktail, Roche).
Les lysats cellulaires solubles ont été dénaturés 5 min à 37 ° C dans du tampon de Laemmli 2
x contenant (en mM): Tris-HCl 125 pH 6,8, SDS 4%, glycerol 20%, bleu de bromophénol 0,004%,
β-mercaptoéthanol à 10%). Les échantillons protéiques (entre 20 et 80 μg), obtenus à partir
de cellules W8B2+ ou du tissu atrial humain, ont été séparés par SDS-PAGE en utilisant des gels
de polyacrylamide à 8% et transférés dans des membranes de nitrocellulose. Les membranes
ont été bloquées 90 min dans une solution de blocage « TBS-Tween » contenant (en mM): Tris
20 pH: 7,6, NaCl 150 et Tween-20 0,2% supplémenté avec 5% de lait écrémé à température
ambiante. Les membranes sont ensuite incubées pendant la nuit à 4 °C avec les anticorps
primaires (tableau MM-5) dilués dans du TBST contenant 5% de lait écrémé. Les membranes
sont ensuite lavées avec du TBS-Tween trois fois pendant 10 min, puis incubées pendant 90
minutes à température ambiante avec les anticorps secondaires correspondants couplés à la
peroxydase HRP (Horse-Radish Peroxydase) dilués dans le TBST contenant 5% de lait écrémé.
Les membranes sont révélées par chémoluminescence avec le substrat (ECL) (GE Healthcare,
Velizy-Villa ou la , F a e . Les sultats o te us so t a al s s e utilisa t l’i ageu
GeneGnome (SynGene Ozyme, Montigny-le-Bretonneux, France).
Tableau MM-5 : Liste des anticorps utilisés pour le Western Blot.
Anticorps primaire Espèce Dilution Fournisseur
Polyclonal anti-KCa1.1 Lapin 1/500e Alomone Labs Polyclonal anti-NCX1 Lapin 1/200e Alomone Labs Polyclonal anti-NCX3 Lapin 1/200e Alomone Labs Polyclonal anti-S1PR3 Lapin 1/100e Abcam Polyclonal anti-Vimentine Lapin 1/500e Santa Cruz
Anticorps secondaire Espèce Dilution Fournisseur
Anti-lapin Chèvre 1/5000e Interchim

Matériels et méthodes
108
XVI- Patch clamp
1 Principe :
Le patch-clamp (« patch » pour fragment de membrane et « clamp » pour maintien) est une
te h i ue d’ tude des a au io i ues. Elle est as e su l’utilisatio d’u e fi e pipette de
e e e plie d’u e solutio o du t i e et o e t e à u a plifi ateur) qui une fois en
contact avec la membrane cellulaire (qui contient des canaux ioniques pe et d’e egist e
des courants ioniques. L’a plifi ateu pe et d’i pose des potentiels et de recueillir des
courants (voltage‐ la p ou d’i pose des ou a ts et de e ueilli des a iatio s de pote tiel
(current-clamp).
Selon la configuration choisie, ette te h i ue pe et d’e egist e des ou a ts
a os opi ues sulta t de l’a ti it de plusieu s e tai es ou illie s de a au , ou des
ou a ts u itai es p o o u s pa l’ou e tu e d’u ou de uel ues a au i di iduels < à
canaux). Pour enregistrer les courants portés sur les CSCs W8B2+, nous avons utilisé le voltage-
clamp en configuration cellule entière (whole cell).
2 Protocole expérimental :
Les courants ioniques ont été enregistrés sur les CSCs W8B2+ (ensemencées à une densité de
2.103 cellules/cm2 dans des boites de culture de 35mm de diamètre) par la technique du patch
la p e o figu atio ellule e ti e. Le pat h la p est alis à l’aide d’u a plifi ateu
Axopatch 200A avec une tête CV 202AU (Molecular Devices, CA, USA). Les courants sont
esu s e po se à l’appli atio d’u e te sio ode oltage la p g e pa
l’a plifi ateu o t ol pa u o di ateu pe so el uip d'u o e tisseu a alogi ue-
numérique (Digidata 1322A, Molecular Devices) et à l'aide du logiciel pClamp version 10
(Molecular Devices).
Au ou s de l’e p i e tatio , les ellules so t pe fus es a e u e solutio e t a ellulai e
contenant (en mM) : 136 NaCl, 5 KCl, 1 MgCl2, 1,8 CaCl2, 0,3 NaH2PO4, 10 Glucose, 10 HEPES,
pH 7,3). Les courants ioniques membranaires sont enregistrés via des électrodes de verre de
sista e o p ise e t e et MΩ e o osili ate GC -T fabriquées sur une étireuse
verticale à double chauffage. La solution intrapipette contient (en mM) : 20 KCl, 110 K-asp, 1
MgCl2, 2 CaCl2, 10 HEPES, 2,5 EGTA, 0,1 GTP, 5 Na2-phosphocréatinine, 5 Mg-ATP, pH 7,2 (PCa
= 6,3). Le calcium est absent pour la solution PCa<9. Les ou a ts so t e egist s e l’a se e

Matériels et méthodes
109
de traitement pharmacologique ou en présence de drogues. Après avoir obtenu une
résista e d’a ole e t e a ai e de l’o d e du giga-Ohm par pression négative, la
e a e ellulai e a t o pue pa su io afi d’o te i la o figu atio ellule-entière.
Divers protocoles de stimulation ont été imposés et sont précisés sur les figures
représentatives des traces de courant enregistrées.
XVII- Tests statistiques
Toutes les analyses statistiques ont été réalisées à l'aide de GraphPad Prism (La Jolla, CA,
USA). Les tests utilisés pour les différentes expériences sont spécifiés dans les légendes des
figures.

Résultats
110
RESULTATS
La partie « Résultats » s’a ti ule autou de t ois hapit es visant à mieux caractériser la
population des cellules souches cardiaques exprimant le marqueur W8B2 (CSCs W8B2+) à
l’ tat sou he ais aussi au ou s de sa diff e iatio a dia ue in vitro.
Dans un premier temps, nous nous sommes focalisés sur la caractérisation phénotypique,
génique et fonctionnelle des CSCs W8B2+ soit :
- La caractérisation immuno-phénotypique après isolement et purification
- La caractérisation génique des canaux ioniques et des acteurs clés de la signalisation
calcique
- L’ olutio de ette e p essio g i ue au ou s de leu diff e iatio a dia ue in
vitro
- L’ tude de l’a ti it al ique au cours de la différenciation cardiaque in vitro
Da s u se o d te ps, ous a o s tudi l’i po ta e des a au io i ues plus
précisément le canal BKCa) dans la régulation des propriétés fondamentales (prolifération et
auto-renouvellement) des CSCs W8B2+.
Et e fi ous a o s a al s les effets et le ode d’a tio de la sphi gosi e -phosphate
ou S1P (un important régulateur physiologique et physiopathologique cardiaque) sur les
propriétés des CSCs W8B2+ (prolifération, auto-renouvellement et migration).
Chapitre I : Isolement et caractérisation des CSCs W8B2+
Différents types de cellules souches ont pu être isolés à partir du tissu cardiaque humain
(Guan and Hasenfuss, 2013). Cette di e sit est due à l’e p essio sp ifi ue de e tai s
marqueurs qui peuvent être des récepteurs membranaires, des facteurs de transcription ou
encore des antigènes de surface. En fonction des travaux et données récemment publiés dans
le domaine des progéniteurs cardiaques humains (Zhang et al., 2015a) ot e tude s’est
o ie t e su la a a t isatio d’u t pe de ellules sou hes a dia ues e p i a t le
marqueur W8B2 (CSCs W8B2+). Plusieurs propriétés ont été vérifiées :
- Leu s p op i t s d’auto-renouvellement et de prolifération
- Leur immuno-phénotype

Résultats
111
- Leu p ofil d’e p essio pou e tai s a ueu s a dia ues
- Leu p ofil d’e p essio g i ue pou e tai s a au io i ues
I- Isolement et caractérisation phénotypique des CSCs W8B2+
1 Isolement des CSCs W8B2+
Les CSCs W8B2+ so t isol es à pa ti d’ ha tillo s d’o eillettes d oites hu ai es o te us
lo s d’op atio s hi u gi ales e : pontages coronariens). Dans cette étude les patients
choisis présentent tous un rythme sinusal o al. L’âge o e est de , ± 9 ans (5 femmes
et 17 hommes).
U e populatio ellulai e totale est d’a o d isol e pa la te h i ue de « culture des
explants » à partir des tissus atriaux humains. Cette population a été ensuite purifiée par les
techniques du tri magnétique et du tri par FACS. La 1ère étape de purification par tri cellulaire
magnétique positif montre que les CSCs W8B2+ représentent environ 3% des cellules totales
(figure R-1 A).
Après expansion cellulaire en culture, la proportion des CSCs W8B2+ isolées par tri
magnétique est enrichie mais reste faible et représente 9,7% des cellules totales (figure R-1
B). La e tape de pu ifi atio pa t i ellulai e pa FAC“ ua t à elle, pe et d’o te i u e
population pure (> 98% de pureté) de CSCs W8B2+ (figure R-1 C).
Ces résultats réalisés sur 22 patients démontrent la présence de CSCs W8B2+ dans
l’o eillette hu ai e et la essit , de pa leu a et , d’utilise des te h i ues de t i ellulai e
pour obtenir une population pure.
2 Capa it s d’auto-renouvellement et de prolifération des CSCs W8B2+
Les CSCs W8B2+ pures isolées adhèrent facilement au plastique et possèdent un
phénotype allongé et fusiforme identique à celui des cellules mésenchymateuses humaines
(figure R-2 A). Pour démontrer le caractère souche de ces cellules, il a été nécessaire de
montrer leur capacité de multiplication clonogénique par un test de formation de colonies
CFU-Fs (colony-forming unit-fi o lasts . A t s fai le de sit d’e se e e e t
cellules/cm²), les CSCs W8B2+ prolifèrent et forment de multiples colonies rendues visibles par
la coloration au Crystal violet (figure R-2 B a,b).
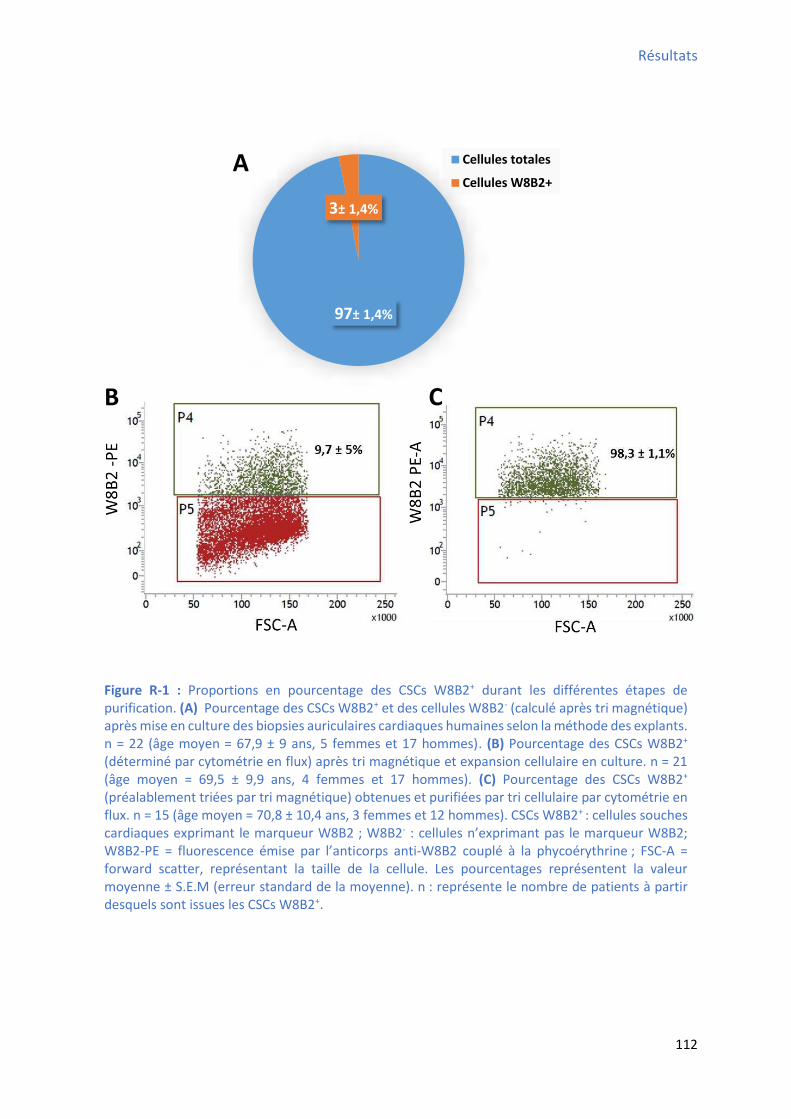
Résultats
112
A
B C
Figure R-1 : Proportions en pourcentage des CSCs W8B2+ durant les différentes étapes de purification. (A) Pourcentage des CSCs W8B2+ et des cellules W8B2- (calculé après tri magnétique) après mise en culture des biopsies auriculaires cardiaques humaines selon la méthode des explants. n = 22 (âge moyen = 67,9 ± 9 ans, 5 femmes et 17 hommes). (B) Pourcentage des CSCs W8B2+ (déterminé par cytométrie en flux) après tri magnétique et expansion cellulaire en culture. n = 21 (âge moyen = 69,5 ± 9,9 ans, 4 femmes et 17 hommes). (C) Pourcentage des CSCs W8B2+ (préalablement triées par tri magnétique) obtenues et purifiées par tri cellulaire par cytométrie en flux. n = 15 (âge moyen = 70,8 ± 10,4 ans, 3 femmes et 12 hommes). CSCs W8B2+ : cellules souches cardiaques exprimant le marqueur W8B2 ; W8B2- : ellules ’e p i a t pas le a ueu W B ; W8B2-PE = fluo es e e ise pa l’a ticorps anti-W8B2 couplé à la phycoérythrine ; FSC-A = forward scatter, représentant la taille de la cellule. Les pourcentages représentent la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir desquels sont issues les CSCs W8B2+.
97± 1,4%
3± 1,4%
Cellules totales
Cellules W8B2+
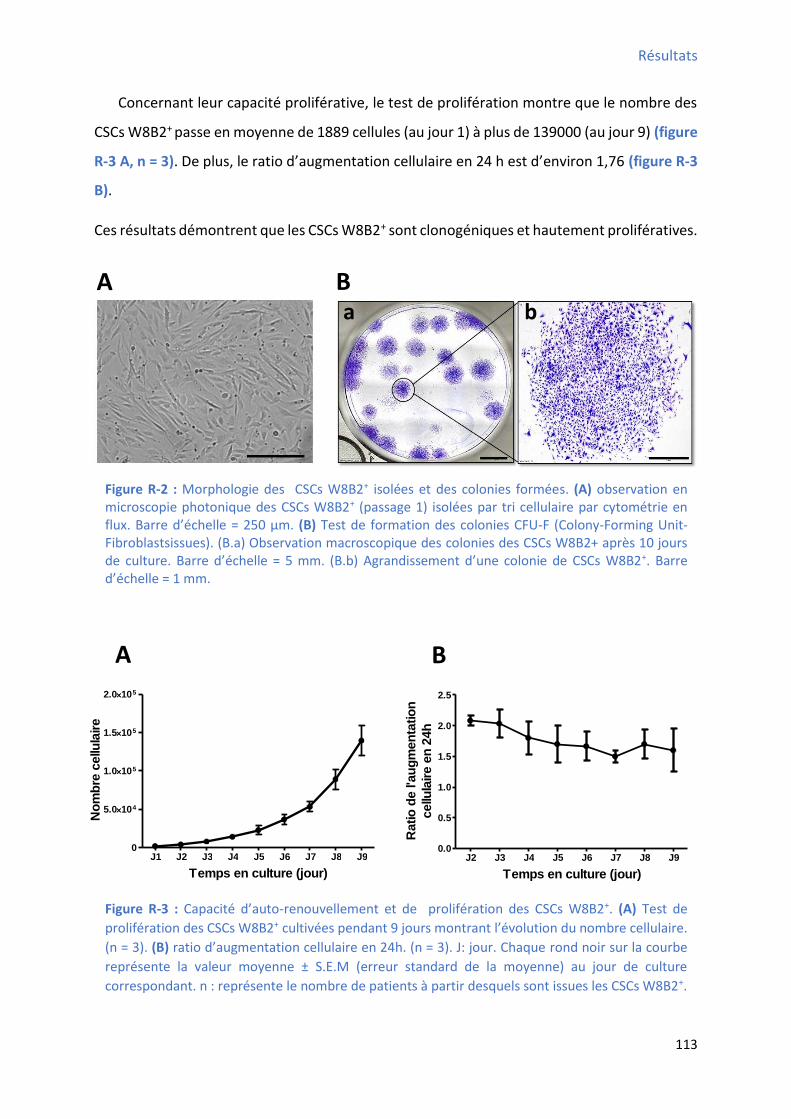
Résultats
113
Concernant leur capacité proliférative, le test de prolifération montre que le nombre des
CSCs W8B2+ passe en moyenne de 1889 cellules (au jour 1) à plus de 139000 (au jour 9) (figure
R-3 A, n = 3). De plus, le atio d’aug e tatio ellulai e e h est d’e i o , (figure R-3
B).
Ces résultats démontrent que les CSCs W8B2+ sont clonogéniques et hautement prolifératives.
A B
B
Figure R-2 : Morphologie des CSCs W8B2+ isolées et des colonies formées. (A) observation en microscopie photonique des CSCs W8B2+ (passage 1) isolées par tri cellulaire par cytométrie en flu . Ba e d’ helle = µ . (B) Test de formation des colonies CFU-F (Colony-Forming Unit-Fibroblastsissues). (B.a) Observation macroscopique des colonies des CSCs W8B2+ après 10 jours de ultu e. Ba e d’ helle = . B. Ag a disse e t d’u e olo ie de C“Cs W B +. Barre d’ helle = .
a b
A
J1 J2 J3 J4 J5 J6 J7 J8 J90
5.0104
1.0105
1.5105
2.0105
Temps en culture (jour)
Nom
bre
cellu
laire
J2 J3 J4 J5 J6 J7 J8 J90.0
0.5
1.0
1.5
2.0
2.5
Temps en culture (jour)
Rat
io d
e l'a
ugm
enta
tion
cellu
laire
en
24h
Figure R-3 : Capa it d’auto-renouvellement et de prolifération des CSCs W8B2+. (A) Test de
prolifération des CSCs W8B2+ ulti es pe da t jou s o t a t l’ olutio du o e ellulai e. (n = 3). (B) atio d’aug e tatio ellulai e e h. = . J: jou . Cha ue rond noir sur la courbe
représente la valeur moyenne ± S.E.M (erreur standard de la moyenne) au jour de culture
correspondant. n : représente le nombre de patients à partir desquels sont issues les CSCs W8B2+.
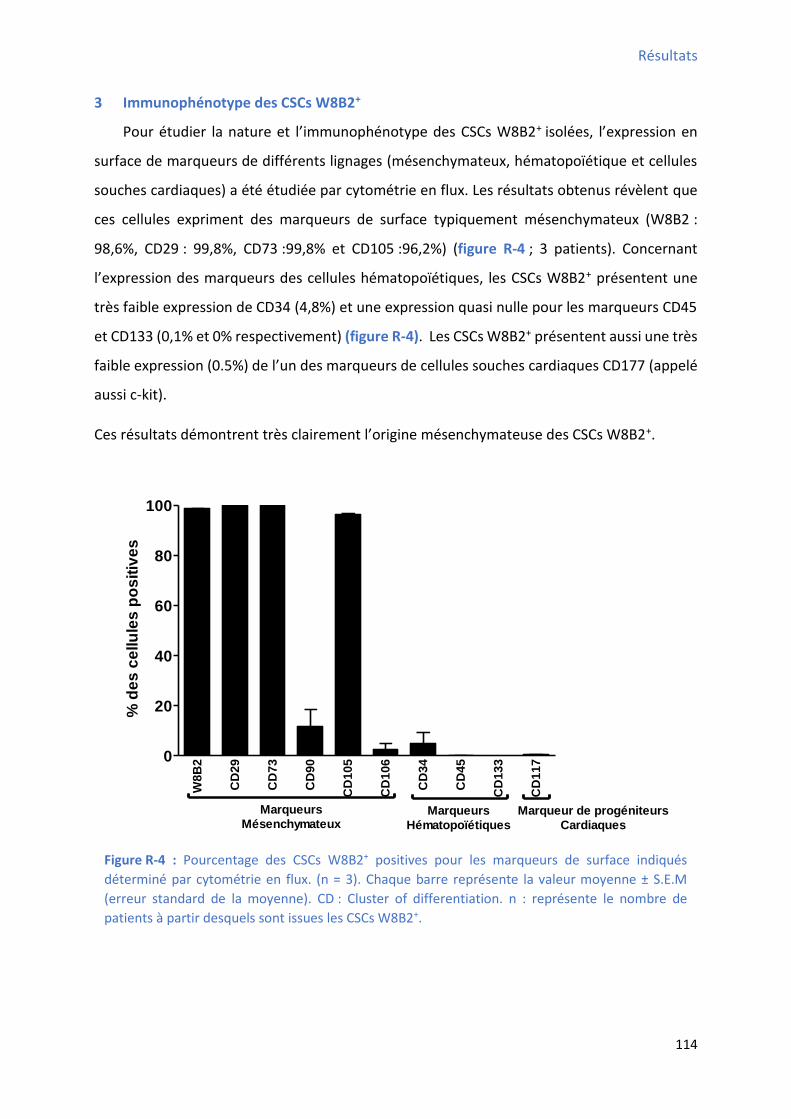
Résultats
114
3 Immunophénotype des CSCs W8B2+
Pour tudie la atu e et l’i u oph ot pe des C“Cs W B + isol es, l’e p essio e
surface de marqueurs de différents lignages (mésenchymateux, hématopoïétique et cellules
souches cardiaques) a été étudiée par cytométrie en flux. Les résultats obtenus révèlent que
ces cellules expriment des marqueurs de surface typiquement mésenchymateux (W8B2 :
98,6%, CD29 : 99,8%, CD73 :99,8% et CD105 :96,2%) (figure R-4 ; 3 patients). Concernant
l’e p essio des a ueu s des ellules h atopoï ti ues, les C“Cs W B + présentent une
très faible expression de CD34 (4,8%) et une expression quasi nulle pour les marqueurs CD45
et CD133 (0,1% et 0% respectivement) (figure R-4). Les CSCs W8B2+ présentent aussi une très
fai le e p essio . % de l’un des marqueurs de cellules souches cardiaques CD177 (appelé
aussi c-kit).
Ces sultats d o t e t t s lai e e t l’o igi e se h ateuse des C“Cs W B +.
W8B
2
CD
29
CD
73
CD
90
CD
105
CD
106
CD
34
CD
45
CD
133
CD
1170
20
40
60
80
100
MarqueursMésenchymateux
MarqueursHématopoïétiques
Marqueur de progéniteursCardiaques
% d
es c
ellu
les
posi
tives
Figure R-4 : Pourcentage des CSCs W8B2+ positives pour les marqueurs de surface indiqués
déterminé par cytométrie en flux. (n = 3). Chaque barre représente la valeur moyenne ± S.E.M
(erreur standard de la moyenne). CD : Cluster of differentiation. n : représente le nombre de
patients à partir desquels sont issues les CSCs W8B2+.

Résultats
115
II-Caractérisation génique des CSCs W8B2+
1 P ofil d’e p essio des a ueu s a dia ues
1.1 P ofil d’e p essio g i ue des fa teu s de t a s iptio a dia ues
Les facteurs de transcription cardiaques contrôlent le programme génétique cardiaque et
jouent un rôle important dans la cardiogenèse et la différenciation cardiaque. Pour
d te i e le p ofil d’e p essio g i ue de e tai s fa teu s de t a s iptio a dia ues,
ous a o s ua tifi leu i eau d’e p essio pa RT-qPCR à haut rendement ou TLDA.
Les facteurs de transcription étudiés sont :
- La famille des facteurs de transcription GATA (protéines à doigts de zinc se fixant sur
la s ue e d’ADN « GATA ») : GATA3, GATA4, GATA5 et GATA6
- La famille des facteurs de transcription HESR (proteins à motif bHLH (basic helix-loop-
helix)) : HEY2
- La famille des facteurs de transcription à homéodomaine « Iroquois homeobox » :
IRX3, IRX4 et IRX5.
- La famille des facteurs de transcription T-box : TBX2, TBX3 et TBX5.
- La famille des facteurs de transcription à homéodomaine NKX : NKX2.5
Les résultats montrent (figure R-5A) que les CSCs W8B2+ expriment fortement les facteurs
de transcription GATA4 et TBX3 mais aussi les facteurs de transcription GATA3, GATA6, IRX3,
TBX2 et TBX5. Une faible expression des facteurs de transcription HEY2 et IRX5 est détecté
dans les CSCs W8B2+. Les résultats montrent aussi que les facteurs de transcription GATA5,
IRX4 et NKX2.5 sont absents dans les CSCs W8B2+.
L’e se le de es o se atio s i di ue ue certains facteurs de transcription cardiaques
précoces sont exprimés (ex : GATA4). Il ressort également une très forte expression de TBX3
connu comme étant impliqué dans les processus de développement cardiaque.
Afi d’ alue le p ofil d’e p essio p otéique de certains facteurs de transcription
cardiaques, nous avons vérifié leur présence par immunocytochimie.
Les résultats illustrés figure R-5B, o fi e t la p se e de la p ot i e GATA et l’a se e
de la protéine NKX2.5 dans les CSCs W8B2+. Il est à noter u’u aut e fa teu de t a s iptio
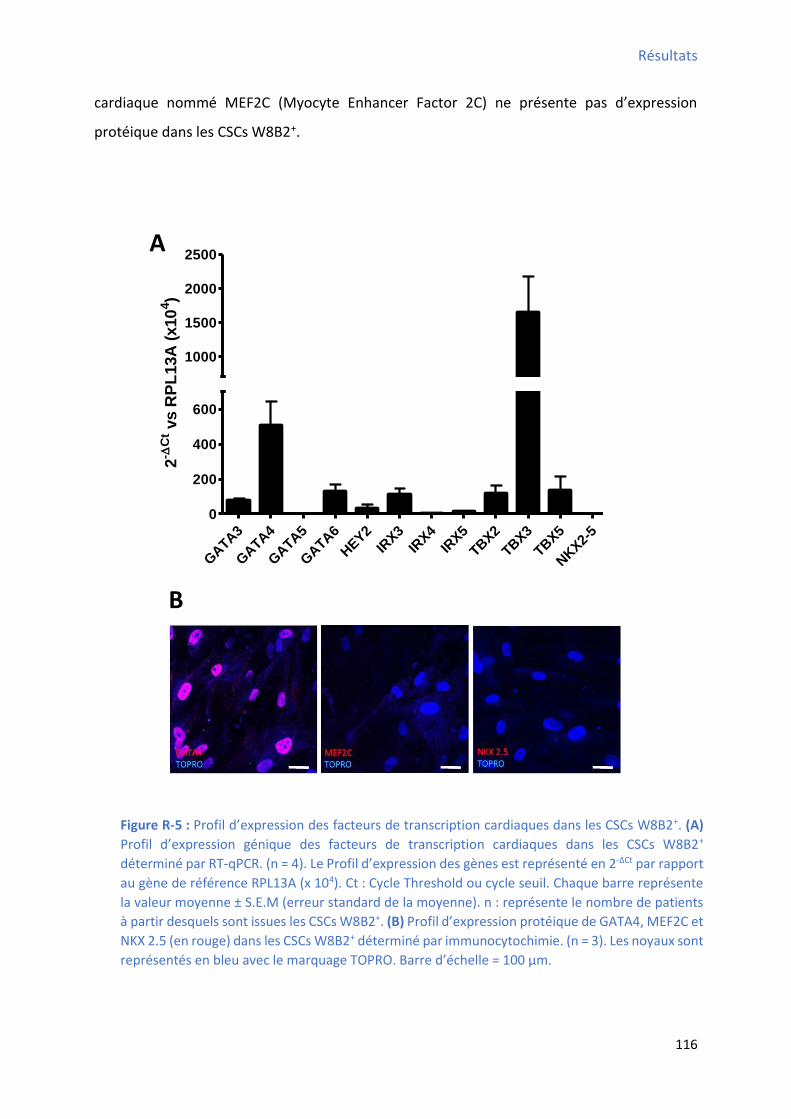
Résultats
116
a dia ue o MEF C M o te E ha e Fa to C e p se te pas d’e p essio
protéique dans les CSCs W8B2+.
GATA3
GATA4
GATA5
GATA6HEY2
IRX3
IRX4
IRX5
TBX2TBX3
TBX5
NKX2-5
0
200
400
600
1000
1500
2000
2500
2- Ct v
s R
PL1
3A (x
104 )
Figure R-5 : P ofil d’e p essio des fa teu s de t a s iptio a dia ues da s les C“Cs W B +. (A)
P ofil d’e p essio g i ue des fa teu s de t a s iptio a dia ues da s les C“Cs W B +
déterminé par RT- PCR. = . Le P ofil d’e p essio des g es est ep se t en 2-ΔCt par rapport
au gène de référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque barre représente
la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients
à partir desquels sont issues les CSCs W8B2+. (B) P ofil d’e p essio p ot i ue de GATA , MEF C et NKX 2.5 (en rouge) dans les CSCs W8B2+ déterminé par immunocytochimie. (n = 3). Les noyaux sont
ep se t s e leu a e le a uage TOPRO. Ba e d’ helle = µ .
A
B
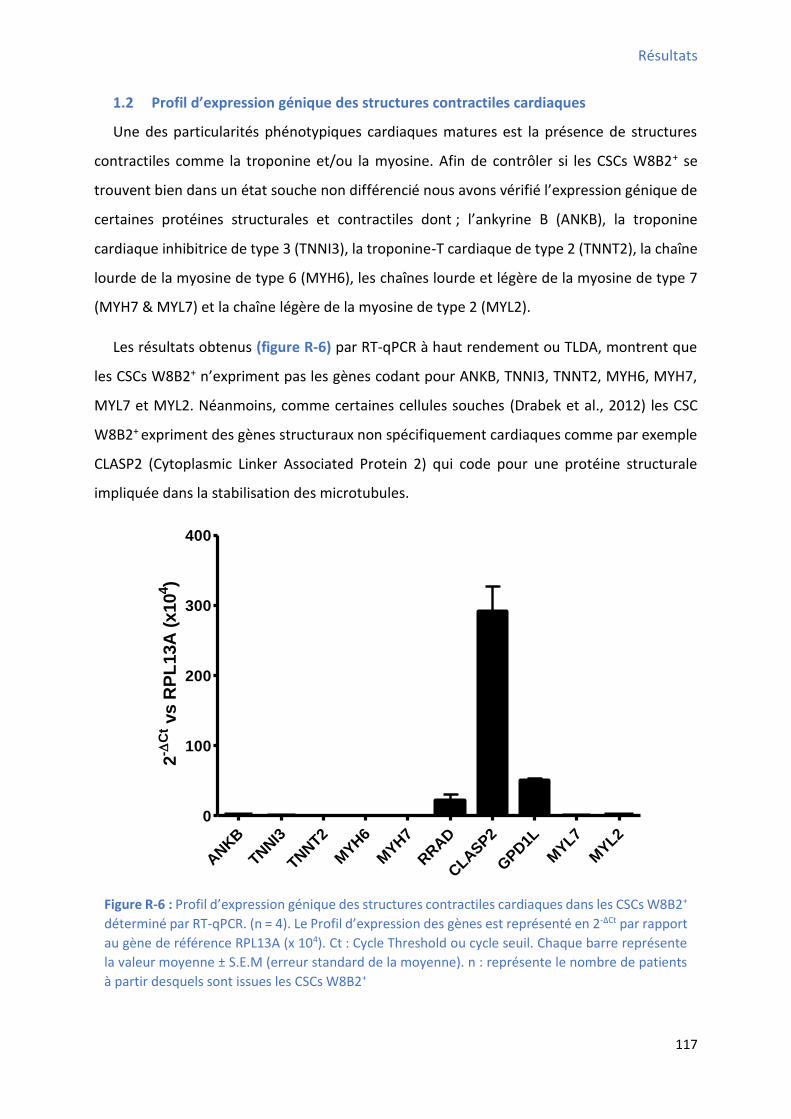
Résultats
117
1.2 P ofil d’e p essio g i ue des structures contractiles cardiaques
Une des particularités phénotypiques cardiaques matures est la présence de structures
contractiles comme la troponine et/ou la myosine. Afin de contrôler si les CSCs W8B2+ se
trouvent bien dans un état souche non diff e i ous a o s ifi l’e p essio g i ue de
certaines protéines structurales et contractiles dont ; l’a k i e B ANKB , la t opo i e
cardiaque inhibitrice de type 3 (TNNI3), la troponine-T cardiaque de type 2 (TNNT2), la chaîne
lourde de la myosine de type 6 (MYH6), les chaînes lourde et légère de la myosine de type 7
(MYH7 & MYL7) et la chaîne légère de la myosine de type 2 (MYL2).
Les résultats obtenus (figure R-6) par RT-qPCR à haut rendement ou TLDA, montrent que
les CSCs W8B2+ ’e p i e t pas les g es oda t pou ANKB, TNNI , TNNT , MYH , MYH ,
MYL7 et MYL2. Néanmoins, comme certaines cellules souches (Drabek et al., 2012) les CSC
W8B2+ expriment des gènes structuraux non spécifiquement cardiaques comme par exemple
CLASP2 (Cytoplasmic Linker Associated Protein 2) qui code pour une protéine structurale
impliquée dans la stabilisation des microtubules.
ANKBTNNI3
TNNT2
MYH6
MYH7
RRAD
CLASP2
GPD1LMYL7
MYL20
100
200
300
400
2- Ct v
s R
PL1
3A (x
104 )
Figure R-6 : P ofil d’e p essio g i ue des st u tu es o t a tiles a dia ues da s les C“Cs W B +
déterminé par RT- PCR. = . Le P ofil d’e p ession des gènes est représenté en 2-ΔCt par rapport
au gène de référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque barre représente
la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients
à partir desquels sont issues les CSCs W8B2+
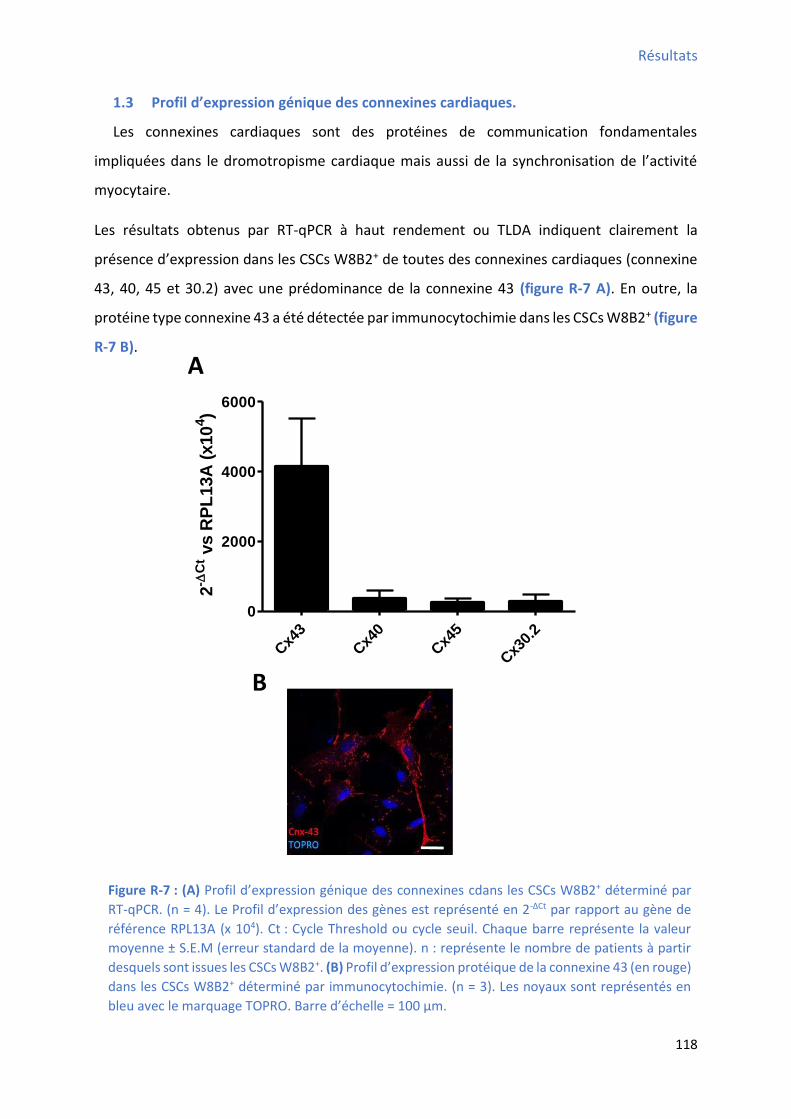
Résultats
118
1.3 P ofil d’e p essio g i ue des o e i es a dia ues.
Les connexines cardiaques sont des protéines de communication fondamentales
i pli u es da s le d o ot opis e a dia ue ais aussi de la s h o isatio de l’a ti it
myocytaire.
Les résultats obtenus par RT-qPCR à haut rendement ou TLDA indiquent clairement la
p se e d’e p essio da s les C“Cs W B + de toutes des connexines cardiaques (connexine
43, 40, 45 et 30.2) avec une prédominance de la connexine 43 (figure R-7 A). En outre, la
protéine type connexine 43 a été détectée par immunocytochimie dans les CSCs W8B2+ (figure
R-7 B).
Figure R-7 : (A) P ofil d’e p essio g i ue des o e i es cdans les CSCs W8B2+ déterminé par
RT- PCR. = . Le P ofil d’e p essio des g es est ep se t e -ΔCt par rapport au gène de
référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque barre représente la valeur
moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir
desquels sont issues les CSCs W8B2+. (B) P ofil d’e p essio p ot i ue de la o e i e e ouge)
dans les CSCs W8B2+ déterminé par immunocytochimie. (n = 3). Les noyaux sont représentés en
leu a e le a uage TOPRO. Ba e d’ helle = µ .
Cx43
Cx40
Cx45
Cx30.2
0
2000
4000
6000
2- Ct v
s R
PL1
3A (x
104 )
B
A
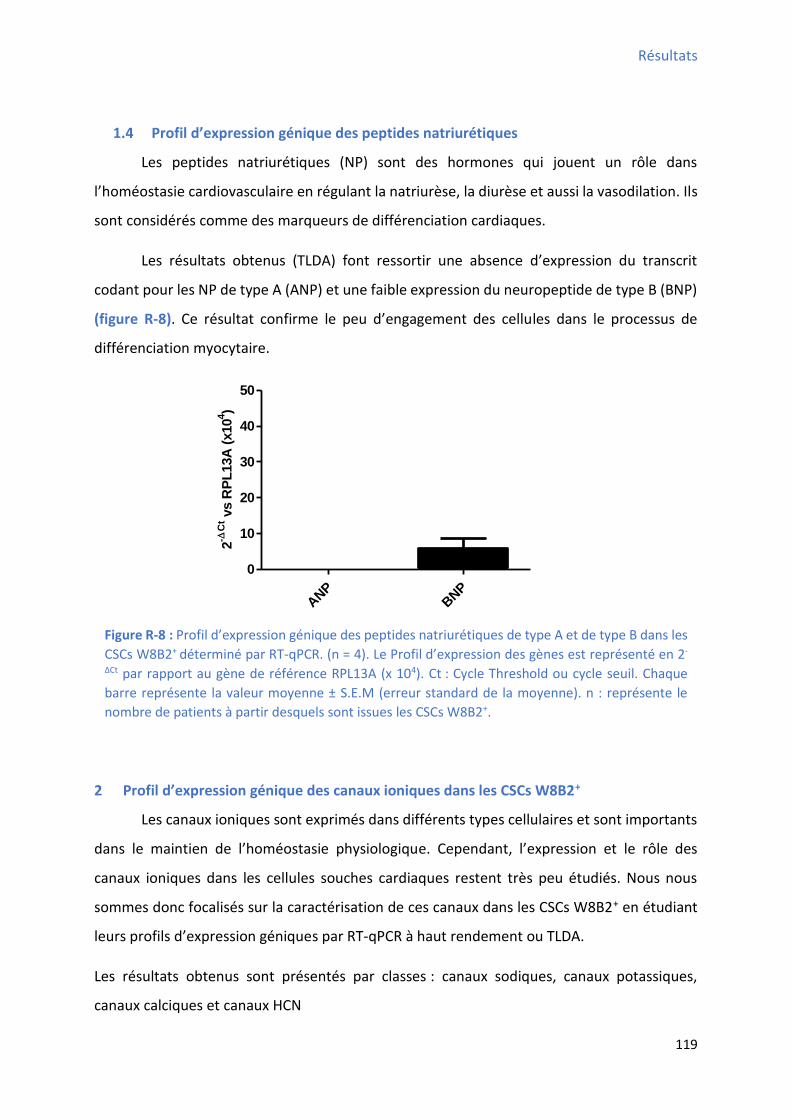
Résultats
119
1.4 P ofil d’e p essio g i ue des peptides at iu ti ues
Les peptides natriurétiques (NP) sont des hormones qui jouent un rôle dans
l’ho ostasie a dio as ulai e e gula t la at iu se, la diu se et aussi la asodilatio . Ils
sont considérés comme des marqueurs de différenciation cardiaques.
Les sultats o te us TLDA fo t esso ti u e a se e d’e p essio du t a s it
codant pour les NP de type A (ANP) et une faible expression du neuropeptide de type B (BNP)
(figure R-8). Ce sultat o fi e le peu d’e gage e t des ellules dans le processus de
différenciation myocytaire.
2 P ofil d’e p essio g i ue des a au io i ues da s les CSCs W B +
Les canaux ioniques sont exprimés dans différents types cellulaires et sont importants
da s le ai tie de l’ho ostasie ph siologi ue. Cepe da t, l’e p essio et le ôle des
canaux ioniques dans les cellules souches cardiaques restent très peu étudiés. Nous nous
sommes donc focalisés sur la caractérisation de ces canaux dans les CSCs W8B2+ en étudiant
leu s p ofils d’e p essio g i ues par RT-qPCR à haut rendement ou TLDA.
Les résultats obtenus sont présentés par classes : canaux sodiques, canaux potassiques,
canaux calciques et canaux HCN
Figure R-8 : P ofil d’e p essio g i ue des peptides at iu ti ues de t pe A et de t pe B da s les CSCs W8B2+ déterminé par RT- PCR. = . Le P ofil d’e p essio des g es est représenté en 2-
ΔCt par rapport au gène de référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque
barre représente la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le
nombre de patients à partir desquels sont issues les CSCs W8B2+.
ANPBNP
0
10
20
30
40
50
2- Ct v
s R
PL1
3A (x
104 )
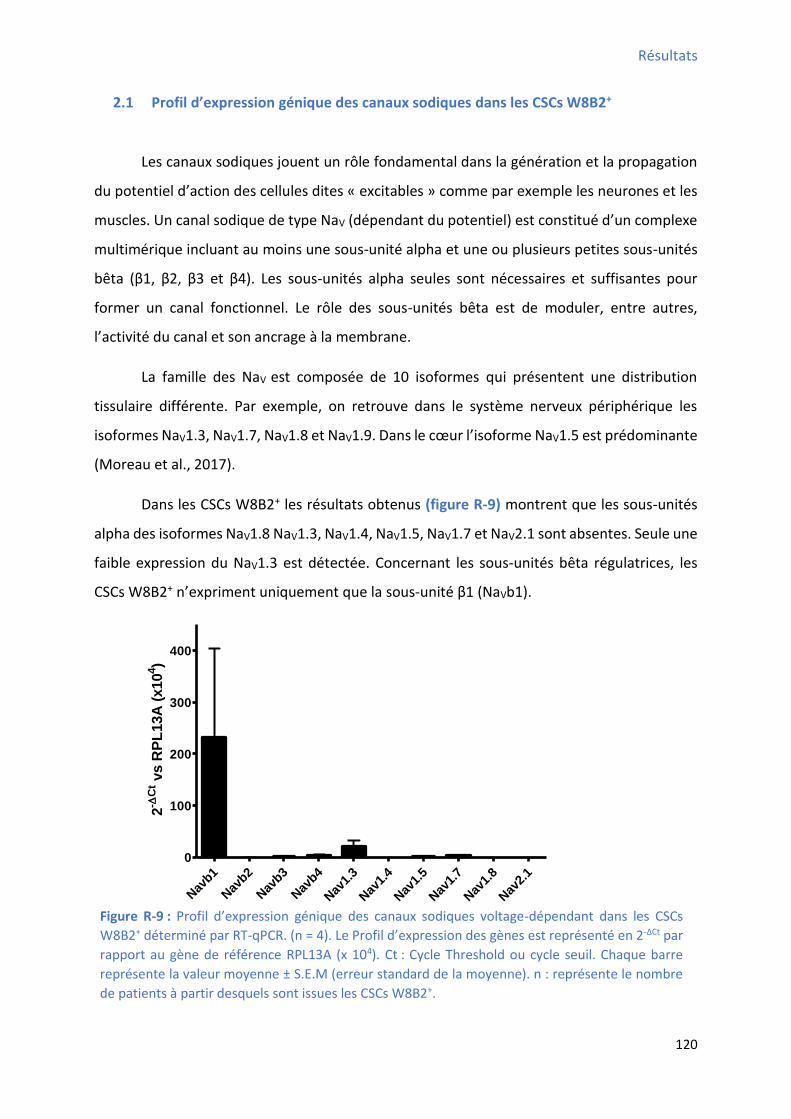
Résultats
120
2.1 P ofil d’e p ession génique des canaux sodiques dans les CSCs W8B2+
Les canaux sodiques jouent un rôle fondamental dans la génération et la propagation
du pote tiel d’a tio des ellules dites « excitables » comme par exemple les neurones et les
muscles. Un canal sodique de type NaV d pe da t du pote tiel est o stitu d’u o ple e
multimérique incluant au moins une sous-unité alpha et une ou plusieurs petites sous-unités
ta β , β , β et β . Les sous-unités alpha seules sont nécessaires et suffisantes pour
former un canal fonctionnel. Le rôle des sous-unités bêta est de moduler, entre autres,
l’a ti it du a al et so a age à la e a e.
La famille des NaV est composée de 10 isoformes qui présentent une distribution
tissulaire différente. Par exemple, on retrouve dans le système nerveux périphérique les
isoformes NaV1.3, NaV1.7, NaV1.8 et NaV . . Da s le œu l’isofo e NaV1.5 est prédominante
(Moreau et al., 2017).
Dans les CSCs W8B2+ les résultats obtenus (figure R-9) montrent que les sous-unités
alpha des isoformes NaV1.8 NaV1.3, NaV1.4, NaV1.5, NaV1.7 et NaV2.1 sont absentes. Seule une
faible expression du NaV1.3 est détectée. Concernant les sous-unités bêta régulatrices, les
CSCs W8B2+ ’expriment uniquement que la sous-u it β NaVb1).
Figure R-9 : P ofil d’e p essio g i ue des a au sodi ues oltage-dépendant dans les CSCs
W8B2+ déterminé par RT- PCR. = . Le P ofil d’e p essio des g es est ep se t e -ΔCt par
rapport au gène de référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque barre
représente la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre
de patients à partir desquels sont issues les CSCs W8B2+.
Navb1
Navb2
Navb3
Navb4
Nav1.3
Nav1.4
Nav1.5
Nav1.7
Nav1.8
Nav2.1
0
100
200
300
400
2- Ct v
s R
PL1
3A (x
104 )

Résultats
121
2.2 P ofil d’e p essio g i ue des a au potassi ues da s les CSCs W B +
Les canaux potassiques participent à de nombreuses fonctions biologiques et sont
notamment responsables du maintien du potentiel membranaire, de la transmission de
l'influx nerveux et de la rythmogenèse.
Les canaux potassiques sont très variés et peuvent être regroupés en 4 grandes familles :
-Les canaux potassiques dépendant du potentiel (Kv), 6 segments transmembranaires (STM)
et d’u e ou le P fo a t le filtre de sélectivité du canal :
Cette famille regroupe la sous-famille : des KV1.x appelée « shaker », des KV2.x appelée
«shab », des KV3.x appelée «shaw », des KV4.x appelée «shal », des KV7.x appelée «KQT » et
la sous-famille des KV10.x KV11.x et KV12.x appelée « eag » (Luneau et al., 1991; Perney and
Kaczmarek, 1991; Stühmer et al., 1989).
Les résultats obtenus par RT-qPCR à haut rendement ou TLDA montrent que les CSCs
W8B2+ expriment fortement le KV3.4 et faiblement le KV4.2, des canaux potassiques
cardiaques impliqués dans le courant transitoire sortant repolarisant Ito (Han et al., 2002;
Nerbonne, 2016) (figure R-10). Concernant les canaux KV1.2, KV1.4, KV1.5, KV1.7, KV2.1, KV4.3,
KV7.1 (KVLQT1) et KV . hERG , au u e e p essio sig ifi ati e ’est d te t e.
De plus, les résultats des RT-PCR (figure R-11) confirment et montrent que les CSCs
W8B2+ ’e p i e t pas le t a s it KV4.3 et modérément le KV4.2.
E effet, l’isofo e KV4.2 (220 pb) est exprimée à la fois dans les CSCs W8B2+ et dans
le tissu atrial humain utilisé comme contrôle positif.
Qua t à l’isofo e KV4.3, nous obtenons une bande de 310 pb seulement dans le tissu
atrial humain (contrôle positif).
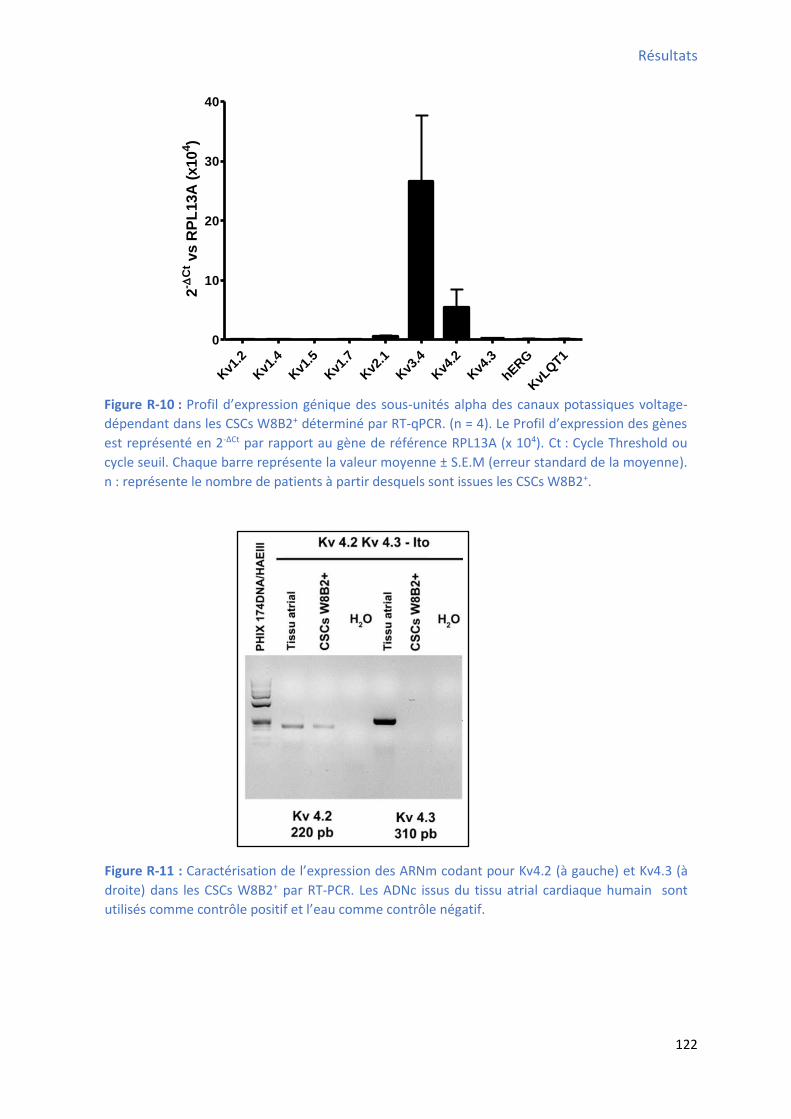
Résultats
122
Figure R-10 : P ofil d’e p essio g i ue des sous-unités alpha des canaux potassiques voltage-
dépendant dans les CSCs W8B2+ déterminé par RT- PCR. = . Le P ofil d’e p essio des g es est représenté en 2-ΔCt par rapport au gène de référence RPL13A (x 104). Ct : Cycle Threshold ou
cycle seuil. Chaque barre représente la valeur moyenne ± S.E.M (erreur standard de la moyenne).
n : représente le nombre de patients à partir desquels sont issues les CSCs W8B2+.
Kv1.2
Kv1.4
Kv1.5
Kv1.7
Kv2.1
Kv3.4
Kv4.2
Kv4.3
hERG
KvLQT1
0
10
20
30
40
2- Ct v
s R
PL1
3A (x
104 )
Figure R-11 : Ca a t isatio de l’e p essio des ARN oda t pou K . (à gauche) et Kv4.3 (à
droite) dans les CSCs W8B2+ par RT-PCR. Les ADNc issus du tissu atrial cardiaque humain sont
utilis s o e o t ôle positif et l’eau o e o t ôle gatif.
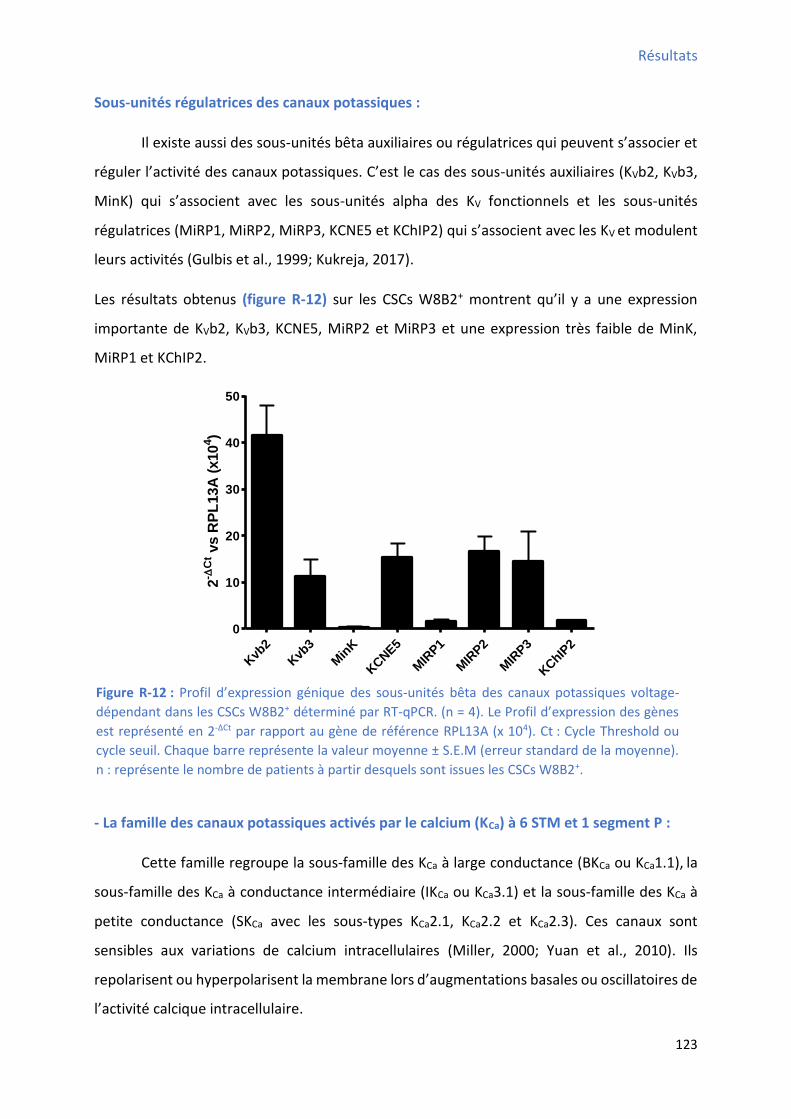
Résultats
123
Sous-unités régulatrices des canaux potassiques :
Il existe aussi des sous-u it s ta au iliai es ou gulat i es ui peu e t s’asso ie et
gule l’a ti it des a au potassi ues. C’est le as des sous-unités auxiliaires (KVb2, KVb3,
Mi K ui s’asso ie t a e les sous-unités alpha des KV fonctionnels et les sous-unités
régulatrices (MiRP1, MiRP2, MiRP3, KCNE et KChIP ui s’asso ie t a e les KV et modulent
leurs activités (Gulbis et al., 1999; Kukreja, 2017).
Les résultats obtenus (figure R-12) sur les CSCs W8B2+ mo t e t u’il a u e e p essio
importante de KVb2, KVb3, KCNE5, MiRP2 et MiRP3 et une expression très faible de MinK,
MiRP1 et KChIP2.
- La famille des canaux potassiques activés par le calcium (KCa) à 6 STM et 1 segment P :
Cette famille regroupe la sous-famille des KCa à large conductance (BKCa ou KCa1.1), la
sous-famille des KCa à conductance intermédiaire (IKCa ou KCa3.1) et la sous-famille des KCa à
petite conductance (SKCa avec les sous-types KCa2.1, KCa2.2 et KCa2.3). Ces canaux sont
sensibles aux variations de calcium intracellulaires (Miller, 2000; Yuan et al., 2010). Ils
repolarisent ou h pe pola ise t la e a e lo s d’aug e tatio s asales ou os illatoi es de
l’a ti it al i ue i t a ellulai e.
Figure R-12 : P ofil d’e p essio g i ue des sous-unités bêta des canaux potassiques voltage-
dépendant dans les CSCs W8B2+ déterminé par RT- PCR. = . Le P ofil d’e p essio des g es est représenté en 2-ΔCt par rapport au gène de référence RPL13A (x 104). Ct : Cycle Threshold ou
cycle seuil. Chaque barre représente la valeur moyenne ± S.E.M (erreur standard de la moyenne).
n : représente le nombre de patients à partir desquels sont issues les CSCs W8B2+.
Kvb2
Kvb3
MinK
KCNE5
MIRP1
MIRP2
MIRP3
KChIP2
0
10
20
30
40
50
2- Ct v
s R
PL1
3A (x
104 )
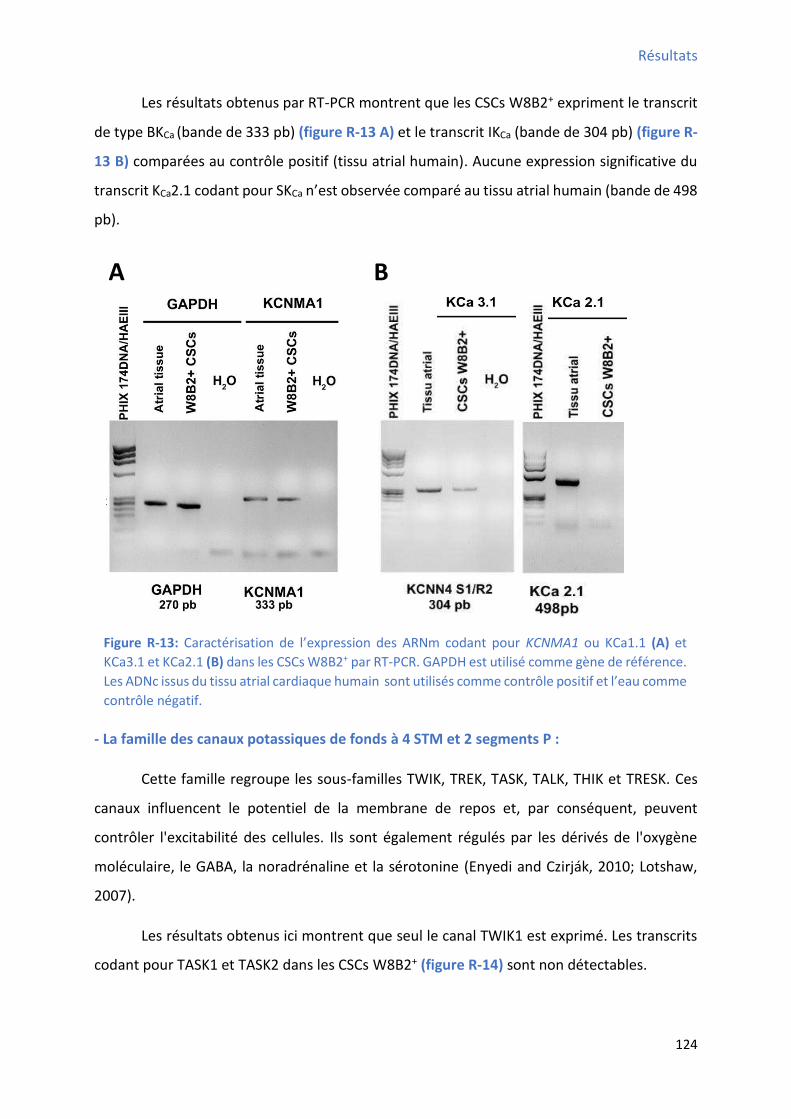
Résultats
124
Les résultats obtenus par RT-PCR montrent que les CSCs W8B2+ expriment le transcrit
de type BKCa (bande de 333 pb) (figure R-13 A) et le transcrit IKCa (bande de 304 pb) (figure R-
13 B) comparées au contrôle positif (tissu atrial humain). Aucune expression significative du
transcrit KCa2.1 codant pour SKCa ’est o se e comparé au tissu atrial humain (bande de 498
pb).
- La famille des canaux potassiques de fonds à 4 STM et 2 segments P :
Cette famille regroupe les sous-familles TWIK, TREK, TASK, TALK, THIK et TRESK. Ces
canaux influencent le potentiel de la membrane de repos et, par conséquent, peuvent
contrôler l'excitabilité des cellules. Ils sont également régulés par les dérivés de l'oxygène
moléculaire, le GABA, la noradrénaline et la sérotonine (Enyedi and Czirják, 2010; Lotshaw,
2007).
Les résultats obtenus ici montrent que seul le canal TWIK1 est exprimé. Les transcrits
codant pour TASK1 et TASK2 dans les CSCs W8B2+ (figure R-14) sont non détectables.
B A
Figure R-13: Ca a t isatio de l’e p essio des ARN oda t pou KCNMA1 ou KCa1.1 (A) et
KCa3.1 et KCa2.1 (B) dans les CSCs W8B2+ par RT-PCR. GAPDH est utilisé comme gène de référence.
Les ADNc issus du tissu atrial cardiaque humain sont utilisés comme contrôle positif et l’eau o e contrôle négatif.
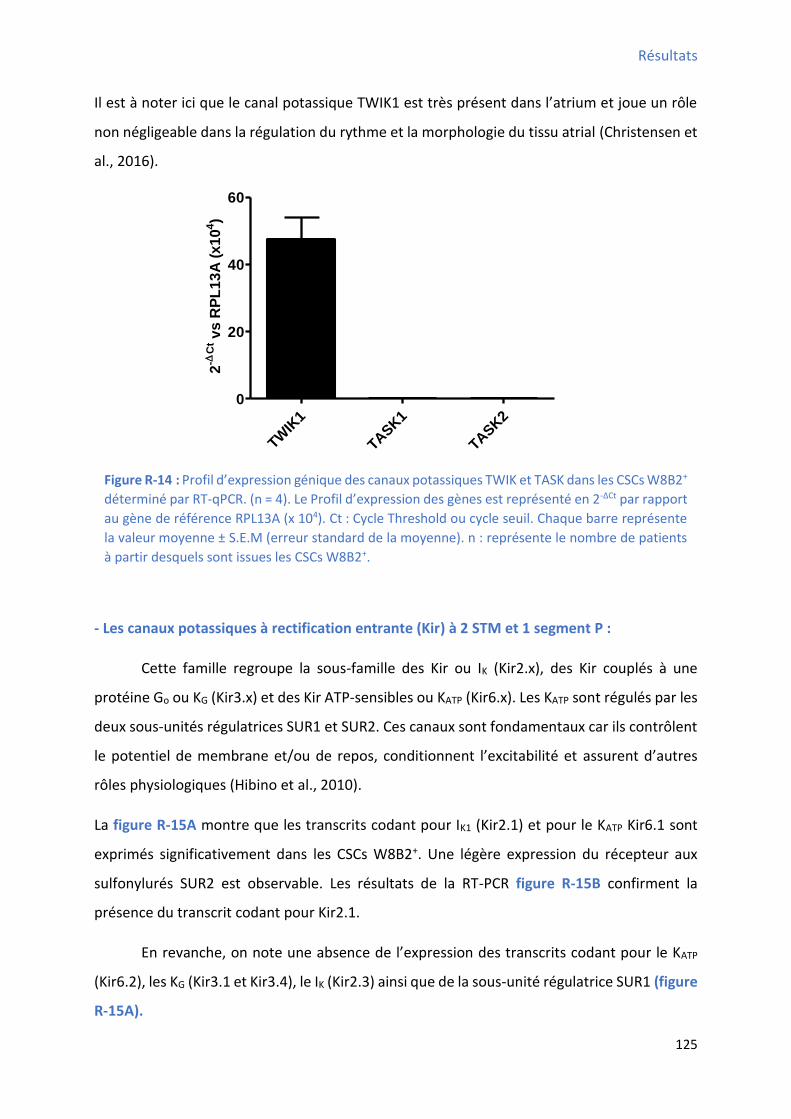
Résultats
125
Il est à ote i i ue le a al potassi ue TWIK est t s p se t da s l’at iu et joue u ôle
non négligeable dans la régulation du rythme et la morphologie du tissu atrial (Christensen et
al., 2016).
- Les canaux potassiques à rectification entrante (Kir) à 2 STM et 1 segment P :
Cette famille regroupe la sous-famille des Kir ou IK (Kir2.x), des Kir couplés à une
protéine Go ou KG (Kir3.x) et des Kir ATP-sensibles ou KATP (Kir6.x). Les KATP sont régulés par les
deux sous-unités régulatrices SUR1 et SUR2. Ces canaux sont fondamentaux car ils contrôlent
le pote tiel de e a e et/ou de epos, o ditio e t l’e ita ilit et assu e t d’aut es
rôles physiologiques (Hibino et al., 2010).
La figure R-15A montre que les transcrits codant pour IK1 (Kir2.1) et pour le KATP Kir6.1 sont
exprimés significativement dans les CSCs W8B2+. Une légère expression du récepteur aux
sulfonylurés SUR2 est observable. Les résultats de la RT-PCR figure R-15B confirment la
présence du transcrit codant pour Kir2.1.
En revanche, on note une absence de l’e p essio des t a s its oda t pou le KATP
(Kir6.2), les KG (Kir3.1 et Kir3.4), le IK (Kir2.3) ainsi que de la sous-unité régulatrice SUR1 (figure
R-15A).
Figure R-14 : P ofil d’e p essio g i ue des a au potassi ues TWIK et TA“K da s les C“Cs W B +
déterminé par RT- PCR. = . Le P ofil d’e p essio des g es est ep se t e -ΔCt par rapport
au gène de référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque barre représente
la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients
à partir desquels sont issues les CSCs W8B2+.
TWIK
1
TASK1
TASK20
20
40
602- C
t vs
RP
L13A
(x10
4 )
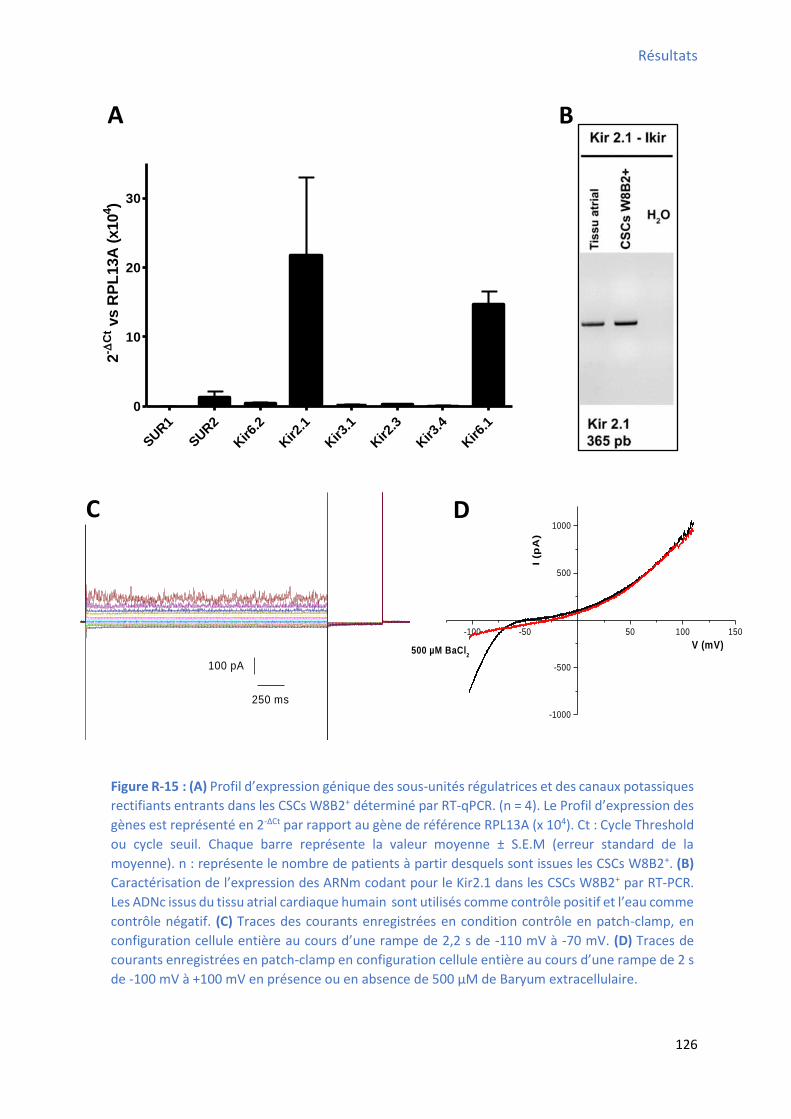
Résultats
126
Figure R-15 : (A) P ofil d’e p essio g i ue des sous-unités régulatrices et des canaux potassiques
rectifiants entrants dans les CSCs W8B2+ déterminé par RT- PCR. = . Le P ofil d’e p essio des gènes est représenté en 2-ΔCt par rapport au gène de référence RPL13A (x 104). Ct : Cycle Threshold
ou cycle seuil. Chaque barre représente la valeur moyenne ± S.E.M (erreur standard de la
moyenne). n : représente le nombre de patients à partir desquels sont issues les CSCs W8B2+. (B)
Ca a t isatio de l’e p essio des ARN odant pour le Kir2.1 dans les CSCs W8B2+ par RT-PCR.
Les ADN issus du tissu at ial a dia ue hu ai so t utilis s o e o t ôle positif et l’eau o e contrôle négatif. (C) Traces des courants enregistrées en condition contrôle en patch-clamp, en
configuration cellule entière au ou s d’u e a pe de , s de -110 mV à -70 mV. (D) Traces de
courants enregistrées en patch- la p e o figu atio ellule e ti e au ou s d’u e a pe de s
de -100 mV à +100 mV en présence ou en absence de 500 µM de Baryum extracellulaire.
SUR1
SUR2
Kir6.2
Kir2.1
Kir3.1
Kir2.3
Kir3.4
Kir6.1
0
10
20
30
2- Ct v
s R
PL1
3A (x
104 )
B A
100 pA
250 ms
-100 -50 50 100 150
-1000
-500
500
1000
500 µM BaCl 2
I (p
A)
V (mV)
C D
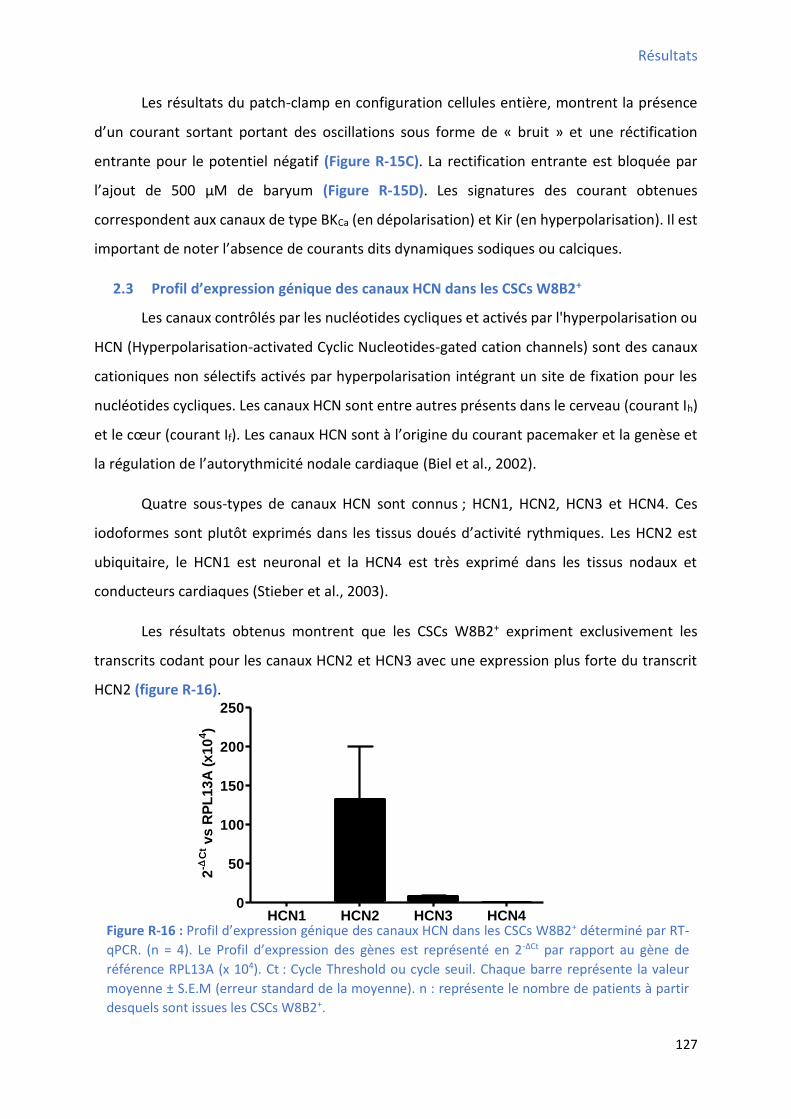
Résultats
127
Les résultats du patch-clamp en configuration cellules entière, montrent la présence
d’u ou a t sortant portant des oscillations sous forme de « bruit » et une réctification
entrante pour le potentiel négatif (Figure R-15C). La rectification entrante est bloquée par
l’ajout de µM de a u (Figure R-15D). Les signatures des courant obtenues
correspondent aux canaux de type BKCa (en dépolarisation) et Kir (en hyperpolarisation). Il est
i po ta t de ote l’a se e de ou ants dits dynamiques sodiques ou calciques.
2.3 P ofil d’e p essio g i ue des a au HCN da s les CSCs W B +
Les canaux contrôlés par les nucléotides cycliques et activés par l'hyperpolarisation ou
HCN (Hyperpolarisation-activated Cyclic Nucleotides-gated cation channels) sont des canaux
cationiques non sélectifs activés par hyperpolarisation intégrant un site de fixation pour les
nucléotides cycliques. Les canaux HCN sont entre autres présents dans le cerveau (courant Ih)
et le œu ou a t If). Les canaux HCN so t à l’o igi e du ou a t pa e ake et la ge se et
la gulatio de l’auto th i it odale a dia ue (Biel et al., 2002).
Quatre sous-types de canaux HCN sont connus ; HCN1, HCN2, HCN3 et HCN4. Ces
iodofo es so t plutôt e p i s da s les tissus dou s d’a ti it th i ues. Les HCN est
ubiquitaire, le HCN1 est neuronal et la HCN4 est très exprimé dans les tissus nodaux et
conducteurs cardiaques (Stieber et al., 2003).
Les résultats obtenus montrent que les CSCs W8B2+ expriment exclusivement les
transcrits codant pour les canaux HCN2 et HCN3 avec une expression plus forte du transcrit
HCN2 (figure R-16).
Figure R-16 : P ofil d’e p essio g i ue des a au HCN da s les C“Cs W B + déterminé par RT-
PCR. = . Le P ofil d’e p essio des g es est ep se t e -ΔCt par rapport au gène de
référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque barre représente la valeur
moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir
desquels sont issues les CSCs W8B2+.
HCN1 HCN2 HCN3 HCN40
50
100
150
200
250
2- Ct v
s R
PL1
3A (x
104 )

Résultats
128
2.4 P ofil d’e p essio g i ue des a au al i ues da s les CSCs W B +
Les canaux calciques sont impliqués dans de nombreuses fonctions cellulaires allant du
couplage excitation- o t a tio us ulai e à la li atio d’ho o es ou de
neurotransmetteurs.
On décrit deux types de canaux calciques ; les canaux calciques récepteurs
dépendants do t l’ou e tu e est o te ue pa l'a tio des diateu s hi i ues et/ou les
canaux calciques potentiel-dépendants qui s'ouvrent grâce aux variations de voltage.
Les canaux calciques potentiel-dépendants (CaV so t o stitu s d’u e sous-unité
canalaire alpha (qui apporte la diversité fonctionnelle) associée aux sous-unités régulatrices
bêta et alpha 2-delta.
Les CaV sont eux-mêmes classés en plusieurs types (Catterall, 2011) ;
-Les CaV de type L (CaV 1.x) : exprimés dans les muscles (lisse, squelettique et cardiaque), les
neurones, les cellules endocrines et la rétine.
-Les CaV de type P/Q (CaV 2.1) : e p i s da s les eu o es et les ellules β pa ati ues.
-Les CaV de type N (CaV 2.2) : exprimés dans les neurones.
-Les CaV de type R (CaV 2.3) : exprimés dans les neurones et les cellules endocrines.
-Les CaV de type T (CaV 3.x) : exprimés dans les muscles (lisse et cardiaque), les neurones et
les cellules endocrines.
Dans notre étude, nous avons ciblé les CaV a dia ues à as et haut seuil d’a ti atio
(CaV 1.2, CaV 1.3, CaV 3.1 et CaV 3.2) et à leurs sous-u it s gulat i es α , α et β .
Les résultats figure R-17 montrent que les CSCs W8B2+ ’e p i e t au u t a s it
codant pour les CaV cardiaques et les sous-u it s gulat i es α et β . La sous-unité
gulat i e α ua t à elle, p se te u e e p essio t s fo te du t a s it. Ces sultats
sugg e t u’au u e pe a ilit fo tio elle al i ue d pe da te du oltage ’est
présente dans les progéniteurs cardiaques.
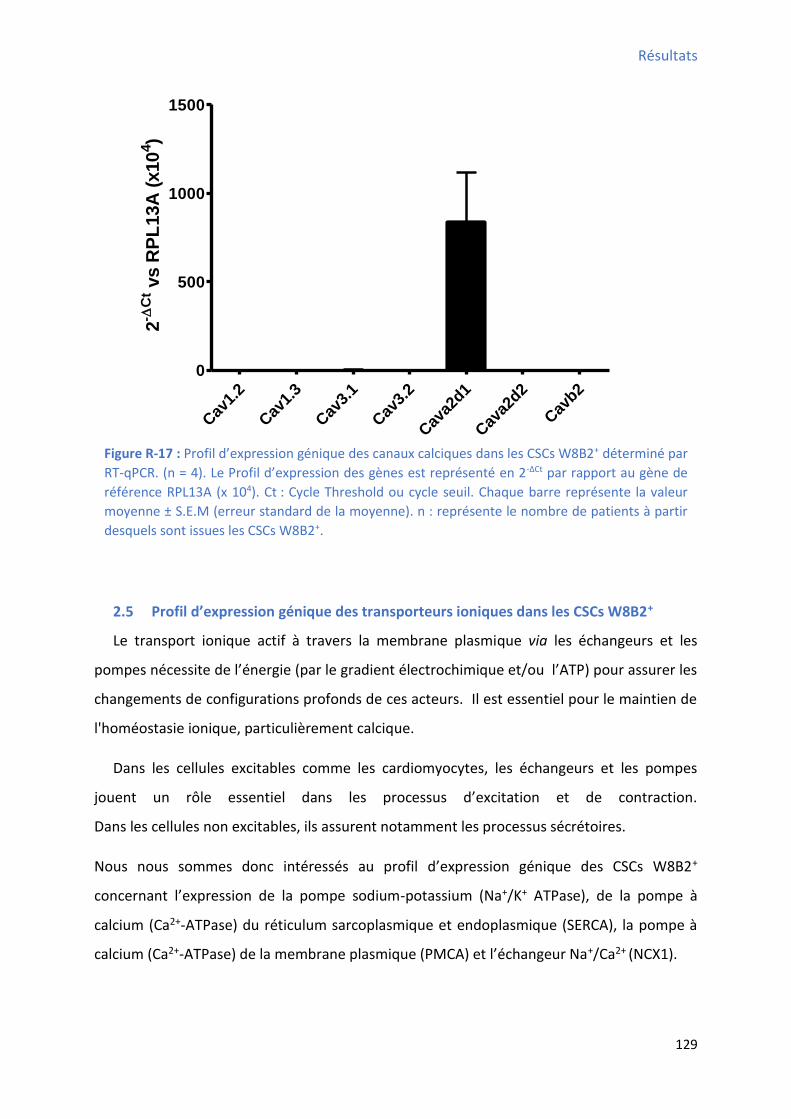
Résultats
129
2.5 P ofil d’e p essio g i ue des t a spo teu s io i ues da s les CSCs W B +
Le transport ionique actif à travers la membrane plasmique via les échangeurs et les
po pes essite de l’ e gie pa le g adie t le t o hi i ue et/ou l’ATP pou assu e les
changements de configurations profonds de ces acteurs. Il est essentiel pour le maintien de
l'homéostasie ionique, particulièrement calcique.
Dans les cellules excitables comme les cardiomyocytes, les échangeurs et les pompes
joue t u ôle esse tiel da s les p o essus d’e itatio et de o t a tio .
Dans les cellules non excitables, ils assurent notamment les processus sécrétoires.
Nous nous so es do i t ess s au p ofil d’e p essio g i ue des C“Cs W B +
o e a t l’e p essio de la po pe sodiu -potassium (Na+/K+ ATPase), de la pompe à
calcium (Ca2+-ATPase) du réticulum sarcoplasmique et endoplasmique (SERCA), la pompe à
calcium (Ca2+-ATPase de la e a e plas i ue PMCA et l’ ha geu Na+/Ca2+ (NCX1).
Figure R-17 : P ofil d’e p essio g i ue des canaux calciques dans les CSCs W8B2+ déterminé par
RT- PCR. = . Le P ofil d’e p essio des g es est ep se t e -ΔCt par rapport au gène de
référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque barre représente la valeur
moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir
desquels sont issues les CSCs W8B2+.
Cav1.2
Cav1.3
Cav3.1
Cav3.2
Cava2
d1
Cava2
d2
Cavb2
0
500
1000
1500
2- Ct v
s R
PL1
3A (x
104 )
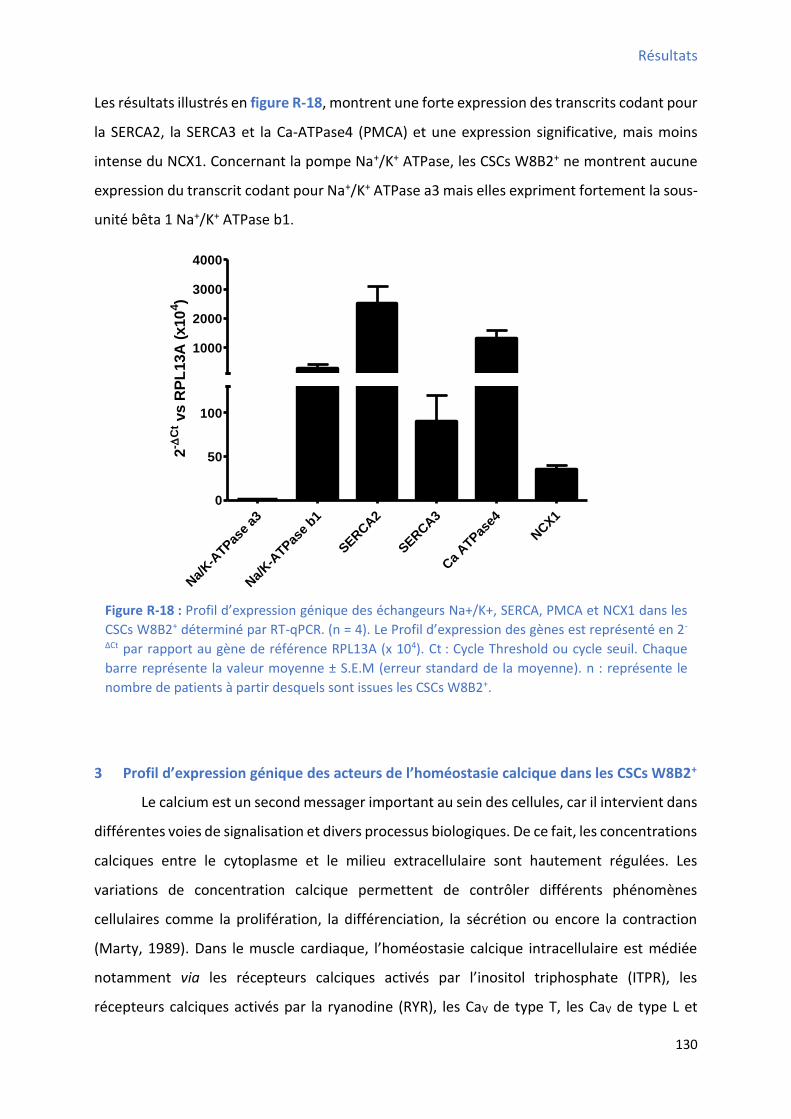
Résultats
130
Les résultats illustrés en figure R-18, montrent une forte expression des transcrits codant pour
la SERCA2, la SERCA3 et la Ca-ATPase4 (PMCA) et une expression significative, mais moins
intense du NCX1. Concernant la pompe Na+/K+ ATPase, les CSCs W8B2+ ne montrent aucune
expression du transcrit codant pour Na+/K+ ATPase a3 mais elles expriment fortement la sous-
unité bêta 1 Na+/K+ ATPase b1.
3 P ofil d’e p essio g i ue des a teu s de l’ho ostasie al i ue da s les CSCs W B +
Le calcium est un second messager important au sein des cellules, car il intervient dans
différentes voies de signalisation et divers processus biologiques. De ce fait, les concentrations
calciques entre le cytoplasme et le milieu extracellulaire sont hautement régulées. Les
variations de concentration calcique permettent de contrôler différents phénomènes
cellulaires comme la prolifération, la différenciation, la sécrétion ou encore la contraction
(Marty, 1989). Da s le us le a dia ue, l’ho ostasie al i ue i t a ellulai e est di e
notamment via les epteu s al i ues a ti s pa l’i ositol t iphosphate ITPR , les
récepteurs calciques activés par la ryanodine (RYR), les CaV de type T, les CaV de type L et
Figure R-18 : P ofil d’e p essio g i ue des ha geu s Na+/K+, “ERCA, PMCA et NCX da s les CSCs W8B2+ déterminé par RT- PCR. = . Le P ofil d’e p essio des g es est ep se t e -
ΔCt par rapport au gène de référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque
barre représente la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le
nombre de patients à partir desquels sont issues les CSCs W8B2+.
Na/K-A
TPase a
3
Na/K-A
TPase b
1
SERCA2
SERCA3
Ca ATPas
e4NCX1
0
50
100
1000
2000
3000
4000
2- Ct v
s R
PL1
3A (x
104 )
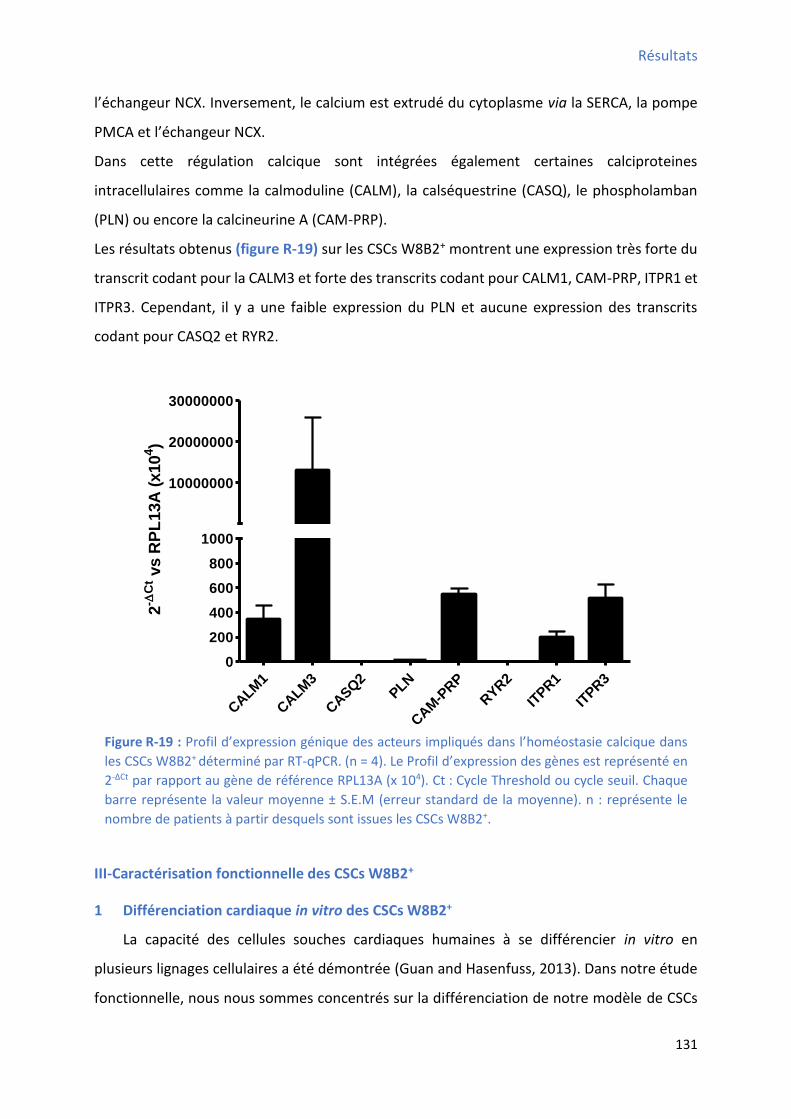
Résultats
131
l’ ha geu NCX. I e se e t, le al iu est e t ud du toplas e via la SERCA, la pompe
PMCA et l’ ha geu NCX.
Dans cette régulation calcique sont intégrées également certaines calciproteines
intracellulaires comme la calmoduline (CALM), la calséquestrine (CASQ), le phospholamban
(PLN) ou encore la calcineurine A (CAM-PRP).
Les résultats obtenus (figure R-19) sur les CSCs W8B2+ montrent une expression très forte du
transcrit codant pour la CALM3 et forte des transcrits codant pour CALM1, CAM-PRP, ITPR1 et
ITPR3. Cependant, il y a une faible expression du PLN et aucune expression des transcrits
codant pour CASQ2 et RYR2.
III-Caractérisation fonctionnelle des CSCs W8B2+
1 Différenciation cardiaque in vitro des CSCs W8B2+
La capacité des cellules souches cardiaques humaines à se différencier in vitro en
plusieurs lignages cellulaires a été démontrée (Guan and Hasenfuss, 2013). Dans notre étude
fonctionnelle, nous nous sommes concentrés sur la différenciation de notre modèle de CSCs
Figure R-19 : P ofil d’e p essio g i ue des a teu s i pli u s da s l’ho ostasie al i ue da s les CSCs W8B2+ déterminé par RT-qPCR. (n = 4). Le Profil d’e p essio des g es est ep se t e 2-ΔCt par rapport au gène de référence RPL13A (x 104). Ct : Cycle Threshold ou cycle seuil. Chaque
barre représente la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le
nombre de patients à partir desquels sont issues les CSCs W8B2+.
CALM1
CALM3
CASQ2PLN
CAM-PRP
RYR2
ITPR1
ITPR3
0
200
400
600
800
1000
10000000
20000000
30000000
2- Ct v
s R
PL1
3A (x
104 )

Résultats
132
W8B2+ e lig age a dio o tai e. L’ tude des a is es i pli u s da s ette
diff e iatio e a dio o tes est fo da e tale a elle pe et d’o ie te et d’i agi e
des solutio s th apeuti ues de de i e g at i e. Le ut ulti e est d’obtenir des
ellules p og it i es ui u e fois g eff es, se aie t apa les de pa e le œu pa
e pla e e t ou e fo e e t des a dio o tes da s les as d’i suffisa e a dia ue
grave.
Les cardiomyocytes dérivés de cellules souches cardiaques après différenciation in vitro
décrits à ce jour, présentent des caractéristiques fonctionnelles et structurelles plutôt
immatures par rapport aux cardiomyocytes adultes natifs. Ainsi, un défi futur sera de
développer des stratégies pour atteindre un degré plus élevé de maturation des
cardiomyocytes in vitro.
Dans cette partie, nous avons suivi le processus de différenciation cardiaque in vitro des
CSCs W8B2+ e a al sa t l’e p essio g i ue des a ueu s a dia ues ou a e t utilis s
dans la littérature (structures contractiles, facteurs de transcription cardiaque, connexines
et … ais aussi le p ofil g i ue des a au io i ues et des a teu s de l’ho ostasie calcique
a au sodi ues, potassi ues, al i ues, po pes, ha geu s et … .
1.1 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des marqueurs cardiaques
1.1.1 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des facteurs de transcription cardiaques
Les facteurs de transcriptions cardiaques sont primordiaux pour le développement et
la morphogenèse cardiaque et sont utilisés pour caractériser les différents types de cellules
souches ca dia ues. Il est do i po ta t d’a al se l’ olutio de es fa teu s ap s
différenciation.
Les résultats montrent que les transcrits codant pour GATA5 et GATA6 sont fortement
augmentés (+ 55,3 fois pour GATA5 et + 14 ,4 fois pour GATA6) (figure R-20A). L’e p essio
de GATA3 et TBX5 est légèrement induite (+ 2,6 fois pour GATA3 et + 2,9 fois pour TBX5) et
au u e a iatio de l’e p essio g i ue des fa teu s de t a s iptio GATA , HEY , IRX ,
IRX , TBX et NKX . ’est o se e. Co e a t les deu fa teurs de transcription IRX3 et
TBX3, une légère répression de leurs transcrits est observée (- 3,8 fois pour IRX3 et -2,5 fois
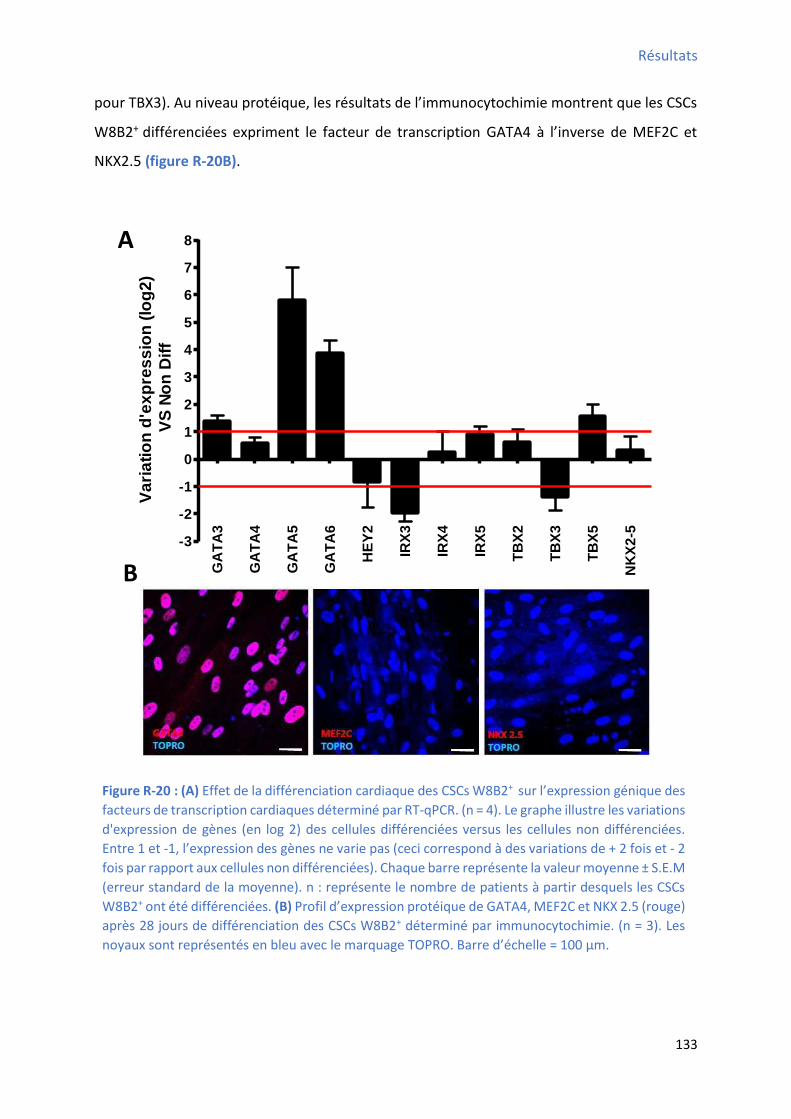
Résultats
133
pou TBX . Au i eau p ot i ue, les sultats de l’i u o to hi ie o t e t ue les C“Cs
W8B2+ diff e i es e p i e t le fa teu de t a s iptio GATA à l’i e se de MEF C et
NKX2.5 (figure R-20B).
GA
TA
3
GA
TA
4
GA
TA
5
GA
TA
6
HE
Y2
IRX
3
IRX
4
IRX
5
TB
X2
TB
X3
TB
X5
NK
X2-
5
-3
-2
-1
0
1
2
3
4
5
6
7
8
Var
iatio
n d'
expr
essi
on (l
og2)
VS
Non
Diff
Figure R-20 : (A) Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des facteurs de transcription cardiaques déterminé par RT-qPCR. (n = 4). Le graphe illustre les variations
d'expression de gènes (en log 2) des cellules différenciées versus les cellules non différenciées.
Entre 1 et - , l’e p essio des g es e a ie pas e i o espo d à des a iatio s de + fois et - 2
fois par rapport aux cellules non différenciées). Chaque barre représente la valeur moyenne ± S.E.M
(erreur standard de la moyenne). n : représente le nombre de patients à partir desquels les CSCs
W8B2+ ont été différenciées. (B) P ofil d’e p ession protéique de GATA4, MEF2C et NKX 2.5 (rouge)
après 28 jours de différenciation des CSCs W8B2+ déterminé par immunocytochimie. (n = 3). Les
o au so t ep se t s e leu a e le a uage TOPRO. Ba e d’ helle = µ .
B
A

Résultats
134
1.1.2 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des structures contractiles cardiaques
La mise en place des structures contractiles cardiaques après les protocoles de
différenciation cardiaque in vitro est souvent observée pour confirmer la présence ou non
d’u ph ot pe a dia ue ap s diff e iatio des ellules sou hes a dia ues.
Nous nous sommes donc intéressés aux marqueurs de différenciation cardiaque
o e la t opo i e, la osi e, l’a k i e ou e o e les peptides natriurétiques.
Les résultats obtenus par RT-qPCR à haut rendement ou TLDA (figure R-21A), montrent
ue l’e p essio des t a s its oda t pou l’ANKB et MYL est fo te e t i duite au ou s de
la différenciation cardiaque (induction de + 137 fois pour ANKB et + 13 fois pour MYL7). A
moindre intensité il en est de même pour les transcrits TNNT2 et e GPD1L (induction de + 3,3
fois pour la TNNT2 et + 2,9 fois pour GPD1L).
Au u e odifi atio de l’e p essio de CLA“P et de MYL ’est isi le ; en effet, entre + 2
fois et – 2 fois par rapport aux CSCs W8B2+ o diff e i es. E e a he, l’e p essio des
transcrits codant pour la TNNI3, MYH6, MYH7 et RRAD est réprimé au cours de la
différenciation (répression de – 5,2 fois pour la TNNI3, - 4,7 fois pour MYH6, - 3.1 fois pour
MYH7 et - 3,2 fois pour RRAD).
Les i u o a uages i di ue t aussi u’il ’ a pas de st u tu es o t a tiles
matures (absence de striations typiques des a dio o te o e l’i di ue le marquage de
TNNT2, a-actinine et b-MHC (figure R-21B).
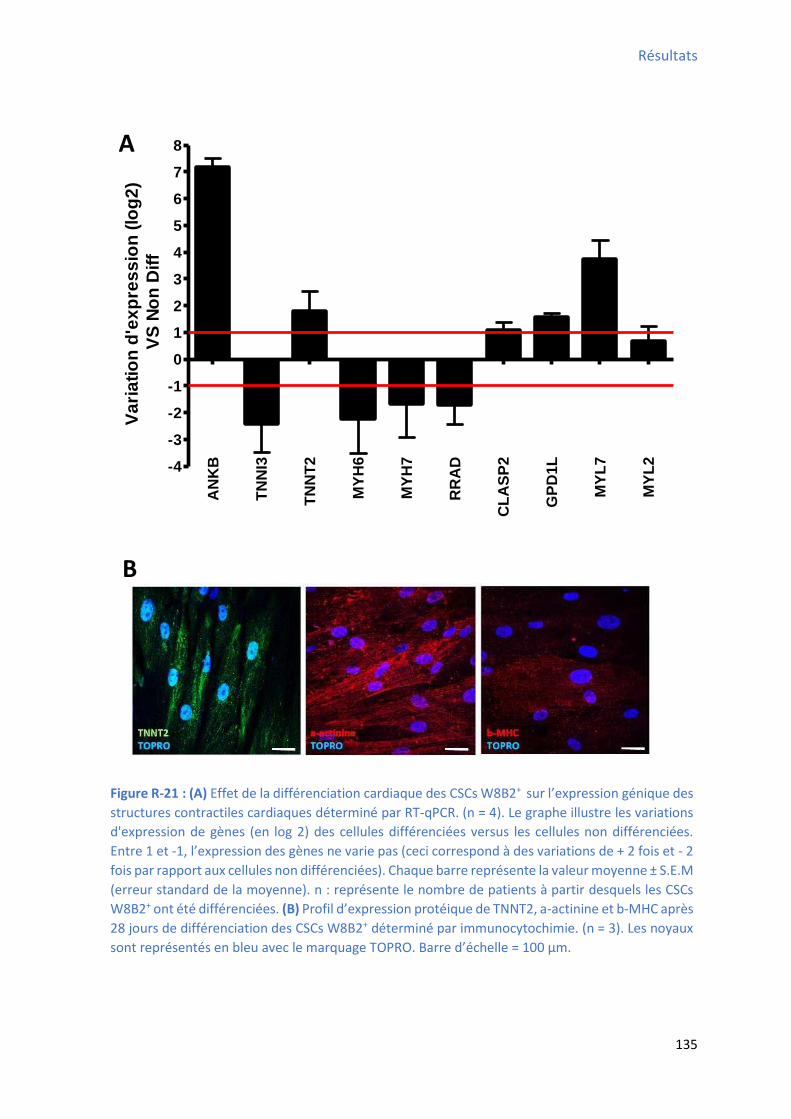
Résultats
135
AN
KB
TN
NI3
TN
NT
2
MY
H6
MY
H7
RR
AD
CLA
SP
2
GP
D1L
MY
L7
MY
L2-4
-3
-2
-1
0
1
2
3
4
5
6
7
8V
aria
tion
d'ex
pres
sion
(log
2) V
S N
on D
iff
Figure R-21 : (A) Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des structures contractiles cardiaques déterminé par RT-qPCR. (n = 4). Le graphe illustre les variations
d'expression de gènes (en log 2) des cellules différenciées versus les cellules non différenciées.
Entre 1 et - , l’e p ession des gènes ne varie pas (ceci correspond à des variations de + 2 fois et - 2
fois par rapport aux cellules non différenciées). Chaque barre représente la valeur moyenne ± S.E.M
(erreur standard de la moyenne). n : représente le nombre de patients à partir desquels les CSCs
W8B2+ ont été différenciées. (B) P ofil d’e p essio p ot i ue de TNNT , a-actinine et b-MHC après
28 jours de différenciation des CSCs W8B2+ déterminé par immunocytochimie. (n = 3). Les noyaux
sont représentés en bleu avec le marquage TOPRO. Ba e d’ helle = µ .
A
B
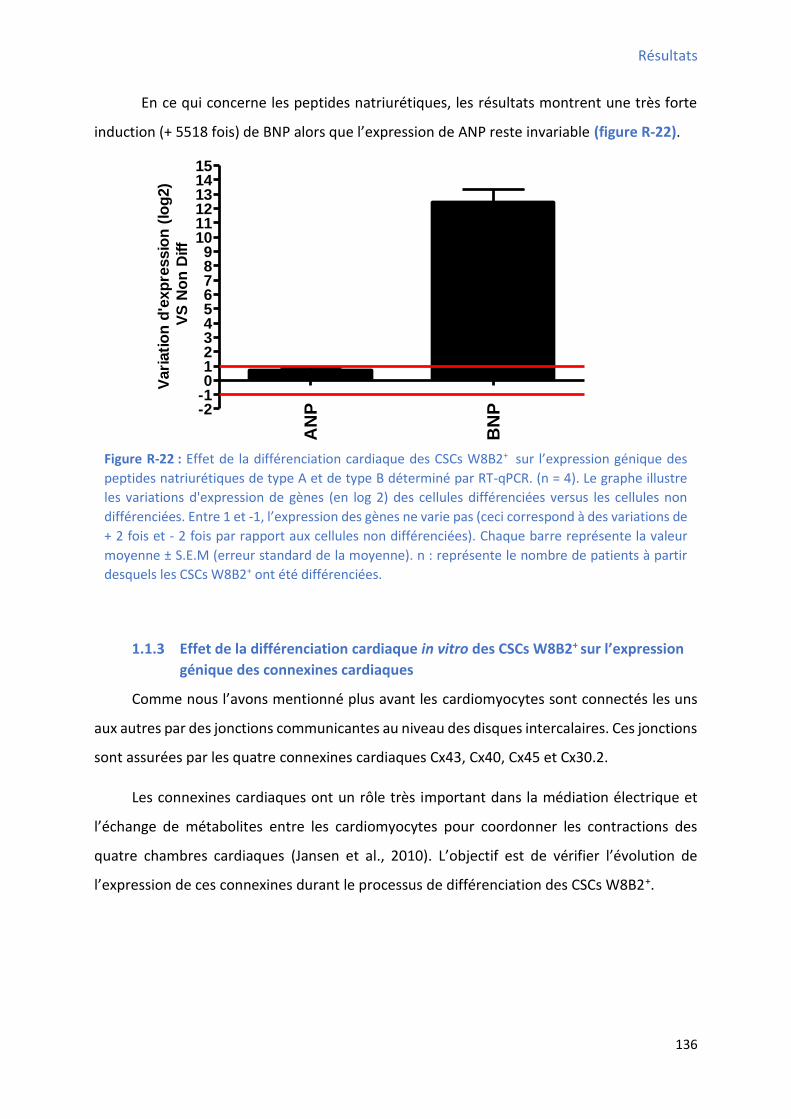
Résultats
136
En ce qui concerne les peptides natriurétiques, les résultats montrent une très forte
indu tio + fois de BNP alo s ue l’e p essio de ANP este i a ia le (figure R-22).
1.1.3 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des connexines cardiaques
Co e ous l’a o s e tio plus a a t les cardiomyocytes sont connectés les uns
aux autres par des jonctions communicantes au niveau des disques intercalaires. Ces jonctions
sont assurées par les quatre connexines cardiaques Cx43, Cx40, Cx45 et Cx30.2.
Les connexines cardiaques ont un rôle très important dans la médiation électrique et
l’ ha ge de ta olites entre les cardiomyocytes pour coordonner les contractions des
quatre chambres cardiaques (Jansen et al., 2010). L’o je tif est de ifie l’ olutio de
l’e p essio de es o e i es du a t le p o essus de diff e iatio des C“Cs W B +.
Figure R-22 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des peptides natriurétiques de type A et de type B déterminé par RT-qPCR. (n = 4). Le graphe illustre
les variations d'expression de gènes (en log 2) des cellules différenciées versus les cellules non
différenciées. Entre 1 et - , l’e p essio des g es e a ie pas e i o espo d à des a iatio s de + 2 fois et - 2 fois par rapport aux cellules non différenciées). Chaque barre représente la valeur
moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir
desquels les CSCs W8B2+ ont été différenciées.
AN
P
BN
P-2-10123456789
101112131415
Var
iatio
n d'
expr
essi
on (
log2
) V
S N
on D
iff
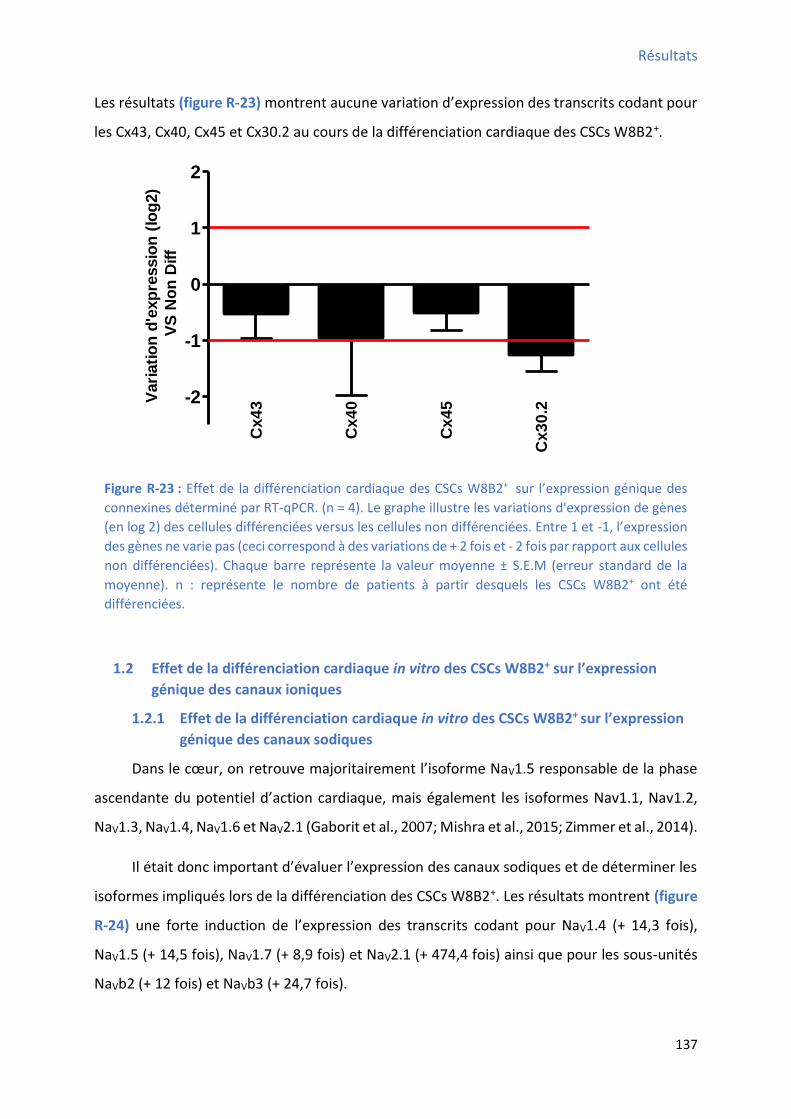
Résultats
137
Les résultats (figure R-23) montre t au u e a iatio d’e p essio des t a s its oda t pou
les Cx43, Cx40, Cx45 et Cx30.2 au cours de la différenciation cardiaque des CSCs W8B2+.
1.2 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des canaux ioniques
1.2.1 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des canaux sodiques
Da s le œu , o et ou e ajo itai e e t l’isofo e NaV1.5 responsable de la phase
as e da te du pote tiel d’a tio a dia ue, ais gale e t les isoformes Nav1.1, Nav1.2,
NaV1.3, NaV1.4, NaV1.6 et NaV2.1 (Gaborit et al., 2007; Mishra et al., 2015; Zimmer et al., 2014).
Il tait do i po ta t d’ alue l’e p essio des a au sodi ues et de d te i e les
isoformes impliqués lors de la différenciation des CSCs W8B2+. Les résultats montrent (figure
R-24) u e fo te i du tio de l’e pression des transcrits codant pour NaV1.4 (+ 14,3 fois),
NaV1.5 (+ 14,5 fois), NaV1.7 (+ 8,9 fois) et NaV2.1 (+ 474,4 fois) ainsi que pour les sous-unités
NaVb2 (+ 12 fois) et NaVb3 (+ 24,7 fois).
Cx4
3
Cx4
0
Cx4
5
Cx3
0.2-2
-1
0
1
2V
aria
tion
d'ex
pres
sion
(lo
g2)
VS
Non
Diff
Figure R-23 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des connexines déterminé par RT-qPCR. (n = 4). Le graphe illustre les variations d'expression de gènes
(en log 2) des cellules différenciées versus les cellules non différenciées. Entre 1 et - , l’e p essio des gènes ne varie pas (ceci correspond à des variations de + 2 fois et - 2 fois par rapport aux cellules
non différenciées). Chaque barre représente la valeur moyenne ± S.E.M (erreur standard de la
moyenne). n : représente le nombre de patients à partir desquels les CSCs W8B2+ ont été
différenciées.
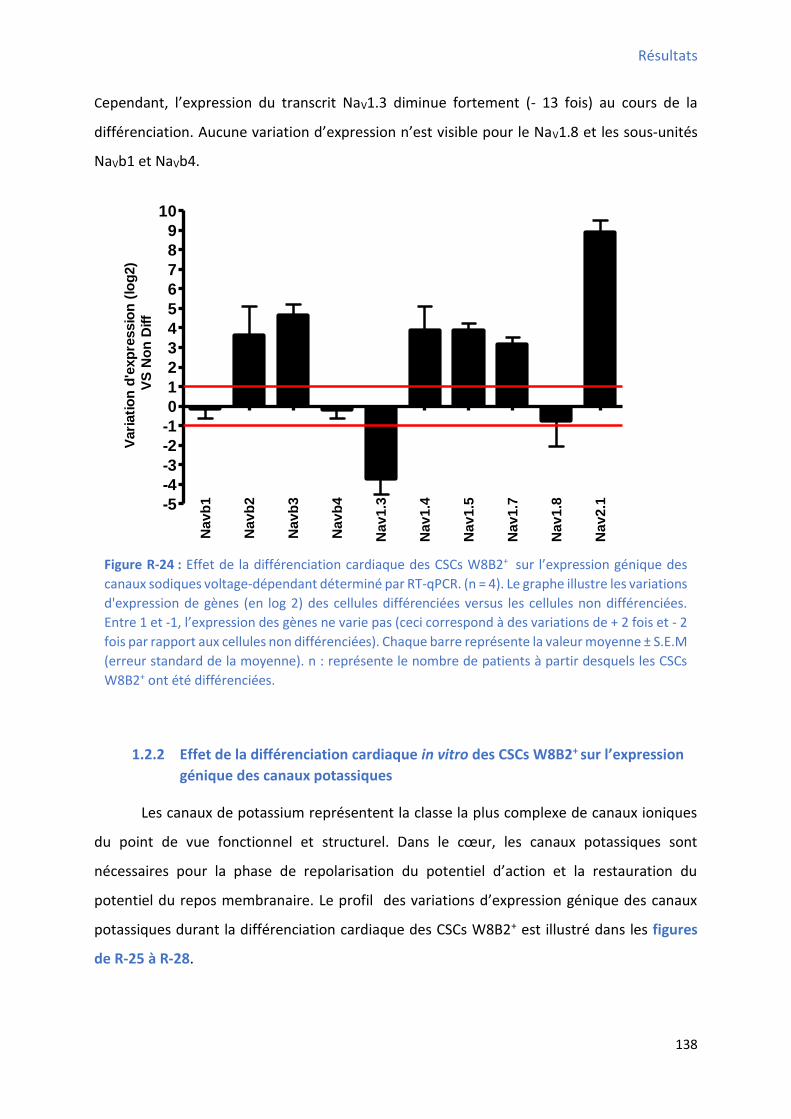
Résultats
138
Cepe da t, l’e p essio du t a s it NaV1.3 diminue fortement (- 13 fois) au cours de la
diff e iatio . Au u e a iatio d’e p essio ’est isi le pou le NaV1.8 et les sous-unités
NaVb1 et NaVb4.
1.2.2 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des canaux potassiques
Les canaux de potassium représentent la classe la plus complexe de canaux ioniques
du point de vue fonction el et st u tu el. Da s le œu , les a au potassi ues so t
essai es pou la phase de epola isatio du pote tiel d’a tio et la estau atio du
pote tiel du epos e a ai e. Le p ofil des a iatio s d’e p essio g i ue des a au
potassiques durant la différenciation cardiaque des CSCs W8B2+ est illustré dans les figures
de R-25 à R-28.
Figure R-24 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux sodiques voltage-dépendant déterminé par RT-qPCR. (n = 4). Le graphe illustre les variations
d'expression de gènes (en log 2) des cellules différenciées versus les cellules non différenciées.
Entre 1 et -1, l’e p essio des g es e a ie pas e i o espo d à des a iatio s de + fois et - 2
fois par rapport aux cellules non différenciées). Chaque barre représente la valeur moyenne ± S.E.M
(erreur standard de la moyenne). n : représente le nombre de patients à partir desquels les CSCs
W8B2+ ont été différenciées.
Nav
b1
Nav
b2
Nav
b3
Nav
b4
Nav
1.3
Nav
1.4
Nav
1.5
Nav
1.7
Nav
1.8
Nav
2.1-5
-4-3-2-10123456789
10
Var
iatio
n d'
expr
essi
on (
log2
) V
S N
on D
iff
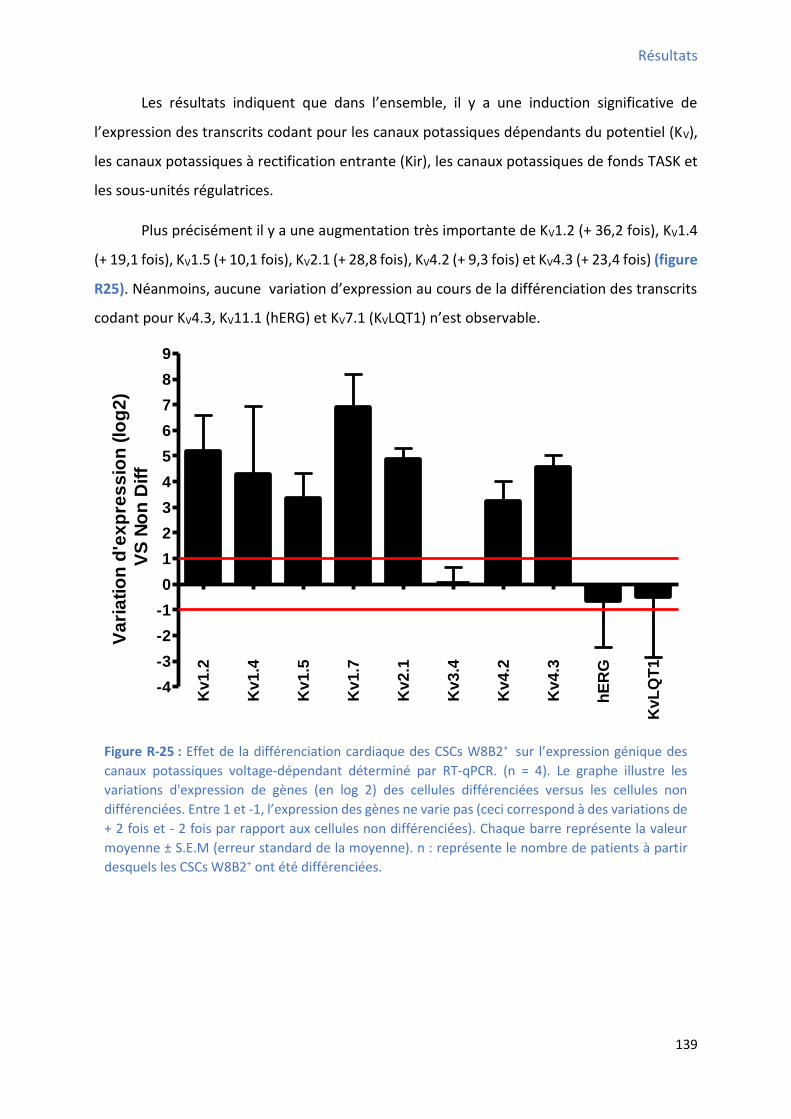
Résultats
139
Les résultats indiquent ue da s l’e se le, il a u e i du tio sig ifi ati e de
l’e p essio des t a s its oda t pou les a au potassi ues d pe da ts du pote tiel KV),
les canaux potassiques à rectification entrante (Kir), les canaux potassiques de fonds TASK et
les sous-unités régulatrices.
Plus précisément il y a une augmentation très importante de KV1.2 (+ 36,2 fois), KV1.4
(+ 19,1 fois), KV1.5 (+ 10,1 fois), KV2.1 (+ 28,8 fois), KV4.2 (+ 9,3 fois) et KV4.3 (+ 23,4 fois) (figure
R25). N a oi s, au u e a iatio d’e p essio au ou s de la diff e iatio des t a s its
codant pour KV4.3, KV11.1 (hERG) et KV7.1 (KVLQT ’est o se a le.
Figure R-25 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux potassiques voltage-dépendant déterminé par RT-qPCR. (n = 4). Le graphe illustre les
variations d'expression de gènes (en log 2) des cellules différenciées versus les cellules non
différenciées. Entre 1 et - , l’e p ession des gènes ne varie pas (ceci correspond à des variations de
+ 2 fois et - 2 fois par rapport aux cellules non différenciées). Chaque barre représente la valeur
moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir
desquels les CSCs W8B2+ ont été différenciées.
Kv1
.2
Kv1
.4
Kv1
.5
Kv1
.7
Kv2
.1
Kv3
.4
Kv4
.2
Kv4
.3
hER
G
KvL
QT
1
-4
-3
-2
-1
0
1
2
3
4
5
6
7
8
9
Var
iatio
n d'
expr
essi
on (l
og2)
VS
Non
Diff
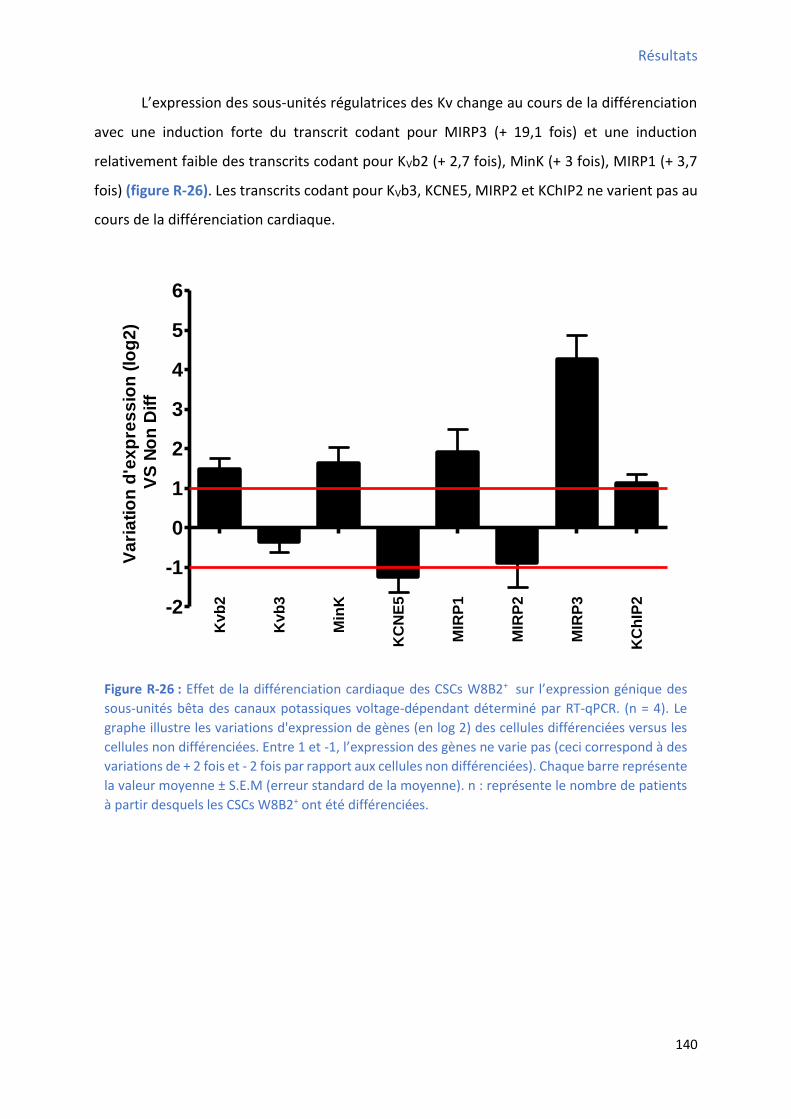
Résultats
140
L’e p essio des sous-unités régulatrices des Kv change au cours de la différenciation
avec une induction forte du transcrit codant pour MIRP3 (+ 19,1 fois) et une induction
relativement faible des transcrits codant pour KVb2 (+ 2,7 fois), MinK (+ 3 fois), MIRP1 (+ 3,7
fois) (figure R-26). Les transcrits codant pour KVb3, KCNE5, MIRP2 et KChIP2 ne varient pas au
cours de la différenciation cardiaque.
Figure R-26 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des sous-unités bêta des canaux potassiques voltage-dépendant déterminé par RT-qPCR. (n = 4). Le
graphe illustre les variations d'expression de gènes (en log 2) des cellules différenciées versus les
cellules non différenciées. Entre 1 et - , l’e p essio des g es e a ie pas e i o espo d à des variations de + 2 fois et - 2 fois par rapport aux cellules non différenciées). Chaque barre représente
la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients
à partir desquels les CSCs W8B2+ ont été différenciées.
Kvb
2
Kvb
3
Min
K
KC
NE
5
MIR
P1
MIR
P2
MIR
P3
KC
hIP
2-2
-1
0
1
2
3
4
5
6
Var
iatio
n d'
expr
essi
on (l
og2)
VS
Non
Diff
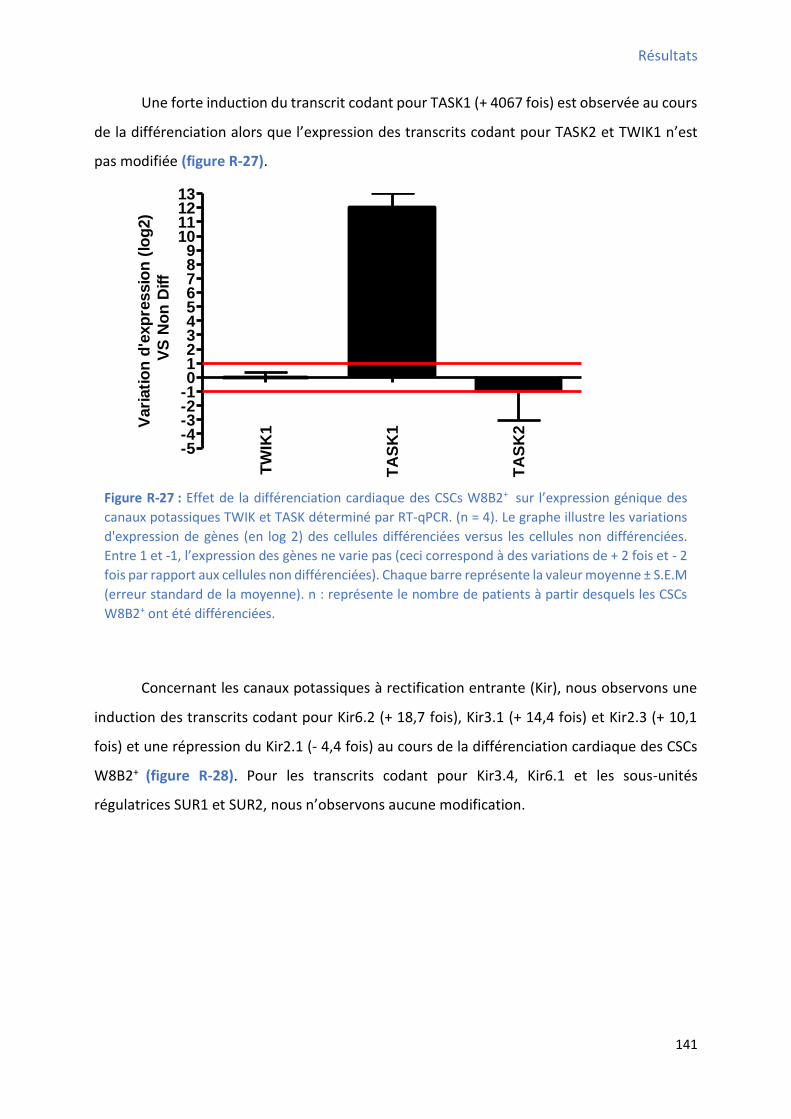
Résultats
141
Une forte induction du transcrit codant pour TASK1 (+ 4067 fois) est observée au cours
de la diff e iatio alo s ue l’e p essio des t a s its oda t pou TA“K et TWIK ’est
pas modifiée (figure R-27).
Concernant les canaux potassiques à rectification entrante (Kir), nous observons une
induction des transcrits codant pour Kir6.2 (+ 18,7 fois), Kir3.1 (+ 14,4 fois) et Kir2.3 (+ 10,1
fois) et une répression du Kir2.1 (- 4,4 fois) au cours de la différenciation cardiaque des CSCs
W8B2+ (figure R-28). Pour les transcrits codant pour Kir3.4, Kir6.1 et les sous-unités
gulat i es “UR et “UR , ous ’o se o s au u e odifi atio .
TW
IK1
TA
SK
1
TA
SK
2
-5-4-3-2-10123456789
10111213
Var
iatio
n d'
expr
essi
on (
log2
) V
S N
on D
iff
Figure R-27 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux potassiques TWIK et TASK déterminé par RT-qPCR. (n = 4). Le graphe illustre les variations
d'expression de gènes (en log 2) des cellules différenciées versus les cellules non différenciées.
Entre 1 et - , l’e p essio des g es e a ie pas e i o espo d à des a iatio s de + fois et - 2
fois par rapport aux cellules non différenciées). Chaque barre représente la valeur moyenne ± S.E.M
(erreur standard de la moyenne). n : représente le nombre de patients à partir desquels les CSCs
W8B2+ ont été différenciées.
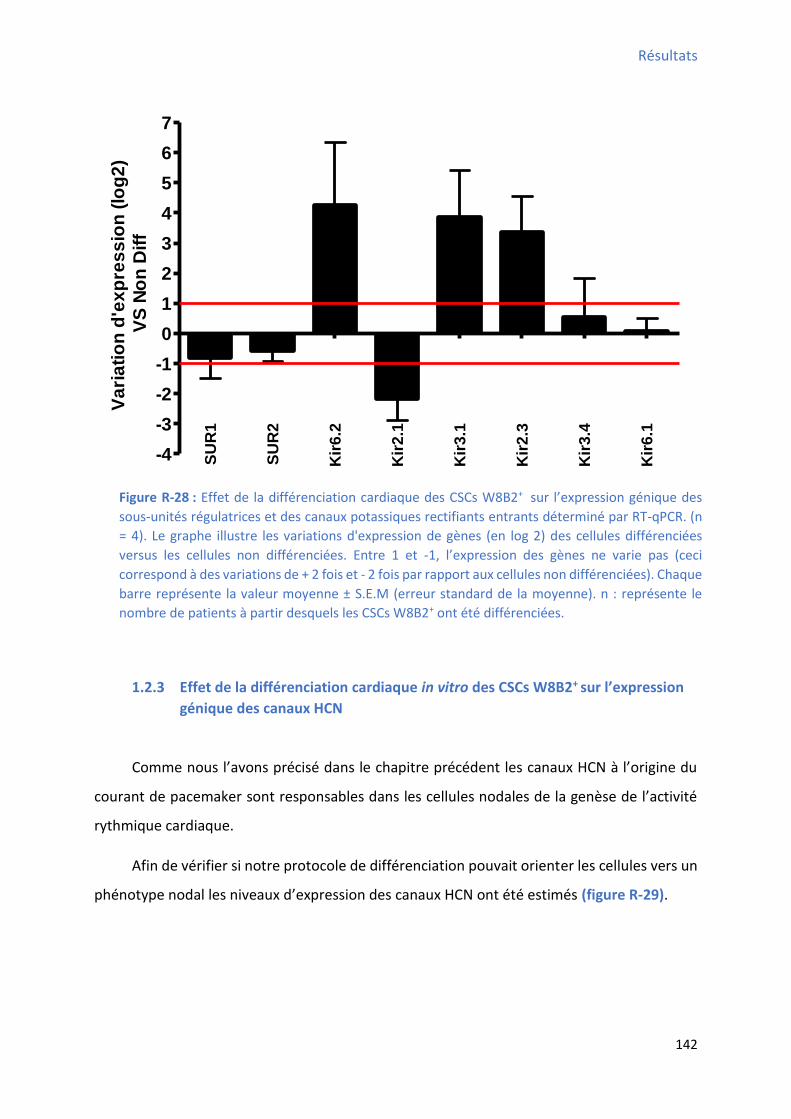
Résultats
142
1.2.3 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des canaux HCN
Comme nous l’a o s p is da s le hapit e p de t les a au HCN à l’o igi e du
ou a t de pa e ake so t espo sa les da s les ellules odales de la ge se de l’a ti it
rythmique cardiaque.
Afin de vérifier si notre protocole de différenciation pouvait orienter les cellules vers un
ph ot pe odal les i eau d’e p essio des a au HCN o t t esti s (figure R-29).
SU
R1
SU
R2
Kir6
.2
Kir2
.1
Kir3
.1
Kir2
.3
Kir3
.4
Kir6
.1
-4
-3
-2
-1
0
1
2
3
4
5
6
7V
aria
tion
d'ex
pres
sion
(log
2) V
S N
on D
iff
Figure R-28 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des sous-unités régulatrices et des canaux potassiques rectifiants entrants déterminé par RT-qPCR. (n
= 4). Le graphe illustre les variations d'expression de gènes (en log 2) des cellules différenciées
versus les cellules non différenciées. Entre 1 et - , l’e p essio des g es e a ie pas e i correspond à des variations de + 2 fois et - 2 fois par rapport aux cellules non différenciées). Chaque
barre représente la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le
nombre de patients à partir desquels les CSCs W8B2+ ont été différenciées.
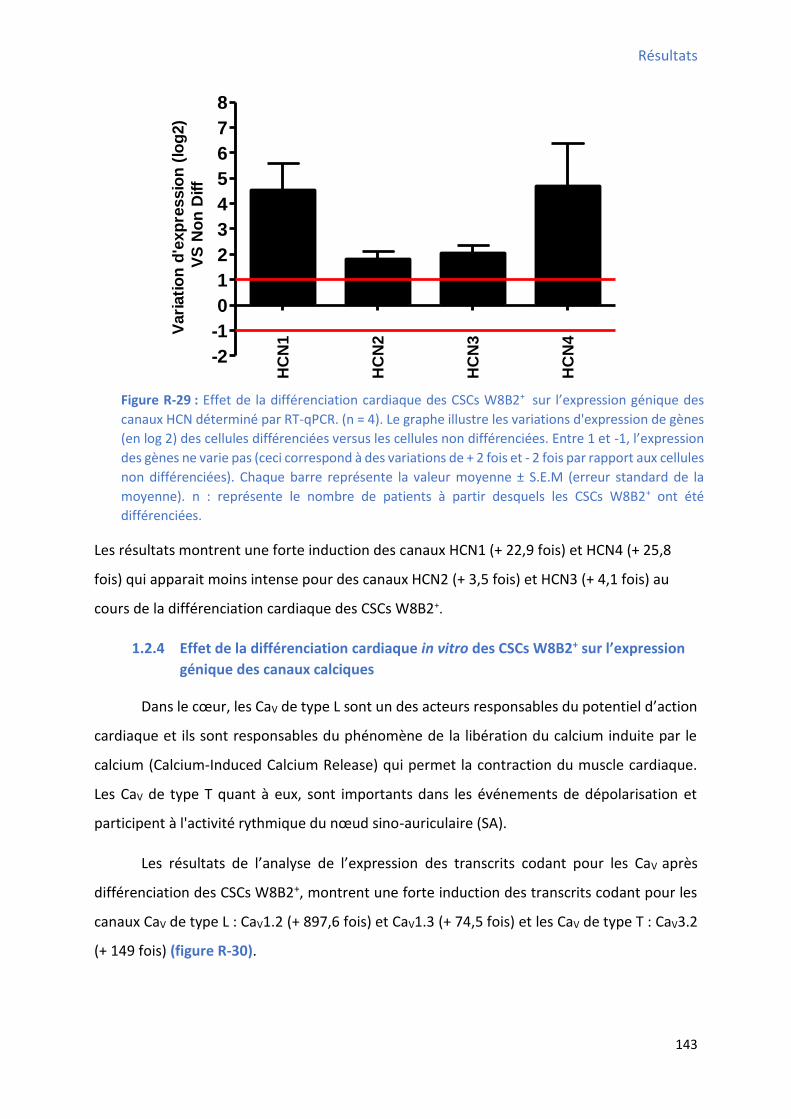
Résultats
143
Les résultats montrent une forte induction des canaux HCN1 (+ 22,9 fois) et HCN4 (+ 25,8
fois) qui apparait moins intense pour des canaux HCN2 (+ 3,5 fois) et HCN3 (+ 4,1 fois) au
cours de la différenciation cardiaque des CSCs W8B2+.
1.2.4 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des canaux calciques
Da s le œu , les CaV de t pe L so t u des a teu s espo sa les du pote tiel d’a tio
cardiaque et ils sont responsables du phénomène de la libération du calcium induite par le
calcium (Calcium-Induced Calcium Release) qui permet la contraction du muscle cardiaque.
Les CaV de type T quant à eux, sont importants dans les événements de dépolarisation et
pa ti ipe t à l'a ti it th i ue du œud si o-auriculaire (SA).
Les sultats de l’a al se de l’e p essio des transcrits codant pour les CaV après
différenciation des CSCs W8B2+, montrent une forte induction des transcrits codant pour les
canaux CaV de type L : CaV1.2 (+ 897,6 fois) et CaV1.3 (+ 74,5 fois) et les CaV de type T : CaV3.2
(+ 149 fois) (figure R-30).
HC
N1
HC
N2
HC
N3
HC
N4
-2-1012345678
Var
iatio
n d'
expr
essi
on (
log2
) V
S N
on D
iff
Figure R-29 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux HCN déterminé par RT-qPCR. (n = 4). Le graphe illustre les variations d'expression de gènes
(en log 2) des cellules différenciées versus les cellules non différenciées. Entre 1 et - , l’e p essio des gènes ne varie pas (ceci correspond à des variations de + 2 fois et - 2 fois par rapport aux cellules
non différenciées). Chaque barre représente la valeur moyenne ± S.E.M (erreur standard de la
moyenne). n : représente le nombre de patients à partir desquels les CSCs W8B2+ ont été
différenciées.
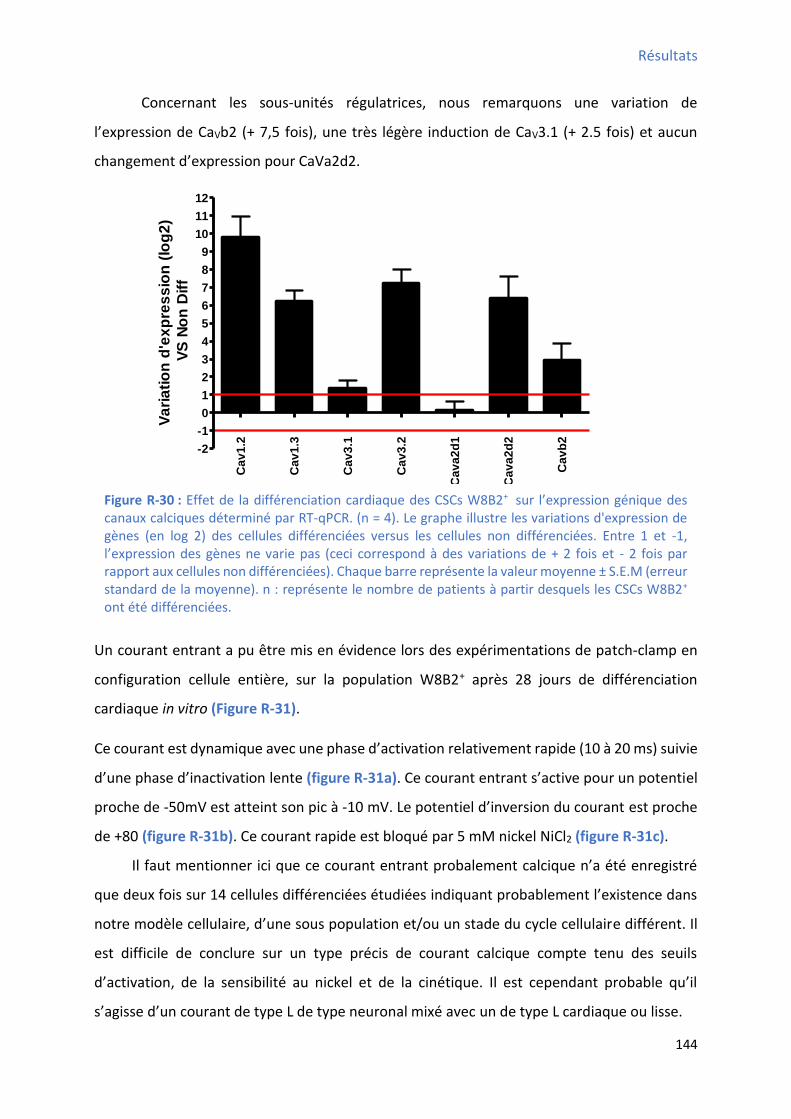
Résultats
144
Concernant les sous-unités régulatrices, nous remarquons une variation de
l’e p essio de CaVb2 (+ 7,5 fois), une très légère induction de CaV3.1 (+ 2.5 fois) et aucun
ha ge e t d’e p essio pou CaVa d .
Un courant entrant a pu être mis en évidence lors des expérimentations de patch-clamp en
configuration cellule entière, sur la population W8B2+ après 28 jours de différenciation
cardiaque in vitro (Figure R-31).
Ce courant est d a i ue a e u e phase d’a ti atio relativement rapide (10 à 20 ms) suivie
d’u e phase d’i a ti atio lente (figure R-31a). Ce ou a t e t a t s’a ti e pou u pote tiel
proche de -50mV est atteint son pic à - V. Le pote tiel d’i e sio du ou a t est proche
de +80 (figure R-31b). Ce courant rapide est bloqué par 5 mM nickel NiCl2 (figure R-31c).
Il faut e tio e i i ue e ou a t e t a t p o ale e t al i ue ’a t e egist
ue deu fois su ellules diff e i es tudi es i di ua t p o a le e t l’e iste e da s
ot e od le ellulai e, d’u e sous populatio et/ou u stade du le ellulai e différent. Il
est difficile de conclure sur un type précis de courant calcique compte tenu des seuils
d’a ti atio , de la se si ilit au i kel et de la i ti ue. Il est epe da t p o a le u’il
s’agisse d’u ou a t de t pe L de t pe eu o al i a e un de type L cardiaque ou lisse.
Cav
1.2
Cav
1.3
Cav
3.1
Cav
3.2
Cav
a2d1
Cav
a2d2
Cav
b2-2
-1
0
1
2
3
4
5
6
7
8
9
10
11
12
Var
iatio
n d'
expr
essi
on (
log2
) V
S N
on D
iff
Figure R-30 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des canaux calciques déterminé par RT-qPCR. (n = 4). Le graphe illustre les variations d'expression de gènes (en log 2) des cellules différenciées versus les cellules non différenciées. Entre 1 et -1, l’e p essio des g es e a ie pas e i o espo d à des a iatio s de + fois et - 2 fois par rapport aux cellules non différenciées). Chaque barre représente la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir desquels les CSCs W8B2+ ont été différenciées.
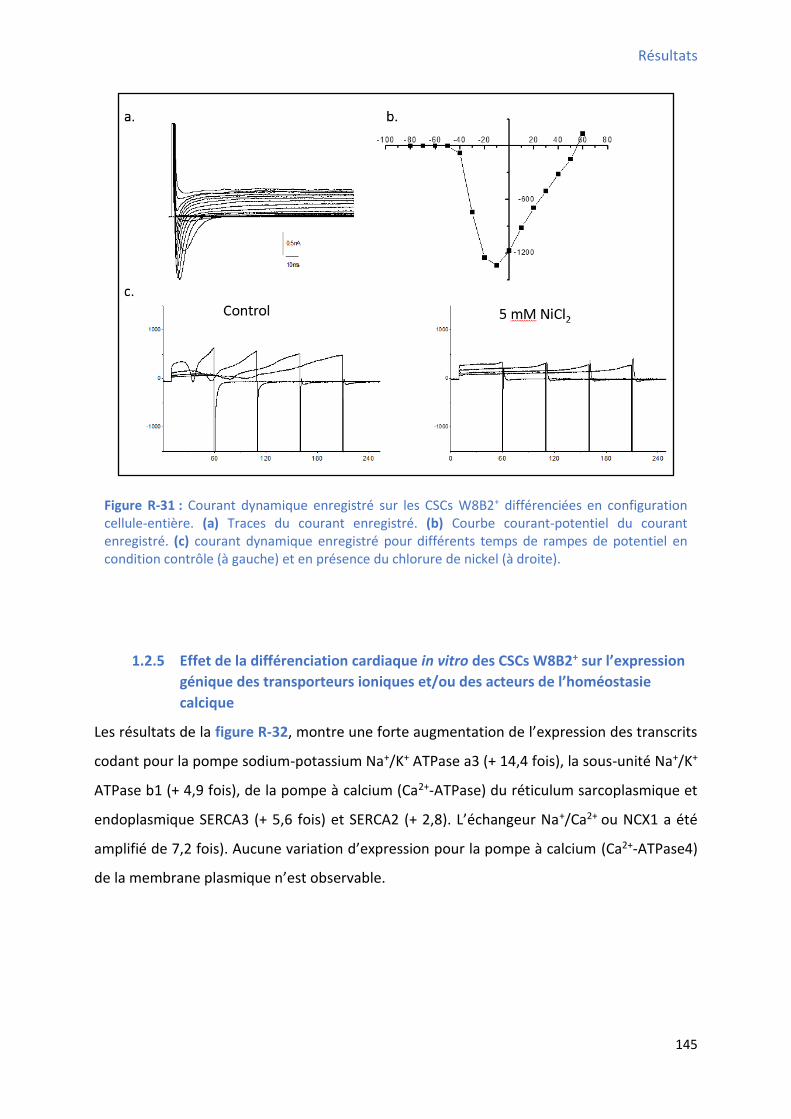
Résultats
145
1.2.5 Effet de la différenciation cardiaque in vitro des CSCs W8B2+ su l’e p essio génique des t a spo teu s io i ues et/ou des a teu s de l’ho ostasie calcique
Les résultats de la figure R-32, o t e u e fo te aug e tatio de l’e p essio des t a s its
codant pour la pompe sodium-potassium Na+/K+ ATPase a3 (+ 14,4 fois), la sous-unité Na+/K+
ATPase b1 (+ 4,9 fois), de la pompe à calcium (Ca2+-ATPase) du réticulum sarcoplasmique et
e doplas i ue “ERCA + , fois et “ERCA + , . L’ ha geu Na+/Ca2+ ou NCX1 a été
a plifi de , fois . Au u e a iatio d’e p essio pou la po pe à al iu (Ca2+-ATPase4)
de la e a e plas i ue ’est o se a le.
Figure R-31 : Courant dynamique enregistré sur les CSCs W8B2+ différenciées en configuration cellule-entière. (a) Traces du courant enregistré. (b) Courbe courant-potentiel du courant enregistré. (c) courant dynamique enregistré pour différents temps de rampes de potentiel en condition contrôle (à gauche) et en présence du chlorure de nickel (à droite).
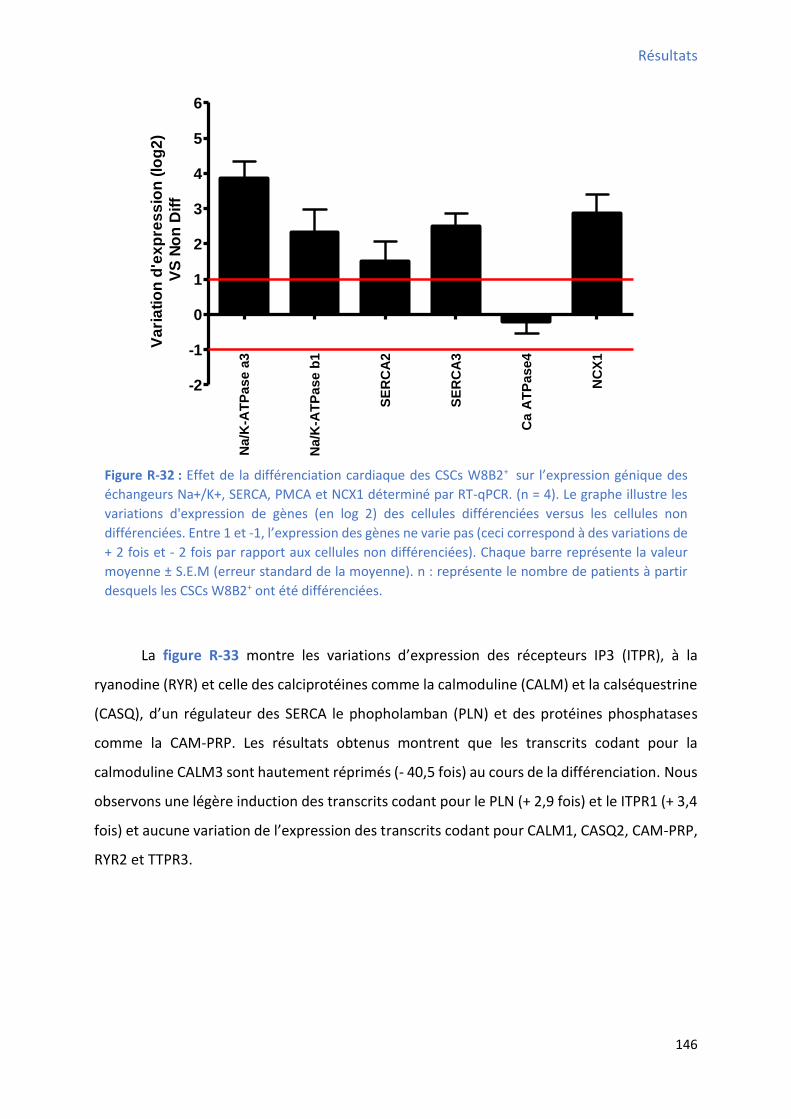
Résultats
146
La figure R-33 o t e les a iatio s d’e p essio des epteu s IP (ITPR), à la
ryanodine (RYR) et celle des calciprotéines comme la calmoduline (CALM) et la calséquestrine
CA“Q , d’u gulateu des “ERCA le phophola a PLN et des p ot i es phosphatases
comme la CAM-PRP. Les résultats obtenus montrent que les transcrits codant pour la
calmoduline CALM3 sont hautement réprimés (- 40,5 fois) au cours de la différenciation. Nous
observons une légère induction des transcrits codant pour le PLN (+ 2,9 fois) et le ITPR1 (+ 3,4
fois et au u e a iatio de l’e p essio des transcrits codant pour CALM1, CASQ2, CAM-PRP,
RYR2 et TTPR3.
Figure R-32 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des échangeurs Na+/K+, SERCA, PMCA et NCX1 déterminé par RT-qPCR. (n = 4). Le graphe illustre les
variations d'expression de gènes (en log 2) des cellules différenciées versus les cellules non
différenciées. Entre 1 et - , l’e p essio des g es e a ie pas e i o espo d à des a iatio s de + 2 fois et - 2 fois par rapport aux cellules non différenciées). Chaque barre représente la valeur
moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir
desquels les CSCs W8B2+ ont été différenciées.
Na/
K-A
TP
ase
a3
Na/
K-A
TP
ase
b1
SE
RC
A2
SE
RC
A3
Ca
AT
Pas
e4
NC
X1
-2
-1
0
1
2
3
4
5
6
Var
iatio
n d'
expr
essi
on (l
og2)
VS
Non
Diff
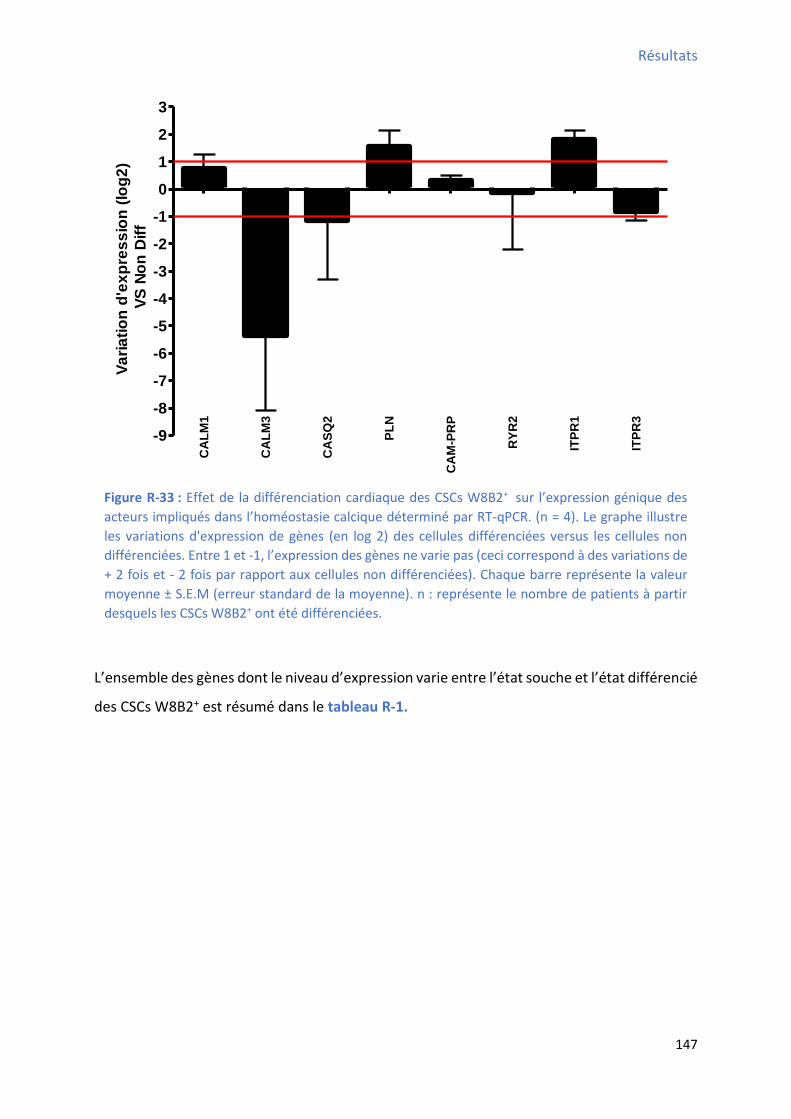
Résultats
147
L’e se le des g es do t le i eau d’e p ession varie entre l’ tat sou he et l’ tat diff e i
des CSCs W8B2+ est résumé dans le tableau R-1.
Figure R-33 : Effet de la différenciation cardiaque des CSCs W8B2+ su l’e p essio g i ue des a teu s i pli u s da s l’ho ostasie al i ue d te i pa RT-qPCR. (n = 4). Le graphe illustre
les variations d'expression de gènes (en log 2) des cellules différenciées versus les cellules non
différenciées. Entre 1 et - , l’e p essio des g es e a ie pas e i o espo d à des a iatio s de + 2 fois et - 2 fois par rapport aux cellules non différenciées). Chaque barre représente la valeur
moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir
desquels les CSCs W8B2+ ont été différenciées.
CA
LM1
CA
LM3
CA
SQ
2
PLN
CA
M-P
RP
RY
R2
ITP
R1
ITP
R3
-9
-8
-7
-6
-5
-4
-3
-2
-1
0
1
2
3V
aria
tion
d'ex
pres
sion
(lo
g2)
VS
Non
Diff
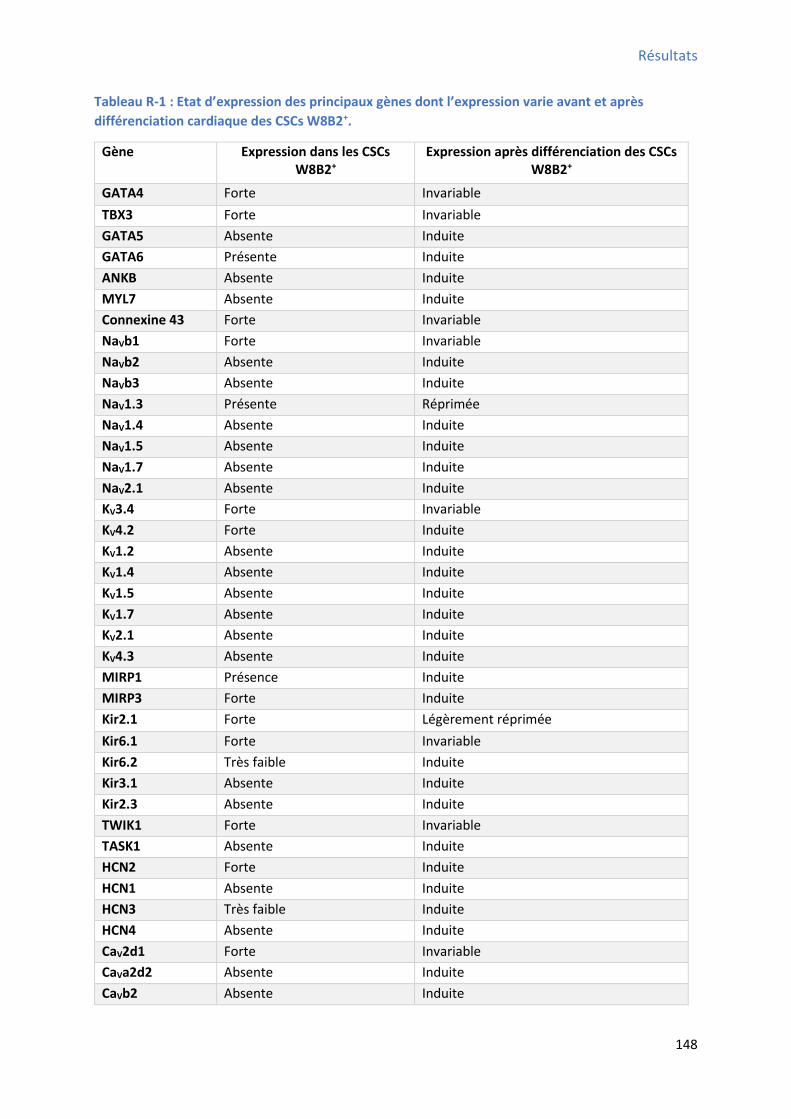
Résultats
148
Tableau R-1 : Etat d’e p essio des p i ipau g es do t l’e p essio va ie ava t et ap s différenciation cardiaque des CSCs W8B2+.
Gène Expression dans les CSCs
W8B2+
Expression après différenciation des CSCs
W8B2+
GATA4 Forte Invariable
TBX3 Forte Invariable
GATA5 Absente Induite
GATA6 Présente Induite
ANKB Absente Induite
MYL7 Absente Induite
Connexine 43 Forte Invariable
NaVb1 Forte Invariable
NaVb2 Absente Induite
NaVb3 Absente Induite
NaV1.3 Présente Réprimée
NaV1.4 Absente Induite
NaV1.5 Absente Induite
NaV1.7 Absente Induite
NaV2.1 Absente Induite
KV3.4 Forte Invariable
KV4.2 Forte Induite
KV1.2 Absente Induite
KV1.4 Absente Induite
KV1.5 Absente Induite
KV1.7 Absente Induite
KV2.1 Absente Induite
KV4.3 Absente Induite
MIRP1 Présence Induite
MIRP3 Forte Induite
Kir2.1 Forte Légèrement réprimée
Kir6.1 Forte Invariable
Kir6.2 Très faible Induite
Kir3.1 Absente Induite
Kir2.3 Absente Induite
TWIK1 Forte Invariable
TASK1 Absente Induite
HCN2 Forte Induite
HCN1 Absente Induite
HCN3 Très faible Induite
HCN4 Absente Induite
CaV2d1 Forte Invariable
CaVa2d2 Absente Induite
CaVb2 Absente Induite
Résultats
149
CaV1.2 Absente Induite
CaV1.3 Absente Induite
CaV3.2 Absente Induite
Pompe Na+/K+ b1 Forte Induite
Pompe Na+/K+ a3 Absente Induite
SERCA2 Forte Invariable
SERCA3 Forte Induite
Pompe calcique Forte Invariable
NXC1 Forte Induite
CALM1 Forte Invariable
CALM3 Forte Réprimée
Calcineurine
(CAM-PRP)
Forte Invariable
ITPR1 Forte Induite
ITPR3 Forte Invariable
BNP Forte Induite
KCa1.1 Présente Non étudiée
KCa3.1 présente Non étudiée
Da s le ut d’a lio e la diff e iatio e a dio o tes des C“Cs W B +, nous
avons opté pour une approche optogénétique par expression du canal rhodopsine 2 (CHR2)
dépolarisant capable de transporter du sodium et des protons activé par flash lumineux dans
le bleu.
Cette dépolarisation pourrait être un puissant déclencheur de la différenciation
cardiaque tel u’il a d jà t o t da s d’aut e t pe de ellules sou hes (Stroh et al., 2011).
Nos résultats préliminaires ont montré que cette stimulation par optogénétique au
cours de la différenciation des CSCs W8B2+, ’a lio e pa la diff e iatio a dia ue ie
au contraire comme le montre la figure R-34. En effet, le profil génique des CSCs W8B2+
différenciées avec stimulation optogénétique est très similaire à celui des CSCs W8B2+ non
différencié.
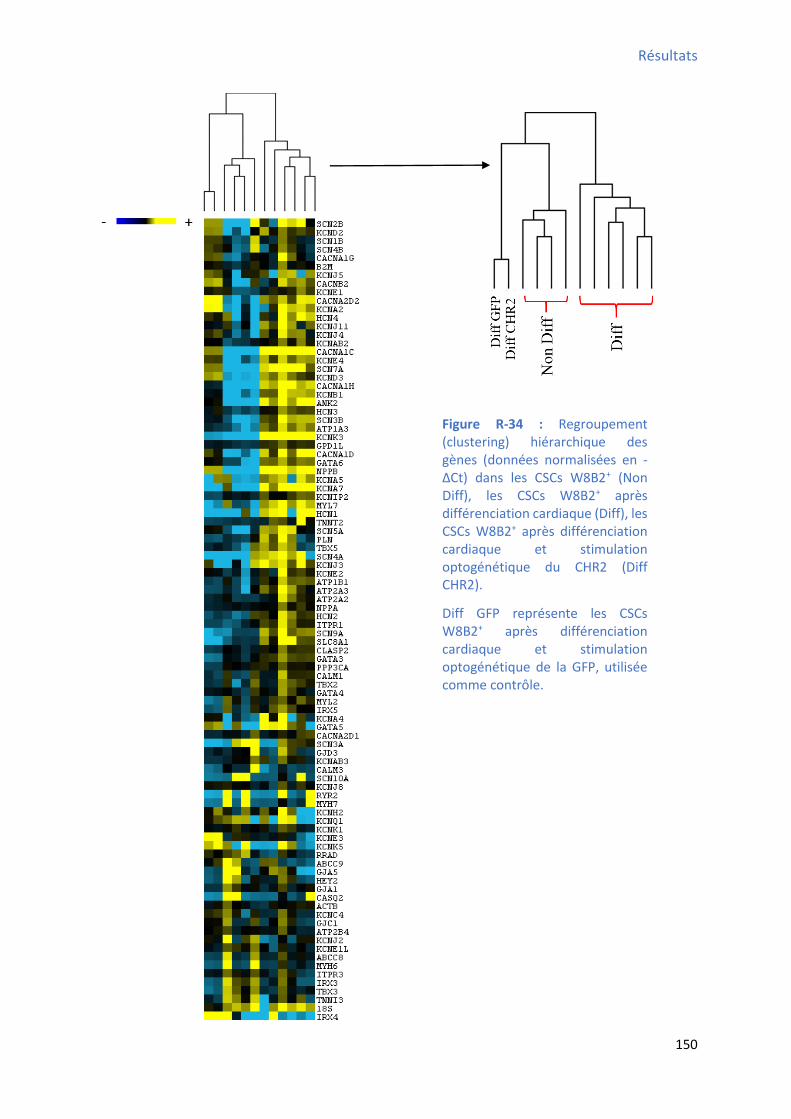
Résultats
150
Figure R-34 : Regroupement (clustering) hiérarchique des gènes (données normalisées en -ΔCt da s les C“Cs W B + (Non Diff), les CSCs W8B2+ après différenciation cardiaque (Diff), les CSCs W8B2+ après différenciation cardiaque et stimulation optogénétique du CHR2 (Diff CHR2).
Diff GFP représente les CSCs W8B2+ après différenciation cardiaque et stimulation optogénétique de la GFP, utilisée comme contrôle.

Résultats
151
2 Etude de l’a tivit al i ue au ou s de la diff e iatio a dia ue in vitro des CSCs
W8B2+
La bibliographie actuelle montre que très peu de travaux sont publiés sur le décryptage
de la sig atu e de l’a ti it et la sig alisatio al i ue da s les ellules sou hes a dia ues.
Seule une étude réalisée sur les CSCs c-kit+ a montré que ces progéniteurs cardiaques
pouvaient présenter des oscillations calciques spontanées qui semblaient jouer un rôle
essentiel dans la régulation de la croissance cellulaire (Ferreira-Martins et al., 2009).
Cepe da t, au u e a a t isatio de l’a ti it al i ue au ou s de la diff e iatio des
C“Cs ’a t appo t e. Not e o je tif est do de sui e l’a ti it al i ue des C“Cs W B +
au cours de leur différenciation cardiaque in vitro et d’appo te des léments de réponse qui
pou aie t ous aide à ieu o p e d e l’i pli atio du al iu da s le p o essus de
différenciation.
2.1 Sig atu e de l’a tivit spo ta e al i ue au ou s de la diff e iatio a dia ue in vitro des CSCs W8B2+
Pour étudier et suivre l’ olutio de l’a ti it al i ue spo ta e au ou s de la
différenciation in vitro des CSCs W8B2+, nous avons utilisé la sonde calcique GCaMP6s comme
senseur des changements de concentrations calciques intracellulaires. Via cette sonde, nous
pouvons sui e l’ olutio de l’a ti it al i ue tout au lo g du p oto ole de diff e iatio
cardiaque in vitro (4 semaines) des CSCs W8B2+.
L’e egist e e t de l’a ti it al i ue a t alis pa i os opie o fo ale à °C. Les
variations de fluorescence de cette dernière reflètent les variations de calcium intracellulaires.
Les enregistrements calciques bruts obtenus par microscopie confocale sont ensuite analysés
à l’aide d’u e a o appli u e sous I age J ui pe et de ua tifie les a iatio s de
fluorescence globales dans chaque cellule étudiée. Ces variations de fluorescence peuvent
être présentées sous forme de courbes en fonction du temps.
Les sultats o te us o t e t ue l’a ti it al i ue e egist e su les C“Cs W B + en
début de différenciation (2ème jour de différenciation) et à la fin du protocole de
différenciation (28ème jou se p se te sous fo e d’os illatio s al i ues (figure R-35).
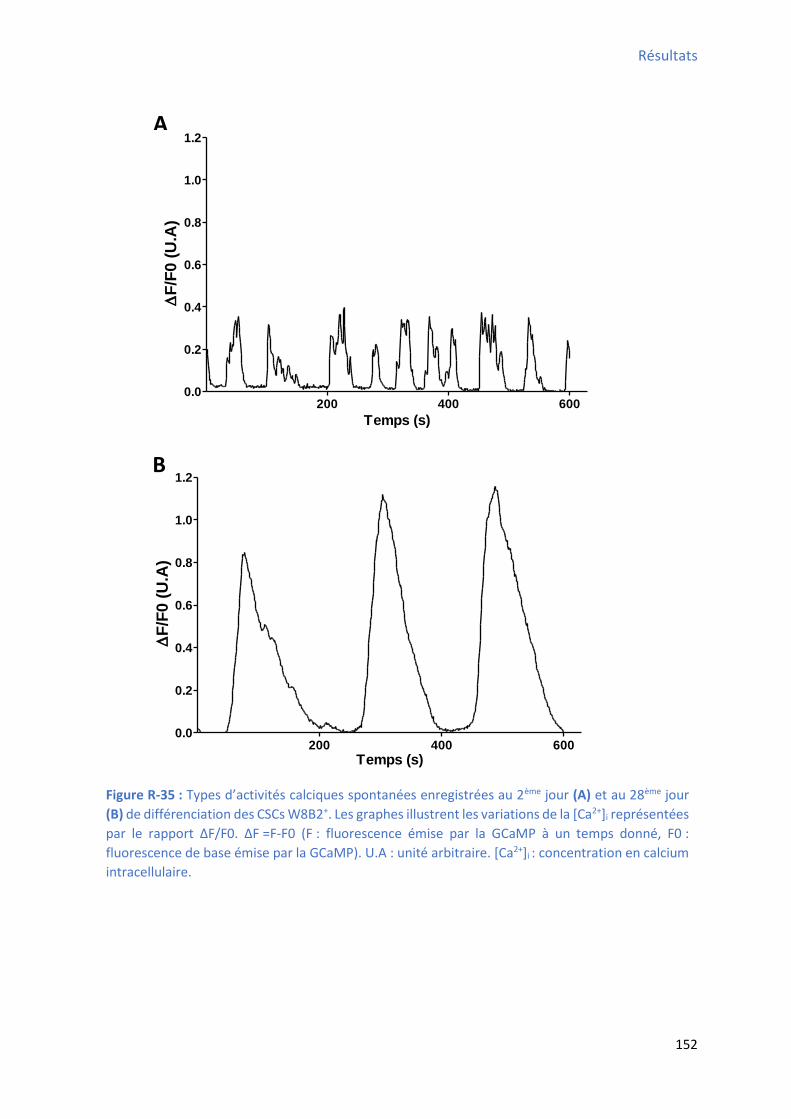
Résultats
152
200 400 6000.0
0.2
0.4
0.6
0.8
1.0
1.2
Temps (s)
F/F0
(U.A
)
200 400 6000.0
0.2
0.4
0.6
0.8
1.0
1.2
Temps (s)
F/F0
(U.A
)
Figure R-35 : T pes d’a ti it s al i ues spo ta es e egist es au ème jour (A) et au 28ème jour
(B) de différenciation des CSCs W8B2+. Les graphes illustrent les variations de la [Ca2+]i représentées
pa le appo t ΔF/F . ΔF =F-F0 (F : fluorescence émise par la GCaMP à un temps donné, F0 :
fluorescence de base émise par la GCaMP). U.A : unité arbitraire. [Ca2+]i : concentration en calcium
intracellulaire.
A
B

Résultats
153
Les oscillations obtenues au 2ème jour de différenciation cardiaque sont irrégulières, plus
fréquentes, et présentent une amplitude nettement plus faible que les oscillations obtenues au 28ème
jour de différenciation.
2.2 Evolution des paramètres calciques au cours de la différenciation cardiaque in
vitro des CSCs W8B2+
Afi d’ tudie e tai s pa a t es de l’a ti it al i ue e egist e au ou s de la
différenciation cardiaque in vitro des CSCs W8B2+, dans chaque cellule, les variations de
globales fluorescence obtenues sous Image J ont été analysées grâce à une application
développée au laboratoire sous le langage IDL (Interactive Data Language). Cette application
permet de mesurer différents paramètres cinétiques des oscillations calciques obtenues
comme ; l’a plitude, la f ue e, la du e, l’ai e sous la ou e, le TTP ti e to peak , le TTR
ti e to e o e ou e o e le pou e tage du te ps d’a ti it oi at iels et thodes .
L’ olutio de es pa a t es est ep se t e pa se ai e pe da t uat e se ai es de
différenciation :
1ère semaine de différenciation : regroupant toutes les oscillations calciques obtenues entre
le 2ème et le 7ème jour post-différenciation.
2ème semaine de différenciation : regroupant toutes les oscillations calciques obtenues entre
le 8ème et le 14ème jour pos-différenciation.
3ème semaine de différenciation : regroupant toutes les oscillations calciques obtenues entre
le 15ème et le 21ème jour post-différenciation.
4ème semaine de différenciation : regroupant toutes les oscillations calciques obtenues entre
le 22ème et le 28ème jour de différenciation.
2.2.1 Evolution de la fréquence des oscillations calciques au cours de la
différenciation cardiaque in vitro des CSCs W8B2+
Les résultats représentés dans la figure R-36A, montrent que la fréquence des
oscillations calcique diminue progressivement au cours des 4 semaines de différenciation.
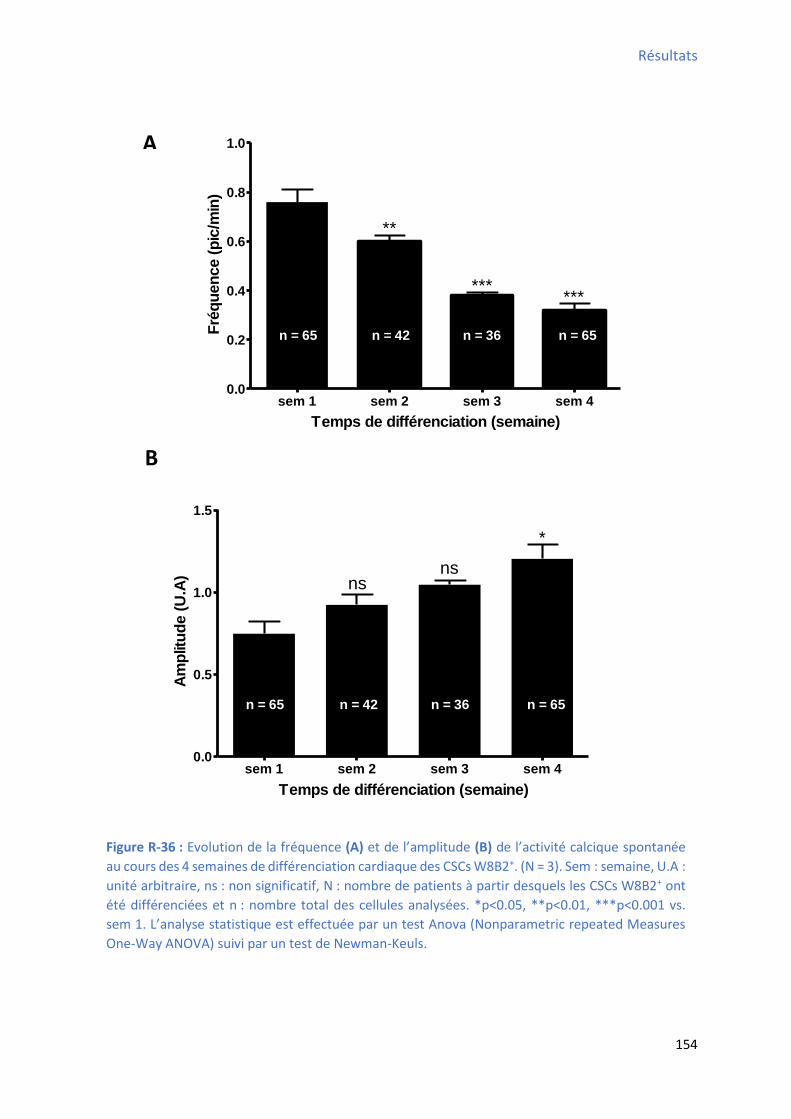
Résultats
154
Figure R-36 : Evolution de la fréquence (A) et de l’a plitude (B) de l’a ti ité calcique spontanée
au cours des 4 semaines de différenciation cardiaque des CSCs W8B2+. (N = 3). Sem : semaine, U.A :
unité arbitraire, ns : non significatif, N : nombre de patients à partir desquels les CSCs W8B2+ ont
été différenciées et n : nombre total des cellules analysées. *p<0.05, **p<0.01, ***p<0.001 vs.
se . L’a al se statisti ue est effe tu e pa u test A o a No pa a et i epeated Measu es One-Way ANOVA) suivi par un test de Newman-Keuls.
B
A
sem 1 sem 2 sem 3 sem 40.0
0.5
1.0
1.5
*
nsns
n = 65 n = 42 n = 36 n = 65
Temps de différenciation (semaine)
Am
plitu
de (U
.A)
sem 1 sem 2 sem 3 sem 40.0
0.2
0.4
0.6
0.8
1.0
**
******
n = 65 n = 42 n = 36 n = 65
Temps de différenciation (semaine)
Fréq
uenc
e (p
ic/m
in)

Résultats
155
En effet, cette fréquence baisse significativement et diminue de 0,75 pic/min en
semaine 1 à 0,31 pic/min en semaine 4 de différenciation (soit une diminution de 58,7 %).
Cette diminution de la fréquence est également significative en comparant la semaine 1 avec
la semaine 2 (une baisse de 20%) et la semaine 1 avec la semaine 3 (une baisse de 50,6%).
2.2.2 Evolutio de l’a plitude des os illatio s al i ues au ou s de la différenciation cardiaque in vitro des CSCs W8B2+
L’a plitude a i ale de fluo es e e appo te des i di atio s su la ua tit de
calcium libéré. Les résultats représentés dans la figure R-36B, o t e t ue l’a plitude des
oscillations calciques augmente progressivement à chaque semaine de différenciation jus u’à
atteindre une significativité statistique à la semaine 4. Cette amplitude passe de 0,7 U.A en
semaine 1 à 1,2 U.A en semaine 4. Ceci représente une augmentation significative de
l’a plitude de 1,4%.
2.2.3 Evolution de la durée des oscillations calciques au cours de la différenciation
cardiaque in vitro des CSCs W8B2+
La figure R-37A montre que la durée des oscillations calciques augmente
progressivement au cours des quatre semaines de différenciation. Cette durée est de 36,8 s
en semaine 1 contre 88,7 s en semaine 4 soit une augmentation significative de 141%.
Cette augmentation est aussi significative entre la semaine 1 (36,8 s) et la semaine 3 (74,2 s)
soit 101,6% d’aug e tatio .
2.2.4 Evolutio de l’ai e des os illatio s al i ues au ou s de la diff e iation
cardiaque in vitro des CSCs W8B2+
L’ai e sous la ou e appo te des i di atio s su l’i po ta e des os illatio s al i ues
puis u’elle p e d e o pte à la fois l’a plitude du ph o e et sa du e. La figure R-37B
o t e ue l’ai e sous la ou e augmente progressivement au cours des quatre semaines de
diff e iatio . L’ai e aug e te sig ifi ati e e t de , U.A.s e se ai e à , U.A.s e
semaine 4 (soit une augmentation de 210,9%).
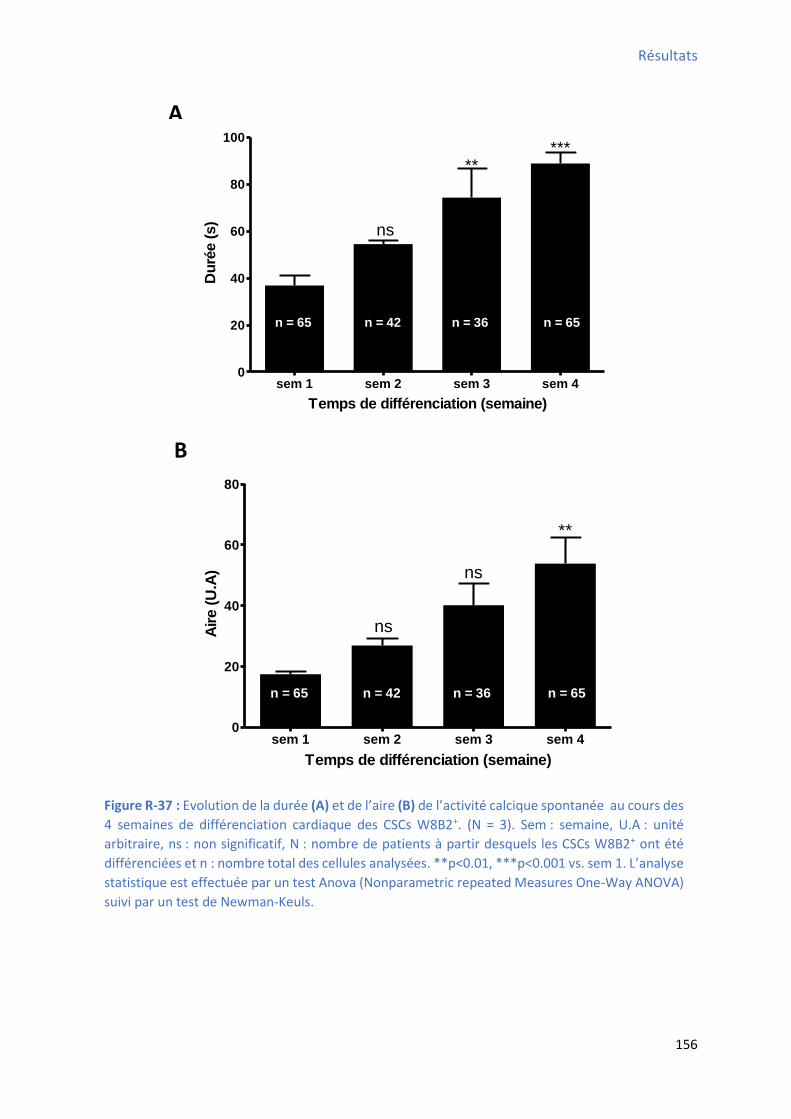
Résultats
156
Figure R-37 : Evolution de la durée (A) et de l’ai e (B) de l’a ti it al i ue spo ta e au ou s des 4 semaines de différenciation cardiaque des CSCs W8B2+. (N = 3). Sem : semaine, U.A : unité
arbitraire, ns : non significatif, N : nombre de patients à partir desquels les CSCs W8B2+ ont été
différenciées et n : nombre total des cellules analysées. **p< . , ***p< . s. se . L’a al se statistique est effectuée par un test Anova (Nonparametric repeated Measures One-Way ANOVA)
suivi par un test de Newman-Keuls.
A
B
sem 1 sem 2 sem 3 sem 40
20
40
60
80
100***
**
ns
n = 65 n = 42 n = 36 n = 65
Temps de différenciation (semaine)
Dur
ée (s
)
sem 1 sem 2 sem 3 sem 40
20
40
60
80
**
ns
ns
n = 65 n = 42 n = 36 n = 65
Temps de différenciation (semaine)
Aire
(U.A
)

Résultats
157
2.2.5 Evolution du TTP (time to peak) des oscillations calciques au cours de la
différenciation cardiaque in vitro des CSCs W8B2+
Le TTP est le temps mis pour atteindre le pic ou le maximum de fluorescence « F ». Le
TTP apporte des indications sur le temps des libérations calciques intracellulaire.
La figure R-38A montre que le TTP augmente progressivement au cours des quatre
semaines de différenciation cardiaque et il passe de 9,8 s en semaine 1 à 26 s en semaine 4
(soit une augmentation significative de 165,3%). Cette augmentation est également
significative entre la semaine 1 et la semaine 3 post-différenciation (augmentation
significative de 120,1%).
2.2.6 Evolution du TTR (time to recovery) des oscillations calciques au cours de la
différenciation cardiaque in vitro des CSCs W8B2+
Le TTR est le temps mis pour atteindre la fluorescence de base « F0 ». Le TTR apporte
des indications sur le temps des recaptures et extrusions calciques intracellulaires.
La figure R.38B montre que le TTR augmente progressivement entre la semaine 1, la
semaine 2 et la semaine 4 post-différenciation. Entre la semaine 2 et la semaine 3 post-
différenciation, le TTR reste stable.
Le TTR passe de 27 s en semaine 1 à 62,7 s en semaine 4 (soit une augmentation
significative de 132,2%). Cette augmentation est également significative entre la semaine 1 et
la semaine 3 (augmentation significative de 52,9%) et la semaine 1 et la semaine 2 post-
différenciation (augmentation significative de 50,3%).
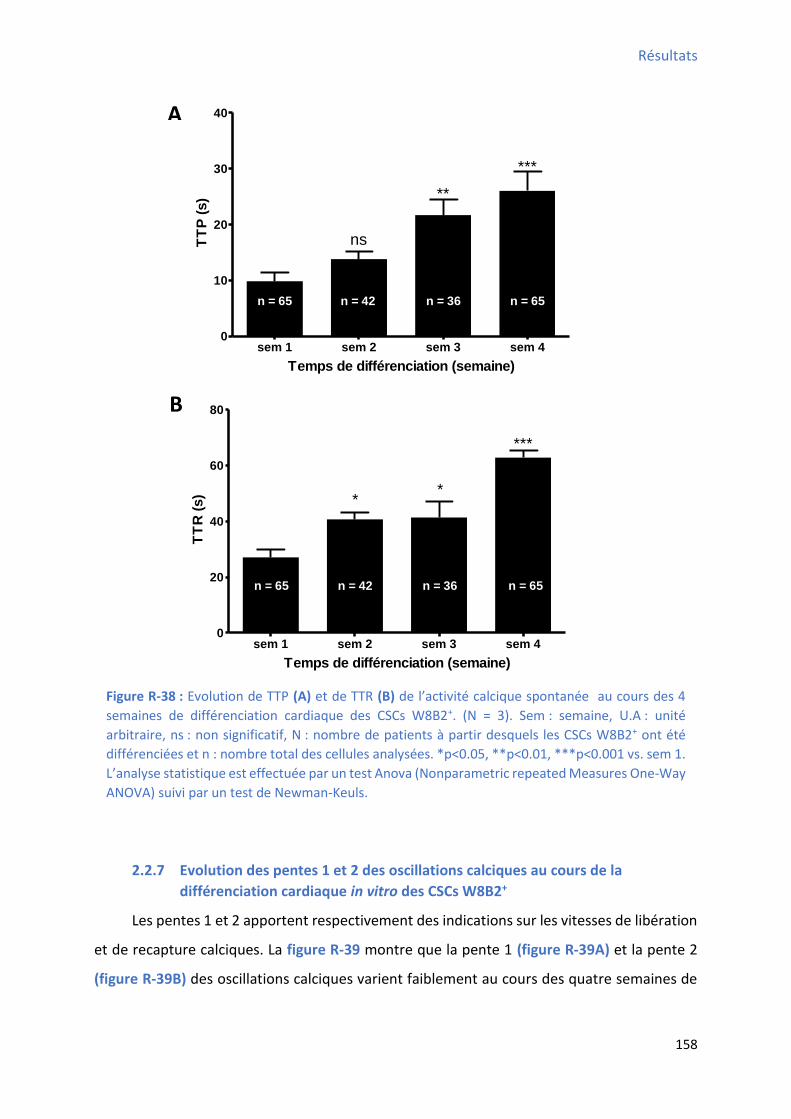
Résultats
158
2.2.7 Evolution des pentes 1 et 2 des oscillations calciques au cours de la
différenciation cardiaque in vitro des CSCs W8B2+
Les pentes 1 et 2 apportent respectivement des indications sur les vitesses de libération
et de recapture calciques. La figure R-39 montre que la pente 1 (figure R-39A) et la pente 2
(figure R-39B) des oscillations calciques varient faiblement au cours des quatre semaines de
sem 1 sem 2 sem 3 sem 40
10
20
30
40
***
ns
**
n = 65 n = 42 n = 36 n = 65
Temps de différenciation (semaine)
TT
P (s
)
Figure R-38 : Evolution de TTP (A) et de TTR (B) de l’a ti it al i ue spo ta e au ou s des semaines de différenciation cardiaque des CSCs W8B2+. (N = 3). Sem : semaine, U.A : unité
arbitraire, ns : non significatif, N : nombre de patients à partir desquels les CSCs W8B2+ ont été
différenciées et n : nombre total des cellules analysées. *p<0.05, **p<0.01, ***p<0.001 vs. sem 1.
L’a al se statisti ue est effe tu e pa u test A o a No pa a et i epeated Measu es O e-Way
ANOVA) suivi par un test de Newman-Keuls.
A
B
sem 1 sem 2 sem 3 sem 40
20
40
60
80
***
**
n = 65 n = 42 n = 36 n = 65
Temps de différenciation (semaine)
TT
R (s
)
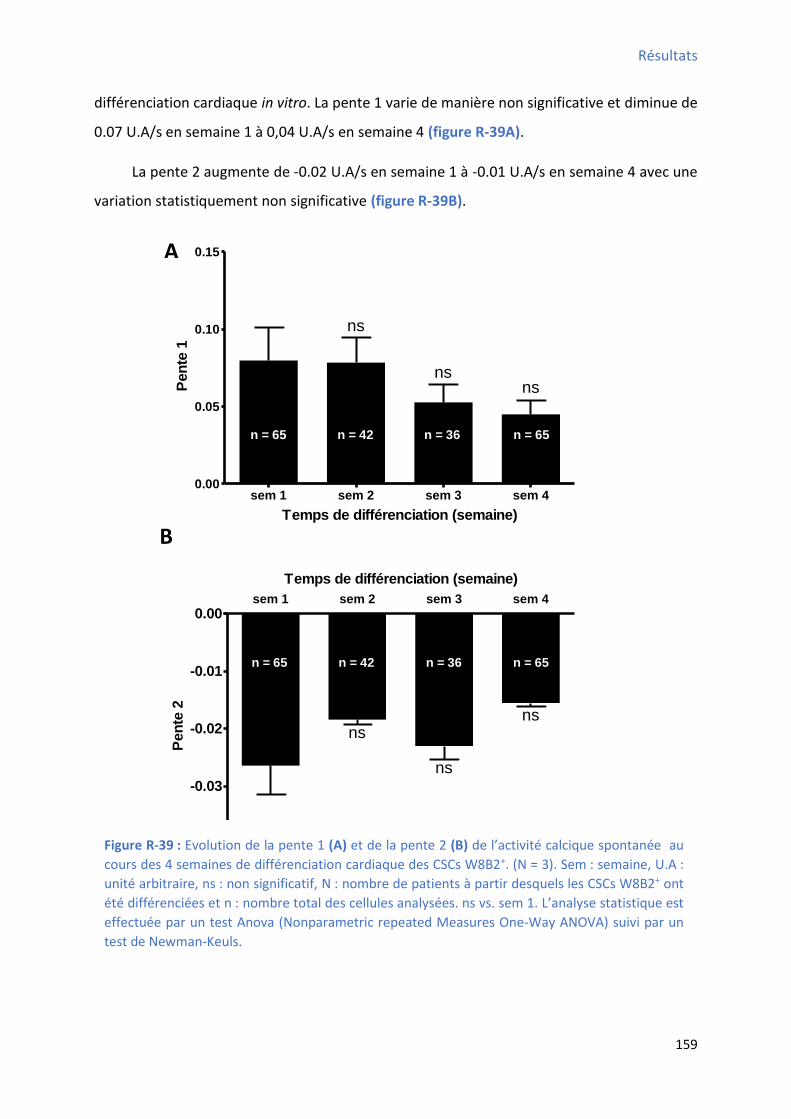
Résultats
159
différenciation cardiaque in vitro. La pente 1 varie de manière non significative et diminue de
0.07 U.A/s en semaine 1 à 0,04 U.A/s en semaine 4 (figure R-39A).
La pente 2 augmente de -0.02 U.A/s en semaine 1 à -0.01 U.A/s en semaine 4 avec une
variation statistiquement non significative (figure R-39B).
sem 1 sem 2 sem 3 sem 40.00
0.05
0.10
0.15
ns
nsns
n = 65 n = 42 n = 36 n = 65
Temps de différenciation (semaine)
Pen
te 1
A
B
Figure R-39 : Evolution de la pente 1 (A) et de la pente 2 (B) de l’a ti it al i ue spo ta e au cours des 4 semaines de différenciation cardiaque des CSCs W8B2+. (N = 3). Sem : semaine, U.A :
unité arbitraire, ns : non significatif, N : nombre de patients à partir desquels les CSCs W8B2+ ont
été différenciées et n : nombre total des cellules analysées. s s. se . L’a al se statisti ue est effectuée par un test Anova (Nonparametric repeated Measures One-Way ANOVA) suivi par un
test de Newman-Keuls.
sem 1 sem 2 sem 3 sem 4
-0.03
-0.02
-0.01
0.00
ns
ns
ns
n = 65 n = 42 n = 36 n = 65
Temps de différenciation (semaine)
Pen
te 2
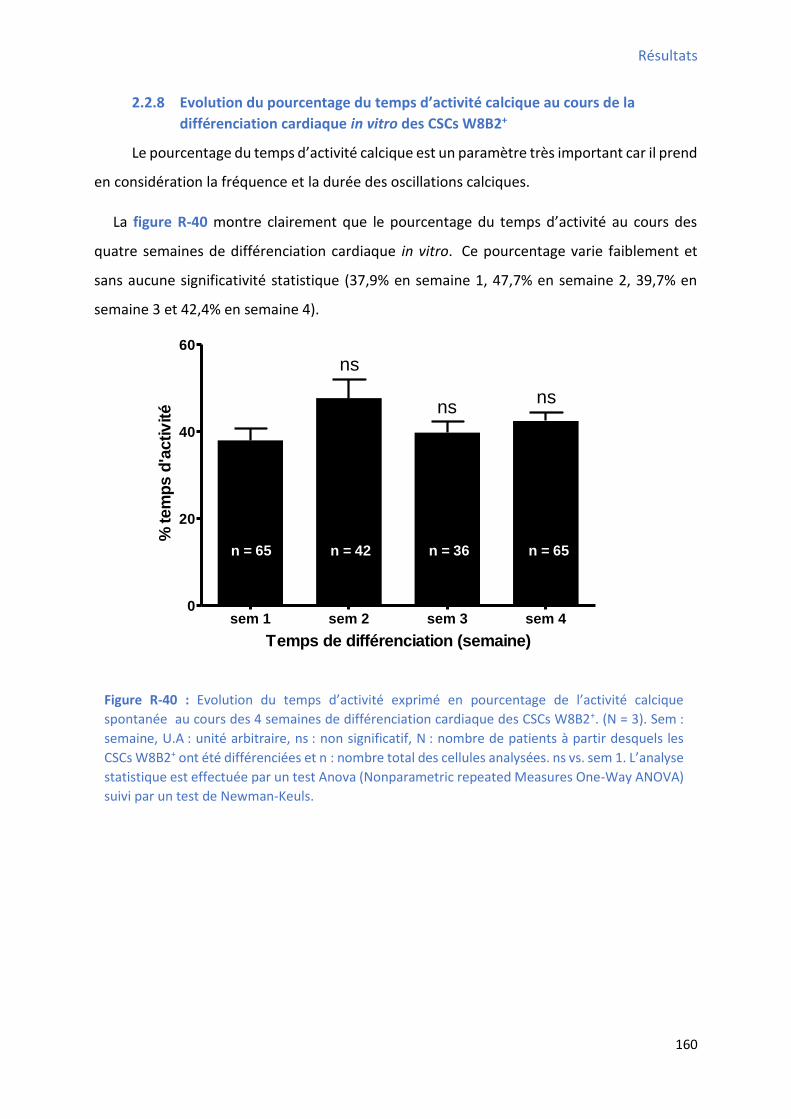
Résultats
160
2.2.8 Evolutio du pou e tage du te ps d’a tivit alcique au cours de la
différenciation cardiaque in vitro des CSCs W8B2+
Le pou e tage du te ps d’a ti it al i ue est u pa a t e t s i po ta t a il p e d
en considération la fréquence et la durée des oscillations calciques.
La figure R-40 o t e lai e e t ue le pou e tage du te ps d’a ti it au ou s des
quatre semaines de différenciation cardiaque in vitro. Ce pourcentage varie faiblement et
sans aucune significativité statistique (37,9% en semaine 1, 47,7% en semaine 2, 39,7% en
semaine 3 et 42,4% en semaine 4).
sem 1 sem 2 sem 3 sem 40
20
40
60ns
nsns
n = 65 n = 42 n = 36 n = 65
Temps de différenciation (semaine)
% te
mps
d'a
ctiv
ité
Figure R-40 : E olutio du te ps d’a ti it e p i e pou e tage de l’a ti it al i ue spontanée au cours des 4 semaines de différenciation cardiaque des CSCs W8B2+. (N = 3). Sem :
semaine, U.A : unité arbitraire, ns : non significatif, N : nombre de patients à partir desquels les
CSCs W8B2+ ont été différenciées et n : nombre total des cellules analysées. s s. se . L’a al se statistique est effectuée par un test Anova (Nonparametric repeated Measures One-Way ANOVA)
suivi par un test de Newman-Keuls.

Résultats
161
2.3 Identification des acteurs impliqués dans la génération des oscillations calciques
dans les CSCs W8B2+ différenciées
Pou alue le ôle des diff e ts a teu s ajeu s de l’ho ostasie al i ue da s la
génération des oscillations calciques observées dans les CSCs W8B2+ après 28 jours de
différenciation cardiaque in vitro, ous a o s tudi l’effet de l’i hi itio sp ifi ue de ha u
de es a teu s à l’aide d’age ts pha a ologi ues.
Soit :
L’ ha geu Na+/Ca2+ ou NCX: i hi pa l’age t pha a ologi ue “EA
La pompe à calcium du réticulum sarco-endoplasmique (SERCA) : inhibée par la
thapsigargine
Les epteu s à l’IP : inhibés par la xéstospongine C et le 2-APB (2-
Aminoethoxydiphenyl borate)
Les canaux calciques dépendants du voltage de type L : inhibés par la nifédipine
Les récepteurs à la ryanodine : inhibés par la ryanodine à forte concentration
Nous nous également testé :
L’effet de la sphi gosi e -phosphate, un second messager lipidique connu pour
moduler les stocks calciques intracellulaires
L’effet de l’i hi itio du a al BKCa ; u a al potassi ue do t l’a ti atio est
d pe da te de l’ho ostasie al i ues i t a ellulai e.
Les effets observés lors de cette étude pharmacologique sur les oscillations calciques sont
analysés grâce à Image J et les paramètres calciques décrits précédemment sont analysés
grâce à une application développée au laboratoire sous le langage IDL (Interactive Data
Language).
2.3.1 Effet de l’i hi itio de l’ ha geu NCX su les os illations calciques dans les
CSCs W8B2+ différenciées pendant 28 jours
Le “EA i hi iteu s le tif de l’ ha geu Na+/Ca2+ ou NCX, utilisé à 10 µM abolit
totalement les oscillations calciques observées sur les CSCs W8B2+ après 28 jours de
différenciation (figure R-41).
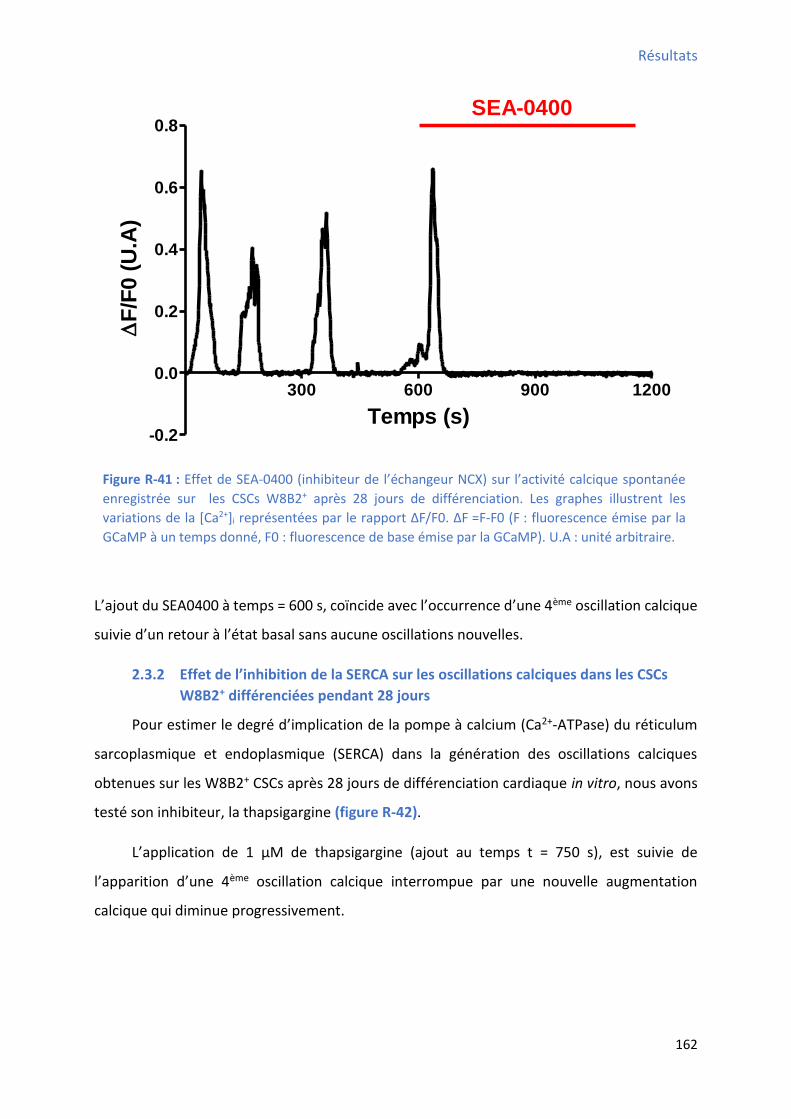
Résultats
162
L’ajout du “EA à te ps = s, oï ide a e l’o u e e d’u e ème oscillation calcique
suivie d’u etou à l’ tat asal sa s au u e os illatio s ou elles.
2.3.2 Effet de l’i hi itio de la SERCA su les os illatio s al i ues da s les CSCs W8B2+ différenciées pendant 28 jours
Pou esti e le deg d’i pli atio de la po pe à calcium (Ca2+-ATPase) du réticulum
sarcoplasmique et endoplasmique (SERCA) dans la génération des oscillations calciques
obtenues sur les W8B2+ CSCs après 28 jours de différenciation cardiaque in vitro, nous avons
testé son inhibiteur, la thapsigargine (figure R-42).
L’appli atio de µM de thapsigargine (ajout au temps t = 750 s), est suivie de
l’appa itio d’u e ème oscillation calcique interrompue par une nouvelle augmentation
calcique qui diminue progressivement.
Figure R-41 : Effet de SEA- i hi iteu de l’ ha geu NCX su l’a ti it al i ue spo ta e enregistrée sur les CSCs W8B2+ après 28 jours de différenciation. Les graphes illustrent les
variations de la [Ca2+]i ep se t es pa le appo t ΔF/F . ΔF =F-F0 (F : fluorescence émise par la
GCaMP à un temps donné, F0 : fluorescence de base émise par la GCaMP). U.A : unité arbitraire.
300 600 900 1200
-0.2
0.0
0.2
0.4
0.6
0.8SEA-0400
Temps (s)
F/F0
(U.A
)
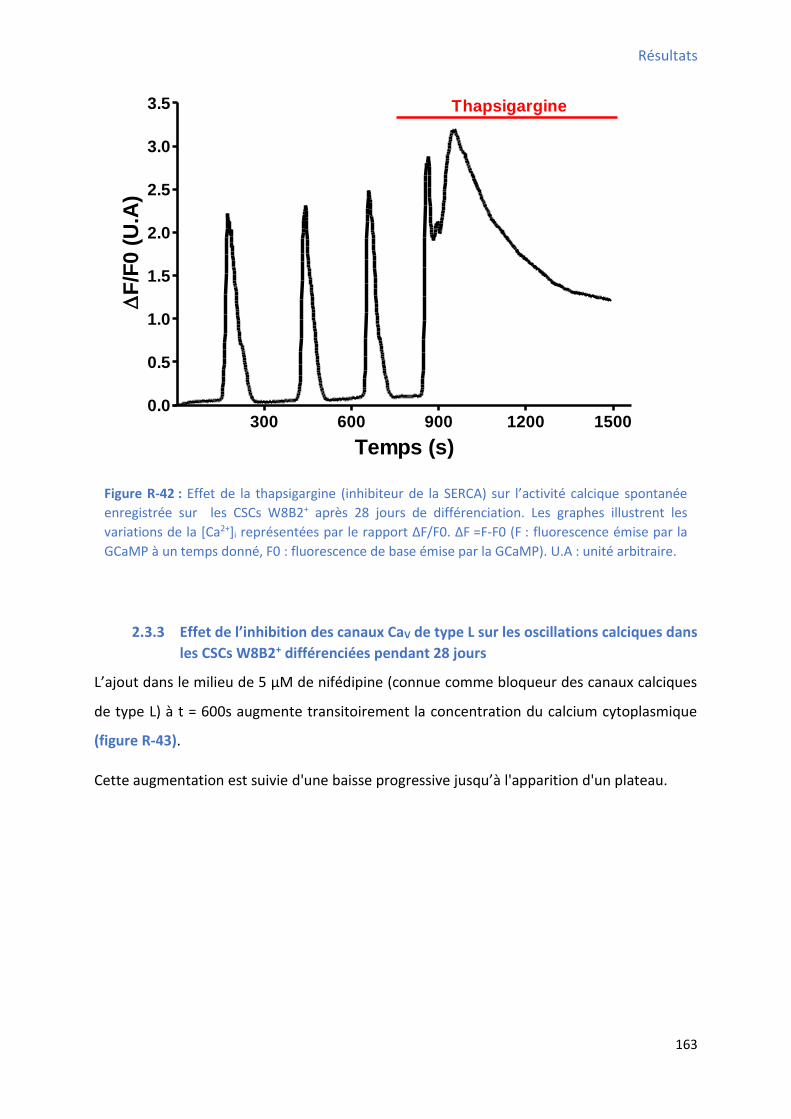
Résultats
163
2.3.3 Effet de l’i hi itio des a au CaV de type L sur les oscillations calciques dans
les CSCs W8B2+ différenciées pendant 28 jours
L’ajout da s le ilieu de µM de if dipi e o ue o e lo ueu des a au al i ues
de type L) à t = 600s augmente transitoirement la concentration du calcium cytoplasmique
(figure R-43).
Cette augmentation est suivie d'u e aisse p og essi e jus u’à l'appa itio d'u plateau.
300 600 900 1200 15000.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5 Thapsigargine
Temps (s)
F/F0
(U.A
)
Figure R-42 : Effet de la thapsiga gi e i hi iteu de la “ERCA su l’a ti it al i ue spo ta e enregistrée sur les CSCs W8B2+ après 28 jours de différenciation. Les graphes illustrent les
variations de la [Ca2+]i ep se t es pa le appo t ΔF/F . ΔF =F-F0 (F : fluorescence émise par la
GCaMP à un temps donné, F0 : fluorescence de base émise par la GCaMP). U.A : unité arbitraire.
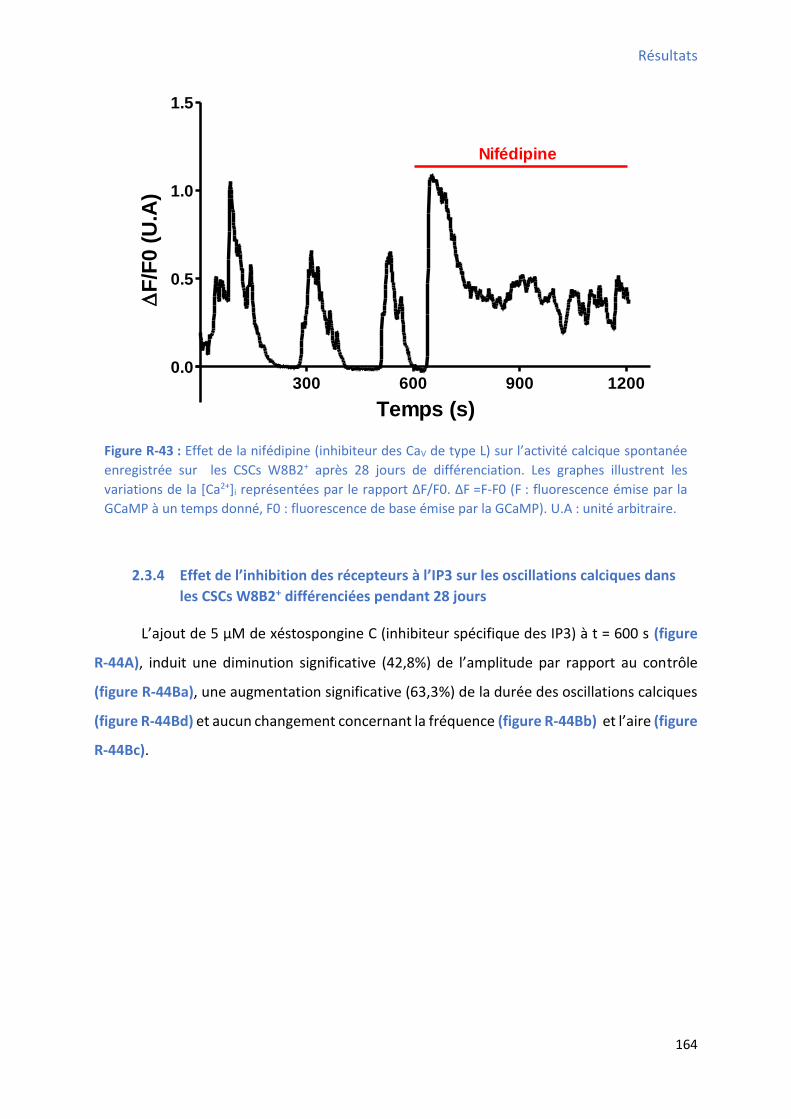
Résultats
164
2.3.4 Effet de l’i hi itio des epteu s à l’IP sur les oscillations calciques dans
les CSCs W8B2+ différenciées pendant 28 jours
L’ajout de µM de stospo gi e C i hi iteu sp ifi ue des IP à t = s (figure
R-44A), i duit u e di i utio sig ifi ati e , % de l’a plitude pa appo t au o trôle
(figure R-44Ba), une augmentation significative (63,3%) de la durée des oscillations calciques
(figure R-44Bd) et aucun changement concernant la fréquence (figure R-44Bb) et l’ai e (figure
R-44Bc).
300 600 900 12000.0
0.5
1.0
1.5
Nifédipine
Temps (s)
F/F0
(U.A
)
Figure R-43 : Effet de la nifédipine (inhibiteur des CaV de t pe L su l’a ti it al i ue spo ta e enregistrée sur les CSCs W8B2+ après 28 jours de différenciation. Les graphes illustrent les
variations de la [Ca2+]i ep se t es pa le appo t ΔF/F . ΔF =F-F0 (F : fluorescence émise par la
GCaMP à un temps donné, F0 : fluorescence de base émise par la GCaMP). U.A : unité arbitraire.
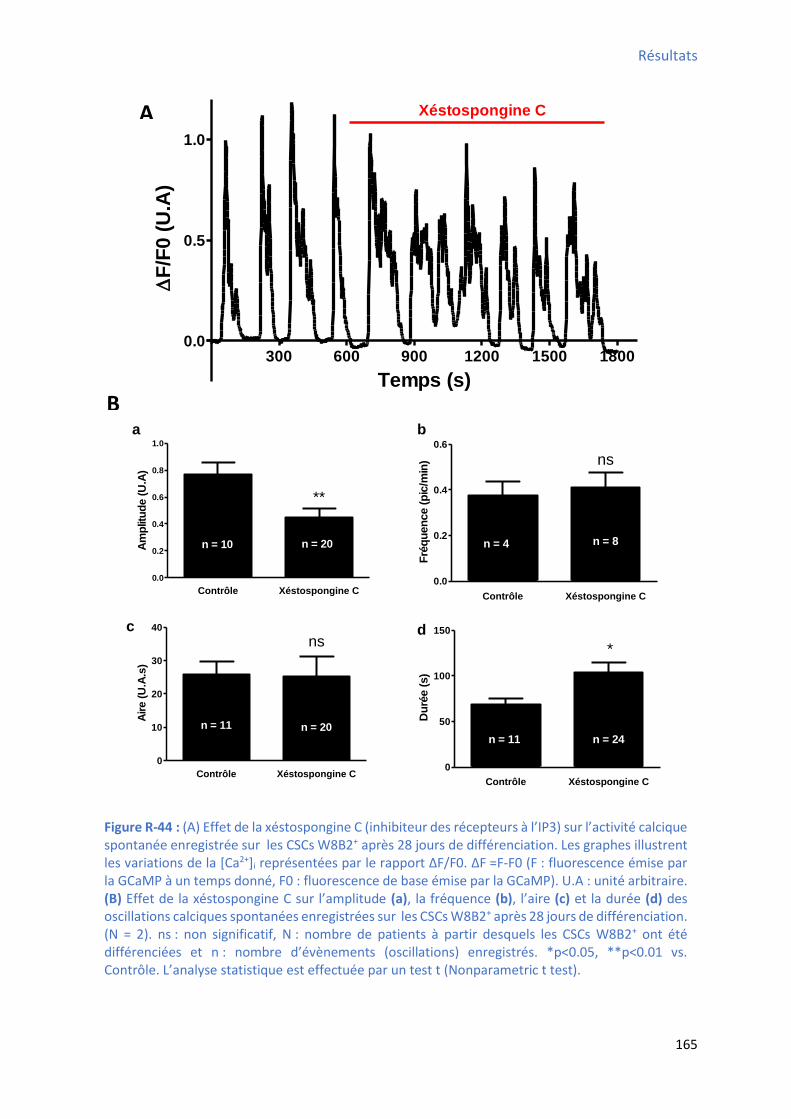
Résultats
165
300 600 900 1200 1500 18000.0
0.5
1.0
Xéstospongine C
Temps (s)
F/F0
(U.A
)
Figure R-44 : (A) Effet de la stospo gi e C i hi iteu des epteu s à l’IP su l’a ti it al i ue spontanée enregistrée sur les CSCs W8B2+ après 28 jours de différenciation. Les graphes illustrent les variations de la [Ca2+]i ep se t es pa le appo t ΔF/F . ΔF =F-F0 (F : fluorescence émise par la GCaMP à un temps donné, F0 : fluorescence de base émise par la GCaMP). U.A : unité arbitraire. (B) Effet de la stospo gi e C su l’a plitude (a), la fréquence (b), l’ai e (c) et la durée (d) des oscillations calciques spontanées enregistrées sur les CSCs W8B2+ après 28 jours de différenciation. (N = 2). ns : non significatif, N : nombre de patients à partir desquels les CSCs W8B2+ ont été différenciées et n : o e d’ e e ts (oscillations) enregistrés. *p<0.05, **p<0.01 vs. Co t ôle. L’a al se statisti ue est effe tu e pa u test t No pa a et i t test .
Contrôle Xéstospongine C0.0
0.2
0.4
0.6
0.8
1.0
**
n = 10 n = 20Am
plitu
de (U
.A)
Contrôle Xéstospongine C
0.0
0.2
0.4
0.6
ns
n = 4 n = 8
Fréq
uenc
e (p
ic/m
in)
Contrôle Xéstospongine C0
10
20
30
40
ns
n = 11 n = 20
Aire
(U.A
.s)
Contrôle Xéstospongine C0
50
100
150
*
n = 11 n = 24
Dur
ée (s
)a b
c d
B
A
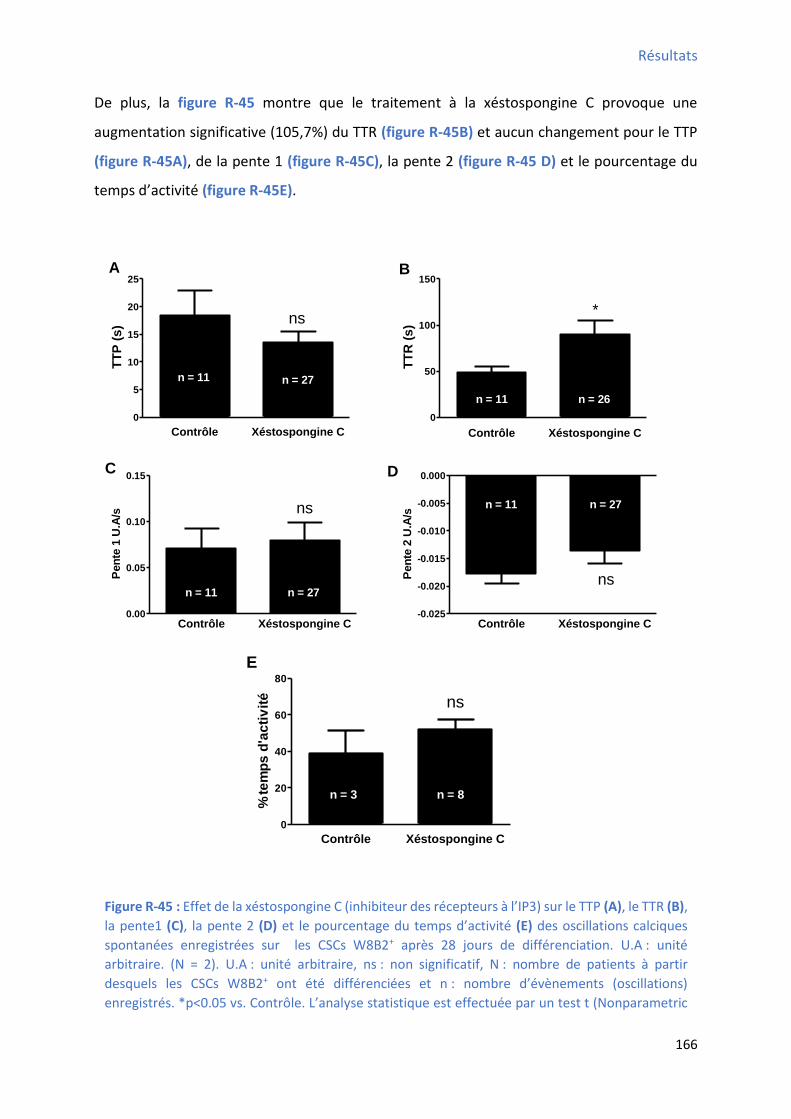
Résultats
166
De plus, la figure R-45 montre que le traitement à la xéstospongine C provoque une
augmentation significative (105,7%) du TTR (figure R-45B) et aucun changement pour le TTP
(figure R-45A), de la pente 1 (figure R-45C), la pente 2 (figure R-45 D) et le pourcentage du
te ps d’a ti it (figure R-45E).
Figure R-45 : Effet de la stospo gi e C i hi iteu des epteu s à l’IP su le TTP (A), le TTR (B),
la pente1 (C), la pente 2 (D) et le pou e tage du te ps d’a ti it (E) des oscillations calciques
spontanées enregistrées sur les CSCs W8B2+ après 28 jours de différenciation. U.A : unité
arbitraire. (N = 2). U.A : unité arbitraire, ns : non significatif, N : nombre de patients à partir
desquels les CSCs W8B2+ ont été différenciées et n : o e d’ e e ts os illatio s e egist s. *p< . s. Co t ôle. L’a alyse statistique est effectuée par un test t (Nonparametric
t test).
Contrôle Xéstospongine C0
5
10
15
20
25
ns
n = 11 n = 27
TT
P (
s)
Contrôle Xéstospongine C0
50
100
150
*
n = 11 n = 26T
TR
(s)
Contrôle Xéstospongine C0.00
0.05
0.10
0.15
ns
n = 11 n = 27
Pen
te 1
U.A
/s
Contrôle Xéstospongine C-0.025
-0.020
-0.015
-0.010
-0.005
0.000
ns
n = 11 n = 27
Pen
te 2
U.A
/s
A B
C D
Contrôle Xéstospongine C0
20
40
60
80
ns
n = 3 n = 8
% te
mps
d'a
ctiv
ité
E

Résultats
167
Nous avons ensuite testé le 2-APB connu comme étant est un autre inhibiteur des
epteu s à l’IP ais a e oi s de sp ifi it ue la stospo gi e C.
En effet, selon la concentration utilisée, le 2-APB agit sur des cibles différentes. Le 2-
APB est u a tago iste des epteu s à l’IP IC50 = μM et u odulateu des a au “OCs
(store-operated calcium channels) (Diver et al., 2001; Ma et al., 2002; Prakriya and Lewis,
2001). Le 2-APB stimule les libérations calciques à partir des SOCs pour de faibles
concentrations (< µM et les i hi e à fo tes o e t atio s jus u’à µM .
Les sultats o t e t ue l’ajout de et µM de 2-APB ’i pa te isi le e t pas
les oscillations calciques observées dans les CSCs W8B2+ après 28 jours de différenciation
cardiaque in vitro (figure R-46 A).
Cepe da t, l’a al se des pa a t es i ti ues des os illatio s calciques après traitement au
2-APB, o t e des effets su e tai s pa a t es o e l’a plitude, l’ai e, la du e, le TTP
et le TTR.
Le 2-APB à µM, aug e te l’a plitude de , % pa appo t au o t ôle et à l’i e se, la
diminue de 26% quand il est utilisé à 50 µM (figure R-46 Ba). Le 2-APB utilis à µM ’a pas
d’effet sig ifi atif su la f ue e (figure R-46 Bb), l’ai e (figure R-46 Bc), la durée (figure R-
46 Bd), le TTP (figure R-47 A), le TTR (figure R-47 B), la pente 1 (figure R-47 C), la pente 2
(figure R-47 D) et le pource tage du te ps d’a ti it (figure R-47 E).
Le 2-APB à µM, ’a pas d’effet su la f ue e (figure R-46 Bb) mais diminue de 61,5%
l’ai e (figure R-46 Bc) et diminue de 45,1% la durée (figure R-46 Bd). De plus, le 2-APB à 50
µM ’a pas d’effet su le TTP (figure R-47 A), la pente 1 (figure R-47 C) et la pente 2 (figure R-
47 D) mais diminue de 56,3% le TTR (figure R-45 B) et de 48,4% le pourcentage du temps
d’a ti it (figure R-47 E).
L’e se le de es sultats sugg e t u e pa ti ipatio sig ifi ati e de la voie IP3
se si le ui se ait i pli u e plus sp ifi ue e t da s l’i te sit du sig al al i ue sa s
altérer sa fréquence.
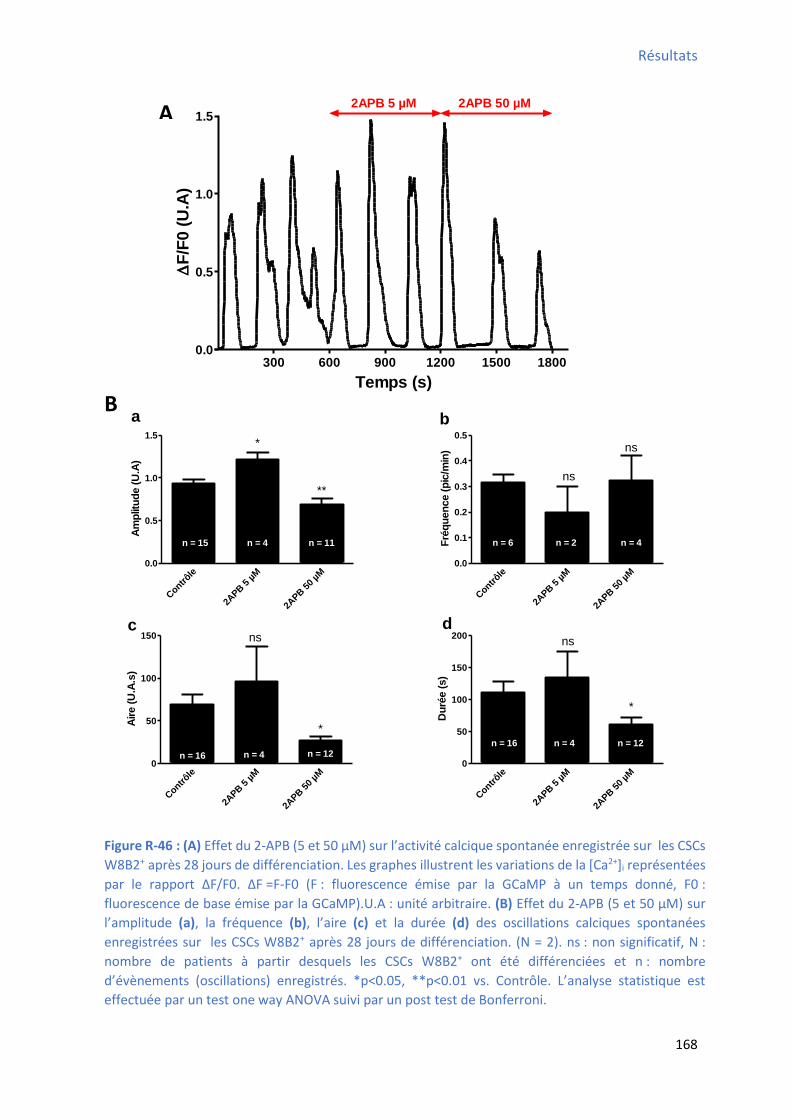
Résultats
168
300 600 900 1200 1500 18000.0
0.5
1.0
1.52APB 5 µM 2APB 50 µM
Temps (s)
F/F0
(U.A
)
A
Figure R-46 : (A) Effet du 2-APB et µM su l’a ti it al i ue spo ta e e egist e su les C“Cs W8B2+ après 28 jours de différenciation. Les graphes illustrent les variations de la [Ca2+]i représentées
pa le appo t ΔF/F . ΔF =F-F0 (F : fluorescence émise par la GCaMP à un temps donné, F0 :
fluorescence de base émise par la GCaMP).U.A : unité arbitraire. (B) Effet du 2-APB (5 et 50 µM) sur
l’a plitude (a), la fréquence (b), l’ai e (c) et la durée (d) des oscillations calciques spontanées
enregistrées sur les CSCs W8B2+ après 28 jours de différenciation. (N = 2). ns : non significatif, N :
nombre de patients à partir desquels les CSCs W8B2+ ont été différenciées et n : nombre
d’ e e ts os illatio s e egist s. *p< . , **p< . s. Co t ôle. L’a al se statisti ue est effectuée par un test one way ANOVA suivi par un post test de Bonferroni.
B
Contrô
le
2APB 5
µM
2APB 50
µM0.0
0.5
1.0
1.5*
n = 15 n = 11n = 4
**
Am
plitu
de (U
.A)
Contrô
le
2APB 5
µM
2APB 50
µM0.0
0.1
0.2
0.3
0.4
0.5
ns
ns
n = 6 n = 2 n = 4Fréq
uenc
e (p
ic/m
in)
Contrô
le
2APB 5
µM
2APB 50
µM0
50
100
150 ns
*
n = 16 n = 12n = 4
Aire
(U.A
.s)
Contrô
le
2APB 5
µM
2APB 50
µM0
50
100
150
200 ns
*
n = 16 n = 12n = 4
Dur
ée (s
)
a b
c d
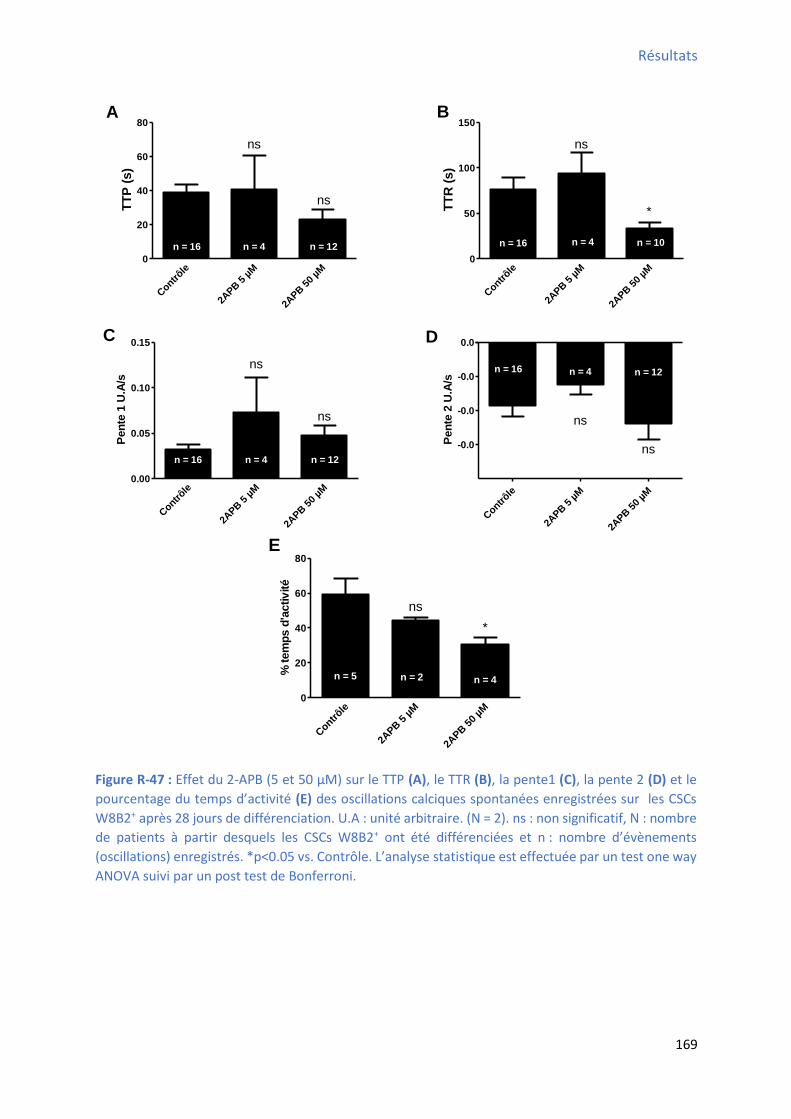
Résultats
169
Contrô
le
2APB 5
µM
2APB 50
µM0
20
40
60
80
ns
*
n = 5 n = 4n = 2% te
mps
d'a
ctiv
ité
E
Contrô
le
2APB 5
µM
2APB 50
µM0
20
40
60
80
ns
ns
n = 4n = 16 n = 12
TTP
(s)
Contrô
le
2APB 5
µM
2APB 50
µM0
50
100
150
ns
*
n = 16 n = 4 n = 10
TTR
(s)
Contrô
le
2APB 5
µM
2APB 50
µM0.00
0.05
0.10
0.15
ns
ns
n = 16 n = 4 n = 12
Pen
te 1
U.A
/s
Contrô
le
2APB 5
µM
2APB 50
µM
-0.0
-0.0
-0.0
0.0
ns
ns
n = 16 n = 4 n = 12
Pen
te 2
U.A
/s
A B
C D
Figure R-47 : Effet du 2-APB (5 et 50 µM) sur le TTP (A), le TTR (B), la pente1 (C), la pente 2 (D) et le
pou e tage du te ps d’a ti it (E) des oscillations calciques spontanées enregistrées sur les CSCs
W8B2+ après 28 jours de différenciation. U.A : unité arbitraire. (N = 2). ns : non significatif, N : nombre
de patients à partir desquels les CSCs W8B2+ ont été différenciées et n : o e d’ e e ts os illatio s e egist s. *p< . s. Co t ôle. L’a al se statistique est effectuée par un test one way
ANOVA suivi par un post test de Bonferroni.
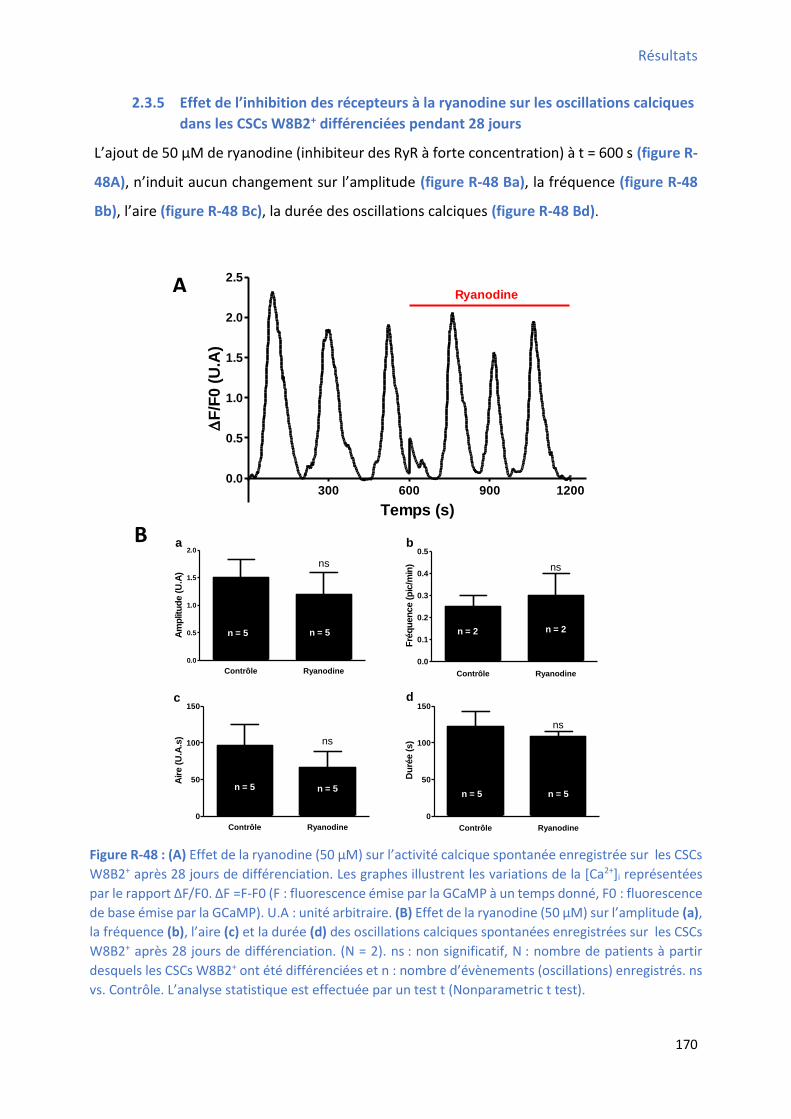
Résultats
170
2.3.5 Effet de l’i hi itio des epteu s à la a odi e su les os illatio s al i ues dans les CSCs W8B2+ différenciées pendant 28 jours
L’ajout de µM de a odi e i hi iteu des R R à fo te o e t atio à t = s (figure R-
48A), ’i duit au u ha ge e t su l’a plitude (figure R-48 Ba), la fréquence (figure R-48
Bb), l’ai e (figure R-48 Bc), la durée des oscillations calciques (figure R-48 Bd).
300 600 900 12000.0
0.5
1.0
1.5
2.0
2.5Ryanodine
Temps (s)
F/F0
(U.A
)
Figure R-48 : (A) Effet de la a odi e µM su l’a ti it al i ue spo ta e e egist e su les C“Cs W8B2+ après 28 jours de différenciation. Les graphes illustrent les variations de la [Ca2+]i représentées
pa le appo t ΔF/F . ΔF =F-F0 (F : fluorescence émise par la GCaMP à un temps donné, F0 : fluorescence
de base émise par la GCaMP). U.A : unité arbitraire. (B) Effet de la a odi e µM su l’a plitude (a),
la fréquence (b), l’ai e (c) et la durée (d) des oscillations calciques spontanées enregistrées sur les CSCs
W8B2+ après 28 jours de différenciation. (N = 2). ns : non significatif, N : nombre de patients à partir
desquels les CSCs W8B2+ ont été différenciées et n : o e d’ e e ts (oscillations) enregistrés. ns
s. Co t ôle. L’a al se statisti ue est effe tu e pa u test t No pa a et i t test .
Contrôle Ryanodine0.0
0.5
1.0
1.5
2.0
n = 5 n = 5
ns
Am
plitu
de (U
.A)
Contrôle Ryanodine
0.0
0.1
0.2
0.3
0.4
0.5
n = 2 n = 2
ns
Fréq
uenc
e (p
ic/m
in)
Contrôle Ryanodine0
50
100
150
n = 5 n = 5
ns
Aire
(U.A
.s)
Contrôle Ryanodine0
50
100
150
n = 5 n = 5
ns
Dur
ée (s
)
a b
c d
A
B
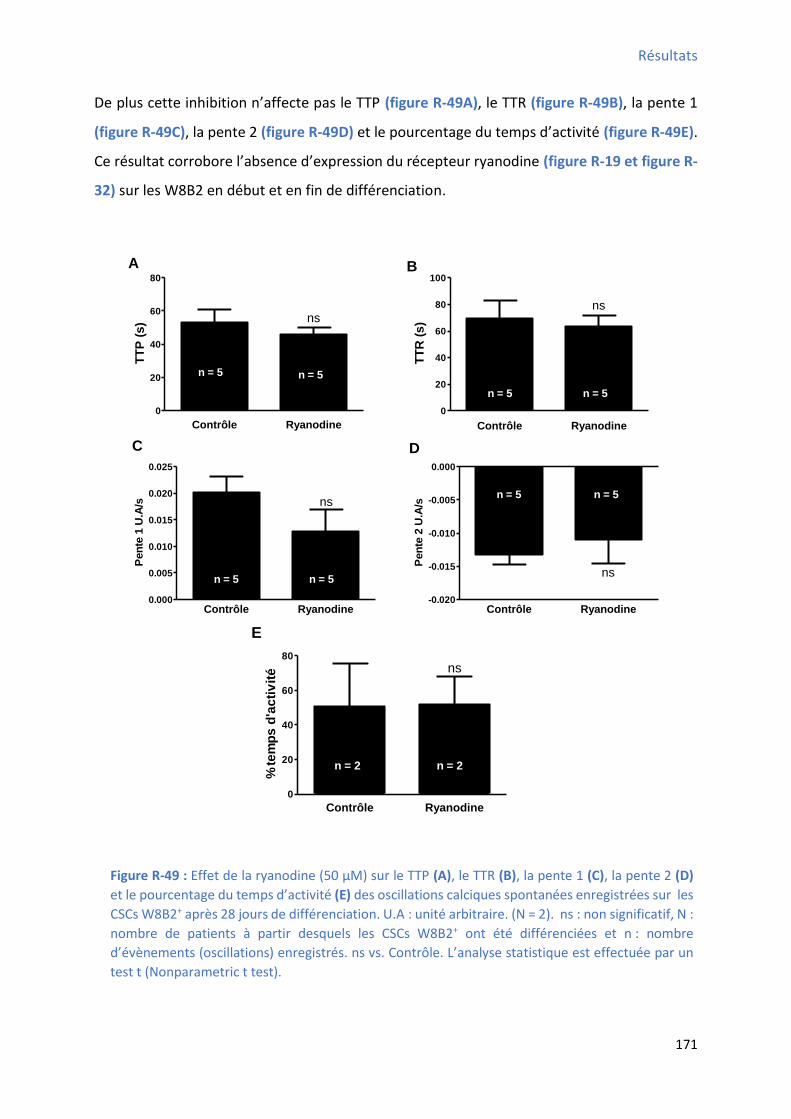
Résultats
171
De plus ette i hi itio ’affe te pas le TTP (figure R-49A), le TTR (figure R-49B), la pente 1
(figure R-49C), la pente 2 (figure R-49D) et le pou e tage du te ps d’a ti it (figure R-49E).
Ce sultat o o o e l’a se e d’e p essio du epteu a odi e (figure R-19 et figure R-
32) sur les W8B2 en début et en fin de différenciation.
Contrôle Ryanodine0
20
40
60
80
n = 5 n = 5
ns
TT
P (
s)
Contrôle Ryanodine0
20
40
60
80
100
n = 5 n = 5
ns
TT
R (s
)
Contrôle Ryanodine0.000
0.005
0.010
0.015
0.020
0.025
n = 5 n = 5
ns
Pen
te 1
U.A
/s
Contrôle Ryanodine-0.020
-0.015
-0.010
-0.005
0.000
n = 5 n = 5
ns
Pen
te 2
U.A
/s
A B
C D
Contrôle Ryanodine0
20
40
60
80
n = 2 n = 2
ns
% te
mps
d'a
ctiv
ité
E
Figure R-49 : Effet de la ryanodine (50 µM) sur le TTP (A), le TTR (B), la pente 1 (C), la pente 2 (D)
et le pourcentage du te ps d’a ti it (E) des oscillations calciques spontanées enregistrées sur les
CSCs W8B2+ après 28 jours de différenciation. U.A : unité arbitraire. (N = 2). ns : non significatif, N :
nombre de patients à partir desquels les CSCs W8B2+ ont été différenciées et n : nombre
d’ e e ts os illatio s e egist s. s s. Co t ôle. L’a al se statistique est effectuée par un
test t (Nonparametric t test).

Résultats
172
Chapitre II : Rôle du canal BKCa dans la régulation de la prolifération et de
l’auto-renouvellement des CSCs W8B2+
Les résultats présentés dans ce chapitre sont présentés sous format article scientifique.
Ce dernier est soumis dans le journal « The FEBS Journal » et il est actuellement en révision.
L’a ti le s ie tifi ue s’i titule « FUNCTIONAL BKCa CHANNEL IN HUMAN RESIDENT CARDIAC
STEM CELLS EXPRESSING W8B2».
1 Cad e de l’ tude
Le œu hu ai adulte a ite plusieu s populatio s de ellules sou hes a dia ues ui
expriment des marqueurs spécifiques. Récemment, une nouvelle population positive pour le
marqueur W8B2 a été identifiée. Ces cellules souches cardiaques exprimant W8B2 (CSCs
W8B2+) ont une origine mésenchymateuse et présentent un fort potentiel thérapeutique
(Zhang et al., 2015a). Cependant, le profil électrophysiologique de ces cellules n'a pas encore
été caractérisé.
Nous nous sommes donc focalisés sur la caractérisation des canaux fonctionnels
exprimés par les CSCs W8B2+ et leurs éventuels rôles dans la régulation de leurs propriétés
(auto-renouvellement, prolifération et migration cellulaire) qui sont importantes pour le
maintien de leur nombre et de leur mobilité pour la réparation tissulaire tout au long de la vie.
Grâce à la technique du patch-clamp, nous avons montré pour la première fois la signature
le t oph siologi ue d’u ou a t BKCa dans ces cellules.
Les canaux potassiques à large conductance activés par le calcium, ou BKCa, sont des
a au ui s’ou e t suite aux changements du potentiel électrique membranaire et/ou suite
à l’aug e tatio du calcium intracellulaire. Ces canaux se trouvent dans de nombreux tissus
et participent à divers processus cellulaires (Toro et al., 2014). Les canaux BKCa sont impliqués
dans la modulation de l'excitabilité vasomotrice et nerveuse en régulant le potentiel de
membrane et la signalisation calcique (Nelson and Quayle, 1995). En outre, cette large
conductance potassique contribue au maintien du potentiel de membrane dans les petits
vaisseaux myogéniques (Waldron and Cole, 1999).
Il est intéressant de noter que ce canal joue un rôle clé dans de nombreux processus
biologiques tels que le métabolisme cellulaire, la prolifération, la migration et l'expression des
gènes. En plus de ses rôles physiologiques, le canal BKCa est impliqué dans plusieurs

Résultats
173
pathologies comme l'obésité ou le cancer (Toro et al., 2014). Cepe da t, l’e p essio
fonctionnelle et le rôle des canaux BKCa dans les cellules souches cardiaques humaines,
restent peu étudiés.
Après avoir identifié le canal fonctionnel BKCa dans notre modèle de CSCs W8B2+, nous
avons donc voulu étudier son rôle physiologique dans la régulation des processus
fondamentaux de ces cellules souches comme l'auto-renouvellement, la prolifération et la
migration.
2 Résumé des résultats
Co e ous l’a o s d jà d it p de e t hapit e I résultat (p 114) Les CSCs
W8B2+ isolées par tri magnétique suivi du tri par FACS, ont une morphologie étalée et
fusifo e o e elle des fi o lastes et poss de t des apa it s d’auto-renouvellement
(démontrées par le test de formation de colonies « CFU-Fs »). De plus, ces cellules
p olif aie t apide e t a e u te ps de d dou le e t d’e i o h. L'a al se pa
cytométrie de flux a révélé que les CSCs W8B2+ expriment fortement les marqueurs de surface
associés aux cellules souches mésenchymateuses (W8B2, CD29, CD73 et CD105) et sont
négatives pour les marqueurs hématopoïétiques (CD34, CD45 et CD133). Au niveau génique,
les CSCs W8B2+ ont montré une forte expression des facteurs de transcription cardiaque
spécifiques précoces comme GATA4 et MEF2C mais pas Nkx2.5. De plus, ces cellules expriment
les transcrits codant pour la connexine 43 (isoforme cardiaque) mais pas les transcrits codant
pour les structures contractiles cardiaques (ACTC1, TNNT2, b-MHC).
Les enregistrements de courants obtenus par la technique de patch clamp (en
configu atio ellule e ti e , o t l la p se e d’u ou a t potassi ue de t pe BKCa
bloqué par la paxilline. L'identité moléculaire du canal BKCa a été confirmée au niveau
transcriptionnel par RT-PCR et au niveau protéique par Western Blot dans les CSCs W8B2+.
Pour évaluer le rôle du canal BKCa dans la prolifération cellulaire des CSCs W8B2+, la paxilline
a t utilis e pou lo ueu sp ifi ue e t e a al. La pa illi e utilis e à μM et μM a
progressivement inhibé la prolifération à partir du 4è e jou e ultu e. D’aut e pa t, l'a al se
du le ellulai e pa to t ie de flu , a o t ue la pa illi e μM et pa ti uli e e t
à 10 µM) a significativement augmenté la fraction de la population cellulaire en phase G1 et
considérablement diminu la f a tio de la populatio ellulai e e phase “/G . L’i hi itio
du canal BKCa a affecté l'auto-renouvellement des CSCs W8B2+. En effet, la paxilline (utilisée à

Résultats
174
, et μM a o sid a le e t duit le o e de olo ies CFU-Fs après dix jours de
culture.
Concernant la migration cellulaire évaluée par le test de blessure (wound healing), les
résultats ont montré que le blocage du canal BKCa par la paxilline ne réprime pas les capacités
migratoires des cellules.
L’e se le de es sultats d o t e que le canal BKCa est probablement un
régulateur clé de la prolifération et de l'auto-renouvellement des CSCs W8B2+ et fournit une
base pour une meilleure compréhension de la physiologie de ces cellules. De nouvelles
recherches, basées sur la relation ent e le pote tiel de e a e et l’a ti it al i ue via les
canaux BKCa, semblent primordiales pour ouvrir de nouvelles voies thérapeutiques.
3 Article scientifique: « FUNCTIONAL BKCa CHANNEL IN HUMAN RESIDENT CARDIAC
STEM CELLS EXPRESSING W8B2».

Résultats
175
FUNCTIONAL BKCa CHANNEL IN HUMAN RESIDENT CARDIAC STEM CELLS EXPRESSING
W8B2
Ayad O , Magaud C, Sebille S, Bescond J, Cognard C, Faivre JF, Bois P* and A Chatelier*
Equipe Transferts Ioniques et Rythmicité Cardiaque, Laboratoire Signalisation et Transports Ioniques
Membranaires, ERL CNRS 7368, EA 7349, Université de Poitiers *These authors have contributed equally to this work
ABSTRACT
Recently, a new population of resident cardiac stem cells positive for W8B2 marker has been
identified. These cardiac stem cells (CSCs) are considered an ideal cellular source to repair
myocardial damage after infarction. However, the electrophysiological profile of these cells
has not been characterized yet. We first establish the conditions of isolation and expansion of
W8B2+ CSCs from human heart biopsies using magnetic sorting system followed by flow
cytometry cell sorting. These cells display a spindle-shaped morphology, are highly
proliferative and possess self-renewal capacity demonstrated by their ability to form colonies.
Besides, W8B2+ CSCs are positive for mesenchymal markers but negative for hematopoietic
and endothelial ones. RT-qPCR and immunostaining experiments show that W8B2+ CSCs
express some early cardiac-specific transcription factors but lack the expression of cardiac-
specific structural genes. Using patch-clamp in whole cell configuration, we show for the first
time the electrophysiological signature of BKCa current in these cells. Accordingly, RT-PCR and
western blotting analysis confirm the presence of BKCa at both mRNA and protein levels in
W8B2+ CSCs. Interestingly, BKCa channel inhibition by paxilline decreases cell proliferation in
a concentration-dependent manner and cell cycling progression by accumulating the cells at
G0/G1 phase. The inhibition of BKCa also decreased self-renewal capacity but did not affect
W8B2+ CSCs migration. Taken together, our results are consistent with an important role of
BKCa channels on cell cycle progression and self-renewal in human cardiac stem cells.
Key words: W8B2+ CSCs, BKCa channel, cell proliferation, cell cycle; migration; patch-clamp;
self-renewal.
INTRODUCTION
The adult human heart harbors several populations of cardiac stem/progenitor cells that
express specific markers. Recently a new population was identified which is positive for W8B2
marker (also called mesenchymal stem cell antigen-1) and shows high expression of
mesenchymal but not hematopoietic nor endothelial markers. W8B2 positive cardiac stem
cells (W8B2+ CSCs) exhibit a strong therapeutic potential when transplanted into a chronic
myocardial infarction rat model [1]. Self-renewal, proliferation and migration are important
properties that allow the perpetuation of progenitor cells and their mobility for repairing
processes during life. However, the mechanisms that control these properties still have to be
characterized in W8B2+ CSC.

Résultats
176
High-conductance calcium-activated potassium channels (BKCa) are found in many tissues and
participate in variety of cellular processes [2]. These channels are gated open by both binding
of intracellular calcium and membrane depolarization. BKCa channels are involved in the
modulation of vasomotor and nerve excitability by regulating membrane potential and
calcium signaling [3]. Moreover, this large potassium conductance contributes to the
maintenance of the membrane potential in small myogenic vessels [3-5]. Interestingly,
increasing evidence indicates that the channel plays a key role in numerous biological
processes such as cell metabolism, proliferation, migration, and gene expression. It strongly
impacts physiological cell functions, for example in human primary skeletal myoblast [6] or in
pathologies like obesity or brain, prostate, and mammary cancers [2]. However, whether
these high-conductance calcium-activated potassium channels are functionally expressed in
human cardiac progenitor cells remains poorly understood. Although human cardiac W8B2+
progenitor cells have been well characterized, the physiological role of BKCa channel in these
cells type has never been reported.
In the present study, we identify BKCa channel current expressed in human cardiac W8B2+
progenitor cells isolated from human atrial appendage. We also investigate the role of BKCa
on proliferation, migration and self-renewal processes in these progenitor cells.
MATERIALS AND METHODS
Isolation of W8B2+ CSCs
Human right atrial specimens were obtained from 10 adult patients (mean age 72.7 ± 3.2
years, 9 males and 1 female). The right human atrium samples are operational waste resulting
from the implementation of extracorporeal cardiac surgery, such as coronary artery bypass
surgery, and are obtained in cooperation with the University hospital of Poitiers. All
procedures were carried out in accordance with the Declaration of Helsinki. Freshly harvested
specimens were rinsed with PBS and manually minced into 1–2 mm3 fragments and subjected
to enzymatic digestion with collagenase A (1 mg/mL, Sigma-Aldrich) for 20 minutes at 37°C.
The tissue fragments were plated onto fibronectin- oated dishes μg/ L, Ro he Diagnostics) and cultured in explant medium containing IMDM medium (Lonza) supplemented
with 20% fetal bovine serum (Biowest), 0. M β-mercaptoethanol (Sigma-Aldrich), 2 mM L-
gluta i e, μg/ L st epto i “ig a-Ald i h , μg/ L pe i illi “ig a-Aldrich) and
. μg/ L a phote i i B “ig a-Aldrich), at 37 °C and 5% CO2.
Magnetic-activated cell sorting
After 2–3 weeks, monolayers of adherent cells outgrowth from the adherent tissue fragments
were collected using enzymatic digestion with Accumax® (Sigma-Aldrich) and partially
enriched for W8B2 marker by magnetic Cell Sorting System (Miltenyi Biotec, Bergisch
Gladbach, Germany) using W8B2 antibody. The cells were incubated for 15 minutes at 4-8°C
in PBS-BSA 0.5% -2 mM EDTA buffer containing FCR Blocking Reagent® (Miltenyi Biotec,
Bergisch Gladbach, Germany), mouse anti-MSCA-1 monoclonal antibody coupled to magnetic
micro-beads (Miltenyi Biotec, Bergisch Gladbach, Germany). After being washed, the cells
e e passed th ough a μ po osit lo ell sie e Co i g pla ed i to a MAC“ ell separation column (Miltenyi Biotec, Bergisch Gladbach, Germany) positioned on a magnet.

Résultats
177
Inside the column, the retained cells were washed with PBS-BSA buffer 0.5% - 2 mM EDTA and
then harvested in this same buffer by separating the column from the magnet. The positive
fraction (W8B2+ cells) were seeded in growth medium containing 25% EGM-2 (Lonza) and 75%
M Lo za , supple e ted ith % fetal o i e se u Bio est , μg/ L st epto i (Sigma-Ald i h a d . μg/ L a phote i i B “ig a-Aldrich), 0.1 mM nonessential amino
acids, 100 U/mL penicillin, cultured at 37°C 5% CO2.
Flow cytometry cell sorting
After magnetic cell sorting, confluent cells were labeled with W8B2 PE-conjugated antibody
and sorted using a FACS Aria Flow cytometer and sorter. Cells were first rinsed with PBS and
then harvested by Accumax® (Sigma-Aldrich) enzymatic digestion. The cells were taken up in
PBS buffer - BSA 0.5% - 2 mM EDTA and labeled with the isotypic antibody coupled to
phycoerythrin (1:11, Miltenyi Biotec, Bergisch Gladbach, Germany) or MSCA-1 (W8B2)
antibody coupled to phycoerythrin (1:11, Miltenyi Biotec, Bergisch Gladbach, Germany) for 10
min, in the dark, at 2-8 ° C. After labelling, the cells were washed with PBS buffer - 0.5% BSA -
2 mM EDTA and sorted by a flow cytometer (FACS Aria III, BD). After cell sorting, the collected
W8B2+ cells were seeded into flasks with growth medium.
Cell surface marker analysis
The cell surface markers of sorted human W8B2+ CSCs were analyzed with flow cytometry.
The cells were rinsed twice with pre-cooled PBS and re-suspended in cold PBS containing
specific human antibodies from Miltenyi Biotec, Bergisch Gladbach, Germany: Mouse IgG1
CD117-APC (1:11), Mouse IgG2a CD45-VioBlue (1:11), Mouse IgG1 CD133/1-VioBright FITC
(1:11), Mouse IgG1 CD105-PE(1:11), Mouse IgG2a CD34-PerCP-Vio700 (1:11), Mouse IgG1
CD90-PE-Vio770(1:11), Rec Human IgG1 (S) CD106-FITC : , Mouse IgG κ CD -PE-Vio770
(1:11), Mouse IgG1 CD73-APC (1:11). For each antibody, isotype control antibodies were used:
Mouse IgG2a-VioBlue (1:11), REA Control (S)-FITC (1:11), Mouse IgG1-VioBright FITC (1:11),
Mouse IgG1-PE (1:11), Mouse IgG2a-PerCP-Vio700 (1:11), Mouse IgG1-PE-Vio770 (1:11),
Mouse IgG1-APC (1:11). For flow cytometry florescence compensation, MACS Comp Bead Kit,
anti-mouse Igk (Miltenyi Biotec, Bergisch Gladbach, Germany) and MACS Comp Bead Kit, anti-
REA (Miltenyi Biotec, Bergisch Gladbach, Germany) were used.
The cells were incubated at 4 °C in the dark for 10 min, then washed with PBS, and re-
suspended for flow cytometry analysis. The percentage of positive cells was analyzed with
FACS Aria Flow cytometer based on over 20000 events.
Colony formation assay (colony-forming unit fibroblasts)
W8B2+ CSCs were seeded at a density of 5 cells/cm² in a 6-well plate, in growth medium. The growth medium was renewed every 3-4 days. After 10 days of culture, the cells were rinsed with PBS, fixed for 10 min with 4% PFA and finally stained with 0.5% Crystal violet for 30 min (Sigma-Aldrich). The colonies were photographed under a phase contrast microscope and the colonies were counted. Proliferation assay

Résultats
178
Cells were plated at a density of 133 cells/cm2 in 35mm dishes with growth medium for 9 days.
Cells were then synchronized to G0/G1 phase with a culture medium containing 1% FBS for 12
h. Every day, cell number is counted for each condition (control vs paxilline treated) using
Malassez cell chamber.
Cell cycle analysis with propidium iodide incorporation
Cells were plated and cultured in 35 mm cell culture dishes at a density of 5×102 cells/cm2.
Cells were then synchronized to G0/G1 phase with a culture medium containing 1% FBS for 12
h. Briefly, the cells were cultured in normal culture medium with ion channel blocker paxilline
for 9 days. The cells were lifted using 0.125% trypsin, washed with PBS, and fixed with 70%
ethanol for 30 minutes, cells were incubated with the staining solution (0.1% triton, 50µg/mL
of RNase A (Sigma-Aldrich) and 5 µg/mL of propidium iodide (Sigma-Aldrich) in PBS in the dark
for 90 minutes. Data were acquired using FACS VERSE cytometer (BD) and the percentage of
cell cycle phases was determined using and FLOWJO software (Flowjo LLC).
Immunostaining
W8B2+ cells were seeded on gelatin-fibronectin coated glass slide. Cells were then fixed,
permeabilized and incubated overnight with primary antibodies directed against connexin-43
(1:500, mouse monoclonal antibody, BD Transduction Laboratories, San Jose, CA), GATA4
(1:250, mouse monoclonal antibody, Santa Cruz Biotech,Texas,U.S.A.) and Nkx2.5 (1:250, goat
polyclonal antibody, R & D Systems Minneapolis,U.S.A). Cells were washed three times with
PBS and incubated for 2 h with the secondary antibody against connexin-43 (1:250, goat anti-
mouse alexa 555, Invitrogen,California , U.S.A), GATA-4 (1:250, goat anti-mouse alexa 555,
Invitrogen,California , U.S.A) and NKX2.5 (1:250, donkey anti-goat, alexa 555,
Invitrogen,California , U.S.A). TOPRO (1:1000, Invitrogen,California , U.S.A) was used for nuclei
labelling. Finally, the slides were slide-mounted in Mowiol® (Sigma-Aldrich) and visualized
using confocal microscopy.
Patch clamp
Cells were seeded at a density of 2.103 cells/cm2 in 35mm diameter culture dish 24 h prior to
the patch clamp experiments. The ionic currents were recorded on human W8B2+ CSCs in the
whole-cell configuration using an Axopatch 200A amplifier with a CV 202AU headstage
(Molecular Devices, CA, USA). Voltage clamp were generated by a personal computer
equipped with an analog-digital converter (Digidata 1322a, Molecular Devices) using pClamp
software v10 (Molecular Devices). During recordings, cells were superfused with an
extracellular solution containing (in mM) : 136 NaCl, 5 KCI, 1 MgCl2, 1.8 CaCl2, 0.3 NaH2PO4,
10 glucose , 10 HEPES, pH 7.3. Membrane ion currents were recorded via glass electrodes of
borosilicate GC150-T manufactured on a vertical twin heating puller. Pipette resistance was
et ee a d MΩ he filled ith the i t apipette solutio hi h o tai ed in mM): 20
KCl, 110 K-asp, 1 MgCl2, 10 HEPES, 2 CaCl2, 2.5 EGTA, 0.1 GTP, 5 Na2-phosphocreatine, 5 Mg-
ATP, pH 7.2 (PCa = 6.3). The currents were recorded in control (with 0.1% DMSO as vehicle) or
i pa illi e μM o ditio s. A al ses e e pe fo ed usi g Cla pfit soft a e. Cell

Résultats
179
capacitance was measured by integrating the area under the capacitive transient elicited by 5
mV depolarizing steps from holding potential of 0 mV.
Reverse transcription and polymerase chain reaction
Total RNA from W8B2+ cells was isolated using Rnable reagent (Eurobio) followed by
chloroform extraction and isopropanol precipitation. RNA integrity was evaluated by
ethydium bromide staining on a 1% agarose gel. Total RNA was quantified by assessing optical
density at 260 and 280 nm (NanoDrop ND-100 Labtech France). cDNA was synthesized using
the Pd N a do he a e p i e I it oge . μL of total RNA µg as added to μL of reaction mixture (100 mM Tris–HCl pH 8.3, 150 mM KCl, 6.25 mM MgCl2, 20 mM DTT, 2
M dNTPs I it oge a d . μg Ra do P i e Pd N I it oge . RNA e e de atu ed at 65 °C during 2 min and then added to 40 U RNAse inhibitor (RNaseOUT-Invitrogen) and 400
U M-MLV Re e se T a s iptase I it oge to μL fi al olu e. DNA as s thesized at °C fo h a d the added ith μL ste ile ate . Re ai i g e z es e e heat-
deactivated (100 °C, 2 min). After the RT p o edu e, μL of DNA ≈ g as added to μL of PCR ea tio i tu e M T is–HCl pH 8.4, 55 mM KCl, 2.2 mM MgCl2, 277.8 µM
dNTPs, 12 pmol forward and reverse primers and 1.25 U of Taq Polymerase (Invitrogen). The
PCR reaction was performed with a thermocycler (denaturation at 95 °C for 5 min, followed
by 38 cycles at 57 °C for 30 s, 72 °C for 30 s, and 95 °C for 30 s. After the last cycle, samples
were incubated at 57 °C for 2 min and at 72 °C for 10 min to ensure complete product
extension. All primers are described in table 1. GAPDH cDNA was used as housekeeping. cDNA
extracted from cardiac atrial tissue was used as a positive control and the negative control
consisted of the PCR reaction without the addition of the template. Amplified products were
separated by electrophoresis on 2% agarose gels (containing 0.01% ethidium bromide) in Tris-
Acetate-EDTA buffer and visualised using a UV Transilluminator.
Quantitative RT-qPCR
After reverse transcription the quantitative PCR reaction was carried out on a 96 well- plate,
with the sense and antisense primers allowing the amplification of GAPDH (used as reference
ge e , GATA , MEF C, ACTC , TNNT , a d β-MHC (all primers are described in table 1). Each
well contained a 25 mM primer pair (sense and anti-sense), cDNA at 100 ng/well final , as well
as Power SYBR® Green PCR reagent Master Mix (Thermo Fisher Scientific) containing AmpliTaq
Gold® DNA Polymerase. Each sample was polymerized in triplicate to best estimate the
threshold cycle of fluorescence appearance and water was used as a negative control. The
amplification reaction begins with initial denaturation of the cDNA for 10 min at 95 °C,
allowing the activation of the DNA polymerase, and is followed by 40 successive cycles
consisting of a denaturation step for 15 sec at 95 °C followed by a step of hybridization of the
primers and elongation for 1 min at 60 ° C. The results were obtained and analyzed by the
7500 Fast Real-Time PCR System (Applied Biosystems USA).
Western blotting analysis
To determine KCa1.1 protein expression, cultured W8B2+ cells were washed with cold
phosphate-buffered saline (PBS) and lysed by scraping the cells into a

Résultats
180
radioimmunoprecipitation assay buffer (in mM: Tris-HCl 50 pH:8, NaCl 150, EDTA 5, 0.05%
Igepal, 1% deoxycholic acid, 1% Triton X-100, 0.1% SDS) containing protease and phosphatase
inhibitors (Protease Inhibitor Cocktail, Sigma-Aldrich - PhosSTOP Phosphatase Inhibitor
Cocktail, Roche).
Soluble cell lysates were denatured 5 min at 37°C in 2× Laemmli sample buffer (in mM): Tris-
HCl pH . , “D“ %, gl e ol %, o ophe ol lue . %, β-mercaptoethanol 10%).
P otei sa ples a d μg , o tai ed f o ultu ed W B + cells, were separated by SDS-
PAGE using 8% polyacrylamide gels and transferred to nitrocellulose membranes. Membranes
were blocked 90 min in TBS-Tween blocking solution (in mM: Tris 20 pH : 7.6, NaCl 150 and
Tween-20 0.2%) with 5% non-fat milk at room temperature. Blots were then incubated
overnight at 4°C with primary antibodies diluted in TBST-5% nonfat dry milk. We used
polyclonal anti-rabbit KCa1.1 (1:500, Alomone Labs, Jerusalem, Israel). Vimentine was probed
by rabbit polyclonal anti-vimentine (1:500, Santa Cruz Biotechnology, Dallas, U.S.A).
Membranes were washed with TBS-Tween three times for 10 min and then incubated for 90
min at room temperature with specific anti-rabbit horseradish peroxidase-conjugated
secondary antibodies (1:5000, Interchim, Montluçon, France). Membranes were revealed
with enhanced chemiluminescence (ECL) chemiluminescent substrate (GE Healthcare, Velizy-
Villacoublay, France). The results were analyzed by using the GeneGnome Imager (SynGene
Ozyme, Montigny - le - Bretonneux, France).
Cell mobility determination
Cell migration was determined using a wound healing method to investigate the potential
effect of BKCa channels on cell mobility in human W8B2+ CSCs. The wound healing assay was
conducted when the cells grew to total confluence in 6-well plates in 0% FBS culture medium.
A sta da d ou d as eated s at hi g the ell o ola e ith a ste ile μL plasti pipette tip. After removing cell fragments by washing cell monolayer gently with PBS, the cells
were incubated at 37°C with the medium containing 0% FBS and 10 µM paxilline for 7 h. The
cells were then fixed and nuclei were labelled with DAPI. Transmission images were taken just
after the wound (time 0h) and after 7h of paxilline treatment. The images were generated
with a spinning disk confocal station (Revolution, Andor) equipped with a high precision
motorized XY stage (Marzhauser) and IQ3 software (Revolution, Andor) that allowed to
memorize positions of each sample. The number of migrated cells on the images were counted
(based on the nuclei labelling) to assess cell mobility under different conditions of treatments.
Statistical analysis Results were expressed as mean ± SEM. All statistical analysis were performed using GraphPad
Prism (La Jolla, CA, USA). The tests used for the different assays are provided in the figure
legends.
RESULTS
W8B2+ CSCs are clonogenic and proliferative Purified W8B2+ CSCs displayed spindle-shape and fibroblastic morphology during early
passages of culture (Fig. 1A). To demonstrate their stem character and their self-renewal

Résultats
181
capacity, their ability to elicit clonogenic multiplication was checked using a colony-forming
unit fibroblasts (CFU-Fs) test carried out by seeding pure W8B2+ CSCs at low density. After 10
days of growth, the cells were able to form several colonies confirming self-renewal ability
(Fig. 1B). Cells were rapidly proliferating and about 1.4 105 cells were obtained after 9 days of
culture from 2.103 cells at day 1 (Fig. 1C left). The cell increase fold in 24 hours was about 1.76
± 0.2 (Fig. 1C right).
Immunophenotype of W8B2+ CSCs Flow cytometry analysis revealed that W8B2+ CSCs highly express surface markers associated
with mesenchymal stem cells (W8B2, CD29, CD73 and CD105) (Fig. 1D). Regarding CD90 and
CD106, two others mesenchymal markers, W8B2+ CSCs showed very little expression. W8B2+
cells were negative for hematopoietic lineage markers (CD34and CD133), CD45, excluding the
possible contribution of bone marrow cells and mast cells, and CD117, a marker of another
type of cardiac stem/progenitor cells. (Fig. 1D).
At gene level, W8B2+ cells showed strong expression of early specific cardiac transcription
factors GATA4 and MEF2C but not Nkx2.5. They did express connexin 43 transcript but lacked
expression of cardiac-specific structural genes ACTC1, TNNT2, b-MHC (Fig. 1E). At the protein
level, immunostaining show that W8B2+ cells did express GATA4 and connexin-43 but no
specific labelling of Nkx2.5 was detected (Fig. 1F).
BKCa current can be recorded and identified in W8B2+ CSCs Ionic current recordings were performed on W8B2+ CSCs using patch-clamp in the whole cell
configuration. Representative traces, shown in Fig. 2, exhibit voltage–dependent currents
ith ois os illatio s du i g st o g depola izatio s et ee + a d + V Fig. Aa . Accordingly, the I/V curve obtained by voltage-ramps ranging from -110 to +110 mV, shows a
strong outward going rectification with a significant increase in noise at high depolarization
voltages (Fig. 2B). This current signature, which is characteristic for BKCa current, was present
in 8 out of 9 human W8B2 progenitor cells. When cells were superfused with 10 µM paxilline
(a BKCa selective blocker the out a d u e t as de eased a d ois u e t os illatio s were abolished (Fig. 2Ab, 2B and 2C), as expected for BKca inhibition. Similar inhibitory effect
was obtained with 5 mM TEA (not shown, n = 3).
W8B2+ CSCs express BKCa channel at both transcriptional and protein levels To investigate the molecular identity of the functional ionic channel in W8B2+ CSCs, gene
expression was examined by RT-PCR using the human gene-specific primers targeting genes
for BKCa channel (Table 1). Fig. 3A shows the images of RT-PCR products corresponding to
gene expression of ion channels. A remarkable expression of KCNMA1 gene (coding for BKCa)
was detected. Atrial tissue was used as positive control and GAPDH as reference. At the
protein level, western blot analysis demonstrated an abundant expression of the ionic channel
responsible for BKCa current using the specific antibody anti-KCa1.1 (Fig.3B). Atrial tissue was
used as positive control and vimentin as reference.
Inhibition of BKCa channel by paxilline decreases cell proliferation and cell cycling
progression in a concentration-dependent manner

Résultats
182
To evaluate the role of BKCa channel in cell proliferation, paxilline was used as a specific
blocker of this channel at different concentrations (1, 3 and 10 µM). Whereas paxilline 1 µM
had no effect, the cell proliferation was progressively inhibited by the application of paxilline
at 3 µM and 10 µM until day 9 of culture (Fig. 4A). The inhibitory effect of paxilline 10 µM was
stronger than that of 3 µM, an effect which reached statistical significance from day 4. The
paxilline blocking effect of BKCa on cell cycling progression was evaluated by flow cytometry
analysis in human W8B2+ CSCs. Fig 4B illustrates an example of the mean percentage values
of li g phases i o t ol ‰ DM“O as ehi le o i ells t eated ith pa illi e a d µM) at day 5, showing a shift from cells in S and G2 phases into G1 phase. Taken together,
these experiments show that 10 µM paxilline significantly increased the fraction of G1
population from 56.9 % to 72.4 % (p<0.01) and from 59.8 % to 71.4 % (p<0.01; Fig 4C) at day
5 and 6, respectively. In contrary, fraction of S+G2 population was significantly decreased from
39.8% to 22.8% (p<0.001) at day 5 and from 37.6 % to 24.4 % (p<0.05) at day 6 after treatment
with 10 µM of paxilline (Fig 4C). At 3 µM, the effect of paxilline was lower in amplitude but
reached statistical significance versus control only for the S+G2 population. These results
suggest that BKCa participates in G1/S cell cycling progression in human W8B2+ CSCs.
Inhibition of BKCa channel by paxilline decreases self-renewal in human W8B2+ CSCs
To investigate further the role of BKCa channel, paxilline was used at different concentrations
(1, 3 and 10 µM) to block the activity of BKCa channel on self-renewal of W8B2+ CSCs. Paxilline
at 1, 3 and 10 µM significantly reduced the number of colony-forming unit W8B2+ CSCs after
ten days of culture (Fig. 5). This decrease reached about 15 % (p<0.05), 34 % (p<0.001) and
50% (p<0.001) with 1 ,3 and 10 µM of paxilline, respectively. This result suggests that BKCa
channel is important in the stem cells self-renewal process by which stem cells divide to
perpetuate their pool throughout life and maintain their undifferentiated state.
Inhibition of BKCa channel by paxilline did not affect cell migration To examine whether BKCa channel can regulate cell migration in human W8B2+ CSCs, wound
healing assay was conducted in cells treated with different concentrations of paxilline. Figure
A sho s the ou d heali g i ages i o t ol ‰ DM“O as ehi le a d pa illi e o ditio s (1, 3 and 10 µM). For each condition, images were taken just after the wound (time 0h) and
after 7 h of migration. To estimate the cell migration, nuclei of cells were fixed and colored
with DAPI (green) after 7 hours as illustrated (fig. 6A, right). Fig. 6B illustrates the ratio of
migrated cells into the acellular area after 7 h in the different treatments. Blockade of BKCa
channels with paxilline did not impact cell migration compared to the control (ns) suggesting
that BKCa channel may not be involved in cell migration regulation of W8B2+ CSCs.
DISCUSSION
In the present study, human W8B2+ cardiac progenitor cells isolated from right atrial
specimens highly expressed surface markers associated with mesenchymal stem cells (W8B2,
CD29, CD73, and CD105) but not hematopoietic markers. Moreover, human W8B2+ CSCs
exhibited an important clonogenic efficiency. These human cardiac stem cell properties
assessed in our experimental conditions are consistent with those reported previously [1].

Résultats
183
In stem cells, the presence of BKCa current has been reported in some cell types like human
embryonic stem cells [7], human mesenchymal stem cells from different origins [8-10], cancer
stem cells [11] and human cardiac progenitor cells [12]. In the present study,
electrophysiological properties of W8B2+ CSCs, paxilline sensitivity, RT-PCR and Western
immunoblot analysis, all converge on the idea that BKCa current exists in these cells and that
the ionic channel responsible for this current is encoded by KCa1.1 gene.
In non-excitable cells, high-conductance calcium-activated potassium channel has been
reported to be involved in the regulation on cell proliferation. For example, blocking BKCa by
the selective channel blocker inhibits cell proliferation in human endothelial cells [13-14], in
mouse ES cells [7] and in human preadipocytes [15]. In human cardiac fibroblast, BKCa
inhibition by paxilline results in a cell proliferation reduction [16]. Other studies report that
inhibition of BKCa has little effect on cell proliferation in human bronchial smooth muscle cells
[17], or MCF-7 cells [18]. Recently it was reported that blocking of KCa1.1 in normal myoblasts
induced an increase in proliferation [6], suggesting that regulation of cell proliferation by BKCa
channels may be cell-type dependent. In the present study, we report that inhibition of BKCa
by paxilline significantly reduces human W8B2+ CSCs proliferation as well as the proportion of
cells in the G0/G1 phase of the cell cycle. This result is in agreement with that obtained on
human cardiac c-kit+ progenitor cells [19]. Interestingly, it has been reported that membrane
potential hyperpolarized during the cell cycle and that potassium conductance increased
during progress from G1 to S phase in rat mesenchymal stem cells from bone marrow [19].
Accordingly, the effects that we have obtained by inhibiting BKCa could be due to changes in
the membrane potential during the cellular cycle.
The present study also shows that blocking BKCa channels with paxilline decreased the
number of colony-forming unit fibroblasts (CFU-Fs) suggesting that BKCa channel could be
involved in the process of self-renewal and differentiation of W8B2+ CSCs. Self-renewal
consists in cell division with maintenance of the undifferentiated state. This requires cell cycle
control and often maintenance of multipotency or pluripotency, depending on the stem cell
type [21]. We show that mesenchymal stem cells are clonogenic when plated at low densities.
The number and the morphology of the colonies formed by a fixed number of input cells
provide preliminary information about the ability of progenitors to differentiate and
proliferate.
Ca2+-activated potassium channels, including BKCa, are involved in the regulation of
differentiation of several cell types. Similarly to what was reported on cell proliferation (see
above), BKCa modulation can enhance or inhibit the differentiation process depending on the
cell type. It has been shown for example that inhibition of potassium channels influences the
differentiation of endothelial progenitor cells (EPCs) from human peripheral blood [22]. These
cells strongly express BKCa channel, which inhibition disrupts their differentiation into mature
endothelial cells. On another hand, inhibition of BKCa with paxilline negatively regulates the
differentiation of human bone marrow-derived mesenchymal stem cells (MSCs) suggesting
the essential role of BKCa in maintaining bone marrow physiological function [23]. Further
study is required to find out how BKCa channel may contribute to cellular differentiation in
human W8B2+ CSCs.

Résultats
184
In addition to differentiation and proliferation, previous studies have demonstrated that the
Ca2+-activated potassium channel is closely involved in cell migration but its implication is
again cell-type specific. For example, KCa1.1 channels were shown to be required for
migration in gloma cells, but not in microglia cells [24-25]. Interestingly, blockade of BKCa by
paxilline inhibited cell migration in human cardiac c-kit+ progenitor cells [19]. In the present
study, inhibition of BKCa did not affect W8B2+ CSCs migration suggesting that the regulation
of mobility differs between the two types of human cardiac progenitor cells. This discrepancy
could be due to the various calcium signaling parameters such as amplitude and frequencies
of calcium oscillations, which regulate directly and/or indirectly BKCa channel (preliminary
data not shown). An estimation of calcium subcellular domains within cells is required to
investigate further this hypothesis, for example using green fluorescence protein (GFP)-linked
calcium calmoduline (CaM) probes [25].
In conclusion, we demonstrate that BKCa channel is likely a key regulator of proliferation and
self-renewal of human cardiac stem cells and provides a basis for a better understanding of
the physiology of these cells. Further investigations are required to open new avenues for
cellular therapeutic strategies since W8B2+ cells have been described as having powerful
regenerative and functional effects when injected into the hearts of rats after attack [1].
Figure Legends
Figure 1. Growth and phenotype characteristics of W8B2+ CSCs. (A) Cell morphology at
passage 2 of isolated W8B2+ cells. (B) colony-forming unit fibroblasts (CFU-Fs) assay after 10
days post-plating. (C) Proliferation assay of W8B2+ cells cultured for 9 days. (n=3). D: day. (D)
Percentage of positive cells for the indicated surface markers in W8B2+ CSCs determined by
flow cytometry. (n=3). (E) Gene expression of cardiac markers in W8B2+ CSCs determined by
RT-qPCR (n=3). Ct: cycle threshold. Bar graph represents the 1/Ct ratio for each gene. A value
of /Ct hi h is ≤ . Ct= o espo ds to a egligible gene expression. (F)
Immunostaining of cardiac markers in W8B2+ CSCs. Left: GATA4; Center: Nkx2.5; Right:
connexin 43. Nuclei are marked in blue with TOPRO staining. (n=3). Scale bare = 50 µm.
Figure 2. Inhibition of membrane current by BKCa ion channel blocker in W8B2+ CSCs. (A)
Voltage dependent current recorded in whole cell configuration during 2 s pulses at potentials
from -110 to +90 mV (10 mV increment) with a holding potential of -80 mV in the absence (Aa)
and in the presence (Ab) of 10 µM paxilline (arrow indicates 0 current level). (B) I/V curves
obtained during a ramp protocol stimulation from -110 to 110 mV (duration 2 sec) in the
absence and in the presence of 10 µM paxilline. In A and B, note the presence of large noisy
oscillations (typical signature of BKCa channels) on the current traces obtained for high
positive potentials in control condition. (C) Current density estimated at +110 mV in control
and in the presence of paxilline (10 µM) (P<0.001, n= 5).
Figure 3. Molecular scanning of functional channel in W8B2+ CSCs. (A) Characterization of the
expression of KCNMA1 mRNA in cultured W8B2+ cells by RT-PCR. Expression of KCNMA1
mRNA was analyzed in cultured W8B2+ Cells using RT-PCR primers specific. Amplification
products were resolved on 2% agarose gels alongside a X174 RF DNA/HaeIII fragments

Résultats
185
ladder marker. GAPDH cDNA was used as housekeeping. cDNA extracted from cardiac atrial
tissue was used as a positive control and the negative control consisted of the PCR reaction
without the addition of the template. Fragment of the expected molecular size indicated the
expression of KCNMA1 in cultured W8B2+ Cells. The top and the down arrows indicate
amplicons with 310 and 271-281 bp, respectively. (B) Characterization of the expression of
KCa1.1 protein in cultured W8B2+ cells. Western blotting analysis were performed from cell
l sates o tai i g o μg of total p otei s. As sho , W B + cells displayed a band with
a molecular mass of 100 kDa approximately, as expected for KCa1.1 protein channel.
Figure 4. Effect of BKCa inhibition by paxilline on cell proliferation and cell cycling in W8B2+
CSCs. (A) Cell proliferation assay expressed as percentage of cell number obtained with vehicle
(control) or paxilline at different concentrations. (n=3,*p<0.05, **p<0.01, ***p<0.001 vs.
control, Two-Way ANOVA with Bonferroni post-test).
(B) Examples of flow cytometry graphs with percentage of cycling phases in cells treated with
vehicle (control, left) or paxilline (3 (middle) and 10 µM (right)) at day 5 post treatment. (C)
Distribution of cell cycle phases in cells treated with vehicle (control) or paxilline at 3 and 10
µM after 5 and 6 days post-plating. (n=3,*p<0.05, **p<0.01, ***p<0.001 vs. control, Two-Way
ANOVA with Bonferroni post-test).
Figure 5. Effect of BKCa inhibition by paxilline on self-renewal of W8B2+ CSCs. (A) Images of
colony-forming unit W8B2+ CSCs in control or paxilline conditions after 10 days post-plating.
(B) Number of CFU-Fs in cells treated with vehicle (control) or paxilline at different
concentrations for 10 days post-plating. (n=3,*p<0.05, ***p<0.001 vs. control, Nonparametric
One-Way ANOVA).
Figure 6. Effect of BKCa inhibition by paxilline on cell migration in W8B2+ CSCs. Cell images
with a cellular wound at time 0 (left) and after 7 hours (right) in cells treated with vehicle
(control) or paxilline at different concentrations. Green dots represent nuclei of cells fixed and
colored with DAPI after 7 hours. (B) % of migrated cells in W8B2+ CSCs treated with vehicle
(control) or paxilline at different concentrations. (n=3, Nonparametric One-Way ANOVA).
Scale bare = 100 µm.
ACKNOWLEDGMENT
This work has benefited from the facilities and expertise of ImageUP (University of Poitiers)
and was supported by AFM.
References
1. Zhang, Y., Sivakumaran, P., Newcomb, A.E., Hernandez, D., Harris, N., Khanabdali, R., Liu, G.-S., Kelly, D.J., Pébay, A., Hewitt, A.W., et al. (2015). Cardiac Repair With a Novel Population of Mesenchymal Stem Cells Resident in the Human Heart. Stem Cells 33, 3100–3113. 2.Toro, L., Li, M., Zhang, Z., Singh, H., Wu, Y., and Stefani, E. (2014). MaxiK channel and cell signalling. Pflugers Arch. 466, 875–886. 3.Nelson, M.T., and Quayle, J.M. (1995). Physiological roles and properties of potassium channels in
arterial smooth muscle. Am. J. Physiol. 268, C799-822.
4.Waldron, G.J., and Cole, W.C. (1999). Activation of vascular smooth muscle K+ channels by
endothelium-derived relaxing factors. Clin. Exp. Pharmacol. Physiol. 26, 180–184.

Résultats
186
5.Ko, E.A., Han, J., Jung, I.D., and Park, W.S. (2008). Physiological roles of K+ channels in vascular
smooth muscle cells. J Smooth Muscle Res 44, 65–81.
6.Tajhya, R.B., Hu, X., Tanner, M.R., Huq, R., Kongchan, N., Neilson, J.R., Rodney, G.G., Horrigan, F.T., Timchenko, L.T., and Beeton, C. (2016). Functional KCa1.1 channels are crucial for regulating the proliferation, migration and differentiation of human primary skeletal myoblasts. Cell Death Dis 7, e2426. 7.Wang, K., Xue, T., Tsang, S.-Y., Van Huizen, R., Wong, C.W., Lai, K.W., Ye, Z., Cheng, L., Au, K.W., Zhang, J., et al. (2005). Electrophysiological properties of pluripotent human and mouse embryonic stem cells. Stem Cells 23, 1526–1534. 8.Heubach, J.F., Graf, E.M., Leutheuser, J., Bock, M., Balana, B., Zahanich, I., Christ, T., Boxberger, S.,
Wettwer, E., and Ravens, U. (2004). Electrophysiological properties of human mesenchymal stem cells.
J. Physiol. (Lond.) 554, 659–672.
9.Li, G.-R., Sun, H., Deng, X., and Lau, C.-P. (2005). Characterization of ionic currents in human
mesenchymal stem cells from bone marrow. Stem Cells 23, 371–382.
10.Park, K.S., Jung, K.H., Kim, S.H., Kim, K.S., Choi, M.R., Kim, Y., and Chai, Y.G. (2007). Functional expression of ion channels in mesenchymal stem cells derived from umbilical cord vein. Stem Cells 25, 2044–2052. 11.Park, J.H., Park, S.J., Chung, M.K., Jung, K.H., Choi, M.R., Kim, Y., Chai, Y.G., Kim, S.J., and Park, K.S. (2010). High expression of large-conductance Ca2+-activated K+ channel in the CD133+ subpopulation of SH-SY5Y neuroblastoma cells. Biochem. Biophys. Res. Commun. 396, 637–642. 12.Zhang, Y.-Y., Li, G., Che, H., Sun, H.-Y., Li, X., Au, W.-K., Xiao, G.-S., Wang, Y., and Li, G.-R. (2014). Characterization of functional ion channels in human cardiac c-kit+ progenitor cells. Basic Res Cardiol 109, 407. 13.Wiecha, J., Münz, B., Wu, Y., Noll, T., Tillmanns, H., and Waldecker, B. (1998). Blockade of Ca2+-Activated K+ Channels Inhibits Proliferation of Human Endothelial Cells Induced by Basic Fibroblast Growth Factor. JVR 35, 363–371. 14.Kuhlmann, C.R.W., Most, A.K., Li, F., Münz, B.M., Schaefer, C.A., Walther, S., Raedle-Hurst, T.,
Waldecker, B., Piper, H.M., Tillmanns, H., and Wiecha J (2005). Endothelin-1-induced proliferation of
human endothelial cells depends on activation of K+ channels and Ca+ influx. Acta Physiol. Scand. 183,
161–169.
15.Hu, H., He, M.-L., Tao, R., Sun, H.-Y., Hu, R., Zang, W.-J., Yuan, B.-X., Lau, C.-P., Tse, H.-F., and Li, G.-R. (2009). Characterization of ion channels in human preadipocytes. J. Cell. Physiol. 218, 427–435. 16.He, M.-L., Liu, W.-J., Sun, H.-Y., Wu, W., Liu, J., Tse, H.-F., Lau, C.-P., and Li, G.-R. (2011). Effects of
ion channels on proliferation in cultured human cardiac fibroblasts. Journal of Molecular and Cellular
Cardiology 198–206.
17.Shepherd, M.C., Duffy, S.M., Harris, T., Cruse, G., Schuliga, M., Brightling, C.E., Neylon, C.B.,
Bradding, P., and Stewart, A.G. (2007). KCa3.1 Ca2+ activated K+ channels regulate human airway
smooth muscle proliferation. Am. J. Respir. Cell Mol. Biol. 37, 525–531.
18.Ouadid-Ahidouch, H., Roudbaraki, M., Delcourt, P., Ahidouch, A., Joury, N., and Prevarskaya, N. (2004). Functional and molecular identification of intermediate-conductance Ca(2+)-activated K(+) channels in breast cancer cells: association with cell cycle progression. Am. J. Physiol., Cell Physiol. 287, C125-134. 19.Zhang, Y.-Y., Li, G., Che, H., Sun, H.-Y., Xiao, G.-S., Wang, Y., and Li, G.-R. (2015). Effects of BKCa and
Kir2.1 Channels on Cell Cycling Progression and Migration in Human Cardiac c-kit+ Progenitor Cells.
PLoS ONE 10, e0138581.
20.Deng, X.L., Lau, C.P., Lai, K., Cheung, K.F., Lau, G.K., and Li, G.R. (2007). Cell cycle-dependent
expression of potassium channels and cell proliferation in rat mesenchymal stem cells from bone
marrow. Cell Proliferation 40, 656–670.
21.He, S., Nakada, D., and Morrison, S.J. (2009). Mechanisms of stem cell self-renewal. Annu. Rev. Cell
Dev. Biol. 25, 377–406.

Résultats
187
22.Ye, G., Guan, H., Karush, J., Wang, F., Xu, X., Mao, H., Huang, X., Yang, X., Peng, P., Ba, Y., et al. (2014). Effects of Ca2+-activated potassium and inward rectifier potassium channel on the differentiation of endothelial progenitor cells from human peripheral blood. Mol Biol Rep 41, 3413–3423. 23.Zhang, Y.-Y., Yue, J., Che, H., Sun, H.-Y., Tse, H.-F., and Li, G.-R. (2014). BKCa and hEag1 channels regulate cell proliferation and differentiation in human bone marrow-derived mesenchymal stem cells. J. Cell. Physiol. 229, 202–212. 24.Schilling, T., Stock, C., Schwab, A., and Eder, C. (2004). Functional importance of Ca2+-activated K+ channels for lysophosphatidic acid-induced microglial migration. Eur. J. Neurosci. 19, 1469–1474. 25.Weaver, A.K., Bomben, V.C., and Sontheimer, H. (2006). Expression and function of calcium-
activated potassium channels in human glioma cells. Glia 54, 223–233.
26.Li, C.J., Heim, R., Lu, P., Pu, Y., Tsien, R.Y., and Chang, D.C. (1999). Dynamic redistribution of calmodulin in HeLa cells during cell division as revealed by a GFP-calmodulin fusion protein technique. J. Cell. Sci. 112 ( Pt 10), 1567–1577.
Table 1: Human gene-specific primers for RT-PCR and RT-qPCR
Gene name fo a d ’ ’ e e se ’ ’
GAPDH AAGGTCATCCCTGAGCTGAA TGACAAAGTGGTCGTTGAGG
MEF2C AAGAAGGAGATTTGTTTGGAGG CTTTGTAAATGTCACCTGTCTG
GATA4 CCCAATCTCGATATGTTTGAC CGTTCATCTTGTGGTAGAGG
TNNT2 CTGCTGTTCTGAGGGAGAGC CACCAAGTTGGGATGAACG
ACTC1 CCTTCATTGGTATGGAATCTG AATCTTCATGGTGCTTAGGAG
β-MHC GAGCTACGGGTTCCCCTCT GGAAGCTCTGCTACCCCTTTT
NKX2.5 AAAGAAAGGGTGTGAACTGC CGCACAGTAATGGTAAGGGA
KCNMA1 CCTGCAGCGAAGTATCATCA ACTCGAAGTGAAGCTGCCAT
Connexin43 GGGCGTTAAGGATCGGGTTAAGG GTTGGTGAGGAGCAGCCATTGAA
Résultats
188
Figure 1
Résultats
189
Figure 2
Figure 3
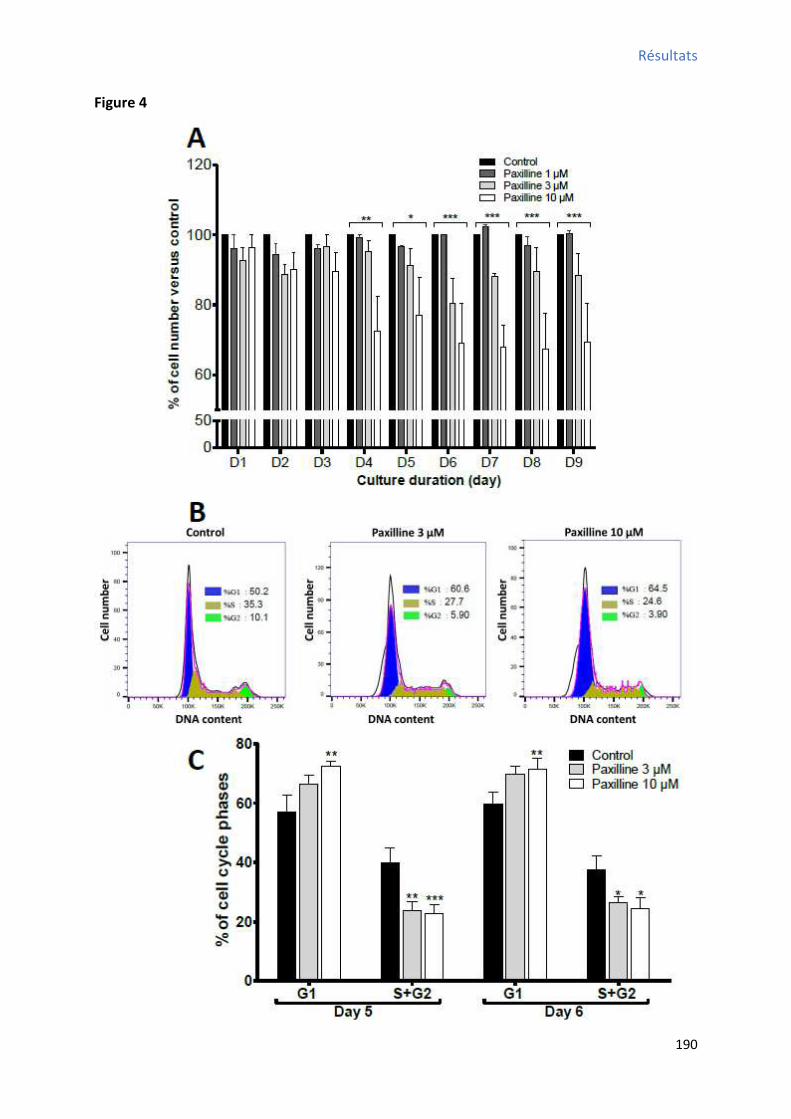
Résultats
190
Figure 4
Résultats
191
Figure 5
Figure 6
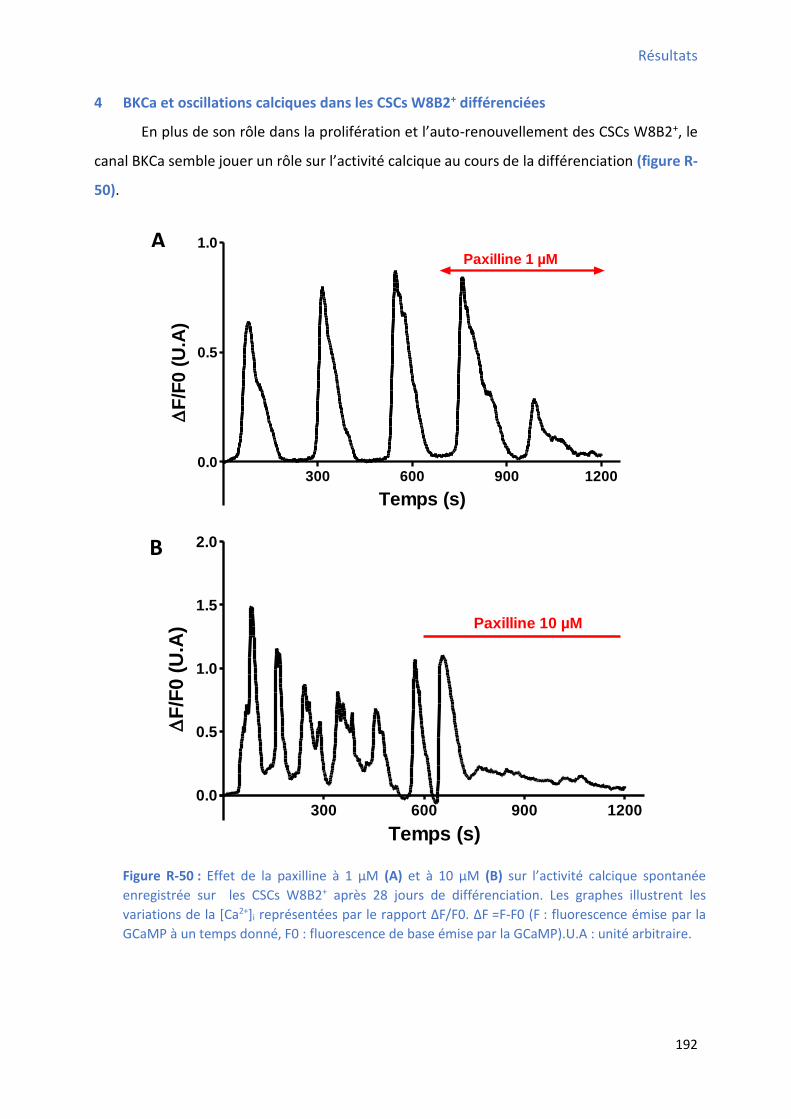
Résultats
192
4 BKCa et oscillations calciques dans les CSCs W8B2+ différenciées
En plus de so ôle da s la p olif atio et l’auto-renouvellement des CSCs W8B2+, le
canal BKCa semble joue u ôle su l’a ti it al i ue au ou s de la différenciation (figure R-
50).
300 600 900 12000.0
0.5
1.0Paxilline 1 µM
Temps (s)
F/F0
(U.A
)
300 600 900 12000.0
0.5
1.0
1.5
2.0
Paxilline 10 µM
Temps (s)
F/F0
(U.A
)
Figure R-50 : Effet de la paxilline à 1 µM (A) et à 10 µM (B) su l’a ti it al i ue spo ta e enregistrée sur les CSCs W8B2+ après 28 jours de différenciation. Les graphes illustrent les
variations de la [Ca2+]i ep se t es pa le appo t ΔF/F . ΔF =F-F0 (F : fluorescence émise par la
GCaMP à un temps donné, F0 : fluorescence de base émise par la GCaMP).U.A : unité arbitraire.
B
A

Résultats
193
La perfusion de 1 µM de paxilline (inhibiteur spécifique du canal BKCa) à t = 700 s
(figure R-50A), ’i duit au u ha ge e t su l’a plitude (figure R-51A), la fréquence (figure
R-51B), l’ai e (figure R-51C) , la durée des oscillations calciques (figure R-51D), le TTP (figure
R-52A), le TTR (figure R-52B), la pente 1 (figure R-52C) , la pente 2 (figure R-52D) et le
pou e tage du te ps d’a ti it (figure R-52E).
Cependant, à 10 µM (figure R-50B) la pa illi e di i ue sig ifi ati e e t l’a plitude
(diminution de 83,3%, figure R-51A), la fréquence (diminution de 83,3%, figure R-51B , l’ai e
(diminution de 81,3%, figure R-51C) et la durée (diminution de 67,7%, figure R-51D) des
oscillations calciques par rapport au contrôle.
De plus, la figure R-52 montre que le traitement la paxilline à 10 µM, provoque une
diminution significative (64.1%) du TTR (figure R-52B), de la pente 1 (81,4%) (figure R-52C), et
du pou e tage du te ps d’a ti it % (figure R-52E) mais ne modifie pas le TTP (figure R-
50A) et la pente 2 (figure R-52D).
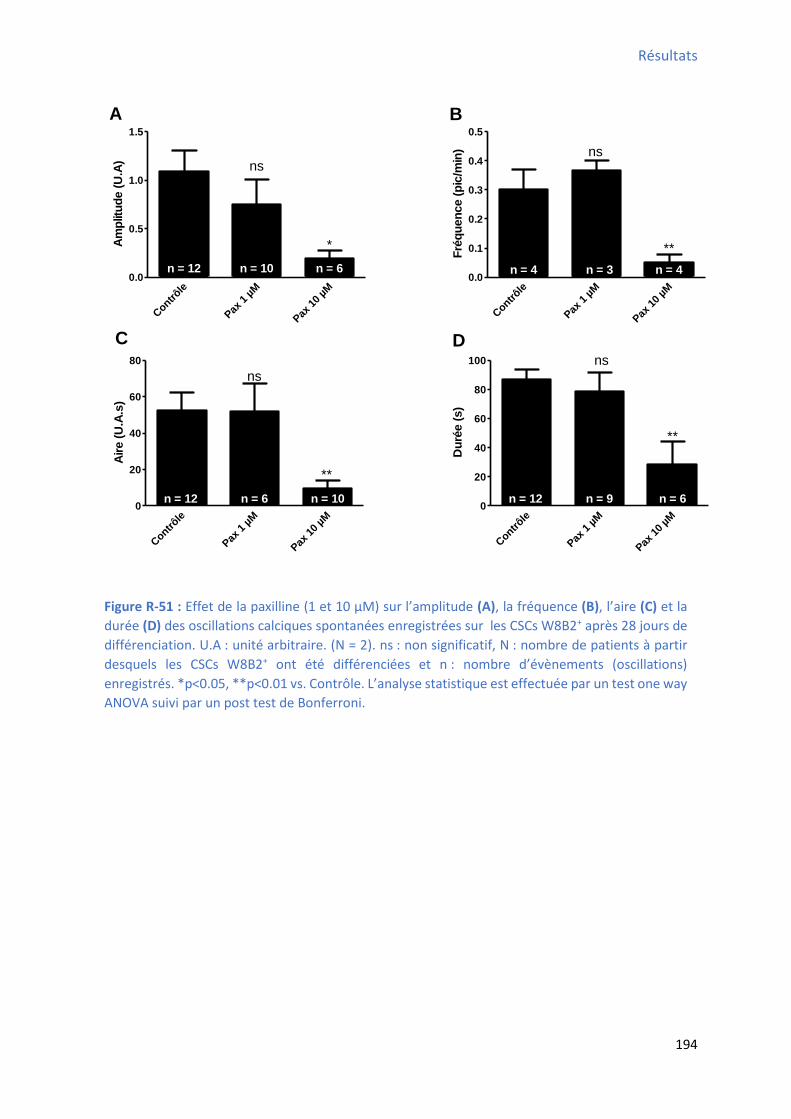
Résultats
194
Contrô
le
Pax 1
µM
Pax 10
µM0.0
0.5
1.0
1.5
n = 12
ns
*
n = 10 n = 6
Am
plitu
de (U
.A)
Contrô
le
Pax 1
µM
Pax 10
µM0.0
0.1
0.2
0.3
0.4
0.5
ns
ns
**
n = 4 n = 3 n = 4
Fréq
uenc
e (p
ic/m
in)
Contrô
le
Pax 1
µM
Pax 10
µM0
20
40
60
80
**
ns
n = 12 n = 6 n = 10
Aire
(U.A
.s)
Contrô
le
Pax 1
µM
Pax 10
µM0
20
40
60
80
100 ns
**
n = 12 n = 9 n = 6
Dur
ée (s
)
A B
C D
Figure R-51 : Effet de la pa illi e et µM su l’a plitude (A), la fréquence (B), l’ai e (C) et la
durée (D) des oscillations calciques spontanées enregistrées sur les CSCs W8B2+ après 28 jours de
différenciation. U.A : unité arbitraire. (N = 2). ns : non significatif, N : nombre de patients à partir
desquels les CSCs W8B2+ ont été différenciées et n : o e d’ e e ts os illatio s e egist s. *p< . , **p< . s. Co t ôle. L’a al se statisti ue est effe tu e pa u test o e a ANOVA suivi par un post test de Bonferroni.
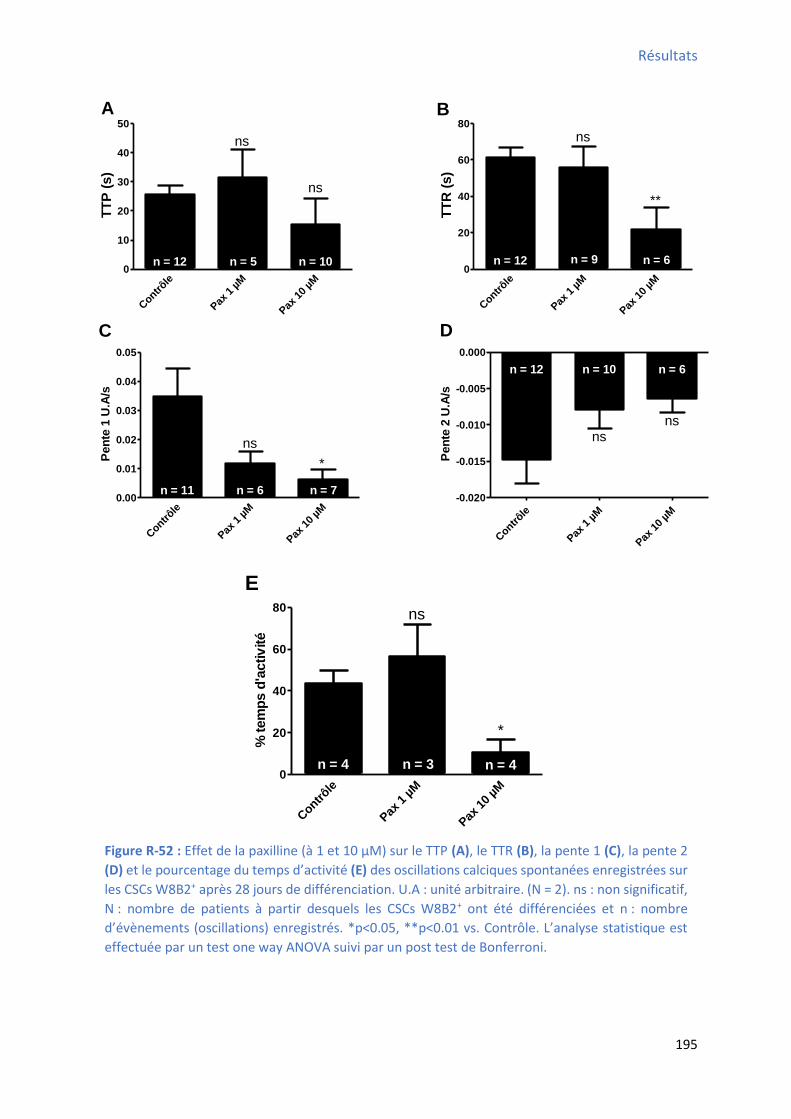
Résultats
195
Contrô
le
Pax 1
µM
Pax 10
µM0
20
40
60
80
ns
*
ns
n = 4 n = 3 n = 4
% te
mps
d'a
ctiv
ité
E
Contrô
le
Pax 1
µM
Pax 10
µM0
10
20
30
40
50
ns
ns
n = 12 n = 5 n = 10
TT
P (
s)
Contrô
le
Pax 1
µM
Pax 10
µM0
20
40
60
80ns
**
n = 12 n = 9 n = 6
TT
R (
s)
Contrô
le
Pax 1
µM
Pax 10
µM0.00
0.01
0.02
0.03
0.04
0.05
ns
*
n = 11 n = 6 n = 7
Pen
te 1
U.A
/s
Contrô
le
Pax 1
µM
Pax 10
µM
-0.020
-0.015
-0.010
-0.005
0.000
nsns
n = 12 n = 10 n = 6
Pen
te 2
U.A
/s
A B
C D
Figure R-52 : Effet de la paxilline (à 1 et 10 µM) sur le TTP (A), le TTR (B), la pente 1 (C), la pente 2
(D) et le pou e tage du te ps d’a ti it (E) des oscillations calciques spontanées enregistrées sur
les CSCs W8B2+ après 28 jours de différenciation. U.A : unité arbitraire. (N = 2). ns : non significatif,
N : nombre de patients à partir desquels les CSCs W8B2+ ont été différenciées et n : nombre
d’ e e ts os illatio s e egist s. *p< . , **p< . s. Co t ôle. L’a al se statisti ue est effectuée par un test one way ANOVA suivi par un post test de Bonferroni.
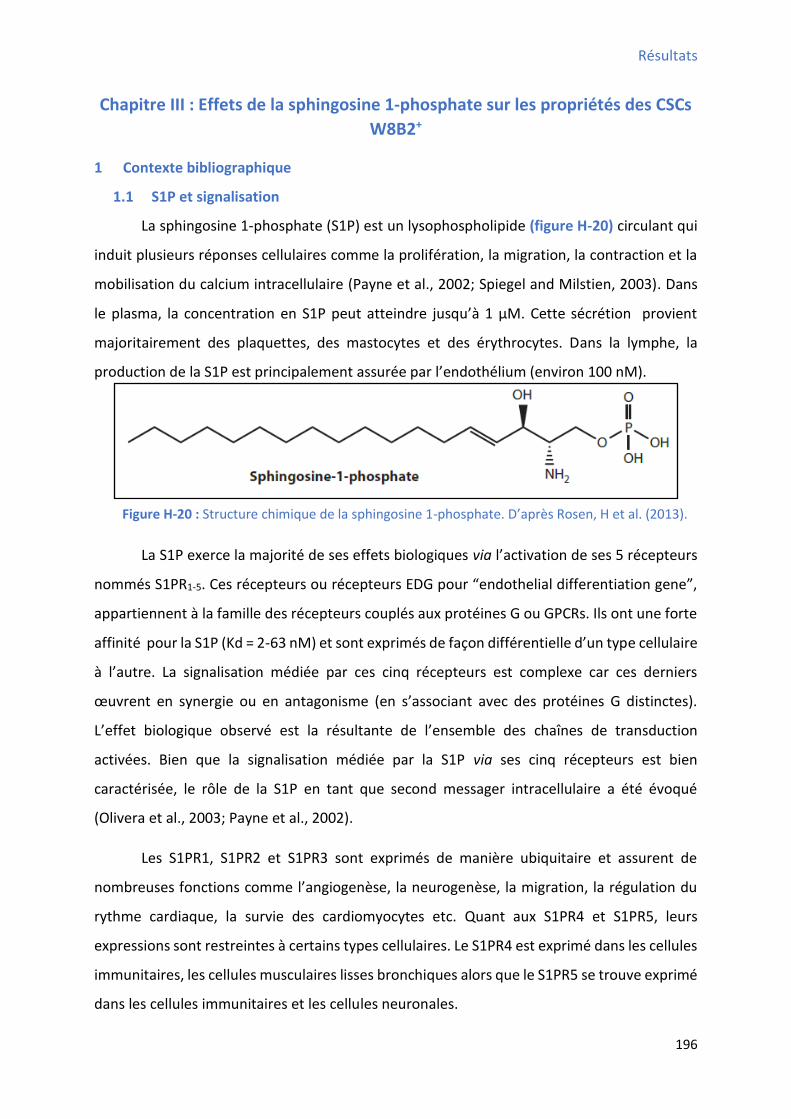
Résultats
196
Chapitre III : Effets de la sphingosine 1-phosphate sur les propriétés des CSCs
W8B2+
1 Contexte bibliographique
1.1 S1P et signalisation
La sphingosine 1-phosphate (S1P) est un lysophospholipide (figure H-20) circulant qui
induit plusieurs réponses cellulaires comme la prolifération, la migration, la contraction et la
mobilisation du calcium intracellulaire (Payne et al., 2002; Spiegel and Milstien, 2003). Dans
le plas a, la o e t atio e “ P peut attei d e jus u’à µM. Cette s tio p o ie t
majoritairement des plaquettes, des mastocytes et des érythrocytes. Dans la lymphe, la
p odu tio de la “ P est p i ipale e t assu e pa l’e doth liu e i o M .
La S1P exerce la majorité de ses effets biologiques via l’a ti atio de ses epteu s
nommés S1PR1-5. Ces epteu s ou epteu s EDG pou e dothelial diffe e tiatio ge e ,
appartiennent à la famille des récepteurs couplés aux protéines G ou GPCRs. Ils ont une forte
affinité pour la S1P (Kd = 2- M et so t e p i s de faço diff e tielle d’u t pe cellulaire
à l’aut e. La sig alisatio di e pa es i epteu s est o ple e a es de ie s
œu e t e s e gie ou e a tago is e e s’asso ia t a e des p ot i es G disti tes .
L’effet iologi ue o se est la sulta te de l’e se le des chaînes de transduction
activées. Bien que la signalisation médiée par la S1P via ses cinq récepteurs est bien
caractérisée, le rôle de la S1P en tant que second messager intracellulaire a été évoqué
(Olivera et al., 2003; Payne et al., 2002).
Les S1PR1, S1PR2 et S1PR3 sont exprimés de manière ubiquitaire et assurent de
o euses fo tio s o e l’a gioge se, la eu oge se, la ig atio , la gulatio du
rythme cardiaque, la survie des cardiomyocytes etc. Quant aux S1PR4 et S1PR5, leurs
expressions sont restreintes à certains types cellulaires. Le S1PR4 est exprimé dans les cellules
immunitaires, les cellules musculaires lisses bronchiques alors que le S1PR5 se trouve exprimé
dans les cellules immunitaires et les cellules neuronales.
Figure H-20 : Structure chimique de la sphingosine 1-phosphate. D’ap s Rose , H et al. .

Résultats
197
1.2 S1P et tissu cardiaque
Pa i les i epteu s à la “ P, le œu e e p i e t ois ; le S1PR1, le S1PR2 et le
S1PR3. De par leur expression et localisation ces récepteurs sont impliqués dans différentes
fonctions cardiaques. En effet, dans les cardiomyocytes, on retrouve les récepteurs S1PR1,
S1PR2 et S1PR3 avec une prédominance du S1PR1 (Means et al., 2008; Zhang et al., 2007).
L’a ti atio de es epteu s da s les a dio o tes est asso i e au th e et contractilité
cardiaque (Budde et al., 2002; Sanna et al., 2004), à l’i du tio d’h pe t ophie (Robert et al.,
2001), à la p ote tio de l’is h ie (Jin et al., 2004; Zhang et al., 2007) et à la mobilisation du
calcium intracellulaire (Egom et al., 2016).
Les cellules musculaires lisses, quant à elles, expriment les S1PR1, S1PR2 et S1PR3 avec
une prédominance des récepteurs S1PR2 et S1PR3 (Alewijnse et al., 2004; Kluk and Hla, 2001).
L’a ti ation de ces récepteurs dans le muscle lisse est associée à la vasoconstriction (Ryu et
al., 2002) et la migration (Alewijnse et al., 2004).
Dans les cellules endothéliales vasculaires, on retrouve majoritairement le S1PR1 et
des niveaux faibles d’e p essio de S1PR2 et de “ PR . L’a ti atio de es p teu s est
asso i e à l’a gioge se (Allende et al., 2003), à la migration (Wang et al., 1999a), l’adh e e
(Windh et al., 1999) et l’i t g it de l’e doth liu (Brindley et al., 2002; Garcia et al., 2001).
Les fibroblastes cardiaques expriment majoritairement le S1PR3 et à des niveaux
faibles des récepteurs S1PR1 et S1PR2 (Landeen et al., 2008). La signalisation par ces
récepteurs régule la prolifération des fibroblastes cardiaques (Benamer et al., 2011; Gellings
Lowe et al., 2009).
En plus des rôles physiologiques cardiaques, il a été montré que les quantités de S1P
circulante augmentent chez les patie ts attei ts d’i suffisa e a dia ue (Polzin et al., 2017),
d’i fa tus du o a de aigu (Klyachkin et al., 2015) et de fibrose cardiaque (Takuwa et al.,
2013). La “ P est aussi i pli u e da s d’aut es pathologies de t pe i fla atoi es,
immunitaires, métaboliques ou encore cancéreuses (Maceyka et al., 2012).

Résultats
198
1.3 S1P et cellules souches
Chez la souris, il a été montré que les cellules souches mésenchymateuse (CSMs) de la
moelle osseuse secrétaient la S1P et que cette dernière stimulait la prolifération des cellules
satellites du muscle squelettique (Sassoli et al., 2014).
D’aut es tudes i di ue t ue la “ P pou ait sti ule la p olif atio et la ig atio
des cellules satellites(Calise et al., 2012; Donati et al., 2007) et des cellules souches
mésenchymateuse de la moelle osseuse (Lu et al., 2015; Price et al., 2015). La S1P participe
également à la régulation de la différenciation des CSMs murines de la moelle osseuse. En
effet, la S1P améliorait la différenciation des CSMs murines en adipocytes, en ostéoblastes et
en cellules endothéliales vasculaires (Lu et al., 2015; Price et al., 2015).
Chez l’Ho e, la “ P joue u ôle i po ta t da s la gulatio de la p olifération, de
la différenciation et la migration. Certaines études montrent que la S1P intervient dans la
survie cellulaire des CSEs humaines (Inniss and Moore, 2006) et module la migration des CSMs
humaines de la moelle osseuse (Kong et al., 2014). De plus, la S1P oriente la différenciation
des CSMs du tissu adipeux vers un phénotype musculaire lisse (Nincheri et al., 2009) et
favorise la différenciation des CSMs humains du cordon ombilical vers un phénotype
cardiaque (Zhao et al., 2011).
2 Co te te de l’ tude
La “ P gule l’a ti it a dio-vasculaire (via ses récepteurs) par la médiation de voies
de signalisation intervenant dans la régulation du tonus vasculaire et dans l’i t g it de la
a i e e doth liale. Il a lio e la sista e des a dio o tes à l’is h ie-reperfusion.
Cependant, le rôle de ce lysophospholipide dans la régulation des propriétés des cellules
sou hes a dia ue ’a pas t tudi .
Nous nous sommes donc focalisés dans notre étude sur les points suivants :
Ca a t isatio de l’e p essio des i epteu s à la “ P da s les C“Cs W B +
Effet de la S1P utilisé à 1µM (concentration plasmatique normale) sur les propriétés
d’auto-renouvellement, de prolifération et de migration des CSCs W8B2+.
Implication des récepteurs à la S1P (exprimés dans CSCs W8B2+) dans la régulation
des propriétés fondamentales de ces cellules (auto-renouvellement, prolifération et
migration).

Résultats
199
3 Résultats
3.1 P ofil d’e p essio des i epteu s à la S P da s les CSCs W B +
La S1P assure la majorité de ses fonctions en agissant sur ces cinq récepteurs qui sont
exprimés de manière différente selon le tissu et le type cellulaire. Pour identifier les récepteurs
à la S1P (S1PR) exprimés dans les CSCs W8B2+, nous avons vérifié la présence ou non des
transcrits codant pour les 5 récepteurs par la technique de RT-PCR.
Les résultats obtenus montrent que les CSCs W8B2+ expriment les transcrits codant
pour les récepteurs S1PR1 (variant 1) et S1PR2 (figure R-53). Le tissu atrial humain utilisé
comme contrôle positif, présente aussi une expression des transcrits codant pour S1PR1
(variant 1 et 2) et S1PR2.
La figure R-54 A, montre que les CSCs W8B2+ expriment faiblement les transcrits
codant pour le S1PR3 comparées aux cellules issues de glioblastome humain (1) utilisées
comme contrôle positif. Concernant le S1PR4, les transcrits codant pour ce récepteur sont
totalement absents dans les CSC W8B2+ et le tissu atrial humain comparés aux cellules issues
de glioblastome humain (2).
Figure R-53 : Ca a t isatio de l’e p essio des ARN oda t pou les epteu s à la sphi gosi e 1 phosphate de type 1 (S1PR1) et de type 2 (S1PR2) dans les CSCs W8B2+ par RT-PCR. GAPDH est
utilisé comme gène de référence. Les ADNc issus du tissu atrial cardiaque humain sont utilisés
o e o t ôle positif et l’eau o e o t ôle gatif.
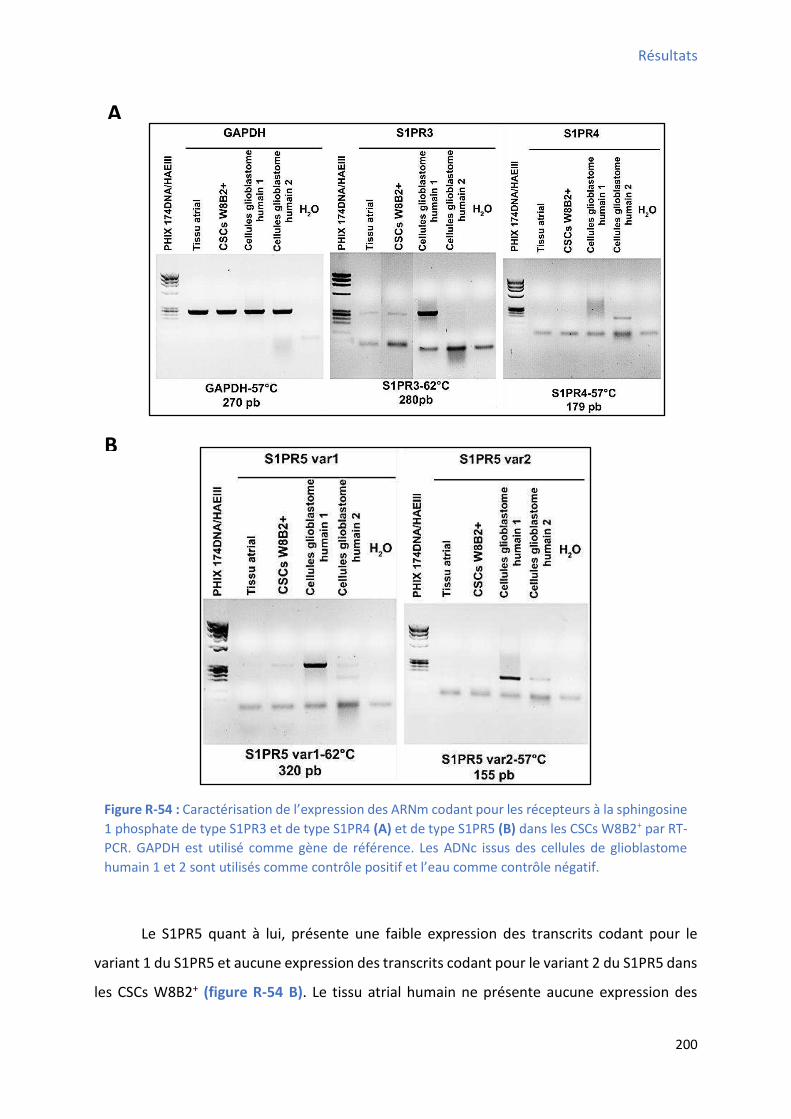
Résultats
200
Le S1PR5 quant à lui, présente une faible expression des transcrits codant pour le
variant 1 du S1PR5 et aucune expression des transcrits codant pour le variant 2 du S1PR5 dans
les CSCs W8B2+ (figure R-54 B). Le tissu atrial humain ne présente aucune expression des
Figure R-54 : Ca a t isatio de l’e p essio des ARN oda t pou les epteu s à la sphi gosi e 1 phosphate de type S1PR3 et de type S1PR4 (A) et de type S1PR5 (B) dans les CSCs W8B2+ par RT-
PCR. GAPDH est utilisé comme gène de référence. Les ADNc issus des cellules de glioblastome
hu ai et so t utilis s o e o t ôle positif et l’eau o e o t ôle gatif.
A
B
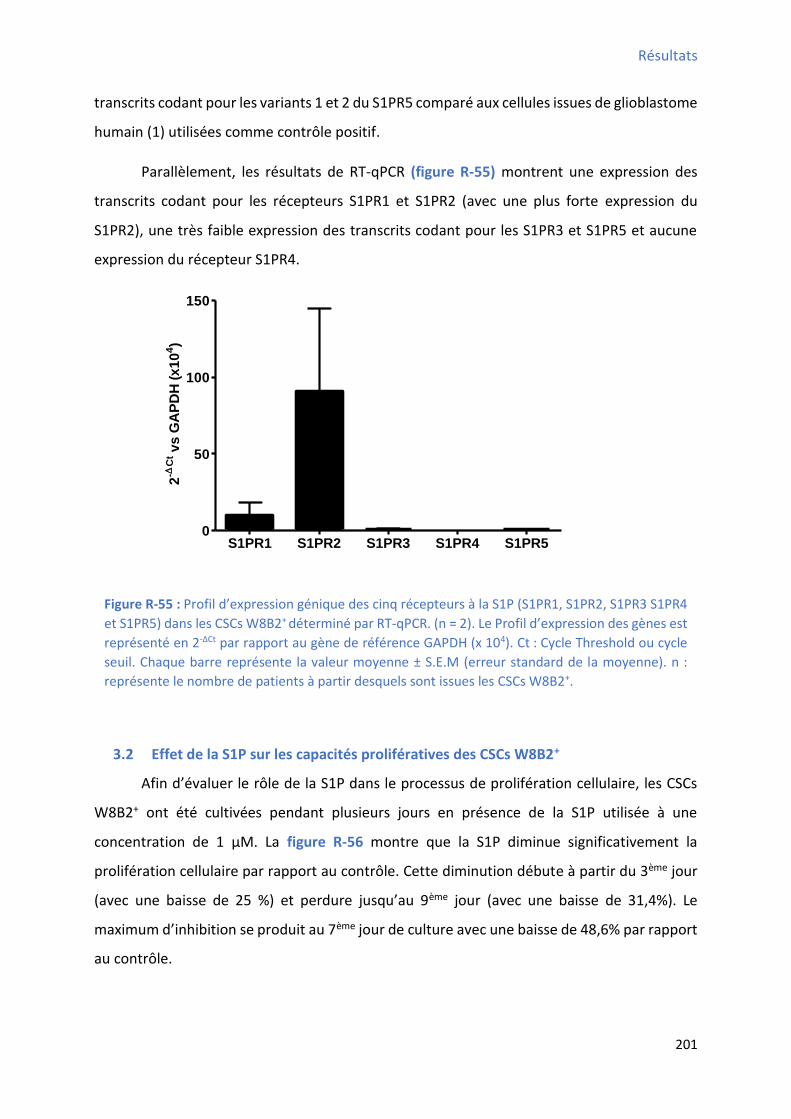
Résultats
201
transcrits codant pour les variants 1 et 2 du S1PR5 comparé aux cellules issues de glioblastome
humain (1) utilisées comme contrôle positif.
Parallèlement, les résultats de RT-qPCR (figure R-55) montrent une expression des
transcrits codant pour les récepteurs S1PR1 et S1PR2 (avec une plus forte expression du
S1PR2), une très faible expression des transcrits codant pour les S1PR3 et S1PR5 et aucune
expression du récepteur S1PR4.
3.2 Effet de la S1P sur les capacités prolifératives des CSCs W8B2+
Afi d’ alue le ôle de la “ P da s le p o essus de p olif atio ellulai e, les C“Cs
W8B2+ ont été cultivées pendant plusieurs jours en présence de la S1P utilisée à une
o e t atio de μM. La figure R-56 montre que la S1P diminue significativement la
prolifération cellulaire par rapport au contrôle. Cette diminution débute à partir du 3ème jour
a e u e aisse de % et pe du e jus u’au ème jour (avec une baisse de 31,4%). Le
a i u d’i hi itio se p oduit au ème jour de culture avec une baisse de 48,6% par rapport
au contrôle.
Figure R-55 : Profil d’e p essio g i ue des i epteu s à la “ P “ PR , “ PR , “ PR “ PR et S1PR5) dans les CSCs W8B2+ déterminé par RT-qPCR. (n = 2). Le Profil d’e p essio des g es est représenté en 2-ΔCt par rapport au gène de référence GAPDH (x 104). Ct : Cycle Threshold ou cycle
seuil. Chaque barre représente la valeur moyenne ± S.E.M (erreur standard de la moyenne). n :
représente le nombre de patients à partir desquels sont issues les CSCs W8B2+.
S1PR1 S1PR2 S1PR3 S1PR4 S1PR50
50
100
150
2- Ct v
s G
AP
DH
(x10
4 )
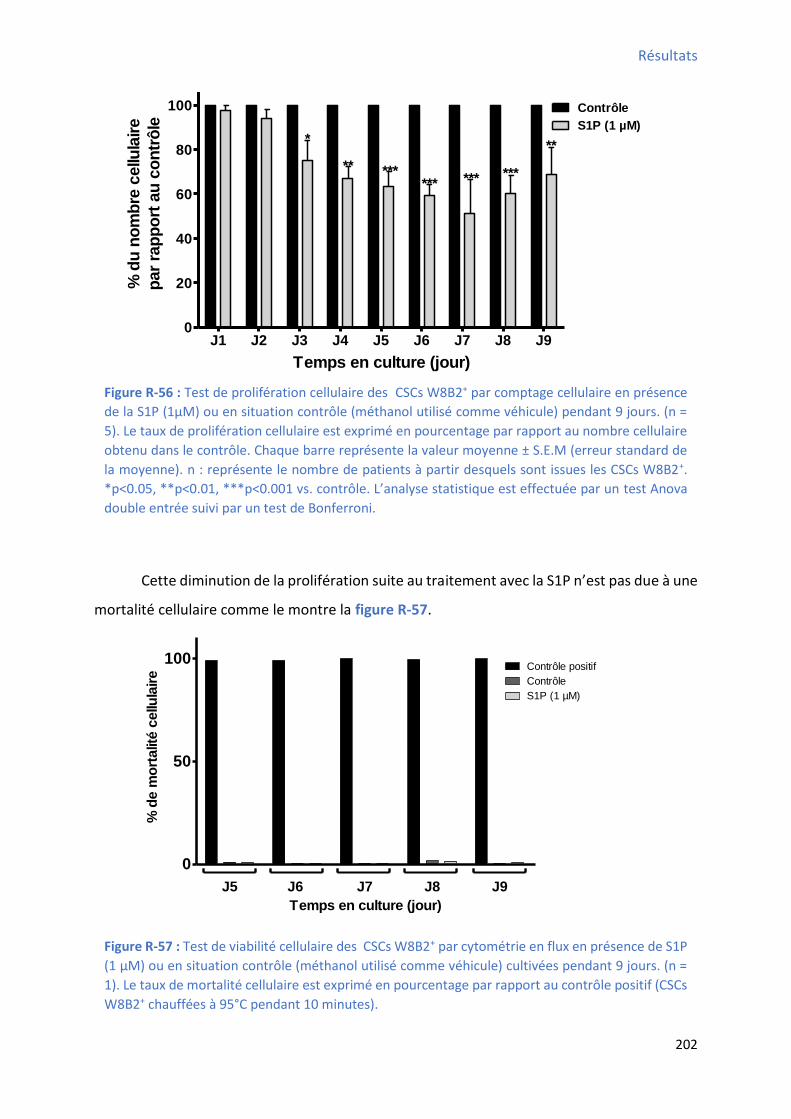
Résultats
202
Cette diminution de la prolifération suite au traite e t a e la “ P ’est pas due à une
mortalité cellulaire comme le montre la figure R-57.
Figure R-56 : Test de prolifération cellulaire des CSCs W8B2+ par comptage cellulaire en présence
de la S1P (1µM) ou en situation contrôle (méthanol utilisé comme véhicule) pendant 9 jours. (n =
5). Le taux de prolifération cellulaire est exprimé en pourcentage par rapport au nombre cellulaire
obtenu dans le contrôle. Chaque barre représente la valeur moyenne ± S.E.M (erreur standard de
la moyenne). n : représente le nombre de patients à partir desquels sont issues les CSCs W8B2+.
*p< . , **p< . , ***p< . s. o t ôle. L’a al se statisti ue est effe tu e pa u test Anova
double entrée suivi par un test de Bonferroni.
J1 J2 J3 J4 J5 J6 J7 J8 J90
20
40
60
80
100 ContrôleS1P (1 µM)
*
** ****** *** ***
**
Temps en culture (jour)
% d
u no
mbr
e ce
llula
irepa
r rap
port
au c
ontrô
le
J5 J6 J7 J8 J9
0
50
100Contrôle positifContrôleS1P (1 µM)
Temps en culture (jour)
% d
e m
orta
lité
cellu
laire
Figure R-57 : Test de viabilité cellulaire des CSCs W8B2+ par cytométrie en flux en présence de S1P
(1 µM) ou en situation contrôle (méthanol utilisé comme véhicule) cultivées pendant 9 jours. (n =
1). Le taux de mortalité cellulaire est exprimé en pourcentage par rapport au contrôle positif (CSCs
W8B2+ chauffées à 95°C pendant 10 minutes).
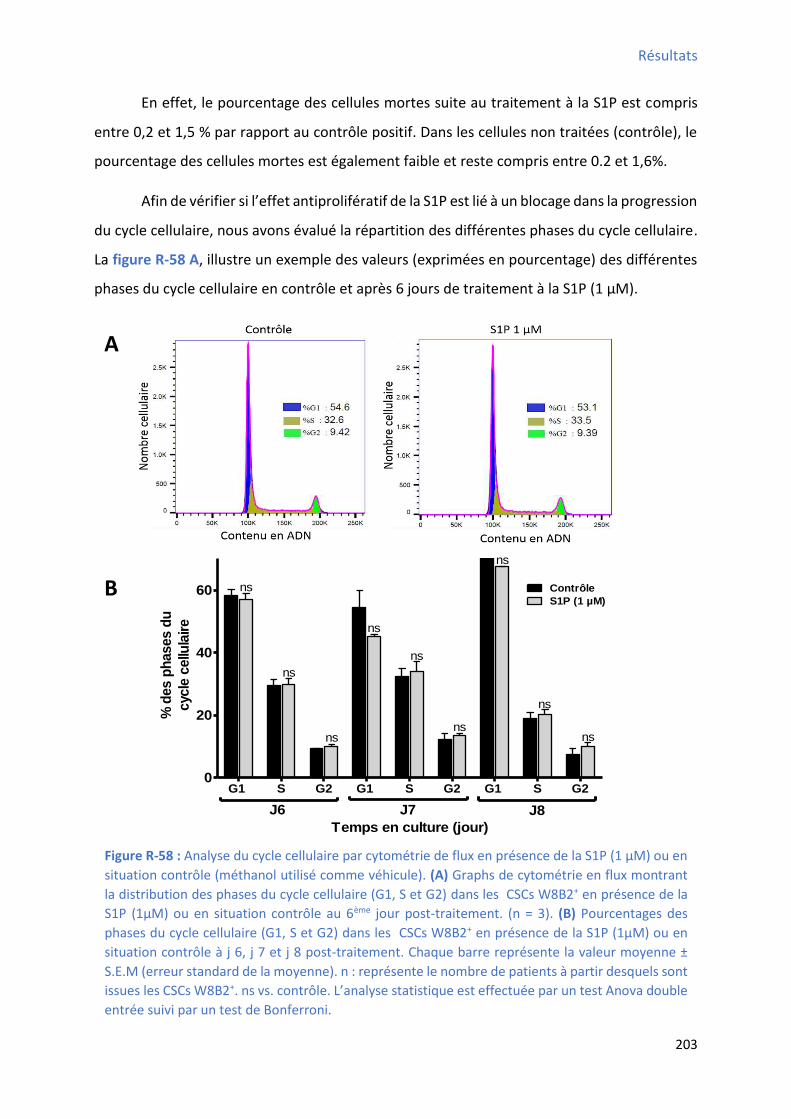
Résultats
203
En effet, le pourcentage des cellules mortes suite au traitement à la S1P est compris
entre 0,2 et 1,5 % par rapport au contrôle positif. Dans les cellules non traitées (contrôle), le
pourcentage des cellules mortes est également faible et reste compris entre 0.2 et 1,6%.
Afi de ifie si l’effet a tip olif atif de la “ P est li à u lo age da s la p og essio
du cycle cellulaire, nous avons évalué la répartition des différentes phases du cycle cellulaire.
La figure R-58 A, illustre un exemple des valeurs (exprimées en pourcentage) des différentes
phases du le ellulai e e o t ôle et ap s jou s de t aite e t à la “ P μM .
G1 S G2 G1 S G2 G1 S G20
20
40
60 ContrôleS1P (1 µM)
J6 J7 J8
ns
ns
ns
ns
ns
ns
ns
ns
ns
Temps en culture (jour)
% d
es p
hase
s du
cycl
e ce
llula
ire
Figure R-58 : Analyse du cycle cellulaire par cytométrie de flux en présence de la S1P (1 µM) ou en
situation contrôle (méthanol utilisé comme véhicule). (A) Graphs de cytométrie en flux montrant
la distribution des phases du cycle cellulaire (G1, S et G2) dans les CSCs W8B2+ en présence de la
S1P (1µM) ou en situation contrôle au 6ème jour post-traitement. (n = 3). (B) Pourcentages des
phases du cycle cellulaire (G1, S et G2) dans les CSCs W8B2+ en présence de la S1P (1µM) ou en
situation contrôle à j 6, j 7 et j 8 post-traitement. Chaque barre représente la valeur moyenne ±
S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir desquels sont
issues les CSCs W8B2+. s s. o t ôle. L’a al se statisti ue est effe tu e pa u test A o a dou le entrée suivi par un test de Bonferroni.
A
B
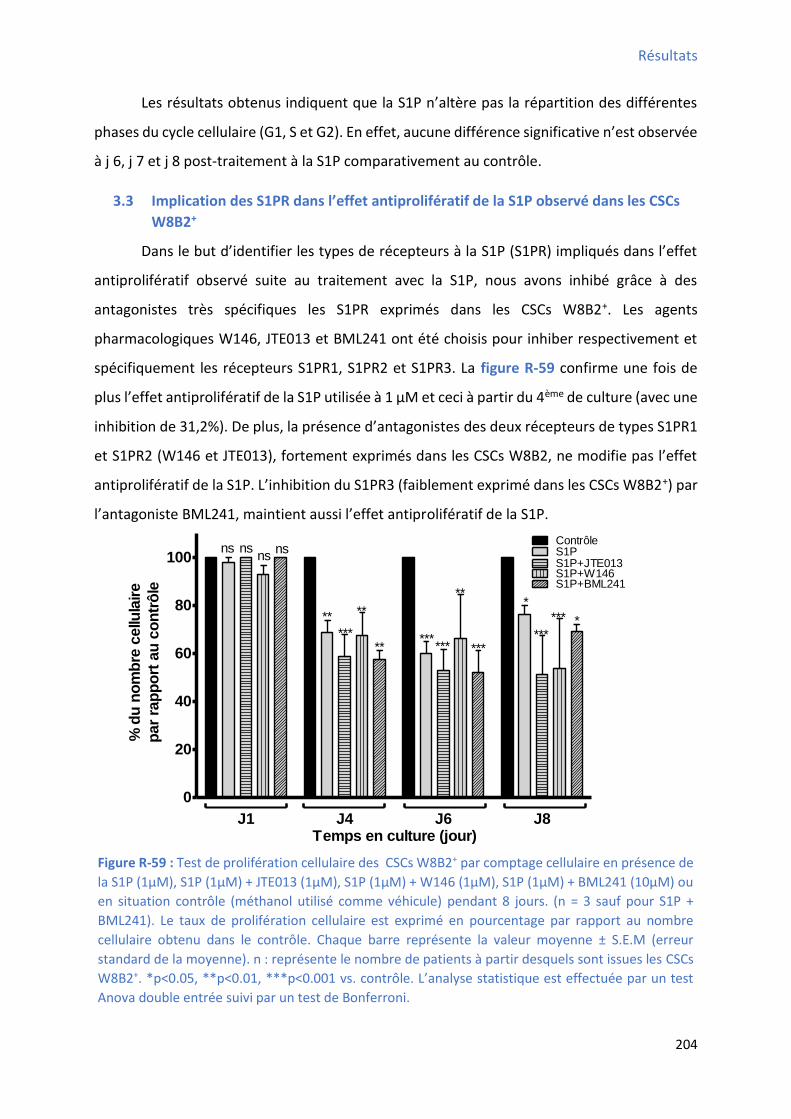
Résultats
204
Les sultats o te us i di ue t ue la “ P ’alt e pas la pa titio des diff e tes
phases du le ellulai e G , “ et G . E effet, au u e diff e e sig ifi ati e ’est o se e
à j 6, j 7 et j 8 post-traitement à la S1P comparativement au contrôle.
3.3 I pli atio des S PR da s l’effet a tip olif atif de la S P o se v da s les CSCs W8B2+
Da s le ut d’ide tifie les t pes de epteu s à la “ P “ PR i pli u s da s l’effet
antiprolifératif observé suite au traitement avec la S1P, nous avons inhibé grâce à des
antagonistes très spécifiques les S1PR exprimés dans les CSCs W8B2+. Les agents
pharmacologiques W146, JTE013 et BML241 ont été choisis pour inhiber respectivement et
spécifiquement les récepteurs S1PR1, S1PR2 et S1PR3. La figure R-59 confirme une fois de
plus l’effet a tip olif atif de la “ P utilis e à µM et e i à pa ti du ème de culture (avec une
i hi itio de , % . De plus, la p se e d’a tago istes des deux récepteurs de types S1PR1
et “ PR W et JTE , fo te e t e p i s da s les C“Cs W B , e odifie pas l’effet
a tip olif atif de la “ P. L’i hi itio du “ PR fai le e t e p i da s les C“Cs W B +) par
l’a tago iste BML , ai tie t aussi l’effet a tip olifératif de la S1P.
J1 J4 J6 J80
20
40
60
80
100ContrôleS1PS1P+JTE013S1P+W146S1P+BML241
ns
**
***
*
ns
******
***
ns
****
***
ns
** ***
*
Temps en culture (jour)
% d
u no
mbr
e ce
llula
irepa
r rap
port
au c
ontrô
le
Figure R-59 : Test de prolifération cellulaire des CSCs W8B2+ par comptage cellulaire en présence de
la S1P (1µM), S1P (1µM) + JTE013 (1µM), S1P (1µM) + W146 (1µM), S1P (1µM) + BML241 (10µM) ou
en situation contrôle (méthanol utilisé comme véhicule) pendant 8 jours. (n = 3 sauf pour S1P +
BML241). Le taux de prolifération cellulaire est exprimé en pourcentage par rapport au nombre
cellulaire obtenu dans le contrôle. Chaque barre représente la valeur moyenne ± S.E.M (erreur
standard de la moyenne). n : représente le nombre de patients à partir desquels sont issues les CSCs
W8B2+. *p< . , **p< . , ***p< . s. o t ôle. L’a al se statisti ue est effe tu e pa u test Anova double entrée suivi par un test de Bonferroni.
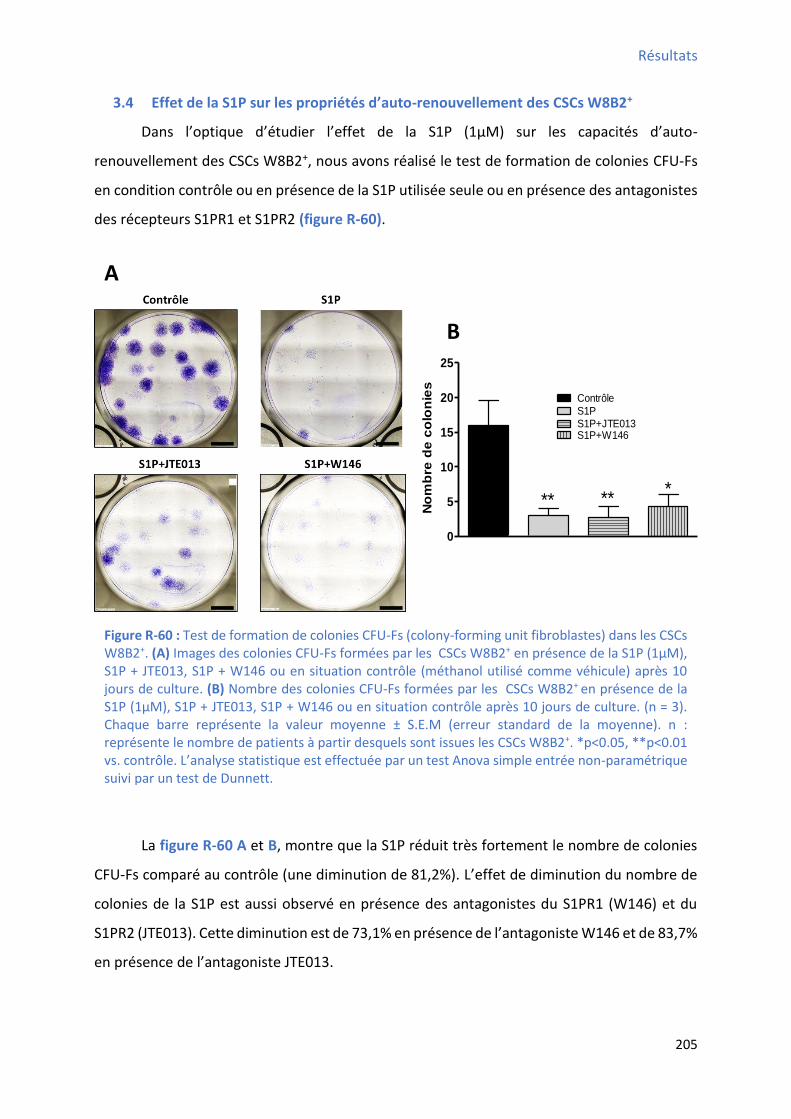
Résultats
205
3.4 Effet de la S P su les p op i t s d’auto-renouvellement des CSCs W8B2+
Da s l’opti ue d’ tudie l’effet de la “ P µM su les apa it s d’auto-
renouvellement des CSCs W8B2+, nous avons réalisé le test de formation de colonies CFU-Fs
en condition contrôle ou en présence de la S1P utilisée seule ou en présence des antagonistes
des récepteurs S1PR1 et S1PR2 (figure R-60).
La figure R-60 A et B, montre que la S1P réduit très fortement le nombre de colonies
CFU-Fs comparé au contrôle (une diminution de 81,2%). L’effet de di i utio du o e de
colonies de la S1P est aussi observé en présence des antagonistes du S1PR1 (W146) et du
S1PR2 (JTE013). Cette di i utio est de , % e p se e de l’a tago iste W et de , %
e p se e de l’a tago iste JTE .
0
5
10
15
20
25
** ** *
ContrôleS1PS1P+JTE013S1P+W146
No
mb
re d
e c
olo
nie
s
Figure R-60 : Test de formation de colonies CFU-Fs (colony-forming unit fibroblastes) dans les CSCs W8B2+. (A) Images des colonies CFU-Fs formées par les CSCs W8B2+ en présence de la S1P (1µM), S1P + JTE013, S1P + W146 ou en situation contrôle (méthanol utilisé comme véhicule) après 10 jours de culture. (B) Nombre des colonies CFU-Fs formées par les CSCs W8B2+ en présence de la S1P (1µM), S1P + JTE013, S1P + W146 ou en situation contrôle après 10 jours de culture. (n = 3). Chaque barre représente la valeur moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir desquels sont issues les CSCs W8B2+. *p<0.05, **p<0.01
s. o t ôle. L’a al se statisti ue est effe tu e pa u test A o a si ple e t e o -paramétrique suivi par un test de Dunnett.
A
B
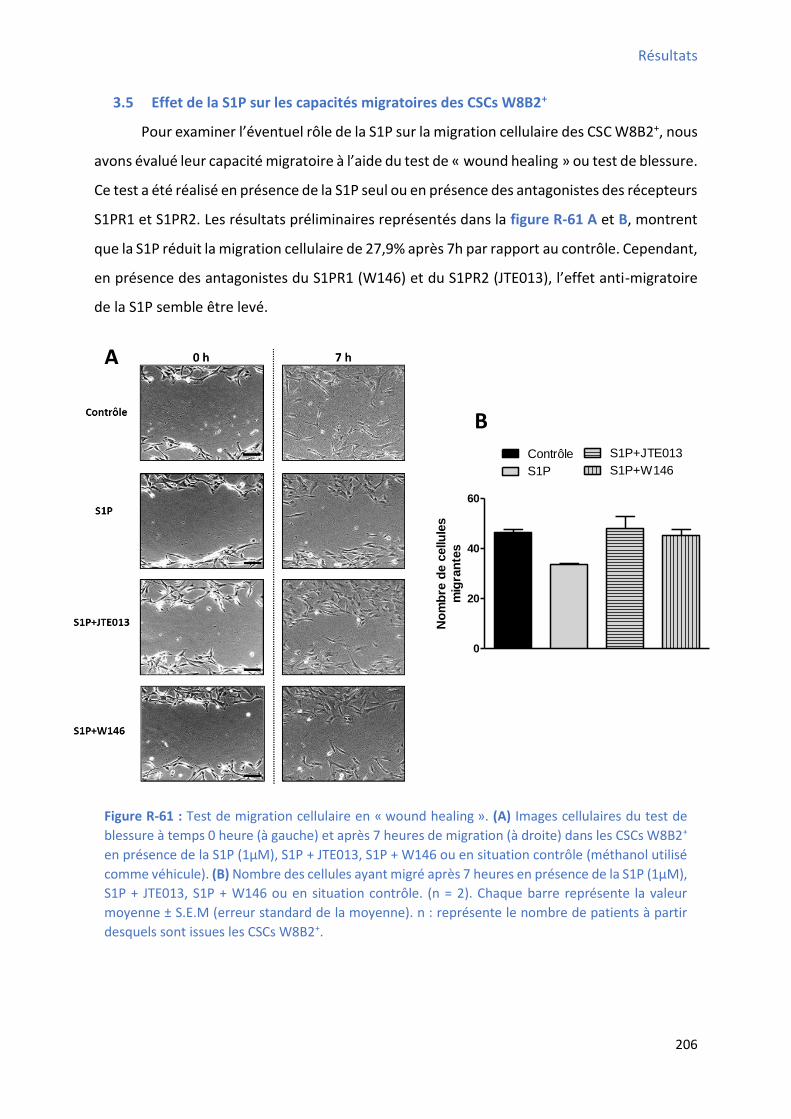
Résultats
206
3.5 Effet de la S1P sur les capacités migratoires des CSCs W8B2+
Pou e a i e l’ e tuel ôle de la “ P su la ig atio ellulai e des C“C W8B2+, nous
a o s alu leu apa it ig atoi e à l’aide du test de « wound healing » ou test de blessure.
Ce test a été réalisé en présence de la S1P seul ou en présence des antagonistes des récepteurs
S1PR1 et S1PR2. Les résultats préliminaires représentés dans la figure R-61 A et B, montrent
que la S1P réduit la migration cellulaire de 27,9% après 7h par rapport au contrôle. Cependant,
e p se e des a tago istes du “ PR W et du “ PR JTE , l’effet a ti-migratoire
de la S1P semble être levé.
A
Figure R-61 : Test de migration cellulaire en « wound healing ». (A) Images cellulaires du test de
blessure à temps 0 heure (à gauche) et après 7 heures de migration (à droite) dans les CSCs W8B2+
en présence de la S1P (1µM), S1P + JTE013, S1P + W146 ou en situation contrôle (méthanol utilisé
comme véhicule). (B) Nombre des cellules ayant migré après 7 heures en présence de la S1P (1µM),
S1P + JTE013, S1P + W146 ou en situation contrôle. (n = 2). Chaque barre représente la valeur
moyenne ± S.E.M (erreur standard de la moyenne). n : représente le nombre de patients à partir
desquels sont issues les CSCs W8B2+.
0
20
40
60
ContrôleS1P
S1P+JTE013S1P+W146
Nom
bre
de c
ellu
les
mig
rant
es
B

Discussion
207
DISCUSSION
I-Isolement et caractérisation des CSCs W8B2+
Dans la première partie des résultats, nous avons mis en place un modèle de cellules
souches cardiaques exprimant le marqueur W8B2 (CSCs W8B2+ à pa ti d’ ha tillo s
auriculaires humains en collaboration avec le CHU de Poitiers. Ces CSCs W8B2+ isolées dans
un premier temps par tri cellulaire magnétique ne représentaient que 3% de la population
cellulaire totale. Ce faible pourcentage est proche de celui obtenu par BEARZI et al., 2007 pour
la population des CSCs c-kit+ hu ai es. Afi d’a lio e le e de e t ous a o s i s u e
étape de tri cellulaire par cytométrie de flux, plus fiable et efficace, et qui nous a permis
d’o te i u e populatio pu e de C“Cs W B + (plus de 98% de pureté).
Les CSCs W8B2+ isolées sont apa les d’auto-renouvellement et hautement
prolifératives. Elles expriment des marqueurs typiques des cellules souches
se h ateuses W B , CD , CD et CD o e l’o t d jà o t )ha g et ses
collègues (Zhang et al., 2015a). Cependant, les CSCs W8B2+ isolées dans nos conditions
expérimentales, présentent une faible expression des marqueurs mésenchymateux CD90 et
CD106. Néanmoins, une faible expression du CD90 est fréquemment observée sur les CSCs
hu ai es d’o igi e se h ateuse Rossi i et al., 1 ; Reus et al., 2016). De plus, Yang
et ses oll gues, o t o t u’au sei e des C“Ms hu ai es, l’e p essio de CD
variait selon la nature tissulaire (0,73% dans les CSMs du tissu adipeux, 7,44% dans les CSMs
du cordon ombilical, 32,04% dans les CSMs de la moelle osseuse et 65,01% dans les CSMs du
placenta) (Yang et al., 2013b). Cette diff e e d’e p essio du CD et CD , peut aussi t e
due au diff e tes sou es d’a ti o ps utilis s pou la to t ie. Les C“Cs W B + présentent
une très faible expression des marqueurs de cellules hématopoïétiques (CD34, CD45, CD133)
et du marqueur de CSCs CD117 (ou c-kit , o fi a t ai si l’o igi e se h ateuse de
notre population.
Les CSCs W8B2+ non différenciées expriment certains transcrits codant pour des
facteurs de transcription spécifiques du lignage cardiaque comme GATA4 et TBX3. GATA4 et
TBX3 ont un rôle clé dans le développement et la fonction cardiaque (Ang et al., 2016; Yang et
al., 2013a). TBX3 est essentiel dans la régulation génique et phénotypique des cellules nodales

Discussion
208
(Hoogaars et al., 2007). Les travaux de Bakker et al. ont mont ue l’i du tio de TBX3
favorisait la reprogrammation des cardiomyocytes matures de souris en cellules possédant un
phénotype nodale (Bakker et al., 2012). Co e da s d’aut es od les de ellules souches
cardiaques, les CSCs W8B2+ ’e p i e t pas tous les fa teu s de t a s iptio a dia ues tels
que GATA5, IRX4 ou encore NKX2.5. Par exemple, dans les CSCs c-kit+ humaines, seulement
0,4% des cellules sont positives pour GATA4 et NKX2.5 (Bearzi et al., 2007). Dans les CSCs Isl-
1+ fœtales hu ai es, deu populatio s a e u pou oi de diff e iatio diff e t o t t
identifiées; une population exprimant uniquement Isl-1 et une autre qui expriment Isl-1 avec
Nkx2.5 et TBX1 (Bu et al., 2009). Il est do p o a le ue l’e p ession des facteurs de
transcription cardiaques soit dépendante du type des CSCs étudié.
La nature non différencié des CSCs W8B2+ isolées dans nos conditions expérimentales
est e fo e pa l’a se e d’e p essio g i ue des st u tu es o t a tiles a dia ues
(comme par exemple ANKB, TNNI3, TNNT2).
Le œu e p i e uat e isofo es de la o e i e C , C , C et C . a e
u e dist i utio pa ti uli e selo la gio du œu Despla tez, . L’e p essio des
quatre isoformes de la connexine cardiaque (avec une prédominance de la connexine 43) est
détectée dans les CSCs W8B2+. Leu p se e sugg e u’elles pou aie t joue u ôle da s
le contact intracellulaire observé dans les niches intra tissulaires des cellules souches
cardiaques.
Le profil d’e p essio g i ue des peptides at iu ti ues NP o t e ue les C“Cs W B +
e p i e t le NP de t pe B. Ce de ie ui est u i di ateu d’i suffisa e a dia ue, a t
récemment montré comme modulateur important de la prolifération des CSCs Sca-1+ murines
(Rignault-Clerc et al., 2017).
II-P ofil d’e p essio g i ue des a au io i ues da s les CSCs W B +
L’a al se des t a s its oda t pou les a au io i ues o t e ue les C“Cs W B +
p se te t u ph ot pe de ellules o e ita les ui se t aduit pa l’a se e des t a s its
oda t pou les a au sodi ues ota e t l’isofo e a dia ue NaV1.5) et les canaux
calciques (notamment les isoformes cardiaques CaV 1.2, CaV 1.3, CaV 3.1 et CaV 3.2). Dans la
littérature, seule une petite population de CSMs humaines de la moelle osseuse présentait
des canaux sodiques tetrodotoxine (TTX) sensibles (codés par NaV1.7 dans 29% des cellules)

Discussion
209
et des canaux de type ICa.L (codés par CaV1.2 dans 15% des cellules) (Li et al., 2005).
N a oi s, u e aut e tude alis e su le e t pe ellulai e i di ue l’a se e des
transcrits codant pour le canal sodique NaV . o l e a e l’a se e des ou a t sodi ues
(Heubach et al., 2004). Dans les CSMs humaines du tissu adipeux, seulement des courants INa
ont pu être enregistrés dans une petite population de cellules (8%) (Bai et al., 2007) alors que
dans les CSMs de la veine du cordon ombilical humain, 30% des cellules présentaient des
courants INa TTX sensibles (codé par hNE-Na) (Park et al., 2007). Dans les CSC c-kit+ humaines,
des courants Na+ TTX sensibles codés par SCN3A et SCN8A ont été observés dans 61% des
cellules (Zhang et al., 2014a). Ces études suggèrent que le profil d'expression des canaux
ioniques dans les CSMs peut être tissu-spécifique.
Contrairement aux canaux sodiques, les CSCs W8B2+ montre une expression génique
de certains canaux potassiques codant pour les canaux K dépendant du voltage KV3.4 et KV4.2
(Ito), les canaux K activés par le calcium intracellulaire BKCa et SK4, les canaux K à rectification
entrante Kir2.1 et Kir6.1 et les canaux K TWIK1. Ces résultats sont en accord avec ceux décrits
dans la littérature notamment dans les CSMs humaines de la moelle osseuse où des courants
KCa (codé par KCa1.1) ont été observés (Heubach et al., 2004).
D’aut es tudes alis es da s les CSMs de la veine du cordon ombilical humain (Park
et al., o t o t l’e p essio de plusieu s ou a ts potassi ues BKCa (codé par KCa1.1)
dans 92% des cellules, Ito (codé par KV1.4 et KV4.2) dans 50% des cellules, et des courants IKir
(codé par TWIK et Kir2.1) dans 5% des cellules. En outre, aucun courant IKdr typique n'a été
enregistré, bien que les transcrits KV1.1 et KV10.1 soient détectés dans ces cellules. Dans les
CSMs humaines du tissu adipeux, des courants IKdr et BKCa ont pu être enregistrés (73% des
cellules) ainsi que des courants Ito (19% des cellules) (Bai et al., 2007). Dans les cellules
progénitrices cardiaques humaines c-kit+, plusieurs courants potassiques ont été également
caractérisés (des courants BKCa codés par KCa.1.1 (dans 86% des cellules), des courants IKir
codés par Kir2.1 (dans 84% des cellules), des courants Ito codés par Kv4.2 et Kv4.3 (dans 47%
des cellules) (Zhang et al., 2014).
III-Rôle des canaux ioniques dans les CSCs W8B2+
L’e se le de es tudes o t e t u e e p essio diff e tielle des a au io i ues
dans les cellules souches qui serait liée à leur origine tissulaire et/ou à des fonctions

Discussion
210
spécifiques. Le rôle de ces canaux dans les cellules souches adultes reste peu étudié mais les
do es dispo i les o t e t u’ils so t t oite e t i pli u s da s la gulatio de la
prolifération et la migration cellulaire.
Le blocage du courant IKdr réduit considérablement la prolifération des CSMs
hu ai es du tissu adipeu alo s ue l'i hi itio d’IKCa 'a pas d'effet Bai et al., . De
plus, l’i hi itio des ou a ts BKCa et KV4.2 (Ito) réduit la prolifération cellulaire des pré-
adipocytes humains (Hu et al., 2009). Une autre étude a montré que l’i hi itio de BKCa réduit
la prolifération cellulaire des CSMs hu ai es de la oelle osseuse e p o o ua t l’a t du
cycle en phase G0/G1 (Zhang et al., 2014b). Dans les CSCs c-kit+ hu ai es, l’i hi itio de BKCa
réduit la prolifération (en accumulant les cellules en phase G0/G1) et réduit également la
ig atio ellulai e. Cepe da t, l’i hi itio du a al Ki . ’affe te pas la p olif atio
cellulaire mais stimule la migration cellulaire (Zhang et al., 2015b).
Dans notre modèle de CSCs W8B2+, nous avons observé deux courants majoritaires, le courant
BKCa (codé par KCa1.1) et le courant IK1 od p o a le e t pa Ki . . L’appli atio de
l’apa i e lo ueu du a al SK1, 2, 3) ne provoque aucun effet suggérant que les canaux
SK1, 2 et 3 ne sont pas des canaux fonctionnels dans les CSCs W8B2+ (résultats non présentés).
Quant au canal BKCa il joue un rôle important dans notre modèle cellulaire car son inhibition
affecte les propriétés fondamentales (prolifération et auto-renouvellement) des CSCs W8B2+
(Résultats, chapitre II « article scientifique »).
Le rôle précis des canaux Na+ TTX sensibles et des canaux Na+ TTX résistants (NaV1.5),
’est pas totale e t lu id da s les ellules sou hes adultes. Da s les CSMs humaines du
tissu adipeux, le blocage de l'INa n'affecte pas la prolifération cellulaire (Bai et al., 2007). De
e, l’i hi itio des canaux Na+ TTX sensibles dans les CSCs c-kit+ ’a pas d’i pa t su leu
prolifération (Zhang et al., 2015b).
Nos résultats ont également montré que les CSCs W8B2+ expriment les transcrits
codant pour les canaux potassiques TWIK1 de la famille des canaux K à deux pores (K2P) qui
sont entre autres des canaux sensibles au pH (Chatelain et al., 2012). Les données de la
littérature ne cessent de démontrer que les acteurs de cette famille possèdent divers rôles
physiologiques et pathologiques (Feliciangeli et al., 2015). Un étude récente montre que le

Discussion
211
a al TWIK est fo te e t e p i da s les o eillettes et u’il est di e te e t i pli u da s
la régulation de la fréquence cardiaque et de la taille des oreillettes (Christensen et al., 2016).
Malg la p se e des t a s its oda t pou HCN , ous ’a o s pas o se de
courant If dans les CSCs W8B2+. Heubach et ses collègues ont obtenus les mêmes résultats
dans les CSMs humaines de la moelle osseuse, qui exp i e t u i ue e t l’ARN de
l’isofo e HCN sa s g e de ou a t a ti e h pe pola isatio Heu a h et al., .
Au u e tude à l’heu e a tuelle ’a o t l’e iste e de ou a ts If dans les CSCs humaines
adultes.
Concernant les canaux calciques, nous ’a o s e egist au u ou a t al i ue da s
notre modèle de CSCs W8B2+. Ce constat est peu surprenant car les résultats obtenus en RT-
qPCR ne montrent aucune expression des transcrits codant pour les CaV de type L (Cav1.2,
Cav1.3) et les CaV de type T (CaV3.1 et CaV3.2). Cependant, plusieurs équipes ont reporté
l’e p essio des t a s its oda ts pou les CaV de t pe L et T et la p se e de ou a t CaV
de type L dans les CSMs humaines de la moelle osseuse (Kawano et al., 2002 ; Heubach et al.,
2004 ; Zahanich et al., 2005). Il est important de mentionner dans ces travaux une
h t og it e tai e da s l’e p essio fo tio elle des a au al i ues. E effet, % des
CSMs de la moelle osseuse expriment les transcrits codant pour les canaux Ca2+ de type L et
seulement une petite population (13%) de ces cellules présentent des courant fonctionnels
(Zahanich et al., 2005). Dans les CSMs humaines du tissu adipeux, les ARNm des CaV de type L
et T ont été détectés sans aucune fonctionnalité (Bai et al., 2007). De même, dans les CSCs c-
kit+ hu ai es, au u ou a t al i ue ’a t o se (Zhang et al., 2014a).
IV-P ofil d’e p essio g i ue des a teurs calciques dans les CSCs W8B2+
Bien que les CSCs W8B2+ ’e p i e t pas de t a s its oda t pou les a au
calciques, nos résultats montrent une présence de plusieurs transporteurs ioniques (la pompe
Na+/K+, la pompe calcique SERCA, la pompe à calcium PMCA et l’ ha geu Na+/Ca2+) et des
a teu s de l’ho ostasie al i ue la CALM / et les epteu s à l’IP ITPR / .
Ce p ofil d’e p essio est se la le à e ui a t d jà d it su les CSMs humaines
et ui se ait à l’o igi e de ph o es d’os illations calciques spontanées (Ye, 2010). En effet,
les CSMs humaines possèdent des oscillations calciques spontanées qui sont régulées
ajo itai e e t pa les IP R, la “ERCA, l’ ha geu Na+/Ca2+ et la pompe PMCA (Kawano et

Discussion
212
al., 2002, 2003, 2006; Orciani et al., 2010). Ces oscillations induiraient des fluctuations du
potentiel membranaire via le canal potassique IKCa (Kawano et al., 2003) et seraient associées
à la translocation nucléaire de NFAT; un facteur de transcription connu comme régulateur
d’e p essio g i ue Ka a o et al., .
Ces oscillations calciques sont aussi présentes dans les CSCs c-kit+ humaines (Ferreira-
Martins et al., 2009; Maxwell et al., 2016) et sont régulées par les IP3R et la SERCA sans
impliquer la pompe Ca2+ membranaire (Ferreira-Martins et al., 2009). D’aut e pa t, il appa ait
que ces oscillations calciques jouent un rôle essentiel dans la régulation de la prolifération
cellulaire. L'induction des oscillations calciques dans les CSCs c-kit+ avant leur injection dans
des œu s i fa is de sou is, a lio e ait la g effe et l’e pa sio de es ellules sou hes.
(Ferreira-Martins et al., 2009).
Il est à noter que notre modèle de CSCs W8B2+, nous retrouvons aussi ces oscillations
al i ues sultats o o t s ais ous ’a o s pas e o e a a t is les a teu s
calciques impliqués dans leur genèse.
Pa all le e t à l’e p essio des odulateu s i pli u s da s l’ho ostasie al i ue
nous avons pu mettre en évidence, sur notre modèle de CSCs W8B2+, l’e p essio des
transcrits codants pour la calmoduline et la calcineurine qui sont deux principaux médiateurs
des effets moléculaires du calcium. La calmoduline, qui est un acteur fondamental de
t adu tio ellulai e li à l’a ti it al i ue, gule plusieu s p o essus ellulai es do t la
régulation du cycle cellulaire et la cytokinèse (Tsang et al., 2006). E a ti a t d’aut es p ot i e
(comme la CaMKII ou Ca2+/calmodulin-dependant protein kinase II), la calmoduline promeut
la survie des CSCs de souris et leur engagement dans le lignage cardiaque (Quijada et al.,
2015). La calcineurine intervient également dans diverses fonctions notamment le
développement du muscle cardiaque et squelettique via le facteur de transcription NFAT
(Schulz and Yutzey, 2004; Seok et al., 2014; Wu et al., 2009) et le cycle cellulaire (Afroze et al.,
2003).
Il est donc très probable que la calmoduline et la calcineurine jouent un rôle
fondamental dans la régulation de prolifération des CSCs W8B2+ en relation directe ou
indirecte avec la signature calcique.

Discussion
213
En conclusion, de cette partie nous avons pu isoler et purifier à partir de biopsie de
œu hu ai des ellules d’o igi e se h ateuse qui possèdent toutes les
caractéristiques des cellules sou hes. L’a al se glo ale de l’e p essio g i ue i di ue
lai e e t u’elles e so t pas « strictement excitables » par analogie aux cardiomyocytes
(absence de canaux dépolarisants de type sodique et calcique) mais auraient une activité
intracellulaire liée probablement aux IP3 et à certaines conductances potassiques calcium-
dépendante de type BKCa. Ces cellules possèdent un potentiel de différenciation certain
orienté prioritairement vers un génotype et un phénotype myocytaire.
V-Différenciation cardiaque in vitro des CSCs W8B2+
1 Profil d’e p essio des marqueurs cardiaques
Les cellules souches adultes ont la propriété de se différencier in vivo et générer des
cellules différenciées du tissu dans lequel elles se trouvent (Ghosh et al., 2011; Guilak et al.,
2010). L’o je tif i i est d’ tudie le pouvoir de différenciation cardiaque de notre modèle de
CSCs W8B2+. Ce choix de différenciation est particulièrement intéressant car il permet de
proposer des thérapies cellulaires de type autologue qui permettraient de remplacer des
tissus nécrosés par de nouveaux cardiomyocytes lors de pathologies cardiaques comme
l’i fa tus pa e e ple.
Pou i dui e ette diff e iatio a dia ue et da s le ut d’u e tude o pa ati e
nous avons utilisé des inducteurs chimiques de différenciation en se basant sur les travaux de
Zhang (Zhang et al., 2015a).
Nos résultats de RT-qPCR montrent u’ap s jou s de diff e iatio in vitro, les
CSCs W8B2+ présentent un profil génique très différent de celui avant différenciation.
Néanmoins, les cellules différenciées ne présentent pas le phénotype strict des
a dio o tes. Ce i se t aduit à l’ helle p ot i ue pa l’a se e de st iatio s t pi ues des
structures contractiles (la troponine T cardiaque, l’α-actinine, la chaîne lourde la myosine
a dia ue et à l’ helle t a s ipto i ue pa l’a se e d’i du tio des t a s its odant
notamment pour les chaînes lourdes de la myosine cardiaques. En revanche, certains
t a s its oda t pou des a ueu s de diff e iatio a dia ue o e l’a k i e B, la
chaîne légère de la myosine, le facteur natriurétique de type B sont surexprimés au cours de
du processus de différenciation.

Discussion
214
L’a k i e B ANKB a t o t e o e u e p ot i e gulat i e de l’auto atis e
du œud si o-auriculaire humain (Hund and Mohler, 2008; Le Scouarnec et al., 2008)
probablement via la régulation du canal calcique CaV1.3 (Cunha et al., 2011). Parmi les chaînes
légères de la osi e, l’isofo e au i ulai e MYL7 est surexprimé au cours de la
diff e iatio a dia ue o e l’o t o t )ha g et ses oll gues (Zhang et al., 2015a).
Ces résultats ne sont aucunement surprenant car les cardiomyocytes différenciés à partir des
cellules souches in vitro ont été largement montrés comme ayant un phénotype cardiaque
immature sur les plans morphologiques et électrophysiologiques (Shi et al., 2016; Yang et al.,
2014; Zhang et al., 2015a).
Il est probable que les protéines contractiles aient une localisation nucléaire immature
dans les « cardiomyocytes » issus des CSCs W8B2+. En effet, plusieurs études ont montré
l’e p essio de structures contractiles (comme la tropomyosine et la troponine) dans le noyau
des CSMs de rat à un stade précoce de différenciation (Asumda and Chase, 2012). D’aut es
marqueurs cardiaques comme les facteurs de transcription GATA5 et GATA6 se trouvent
également surexprimés au cours de la différenciation cardiaque des CSCs W8B2+. Ces deux
facteurs jouent un rôle fondamental dans la morphogenèse cardiaque (Charron and Nemer,
1999; Peterkin et al., 2005) particulièrement vers le développe e t d’u ph ot pe odal ,
comme par exemple, sur les CSEs de souris (Hashem and Claycomb, 2013; Wiese et al., 2011).
GATA6 est également surexprimé au cours de la différenciation vers un phénotype
cardiomyocytaire des cellules humaines du cordon ombilical exprimant c-kit (Iachininoto et
al., 2015) mais aussi lors la reprogrammation des CSMs humaines en précurseurs
cardiovasculaires (Vecellio et al., 2012). GATA6 serait également essentiel dans le processus
de différenciation en cellules musculaires lisses vasculaires (Lepore et al., 2005; Losa et al.,
2017).
Dans ce contexte, il est a noter u’au u e i du tio de GATA ’a t o se e à
l’ helle t a s ipto i ue da s ot e od le, o e peu e t le o fi e deu aut es
études (Reus et al., 2016; Zhang et al., 2015a).
Nos sultats soulig e t gale e t u e a plifi atio de l’e p essio du fa teu
natriurétique de type B (BNP) au cours de la différenciation des CSCs W8B2+. Cette induction
a été également retrouvée lors de la différenciation en cardiomyocytes des CSMs de rat et des
CSEs de singe (Abdelalim et al., 2006; Fukuda, 2002; He et al., 2011). Ce facteur natriurétique

Discussion
215
est apa le d’i dui e la diff e iatio a dio o tai e des C“Cs de sou is (Bielmann et al.,
2015) mais également d’améliorer la fonction cardiaque lors de transplantation de CSMs dans
u od le u i d’i suffisa e a dia ue (Zhang et al., 2013). Récemment, Al-Maqtari et ses
collègues ont montré que les transcrits codant pour le BNP sont surexprimés lors de
l’i du tio des C“Cs -kit+ humaines par certains facteurs de transcription cardiaques (Al-
Maqtari et al., 2017).
Cependant, nous ’a o s pas o se de ha ge e ts d’e p essio de t a s its
oda t pou le fa teu at iu ti ue de t pe A ANP d’auta t plus ue e de ie e p se te
pas d’e p essio da s les C“Cs W B + a a t diff e iatio . Il est à ote u’il a pu t e is
en évidence, notamment par notre équipe, que l’ANP joue u ôle i po ta t lo s de la
différenciation des fibroblastes de rat en myofibroblastes (Cameron et al., 2000; Moubarak et
al., 2015). L’e p essio et le ôle de es deu t pes de fa teu s at iu ti ues pou aie t
dépendre du type cellulaire.
L’e se le de es sultats o fi e t le ph ot pe i atu e des a dio o tes o te us
après différenciation in vitro des CSCs W8B2+.
L’utilisatio de l’optog ti ue à l’aide du ChR o e o e pe etta t
l’a lio atio de ette diff e iatio s’est l i effi a e a le « clustering hiérarchique »
des gènes se rapproche plus des CSCs W8B2+ non différenciées que celui des CSCs W8B2+
après différenciation cardiaque. Nous pensons que ce constat viendrait de la dégradation par
la lumière bleue (utilisée en pulse de 50 ms à une fréquence de 1Hz pour stimuler le ChR2
tout au long du protocole de différenciation) des agents de différenciation utilisés dans notre
protocole comme par exemple la vitamine A (acide rétinoïque).
2 Profil d’e p essio des transcrits codant pour les canaux ioniques
Les résultats des RT- PCR ous o t pe is d’ide tifie plusieu s t a s its oda ts pou
des a au io i ues sodi ues, potassi ues, al i ues, HCN do t d’e p essio appa ait au
cours de la différenciation cardiaque in vitro de notre modèle de CSCs W8B2+.
En effet, plusieurs transcrits codants pour les canaux sodiques apparaissent au cours
de la diff e iatio ota e t, l’ e ge e de l’e p essio de l’isofo e a dia ue
NaV1.5 ; un canal sodique responsable de la phase de dépolarisation (phase 0) du potentiel
d’a tio a dia ue. E plus de l’isofo e a dia ue, les t a s its oda ts pou les isofo es

Discussion
216
neuronaux (NaV1.7) et du muscle squelettique (NaV1.4) émergent au cours de la
diff e iatio . Cette o se atio est oupl e à l’appa itio des t a s its codants pour les
sous-unités β régulatrices (NaV β et NaV β . Ces sultats so t t s se la les à eu d its
au cours de la différenciation cardiomyocytaire des iPSC humaines (Moreau et al., 2017). Une
autre isoforme (NaV . e p i da s le œu hu ai Ga o it et al., appa ait
également durant la différenciation. Une étude réalisée sur les CSCs c-kit+ de souris a montré
l’e p essio p ot i ue de a au sodi ues de e atu e ais o fo tio els a se e
de courant sodique) (Lagostena et al., 2005).
Après différenciation « cardiaque » des CSCs W8B2+, nous avons également observé
l’appa itio des t a s its KV1.4 et KV4.3 et la surexpression du transcrits KV4.2 codant pour
les canaux potassiques cardiaques impliqués dans le courant transitoire sortant repolarisant
Ito du a t la phase du pote tiel d’a tio a dia ue. Il est i po ta t de p ise ue des
transcrits codant pour des canaux potassiques neuronaux apparaissent au cours de la
différenciation cardiaque. Ceux-ci incluent le KV1.2 et le KV2.1 ; des canaux responsables du
courant IKDR essai e à la epola isatio du pote tiel d’a tio eu o al.
Les transcrits codant pour les canaux KV1.5 et KV1.7 responsables des courants sortants
repolarisants ultra-rapides IKur apparaissent au cours de la différenciation. Cependant les
transcrits codant pour les canaux hERG (responsables du courant IKr) et KVLQT1 (responsables
du courant IKs) essentiels pou la epola isatio phase du pote tiel d’a tio a diaque sont
absents. Parallèlement, nous avons observé la surexpression des sous-unités β régulatrices
des canaux potassiques après différenciation cardiaque.
Nous retrouvons également une induction très forte des transcrits codant pour TWIK1
(canal potassique à deux domaines P ou K2P). Ce canal serait impliqué dans les processus
fréquentiels et sécrétoires de certaines cellules neurales (Buckler, 2015; Ryoo and Park, 2016)
et semble jouer un rôle important dans la régulation du tonus musculaire vasculaire lisse
(Olschewski et al., 2006; Tang et al., 2009).
Nos sultats o t e t gale e t l’appa itio des t a s its oda t pou le a al
calcique CaV1.2 responsable du courant ICaL essentiel pour la phase du plateau (phase 3) du
pote tiel d’a tio a dia ue. De e ue les t a s its oda ts pou le a al CaV1.3 ; un
canal responsable du courant ICaL neuronal et qui serait également impliqué dans la

Discussion
217
g atio de l’a ti it pa e aker cardiaque chez la souris (Mangoni et al., 2003; Zhang et
al., 2005). D’aut es t a s its, oda t pou le a al al i ue CaV3.2 dont le courant (ICaT) est
responsable d’u e pa tie de la phase le te de d pola isatio du pote tiel d’a tio si usale
apparaissent au cours de la différenciation cardiaque. L’appa itio de l’e p essio de es
canaux calciques (de type L et T) est corrélée avec celle des sous-unités régulatrices
importantes pour la modulation de leur activité.
Grajales et ses collègues ont montré que durant la différenciation myogénique des
MSC murines, les transcrits codant pour les canaux calciques CaV1.2 étaient induits alors que
les transcrits codant pour les canaux CaV3.2 ne présentaient aucune variation (Grajales et al.,
2015). Il faut mentionner que ces conductances calciques joueraient un rôle important dans
les p op i t s ellulai es des ellules sou hes. E effet, l’i hi itio des a au al i ues de
type L par la nifédipine, diminuait la prolifération et la différenciation cardiaque des CSCs de
souris (Hotchkiss et al., 2014).
La différenciation cardiaque in vitro de notre modèle de CSCs W8B2+ est aussi
accompagnée de l’appa itio des t a s its oda ts pou HCN isofo e eu o ale et HCN
(isoforme cardiaque) et d’une surexpression de HCN2 et HCN3. Ces canaux HCN, notamment
le HCN4 so t à l’o igi e des ou a ts If espo sa les de l’a ti it th i ue a dia ue des
cellules nodales. Plusieurs études réalisées sur les CSMs hu ai es et hez d’aut es esp es,
ont montré clairement que la différenciation en cellules pacemaker fonctionnelles était
corrélée avec la surexpression génique des isoformes HCN1 ou HCN4 (Bruzauskaite et al.,
2016; Lu et al., 2013; Zhou et al., 2013).
Concernant les canaux potassiques rectifiant entrants non dépendants du temps, nous
a o s o se l’appa itio des t a s its Ki . oda t pou le a al esponsable du courant
potassique IK1, e tifia t e t a t. Ce ou a t est fo da e tal puis u’il o ditio e le
potentiel dit de « repos » à u e aleu p o he du pote tiel d’ uili e au potassiu . D’aut es
t a s its oda t pou des a au potassi ues e tifia t e t a ts se si les à l’a t l holi e
Ki . et se si les à l’ATP Ki . appa aisse t gale e t au ou s de la différenciation des
CSCs W8B2. Les canaux Kir3.x ont une localisation nodale et atriale et sont impliqués dans la
régulation du rythme cardiaque via les récepteurs muscariniques de type M2 (Corey et al.,
1998; Krapivinsky et al., 1995). Quant aux canaux Kir6.x couplés aux récepteurs aux
sulfonylurés (SUR2A ; qui est maintenu), ils sont fortement impliqués dans la régulation de la
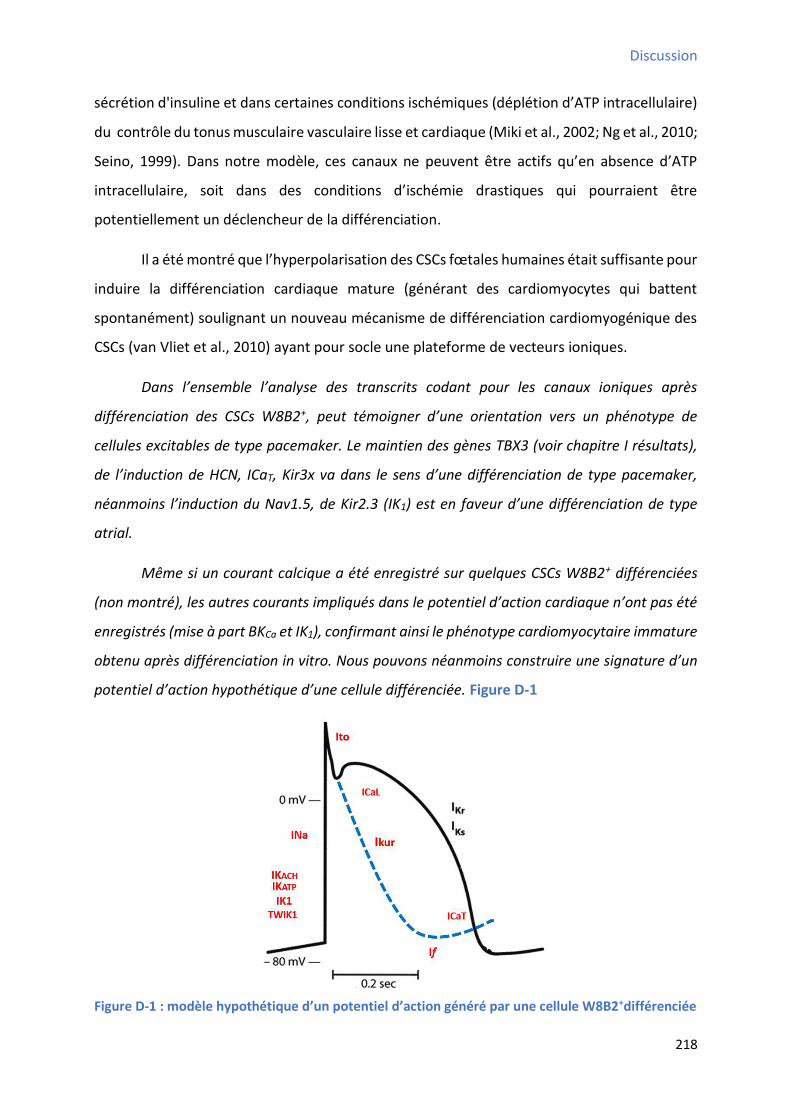
Discussion
218
s tio d'i suli e et da s e tai es o ditio s is h i ues d pl tio d’ATP i t a ellulai e
du contrôle du tonus musculaire vasculaire lisse et cardiaque (Miki et al., 2002; Ng et al., 2010;
Seino, 1999). Da s ot e od le, es a au e peu e t t e a tifs u’e a se e d’ATP
i t a ellulai e, soit da s des o ditio s d’is h ie d asti ues ui pou aie t t e
potentiellement un déclencheur de la différenciation.
Il a t o t ue l’h pe pola isatio des C“Cs fœtales hu ai es tait suffisa te pou
induire la différenciation cardiaque mature (générant des cardiomyocytes qui battent
spontanément) soulignant un nouveau mécanisme de différenciation cardiomyogénique des
CSCs (van Vliet et al., 2010) ayant pour socle une plateforme de vecteurs ioniques.
Da s l’e se le l’a al se des t anscrits codant pour les canaux ioniques après
différenciation des CSCs W8B2+, peut t oig e d’u e o ie tatio ve s u ph ot pe de
cellules excitables de type pacemaker. Le maintien des gènes TBX3 (voir chapitre I résultats),
de l’i du tio de HCN, ICaT, Ki va da s le se s d’u e diff e iatio de t pe pa e ake ,
a oi s l’i du tio du Nav .5, de Ki . IK1 est e faveu d’u e diff e iatio de t pe
atrial.
Même si un courant calcique a été enregistré sur quelques CSCs W8B2+ différenciées
(non o t , les aut es ou a ts i pli u s da s le pote tiel d’a tio a dia ue ’o t pas t
enregistrés (mise à part BKCa et IK1), confirmant ainsi le phénotype cardiomyocytaire immature
obtenu après différenciation in vitro. Nous pouvons néanmoins construire u e sig atu e d’u
pote tiel d’a tio h poth ti ue d’u e ellule diff e i e. Figure D-1
Figure D-1 : od le h poth ti ue d’u pote tiel d’a tio g pa u e ellule W B +différenciée

Discussion
219
3 Oscillations calciques au cours de la différenciation cardiaque in vitro des CSCs W8B2+
E plus de l’appa itio de l’e p essio des transcrits codant pour les canaux calciques
de t pe L et T , os sultats o t o t u ha ge e t da s l’e p essio des a teu s de
l’ho ostasie al i ue i pli u s da s le ouplage excitation-contraction au cours de la
différenciation des CSCs W8B2+. E effet, ous a o s o se u e appa itio de l’e p essio
des transcrits codant pour la pompe Na+/K+, une surexpression des transcrits codant pour la
“ERCA, l’ ha geu NCX et les epteu s à l’IP et u e fo te sous-expression de la CALM3. En
pa all le, l’i age ie al i ue via l’utilisatio de la so de al i ue GCaMP a l des
ha ge e ts sig ifi atifs da s la f ue e, l’a plitude, l’ai e, le TTP, le TTR et les pe tes des
oscillatio s al i ues o se es au ou s de ette diff e iatio a dia ue. De plus, l’ tude
pharmacologique réalisée sur les CSCs après différenciation a permis de déterminer le rôle
ajeu de l’ ha geu NCX da s la g atio de es os illatio s. Les sultats de l’i hi itio
de e tai s a teu s al i ues sugg e t ue es os illatio s so t g es d’u e pa t pa les
canaux calciques CaV de t pe L et les epteu s à l’IP espo sa les de l’aug e tatio
al i ue i t a ellulai e et d’aut e pa t pa la “ERCA pour la recapture calcique responsable de
la baisse de la concentration calcique intracellulaire. Ces oscillations apparaissent insensibles
à la ryanodine, un constat logique car les transcrits qui la codent sont absents dans les CSCs
non différenciées et après différenciation cardiaque.
Dans les cellules souches les oscillations calciques ont été enregistrées et analysées
principalement dans les CSMs humaines (Orciani et al., 2010 ; Kawano et al., 2006 ; Kawano
et al., 2003) . Elles ont été néanmoins obse es et a a t is es da s d’aut es t pes
cellulaires comme les fibroblastes cardiaques humains (Chen et al., 2010), les cellules
endothéliales aortiques et cérébrovasculaires humaines (Hu et al., 2002; Scharbrodt et al.,
2009) et les lymphocytes (Lewis, 2003). Toutefois, leurs rôles dans ces différents types
cellulaires restent à élucider.
Il est i po ta t aussi de sig ale le a ue uel de do es su l’ olutio et le ôle
de ces oscillations lors de processus de différenciation notamment cardiaque.
Au cours de la différenciation cardiaque in vitro des CSCs W8B2+, nous avons observé
u e di i utio de la f ue e et u e aug e tatio de l’a plitude des os illatio s al i ues
ce qui pourrait suggérer un rôle de ces oscillations dans le processus de différenciation. Une
étude sur la différenciation ostéogénique des CSMs humaines a rapporté également une

Discussion
220
diminution de la fréquence des oscillations calciques au cours de la différenciation mais aucun
ha ge e t o e a t l’a plitude (Sun et al., 2007). Dans ce travail, la différenciation
ostéogénique des CSMs conjuguée à la réduction de la fréquence des oscillations (induites par
sti ulatio le t i ue aug e tait l’effi a it de la diff e iatio , e ui sugg e u ôle
important de la fréquence dans ce p o essus. D’aut e pa t, il a t o t ue des os illatio s
calciques apparaissent dans les cellules souches hépatiques de rat et les MSC humaines quand
celles-ci sont mises en culture avec des cardiomyocytes de rats néonataux (Muller-Borer et
al., 2012, 2007). Ces oscillations dans ces cellules souches sont synchrones avec celles des
a dio o tes et p o o ue t l’a uisitio d’u ph ot pe a dia ue pa l’i du tio
transcriptomique de gènes cardiaques. Dolmetsch et ses collègues indiquent que la fréquence
des os illatio s al i ues gulait diff e e t l’e p essio g i ue et seul NF-kB répondait
aux oscillations de faibles fréquences (Dolmetsch et al., 1998).
Nos résultats montrent une sous-expression significative des transcrits codants pour la
calmoduline 3 supposant son a se e d’implication durant le processus de différenciation
cardiaque.
Cepe da t, au u ha ge e t d’e p essio des t a s its oda ts pou la al i eu i e ’a
été observé. La calmoduline (CaM) et la calcineurine font partie des principaux médiateurs
des effets moléculaires du calcium en activant plusieurs cibles moléculaires comme NF-AT,
NF-KB, CaMKII. La odulatio de l’e p essio sp ifi ue de fa teu s de t a s iptio (Smedler
and Uhlén, 2014) serait en relation avec les paramètres cinétiques, d’a plitude et de
f ue e de l’ho loge al i ue i t i s ue.
Le facteur de transcription NFAT activé par la calcineurine jouerait le rôle de décodeur des
oscillations calciques (Dolmetsch et al., 1998). Par exemple, dans les cellules endothéliales
humai es de la ei e du o do , la f ue e, l’a plitude et la du e des os illatio s al i ues
gulaie t l’a ti it du fa teu de t a s iptio NF-KB (Song et al., 2012). Ce facteur de
transcription est activé seule e t si l’a plitude des os illatio s attei t u e aleu seuil. “o
activité est plus intense au fur et à mesure que la fréquence et/ou la durée des oscillations
augmentent. Plusieurs études ont également souligné que la fréquence et la durée des
oscillatio s gulaie t l’a ti it de NFAT da s plusieu s od les ellulai es (Onohara et al.,
2006; Tomida et al., 2003; Uhlén et al., 2006). Des travaux récents indiquent (via un contrôle
des oscillations calciques par optogénétique) que le facteur de transcription NFAT est aussi

Discussion
221
se si le au a iatio s de l’ai e des os illatio s al i ues (Hannanta-Anan and Chow, 2016).
L’e se le des données disponibles montre que NFAT est activé par les oscillations calciques
a a t u e f ue e de à Hz et d’u e du e de , à s (Smedler and Uhlén, 2014).
Le facteur de transcription NF-KB (activé par la calcineurine) connu comme étant
se si le à la f ue e, la du e et l’ai e des os illatio s al i ues da s plusieu s t pes
cellulaires (Hu et al., 1999; Jin et al., 2006; Scharbrodt et al., 2009; Zhu et al., 2008, 2011).
NF-KB se ait a ti pa les os illatio s al i ues a a t u e f ue e de à Hz et d’u e
durée de 36 à 52 s (Smedler and Uhlén, 2014).
L’a ti atio de la al iu al oduli e ki ase de t pe II CaMKII est aussi sensible aux
oscillations calciques (De Koninck and Schulman, 1998; Dupont et al., 2003). Elle est activée
par les oscillations calciques ayant une fréquence de 100 à 4000 mHz et une durée de 200 ms
(Smedler and Uhlén, 2014).
La protéase calpaïne a été également considérée comme un décodeur calcique (Tompa
et al., 2001) et activée par les oscillations calciques ayant une fréquence de 1 à 50 Hz et une
durée de 20 ms (Smedler and Uhlén, 2014).
Durant la différenciation des cellules souches en cardiomyocytes, la voie MAPK
(Mitogen-activated protein kinases) joue un rôle central (Grajales et al., 2010; Park et al., 2009;
Tonelli et al., 2012). Elle serait également sensible aux oscillations calciques (Lockyer et al.,
2001; Tognon et al., 1998; Walker et al., 2004). L’a ti atio de ette oie via la
phosphorylation de ERK) est régulée par des oscillations calciques ayant une fréquence de 1,7
à 17 mHz et une durée approximative de 50 s (Kupzig et al., 2005). Ces résultats ont été
confirmés dans une étude de modélisation théorique montrant une corrélation négative entre
la di i utio de la f ue e os illatoi e et l’a ti atio de la oie MAPK (Yi et al., 2011).
Il est très probable que les changements dans les paramètres des oscillations calciques
f ue e, a plitude, du e, ai e … o se v es ap s diff e iatio de ot e od le de CSCs
W8B2+ soie t li s à l’a uisitio du ph ot pe a dia ue i atu e o se v .
De plus, les changements des paramètres des oscillations calciques pourraient bien agir sur
des a teu s ol ulai es pou odule l’e p essio de g es a dia ues. Co pte te u des
données bibliographiques, la voie MAPK serait un décodeur potentiel des changements de

Discussion
222
fréquence et de durée des oscillations observées dans notre modèle de CSCs W8B2+
différenciées. Dans ces dernières, la fréquence est de 5 mHz et la durée est de 92 s, des valeurs
proches de celles auxquelles la MAPK est sensible (Smedler and Uhlén, 2014). De plus, nous
observons une diminution de la fréquence au cours de la différenciation cardiaque, des
sultats se la les à l’ tude de od lisatio th o i ue (Yi et al., 2011) selon laquelle la
di i utio de la f ue e os illatoi e et li e à l’a tivatio de la voie MAPK.
Selon Smedler et Uhlén, la MAPK est également sensible aux oscillations dans les
cardiomyocytes d’auta t plus ue ette oie est i pli u e da s la diff e iatio a dia ue.
Il est très probable que les facteurs de transcription NFAT et NF-KB (régulés par la calcineurine)
ne seraient pas des décodeurs des oscillations observées dans les W8B2+ différenciées car
l’e p essio des t a s its oda t pou la al i eu i e e ha ge t pas au ou s de la
différenciation. De même, la CaMKII ne serait pas directement impliquée car les transcrits
codant pour la calmoduline se trouvent sous-exprimés au cours de la différenciation. De plus,
la alpaï e, la CaMKII ’est pas se si le au valeu s de f ue e et de du e o se v es su les
CSCs W8B2+ différenciées.
Il est i po ta t de ote u’e plus de so ôle esse tiel da s la p olif atio et l’auto-
renouvellement des CSCs W8B2+, le canal potassique activé par le calcium BKCa semble jouer
u ôle da s la diff e iatio . L’i hi itio du a al BKCa diminuait drastiquement les
pa a t es des os illatio s al i ues f ue e, a plitude, du e… o se es su les C“Cs
W8B2+ diff e i es. D’aut e pa t, les os illatio s al i ues spo ta es i duise t des
fluctuations du potentiel membranaire via le canal potassique IKCa (Kawano et al., 2003) et
est associées à la translocation nucléaire de NFAT (Kawano et al., 2006). Une autre étude a
o t ue le pote tiel e a ai e gulait les os illatio s al i ues o t ôl es pa l’IP
(Kukuljan et al., 1994) suggérant une implication du canal BKCa dans la régulation ce ces
oscillations.

Discussion
223
4 La sphingosine 1-phosphate (S1P) joue un rôle important dans les CSCs W8B2+
La signalisation par les récepteurs à la S1P (S1PR) joue un rôle important dans la
gulatio des p op i t s fo da e tales p olif atio , diff e iatio … des ellules
cardiaques (Means and Brown, 2009). Cepe da t au u e tude à l’heu e a tuelle su les C“Cs
et notamment sur les CSCs W8B2+ n’a appo t des effets de la “ P. Nos sultats o t o t
sur ce modèle de cellules souches cardiaques, l’e p essio t a s ipto i ue des epteu s
S1PR1, S1PR2 et S1PR3 (avec une prédominance du S1PR2). Ce p ofil d’e p essio est
semblable à celui des MSC humaines de la moelle osseuse (Kong et al., 2014; Quint et al.,
2013). Da s le œu hu ai , Ah ed et ses oll gues o t ide tifi l’e p essio des “ PR ,
S1PR2 et S1PR3 au niveau transcriptomique et S1PR1 et S1PR3 au niveau protéique, un profil
quasi identique, da s les œu s de ats (Ahmed et al., 2017).
Dans notre étude, l’ajout de la “ P à µM, u e o e t atio plas ati ue
physiologique) diminue significativement la prolifération des CSCs W8B2+. Cependant
l’a al se du le ellulai e ap s et jou s de t aite e t à la “ P e o t e pas de
différence dans les phases du cycle comparé au contrôle contrairement à la paxilline et
l’i ioto i e. De plus, l’i hi itio pha a ologi ue sp ifi ue des epteu s “ PR , “ PR
et S1PR3 (candidats moléculaires pote tiels e ous a pas pe is d’ide tifie le ou les
epteu s i pli u s da s l’effet a tip olif atif di pa la “ P.
Il est important de signaler que la majorité des données de la littérature rapporte un
effet pro-prolifératif de la S1P et ceci dans plusieurs modèles cellulaire tels que les fibroblastes
cardiaques (via le S1PR3) (Benamer et al., 2011), les cellules satellites (via le S1PR2 et le S1PR3)
(Calise et al., 2012; Donati et al., 2013), les cellules endothéliales (via le S1PR1 et le S1PR3)
(Kimura et al., 2000) et les CSMs humaines du chorion (Innamorati et al., 2017).
Cependant, la S1P peut avoir un effet antiprolifératif notamment dans les lymphocytes
T humaines (Jin et al., 2003), les myofibroblastes hépatiques humains (Davaille et al., 2000),
les hépatocytes (Ikeda et al., 2003) et dans les cellules thyroïdiennes de rats (Björklund et al.,
2005). Da s les l pho tes T, le a is e ’a pas t tudi ais l’effet a tip olif atif
dans les hépatocytes de rats serait médié par le S1PR2 (via la voie Rho) (Ikeda et al., 2003).
Les auteurs ont supposé que la S1P est un régulateur négatif de la régénération hépatique.
Da s les ofi o lastes h pati ues hu ai s, l’effet a tip olif atif de la “ P sti ulait la oie
de l’AMP suite à u e aug e tatio du taux des protaglandine E2 (Davaille et al., 2000). Dans

Discussion
224
les cellules thyroïdiennes de rats, la S1P augmente la concentration de calcium intracellulaire
(via la phoshoplipase C et l’IP et de l’AMP . D’aut e pa t, la “ P p o o ue u e aug e tatio
transitoirement des transcrits codant pour son propre récepteur S1PR2 (Björklund et al.,
. De plus, Ji et ses oll gues o t o t ue l’a ti atio des l pho tes T par des
stimuli chimiques i pa tait o sid a le e t l’e p essio des “ PR Ji et al., 2003).
L’e se le de es do es o t e t lai e e t ue l’effet a tip olif atif de la S P est
di pa le S PR faisa t i te ve i le al iu . Le ha ge e t de l’e p essio des S PR pa
la S1P elle-même ajoute u aut e iveau d’auto-régulation par la S1P.
Il a été montré récemment que la S1P pouvait être secrétée par les CSMs de souris
(Sassoli et al., 2014) et humaines dérivées des iPS (Du et al., 2017), ajouta t ai si d’aut es
niveaux de complexité de régulation para et autocrine. En effet, cette S1P secrétée par les
CSMs u i es tait apa le d’i dui e la p olif atio des cellules souches satellites et des
cellules myoblastiques C2C12 (Sassoli et al., 2014). Quant aux MSC humaines dérivées des iPS,
les auteurs observent pour la première fois que la sécrétion de la S1P passait par un processus
exosomique via la membrane plasmique des hépatocytes (Du et al., 2017). Ces exosomes
taie t apa les d’i dui e la p olif atio des h pato tes p o a le e t e i duisa t la
synthèse et la libération de la S1P intracellulaire.
La S1P agit principalement via ses cinq récepteurs mais une voie intracellulaire
déclenchée indépendamment des récepteurs est de plus en plus décrite (Dupont et al., 2003;
Payne et al., 2002). Par exemple, Xiu et ses collègues ont montré que la S1P intracellulaire
i duit l’e p essio du ollag e pa les ofi o lastes lo s de la fi ose h pati ue,
indépendamment de l’a ti atio de ses récepteurs (Xiu et al., 2015).
Ces données viennent encore une fois ajouter un autre niveau de régulation
indépendant de la signalisation médiée par les récepteurs à la S1P.
Nos sultats o t gale e t o t ue la “ P di i ue d asti ue e t l’auto-
e ou elle e t, u a is e de di isio a e p se atio de l’ tat sou he et ui peut t e
odul pa l’e i o e e t (He et al., 2009). L’i hi itio des “ PR1 et S1PR2 dans notre
modèle de CSCs W8B2+, a o t ue ette p essio de l’auto- e ou elle e t ’est pas
médiée par les récepteurs S1PR1 et S1PR2.

Discussion
225
Il a été montré que la S1P joue un rôle très important dans la régulation des cellules
souches neurales (CSNs) de souris (Codega et al., 2014). Ces CSNs se trouvent dans deux états ;
un état quiescent (qCNSs) et un état activé (aCNSs). Quand elles sont activées, les CNSs
deviennent hautement prolifératives. Ces chercheurs ont montré que la S1P agit en diminuant
l’a ti atio des CN“s sa s alt e le o e des aCN“s alo s ue les p ostagla di es D
di i uaie t à la fois l’a ti atio des CN“s et le o e des aCNSs (Codega et al., 2014). Ces
travaux soulignent un rôle important de la S1P dans la physiologie des cellules souches.
Une autre étude décrite sur les MSC humaines du chorion a montré que la S1P
di i uait l’auto-renouvellement seulement si elle est combinée avec des agents qui
aug e te t la o e t atio i t a ellulai e de l’AMP (Innamorati et al., 2017).
Ces do es de la litt atu e sugg e t ue la S P gule gative e t l’auto-
renouvellement des cellules souches en maintenant leur état quiescent ou en induisant leur
engagement vers un processus de différenciation.
Enfin, nos résultats préliminaires ont montré un effet plutôt anti-migratoire de la S1P
dans les CSCs W8B2+.
La migration joue un rôle essentiel dans la régulation des propriétés des cellules
souches. En effet, la majorité des cellules souches est retenue dans une niche mais peuvent la
quitter en réponse à des stimuli spécifiques, un processus appelé « mobilisation ». Les cellules
souches sont également capables de regagner la niche par un processus opposé nommé
domiciliation ou « homing ». Il s’a e ue la “ P est u sti ulus t s i po ta t qui régule la
mobilisation de différents types cellulaire comme les CSMs, les progéniteurs endothéliaux, les
CSHs etc. (Ratajczak et al., 2014).
Le rôle de la S1P dans la mobilisation des CSHSs a été bien étudié et les études ont
soulig l’i po ta e du g adie t “ P e t e le sa g, la l phe et le tissu da s la gulatio
de la mobilisation de ces cellules (Golan et al., 2013; Liu et al., 2011).
L’e se le des do es de la litt atu e su l’effet de la “ P su la ig atio ellulai e in vitro
montre des effets pro et anti- ig atoi es. L’effet p o-migratoire a été signalé par exemple
dans les cellules endothéliales humaines (probablement via S1PR1 et S1PR3) (Kimura et al.,
2000), les cellules satellites de souris (Calise et al., 2012), les CSMs humaines de la moelle
osseuse (via S1PR1 et S1PR3)(Kong et al., 2014) et les fibroblastes cardiaques de souris (via le

Discussion
226
S1PR3) Be a e et al., . Qua t à l’effet a ti-migratoire, la S1P inhibait la migration des
CSMs humaines du chorion (via l’aug e tatio de l’AMP I a o ati et al., , les
cellules musculaires lisses bronchiques humaines (via S1PR2)(Kawata et al., 2005), les
myoblastes C2C12 (via S1PR2) (Becciolini et al., 2006) et les p u seu s d’oligode d o tes
de rat (via S1PR5) (Novgorodov et al., 2007).
E su , l’e se le de os sultats o t e u e e p essio des t a s its oda t
pour S1PR1, S1PR2 et S1PR3 avec une prédominance du S1PR2 dans notre modèle de CSCs
W8B2+. De plus dans ce modèle cellulaire, la S1P régule négativement la prolifération et/ou
l’auto-renouvellent et la migration. Les données de la littérature suggèrent que la S1P
augmenterait la quiescence (cellules en phase G0, sorties du le ellulai e ou l’e gage e t
des CSCs W8B2+ dans une voie de différenciation.
Quant au mécanisme, les données de la littérature nous orientent vers un effet de la
S1P médié par le S1PR2 faisant intervenir le Ca2+ et l’AMP . Ce i peut e ett e e uestion la
sp ifi it de so i hi itio S PR pa l’age t JTE . E effet, l’i hi itio du S PR pa le
JTE s’est v l e i effi a e (Salomone and Waeber, 2011) et su tout su l’aug e tatio du
calcium intracellulaire (Long et al., 2010).
Si o , d’aut es a is es e : autorégulation par la S1P de ses propres récepteurs,
sécrétion de la S1P par les CSCs W8B2+, signalisation intracellulaire etc.) peuvent être à
l’origine des effets observés de la S1P dans notre modèle.

Conclusion générale et perspectives
227
CONCLUSION GENERALE ET PERSPECTIVES
Nous a o s pu isole et pu ifie à pa ti de iopsies de œu hu ai des ellules
d’o igi e se h ateuse e p i a t le a ueu W B2, ayant des caractéristiques de
cellules souches. Ces clones apparaissent homogènes de par leur génotype et leur phénotype.
L’a al se glo ale de l’e p essio g i ue e pa ti ulie des a au io i ues, i di ue
que les CSCs W8B2+ possèdent un phénotype non-e ita le o e ’est le as pou les aut es
types de cellules souches. Ces cellules expriment quelques courants potassiques, notamment
le canal BKCa ui s’a e esse tiel pou gule leu s p op i t s fo da e tales o e la
p olif atio et l’auto-renouvellement. De plus, ces cellules présentent une signature calcique
ou a ti it al i ue i t a ellulai e p o a le e t li e à l’ho loge al i ue de t pe IP et au
conductances potassiques calcium-dépendantes de type BKCa. L’a al se t a s ipto i ue
mont e ue es ellules peu e t s’o ie te e s u ph ot pe a dia ue de t pe odal et/ou
auriculaire immature quand elles sont différenciées in vitro (figure C-1).
Cette diff e iatio s’a o pag e d’i po ta ts ha ge e ts de l’a ti it al i ue et
peut s’appa e te à l’appa itio d’u e ho loge al i ue e a ai e gulat i e de l’ho loge
calcique interne. Nos expérimentations montrent que la différenciation in vitro, par analogie
à la différenciation des cellules souches pluripotentes induites, ne peut en aucun cas conduire
à l’o te tio de a dio o tes t pi ues. Nous o t o s gale e t ue la diff e iatio in
vitro ne semble pas la meilleure orientation pour la réalisation de greffe cellulaire cardiaque.
Les approches génomiques et fonctionnelles des progéniteurs cardiaques semblent les plus
pertinentes pour en intégrant la notion de niches cardiaques et les processus
autocrine/paracrine liés à leur environnement multicellulaire. La cible principale apparait
o e ta t l’ho loge al i ue i te e p i itive intégrant tous les facteurs de régulation.
Comme par exemple les processus inflammatoires, hormonaux, ischémiques, mécaniques, de
o ta tes… sus epti les d’ eille es p og iteu s pou u e pa atio tissulai e op a te.
Dans ce cadre et comme exemple, les CSCs W8B2+ sont particulièrement sensibles à la
sphingosine 1-phosphate (S1P), un lipide plasmatique extrêmement important qui agit sur ces
ellules e di i ua t leu s apa it s de p olif atio , d’auto-renouvellement et de migration.

Conclusion générale et perspectives
228
Dans son e se le e t a ail appo te d’i po ta tes i fo atio s su la ph siologie
des ellules sou hes a dia ues, ota e t su l’i pli atio des a au io i ues et l’a ti it
dynamique calcique intrinsèque dans la régulation de leurs propriétés.
Néanmoins, cette étude doit être complétée par :
U e tude plus app ofo die de l’a ti it al i ue spo ta e des ellules sou hes
W B pa u e a a t isatio pha a ologi ue, afi d’ide tifie les a teu s i pli u s
da s la g atio de leu l’ho loge ou os illateu al ique.
L’a al se à l’ helle p ot i ue des a au io i ues et a teu s al i ues do t
l’e p essio ha ge au ou s de la diff e iatio o e pa e e ple les a au de
la K2P et TRP, ASIC, les récepteurs purinergiques ou SOCs ...
Une optimisation du protocole de différenciation cardiaque de type chimique en
ajustant notre mode de stimulation par optogénétique ou par des protocoles de
différenciation plus performants (par exemple en 3 dimensions ou en mimant le
microenvironnement cardiaque).
Une étude plus poussée pour décrypter au niveau moléculaire les cibles impliquées
dans les modifications des paramètres des oscillations calciques au cours de la
différenciation.
Un examen plus approfondi du ode d’a tio de la “ P ui peut t e u odulateu
négatif de la régénération cardiaque en impactant les propriétés fondamentales des
C“Cs. Plusieu s poi ts doi e t t e solus pou d te i e le a is e d’a tio de
ce lipide (expression protéique des récepteurs, étude de la quiescence, inhibition plus
ciblée des ré epteu s et …
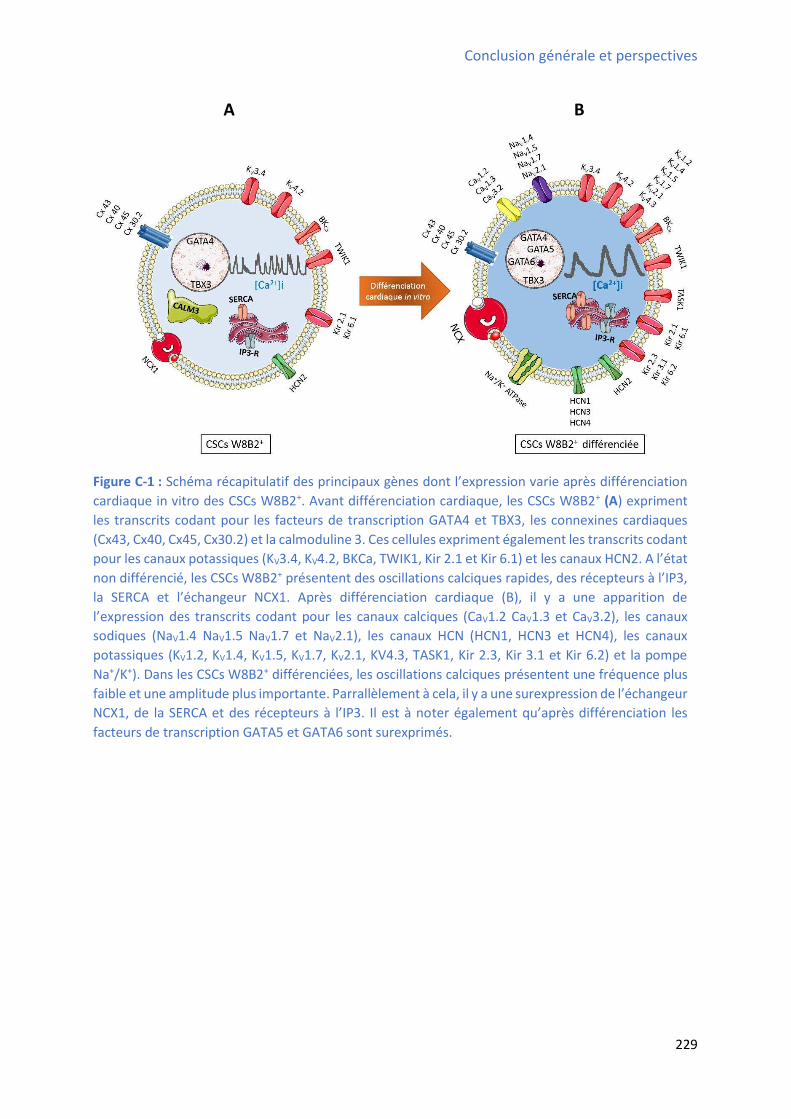
Conclusion générale et perspectives
229
Figure C-1 : Sch a apitulatif des p i ipau g es do t l’e p essio a ie ap s diff e iatio cardiaque in vitro des CSCs W8B2+. Avant différenciation cardiaque, les CSCs W8B2+ (A) expriment
les transcrits codant pour les facteurs de transcription GATA4 et TBX3, les connexines cardiaques
(Cx43, Cx40, Cx45, Cx30.2) et la calmoduline 3. Ces cellules expriment également les transcrits codant
pour les canaux potassiques (KV3.4, KV4.2, BKCa, TWIK1, Kir 2.1 et Kir 6.1) et les canau HCN . A l’ tat non différencié, les CSCs W8B2+ p se te t des os illatio s al i ues apides, des epteu s à l’IP , la “ERCA et l’ ha geu NCX . Ap s diff e iatio a dia ue B , il a u e appa itio de l’e p essio des t a s its oda t pou les canaux calciques (CaV1.2 CaV1.3 et CaV3.2), les canaux
sodiques (NaV1.4 NaV1.5 NaV1.7 et NaV2.1), les canaux HCN (HCN1, HCN3 et HCN4), les canaux
potassiques (KV1.2, KV1.4, KV1.5, KV1.7, KV2.1, KV4.3, TASK1, Kir 2.3, Kir 3.1 et Kir 6.2) et la pompe
Na+/K+). Dans les CSCs W8B2+ différenciées, les oscillations calciques présentent une fréquence plus
faible et u e a plitude plus i po ta te. Pa all le e t à ela, il a u e su e p essio de l’ ha geu NCX , de la “ERCA et des epteu s à l’IP . Il est à ote gale e t u’ap s diff e iatio les facteurs de transcription GATA5 et GATA6 sont surexprimés.
A B

Références bibliographiques
230
REFERENCES BIBLIOGRAPHIQUES
Abdelalim, E.M., Takada, T., Toyoda, F., Omatsu-Kanbe, M., Matsuura, H., Tooyama, I., and Torii, R. (2006). In vitro expression of natriuretic peptides in cardiomyocytes differentiated from monkey embryonic stem cells. Biochem. Biophys. Res. Commun. 340, 689–695.
Adewumi, Adewumi, O., Aflatoonian, B., Ahrlund-Richter, L., Amit, M., Andrews, P.W., Beighton, G., Bello, P.A., Benvenisty, N., Berry, L.S., et al. (2007). Characterization of human embryonic stem cell lines by the International Stem Cell Initiative. Nat. Biotechnol. 25, 803–816.
Afroze, T., Yang, L.L., Wang, C., Gros, R., Kalair, W., Hoque, A.N., Mungrue, I.N., Zhu, Z., and Husain, M. (2003). Calcineurin-independent regulation of plasma membrane Ca2+ ATPase-4 in the vascular smooth muscle cell cycle. Am. J. Physiol. Cell Physiol. 285, C88-95.
Agbulut, O., Menot, M.-L., Li, Z., Marotte, F., Paulin, D., Hagège, A.A., Chomienne, C., Samuel, J.-L., and Menasché, P. (2003). Temporal patterns of bone marrow cell differentiation following transplantation in doxorubicin-induced cardiomyopathy. Cardiovasc. Res. 58, 451–459.
Aguiar, A.M. de, Kuligovski, C., Costa, M.T.B.A. da, Stimamiglio, M.A., Rebelatto, C.L.K., Senegaglia, A.C., Brofman, P.R.S., Dallagiovanna, B., Goldenberg, S., and Correa, A. (2011). Alkaline phosphatase-positive cells isolated from human hearts have mesenchymal stem cell characteristics. Stem Cell Discov. 01, 71.
Ahmed, N., Linardi, D., Decimo, I., Mehboob, R., Gebrie, M.A., Innamorati, G., Luciani, G.B., Faggian, G., and Rungatscher, A. (2017). Characterization and Expression of Sphingosine 1-Phosphate Receptors in Human and Rat Heart. Front. Pharmacol. 8.
Al Attar, N., Carrion, C., Ghostine, S., Garcin, I., Vilquin, J.-T., Hagège, A.A., and Menasché, P. (2003). Long-term (1 year) functional and histological results of autologous skeletal muscle cells transplantation in rat. Cardiovasc. Res. 58, 142–148.
Alewijnse, A.E., Peters, S.L.M., and Michel, M.C. (2004). Cardiovascular effects of sphingosine-1-phosphate and other sphingomyelin metabolites. Br. J. Pharmacol. 143, 666–684.
Alexander, D., Rieger, M., Klein, C., Ardjomandi, N., and Reinert, S. (2013). Selection of osteoprogenitors from the jaw periosteum by a specific animal-free culture medium. PloS One 8, e81674.
Alfarano, C., Roubeix, C., Chaaya, R., Ceccaldi, C., Calise, D., Mias, C., Cussac, D., Bascands, J.L., and Parini, A. (2012). Intraparenchymal injection of bone marrow mesenchymal stem cells reduces kidney fibrosis after ischemia-reperfusion in cyclosporine-immunosuppressed rats. Cell Transplant. 21, 2009–2019.
Allende, M.L., Yamashita, T., and Proia, R.L. (2003). G-protein-coupled receptor S1P1 acts within endothelial cells to regulate vascular maturation. Blood 102, 3665–3667.
Al-Maqtari, T., Hong, K.U., Vajravelu, B.N., Moktar, A., Cao, P., Moore, J.B., and Bolli, R. (2017). Transcription factor-induced activation of cardiac gene expression in human c-kit+ cardiac progenitor cells. PloS One 12, e0174242.

Références bibliographiques
231
Altman, J. (1963). Autoradiographic investigation of cell proliferation in the brains of rats and cats. Anat. Rec. 145, 573–591.
Altman, J. (1969). Autoradiographic and histological studies of postnatal neurogenesis. IV. Cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb. J. Comp. Neurol. 137, 433–457.
Altman, J., and Das, G.D. (1965). Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 124, 319–335.
Amoh, Y., Li, L., Campillo, R., Kawahara, K., Katsuoka, K., Penman, S., and Hoffman, R.M. (2005). Implanted hair follicle stem cells form Schwann cells that support repair of severed peripheral nerves. Proc. Natl. Acad. Sci. U. S. A. 102, 17734–17738.
Ang, Y.-S., Rivas, R.N., Ribeiro, A.J.S., Srivas, R., Rivera, J., Stone, N.R., Pratt, K., Mohamed, T.M.A., Fu, J.-D., Spencer, C.I., et al. (2016). Disease Model of GATA4 Mutation Reveals Transcription Factor Cooperativity in Human Cardiogenesis. Cell 167, 1734–1749.e22.
Antonitsis, P., Ioannidou-Papagiannaki, E., Kaidoglou, A., and Papakonstantinou, C. (2007). In vitro cardiomyogenic differentiation of adult human bone marrow mesenchymal stem cells. The role of 5-azacytidine. Interact. Cardiovasc. Thorac. Surg. 6, 593–597.
Anversa, P., Kajstura, J., Leri, A., and Bolli, R. (2006). Life and death of cardiac stem cells: a paradigm shift in cardiac biology. Circulation 113, 1451–1463.
Asahara, T., Murohara, T., Sullivan, A., Silver, M., van der Zee, R., Li, T., Witzenbichler, B., Schatteman, G., and Isner, J.M. (1997). Isolation of putative progenitor endothelial cells for angiogenesis. Science 275, 964–967.
Asumda, F.Z., and Chase, P.B. (2012). Nuclear cardiac troponin and tropomyosin are expressed early in cardiac differentiation of rat mesenchymal stem cells. Differ. Res. Biol. Divers. 83, 106–115.
Attali, B., Wang, N., Kolot, A., Sobko, A., Cherepanov, V., and Soliven, B. (1997). Characterization of delayed rectifier Kv channels in oligodendrocytes and progenitor cells. J. Neurosci. Off. J. Soc. Neurosci. 17, 8234–8245.
Azhar, M., Schultz, J.E.J., Grupp, I., Dorn, G.W., Meneton, P., Molin, D.G.M., Gittenberger-de Groot, A.C., and Doetschman, T. (2003). Transforming growth factor beta in cardiovascular development and function. Cytokine Growth Factor Rev. 14, 391–407.
Bai, X., Ma, J., Pan, Z., Song, Y.-H., Freyberg, S., Yan, Y., Vykoukal, D., and Alt, E. (2007). Electrophysiological properties of human adipose tissue-derived stem cells. Am. J. Physiol. Cell Physiol. 293, C1539-1550.
Bakker, M.L., Boink, G.J.J., Boukens, B.J., Verkerk, A.O., van den Boogaard, M., den Haan, A.D., Hoogaars, W.M.H., Buermans, H.P., de Bakker, J.M.T., Seppen, J., et al. (2012). T-box transcription factor TBX3 reprogrammes mature cardiac myocytes into pacemaker-like cells. Cardiovasc. Res. 94, 439–449.
Balsam, L.B., Wagers, A.J., Christensen, J.L., Kofidis, T., Weissman, I.L., and Robbins, R.C. (2004). Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. Nature 428, 668–673.

Références bibliographiques
232
Barile, L., Messina, E., Giacomello, A., and Marbán, E. (2007). Endogenous cardiac stem cells. Prog. Cardiovasc. Dis. 50, 31–48.
Bayer, S.A. (1982). Changes in the total number of dentate granule cells in juvenile and adult rats: A correlated volumetric and 3H-thymidine autoradiographic study. Exp. Brain Res. 46, 315–323.
Bearzi, C., Rota, M., Hosoda, T., Tillmanns, J., Nascimbene, A., De Angelis, A., Yasuzawa-Amano, S., Trofimova, I., Siggins, R.W., Lecapitaine, N., et al. (2007). Human cardiac stem cells. Proc. Natl. Acad. Sci. U. S. A. 104, 14068–14073.
Bearzi, C., Leri, A., Lo Monaco, F., Rota, M., Gonzalez, A., Hosoda, T., Pepe, M., Qanud, K., Ojaimi, C., Bardelli, S., et al. (2009). Identification of a coronary vascular progenitor cell in the human heart. Proc. Natl. Acad. Sci. U. S. A. 106, 15885–15890.
Becciolini, L., Meacci, E., Donati, C., Cencetti, F., Rapizzi, E., and Bruni, P. (2006). Sphingosine 1-phosphate inhibits cell migration in C2C12 myoblasts. Biochim. Biophys. Acta 1761, 43–51.
Behfar, A., Zingman, L.V., Hodgson, D.M., Rauzier, J.-M., Kane, G.C., Terzic, A., and Pucéat, M. (2002). Stem cell differentiation requires a paracrine pathway in the heart. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 16, 1558–1566.
Belin, S., Kaya, F., Duisit, G., Giacometti, S., Ciccolini, J., and Fontés, M. (2009). Antiproliferative effect of ascorbic acid is associated with the inhibition of genes necessary to cell cycle progression. PloS One 4, e4409.
Beltrami, A.P., Urbanek, K., Kajstura, J., Yan, S.M., Finato, N., Bussani, R., Nadal-Ginard, B., Silvestri, F., Leri, A., Beltrami, C.A., et al. (2001). Evidence that human cardiac myocytes divide after myocardial infarction. N. Engl. J. Med. 344, 1750–1757.
Beltrami, A.P., Barlucchi, L., Torella, D., Baker, M., Limana, F., Chimenti, S., Kasahara, H., Rota, M., Musso, E., Urbanek, K., et al. (2003). Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 114, 763–776.
Benamer, N., Fares, N., Bois, P., and Faivre, J.-F. (2011). Electrophysiological and functional effects of sphingosine-1-phosphate in mouse ventricular fibroblasts. Biochem. Biophys. Res. Commun. 408, 6–11.
Bergmann, O., Zdunek, S., Felker, A., Salehpour, M., Alkass, K., Bernard, S., Sjostrom, S.L., Szewczykowska, M., Jackowska, T., Dos Remedios, C., et al. (2015). Dynamics of Cell Generation and Turnover in the Human Heart. Cell 161, 1566–1575.
Berridge, M.J., Bootman, M.D., and Lipp, P. (1998). Calcium--a life and death signal. Nature 395, 645–648.
Berridge, M.J., Lipp, P., and Bootman, M.D. (2000). The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 1, 11–21.
Bers, D.M. (2002). Cardiac excitation-contraction coupling. Nature 415, 198–205.
Biel, M., Schneider, A., and Wahl, C. (2002). Cardiac HCN channels: structure, function, and modulation. Trends Cardiovasc. Med. 12, 206–212.

Références bibliographiques
233
Bielmann, C., Rignault-Clerc, S., Liaudet, L., Li, F., Kunieda, T., Sogawa, C., Zehnder, T., Waeber, B., Feihl, F., and Rosenblatt-Velin, N. (2015). Brain natriuretic peptide is able to stimulate cardiac progenitor cell proliferation and differentiation in murine hearts after birth. Basic Res. Cardiol. 110, 455.
Biteau, B., Hochmuth, C.E., and Jasper, H. (2008). JNK activity in somatic stem cells causes loss of tissue homeostasis in the aging Drosophila gut. Cell Stem Cell 3, 442–455.
Bjerknes, M., and Cheng, H. (2002). Multipotential stem cells in adult mouse gastric epithelium. Am. J. Physiol. Gastrointest. Liver Physiol. 283, G767-777.
Björklund, S., Palmberg, S., Rask, S., Westerdahl, A.-C., and Törnquist, K. (2005). Effects of sphingosine 1-phosphate on calcium signaling, proliferation and S1P2 receptor expression in PC Cl3 rat thyroid cells. Mol. Cell. Endocrinol. 231, 65–74.
Blackiston, D.J., McLaughlin, K.A., and Levin, M. (2009). Bioelectric controls of cell proliferation: ion channels, membrane voltage and the cell cycle. Cell Cycle Georget. Tex 8, 3527–3536.
Blatt, A., Robinson, D., Cotter, G., Efrati, S., Simantov, Y., Bar, I., Kaluski, E., Krakover, R., Sidenko, S., Evron, Z., et al. (2003). Improved regional left ventricular function after successful satellite cell grafting in rabbits with myocardial infarction. Eur. J. Heart Fail. 5, 751–757.
Bolli, R., Chugh, A.R., D’A a io, D., Lough a , J.H., “todda d, M.F., Ik a , “., Bea he, G.M., Wag e , S.G., Leri, A., Hosoda, T., et al. (2011). Cardiac stem cells in patients with ischaemic cardiomyopathy (SCIPIO): initial results of a randomised phase 1 trial. Lancet Lond. Engl. 378, 1847–1857.
Bollini, S., Smart, N., and Riley, P.R. (2011). Resident cardiac progenitor cells: at the heart of regeneration. J. Mol. Cell. Cardiol. 50, 296–303.
Bond, A.M., Ming, G.-L., and Song, H. (2015). Adult Mammalian Neural Stem Cells and Neurogenesis: Five Decades Later. Cell Stem Cell 17, 385–395.
Boni, A., Urbanek, K., Nascimbene, A., Hosoda, T., Zheng, H., Delucchi, F., Amano, K., Gonzalez, A., Vitale, S., Ojaimi, C., et al. (2008). Notch1 regulates the fate of cardiac progenitor cells. Proc. Natl. Acad. Sci. U. S. A. 105, 15529–15534.
Brand, T., and Schneider, M.D. (1995). The TGF beta superfamily in myocardium: ligands, receptors, transduction, and function. J. Mol. Cell. Cardiol. 27, 5–18.
Brindley, D.N., English, D., Pilquil, C., Buri, K., and Ling, Z.C. (2002). Lipid phosphate phosphatases regulate signal transduction through glycerolipids and sphingolipids. Biochim. Biophys. Acta 1582, 33–44.
Bruzauskaite, I., Bironaite, D., Bagdonas, E., Skeberdis, V.A., Denkovskij, J., Tamulevicius, T., Uvarovas, V., and Bernotiene, E. (2016). Relevance of HCN2-expressing human mesenchymal stem cells for the generation of biological pacemakers. Stem Cell Res. Ther. 7, 67.
Bu, L., Jiang, X., Martin-Puig, S., Caron, L., Zhu, S., Shao, Y., Roberts, D.J., Huang, P.L., Domian, I.J., and Chien, K.R. (2009). Human ISL1 heart progenitors generate diverse multipotent cardiovascular cell lineages. Nature 460, 113–117.
Buckler, K.J. (2015). TASK channels in arterial chemoreceptors and their role in oxygen and acid sensing. Pflugers Arch. 467, 1013–1025.

Références bibliographiques
234
Budde, K., Schmouder, R.L., Brunkhorst, R., Nashan, B., Lücker, P.W., Mayer, T., Choudhury, S., Skerjanec, A., Kraus, G., and Neumayer, H.H. (2002). First human trial of FTY720, a novel immunomodulator, in stable renal transplant patients. J. Am. Soc. Nephrol. JASN 13, 1073–1083.
Bühring, H.-J., Battula, V.L., Treml, S., Schewe, B., Kanz, L., and Vogel, W. (2007). Novel markers for the prospective isolation of human MSC. Ann. N. Y. Acad. Sci. 1106, 262–271.
Bujak, M., and Frangogiannis, N.G. (2007). The role of TGF-beta signaling in myocardial infarction and cardiac remodeling. Cardiovasc. Res. 74, 184–195.
Burger, P.E., Xiong, X., Coetzee, S., Salm, S.N., Moscatelli, D., Goto, K., and Wilson, E.L. (2005). Sca-1 expression identifies stem cells in the proximal region of prostatic ducts with high capacity to reconstitute prostatic tissue. Proc. Natl. Acad. Sci. U. S. A. 102, 7180–7185.
Burridge, P.W., Thompson, S., Millrod, M.A., Weinberg, S., Yuan, X., Peters, A., Mahairaki, V., Koliatsos, V.E., Tung, L., and Zambidis, E.T. (2011). A universal system for highly efficient cardiac differentiation of human induced pluripotent stem cells that eliminates interline variability. PloS One 6, e18293.
Bussolati, G., and Cassoni, P. (2001). Editorial: the oxytocin/oxytocin receptor system-expect the unexpected. Endocrinology 142, 1377–1379.
Cai, C.-L., Liang, X., Shi, Y., Chu, P.-H., Pfaff, S.L., Chen, J., and Evans, S. (2003). Isl1 identifies a cardiac progenitor population that proliferates prior to differentiation and contributes a majority of cells to the heart. Dev. Cell 5, 877–889.
Cai, J., Cheng, A., Luo, Y., Lu, C., Mattson, M.P., Rao, M.S., and Furukawa, K. (2004). Membrane properties of rat embryonic multipotent neural stem cells. J. Neurochem. 88, 212–226.
Calise, S., Blescia, S., Cencetti, F., Bernacchioni, C., Donati, C., and Bruni, P. (2012). Sphingosine 1-phosphate stimulates proliferation and migration of satellite cells: role of S1P receptors. Biochim. Biophys. Acta 1823, 439–450.
Cameron, V.A., Rademaker, M.T., Ellmers, L.J., Espiner, E.A., Nicholls, M.G., and Richards, A.M. (2000). Atrial (ANP) and brain natriuretic peptide (BNP) expression after myocardial infarction in sheep: ANP is synthesized by fibroblasts infiltrating the infarct. Endocrinology 141, 4690–4697.
Campagnoli, C., Roberts, I.A., Kumar, S., Bennett, P.R., Bellantuono, I., and Fisk, N.M. (2001). Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood 98, 2396–2402.
Caplan, A.I. (1986). Molecular and cellular differentiation of muscle, cartilage, and bone in the developing limb. Prog. Clin. Biol. Res. 217B, 307–318.
Carr, C.A., Stuckey, D.J., Tan, J.J., Tan, S.C., Gomes, R.S.M., Camelliti, P., Messina, E., Giacomello, A., Ellison, G.M., and Clarke, K. (2011). Cardiosphere-derived cells improve function in the infarcted rat heart for at least 16 weeks--an MRI study. PloS One 6, e25669.
Cassoni, P., Sapino, A., Munaron, L., Deaglio, S., Chini, B., Graziani, A., Ahmed, A., and Bussolati, G. (2001). Activation of functional oxytocin receptors stimulates cell proliferation in human trophoblast and choriocarcinoma cell lines. Endocrinology 142, 1130–1136.
Castaldo, C., Di Meglio, F., Nurzynska, D., Romano, G., Maiello, C., Bancone, C., Müller, P., Böhm, M., Cotrufo, M., and Montagnani, S. (2008). CD117-positive cells in adult human heart are localized in the

Références bibliographiques
235
subepicardium, and their activation is associated with laminin-1 and alpha6 integrin expression. Stem Cells Dayt. Ohio 26, 1723–1731.
Catterall, W.A. (2011). Voltage-Gated Calcium Channels. Cold Spring Harb. Perspect. Biol. 3.
Catterall, W.A., and Swanson, T.M. (2015). Structural Basis for Pharmacology of Voltage-Gated Sodium and Calcium Channels. Mol. Pharmacol. 88, 141–150.
Chamuleau, S.A.J., van Belle, E., and Doevendans, P.A. (2009). Enhancing cardiac stem cell differentiation into cardiomyocytes. Cardiovasc. Res. 82, 385–387.
Chan, S.S.-K., Chen, J.-H., Hwang, S.-M., Wang, I.-J., Li, H.-J., Lee, R.T., and Hsieh, P.C.H. (2009). Salvianolic acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells. Biochem. Biophys. Res. Commun. 387, 723–728.
Charron, F., and Nemer, M. (1999). GATA transcription factors and cardiac development. Semin. Cell Dev. Biol. 10, 85–91.
Chatelain, F.C., Bichet, D., Douguet, D., Feliciangeli, S., Bendahhou, S., Reichold, M., Warth, R., Barhanin, J., and Lesage, F. (2012). TWIK1, a unique background channel with variable ion selectivity. Proc. Natl. Acad. Sci. U. S. A. 109, 5499–5504.
Che, H., Xiao, G., Sun, H., Wang, Y., and Li, G. (2016). Functional TRPV2 and TRPV4 channels in human a dia ‐kit+ p oge ito ells. J. Cell. Mol. Med. 20, 1118–1127.
Chen, J., Liu, H., Liu, J., Qi, J., Wei, B., Yang, J., Liang, H., Chen, Y., Chen, J., Wu, Y., et al. (2013). H3K9 methylation is a barrier during somatic cell reprogramming into iPSCs. Nat. Genet. 45, 34–42.
Chen, J.-B., Tao, R., Sun, H.-Y., Tse, H.-F., Lau, C.-P., and Li, G.-R. (2010). Multiple Ca2+ signaling pathways regulate intracellular Ca2+ activity in human cardiac fibroblasts. J. Cell. Physiol. 223, 68–75.
Cheng, J.C., Matsen, C.B., Gonzales, F.A., Ye, W., Greer, S., Marquez, V.E., Jones, P.A., and Selker, E.U. (2003). Inhibition of DNA methylation and reactivation of silenced genes by zebularine. J. Natl. Cancer Inst. 95, 399–409.
Chien, K.R., and Olson, E.N. (2002). Converging pathways and principles in heart development and disease: CV@CSH. Cell 110, 153–162.
Chimenti, I., Smith, R.R., Li, T.-S., Gerstenblith, G., Messina, E., Giacomello, A., and Marbán, E. (2010). Relative roles of direct regeneration versus paracrine effects of human cardiosphere-derived cells transplanted into infarcted mice. Circ. Res. 106, 971–980.
Chong, J.J.H., Chandrakanthan, V., Xaymardan, M., Asli, N.S., Li, J., Ahmed, I., Heffernan, C., Menon, M.K., Scarlett, C.J., Rashidianfar, A., et al. (2011). Adult cardiac-resident MSC-like stem cells with a proepicardial origin. Cell Stem Cell 9, 527–540.
Chong, J.J.H., Forte, E., and Harvey, R.P. (2014). Developmental origins and lineage descendants of endogenous adult cardiac progenitor cells. Stem Cell Res. 13, 592–614.
Christensen, A.H., Chatelain, F.C., Huttner, I.G., Olesen, M.S., Soka, M., Feliciangeli, S., Horvat, C., Santiago, C.F., Vandenberg, J.I., Schmitt, N., et al. (2016). The two-pore domain potassium channel, TWIK-1, has a role in the regulation of heart rate and atrial size. J. Mol. Cell. Cardiol. 97, 24–35.

Références bibliographiques
236
Christov, C., Chrétien, F., Abou-Khalil, R., Bassez, G., Vallet, G., Authier, F.-J., Bassaglia, Y., Shinin, V., Tajbakhsh, S., Chazaud, B., et al. (2007). Muscle Satellite Cells and Endothelial Cells: Close Neighbors and Privileged Partners. Mol. Biol. Cell 18, 1397–1409.
Chugh, A.R., Beache, G.M., Loughran, J.H., Mewton, N., Elmore, J.B., Kajstura, J., Pappas, P., Tatooles, A., Stoddard, M.F., Lima, J.A.C., et al. (2012). Administration of cardiac stem cells in patients with ischemic cardiomyopathy: the SCIPIO trial: surgical aspects and interim analysis of myocardial function and viability by magnetic resonance. Circulation 126, S54-64.
Ciccolini, F., Collins, T.J., Sudhoelter, J., Lipp, P., Berridge, M.J., and Bootman, M.D. (2003). Local and global spontaneous calcium events regulate neurite outgrowth and onset of GABAergic phenotype during neural precursor differentiation. J. Neurosci. Off. J. Soc. Neurosci. 23, 103–111.
Clapham, D.E. (2007). Calcium signaling. Cell 131, 1047–1058.
Codega, P., Silva-Vargas, V., Paul, A., Maldonado-Soto, A.R., Deleo, A.M., Pastrana, E., and Doetsch, F. (2014). Prospective identification and purification of quiescent adult neural stem cells from their in vivo niche. Neuron 82, 545–559.
Coppi, E., Pugliese, A.M., Urbani, S., Melani, A., Cerbai, E., Mazzanti, B., Bosi, A., Saccardi, R., and Pedata, F. (2007). ATP modulates cell proliferation and elicits two different electrophysiological responses in human mesenchymal stem cells. Stem Cells Dayt. Ohio 25, 1840–1849.
Corey, S., Krapivinsky, G., Krapivinsky, L., and Clapham, D.E. (1998). Number and stoichiometry of subunits in the native atrial G-protein-gated K+ channel, IKACh. J. Biol. Chem. 273, 5271–5278.
Cunha, S.R., Hund, T.J., Hashemi, S., Voigt, N., Li, N., Wright, P., Koval, O., Li, J., Gudmundsson, H., Gumina, R.J., et al. (2011). Defects in ankyrin-based membrane protein targeting pathways underlie atrial fibrillation. Circulation 124, 1212–1222.
Cu le , G.F., Ha es, M., A sa i, B., “ha , G., R a , A., Ba , F., O’B ie , T., O’Toole, D., a d Laffe , J.G. (2012). Mesenchymal stem cells enhance recovery and repair following ventilator-induced lung injury in the rat. Thorax 67, 496–501.
D’A a io, D., Fio i i, C., Ca p ell, P.M., Goi h e g, P., “a ada, F., )he g, H., Hosoda, T., Rota, M., Connell, J.M., Gallegos, R.P., et al. (2011). Functionally-Competent Cardiac Stem Cells Can Be Isolated from Endomyocardial Biopsies of Patients with Advanced Cardiomyopathies. Circ. Res. 108, 857–861.
Das, A., James, J., Rahnenführer, J., Thoreson, W., Bhattacharya, S., Zhao, X., and Ahmad, I. (2005). Retinal properties and potential of the adult mammalian ciliary epithelium stem cells. Vision Res. 45, 1653–1666.
D’As e zo, M., Pia e ti i, R., Casal o e, P., Budo i, M., Palli i, R., Azze a, G.B., a d G assi, C. . Role of L-type Ca2+ channels in neural stem/progenitor cell differentiation. Eur. J. Neurosci. 23, 935–944.
Davaille, J., Gallois, C., Habib, A., Li, L., Mallat, A., Tao, J., Levade, T., and Lotersztajn, S. (2000). Antiproliferative properties of sphingosine 1-phosphate in human hepatic myofibroblasts. A cyclooxygenase-2 mediated pathway. J. Biol. Chem. 275, 34628–34633.
Davis, E.P., Waffarn, F., and Sandman, C.A. (2011). Prenatal treatment with glucocorticoids sensitizes the hpa axis response to stress among full-term infants. Dev. Psychobiol. 53, 175–183.

Références bibliographiques
237
De Koninck, P., and Schulman, H. (1998). Sensitivity of CaM kinase II to the frequency of Ca2+ oscillations. Science 279, 227–230.
Dellavalle, A., Sampaolesi, M., Tonlorenzi, R., Tagliafico, E., Sacchetti, B., Perani, L., Innocenzi, A., Galvez, B.G., Messina, G., Morosetti, R., et al. (2007). Pericytes of human skeletal muscle are myogenic precursors distinct from satellite cells. Nat. Cell Biol. 9, 255–267.
Deng, F., Lei, H., Hu, Y., He, L., Fu, H., Feng, R., Feng, P., Huang, W., Wang, X., and Chang, J. (2016). Combination of retinoic acid, dimethyl sulfoxide and 5-azacytidine promotes cardiac differentiation of human fetal liver-derived mesenchymal stem cells. Cell Tissue Bank. 17, 147–159.
Deng, X.-L., Sun, H.-Y., Lau, C.-P., and Li, G.-R. (2006). Properties of ion channels in rabbit mesenchymal stem cells from bone marrow. Biochem. Biophys. Res. Commun. 348, 301–309.
Deng, X.L., Lau, C.P., Lai, K., Cheung, K.F., Lau, G.K., and Li, G.R. (2007). Cell cycle-dependent expression of potassium channels and cell proliferation in rat mesenchymal stem cells from bone marrow. Cell Prolif. 40, 656–670.
Dennis, J.E., Merriam, A., Awadallah, A., Yoo, J.U., Johnstone, B., and Caplan, A.I. (1999). A Quadripotential Mesenchymal Progenitor Cell Isolated from the Marrow of an Adult Mouse. J. Bone Miner. Res. 14, 700–709.
Deten, A., Hölzl, A., Leicht, M., Barth, W., and Zimmer, H.G. (2001). Changes in extracellular matrix and in transforming growth factor beta isoforms after coronary artery ligation in rats. J. Mol. Cell. Cardiol. 33, 1191–1207.
Deten, A., Volz, H.C., Clamors, S., Leiblein, S., Briest, W., Marx, G., and Zimmer, H.-G. (2005). Hematopoietic stem cells do not repair the infarcted mouse heart. Cardiovasc. Res. 65, 52–63.
Devito, L., Badraiq, H., Galleu, A., Taheem, D.K., Codognotto, S., Siow, R., Khalaf, Y., Briley, A., Shennan, A., Posto , L., et al. . Wha to ’s jell ese h al st o al/ste ells de i ed u der chemically defined animal product-free low oxygen conditions are rich in MSCA-1(+) subpopulation. Regen. Med. 9, 723–732.
Di Nicola, M., Carlo-Stella, C., Magni, M., Milanesi, M., Longoni, P.D., Matteucci, P., Grisanti, S., and Gianni, A.M. (2002). Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 99, 3838–3843.
Diver, J.M., Sage, S.O., and Rosado, J.A. (2001). The inositol trisphosphate receptor antagonist 2-aminoethoxydiphenylborate (2-APB) blocks Ca2+ entry channels in human platelets: cautions for its use in studying Ca2+ influx. Cell Calcium 30, 323–329.
Dolmetsch, R.E., Xu, K., and Lewis, R.S. (1998). Calcium oscillations increase the efficiency and specificity of gene expression. Nature 392, 933–936.
Dominici, M., Le Blanc, K., Mueller, I., Slaper-Cortenbach, I., Marini, F., Krause, D., Deans, R., Keating, A., Prockop, D., and Horwitz, E. (2006). Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 8, 315–317.
Donati, C., Cencetti, F., Nincheri, P., Bernacchioni, C., Brunelli, S., Clementi, E., Cossu, G., and Bruni, P. (2007). Sphingosine 1-phosphate mediates proliferation and survival of mesoangioblasts. Stem Cells Dayt. Ohio 25, 1713–1719.

Références bibliographiques
238
Donati, C., Cencetti, F., and Bruni, P. (2013). Sphingosine 1-phosphate axis: a new leader actor in skeletal muscle biology. Front. Physiol. 4, 338.
Dorfman, J., Duong, M., Zibaitis, A., Pelletier, M.P., Shum-Tim, D., Li, C., and Chiu, R.C. (1998). Myocardial tissue engineering with autologous myoblast implantation. J. Thorac. Cardiovasc. Surg. 116, 744–751.
Drabek, K., Gutiérrez, L., Vermeij, M., Clapes, T., Patel, S.R., Boisset, J.-C., van Haren, J., Pereira, A.L., Liu, Z., Akinci, U., et al. (2012). The microtubule plus-end tracking protein CLASP2 is required for hematopoiesis and hematopoietic stem cell maintenance. Cell Rep. 2, 781–788.
Du, Y., Li, D., Han, C., Wu, H., Xu, L., Zhang, M., Zhang, J., and Chen, X. (2017). Exosomes from Human-Induced Pluripotent Stem Cell-Derived Mesenchymal Stromal Cells (hiPSC-MSCs) Protect Liver against Hepatic Ischemia/ Reperfusion Injury via Activating Sphingosine Kinase and Sphingosine-1-Phosphate Signaling Pathway. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 43, 611–625.
Duester, G. (2008). Retinoic acid synthesis and signaling during early organogenesis. Cell 134, 921–931.
Dupont, G., Houart, G., and De Koninck, P. (2003). Sensitivity of CaM kinase II to the frequency of Ca2+ oscillations: a simple model. Cell Calcium 34, 485–497.
Eckfeldt, C.E., Mendenhall, E.M., and Verfaillie, C.M. (2005). The ole ula epe toi e of the al ight stem cell. Nat. Rev. Mol. Cell Biol. 6, 726–737.
Edalatmanesh, M.A., Bahrami, A.R., Hosseini, E., Hosseini, M., and Khatamsaz, S. (2011). Bone marrow derived mesenchymal stem cell transplantation in cerebellar degeneration: a behavioral study. Behav. Brain Res. 225, 63–70.
Egom, E.E.-A., Bae, J.S.H., Capel, R., Richards, M., Ke, Y., Pharithi, R.B., Maher, V., Kruzliak, P., and Lei, M. (2016). Effect of sphingosine-1-phosphate on L-type calcium current and Ca(2+) transient in rat ventricular myocytes. Mol. Cell. Biochem. 419, 83–92.
Eisenberg, L.M., Burns, L., and Eisenberg, C.A. (2003). Hematopoietic cells from bone marrow have the potential to differentiate into cardiomyocytes in vitro. Anat. Rec. A. Discov. Mol. Cell. Evol. Biol. 274, 870–882.
Ellison, G.M., Vicinanza, C., Smith, A.J., Aquila, I., Leone, A., Waring, C.D., Henning, B.J., Stirparo, G.G., Papait, R., Scarfò, M., et al. (2013). Adult c-kit(pos) cardiac stem cells are necessary and sufficient for functional cardiac regeneration and repair. Cell 154, 827–842.
Emmert, M.Y., Emmert, L.S., Martens, A., Ismail, I., Schmidt-Richter, I., Gawol, A., Seifert, B., Haverich, A., Martin, U., and Gruh, I. (2013). Higher frequencies of BCRP+ cardiac resident cells in ischaemic human myocardium. Eur. Heart J. 34, 2830–2838.
Engelhardt, J.F. (2001). Stem cell niches in the mouse airway. Am. J. Respir. Cell Mol. Biol. 24, 649–652.
E o , R.M., O’Co o , K.C., “o g, H., La ks, D.J., a d “ h a tz, D.K. . Agg egatio kinetics of well and poorly differentiated human prostate cancer cells. Biotechnol. Bioeng. 80, 580–588.
Enyedi, P., and Czirják, G. (2010). Molecular background of leak K+ currents: two-pore domain potassium channels. Physiol. Rev. 90, 559–605.

Références bibliographiques
239
Esteban, M.A., Wang, T., Qin, B., Yang, J., Qin, D., Cai, J., Li, W., Weng, Z., Chen, J., Ni, S., et al. (2010). Vitamin C enhances the generation of mouse and human induced pluripotent stem cells. Cell Stem Cell 6, 71–79.
Fadini, G.P., Boscaro, E., de Kreutzenberg, S., Agostini, C., Seeger, F., Dimmeler, S., Zeiher, A., Tiengo, A., and Avogaro, A. (2010). Time Course and Mechanisms of Circulating Progenitor Cell Reduction in the Natural History of Type 2 Diabetes. Diabetes Care 33, 1097–1102.
Fan, C.G., Tang, F.W., Zhang, Q.J., Lu, S.H., Liu, H.Y., Zhao, Z.M., Liu, B., Han, Z.B., and Han, Z.C. (2005). Characterization and neural differentiation of fetal lung mesenchymal stem cells. Cell Transplant. 14, 311–321.
Fang, S., Wei, J., Pentinmikko, N., Leinonen, H., and Salven, P. (2012). Generation of Functional Blood Vessels from a Single c-kit+ Adult Vascular Endothelial Stem Cell. PLoS Biol. 10.
Faroni, A., Rothwell, S.W., Grolla, A.A., Terenghi, G., Magnaghi, V., and Verkhratsky, A. (2013). Differentiation of adipose-derived stem cells into Schwann cell phenotype induces expression of P2X receptors that control cell death. Cell Death Dis. 4, e743.
Feliciangeli, S., Chatelain, F.C., Bichet, D., and Lesage, F. (2015). The family of K2P channels: salient structural and functional properties. J. Physiol. 593, 2587–2603.
Fernandes, K.J.L., McKenzie, I.A., Mill, P., Smith, K.M., Akhavan, M., Barnabé-Heider, F., Biernaskie, J., Junek, A., Kobayashi, N.R., Toma, J.G., et al. (2004). A dermal niche for multipotent adult skin-derived precursor cells. Nat. Cell Biol. 6, 1082–1093.
Ferreira-Martins, J., Rondon-Clavo, C., Tugal, D., Korn, J.A., Rizzi, R., Padin-Iruegas, M.E., Ottolenghi, S., De Angelis, A., Urbanek, K., Ide-Iwata, N., et al. (2009). Spontaneous calcium oscillations regulate human cardiac progenitor cell growth. Circ. Res. 105, 764–774.
Fiorio Pla, A., Maric, D., Brazer, S.-C., Giacobini, P., Liu, X., Chang, Y.H., Ambudkar, I.S., and Barker, J.L. (2005). Canonical transient receptor potential 1 plays a role in basic fibroblast growth factor (bFGF)/FGF receptor-1-induced Ca2+ entry and embryonic rat neural stem cell proliferation. J. Neurosci. Off. J. Soc. Neurosci. 25, 2687–2701.
Forostyak, O., Romanyuk, N., Verkhratsky, A., Sykova, E., and Dayanithi, G. (2013). Plasticity of calcium signaling cascades in human embryonic stem cell-derived neural precursors. Stem Cells Dev. 22, 1506–1521.
Forostyak, O., Forostyak, S., Kortus, S., Sykova, E., Verkhratsky, A., and Dayanithi, G. (2016). Physiology of Ca2+ signalling in stem cells of different origins and differentiation stages. Cell Calcium 59, 57–66.
Frey, N., McKinsey, T.A., and Olson, E.N. (2000). Decoding calcium signals involved in cardiac growth and function. Nat. Med. 6, 1221–1227.
Fukuda, K. (2002). Reprogramming of bone marrow mesenchymal stem cells into cardiomyocytes. C. R. Biol. 325, 1027–1038.
Gaborit, N., Le Bouter, S., Szuts, V., Varro, A., Escande, D., Nattel, S., and Demolombe, S. (2007). Regional and tissue specific transcript signatures of ion channel genes in the non-diseased human heart. J. Physiol. 582, 675–693.

Références bibliographiques
240
Gambini, E., Pompilio, G., Biondi, A., Alamanni, F., Capogrossi, M.C., Agrifoglio, M., and Pesce, M. (2011). C-kit+ cardiac progenitors exhibit mesenchymal markers and preferential cardiovascular commitment. Cardiovasc. Res. 89, 362–373.
Gang, E.J., Bosnakovski, D., Figueiredo, C.A., Visser, J.W., and Perlingeiro, R.C.R. (2007). SSEA-4 identifies mesenchymal stem cells from bone marrow. Blood 109, 1743–1751.
Garcia, J.G., Liu, F., Verin, A.D., Birukova, A., Dechert, M.A., Gerthoffer, W.T., Bamberg, J.R., and English, D. (2001). Sphingosine 1-phosphate promotes endothelial cell barrier integrity by Edg-dependent cytoskeletal rearrangement. J. Clin. Invest. 108, 689–701.
Gardner, S., Alzhanov, D., Knollman, P., Kuninger, D., and Rotwein, P. (2011). TGF-β inhibits muscle differentiation by blocking autocrine signaling pathways initiated by IGF-II. Mol. Endocrinol. Baltim. Md 25, 128–137.
Gargett, C.E. (2007). Uterine stem cells: what is the evidence? Hum. Reprod. Update 13, 87–101.
Gassanov, N., Er, F., Zagidullin, N., Jankowski, M., Gutkowska, J., and Hoppe, U.C. (2008). Retinoid acid-induced effects on atrial and pacemaker cell differentiation and expression of cardiac ion channels. Differ. Res. Biol. Divers. 76, 971–980.
Gay, M.S., Li, Y., Xiong, F., Lin, T., and Zhang, L. (2015). Dexamethasone Treatment of Newborn Rats Decreases Cardiomyocyte Endowment in the Developing Heart through Epigenetic Modifications. PloS One 10, e0125033.
Gellings Lowe, N., Swaney, J.S., Moreno, K.M., and Sabbadini, R.A. (2009). Sphingosine-1-phosphate and sphingosine kinase are critical for transforming growth factor-β-stimulated collagen production by cardiac fibroblasts. Cardiovasc. Res. 82, 303–312.
Ghosh, M., Helm, K.M., Smith, R.W., Giordanengo, M.S., Li, B., Shen, H., and Reynolds, S.D. (2011). A single cell functions as a tissue-specific stem cell and the in vitro niche-forming cell. Am. J. Respir. Cell Mol. Biol. 45, 459–469.
Gimpl, G., and Fahrenholz, F. (2001). The oxytocin receptor system: structure, function, and regulation. Physiol. Rev. 81, 629–683.
Gojo, S., Gojo, N., Takeda, Y., Mori, T., Abe, H., Kyo, S., Hata, J., and Umezawa, A. (2003). In vivo cardiovasculogenesis by direct injection of isolated adult mesenchymal stem cells. Exp. Cell Res. 288, 51–59.
Golan, K., Kollet, O., and Lapidot, T. (2013). Dynamic Cross Talk between S1P and CXCL12 Regulates Hematopoietic Stem Cells Migration, Development and Bone Remodeling. Pharmaceuticals 6, 1145–1169.
Goodell, M.A., Nguyen, H., and Shroyer, N. (2015). Somatic stem cell heterogeneity: diversity in the blood, skin and intestinal stem cell compartments. Nat. Rev. Mol. Cell Biol. 16, 299–309.
Grajales, L., García, J., Banach, K., and Geenen, D.L. (2010). Delayed enrichment of mesenchymal cells promotes cardiac lineage and calcium transient development. J. Mol. Cell. Cardiol. 48, 735–745.
Grajales, L., Lach, L.E., Janisch, P., Geenen, D.L., and García, J. (2015). Temporal expression of calcium channel subunits in satellite cells and bone marrow mesenchymal cells. Stem Cell Rev. 11, 408–422.

Références bibliographiques
241
Gronthos, S., Mankani, M., Brahim, J., Robey, P.G., and Shi, S. (2000). Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. U. S. A. 97, 13625–13630.
Gruber, P.J., Kubalak, S.W., Pexieder, T., Sucov, H.M., Evans, R.M., and Chien, K.R. (1996). RXR alpha deficiency confers genetic susceptibility for aortic sac, conotruncal, atrioventricular cushion, and ventricular muscle defects in mice. J. Clin. Invest. 98, 1332–1343.
Guan, K., and Hasenfuss, G. (2013). Cardiac resident progenitor cells: evidence and functional significance. Eur. Heart J. 34, 2784–2787.
Guilak, F., Estes, B.T., Diekman, B.O., Moutos, F.T., and Gimble, J.M. (2010). 2010 Nicolas Andry Award: Multipotent adult stem cells from adipose tissue for musculoskeletal tissue engineering. Clin. Orthop. 468, 2530–2540.
Gulbis, J.M., Mann, S., and MacKinnon, R. (1999). Structure of a Voltage-Dependent K+ Channel β Subunit. Cell 97, 943–952.
Gussoni, E., Soneoka, Y., Strickland, C.D., Buzney, E.A., Khan, M.K., Flint, A.F., Kunkel, L.M., and Mulligan, R.C. (1999). Dystrophin expression in the mdx mouse restored by stem cell transplantation. Nature 401, 390–394.
Gutkowska, J., Jankowski, M., and Antunes-Rodrigues, J. (2014). The role of oxytocin in cardiovascular regulation. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Medicas E Biol. 47, 206–214.
Han, W., Bao, W., Wang, Z., and Nattel, S. (2002). Comparison of ion-channel subunit expression in canine cardiac Purkinje fibers and ventricular muscle. Circ. Res. 91, 790–797.
Han, X.D., Chung, S.W., and Wong, P.M. (1995). Identification of a unique membrane-bound molecule on a hemopoietic stem cell line and on multipotent progenitor cells. Proc. Natl. Acad. Sci. U. S. A. 92, 11014–11018.
Han, Y., Chen, J.-D., Liu, Z.-M., Zhou, Y., Xia, J.-H., Du, X.-L., and Jin, M.-W. (2010). Functional ion channels in mouse cardiac c-kit(+) cells. Am. J. Physiol. Cell Physiol. 298, C1109-1117.
Hannanta-Anan, P., and Chow, B.Y. (2016). Optogenetic Control of Calcium Oscillation Waveform Defines NFAT as an Integrator of Calcium Load. Cell Syst. 2, 283–288.
Hashem, S.I., and Claycomb, W.C. (2013). Genetic isolation of stem cell-derived pacemaker-nodal cardiac myocytes. Mol. Cell. Biochem. 383, 161–171.
Hawke, T.J., and Garry, D.J. (2001). Myogenic satellite cells: physiology to molecular biology. J. Appl. Physiol. Bethesda Md 1985 91, 534–551.
Haynesworth, S.E., Baber, M.A., and Caplan, A.I. (1992). Cell surface antigens on human marrow-derived mesenchymal cells are detected by monoclonal antibodies. Bone 13, 69–80.
He, S., Nakada, D., and Morrison, S.J. (2009). Mechanisms of stem cell self-renewal. Annu. Rev. Cell Dev. Biol. 25, 377–406.
He, Z., Li, H., Zuo, S., Pasha, Z., Wang, Y., Yang, Y., Jiang, W., Ashraf, M., and Xu, M. (2011). Transduction of Wnt11 promotes mesenchymal stem cell transdifferentiation into cardiac phenotypes. Stem Cells Dev. 20, 1771–1778.

Références bibliographiques
242
Heineke, J., Auger-Messier, M., Xu, J., Oka, T., Sargent, M.A., York, A., Klevitsky, R., Vaikunth, S., Duncan, S.A., Aronow, B.J., et al. (2007). Cardiomyocyte GATA4 functions as a stress-responsive regulator of angiogenesis in the murine heart. J. Clin. Invest. 117, 3198–3210.
Heubach, J.F., Graf, E.M., Leutheuser, J., Bock, M., Balana, B., Zahanich, I., Christ, T., Boxberger, S., Wettwer, E., and Ravens, U. (2004). Electrophysiological properties of human mesenchymal stem cells. J. Physiol. 554, 659–672.
Hibino, H., Inanobe, A., Furutani, K., Murakami, S., Findlay, I., and Kurachi, Y. (2010). Inwardly rectifying potassium channels: their structure, function, and physiological roles. Physiol. Rev. 90, 291–366.
Hierlihy, A.M., Seale, P., Lobe, C.G., Rudnicki, M.A., and Megeney, L.A. (2002). The post-natal heart contains a myocardial stem cell population. FEBS Lett. 530, 239–243.
Hochgreb, T., Linhares, V.L., Menezes, D.C., Sampaio, A.C., Yan, C.Y.I., Cardoso, W.V., Rosenthal, N., and Xavier-Neto, J. (2003). A caudorostral wave of RALDH2 conveys anteroposterior information to the cardiac field. Dev. Camb. Engl. 130, 5363–5374.
Hoogaars, W.M.H., Engel, A., Brons, J.F., Verkerk, A.O., de Lange, F.J., Wong, L.Y.E., Bakker, M.L., Clout, D.E., Wakker, V., Barnett, P., et al. (2007). Tbx3 controls the sinoatrial node gene program and imposes pacemaker function on the atria. Genes Dev. 21, 1098–1112.
Horackova, M., Arora, R., Chen, R., Armour, J.A., Cattini, P.A., Livingston, R., and Byczko, Z. (2004). Cell transplantation for treatment of acute myocardial infarction: unique capacity for repair by skeletal muscle satellite cells. Am. J. Physiol. Heart Circ. Physiol. 287, H1599-1608.
Hotchkiss, A., Feridooni, T., Zhang, F., and Pasumarthi, K.B.S. (2014). The effects of calcium channel blockade on proliferation and differentiation of cardiac progenitor cells. Cell Calcium 55, 238–251.
Houser, S.R., and Molkentin, J.D. (2008). Does contractile Ca2+ control calcineurin-NFAT signaling and pathological hypertrophy in cardiac myocytes? Sci. Signal. 1, pe31.
Hu, H., He, M.-L., Tao, R., Sun, H.-Y., Hu, R., Zang, W.-J., Yuan, B.-X., Lau, C.-P., Tse, H.-F., and Li, G.-R. (2009). Characterization of ion channels in human preadipocytes. J. Cell. Physiol. 218, 427–435.
Hu, Q., Deshpande, S., Irani, K., and Ziegelstein, R.C. (1999). [Ca2+] i Oscillation Frequency Regulates Agonist-stimulated NF-κB Transcriptional Activity. J. Biol. Chem. 274, 33995–33998.
Hu, Q., Natarajan, V., and Ziegelstein, R.C. (2002). Phospholipase D regulates calcium oscillation frequency and nuclear factor-kappaB activity in histamine- stimulated human endothelial cells. Biochem. Biophys. Res. Commun. 292, 325–332.
Hund, T.J., and Mohler, P.J. (2008). Ankyrin-based targeting pathway regulates human sinoatrial node automaticity. Channels Austin Tex 2, 404–406.
Iachininoto, M.G., Capodimonti, S., Podda, M.V., Valentini, C.G., Bianchi, M., Leone, A.M., Teofili, L., and Leone, G. (2015). In vitro cardiomyocyte differentiation of umbilical cord blood cells: crucial role for c-kit(+) cells. Cytotherapy 17, 1627–1637.
Ia u, C.B., Ia u, D., Re ţea, I., Hostiu , “., De e giu, D., a d Rusu, M.C. . Mole ula signatures of cardiac stem cells. Romanian J. Morphol. Embryol. Rev. Roum. Morphol. Embryol. 56, 1255–1262.

Références bibliographiques
243
Ikeda, H., Satoh, H., Yanase, M., Inoue, Y., Tomiya, T., Arai, M., Tejima, K., Nagashima, K., Maekawa, H., Yahagi, N., et al. (2003). Antiproliferative property of sphingosine 1-phosphate in rat hepatocytes involves activation of Rho via Edg-5. Gastroenterology 124, 459–469.
I ’t A ke , P.“., “ he jo , “.A., Kleijburg-van der Keur, C., Noort, W.A., Claas, F.H.J., Willemze, R., Fibbe, W.E., and Kanhai, H.H.H. (2003). Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood 102, 1548–1549.
Innamorati, G., Fontana, E., Steccanella, F., Gandhi, K., Bassi, G., Zandonà, V., and Giacomello, L. (2017). Pleiotropic effects of sphingosine-1-phosphate signaling to control human chorionic mesenchymal stem cell physiology. Cell Death Dis. 8, e2930.
Inniss, K., and Moore, H. (2006). Mediation of apoptosis and proliferation of human embryonic stem cells by sphingosine-1-phosphate. Stem Cells Dev. 15, 789–796.
Itzhaki-Alfia, A., Leor, J., Raanani, E., Sternik, L., Spiegelstein, D., Netser, S., Holbova, R., Pevsner-Fischer, M., Lavee, J., and Barbash, I.M. (2009). Patient characteristics and cell source determine the number of isolated human cardiac progenitor cells. Circulation 120, 2559–2566.
Ivanyuk, D., Budash, G., Zheng, Y., Gaspar, J.A., Chaudhari, U., Fatima, A., Bahmanpour, S., Grin, V.K., Popandopulo, A.G., Sachinidis, A., et al. (2015). Ascorbic Acid-Induced Cardiac Differentiation of Murine Pluripotent Stem Cells: Transcriptional Profiling and Effect of a Small Molecule Synergist of Wnt/β-Catenin Signaling Pathway. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 36, 810–830.
Jackson, K.A., Majka, S.M., Wang, H., Pocius, J., Hartley, C.J., Majesky, M.W., Entman, M.L., Michael, L.H., Hirschi, K.K., and Goodell, M.A. (2001). Regeneration of ischemic cardiac muscle and vascular endothelium by adult stem cells. J. Clin. Invest. 107, 1395–1402.
Jäderstad, J., Jäderstad, L.M., Li, J., Chintawar, S., Salto, C., Pandolfo, M., Ourednik, V., Teng, Y.D., Sidman, R.L., Arenas, E., et al. (2010). Communication via gap junctions underlies early functional and beneficial interactions between grafted neural stem cells and the host. Proc. Natl. Acad. Sci. U. S. A. 107, 5184–5189.
Jang, S., Cho, H.-H., Cho, Y.-B., Park, J.-S., and Jeong, H.-S. (2010). Functional neural differentiation of human adipose tissue-derived stem cells using bFGF and forskolin. BMC Cell Biol. 11, 25.
Jansen, J.A., van Veen, T.A.B., de Bakker, J.M.T., and van Rijen, H.V.M. (2010). Cardiac connexins and impulse propagation. J. Mol. Cell. Cardiol. 48, 76–82.
Jin, S., Tian, D., Chen, J., Zhu, L., Liu, S., and Wang, D. (2006). Passive sensitization increases histamine-stimulated calcium signaling and NF-κB transcription activity in bronchial epithelial cells. Acta Pharmacol. Sin. 27, 708–714.
Jin, Y., Knudsen, E., Wang, L., Bryceson, Y., Damaj, B., Gessani, S., and Maghazachi, A.A. (2003). Sphingosine 1-phosphate is a novel inhibitor of T-cell proliferation. Blood 101, 4909–4915.
Jin, Z.-Q., Goetzl, E.J., and Karliner, J.S. (2004). Sphingosine Kinase Activation Mediates Ischemic Preconditioning in Murine Heart. Circulation 110, 1980–1989.
Johnston, P.V., Sasano, T., Mills, K., Evers, R., Lee, S.-T., Smith, R.R., Lardo, A.C., Lai, S., Steenbergen, C., Gerstenblith, G., et al. (2009). Engraftment, differentiation, and functional benefits of autologous

Références bibliographiques
244
cardiosphere-derived cells in porcine ischemic cardiomyopathy. Circulation 120, 1075–1083, 7 p following 1083.
Jüttermann, R., Li, E., and Jaenisch, R. (1994). Toxicity of 5-aza- ’-deoxycytidine to mammalian cells is mediated primarily by covalent trapping of DNA methyltransferase rather than DNA demethylation. Proc. Natl. Acad. Sci. U. S. A. 91, 11797–11801.
Kajstura, J., Leri, A., Finato, N., Di Loreto, C., Beltrami, C.A., and Anversa, P. (1998). Myocyte proliferation in end-stage cardiac failure in humans. Proc. Natl. Acad. Sci. U. S. A. 95, 8801–8805.
Kajstura, J., Rota, M., Whang, B., Cascapera, S., Hosoda, T., Bearzi, C., Nurzynska, D., Kasahara, H., Zias, E., Bonafé, M., et al. (2005). Bone marrow cells differentiate in cardiac cell lineages after infarction independently of cell fusion. Circ. Res. 96, 127–137.
Kaplan, M.S., and Hinds, J.W. (1977). Neurogenesis in the adult rat: electron microscopic analysis of light radioautographs. Science 197, 1092–1094.
Kattman, S.J., Witty, A.D., Gagliardi, M., Dubois, N.C., Niapour, M., Hotta, A., Ellis, J., and Keller, G. (2011). Stage-specific optimization of activin/nodal and BMP signaling promotes cardiac differentiation of mouse and human pluripotent stem cell lines. Cell Stem Cell 8, 228–240.
Kawamoto, A., and Losordo, D.W. (2008). Endothelial Progenitor Cells for Cardiovascular Regeneration. Trends Cardiovasc. Med. 18, 33–37.
Kawano, S., Shoji, S., Ichinose, S., Yamagata, K., Tagami, M., and Hiraoka, M. (2002). Characterization of Ca(2+) signaling pathways in human mesenchymal stem cells. Cell Calcium 32, 165–174.
Kawano, S., Otsu, K., Shoji, S., Yamagata, K., and Hiraoka, M. (2003). Ca(2+) oscillations regulated by Na(+)-Ca(2+) exchanger and plasma membrane Ca(2+) pump induce fluctuations of membrane currents and potentials in human mesenchymal stem cells. Cell Calcium 34, 145–156.
Kawano, S., Otsu, K., Kuruma, A., Shoji, S., Yanagida, E., Muto, Y., Yoshikawa, F., Hirayama, Y., Mikoshiba, K., and Furuichi, T. (2006). ATP autocrine/paracrine signaling induces calcium oscillations and NFAT activation in human mesenchymal stem cells. Cell Calcium 39, 313–324.
Kawata, T., Ishizuka, T., Tomura, H., Hisada, T., Dobashi, K., Tsukagoshi, H., Ishiwara, M., Kurose, H., Mori, M., and Okajima, F. (2005). Sphingosine 1-phosphate inhibits migration and RANTES production in human bronchial smooth muscle cells. Biochem. Biophys. Res. Commun. 331, 640–647.
Keating, G.M. (2009). Azacitidine: a review of its use in higher-risk myelodysplastic syndromes/acute myeloid leukaemia. Drugs 69, 2501–2518.
Keith, M.C.L., Tang, X.-L., Tokita, Y., Li, Q.-H., Ghafghazi, S., Moore Iv, J., Hong, K.U., Elmore, B., Amraotkar, A., Ganzel, B.L., et al. (2015). Safety of intracoronary infusion of 20 million C-kit positive human cardiac stem cells in pigs. PloS One 10, e0124227.
Kelly, B.A., Lewandowski, A.J., Worton, S.A., Davis, E.F., Lazdam, M., Francis, J., Neubauer, S., Lucas, A., Singhal, A., and Leeson, P. (2012). Antenatal glucocorticoid exposure and long-term alterations in aortic function and glucose metabolism. Pediatrics 129, e1282-1290.
Khakoo, A.Y., and Finkel, T. (2005). Endothelial progenitor cells. Annu. Rev. Med. 56, 79–101.

Références bibliographiques
245
Kim, Y.S., and Ahn, Y. (2012). A long road for stem cells to cure sick hearts: update on recent clinical trials. Korean Circ. J. 42, 71–79.
Kim, C.F.B., Jackson, E.L., Woolfenden, A.E., Lawrence, S., Babar, I., Vogel, S., Crowley, D., Bronson, R.T., and Jacks, T. (2005). Identification of bronchioalveolar stem cells in normal lung and lung cancer. Cell 121, 823–835.
Kimura, T., Watanabe, T., Sato, K., Kon, J., Tomura, H., Tamama, K., Kuwabara, A., Kanda, T., Kobayashi, I., Ohta, H., et al. (2000). Sphingosine 1-phosphate stimulates proliferation and migration of human endothelial cells possibly through the lipid receptors, Edg-1 and Edg-3. Biochem. J. 348 Pt 1, 71–76.
Kitisin, K., Saha, T., Blake, T., Golestaneh, N., Deng, M., Kim, C., Tang, Y., Shetty, K., Mishra, B., and Mishra, L. (2007). Tgf-Beta signaling in development. Sci. STKE Signal Transduct. Knowl. Environ. 2007, cm1.
Kluk, M.J., and Hla, T. (2001). Role of the sphingosine 1-phosphate receptor EDG-1 in vascular smooth muscle cell proliferation and migration. Circ. Res. 89, 496–502.
Klyachkin, Y.M., Nagareddy, P.R., Ye, S., Wysoczynski, M., Asfour, A., Gao, E., Sunkara, M., Brandon, J.A., Annabathula, R., Ponnapureddy, R., et al. (2015). Pharmacological Elevation of Circulating Bioactive Phosphosphingolipids Enhances Myocardial Recovery After Acute Infarction. Stem Cells Transl. Med. 4, 1333–1343.
Kolwicz, S.C., MacDonnell, S.M., Renna, B.F., Reger, P.O., Seqqat, R., Rafiq, K., Kendrick, Z.V., Houser, S.R., Sabri, A., and Libonati, J.R. (2009). Left ventricular remodeling with exercise in hypertension. Am. J. Physiol. Heart Circ. Physiol. 297, H1361-1368.
Kong, Y., Wang, H., Lin, T., and Wang, S. (2014). Sphingosine-1-phosphate/S1P Receptors Signaling Modulates Cell Migration in Human Bone Marrow-Derived Mesenchymal Stem Cells. Mediators Inflamm. 2014.
Kotova, P.D., Sysoeva, V.Y., Rogachevskaja, O.A., Bystrova, M.F., Kolesnikova, A.S., Tyurin-Kuzmin, P.A., Fadeeva, J.I., Tkachuk, V.A., and Kolesnikov, S.S. (2014). Functional expression of adrenoreceptors in mesenchymal stromal cells derived from the human adipose tissue. Biochim. Biophys. Acta 1843, 1899–1908.
K api i sk , G., Go do , E.A., Wi k a , K., Veli i o ić, B., K api i sk , L., a d Clapha , D.E. . The G-protein-gated atrial K+ channel IKACh is a heteromultimer of two inwardly rectifying K(+)-channel proteins. Nature 374, 135–141.
Kubo, H., and Alitalo, K. (2003). The bloody fate of endothelial stem cells. Genes Dev. 17, 322–329.
Kubo, H., Jaleel, N., Kumarapeli, A., Berretta, R.M., Bratinov, G., Shan, X., Wang, H., Houser, S.R., and Margulies, K.B. (2008). Increased cardiac myocyte progenitors in failing human hearts. Circulation 118, 649–657.
Kuçi, S., Kuçi, Z., Kreyenberg, H., Deak, E., Pütsch, K., Huenecke, S., Amara, C., Koller, S., Rettinger, E., Grez, M., et al. (2010). CD271 antigen defines a subset of multipotent stromal cells with immunosuppressive and lymphohematopoietic engraftment-promoting properties. Haematologica 95, 651–659.

Références bibliographiques
246
Kuhlmann, C.R.W., Most, A.K., Li, F., Münz, B.M., Schaefer, C.A., Walther, S., Raedle-Hurst, T., Waldecker, B., Piper, H.M., Tillmanns, H., et al. (2005). Endothelin-1-induced proliferation of human endothelial cells depends on activation of K+ channels and Ca+ influx. Acta Physiol. Scand. 161–169.
Kuhn, H.G., Dickinson-Anson, H., and Gage, F.H. (1996). Neurogenesis in the dentate gyrus of the adult rat: age-related decrease of neuronal progenitor proliferation. J. Neurosci. Off. J. Soc. Neurosci. 16, 2027–2033.
Kukreja, R.C. (2017). Myriad roles of voltage-activated potassium channel subunit Kvβ1.1 in the heart. Am. J. Physiol. - Heart Circ. Physiol. 312, H546–H548.
Kukuljan, M., Rojas, E., Catt, K.J., and Stojilkovic, S.S. (1994). Membrane potential regulates inositol 1,4,5-trisphosphate-controlled cytoplasmic Ca2+ oscillations in pituitary gonadotrophs. J. Biol. Chem. 269, 4860–4865.
Kuznetsov, S.A., Krebsbach, P.H., Satomura, K., Kerr, J., Riminucci, M., Benayahu, D., and Robey, P.G. (1997). Single-colony derived strains of human marrow stromal fibroblasts form bone after transplantation in vivo. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 12, 1335–1347.
Kuznetsov, S.A., Mankani, M.H., Gronthos, S., Satomura, K., Bianco, P., and Robey, P.G. (2001). Circulating Skeletal Stem Cells. J. Cell Biol. 153, 1133–1140.
Lagostena, L., Avitabile, D., De Falco, E., Orlandi, A., Grassi, F., Iachininoto, M.G., Ragone, G., Fucile, S., Pompilio, G., Eusebi, F., et al. (2005). Electrophysiological properties of mouse bone marrow c-kit+ cells co-cultured onto neonatal cardiac myocytes. Cardiovasc. Res. 66, 482–492.
Landeen, L.K., Dederko, D.A., Kondo, C.S., Hu, B.S., Aroonsakool, N., Haga, J.H., and Giles, W.R. (2008). Mechanisms of the negative inotropic effects of sphingosine-1-phosphate on adult mouse ventricular myocytes. Am. J. Physiol. Heart Circ. Physiol. 294, H736-749.
Lang, F., Shumilina, E., Ritter, M., Gulbins, E., Vereninov, A., and Huber, S.M. (2006). Ion channels and cell volume in regulation of cell proliferation and apoptotic cell death. Contrib. Nephrol. 152, 142–160.
Langer, D., Ikehara, Y., Takebayashi, H., Hawkes, R., and Zimmermann, H. (2007). The ectonucleotidases alkaline phosphatase and nucleoside triphosphate diphosphohydrolase 2 are associated with subsets of progenitor cell populations in the mouse embryonic, postnatal and adult neurogenic zones. Neuroscience 150, 863–879.
Laugwitz, K.-L., Moretti, A., Lam, J., Gruber, P., Chen, Y., Woodard, S., Lin, L.-Z., Cai, C.-L., Lu, M.M., Reth, M., et al. (2005). Postnatal isl1+ cardioblasts enter fully differentiated cardiomyocyte lineages. Nature 433, 647–653.
Laurie, G. (2004). Patenting Stem Cells of Human Origin.
Lázár, E., Sadek, H.A., and Bergmann, O. (2017). Cardiomyocyte renewal in the human heart: insights from the fall-out. Eur. Heart J. 38, 2333–2342.
Le Scouarnec, S., Bhasin, N., Vieyres, C., Hund, T.J., Cunha, S.R., Koval, O., Marionneau, C., Chen, B., Wu, Y., Demolombe, S., et al. (2008). Dysfunction in ankyrin-B-dependent ion channel and transporter targeting causes human sinus node disease. Proc. Natl. Acad. Sci. U. S. A. 105, 15617–15622.
LeClair, K.P., Palfree, R.G., Flood, P.M., Hammerling, U., and Bothwell, A. (1986). Isolation of a murine Ly-6 cDNA reveals a new multigene family. EMBO J. 5, 3227–3234.

Références bibliographiques
247
Lee, J.-H., Bhang, D.H., Beede, A., Huang, T.L., Stripp, B.R., Bloch, K.D., Wagers, A.J., Tseng, Y.-H., Ryeom, S., and Kim, C.F. (2014). Lung stem cell differentiation in mice directed by endothelial cells via a BMP4-NFATc1-thrombospondin-1 axis. Cell 156, 440–455.
Lee, J.Y., Qu-Petersen, Z., Cao, B., Kimura, S., Jankowski, R., Cummins, J., Usas, A., Gates, C., Robbins, P., Wernig, A., et al. (2000). Clonal isolation of muscle-derived cells capable of enhancing muscle regeneration and bone healing. J. Cell Biol. 150, 1085–1100.
Lee, K.-D., Kuo, T.K.-C., Whang-Peng, J., Chung, Y.-F., Lin, C.-T., Chou, S.-H., Chen, J.-R., Chen, Y.-P., and Lee, O.K.-S. (2004). In vitro hepatic differentiation of human mesenchymal stem cells. Hepatol. Baltim. Md 40, 1275–1284.
Leobon, B., Garcin, I., Menasche, P., Vilquin, J.-T., Audinat, E., and Charpak, S. (2003). Myoblasts transplanted into rat infarcted myocardium are functionally isolated from their host. Proc. Natl. Acad. Sci. U. S. A. 100, 7808–7811.
Lepore, J.J., Cappola, T.P., Mericko, P.A., Morrisey, E.E., and Parmacek, M.S. (2005). GATA-6 regulates genes promoting synthetic functions in vascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 25, 309–314.
Leri, A., Kajstura, J., and Anversa, P. (2005). Cardiac stem cells and mechanisms of myocardial regeneration. Physiol. Rev. 85, 1373–1416.
Lewis, R.S. (2003). Calcium oscillations in T-cells: mechanisms and consequences for gene expression. Biochem. Soc. Trans. 31, 925–929.
Li, G.-R., and Deng, X.-L. (2011). Functional ion channels in stem cells. World J. Stem Cells 3, 19–24.
Li, L., and Xie, T. (2005). Stem cell niche: structure and function. Annu. Rev. Cell Dev. Biol. 21, 605–631.
Li, G.-R., Sun, H., Deng, X., and Lau, C.-P. (2005). Characterization of ionic currents in human mesenchymal stem cells from bone marrow. Stem Cells Dayt. Ohio 23, 371–382.
Li, G.-R., Deng, X.-L., Sun, H., Chung, S.S.M., Tse, H.-F., and Lau, C.-P. (2006). Ion Channels in Mesenchymal Stem Cells from Rat Bone Marrow. STEM CELLS 24, 1519–1528.
Li, L., Mignone, J., Yang, M., Matic, M., Penman, S., Enikolopov, G., and Hoffman, R.M. (2003). Nestin expression in hair follicle sheath progenitor cells. Proc. Natl. Acad. Sci. U. S. A. 100, 9958–9961.
Li, M., Na i, N., Yahi o, E., Liu, K., Po ell, P.C., B adle , W.E., Ma ti , D.I.K., G aha , R.M., Dell’Italia, L.J., and Husain, A. (2008). c-kit is required for cardiomyocyte terminal differentiation. Circ. Res. 102, 677–685.
Li, W., Hayashida, Y., Chen, Y.-T., and Tseng, S.C.G. (2007). Niche regulation of corneal epithelial stem cells at the limbus. Cell Res. 17, 26–36.
Liao, R., Pfister, O., Jain, M., and Mouquet, F. (2007). The bone marrow--cardiac axis of myocardial regeneration. Prog. Cardiovasc. Dis. 50, 18–30.
Liebau, S., Pröpper, C., Böckers, T., Lehmann-Horn, F., Storch, A., Grissmer, S., and Wittekindt, O.H. (2006). Selective blockage of Kv1.3 and Kv3.1 channels increases neural progenitor cell proliferation. J. Neurochem. 99, 426–437.

Références bibliographiques
248
Liggins, G.C., and Howie, R.N. (1972). A controlled trial of antepartum glucocorticoid treatment for prevention of the respiratory distress syndrome in premature infants. Pediatrics 50, 515–525.
Lim, C.G., Kim, S.-S., Suh-Kim, H., Lee, Y.-D., and Ahn, S.C. (2008). Characterization of ionic currents in human neural stem cells. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 12, 131–135.
Linke, A., Müller, P., Nurzynska, D., Casarsa, C., Torella, D., Nascimbene, A., Castaldo, C., Cascapera, S., Böhm, M., Quaini, F., et al. (2005). Stem cells in the dog heart are self-renewing, clonogenic, and multipotent and regenerate infarcted myocardium, improving cardiac function. Proc. Natl. Acad. Sci. U. S. A. 102, 8966–8971.
Liu, J., Hsu, A., Lee, J.-F., Cramer, D.E., and Lee, M.-J. (2011). To stay or to leave: Stem cells and progenitor cells navigating the S1P gradient. World J. Biol. Chem. 2, 1–13.
Liu, Y.-J., Wang, X.-G., Tang, Y.-B., Chen, J.-H., Lv, X.-F., Zhou, J.-G., and Guan, Y.-Y. (2010). Simvastatin ameliorates rat cerebrovascular remodeling during hypertension via inhibition of volume-regulated chloride channel. Hypertens. Dallas Tex 1979 56, 445–452.
Llucià-Valldeperas, A., Sanchez, B., Soler-Botija, C., Gálvez-Montón, C., Roura, S., Prat-Vidal, C., Perea-Gil, I., Rosell-Ferrer, J., Bragos, R., and Bayes-Genis, A. (2014). Physiological conditioning by electric field stimulation promotes cardiomyogenic gene expression in human cardiomyocyte progenitor cells. Stem Cell Res. Ther. 5, 93.
Lockyer, P.J., Kupzig, S., and Cullen, P.J. (2001). CAPRI regulates Ca2+-dependent inactivation of the Ras-MAPK pathway. Curr. Biol. 11, 981–986.
Long, J.S., Fujiwara, Y., Edwards, J., Tannahill, C.L., Tigyi, G., Pyne, S., and Pyne, N.J. (2010). Sphingosine 1-phosphate receptor 4 uses HER2 (ERBB2) to regulate extracellular signal regulated kinase-1/2 in MDA-MB-453 breast cancer cells. J. Biol. Chem. 285, 35957–35966.
Losa, M., Latorre, V., Andrabi, M., Ladam, F., Sagerström, C., Novoa, A., Zarrineh, P., Bridoux, L., Hanley, N.A., Mallo, M., et al. (2017). A tissue-specific, Gata6-driven transcriptional program instructs remodeling of the mature arterial tree. ELife 6.
Lotshaw, D.P. (2007). Biophysical, pharmacological, and functional characteristics of cloned and native mammalian two-pore domain K+ channels. Cell Biochem. Biophys. 47, 209–256.
Lu, W., Yaoming, N., Boli, R., Jun, C., Changhai, Z., Yang, Z., and Zhiyuan, S. (2013). mHCN4 genetically modified canine mesenchymal stem cells provide biological pacemaking function in complete dogs with atrioventricular block. Pacing Clin. Electrophysiol. PACE 36, 1138–1149.
Lu, W., Xiu, X., Zhao, Y., and Gui, M. (2015). Improved Proliferation and Differentiation of Bone Marrow Mesenchymal Stem Cells Into Vascular Endothelial Cells With Sphingosine 1-Phosphate. Transplant. Proc. 47, 2035–2040.
Luneau, C., Wiedmann, R., Smith, J.S., and Williams, J.B. (1991). Shaw-like rat brain potassium channel DNA’s ith di e ge t ’ e ds. FEB“ Lett. 288, 163–167.
Luskin, M.B. (1993). Restricted proliferation and migration of postnatally generated neurons derived from the forebrain subventricular zone. Neuron 11, 173–189.
Ma, T. (2010). Mesenchymal stem cells: From bench to bedside. World J. Stem Cells 2, 13–17.

Références bibliographiques
249
Ma, H.-T., Venkatachalam, K., Parys, J.B., and Gill, D.L. (2002). Modification of Store-operated Channel Coupling and Inositol Trisphosphate Receptor Function by 2-Aminoethoxydiphenyl Borate in DT40 Lymphocytes. J. Biol. Chem. 277, 6915–6922.
Maceyka, M., Harikumar, K.B., Milstien, S., and Spiegel, S. (2012). Sphingosine-1-phosphate signaling and its role in disease. Trends Cell Biol. 22, 50–60.
MacFarlane, S.N., and Sontheimer, H. (2000). Changes in ion channel expression accompany cell cycle progression of spinal cord astrocytes. Glia 30, 39–48.
MacLellan, W.R., and Schneider, M.D. (2000). Genetic dissection of cardiac growth control pathways. Annu. Rev. Physiol. 62, 289–319.
Macrin, D., Joseph, J.P., Pillai, A.A., and Devi, A. (2017). Eminent Sources of Adult Mesenchymal Stem Cells and Their Therapeutic Imminence. Stem Cell Rev.
Majumdar, M.K., Thiede, M.A., Mosca, J.D., Moorman, M., and Gerson, S.L. (1998). Phenotypic and functional comparison of cultures of marrow-derived mesenchymal stem cells (MSCs) and stromal cells. J. Cell. Physiol. 176, 57–66.
Majumdar, M.K., Keane-Moore, M., Buyaner, D., Hardy, W.B., Moorman, M.A., McIntosh, K.R., and Mosca, J.D. (2003). Characterization and functionality of cell surface molecules on human mesenchymal stem cells. J. Biomed. Sci. 10, 228–241.
Makkar, R.R., Smith, R.R., Cheng, K., Malliaras, K., Thomson, L.E.J., Berman, D., Czer, L.S.C., Marbán, L., Mendizabal, A., Johnston, P.V., et al. (2012). Intracoronary cardiosphere-derived cells for heart regeneration after myocardial infarction (CADUCEUS): a prospective, randomised phase 1 trial. Lancet Lond. Engl. 379, 895–904.
Malmersjö, S., Rebellato, P., Smedler, E., Planert, H., Kanatani, S., Liste, I., Nanou, E., Sunner, H., Abdelhady, S., Zhang, S., et al. (2013). Neural progenitors organize in small-world networks to promote cell proliferation. Proc. Natl. Acad. Sci. U. S. A. 110, E1524-1532.
Mangoni, M.E., Couette, B., Bourinet, E., Platzer, J., Reimer, D., Striessnig, J., and Nargeot, J. (2003). Functional role of L-type Cav1.3 Ca2+ channels in cardiac pacemaker activity. Proc. Natl. Acad. Sci. U. S. A. 100, 5543–5548.
Martin, C.M., Meeson, A.P., Robertson, S.M., Hawke, T.J., Richardson, J.A., Bates, S., Goetsch, S.C., Gallardo, T.D., and Garry, D.J. (2004). Persistent expression of the ATP-binding cassette transporter, Abcg2, identifies cardiac SP cells in the developing and adult heart. Dev. Biol. 265, 262–275.
Massagué, J., Cheifetz, S., Endo, T., and Nadal-Ginard, B. (1986). Type beta transforming growth factor is an inhibitor of myogenic differentiation. Proc. Natl. Acad. Sci. U. S. A. 83, 8206–8210.
Matsuura, K., Nagai, T., Nishigaki, N., Oyama, T., Nishi, J., Wada, H., Sano, M., Toko, H., Akazawa, H., Sato, T., et al. (2004). Adult cardiac Sca-1-positive cells differentiate into beating cardiomyocytes. J. Biol. Chem. 279, 11384–11391.
Maxwell, J.T., Wagner, M.B., and Davis, M.E. (2016). Electrically Induced Calcium Handling in Cardiac Progenitor Cells. Stem Cells Int. 2016, 8917380.
McCullagh, K.J.A., and Perlingeiro, R.C.R. (2015). Coaxing stem cells for skeletal muscle repair. Adv. Drug Deliv. Rev. 84, 198–207.

Références bibliographiques
250
McDonnell, T.J., and Oberpriller, J.O. (1984). The response of the atrium to direct mechanical wounding in the adult heart of the newt, Notophthalmus viridescens. An electron-microscopic and autoradiographic study. Cell Tissue Res. 235, 583–592.
Means, C.K., and Brown, J.H. (2009). Sphingosine-1-phosphate receptor signalling in the heart. Cardiovasc. Res. 82, 193–200.
Means, C.K., Miyamoto, S., Chun, J., and Brown, J.H. (2008). S1P1 receptor localization confers selectivity for Gi-mediated cAMP and contractile responses. J. Biol. Chem. 283, 11954–11963.
Menasché, P., Hagège, A.A., Scorsin, M., Pouzet, B., Desnos, M., Duboc, D., Schwartz, K., Vilquin, J.T., and Marolleau, J.P. (2001). Myoblast transplantation for heart failure. Lancet Lond. Engl. 357, 279–280.
Menasché, P., Hagège, A.A., Vilquin, J.-T., Desnos, M., Abergel, E., Pouzet, B., Bel, A., Sarateanu, S., Scorsin, M., Schwartz, K., et al. (2003). Autologous skeletal myoblast transplantation for severe postinfarction left ventricular dysfunction. J. Am. Coll. Cardiol. 41, 1078–1083.
Messina, E., De Angelis, L., Frati, G., Morrone, S., Chimenti, S., Fiordaliso, F., Salio, M., Battaglia, M., Latronico, M.V.G., Coletta, M., et al. (2004). Isolation and expansion of adult cardiac stem cells from human and murine heart. Circ. Res. 95, 911–921.
Miki, T., Suzuki, M., Shibasaki, T., Uemura, H., Sato, T., Yamaguchi, K., Koseki, H., Iwanaga, T., Nakaya, H., and Seino, S. (2002). Mouse model of Prinzmetal angina by disruption of the inward rectifier Kir6.1. Nat. Med. 8, 466–472.
Miller, C. (2000). An overview of the potassium channel family. Genome Biol. 1, reviews0004.1-reviews0004.5.
Mimeault, M., and Batra, S.K. (2006). Concise review: recent advances on the significance of stem cells in tissue regeneration and cancer therapies. Stem Cells Dayt. Ohio 24, 2319–2345.
Mishra, R., Vijayan, K., Colletti, E.J., Harrington, D.A., Matthiesen, T.S., Simpson, D., Goh, S.K., Walker, B.L., Almeida-Porada, G., Wang, D., et al. (2011). Characterization and functionality of cardiac progenitor cells in congenital heart patients. Circulation 123, 364–373.
Mishra, S., Reznikov, V., Maltsev, V.A., Undrovinas, N.A., Sabbah, H.N., and Undrovinas, A. (2015). Contribution of sodium channel neuronal isoform Nav1.1 to late sodium current in ventricular myocytes from failing hearts. J. Physiol. 593, 1409–1427.
Molofsky, A.V., Pardal, R., Iwashita, T., Park, I.-K., Clarke, M.F., and Morrison, S.J. (2003). Bmi-1 dependence distinguishes neural stem cell self-renewal from progenitor proliferation. Nature 425, 962–967.
Montoro, R.J., and Yuste, R. (2004). Gap junctions in developing neocortex: a review. Brain Res. Brain Res. Rev. 47, 216–226.
Moreau, A., Mercier, A., Thériault, O., Boutjdir, M., Burger, B., Keller, D.I., and Chahine, M. (2017). Biophysical, Molecular, and Pharmacological Characterization of Voltage-Dependent Sodium Channels From Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Can. J. Cardiol. 33, 269–278.

Références bibliographiques
251
Moretti, A., Caron, L., Nakano, A., Lam, J.T., Bernshausen, A., Chen, Y., Qyang, Y., Bu, L., Sasaki, M., Martin-Puig, S., et al. (2006). Multipotent embryonic isl1+ progenitor cells lead to cardiac, smooth muscle, and endothelial cell diversification. Cell 127, 1151–1165.
Morrison, S.J., and Spradling, A.C. (2008). Stem cells and niches: mechanisms that promote stem cell maintenance throughout life. Cell 132, 598–611.
Moubarak, M., Magaud, C., Saliba, Y., Chatelier, A., Bois, P., Faivre, J.-F., and Fares, N. (2015). Effects of atrial natriuretic peptide on rat ventricular fibroblasts during differentiation into myofibroblasts. Physiol. Res. 64, 495–503.
Mukaddam-Daher, S., Jankowski, M., Wang, D., Menaouar, A., and Gutkowska, J. (2002). Regulation of cardiac oxytocin system and natriuretic peptide during rat gestation and postpartum. J. Endocrinol. 175, 211–216.
Muller-Borer, B., Esch, G., Aldina, R., Woon, W., Fox, R., Bursac, N., Hiller, S., Maeda, N., Shepherd, N., Jin, J.P., et al. (2012). Calcium dependent CAMTA1 in adult stem cell commitment to a myocardial lineage. PloS One 7, e38454.
Muller-Borer, B.J., Cascio, W.E., Esch, G.L., Kim, H.-S., Coleman, W.B., Grisham, J.W., Anderson, P.A.W., and Malouf, N.N. (2007). Mechanisms controlling the acquisition of a cardiac phenotype by liver stem cells. Proc. Natl. Acad. Sci. U. S. A. 104, 3877–3882.
Murry, C.E., Soonpaa, M.H., Reinecke, H., Nakajima, H., Nakajima, H.O., Rubart, M., Pasumarthi, K.B.S., Virag, J.I., Bartelmez, S.H., Poppa, V., et al. (2004). Haematopoietic stem cells do not transdifferentiate into cardiac myocytes in myocardial infarcts. Nature 428, 664–668.
Naeem, N., Haneef, K., Kabir, N., Iqbal, H., Jamall, S., and Salim, A. (2013). DNA methylation inhibitors, 5-azacytidine and zebularine potentiate the transdifferentiation of rat bone marrow mesenchymal stem cells into cardiomyocytes. Cardiovasc. Ther. 31, 201–209.
Nakagami, H., Maeda, K., Morishita, R., Iguchi, S., Nishikawa, T., Takami, Y., Kikuchi, Y., Saito, Y., Tamai, K., Ogihara, T., et al. (2005). Novel autologous cell therapy in ischemic limb disease through growth factor secretion by cultured adipose tissue-derived stromal cells. Arterioscler. Thromb. Vasc. Biol. 25, 2542–2547.
Nelson, M.T., and Quayle, J.M. (1995). Physiological roles and properties of potassium channels in arterial smooth muscle. Am. J. Physiol. 268, C799-822.
Nerbonne, J.M. (2016). Molecular Basis of Functional Myocardial Potassium Channel Diversity. Card. Electrophysiol. Clin. 8, 257–273.
Ng, K.-E., Schwarzer, S., Duchen, M.R., and Tinker, A. (2010). The intracellular localization and function of the ATP-sensitive K+ channel subunit Kir6.1. J. Membr. Biol. 234, 137–147.
Niederreither, K., and Dollé, P. (2008). Retinoic acid in development: towards an integrated view. Nat. Rev. Genet. 9, 541–553.
Niederreither, K., Subbarayan, V., Dollé, P., and Chambon, P. (1999). Embryonic retinoic acid synthesis is essential for early mouse post-implantation development. Nat. Genet. 21, 444–448.

Références bibliographiques
252
Niederreither, K., Vermot, J., Le Roux, I., Schuhbaur, B., Chambon, P., and Dollé, P. (2003). The regional pattern of retinoic acid synthesis by RALDH2 is essential for the development of posterior pharyngeal arches and the enteric nervous system. Dev. Camb. Engl. 130, 2525–2534.
Nigro, P., Perrucci, G.L., Gowran, A., Zanobini, M., Capogrossi, M.C., and Pompilio, G. (2015). c-kit(+) cells: the tell-tale heart of cardiac regeneration? Cell. Mol. Life Sci. CMLS 72, 1725–1740.
Nincheri, P., Luciani, P., Squecco, R., Donati, C., Bernacchioni, C., Borgognoni, L., Luciani, G., Benvenuti, S., Francini, F., and Bruni, P. (2009). Sphingosine 1-phosphate induces differentiation of adipose tissue-derived mesenchymal stem cells towards smooth muscle cells. Cell. Mol. Life Sci. CMLS 66, 1741–1754.
Nishino, J., Kim, I., Chada, K., and Morrison, S.J. (2008). Hmga2 promotes neural stem cell self-renewal in young but not old mice by reducing p16Ink4a and p19Arf Expression. Cell 135, 227–239.
Noo t, W.A., K uissel i k, A.B., i ’t A ke , P.“., K uge , M., a Bezooije , R.L., de Paus, R.A., Heemskerk, M.H.M., Löwik, C.W.G.M., Falkenburg, J.H., Willemze, R., et al. (2002). Mesenchymal stem cells promote engraftment of human umbilical cord blood-derived CD34(+) cells in NOD/SCID mice. Exp. Hematol. 30, 870–878.
Noseda, M., Harada, M., McSweeney, S., Leja, T., Belian, E., Stuckey, D.J., Abreu Paiva, M.S., Habib, J., Macaulay, I., de Smith, A.J., et al. (2015). PDGFRα demarcates the cardiogenic clonogenic Sca1+ stem/progenitor cell in adult murine myocardium. Nat. Commun. 6, 6930.
Novgorodov, A.S., El-Alwani, M., Bielawski, J., Obeid, L.M., and Gudz, T.I. (2007). Activation of sphingosine-1-phosphate receptor S1P5 inhibits oligodendrocyte progenitor migration. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 21, 1503–1514.
O’Co o , M.D., Ka del, M.D., Iosfi a, I., Youssef, D., Lu, M., Li, M.M., Ve aute e , “., Nag , A., a d Eaves, C.J. (2008). Alkaline phosphatase-positive colony formation is a sensitive, specific, and quantitative indicator of undifferentiated human embryonic stem cells. Stem Cells Dayt. Ohio 26, 1109–1116.
Oh, H., Bradfute, S.B., Gallardo, T.D., Nakamura, T., Gaussin, V., Mishina, Y., Pocius, J., Michael, L.H., Behringer, R.R., Garry, D.J., et al. (2003). Cardiac progenitor cells from adult myocardium: homing, differentiation, and fusion after infarction. Proc. Natl. Acad. Sci. U. S. A. 100, 12313–12318.
Oh, H., Chi, X., Bradfute, S.B., Mishina, Y., Pocius, J., Michael, L.H., Behringer, R.R., Schwartz, R.J., Entman, M.L., and Schneider, M.D. (2004). Cardiac muscle plasticity in adult and embryo by heart-derived progenitor cells. Ann. N. Y. Acad. Sci. 1015, 182–189.
Olivera, A., Rosenfeldt, H.M., Bektas, M., Wang, F., Ishii, I., Chun, J., Milstien, S., and Spiegel, S. (2003). Sphingosine kinase type 1 induces G12/13-mediated stress fiber formation, yet promotes growth and survival independent of G protein-coupled receptors. J. Biol. Chem. 278, 46452–46460.
Olschewski, A., Li, Y., Tang, B., Hanze, J., Eul, B., Bohle, R.M., Wilhelm, J., Morty, R.E., Brau, M.E., Weir, E.K., et al. (2006). Impact of TASK-1 in human pulmonary artery smooth muscle cells. Circ. Res. 98, 1072–1080.
Onohara, N., Nishida, M., Inoue, R., Kobayashi, H., Sumimoto, H., Sato, Y., Mori, Y., Nagao, T., and Ku ose, H. . TRPC a d TRPC a e esse tial fo a giote si II‐i du ed a dia h pe t oph . EMBO J. 25, 5305–5316.

Références bibliographiques
253
Orciani, M., Mariggiò, M.A., Morabito, C., Di Benedetto, G., and Di Primio, R. (2010). Functional characterization of calcium-signaling pathways of human skin-derived mesenchymal stem cells. Skin Pharmacol. Physiol. 23, 124–132.
Orlic, D., Kajstura, J., Chimenti, S., Bodine, D.M., Leri, A., and Anversa, P. (2001a). Transplanted adult bone marrow cells repair myocardial infarcts in mice. Ann. N. Y. Acad. Sci. 938, 221-229; discussion 229-230.
Orlic, D., Kajstura, J., Chimenti, S., Limana, F., Jakoniuk, I., Quaini, F., Nadal-Ginard, B., Bodine, D.M., Leri, A., and Anversa, P. (2001b). Mobilized bone marrow cells repair the infarcted heart, improving function and survival. Proc. Natl. Acad. Sci. U. S. A. 98, 10344–10349.
Orrenius, S., Zhivotovsky, B., and Nicotera, P. (2003). Regulation of cell death: the calcium-apoptosis link. Nat. Rev. Mol. Cell Biol. 4, 552–565.
Osmond, M.K., Butler, A.J., Voon, F.C., and Bellairs, R. (1991). The effects of retinoic acid on heart formation in the early chick embryo. Dev. Camb. Engl. 113, 1405–1417.
Padayatty, S.J., Sun, H., Wang, Y., Riordan, H.D., Hewitt, S.M., Katz, A., Wesley, R.A., and Levine, M. (2004). Vitamin C pharmacokinetics: implications for oral and intravenous use. Ann. Intern. Med. 140, 533–537.
Pajoohesh-Ganji, A., and Stepp, M.A. (2005). In search of markers for the stem cells of the corneal epithelium. Biol. Cell 97, 265–276.
Paquin, J., Danalache, B.A., Jankowski, M., McCann, S.M., and Gutkowska, J. (2002). Oxytocin induces differentiation of P19 embryonic stem cells to cardiomyocytes. Proc. Natl. Acad. Sci. U. S. A. 99, 9550–9555.
Pardo, L.A. (2004). Voltage-gated potassium channels in cell proliferation. Physiol. Bethesda Md 19, 285–292.
Park, J.-S., Kim, Y.-S., and Yoo, M.-A. (2009). The role of p38b MAPK in age-related modulation of intestinal stem cell proliferation and differentiation in Drosophila. Aging 1, 637–651.
Park, K.S., Jung, K.H., Kim, S.H., Kim, K.S., Choi, M.R., Kim, Y., and Chai, Y.G. (2007). Functional expression of ion channels in mesenchymal stem cells derived from umbilical cord vein. Stem Cells Dayt. Ohio 25, 2044–2052.
Pasha, Z., Wang, Y., Sheikh, R., Zhang, D., Zhao, T., and Ashraf, M. (2008). Preconditioning enhances cell survival and differentiation of stem cells during transplantation in infarcted myocardium. Cardiovasc. Res. 77, 134–142.
Patterson, J.M., Johnson, M.H., Zimonjic, D.B., and Graubert, T.A. (2000). Characterization of Ly-6M, a novel member of the Ly-6 family of hematopoietic proteins. Blood 95, 3125–3132.
Payne, S.G., Milstien, S., and Spiegel, S. (2002). Sphingosine-1-phosphate: dual messenger functions. FEBS Lett. 531, 54–57.
Pelton, R.W., Saxena, B., Jones, M., Moses, H.L., and Gold, L.I. (1991). Immunohistochemical localization of TGF beta 1, TGF beta 2, and TGF beta 3 in the mouse embryo: expression patterns suggest multiple roles during embryonic development. J. Cell Biol. 115, 1091–1105.

Références bibliographiques
254
Perney, T.M., and Kaczmarek, L.K. (1991). The molecular biology of K+ channels. Curr. Opin. Cell Biol. 3, 663–670.
Peterkin, T., Gibson, A., Loose, M., and Patient, R. (2005). The roles of GATA-4, -5 and -6 in vertebrate heart development. Semin. Cell Dev. Biol. 16, 83–94.
Petersen, B.E., Grossbard, B., Hatch, H., Pi, L., Deng, J., and Scott, E.W. (2003). Mouse A6-positive hepatic oval cells also express several hematopoietic stem cell markers. Hepatol. Baltim. Md 37, 632–640.
Pfister, O., Mouquet, F., Jain, M., Summer, R., Helmes, M., Fine, A., Colucci, W.S., and Liao, R. (2005). CD31- but Not CD31+ cardiac side population cells exhibit functional cardiomyogenic differentiation. Circ. Res. 97, 52–61.
Piersma, A.H., Brockbank, K.G., Ploemacher, R.E., van Vliet, E., Brakel-van Peer, K.M., and Visser, P.J. (1985). Characterization of fibroblastic stromal cells from murine bone marrow. Exp. Hematol. 13, 237–243.
Pittenger, M.F., Mackay, A.M., Beck, S.C., Jaiswal, R.K., Douglas, R., Mosca, J.D., Moorman, M.A., Simonetti, D.W., Craig, S., and Marshak, D.R. (1999). Multilineage potential of adult human mesenchymal stem cells. Science 284, 143–147.
Planat-Benard, V., Silvestre, J.-S., Cousin, B., André, M., Nibbelink, M., Tamarat, R., Clergue, M., Manneville, C., Saillan-Barreau, C., Duriez, M., et al. (2004). Plasticity of human adipose lineage cells toward endothelial cells: physiological and therapeutic perspectives. Circulation 109, 656–663.
Polzin, A., Piayda, K., Keul, P., Dannenberg, L., Mohring, A., Gräler, M., Zeus, T., Kelm, M., and Levkau, B. (2017). Plasma sphingosine-1-phosphate concentrations are associated with systolic heart failure in patients with ischemic heart disease. J. Mol. Cell. Cardiol. 110, 35–37.
Prakriya, M., and Lewis, R.S. (2001). Potentiation and inhibition of Ca2+ release-activated Ca2+ channels by 2-aminoethyldiphenyl borate (2-APB) occurs independently of IP3 receptors. J. Physiol. 536, 3–19.
Price, S.T., Beckham, T.H., Cheng, J.C., Lu, P., Liu, X., and Norris, J.S. (2015). Sphingosine 1-Phosphate Receptor 2 Regulates the Migration, Proliferation, and Differentiation of Mesenchymal Stem Cells. Int. J. Stem Cell Res. Ther. 2.
Psaltis, P.J., Zannettino, A.C.W., Worthley, S.G., and Gronthos, S. (2008). Concise review: mesenchymal stromal cells: potential for cardiovascular repair. Stem Cells Dayt. Ohio 26, 2201–2210.
Pucéat, M. (2007). TGFbeta in the differentiation of embryonic stem cells. Cardiovasc. Res. 74, 256–261.
Qian, Q., Qian, H., Zhang, X., Zhu, W., Yan, Y., Ye, S., Peng, X., Li, W., Xu, Z., Sun, L., et al. (2012). 5-Azacytidine induces cardiac differentiation of human umbilical cord-derived mesenchymal stem cells by activating extracellular regulated kinase. Stem Cells Dev. 21, 67–75.
Quaini, F., Urbanek, K., Beltrami, A.P., Finato, N., Beltrami, C.A., Nadal-Ginard, B., Kajstura, J., Leri, A., and Anversa, P. (2002). Chimerism of the transplanted heart. N. Engl. J. Med. 346, 5–15.
Quijada, P., Hariharan, N., Cubillo, J.D., Bala, K.M., Emathinger, J.M., Wang, B.J., Ormachea, L., Bers, D.M., Sussman, M.A., and Poizat, C. (2015). Nuclear Calcium/Calmodulin-dependent Protein Kinase II

Références bibliographiques
255
Signaling Enhances Cardiac Progenitor Cell Survival and Cardiac Lineage Commitment. J. Biol. Chem. 290, 25411–25426.
Quint, P., Ruan, M., Pederson, L., Kassem, M., Westendorf, J.J., Khosla, S., and Oursler, M.J. (2013). Sphingosine 1-Phosphate (S1P) Receptors 1 and 2 Coordinately Induce Mesenchymal Cell Migration through S1P Activation of Complementary Kinase Pathways. J. Biol. Chem. 288, 5398–5406.
Rahman, F., Al Frouh, F., Bordignon, B., Fraterno, M., Landrier, J.-F., Peiretti, F., and Fontes, M. (2014). Ascorbic acid is a dose-dependent inhibitor of adipocyte differentiation, probably by reducing cAMP pool. Front. Cell Dev. Biol. 2, 29.
Rahman, F., Bordignon, B., Culerrier, R., Peiretti, F., Spicuglia, S., Djabali, M., Landrier, J.F., and Fontes, M. (2017). Ascorbic acid drives the differentiation of mesoderm-derived embryonic stem cells. Involvement of p38 MAPK/CREB and SVCT2 transporter. Mol. Nutr. Food Res. 61.
Rastegar, F., Shenaq, D., Huang, J., Zhang, W., Zhang, B.-Q., He, B.-C., Chen, L., Zuo, G.-W., Luo, Q., Shi, Q., et al. (2010). Mesenchymal stem cells: Molecular characteristics and clinical applications. World J. Stem Cells 2, 67–80.
Ratajczak, M.Z., Suszynska, M., Borkowska, S., Ratajczak, J., and Schneider, G. (2014). The role of sphingosine-1-phosphate (S1P) and ceramide-1-phosphate (C1P) in the trafficking of normal and malignant cells. Expert Opin. Ther. Targets 18, 95–107.
Reik, W. (2007). Stability and flexibility of epigenetic gene regulation in mammalian development. Nature 447, 425–432.
Reinecke, H., Poppa, V., and Murry, C.E. (2002). Skeletal muscle stem cells do not transdifferentiate into cardiomyocytes after cardiac grafting. J. Mol. Cell. Cardiol. 34, 241–249.
Reiss, K., Kajstura, J., Zhang, X., Li, P., Szoke, E., Olivetti, G., and Anversa, P. (1994). Acute myocardial infarction leads to upregulation of the IGF-1 autocrine system, DNA replication, and nuclear mitotic division in the remaining viable cardiac myocytes. Exp. Cell Res. 213, 463–472.
Resende, R.R., da Costa, J.L., Kihara, A.H., Adhikari, A., and Lorençon, E. (2010). Intracellular Ca2+ regulation during neuronal differentiation of murine embryonal carcinoma and mesenchymal stem cells. Stem Cells Dev. 19, 379–394.
Reus, T.L., Robert, A.W., Da Costa, M.B.A., de Aguiar, A.M., and Stimamiglio, M.A. (2016). Secretome from resident cardiac stromal cells stimulates proliferation, cardiomyogenesis and angiogenesis of progenitor cells. Int. J. Cardiol. 221, 396–403.
Reya, T., and Clevers, H. (2005). Wnt signalling in stem cells and cancer. Nature 434, 843–850.
Rignault-Clerc, S., Bielmann, C., Liaudet, L., Waeber, B., Feihl, F., and Rosenblatt-Velin, N. (2017). Natriuretic Peptide Receptor B modulates the proliferation of the cardiac cells expressing the Stem Cell Antigen-1. Sci. Rep. 7, 41936.
van de Rijn, M., Heimfeld, S., Spangrude, G.J., and Weissman, I.L. (1989). Mouse hematopoietic stem-cell antigen Sca-1 is a member of the Ly-6 antigen family. Proc. Natl. Acad. Sci. U. S. A. 86, 4634–4638.
Risau, W., and Flamme, I. (1995). Vasculogenesis. Annu. Rev. Cell Dev. Biol. 11, 73–91.

Références bibliographiques
256
Robert, P., Tsui, P., Laville, M.P., Livi, G.P., Sarau, H.M., Bril, A., and Berrebi-Bertrand, I. (2001). EDG1 receptor stimulation leads to cardiac hypertrophy in rat neonatal myocytes. J. Mol. Cell. Cardiol. 33, 1589–1606.
Rodríguez-Gómez, J.A., Levitsky, K.L., and López-Barneo, J. (2012). T-type Ca2+ channels in mouse embryonic stem cells: modulation during cell cycle and contribution to self-renewal. Am. J. Physiol. Cell Physiol. 302, C494-504.
Rosen, H., Stevens, R., Hanson, M., Roberts, E. and Oldstone, M. (2013). Sphingosine-1-Phosphate
and Its Receptors: Structure, Signaling, and Influence. Annual Review of Biochemistry. 82, 637-662.
Rosenblatt-Velin, N., Lepore, M.G., Cartoni, C., Beermann, F., and Pedrazzini, T. (2005). FGF-2 controls the differentiation of resident cardiac precursors into functional cardiomyocytes. J. Clin. Invest. 115, 1724–1733.
Rossini, A., Frati, C., Lagrasta, C., Graiani, G., Scopece, A., Cavalli, S., Musso, E., Baccarin, M., Di Segni, M., Fagnoni, F., et al. (2011). Human cardiac and bone marrow stromal cells exhibit distinctive properties related to their origin. Cardiovasc. Res. 89, 650–660.
Rubart, M., Soonpaa, M.H., Nakajima, H., and Field, L.J. (2004). Spontaneous and evoked intracellular calcium transients in donor-derived myocytes following intracardiac myoblast transplantation. J. Clin. Invest. 114, 775–783.
Rumyantsev, P.P., and Borisov, A. (1987). DNA synthesis in myocytes from different myocardial compartments of young rats in norm, after experimental infarction and in vitro. Biomed. Biochim. Acta 46, S610-615.
Ryoo, K., and Park, J.-Y. (2016). Two-pore Domain Potassium Channels in Astrocytes. Exp. Neurobiol. 25, 222–232.
Ryu, Y., Takuwa, N., Sugimoto, N., Sakurada, S., Usui, S., Okamoto, H., Matsui, O., and Takuwa, Y. (2002). Sphingosine-1-phosphate, a platelet-derived lysophospholipid mediator, negatively regulates cellular Rac activity and cell migration in vascular smooth muscle cells. Circ. Res. 90, 325–332.
Sackstein, R. (2004). The Bone Marrow Is Akin to Skin: HCELL and the Biology of Hematopoietic Stem Cell Homing11Reprinted from J Invest Dermatol 122:1061-1069, 2004. J. Investig. Dermatol. Symp. Proc. 9, 215–223.
Sakaue, M., and Sieber-Blum, M. (2015). Human epidermal neural crest stem cells as a source of Schwann cells. Dev. Camb. Engl. 142, 3188–3197.
Salomone, S., and Waeber, C. (2011). Selectivity and specificity of sphingosine-1-phosphate receptor ligands: caveats and critical thinking in characterizing receptor-mediated effects. Front. Pharmacol. 2, 9.
Sanna, M.G., Liao, J., Jo, E., Alfonso, C., Ahn, M.-Y., Peterson, M.S., Webb, B., Lefebvre, S., Chun, J., Gray, N., et al. (2004). Sphingosine 1-phosphate (S1P) receptor subtypes S1P1 and S1P3, respectively, regulate lymphocyte recirculation and heart rate. J. Biol. Chem. 279, 13839–13848.
Sassoli, C., Frati, A., Tani, A., Anderloni, G., Pierucci, F., Matteini, F., Chellini, F., Zecchi Orlandini, S., Formigli, L., and Meacci, E. (2014). Mesenchymal stromal cell secreted sphingosine 1-phosphate (S1P) exerts a stimulatory effect on skeletal myoblast proliferation. PloS One 9, e108662.

Références bibliographiques
257
Sato, H., Takahashi, M., Ise, H., Yamada, A., Hirose, S., Tagawa, Y., Morimoto, H., Izawa, A., and Ikeda, U. (2006). Collagen synthesis is required for ascorbic acid-enhanced differentiation of mouse embryonic stem cells into cardiomyocytes. Biochem. Biophys. Res. Commun. 342, 107–112.
Schaarschmidt, G., Wegner, F., Schwarz, S.C., Schmidt, H., and Schwarz, J. (2009). Characterization of voltage-gated potassium channels in human neural progenitor cells. PloS One 4, e6168.
Scharbrodt, W., Abdallah, Y., Kasseckert, S.A., Gligorievski, D., Piper, H.M., Böker, D.-K., Deinsberger, W., and Oertel, M.F. (2009). Cytosolic Ca2+ oscillations in human cerebrovascular endothelial cells after subarachnoid hemorrhage. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 29, 57–65.
Schiller, M., Javelaud, D., and Mauviel, A. (2004). TGF-beta-induced SMAD signaling and gene regulation: consequences for extracellular matrix remodeling and wound healing. J. Dermatol. Sci. 35, 83–92.
Schriefer, J.A., Lewis, P.R., and Miller, J.W. (1982). Role of fetal oxytocin in parturition in the rat. Biol. Reprod. 27, 362–368.
Schulz, R.A., and Yutzey, K.E. (2004). Calcineurin signaling and NFAT activation in cardiovascular and skeletal muscle development. Dev. Biol. 266, 1–16.
Sedan, O., Dolnikov, K., Zeevi-Levin, N., Leibovich, N., Amit, M., Itskovitz-Eldor, J., and Binah, O. (2008). 1,4,5-Inositol trisphosphate-operated intracellular Ca(2+) stores and angiotensin-II/endothelin-1 signaling pathway are functional in human embryonic stem cell-derived cardiomyocytes. Stem Cells Dayt. Ohio 26, 3130–3138.
Seino, S. (1999). ATP-sensitive potassium channels: a model of heteromultimeric potassium channel/receptor assemblies. Annu. Rev. Physiol. 61, 337–362.
Seki, T., and Arai, Y. (1993). Highly polysialylated neural cell adhesion molecule (NCAM-H) is expressed by newly generated granule cells in the dentate gyrus of the adult rat. J. Neurosci. Off. J. Soc. Neurosci. 13, 2351–2358.
Seok, H.Y., Chen, J., Kataoka, M., Huang, Z.-P., Ding, J., Yan, J., Hu, X., and Wang, D.-Z. (2014). Loss of MicroRNA-155 protects the heart from pathological cardiac hypertrophy. Circ. Res. 114, 1585–1595.
Serradifalco, C., Catanese, P., Rizzuto, L., Cappello, F., Puleio, R., Barresi, V., Nunnari, C.M., Zummo, G., and Di Felice, V. (2011). Embryonic and foetal Islet-1 positive cells in human hearts are also positive to c-Kit. Eur. J. Histochem. EJH 55, e41.
Shi, S., Wu, X., Wang, X., Hao, W., Miao, H., Zhen, L., and Nie, S. (2016). Differentiation of Bone Marrow Mesenchymal Stem Cells to Cardiomyocyte-Like Cells Is Regulated by the Combined Low Dose Treatment of Transforming Growth Factor-β1 and 5-Azacytidine. Stem Cells Int. 2016, 3816256.
Shin, H.Y., Hong, Y.H., Jang, S.S., Chae, H.G., Paek, S.L., Moon, H.E., Kim, D.G., Kim, J., Paek, S.H., and Kim, S.J. (2010). A role of canonical transient receptor potential 5 channel in neuronal differentiation from A2B5 neural progenitor cells. PloS One 5, e10359.
Short, B., Brouard, N., Driessen, R., and Simmons, P.J. (2001). Prospective isolation of stromal progenitor cells from mouse BM. Cytotherapy 3, 407–408.

Références bibliographiques
258
Si, Y., Zhao, Y., Hao, H., Liu, J., Guo, Y., Mu, Y., Shen, J., Cheng, Y., Fu, X., and Han, W. (2012). Infusion of mesenchymal stem cells ameliorates hyperglycemia in type 2 diabetic rats: identification of a novel role in improving insulin sensitivity. Diabetes 61, 1616–1625.
“i i iak, T., Kala ski, R., Fisze , D., Je z ko ska, O., Rzeź i zak, J., Roz ado ska, N., a d Ku pisz, M. (2004). Autologous skeletal myoblast transplantation for the treatment of postinfarction myocardial injury: phase I clinical study with 12 months of follow-up. Am. Heart J. 148, 531–537.
Simpson, D.L., Mishra, R., Sharma, S., Goh, S.K., Deshmukh, S., and Kaushal, S. (2012). A strong regenerative ability of cardiac stem cells derived from neonatal hearts. Circulation 126, S46-53.
Singla, D.K., Kumar, D., and Sun, B. (2005). Transforming growth factor-beta2 enhances differentiation of cardiac myocytes from embryonic stem cells. Biochem. Biophys. Res. Commun. 332, 135–141.
Slager, H.G., Van Inzen, W., Freund, E., Van den Eijnden-Van Raaij, A.J., and Mummery, C.L. (1993). Transforming growth factor-beta in the early mouse embryo: implications for the regulation of muscle formation and implantation. Dev. Genet. 14, 212–224.
Smedler, E., and Uhlén, P. (2014). Frequency decoding of calcium oscillations. Biochim. Biophys. Acta 1840, 964–969.
Smith, D.O., Rosenheimer, J.L., and Kalil, R.E. (2008). Delayed rectifier and A-type potassium channels associated with Kv 2.1 and Kv 4.3 expression in embryonic rat neural progenitor cells. PloS One 3, e1604.
Smith, R.R., Barile, L., Cho, H.C., Leppo, M.K., Hare, J.M., Messina, E., Giacomello, A., Abraham, M.R., and Marbán, E. (2007). Regenerative potential of cardiosphere-derived cells expanded from percutaneous endomyocardial biopsy specimens. Circulation 115, 896–908.
Sobiesiak, M., Sivasubramaniyan, K., Hermann, C., Tan, C., Orgel, M., Treml, S., Cerabona, F., de Zwart, P., Ochs, U., Müller, C.A., et al. (2010). The mesenchymal stem cell antigen MSCA-1 is identical to tissue non-specific alkaline phosphatase. Stem Cells Dev. 19, 669–677.
Solanas, G., and Benitah, S.A. (2013). Regenerating the skin: a task for the heterogeneous stem cell pool and surrounding niche. Nat. Rev. Mol. Cell Biol. 14, 737–748.
Song, S., Li, J., Zhu, L., Cai, L., Xu, Q., Ling, C., Su, Y., and Hu, Q. (2012). Irregular Ca(2+) oscillations regulate transcription via cumulative spike duration and spike amplitude. J. Biol. Chem. 287, 40246–40255.
Soonpaa, M.H., and Field, L.J. (1998). Survey of studies examining mammalian cardiomyocyte DNA synthesis. Circ. Res. 83, 15–26.
Spiegel, S., and Milstien, S. (2003). Sphingosine-1-phosphate: an enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 4, 397–407.
Stadtfeld, M., Apostolou, E., Ferrari, F., Choi, J., Walsh, R.M., Chen, T., Ooi, S.S.K., Kim, S.Y., Bestor, T.H., Shioda, T., et al. (2012). Ascorbic acid prevents loss of Dlk1-Dio3 imprinting and facilitates generation of all-iPS cell mice from terminally differentiated B cells. Nat. Genet. 44, 398–405, S1-2.
Stainier, D.Y., and Fishman, M.C. (1992). Patterning the zebrafish heart tube: acquisition of anteroposterior polarity. Dev. Biol. 153, 91–101.

Références bibliographiques
259
Stewart, R.R., Zigova, T., and Luskin, M.B. (1999). Potassium currents in precursor cells isolated from the anterior subventricular zone of the neonatal rat forebrain. J. Neurophysiol. 81, 95–102.
Stieber, J., Herrmann, S., Feil, S., Löster, J., Feil, R., Biel, M., Hofmann, F., and Ludwig, A. (2003). The hyperpolarization-activated channel HCN4 is required for the generation of pacemaker action potentials in the embryonic heart. Proc. Natl. Acad. Sci. U. S. A. 100, 15235–15240.
Stroh, A., Tsai, H.-C., Wang, L.-P., Zhang, F., Kressel, J., Aravanis, A., Santhanam, N., Deisseroth, K., Konnerth, A., and Schneider, M.B. (2011). Tracking stem cell differentiation in the setting of automated optogenetic stimulation. Stem Cells Dayt. Ohio 29, 78–88.
Studeny, M., Marini, F.C., Dembinski, J.L., Zompetta, C., Cabreira-Hansen, M., Bekele, B.N., Champlin, R.E., and Andreeff, M. (2004). Mesenchymal stem cells: potential precursors for tumor stroma and targeted-delivery vehicles for anticancer agents. J. Natl. Cancer Inst. 96, 1593–1603.
Stühmer, W., Ruppersberg, J.P., Schröter, K.H., Sakmann, B., Stocker, M., Giese, K.P., Perschke, A., Baumann, A., and Pongs, O. (1989). Molecular basis of functional diversity of voltage-gated potassium channels in mammalian brain. EMBO J. 8, 3235–3244.
Sun, S., Liu, Y., Lipsky, S., and Cho, M. (2007). Physical manipulation of calcium oscillations facilitates osteodifferentiation of human mesenchymal stem cells. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 21, 1472–1480.
Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M., Ichisaka, T., Tomoda, K., and Yamanaka, S. (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131, 861–872.
Takuwa, Y., Ikeda, H., Okamoto, Y., Takuwa, N., and Yoshioka, K. (2013). Sphingosine-1-phosphate as a mediator involved in development of fibrotic diseases. Biochim. Biophys. Acta 1831, 185–192.
Tallini, Y.N., Greene, K.S., Craven, M., Spealman, A., Breitbach, M., Smith, J., Fisher, P.J., Steffey, M., Hesse, M., Doran, R.M., et al. (2009). c-kit expression identifies cardiovascular precursors in the neonatal heart. Proc. Natl. Acad. Sci. U. S. A. 106, 1808–1813.
Tang, B., Li, Y., Nagaraj, C., Morty, R.E., Gabor, S., Stacher, E., Voswinckel, R., Weissmann, N., Leithner, K., Olschewski, H., et al. (2009). Endothelin-1 inhibits background two-pore domain channel TASK-1 in primary human pulmonary artery smooth muscle cells. Am. J. Respir. Cell Mol. Biol. 41, 476–483.
Tao, R., Lau, C.-P., Tse, H.-F., and Li, G.-R. (2007). Functional ion channels in mouse bone marrow mesenchymal stem cells. Am. J. Physiol. Cell Physiol. 293, C1561-1567.
Tao, R., Lau, C.-P., Tse, H.-F., and Li, G.-R. (2008). Regulation of cell proliferation by intermediate-conductance Ca2+-activated potassium and volume-sensitive chloride channels in mouse mesenchymal stem cells. Am. J. Physiol. Cell Physiol. 295, C1409-1416.
Tarnowski, M., and Sieron, A.L. (2006). Adult stem cells and their ability to differentiate. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 12, RA154-163.
Taupin, P. (2006). Neural progenitor and stem cells in the adult central nervous system. Ann. Acad. Med. Singapore 35, 814–820.
Thomson, J.A., Itskovitz-Eldor, J., Shapiro, S.S., Waknitz, M.A., Swiergiel, J.J., Marshall, V.S., and Jones, J.M. (1998). Embryonic stem cell lines derived from human blastocysts. Science 282, 1145–1147.

Références bibliographiques
260
Tognon, C.E., Kirk, H.E., Passmore, L.A., Whitehead, I.P., Der, C.J., and Kay, R.J. (1998). Regulation of RasGRP via a Phorbol Ester-Responsive C1 Domain. Mol. Cell. Biol. 18, 6995–7008.
Toma, C., Pittenger, M.F., Cahill, K.S., Byrne, B.J., and Kessler, P.D. (2002). Human mesenchymal stem cells differentiate to a cardiomyocyte phenotype in the adult murine heart. Circulation 105, 93–98.
Toma, J.G., Akhavan, M., Fernandes, K.J., Barnabé-Heider, F., Sadikot, A., Kaplan, D.R., and Miller, F.D. (2001). Isolation of multipotent adult stem cells from the dermis of mammalian skin. Nat. Cell Biol. 3, 778–784.
Tomida, T., Hirose, K., Takizawa, A., Shibasaki, F., and Iino, M. (2003). NFAT functions as a working memory of Ca2+ signals in decoding Ca2+ oscillation. EMBO J. 22, 3825–3832.
Tomita, Y., Matsumura, K., Wakamatsu, Y., Matsuzaki, Y., Shibuya, I., Kawaguchi, H., Ieda, M., Kanakubo, S., Shimazaki, T., Ogawa, S., et al. (2005). Cardiac neural crest cells contribute to the dormant multipotent stem cell in the mammalian heart. J. Cell Biol. 170, 1135–1146.
Tompa, P., Töth-Boconádi, R., and Friedrich, P. (2001). Frequency decoding of fast calcium oscillations by calpain. Cell Calcium 29, 161–170.
Tonelli, F.M.P., Santos, A.K., Gomes, D.A., da Silva, S.L., Gomes, K.N., Ladeira, L.O., and Resende, R.R. (2012). Stem cells and calcium signaling. Adv. Exp. Med. Biol. 740, 891–916.
Toro, L., Li, M., Zhang, Z., Singh, H., Wu, Y., and Stefani, E. (2014). MaxiK channel and cell signalling. Pflugers Arch. 466, 875–886.
Tran, T.D.N., Zolochevska, O., Figueiredo, M.L., Wang, H., Yang, L.-J., Gimble, J.M., Yao, S., and Cheng, H. (2014). Histamine-induced Ca2+ signalling is mediated by TRPM4 channels in human adipose-derived stem cells. Biochem. J. 463, 123–134.
Tran, T.D.N., Yao, S., Hsu, W.H., Gimble, J.M., Bunnell, B.A., and Cheng, H. (2015). Arginine vasopressin inhibits adipogenesis in human adipose-derived stem cells. Mol. Cell. Endocrinol. 406, 1–9.
Tropea, K.A., Leder, E., Aslam, M., Lau, A.N., Raiser, D.M., Lee, J.-H., Balasubramaniam, V., Fredenburgh, L.E., Alex Mitsialis, S., Kourembanas, S., et al. (2012). Bronchioalveolar stem cells increase after mesenchymal stromal cell treatment in a mouse model of bronchopulmonary dysplasia. Am. J. Physiol. Lung Cell. Mol. Physiol. 302, L829-837.
Tsang, W.Y., Spektor, A., Luciano, D.J., Indjeian, V.B., Chen, Z., Salisbury, J.L., Sánchez, I., and Dynlacht, B.D. (2006). CP110 cooperates with two calcium-binding proteins to regulate cytokinesis and genome stability. Mol. Biol. Cell 17, 3423–3434.
Uhlén, P., Burch, P.M., Zito, C.I., Estrada, M., Ehrlich, B.E., and Bennett, A.M. (2006). Gain-of-function/Noonan syndrome SHP-2/Ptpn11 mutants enhance calcium oscillations and impair NFAT signaling. Proc. Natl. Acad. Sci. U. S. A. 103, 2160–2165.
Ulloa-Montoya, F., Verfaillie, C.M., and Hu, W.-S. (2005). Culture systems for pluripotent stem cells. J. Biosci. Bioeng. 100, 12–27.
Urbanek, K., Quaini, F., Tasca, G., Torella, D., Castaldo, C., Nadal-Ginard, B., Leri, A., Kajstura, J., Quaini, E., and Anversa, P. (2003). Intense myocyte formation from cardiac stem cells in human cardiac hypertrophy. Proc. Natl. Acad. Sci. U. S. A. 100, 10440–10445.

Références bibliographiques
261
Urbanek, K., Rota, M., Cascapera, S., Bearzi, C., Nascimbene, A., De Angelis, A., Hosoda, T., Chimenti, S., Baker, M., Limana, F., et al. (2005). Cardiac stem cells possess growth factor-receptor systems that after activation regenerate the infarcted myocardium, improving ventricular function and long-term survival. Circ. Res. 97, 663–673.
Vacanti, V., Kong, E., Suzuki, G., Sato, K., Canty, J.M., and Lee, T. (2005). Phenotypic changes of adult porcine mesenchymal stem cells induced by prolonged passaging in culture. J. Cell. Physiol. 205, 194–201.
Valente, M., Nascimento, D.S., Cumano, A., and Pinto-do-Ó, P. (2014). Sca-1+ cardiac progenitor cells and heart-making: a critical synopsis. Stem Cells Dev. 23, 2263–2273.
Van Haele, M., and Roskams, T. (2017). Hepatic Progenitor Cells: An Update. Gastroenterol. Clin. North Am. 46, 409–420.
Vecellio, M., Meraviglia, V., Nanni, S., Barbuti, A., Scavone, A., DiFrancesco, D., Farsetti, A., Pompilio, G., Colombo, G.I., Capogrossi, M.C., et al. (2012). In vitro epigenetic reprogramming of human cardiac mesenchymal stromal cells into functionally competent cardiovascular precursors. PloS One 7, e51694.
van Vliet, P., Roccio, M., Smits, A.M., van Oorschot, A. a. M., Metz, C.H.G., van Veen, T. a. B., Sluijter, J.P.G., Doevendans, P.A., and Goumans, M.-J. (2008). Progenitor cells isolated from the human heart: a potential cell source for regenerative therapy. Neth. Heart J. Mon. J. Neth. Soc. Cardiol. Neth. Heart Found. 16, 163–169.
van Vliet, P., de Boer, T.P., van der Heyden, M.A.G., El Tamer, M.K., Sluijter, J.P.G., Doevendans, P.A., and Goumans, M.-J. (2010). Hyperpolarization induces differentiation in human cardiomyocyte progenitor cells. Stem Cell Rev. 6, 178–185.
Wakitani, S., Saito, T., and Caplan, A.I. (1995). Myogenic cells derived from rat bone marrow mesenchymal stem cells exposed to 5-azacytidine. Muscle Nerve 18, 1417–1426.
Waldron, G.J., and Cole, W.C. (1999). Activation of vascular smooth muscle K+ channels by endothelium-derived relaxing factors. Clin. Exp. Pharmacol. Physiol. 26, 180–184.
Walker, S.A., Kupzig, S., Bouyoucef, D., Davies, L.C., Tsuboi, T., Bivona, T.G., Cozier, G.E., Lockyer, P.J., Buckler, A., Rutter, G.A., et al. (2004). Identification of a Ras GTPase‐a ti ati g p otei egulated e epto ‐ ediated Ca + os illatio s. EMBO J. 23, 1749–1760.
Walkup, M.H., and Gerber, D.A. (2006). Hepatic stem cells: in search of. Stem Cells Dayt. Ohio 24, 1833–1840.
Wang, B., Borazjani, A., Tahai, M., Curry, A.L. de J., Simionescu, D.T., Guan, J., To, F., Elder, S.H., and Liao, J. (2010). Fabrication of cardiac patch with decellularized porcine myocardial scaffold and bone marrow mononuclear cells. J. Biomed. Mater. Res. A 94, 1100–1110.
Wang, F., Van Brocklyn, J.R., Hobson, J.P., Movafagh, S., Zukowska-Grojec, Z., Milstien, S., and Spiegel, S. (1999a). Sphingosine 1-phosphate stimulates cell migration through a G(i)-coupled cell surface receptor. Potential involvement in angiogenesis. J. Biol. Chem. 274, 35343–35350.
Wang, G., Dayanithi, G., Newcomb, R., and Lemos, J.R. (1999b). An R-type Ca(2+) current in neurohypophysial terminals preferentially regulates oxytocin secretion. J. Neurosci. Off. J. Soc. Neurosci. 19, 9235–9241.

Références bibliographiques
262
Wang, K., Xue, T., Tsang, S.-Y., Van Huizen, R., Wong, C.W., Lai, K.W., Ye, Z., Cheng, L., Au, K.W., Zhang, J., et al. (2005). Electrophysiological properties of pluripotent human and mouse embryonic stem cells. Stem Cells Dayt. Ohio 23, 1526–1534.
Wang, S.-P., Wang, J.-A., Luo, R.-H., Cui, W.-Y., and Wang, H. (2008). Potassium channel currents in rat mesenchymal stem cells and their possible roles in cell proliferation. Clin. Exp. Pharmacol. Physiol. 35, 1077–1084.
Wang, T., Chen, K., Zeng, X., Yang, J., Wu, Y., Shi, X., Qin, B., Zeng, L., Esteban, M.A., Pan, G., et al. (2011). The histone demethylases Jhdm1a/1b enhance somatic cell reprogramming in a vitamin-C-dependent manner. Cell Stem Cell 9, 575–587.
Wang, X., Hu, Q., Nakamura, Y., Lee, J., Zhang, G., From, A.H.L., and Zhang, J. (2006). The role of the sca-1+/CD31- cardiac progenitor cell population in postinfarction left ventricular remodeling. Stem Cells Dayt. Ohio 24, 1779–1788.
Waring, C.D., Vicinanza, C., Papalamprou, A., Smith, A.J., Purushothaman, S., Goldspink, D.F., Nadal-Ginard, B., Torella, D., and Ellison, G.M. (2014). The adult heart responds to increased workload with physiologic hypertrophy, cardiac stem cell activation, and new myocyte formation. Eur. Heart J. 35, 2722–2731.
Watt, F.M. (2002). Role of integrins in regulating epidermal adhesion, growth and differentiation. EMBO J. 21, 3919–3926.
Weick, J.P., Austin Johnson, M., and Zhang, S.-C. (2009). Developmental regulation of human embryonic stem cell-derived neurons by calcium entry via transient receptor potential channels. Stem Cells Dayt. Ohio 27, 2906–2916.
Welm, B.E., Tepera, S.B., Venezia, T., Graubert, T.A., Rosen, J.M., and Goodell, M.A. (2002). Sca-1(pos) cells in the mouse mammary gland represent an enriched progenitor cell population. Dev. Biol. 245, 42–56.
Wiese, C., Nikolova, T., Zahanich, I., Sulzbacher, S., Fuchs, J., Yamanaka, S., Graf, E., Ravens, U., Boheler, K.R., and Wobus, A.M. (2011). Differentiation induction of mouse embryonic stem cells into sinus node-like cells by suramin. Int. J. Cardiol. 147, 95–111.
Williams, A.R., and Hare, J.M. (2011). Mesenchymal stem cells: Biology, patho-physiology, translational findings, and therapeutic implications for cardiac disease. Circ. Res. 109, 923–940.
Windh, R.T., Lee, M.J., Hla, T., An, S., Barr, A.J., and Manning, D.R. (1999). Differential coupling of the sphingosine 1-phosphate receptors Edg-1, Edg-3, and H218/Edg-5 to the G(i), G(q), and G(12) families of heterotrimeric G proteins. J. Biol. Chem. 274, 27351–27358.
Winitsky, S.O., Gopal, T.V., Hassanzadeh, S., Takahashi, H., Gryder, D., Rogawski, M.A., Takeda, K., Yu, Z.X., Xu, Y.H., and Epstein, N.D. (2005). Adult murine skeletal muscle contains cells that can differentiate into beating cardiomyocytes in vitro. PLoS Biol. 3, e87.
Wobus, A.M., Kaomei, G., Shan, J., Wellner, M.C., Rohwedel, J., Ji Guanju, null, Fleischmann, B., Katus, H.A., Hescheler, J., and Franz, W.M. (1997). Retinoic acid accelerates embryonic stem cell-derived cardiac differentiation and enhances development of ventricular cardiomyocytes. J. Mol. Cell. Cardiol. 29, 1525–1539.

Références bibliographiques
263
Woodbury, D., Schwarz, E.J., Prockop, D.J., and Black, I.B. (2000). Adult rat and human bone marrow stromal cells differentiate into neurons. J. Neurosci. Res. 61, 364–370.
Wu, X., Chang, B., Blair, N.S., Sargent, M., York, A.J., Robbins, J., Shull, G.E., and Molkentin, J.D. (2009). Plasma membrane Ca2+-ATPase isoform 4 antagonizes cardiac hypertrophy in association with calcineurin inhibition in rodents. J. Clin. Invest. 119, 976–985.
Wulf, G.G., Luo, K.-L., Jackson, K.A., Brenner, M.K., and Goodell, M.A. (2003). Cells of the hepatic side population contribute to liver regeneration and can be replenished with bone marrow stem cells. Haematologica 88, 368–378.
Xin, L., Lawson, D.A., and Witte, O.N. (2005). The Sca-1 cell surface marker enriches for a prostate-regenerating cell subpopulation that can initiate prostate tumorigenesis. Proc. Natl. Acad. Sci. U. S. A. 102, 6942–6947.
Xiu, L., Chang, N., Yang, L., Liu, X., Yang, L., Ge, J., and Li, L. (2015). Intracellular sphingosine 1-phosphate contributes to collagen expression of hepatic myofibroblasts in human liver fibrosis independent of its receptors. Am. J. Pathol. 185, 387–398.
Xu, W., Zhang, X., Qian, H., Zhu, W., Sun, X., Hu, J., Zhou, H., and Chen, Y. (2004). Mesenchymal stem cells from adult human bone marrow differentiate into a cardiomyocyte phenotype in vitro. Exp. Biol. Med. Maywood NJ 229, 623–631.
Yanagida, E., Shoji, S., Hirayama, Y., Yoshikawa, F., Otsu, K., Uematsu, H., Hiraoka, M., Furuichi, T., and Kawano, S. (2004). Functional expression of Ca2+ signaling pathways in mouse embryonic stem cells. Cell Calcium 36, 135–146.
Yang, L., Soonpaa, M.H., Adler, E.D., Roepke, T.K., Kattman, S.J., Kennedy, M., Henckaerts, E., Bonham, K., Abbott, G.W., Linden, R.M., et al. (2008). Human cardiovascular progenitor cells develop from a KDR+ embryonic-stem-cell-derived population. Nature 453, 524–528.
Yang, X., Pabon, L., and Murry, C.E. (2014). Engineering adolescence: maturation of human pluripotent stem cell-derived cardiomyocytes. Circ. Res. 114, 511–523.
Yang, Y.-Q., Gharibeh, L., Li, R.-G., Xin, Y.-F., Wang, J., Liu, Z.-M., Qiu, X.-B., Xu, Y.-J., Xu, L., Qu, X.-K., et al. (2013a). GATA4 loss-of-function mutations underlie familial tetralogy of fallot. Hum. Mutat. 34, 1662–1671.
Yang, Z.X., Han, Z.-B., Ji, Y.R., Wang, Y.W., Liang, L., Chi, Y., Yang, S.G., Li, L.N., Luo, W.F., Li, J.P., et al. (2013b). CD106 identifies a subpopulation of mesenchymal stem cells with unique immunomodulatory properties. PloS One 8, e59354.
Yarden, Y., Kuang, W.J., Yang-Feng, T., Coussens, L., Munemitsu, S., Dull, T.J., Chen, E., Schlessinger, J., Francke, U., and Ullrich, A. (1987). Human proto-oncogene c-kit: a new cell surface receptor tyrosine kinase for an unidentified ligand. EMBO J. 6, 3341–3351.
Yasuda, T., and Adams, D.J. (2010). Physiological roles of ion channels in adult neural stem cells and their progeny. J. Neurochem. 114, 946–959.
Yasuda, T., Bartlett, P.F., and Adams, D.J. (2008). K(ir) and K(v) channels regulate electrical properties and proliferation of adult neural precursor cells. Mol. Cell. Neurosci. 37, 284–297.

Références bibliographiques
264
Ye, B. (2010). Ca2+ oscillations and its transporters in mesenchymal stem cells. Physiol. Res. 59, 323–329.
Ye, N.-S., Zhang, R.-L., Zhao, Y.-F., Feng, X., Wang, Y.-M., and Luo, G.-A. (2006). Effect of 5-azacytidine on the protein expression of porcine bone marrow mesenchymal stem cells in vitro. Genomics Proteomics Bioinformatics 4, 18–25.
Yi, M., Zhao, Q., Tang, J., and Wang, C. (2011). A theoretical modeling for frequency modulation of Ca2+ signal on activation of MAPK cascade. Biophys. Chem. 157, 33–42.
Yoo, J.U., Barthel, T.S., Nishimura, K., Solchaga, L., Caplan, A.I., Goldberg, V.M., and Johnstone, B. (1998). The chondrogenic potential of human bone-marrow-derived mesenchymal progenitor cells. J. Bone Joint Surg. Am. 80, 1745–1757.
Young, R.G., Butler, D.L., Weber, W., Caplan, A.I., Gordon, S.L., and Fink, D.J. (1998). Use of mesenchymal stem cells in a collagen matrix for Achilles tendon repair. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 16, 406–413.
Yu, H.-M., Wen, J., Wang, R., Shen, W.-H., Duan, S., and Yang, H.-T. (2008). Critical role of type 2 ryanodine receptor in mediating activity-dependent neurogenesis from embryonic stem cells. Cell Calcium 43, 417–431.
Yuan, P., Leonetti, M.D., Pico, A.R., Hsiung, Y., and MacKinnon, R. (2010). Structure of the Human BK Channel Ca2+-Activation Apparatus at 3.0 Å Resolution. Science 329, 182–186.
Yutzey, K.E., Rhee, J.T., and Bader, D. (1994). Expression of the atrial-specific myosin heavy chain AMHC1 and the establishment of anteroposterior polarity in the developing chicken heart. Dev. Camb. Engl. 120, 871–883.
Zacharek, S.J., Fillmore, C.M., Lau, A.N., Gludish, D.W., Chou, A., Ho, J.W.K., Zamponi, R., Gazit, R., Bock, C., Jäger, N., et al. (2011). Lung stem cell self-renewal relies on BMI1-dependent control of expression at imprinted loci. Cell Stem Cell 9, 272–281.
Zahanich, I., Graf, E.M., Heubach, J.F., Hempel, U., Boxberger, S., and Ravens, U. (2005). Molecular and functional expression of voltage-operated calcium channels during osteogenic differentiation of human mesenchymal stem cells. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 20, 1637–1646.
Zammit, P.S., Partridge, T.A., and Yablonka-Reuveni, Z. (2006). The skeletal muscle satellite cell: the stem cell that came in from the cold. J. Histochem. Cytochem. Off. J. Histochem. Soc. 54, 1177–1191.
Zeineddine, D., Papadimou, E., Chebli, K., Gineste, M., Liu, J., Grey, C., Thurig, S., Behfar, A., Wallace, V.A., Skerjanc, I.S., et al. (2006). Oct-3/4 dose dependently regulates specification of embryonic stem cells toward a cardiac lineage and early heart development. Dev. Cell 11, 535–546.
Zhang, J., Honbo, N., Goetzl, E.J., Chatterjee, K., Karliner, J.S., and Gray, M.O. (2007). Signals from type 1 sphingosine 1-phosphate receptors enhance adult mouse cardiac myocyte survival during hypoxia. Am. J. Physiol. Heart Circ. Physiol. 293, H3150-3158.
Zhang, J., Zhang, L., Wu, Q., Liu, H., and Huang, L. (2013). Recombinant human brain natriuretic peptide therapy combined with bone mesenchymal stem cell transplantation for treating heart failure in rats. Mol. Med. Rep. 7, 628–632.

Références bibliographiques
265
Zhang, Y., Sivakumaran, P., Newcomb, A.E., Hernandez, D., Harris, N., Khanabdali, R., Liu, G.-S., Kelly, D.J., Pébay, A., Hewitt, A.W., et al. (2015a). Cardiac Repair With a Novel Population of Mesenchymal Stem Cells Resident in the Human Heart. Stem Cells Dayt. Ohio 33, 3100–3113.
Zhang, Y.-Y., Yue, J., Che, H., Sun, H.-Y., Tse, H.-F., and Li, G.-R. (2014b). BKCa and hEag1 channels regulate cell proliferation and differentiation in human bone marrow-derived mesenchymal stem cells. J. Cell. Physiol. 229, 202–212.
Zhang, Y.-Y., Li, G., Che, H., Sun, H.-Y., Li, X., Au, W.-K., Xiao, G.-S., Wang, Y., and Li, G.-R. (2014a). Characterization of functional ion channels in human cardiac c-kit+ progenitor cells. Basic Res. Cardiol. 109, 407.
Zhang, Y.-Y., Li, G., Che, H., Sun, H.-Y., Xiao, G.-S., Wang, Y., and Li, G.-R. (2015b). Effects of BKCa and Kir2.1 Channels on Cell Cycling Progression and Migration in Human Cardiac c-kit+ Progenitor Cells. PloS One 10, e0138581.
Zhang, Z., He, Y., Tuteja, D., Xu, D., Timofeyev, V., Zhang, Q., Glatter, K.A., Xu, Y., Shin, H.-S., Low, R., et al. (2005). Functional roles of Cav1.3(alpha1D) calcium channels in atria: insights gained from gene-targeted null mutant mice. Circulation 112, 1936–1944.
Zhao, W., Li, J.-J., Cao, D.-Y., Li, X., Zhang, L.-Y., He, Y., Yue, S.-Q., Wang, D.-S., and Dou, K.-F. (2012). Intravenous injection of mesenchymal stem cells is effective in treating liver fibrosis. World J. Gastroenterol. 18, 1048–1058.
Zhao, Z., Chen, Z., Zhao, X., Pan, F., Cai, M., Wang, T., Zhang, H., Lu, J.R., and Lei, M. (2011). Sphingosine-1-phosphate promotes the differentiation of human umbilical cord mesenchymal stem cells into cardiomyocytes under the designated culturing conditions. J. Biomed. Sci. 18, 37.
Zhou, Y.-F., Yang, X.-J., Li, H.-X., Han, L.-H., and Jiang, W.-P. (2013). Genetically-engineered mesenchymal stem cells transfected with human HCN1 gene to create cardiac pacemaker cells. J. Int. Med. Res. 41, 1570–1576.
Zhu, L., Luo, Y., Chen, T., Chen, F., Wang, T., and Hu, Q. (2008). Ca2+ oscillation frequency regulates agonist-stimulated gene expression in vascular endothelial cells. J. Cell Sci. 121, 2511–2518.
Zhu, L., Song, S., Pi, Y., Yu, Y., She, W., Ye, H., Su, Y., and Hu, Q. (2011). Cumulated Ca2+ spike duration underlies Ca2+ oscillation frequency-regulated NFκB transcriptional activity. J. Cell Sci. 124, 2591–2601.
Zimmer, T., Haufe, V., and Blechschmidt, S. (2014). Voltage-gated sodium channels in the mammalian heart. Glob. Cardiol. Sci. Pract. 2014, 449–463.
Zuk, P.A., Zhu, M., Ashjian, P., De Ugarte, D.A., Huang, J.I., Mizuno, H., Alfonso, Z.C., Fraser, J.K., Benhaim, P., and Hedrick, M.H. (2002). Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 13, 4279–4295.

Résumé
266
RESUME
Résumé
L’o je tif de ette th se tait de d eloppe et de a a t ise u od le de ellules sou hes
cardiaques humaines dans un contexte de thérapie cellulaire.
Ap s a oi s le tio et a a t is u e populatio de ellules sou hes d’o igi e
se h ateuse, isol e à pa ti d’au i ules hu ai es, e p i a t le a ueu W B C“Cs
W8B2+), nous nous sommes focalisés (par les techniques de RT-qPCR à haut rendement,
d’i u o-marquage, de western-blot et de fluorescence calcique) sur ; 1) la caractérisation
g i ue des a au io i ues et des a teu s de la sig alisatio al i ue et l’ tude de leu
diff e iatio i it o e pa all le à l’a ti it al i ue i t a ellulaire.
Les résultats montrent que CSCs W8B2+ tendent à se différencier en cellules pacemaker.
Certains gènes spécifiques nodaux, comme Tbx3, HCN, ICaT,L, KV, NCX, s’e p i e t du a t la
diff e iatio . L’e egist e e t de l’a ti it al i ue ia u e so de optogénétique) montre
la p se e d’os illatio s al i ues ui olue t e f ue e et e i te sit pe da t la
différenciation. Les stocks-IP se si les et l’ ha geu NCX joue aie t u ôle fo da e tal.
Nous a o s e suite tudi l’i po ta e du a al BKCa et des récepteurs sphingosine 1-
phosphate (S1P) dans la régulation des propriétés fondamentales des CSCs W8B2+.
L'inhibition du BKCa diminue la prolifération cellulaire en accumulant les cellules à la phase
G0/G1, réprime l'auto- e ou elle e t ais ’affe te pas la ig atio . Qua t à la “ P elle
f ei e la p olif atio et l’auto-renouvellement via une voie différente de celles des
récepteurs S1P1,2,3.
Ce travail fait ressortir des cibles moléculaires fondamentales dans un contexte de thérapie
cellulaire cardiaque.
Mots-clés : thérapie cellulaire, cellules souches cardiaques humaines, cellules
mésenchymateuses, différenciation, canaux ioniques, signalisation calcique, canal BKCa,
sphingosine 1-phosphate, optogénétique.

Résumé
267
Abstract
The aim of this thesis was to develop and characterize a model of human heart stem cells in a
context of cell therapy.
A population of mesenchymal stem cells, expressing the W8B2 marker (CSCs W8B2+), was first
isolated from human auricles and characterized using high-throughput RT-qPCR techniques,
immuno-labeling, western-blot and calcium fluorescence imaging. These experiments were
focused on 1) the gene expression of ion channels and calcium signaling proteins; and 2) the
study of CSCs W8B2+ in vitro differentiation and associated intracellular calcium activity
changes.
The results show that CSCs W8B2+ tend to differentiate into pacemaker cells. Some nodal
specific genes such as Tbx3, HCN, ICaT, L, KV, NCX, are expressed during differentiation. The
recording of calcium activity (via an optogenetic probe) shows the presence of calcium
oscillations that change in frequency and intensity during differentiation. IP3 sensitive calcium
stocks and the NCX exchanger would play a fundamental role in these variations.
Then we studied the importance of the BKCa channel and the sphingosine 1-phosphate (S1P)
receptors in the regulation of the fundamental properties of the W8B2+ CSCs. Inhibition of
BKCa reduces cell proliferation by accumulating cells in the G0 / G1 phase, suppresses cell self-
renewal but does not affect migration properties. Concerning S1P, it decreases proliferation
and self-renewal without stimulate S1P1,2,3 receptors.
This work highlights fundamental potential molecular targets in a context of cardiac cell
therapy
Keywords : cell therapy, human cardiac stem cells, mesenchymal cells, differentiation, ion
channels, calcium signaling, BKCa channel, sphingosine 1-phosphate, optogenetic.

Annexes
268
ANNEXES

Annexes
269
Annexes
270
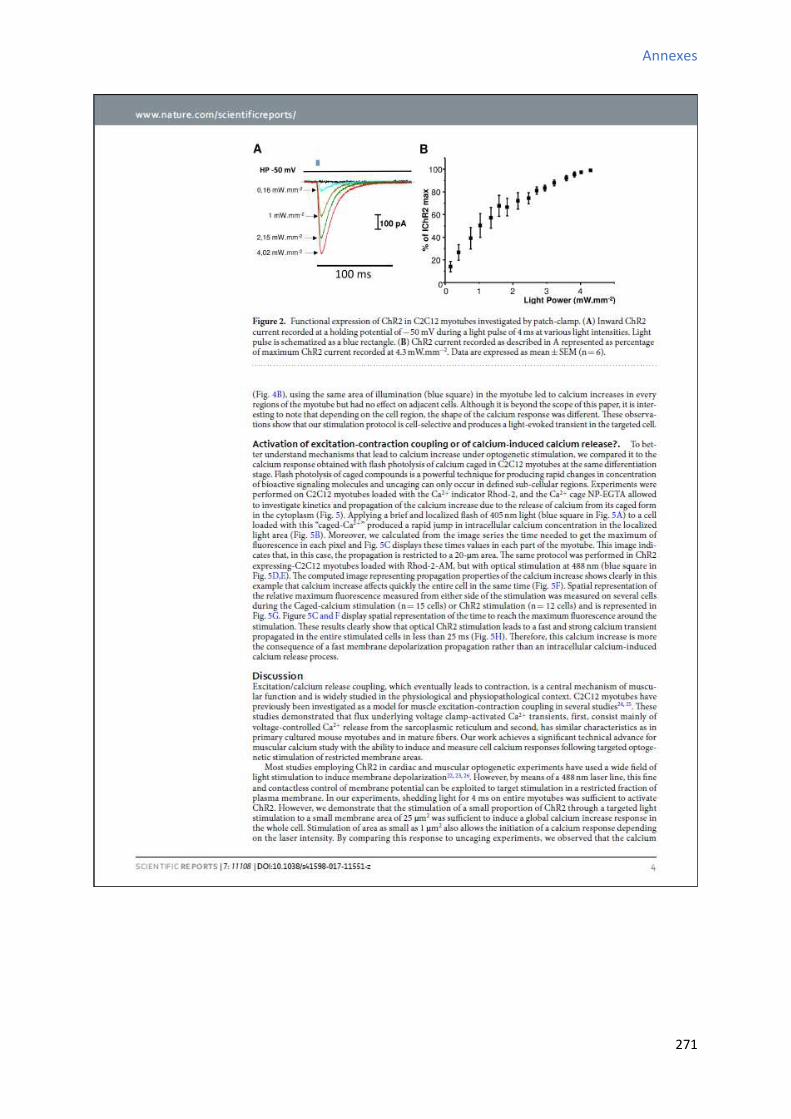
Annexes
271
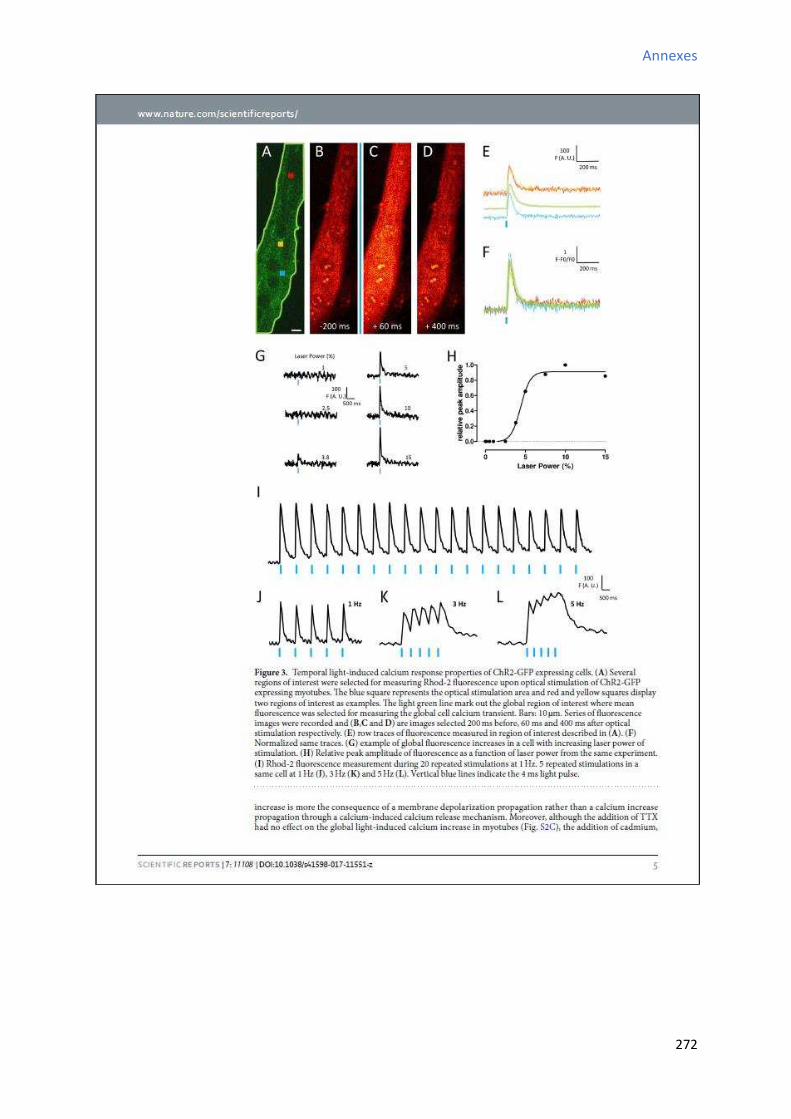
Annexes
272

Annexes
273
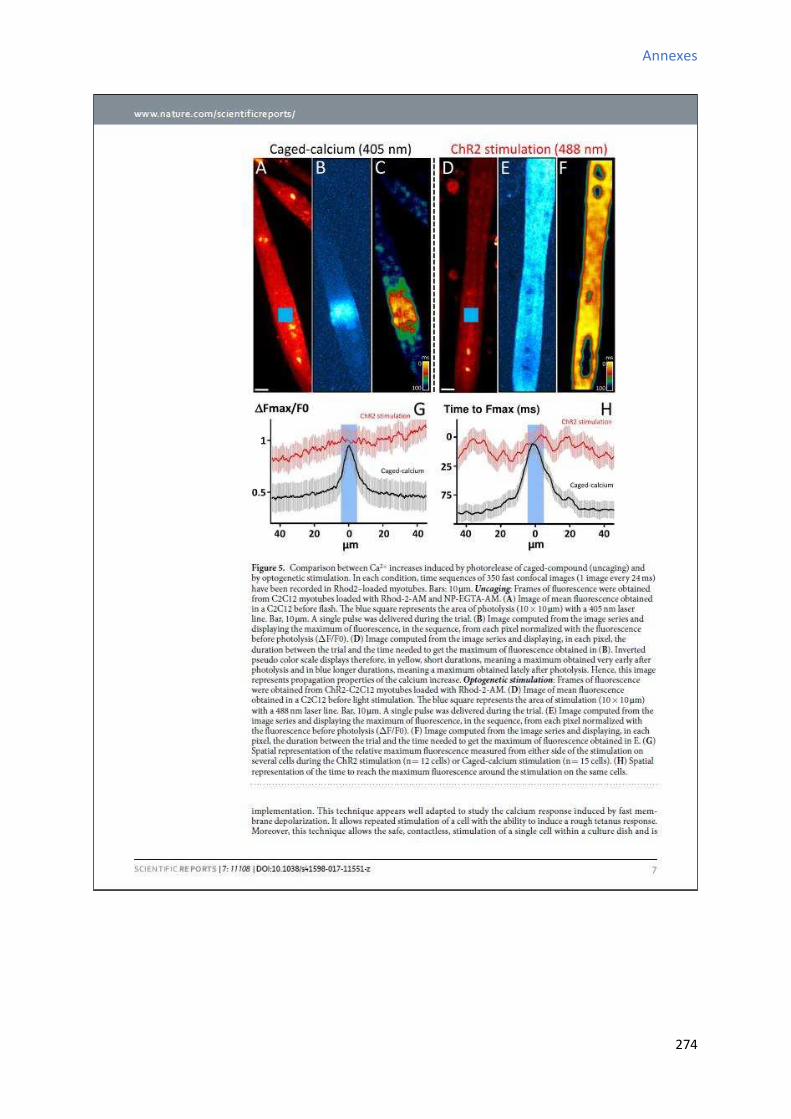
Annexes
274

Annexes
275

Annexes
276

Annexes
277