Embed Size (px)
Citation preview

1
Programme & résumés des JER
Bordeaux, 7 & 8 mars 2017
https://www.r4p-inra.fr/

2
SOMMAIRE
Remerciements ............................................................................................. 3
Organisation .................................................................................................. 4
Programme .................................................................................................... 6-7
Résumés des interventions ........................................................................... 8

3
Remerciements : Nous remercions ici tous nos partenaires financiers :
INRA – Département de santé des plantes
et UMR Santé et Agroécologie du Vignoble
Site Web : http://www.smach.inra.fr/ Site web UMR : UMR Santé et Agroécologie du vignoble
ANSES
(Agence nationale de sécurité sanitaire de l’alimentation, de
l'environnement et du travail)
27-31, avenue du Général Leclerc 94701 Maisons-Alfort Cedex
Site Web : http://www.anses.fr/fr
CIVB
Conseil interprofessionnel du vin de bordeaux
1, Cours du XXX juillet 33075 Bordeaux Cedex
FRANCE
RMT Florad Gestion de la flore adventice en grandes cultures et en vigne
www.florad.org
BNIC
Bureau National Interprofessionnel du Cognac (BNIC)
23 allées Bernard Guionnet BP 90018
16100 Cognac Cedex
ISVV
Institut des Sciences de la Vigne et du Vin
210 chemin de Leysotte, CS 50008 33882 Villenave d'Ornon
Labex COTE
Labex COTE : Evolution, adaptation et gouvernance des écosystèmes
continentaux et côtiers
Co-direction : Antoine Kremer (INRA) et Hélène Budzinski (CNRS, UMR
EPOC)
Les ODG : Médoc, Haut-Médoc, Listrac-Médoc
et Bordeaux-Bordeaux supérieur,
et les Châteaux Montrose, Tronquoy-Lalande
et De Fargues qui ont gracieusement offert les
vins

4
Nous souhaitons aussi remercier les personnes de l’UMR Save (MC Médalin, MC
Dufour, S. Chamont, JM Armand, L. Druelle, G Taris, S. Richard, A. Nivault, P. Sauris, A. Bellée,
G. Comont), et Cécile Lefflet de l’ISVV qui ont rendu possible la tenue de ces Journées par leur
implication dans l’organisation et la logistique et nos invités P. Mauranx, JB Thibord et JP
Guillemin
MF Corio-Costet J Grosman
D. Debieu M. Siegwart C. Délye
S. Fillinger
A. Micoud
B. Barres AS. Walker

5
Les objectifs du réseau
• Prédire les résistances
• Diffuser les méthodologies d'étude et de diagnostic
• Organiser la recherche publique pour optimiser l'efficience des pesticides
• Favoriser les transferts recherche ↔ profession
Marie-France Corio-Costet
INRA Bordeaux Aquitaine Unité : Santé et agroécologie du vignoble
www6.bordeaux-aquitaine.inra.fr/sante-agroecologie-vignoble/
Danièle Debieu
INRA Versailles – Grignon Unité : BIOlogie GEstion des Risques en agriculture Équipe : Antifongiques, Mode d'Action et Résistances
Christophe Délye
INRA Dijon Unité : Agroécologie Équipe : Adaptation des Adventices aux Agroécosystèmes
Sabine Fillinger
INRA Versailles - Grignon Unité : BIOlogie GEstion des Risques en agriculture Équipe : Antifongiques, Mode d'Action et Résistances
Jacques Grosman
DGAL – SDQSP Expert national filière Vigne Animateur du réseau des experts de la protection des végétaux
Benoit Barres
ANSES - Lyon Équipe : RPP (Résistance aux Produits Phytosanitaires
Myriam Siegwart
INRA PACA- Avignon Unité Plantes et Systèmes de cultures Horticoles Équipe : Écologie de la Production Intégrée
Anne Sophie Walker
INRA Versailles - Grignon Unité : BIOlogie GEstion des Risques en agriculture Équipe : Antifongiques, Mode d'Action et Résistances http://www6.versailles-grignon.inra.fr/bioger

6
Mardi 7 mars 2017
• 9h30-10h00 : Accueil des participants • 10h00-10h20 : Présentation du réseau R4P par AS Walker • 10h20-12h10 : Session 1 - Bases de la résistance aux Produits de Protection des
Plante s (PPP) o Généralités sur les résistances aux pesticides, par C Délye o Outil de diagnostic et de détection de la résistance, par S Fillinger o Suivi des résistances et réglementation : qui fait quoi ? par
J Grosman et V. Mironet
• 12h15 : Cocktail-déjeuner
• 14h00-17h40 : Session 2 - État des lieux des résistances aux fong icides en France
o Mode d’action et classification des fongicides agricoles par D Debieu o Situation au vignoble
� Introduction : pathogènes et traitements sur vigne, par MF Corio-Costet � Evolution de la résistance et rôle de la diversité chez l’oïdium, par MF
Corio-Costet � Évolution de la résistance du mildiou, par B Barrès � État des lieux de la résistance de Botrytis, par AS Walker
• Pause (15 min)
o Résistance aux fongicides en maraîchage et arboriculture, B Barrès o Résistance aux fongicides en grandes cultures AS Walker
• 17h40-18h25: Session 2 - État des lieux des résistances aux inse cticides
en France o Mode d’action et classification des insecticides agricoles, par M Siegwart o Résistance aux insecticides des hémiptères, par B Barrès
• 18h25-18h45: Présentation du site web de R4P, par AS Walker
• 18h55 : Fin de la première journée avec remise des questionnaires-Enquête
• 19h00 Départ à pied pour le dîner dégustation et animation musicale
Merci de penser à rendre le bulletin jaune à la fin de cette session.

7
Mercredi 8 mars 2017
• 9h00-9h40 : Session 2 - État des lieux des résistances aux inse cticides en
France (suite)
o Résistance aux insecticides de lépidoptères et diptères dans d’autres cultures, par M Siegwart
o Résistance aux insecticides des coléoptères du colza, par M Siegwart
• 9h40- 10h40: Session 2 - État des lieux des résistances aux herb icides en France
o Mode d’action et classification des herbicides, par C Délye
o Résistances aux herbicides en vigne, par C Délye et J Grosman
o Résistances aux herbicides en grandes cultures et maraîchage, par C Délye
• 10h40 : Pause
• 11h00-12h10 : Session 3 - Prévenir et gérer la résistance aux PPP
o Réduire les risques de sélection des résistances : de la théorie à la pratique, des pistes pour améliorer la durabilité des PPP, par AS Walker, C Délye et M Siegwart
• 12h15: Cocktail-déjeuner
• 13h45-15h20 : Session 3 - Gérer la résistance aux PPP, exemples d e succès de gestion des pesticides
o Fongicides : évolution de la résistance de l’oïdium de la vigne aux QoIs et gestion, par P Mauranx (Acteo-Vivadour)
o Insecticides : détection et gestion de la résistance de la pyrale du maïs à la famille des pyréthrinoïdes, par JB Thibord (Arvalis)
o Herbicides : le désherbage non chimique, par JP Guillemin (INRA Dijon)
• 15h20-16h20 : Questions et débat (sur la base du retour des questionnaires remis aux participants à l’accueil)
• 16h20- 16h30 : Conclusion
o Synthèse des JÉR 2017 : vers le futur et au-delà, par MF Corio-Costet
o Clôture vers 16h30-17h00
Merci de penser à rendre la fiche de satisfaction avant de partir

8
Généralités sur la résistance aux Produits de Protection des Plantes (PPP)
Anne-Sophie WALKER, Christophe DELYE INRA, UMR1290 BIOGER, Avenue L. Brétignières, 78850 Thiverval-Grignon, France. INRA, Agroécologie, 17 rue Sully, 21000 Dijon, France. [email protected], [email protected] Résumé :
La résistance « biologique » correspond à une moindre sensibilité d’un individu à un PPP, mesurée au laboratoire par comparaison avec des individus sensibles de référence. Un individu résistant peut être caractérisé par son facteur de résistance (l’« intensité » ou la « force » de la résistance, correspondant au rapport entre la quantité de PPP requise pour avoir le même effet sur un individu résistant que sur un individu sensible), et son profil (ou spectre) de résistance (la gamme de substances auquel l’individu est résistant). Le caractère de résistance est transmis aux générations suivantes de bio-agresseurs, car déterminé par une ou des modifications du génome. En fonction de son contrôle génétique, on distingue :
- la résistance croisée, qui est une résistance à plusieurs substances due à une seule modification du génome.
- la résistance multiple, qui est due à l’accumulation, au sein d’un même individu, de plusieurs modifications du génome induisant des résistances à plusieurs substances. Le profil de résistance d’un individu dépend de ses mécanismes de résistance. On distingue globalement deux types de résistances, selon la nature du mécanisme en cause :
- la résistance liée à la cible du PPP. Ce type de résistance ne concerne qu’un seul mode d’action, mais n’affecte pas forcément toutes les substances ayant ce mode d’action.
- la résistance non liée à la cible du PPP, qui fait appel à d’autres mécanismes comme la métabolisation (« détoxication »), la séquestration ou l’efflux accru du PPP. Ce type de résistance est complexe car nécessitant souvent plusieurs modifications du génome. Il peut concerner des substances ayant des modes d’action différents. Certains mécanismes de résistance peuvent avoir des effets secondaires (« pléiotropes ») sur le cycle de vie du bio-agresseur. On peut éventuellement observer une pénalité en l’absence de l’application de PPP chez des individus résistants (« coût génétique » ou « altération de la fitness » : individus moins vigoureux, se reproduisant moins...).
À l’échelle de la population de bio-agresseurs, la résistance « en pratique » s’observe au champ. Elle est due à la présence dans les populations d’individus présentant une résistance biologique en fréquence suffisante et avec des facteurs de résistance suffisamment forts pour entraîner une perte d’efficacité des PPP mesurable par l’agriculteur.
La résistance aux PPP est un phénomène évolutif. La pression de sélection exercée par les PPP favorise la reproduction des individus les moins sensibles (les plus résistants), qui préexistent en faible fréquence dans les populations de bio-agresseurs. Sous l’effet des applications, la fréquence des individus résistants augmente dans les populations, jusqu’à causer éventuellement une perte de contrôle. L’enjeu des stratégies anti-résistance est donc de ralentir la sélection de résistances par les PPP. Il est cependant impossible de l’éviter totalement à partir du moment où des populations de bio-agresseurs sont exposées à des PPP.

9
Mots-clefs: résistance biologique, résistance en pratique, sélection, résistance croisée, résistance multiple, facteur de résistance, coût génétique, populations, mécanismes de résistance.

10
Outils de diagnostic et de détection de la résistance Sabine FILLINGER UMR BIOGER, INRA, AgroParisTech, Université Paris-Saclay, Thiverval-Grignon. [email protected] Résumé : Le suivi de la résistance des bio-agresseurs aux produits phytosanitaires est le point clé dans l’évaluation de l’efficacité et des risques associés aux traitements des cultures. Quel que soit le laboratoire concerné, la plupart des analyses sont faites par des tests biologiques qui varient selon le bio-agresseur étudié (champignon, insecte, adventice). Ces tests renseignent généralement les niveaux de résistance aux produits testés et, selon l’organisme, les spectres de résistance (croisées ou non entre modes d’action). Ils sont robustes, ne nécessitent pas de grande technicité et ni de connaissance sur d’éventuels mécanismes de résistance. Leur inconvénient majeur est la durée expérimentale.
D’autres tests, plus techniques mais souvent plus rapides sont possibles dans les laboratoires équipées et formées. Qu’il s’agisse de tests biochimiques ou moléculaires (PCR, LAMP ou HRM), ils nécessitent cependant la connaissance du mode d’action du pesticide voire du mécanisme de résistance impliqué. Il existe également des méthodes moléculaires quantitatives telles que la qPCR ou le séquençage haut débit qui permettent d’évaluer les fréquences d’allèles de résistance au sein de populations de bio-agresseurs.
Le choix du type d’analyse doit prendre en compte la biologie du bio-agresseur, les connaissances existantes sur le produit phytosanitaire et la/les résistance(s) concernées, l’information recherchée (résistance : oui/non, à quelle fréquence ?), l’équipement nécessaire et le coût du test. Dans cet exposé, nous aborderons les principes des tests majeurs, leurs avantages et inconvénients ainsi que les applications potentielles.
Mots-clefs: résistance, analyses biologiques, tests enzymatiques, diagnostic moléculaire.

11

12
Suivi des résistances et réglementation : qui fait quoi ? Jacques GROSMAN1 et Véronique MIRONET2
1DGAL-SDQSPV, Draaf-Sral Auvergne- Rhône-Alpes, 165 rue de Garibaldi, 69401 Lyon CEDEX 03, France ; 2Anses – Direction des produits Règlementés (DEPR), Unité Efficacité, 14 rue Pierre et Marie Curie, 94 700 Maisons-Alfort [email protected], [email protected] Résumé :
Dans le cadre de l'autorisation de mise sur le marché des produits, les sociétés phytosanitaires sont tenues de donner toute information sur le risque d'apparition de phénomènes de résistance liée à l'utilisation des produits. En cas de risque, les sociétés doivent proposer des mesures de gestion adaptées et mettre en place une surveillance en post-autorisation afin de décrire l'évolution de la résistance et signaler une éventuelle baisse de l'efficacité au terrain. La décision d'autorisation de mise sur le marché tient compte du risque lié à la résistance.
Par ailleurs, depuis de nombreuses années, un plan de surveillance des phénomènes de résistance est piloté par le Ministère en charge de l'agriculture. Depuis 2012, il est intégré au suivi des effets non intentionnels des produits phytopharmaceutiques du Plan Ecophyto dont la version 2 préconise un renforcement de la surveillance des résistances. Il a pour objet de recueillir des informations sur d'éventuelles dérives de sensibilité des pathogènes et ravageurs vis-à-vis des principales familles de pesticides. Le choix des couples bio-agresseurs/substances active est décidé en fonction des enjeux liés à l'utilisation des modes d'action dans différentes filières : il s'agit de détecter les premières émergences de résistance et mettre au point des méthodes de diagnostic.
Enfin, les phénomènes de résistances sont pris en compte dans le dispositif récent (loi du 13/10/2014) de phyto-pharmacovigilance
En fonction des résultats et éventuellement d'investigations complémentaires sur les pertes d'efficacité susceptibles d'être engendrées par le développement de la résistance, notamment via des réseaux d'essais d'efficacité, des mesures de gestion pourront être prises, allant des recommandations dans les notes techniques jusqu'à des modifications des conditions d'applications liées aux décisions d'autorisations de mise sur le marché.
Mots-clefs: résistance, produits phytopharmaceutiques, autorisation de mise sur le marché, plan de surveillance, plan ECOPHYTO, notes techniques communes, phyto-pharmacovigilance.

13
14
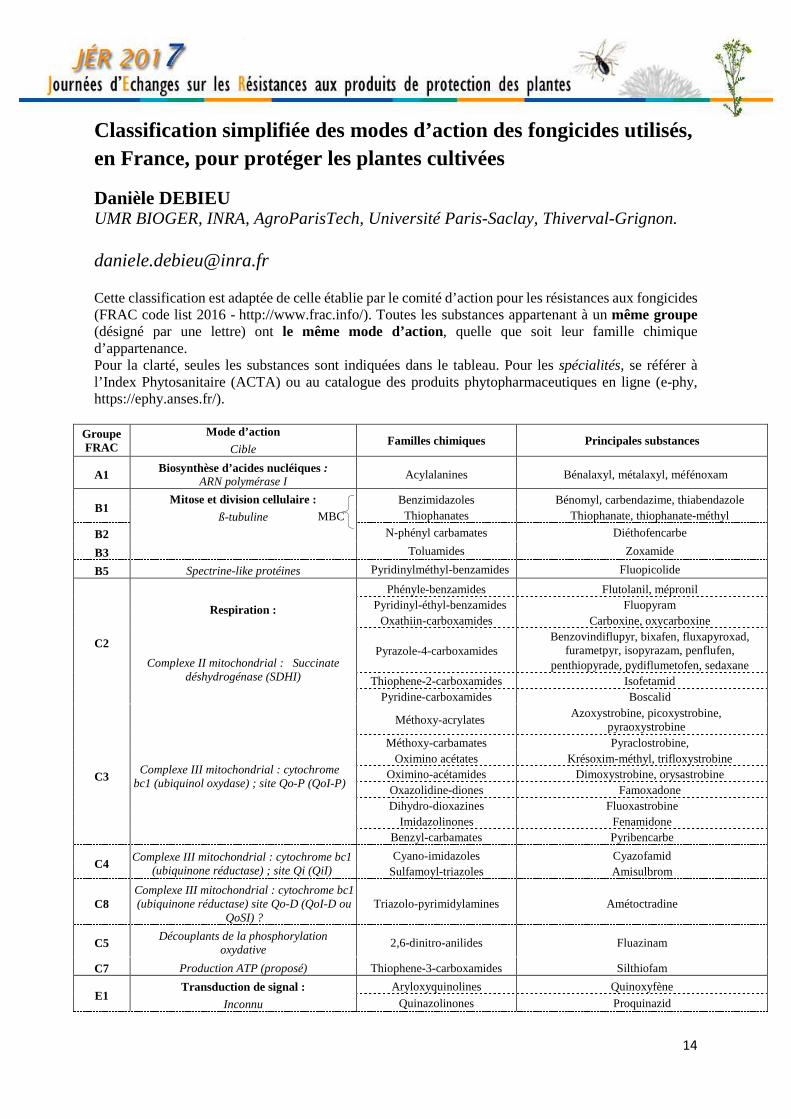
Classification simplifiée des modes d’action des fongicides utilisés, en France, pour protéger les plantes cultivées
Danièle DEBIEU UMR BIOGER, INRA, AgroParisTech, Université Paris-Saclay, Thiverval-Grignon. [email protected] Cette classification est adaptée de celle établie par le comité d’action pour les résistances aux fongicides (FRAC code list 2016 - http://www.frac.info/). Toutes les substances appartenant à un même groupe (désigné par une lettre) ont le même mode d’action, quelle que soit leur famille chimique d’appartenance. Pour la clarté, seules les substances sont indiquées dans le tableau. Pour les spécialités, se référer à l’Index Phytosanitaire (ACTA) ou au catalogue des produits phytopharmaceutiques en ligne (e-phy, https://ephy.anses.fr/).
Groupe FRAC
Mode d’action
Cible Familles chimiques Principales substances
A1 Biosynthèse d’acides nucléiques : ARN polymérase I
Acylalanines Bénalaxyl, métalaxyl, méfénoxam
B1 Mitose et division cellulaire :
ß-tubuline
Benzimidazoles Bénomyl, carbendazime, thiabendazole Thiophanates Thiophanate, thiophanate-méthyl
B2 N-phényl carbamates Diéthofencarbe
B3 Toluamides Zoxamide
B5 Spectrine-like protéines Pyridinylméthyl-benzamides Fluopicolide
C2
Respiration :
Complexe II mitochondrial : Succinate déshydrogénase (SDHI)
Phényle-benzamides Flutolanil, mépronil Pyridinyl-éthyl-benzamides Fluopyram
Oxathiin-carboxamides Carboxine, oxycarboxine
Pyrazole-4-carboxamides Benzovindiflupyr, bixafen, fluxapyroxad,
furametpyr, isopyrazam, penflufen, penthiopyrade, pydiflumetofen, sedaxane
Thiophene-2-carboxamides Isofetamid Pyridine-carboxamides Boscalid
C3 Complexe III mitochondrial : cytochrome
bc1 (ubiquinol oxydase) ; site Qo-P (QoI-P)
Méthoxy-acrylates Azoxystrobine, picoxystrobine,
pyraoxystrobine Méthoxy-carbamates Pyraclostrobine,
Oximino acétates Krésoxim-méthyl, trifloxystrobine Oximino-acétamides Dimoxystrobine, orysastrobine Oxazolidine-diones Famoxadone Dihydro-dioxazines Fluoxastrobine
Imidazolinones Fenamidone Benzyl-carbamates Pyribencarbe
C4 Complexe III mitochondrial : cytochrome bc1
(ubiquinone réductase) ; site Qi (QiI) Cyano-imidazoles Cyazofamid
Sulfamoyl-triazoles Amisulbrom
C8 Complexe III mitochondrial : cytochrome bc1 (ubiquinone réductase) site Qo-D (QoI-D ou
QoSI) ? Triazolo-pyrimidylamines Amétoctradine
C5 Découplants de la phosphorylation
oxydative 2,6-dinitro-anilides Fluazinam
C7 Production ATP (proposé) Thiophene-3-carboxamides Silthiofam
E1 Transduction de signal :
Inconnu
Aryloxyquinolines Quinoxyfène
Quinazolinones Proquinazid
MBC
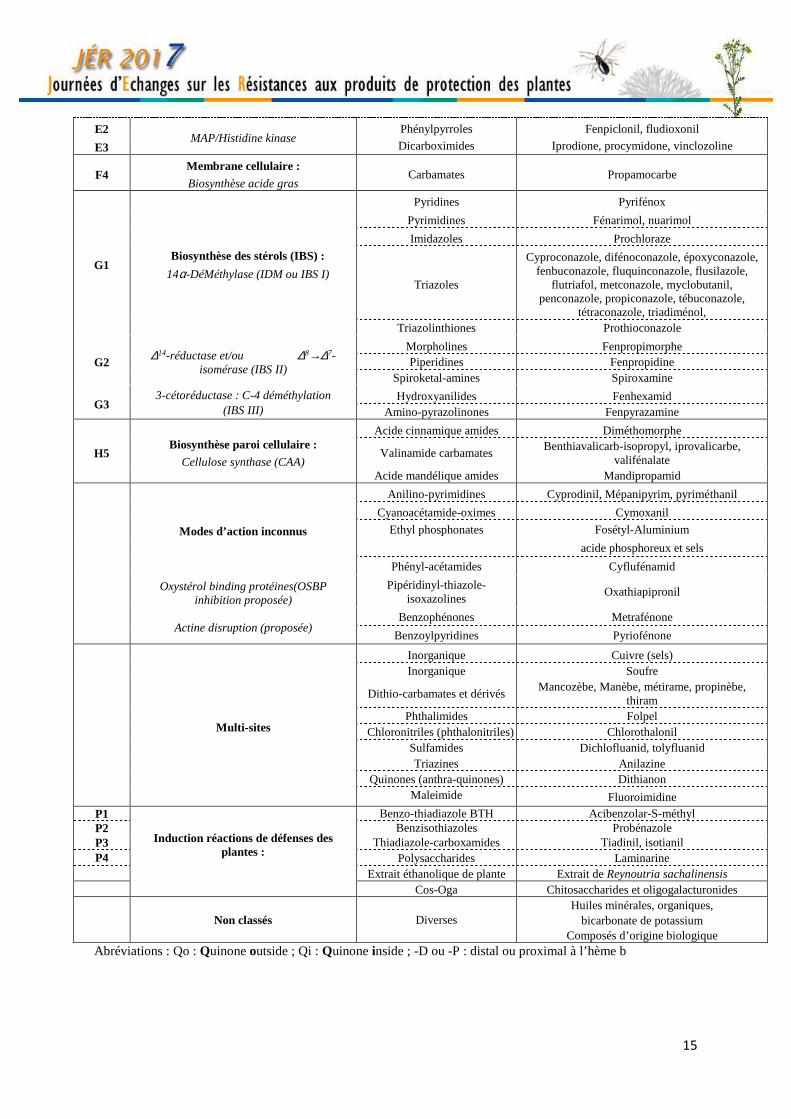
15
E2 MAP/Histidine kinase
Phénylpyrroles Fenpiclonil, fludioxonil
E3 Dicarboximides Iprodione, procymidone, vinclozoline
F4 Membrane cellulaire :
Biosynthèse acide gras Carbamates Propamocarbe
G1 Biosynthèse des stérols (IBS) :
14α-DéMéthylase (IDM ou IBS I)
Pyridines Pyrifénox
Pyrimidines Fénarimol, nuarimol
Imidazoles Prochloraze
Triazoles
Cyproconazole, difénoconazole, époxyconazole, fenbuconazole, fluquinconazole, flusilazole,
flutriafol, metconazole, myclobutanil, penconazole, propiconazole, tébuconazole,
tétraconazole, triadiménol, Triazolinthiones Prothioconazole
G2 ∆14-réductase et/ou ∆8→∆7-isomérase (IBS II)
Morpholines Fenpropimorphe Piperidines Fenpropidine
Spiroketal-amines Spiroxamine
G3 3-cétoréductase : C-4 déméthylation
(IBS III) Hydroxyanilides Fenhexamid
Amino-pyrazolinones Fenpyrazamine
H5 Biosynthèse paroi cellulaire :
Cellulose synthase (CAA)
Acide cinnamique amides Diméthomorphe
Valinamide carbamates Benthiavalicarb-isopropyl, iprovalicarbe,
valifénalate Acide mandélique amides Mandipropamid
Anilino-pyrimidines Cyprodinil, Mépanipyrim, pyriméthanil
Cyanoacétamide-oximes Cymoxanil
Modes d’action inconnus Ethyl phosphonates Fosétyl-Aluminium
acide phosphoreux et sels
Phényl-acétamides Cyflufénamid
Oxystérol binding protéines(OSBP
inhibition proposée) Pipéridinyl-thiazole-
isoxazolines Oxathiapipronil
Actine disruption (proposée)
Benzophénones Metrafénone
Benzoylpyridines Pyriofénone
Multi-sites
Inorganique Cuivre (sels) Inorganique Soufre
Dithio-carbamates et dérivés Mancozèbe, Manèbe, métirame, propinèbe,
thiram Phthalimides Folpel
Chloronitriles (phthalonitriles) Chlorothalonil Sulfamides Dichlofluanid, tolyfluanid Triazines Anilazine
Quinones (anthra-quinones) Dithianon Maleimide Fluoroimidine
P1
Induction réactions de défenses des plantes :
Benzo-thiadiazole BTH Acibenzolar-S-méthyl P2 Benzisothiazoles Probénazole P3 Thiadiazole-carboxamides Tiadinil, isotianil P4 Polysaccharides Laminarine Extrait éthanolique de plante Extrait de Reynoutria sachalinensis Cos-Oga Chitosaccharides et oligogalacturonides
Non classés Diverses Huiles minérales, organiques,
bicarbonate de potassium Composés d’origine biologique
Abréviations : Qo : Quinone outside ; Qi : Quinone inside ; -D ou -P : distal ou proximal à l’hème b

16
Résistance et diversité chez l’oïdium de la vigne
Marie-France CORIO-COSTET INRA, UMR1065 SAVE, ISVV, CS 20032, 33882 Villenave d’Ornon, France. [email protected], Résumé :
La vigne, de par son environnement et en tant que culture pérenne est soumise à la pression de nombreux bio-agresseurs qui nécessite l’utilisation de produits de protection des plantes, dont les fongicides, lesquels représentent environ 80% des produits utilisés en viticulture.
Un panorama de l’état des résistances aux fongicides de l’agent de l’oïdium (Erysiphe necator), du mildiou (Plasmopara viticola) France sera présenté avec quelques cas particuliers démontrant l’importance des connaissances sur les populations de bioagresseurs auxquels l’on s’adresse et les modes d’action des fongicides.
Un focus sera fait sur la résistance aux fongicides inhibiteurs de la biosynthèse des stérols présente en France depuis 1987 et de l’importance de la diversité (groupes génétiques A et B) dans l’expression de la résistance. L’apparition en 2008 et l’évolution de la résistance aux fongicides inhibiteurs de la respiration mitochondriales de type QoI sera abordée, ainsi que l’apparition de nouvelles résistances aux inhibiteurs de la succinate deshydrogénase (SDHI), au quinoxyfène et à la métrafénone. À noter que des différences de sensibilité aux défenses émises par des variétés résistantes de Vitis ont également été mises en évidence, en absence de pression de sélection.
En conclusion, pour gérer la résistance et la diminution des modes d’action disponibles, quelques propositions seront faites pour mieux connaitre ces phénomènes de résistance, en limiter l’apparition et aller vers la résilience de ces phénomènes.
Mots-clefs: résistance, fongicides, environnement, diversité, Oïdium, IDM, QoI, SDHIs, quinoxyfène, métrafénone.

17

18
Évolution des résistances du mildiou de la vigne Benoit BARRÈS Anses, Laboratoire de Lyon - Unité RPP, 31 avenue Tony Garnier, 69364 Lyon Cedex 07 [email protected] Le mildiou de la vigne est une des maladies majeures en viticulture. Elle est causée par un oomycète, Plasmopara viticola et nécessite de nombreux traitements phytosanitaires. Le caractère pérenne de la culture hôte et les traitements répétés font de ce pathogène un parfait candidat à l’évolution de résistances aux produits phytosanitaires. La plupart des familles de produits autorisées en vigne sont aujourd’hui concernées par les phénomènes de résistances, à l’exception notable des produits multi-sites. Ces phénomènes de résistances sont particulièrement importants pour les phénylamides, les cyanoacétamide-oximes, les CAA et les QoI-P. De nouvelles résistances sont également en train d’évoluer pour les familles QoI-D (ou QoSI), QiI et les benzamides. Ces nouvelles résistances sont préoccupantes et concernent notamment des familles de produits qui inhibent le complexe III de la chaine respiratoire. Un bref récapitulatif de la situation actuelle des résistances aux produits phytosanitaires utilisés pour lutter contre le mildiou de la vigne sera exposé. Nous présenterons des données historiques sur l’évolution des résistances et les résultats obtenus plus récemment. Un état des lieux plus précis sera fait sur nos connaissances actuelles des inhibiteurs du complexe III de la chaine respiratoire (QoIs et QiI). Deux types de résistances ont évolué contre ces inhibiteurs : des résistances spécifiques causées par des mutations de la séquence codant pour la protéine cible et une résistance non spécifique via une respiration alternative (aussi appelée AOX).
Mots-clés : Plasmopara viticola, vigne, mildiou, QoI-D, QoI-P, QiI, AOX.

19

20
Etat des lieux de la résistance de Botrytis Anne-Sophie WALKER INRA, UR1290 BIOGER, Avenue L. Brétignières, 78850 Thiverval-Grignon, France [email protected]
La pourriture grise est causée par un complexe de deux espèces cryptiques, Botrytis cinerea et Botrytis pseudocinerea. Cette dernière espèce est minoritaire dans les populations mais est naturellement résistante au fenhexamid. Chez B. cinerea, la résistance acquise est détectée dans les populations françaises pour 5 modes d’action anti-Botrytis unisites (par ordre chronologique de détection) : les benzimidazoles et thiophanates, les dicarboximides, les anilinopyrimidines, les hydroxyanilides et les SDHI. La résistance aux QoI-P a été sélectionnée non intentionnellement chez cette espèce. Pour tous ces modes d’action, la résistance est déterminée par une modification de la cible de ces anti-Botrytis. Ces résistances sont souvent associées (résistance multiple). La fréquence de ces différentes résistances reste actuellement faible à modérée dans les différents vignobles. En effet, la diversité des modes d’action disponibles pour lutter contre cette maladie permet une alternance intra-, voire inter-annuelle des molécules, ce qui contribue, conjointement à l’expression du cout de la résistance caractérisée pour plusieurs modes d’action, à limiter la sélection de la résistance et à éviter globalement les échecs en pratique. La situation est plus difficile pour les cultures pour lesquelles la pression épidémique nécessite un plus grand nombre de traitements (cultures maraîchères et fraisier par exemple). Enfin, la résistance multidrogues (MDR), caractérisée par un efflux accru de fongicides, est relativement fréquente au vignoble et induit une résistance croisée de faible intensité à l’ensemble des anti-Botrytis.
Mots-clés : Botrytis cinerera, vignoble, anti-Botrytis, mutation de cible, résistance multiple, résistance multidrogues (MDR), fenhexamid, inhibiteurs de la succinate deshydrogénase (SDHI).

21

22
Résistances aux fongicides en arboriculture et en maraîchage Benoit BARRÈS Anses, Laboratoire de Lyon - Unité RPP, 31 avenue Tony Garnier, 69364 Lyon Cedex 07 [email protected] Produire un état des lieux des problèmes de résistance en arboriculture et en cultures maraichères est particulièrement difficile, compte tenu de la diversité des cultures et des pathogènes que recouvrent ces deux domaines. Même en s’en tenant aux parasites les plus importants, de nombreux cas de résistance peuvent être cités dont l’impact est plus ou moins conséquent au terrain, en fonction des mécanismes de résistance mis en jeu ainsi que de la fréquence des individus résistants dans les populations en présence. Plusieurs familles de fongicides aux modes d’action très différents, sont concernées par ces phénomènes. Parmi les plus fréquemment mentionnées, se trouvent : les benzimidazoles (thiophanate-méthyl), les dicarboximides (iprodione), les inhibiteurs de la biosynthèse des stérols (IBS), les strobilurines (azoxystrobine, krésoxim-méthyl, …). Après avoir présenté un point sur les cas identifiés en France de résistances aux produits phytosanitaires pour les cultures maraichères et l’arboriculture, nous présenterons plus en détail trois exemples de pathogènes qui illustrent l’évolution des résistances aux pesticides dans ces filières. En premier lieu, nous nous intéresserons à l’agent de la pourriture grise, Botrytis cinerea. Très ubiquiste et présent en maraîchage comme en arboriculture, B. cinerea a développé des mécanismes de résistance vis-à-vis de plusieurs familles de fongicides. Nous verrons ensuite l’exemple d’une des maladies majeures en arboriculture, la tavelure du pommier causée par Venturia inaequalis. Enfin, nous exposerons le cas particulier des monilioses dont les espèces indigènes françaises (Monilinia laxa, Monilinia fructigena) ne montrent pas, à ce jour, de résistances aux principaux fongicides utilisés pour lutter contre elles, alors que l’espèce Monilinia fructicola, introduite en France au début des années 2000, s’est avérée dès sa détection résistante aux benzimidazoles. Mots-clefs: résistance, fongicides, arboriculture, maraîchage, benzimidazoles, dicarboximides, IBS, phénylamides, strobilurines, Venturia inaequalis, Botrytis cinerea, Monilia spp.

23

24
Résistances aux fongicides en grandes cultures
Anne-Sophie WALKER INRA, UR1290 BIOGER, Avenue L. Brétignières, 78850 Thiverval-Grignon, France [email protected] Résumé : Les grandes cultures, et en particulier les céréales, représentent la part majeure de l’assolement français et font, à ce titre, l’objet de 1 à 3 traitements fongicides par an. Un certain nombre de modes d’action sont concernés par des résistances spécifiques, et en particulier :
- Les inhibiteurs de la 14α-déméthylation des stérols (IDM), chez les agents de la septoriose du blé et du piétin-verse. Cette résistance complexe est majoritairement déterminée par des mutations affectant la cible de ces fongicides, dont les combinaisons multiples induisent une grande diversité de phénotypes, ce qui complique leur gestion
- Les inhibiteurs de la respiration comme les QoIs (résistances très implantées chez la septoriose, les helminthosporioses, la ramulariose, Microdochium, les oïdiums) et les SDHIs (résistance en émergence chez la septoriose et installée chez l’helminthosporiose de l’orge et le sclérotinia du colza).
De plus, des résistances non liées à la modification de la cible sont détectées chez les agents de la septoriose et du piétin verse. La résistance multidrogues (ou MDR) résulte d’un efflux accru de fongicides par la cellule du champignon) et induit des spectres de résistance croisée entre modes d’action indépendants, avec des niveaux de résistance généralement faibles. Cependant, ce mécanisme, associé à des résistances spécifiques, peut contribuer à constituer des phénotypes très résistants aux IDM, chez l’agent de la septoriose notamment, et à augmenter le risque de résistance en pratique. Mots-clefs: céréales, colza, inhibiteurs de la déméthylation des stérols (IDM), inhibiteurs de la succinate deshydrogénase (SDHI), résistance multidrogues (MDR), septoriose, piétin verse, oïdium, sclerotiniose.

25
26
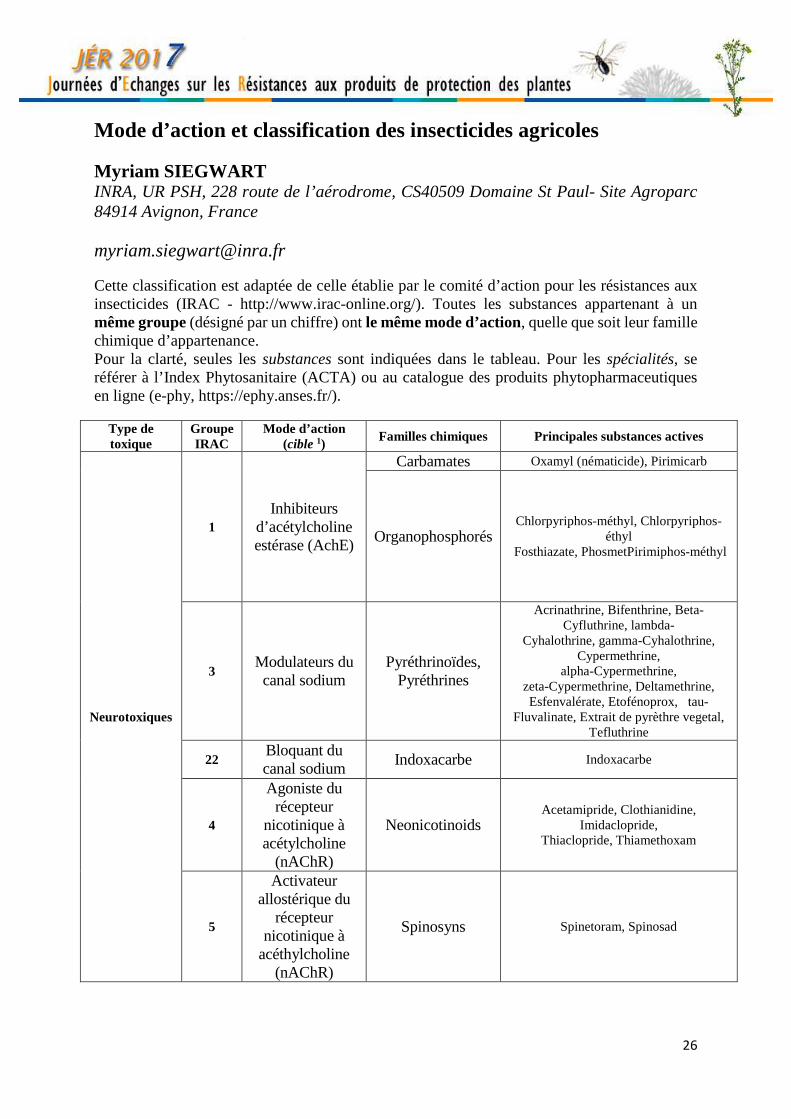
Mode d’action et classification des insecticides agricoles Myriam SIEGWART INRA, UR PSH, 228 route de l’aérodrome, CS40509 Domaine St Paul- Site Agroparc 84914 Avignon, France [email protected] Cette classification est adaptée de celle établie par le comité d’action pour les résistances aux insecticides (IRAC - http://www.irac-online.org/). Toutes les substances appartenant à un même groupe (désigné par un chiffre) ont le même mode d’action, quelle que soit leur famille chimique d’appartenance. Pour la clarté, seules les substances sont indiquées dans le tableau. Pour les spécialités, se référer à l’Index Phytosanitaire (ACTA) ou au catalogue des produits phytopharmaceutiques en ligne (e-phy, https://ephy.anses.fr/).
Type de toxique
Groupe IRAC
Mode d’action (cible 1)
Familles chimiques Principales substances actives
Neurotoxiques
1 Inhibiteurs
d’acétylcholine estérase (AchE)
Carbamates Oxamyl (nématicide), Pirimicarb
Organophosphorés Chlorpyriphos-méthyl, Chlorpyriphos-
éthyl Fosthiazate, PhosmetPirimiphos-méthyl
3 Modulateurs du canal sodium
Pyréthrinoïdes, Pyréthrines
Acrinathrine, Bifenthrine, Beta-Cyfluthrine, lambda-
Cyhalothrine, gamma-Cyhalothrine, Cypermethrine,
alpha-Cypermethrine, zeta-Cypermethrine, Deltamethrine, Esfenvalérate, Etofénoprox, tau-
Fluvalinate, Extrait de pyrèthre vegetal, Tefluthrine
22 Bloquant du canal sodium
Indoxacarbe Indoxacarbe
4
Agoniste du récepteur
nicotinique à acétylcholine
(nAChR)
Neonicotinoids Acetamipride, Clothianidine,
Imidaclopride, Thiaclopride, Thiamethoxam
5
Activateur allostérique du
récepteur nicotinique à
acéthylcholine (nAChR)
Spinosyns Spinetoram, Spinosad
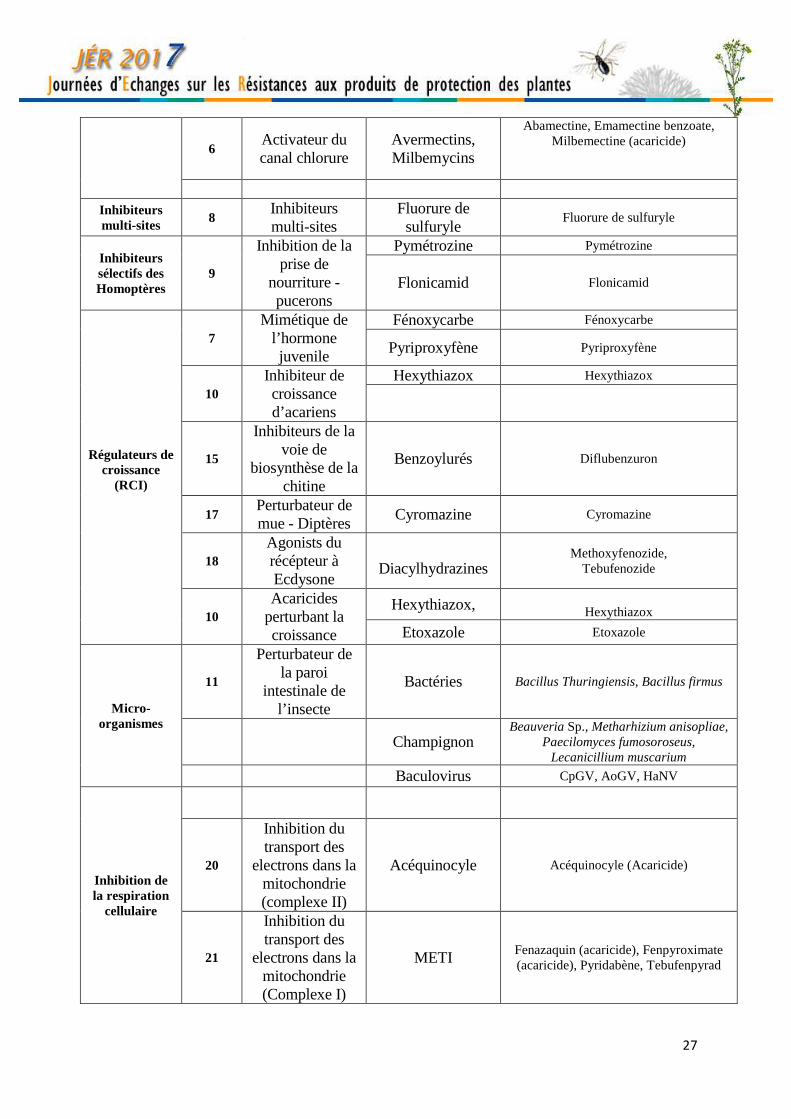
27
6 Activateur du canal chlorure
Avermectins, Milbemycins
Abamectine, Emamectine benzoate, Milbemectine (acaricide)
Inhibiteurs multi-sites
8 Inhibiteurs multi-sites
Fluorure de sulfuryle
Fluorure de sulfuryle
Inhibiteurs sélectifs des Homoptères
9
Inhibition de la prise de
nourriture -pucerons
Pymétrozine Pymétrozine
Flonicamid Flonicamid
Régulateurs de croissance
(RCI)
7 Mimétique de
l’hormone juvenile
Fénoxycarbe Fénoxycarbe
Pyriproxyfène Pyriproxyfène
10 Inhibiteur de croissance d’acariens
Hexythiazox Hexythiazox
15
Inhibiteurs de la voie de
biosynthèse de la chitine
Benzoylurés Diflubenzuron
17 Perturbateur de mue - Diptères
Cyromazine Cyromazine
18 Agonists du récépteur à Ecdysone
Diacylhydrazines
Methoxyfenozide, Tebufenozide
10 Acaricides
perturbant la croissance
Hexythiazox, Hexythiazox
Etoxazole Etoxazole
Micro-organismes
11
Perturbateur de la paroi
intestinale de l’insecte
Bactéries Bacillus Thuringiensis, Bacillus firmus
Champignon Beauveria Sp., Metharhizium anisopliae,
Paecilomyces fumosoroseus, Lecanicillium muscarium
Baculovirus CpGV, AoGV, HaNV
Inhibition de la respiration
cellulaire
20
Inhibition du transport des
electrons dans la mitochondrie (complexe II)
Acéquinocyle Acéquinocyle (Acaricide)
21
Inhibition du transport des
electrons dans la mitochondrie (Complexe I)
METI Fenazaquin (acaricide), Fenpyroximate (acaricide), Pyridabène, Tebufenpyrad
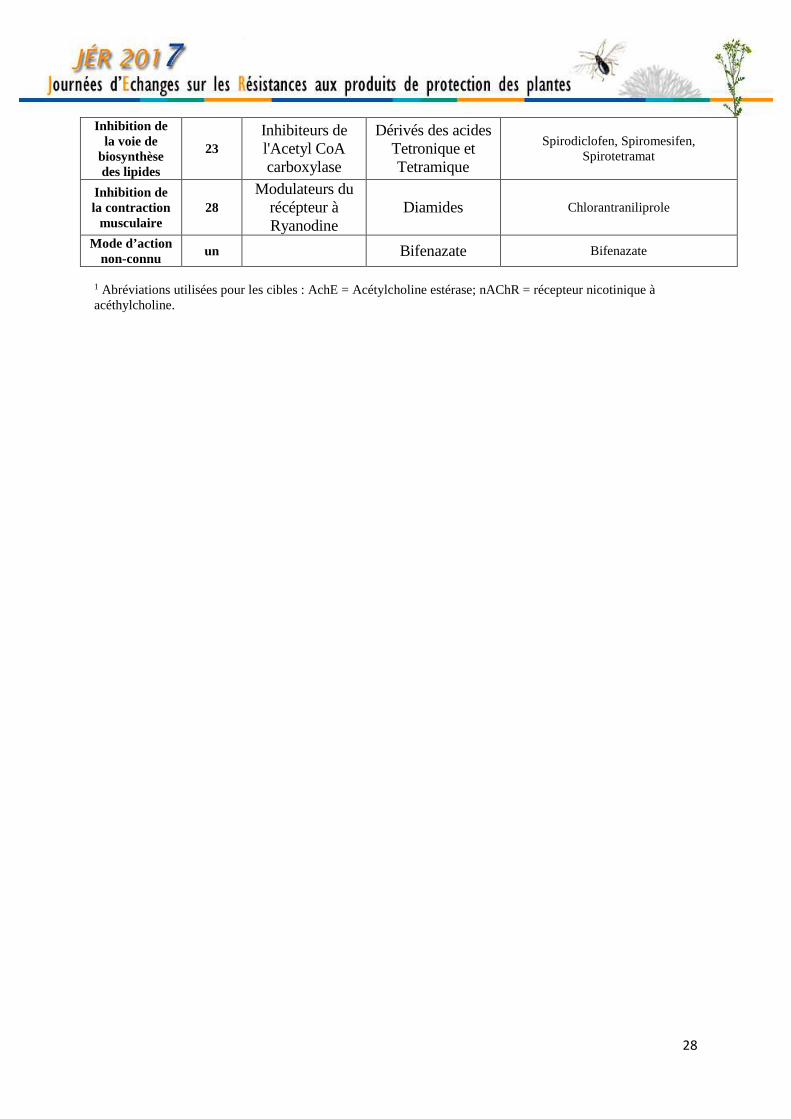
28
Inhibition de la voie de
biosynthèse des lipides
23 Inhibiteurs de l'Acetyl CoA carboxylase
Dérivés des acides Tetronique et Tetramique
Spirodiclofen, Spiromesifen, Spirotetramat
Inhibition de la contraction
musculaire 28
Modulateurs du récépteur à Ryanodine
Diamides Chlorantraniliprole
Mode d’action non-connu
un Bifenazate Bifenazate
1 Abréviations utilisées pour les cibles : AchE = Acétylcholine estérase; nAChR = récepteur nicotinique à acéthylcholine.

29

30
Résistances aux insecticides des Hémiptères Benoit BARRÈS Anses, Laboratoire de Lyon - Unité RPP, 31 avenue Tony Garnier, 69364 Lyon Cedex 07 [email protected]
Les insectes de l’ordre des hémiptères sont des insectes suceurs qui possèdent un rostre et sont caractérisés par de longues antennes et une paire d’aile transformée en hémiélytre (en partie cornée). Au sein de cet ordre, il existe de nombreuses espèces de ravageurs économiquement importants. Les pucerons et les aleurodes appartiennent à cet ordre ainsi que les punaises ou les cicadelles. Ces ravageurs causent parfois des dégâts directs mais sont aussi souvent des vecteurs de maladies virales (par exemple, Scaphoideus titanus, vecteur de la flavescence doréee). C’est pourquoi ils font l’objet de nombreux traitements et qu’en conséquence des résistances aux pesticides ont évolués et évoluent au sein de leurs populations. Les mécanismes de résistance développés par ces insectes sont de deux types. Le premier type de résistance est une résistance dite de « cible » : une mutation du site ciblé par le produit phytosanitaire permet l’acquisition de hauts niveaux de résistance contre celui-ci. Le second type de résistance est qualifié de « métabolique ». Il repose généralement sur une meilleure dégradation du pesticide dans le corps de l’insecte. Ce type de résistance confère souvent un niveau moins élevé de résistance qu’une résistance « cible ». Mais cette résistance peut être efficace contre différentes familles chimiques (on parle alors de résistance croisée). Les principales familles d’insecticides utilisées et concernées par l’évolution de résistances sont des neurotoxiques. Ce sont les carbamates (pyrimicarbe), les organo-phosphorés (chlorpyriphos, phosmet), les pyréthrinoïdes (deltaméthrine, lambda-cyhalothrine, …) et plus récemment, les néonicotinoïdes (imidaclopride, thiaclopride, …).
Après avoir présenté un point sur les cas identifiés en France de résistances aux produits phytosanitaires chez les hémiptères, nous présenterons plus en détails l’exemple de Myzus persicae, le puceron vert du pêcher. Ce puceron très polyphage a développé au cours des années un arsenal de résistances à des groupes d’insecticides de modes d’action différents. Cette espèce a évolué non seulement des résistances de cible à divers produits phytosanitaires, mais également une résistance métabolique. A ce jour cependant, toutes les résistances ne sont pas présentes dans toutes les cultures hôtes. Mots-clefs: résistance, insecticides, hémiptères, carbamates, organo-phosphorés, pyréthrinoïdes, néonicotinoïdes, Myzus persicae.

31

32
Résistance aux insecticides de lépidoptères et diptères dans d’autres cultures Myriam SIEGWART INRA, UR PSH, 228 route de l’aérodrome, CS40509 Domaine St Paul- Site Agroparc 84914 Avignon, France. [email protected] Résumé : Comme pour les Coléoptères et les Hémiptères, les Lépidoptères et les Diptères ont développé principalement des résistances aux insecticides neurotoxiques. Cependant, chez certains Lépidoptères on trouve également des cas de résistance à des familles insecticides pourtant réputées difficilement contournables comme des régulateurs de croissance d’insectes ou encore des bio-insecticides. Afin, d’illustrer la diversité des cas de résistance au sein de ces deux ordres, nous donnerons ici des exemples variés allant d’un ravageur qui a acquis des résistances successives au cours du temps et des mises sur le marché de nouvelles familles chimiques: le carpocapse des pommes et des poires (Cydia pomonella) à un ravageur pour lequel le premier cas de résistance aux pyréthrinoïdes a été détecté en 2012 : la pyrale du maïs (Ostrinia nubilalis).
Le carpocapse des pommes a commencé par développer des résistances aux organo-phosporés, puis aux pyréthrinoïdes, suivi des régulateurs de croissance d’insectes, pour finir avec des biopesticides comme le spinosad et le virus de la granulose (un bio-pesticide micro-organique). Certains individus ne sont résistants qu’à un seul mode d’action mais d’autres peuvent développer des résistances à plusieurs de ces groupes d’insecticides, on parle alors de résistance croisée. D’où la nécessité de suivre l’évolution de ce ravageur afin d’adapter les moyens de lutte en fonction de l’émergence et de la progression des nouveaux mécanismes de résistance. La pyrale du maïs quant à elle commence tout juste à montrer des cas de résistances aux pyréthrinoïdes et plus précisément à la lambda-cyalothrine dans des régions du territoire français bien définies. Cet exemple nous permettra d’illustrer l’importance de la répartition spatiale et de la dynamique de propagation de populations résistantes. Dans la famille des Diptères, ce sont surtout les vecteurs de maladies humaines qui posent un réel problème de santé publique car ils sont très résistants à de nombreuses familles chimiques. En agriculture, les exemples de moindre impact que nous avons choisi de traiter ici portent sur la résistance de la mouche de l’olive à la famille des organo-phosphorés. Nous allons aussi illustrer la notion de risque d’apparition de résistance à travers l’exemple de la Drosophile : Drosophila suzukii récemment introduite en France. Enfin, le dernier couple présenté est composé du Thrips et du Spinosad. Cet insecte appartient à la famille des Thysanoptères. Il commence à présenter des dérives de sensibilité à cet insecticide biologique. Une mutation de la cible moléculaire : le récepteur nicotinique à acéthylcholine (nAChR) est suspecté dans ce cas de figure. Mots-clefs: résistance, insecticides, lépidoptères, diptères, organo-phosphorés, pyréthrinoïdes, régulateur de croissance d’insectes, virus de la granulose.

33

34
Résistance aux insecticides de coléoptères du colza Céline Robert1, Laurent Ruck2, Julien Carpezat1, Martine Leflon 1, Myriam Siegwart3 1 Terres Inovia, Avenue Lucien Brétignières, 78850 Thiverval-Grignon, France ; 2 Terres Inovia, Complexe agricole du Mont Bernard - route de Suippes, 51000 Chalons-en-Champagne, France ;
3 UMR , INRA, 228 route de l'aérodrome - CS40509-Domaine St Paul - Site Agropar, 84914 Avignon Cedex 09, France
[email protected] Résumé : L’altise du colza (Psylliodes chrysocephala) dite grosse altise et le charançon du bourgeon terminal du colza (Ceuthorhynchus picitarsis) sont deux ravageurs majeurs du colza d'hiver en France. Depuis 2009/2010, les infestations de charançon du bourgeon terminal dans un premier temps puis de la grosse altise sont devenues plus difficiles à contrôler malgré les traitements répétés. Le phénomène a augmenté d'année en année. Terres Inovia et ses partenaires, par le biais du groupe AFPP "Insect resistance", ont confirmé que les populations de ces deux ravageurs sont résistantes aux pyréthrinoïdes et que plusieurs mécanismes sont impliqués. Pour la grosse altise, la résistance semble due à une mutation (kdr) dans la cible moléculaire des pyréthrinoïdes. Cette dernière semble bien répartie sur l’ensemble du territoire français. Une deuxième mutation (super-kdr) a également été découverte dans le même gène, mais celle-ci n’est présente que dans l’Yonne et quelques cas ont été trouvés dans des départements voisins. Pour le charançon du bourgeon terminal, la résistance semble bien installée dans le Centre et une partie du Nord Est de la France mais semble absente dans le Sud-Ouest. La première mutation (kdr) a été détectée mais pas la seconde (super kdr). De plus, sur quelques populations, d'autres mécanismes de résistance métabolique dites « non liés à la cible » semblent également impliqués, dans ces deux cas de résistance. En effet une augmentation des capacités de détoxification des insecticides a été mesuré chez les insectes résistants comparés aux sensibles dans quelques populations. Plusieurs mécanismes peuvent co-exister au sein d’une même population. Il n’est pas possible à ce jour de déterminer le niveau de résistance de chaque mécanisme. Plus que jamais, la gestion de ces deux coléoptères ravageurs du colza ne peut reposer sur la seule utilisation d’insecticides chimiques. Il faut remettre les bonnes pratiques agronomiques au cœur du système afin de limiter le plus possible le recours aux insecticides. Mots-clefs: Psylliodes chrysocephala, Ceutorhynchus picitarsis, resistance, pyréthrinoides pratiques agronomiques

35
36
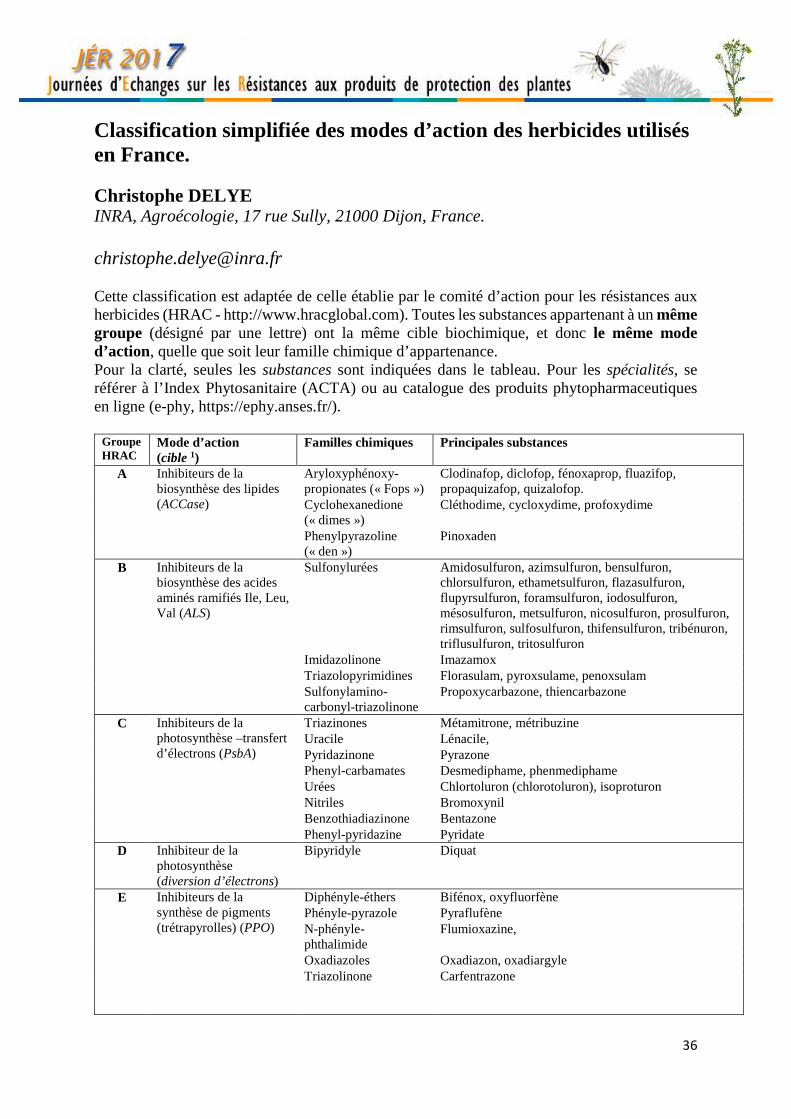
Classification simplifiée des modes d’action des herbicides utilisés en France. Christophe DELYE INRA, Agroécologie, 17 rue Sully, 21000 Dijon, France. [email protected] Cette classification est adaptée de celle établie par le comité d’action pour les résistances aux herbicides (HRAC - http://www.hracglobal.com). Toutes les substances appartenant à un même groupe (désigné par une lettre) ont la même cible biochimique, et donc le même mode d’action, quelle que soit leur famille chimique d’appartenance. Pour la clarté, seules les substances sont indiquées dans le tableau. Pour les spécialités, se référer à l’Index Phytosanitaire (ACTA) ou au catalogue des produits phytopharmaceutiques en ligne (e-phy, https://ephy.anses.fr/).
Groupe HRAC
Mode d’action (cible 1)
Familles chimiques Principales substances
A Inhibiteurs de la biosynthèse des lipides (ACCase)
Aryloxyphénoxy-propionates (« Fops »)
Clodinafop, diclofop, fénoxaprop, fluazifop, propaquizafop, quizalofop.
Cyclohexanedione (« dimes »)
Cléthodime, cycloxydime, profoxydime
Phenylpyrazoline (« den »)
Pinoxaden
B Inhibiteurs de la biosynthèse des acides aminés ramifiés Ile, Leu, Val (ALS)
Sulfonylurées Amidosulfuron, azimsulfuron, bensulfuron, chlorsulfuron, ethametsulfuron, flazasulfuron, flupyrsulfuron, foramsulfuron, iodosulfuron, mésosulfuron, metsulfuron, nicosulfuron, prosulfuron, rimsulfuron, sulfosulfuron, thifensulfuron, tribénuron, triflusulfuron, tritosulfuron
Imidazolinone Imazamox Triazolopyrimidines Florasulam, pyroxsulame, penoxsulam Sulfonylamino-carbonyl-triazolinone
Propoxycarbazone, thiencarbazone
C Inhibiteurs de la photosynthèse –transfert d’électrons (PsbA)
Triazinones Métamitrone, métribuzine Uracile Lénacile, Pyridazinone Pyrazone Phenyl-carbamates Desmediphame, phenmediphame Urées Chlortoluron (chlorotoluron), isoproturon Nitriles Bromoxynil Benzothiadiazinone Bentazone Phenyl-pyridazine Pyridate
D Inhibiteur de la photosynthèse (diversion d’électrons)
Bipyridyle Diquat
E Inhibiteurs de la synthèse de pigments (trétrapyrolles) (PPO)
Diphényle-éthers Bifénox, oxyfluorfène Phényle-pyrazole Pyraflufène N-phényle-phthalimide
Flumioxazine,
Oxadiazoles Oxadiazon, oxadiargyle Triazolinone Carfentrazone
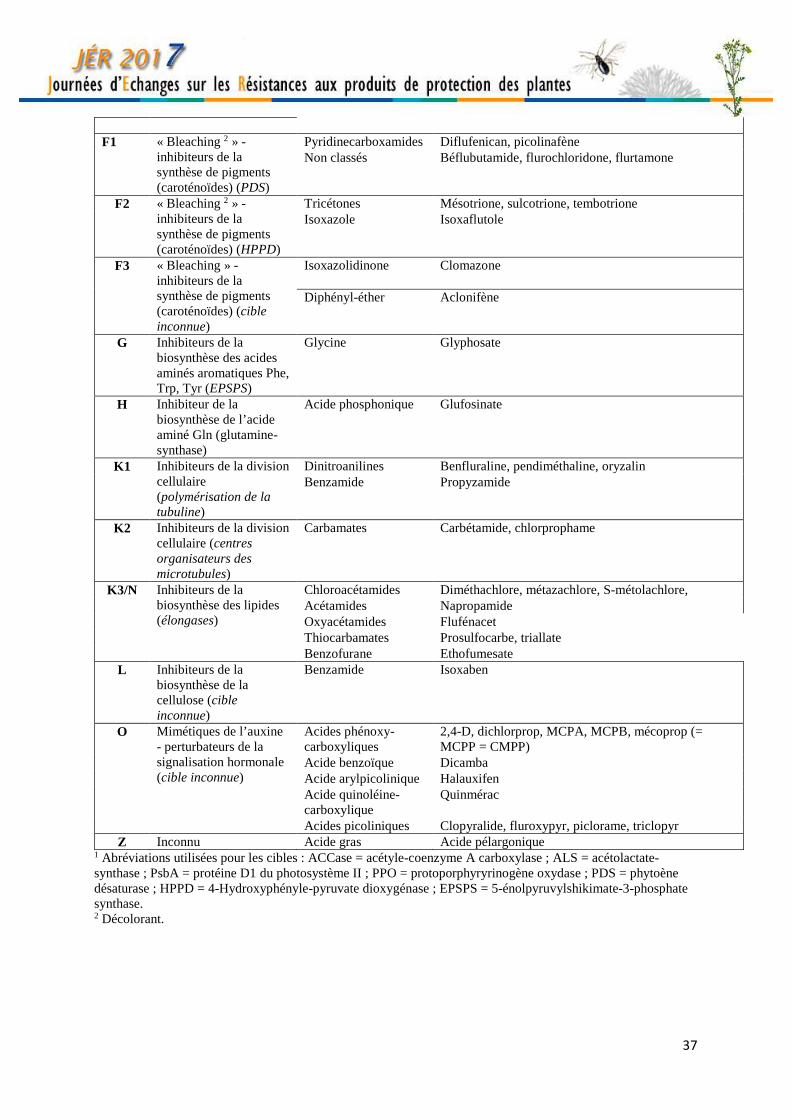
37
F1 « Bleaching 2 » -
inhibiteurs de la synthèse de pigments (caroténoïdes) (PDS)
Pyridinecarboxamides Diflufenican, picolinafène Non classés Béflubutamide, flurochloridone, flurtamone
F2 « Bleaching 2 » - inhibiteurs de la synthèse de pigments (caroténoïdes) (HPPD)
Tricétones Mésotrione, sulcotrione, tembotrione Isoxazole Isoxaflutole
F3 « Bleaching » - inhibiteurs de la synthèse de pigments (caroténoïdes) (cible inconnue)
Isoxazolidinone Clomazone
Diphényl-éther Aclonifène
G Inhibiteurs de la biosynthèse des acides aminés aromatiques Phe, Trp, Tyr (EPSPS)
Glycine Glyphosate
H Inhibiteur de la biosynthèse de l’acide aminé Gln (glutamine-synthase)
Acide phosphonique Glufosinate
K1 Inhibiteurs de la division cellulaire (polymérisation de la tubuline)
Dinitroanilines Benfluraline, pendiméthaline, oryzalin Benzamide Propyzamide
K2 Inhibiteurs de la division cellulaire (centres organisateurs des microtubules)
Carbamates Carbétamide, chlorprophame
K3/N Inhibiteurs de la biosynthèse des lipides (élongases)
Chloroacétamides Diméthachlore, métazachlore, S-métolachlore, Acétamides Napropamide Oxyacétamides Flufénacet Thiocarbamates Prosulfocarbe, triallate Benzofurane Ethofumesate
L Inhibiteurs de la biosynthèse de la cellulose (cible inconnue)
Benzamide Isoxaben
O Mimétiques de l’auxine - perturbateurs de la signalisation hormonale (cible inconnue)
Acides phénoxy-carboxyliques
2,4-D, dichlorprop, MCPA, MCPB, mécoprop (= MCPP = CMPP)
Acide benzoïque Dicamba Acide arylpicolinique Halauxifen Acide quinoléine-carboxylique
Quinmérac
Acides picoliniques Clopyralide, fluroxypyr, piclorame, triclopyr Z Inconnu Acide gras Acide pélargonique
1 Abréviations utilisées pour les cibles : ACCase = acétyle-coenzyme A carboxylase ; ALS = acétolactate-synthase ; PsbA = protéine D1 du photosystème II ; PPO = protoporphyryrinogène oxydase ; PDS = phytoène désaturase ; HPPD = 4-Hydroxyphényle-pyruvate dioxygénase ; EPSPS = 5-énolpyruvylshikimate-3-phosphate synthase. 2 Décolorant.

38
La résistance aux herbicides en viticulture
Jacques GROSMAN1, Christophe DÉLYE2 1Draaf-Sral Rhône-Alpes, 165 rue de Garibaldi, 69401 Lyon CEDEX 03, France 2INRA, Agroécologie, 17 rue Sully, 21000 Dijon, France
[email protected], [email protected] Résumé : Le premier cas de résistance au glyphosate (groupe HRAC G) en France concerne l'ivraie raide (Lolium rigidum). Il a été découvert dans le Gard sur une parcelle de vigne et confirmé par des tests de laboratoire à l'INRA de Dijon en 2006. Depuis, cette résistance a été détectée dans 5 régions viticoles (Pays de Loire, Charente, Aquitaine, Midi-Pyrénées et Languedoc-Roussillon) et déclarée par la firme Mosanto.
Dans le cadre de la gestion de cette résistance, un dispositif de surveillance a été mis en place. Il donne lieu à des prélèvements de graines sur le terrain dans des parcelles présentant des difficultés de désherbage du ray-grass. Les tests de résistance sont réalisés à l’Unité résistance aux produits phytosanitaires de l’ANSES, selon un protocole établi par l’INRA de Dijon. Ce plan de surveillance montre que la résistance est bien implantée dans les zones où de mauvaises pratiques d'utilisation du glyphosate en viticulture ont pu être mises en évidence par le biais d'enquêtes de terrain. Ces pratiques sont notamment la mono-utilisation de glyphosate, le non-respect des doses, des conditions et des périodes d’application.
Ce premier cas de résistance aux herbicides en vigne a été suivi par la détection et la confirmation en 2009 d'un cas de résistance de la Vergerette de Sumatra (Conyza sumatrensis) à la même substance : le glyphosate.
Sur les deux espèces concernées par la résistance au glyphosate (et les espèces voisines des genres Lolium et Conyza), la recherche de solutions a fait l'objet de l'établissement de protocoles d’essais mis en place dans le cadre d'un groupe de travail AFPP-COLUMA-Vigne et a débouché sur des recommandations techniques.
Plus récemment, deux espèces sont venues enrichir la liste des cas de résistance en vigne. Il s’agit cette fois de résistance au groupe B (inhibiteurs de l’ALS). Cette résistance est largement présente chez le Séneçon commun (Senecio vulgaris) depuis au moins 2009, et concerne a minima les vignobles du Bordelais, du Centre-Loire, du Sud-Ouest et du Lyonnais-Beaujolais. Enfin, en 2016, une résistance à des inhibiteurs de l’ALS a été découverte chez la Vergerette du Canada (Conyza canadensis) dans les vignobles du Gard.
Mots-clefs: résistance, vigne, herbicides, désherbage, glyphosate (groupe HRAC G), inhibiteurs de l’ALS (groupe HRAC B), recommandations.

39

40
État des lieux des résistances aux herbicides en France : grandes cultures et maraîchage
Christophe DÉLYE INRA, Agroécologie, 17 rue Sully, 21000 Dijon, France [email protected]
Résumé : En grandes cultures, la résistance aux herbicides a d’abord concerné des inhibiteurs du photosystème 2 (triazines, groupe C, désormais interdites, ce qui a résolu le problème), puis les inhibiteurs de l’ACCase (anti-graminées : « dimes », « fops », « den ») dans les années 1990-2000. Depuis le milieu des années 2000, la résistance touche aussi les « sulfos », un terme trop restrictif qui désigne en fait les 4 familles chimiques d’inhibiteurs de l’ALS (groupe B). Un nouveau groupe vient d’être touché tout récemment : le groupe O (« auxiniques »). Cet exposé commencera par un rappel sur la classification des herbicides en fonction de leur mode d’action (groupes HRAC). Les cas publiés de résistance en France seront décrits :
� Résistances installées : Groupes A et/ou B : Vulpin des champs (Alopecurus myosuroides) et Ivraies (Lolium sp.). Groupe B : Grand Coquelicot (Papaver rhoeas).
� Résistances modérées : Groupes A et/ou B : Folles-Avoines (Avena sp) et Agrostis jouet-du-vent (Apera spica-venti). Groupe B : Bromes (Bromus spp.), Matricaires (Matricaria sp.), Séneçon commun (Senecio vulgaris) et Laiteron épineux (Sonchus asper, en maraîchage).
� Résistances en émergence : Groupes A et/ou B : Digitaire sanguine (Digitaria sanguinalis) Groupe B : Panics (Echinochloa crus-galli et E. phyllopogon), Sétaire verte (Setaria viridis), Tournesol adventice (Helianthus annuus), Stellaire intermédiaire (Stellaria media) et Ambroisie à feuilles d’armoise (Ambrosia artemisiifolia). Groupe O : Grand Coquelicot (Papaver rhoeas). Lorsque l’on considère ces cas de résistance, deux tendances lourdes émergent :
� Une prédominance de la résistance non liée à la cible chez les graminées, très problématique car pouvant concerner des modes d’action différents (par exemple, à la fois groupes A et B). Ce type de résistance a aussi été trouvé chez des dicotylédones (Coquelicot, groupe B), mais semble moins important que chez les graminées.
� La multiplication des cas de dicotylédones (surtout Astéracées) résistant au groupe B.
Les espèces « à risque » (et donc à surveiller) en terme de résistance seront évoqués :
� Crucifères ou Géraniums dans le colza, Chénopodes dans la betterave : risque de résistance au groupe B (lié notamment au déploiement des Variétés

41
Tolérantes aux Herbicides [VTH] et à l’augmentation de l’emploi de ces substances).
� Graminées chez lesquelles la résistance non liée à la cible a évolué (Vulpin des champs et Ivraies) : risque de résistance aux herbicides « racinaires » / de pré-levée (groupes K, K3, N).
Mots-clefs: résistance, herbicides, inhibiteurs de l’ACCase (groupe HRAC A), inhibiteurs de l’ALS (groupe HRAC B), graminées, dicotylédones, grandes cultures, variétés tolérantes aux herbicides (VTH), « auxiniques » (groupe HRAC O), herbicides racinaires (groupes HRAC K1, K3 et N).

42
Prévenir et réduire les risque de sélection des résistances : de la théorie à la pratique, des pistes pour améliorer la durabilité des PPP
Christophe DÉLYE1, Myriam Siegwart2, Anne-Sophie WALKER3 1INRA, Agroécologie, 17 rue Sully, 21000 Dijon, France. 2INRA, UR PSH, Domaine St Paul- Site Agroparc, 84914 Avignon, France. 3INRA, UMR 1290 BIOGER, Avenue L. Brétignières, 78850 Thiverval-Grignon, France. [email protected], [email protected], [email protected] Résumé :
La résistance est le résultat inéluctable de la sélection exercée par les PPP sur les populations de bio-agresseurs. On ne peut pas empêcher une résistance d’évoluer, mais on peut retarder substantiellement son évolution en mettant en place des stratégies appropriées avant (prévention) et après (gestion) la détection des premiers individus résistants. Le point-clef à retenir pour cela est que moins le contrôle d’un bio-agresseur est basé sur une chimie diversifiée, plus le risque de sélectionner rapidement une résistance est élevé. Un exemple type de ce qu’il ne faut pas faire est de contrôler un bio-agresseur présent sur de grandes surfaces en utilisant exclusivement des PPP ayant un même mode d’action, voire un seul PPP. Cette stratégie peut fonctionner à court terme, mais à moyen et long terme, elle est extrêmement risquée du point de vue de la sélection des résistances.
Pour limiter le risque de sélection de résistances, il faut une stratégie de contrôle des bio-agresseurs qui soit diversifiée et efficace. Ceci implique de limiter le plus possible la prolifération des bio-agresseurs en utilisant une diversité de moyens (chimiques et non chimiques) dont la combinaison soit la plus efficace possible : rotations culturales (quand c’est possible), prophylaxie (destruction des sources d’inoculum, du stock semencier ou des formes hivernantes...), et mise en œuvre de toute pratique de contrôle non chimique efficace et acceptable dans un système de production donné. L’emploi des PPP ne devrait idéalement se faire qu’après la mise en œuvre de ces moyens, pour « finir le travail » : l’idée est de réduire le risque de sélection de résistances en réduisant l’infestation sur laquelle on applique un PPP. Et bien entendu, il faut aussi diversifier à la fois dans l’espace et dans le temps la partie chimique du contrôle : alterner les modes d’action, et/ou utiliser des mélanges contenant des substances ayant des modes d’action différents, tout en se plaçant dans des conditions d’application qui permettront une efficacité maximale de l’application. La dose appliquée (pleine ou réduite) peut également fortement influencer l’évolution de la résistance. Au vu de la facilité d’emploi des PPP (« un pt’it coup de pulvé, et c’est plié »), ces recommandations peuvent sembler contraignantes. À court terme, il semble logique de privilégier la simplicité dans le contrôle des bio-agresseurs. Mais le contrôle des bio-agresseurs ne peut se raisonner que dans la durée et collectivement, en considérant l’efficacité des PPP comme un bien commun. En cas de généralisation de la résistance et sans efficacité pratique résiduelle du PPP concerné, il est souvent inutile de continuer à utiliser le PPP concerné. Le contrôle du bio-agresseur résistant devra donc être recherché par d’autres méthodes, chimiques ou non. Une fois

43
la résistance établie, mettre en place des mesures de gestion est en général bien plus contraignant, moins efficace, voire plus coûteux, qu’utiliser une stratégie de prévention des résistances, surtout dans le contexte actuel de réduction de l’utilisation des intrants chimiques et de manque de nouveaux modes d’action.

44
Comment faire face à la résistance aux fongicides chez la vigne pour un distributeur ? Le cas de VIVADOUR
P. Mauranx 1VIVADOUR, Rue de la Menoue, F-32400 Riscle [email protected] Résumé :
Depuis plus de 25 ans, le groupe coopératif VIVADOUR s’est engagé dans une démarche de protection raisonnée du vignoble. Cette démarche repose sur une parfaite connaissance du risque épidémique des principales maladies de la vigne (Mildiou, Oïdium et Pourriture grise) et la mise à disposition pour le viticulteur d’un réseau d’Avertissements Agricoles personnalisés. Mais lorsque l’application d’un traitement fongicide devient nécessaire une question cruciale se pose : avec quel(s) mode(s) d’action est-il préférable d’intervenir ? Pour répondre avec pertinence à cette question, une bonne connaissance de l’efficacité des principaux modes d’action est nécessaire pour chaque pathogène mais pas suffisante : la résistance aux fongicides peut remettre en question localement la performance des fongicides. Conscient des conséquences de cette problématique avec la découverte en 1989 de baisses d’efficacité des IDM sur l’Oïdium de la vigne, VIVADOUR réalise, chaque année, depuis cette date une surveillance de l’évolution des phénomènes de résistance dans sa zone d’activité. En moyenne, 100 échantillons de Mildiou, 40 d’Oïdium et 60 de Botrytis sont prélevés chaque année et étudiés vis-à-vis de 4 à 6 modes d’action différents. En complément, des essais d’efficacité sont réalisés au vignoble pour évaluer l’impact de chaque cas de résistance et déterminer quels sont les meilleurs programmes anti-résistance.
L’objet de cette présentation est d’illustrer la démarche retenue par VIVADOUR en prenant un exemple concret : l’Oïdium de la vigne (Erysiphe necator) et la résistance aux fongicides QoI.
La détection en 2008 de phénotypes d’E. necator résistants aux QoIs dans le Gers et leur forte progression dans les populations entre 2009 et 2010 dans cette région a considérablement réduit les performances des programmes de lutte contre l’oïdium. Les fréquences moyennes de phénotypes résistants dépassent 75% à compter de 2011, sans baisse significative.
Des essais ont rapidement été mis en place et comparent l’efficacité de quelques programmes à base de QoIs dans cette nouvelle situation de résistance. La baisse d’efficacité des programmes à base de 2 traitements QoIs est de 40% à 53% par rapport au programme sans QoI en présence de 10% ou plus de phénotypes résistants en début d’expression de l’épidémie d’oïdium. Cette perte reste très élevée même après l’utilisation d’un seul traitement QoI dans le programme (34% et 41%).
Le choix d’une association entre un QoI et un SDHI limite fortement cette baisse d’efficacité dans un programme à 2 applications consécutives (5% et 8%). Par contre, l’utilisation d’un traitement avec un autre mode d’action entre les deux applications de l’association entre QoI et SDHI se traduit systématiquement par une perte d’efficacité comprise entre 20% et 30%. Le maintien d’un traitement à base de QoI mais associé à un SDHI et appliqué au stade grossissement de baies BBCH75 dans les programmes de traitement procure les meilleures performances en combinaison avec d’autres modes d’action (IBS groupe II et APK). Mots clef: Gestion de la résistance, Fongicides, Vigne, Distribution

45

46
Détection et gestion de la résistance de la pyrale du maïs à la famille des pyréthrinoïdes
Jean-Baptiste THIBORD 1ARVALIS – Institut du végétal, Chemin de Pau, 64 121 Montardon, France [email protected]
Résumé : À partir du milieu des années 2000, quelques parcelles situées en région Centre ont rencontré des difficultés à contenir les attaques de pyrale du maïs malgré la mise en œuvre d’une protection insecticide. Des expérimentations ont alors été mises en œuvre. Elles ont permis de confirmer la perte d’efficacité de plusieurs produits dont la substance active appartient à la famille des pyréthrinoïdes dans les essais conduits en région Centre alors que ces produits conservent une bonne efficacité dans les essais réalisés dans d’autres régions de France.
Des larves de pyrale du maïs ont alors été prélevées dans deux parcelles distinctes : l’une issue de la région à problème et l’autre dans une région témoin où aucune perte d’efficacité n’avait été constatée. Les populations ont été élevées en conditions contrôlées au laboratoire de l’INRA – UR PSH en vue de procéder à des tests de résistance sur néonates. En 2012, les résultats des tests montrent que les deux populations répondent différemment à la lambda-cyhalothrine. La population provenant de la région Centre est moins sensible que la population témoin. La différence de DL50 est significative et le rapport de résistance à la dose létale 50 est de 119. Ces résultats expliquent les échecs de protection insecticide constatés au champ dans cette zone géographique.
La pyrale du maïs a donc développé une résistance à une substance insecticide dans une situation qui présentait un risque a priori limité (une seule génération par an, maïs assolé). Cependant, la faible diversité de solution insecticide a conduit les producteurs à recourir trop fréquemment à la famille des pyréthrinoïdes, seule famille chimique autorisée pour lutter contre la pyrale du maïs depuis le milieu des années 80.
Par chance cette résistance a été mise en évidence au moment où d’autres solutions de lutte sont arrivées sur le marché. Ainsi, plusieurs familles chimiques ayant des modes d’action différents ont pu être utilisées depuis 2011 comme par exemple les diamides anthraniliques avec la substance active chlorantraniliprole (Coragen®). Après plusieurs campagnes d’utilisation dans la région Centre, les populations de pyrale du maïs ont de nouveau été contenues grâce à l’utilisation d’insecticide efficace combinées à des mesures prophylactiques habituelles.
Désormais, l’objectif est d’intégrer ce risque de résistance dans les stratégies de protection à la fois en région Centre pour maintenir une efficacité des nouvelles solutions, mais aussi partout ailleurs pour éviter qu’une résistance se développe sur les produits disponibles, y compris ceux appartenant à la famille des pyréthrinoïdes dont l’efficacité reste satisfaisante à ce jour.
Mots-clefs: Pyrale du maïs, Ostrinia nubilalis, résistance aux insecticides, lambda-cyhalothrine, pyréthrinoïdes.

47

48
Le désherbage non chimique
J.P. Guillemin1 & B. Chauvel2 1AgroSup Dijon, UMR1347 Agroécologie, 26 bd Docteur Petijean, BP 87999, F-21079 Dijon 2INRA, UMR1347 Agroécologie, 17 rue Sully, BP 86510, F-21065 Dijon [email protected] Résumé :
L'apparition de mauvaises herbes résistantes aux herbicides a conduit à développer de nouvelles solutions pour gérer la flore adventice. Un des leviers est de diminuer la pression exercée par les molécules herbicides sur les espèces d’adventices en optimisant le désherbage chimique. Cet ajustement doit passer par une alternance des modes d’action des herbicides appliqués sur une parcelle aux échelles annuelle et pluriannuelle. Il peut également passer par des réductions de quantités d’herbicides appliquée à l’échelle de la parcelle en ne traitant que les zones où sont très présentes les mauvaises herbes. La baisse significative de l’usage des herbicides pour lutter contre les mauvaises herbes nécessite de substituer ces intrants par d’autres interventions dans la parcelle. Plusieurs techniques agronomiques sont maintenant bien identifiées pour agir sur le contrôle de la flore adventice. Les stratégies préventives visent à réduire les possibilités d’installation des plantes adventices à l’implantation de la culture. La modification de la date de semis de la culture peut créer un décalage entre la période de levée de la culture et les périodes optimales de certaines adventices. Cette technique peut être étendue à l’échelle de la rotation en diversifiant cette dernière par une succession de cultures dont les dates d’implantation sont différentes. Une seconde technique consiste à travailler le sol avant le semis de la culture afin de créer les conditions optimales pour la levée des semences de mauvaises herbes. Les adventices ainsi levées seront détruites avant l’implantation de la culture. Il s’agit de la technique du faux semis qui, associée au décalage de la date de semis, peut en conditions favorables se révéler très efficace. Le labour peut être utilisé pour enfouir les semences de mauvaises herbes ; il s’avère très efficace pour détruire les semences dont la persistance est courte (moins de 1 an à 2 ans) dans le stock semencier. Une autre stratégie est d’intervenir au cours de la culture. Dans ce cadre, il est envisageable d’augmenter l’aptitude concurrentielle de la culture afin de réduire la présence et l’impact des mauvaises herbes. Ceci peut passer par l’augmentation de la densité de semis de la culture ou par des choix variétaux. Une autre approche est de détruire les adventices pendant le cycle cultural par le désherbage mécanique (passage d’outils type herse étrille, bineuse, …). Cependant l’efficacité de chaque technique agronomique sur le contrôle des adventices est partielle, et tout cas plus faible que celle attendue d’un traitement de type herbicide. Il est donc nécessaire d’associer et combiner plusieurs de ces techniques, en recherchant des complémentarités, afin de maîtriser la présence des mauvaises herbes dans les parcelles cultivées. Des essais ‘longue durée’ permettant de tester un ensemble de pratiques culturales à long terme se révèlent nécessaires pour obtenir des résultats fiables et reproductibles. Mots clef: Gestion de la résistance, Herbicides, Grandes cultures

49