Embed Size (px)
Citation preview

Eur. J. Biochem. 35,372-379 (1973)
Pr oprikt ks des formes allongkes de l’acktylcholinestkrase en solution Rayon de Stokes, densit6 et masse
Suzanne BON, Franpois RIEGER et Jean MASSOULI~ Institut de Biologie Physico-Chimique, Paris
( R e p le 26 dbcembre 1972/13 mars 1973)
We present here several molecular properties of the asymmetric acetylcholinesterase forms, which we described in our previous publications [l-31. From molecular sieve experiments, we determined the Stokes radius and compared its value with that calculated from micrograph measurements [3] for the A form which is a tetramer associated to an elongated element, called the “tail“ (structure en grappe). A good agreement is obtained with a prolate ellipsoid of identical volume and length ; this indicates that the acetylcholinesterase is, in solution, a rigid structure.
We also estimated the partial specific volume, @, of acetylcholinesterase, by two independent methods: first, by the difference of sedimentation velocity in gradients of different overall density (H,O, ,H,O) ; secondly by absolute density measurements in CsCl equilibrium gradients. Both methods are in reasonable agreement and yield a value of 0.74-0.75 for all forms of the enzyme. This enabled us to calculate, from sucrose gradient data, the sedimentation constant in water a t 20 “C. Combining the values of the Stokes radius, sedimentation constant, and partial specific volume, we calculated the molecular mass of A, C, and D as 430000, 780000, and 1100000. These values justify our hypothesis that these forms possess one, two, and three tetramers, associated with the “tail”.
The methods used in this work are all based on activity measurements, which is a major advantage since only minute quantities of protein are needed. However, these are mainly comparative determinations. and it is difficult to ascertain the precision of the published data for the reference proteins.
Parmi les d8brentes formes molhculaires de l’acbtylcholinestbrase que nous obtenons B partir des organes Blectriques de gymnote, e t que nous avons caractkrisbes par leur constante de sbdimen- tation[l], nous avons distingub par la suite des formes globulaires, G et G‘, et des formes asymb- triques, A, C et D (sue nous avions appelbes ((natives o). En effet, celles-ci sont susceptibles d’etre brishes par les ultrasons et elles sont exclues de tamis molbculaires tels que le Sephadex G-200 ou le Biogel P 300 [2]. Cette derniere propribtb montre que le rayon de Stokes de ces formes emyma- tiques est plus grand qu’il ne le serait pour des protbines globulaires de meme constante de sbdi- mentation. Cependant, cette observation ne permet ni de distinguer entre elles les formes A (8,5S), C (14,2S) et D (18,5S), ni d’estimer la valeur de leur rayon de Stokes.
Une telle donnee fournirait sur la structure de la molbcule des renseignements qu’il serait intbressant de comparer avec ceux dbduits des micrographies
de l’enzyme que nous avons obtenues rbcemment [3]. Alors que les formes globulaires G et G’ appa- raissent respectivement comme des tbtrambres et des dimbres, les formes A, C et D ont une structure en ccgrappe,: B une ((queue)) en bhtonnet est fixhe la cct6te)), ensemble de globules qu’il est possible de dbnombrer approximativement (on en compte 3 B 4 pour A, 6 A 8 pour C, 10 B 12 pour D).
Sous l’action de la trypsine, toutes lea formes asymbtriques sont converties en une forme globu- laire tbtrambrique, Gt. Puisque cette forme t6tra- mhrique semble particulierement stable, nous avons fait l’hypothbse que le tetramere constitue une tunitb d’interaction)) dans les structures plus com- plexes, en ce sens que des liaisons privilbgibes prb- existeraient entre quatre sous-unit& au sein de ces structures. Cette hypothese ambne B supposer que les formes A, C et D contiennent respectivement un, deux et trois tbtrameres; elle explique donc l’existence de ces btats d’association discrets de sous-unit&. Cependant, l’existence du dimere et

Vo1.35, N0.2.1973 S. Bos, F. RIEQER et J. MAssom~fc 373
son obtention par action des ultrasons sur la forme A, prouvent que le tetramere n’est pas l’unit6 de liaison la plus petite.
I1 est clair qu’une etude de comportement hydrodynamique est indispensable pour prBciser ces structures. Nous avons donc cherche B dBter- miner les rayons de Stokes des formes asymhtriques de l’acBtylcholinest6rase en utilisant la methode des tamis moleculaires ; nous avons Bgalement cherche B determiner le volume partiel specifique de ces enzymes, par sedimentation en gradient de sac- charose en presence de ,H,O et H,O, et par centrifugation B 1’Bquilibre dans un gradient de densite (CsC1). L’ensemble de ces deux types de mesures nous a permis d’estimer les masses mole- culaires de ces structures.
MATGRIEL ET MaTHODES
ProtLines
Les methodes de preparation e t de dosage de l’ac6tylcholinestBrase ont d6jQ 6th decrites en d& tail [I]. Les differents marqueurs ont 6th deter- minBs, soit par leur activatite enzymatique: alcool dbshydrogenase de levure, catalase, /?-galactosidase d‘Escherichia coli [I], urBase [4]; soit par absorption: ferritine Q 415 nm, ferricyanure de potassium B 290nm, fibrinoghne de bceuf (fraction l ) , myosine de lapin, collagene de tendon de bamf, et phage T7 entier, Q 280 nm.
Chromatographies sur tamis moldculaires
Nous avons utilise une colonne, de diametre 2,l cm et de longeur 1 m. Apres l’avoir traitbe au dimBthyl-dichlorosiane afin de la, rendre non mouillable, nous l’avons remplie sur une hauteur de 95 cm de Bio-Gel A 15m 200-400 mesh (Biorad), et l a d e B 4°C avec 5 fois son volume de tampon Tris 0,Ol M (pH 7), NaCl 1 M, MgC1, 0,05 M, azide 0,001 M, pour assurer son Bquilibration. Les WB- rents produits chromatographier sont alors dBposes au sommet du gel dans un volume d’environ 1 ml, tamponne comme prBc8demment. Aprhs pen& tration de l’echantillon, la surface du gel est rapidement lavBe de fagon B Bviter la formation de trainBes de produits. L’Blution de la colonne est faite Q 4 “C avec le m6me tampon, B un debit de 4ml par heure, et des fractions de 1 ml sont recueil- lies dans un collecteur Gilson.
Par rapport aux autres types de gels que nous avons essayes (P300, Sepharose 4B et 2B, Agarose 1,5 m et 5 m), le Biogel A 15 m permet la meilleure separation de nos formes enzymatiques dans la zone oh la constante d’elution K D varie lineairement avec le logarithme de la constante de diffusion.
Tableau 1. Donnkes relatives aux protkines de rifjerence Les valeurs soulignks ne correspondent pas aux references, mais It nos mesures d‘bquilibre dans un gradient de chlorure de cbsium. Les valeurs donnees dam la littbrature varient parfois sensiblement suivant les auteurs et les
mbthodes utiliskes ~~~~ ~~
Masse xayon Constante - mol6culaire de Stokes de s6dimen- ’ Proteine
tation nm S
Ferritine [5] 1300000 7,9 65 0,73 Myosine [6] 493000 21,5 6,4 0,73 B-Galaetosidase [7] 540000 8 2 16 0,77 Urease [a] 483 000 691 18,6
Catalase [6] 250 000 5 2 Alcool dbshydro-
Tropomyosine [a] 93000 9,6 2,36 SBrum albumine
Fibrinoghe [6,8] 330000 10,7 7,9 0,71 11,3 0,77
ghase [5] 150000 496 7,4 0,78
-
de bmuf [5] 65 000 395 4,36 0,73
RI~SULTATS Rayon de Stokes
On sait que le retard subi par une proteine dans une colonne de tamis molBculaire ne dBpend pas de sa masse molhculaire, mais plutBt de sa constante de diffusion, ou de son rayon de Stokes Re. Ces deux constantes sont liees par la relation suivante:
kT 6x71D0 Re =
oh k est la constante de Boltzman, T la temperature absolue, 17 la viscositi: et Do la constante ne diffusion, extrapolBe jusqu’h concentration nulle. I1 est donc en principe necessaire d’extrapoler une sBrie de deter- minations en fonction de la concentration. NBan- moins, la quantitB de proteine disponible Btant limitke, nous n’avons utilise qu’une seule concentra- tion pour chaque protitine ; cette concentration Btait d’ailleurs souvent trhs faible (de l’ordre de la nanomole) puisque nous pouvions doser les activitbs enz ymatiques.
relatives aux proteines que nous avons utilisbes comme marqueurs sont rappor- tees dans le Tableau 1.
Le comportement d’une protBine dans le tamis molBculaire est caractBrisb par le parametre d’Blu- tion K D :
Certaines donnBes
oh V , est le volume d’6lution de la proteine, V,, le volume d’exclusion, et V f le volume d’Blution des
Notons que ces donnees sont relatives It une temp& rature de 20 “C, alors que nos experiences ont ktb realides It 4 “C. Nous admettons que le fachur de temperature est Ie m6me pour toutes les protkies.
374 Rayon de Stokes, densite et masse de I’ac&ylcholinestt%ase
I I I I i I I I I I I I
T7 Myosine
+ I
100 Nurnkro des fractions
150
Eur. J. Biochem.
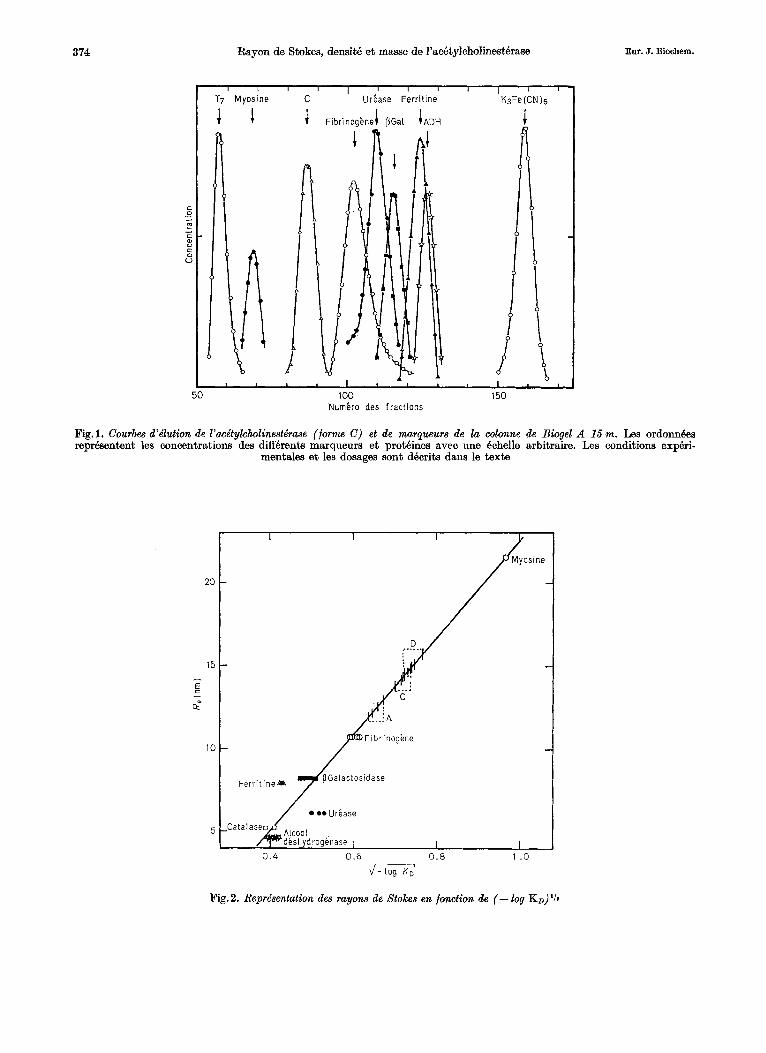
Fig.1. Courbes d’dlution de l’acdtylcholinest6rme (forme C ) et de margueurs de la wlonne de Biogel A 15 m. Les ordonn6es reprbsentent les concentrations des diffbrents marqueurs et protBines avec m e Bchelle arbitraire. Les conditions expbri-
mentales et les dosages sont dkcrits dans le texte
** UrQase
Alcool dkhydroggnase I I
0.4 0.6 0.8 L
1 .o m Fig. 2. Reprbentation des rayons de Stokes en fonction de (- log KD)’Ir

Vol.35, No.2,1973 S. BON, F. RIEGER et J. MASSOULI~ 375
petites molhcules. Nous avons dans chaque experience mesure V , Q l’aide de phages T7 entiers, et Vf b l’aide de ferricyanure de potassium. La limite d’ex- clusion correspond au collagbne, dont le rayon de Stokes est de 31 nm [6].
Pour determiner la constante de diffusion, ou rayon de Stokes, des diffkrentes formes molkcu- laires de l’achtylcholinestbrase, nous avons comparb leurs constantes ( K D ) d’6lution d’un tamis mole- culaire, Q celles des proteines utilisees comme mar- queurs. La Fig.l montre comment s’eluent d’une colonne de Biogel A 15 m diffhrentes prothines ainsi que la forme A de l’ac8tylcholinest6rase. Nous avons vhrifih, par centrifugation en gradient de saccharose, qu’aucune transformation des formes enzymatiques ne s’est produite au cours du passage sur la colonne.
La relation entre le parametre experimental KD et les valeurs des rayons de Stokes des marqueurs donnkes dans la litterature, est indiqube graphique- ment dans la Fig.2, selon la representation empiri- que :
Re =
qui permet d’obtenir une droite [5]. Nous avons tenu surtout compte, pour la calibration de la colonne, du fibrhogene et de la myosine qui se repartissent dans la fraction intermediaire d’klution, oh cette relation lineaire se trouve la mieux v6rifi6e. Elles prbsentent, par ailleurs, un autre avantage : leur structure posskde b la fois des parties globulaires et des parties en bbtonnet, et rappelle celle que nous avons mise en evidence par microscopie Blectro- nique pour les formes A, C et D [3]. En faisant oette interpoIation avec la myosine et le fibrinogene, nous obtenons, pour A, C et D les valeurs Re indi- quQes dans le Tableau 2.
On peut Bgalement analyser les rbsultats exp6ri- mentaux en utilisant un seul marqueur. En effet, Ackers a donne une fonction reliant le parametre d’hlution, KD, au rapport du rayon de Stokes equivalent et d’un parametre caractbristique du gel: ctle rayon de pore effectif)), r [9]. D’aprhs les tables numeriques de cette fonction, il est possible, connaissant K D et Re pour m e protbine de reference, de trouver la valeur de ce parambtre, r, et par suite, de determiner le rayon de Stokes d’autres proteines B partir le leur parametre KD. Nous avons fait ces calculs par rapport A la myosine et au fibrinogene: les valeurs obtenues pour le rayon de pore effectif du Biogel A 15m sont dSerentes, respectivement 56,2 nm et 60,s nm. Cet &art montre que la relation htablie par Ackers ne s’applique que de fagon approchbe au Biogel A 15 m. Nous prkfererons donc la determination graphique empirique de la Fig.2, les valeurs de Re ainsi obtenues s’inserant d’ailleurs assez bien entre les couples des K D calcules selon la
Tableau 2. Valeurs expbimentales du paramLtre d’ilution Ko et valeurs wrrespondantes du rayon dquivalent de Stokes pour
les formes ctnatives B de l’acktylcholinesthsrase
Valcur des rayons Bquivalents de Stokes Re d‘aprhs:
Borme Valeiir ~~. - .~. mol6cu- expBrimentale la relation la mhthode d‘Ackers [91
laire de KD ’:< r = 56,2 nm r = 60,s nm (c- log xD) (myosine) (fibrinonhe)
nm nm nm A 0,352-0,367
-0,383-0,392 12,4 & 0,6 11,4 12,4 C 0,282-0,291
- 0,302 - 0,306 -0,320 14,4 & 0,6 13,5 14’7
D 0,258-0,276 -0,281-0,300 15,O f 1,0 14,2 15,4
methode d’Ackers (Tableau 2) . La principale cause d’incertitude provient indubitablement du manque de proteines de reference dans la region utile de la courbe.
Ces rhsultats mettent en hvidence le caractere non globulaire des formes enzymatiques. A partir des rayons de Stokes obtenus, on peut d’ailleurs essayer de repondre Q la question suivante: la ((queue)), telle qu’on la voit sur les micrographies [3] est-elle rigide, dans la solution? Pour le savoir, nous avons calculQ, d’aprhs les mesures effectubes sur les micrographies, ce que vaudrait le rayon de Stokes de la forme A, supposhe rigide. Cette forme moleculaire est constitutee de 4 globules associes B une ((queue)). Nous avons donc calcule le volume de la molecule et les dimensions d’un ellipso’ide de revolution allong6 de volume Qgal, et dont le grand axe a la m6me longueur que la molhcule. A partir du rapport des axes de l’ellipsoide, on obtient le rapport (/ifo) entre le rayon de Stokes de la molecule e t le rayon d’une sphere de m6me volume [S].
Les resultats sont donnes dans le Tableau 3, pour des estimations extr6mes des dimensions molbculaires: on obtient des valeurs de Re comprises entre 8,5 et 12 nm, ce qui est en bon accord avec les rbsultats expbrimentaux (11,4- 12,4 nm), puisque l’assimilation de la structure ten grappe)) de la forme A b celle d‘une ellipsoide est tres approch8e2. Pour une structure globulaire de meme volume, on obtient un rayon de 5,l-6,6 nm. On peut conclure que la, ((queue)) se comporte en solution comme un bBtonnet rigide. On arrive b la m6me conclusion pour la myosine, en analysant la diffusion de la lumiere [ 121.
L’accord est d’autant meilleur qu’il s’agit sans doute d‘une sous-estimation, car en effectuant le m6me calcul pour la myosine (longueur 150 nm, diambtre 3 nm) [ll], on obtient 17,5nm, au lieu de 21,5nm. Cet effet est probablement dfi en partie L l’hydratation des protbines.

376 Rayon de Stokes, densitk et masse de l’ac6tylcholinesGrase Eur. 3. Biochem.
Tableau 3. Calcul du rayon de Stokes de la forme moldculaire A , d’ap2s les donnth des micrographiea Le rapport entre le rayon de Stokes et celui de la sphbre de mbme volume est obtenu ti partir de la ref. [6]
Diametre Longueur Diametre Rapport d’asymbtrie Rayon de la Rayon de d‘un de la de la a/b d‘un ellipsoide sphere de f/fo Stokes
globule * queue )) u queue a de meme volume m&me volumc
nm nm nm nm nm
Valeur minimum 5,5 43 2 3 1,22 5,i 1,65 8,4 Valeur maximum 6 5 67 325 196 683 1,82 12,o
20
15
10 5
5
0
Nurnhro des fractions
Fig. 3. Gradient de saccharose en zH,O. Les constantes de sedimentation de C et D sont dbterminbes d’aprbs oelles de I’alcool d6shydrog6nase (ADH) et de la /?-galactosidase (/?-Gal)
Constante de ddimentation et volume spkci fique partiel Dans nos precedentes publications, nous avons
caracthrisi? lee diffhrentes esphces d‘ac6tylcholi- nesterase par leurs constantes de st5dimentation d6terminhes par comparaison avec des enzymes marqueurs (alcool dhshydroghase et p-galactosidase). La methode utilisee est la centifugation dans un gradient lineaire de saccharose, selon Martin et Ames [lo]. Nous allons voir gue cette methode permet m e determination approximative de la constante de sbdimentation dans l’eau et du volume partiel sp6- cifique, 17, de I’ac8tylcholinest&rase, en utilisant les resultats d’exphiences r6alisees dans deux milieux de densit6 Wbrente (eau et eau lourde) (Fig.3).
Nous avons utilise une methode d’analyse lbghrement diffkrente de celle utilisee par Meunier, Olsen et Changeux 11131 pour la prothine receptrice de l’sc6tylcholinest8rase; nous ne mesurons pas les densites des dif€&entes fractions des gradients de saccharose.
Nous ferons les approximations suivantes: a) Bien gue les valeurs de @ de l’aleool dbshydro-
genase et de la b-galactosidase soient diffhentes (Tableau l), nous utiliserons une valeur moyenne.
b) Nous supposerons [bien gue nous utilisions des milieux de densite Mbrente (H,O et ,H,O), et mal- gr6 les diffhrences de volume partiel des prot4ines consid6r15es] (rue l’on peut admettre dans tous les cas que la vitesse de migration dans le gradient est constante. C’est cette observation de Martin et Ames [lo] qui est B la base de l’utilisation des gradients de densite. Elle permet d’6crire une rela- tion de la forme:
s = a x (3) oh x est la distance parcourue, proportionnelle B la vitesse, et oh s represente une constante de shdimen- tation, dans les conditions du gradient ; celle-ci depend de la densit6 du milieu et du volume partiel sphifique de la prot6ine. Le parametre 01 depend des conditions de centrifugation (durbe, vitesse).

Vo1.35, No.2,1973 S. BON, F. R I E ~ E R et J. MAssoa~d 377
La mesure de x ne peut nous donner qu’une cons- tante de s6dimentation apparente : nous normalise- rons ces mesures arbitrairement b l’aide des cons- tantes de sedimentation des marqueurs, determinees dans les conditions standard (sw). Les indices 0 et I se rhfkrent respectivement aux marqueurs et b l’ac6tylcholinestbrase, les donnkes relatives aux centrifugations dans l’eau lourde sont notees s’, XI, etc. Les constantes de sedimentation apparentes sont donc :
81 bPP) = (XllXO) sow (4 a)
sl‘(aPP) = (%’/XO? sow. (4b) et
c) La relation (3) definit une constante de sedimentation a. L’existence du gradient de densit6 permet de realiser la condition de Martin et Ames, oh la vitesse de sedimentation est approximative- ment constante [2]. Nous admettrons donc qu’entre la constante de sedimentation dans le gradient et les paramktres moleculaires, on peut h i r e la rela- tion :
dans laquelle Q sera la densite du milieu en absence de saccharoses.
Dans l’eau lourde, la relation (5) devient :
a = - DM (k - 8e’) RT
oh k repr6sente la variation de la masse due It 1’6change protons-deuterons dans la proteine. On admet generalement que k a la mhme valeur pour toutes les prot6ines; la valeur mesuree pour la serum albumine est k = 1,0155 [14].
Comparons alors les resultats obtenus dans les centrifugations en H,O et ,H,O, et pour cela, Bcrivons le rapport, r, des constantes de skdimenta- tion apparentes de l’ac6tylcholinest6rase qui, d’aprks (3) et (4) est &gal a :
(7)
On obtient alors, en reportant (5) et (6) dans (7), une relation oh le rapport, r, ne depend gue des volumes partiels des proteines :
k - El@‘ . I - E o @ k - Go@’ 1 - E l @ - r =
Connaissant r et do, il est possible de calculer 8,. C’est ce gue nous avons fait avee les donnees du Tableau 4. Pour e et c’, nous avons pris les densites de solutions molaires de chlorure de sodium b 4 “C,
L’approximation supplBmentaire que nous intro- duisons ainsi se justifie d’autant mieux que des termes correspondants se retrouvent Q la fois en dhominateur et numhrateur dans lea relations que nous utiliserons.
25 Eur. J. Biochem., Vo1.35
dans H,O et ,H,O, 99O/,, soit e = 1,039 et e’ = 1,14
Puisque la valeur de r est la meme, aux erreurs d’experiences prks, pour toutes les formes mole- culaires de l’acetylcholinest6rase (Tableau 4) leurs volumes specifiques partiels sont indiscernables d’aprhs ces donnees. Avec 8, = 0,775 (cf. ci-dessus), nous obtenons 8, = 0,755 pour l’ac6tylcholinest6- rase.
~ 5 1 .
Dktermination de 7 en Lquilibre de sbdimentation L’Bgaliti? des volumes partiels des formes
moleculaires acetylcholinesterasiques est mise en evidence Bgalement par centrifugation b l’equilibre dans un gradient de chlorure de cesium: toutes les formes de l’ac6tylcholinestkrase se placent au meme niveau dans le gradient, leur densite d’6quilibre de flottaison est done la meme. Rappelons que les proprietes (activib.5 enzymatique, sedimentation) de l’ac6tylcholinest6rase ne sont pas affectees par ce traitement .
Nous avons cherche B determiner la valeur de 8 en utilisant cette methode d’equilibre. Nous avons, pour cela, realis6 un Bquilibre de flottaison de la forme C, en presence de marqueurs (urease, catalase, 8-galactosidase). L’equilibre est atteint B 20 “C et les densites sont mesurees 8, cette temperature. La densite d’bquilibre isopycnique de la B-galactosidase est de 1,301 (A 20 “C), alors que celle de l’acetyl- cholinesterase est de 1,330. Les valeurs correspon- dantes de B sont 0,769 et 0,752 (en admettant qu’il n’y a ni solvatation, ni hydratation prkf6rentielle pour ces protkines, ce qui est vraisemblable en prdsence de NaCl M et MgC1,0,05 M).
Pour comparer avec la determination pr6c6dente, nous avons calcul6 les marges d’incertitude des deux methodes. La m6thode de centrifugation b 1’6quilibre ne depend que de pesees et de la localisation du pic d’activit6 enzymatique : 0,751 < 8 < 0,753. En ce qui concerne la methode de sedimentation en H,O et aH,O, il y a une incertitude sur lea valeurs experimentales de s(app.) (0,2S) qui donne 1,049 < r < 1,066, et une approxi- mation que nous estimons en prenant, dans la relation (8), les valeurs de 8, extrbmes pour les marqueurs, au lieu de la valeur moyenne. On obtient ainsi 0,745 < B, < 0,765. Nous constatons gue ces deux determinations sont parfaitement compatibles, ce qui confirme que la presence de chlorure de cesium ne modifie probablement pas 8: une fixation d’ions Csf en diminuerait la valeur. En effet, les 6 des marqueurs ainsi determines sont superieurs A ceux donn6s dans la litterature (8 de la 8-galactosidase = 0,76 [7]). Peut-&re cela est il dii en partie B l’effet de la pression dans les conditions de la centrifugation, entrainant une augmentation de l’ordre de 2,/, de la densite du milieu (on peut

37s Rayon de Stokes, densite et masse de I’ac6tylcholinesthrase Eur. J. Bioohem.
Tableau 4. Constantes de sddimentation des formes moldculaires de l’ac~tylcholinestdrase
Les constantes apparentes sont determinks par centri- fugation dans un gradient lineaire de saccharose, en presence de NaCI, 1 M; MgCI,, 0,05 M; tampon Tris, 0,Ol M (pH 7), b 6 “C. La constante apparente s‘ est determinee dane 2H,0
(9Qo/o) Constante de Constante de Constante de
pome sedimentation sedimentation sedimentation mol6cu- apparente apparente dam rapport 7 ramenee a
laire dans l’eau l’eau lourde 20 “C dans [s (app.)l [s (app.)l l’eau ( 8 8 0 . ~ )
S S S
A 9 2 998 1,06 9 C 14,2 14,9 1,05 14 D 18,4 19,4 1,06 18,l G 11,8 12,4 1,05 11,6
estimer la pression au milieu du gradient, oh s’equi- Iibrent les protkines, B 500 bars environ).
Calcul des s20,
AprAs avoir obtenu cette valeur de 8,, nous pou- vons calculer la constante de sedimentation szo, des formes A, C et D de I’acetylcholinesthase. Nous avons donc4 :
Cette relation nous permet de calculer slw A partir ds s,(app.) avec e = 1,039 et ew = 1. On obtient les valeurs indiquees dans le Tableau 4.
Masses molkculuires Connaissant la constante de diffusion (ou le
rayon de Stokes equivalent), grBce aux tamis
Explicitons, B partir de (3) et (6), les relations entre la migration dans le gradient de saccharose, x, et les cons- tantes de sedimentation dans le milieu de densite e:
et
et
Dana l’eau
On en tire:
Or, nous avons determine expbrimentalement : 21
4 sl(app.) = s o w . -.
1
a Y)
‘f 2 a5
0
r C
Fi brinogkne
0 500 1000 1500 R,S / ( l - F p ) ,
Fig.4. DBermination de la masse en fonctwn de Re s / ( l - ? e ) . Les valeurs utilisees sont celles des Tableaux 1, 3 et 4. Les constantes de sedimentation sont prises dans l’eau, et nous
avons pris e = 1
molkculaires, la constante de sedimentation et le volume speciiique partiel, il est possible de dhter- miner la masse moleculaire des diffhrentes formes de l’ac8tylcholinestkrase. Celle-ci est en effet propor- tionelle au rapport Re s/(1 - 8 e).
Nous procederons B une determination graphi- que, par comparaison avec les proteines qui nous ont deja servi de marqueurs et pour les- quelles nous garderons les valeurs de masse, 8 et rayon de Stokes indiquees dans le Tableau 1. Pour l’ac8tylcholinest&rase, nous utiliserons les valeurs experimentales determinees prec6demment (Fig.4). Nous obtenons ainsi des masses de 430000 pour A, 780000 pour C et 1100000 pour D.
CONCLUSION Les resultats present& ici peuvent &re obtenus
avec de faibles quantites d’enzyme, puisque nous utilisons toujours les mesures d’activite. Mais, B part la mesure de la densite dans un gradient de chlorure de cesium A l’equilibre, il s’agit de deter- minations comparatives. Or, les donnhes relatives aux proteines de reference (8, rayon de Stokes, et m6me masse moleculaire) sont assez incertaines et souvent difficilement comparables. C’est pour- quoi il est delicat d‘estimer la precision des readtats obtenus.
Nbanmoins, ils nous permettent de prbciser les proprihtes des differentes formes molkculaires de

Vo1.35, No.2,1973 S. BON, F. RIEQER et J. MASSOULI~ 379
1’acBtylcholinestBrase. A condition d’assimiler la forme A il un ellipsoide, dont les dimensions corres- pondent Q celles des rnicrographies [3], on calcule pour cette structure rigide un rayon de Stokes qui correspond bien & la valeur experimentale. I1 est donc probable que la ((queue)) observhe par micro- scopie Blectronique, est m e structure aussi rigide que celle de la myosine, en bhtonnet.
D’autre part, en combinant les valeurs obtenues pour les rayons de Stokes, les constantes de s&hentation, et la densit6 (l/6), nous pouvons estimer les masses molhculaires des formes A, C et D: soit 430000, 780000 et 1100000. Les diffhrences entre A et C, et C et D, sont de 350000 et 320000, respectivement, ce qui est proche de la masse d’un tetramhre, et justifie partiellement l’hypothhse que nous avons faite [3] sup la structure des formes asymhtriques de l’acBtylcholinestBrase, d‘aprds laquelle la ccti3te)) de A contient un tetramhre, celle de C deux thtramhres, et celle de D trois tBtra- mkres.
Nous remercions Janet Powell pour les discussions cri- tiques et constructives que nous avons eues avec elle au cours de la Aalisation de ce travail. I1 a bbnbficib de sub- ventions du Centre National de la Recherche Scientifique (G.R. No 18) et de la DBlBgation GBnBrale B la Recherche Scientifique et Technique (Convention 71.7.3049).
I.
2.
3.
4.
5.
6.
7. 8.
9.
10.
11. 12.
13.
14.
15.
BIBLIOGRAPHIE MassouliB, J. & Rieger, F. (1969) Eur. J . Biochem. 11,
Massoulib, J., Rieger, F. & Bon, S. (1971) Eur. J . Bio- c k m . 21, 542-551.
Rieger, F., Bon, S., MassouliB, J. & Cartaud, J. (1973) Eur. J . Biochem. sous presse.
Siegel, L. M. & Monty, K. J. (1965) Bwchem. Biophys. Res. Comrnun. 19, 494-499.
Siegel, L. M. & Monty, K. J. (1966) Bwchim. Biophys. Acta, 112, 346-362.
Tanford, C. (1961) Physical Chemistry of Macromolecules, 5th ed., pp. 317-456, I. Wiley and Sons, New York.
Sund, H. & Weber, K. (1963) Biochem. 8. 237, 24-34. Steiner, R. F. & Laki, K. (1951) Arch. Biochem. Biophys.
Ackers, G. K. (1964) in Molecular Sieve Processes,
Martin, R. G. & Ames, B. N. (1061) J . Bwl. Chem. 236,
Huxley, H . E. (1963) J . Mol. BioZ. 7, 281-308. Holtzer, A. & Rice, S. A. (1957) J . Am. Chem. 800. 79,
Meunier, J . C., Olsen, R. W. & Changeux, J. P. (1972)
Edelstein, S. J. & Schachman, H. K. (1967) J . Biol.
Weast, R. C. (1972) Handbook of Chemistry and Physics,
441 -455.
34, 24-37.
V O ~ . 3, pp. 723-730.
1372 - 1379.
4847 -4851.
FEBS Lett. 24, 63-68.
Chem. 242, 306-311.
The Chemical Rubber Co., Cleveland, Ohio.
S. Bon, F. Rieger et J. Massoulib Institut de Biologie Physico-Chimique 13 Rue Pierre-et-Marie-Curie, F-75005 Paris, France


![Time-Periodic Solutions to the Full Navier–Stokes–Fourier ...pbmucha/publ/2012/fmnp.pdfNavier–Stokes–Fourier system developed in the first part of the monograph [4], although](https://img.pdfslide.fr/doc/110x75/61360c4c0ad5d2067647c48a/time-periodic-solutions-to-the-full-navierastokesafourier-pbmuchapubl2012fmnppdf.jpg)