Embed Size (px)
Citation preview

BIOCHIMIE, 1973, 55, 1137-1151.
Recherches biochimiques sur l'oogen se. 7. Synth~se et maturation du RNA 5 S dans les petits oocytes de Xenopus laevis.
H e r m a n DENIS et M a u r i c e WE6NEZ ( ' ) .
Laboratoire de Biochimie, Facult~ des Sciences, Universit~ de Liege, Place Delcour, 17, B-4000 Liege, Belgique.
(5-6-1973).
S u m m a r y . - As shown previously, 5 S RNA synthesized by small oocytes of Xenopus :aevis is stored for several months before being incorporated into the r ibosomes. In this paper we compare the propert ies of newly - fo rmed 5 S RNA with those of 5 S RNA stored in the oocyte. Ne'wly-synthesized 5 S RNA and bulk 5 S RNA differ both by the i r electro- phoret ic m.obility and by the i r chromatographic behaviour .
We propose the fol lowing scheme for pos t - t ranscr ip t ional processing of 5 S RNA in oocytes. 5 S RNA first appears as a precursor (form A) containing more than 135 nu- cleotides. The precursor is then quickly shor tened at the 3'-end and gives rise to a second form (B) conta ining 80 p. cent molecules wi th 120 nucleotides and 20 p. cent wi th 121 or 122 nucleotides. Form B is very slowly converted into the storage fo rm (C) of 5 S RNA. Most molecules of form C contain 120 nueteotides, but 15 to 2{) p. cent of them contain 119 or 118 nucleotides. In somatic ceils, ~'e lind no indicat ion of a long- term modification s imi lar to tha t observed in oocytes.
The scheme out l ined above is supported by the fol lowing observations. (1) 32P-labelled 5 S RNA contains a small propor t ion of mo]eeules which are longer
than bulk 5 S RNA by at least 15 nucleotides. These ext ra-nueleot ides are located at the 3'-end of the molecule.
(2) Newly- fo rmed 5 S RNA (form B) is s l ight ly longer at the 3'-end than bulk 5 S RNA. This could be demons t ra ted by measur ing the e leet rophoret ie mobi l i ty of the 3 ' - terminal f r agmen t (nucleotides 90-120) obtained a f te r par t ia l hydro lys i s wi th T~ ribonuetease. After excision of the nucleot ides tha t fol low res idue nr 118 by means of snake venom phospho- diesterase the chromatographic behav iour of newly-synthes ized 5 S RNA becomes iden- lical to tha t of bulk 5 S RNA.
(3) Three different 3 ' - terminal ol igonucleotides were detected in 32P-labelled 5 S RNA (CpUpU, CpUpUpU and CpUpUpUpU) a f te r hydro lys is wi th T, r ibonuelease. CpUpU is the most f r equen t t e rmina l oligonueleotide. As m a n y as 8 different 3'-ends were observed in bulk 5 S RNA. The most f requen t one is CpUpU, as in near ly- formed 5 S RNA (63 p. cent of all ends). The CpUpUpU and CpUpUpUpU ends occur in very few molecules (3 p. cent). Two 3'-ends (C.pU and C : 16 p. cent of all ends) correspond to molecules shor ter by 1
to 4 nueleotides than newly -made 5 S RNA. The remain ing 3 ends (Gp~pApApU, Gp~pApV and CpGpU : 18 p. cent of all ends) were detected only in bulk 5 S RNA. These cannot
derive f rom the 3'-ends observed in newly - fo rmed 5 S RNA by a mere loss of t e rmina l nueleotides.
(4) After labelling wi th 32p, the ur idyl ie residues of the 3 ' - terminal ol igonueleotides (CpUpU, CpUpUpU and CpUpUpUpU) a lways have a higher specitie act ivi ty than the cyt idyl ie residue. This suggests a t u rnove r of the 3'-end ~vhieh migh t he related to the very slow shortening tha t 5 S RNA undergoes during its storage in the oocyte.
Abrdoiations : SDS : dod6cylsulfate de sodium. tRNA : RNA de t ransfer t . A : acide ad~nylique. C : cyt id ine ou acide cyt idyl ique. C ' : hydroxym6thy le -d iOhyl~ne-g lyco i cytosine. G : aeide guanyl ique. G! : acide guanyl ique 2', 3' eyelique. U : ur id ine ou acide ur idyl ique. U ' : hydroxymdthyle -d i~ thy l6ne-g lyeo l uracile. Pour reprdsenter la s6quence des oligonucl~otides
t e rminaux du RNA, nous ut i l isons la no ta t ion : CpUpU. Pour les oligonucldotides int6rieurs et la s6quence compl6te, nous uti l isons la nota t ion : UUAG etc.
( ' ) Adresse actuel le des auteurs : Centre de G6n6ti- que mol6eulaire, CNRS, F-91190 Gif-sur-Yvette, France.
I N T R O D U C T I O N .
Au c o u r s de son a c c r o i s s e m e n t , l ' o o c y t e de
x 6 n o p e a c c u m u l e 30.0.00.0 fo is p l u s de RNA q u ' u n e
cc l lu le s o m a t i q u e . L ' a c c u m u l a t i o n d u RNA 28 S et 18 S est r e n d u e p o s s i b l e p a r u n e a m p l i f i c a t i o n
des g6nes c o r r e s p o n d a n t s [1, 2, 3]. E n r e v a n c h e ,
les gbnes o r g a n i s a t e u r s du RNA 5 S e t d u tRNA ne s o n t pas ampl i f i6s [3, 4}. B ien qu ' i l p o s s ~ d e b e a u c o u p m o i n s de g6nes 5 S que de g~nes 28 S
et 18 S, l ' o o c y t e a c c u m u l e a u t a n t de mo16eules 5 S
que de m o l 6 c u l e s 28 S et 18 S. I I s ' e n s u i t que 1¢~

1138 H. D e n i s el M. W e g n e z .
g~nes 5 S doivent p rodui re plus de mol6cules de BNA que les gSnes 28 S et 18 S. Geci explique pourquoi les gSnes 5 S sont actifs pendan t route la dur6e de l'oogen~se, alors que Ies gbnes 28 S e t 18 S ne le sont que pendan t la part ie terminale de ce processus (vitellogenbse ou grand aceroisse- m e n t ; r6f. [5, 6]). Par ailleurs, les oocytes aug- mentent leur p roduc t ion de RNA 5 S par rappor t aux cellules somatiques en expr imant la totalit6, ou tout au moins la majeure par t ie des gbnes 5 S qu'ils possbdent, alors que les cellules somatiques n 'ut i l isent qu'une par t ie de ces g/rues [7, 8].
Pendan t la phase init iale de l 'oogenbse (petit accroissement) , l 'oocyte synth6tise surtout du RNA 5 S e t du tRNA [5, 6]. I1 faut at tendre plu- sieurs mois pour que le RNA 5 S produi t en excbs durant la premibre phase de l 'oogenbse soit int6gr6 dans les ribosomes [9]. Le RNA 5 S reste done en attente pendan t un laps de temps consid6rable. Durant cette p6riode, le RNA 5 S se trouve en part ie clans le suc cellulaire et en par t ie dans des part icules nucl6oprot6iques pr6sentant un coeffi- cient de s6dimentat ion de 42 S [6, 10]. On peut se demander si le RNA 5 S subit des modificat ions pendant son s6jour prolong6 dans l 'oocyte. Pour r6pondre h cette question, nous avons compar6 les propri6t6s du RNA 5 S stock6 dans l 'oocyte avec celles du RNA 5 S nouvellement synth6tis6.
MATERIEL ET METHODES.
Marquage et puri f icat ion du RNA 5 S,
Le RNA 5 S d 'oocyte est extrait h par t i r d'ovai- res entiers de femelles immatures, mesurant de 2 h 3 cm. Ces ovaires ne cont iennent que des oocy- tes en peti t accroissement et de nombreuses cel- lules somatiques, dont le contenu en RNA est n6gli- geable par rappor t h celui des oocytes. Pour les marquages de courte dur6e (2 heures h 3 jours), les ovaires sont incub6s dans du Ringer ou dans du mil ieu F 12 [11], addi t ionn6 d 'ur id ine [ ~ J (26 Ci /mmol ; 1,00 t, Ci/ml), de guanosine [atI] (3,5 Ci /mmol ; 10,0 I, Ci/ml) ou de a,2p (,1 mCi/ml) . Pour les marquages de longue dur6e, on in]ecte le pr6curseur dans la cavit6 abdominale de la femelle. Le RNA est extrait par Ia m6thode au SDS-ph6nol froid, puts purifi6 par fil tration sur colonne de Sephadex G-10J0 [5] ou par 6Iectro- phorbse en gel de polyacrylamide . Dans quelques cas, le contenu des oocytes a 6t6 centrifug6 en gradient de saccharose, de manibre h s6parer les part icules 42 S du suc cellulaire [10]. Le RNA 5 S
BIOCHIMIE, 1973, 55, n ° 9.
contenu dans ces deux compart iments cellulaires est purifi6 comme d6crit ci-dessous.
Analyse du RNA 5 S.
Pour l 'analyse de la s tructure p r imai re du RNA 5 S, nous avons utilis6 la technique de BrownIee el al. [12]. Les proc6d6s employ6s pour l '61ectrophor5se en gel de poIyacry lamide h 12,3 p. cent et la chromatographie sur colonne d 'a lbumine in6thyl6e- Kieselguhr ont d6jh 6t6 d6crits [13].
Nous avons ~tudi6 la structure secondaire du RNA 5 S e n soumettant celui-ci h une hydrolyse mod6r6e par la r ibonucl6ase T 1 et en analysant les fragments de mol6cule obtenus par 2, 61ectropho- r6.ses successives en gel de po lyacry lamide El4]. Le RNA est dissous dans 1 ml de NaC1 0,2 M, MgCI u 0,02 M, Tris 0,05 M pH 7,5, puts dig6r6 h 5°C au moyen de 5 unit6s de r ibonucl6ase T 1 par mg de RNA. Le RNA est ensuite soumis h l'61ectro- phorSse pendan t 15 heures ~ 5°C dans un bloc de po lyacry lamide h 12,3 p. cent [15!. Le RNA est localis6 dans le gel soit par autoradiographie si le marquage a 6t6 fair au a:,p, soit par colorat ion d'une aliquote migrant en parallble, si Ie marquage a 6t6 fait au tr i t imn. On r6cup~re Ie RNA et pro- cSde h une seconde 61ectrophorSse en pr6sence d'ur6e 7 M [14]. Les coupures introdui les dans le RNA par la ribonucl6ase, qui n 'appara issa ient pas lors de la premibre 61ectrophor~se parce que ta mol6cule conservai t sa configuration native, sont r6v616es par la seconde 61ectrophorbse [14]. Les fragments de mol6cule sont mis en 6vidence par autoradiographie ou par colorat ion au moyen de py ron ine [10].
D~termination de l'extr~mit~ 3' du RNA 5 S.
Pour identifier le nucl6otide 3 ' - t e r m i n a l du R,NA 5 S non marqu6, on emploie la m6thode de Hatlen et ah [16]. Deux mg de RNA 5 S purifi6 sont hydrolys6s au moyen de KOH 0,3 N pendan t 15 heures h 37°C, puts fix6s sur une colonne de Dowex 1 × 8 (forme formiate ; 3 x 1 cm). On passe sur la colonne 20 ml d 'acide formique 5 raM, ce qui 61ue le nucl6oside 3'-terminal. Celui- ci est concentr6 par 6vaporation, pu ts identifi6 par chromatographie sur papier Whatman n ° 1 dans Ie systbme eau-6thanoI-(NH4)2SO 4 propos8 par Lane [17].
I, 'extr6mit6 3' du RNA 5 S stock6 dans l 'oocyte est marqu6e par le proc(~d6 de RajBhandary [183 et de De Wachter et Fiers [19]. I1 s'agit de rendre radioact i f le nucleotide situ6 h l 'extr6mit6 3' de !a mol6cule par oxydat ion au moyen de periodate,

Recherches biochimiques sur l'oogenbse (7). 1139
puts pa r rdduc t ion au m o y e n de b o r o h y d r u r e [ZH]. Nous avons suiv i h la let t re le proc6dd de De W a c h t e r et Fiers . La rdduc t ion finale est fai te pa r du b o r o h y d r u r e de po ta s s ium (20 C i / m m o l ; CEA, Saclay) . Une no.uvelle pu r i f i ca t ion du RNA est n dcessaire aprbs le marquage . Nous avons sou- mis le RNA success ivement h une f i l t ra t ion sur co lonne de Sephadex G-100 et h 2 61ectrophorbses en gel de p o l y a c r y l a m i d e , d ' a b o r d en l ' absence , puts en p rdsence d 'urde 7 M. Le RNA est finale- men t 61u6 du gel au m o y e n de NaC1 0,3 M.
P o u r iden t i f i e r le ou les o l igonuel6ot ides qui po r t en t le m a r q u a g e t e rmina l , on h y d r o l y s e le RNA au m o y e n de r ibonuc l6ase T 1 ou de r ibo- nucl6ase A, puts on ana lyse l ' h y d r o l y s a t p a r chro- m a t o g r a p h i e sur co lonne de DEAE-cel lulose ou p a r f i l t ra t ion sur co lonne de Sephadex G-25. Le p r e m i e r procdd6 s6pare Ies o l igonueldot ides d 'aprbs leur charge et l eur compos i t ion , t and i s que le second les sdpare d 'apr~s leur tai l le , leur compos i t ion et leur charge [201. La DEAE-cel lu- lose (Macherey, Nagel el Co) est ca l ibrde et lavde avec du NaCt 4 M, puts tassde dans une co lonne de c h r o m a t o g r a p h i e (20 × 1 era) et enfin dqui l ibrde dans du t a m p o n Tris-HC1 2~0 mM pH 7,5, urde 7 M [21]. L ' h y d r o l y s a t est appl iqud sur la co lonne dans le m6me tampon , puts dlud p a r un g rad ien t l inda i re de NaC1 0 h 0,3 M dans l 'urde 7 M. On me- sure Ia densi td op t ique h 26:0 nm de chaque frac- t ion recue i l l ie h la sor t ie de la co lonne et la radio- activitd d 'une a l iquote aprbs y avoi r ajoutd 10 vo- lumes d ' Ins ta-Gel (Packard) . Pou r la f i l t ra t ion sur S e p h a d e x G-25, on ut i l ise une co lonne de 125 × 1,5 era. L'61ution est assur6e p a r un flux cons tan t (5 ml pa r heure) de t a m p o n acide ac6t ique-acdtate de sod ium 10 mM p H 5,0, NaN 3 0,02 p. cent.
Le marquage p a r le b o r o h y d r u r e modif ie le nucldot ide 3 ' - terminal . Aprbs une h y d r o l y s e com- p lOe pa r le KOH, on obt ien t le ddrivd h y d r o x y - m6thy le -d id thy lbne-g lyco l de la base te rmina le . Celni-ci est ident if i6 p a r c h r o m a t o g r a p h i c sur p a p i e r W h a t m a n n ° 1 [17], en p rdsence de nucl6o- s ides modif ids p a r o x y d a t i o n au m o y e n de per io- date, puts p a r rdduc t ion au m o y e n de bo rohy- d ru re [19].
RESU.LTATS.
Mise en ~vidence d'une forme << longue>> du RNA 5 S d'oocyte.
Quand on soumet h l 'd lec t rophor6se en gel de p o l y a c r y l a m i d e h 12,3 p. cent le RNA d 'ova i res marquds au z2p p e n d a n t 2 h 3 jours, on ob t ien t
BIOCHIM1E, 1973, 55, n ° 9.
2 bandes p r i nc ipa l e s , c o r r e s p o n d a n t au RNA 5 S et au tRNA (fig. 1). P lus i eu r s bandes rn ineures s ' i n t e r ca l en t ent re la l igne de ddpar t et le RNA 5 S. L 'une d ' en t re elIes (marqude 2 sur la fig. 1) pr6- sente, apr6s h y d r o l y s e p a r la r ibonuc l~ase T1, le m~me << f i n g e r p r i n t >> que le RNA 5 S d 'oocy te (fig. 2 et Tableau I). Toutefois , les o l igonucldot ides 3 ' - t e rminaux (X1, X 2 et X 3) sont absents . Il y a p a r a i l leurs un o l igonucldot ide nouveau (Z), don t nous n ' avons pas pu dd t e rmine r la sdquence complbte, faute de matdr ie l ( tableau I). Cet o l igonucl6ot ide p ro longe de toute 6vidence le RNA 5 S du cStd 3'. En plus de l 'o l igonucl6ot ide Z, le RNA de la bande 2 fourn i t un AG et un UG supp ldmen ta i r e s ( tableau I). La t ache 5 (CAG) est r en fo rcde et i l exis te 2 l aches non numdrotdes, dont ]a molar i t6 est i n f6 r i eu re h 0,5. I1 est diff ici le de d i re si les o l igonucl6ot ides c o r r e s p o n d a n t s p r o v i e n n e n t de c o n t a m i n a n t s ou sont le reflet d 'une hdtdrog6ndit~ du RNA con tenu dans la h a n d e 2 (fig. 1).
FI6. I. - - Electrophor$se en gel de polyacrylamide 12,3 p. cent du RNA extrait d'ovaires entiers de femel- les immatures, marquds au a2p pendant 3 jours. La figure reprdsente le clich~ obtenu en appliquant un film de radiographic eontre le gel pendant 5 heures. Environ 100 /xg de RNA ont ~td soumis /t I'dlectropho- r~se h 5°C pendant 20 heures sous i0 volts/cm.
Tout ceei m o n t r e que la b a n d e 2 (fig. 1) corres- p o n d /t une forme << longue >) du RNA 5 S, conte- nan t au moins 135 nucldot ides . I1 p o u r r a i t s 'agi r d 'un p r d c u r s e u r du RNA 5 S, qui se ra i t recoupd
1140 H. D e n i s et M. W e g n e z .
p o u r d o n n e r na i s s anee h la fo rme de la mol6cu le qui con t i en t 129 nuc l6ot ides . Dans les c o n d i t i o n s hab i tue l les de marquage , la fo rme ¢ l o n g u e a r ep r6sen t e h peu prbs 1 p. cen t du R,NA 5 S total. Nous n ' avons pas jusqu'~ p r6 sen t t rouv6 le m o y e n
con t i en t 120 h 122 nuc l6o t ides (bande 3 de la fig. 1). Nous savons d6jh que le RNA 5 S n6oform6 est t r i p h o s p h o r y l 6 h l ' ex t r6mit6 5' et qu ' i l p e r d peu h peu ses g roupes p h o s p h a t e s p e n d a n t son s6jour p ro long6 dans l ' oocy te [26]. Le RNA 5 S
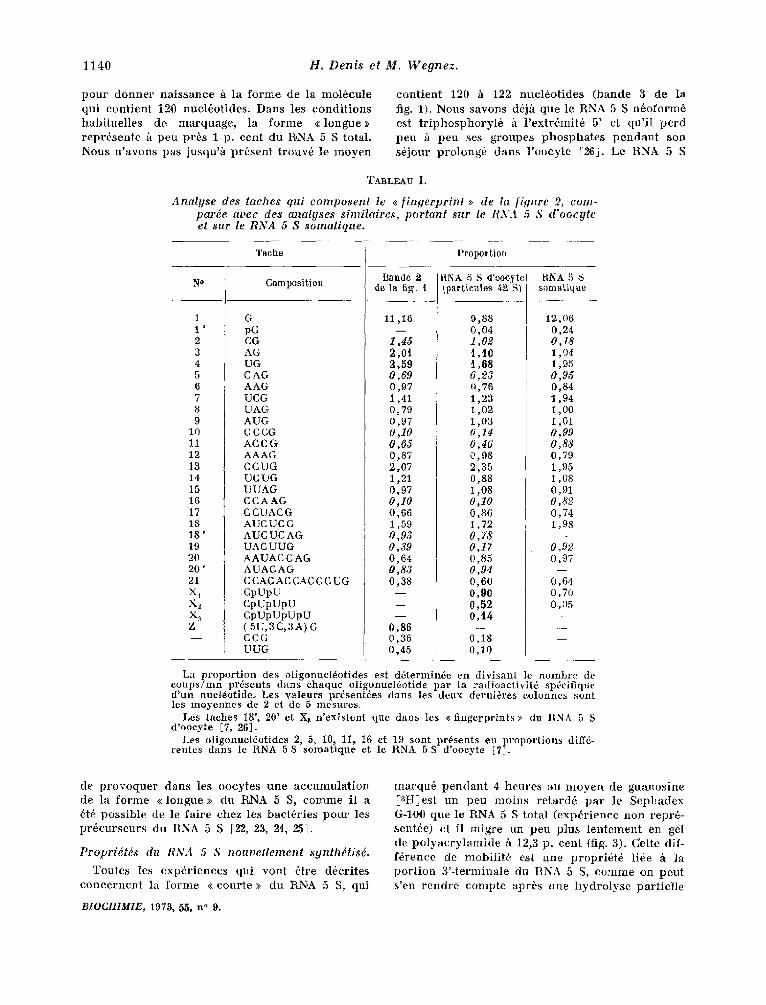
TABLEAU I.
Analyse des taches qui composenl le ~ f ingerprint ~> de Ia figure 2, com- parde avec des analyses similaires, portant sur le RNA 5 S d'oocyte et sar le RNA 5 S somatique.
Tache Proportion
Bande 2 I/NA 5 S d'oocyte RNA 5 S N ° Composition de la fig. I (partieules 42 S) somatique
1 1 ' 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 18' 19 20 20' 21 Xt X~ X3 Z
G pG CG AG UG CAG AAG UCG UAG AUG C CCG ACC G AAAG CCUG UC UG UUAG CCAAG CCUACG AUCUCG AUC UC AG UA C UUG AAUAC C AG AUACAG C CA(: AC CAC C C U G CpUpU CpUpUpU CpUpUpUpU (5U,3C,3A) G CCG UUG
11,16
1,45 2,01 2,59 0,69 0,97 1,41 0,79 0,97 0,10 0,65 0,87 2,07 1,21 0,97 0,10 0,66 1,59 0,93 0,39 0,64 0,83 0,38
0,86 0,36 0,45
9,88 0,04 1,02 i , i0 t ,68 0,23 0,76 1,23 1,02 1,03 0,14 0,46 0,98 2,35 0,88 1,08 0,10 0,86 1,72 O, 78 0,17 0,85 0,94 0,60 0,90 0,52 0,14
0,18 0,10
12,06 0,24 0,18 1,04 1,95 0,95 0,84 1,94 1,00 1,01 0,99 0,88 0,79 1,95 1,08 0,91 0,82 0,74 1,98
0,92 0,97
0,64 0,70 0,05
La proportion des oligonuel6otides est d6termin6e en divisant le nombre de coups/ran pr6sents darts chaque oligonucl6otide par la radioactivit6 sp6eifique d'un nucl6otide. Les valeurs prdsent~es dans les deux derni~res colonnes sont les moyennes de 2 et de 5 mesures.
Les taehes 18', 20' et Xs n'existent que darts les << fingerprints >> du RNA 5 S d'oocyte [7, 26].
Les oligonuel6otides 2, 5, 10, 11, 16 et 19 sont pr6sents eu proportions diff6- rentes darts le RNA 5 S somatique et le RNA 5 S d'ooeyte [7J.
de p r o v o q u e r dans les oocytes une a c c u m u l a t i o n de Ia fo rme ¢longue>> du RNA 5 S, c o m m e il a 6t6 poss ib le de le fa i re chez les bact6r ies p o u r les p r 6 c u r s e u r s du RNA 5 S [22, 23, 24, 25].
Propridtds du RNA 5 S nouvelIement synthHisd.
Toutes les exp6r i ences qui vont 6tre d6cr i tes c o n c e r n e n t la f o r m e (< cour t e >> du RNA 5 S, qui
BIOCHIMIE, 1973, 55, n ° 9.
m a r q u 6 p e n d a n t 4 heu re s au m o y e n de guanos ine [!artiest un peu m o i n s r e t a rd6 pa r le Sephadex G-10@ que le RNA 5 S total ( exp6r ience non repr6- sent6e) et il m i g r e un peu plus l e n t e m e n t en gel de p o l y a c r y l a m i d e h 12,3 p. cen t (fig. 3). Cette dif- f6 rence de mobi l i t6 est une p ropr i6 t6 li6e h ]a p o r t i o n 3 ' - t e rmina le du RNA 5 S, c o m m e on peut s 'en r e n d r e compte apr~s une h y d r o l y s e par t i e l l e
R e c h e r c h e s b i o c h i m i q u e s sur l 'oogendse (7). 1141
du RNA 5 S par la r i bonuc I6as t Tt, suivie d 'une s6para t ion des f ragments obtenus par 2 61ectro- phor6ses successives en gel de p o l y a c r y l a m i d e (fig. 10).
Le RNA 5 S n o u v e l l e m e n t synth6t is6 est 61u6 de l ' a l bumine m6thyl6e apr6s le R:NA 5 S total (fig. 4). Dans les par t i cu les 42 S, le d6calage ent re le profi l de densi t6 op t ique et I t profil de rad ioac t iv i t6 ne d i spara l t que tr6s l e n t e m e n t : il t'aut a t t end re 1 mois pou r qu ' i l s ' e s tompe tout ~ fair (fig. 4). Le
RNA par la cha leur , ni l ' en l6vemen t des groupes phospha t e s 5 ' - t e rminaux pa r Ia phospha t a se alca- l ine ne s u p p r i m e n t le d6calage. La d i f f6rence de c o m p o r t e m e n t c h r o m a t o g r a p h i q u e en t re le RNA 5 S n6oform6 et le R NA 5 S total ne s 'observe que clans l 'oocyte : le RNA 5 S de rate marqu6 pen- dant 5 heures ~ 2 jours est 61u6 de l ' a lbumine m6- thyl6e en mSme temps que Ie RNA 5 S de foie (fig. 6).
Ces p r e m i e r s r6sultats p e u v e n t s ' i n t e rp r6 te r de la man i6 re suivante. Le RNA 5 S n6oform~ serai t 16g6rement plus long h l 'ext r6mit6 3' que le RNA 5 S stock6 dans l 'oocyte , c o m m e l ' i n d i q u e n t sa mobi l i t$ plus faible en gel de p o l y a c r y ] a m i d e (fig. 3 et 10) et son 61ution plus r ap ide du Sepha- dex G-100. Chez Escherichia colt, on sait que l ' a l bumine m6thyl6e re t ien t m o i n s f e r m e m e n t le RNA 5 S total que le RNA 5 S n o u v e l l e m e n t syn- th6tis6, qui poss6de 1 h 3 nuc l6o t ides en plus [23]. Ces nucl~ol ides se t rouven t h l ' ex t r6mit6 d ' une ¢ tige >) bif i laire pu i sque l 'ext r6mit6 5' du RNA 5 S est appar i6e ~ l 'extr6mit6 3' [27]. La pr6sence d 'un bout uni f i la i re p r o l o n g e a n t une <(tige)) bif i laire e n t r a i n e donc une plus forte r6 ten l ion du RNA 5 S par l ' a l bumine m6thyl6e. Nous pensons que le RNA 5 S de x6nope s t c o m p o r t e de la m~me man i6 re que celui d'Escherichia colt. Pour prou- ver qu ' i l e n e s t b ien ainsi, nous devons : 1) mon- t rer que l 'extr6mit6 5' du RNA 5 S est appar i6e h l 'extr6mit6 3' ; 2) m o n t r e r que le r a c c o u r c i s s e m e n t de la mol6cule h l 'ext r6mit6 3' s u p p r i m e le d6ca- lage ent re les profils de densi t6 op t ique et de rad ioac t iv i t6 observ6 sur les fig. 4 et 5 ; 3) iden-
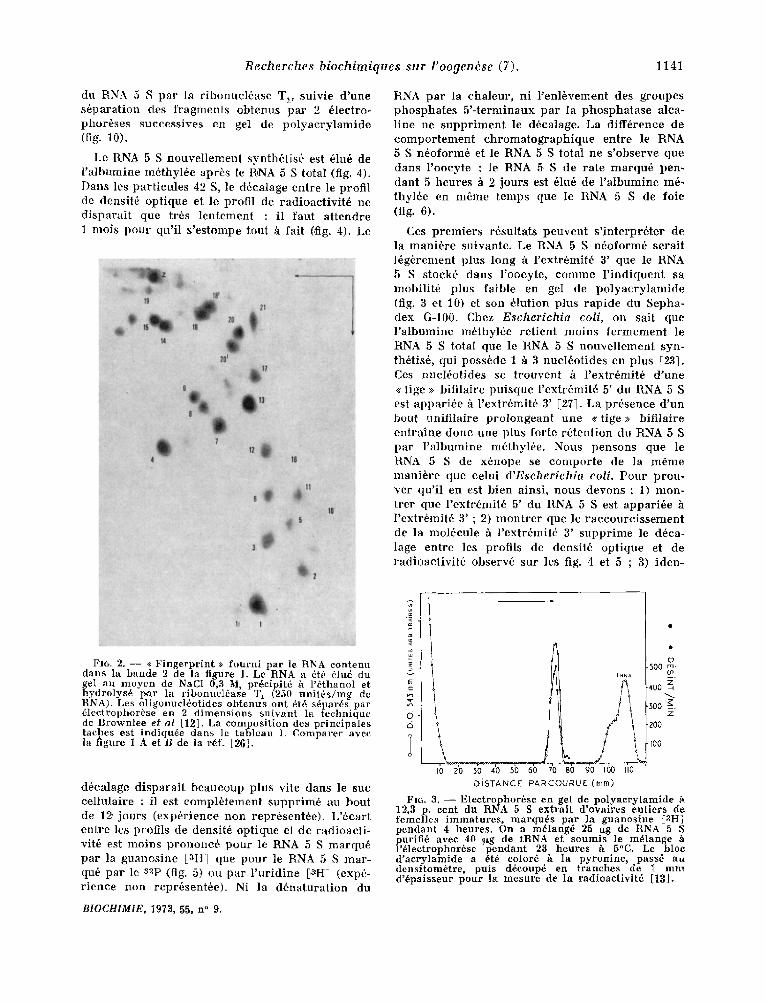
Fro. 2. - - <( Fingerprint >> fourni par le RNA contenu dans la bande 2 de la figure 1. Le RNA a 6t6 5lud du gel au moyen de NaC1 0,3 M, pr6cipit6 h l'~thanol et hydrolys6 par la ribonucl6ase T, (250 unit6s/mg de RNA). Les oligonuel6otides obtenus ont 6t6 s6par6s par 61ectrophor6se en 2 dimer~sions suivant la technique de Brownlee et aI [12]. La composition des principales taches est indiqu6e dans le tableau I. Comparer avec la figure 1 A et B de la rdf. [2fi].
d6calage d i spara i t beaucoup plus vite dans le suc cel lula i re : il est compl6 t emen t suppr im6 au bout de 12 jours (exp6r ience non repr6sent6e) . L '6cart en t re les profils de densi t6 op t ique et de rad ioac t i - vit6 est mo ins p r o n o n c 6 p o u r le RNA 5 S marqu6 par la guanos ine [ZHI que pour le RNA 5 S mar - qu6 pa r le 32p (fig. 5) ou par l ' u r i d ine EZH] (exp6- r i ence non repr6sent6e) . Ni la d6na tu ra t ion du
BIOCHIMIE, 1973, 55, n ° 9.
[ tRNA .500 m
l// I t .,oo
i t / i" I0 20 50 40 50 60 70 80 90 I00 I10
DJSTANCE PARCOURUE (ram)
FIG. 3. - - Electrophor~se en gel de polyacrylamide ~. 12,3 p. cent du RNA 5 S extrait d'ovaires critters de femelles immatures, marqu6s par l a guanosine [aH] pendant 4 heures. On a m61ang6 25 ug de RNA 5 S purifi6 avee 40 t~xg de tRNA et soumis le m@lange l'61ectrophor6se pendant 23 heures h 5°C. Le bloc d'acrylamide a 6t6 color6 h la pyronine, pass6 au densitom6tre, puts d6eoup6 en tranches de I mm d'6paisseur pour la mesure de la radioactivit6 [13].
1142 H, Denis et M. Wegnez.
0,3l A 4 hcur¢s
f J J'L 1 t
10 20 30 40 o ,r..;,
6
0.4.
0,3 ̧
0,2
0,1 ̧
C 12 j o u r s
f --==ec,_ e~e-,1 _~_: ccc _~,e _ , ~
lO
! NoCt M
1,0
• 0.5
,
20 50 40 TUBE N °
0,
3OO
0
200
0. 100
.1000
) , 4 I
• 750
).5
.500 0,2.
- 250 O,H
£i 20 hcures
NoCL
i l l NCM, I.0
0,5
I0 20 50 40
D I mois
10 20 30 40 TUBE N °
3000
-200O
-1000
_! WI. U~
--t
-400 g Z
-300
-200
!00
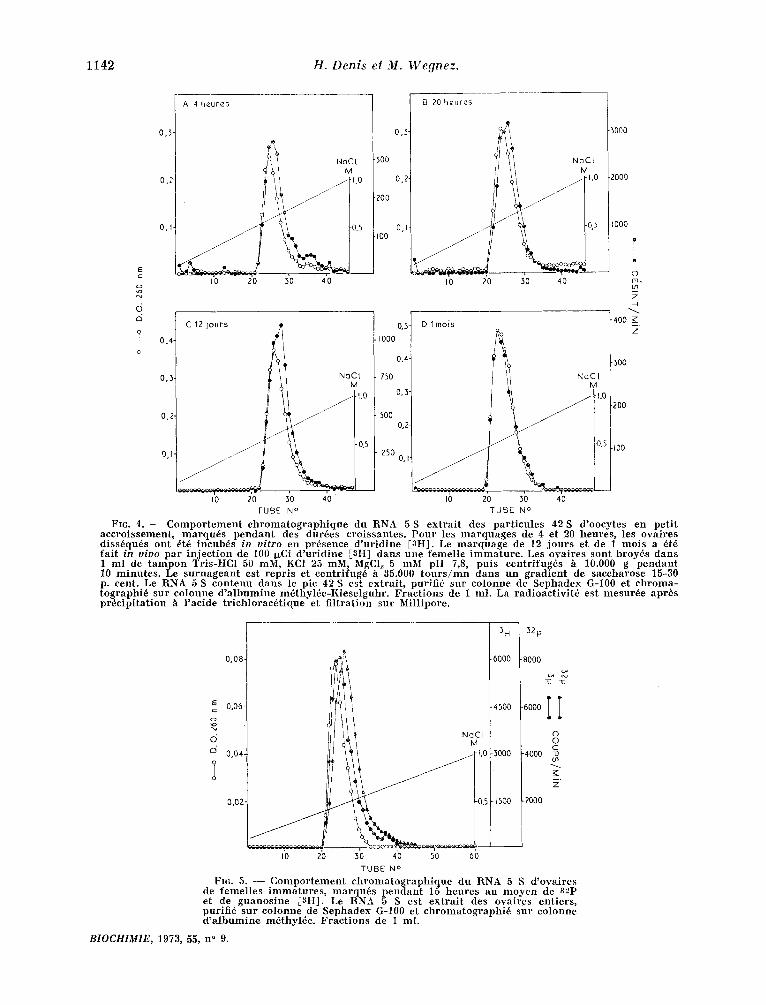
FIG. 4 . - Co m p o r t e m e n t c h r o m a t o g r a p h i q u e du RNA 5 S ex tra i t des par t i cu le s 42 S d 'oocytes en pet i t accro i s sement , m a r q u i s p e n d a n t des dur6es cro i s santes . P o u r les m a r q u a g e s de 4 et 20 heures , les ova ires d i s s6qu6s out ~t6 incub6s in vi tro en presence d 'ur id ine [3HI. Le m a r q u a g e de 12 j o u r s et de 1 m o i s a 6t6
r3 fa i t in vivo par i n j e c t i o n de 100 i~Ci d 'ur id ine , HI darts une f e m e l i e i m m a t u r e . Les ova ires sont broy6s dans 1 ml de t a m p o n Tris -HCl 50 mM, KC] 25 mM, MgCl~ 5 mM pH 7,8, pu i s centr i fugSs & 10.000 g pendant 70 m i n u t e s . Le s u r n a g e a n t est repris et centr i fug6 h 35.000 t o u r s / m n dans un grad ient de saccharose 15-30 p. cent. Le RNA 5 S c o n t e n u dans le pic 42 S est extrai t , p,urifi6 sur co lonne de S e p h a d e x G-100 et c h r o m a - tographi6 sur c o l o n n e d ' a l b u m i n e m~thy lde -Kiese lguhr . Frac t ions de 1 ml. La rad ioac t iv i t6 est mesur6e apr$s pr6c ip i ta t ion & l 'acide t r i ch lorac6t ique et f i l trat ion sur Mil l ipore.
0.08. i 6oo~[8ooo ~
~ 0,06- 450C 6000 [ I
0,0 4 ,I0 4000
I I _
10 20 30 4'0 50 60 TUBE N °
FI6. 5. - - C o m p o r t e m e n t c h r o m a t o g r a p h i q u e du RNA 5 S d 'ovaires de f e m e l l e s i m m a t u r e s , m a r q u 6 s p e n d a n t 15 heures au m o y e n de a2p et de g u a n o s i n e [3H]. Le RNA 5 S est ex tra i t des ova ircs ent iers , purif i6 sur c o l o n n e de S e p h a d e x G-100 et c h r o m a t o g r a p h i 6 sur eo lonne d ' a l b u m i n e mdthyl6e . F r a c t i o n s de 1 ml .
BIOCHIMIE, 1973, 55, n ° 9.
R e c h e r c h e s b i o c h i m i q u e s sur l 'oogendse (7) . 1143
tifier l ' ext r6mit6 3' du RNA 5 S total et du RNA 5 S n6oform6. Nous al lons r e p r e n d r e ces 3 po in t s s6pa ramen t .
Structure secondaire du RNA 5 S.
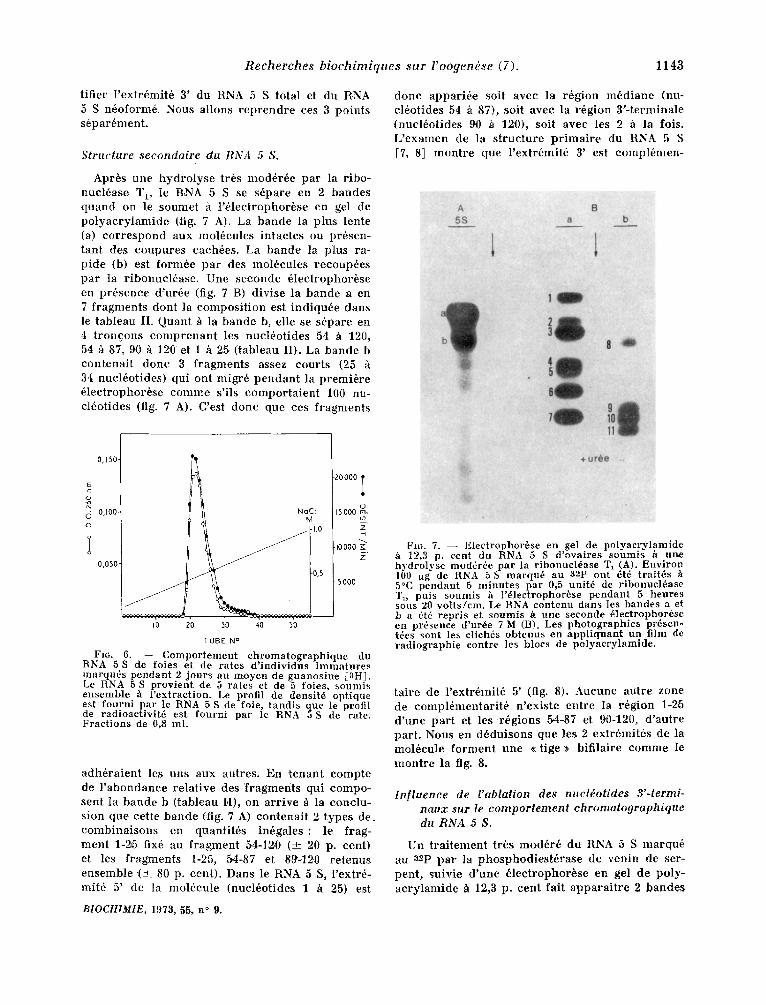
d o n e appar i6e soit avec la r6gion m 6 d i a n e (nu- cl6ot ides 54 h 87), soit avec la r6gion 3 ' - t e rmina le (nuc l6ot ides 90 h 120), soit avec les 2 ~ la fois. L ' e x a m e n de la s t r uc tu r e p r i m a i r e du RNA 5 S [7, 8] m o n t r e que l ' ex t r6mit6 3' est compl6men-
Apr6s une h y d r o l y s e tr6s mod6r6e pa r la r ibo- nucI6ase Tt, Ie R, NA 5 S se s6pare en 2 bandes q u a n d on le soumet h l '61ectrophor6se en gel de p o l y a c r y l a m i d e (fig. 7 A). La b a n d e la plus lente (a) c o r r e s p o n d aux mol6cu les in tac tes ou pr6sen- tant des c o u p u r e s cach6es. La b a n d e la p lus ra- p ide (b) est form6e p a r des mol6cules r ecoup6es pa r la r ibonuc l6ase . Une s econde 61ectrophor/~se en p r6sence d 'ur6e (fig. 7 B) d ivise la b a n d e a en 7 f r agmen t s don t la c o m p o s i t i o n est i nd iqu6e dans le t ab leau II. Quant h la b a n d e b, elle se s6pare en 4 t r o n c o n s c o m p r e n a n t les nuc l6o t ides 54 h 120, 54 h 87, 90 h 120' et I h 25 ( tableau II). La b a n d e b con t ena i t d o n e 3 f r agmen t s assez cour ts (25 34 nucl6ot ides) qui ont migr6 p e n d a n t la p r e m i 6 r e 61ectrophor6se c o m m e s'ils c o m p o r t a i e n t 100 nn- c l6ot ides (fig. 7 A). C'est d o n e que ces fragunents
0 , 1 5 0 .
o
6 o.loo,
~5
0.050-
L0
• N o C I
r MIOI. 5
20 50 40 50
T U B E N °
-20000 I
5 000 m. o3 _. z -4
~0000
5000
Fro. 6. - - Comportement chromatographique du RNA 5 S de foies et de rates d' individus immatures marqu6s pendant 2 jours au m.oyen de guanosine [3HI. Le RNA 5 S provient de 5 rates et de 5 foies, soumi,; ensemble h 1'extraction. Le profil de densit6 optique est fourni par le RNA 5 S de foie, tandis que le profil de radioactivit6 est fourni par le RNA 5 S de rate. Fractions de 0,8 ml.
a d h 6 r a i e n t les uns aux autres. En t e n a n t compte de l ' a b o n d a n c e re la t ive des f r a g m e n t s qui compo- sent la b a n d e b ( tableau I,I), on a r r ive h la conc lu - sion que cet te b a n d e (.fig. 7 A) con t ena i t 2 types de. c o m b i n a i s o n s en quant i t6s in6gales : le frag- men t 1-25 fix6 au f r a g m e n t 54-120 (--4-_ 20 p. cent) et les f r agmen t s 1-25, 5.4-87 et 89-12.0 r e t e n u s ensemble ( ± 80 p. cent ) . Dans le RNA 5 S, l 'extr6- rnit6 5' de Ia mol6cu le (nuc16otides 1 h 25) est
Fro. 7. - - Electrophor6se en gel de polyacrvlamide h 12,3 p. cent dn RNA 5 S d'ovaires soumis h urm hydrolyse mod6r6e par la ribonuel6ase T1 (A). Environ 100 :~g de RNA 5 S marqu6 au a2p ont 6t6 traitds h 5°C pendant 5 minutes par 0,5 unit6 de ribonucl6ase Tj, puts soumis h l'61ectrophori~se pendant 5 heures sous 20 volts/em. Le RNA contenu dans les bandes a et b a 6t6 repris et soumis h une seconde 61eetrophorbse en pr6sence d'ur6e 7 M (B). Les photographies pr6sen- t6es sont les clich6s obtenus en appliquant un film de radiographie contre les blocs de polyacrylamide.
t a i re de l ' ext r6mit6 5' (fig. 8). Aucune au t re zone de compl6men ta r i t 6 n 'ex is te en t re la r6gion 1-25 d ' u n e pa r t et les r 6 g i o n s 54-87 et 90-120, d ' au t r e par t . Nous en d6du i sons que les 2 extr6mit6s de la mol6cule f o r m e n t une • t ige • bif i la ire c o m m e le
m o n t r e la fig. 8.
Inf luence de l'ablation des nucldotides Y-termi- naux sur le comportement chromatographique du RNA 5 S.
Un t r a i t e m e n t tr6s mod6r6 du RNA 5 S marqu6 au a2p p a r la p h o s p h o d i e s t 6 r a s e de v e n i n de ser- pen t , suivie d ' u n e 61ectrophorbse en gel de poly- a c r y l a m i d e h 12,3 p. cen t fair a p p a r a i t r e 2 b a n d e s
BIOCHIMIE, 1973, 55, n ° 9.

1144 H. D e n i s et M. W e g n e z .
d ' in tens i t6 in6gale. La plus i m p o r t a n t e a la m6me mobi l i t6 que le RNA 5 S intact . L ' au t r e est un peu plus r ap ide . Elle c o r r e s p o n d & une fo rme de la
cons ta te que le d6calage en t re le profi l de r ad io - act ivi t6 et le p ro ill de densi t6 op t ique a d i spa ru , alors qu ' i l subsis te p o u r les mol6cules Iaiss6es
TABLEAU II.
Composit ion des fragments obtenus apr~s une hfldrolyse partielle dn RNA 5 S marqud a~ szP.
Bande (fig. 7)
8 9
10 11
Composition (n o des nucl6otides)
1-120 38-120
1-75 -] 49-120J 38-89
au moins deux fragments 1-37
90-120
54-120 54-87 90-120
Proportion
0,21 0,17
0,55
0,29 0,29 0,70 1 ,00
0,19 0,86 0,86 1 ,nO
• La composition des bandes a $t6 dfiterminSe par hydrolyse au moyen de la ribonueldase T~ et s6paration des oligonucl&otides par 61ectrophor6se en 2 dimensions.
La proportion des diff4rents fragments est mesur~e en divisant la radioacti- vit5 de chaque fragment par le hombre de nucldotides qu'il eomp.orte. Pour les bandes 1 h 7, la radioactivit6 du fragment 90-120 est prise comme r6fd- rence. Pour les bandes 8 h 11, la radioactivit6 du fragment 1~25 est prise comme r6f~rence.
La bande 5 fourni t ~ l'hydro.lyse tous les oligonucI6otides correspondant aux r~s idus 1-89. Elle contient donc au moins 2 fragments &environ 45 nuci6otides dont les limites exactes n'ont pas pu ~tre d~termin6es.
mol6cule r a c c o u r e i e d ' e n v i r o n 3 nuc l6o t ides h l ' ex t r6mit6 3'. O n observe aussi u n e t ra in6e de m a t 6 r i e l de mobi l i t6 p lus 61ev6e, et d o n c plus f o r t e m e n t en tam6 p a r l ' enzyme . La b a n d e r a p i d e
in tae tes p a r la p h o s p h o d i e s t 6 r a s e (fig. 9). On re- m a r q u e r a e e p e n d a n t que la quant i t6 de RNA r a c e o u r c i qui a pu 6tre ex t ra i t du gel est tr6s fai-
ble (fig. 9).
IO 15 20 23
r 5 C - A . A - C - C - A - C - C - C - U - G - A - A - A - G
G - . C - C - U - A - C - G - G - C / C I I [ } [ I _ / I I
, o ( U - U ) - U - u - c - G - G - A - u - G . - - u - ~ G / \
120 115 I IO" U - G A - C - C - A - U - A - A - G - G - G - U - C - C
I 0 5 I 0 0 95
Fro. 8. - - Mode d 'appariement probable de l'extr~mit~ 5" et de l'extr~mit5 3' du RNA 5 S de x~nope. La s~quence nucl~otidique a ~t~ ~tablie p a r Wegnez et al [7] et Ford et Southern [8]. Cette s~quence est eelle du RNA 5 S nouvelte- ment form~.
e n g e n d r 6 e p a r le t r a i t e m e n t f la phosphod ies t~ - rase a p e r d u le bout gni f i Ia i re qui fa isai t sai l l ie fi l ' ex t r~mit6 3 '~ de la mo lecu l e (fig. 8). C'est la << t ige >> bif i la i re qui a f re in6 la progressio.n de l ' e n z y m e e t : p r o v o q u 6 l ' a p p a r i t i o n en quant i t6 d6- ce lable de mol6cules 16g6rement r a c c o u r c i e s . Si on 61ue ces de rn i~res du gel et e x a m i n e l eu r eom- p o r t e m e n t sur co lonne d ' a l b u m i n e m6thyl6e, on
BIOCHIMIE, 1973, 55, n ° 9.
Extr~mit~ 3" du RNA 5 S rnarqud au s,p.
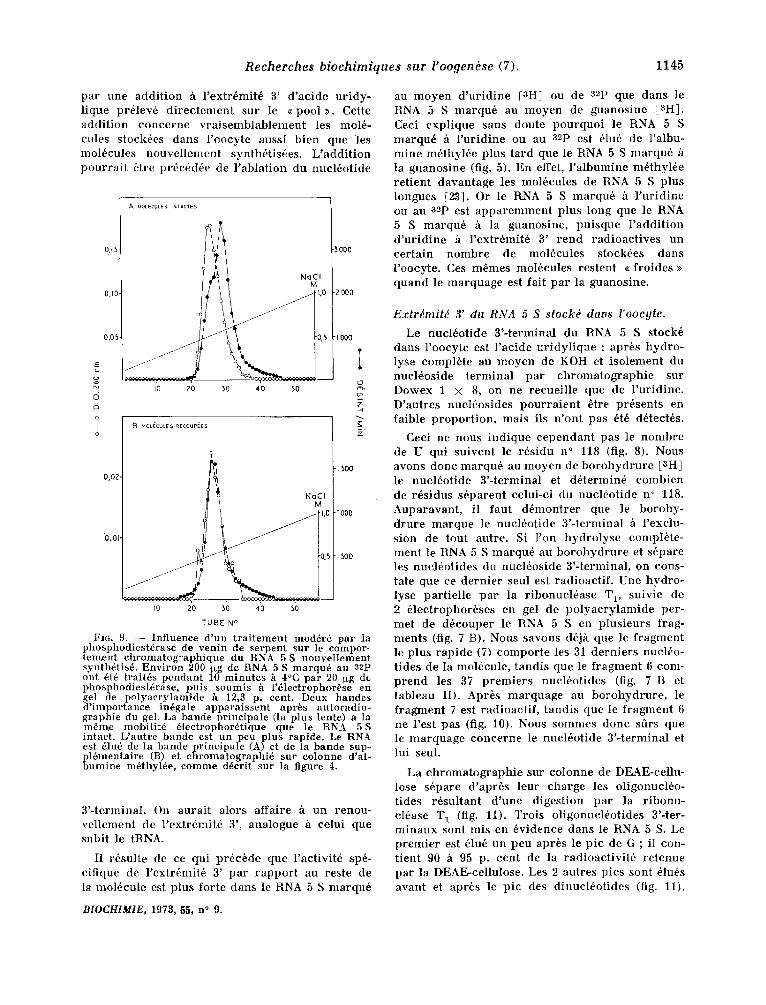
Trois ol igonuch~ot ides di f f6rents (X l, X 2 et Xs) t e r m i n e n t le I~NA 5, S m a r q u 6 au ~2p ( tabIeau I). Le t ab leau I I I d o n n e la c o m p o s i t i o n de ces oligo- nucl6ot ides . On voit que la r ad ioac t iv i t6 p r6sen te dans l ' a c ide u r i d y l i q u e est p lus ~]ev6e qu 'on ne devra i t s 'y a t t endre . C,eci ne peu t s ' exp l ique r que
Recherches b ioch imiques sur l'oogen@se (7). 1 1 4 5
par une add i t ion & l ' ex t r tmi t6 3' d ' a c ide u r idy- l ique pr61ev6 d i r e c t e m e n t sur le << pool >>. Cette add i t i on c o n c e r n e v r a i s e m b l a b l e m e n t les mol t - cules s tock@s darts l 'oocyte aussi b ien que les mo l t cu l e s nouvel le rnent synth t t i s6es . L ' add i t i on pou r r a i t 6tre p r 6 c t d t e de r a b l a t i o n du n u c l t o t i d e
t A MOII~CULES iNTACIES ]
0.0 5 l ~ ~ ~ / ~ 0 " 5 [ 000
I0 20 5'0 ! 4'o s'o ~. & z
22 B MOL£CULES RECOUPEES
1500 0.02.,
1.0 1000
"0, 01' .5 ,500
lb z'o 3o 4'o .so TUBE N °
Fro. 9. - - Influence d'un traitement mud@r6 par la phosphodiesttrase de venin de serpent sur le cornpor- tement chromatographique du RNA 5 S nouvellement synthttist . Environ 200 tlxg de RNA 5 S marqu6 au 82p ont dt6 traitts pendant 10 minutes h 4°C par 20 ug de phosphodiestdrase, puis soumis ~ l'~lectrophorbse en get de polyacrylamide h 12,3 p. cent. Deux bandes d'importance intgale apparaissent aprbs autoradio- graphie du gel. La bande principalc (la plus tente) a la rn~mr mobilit6 6lectrophorttique que le RNA 5S intact. L'autre bande est un peu plus rapide. Le RNA est dlu@ de la bandc principale (A) et de la bande sup- pl tmentaire (B) et chromatographi6 sur eolonne d'al- bumine m~thylte, comme d~crit sur la figure 4.
3 ' - terminal . On aurai t alors affaire h u n renou- ve l l emen t de l ' ex t r tmi t6 3', analogue ~ celui que subit le tRNA.
I1 r t su l t e de ce qui p r t c t d e que l 'act ivi t6 sp t - ci,fique de r e x t r t m i t 6 3' par r appor t au reste de la m o l t c u l e est plus forte dans le RNA 5 S marqu6
au m o y e n d ' u r i d i n e [ Z H ] ou de 32p que dans le RNA 5 S marqu6 au m o y e n de guanos ine [3H]. Ceci exp l ique sans doute p o u r q u o i le RNA 5 S marqu6 & l ' u r i d i n e ou au 32p est 61u6 de l 'albu- m i n e m t t h y l 6 e plus t a rd que le RNA 5 S marqu6 h ta guanos ine (fig. 5). En effet, l ' a lbumine m 6 t h y l t e re t i en t davantage les mo l t cu l e s de RNA 5 S plus longues [23]. Or le RNA 5 S marqu6 h l ' u r i d ine ou an 32.13 est a p p a r e m m e n t plus long que le RNA 5 S marqu6 h la guanosine , pu i sque l ' add i t i on d ' u r i d i n e h l ' ex t r tmi t6 3' r e n d rad ioac t ives un cer ta in nombre de m o l t c u l e s stock6es clans l 'oocyte. Ces m~mes m o l t c u l e s res tent ¢ f ro ides >> q u a n d le marquage est fait pa r la guanosine .
Extr~mit~ 3' dn RNA 5 S stock#, darts l'oocyte.
Le n u c l t o t i d e 3 ' - te rminal du RNA 5 S stock6 dans l 'oocyte est l ' ac ide u r i d y l i q u e : apr~s h y d r o - lyse c o m p l t t e au m o y e n de KOH et i so l emen t du n u c l t o s i d e t e r m i n a l par c h r o m a t o g r a p h i c sur Dowex 1 X 8, on ne recuei l le que de l ' u r id ine . D 'autres n u c l t o s i d e s p o u r r a i e n t ~tre p r t s e n t s en faible p ropo r t i on , mais ils n 'on t pas 6t6 d t t ec t t s .
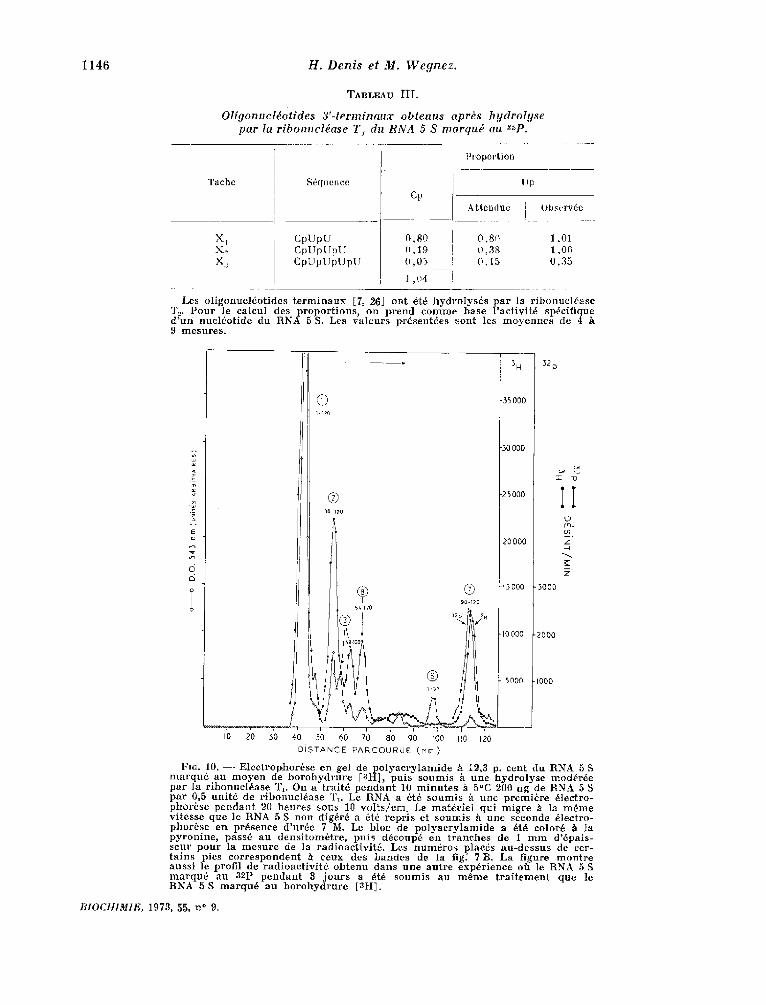
Ceci ne nous i n d i q u e c e p e n d a n t pas le n o m b r e de U qui su ivent le r t s i d u n ° 118 (fig. 8). Nous avons donc marqu6 au m o y e n de b o r o h y d r u r e [~H] le nucl~ot ide 3 ' - te rminal et d t t e r m i n 6 combien de r t s i d u s s t p a r e n t celui-ci du nuc l6ot ide n ° 118. Auparavant , il faut d t m o n t r e r que le borohy- d ru re marque le n u c l t o t i d e 3 ' - te rminal h l 'exclu- sion de tout autre. Si l 'on h y d r o l y s e complbte- merit le RNA 5 S marqu6 au b o r o h y d r u r e et s t pa r e les nucl~ot ides du n u c l t o s i d e 3 ' - terminal , on cons- rate que ce d e r n i e r seul est rad ioac t i f . Une h y d r o - lyse par t ie l le pa r la r i b o n u c l t a s e T1, suivie de 2 61ectrophor~ses en gel de p o l y a c r y l a m i d e per- met de d6couper le RNA 5 S e n p lus ieurs frag- men t s (fig. 7 B). Nous savons dtj& que le f r agmen t le plus r ap ide (7) c o m p o r t e les 31 d e r n i e r s nucl~o- t ides de la molecule , t and i s que le f r agment 6 com- p r e n d les 37 p r e m i e r s n u c l t o t i d e s (fig. 7 B et tableau II). A p r t s m a r q u a g e au b o r o h y d r u r e , le f r agmen t 7 est radioact i f , t and i s que Ie f r agmen t 6 ne l 'est pas (fig. 10). Nous sommes donc stirs que le m a r q u a g e c o n c e r n e le n u c l t o t i d e 3 ' - te rminal et lui seul.
La c h r o m a t o g r a p h i e sur co lonne de DEAE-cellu- lose s t p a r e d ' a p r t s leur charge les o l igonuc l to - t ides r t su l t an t d ' une d iges t ion par la r ibonu- c l tase T 1 (fig. 11). Trois o l igonucl~ot ides 3'-ter- m i n a u x sont mis en 6v idence dans le RNA 5 S. Le p r e m i e r est 61u6 un peu apr~s le p ic de G ; il con- t ient 90 h 9,5 p. cen t de la rad ioac t iv i t6 r e t enue par la DEAE-cellulose. Les 2 autres pics sont 61uts avant et ap r t s le p ic des d i n u c l t o t i d e s (fig. 11).
BIOCHIMIE, 1973, 55, n ° 9.
1146 H. Denis et M. Wegnez.
TABLEAU I I I .
Oligonucldotides 3'-terminaux obtenus aprbs hydroIyse par Ia ribonucl~ase T~ du RNA 5 S marqu~ au s2p.
Taehe
X I X~
Sdquence
CpUpU CpUpUpU CpUpUpUpU
Cp
{i.80 Ii , I9 0,05
1 ,(~4
Proportion
Attendue
0,81) o, 38 O. 15
Up
Observde
1,01 1,06 0,35
Les o l igonuc ldo t ides t e r m i n a u x [7, 2~] ont 6t6 h y d r o l y s d s pa r la r ibonuc l6ase T._,. P o u r le ca lcul des p ropor t ions , on p rend c o m m e base l ' ac t iv i t6 spdcifique d ' u n nuc l6ot ide du RNA 5 S. Les va l eu r s prdsent6es son t les m o y e n n e s de 4 h 9 mesures .
© 1-120
@ ~J8 120
~'o 2'o ~'o
90~120 5 ~,-I 2'0
4942~
;'0 ,oo ,,o DISTANCE PARCOURUE (mm)
3 H 32p
-55 00(
-50 OOC
c.w r-J
I 13
C? c,el.
-20 ooi z
45000 5000
'10000 '2000
• .5000 IlO00
Fro. 10. - - E lec t rophor6se en gel de p o l y a c r y l a m i d e h 12,3 p. cent du RNA 5 S m a r q u 6 au m o y e n de b o r o h y d r u r e [SH], puis soumis & une h y d r o l y s e moddrde pa r la r ibonuc ldase T1. On a t r a i t6 p e n d a n t 10 m i n u t e s h 5°C 200 ug de RNA 5 S pa r 0,5 un i t6 de r ibonue ldase T1. Le RNA a 6t6 soumis & une p remi6re 61ectro- phor6se p e n d a n t 20 henres sous 10 vo ! t s / em. Le ma te r i e l qui migre h la m~me vi tesse que le RNA 5 S non dig6rd a dtd repr i s et s o u m i s h une seconde 61ectro- phorbse en presence d 'ur~e 7 M. Le bloc de p o l y a c r y l a m i d e a dr6 color6 h ta p y r o n i n e , pass6 au dens i tom6t re , puis ddcoup6 en t r anches de I m m d ' @ a i s - seur p o u r la m e s u r e de la r ad ioac t iv i td . Les n u m 6 r o s plac6s au -des sus de cer- t a in s pics c o r r e s p o n d e n t A ceux des bandes de la fig. 7 B. La figure m on t re auss i le profil de r ad ioac t iv i t 6 o b t e n u dans une au t r e exp6r ience off le RNA 5 S m a r q u 6 au 32p p e n d a n t 3 j o u r s a 6t6 soumis au m6me t r a i t e m e n t que le RNA 5 S m a r q u 6 au b o r o h y d r u r e [al l ] .
BIOCHIMIE, 1973, 55, n ° 9,
Recherches biochimiques sur l'oogen~se (7). 1147
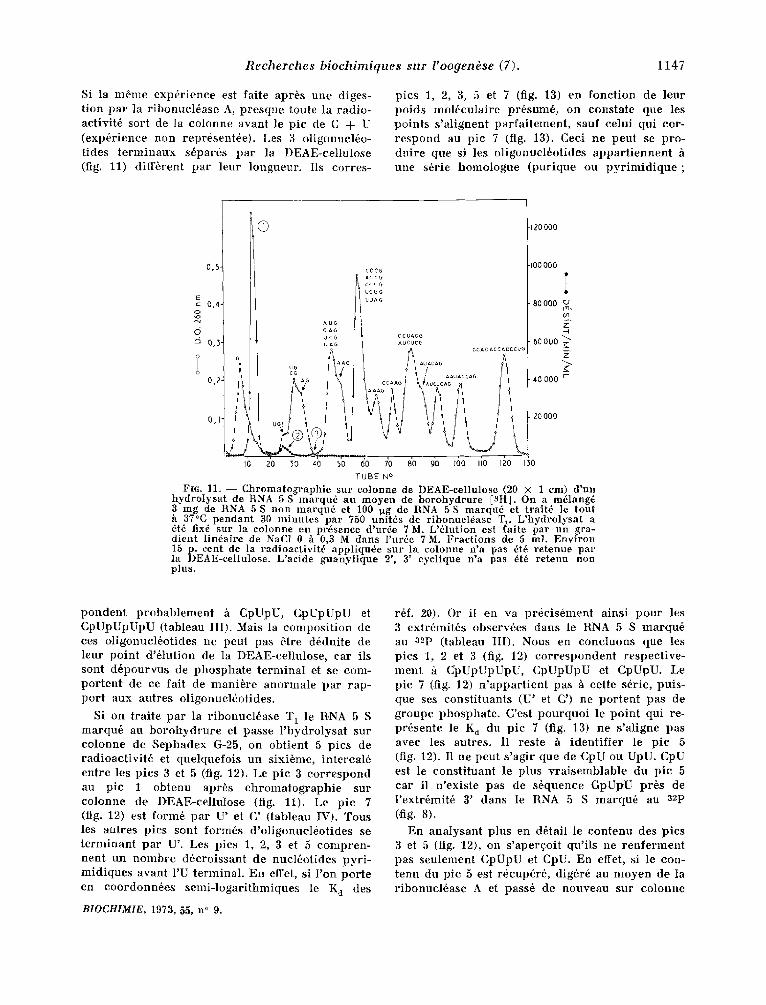
Si la m6me exp6rience est faite apr6s une diges- tion par la r ibonucl6ase A, presque toute la radio- activit6 sort de la colonne avant le pic de C + U (exp6rience non repr6sent6e). Les 3 oligonucl6o- tides te rminaux s6par6s par la DEAE-cellulosc (fig. 11) diff6rent par leur longueur. Ils corres-
pics 1, 2, 3, 5 et 7 (fig. 13) en fonct ion de leur poids mol6culaire pr6sum6, on constate que les points s 'al ignent parfa i tement , sauf celui qui cor- respond au pic 7 (fig. 13). Ceci ne peut se pro- duire que si les oligonucl6otides appar t i ennent h une s6rie homologue (purique ou pyr imid ique ;
© -I 20 000
0.5 -100 000 CCCG ACCG [ CCUC~ UCUG
E UUACz ,- 0,4- 80000 O Frl.
AUG (~ CAG --I
UCG CCUACG f-Ct 0.3- A5 AuCUCG 60 000 TM
Oo,.2- 7 ~ oo, / ....... ~ 4020 ooo ooo ~
. . . . ~ . . . . - ~ . . . . ~ . r . . ~ . _ ; , . ~ . ~ l 10 20 30 40 50 60 70 80 .90 100 II0 120 130
TUBE N °
FIG. 11. - - C h r o m a t o g r a p h i e sur co lonne de DEAE-eel lu lose (20 × 1 em) d 'un h y d r o l y s a t de RNA 5 S m a r q u 6 au m o y e n de b o r o h y d r u r e [aH]. On a mdlang4 3 mg de RNA 5S non m a r q u 6 et I00 .~g de RNA 5S m a r q u 6 et t ra i t6 le tou t h 37°C p e n d a n t 30 m i n u t e s pa r 750 uni t6s de r ibonuc l6ase Tx. L ' h y d r o l y s a t a 6t5 fix6 sur la co lonne en pr6sence d 'ur6e 7 M. L'61ution est f a i t e par u n gra- d i en t l indal re de NaG1 0 h 0,3 M dans l 'u rde 7M. F r a c t i o n s de 5 ml. EnviroJt 15 p. cent de la r ad ioae t iv i t d app l iqu4e sur la co lonne n 'a pas 6t6 r e t enue par la I )EAE-cel lulose. L 'ac ide g u a n y l i q u e 2', 3' cyc l ique n 'a pas 6t6 r e t e n u non plus.
pondent probablement h CpUpU, CpUpUpU et CpUpUpUpU (tableau III). Mais la composi t ion de ces oligonucl6otides ne peut pas 6tre d6duite de leur point d '6lution de la DEAE-cellulose, car ils sont d6pourvus de phosphate te rminal et se corn- por tent de ce fait de manibre anormale par rap- por t aux autres oligonucl6otides.
Si on trai te par la r ibonucl6ase T 1 le R NA 5 S marqu6 au borohydrure et passe l 'hydro lysa t sur colonne de Sephadex G-25, on obtient 5 pics de radioactivi t6 et quelquefois un sixi6me, intercal6 entre les pics 3 et 5 (fig. 1,2). Le pic 3 correspond au pic 1 obtenu apr6s chromatographie snr colonne de DEAE-cellulose (fig. 11). Le pic 7 (fig. 12) est form6 par U' et C' (tableau IV). Tous les autres pics sont form6s d 'oligonucl6otides se te rminant par U'. Les pics 1, 2, 3 et 5 compren- nent un hombre d6croissant de nucI6otides pyri- midiques avant I'U terminal . En effet, si l 'on porte en coordonn6es semi-logari thmiques le K d des
BIOCHIMIE, 1973, 55, n ° 9.
r6f. 20). Or il en va pr6cis6ment ainsi pour les 3 extr6mit6s observ6es darts le RNA 5 S marqu6 au a2p (tableau III). Nous en concluons que les pics 1, 2 et 3 (fig. 12) cor respondent respective- merit /~ C'pUpUpUpU, CpUpUpU et CpUpU. Le pic 7 (fig. 12) n ' appar t ien t pas ~ cette s6rie, puis- que ses const i tuants (U' et C') ne por tent pas de groupe phosphate. C'est pourquoi It point qui re- pr6sente le tK d du pic 7 (fig. 13) ne s 'aligne pas avec les autres. I1 reste ~ ident i f ier le pic 5 (fig. 12). I1 ne peut s 'agir que de CpU ou UpU. CpU est le const i tuant I t plus vraisemblable du pic 5 car il n 'existe pas de s6quence GpUpU prbs de l 'extr6mit6 3' darts I t RNA 5 S marqu6 au a2p (fig. 8).
En analysant plus en d6tail le contenu des pics 3 et 5 (fig. 12), on s 'aper¢oit qu'ils ne renferment pas seulement CpUpU et CpU. En effet, si le con- tenu du pic 5 est r6cup6r6, dig6r6 au moyen de la r ibonucl6ase A e t pass6 de nouveau sur colonne
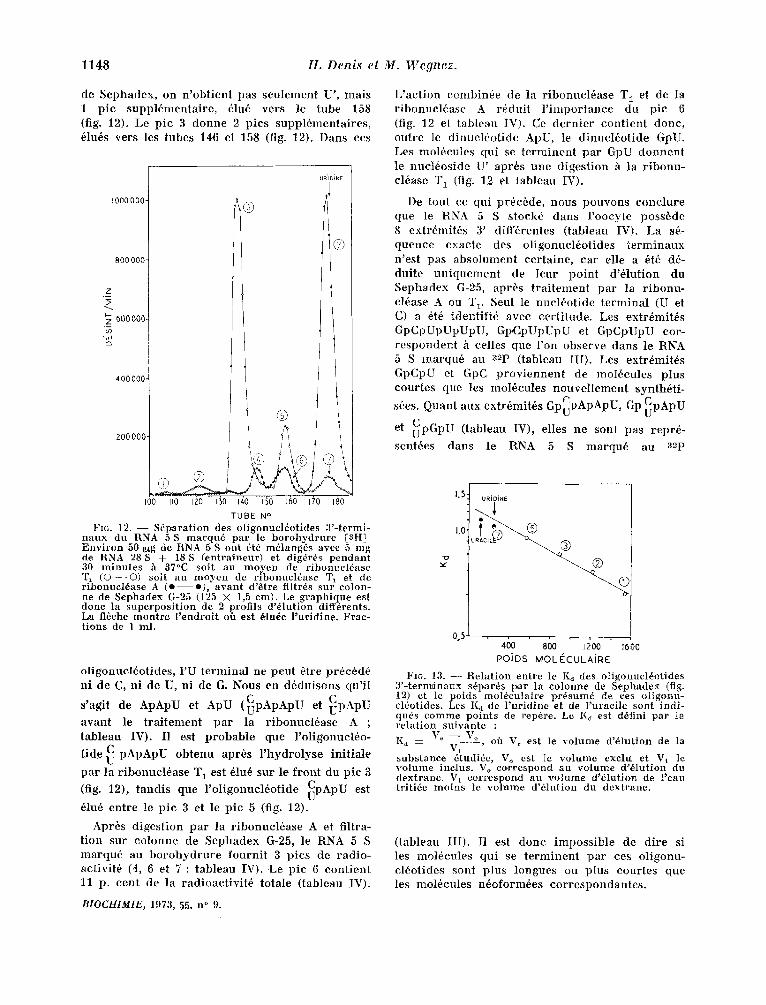
1148 H. Denis el M. Wegnez.
de Sephadex , on n 'ob t ien t pas seu lemen t U', mats 1 p ic supp l6menta i re , 61u6 vers le tube 158 (fig. 12). Le p ic 3 d o n n e 2 pics suppl6menta i res , 61u6s vers les tubes 1~6 et 158 (fig. 12). Darts ces
1000000-
B00000-
Z
600 000-
400000-
200000.
@
IO0 IIO 120 150 14o {50 160 170
TUBE N °
URiDiNE
C
I
I
I
I
Fro. 12. - - Sdparation des oligonuel~otides 3'-termi- naux du RNA 5 S marqn6 par le borohydrure [aH! Environ 50 I~g de RNA 5 S ont 6t~ m61ang~s avec 5 mg de RNA 28 S + 18 S (entraineur) et digdr6s pendant 30 minutes h 37°C soit au moyen de ribonucl6ase % (©- -©) soit au moyen de ribonucl6ase T~ et de ribonucldase A ( o - - e ) , avant d'dtre filtrds sur colon- ne de Sephadex G-25 (125 X 1,5 cm). Le graphique est dour la superposition de 2 profils d'dlution diff6rents. La fl6che montre l 'endroit off est 61ude l 'uridine. Frac- tions de 1 ml.
o l igonuel6ot ides , I'U t e rmina l ne peut 6tre pr6c6d6 ni de C, ni de U, ni de G. Nous en d6duisons qu ' i l
s 'agit de ApApU et ApU ( ~ p A p A p U et ~ p A p U
avant le traitement par la ribonucl6ase A ; tableau IV). I1 est probable que l'oligonucl6o-
tide C pApApU obtenu apr6s l 'hydrolyse initiale
par la ribonucl6ase T~ est 61u6 sur le front du pic 3
(fig. 12), tandis que l 'ol igonucl6otide CpApU est
61u6 entre le pic 3 et le pic 5 (fig. 12).
Aprbs digestion par la ribonucl6ase A et filtra- tion sur colonne de Sephadex 6-25, le RNA 5 S marqu6 au borohydrure fournit 3 pics de radio- activit6 (4, 6 et 7 : tableau IV). ,Le pic 6 contient 11 p. c e n t de la radioactivit6 totale (tableau IV).
L'ae t ion combin6e de la r ibonuc lbase T1. et de la r ibonue l6ase A r6dui t l ' i m p o r t a n c e du pic 6 (fig. 12 et tableau IV). Ce d e r n i e r con t i en t donc, outre le d inuc l6o t ide ApU, le d inuc l6o t ide GpU. L es mol6cules qui se t e r m i n e n t pa r GpU d o n n e n t Ie nuc l6os ide U' apr6s une d iges t ion ',k la r ibonu- cl6ase T~ (fig. 12 et tableau IV).
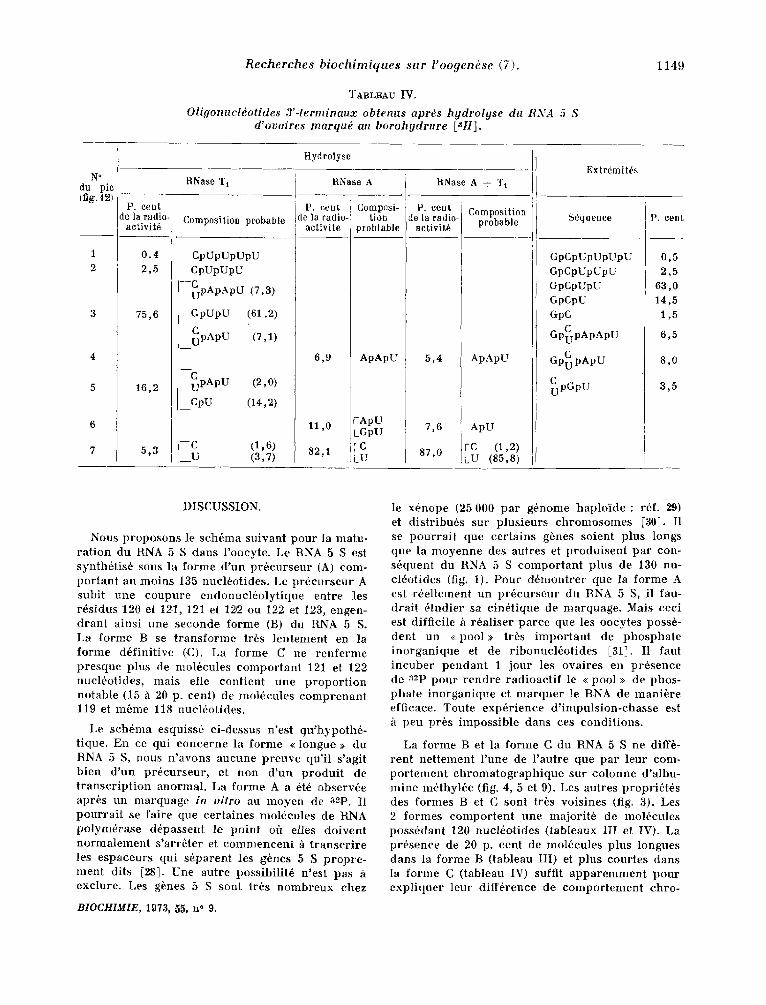
De tout ce qui pr6e6de, nous pouvons conc lu re que le RNA 5 S stock6 dans l 'oocyte poss6de 8 extr~mit6s 3' diff6rentes ( tableau IV). La s6- quence exacte des o l igonuel6ot ides "ierminaux n 'est pas abso lumen t cer ta ine , ear elle a 6i6 d6- dui te u n i q u e m e n t de Ieur po in t d'61ution du Sephadex G-25, aprbs t r a i t e m e n t par la r ibonu- el6ase A ou T 1. Seul le nuc l6o t ide t e r m i n a l (U et C) a 6t6 ident i f i6 avee cer t i tude. Les extr6mit~s GpCpUpUpUpU, Gp,CpUpUpU et GpCpUpU cor- r e s p o n d e n t h celles que l 'on observe dans le RNA 5 S marqu6 au 32p ( tableau III). Les extr6mit6s GpCpU et GpC p r o v i e n n e n t de mol6cules plus cour tes que les moI6cules n o u v e l l e m e n t synth6t i -
s6es. Quant aux extr6mit6s Gp~pApApU, Gp CpApU
et ~ p G p U (tableau IV), elles ne sont pas repr(%
sent6es dans le RNA 5 S marqu6 au 32p
].5~ URiOiNE
i .o ~ ®
0,5 4;0 s ; o ~ ,~oo ' ,600
PO]DS MOL ECULAiRE
]qG. 13. - - Relation entre ]e Kd des oligonuc!~otides 3'-terrr~inaux s4p,ar~s par la colonne de Sephade.~ (fig. 12) et le poids moI4cutaire pr6sum6 de ces oligonu- cl6otides. Les K,~ de l 'uridine et de l'uracile sont indi- quds comme points de rep6re. Le Ka est d6fini par ta relation suivante : Kd -- Ve - - Vo, oh V~ est le volume d'Siution de la
Vl substance 6tudide, Vo est le volume exclu et V~ le volume indus. Vo co.rrespond au volume d'61ution dt~ dextrane. V~ correspond au volume d'61ution de l'eau triti6e moins le volume d'61ution du dextrane.
( tableau III). I1 est donc imposs ib le de d i re si les mol6cules qui se t e r m i n e n t pa r ces ol igonu- cl6otides sont plus longues ou plus courtes que les mol6cules n6oforn16es co r r e spondan te s .
BIOCHIMIE, 1973, 55, n ° 9.
Recherches biochimiques sur l'oogendse (7).
TABLEAU I V .
Oligonucldotides 3'-terminaux obtenus apr~s hgdrolyse du RNA 5 S d'ovaires marqu~ au borohydrure [~H].
1149
N a du pie (fig. 121
Hydrolyse
P. cent de la radio-
aetivit6
0 . 4
2,5
75,6
16,2
5,3
RNase T~
Composition probable
CpUpUpUpU CpUpUpU
--~pApApU (7,3)
CpUpU !61,2)
~ p A p U (7,1)
RNase A
P. ceUL de la radio-
activite
Composi- tion
problable
ApApU
RNase A + "1"~
P. cent de la radio-
activit6
---•pApU (2,0)
I CpU (14,2)
( , ,6, (3,7)
6 , 9 5,4
11,0
82,1
[ ApU GpU 7,6
87,0
Composition probable
ApApU
ApU
(85,8)
Extrdmitds
S6quence
GpCpUpUpUpU GpCpUpUpU GpCpUpU GpCpU GpC
C GPuPApApU
C GPuPApU
~pGpU
p. cent
0,5 2,5
63,0 14,5 1,5
6,5
8,0
3,5
DilSCUSSION.
Nous proposons le sch6ma suivant pour la matu- ra t ion du RNA 5 S dans l 'oocyte. Le RNA 5 S est synth6tis6 sous la forme d'un pr6curseur (A) com- por tant au moths 135 nucl6otides. Le pr6curseur A subit une coupure endonucl6olyt ique entre les r6sidus 120 et 12,1, 121 et 12~2 ou 122 et 123, engen- drant ainsi une seconde forme (B) du RNA 5 S. La forme B se t ransforme tr6s lentement en la forme d6finitive (C). La forme C ne renferme presque plus de mol6cules compor tant 121 et 122 nucl6otides, mats d ie contient une propor t ion notable (15 h 20 p. cent) de mol6cules comprenant 119 et m6me 118 nucl6otides.
Le sch6ma esquiss6 ci-dessus n'est qu 'hypoth6- tique. En ce qui concerne la forme << longue >> du RNA 5 S, nous n 'avons aucune preuve qu'il s'agit bien d 'un pr6curseur, et non d 'un produi t de t ranscr ip t ion anormal. La forme A a 6t6 observ6e apr~s un marquage in vitro au moyen de 32p. I1 pour ra i t se faire que certaines mol6cules de RNA polym6rase d6passent le point o6 elles doivent normalement s 'arr6ter et commencent /L t ranscr i re les espaceurs qui s6parent les g6nes 5 S propre- merit dits [28]. Une autre possibilit6 n'est pas /~ exclure. Les g6nes 5 S s0nt tr6s nombreux chez
BIOCHIMIE, 1973, 55, n ° 9.
le x6nope (25 000 par g6nome haploide : r6f. 29) et distribu6s sur plusieurs chromosomes [30]. I1 se pour ra i t que certains g6nes soient plus longs que la moyenne des autres et produisent par con- s6quent du RNA 5 S compor tant plus de 130 nu- cl6otides (fig. 1). Pour d6montrer que la forme A est r6ellement un pr6curseur du RNA 5 S, il fau- drai t 6tudier sa cin6tique de marquage. Mats ceci est difficile /~ r6aliser parce que les oocytes poss6- dent un <<pool >> trbs impor tant de phosphate inorganique et de r ibonucl6otides [31]. I1 faut incuber pendant 1 jour les ovaires en pr6sence de 32p pour rendre radioact i f l e ¢ pool >> de phos- phate inorganique et marquer le RNA de manibre efficace. Toute exp6rience d ' impulsion-chasse est h peu pr6s impossible darts ces condit ions.
La forme B e t la forme C du R, NA 5 S n e diff6- rent nettement l 'une de l 'autre que par leur com- por tement chromatographique sur colonne d'albu- mine m6thyl6e (fig. 4, 5 et 9). Les autres propri6t6s des formes B e t C sont tr6s voisines (fig. 3). Les 2 formes compor tent une majorit6 de mol6cules possedant 120 nucldotides (tableaux III et IV). La pr6sence de 20 p. cent de mol6cules plus longues dans la forme B (tableau III) et plus courtes dans la forme C (tableau IV) suffit apparemment pour expliquer leur diff6rence de comportement chro-

1150 H. Denis et M. Wegnez .
m a t o g r a p h i q u e (fig. 9'). Le s6jour pro long6 du R,NA 5 S dans l ' oocy te e n t r a l n e donc le r accour - e i s sement d 'une pa r t i e des mol$cules h l 'extr6- mit6 3', de m~me qu 'nne per te des groupes phos- pha tes h l ' ex t r6mi t6 5' [26].
Le r a c e o u r e i s s e m e n t du R:N.A 5 S est p robab le - m e n t p r o d u i t p a r une exonucI6ase. C'est peut-~tre Ie mSme enzyme qui ajoute des r6s idus u r idy l i - ques h l ' ex t r6mi t6 3' ( tableau III) . Ce p h 6 n o m 6 n e n 'es t pas i n c o m p a t i b l e avec le r a c c o u r c i s s e m e n t de la mol6eule observ6 h long terme. Ce qui se p r o d u i t v ra i semblab lemen t , c 'est une ab la t ion de I'U t e rmina l , suivie de son r e m p l a e e m e n t p a r un nue l6ot ide pr61ev6 duns le ~< pool >~. Le remplace - men t n 'es t sans doute pus tou t h fa i t aussi efficace que l ' ab la t ion , ce qui en t r a ine pou r ee r ta ines mo- 16cules la per te de 1 on de p lus ieu r s nucl6ot ides .
La pr6sence des extr6mit6s GpCpApApU,
G p C p A p U et ~ipGpU (tableau IV) dans le RNA 5 S
s tock6 est diff ic i le :k c o m p r e n d r e p u i s q u ' a u c u n e extr6mit6 c o r r e s p o n d a n t e n 'ex is te duns le RNA 5 S nouve l l emen t syn th6 t i s6 (fig. 8). P lus i eu r s expl ica- t ions sont possibles . 1) On b ien ces ol igonucl6o- t ides n 'on t pas 6t6 d~tect6s duns le RNA 5 S n~o- form6 ( tableau III). Cect p a r a i t peu v ra i semblab le , pu i sque la molaritO add i t i onn6e des ol igonucl6o- t ides Xx, X2 et X 3 est sup6r ieure h 1 ( tableau III) .
2) Ou bien les extr6mitOs GpCpApApU, Gp~,pApU
et ~ p G p U p r o v i e n n e n t d 'une c o n t a m i n a t i o n du
RNA 5 S p a r d ' au t res esp~ces de RNA. C e t t e deux i&ne poss ib i l i t6 est aussi peu probable . Le RNA 5 S a 6t6 pur i f i6 p a r f i l t ra t ion sur co lonne de S e p h a d e x G-100, suivie de 2 61ectrophor6ses en gel de p o l y a e r y l a m i d e , don t l ' une en prSsenee d 'urSe 7 M. Tout o l igonuel6ot ide qui a d h 6 r e r a i t an RNA 5 S devra i t 6tre 61imin6 au eours de la pu r i f i ca - t ion. 3) I1 reste une t ro i s i6me poss ib i l i t6 : une pa r t i e du RNA 5 S ,(___ 15 p. cent) p o u r r a i t 6tre modi f i6e h l ' ex t r6mi t6 3'. Une ab la t ion se p rodu i - r a i t j u squ ' au nue l6o t ide .118 ( C ; fig. 8) et se ra i t suivie de l ' a d d i t i o n soit de GpU, soit de ApU, soi t de ApApU (tableau IV). A p r e m i 6 r e rue , eette sui te d '6v6nements p a r a l t peu probable . Mats il ne tau t pus oubl ie r que le RNA 5 S s6 journe duns l 'oocyte p e n d a n t de n o m b r e u x mois [9] et peu t sub i r des r e m a n i e m e n t s i m p o r t a n t s qui n ' a p p a r a i s s e n t pus q u a n d l ' i n c o r p o r a t i o n duns Ie r ibosome a l ieu tout de sui te apr6s la synth6se , eomme c 'est le cas duns les cel lules somat iques (fig. 6).
Les h y d r o l y s e s mod6r6es p a r la r ibonuc l6ase T~ a p p o r t e n t ce r ta ines i n f o r m a t i o n s c o n c e r n a n t la s t ruc tu re s e c o n d a i r e du RNA 5 S de x6nope. Six coupures i n t rodu i t e s p a r la r ibonuc l6ase Tx ont pu
BIOCHIMIE, 1973, 55, n ° 9.
8tre lo.calis6es ( tableau II) : aprbs les nuc l6ot ides 25, 37, 48, 53, 75 et 89. I1 y a donc 2 r6gions par t i - cu l i6 rement expos6es dans la mol6cule : celle qui est compr i se en t re les nuc l6ot ides 25 et 53 et celle qui est compr i se ent re les nuc ldot ides 75 et 89. Aucune coupure ne se p r o d u i t avan t le r~sidu 25 et apr~s le r~sidu 89. Ceci conf i rme que les 2 extr&ni t6s de la mol6cule sont appar i6es en une <( tige >~ bifflaire, peu sens ib le h la r ibonuc l6ase (fig. 8). Les zones c o r r e s p o n d a n t aux r6s idus 25 53 et 75 h 89 t o r m e n t p r o b a b I e m e n t des boucles expos6es ~ l ' enzyme. La f r6quence de ce r t a ines coupures var ie d 'une h y d r o l y s e fi l ' au t re (fig. 7 et 10 et t ab leau II). Pa r exemple , de nombreuses moI6cules ont 0 6 coup6es apr6s le r6s idu 53 dans l ' exp6 r i ence de la f igure 10, mats beaucoup moths l 'ont 0 6 dans l ' exp6r i ence d6cri te sur la fig. 7. I1 est diff ici le d ' exp l i que r ces var ia t ions .
Remerciements.
Nous remereions le Pro.fesseur R. Monier d'avoir bien voulu aecueillir M. Wegnez pendant la r~alisation d'une pattie de ce travail. Nous remereions aussi le Fonds National de la Recherche Scientifique pour son aide fin,anci6re. M. Wegnez est boursier du Patrimoiue de l'Universit~ de Libge.
R~SUM~.
Comme nous l'avons montr6 prdc6demment, le RNA 5 S synthdtis4 par les petits oocytes de Xenopus laevis est mis en r6serve pendant plusieurs mois avant d'etre irLcorpor6 dans les ribosomes. Duns cet article, nous comparons les pro~pri6t6s du RNA 5 S nouvelle- ment synthdtis6 avec eelles du RNA 5 S stock6 dana l'oocyte. Le RNA 5 S ndoform6 diffSre du RNA 5 S total par sa mobilit6 61ectrophor6tique et par son comportement chromatographique.
Nous p.roposons le schdma suivant p o u r la matu- ration du RNA 5 S dans l'oocyte. Le RNA 5 S apparatt to,at d'abord sons la forme d'un prdeurseur (forme A) contenar~t plus de 135 nacldotides. Ce pr&nrseur subit rapidement une ablatiorL h l 'extr6mit6 3' et donne naissance h la deuxi6me forme (B) du RNA 5 S. CelIe- ci comprend 80 p. cent de mol&ules longues de 12:0 uuel~otides et 20 p. cent de mol&ules longues de 121 ou 1~2 nttcl~otides. La fovme B se convertit tr6s lentement en uue troisi&me forme (C), qui est la forme de stockage. La p lupart des molecules de la forme C comportent 12'0 nucI6otides, mats 15 h 20 p. cent d'entre elles comportent 119 ou 118 nucldotides. Dans les cellules somatiques, le RNA 5 S ne subit aucune m,odification h long terme semblable h celle qui s'o.bserve darts les o,ocytes.
Le schema esquiss6 ei-dessus est ap,puy~ par les ob- servations suivantes.
1) Le RNA 5 S marqud an 32p contient une faible proportion de mol6cules qui contiennent au moins 135 aucl6otides. Les nucl~otides suppldmentaires sont localis6s h l 'extr&nit6 3' du RNA 5 S.
2) Le RNA 5 S nouvellement synth~tisd (forme B) est 16g6rement plus long que le RNA 5 S stock& Ceei peat ~tre d6montr5 en mesurant la mobilit6 61ectro-

Recherches biochimiques sur l'oogen~se (7). 1151
phor6tique du f r agmen t 3 ' - terminal (nucl6otides 90- 120) obtenu apr6s nne hydro lyse par t ie l le du RNA 5 S par la r ibonncl6ase T1. Si I 'on en[6ve les nucI6otides qui suivent le r6sidu n ° 118 par un t r a i t emen t & !a phosphodiest6rase de venin de serpent, le comporte- ment ch roma tog raph ique du RNA 5 S nouve l lement form5 devient ident ique h celui du RNA 5 S stock~ dans i 'oocyte.
3) Apr6s hydro tyse par la r ibonucldase T , le RNA 5 S marqu6 au 32p fourn i t 3 oligonucl6otides 3'-termi-- imux (CpUpU, CpUpUpU et CpUpUpUpU). La p lupar t des mol6cules no uve l lement synth6tis~es se t e rminen t par l 'oligonucl6otide CpUpU. Le RNA 5 S stock6 dans l 'oocyte poss6de 8 extr6mit6s 3' diff~rentes. La plus fr~quen~e est CpUpU, comme dans le RNA 5 S nouvel- lement synth6tis6 (63 p. cent de routes les extr6mit6s). Les extrdmit~s CpUpUpU et CpUpUpUpU ne se ren- cont rent que darts 3 p. cent des mol6cules. Deux e x t r ~ mit6s (CpU et C : 16 p. cent de routes les extr6mitds) cor respondent h des molecules con tenan t 1 h 4 nucl~o- tides en moins que le RNA 5 S nouve l lement synth6-
tis~. Les 3 derni~res extr6mit6s (Gp~ pApApU~
Gp~ pApU et ~ pGpU : 18 p. cent de routes les extr~mi-
t~s) n 'ont Std d6tect6es que clans le RNA 5 S stock~ dans l 'oocyte. Ces extr~mit6s ne peuvent pas d6river de celles que l 'on a observ~es dans le RNA 5 S nais- sant par une simple abla t ion des nucldotides t e rmi- naux.
4) Apr6s marquage au 32p, les r6sidus ur idyl iques des oligonucl6otides 3 ' - t e rminanx (CpUpU, CpUpUpU et CpUpUpUpU) ont tou jours une activitd sp6cifique p.lus dlev~e que le r6sidu cyt idyl ique. Ceci suggbre un re~ouve l lement de l 'extr~mit~ 3', qui pour ra i t ~tre en rappor t avec le raceourc issement tr6s lent que le RNA 5 S subit pendan t son stockage dans l 'oocyte.
BIBLIOGRAPHIE.
1. Gall, J. G. (1968) Proc. Nat. Acad. Sci. U.S., 60, 553-560.
2. Evans, D. ,~ Birnstiel, M. L. (1968) Biochim. Bio- phys . Acta, 1'66, 274-276.
3. Bro,wn, D. D. a Dawid, I. B. (1968) Science, 160, 272-280.
4. Wegnez, M. ~ Denis, H. (1972) Biochimie, 54, 1069- 1072.
5. Mairy, M. & Denis, H. (1971) Develop. Biol., 24, 143- 165.
6. Ford, P. J. (1971) Nature, 233, 561-564. 7. Wegnez, M., Monier, R. & Denis, H. (1972) FEBS
Letters, 25, 13-20. 8. Ford, P. J. & Southern, E. M. (1973) Nature N e w
Biol., 241, 7-12. 9. Mairy, M. & Denis, H. (1972) Eur. J. Biochem., 25,
535-543. 10. Denis, H. a Mairy, M. (1972) Eur. J. Biochem., 25,
524-534. 11. Ham, R. G. (1965) Proc. Nat . Acad. Sci. U°S., 53,
288-293. 12. Brownlee, G. G., Sanger, F. & Barrell , B. G. (1968)
J. Mol. Biol., 34, 379-412. 13. Denis, H., Wegnez, M. & Willem, R. (1972) Biochi-
mie, 54, 1189-1196. 14. Vigne, R. & Jordan, B. R. (1971) Biochimie , 53,
981-986. 15. Adams, J. M., Jep,pesen, P. G. N., Sanger, F. & Bar-
rell, B. G. (1969) Nature, 223, 1009-1014. 16. Hatlen, L. E., Amaldi , F. & Attardi, G. (1969) Bio-
chemis try , 8, 4989-5005. 17. Lane, B. G. (1963) Biochim. Biophys . Acta, 72, 110-
112. 18. Ra jBhandary , U. L. (1968) J. Biol. Chem., 243, 556-
564. 19. De Wachter , R. & Fiers, W. (1967) J. MoI. Biol., 30,
507-527. 20. Hohn, T. & Schaller, H. (1967) Biochim. Biophys .
Acta, 138, 466-473. 21. Tomlinson, R. V. ~ Tener, G. M. (1962) J. Am. Chem.
Soc., 84, 2644-2645. 22. Galibert, F., Tiollais, P., Sanfourche, F. & Boiron,
M. (1971) Eur. J. Biochem., 20, 381-391. 23. Monier, R., Feunteun , J., Forget, B., Reynier , M.
Varricchio, F. (1969) Cold Spr. Harb. S~3mp. Quant. Biol., 34, 139-148.
24. Feunteun, J., Jordan , B. R. & Monier, R. (1970) Bull . Soc. Chim. Biol., 52, 979-982.
25. Jordan, B. R., Forget, B. G. & Monier, R. (1971:) J. Mol. Biol., 55, 407-421.
26. SVegnez, M. & Denis, H. Article n ° 6 de la pr6sente s6rie, Biochimie , sous presse.
27. Jordan, B. R. (1971) J. Mol. Biol., 55, 423-439. 28. Brown, D. D., Wensink, P. C. & Jordan, E. (1971)
Proc. Nat. Acad. Sci. U. S., 68, 3175-3179. 29. Brown, D. D. & Weber, C. S. (1968) J. Mol. Biol..
34, 661-680. 30. Pardue, M. L., Brown, D. D. & Birnstiel, M. L.
(1973) Chromosoma, 42, 191-204. 31. Woodland, H. R. ~ Pestell, It. Q. W. (1972) Bio-
chem. J., 127, 597-605.
BIOCHIMIE, 1973, 55, n ° 9. 75