Embed Size (px)
Citation preview

BIOCHIMIE, 1972, 54, 589-595. COLLOQUE C.N.R.S.
BASES M O L I ~ C U L A I R E S D E LA P A T H O L O G I E ,
11-13 septembre 1971.
RNA ct biosynth6se de l'hdmoglobine.
J a c q u e s Knur l .
Ins t i tu t de Pathologic Mol~culaire, 2~, rue du Faubourg S a i n t - J a c q u e s - Paris lit ~.
Abstract. - - In 19~60, we have observed wi th G. Schapira and J. C. Dreyfus t ha t when r ibosomes f rom r a b b i t ret iculocytes were incuba ted w i t h the pH 5 f rac t ion f rom guinea pig .reticulocyte, as wel l as in the reverse system, bo th r abb i t and guinea pig hemoglobins were synthesized. The discovery of the messenger RNA by Jacob and Monod aIIowed us to give an exp lana t ion of thes.e observat ions . The rea l i ty of these synthesis, was ascer ta ined by the m e t h o d of double labe l l ing and peptide analys is .
In order to demons t r a t e the presence of messenger RNA in re t ieuloeytes we have puri- fied RNA f rom these ceils. Hemoglobin synthesi 's was s t imula ted when th is RNA was added to a re t iculocyte cell free system.
This RNA wa,s f rac t iona ted on a sucrose gradient . The f rac t ion wh ich sedimented bet- ween 18S and 4S, and which was called RNA III, s t imula ted hemoglob in cell free syn- thesis. The s t imula t ion is specific, since :
- - The o ther RNA f rac t ions did not s t imula te hemoglob in synthesis . - - RNA III f rom rct icnlocytes f rom other species were e i the r inact ive or inh ib i to ry . These resu l t s arc in favor of the presence of messenger RNA in th i s f rac t ion in ret i-
eulocy~es. RNAs extracted from rabbi t , l iver, kidney, ,i,ntestine nuclei, and the frae~io.n III f rom
these RNAs also s t imula ted hemoglob in synthesis . RNA f rom cytoplasm, RNsA III f rom l iver nuclei f rom o ther species did not s t imula te h, emaglobin synthesis . These resul ts favor the idea tha t nuclei f rom di f ferent ia ted cells conta in messenger RNAs which are not t r ans l a t ed in these cells. We are now s tudying meehan ims involved in the selection of messenger RNAs in dif ferent ia ted cells.
L o r s q u e l ' o n i n c u b e des r 6 t i c u l o c y t e s h 37 ° d a n s u n m i l i e u i o n i q u e c o n v e n a b l e e n p r 6 s e n c e de ~-(14C) g lycoco l l e , o n c o n s t a t e qu ' i l y a u n e syn - t h6se d ' h 6 m o g l o b i n e . La s y n t h 6 s e p o r t e h l a l o i s
s u r l ' h 6 m e et s u r la g l o b i n e [1] , ces d e u x s y n - t h 6 s e s s o n t c o o r d o n n 6 e s . Apr6s que Z a m e c n i k et H o a g l a n d e u r e n t m o n t r 6 q u ' u n s y s t 6 m e a c e l l u l a i r e de fo ie 6 ta i t c a p a b l e d ' i n c o r p o r e r des a c i d e s a m i - n6s dar ts les p r o t 6 i n e s , S c h w e e t , L a l u f r o m el A l l en [2] m i r e n t au p o i n t u n sys t6n le a c e l l u l a i r e de r 6 t i c u l o c y t e s c o m p r e n a n t des r i b o s o m e s el u n e f r a c t i o n c e l l u l a i r e so lub le , la f r a c t i o n p H 5, e t c a p a b l e de s y n t h 6 t i s e r de l ' h 6 m o g l o b i n e , Ce sys- t6me , 16g6remen t m o d i f i 6 , a 6t6 u t i l i s~ clans nos p r e m i e r s t r a v a u x .
INC;UBATIO'N'S ACELrLULAIRES C R O I S E ~ S D E R E T I C U L O C Y T E S .
E n 1960, l ' o p i n i o n g6n6 ra l e 6 ta i t que le RNA r i b o s o m a l 6 ta i t p o r t e u r de l ' i n f o r m a t i o n g 6 n 6 t i q u e et d o n c r e s p o n s a b l e de la n a t u r e des p r o t 6 i n e s s y n t h 6 t i s 6 e s . Afin de c o n f i r m e r ce d o g m e , n o u s a v o n s , a v e c S c h a p i r a , D r e y f u s et Rosa , m i s e n oeuvre des s y s t 6 m e s a c e l l u l a i r e s m i x t e s de r 6 t i cu - l o c y t e s de l a p i n et de c o b a y e , en i n c u b a n t des
r i b o s o m e s d ' u n e e s p b c e et de la f r a c t i o n p H 5 de l ' a u t r e e s p 6 c e [3, 4] . Les h 6 m o g l o b i n e s de ces d e u x e sp6ces s o n t b i e n s 6 p a r 6 e s p a r 61ee t ropho- rbse s u r a m i d o n ~ p H 8,6. L o r s q u e l ' o n i n e u b e les d e u x s y s t 6 m e s m i x t e s , que l ' o n a jou te e n f in d ' i n - c u b a t i o n u n m61ange des d e u x h 6 m o g l o b i n e s se r - v a n t d ' e n t r a i n e u r et q u e l ' o n f r a c t i o n n e e n s u i t e ces h 6 m o g l o b i n e s , o n c o n s t a t e qu ' e l l e s s o n t rou te s les d e u x r a d i o a c t i v e s , a lo r s q u e d a n s u n s y s t b m e h o m o l o g u e , seu le l ' h 6 m o g l o b i n e de l ' e s p 6 e e u t i l i - s6e es t m a r q u 6 e ( t a b l e a u I) .
La c o n c l u s i o n de ce t t e e x p 6 r i e n c c 6 ta i t q u ' u n e p a r t i e de l ' i n f o r m a t i o n g 6 n 6 t i q u e n ' e s t p a s c o n t e - n u e d a n s les r i b o s o m e s e t a [ort iori d a n s le RNA r i b o s o m a l . La t h 6 o r i e d u R N A m e s s a g e r , 6 n o n c 6 c e n 1961 p a r J a c o b et M o n o d [5] , r e t i r a i t le r61e de m a t r i c e au R N A r i b o s o m a l p o u r le d o n n e r u n a u t r e RNA q u i p o u v a i t 6 t re p r 6 s e n t h o r s des r i b o s o m e s , e n a c c o r d a v e c nos r6 su l t a t s . Le RNA m e s s a g e r s e r a i t dor ic p r 6 s e n t n o n s e u l e m e n t darts les r i b o s o m e s , m a t s 6 g a l e m e n t d a n s le s u r n a g e a n t c e l l u l a i r e , ce d e r n i e r m e s s a g e r p o u v a n t c o p r 6 c i p i - t e r h p H 5. C e p e n d a n t ces e x p 6 r i e n c e s n ' 6 t a i e n t p a s c o n c l u a n t e s p o u r t r o i s r a i s o n s :
- - l a p r e u v e n ' 6 t a i t p a s f a i t e que la f r a c t i o n p H 5 6 ta i t e n t i 6 r e m e n t d 6 p o u r v u e de r i b o s o m e s ,
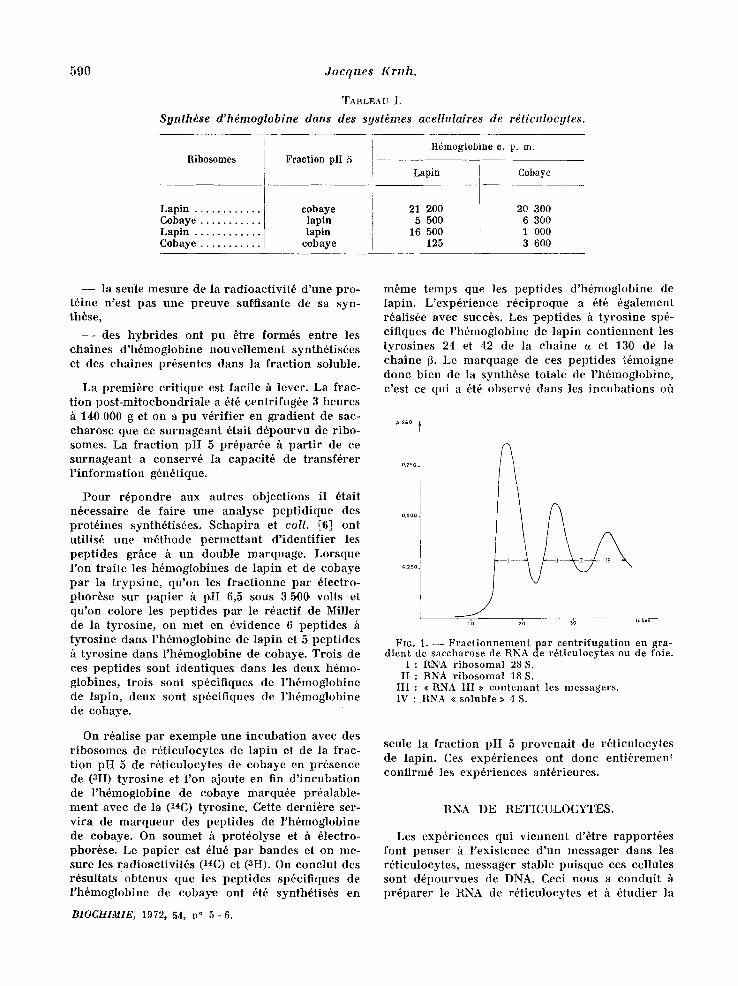
590 Jacques Kruh.
TABLEAU 1.
Synth~se d'hdmoglobine dans des syst~mes acellulaires de r~ticulocytes.
Ribosomes
Lapin . . . . . . . . . . . .
Cobaye . . . . . . . . . . .
Lapin . . . . . . . . . . . .
C o b a y e . . . . . . . . . . .
Fraction pH 5
cobaye lapin lapin
cobaye
H6m0gl0bine e. p. m.
Lapin
21 200 5 500
16 500 125
Cobaye
20 300 6 300 1 000 3 600
- - la seule mesure de la radioact iv i t6 d 'une pro- t6ine n 'est pas une p reuve suffisante de sa syn- th~se,
- - des hybr ides ont pu 6tre form6s entre les chaines d 'h6moglobine nouvel lement synth6tis6es et des chaines prdsenies dans la f rac t ion soluble.
La premi6re cr i t ique est facile h lever. La frac- t ion pos t -mi tochondr ia le a 06 centr ifug6e 3 heures
140 0'04) g e t on a pu v6r i f ier en gradient de sac- charose que ce surnageant 6tait d6pourvu de r ibo- somes. La f rac t ion pH 5 pr6par6e h pa r t i r de ce surnageant a conservd la capacit6 de t ransf6rer l ' i n fo rmat ion g6n6tique.
Pour r6pondre aux autres object ions il 6tait n6cessaire de faire une analyse pep t id ique des prot6ines synth6tis6es. Schap i ra et coll. [6] ont utilis6 une m6thode permet tan t d ' iden t i f ie r les pept ides grace fi un double marquage. Lorsque l 'on t rai te les h6moglobines de lapin et de cobaye par la t rypsine, qu 'on les f rac t ionne par 61ectro- phor6se sur pap ie r /~ pH 6,5 sous 3 500. volts et qu 'on colore les pept ides par le r6actif de Miller de la tyrosine, on met en 6vidence 6 pept ides tyros ine clans l 'h6moglobine de lapin et 5 pept ides
tyros ine dans l 'h6moglobine de cobaye. Trois de ces pept ides sont ident iques darts les deux h6mo- globines, trois sont sp6cifiques de l 'h6moglobine de lapin, deux sont sp6cifiques de l 'h6moglobine de cobaye.
On r6alise par exemple une incubat ion avec des r ibosomes de r6t iculocytes de lapin et de la frac- t ion pH 5 de r6t iculocytes de cobaye en pr6sence de (SH) tyros ine et l 'on ajoute en fin d ' incuba t ion de l 'h6moglobine de cobaye marqu6e prdalable- ment avec de la (1¢C) tyrosine. Cette derni6re ser- v i r a de marqueu r des pept ides de l 'h6moglobine de cobaye. On soumet h prot6olyse et ~ 61ectro- phor6se. Le pap ie r est 61u6 par bandes et on me- sure les radioact ivi t6s (14C) et (~H). On conclut des r6sultats obtenus que les pept ides sp6cifiques de l 'h6moglobine de cobaye ont dtd synth6tis6s en
BIOCHIMIE, 1972, 54, n ° 5- 6.
m6me temps que les pept ides d 'h6moglobine de lapin. L 'exp6r ience r6c iproque a 6t6 6galement r6alis6e avec succ6s. Les pept ides h ty ros ine spd- cifiques de l 'h6moglobine de lapin cont iennent les tyrosines 24 et 42 de la chaine o. et 13~0 de la chalne ~. Le marquage de ces pept ides t6moigne donc bien de la synthbse totale de l 'h6moglobine, c 'est ce qui a 6t6 observ6 dans les incubat ions off
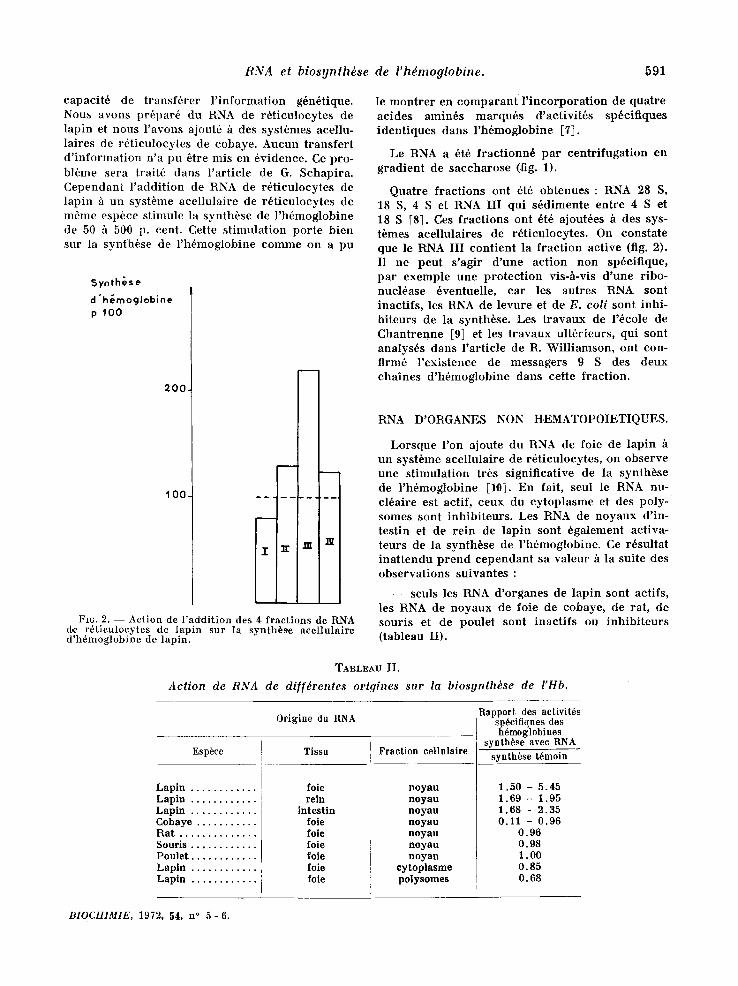
A 260 ]
0,75 (]
0,$00
0.2SO ~
J
FIr. t. - - Fractionnement par centrifugation en gra- dient de saccharose de RNA de r6ticulocytes ou de foie.
I : RN~A ribosomal 28 S. I~: RNA ribosomal 18 S.
III : ¢ I~NA III >> con~enant les messagers. IV : RNA <( solubl'e >> 4 S.
seule la f rac t ion pH 5 p rovena i t de rdt iculocytes de lapin. Ces exp6r iences ont donc ent i6rement confirm6 les exp6r iences ant6rieures.
RNA DE RETICULOCYTES.
Les exp6riences qui v iennent d'6tre rapportdes font penser h l ' ex is tence d 'un messager dans les rdticulocytes, messager stable puisque ces cellules sont ddpourvues de DNA. Ceci nous a condui t h p r6pare r le RNA de r6t icuiocytes et h 6tudier ]a
RNA et biosynthdse de l 'hdmoglobine. 591
capacit6 de t ransf6rer l ' i n fo rmat ion g6n6tique. Nous avons pr6par6 du RNA de r6t iculocytes de lapin et nous l 'avons ajout6 h des syst6mes acellu- laires de r6t iculocytes de cobaye. Aucun t ransfer t d ' in fo rmat ion n 'a pu 6tre mis en 6vidence. Ce pro- blbme sera trait6 dans l 'a r t ic le de G. Schapira . Cependant l ' add i t ion de RNA de r6t iculocytes de lapin h u n systbme acel lulaire de r6t iculocytes de m6me espbce stimule la synth~se de l 'h6moglobine de 5,0 "5 500 p. cent. Cette s t imulat ion por te bien sur la synthbse de l 'h6moglobine comme on a pu
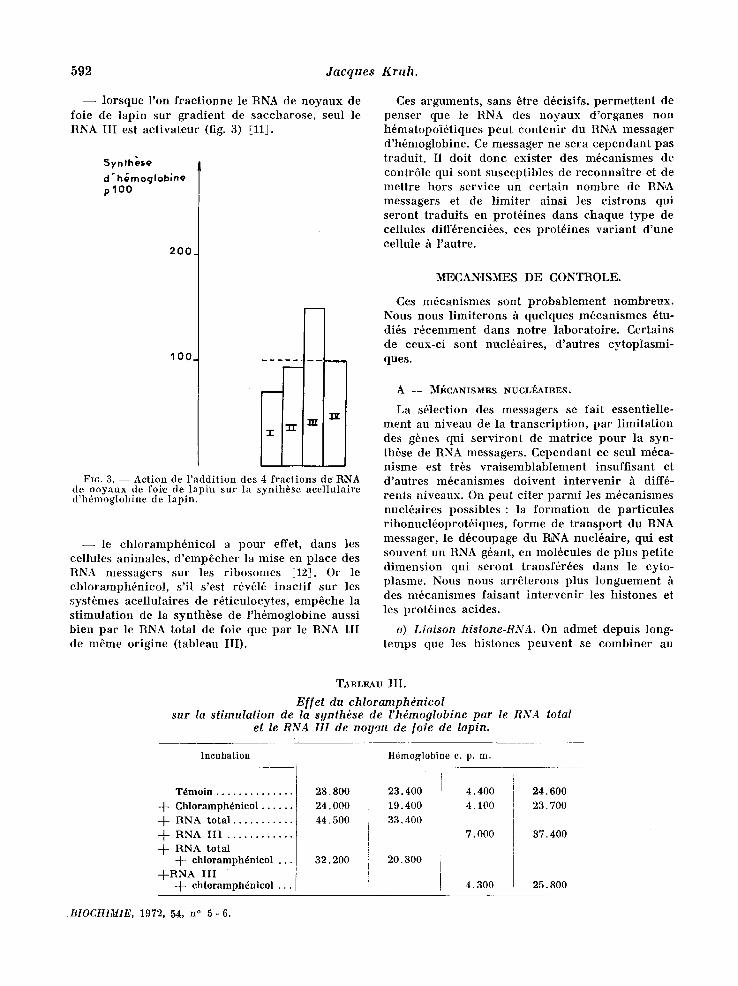
Synthese
d'h~moglobine p 100
I 200 /
'IO0-
FIG. 2 . - -Ac t i on de l'addition des 4 fractions de RNA d,c r6ticulocytes de lapin sur la synth6se acellulaire d'h6moglobine de lapin.
le mont re r en comparan t l ' i nco rpora t ion de quat re acides anain6s marqu6s d 'act ivi t6s sp6cifiques ident iques dans l 'h6moglobine [7].
Le RNA a 6t6 f ract ionn6 par cent r i fugat ion en grad ien t de saccharose (fig. 1).
Quatre f ract ions ont 6t6 obtenues : RNA 28 S, 18 S, 4 S e t RNA III qui s6dimente entre 4 S e t 18 S [81. Ces f ract ions ont 6t6 ajout6es h des sys- t~mes acel lulaires de r6t iculocytes. On constate que le ~ A III cont ient la f rac t ion active (fig. 2). Il ne peut s 'agir d 'une act ion non sp6cifique, par exemple une p ro tec t ion vis-h-vis d 'une ribo- nucl6ase 6ventuelle, car les autres RNA sont inactifs , les RNA de levure et de E. colt sont inhi- bi teurs de la synth~se. Les t ravaux de l '6cole de Chant renne [9] et les t ravaux ult6rieurs, qui sont analys6s dans Part ic le de R. Wil l iamson, ont con- firm6 1'existence de messagers 9 S des deux chaines d 'h6moglobine darts cette fract ion.
RNA D'ORGANES NON HEMATOPOIETIQUES.
Lorsque l 'on ajoute du RNA de foie de lapin h un syst~me acel lulaire de r6ticulocytes, on observe une s t imulat ion tr6s significat ive de la synth~se de l 'h6moglobine [10]. En fait, seuI le RNA nu- el6aire est actif, ceux du cytoplasme et des poly- somes sont inhibi teurs . Les RNA de noyaux d ' in- testin et de re in de lapin sont 6galement activa- teurs de la synthbse de l 'h6moglobine. Ce r6sultat ina t tendu p rend cependant sa valeur h la suite des observat ions suivantes :
- - seuls les RNA d 'organes de lapin sont actifs, les RNA de noyaux de foie de cobaye, de rat, de souris et de poulet sont inact i fs ou inhib i teurs (tableau II).
TABLEAU II.
Action de RNA de diffdrentcs origines sur la biosynth~se de l'Hb.
Esp6ce
Lapin . . . . . . . . . . . . . Lapin . . . . . . . . . . . . Lapin . . . . . . . . . . . . Cobaye . . . . . . . . . . . Rat . . . . . . . . . . . . . . Souris . . . . . . . . . . . . Poulet . . . . . . . . . . . . Lapin . . . . . . . . . . . . Lapin . . . . . . . . . . . . '
0rigine do RNA
Tissu
foie rein
intcstin foie foie foie foie foie foie
Fraction eellolaire
noyau noyau noyau noyau noyau noyau noyan
cytoplasme polysomes
Rapport des activit6s sp6cifiques dos h6moglobiues
synthbse avec RNA synthbse t6moin
1.50 - 5.45 1.69 - 1.95 1.68 - 2.35 0.11 - 0.96
0.96 0.98 1.00 0.85 0.68
I
BIOCH1MIE, 1972, 54, n ° 5- 6.
5 9 2 Jacques Kruh.
- - lorsque l 'on f rac t ionne lc RNA de noyaux de foie de lapin sur gradient de saccharose, seul le RNA III esl ac t ivateur (fig. 3) [11].
5yntht, se d'h~moqlob;ne plO0
2 0 0 .
/ 1 0 0 . . . . . . .~ . . . . .
Fro. 3. - - Action de l'addition des 4 fractions de RI~I,A de nayaux de foi~e de lapin sot la synth6se aeellulaire d'h6moglobine de lapin.
- - l e ch loramph6nico l a pour effet, dans les cellules animales, d ' emp6cher la raise en place des RNA messagers sur les r ibosomes [12]. Or le chloramph6nicol , s ' i l s 'est r6v616 inac t i f sur les systbmes acel lulaires de r6ticulocytes, emp6che la s t imulat ion de la synth6se de l 'h6moglobine aussi bien par le RNA total de foie que par le RNA III de m6me or igine (tableau III).
Ces arguments, sans ~tre d6cisifs, pe rmet ten t de penscr quc l c RNA des noyaux d 'organes non h6matopoiOiques peut conten i r du RNA messager d 'h6moglobine. Ce messager ne sera cepcndan t pas traduit . I1 dolt donc exister des m6canismcs de contrSle qui sont susceptibles de reconna i t r e et de met t re hors service un cer ta in nombre de RN~A messagers et de l imi te r ainsi les c is t rons qui seront t radui ts en prot6ines dans chaque type de cellules diff6renci6es, ces prot6ines var ian t d 'une cellule h l 'autre.
Mt~CAN'ISM~S DE CONTI~OLE.
Ces m6canismes sont p robab lement nombreux. Nous nous l imi te rons h quelques m6canismes 6tu- di6s r6cemment dans notre laboratoire . Certains de ceux-ci sont nucl6aires, d 'autres cytoplasmi- ques.
A - - M~CANISMES NUCL~AIRES.
La s61ection des messagers se fait essentiellc- ment au n iveau de la t ranscr ip t ion , par l imi ta t ion des g~nes qui se rv i ron t de mat r ice pour la syn- th6se de RNA messagers. Cependant ce seul m6ca- nisme est tr6s v ra i semblab lement insuffisant et d 'autres m6canismes doivent in t e rven i r h diff6- rents niveaux. On peui ci ter pa rmi les m6canismes nucl6aires pos s ib l e s : la format ion de par t icules r ibonucl6oprot6iques , forme de t ranspor t du RNA messager, le d6coupage du RNA nucl6aire, qui est souvent un RNA g6ant, en mol6cules de plus peti te d imension qui seront transf6r6es dans le cyto- plasme. Nous nous arr~terons plus longuement h des m6canismes faisant i n t e rvcn i r les histones et les prot6ines acides.
a) Liaison histone-RNA. On admet depuis long- temps que les histoncs peuvent se combine r au
TABLEAU ]II.
Effet du chloramph~nicol sur la stimulation de la synth~se de l'hdmoglobine par le RNA lotal
et le RNA 1H de noyaa de foie de lapin.
Incubation H6moglobine c. p. m.
T~moin . . . . . . . . . . . . . . -~- Chloramph6nicol . . . . . . -~- RNA total . . . . . . . . . . . + RNA m . . . . . . . . . . . .
~- RNA total -~ ehloramph6nicol . . .
-~-RNA I l l I -4- chloramph~nieol . . .
28.800 24.000 44.500
32.200
23.400 4.400 19.400 4.100 33.400
7.000
20.800
4.300
24.600 23.700
37.400
25.800
BIOCH1MIE, 1972, 54, n ° 5- 6.

RNA et biosynth~se de l 'hdmoglobine. 593
DNA, Ies r6gions de DN.A ainsi combin6es sont inactives. Nous avons montr6 que les histones peuvent 6galement agir h u n autre niveau. Lors- que l 'on ajoute des histones h u n syst~me acellu- laire de r6ticulocytes on inh ibe la synth~se de l 'h6moglobine [13]. Ceei est dfi au fair que les histones sont capables de se combiner aux RNA. L'existence d 'une telle l iaison a pu ~tre raise en 6videnee d i rec tement en ut i l i sant du RNA radio- aetif et en reeuei l lant le eomplexe RNA-histones sur un fiItre mil l ipore. Ainsi 100 ~g d 'h is tone peu- vent se combiner h 60 ~g de RNA [14]. Cepen- dant cette l ia ison ne se forme pas si l 'h is tone a 6t6 pr6alablement phosphoryl6e par I 'ATP en pr6- senee d 'une prot6ine kinase.
Ainsi les histones peuvent s 'associer non seule- ment au DNA mats 6galement aux RNA et elles peuvent ainsi jouer un r61e dans la s61ection des messagers. Cependant les histones sont ident iques dans la p lupar t des tissus et il est impossible de faire jouer h e l l e s seules un rSle hautement sp6ci- fique.
b) Prot~ines acides de la chromatine. La chro- marine cont ient 6galement des prot6ines aeides (ou prot6ines non histones). Celles-ci ont donn6 lieu ~ des recherches dans diff6rents plans qui ont permis de mettre en ~videnee leur r61e fonda- menta l dans les cellules diff6renci6es :
les prot6ines aeides sont sp6cifiques du type cellulaire, ee qui a ~t~ 6tabli i nd6pendammen t dans trois laboratoires, h l ' a ide de l'61ectropho- rbse en gel de po lyaery lamide [15, 16, 17].
ces prot6ines, con t ra i rement aux histones, pr6sentent un renouvel lement rapide. Lorsque l 'on compare la vitesse de renouvel lement des pro- t6ines individuel les par la m6thode du double marquage, on trouve des modif icat ions de cer- taines synthbses au cours de la r6g6n6ration du foie aprbs h6pateetomie, et des per turba t ions im- portantes de ces syntheses dans les cellules asci- t iques d 'h6patoma de Zajdela ~18] ;
- - les prot6ines aeides sont capables de se com- b ine r aux histories. Le complexe ainsi form6 pr6- sente vis-h-vis du RNA [14, 19] et vis-h-vis du DNA [20] une affinit6 for tement diminu6e. Ainsi les prot6ines acides, en se combinan t aux histones, peuvent moduler la format ion de la l ia ison histone-RNA aussi b ien que histone-DNA, et modi- fier la synthbse et la raise en service des RNA dans la eellule ;
- - les prot6ines acides ont 6t6 fract ionn6es pa r chromatographies sur D,EAE-cellulose et sur phos- phocellulose. Ceci a permis d ' isoler et d ' ident i f ie r deux de ces prot6ines. L 'une de ces prot6ines est
une phosphoprot6ine qui pr6sente une forte affi- nit6 vis-fi-vis des histones. On a pu mont re r qu'elle 16ve en par t ie Fact ion inh ib i t r i ce des histones vis- h-vis de la synth6se du RNA en pr6senee de R~NA polym6rase (tableau IV). L 'autre prot6ine est une prot6ine kinase qui n ' a pas d 'ac t ion directe sur la synth6se du RNA mats qui est capable de phos- phory le r les histones ainsi que la phosphoprot6ine (tableau Y) [21] ;
- - les prot6ines acides peuvent se combiner aux glucocorticoides : la cort icost6rone se combine aux prot6ines acides du foie de rat. Cette l ia ison est sp6cifique, elle n 'est pas d6plac6e par d 'autres st6rols ou st6ro~des. La constante de dissociat ion est de 5 X 10 .9 M, ce qui cor respond aux concen- t rat ions physiologiques [22].
De l 'ensemble de ces observat ions nous pouvons conclure que les prot6ines acides peuvent inter- ven i r an n iveau du g6nome pour contr61er, par l ' in te rm6dia i re des histones, la synth6se et la raise en service des RNA. Les prot6ines acides qui sont sp6cifiques du type cellulaire, se renouvel lent ra- p idement , la vitesse de nouvel lement 6tant modi- fi6e lors de la r6g6n6ration ou dans le cancer. I1 est de plus vraisemblable que certaines de ces pro- pri6t6s soient influenc6es par divers l igands, en par t icu l ie r hormonaux.
U - - M~CANISMES CYTOPL&SMIQUES.
Nous s ignalerons le r61e qui pout Ore jou6 par les RNA de t ransfer t qui peuvent moduler la syn- th~se de certaines prot6ines, par les facteurs d ' in i t i a t ion ou d 'autres facteurs r ibosomaux. Par ailleurs, certaines observat ions font penser qu ' i l existe dans le cytoplasme certains inh ib i teurs solu- bles sp6cifiques du type cellulaire. L 'addi t ion de fract ion pH 5 de cytoplasme de foie h u n syst~me acellulaire de r6ticulocytes inh ibe la synth~se de l 'h6moglobine, l ' add i t ion de f ract ion pH 5 de r6ti- culocytes h u n syst~me acellulaire de foie inh ibe la synth~se de la s6rumalbumine . Les compos~s responsables de ces inh ib i t ions ont 6t6 identifi6s comme Oant des prot6ines. Ces prot6ines ont 6t6 isol6es par double chromatographie sur CM- et DEAE-cellulose [23, 24, 25]. Elles agissent sur le syst~me h6t6rologue et sont sans act ion sur le systbme homologue (tableau VI).
Le m6canisme d 'ac t ion de ces inh ib i teurs a 0 6 6tudi6. Ces prot6ines sont d6pourvues d 'ac t ion nucl6olytique non sp6eifique et d 'act ion prot6o- lytique. Lorsque l 'on fair agir ehacun de ces inhi - bi teurs sur le RNA de foie et qu 'on analyse celui- ci sur un gradient de saecharose, on constate que l ' i nh ib i t eur de r6ticulocytes a provoqu6 une forte
BIOCHIMIE, 1972, 54, n ° 5- 6.

5 9 4 Jacque s K r u h .
TABLEAU IV.
Action de la phosphoprotdine el de Ia protdine kinase chromatinienne sur la synth~se de RNA en presence de nucldohistone.
I Nature de la matrice
DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . DNA ~- histone . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ] DNA -]- his tone ~- prot6ine kinase . . . . . . . . . . . . ! DNA -~- histone q- phosphoprotd ine . . . . . . . . . . . :
Incorporation de laH) UMP darts le RNA (e. p. m.)
1 240 0 480 0 450 0 802
TABLEAU ¥ .
Activild prot~ine kinasique des prot~ines acides du foie et de deux de ses [factions.
Prot6ine chromatinienne
Prot6ines aeides totates . . . . . . . . . . . . . . . . . . . . . . . Prot6ine kinase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Phosphoprot6 ine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Phosphoprot6ine -~- protdine kinase . . . . . . . . . . . . Phosphoprot6ine -~- histone . . . . . . . . . . . . . . . . . . . Prot~ine kinase ~- histone F2b . . . . . . . . . . . . . . . . .
A ctivitb, kinasique (3,2 p incorpor6 c. p. m.)
2 300 194 290
2 100 180
2 000
TABLEAU V[.
Spdcificitd de l'aciion des inhibiteurs.
Origine de l ' iahibiteur
0 Foie
R~ticulocyte
! Radioactivit6 c. p. m. i I i Seromalbumine (systbme acellulaire du Ioie) I l
| 2 i
1 800 1 625 2 920 1 310
418 252
h6moglobine (systbme acellulaire de
rSticulocytes)
I 2
37 000 44 000 5 450 5 500
34 20O 45 000
TABLEAU VI I .
Action de l ' inhibiteur de foie el du RNA de r~ticulocytes sur la synth~se acellulaire de l'hdmoglobine.
Milieu d'iacubation
T~moin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Jr- 400 l~g R N A r~ticulocyte . . . . . . . . . . . . . . . . . . -~- 200 tzg inhibi teur de foie . . . . . . . . . . . . . . . . . . -~ 400 tzg R N A ~- 200 ~g inhibi teur . . . . . . . . . . ~- 400 ~g RNA incub~ avec l ' inh ib i teur puis purifi~
It6moglobine synth6tis~e
18 000 70 000
3 010 2 950
66 000
BIOCH1MIE, 1972, 54, n ° 5 - 6.

R N A et b io syn th~se de l ' hdmoglob ine .
TABLEAU VIII .
Liaison entre les RNA de tote el l ' inhibiteur de r~ticulocytes.
Nature du BNA
28 S 18 S
RNA I I I 4 S
Radioaclivit6 de {00 ~g de RNA
c. p. m.
7 460 3 250 5 000 2 740
Radioactivit6 du complexe
inhibiteur-RNA I RNA combin6 (en ~g) [fi 100 ~g d'inhibiteur
670 450
1 200 225
9 .0 13.8 24.0
8 .2
595
p e r t u r b a t i o n a l o r s que l ' i n h i b i t e u r de fo ie a u n e a c t i o n b e a u e o u p m o i n s n o t a b l e : la p e r t u r b a t i o n se s i t u e e s s e n t i e l l e m e n t d a n s la r 6 g i o n d u g r a d i e n t o h s o n t l oca l i s6 s la p l u p a r t des RN A m e s s a g e r s . Cec i p e u t ~ t re dfi h u n e a c t i o n n u c l 6 o l y t i q u e sp6c i - f ique de l ' i n h i b i t e u r ou fi u n e c o m b i n a i s o n de l ' i n h i b i t e u r a v e c c e r t a i n s RNA. N o u s a v o n s p r6 - p a r 6 d u RNA de r 6 t i c u l o c y t e s qu i s t i m u l e la syn - t h6se a c e l l u l a i r e de l ' h 6 m o g l o b i n e . Si l ' o n p r6 - i n e u b e ee BNA a v e c l ' i n h i b i t e u r de fo ie et q u ' o n le p u r i f i e , o n c o n s t a t e q u ' i l a g a r d 6 s o n ac t iv i t6 st i- m u l a n t e ( t a b l e a u VII ) , ce qu i n ' a u r a i t p a s 6t6 ]e cas si l ' i n h i b i t e u r a v a i t agi p a r a c t i o n n u e l 6 o l y - t i que . U n e m 6 t h o d e d i r e c t e a p e r m i s de m e t t r e e n 6 v i d e n e e u n e l i a i s o n e n t r e l ' i n h i b i t e u r et le RNA r a d i o a c t i f . L ' i n h i b i t e u r 6 ran t i n s o l u b l e h p H 5, o n p e u t r e c u e i l l i r le c o m p l e x e s u r m i l l i p o r e . L ' i n h i - b i t e u r de r 6 t i e u l o c y t e s a l ' a f f in i t6 la p l u s g r a n d e p o u r le RNA I I I de fo ie qu i e o n t i e n t la p l u p a r t des m e s s a g e r s ( t a b l e a u VI I I ) , 100 ~g d ' i n h i b i t e u r de r 6 t i c u l o c y t e s p e u v e n t s ' a s s o c i e r h 24 ~g de RNA de f o i e [26].
L ' e n s e m b l e de ces t r a v a u x p e r m e t de p e n s e r
q u ' i l ex i s t e d a n s les ce l lu le s d i f f 6 r e n c i 6 e s u n e s6 r i e
de m 6 c a n i s m e s a g i s s a n t s u c c e s s i v e m e n t e n t r e
l ' 6 t a p e de la t r a n s c r i p t i o n et ce l le de l a t r a d u c t i o n
et qu i p e r m e t de s 6 1 e c t i o n n e r les RNA m e s s a g e r s qu i s e r o n t s y n t h 6 t i s 6 s p u t s t r a d u i t s . L a c o n n a i s -
s a n c e que n o u s a v o n s de la s t r u c t u r e de l ' h 6 m o -
g l o b i n e , les p r o p r i 6 t 6 s r e m a r q u a b l e s des s y s t 6 m e s
de s y n t h 6 s e in vitro de ce t t e p r o t 6 i n e o n t p e r m i s d ' u t i l i s e r ce m o d u l e p r i v i l 6 g i 6 d e ce l lu l e h a u t e -
m e n t d i f f 6 r e n c i 6 e c a p a b l e de s y n t h 6 t i s e r a c t i ve - m e n t u n e p r o t 6 i n e p a r f a i t e m e n t c o n n u e .
BIBLIOGRAPHIE.
1. Kruh, J a Borsook, H. (1956) J. Biol. Chem., 220, 905.
2. Sch~cett, R., Lamfrom, H. a Allan, E. (1958) Proc. Natl. Acad. Set. V.S., 44, 1029.
3. Kruh, J., Rosa, J., Dreyfus, J. C. a Schapira, G. (19,61) Biochim. Biophys. Acta, 49, 509.
4. Kruh, J., Dreyfus, J. C., Rosa, J. a Schapira, G. (1962) Biochim. Biophys. Aeta, 55, 690.
5. Jacob, F. a Monod, J. (1961) J. Mol. Biol., 3, 318. 6. Schapira, G., Padieu, P., Mateknia, N., Kruh, J. a
Dreyfus, J. C. (1966) J. Mol .Biol., 20, 427. 7. Kruh, J., Dreyfus, J. G. a Schapira, G. (1964) Bio-
chim. Biophys. Acta, 87, 253. 8. Kruh, J., Schapira, G., Lareau, J. a Dreyfus, J. C.
(1964) Biochim. Biophys. Acta, 87, 669. 9. Chant renne , H., Burny, A. a Marbaix, G. (1967)
Prog. Nucleic Acid Bes., 7, 173. 10. Kruh, J., Dreyfus, J. G. a Schapira, G. (1.964) Bio-
chim. Biophys. Acta, 91, 494. 11. Kruh, J., Dreyfus, J. C. a Schapira, G. (1966) Bio-
ehim. Biophys. Acta, 114, 384. 12. Weisberger, A. S., Wolfe, S. et Armen t rau t , S.
(196.41) J. Expl. Med., 120, 161. 13. Kruh, J. a Labie, D. (1968) Biochim. Biophys. Aeta,
161, 518. t4. Dastugue, B., Tichonieky, L., Hanoune, J. a Kruh,
J. (197'0) FEBS letters, 8, 133. 15. Dastugue, B., Tiehonicky, L., Peni t -Soria , J. a
Kruh, J. (197.0) Ball. Soc. Chim. Biol., 52, 391. 16. Loeb, J. E. a Creuset, C. (1969) FEBS letters, 5, 37. 17. Gilmour, R. S. ,¢ Paul, J. (19'70) FEBS Letters, 9, 242. 18. Dastugue, B., Hanoune, J. a Kruh, J. (1971) FEBS
Letters, 19, 65. 119. Kruh, J., Tiehonicky, L. a Wa jeman , H. (1969) Bio-
chim. Biophys. Acta, 195, 549. 20. Kamiyama, M. a Wang, T. Y. (1971) Biochim. Bio-
phys. Aeta, 228, 563. 2I. Kamiyama, M., Dastugue, B. a Kruh, J. (1971) Bio-
ehim. Biophys. Res. Comm., 44, 1345. 22. Dastugue, B., Defer, N. & Kruh, J. (1971) FEBS Let-
ters, 16, 121. 2'3. Kruh, J. • Levy, F. (1'967) Biochim. Biophys. Acta,
145, 460. 24. Levy, F., Tiehonieky, L. a Kruh, J. (1970) Biochim.
Biophys. Acta, 209, 521. 25. Levy, F., Tichonicky, L. & Kruh, J. (1'971) Biochimie,
53, 671. 26. Levy, F., Tichonieky, L. a Kruh. J. (1972) Bio-
chimie, 54, 63.
BIOCH1MIE, 1972, 54, n ° 5 - 6.





























