Embed Size (px)
Citation preview

Exp Brain Res (1991) 84:229-232
Experimental Brain Research �9 Springer-Verlag 1991
Research Note
Serotonin and cervical respiratory motoneurones: intracellular study in the newborn rat brainstem-spinal cord preparation D. Morin*, R. Monteau, and G. Hilaire
Equipe Biologie des Rythmes et du D6veloppement, URA-CNRS 0205, Facult~ des Sciences et Techniques St J~r6me. F-13397 Marseille C6dex 13, France
Received August 6, 1990 / Accepted October 24, 1990
Summary. Previous experiments performed in the in vi- tro newborn rat brainstem-spinal cord preparation re- ported that the addition of serotonin (5-HT, 30-50 pM) to the bathing medium induced increases in the respira- tory frequency and a large tonic discharge on all the cervical ventral roots. The aim of the present work was to define whether the 5-HT-induced tonic discharge in- volved respiratory or non-respiratory motoneurones. In- tracellular recordings demonstrated that cervical (C2) motoneurones (n=27) were depolarized by 5-HT but that the 5-HT-induced tonic discharge was mainly due to recruitment of silent motoneurones (n=18) which fired permanently (15/18; 17+3 Hz) under 5-HT. The respiratory motoneurones (n = 9) retained a phasic inspi- ratory discharge (5/9), even if some (4./9) occasionally exhibited a few spikes during expiration. Therefore, it is concluded that the 5-HT-induced tonic discharge is unlikely to have functional significance in respiration.
Key words: Respiration - Serotonin Motoneurone - In vitro - Newborn rat
Introduction
The isolated newborn rat brainstem-spinal cord prepara- tion has been shown to retain rhythmic activity in vitro which appears as bursts of action potentials on cranial nerves and cervical ventral roots. This activity is consis- tent with a respiratory function since rhythmic dis- charges i) occur on phrenic roots and thoracic respirato- ry muscles (Suzue 1984), ii) are sensitive to lung vagal afferents (Murakoshi and Otsuka 1985) as well as pH and CO2 levels (Harada etal . 1985; Monteau etal .
Offprint requests to: D. Morin (address see above)
1990c), and iii) involve medullary neurones of the ven- tral respiratory group (Hilaire et al. 1990). This in vitro preparation appears to be a useful tool for analysing the pharmacology of the central respiratory activity since drugs added to the bathing medium diffuse within the nervous tissue and may elicit respiratory changes which are independent from the periphery. Hence, sero- tonin (5-HT) added to the bathing medium (30 ~tM) in- creases the respiratory frequency with a short latency (less than 1 rain) and evokes a large tonic discharge in 2-3 min on C1-C4 ventral roots (Monteau et al. 1990b). Frequency changes involve 5-HT1 receptors located at the medullary level whereas the tonic discharge recorded on cervical ventrol roots involve 5-HT 2 receptors located in the spinal cord (Morin et al. 1990). However, since the recordings originated from the whole ventral roots and splitting the roots into thin filaments was impossible to perform, it was not determined whether 5-HT tonic activation was due to a permanent firing of previously active respiratory motoneurones or to the recruitment of previously silent non-respiratory motoneurones. Al- though such a large tonic discharge could resemble an apneustic breathing pattern (Stella 1938), this hypothesis is unlikely since i) apneustic breathing seldom occurs in rats (Monteau et al. 1989, 1990a), and ii) 5-HT tonic activation is still evoked on an isolated spinal cord (Monteau et al. 1990b; Morin et al. 1990). The aim of the present work was to analyse by intracellular record- ings the pattern of firing of spinal motoneurones in the newborn rat brainstem-spinal cord preparation under 5-HT superfusion. Experiments were performed at the C2 level because this ventral root is easily identified and readily accessible to antidromic stimulation. Results demonstrate that some respiratory motoneurones may occasionally fire a few spikes during expiration but that the 5-HT-induced tonic activation is mainly due to the recruitment of previously silent motoneurones which be- come permanently active.
230
Material and methods
Newborn rats, 0-3 days old, were anaesthetized with ether and decerebrated. Another section was performed at the level of the last cervical vertebra. The skin, muscles and bones surrounding the central nervous system were rapidly removed. The preparation (i.e. pons, medulla and cervical spinal cord) was then placed in a 1.5 ml chamber perfused with artificial cerebrospinal fluid identi- cal to that used previously (Hilaire et al. 1990; Monteau et al. 1989; Morin et al. 1990). warmed to 27_+0.5 ~ C, equilibrated with 95% 02 and 5% CO2 (pH 7.3) and permanently drained by suction (3 ml/min). Cranial and vertebral bones were removed with thin forceps and the brainstem was fixed with the ventral surface up- wards.
Electrical activity of the left C2 ventral root (and occasionaly the right) was recorded using suction electrodes. Signals were fil- tered (5 3000 Hz), amplified (Neurolog System, Digitimer), and fed to a leaky integrator, oscilloscopes and a paper recorder (Gould TA 2000). The changes in the respiratory activity elicited by 5-HT medium appear simultaneously on both left and right cervical ven- tral root recordings which showed the same time course and re- sponse to 5-HT (Monteau et al. 1990b). The right C2 ventral root was sucked into an electrode for stimulating purposes (200 ps, 25 100 gA) via an isolation unit and digital stimulator (WPI 830); the ipsilateral C2 segment was explored with micropipettes (3 M potassium acetate, impedance 60 to 100 Mr!_) moved by a motor- ized microdrive (1 I-tm step) from the ventral surface to a maximal depth of 400 gin. The DC signal was fed to a laboratory made microelectrode amplifier, oscilloscope, paper recorder and stored on tape. Input membrane impedance was calculated fi'om the volt- age changes elicited by injection of positive and negative pulses of current (t00 ms, 100 400 pA). Results were given as mean plus or minus SEM and differences were taken to be statisticaly signifi- cant at P < 0.05, using Student's t-test to make paired and unpaired comparisons.
Serotonin (Sigma) and e-methyl-serotonin (RB[) were dis- solved in the bathing medium (30-50 gM) and applied by super- fusion.
Results
In t race l lu la r record ings were o b t a i n e d f rom 27 cervical neurones ident i f ied as m o t o n e u r o n e s on the basis o f their a n t i d romic ac t iva t ion fo l lowing C2 vent ra l r oo t s t imula- t ion (Fig. 1 B and 2 B). Twin pulse s t imula t ion lead ing to d i s soc ia t ion be tween soma to -den t r i t i c spike and spike o f the ini t ial segment was used to assess the somat ic or igin o f the recordings . Al l the r epo r t ed neurones sat- isfied the three fo l lowing cr i te r ia : i) m e m b r a n e po ten t i a l o f a t least - 55 mV, ii) abi l i ty to del iver full an t i d romic spikes and iii) abi l i ty to emit spike(s) e i ther spon t ane ous - ly or dur ing inject ion o f depo la r i z ing pulses.
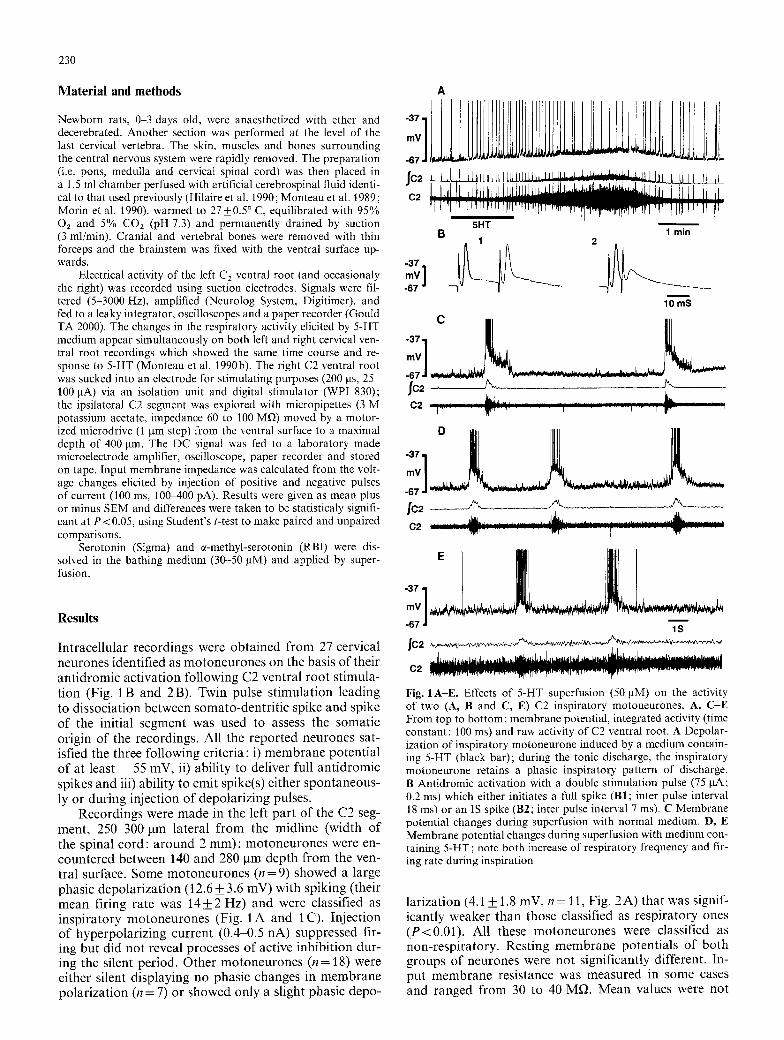
Record ings were m a d e in the left pa r t o f the C2 seg- ment , 250 300 lam la tera l f rom the midl ine (width o f the spinal c o r d : a r o u n d 2 mm) ; m o t o n e u r o n e s were en- coun te red be tween 140 and 280 g m dep th f rom the ven- t ral surface. Some m o t o n e u r o n e s (n = 9) showed a large phas ic d e p o l a r i z a t i o n (12.6 _+ 3.6 mV) wi th sp ik ing ( their mean fir ing ra te was 14_+2 Hz) and were classif ied as i n sp i r a to ry m o t o n e u r o n e s (Fig. 1 A and 1 C). In jec t ion o f hype rpo la r i z ing cur ren t (0.4-0.5 nA) suppressed fir- ing bu t d id not reveal processes o f active inh ib i t ion dur- ing the silent per iod . Othe r m o t o n e u r o n e s (n = 18) were ei ther si lent d i sp lay ing no phasic changes in m e m b r a n e po l a r i za t i on (n = 7) or showed only a slight phas ic depo-
A
=11 llJ - 6 7 a ' ~ r ~ . . . . . . . . . . . . . . .
J'c2 c2 kjCIIIIIIl,-'!)::"':~':!,l, LJ k L_L~L
5 H T B 1 m i n
-37 ~ 2 my] -67
10 mS
/C2 - - / % J~. L . . . . . . . . . . K . . . . . .
C 2 F : : : : : : ' r " " - I ' ] ' [ F '
mV i
-67
0 2 . m m ~ l ~= . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . , 'w ~ . . . . . "m r . . . . . . . . I . . . . . . . . . . . ~ . . . . . .
-67 a 1 S J'C2 ~.~.~.~','~w~.~.~/'~,.~'~'~ ~'~'~J'~-~'~'~'~'~'~~'~
" i ] [ i l l , , n -~ , . . . . ~ , r , !
Fig. 1A-E. Effects of 5-HT superfusion (50 pM) on the activity of two (A, B and C, E) C2 inspiratory motoneurones. A, C-E From top to bottom : membrane potential, integrated activity (time constant: 100 ms) and raw activity of C2 ventral root. A Depolar- ization of inspiratory motoneurone induced by a medium contain- ing 5-HT (black bar); during the tonic discharge, the inspiratory motoneurone retains a phasic inspiratory pattern of discharge. B Antidromic activation with a double stimulation pulse (75 gA; 0.2 ms) which either initiates a full spike (B1; inter pulse interval 18 ms) or an IS spike (B2; inter pulse interval 7 ms). C Membrane potential changes during superfusion with normal medium. D, E Membrane potential changes during superfusion with medium con- taining 5-HT; note both increase of respiratory frequency and fir- ing rate during inspiration
la r iza t ion (4.1 _+ 1.8 mV, n = 11, Fig. 2 A ) tha t was signif- icant ly weaker than those classif ied as r e sp i r a to ry ones ( P < 0 . 0 1 ) . Al l these m o t o n e u r o n e s were classif ied as non- re sp i r a to ry . Res t ing m e m b r a n e po ten t i a l s o f bo th g roups o f neurones were no t s ignif icant ly different . In- pu t m e m b r a n e resis tance was measu red in some cases and ranged f rom 30 to 40 MfL M e a n values were not
231
significantly different between both groups but the sam- ple was small (n = 6).
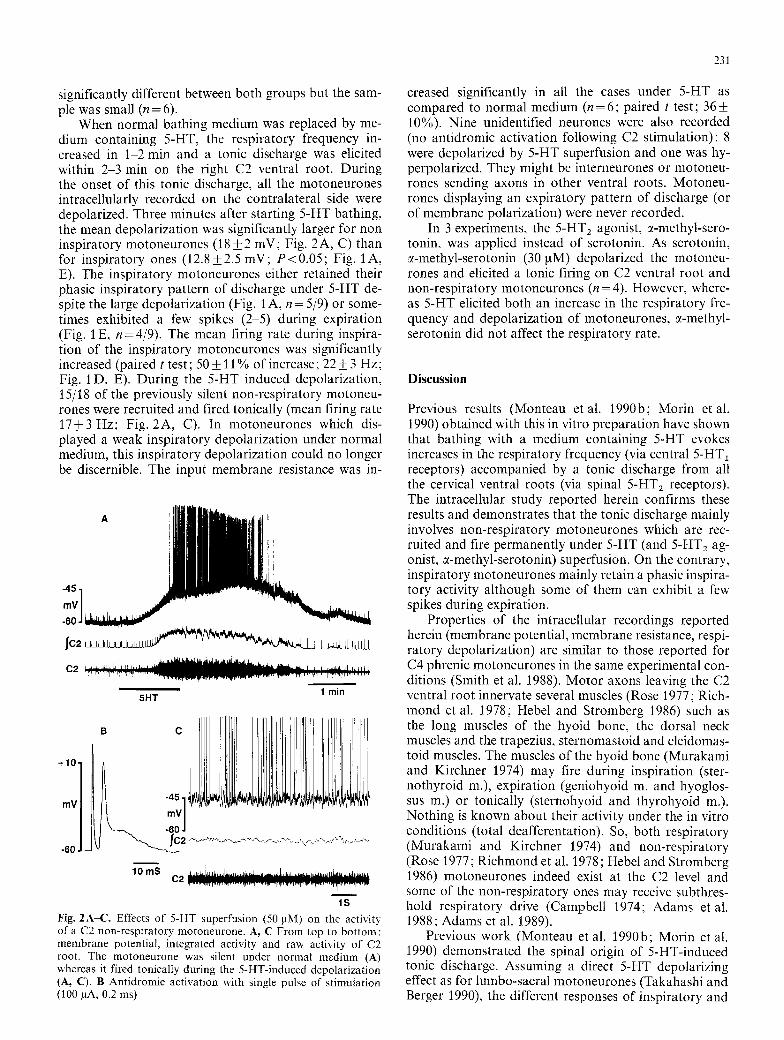
When normal bathing medium was replaced by me- dium containing 5-HT, the respiratory frequency in- creased in 1-2 rain and a tonic discharge was elicited within 2-3 min on the right C2 ventral root. During the onset of this tonic discharge, all the motoneurones intracellularly recorded on the contralateral side were depolarized. Three minutes after starting 5-HT bathing, the mean depolarization was significantly larger for non inspiratory motoneurones (18 4-_ 2 mV; Fig. 2 A, C) than for inspiratory ones (12.8+2.5 mV; P<0.05; Fig. 1A, E). The inspiratory motoneurones either retained their phasic inspiratory pattern of discharge under 5-HT de- spite the large depolarization (Fig. 1 A, n = 5/9) or some- times exhibited a few spikes (2-5) during expiration (Fig. 1 E, n = 4/9). The mean firing rate during inspira- tion of the inspiratory motoneurones was significantly increased (paired t test; 50 _+ 11% of increase; 22 _ 3 Hz; Fig. 1 D, E). During the 5-HT induced depolarization, 15/18 of the previously silent non-respiratory motoneu- rones were recruited and fired tonically (mean firing rate 17+ 3 Hz: Fig. 2A, C). In motoneurones which dis- played a weak inspiratory depolarization under normal medium, this inspiratory depolarization could no longer be discernible. The input membrane resistance was in-
i u
' i i ! ':
..45,
-60,
C2 5HT 1 min
B (
mV] -45.60m~
-60 10 mS C2 """~r"v'"v"'q"~"'~-r'~"r"r"r"r'~"~""nT~"~'~"
1S Fig. 2A-C. Effects of 5-HT superfusion (50 pM) on the activity of a C2 non-respiratory motoneurone. A, C From top to bottom: membrane potential, integrated activity and raw activity of C2 root. The motoneurone was silent under normal medium (A) whereas it fired tonically during the 5-HT-induced depolarization (A, C). B Antidromic activation with single pulse of stimulation (100 laA, 0.2 ms)
creased significantly in all the cases under 5-HT as compared to normal medium (n = 6: paired t test; 36 4- 10%). Nine unidentified neurones were also recorded (no antidromic activation following C2 stimulation): 8 were depolarized by 5-HT superfusion and one was hy- perpolarized. They might be interneurones or motoneu- rones sending axons in other ventral roots. Motoneu- rones displaying an expiratory pattern of discharge (or of membrane polarization) were never recorded.
In 3 experiments, the 5-HT2 agonist, ~.-methyl-sero- tonin, was applied instead of serotonin. As serotonin, :~-methyl-serotonin (30 gM) depolarized the motoneu- rones and elicited a tonic firing on C2 ventral root and non-respiratory motoneurones (n = 4). However, where- as 5-HT elicited both an increase in the respiratory fre- quency and depolarization of motoneurones, c~-methyl- serotonin did not affect the respiratory rate.
Discussion
Previous results (Monteau etal. 1990b; Morin etal. 1990) obtained with this in vitro preparation have shown that bathing with a medium containing 5-HT evokes increases in the respiratory frequency (via central 5-HT1 receptors) accompanied by a tonic discharge from all the cervical ventral roots (via spinal 5-HT 2 receptors). The intracellular study reported herein confirms these results and demonstrates that the tonic discharge mainly involves non-respiratory motoneurones which are rec- ruited and fire permanently under 5-HT (and 5-HT2 ag- onist, c~-methyl-serotonin) superfusion. On the contrary, inspiratory motoneurones mainly retain a phasic inspira- tory activity although some of them can exhibit a few spikes during expiration.
Properties of the intracellular recordings reported herein (membrane potential, membrane resistance, respi- ratory depolarization) are similar to those reported for C4 phrenic motoneurones in the same experimental con- ditions (Smith et al. 1988). Motor axons leaving the C2 ventral root innervate several muscles (Rose 1977; Rich- mond et al. 1978; Hebel and Stromberg 1986) such as the long muscles of the hyoid bone, the dorsal neck muscles and the trapezius, sternomastoid and cleidomas- toid muscles. The muscles of the hyoid bone (Murakami and Kirchner 1974) may fire during inspiration (ster- nothyroid m.), expiration (geniohyoid m. and hyoglos- sus m.) or tonically (sternohyoid and thyrohyoid m.). Nothing is known about their activity under the in vitro conditions (total deafferentation). So, both respiratory (Murakami and Kirchner 1974) and non-respiratory (Rose 1977; Richmond et al. I978; Hebel and Stromberg 1986) motoneurones indeed exist at the C2 level and some of the non-respiratory ones may receive subthres- hold respiratory drive (Campbell 1974; Adams et al. 1988; Adams et al. 1989).
Previous work (Monteau et al. 1990b; Morin et al. 1990) demonstrated the spinal origin of 5-HT-induced tonic discharge. Assuming a direct 5-HT depolarizing effect as for lumbo-sacral motoneurones (Takahashi and Berger 1990), the different responses of inspiratory and

232
n o n - r e s p i r a t o r y C2 m o t o n e u r o n e s a p p e a r diff icult to ex- plain . Both g roups were depo la r i zed by 5 -HT bu t inspi- r a to ry ones s ignif icant ly less t han others. So, i n sp i r a to ry m o t o n e u r o n e s migh t be less sensit ive to 5 -HT in agree- men t wi th the weak 5 -HT effect on phren ic m o t o n e u - rones (Lal ley 1986). I n s p i r a t o r y m o t o n e u r o n e s might be less depo la r i zed and might re ta in a phas ic d ischarge unde r 5 -HT because an act ive inh ib i t ion dur ing expi ra- t ion cou ld coun te rac t 5 -HT depo la r i za t ion . A n inhibi- t ion dur ing exp i r a t ion exists in vivo in the adu l t cat (Berger 1979), bu t is un l ike ly in the in v i t ro p r e p a r a t i o n since i) the ac t iv i ty o f exp i r a to ry neurones was never obse rved in this in v i t ro p r e p a r a t i o n ei ther in the medu l l a (Hi la i re et al. 1990) or in the spinal co rd and ii) impose d h y p e r p o l a r i z a t i o n o f r e sp i r a to ry m o t o n e u r o n e s d id not reveal features o f act ive exp i r a to ry inhibi t ion . The grea ter 5 -HT- induced d e p o l a r i z a t i o n on the non- re sp i r a - to ry m o t o n e u r o n e s might be re la ted to an o rgan i sed p ro - cess involv ing a spinal ne twork in re la t ion to pos tu ra l / l o c o m o t o r funct ions which have been a l r eady r epo r t ed in this p r e p a r a t i o n (Smith and F e l d m a n 1987).
F r o m the results r e p o r t e d herein, it r emains imposs i - ble to ident i fy m o r e precisely the mechan i sms involved in the 5 -HT- induced tonic discharge. However , in t racel- lu lar record ings d e m o n s t r a t e d tha t the ac t iva t ion o f C2 m o t o n e u r o n e s by 5 -HT main ly involved n o n - r e s p i r a t o r y units . So, to conclude , the 5 -HT- induced tonic d i scharge on cervical vent ra l roo ts resul t ing f rom ac t iva t ion o f spinal 5-HT2 receptors ( M o n t e a u et al. 1990b; M o r i n et al. 1990) is un l ike ly to relate specif ical ly to a func t ion in resp i ra t ion .
Acknowledgements. The expert assistance of A.M. Lajard and M. Manneville in this study is gratefully acknowledged. We wish to thank Dr. A.D. Miller for valuable reviewing of the manuscript. This work was supported by the CNRS (URA 205), the INSERM (grant 886006), and a grant from the "Nakre et Vivre" Founda- tion.
References
Adams L, Datta AK, Guz A (1988) Short-term synchronization of motor unit discharge in human sterno-cleidomastoid muscle during different respiratory and postural tasks. J Physiol (Lond) 399 : 34P
Adams L, Datta AK, Guz A (1989) Synchronization of motor firing during different respiratory and postural tasks in human sternocleidomastoid muscle. J Physiol (Lond) 413 : 213-231
Berger AJ (1979) Phrenic motoneurons in the cat: subpopulations and nature of respiratory drive potentials. J Neurophysiol 42 : 76-90
Campbell EJM (1974) Muscular activity in normal and abnormal ventilation. In: Wykc B (ed) Ventilatory and phonatory control systems. Oxford University Press, London, pp 3-11
Harada Y, Wang YZ, Kuno M (1985) Central chemosensitivity to H + and CO2 in the rat respiratory centre in vitro. Brain Res 333:336 339
Hebel R, Stromberg MW (1986) Anatomy and embryology of the laboratory rat. BioMed Verlag, W6rthsee, p 271
Hilaire G, Monteau R, Gauthier P, Rega P, Morin D (1990) Func- tional significance of the dorsal respiratory group in adult and newborn rats: in vivo and in vitro studies. Neurosci Lett 111:133-138
Lalley PM (1986) Serotoninergic and non serotoninergic responses of phrenic motoneurones to raphe stimulation in the cat. J Physiol (Lond) 380:373-385
Monteau R, Errchidi S, Gauthier P, Hilaire G, Rega P (1989) Pneumotaxic centre and apneustic breathing: interspecies dif- ferences between rat and cat. Neurosci Lett 99: 311-316
Monteau R, Gauthier P, Rega P, Hilaire G (1990a) Effects of N-methyl-D-aspartate (NMDA) antagonist MK-801 on breath- ing pattern in rats. Neurosci Lett 109:134-139
Monteau R, Morin D, Hennequin S, Hilaire G (1990b) Differential effects of serotonin on respiratory activity of hypoglossal and cervical motoneurons: an in vitro study on the newborn rat. Neurosci Lett 111 : 127-132
Monteau R, Morin D, Hilaire G (1990c) Ace@choline and central chemosensitivity: in vitro study in the newborn rat. Respir Physiol 81:241 254
Morin D, Hennequin S, Monteau R, Hilaire G (1990) Serotonergic influences on central respiratory activity: an in vitro study in the newborn rat. Brain Res 535:281-287
Murakami Y, Kirchner JA (1974) Respiratory activity of the exter- nal laryngeal muscles: an electromyographic study in the cat. In: Wyke B (ed) Ventilatory and phonatory control systems. Oxford University Press, London, pp 430-448
Murakoshi T, Otsuka M (1985) Respiratory reflexes in an isolated brainstem-lung preparation of the newborn rat: possible in- volvement of gamma-aminobutyric acid and glycine. Neurosci Lett 62:63-68
Richmond FJR, Scott DA~ Abrahams VC (1978) Distribution of motoneurones to the neck muscles, biventer cervicis, splenius and complexus in the cat. J Comp Neurol 181:451 464
Rose PK (1977) Morphology of motoneurones in the upper cervical cord of the adult cat. J Physiol (Lond) 272 : 37P
Smith JC, Feldman JL (1987) In vitro brainstem-spinal cord prepa- ration for study of motor systems for mammalian respiration and locomotion. J Neurosci Meth 21:321-333
Smith JC, Liu G, Feldman JL (1988) Intracellular recording from phrenic motoneurons receiving respiratory drive in vitro. Neu- rosci Lett 88:27-32
Stella G (1938) On the mechanisms of production and the physio- logical significance of 'apneusis ' . J Physiol (Lond) 93:10-23
Suzue T (1984) Respiratory rhythm generation in the in vitro brain- stem-spinal cord preparation of neonatal rat. J Physiol (Lond) 93 : 173-183
Takahashi T, Berger AJ (1990) Direct excitation of rat spinal moto- neurones by serotonin. J Physiol (Lond) 423 : 63-76












![#JP'...-q tE-FN ts-'**;. =(r : *' '''T&-K B] ffi 'd ri,f."f,..ir'L{Sii,::,:::t;,1;,,:"1 Austra ian owned.Snugg ebum creates comiy ara :::,lcepwear for kids aged from newborn](https://img.pdfslide.fr/doc/110x75/604cdeb75ac12142d739a0c3/jp-q-te-fn-ts-r-t-k-b-ffi-d-riffirlsiit11.jpg)











