Embed Size (px)
Citation preview

Eur J Biochem I O Y , 103-108 (1980) ’?j by FEBS 1980
Spectral Characterization of Cytochrome P-450 of a Strain of Cundidu tropicalis Grown on Tetradecane Daniel MANSUY, Maryse CARLIER, Jean-Claude BERTRAND, and Edgard AZOULAY
Laboratoire de Chimie de I’Ecole Normale Superieure, Paris, and Laboratoire de Structure et Fonction des Biomembraneb, Unite d’Enseignement et de Recherche de Luminy, Marseille
(Received January 23/May 12, 1980)
Several properties of the cytochrome P-450 induced in the yeast Candida tropicalis by growth on tetradecane have been studied by differential visible spectroscopy on microsomes. The spectral changes typical of this cytochrome have been obtained by subtraction of an unspecific spectral change, possibly due to the presence of other hemoproteins in microsomes, from the experimental difference spectra. Like the previously described cytochromes P-450 from yeast and mammalian liver, C. tropicalis cytochrome P-450 is in spin-state equilibrium at ambient temperature : about 30 % of the originally low-spin cytochrome is converted to the high-spin state upon increasing the ionic strength of the medium, whereas 30 7; of the originally high-spin cytochrome is converted to the low-spin state upon addition of hydrophobic alcohols. C. tropicalis cytochronie P-450 readily binds nitrogenous ligands, isocyanides and phosphines in the ferric and ferrous state with spectral charac- teristics similar to those reported for other yeast or mammalian cytochromes P-450. It also reacts successively with cumylhydroperoxide and 1,3-benzodioxole to form a high-valent iron-oxo species and an iron-carbene metabolite complex. However it fails to produce any spectral or spin-state change upon addition of hydrophobic non-coordinating compounds such as n-tetradecane, its substrate in vivo
It has been shown that the biosynthesis of a cyto- chrome P-450 is induced in a strain of Cundidu tropi- calis by growth on alkanes as the only source of carbon, under aerobic conditions [l, 21. This induction is obtained with straight-chain alkanes having at least ten carbon atoms, but also by long-chain alkenes and alcohols [3,4].
This cytochrome is functional in the hydroxylation of alkanes and fatty acids [l - 3,5], requires NADPH and oxygen and is inhibited by CO. The hydroxylase system has been resolved into three components : cyto- chrome P-450, NADPH : cytochrome P-450 reductase and a thermostable lipid fraction [5]. On recombina- tion to the active enzyme system, yeast reductase and the lipid fraction may be replaced by corresponding fractions from rat liver microsomes [5]. These results suggest a close similarity between the monooxygenases of C. tropicalis and those of mammalian liver micro- somes. However, it is noteworthy that the former are able to hydroxylate efficiently only long-chain linear alkanes and fatty acids exclusively in Q position [6]. Recently, a detailed study of the structure and
Enzyme. NADPH : cytochrome P-450 reductase (EC 1.6.2.4).
spectral properties of another yeast cytochrome P-450 from Sacchuvornyces crrevisiae has been reported [7-121. Up to now, the only visible difference spec- trum described for the C. tropicalis cytochrome P-450 is that of its CO complex [1,2,5]. The aim of the present work was to characterize more precisely the C. tropicalis cytochrome P-450, from spectral studies, and to compare it to cytochromeP-450 of S.cere- visiar and of mammalian liver microsomes.
MATERIALS AND METHODS
The yeast Candidu tropiculis, strain 101 [I31 (CBS 6947) was used. The minimal medium used and the growth conditions have been previously described [4]. The preparation of the microsomal fraction from the cells in the exponential phase of growth were carried out as previously described [6]. Cytochrome P-450 was assayed by its carbon monoxide difference spec- trum after reduction .by sodium dithionite according to the procedure of Omura and Sat0 [14], assuming an absorption coefficient of 91 mM-’ cm-’ between 450 and 490 nm. The protein concentration was deter- mined by the method described by Lowry et al. [15].
104 Characteristics of Cytochrome P-450 from C. tropicalis
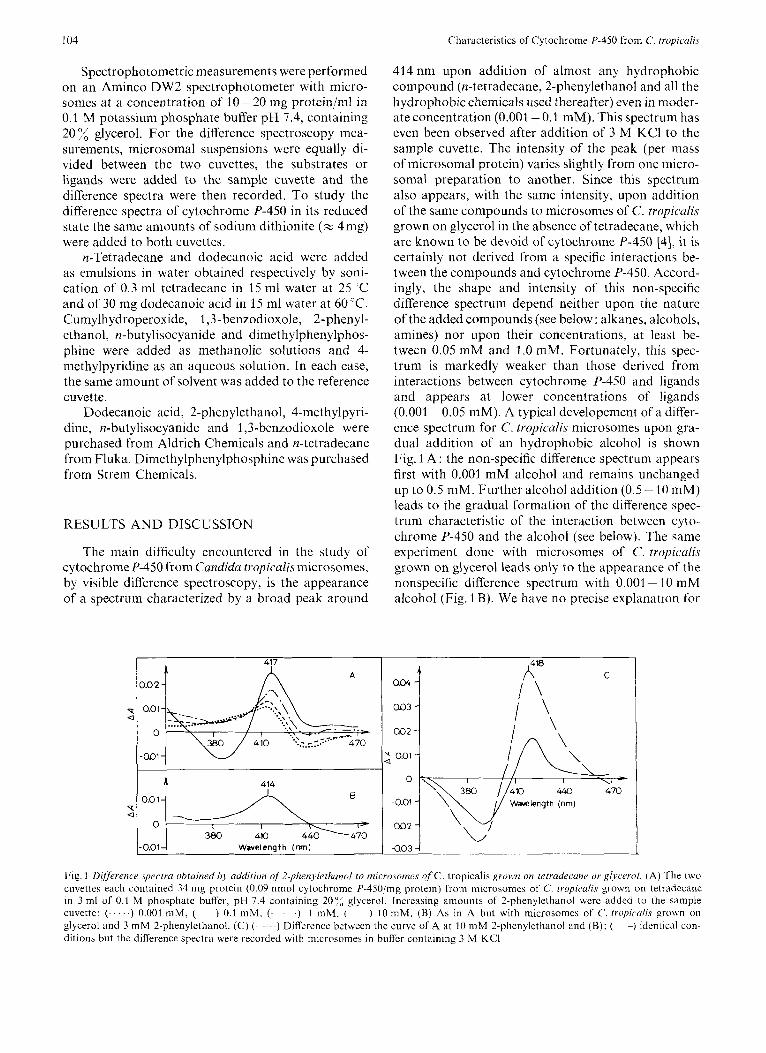
Spectrophotometric measurements were performed on an Aminco DW2 spectrophotometer with micro- somes at a concentration of 10-20 nig protein/ml in 0.1 M potassium phosphate buffer pH 7.4, containing 20 glycerol. For the difference spectroscopy mea- surements, microsomal suspensions were equally di- vided between the two cuvettes, the substrates or ligands were added to the sample cuvette and the difference spectra were then recorded. To study the difference spectra of cytochrome P-450 in its reduced state the same amounts of sodium dithionite (= 4mg) were added to both cuvettes.
n-Tetradecane and dodecanoic acid were added as emulsions in water obtained respectively by soni- cation of 0.3 ml tetradecane in 15 ml water at 25 'C and of 30 mg dodecanoic acid in 15 ml water at 60 "C. Cumylhydroperoxide, 1,3- benzodioxole, 2-phenyl- ethanol, n-butylisocyanide and dimethylphenylphos- phine were added as methanolic solutions and 4- methylpyridine as an aqueous solution. In each case, the same amount of solvent was added to the reference cuvette.
Dodecanoic acid, 2-phenylethanol, 4-methylpyri- dine, n-butylisocyanide and 1,3-benzodioxole were purchased from Aldrich Chemicals and n-tetradecane from Fluka. Dimethylphenylphosphine was purchased from Strem Chemicals.
RESULTS AND DISCUSSION
The main difficulty encountered in the study of cytochrome P-450 from Cundidu tropicalis microsomes, by visible difference spectroscopy, is the appearance of a spectrum characterized by a broad peak around
0.02 -
414nm upon addition of almost any hydrophobic compound (n-tetradecane, 2-phenylethanol and all the hydrophobic chemicals used thereafter) even in nioder- ate concentration (0.001 -0.1 mM). This spectrum has even been observed after addition of 3 M KCI to the sample cuvette. The intensity of the peak (per mass of microsomal protein) varies slightly from one micro- soma1 preparation to another. Since this spectrum also appears, with the same intensity, upon addition of the same compounds to microsomes of C. tropicalis grown on glycerol in the absence of tetradecane, which are known to be devoid of cytochrome P-450 [4], it is certainly not derived from a specific interactions be- tween the compounds and cytochrome P-450. Accord- ingly, the shape and intensity of this non-specific difference spectrum depend neither upon the nature of the added compounds (see below: alkanes, alcohols, amines) nor upon their concentrations, at least be- tween 0.05 mM and 1 .O mM. Fortunately, this spec- trum is markedly weaker than those derived from interactions between cytochrome P-450 and ligands and appears at lower concentrations of ligands (0.001 -0.05 mM). A typical developement of a differ- ence spectrum for C. tropiculis microsomes upon gra- dual addition of an hydrophobic alcohol is shown Fig. 1 A : the non-specific difference spectrum appears first with 0.001 mM alcohol and remains unchanged up to 0.5 mM. Further alcohol addition (0.5 - 10 mM) leads to the gradual formation of the difference spec- trum characteristic of the interaction between cyto- chrome P-450 and the alcohol (see below). The same experiment done with microsomes of C. tropiculis grown on glycerol leads only to the appearance of the nonspecific difference spectrum with 0.001 - 10 mM alcohol (Fig. 1 B). We have no precise explanation for
2 aoi-
440 Wavelength (nrn)
- 0.0 1 Wavelength (nm) a03 7
Fig. 1 Difference .spectra ohtuined by uddition of 2-phenyletltanol to microsomes of C. tropicalis grown on tetrudecune or glycerol. (A) The two cuveltes each contained 34 mg protein (0.09 nmol cytochrome P-450inig protein) [rom microsomes of C. t ropicd is grown on tetradecane in 3 ml of 0.1 M phosphate buffer, pH 7.4 containing 207, glycerol. Increasing amounts of 2-phenylethanol were added to the sample cuvette: (. . . . .) 0.001 mM, ( ) 0.1 mM, (. . .) 1 mM. ( ) 10 m M . (R) As in A but with microsomes of C. tropicalis grown on glycerol and 3 m M 2-phenylethanol. (C) ( ~ ) Difference between the curve of A at 10 mM 2-phenylethanol and (€3); ( --) identical con- ditions but the difference spectra were recorded with microsomes in buffer containing 3 M KCI

D. Mansuy, M. Carlier, J.-C. Bertrdnd, and E. Azoulay 105
this nonspecific difference spectrum but believe that it is due to a modification of a hemoprotein present in C. tropicalis inicrosomes (e.g. catalase, peroxidase or cytochroine oxidase) which has not been removed during the preparation of microsomes [33].
In the other figures of this paper, the non-specific difference spectrum (which can be obtained from microsomes of C. tropicalis grown on glycerol) has been substracted from the experimental difference spectra of microsomes of C. tropicalis grown on tetra- decane. In the following, the expression ‘C. tropicalis microsomes’ will mean microsomes of C. tropicalis grown on tetradecane.
Spectral Clianges Caused by Variation of the Ionic Strength
Upon addition of 3 M KCI to C. tropicalis micro- somes, a characteristic difference spectrum appears with a peak at 386 nm and a trough at 416 nm (Fig. 2) corresponding to the transition of the originally low- spin oxidized cytochrome P-450 to the high-spin state, as previously described in the case of S. cerevisiae cytochrome P-450 [S]. If one uses the difference ab- sorption coefficient At . (390-412 nm) of 130 mM-’ cm I , reported for the conversion of Pseudomonus putida low-spin cytochrome P-450 to the high-spin state [16], one finds that this transition affects 332, of the cytochrome, in the experimental conditions of 25 C, 100 mM phosphate buffer pH 7.4,20% glycerol.
In teruct ion with Hydrop ho hic Non-coordinating Compounds
Addition of n-tetradecane, which is the substrate of C. tropicalis cytochrome P-450 in vivo, to C. tropi- culis microsomes fails to produce any specific spectral change. Even the addition of a large excess of an emulsion of n-tetradecane in water, in the presence of added phospholipids (tripalmitoylphosphatidyl- choline at 0.1 mg/mg microsomal protein, or total lipids extracted from rat liver microsomes at 0.5 mg/mg microsomal proteins), does not produce any expected type-I spectral change (Fig.2). A similar result is ob- tained with dodecanoic acid, which is however hy- droxylated in the w position by C. tropicalis micro- somes and NADPH in aerobic conditions [1,6], and with various well-known substrates of liver cyto- chromes P-450 such as cyclohexane and SKF525A.
Interaction qf C . tropicalis Microsomes with Cytochrorne P-450 Ligunds
Fig. 1 A shows the difference spectra of C. tropicalis microsomes observed upon addition of increasing amounts of 2-phenylethanol. The spectrum corre- sponding specifically to the interaction between cyto-
1 -0.01
0.02 1 Fig. 2. E’ j f i~ t s of’ ionic strerigth utzd Izydropkohic c~ompounds on tlle spectrum uf microsomes o/ C. tropicalis grobvn on tetradecune. Thc two cuvettes contained 34 mg protein (0.09 nmol cytochrome P-450jmg protein). (-) Addition of 3 M KC1 in the sample cuvette; ( ) addition of 0.9 mM tetradecane and rat hepatic microsomal phospholipids (0.5 mgj to the sample cuvette
chromeP-450 and the alcohol (Fig. 1 C), with a peak at 41 8 nm and a trough around 388 nm, gradually appears when the alcohol concentration varies from 0.5 mM to 10 mM. Hydrophobic alcohols thus interact with C. tropicalis cytochrome P-450 in the same manner as with mammalian liver [17- 191 and S. cerevisiae [8] cytochrome P-450. The corresponding spectral change, called reverse type I, reflects the coordination of the alcohol oxygen as a weak-ligand to cytochrome P-450 Fe(II1) originally in the high-spin state. The spectrum is the actual mirror of that obtained for the transition of originally low-spin cytochrome P-450 to the high-spin state. This suggests that the sixth endoge- nous ligand of iron in originally low-spin C. tropicdis cytochrome P-450, as in liver or S. cevevisiue cyto- chromes P-450, is a weak ligand similar to an alcohol, presumably the OH group of water or of an amino acid residue of the protein [18-21]. Assuming that A i: (420 - 390 nm) = 130 mM cm- for the reverse- type-I spectral change, one finds that about 307; of originally high-spin C. tropiculis cytochrome P-450 is able to bind 2-phenylethanol. When the alcohol is added to C. tropicalis microsomes containing 3 M KCI, the same difference spectrum appears but with a greater intensity (Fig. 1 C) indicating that, in this case, about 56 ;< of the cytochrome undergoes a spin- state change. The increase of the proportion of high- spin cytochrome P-450 which binds the alcohol (56- 30 = 26 %) in the presence of 3 M KCI corresponds roughly to the increase of the proportion of high-spin cytochrome upon addition of 3 M KCI (33p/,, see preceding section).
Stronger ligands like 4-methylpyridine (Fig. 3), dimethylphenylphosphine (Fig. 4) and n-butyliso- cyanide (Fig. 5) produce difference spectra with peaks

Characteristics of Cytochrome P-450 from C. tropicalis I06
0.02
0.01
0
T a -0.01 -
-0.02 -
I 392
Fig. 3. Spectral interaction of rytorhrome P-450,from microsornes of C . tropicalis with 4-nzethylpyl-idine. Conditions were similar to those of Fig. 1; 6.6 mg proteinlml, 0.6 pM cytochrome P-450. 4-Methylpyridine was added to the sample cuvette at (-) 4 mM, (---) 20 mM, (- --) 80 mM, (-.-.-) 120 mM
0.06 I 458 A
/ \
0.04
0.02
P O cl
- 0.0 2
- 0.04
I \ -0.06 I \ /
-0.08
Fig. 4. Spectral interaction of cytoclzrome P-450,from rnicrosome.r o j C . tropicalis with ~~imetliylplien~lplzosphine. Conditions of Fig. 3, 0.7 pM cytochrome P-450. (-) Addition of 3 mM dimethyl- phenylphosphine to the sample cuvette; ( ) further addition of 10 mM sodium dithionite to both cuvettes
respectively at 428, 456 and 435 nm, very similar to those obtained with liver microsomal cytochromes P-450 [29,21,22]. A difference spectrum with a peak at 428 nm and a trough around 394nm appears gradually with isosbestic points at 419 nm and 449 nm when the 4-methylpyridine concentration is increased
0.15
0.10
0.05
0
- 0.05
435 h
T'
i50
Fig. 5. Spectral interaction ofc:ytochrorne P-45Vjrom microsomes of C. tropicalis with n-hutylisocyunide. Conditions of Fig. 3 (0.7 pM cytochrome P-450). (--) Addition of 5 mM n-butylisocyanide to the sample cuvette; (---) further addition of 10 mM sodium di- thionite to both cuvettes
from 0.4 mM to 20 mM. The apparent spectral disso- ciation constant, Ks, calculated from these curves (from three experiments) lies around 2 mM and thus is similar to that obtained with rat liver microsomes, 1.2mM (D. Mansuy and M. Carlier, unpublished re- sults). With higher pyridine concentrations (> 20mM), a second trough appears around 407 nm without appreciable variation of the peak position. Similar results have been reported for the interactions of amines with liver [23] and S. cerevisiue [8] micro- somes. Here, 4-methylpyridine (L) in moderate con- centration first binds to originally high-spin C. tropi- calk cytochrome P-450, which causes a high-spin Fe(II1) to low-spin Fe(II1)-L transition corresponding to a difference spectrum with a trough at 394nm. Higher concentrations of 4-methylpyridine are neces- sary for the replacement of the endogenous sixth ligand (L') of originally low-spin C. lropicalis cyto- chrome P-450, which results in a low-spin Fe(II1)-L' to low-spin Fe(II1)-L transition corresponding to a difference spectrum with a trough at 407 nm.
The spectrum produced by dimethylphenylphos- phine is of the hyperporphyrin type with two maxima at 370 and 456 nm. The phosphine also binds to C. tropicalis cytochrome P-450 in the reduced state, the derived complex being characterized by a peak at 458 nm (Fig. 4). The position and very high intensity of this peak, (A458 - A490) per nmol cytochrome P-450 = 0.1, are identical to those obtained with rat liver microsomes [22].
Upon reduction of the complex of C. tropiculis cytochrome P-450 Fe(II1) with n-butylcyanide by di-

D. Mansuy, M. Carlier, J.-C. Bertrand, and 6. Azoulay 107
thionite (Fig. 5), two peaks appear at 435 and 457 nm as in the case of liver [14] or S. cerevisiae [8] micro- somes. It is well known that the relative intensities of these two peaks are pH-dependent [24]. It is note- worthy that the intensity of the 457-nm peak relative to that of the 435-nm peak remains considerably lower (whatever the pH) for C. tropicalis than for S. cere- visiae or liver microsomes.
Reaction o j C . tropicalis Microsomes with Cumylhydroperoxide and 1,3- Benzodioxole
The addition of cumylhydroperoxide to C. tropi- culis microsomes leads to the appearance of a difference spectrum with a peak at 444 nm (Fig.6). A similar result obtained with liver microsomes has been attributed to the formation of an unstable high- valent cytochrome P-450 Fe = 0 ferry1 complex in steady-state concentration [25]. A further addition of 1,3-benzodioxole to these liver microsomes containing cumylhydroperoxide has been shown to lead to the formation of a cytochrome P-450 Fe(II1)-benzodioxole metabolite complex [28] characterized by a difference spectrum with a peak around 440 nm which is very similar to that of the cytochrome P-450 F e z 0 com- plex. This metabolite, which is a strong ligand of cytochrome P-450, appears to be the 1,3-benzodioxoIe- 2-carbene derived from an oxidation of the niethylene group of 1,3-benzodioxole by the active oxygen cyto- chrome P-450 complex [29]. Reduction of this carbene complex by sodium dithionite leads to the corre- sponding cytochrome P-450 Fe(I1)-I .3-benzodioxole- 2-carbene complex characterized by a difference spec- trum with two peaks around 430 and 455 nm [26- 291. As shown in Fig. 6, similar complexes of C. tropicalis cytochrome P-450 in the ferric and ferrous state, characterized respectively by peaks at 444 nm and at 426 and 456 nm are formed in the same conditions. The two peaks at 426 and 456 nm have been obtained reproducibly with several samples of C . tropiculis inicrosonies but with a considerably lower intensity than with liver microsomes. It should also be men- tioned that the formation of the 444-nm peak upon hydroperoxide addition has not been obtained with some samples of C. tropicalis microsomes.
The haem iron of C . tropicalis cytochrome P-450 exhibits several properties similar to those of liver and S. cerevisiae cytochromes P-450, as follows.
a) All of them are able to bind nitrogenous ligands, isocyanides and phosphines, in the ferric state, and, isocyanides, phosphines and carbon monoxide in the ferrous state, leading to the same characteristic spectral variations.
b) All of them are in spin-state equilibrium at am- bient temperature, the low-spin form being an hexaco- ordinated iron complex with presumably and hydroxyl- containing sixth endogenous weak ligand. About 30
1 0.005
, I
470 500 ‘\ . ,i Wavelength (nm)
‘I 4
0.005 -
i 0.010
Fig. 6. Dlffirence spectru of microsomes of C . tropicalis grown on tetrudecune in the presence of cumylhydroperoxide and 1.3-henzo- dioxole. Conditions of Fig. 3 (0.5 pM cytochrorne P-450. (- ~ ) Addition ol‘ 0.2 mM cumylhydroperoxide to the sample cuvette; (---) further addition of 3 mM 1,3-benzodioxole to the sample cuvette; (.-.--.) further addition of 10 mM sodium dithionite to both cuvettes
of the originally low-spin C . tropicalis cytochrome P-450 is converted to the high-spin state upon increas- ing the ionic strength of the medium. The reverse spin- state change occurs upon addition of an hydrophobic alcohol, about 30 of C. tropicalis cytochrome P-450, which was originally high-spin, being affected.
c) C. tropicalis and liver cytochromes P-450 are able to form very reactive iron-oxo species upon reaction with cumylhydroperoxide and iron-carbene complexes [29] upon 1,3-benzodioxole oxidation.
I t is noteworthy that there is an important pro- portion of C. tropicalis cytochrome P-450 in the high- spin state in C . tropicalis microsomes at ambient temperature. This is underlined by the intense reverse- type-] spectral change produced by addition of alcohols to C . tropicalis inicrosomes and by the posi- tion of the through at 394 nm in the difference spectra produced by addition of pyridines, which corresponds to the binding of these ligands to high-spin cyto- chrome P-450 Fe(II1) [18,19,23]. In this respect, C. tropicalis cytochrome P-450 resembles hepatic cyto- chromes P-450 from rabbits pretreated with P-naph- thoflavone or 3-methylcholanthrene [19] or S. cere- visiae cytochrome P-450 [8], which are known to exist largely in the high-spin state in vivo, more than hepatic cytochrome P-450 from phenobarbital-pre- treated rats [19].
The most intriguing feature of microsomal C . tro- picalis cytochrome P-450 is its inability to undergo any conversion from low-spin to high-spin state upon addition of hydrophobic compounds and in particular of tetradecane which is the substrate oxidized by C. tropiculis cytochrome P-450 in vivo. A possible ex- planation for this result could be the presence of n-tetradecane or one of its metabolites remaining bound to the protein of the active site of cytochrome P-450 during the preparation of the microsomes.

1 OX D. Mansuy, M. Carlier, J.-C. Bertrand, and E. Azoulay: Characteristics of Cytochrome P-450 from C. tropicalis
Similar results have been obtained for the interactions of cyclohexane or other hydrophobic compounds with liver cytochromes P-450 from /3-naphthoflavone- treated rabbits [19] or with S. cercvisiue cytochrome P-450 [S] which all exist predominantly in the high- spin state in vivo. However, in the case of C. wopicalis cytochronie P-450, this lack of spectral change upon addition of tetradecane could be related to the absence of hydroxylating activity of C. tropicalis microsomes with tetradecane as substrate [6]. This is not caused by an important denaturation of the active site of C. tropicalis cytochrome P-450 during the preparation of microsomes since C. tropicalis microsomes are ac- tive in the hydroxylation of dodecanoic acid [6].
A possible explanation for the lack of spectral change after addition of tetradecane, and for the lack of tetradecane hydroxylase activity in vitro, could be the loss of a cofactor of transport of the very hydro- phobic tetradecane to the active site of C. tropirulis cytochronie P-450 during isolation of the microsomes. This explanation seems likely for two reasons: first, it is known that more-or-less specific emulsifying substances, such as rhamnolipids, are biosynthesized by yeasts grown on alkanes [30,31]; second, it has recently been reported that an acidic phospholipid together with a protein factor are necessary for ex- ternally added squalene (another very hydrophobic compound) to bind to the catalytic site of squalene epoxidase of rat liver inicrosomes [32]. Experiments for the isolation of such a transport factor of tetra- decane in C. tropicalis are in progress.
REFERENCES 1 .
2.
3.
4.
5 .
6.
Lchcault, J . M., Lode, E. T. & Coon, M. J . (1971) Bioclimz.
Gallo, M., Bertrand, J . C. & Azoulay, E. (1971) FLBS Lett.
Gallo, M., Bertrand, J. C., Rochc, B. & Azoulay, E. (1973)
Gilewicz, M., Zacek, M., Bertrand, J. C. & Azoulay, E. (1979)
Duppel, W., Leheault, J . M. & Coon, M. J. (1973) Enr. J .
Bertrand, J. C., Gilewicz, M., Bazin, H., Zacek, M. & Azoulay.
Biopliq's. Res. Commun. 42, 413-419.
19,45-49.
h c l i i m . Biophys. Actu, 296, 624- 630.
Can. J . Microhiol. 25, 201 -206.
Bioclzem. 36, 583 - 592.
E. (1979) FEBS Lett. 105, 143-146.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16. 17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27. 28.
29.
30.
31.
32.
33.
Yoshida, Y., Kumaoka, H. & Sato, R. (1974) J . Bioclienz.
Yoshida, Y. & Kumaoka, H . (1975) ./. Biochum. f T o k j ~ i ) 78.
Kubota, S., Yoshida, Y. & Kumaoka, H. (1977) ./. Bioclwrn.
Yoshida, Y., Aoyama, Y., Kumaoka, H. & Kubota, S. (1977)
Aoyama, Y., Yoshida, Y., Kubota, S., Kumaoka. H. & Furu-
Aoyama, Y. & Yoshida, Y. (1978) Biochem. Biopliys. Res.
Duvnjak, Z., Roche, B. & Azoulay, E. (1970) Arch. Mikrohiol.
Omura, T. 62 Sato, R. (1964) J . Biol. Chenz. 239. 2370-
Lowry, 0. H., Rosebrough, N. J., Farr. A. L.. & Randall, R . J.
Peterson, J . A. (1971) Arch. Bioclzem. Biopl7y.s. 144. 678-693. Schenkman, J . B., Cinti, D. L., Orrenius, S., Moldeus, P. &
Yoshida, Y. & Kumaoka, H. (1975) J . Bioclicw. (ToXxo) 78,
Kuniaki, K., Salo, M., Kon, H. & Nehert, D. W. (1978) J .
Ullrich, V., Sakurai, H. & Ruf, H. H. (1979) Act0 Biol. Med.
Schenkman, J. B., Remmer, H. & kstahrook. R. W . (1967)
Mansuy, D., Duppcl, W., Ruf, H. H. & Ullrich, V. (1974)
Jcfcoate, C. R. E., Gaylor, J. L. & Calahrese, R. (1969) Bio-
(Tokyo) 75, 1201-1219.
785 - 794.
(Tokyo) 81, 1 78 - 205.
Biocl7em. Biopliys. Res. Commun. 78, 1005 - 1010.
michi, A. (1978) Arch. Biockem. Biophys. 385, 17-24.
Commun. 82, 33 - 38.
72, 135-139.
2378.
(1951) J . Biol. Chem. 193, 265-275.
Kraschniz, R. (1972) Biochemistry, I ! , 4243.
455-466.
B i d . Clzem. 253, 1048-1058.
Germ. 38, 287-297.
Mol. Pharmucol. 3, 113 - 123.
Hoppe-Seyler's Z. Ph.vsiol. Cliem. 355, 1341 - 1349.
chemistry, 8 , 3455 - 3463. Imai, Y. & Sato, R. (1967) J . Bioch~m. (TfiXJYii 62, 464-
473. Rahirntula, A. U., O'Brien, P. J., Hrycay, E. G., Peterson, J .
A . & Estahrook, R . W. (1974) Biochem. Riopliys. R e x Cornmuti. 60, 695 - 702.
Philpot, R . M. & Hodgson, E. (1971) C%rm.-Bio/, Intercrct. 4 . 185 - 195.
Franklin, M. R. (1971) Xenohiotica, I , 581 -588. Elcoinbe, C. R., Bridges, J . W., Nimmo-Smith, R. H. & Wer-
Mansuy, D., Battioni, J. P., Chottard. J . C. & Ullrich. V. (1979)
Hisatsuka, K., Nakahava, T., S a m , N. & Yamada. K. (1971)
Rosenhcrg, E., Zuckerherg, A., Ruhinovitz, C. & Gutnick, D.
Nakamura, M. & Sato, R . (1979) Biocliem. Biophys. Kes.
Kawamoto, S. , Nozaki, C. , Tanaka. A . & Fukui, S. (1978)
ringloer, J. (1975) B i o c l i ~ ~ . Soc. Tram. 3, 967-968.
J . A m . Clzem. Soc. 101, 3971-3973.
A g r . Biol. C'l7em. 35, 686- 692.
L. (1 979) Appl. Lnviron. Microhiol. 37, 402 - 408.
Commun. 89, 900 - 906.
Eur. J . Bioc'lieni. 83, 609-613.
D. Mansuy and M. Carlier, Lahoratoire de Chimie de I'Ecole N o r m I I C Supkrieure, Assock au Centre National de la Recherche Scientilique no. 32, 24 Rue Lhomond, F-75231 Paris-Cedex-05, France
J.-C. Bertrand and E. Azoulay, Lahoratoire de Structure et Function des Biomemhranes, Unite d'tnscignement et de Recherche du Centre Universitaire Marseille-Luminy, Universite d'Aix Marseille, 70 Route Lion-Lachamp, P-I 3288 Marseille-Cedex-2, France














![Paul Desmond (1924-1977) · Blue rondo à la turk. - [7] (1997) avec Paul Desmond (1924-1977) comme Saxophoniste I've grown accustomed to her face. - [7] (1997) avec Paul Desmond](https://img.pdfslide.fr/doc/110x75/60cdac42cca40d79bf3516a4/paul-desmond-1924-1977-blue-rondo-la-turk-7-1997-avec-paul-desmond-1924-1977.jpg)













