Embed Size (px)
Citation preview

490 PRELIMINARY NOTES
We are now studying a new approach to the selective cleavage problem, provided by the reaction of nucleosides with oximes and hydroxamic acids; the results of these experiments will be published elsewhere.
As far as we know, the reaction investigated is the first completely selective route for modification of a single nucleic base which is believed to gain importance in genetic studies and nucleic acids sequence analysis.
Institute /or Chemistry o/ Natural Products, Academy o/ Sciences, Moscow (U.S.S.R)
N. K. KOCHETKOV E. I. BUDOWSKY R. P. SHIBAEVA
t 1~. K. KOCItETKOV, E. I. BUDOWSKY AND N. A. SIMUKOVA, Biokhimia, 27 (1962) lO35. t D. BROWN AND P. SNELL, J. Mol. Biol., 3 (196i) 709 . s H. SCrlUSTER, J. Mol. Biol., 3 (1962) 447. 4 E. FREESE, E. BAUTZ AND E. BAUTZ-FREESE, Proc. Natl. Acad. Sci. U.S., 47 (1961) 845. 6 D. VERWOERD, H. HOHLAGE AND W. ZILLIO, Nature, 192 (1961) lO38. 6 j . j . Fox , D. VAN PRAAG, I. WEMPEN, I. L. DOERR, L. CHEONG, J. E. KNOLL, M. L. EIDINOFF, A.
BENDICH AND G. B. BROWN, J. Am. Chem. Soc., 81 ~I959 ) 178. * R. M. KHOMUTOV, Zh. Obshch. Khim., 31 (1961) 1992.
Received January I8th, 1963
Biochim. Biophys. Acta, 68 (1963) 493-496
PN 6090
Sur la biosynth~$e de 3,4-dihydroxyph6nylalanine h~moglobine
En utilisant des ribosomes de Escherichia coli en prdsence d'un polym6re mixte des aeides uridylique et guanylique (poly-UG), on a observd que l'alanine ou l'acide cys- t~ique, introduits dans le milieu ~ l'dtat d'esters avec le RN'A soluble (s-RNA) sp6- cifique de la cyst6ine, 6taient incorpords A la place de la cystdine dans un mat6riel de nature prot6ique synthdtis61, ~. Ces r6sultats ont montrd que le positionnement de l'acide amind dans la protdine en formation 6tait ddtelTnin6, comme le suggdrait l 'hy- poth6se de 'Tadapta teur ''s, par le s-RN'A portant l'acide amind. Cependant, comme dans le syst~me utilisd le poly-UG jouait le r61e d'un messager artificiel et conduisait
la synth6se de prot6ines elles aussi artificielles, on pouvait se demander si des r6sul- tats analogues pouvaient 8tre obtenus avec un matdriel met tant en oeuvre un rues- sager naturel.
Dans ce travail, on a examind le comportement du syst6me qui synthdtise l'hd- moglobine in vitro* en relation avec le s-RNA sp~cifique de la tyrosine et contenant
sa place la 3,4-dihydroxyph6nyl-L-alanine (DOPA)*. Des ribosomes de rdticulocytes de lapin ont 6td incub6s avec du s-RNA de E.
coli charg6 avec tousles [z*C]amino acides et contenant soit le [l*C]tyrosyl-s-RNA, soit le [I*C]DOPA-s-RNA. L'h6moglobine synthdtisde a dtd isolde et, apr6s digestion trypsique, analysfie par chromatographie-61ectrophor6se. Les r6sultats obtenus ont
" Avec les r ibosomes de E. toll, darts les condi t ions ddcri tes prdc6:lernment, 700 coups / ra in o n t fit6 retrouvds dans le prdcipi td t r iehloroacdt ique , su r xo 4 coups / ra in de l ' C - t y r - s - R N A ajou- t~s e t 28oo si p o l y - U A 6ta i t prf isent dans le mil ieu. Avec le I ' C - D O P A - s - R N A , les rad ioac t iv i tds respectives dta ien t 460 et 18oo coups/ ra in ; c e p e n d a n t avec cer ta ines prf iparat iot ls l ' incorpora t ion
en l ' absence et la s t i m u l a t i o n en prdsence de p o l y - U A fitaient t r~s faibles.
Biochim. Biophys. Acta, 68 (1963) 496-499

PRELIMINARY NOTES 49"]
naontr~ que la [I~C]DOPA 6tait retrouv6e dans les na6naes peptides que ceux contenant ~t l'6tat normal la [14C]tyrosine.
Le s-RNA de E. coli a 6t6 pr6par6 et charg6 avec les acides amines selon NATHANS ET LIPMANN s, s6par6naent avec les [12C]acides anainds et avec la [14C] tyrosine (40o naC/ mnaole). Pour le [14C]tyrosyl-s-RNA on a obtenu o.3 na/~naole de tyrosine par mg de s-RN'A ou 62 ooo coups/rain. Des quantit6s 6gales de [12C]amino acyl-s-RNA et de [14C]tyrosyl-s-RNA ont 6t6 mdlangdes et le m6lange a 6t6 utilis~, soit directement, soit apr~s oxydation du tyrosyl-s-RNA en DOPA-s-RNA.
Cette derni~re a 6t~ r6alis6e dans des conditions analogues ~ celles ddcrites pour l 'oxydation des testes de tyrosine des prot6inese: 0.5 nal de [14C]tyrosyl-s-RNA et 0.5 nal de s-RNA est6rifi6 avec tous les [l~C]anaino acides ont ~t6 incub6s ~ 2o ° sous agitation continue avec o.o3 o nal de solution de polyphdnoloxydase (EC 1.1o.3.1) de chanapignon 7 (45 ooo Unitds Hogeboom-Adams par nal) et un exc~s d'acide as- corbique (16/,naoles). En 9o sec l 'oxydation est conapl~te et, apr~s hydrolyse ~ pH I I et analyse chronaatographique, on observe seulenaent la prdsence de DOPA libre et d'une faible quantitd (inf6rieure A IO %) d'un produit d'oxydation de celle-ci. Les conditions d'incubation d6crites ont ~t~ cependant variables pour chaque lot de s-RN'A utilis~.
Les ribosomes de r6ticulocytes de lapin ont ~t6 prdpar6s selon ALLEN ET SCHWEET 8 et le poly-UA dans les conditions d~crites pour le poly-UG ~.
Conditions expdrimentales et rdsultats. 50 nag de ribosomes de r~ticulocytes, i6 nag de s-RN'Acharg~ avec 19 [~C]acides amin6s et contenant soit du [~4C]tyrosyl-s-RNA (5" IO5 coups/nain) (Milieu I) soit du [ltC]DOPA-s-RNA (5" IO~ coups/min) (Milieu II) ont 6t6 incub6s pendant 3omin A 37 ° avec: 25o/~moles Tris-HC1 (pH 7.4); 15o/~moles KC1; 24/~naoles MgCI~; 42o/~naoles phospho6nolpyruvate; o.2 mg pyruvate- kinase (EC 2.7.1.4o); 1.5/,mole GTP; 37/~moles glutathion r6duit; o.2/~naole h6mine et I nag (exprina6 en prot~ine) du surnageant ~ lO5 ooo×g d'honaog6nat de foie de lapin (volume final 4 ml).
Apr~s l'incubation, 6 nag d'h6moglobine pure de lapin sont ajout6s dans chaque milieu et une pattie aliquote est pr6lev6e pour d6terminer le taux d'incorporation du ~C dans le naat~riel pr6cipitable par l'acide trichloroac6tique ~ chaud. Le reste a 6t6 dialys6 A+2 ° pendant 48 h contre 6 1 de tampon aux phosphates O.olM (pH 7.0). L'h6moglobine a 6t6 ensuite purifi6e sur colonne de DEAE-cellulose selon HENNESSEY et al. 9. La solution d'h6moglobine a dt6 ensuite concentrde par dialyse sous vide, dig6r6e par la trypsine cristallisde et les peptides forna6s ont ~t6 analysds par chromato-~lec- trophor~se selon KATZ et al. ~°. Les chromato-61ectrophor6gramnaes ont dt6 ensuite autoradiographi6s (Films Kodak Kodirex) pendant I naois.
Dans le Milieu I contenant !e [l~C]tyrosine-s-RNA, 45 ooo coups/rain ont 6t6 trouvds dans le prdcipitd trichloroacdtique et 14 I5O coups/nain dans l'h6naoglobine purifi6e. Dans le Milieu I I contenant le [~C]DOPA-s-RNA les radioactivit6s retrou- v~es ont dt6 4 ° 5oo et 13 5oo coups/rain respectivenaent. La Fig. I montre que, dans l'h6naoglobine isol6e du Milieu II, 7 peptides radioactifs sont nais en 6vidence. Leur position chronaato-dlectrophor6tique est comparable A celle des peptides de l'h6nao- globine isol6e du Milieu I, qui contenait le [~C]tyrosyl-s-RNA, et A celle des pep- tides de tyrosine (r6vdlds par r,t-nitroso-/~-naphtol) de l'h6naoglobine purifi6e de lapin. Selon les donn6es analytiques de N'AUGHTON ET DISTZlS 11 et de DIAMOND ET BRAL~NITZERlZ, 6 peptides de tyrosine devraient 6tre trouv6s. La pr6sence d'un peptide
Biochim. Biophys..4eta, 61 (I962) 496-499
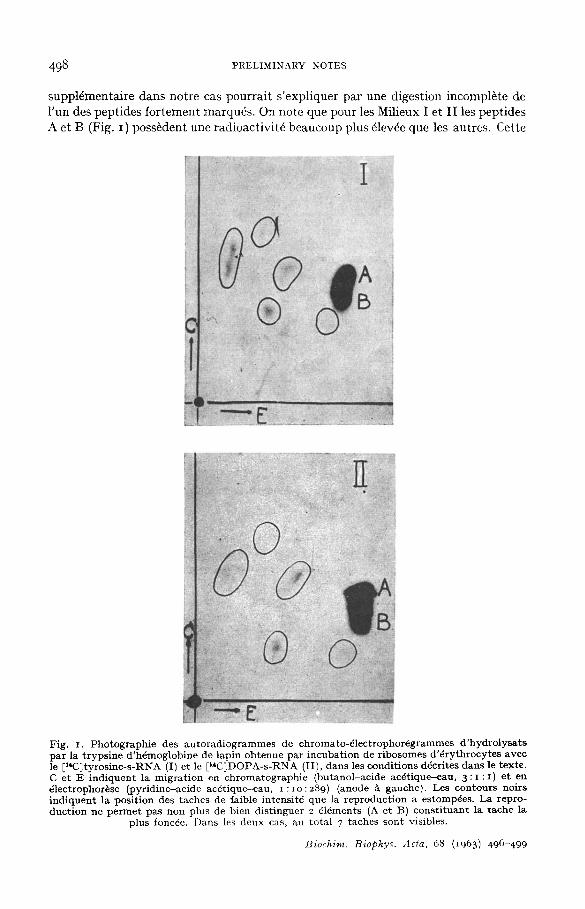
498 PRELIMINARY NOTES
suppl6mentaire dans notre cas pourrait s'expliquer par une digestion incompl6te de l 'un des peptides forternent marqu6s. On note que pour les Milieux I e t II les peptides A et B (Fig. I) poss6dent une radioactivit6 beaucoup plus 4lev6e que les autres. Cette
Fig. I. Pho tog raph i e des a u t o r a d i o g r a m m e s de ch roma to -~ lec t rophorSgrammes d ' h y d r o l y s a t s pa r la t r yps ine d ' h6mog lob ine de lap in ob tenue pa r i ncuba t i on de r ibosomes d ' 6 r y t h r o c y t e s avec le [ l*C]tyros ine-s-RNA (I) et le [ t4C]DOPA-s-RN A (II), dans les cond i t ions d~cri tes d a n s le texte . C et E i n d i q u e n t la m i g r a t i o n en c h r o m a t o g r a p h i e (bu tano l -ac ide ac6tique--eau, 3 : I : I) e t en ~lectrophor~se (pyr id ine-ac ide ac~ t ique-eau , i : i o :289) (anode A gauche) . Les con tou r s noirs i n d i q u e n t la pos i t ion des t aches de faible in tens i t6 que la r ep roduc t ion a es tomp6es . La repro- duc t i on ne p e r m e t pa s non p lus de bien d i s t inguer 2 616ments (A et B) c o n s t i t u a n t la t ache la
p lus fonc~e. D a n s les deux cas, au to ta l 7 t aches son t visibles.
Biochim. Biophys. Acta, 68 (1963) 496-499

PRELIMINARY NOTES 499
diff6rence paralt traduire l'allongement prioritaire, par le syst~me consid6r6, des chalnes incompl~tement form6es les plus longues en raison n (I) de l'existence de 2 r6sidus de tyrosine dans le tiers C-terminal des chalnes x et fl et (2) de la biosynth~se s6quen- tielle des chalnes ~ partir de l'extr~mit6 N--terminale. La position tr~s voisine des peptides contenant la DOPA et de ceux renfermant la tyrosine sur les "fingerprints", peut s'expliquer, soit par des diff6rences de propri6t6s physico-chimiques entre ces deux types de mol6cules, trop faibles pour entrainer une diff6rence de migration dans les conditions de chromato-61ectrophor~se utilis6es, soit par les quantit6s trop petites de DOPA-peptides qui seraient "entrain6es" par les peptides normaux A tyrosine, soit par les deux.
Dans les conditions d6crites, les ribosomes repr6sentaient le facteur limitant de la synth~se. En effet, si leur quantit6 ~tait doubl~e par rapport au s-RN'A, 15-2o % de la tyrosine 6taient retrouv6s dans le pr6cipit6 trichloroac6tique. I1 a 6t6 par ailleurs v6rifi6 que, dans les conditions employ6es, le s-RNA total de E. coli ne pouvait pas 6tre charg6 avec la DOPA libre et que, durant l'incubation avec les ribosomes, la DOPA li6e au s-RN'A n'6tait pas r6duite en tyrosyl-s-RN'A.
Les r6sultats "de ces exp6riences montrent que, dans un syst~me synth6tisant une prot6ine sp6cifique, un acide amin6 inhabituel comme la DOPA, lorsqu'il est introduit combin6 au s-RNA sp6cifique de la tyrosine, est incorpor6 dans cette prot6ine et retrouv6 dans les s6quences peptidiques qui contiennent normalement la tyrosine.
Nous remercions le Professeur KERTESZ et le Dr. ZITO (Tunis, Rome) pour la pr6paration de polyph6noloxydase qu'ils ont mis a notre disposition.
Ddpartement de Biologie, Centre d'Etudes Nucldaires, Gi[-sur-Yvette (Seine et Oise) et Biochimie Mddicale, Facultd de Mddecine et de Pharmacie, Marseille (France)
FRANCOIS CHAPEVILLE
GUY CARTOUZOU
SERGE LISSITZKY
1 F. CHAPEVILLE, F. LIPMANN, G-. VON EHRENSTEIN, B. WEISBLUM, W. J. RAY ET S. BENZER, Proc. Natl. Acad. Sci. U. S., 48 (1962) lO86.
2 F. CHAPEVILLE, Federation Proc., 2I (1962) 414 d. s F. H. C. CRICK, Syrup. Soc. Exptl. Biol., 12 (I958) 138. 4 G. VON EHRENSTEIN ET F. LIPMANN, Proc. Natl. Acad. Sci. U.S., 47 (1961) 941. 6 D. I~ATHANS ET F. LIPMANN, Proc. Natl. Acad. Sci. U.S., 47 (I961) 497. s S. LISSITZKY ET M. ROLLAND, Biochim. Biophys. Acta, 56 (1962) 95- 7 D. KERTESZ ET R. ZITO, Nature, 179 (1957) lO17. 8 E. H. ALLEN ET R. S. SCHWEET, J. Biol. Chem., 237 (I962) 760. 9 M. A. HENNESSEY, A. M. HAFFNER ET B. W. GABRID, Federation Proc., 19 (196o) 65.
xo A. M. KATZ, W. J. DREYER ETC. B. ANFINSEN, J. Biol. Chem., 234 (1959) 2897. 1~ M. A. NAUGHTON ET H. M. DINTZlS, Proc. Natl. Acad. Sci. U.S., 48 (1962) 1822. xi j . M. DIAMOND ET G. BRAUNITZER, Nature, 194 (1962) 1287.
Re~u le 17 Janvier, 1963.
Biochim. Biophys. Acta, 68 (1963) 496--499





























