Embed Size (px)
Citation preview

1
TRAVAUX DIRIGÉS
DE BIOCHIMIE
L1S2
ANNÉE UNIVERSITAIRE 2011-2012 C. LACOMBE
C. MORIN
Sommaire LES ACIDES AMINES ET LES PROTEINES
Propriétés acido-basiques des acides aminés et des peptides
Structures
Masse molaire – Techniques de séparation
p 1
p 1
p 3
p 5
ENZYMOLOGIE
LES ACIDES NUCLEIQUES
Bases hétérocycliques et nucléotides
Détermination de séquence de polynucléotides
Thermodénaturation de l’ADN et test ADN
p 8
p 14
p 14
p 17
p 19
2
LES ACIDES AMINÉS ET LES PROTÉINES PROPRIÉTÉS ACIDO-BASIQUES DES ACIDES AMINÉS ET DES PEPTIDES
1 Parmi les acides aminés constitutifs des protéines, indiquer ceux qui sont chargés positivement ou
négativement à pH 7.
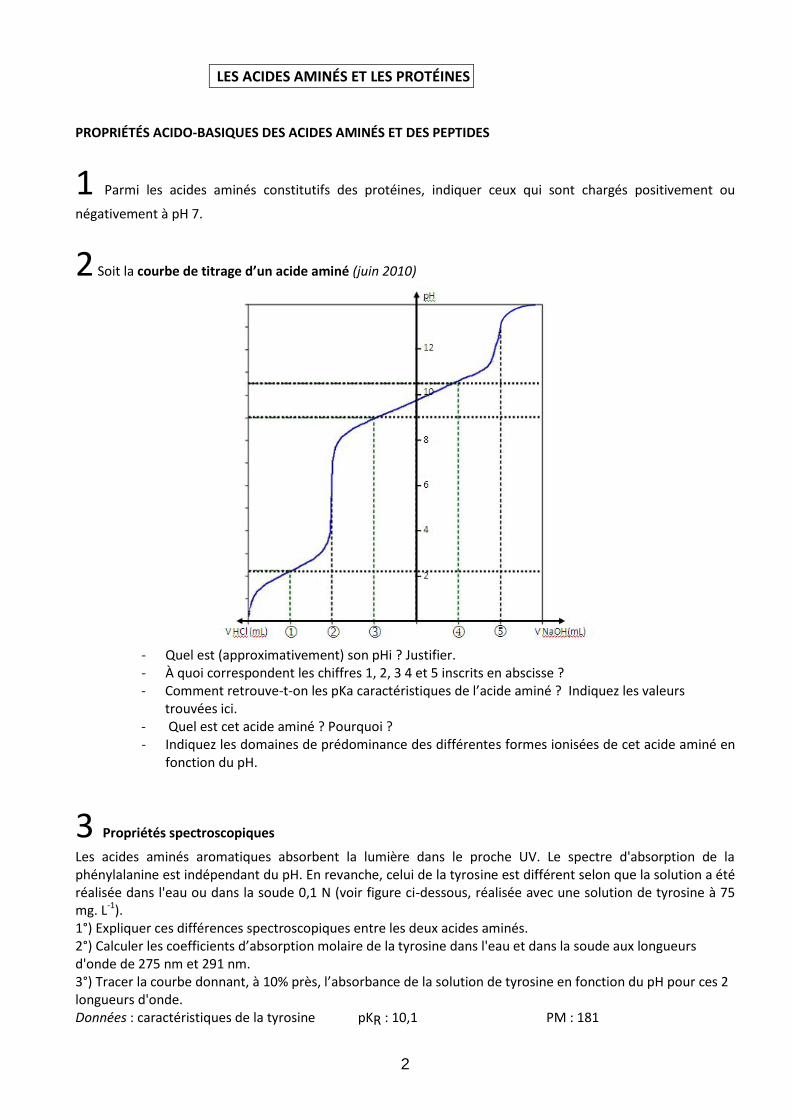
2 Soit la courbe de titrage d’un acide aminé (juin 2010)
- Quel est (approximativement) son pHi ? Justifier. - À quoi correspondent les chiffres 1, 2, 3 4 et 5 inscrits en abscisse ? - Comment retrouve-t-on les pKa caractéristiques de l’acide aminé ? Indiquez les valeurs
trouvées ici. - Quel est cet acide aminé ? Pourquoi ? - Indiquez les domaines de prédominance des différentes formes ionisées de cet acide aminé en
fonction du pH.
3 Propriétés spectroscopiques
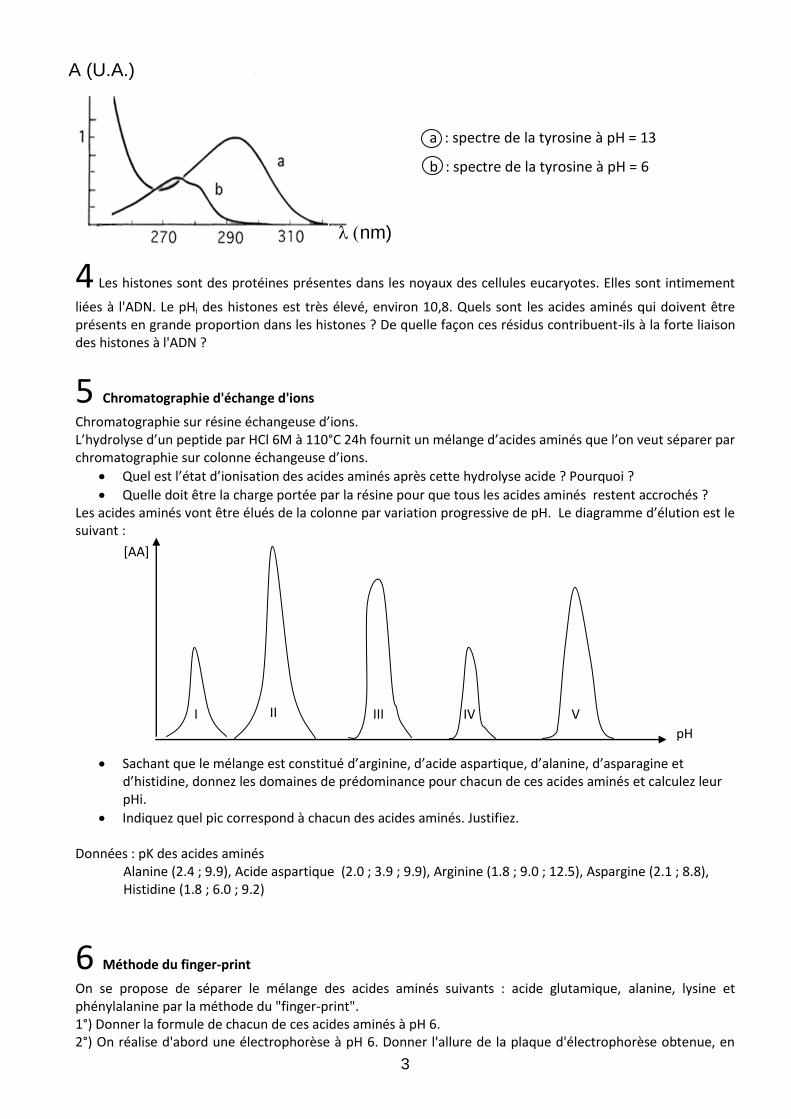
Les acides aminés aromatiques absorbent la lumière dans le proche UV. Le spectre d'absorption de la phénylalanine est indépendant du pH. En revanche, celui de la tyrosine est différent selon que la solution a été réalisée dans l'eau ou dans la soude 0,1 N (voir figure ci-dessous, réalisée avec une solution de tyrosine à 75 mg. L-1). 1°) Expliquer ces différences spectroscopiques entre les deux acides aminés. 2°) Calculer les coefficients d’absorption molaire de la tyrosine dans l'eau et dans la soude aux longueurs d'onde de 275 nm et 291 nm. 3°) Tracer la courbe donnant, à 10% près, l’absorbance de la solution de tyrosine en fonction du pH pour ces 2 longueurs d'onde. Données : caractéristiques de la tyrosine pKR : 10,1 PM : 181
3
4 Les histones sont des protéines présentes dans les noyaux des cellules eucaryotes. Elles sont intimement
liées à l'ADN. Le pHi des histones est très élevé, environ 10,8. Quels sont les acides aminés qui doivent être présents en grande proportion dans les histones ? De quelle façon ces résidus contribuent-ils à la forte liaison des histones à l'ADN ?
5 Chromatographie d'échange d'ions
Chromatographie sur résine échangeuse d’ions. L’hydrolyse d’un peptide par HCl 6M à 110°C 24h fournit un mélange d’acides aminés que l’on veut séparer par chromatographie sur colonne échangeuse d’ions.
Quel est l’état d’ionisation des acides aminés après cette hydrolyse acide ? Pourquoi ?
Quelle doit être la charge portée par la résine pour que tous les acides aminés restent accrochés ? Les acides aminés vont être élués de la colonne par variation progressive de pH. Le diagramme d’élution est le suivant :
Sachant que le mélange est constitué d’arginine, d’acide aspartique, d’alanine, d’asparagine et d’histidine, donnez les domaines de prédominance pour chacun de ces acides aminés et calculez leur pHi.
Indiquez quel pic correspond à chacun des acides aminés. Justifiez.
Données : pK des acides aminés Alanine (2.4 ; 9.9), Acide aspartique (2.0 ; 3.9 ; 9.9), Arginine (1.8 ; 9.0 ; 12.5), Aspargine (2.1 ; 8.8), Histidine (1.8 ; 6.0 ; 9.2)
6 Méthode du finger-print
On se propose de séparer le mélange des acides aminés suivants : acide glutamique, alanine, lysine et phénylalanine par la méthode du "finger-print". 1°) Donner la formule de chacun de ces acides aminés à pH 6. 2°) On réalise d'abord une électrophorèse à pH 6. Donner l'allure de la plaque d'électrophorèse obtenue, en
nm)
A (U.A.)
a : spectre de la tyrosine à pH = 13
b : spectre de la tyrosine à pH = 6
pH
[AA]
I II III IV V

4
précisant le(s) acide(s) aminé(s) correspondant à chaque tache observée. 3°) On effectue ensuite une chromatographie de partage dans la direction perpendiculaire à celle de l'électrophorèse. L'une des taches se dédouble. Préciser laquelle et indiquer, en le justifiant, quel est l'acide aminé qui est entraîné le plus loin.
STRUCTURES
7Soit le peptide AYPKKL, donner à pH 7 et 12 l'état ionique de ce peptide. On rappelle que les pKa des
différents groupements d'un peptide diffèrent au maximum d'une unité de ceux des acides aminés.
8 Donner les structures possibles d'un pentapeptide connaissant les résultats expérimentaux suivants :
1°) L'hydrolyse totale aboutit à la formation des composés suivants : Gly, Phe, Lys, Met, Ala. De plus, l’action ménagée du réactif d'Edman libère un PTH-Gly. 2°) Après l'action de la trypsine ou de la chymotrypsine, il se forme un dipeptide et un tripeptide. 3°) Le bromure de cyanogène n'agit par sur ce composé. Quelle expérience feriez-vous pour lever l'ambiguïté ?
9 Dans le but de déterminer la structure d'un peptide dérivé d'une hormone H, on réalise un certain nombre
d'expériences décrites ci-dessous. Pour chacune des expériences proposées, on indiquera clairement le rôle des réactifs utilisés ainsi que les conclusions que l'on peut tirer du résultat obtenu. (Si un réactif intervient au niveau de plusieurs expériences, on ne précisera qu'une fois son rôle). 1°) Un traitement par HCl 6N à chaud aboutit au mélange d'acides aminés suivants : Glu, Arg, Gly, His, Leu, Pro, Ser, Tyr. Tous ces acides aminés sont présents à la même concentration, sauf la glycine qui est à concentration double des autres. L'action courte d'une aminopeptidase libère un Glu. 2°) L'action de la chymotrypsine aboutit à la formation de trois peptides : P1, P2 et P3.
3°) Un traitement par HCl 6N à chaud sur chacun des peptides ci-dessus permet d'isoler : - pour P1 : Glu et His
- pour P2 : Ser et Tyr
- pour P3 : Gly, Arg, Leu et Pro (avec Gly en concentration double)
4°) L'action de la trypsine sur P3 aboutit à la formation d'un tripeptide P4 et d'un dipeptide P5. L'action du PITC
sur le tripeptide P4 et sur le dipeptide P5 permet de caractériser respectivement PTH Gly et PTH-Pro. L'action
ménagée d'une carboxypeptidase libère la leucine à partir de P3 ainsi qu'à partir de P5. Donner la (ou les)
structure(s) du peptide H initial compatible (s) avec l'ensemble de ces résultats. 5°) En fait, dans l'hormone native, le groupement -COOH terminal est amidifié (-CONH2). Selon les conditions
d'extraction du peptide H, celui-ci pourra donc se terminer soit par une fonction acide carboxylique, soit par une fonction amide. Sachant que lors d'une électrophorèse à pH 7, ce peptide migre vers la cathode, on déduira la nature de la fonction effectivement présente en bout de chaîne. N. B. : On rappelle que le pKa de la chaîne latérale de l'histidine est égal à 6.
10 Après purification d’une protéine, une hydrolyse trypsique est réalisée. L’un des peptides P va être
soumis aux différentes expériences suivantes. Pour chacune de celles-ci, il est demandé de rappeler l’action des réactifs utilisés et d’en tirer le maximum de conclusions.
a) La composition en acides aminés de cet hexapeptide a été déterminée après hydrolyse acide. Elle
permet d’obtenir un mélange des acides aminés suivants en proportions stœchiométriques : Glu, Lys, Met, Ser,
Val.
b) L ‘action courte du réactif d’Edman libère du PHT-Val.
c) Après action du bromure de cyanogène, on obtient un dipeptide P1 et un tétrapeptide P2. L’action
courte de l’aminopeptidase libère une valine à partir de P1 et une sérine à partir de P2.

5
d) Après action de la protéase V8 de Streptococcus aureus libère un tétrapeptide P3 et un dipeptide P4.
L’action courte de l’aminopeptidase libère une valine à partir de P3.
Quelle structure de P est compatible avec l’ensemble de ces résultats ?
11 Dans le but de déterminer la structure primaire d'un peptide A, on réalise les expériences suivantes :
1°) Une hydrolyse totale en milieu HCl 6N à chaud permet d'obtenir un mélange équimoléculaire de Gly, Glu, Arg, Asp, Cys-S-S-Cys, Leu, Lys, Met, Tyr, Val. L’action ménagée du réactif d'Edman libère un PTH-Gly et un PTH-Gln. 2°) L'action du -mercaptoéthanol permet d'isoler deux peptides B et C. 3°) On réalise l'hydrolyse de B par voie enzymatique : a) En présence de chymotrypsine, on obtient un peptide et de la valine. b) En présence de trypsine, on obtient 2 tripeptides B1 et B2 dans lesquels on caractérise, entre autres,
après séparation et action du PITC, du PTH-Gly pour B1 et du PTH-Cys pour B2.
4°) L'action du bromure de cyanogène (BrCN) sur C permet d'obtenir un peptide et de l'acide aspartique; 5°) L'action de la trypsine sur C permet d'isoler un tripeptide C1 et un dipeptide C2. L'action du PITC sur C1
permet d'isoler du PTH-Gln.
Pour chacune des expériences présentées ci-dessus, on indiquera clairement la spécificité de l'enzyme ou du réactif utilisé, ainsi que les conclusions que l'on peut tirer des résultats obtenus. On en déduira la (ou les) structure(s) possible(s) pour chacun des peptides B et C puis A.
12 1°) L'hydrolyse acide d'une millimole d'un pentapeptide fournit 2 millimoles d'acide glutamique et 1
millimole de lysine. L'hydrolysat ne contient pas d'autre acide aminé identifiable. 2°) L'action de la trypsine coupe le pentapeptide en deux peptides dont l'un migre vers la cathode et l'autre vers l'anode lors d'une électrophorèse réalisée à pH 7. Le traitement de l'un de ces fragments par le PITC donne du PTH-Trp . 3°) La chymotrypsine hydrolyse le pentapeptide initial en deux dipeptides et de l'acide glutamique. Proposer une séquence compatible avec l'ensemble de ces données.
13 Un peptide est composé de 30 acides aminés. Sa composition globale, obtenue après hydrolyse acide,
est la suivante : Gly 3 Leu 3 Ser 2 Phe 2 Arg 2 Glu 3 Ala 3 Ile 1 Thr 1 Tyr 1 Lys 1 Met 1 Val 3 Pro 2 Cys 1
Un traitement par la trypsine permet d'isoler 4 peptides A, B, C et D dont les séquences sont analysées indépendamment. 1°) La composition en acides aminés du peptide A est la suivante : 2 Ala, Tyr, Ile, Lys. L'action du PITC libère 1 PTH-Ile et celle de la chymotrypsine 2 peptides, un dipeptide et un tripeptide. A l'aide de ces résultats, déterminer la ou les séquences du peptide A. Quelle autre méthode permettrait de résoudre entièrement cette séquence ? 2°) Les séquences des peptides B, C et D sont obtenues partiellement par la méthode récurrente d'Edman. Les résultats obtenus sont : B : Met - Ala - Ser - (4) - Leu - Thr - Arg C : Gly - Trp - Leu - (4) - Pro - Gly - Phe - Ser - Arg D : Val - Glu - Leu L'action de la chymotrypsine sur le peptide initial libère 5 peptides dont les séquences sont les suivantes : E : Leu - Pro - Gln - Cys - Val - Pro - Gly - Phe F : Val - Glu - Leu - Thr - Arg - Ile - Tyr

6
G : Met - Ala - Ser - Gly - Phe H : Ala - Ala - Lys - Gly - Trp I : Ser - Arg - Val - Glu -Leu Quelle est la séquence du peptide initial ?
MASSE MOLAIRE - TECHNIQUES DE SÉPARATION
14 Concentration in vivo
E. coli est une bactérie approximativement cylindrique d'environ 2 µm de longueur et de 1 µm de diamètre. Si on la fait croître sur lactose, elle contient de la β-galactosidase (masse molaire 540 000). La masse volumique moyenne de la cellule bactérienne est égale à 1,2 g.mL-1. La masse des protéines représente 14% de la masse totale. La masse de la β-galactosidase est 1% de la masse des protéines. Calculer le nombre de molécules de β-galactosidase présentes par cellule.
15 Analyse pondérale et SDS-PAGE (examen juin 2003)
Une protéine impliquée dans les interactions avec l’ADN a été isolée.
1°) Un micro dosage a montré la présence de Zinc dans cette protéine à raison de 40 ng de Zinc pour 55 g de protéine. Le poids atomique du zinc est 65.
Dans l’hypothèse la plus simple, quel est le Poids Moléculaire (PM) de cette protéine ? 2°) On réalise une électrophorèse en conditions dénaturantes (SDS-PAGE) de cette protéine. On obtient le résultat suivant : Ligne de dépôt
Définir le terme SDS-PAGE et décrire brièvement le principe de cette technique. La distance de migration (d) de chacune des protéines a été mesurée.
PM logPM d (cm)
Phosphorylase 97 000 4,99 0,5
Sérumalbumine bovine 66 000 4,82 1,4
Glutamate déshydrogénase 55 000 4,74 1,9
Ovalbumine 45 000 4,65 2,3
Glucose-3-phosphate déshydrogénase 36 000 4,55 2,85
Trypsinogène 24 000 4,38 3,8
X 2,15
Y 2,5
Tracer la droite log(PM) = f(d) en prenant pour abscisse 4 cm = 1cm de migration et en ordonnée 2 cm pour 0,1 unité de log.
X Y
Phosphorylase
Sérumalbumine bovine Glutamate déshydrogénase
Trypsinogène
Glucose-3phosphate déshydrogénase
Ovalbumine

7
Déterminez la masse correspondant aux bandes X et Y observées sur le gel.
X = Y =
Données : tableau de conversion logPM/PM (les valeurs ont été arrondies pour les PM)
logPM 4,56 4,57 4,58 4,59 4,60 4,61 4,62 4,63 4,64
PM 36 300 37 100 38 000 38 900 39 800 40 700 41 700 42 700 43 600
logPM 4,65 4,66 4,67 4,68 4,69 4,70 4,71 4,72 4,73
PM 44 700 45 700 46 800 47 900 49 000 50 100 51 300 52 500 53 700
Qu’en déduisez-vous sur la protéine (taille, composition) ?
3°) Après action de la chymotrypsine sur cette protéine et séparation par chromatographie des peptides
ainsi générés, la structure primaire de l’un des peptides, P1, a été obtenue comme suit. Pour chacune des expériences réalisées, il est demandé de rappeler l’action des réactifs utilisés et d’en tirer le maximum de conclusions.
a - La composition en acides aminés de cet heptapeptide a été déterminée après hydrolyse acide. Elle permet d’obtenir un mélange des acides aminés suivants en proportions stœchiométriques : Met, Lys, Arg, Glu, Ile et Leu.
b - L’action courte de l’aminopeptidase libère une leucine. c - Après action de la trypsine, on obtient 2 dipeptides P2 et P3 et un tripeptide P4.
Une action courte du réactif d’Edman libère du PTH-Leu à partir de P2, PTH-Ile à partir de P3 et PTH-Glu à partir de P4.
d - Après action du bromure de cyanogène sur P1, obtient un tripeptide P5 et un tétrapeptide P6. Une action limitée du réactif d’Edman libère du PTH-Lys à partir de P5 et du PTH-Leu à partir de P6.
Quelle structure de P1 pouvez-vous déduire de l’ensemble de ces résultats ?
16 La plupart des protéines sont insolubles dans l'eau pure, solubles dans les solutions salines diluées, puis
à nouveau insolubles dans les solutions salines concentrées. Expliquer comment on pourrait utiliser cette propriété pour séparer deux protéines A et B dont les courbes de solubilité sont données ci-dessous.
Concentration de (NH4)2SO4 M
Solu
bili
té m
g/m
L
8
CONFORMATION
18Parmi les acides aminés suivants, indiquer ceux qui seront probablement en surface ou à l'intérieur
d'une protéine globulaire : Asp, Leu, Val, Gln, Lys.
19 Structure tertiaire des protéines
L'insuline est une hormone constituée de 2 chaînes peptidiques contenant 3 ponts disulfures (dont 2 unissent les deux chaînes). La réduction par le -mercaptoéthanol donne un mélange de polypeptides inactifs que l'on n'a jamais réussi à renaturer. Quelles conclusions peut-on tirer de cette expérience ? (On a observé que la biosynthèse de l'insuline passe par une pro-insuline monocaténaire).
20 (examen juin 2006) Soit une protéine représentée par le schéma suivant :
Po
uvo
ir r
ota
toir
e sp
écif
iqu
e
0 2 4 6 8 10 12
14
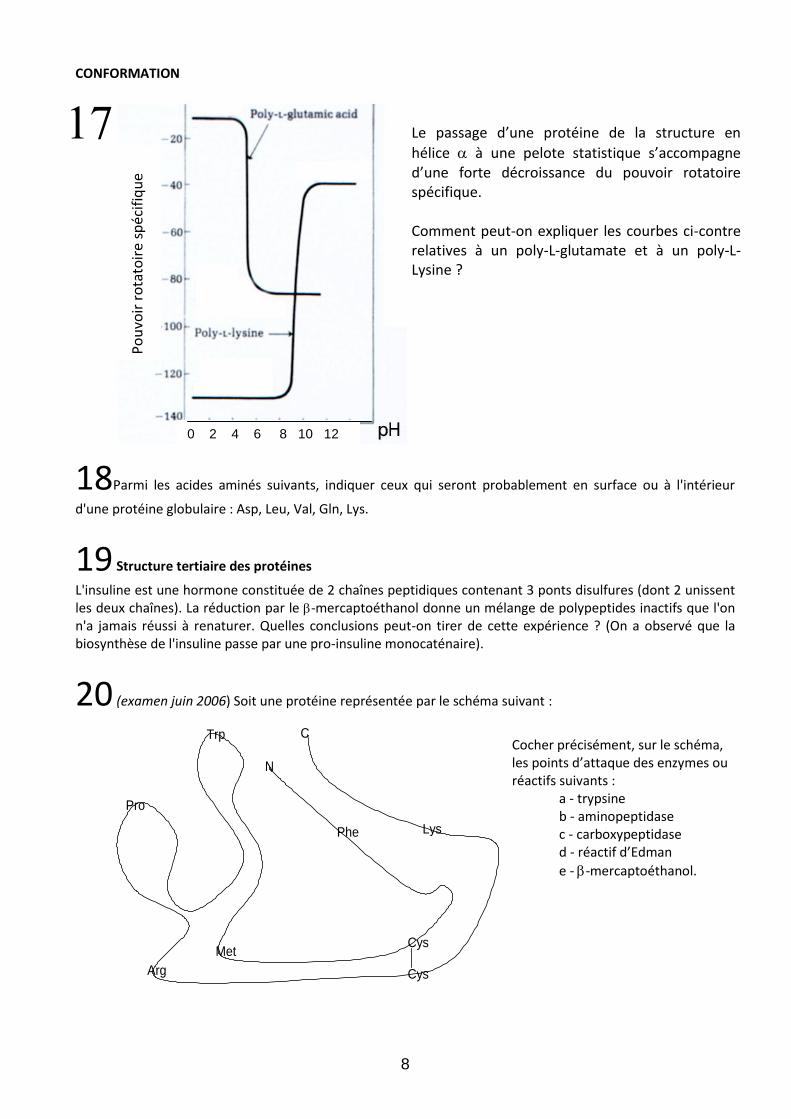
Le passage d’une protéine de la structure en
hélice à une pelote statistique s’accompagne d’une forte décroissance du pouvoir rotatoire spécifique. Comment peut-on expliquer les courbes ci-contre relatives à un poly-L-glutamate et à un poly-L-Lysine ?
17
C
Lys
Cys Cys
Met
Phe
N
Pro
Arg
Trp Cocher précisément, sur le schéma, les points d’attaque des enzymes ou réactifs suivants : a - trypsine b - aminopeptidase c - carboxypeptidase d - réactif d’Edman
e - -mercaptoéthanol.

9
INTRODUCTION À L’ENZYMOLOGIE
21 Examen de janvier 2003
Une enzyme E catalyse la réaction SP. En déterminant l'activité de l'enzyme à différentes concentrations de substrat, avec une quantité constante d'enzyme de 10-4g, on obtient les paires de valeurs suivantes :
S0 (µM) 0 5 10 20 50 100 200 300
Vo (µmol min-1) 0 20 30 40 50 55 60 60
Estimez sans tracer de courbe les valeurs de Km et de Vmax.
En utilisant la linéarisation, déterminer avec précision les valeurs de Km et Vmax. Comparez ces valeurs avec les résultats précédents.
En sachant que le masse molaire de cette enzyme est de 105 g mol-1, calculez son activité spécifique molaire.
Quel est le pourcentage d'enzyme libre par rapport à l'enzyme totale, à une concentration de substrat de 10 µM ? Justifiez.
Quelle sera la vitesse de la réaction en présence de 200 µM de S, si on double la quantité de l'enzyme? Qu'en serait-il à 20 µM de S ?
22 Examen de juin 2010
La vitesse d’une réaction enzymatique en fonction de la concentration du substrat est indiquée dans le tableau suivant :
Vo 33 25 20 12,5 8 [S] 6,6 x 10-2 3,3 x 10-2 2,2 x 10-2 1,1 x 10-2 0,65 x 10-2
1/Vo 3 x 10-2 4 x 10-2 5 x 10-2 8 x 10-2 12 x 10-2 1/[S] 15 30 45 90 150
Vo est exprimée en moles de produit formé par minute et par gramme d’enzyme [S] est exprimée en M.
a - Tracer la courbe de Lineweaver-Burk en utilisant 2 cm pour 1M-1 et 1 cm pour 1 10-2 gramme d’enzyme.minute.mole de produit formé-1 b – Déterminer graphiquement KM et Vmax de cette enzyme c – Tracer la courbe correspondant à la cinétique de la réaction lorsque l’on ajoute au milieu un analogue structural du substrat.
23La trypsine hydrolyse totalement l'ester para-nitrophénolique de la N-carbobenzoxy-lysine (CBZ-Lys-
ONP). La réaction de libération du para-nitrophénol (l'un des produits de la réaction) est suivie par mesure de l'absorbance à 410 nm. A cette longueur d'onde, le para-nitrophénol a une absorbance différente de celle du substrat.
1°) L'hydrolyse enzymatique du substrat est effectuée dans les conditions suivantes : S = 2.10-4 M; pH = 7,8; T = 25°C ; le volume réactionnel est de 3 mL ; les cuves de mesure du spectrophotomètre ont un parcours optique de 1 cm. Le tableau suivant donne les valeurs d'absorbance mesurée à 410 nm, à différents temps et pour différentes concentrations d’enzyme.

10
Temps en min Conditions 1
E0 = 0,24 µg.mL-1 Conditions 2
E0 = 0,12 µg. mL-1 Conditions 3
E0 = 0,06 µg. mL-1
1 0,24 0,12 0,06
2 0,48 0,24 0,12
3 0,72 0,36 0,18
4 0,90 0,48 0,24
5 0,96 0,60 0,30
6 1,00 0,72 0,36
7 1,00 0,81 0,42
8 1,00 0,89 0,48
9 1,00 0,95 0,54
10 1,00 0,98 0,60
20 1,00 1,00 1,00
a) Tracez les courbes correspondant à l'hydrolyse du substrat en fonction du temps, pour les différentes concentrations d'enzyme et en déduire les valeurs des vitesses initiales exprimées en UI en sachant que εM à 410 nm = 5000 M-1cm-1.
b) Quelles sont les concentrations de produits, exprimées en µM, apparues au bout 20 minutes ?
c) Quels sont les nombres d’unités d’enzyme présentes par mL de cuve pour chaque expérience ? Combien y a-t-il d’unités dans la cuve à chaque expérience ?
d) On introduit dans la cuve 0,18 µg.mL-1 d’enzyme. Pour la même concentration en substrat (2.10-4M), quelles sont les concentrations de produits apparus au bout de 2 min et de 20 min ?
e) Tracer la courbe Vo = f(Eo) à partir de ces différentes expériences.
f) Calculer l’activité spécifique en µmol.min-1.mg-1.
2°) Dans une deuxième série d'expériences, on détermine la vitesse initiale d'hydrolyse pour différentes concentrations en substrat (la concentration d'enzyme étant identique dans chacune des expériences et égale à 0,12 µg. mL-1) .
S0 (µM ) 1 2,5 5 7,5 10 25 50 75 100
Vo (10-2 ΔU.A.min-1
) 1,15 2,5 4,2 5,4 6,3 9 10,5 11,1 11,5
a) Exprimer Vo en UI. Tracer la courbe Vo = f (S0). Déterminer les paramètres cinétiques de la préparation: VM et KM.
Calculer kcat en s-1, sachant que la masse molaire de l'enzyme est de 24 000 g.mole-1.
b) Quelles seraient les valeurs de ces paramètres si la concentration d'enzyme utilisée était de 0,06 µg mL-1.
24 1°) L’étude de la cinétique d’une réaction enzymatique montre en particulier que pour une
concentration de substrat (de poids moléculaire 200) égale à 50 µg. mL-1, la vitesse de réaction est la moitié de la vitesse maximale observée en présence d’une concentration saturante de substrat. Est-il possible, à partir
de ces seules données, de définir la constante de Michaelis KM de ce système ? Justifiez votre réponse.
2°) À partir des valeurs suivantes, déterminez graphiquement les constantes du système : Vmax et KM. Justifiez.
[S] M 10-3 1,6 .10-4 10-4 6,6.10-5 4,6. 10-5
Vo * 0,285 0,14 0,10 0,077 0,055
* la vitesse initiale est indiquée en micromoles de produit formées en 10 minutes.

11
3°) La même série de mesures est réalisée en présence d’un inhibiteur à la concentration de
10-5 M. L’analyse graphique des résultats montre que la vitesse maximale est inchangée et que l’intersection de la droite obtenue expérimentalement avec l’axe des abscisses (1/*S+) définit un point à mi-distance entre l’origine et le point d’intersection préalablement déterminé en l’absence d’inhibiteur.
- Tracer cette droite. - De quel type d’inhibition s’agit-il ? Justifier votre réponse.
- Quelle est la valeur de la constante d’affinité (KI) de l’inhiteur pour l’enzyme ?
25 L’acide salicylique inhibe l’action catalytique de la glutamate déshydrogénase. La concentration en
acide salicylique est supposée constante et égale à 40 mM. À partir des données ci-dessous, 1°) déterminez le type d’inhibition (compétitive ou non compétitive). 2°) calculez le KM pour l’enzyme et calculer KI, constante de dissociation du complexe enzyme-
inhibiteur.
S (mM) 1,5 2,0 3,0 4,0 8,0 16
1/S 0,67 0,50 0,33 0,25 0,125 0,062
Sans acide salicylique P (mg.mn-1
) 0,21 0,25 0,3 0,38 0,4 0,44
1/P 4,76 4,0 3,33 3,03 2,5 2,27
Avec acide salicylique P (mg.mn-1) 0,085 0,1 0,12 0,13 0,16 0,18
1/P 11,7 10,0 8,3 7,5 6,25 5,55
26 La malate déshydrogénase catalyse la réduction de l’oxaloacétate en malate. La réaction globale peut
s’écrire : Oxaloacétate + NADH + H+ Malate + NAD+
Les activités correspondant à cette réaction ont été mesurées en absence et en présence d’un inhibiteur dont la concentration était de 2mM. La quantité d’enzyme présente était de 10-4g. Les résultats obtenus ont permis de tracer le graphe de Lineweaver-Burk (1/Vo = f(1/S)) ci-dessous :
-2,0 -1,5 -1,0 -0,5 0,0 0,5 1,0 1,5 2,0
0
1
2
3
4
1/Vo (µmol-1min)
1/S (mM-1)
A
B

12
1- Indiquez quelle est la droite A ou B correspondant à l’expérience en présence de l’inhibiteur et le type d’inhibition correspondant
2- Déterminez Vmax et KM en absence d’inhibiteur et les constantes apparentes Vmaxapp , et KMapp en sa présence.
3- Calculez la constante d’inhibition (KI). 4- La masse molaire de l’enzyme étant de 5.106g mol-1, calculez l’activité spécifique molaire (kcat).
5- Quelles seraient les valeurs de KM, Vmax et kcat que l’on aurait déterminées si l’expérience avait été effectuée en utilisant une quantité d’enzyme de 5.10-5g.
27 À pH 7,7 et à 25°C, les cinétiques d'hydrolyse de l'o-nitrophényl-galactoside par la
–galactosidase de Escherichia coli ont été étudiées en absence ou en présence de l'un des inhibiteurs
suivants: o-nitrophényl--D-thiogalactoside (ONPTG), maltose.
Les mesures de vitesses initiales pour chacune de ces expériences sont données dans le tableau suivant comme la variation de l'absorbance à 410 nm (longueur d'onde d'absorption maximale de l'ONP). Le volume réactionnel est de 1 mL.
Vo (ΔUA.min-1)
S (M) Sans inhibiteur
ONPTG (3.10-4M) Maltose (0,25 M)
2,5 10-5 0,033 0,018 0,0165
5,0 10-5 0,055 0,033 0,0275
1,0 10-4 0,0825 0,055 0,041
2,5 10-4 0,118 0,091 0,059
5,0 10-4 0,138 0,118 0,069
1,0 10-3 0,150 0,138 0,075
a) Tracer les courbes Vo = f(S) et 1/Vo = f (1/S) pour chacune des expériences.
b) Comparer les résultats obtenus avec les inhibiteurs à celui obtenu pour l'expérience réalisée en absence d'inhibiteur, et déterminer la nature de l'inhibition pour chacun des inhibiteurs.
c) Déterminer Vmax et KM pour l'expérience réalisée en absence d'inhibiteur.
d) Calculer kcat en sec-1 sachant que [E0 ]= 1,19 10-9 M et que εM de l'ONP à 410 nm = 12 000 M–1cm-1.
e) Déterminer V'max, K'M, et KI pour chacun des inhibiteurs.
Dans chacun des cas précédents, quelle concentration de substrat utiliseriez-vous pour montrer l'effet de l'inhibiteur sur l'activité enzymatique ?
28 On considère les réactions d’hydrolyse des quatre peptides suivants :
Gly-Ser Gly + Ser
Phe-Ser Phe + Ser
Lys-Ser Lys + Ser
Ser-Phe Ser + Phe Pour augmenter la vitesse de cette hydrolyse, on peut envisager d’utiliser la trypsine ou la chymotrypsine. Quels sont, parmi les peptides ci-dessus, ceux pour lesquels l’une ou l’autre de ces enzymes sera a priori efficace (préciser à chaque fois laquelle de deux en justifiant) On étudie l’hydrolyse trypsique d’un dipeptide formé de deux résidus d’acides aminés identiques :
3HN – CH – CO – NH – CH – COO- 2 3HN – CH – COO- + +
13
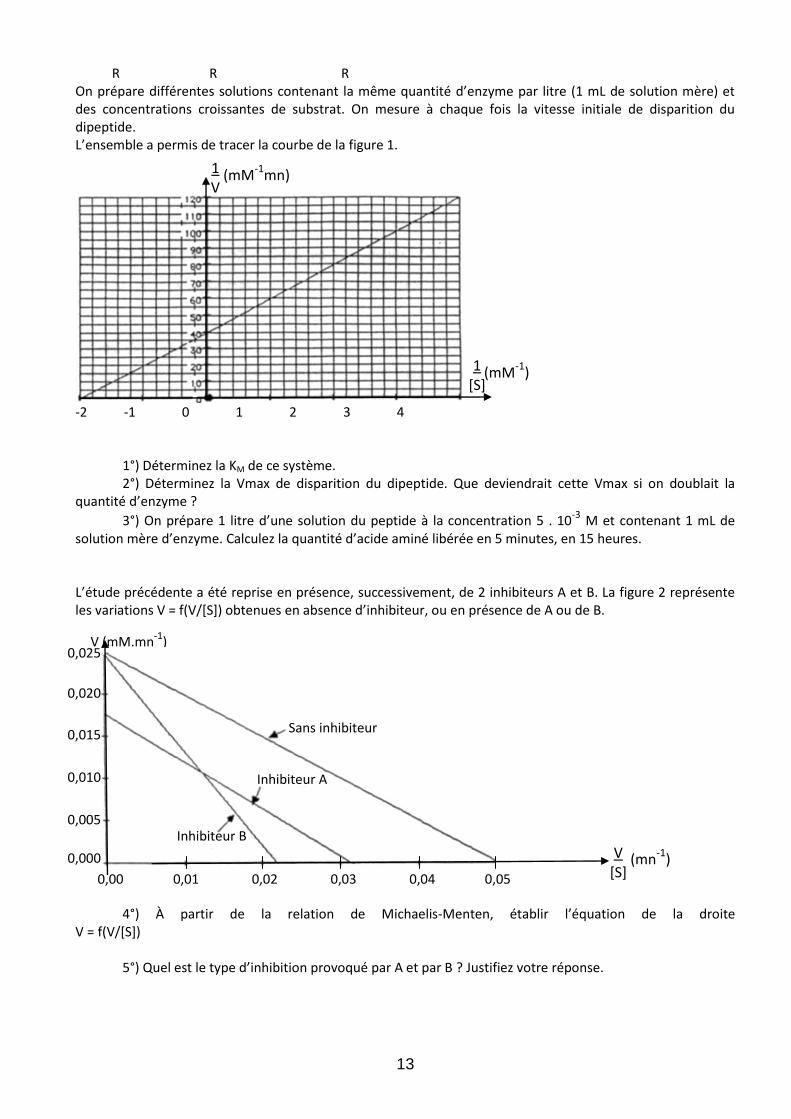
R R R On prépare différentes solutions contenant la même quantité d’enzyme par litre (1 mL de solution mère) et des concentrations croissantes de substrat. On mesure à chaque fois la vitesse initiale de disparition du dipeptide. L’ensemble a permis de tracer la courbe de la figure 1.
-2 -1 0 1 2 3 4
1°) Déterminez la KM de ce système. 2°) Déterminez la Vmax de disparition du dipeptide. Que deviendrait cette Vmax si on doublait la
quantité d’enzyme ?
3°) On prépare 1 litre d’une solution du peptide à la concentration 5 . 10-3 M et contenant 1 mL de solution mère d’enzyme. Calculez la quantité d’acide aminé libérée en 5 minutes, en 15 heures. L’étude précédente a été reprise en présence, successivement, de 2 inhibiteurs A et B. La figure 2 représente les variations V = f(V/*S+) obtenues en absence d’inhibiteur, ou en présence de A ou de B.
4°) À partir de la relation de Michaelis-Menten, établir l’équation de la droite
V = f(V/[S])
5°) Quel est le type d’inhibition provoqué par A et par B ? Justifiez votre réponse.
1 V
(mM-1mn)
1 [S]
(mM-1
)
V (mM.mn-1
)
Sans inhibiteur
Inhibiteur A
Inhibiteur B
0,00 0,01 0,02 0,03 0,04 0,05
0,025
0,020
0,015
0,010
0,005
0,000
V [S]
(mn-1)

14
29 L’aldolase est une enzyme catalysant l’hydrolyse du fructose 1,6-diphosphate :
fructose 1,6-diphosphate → glycéraldéhyde 3-phosphate + dihydroxyacétone 3-phosphate
On dispose d’un sérum humain contenant deux aldolases d’origine tissulaire différente, l’une H d’origine hépatique, l’autre, M d’origine musculaire. Afin d’en déterminer les paramètres cinétiques, on étudie les variations de la vitesse de la réaction, c’est-à-dire l’apparition du nombre de moles de glycéraldéhyde 3-phosphate en une minute en fonction de la concentration en fructose 1,6-diphosphate. Les résultats sont les suivants :
[fructose 1,6-diphosphate]* 0,25.10
-3 0,33.10
-3 0,5.10
-3 1.10
-3
1/[fructose 1,6-diphosphate] 4000 3030 2000 1000
Aldolase M
Nombre de moles de glycéraldéhyde 3-phosphate en 1 mn
0,60.10-3
0,72 .10-3
0,87.10-3
1,11.10-3
1/ N moles glycéraldéhyde 3-phosphate en 1 mn
1667 1389 1149 900
Aldolase H
Nombre de moles de glycéraldéhyde 3-phosphate en 1 mn
0,20.10-3
0,25 .10-3
0,33.10-3
0,50.10-3
1/ N moles glycéraldéhyde 3-phosphate en 1 mn
5000 4000 3030 2000
*Les concentrations sont exprimées de [fructose 1,6-diphosphate] en M
1 – Tracer les courbes V = f([S]) pour les 2 enzymes (en utilisant 1 cm pour 10-4 M et 1 cm pour 10-4 mole.min-1).
En déduire Vmax et KM pour les 2 enzymes. Justifier. 2 – Tracer les représentations de Lineweaver-Burk pour ces 2 enzymes (en utilisant 1cm pour 500 M-1
et 1 cm pour 500 M-1.min-1). En réduire à nouveau Vmax et KM pour les 2 enzymes. Justifier. 3 – Comparer les résultats obtenus par les 2 méthodes. Conclusion ? 4 – Comparer les affinités des 2 aldolases vis-à-vis du fructose 1,6-diphosphate.
30 Influence de la température
On considère l’oxydation d’un alcool catalysée par une alcool déshydrogénase de masse molaire 50 000 :
RCH2OH + NAD+ RCHO + NADH
L’enzyme possède les caractéristiques suivantes à 25 °C :
Activité spécifique moléculaire (turnover) 103 mn-1
Constantes de Michaelis alcool : 2 .10-4 M
NAD+ : 4 .10-5 M
1°) À 10 mL d’une solution d’alcool 4 .10-3 M et de NAD+ 4 .10-4 M, on ajoute 5 µg d’enzyme pure. Quel est le nombre d’unités internationales d’enzyme présentes dans les 10 mL de solution à 25°C ? Quelle est la concentration d’aldéhyde formée en 10 mn ? 2°) On préincube l’enzyme pendant 10 mn à 50°C, puis on reprend l’expérience précédente. Calculer la concentration d’aldéhyde formée en 100 mn dans ce cas, sachant que la constante d’inactivation à cette
température est égale à 2,88 . 10-2 mn-1.

15
31 La vitesse initiale d’une réaction enzymatique est étudiée en fonction de la concentration en substrat.
Quelles sont les propositions compatibles avec les résultats expérimentaux ? A – La droite de l’expérience 1 est obtenue avec une enzyme ayant une plus forte affinité pour le substrat que dans l’expérience 2. B – Dans l’expérience 1, l’enzyme a une Vmax supérieure à celle obtenue dans l’expérience 2. C – La droite de l’expérience 2 est obtenue en présence d’un inhibiteur compétitif absent dans l’expérience 1, l’enzyme étant identique et à la même concentration dans les 2 expériences. D – La droite de l’expérience 2 est obtenue en présence d’un inhibiteur non compétitif absent dans l’expérience 1, l’enzyme étant identique et à la même concentration dans les 2 expériences. E – L’expérience 1 est réalisée avec une concentration d’enzyme double de celle de l’expérience 2.
LES ACIDES NUCLÉIQUES
BASES HÉTÉROCYCLIQUES ET NUCLÉOTIDES
32 Bases hétérocycliques
- Ecrire les formules développées des noyaux puriques et pyrimidiques.
- Donner les formules des 5 bases nucléiques usuelles : Adénine, Guanine, Uracile, Cytosine et Thymine.
- Montrer les possibilités de tautomérie lactime-lactame de ces molécules.
33 Nucléosides
- Mettre en évidence la liaison N-glycosidique. Remarquer que la présence de cette liaison implique des
formes lactame pour l'Uridine, la Cytidine et la Thymidine.
- Quelle est la différence entre les ribonucléosides et les désoxyribonucléosides ?
- Indiquer sur des schémas les interactions lors de l'appariement des bases dans les acides nucléiques.
34 Nucléotides
- Donner la formule développée d'un nucléotide triphosphate.
- Quel est l'état d'ionisation des groupements phosphate à pH 7 ?
1/Vo Expérience 2
Expérience 1
1/[S]

16
35 Structure
- Etant donné un fragment d'un brin d'ADN de structure :
PA
G
C
T
P
P
P
P
P
A
G
1°) Donner la structure du brin complémentaire correspondant.
2°) Donner la formule développée de l’ADN bicaténaire.
36 On donne la composition en bases d'un ADN :
C = 19% A = 25% T = 33% G = 23%
Que peut-on en conclure ?
37 Combien de fonctions OH (alcools) libres contient une molécule d'ADN ? Même question pour une
molécule d'ARN.
38 Une solution contenant un mélange d'AMP et de GMP possède une absorption de 0,652 à 260 mm et
de 0,284 à 280 nm. Connaissant les coefficients d'absorption molaire donnés ci-dessous, calculer les
concentrations respectives de l'AMP et du GMP dans la solution (épaisseur de la cuve = 1 cm).
AMP : ε260 = 15,4 . 103 L.mol-1cm-1 GMP : ε260 = 11,7 . 103 L.mol-1cm-1
AMP : ε280 = 2,5 . 103 L.mol-1cm-1 GMP : ε280 = 7,7 . 103 L.mol-1cm-1
39 L'ADN du corps humain
Calculer la masse en grammes d'une molécule d'ADN bicaténaire s'étirant de la Terre à la Lune
(# 370 000 km). La double hélice a une masse d'environ 10-18g/1 000 paires de nucléotides. Chaque paire de bases s'étend sur environ 0,34 nm. En comparaison, votre corps contient environ 0,5g d'ADN !
17
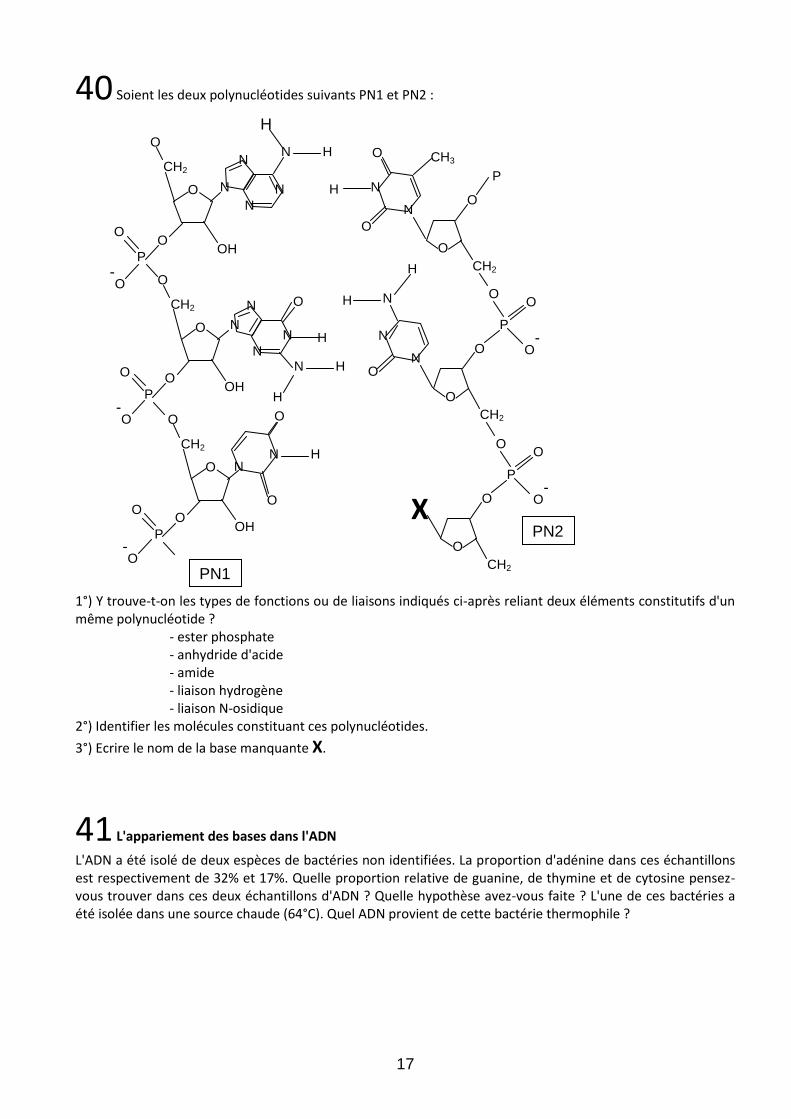
40 Soient les deux polynucléotides suivants PN1 et PN2 :
1°) Y trouve-t-on les types de fonctions ou de liaisons indiqués ci-après reliant deux éléments constitutifs d'un même polynucléotide ? - ester phosphate - anhydride d'acide - amide - liaison hydrogène - liaison N-osidique 2°) Identifier les molécules constituant ces polynucléotides.
3°) Ecrire le nom de la base manquante X.
41 L'appariement des bases dans l'ADN
L'ADN a été isolé de deux espèces de bactéries non identifiées. La proportion d'adénine dans ces échantillons est respectivement de 32% et 17%. Quelle proportion relative de guanine, de thymine et de cytosine pensez-vous trouver dans ces deux échantillons d'ADN ? Quelle hypothèse avez-vous faite ? L'une de ces bactéries a été isolée dans une source chaude (64°C). Quel ADN provient de cette bactérie thermophile ?
H
N H O CH3
N
N
H
O
O
O
P
CH2
O
-O
CH2
O
O
P OH
O
CH2
O
O
N
P OH
O
N
N
N
O N
N
N N
N H
H
H N
O
N
N
H
H
O
O
O
P
CH2
O
O-
CH2
O
O
P OH
O N
O
N H
O
O
O
O
P
CH2
O
PN1
PN2
O
O
-O
-O
O- X

18
DÉTERMINATION DE SÉQUENCE DE POLYNUCLÉOTIDES
42 La technique de séquençage de Sanger a été utilisée pour déterminer la séquence d'un brin d'ADN.
L'autoradiographie obtenue est présentée ci-dessous :
1°) Rappeler le principe de cette méthode 2°) Préciser les positions de l'anode et de la cathode. Justifier. 3°) Déterminer à partir de ce schéma la séquence du brin d'ADN 4°) Préciser où se trouve l'extrémité 5' et l'extrémité 3' de la séquence que vous venez d'écrire.
43 Après l'action d'une enzyme de restriction sur un ADN puis désappariement et séparation des brins, on
obtient un polynucléotide P. Ce brin traité par la méthode biochimique de Sanger a fourni l'autoradiographie suivante :
ddA ddT ddC ddG
1°) Donner brièvement le principe de cette méthode. Lire la séquence. 2°) Quelle est la séquence de P ? 3°) Donner la structure schématique de P en indiquant les liaisons entre chaque nucléotide. (Ne pas expliciter les formules des bases) 4°) Ecrire la structure schématique de l'ADN bicaténaire d'origine en y faisant figurer l'orientation 5'3' et les phosphates.
ddA ddT ddCA
ddG
Sens de la
migration

19
THERMODÉNATURATION DE L'ADN ET TEST ADN
44 (DS mai 2009)
La composition, en bases, d’ADN d’origines différentes a été mesurée à 2% près. ADN %A %C %G %T A 21 28 29 22 B 23 24 28 35 C 18 31 32 19 D 33 17 18 32
1°) Calculer le pourcentage de (A + T) pour chacun de ces ADN. 2°) Dans les mêmes conditions expérimentales, on prépare, pour chacun des ces ADN, une solution dont l’absorbance est de 1,0 U.A. Une dénaturation de ces différents ADN est réalisée par chauffage progressif et, en même temps, l’absorbance de chaque solution est mesurée. Les résultats obtenus sont présentés ci-dessous:
3°) Expliquez le phénomène observé. 4°) A quels ADN correspondent les courbes 1, 2, 3 et 4 ? Justifier votre réponse. Une solution d'ADN bicaténaire est chauffée puis refroidie à la température de la pièce en 2 minutes. Quel sera le changement d'absorption à 260 nm durant le refroidissement dans les trois cas cités ci-dessous ?
si la solution est chauffée juste au-dessous de TM.
si la solution est chauffée juste au-dessus de TM.
45 Une solution d’ADN bicaténaire est chauffée puis refroidie à la température de la pièce en 2 minutes.
Quel sera le changement d’absorption à 260 nm durant le refroidissement dans les 2 cas cités ci-dessous ?
si la solution est chauffée juste au-dessous de TM.
si la solution est chauffée juste au-dessus de TM.
46 - Un échantillon inconnu d'acide nucléique est soumis à diverses expériences :
1°) Une partie de l'échantillon présente un effet hyperchrome important (+ 30%) lorsqu'on le chauffe. Soumis
à un refroidissement rapide, il ne présente alors qu'une faible diminution de l'absorbance à 260 nm.
2°) Une autre partie de l'échantillon est centrifugé en gradient de chlorure de césium. Il forme une bande
unique de densité = 1,770 g.cm-3.
Si on soumet l'acide nucléique à l'expérience (1) (thermodénaturation puis refroidissement rapide) avant la
centrifugation, on obtient une bande de densité = 1,720 g.cm-3 qui ne contient plus que la moitié du matériel
initial. Le contenu du tube de centrifugation est alors mélangé et maintenu à une température intermédiaire
(qui permet l'hybridation), puis re-centrifugé, la bande obtenue a une densité de 1,770 g.cm-3 et correspond à

20
la totalité du matériel.
3°) La même expérience de centrifugation est réalisée sur du matériel incubé dans la soude à pH 12,5. La bande à 1,720 g.cm-3 est retrouvée mais la deuxième partie de l'expérience (hybridation + re-centrifugation) est impossible à réaliser. Quel est la structure de l'acide nucléique initial ?
47 – Les enzymes de restriction sont des endonucléases capables de couper l’ADN double brin en des sites
très spécifiques qui sont généralement des palindromes.
- Retrouver la séquence palindromique dans cet ADN :
5’…ATTGCCGTAGAATTCCGATAGGC…3’
3’…TAACGGCATCTTAAGGCTATCCG…5’
- Indiquer les fragments générés par l’enzyme sachant que ces fragments se termineront par des
extrémités cohésives avec 4 nucléotides qui ‘dépassent’ en 3’.
48 - Dans une recherche récente de paternité, les ADN provenant d’un enfant (I), de sa mère (II) et de son
père présumé (III) ont été traités en même temps que celui provenant d’un individu (IV) qui n’avait aucun lien avec les personnes précédemment citées. L’ADN a été extrait à partir des cellules de la joue prélevées tout simplement avec un coton-tige. Après purification, cet ADN est fragmenté par l’utilisation d’enzymes de restriction. Du fait de la variabilité génétique, les sites de restriction peuvent subir des mutations qui les modifient. Ces sites de restriction ne sont alors plus reconnus par les enzymes, ce qui fait un site de coupure en moins (ou en plus) pour l’ADN. L’ADN de chaque individu sera découpé par plusieurs enzymes qui vont alors générer toute une série de fragments. Ces fragments sont séparés par électrophorèse sur gel. On va comparer la migration des fragments obtenus par les 4 personnes de notre test. Au vu de cette partie du gel présenté, pensez-vous que le père présumé puisse être le père de cet enfant ? N.B. : Il ne faut pas oublier que nous possédons chaque chromosome en double, l’un venant du père, l’autre de la mère.
II I III IV