Embed Size (px)
Citation preview


Universite de sherbrooke
Radiosensibilisation de l’ADN aux electrons hydrates
par les adduits de cisplatine et leur detachement
Par
Behnaz Behmand
Departement de medecine nucleaire et radiobiologie
These presentee a la Faculte de Medecine et des Sciences de la Sante en vue del’obtention du diplome de philosophiae doctor (Ph.D.)en sciences des radiations et imagerie biomedicale
Sherbrooke, Quebec, Canada
Juillet, 2015
Membres du jury d’evaluationPr. Martin Lepage President du jury, Programme de Sciences des radiations
et imagerie biomedicalePr. Leon Sanche Directeur de recherche, Programme de Sciences des radiations
et imagerie biomedicalePr. Darel Hunting Directeur de recherche, Programme de Sciences des
radiations et imagerie biomedicale
Pr. Richard Wagner Evaluateur interne au programme, Programme de Sciencesdes radiations et imagerie biomedicale
Pr. Antonio Conconi Evaluateur externe au programme, Programme demicrobiologie et infectiologie, FMSS
Pr. Christopher J. Wilds Evaluateur externe a l’universite, Department of Chemistry& Biochemistry, Concordia University

Dedicace
A mes parents qui m’ont constamment soutenue ettoujours encouragee.

RESUME
Radiosensibilisation de l’ADN aux electrons hydrates par les adduits decisplatine et leur detachement
Par
Behnaz Behmand
Programme de sciences des radiations et imagerie biomedicaleThese presentee a la Faculte de medecine et des sciences de la sante en vue de
l’obtention du diplome de philosophiae doctor (Ph.D.) en sciences des radiations etimagerie biomedicale, Faculte de medecine et des sciences de la sante, Universite de
Sherbrooke, Sherbrooke, Quebec, Canada, J1H 5N4
Le traitement concomitant de radiotherapie et de chimiotherapie a ameliore le taux desurvie des patients atteints du cancer. Le cisplatine est un des agents chimiotherapeu-tiques les plus utilises, mais les mecanismes d’interaction avec la radiation demeurenttoujours inconnus. Puisqu’une grande partie de la cellule est constituee d’eau, l’effet indi-rect de la radiation est alors important. Parmi les radicaux issus de la radiolyse de l’eau,l’electron hydrate est estime incapable de generer des cassures de brin a l’ADN. Cepen-dant, la modification structurelle et chimique de l’ADN par le cisplatine peut favoriser lacassure de brins. L’etude presentee dans cette these a pour but de comprendre le meca-nisme d’interaction entre les electrons hydrates et l’adduit de cisplatine-oligonucleotide.Pour cela, differents types d’oligonucleotides contenant une ou deux guanines, soit le sited’attachement du cisplatine, sont utilises. Cette interaction induit un detachement del’adduit de cisplatine de l’ADN plutot que des cassures de brin. Des dommages aux basesde l’ADN ont ete observes et quantifies par chromatographie en phase liquide a hauteperformance (HPLC). Les resultats montrent une augmentation des dommages aux basesde l’ADN en presence de cisplatine, soit aux guanines ainsi qu’aux sites qui ne sont pasles sites d’attachement de cisplatine. La constante de vitesse de la reaction des electronshydrates avec le complexe d’oligonucleotide-cisplatine a ete mesuree par la technique deradiolyse pulsee. Les resultats montrent une augmentation de cette constante en presencede cisplatine par rapport au cas d’un oligonucleotide en absence de cisplatine.MOT-CLES : Dommage a l’ADN, cisplatine, electron hydrate, oligonucleotide

TABLES DES MATIERES
RESUME
TABLE DES MATIERES i
LISTE DES FIGURES iii
LISTE DES TABLEAUX v
LISTE DES ABREVIATIONS vi
1 INTRODUCTION 11.1 L’effet concomitant de la radiochimiotherapie . . . . . . . . . . . . . . . 11.2 Adduits de cisplatine-ADN . . . . . . . . . . . . . . . . . . . . . . . . . . 11.3 Interaction des rayonnements ionisants avec la matiere . . . . . . . . . . 5
1.3.1 Effet direct de la radiation sur l’ADN . . . . . . . . . . . . . . . . 61.3.2 Effet indirect de la radiation sur l’ADN . . . . . . . . . . . . . . . 131.3.3 Effet radiosensibilisateur de l’oxygene . . . . . . . . . . . . . . . . 15
1.4 L’echelle de temps des processus chimiques du rayonnement . . . . . . . 171.5 Les dommages induits a l’ADN . . . . . . . . . . . . . . . . . . . . . . . 17
1.5.1 Transfert de charge . . . . . . . . . . . . . . . . . . . . . . . . . . 171.6 Objectifs du projet de recherche . . . . . . . . . . . . . . . . . . . . . . . 21
2 ARTICLE 1 23
3 ARTICLE 2 39
4 ARTICLE 3 56
5 DISCUSSION 715.1 Detachement de l’adduit du cisplatine . . . . . . . . . . . . . . . . . . . . 725.2 Augmentation des dommages aux bases . . . . . . . . . . . . . . . . . . . 745.3 Radiolyse pulsee et la determination de la constante de vitesse . . . . . . 75
6 CONCLUSION ET PERSPECTIVES 79
i

TABLES DES MATIERES ii
REMERCIEMENTS 81
LISTE DES REFERENCES 87

LISTE DES FIGURES
1 INTRODUCTION 11 Avantages theoriques de la radiochimiotherapie . . . . . . . . . . . . . . 22 Structure de l’ADN . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33 Spectre electromagnetique . . . . . . . . . . . . . . . . . . . . . . . . . . 64 Cassures simple et double brins de l’ADN (3≤E≤20 eV) . . . . . . . . . 85 Cassures simple et double brins de l’ADN (0≤E≤5 eV) . . . . . . . . . . 96 Attachement dissociatif dans les differentes bases de l’ADN . . . . . . . . 107 Calcul theorique de l’energie de l’orbitale π∗ et σ∗ en fonction de la lon-
gueur du lien C-O du groupement sucre-phosphate . . . . . . . . . . . . 128 Attachement dissociatif du cisplatine . . . . . . . . . . . . . . . . . . . . 139 Survie cellulaire de CHO dans differentes atmospheres, irradiees par des
rayons X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1610 Differents types de dommage a l’ADN par radiation ionisante . . . . . . . 1911 Transfert d’electron en exces de N, N, N’, N’-tetramethyl-1,5-diaminoaphthalene
(D) a 5-bromo-2’-deoxyuridine (BrU) dans l’ADN . . . . . . . . . . . . . 21
2 ARTICLE 1 231 Gel electrophoresis showing the effect of •OH and hydrated electrons on
oligonucleotides, with and without the cisPt adduct, as a function of irra-diation dose . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
2 Electrophoresis profiles showing the effect of •OH radicals and hydratedelectrons on ODN-GTG and ODN-GTG-cisPt, following irradiation with1000 Gy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
3 Gel electrophoresis showing the effect of hydrated electrons as a functionof the dose of radiation given to the ODN-TGTand given to ODN-TGT-cisPt 31
4 Effect of hydrated electrons as a function of the dose on the ODN-TGT-cisPt and the formation of ODN-TGT . . . . . . . . . . . . . . . . . . . 31
5 Gel electrophoresis showing the effect of hydrated electrons as a functionof the dose to ODN-GTG and ODN-GTG-cisPt . . . . . . . . . . . . . . 32
iii

LISTE DES FIGURES iv
6 Effect of hydrated electrons as a function of the dose to ODN-GTG-cisPtand the formation of ODN-GTG . . . . . . . . . . . . . . . . . . . . 33
7 Dissociative electron-transfer mechanism resulting in bond cleavage bet-ween cisPt and oligonucleotide . . . . . . . . . . . . . . . . . . . . . . . . 34
3 ARTICLE 2 391 HPLC traces for deoxyribonucleosides from the oligonucleotides following
reaction with hydrated electrons, generated by ionizing radiation . . . . . 452 Loss of the dG-cisPt-dG adduct and formation of undamaged dG and
modified dG by hydrated electrons . . . . . . . . . . . . . . . . . . . . . 463 Percentage of modified dG and dT as a function of irradiation dose for
ODN-GTG and ODN-GTG-cisPt following digestion to nucleosides . . . 474 Structure of dG-cisPt-dG following the digestion . . . . . . . . . . . . . . 485 ESI-MS spectrum of dG-cisPt-dG . . . . . . . . . . . . . . . . . . . . . . 486 ESI-MS/MS spectrum of dG-cisPt-dG at m/z 760 . . . . . . . . . . . . . 49
4 ARTICLE 3 561 Nanosecond decay kinetics of e−aq at 600 nm in hydrolyzed cisplatin, GTG,
and GTG-cisPt solutions . . . . . . . . . . . . . . . . . . . . . . . . . . 612 Pseudo-first-order rate constant kobs vs hydrolyzed cisplatin, GTG, and
GTG-cisPt concentration . . . . . . . . . . . . . . . . . . . . . . . . . . . 623 Kinetics of GTG-cisPt complex in presence of tert-butanol scavenger . . 634 Transient absorption of GTG-cisPt complex in presence of tert-butanol
scavenger . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 645 Proposed mechanism of d(GpTpG) release from a GTG-cisPt complex . 65
5 DISCUSSION 7112 Complexe TTTGTTTTTTT-cisPt et TTTGTGTTTTT-cisPt . . . . . . 7313 Courbes de potentiel de molecule AB a l’etat neutre (AB) et a l’etat
d’anion (AB−) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7414 Plus faible orbitale moleculaire non-occupee de cisplatine . . . . . . . . . 7515 Transfert de l’electron d’une base a une autre vers le cisPt . . . . . . . . 76

LISTE DES TABLEAUX
1 INTRODUCTION 11 Deformations de l’ADN par adduit de cisplatine . . . . . . . . . . . . . . 42 Rendement des CSB et CDB par •OH, H• et e−hyd pour l’ADN sans et avec
l’adduit de cisplatine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 153 Differents types de dommage a l’ADN avec une dose de 1 Gy d’une source
a faible TEL et l’effet cellulaire . . . . . . . . . . . . . . . . . . . . . . . 18
4 ARTICLE 3 561 Rate Constant of the e−aq Reaction with Hydrolyzed Cisplatin, GTG, and
GTG-cisPt . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
v

LISTE DES ABREVIATIONS
Acide desoxyribonucleique ADNAcide ethylene-diamine-tetraacetique EDTAAdenine ACassure simple brin CSBCassure double brins CDBChromatographie en phase liquide a haute performance HPLCCytosine CDiamminedichloroplatine (II) cisplatine
Electron hydrate e−hydElectrons secondaires ES
Electron-volt eV
Energie du seuil thermodynamique ΔH
Energie de dissociation DAdduit de cisplatine cisPtAffinite electronique AEElectron Spin Resonance ESRFacteur d’amplification de l’oxygene FAOGray GyGuanine GHighest Occupied Molecular Orbital HOMOLowest Unoccupied Molecular Orbital LUMOOligonucleotide TTTTTGTGTTT ODN-GTGOligonucleotide TTTTTTTGTTT ODN-TGTRadical hydroxyle •OHReparation par excision des nucleotides NERThymine TTransfert dissociatif de l’electron TDETransfert d’energie lineaire TEL
vi

INTRODUCTION
1.1 L’effet concomitant de la radiochimiotherapie
La radiochimiotherapie est une methode tres efficace pour traiter de facon locoregio-
nale les tumeurs et peut etre combinee avec la chirurgie (Seiwert et al., 2007). Ce type
de traitement concomitant a considerablement ameliore le controle des tumeurs ainsi
que la survie des patients (Samant et al., 1999; Boscolo-Rizzo et al., 2011; Peters et al.,
2000; Candelaria et al., 2006; Seiwert et al., 2007). Il y a deux avantages theoriques a la
radiochimiotherapie (voir figure 1), soient : (1) l’additivite des effets antitumoraux, qui
suppose que chaque element du traitement possede son propre effet antitumoral auquel
s’ajoute celui des autres ; (2) l’interaction synergique, qui considere qu’il existe une inter-
action superadditive lorsque l’effet du traitement combine apparaıt superieur a la somme
de l’effet des traitements individuels (Seiwert et al., 2007).
Plusieurs mecanismes sont impliques dans les proprietes de radiosensibilisation des
agents chimiotherapeutiques, tels que l’inhibition de la reparation de l’ADN, l’augmenta-
tion des dommages a l’ADN, la diminution du seuil de l’apoptose ainsi que la perturbation
du cycle cellulaire qui maintient la cellule dans les phases sensibles (G2/M) plus longtemps
(Choy, 2003; Hei and Hall, 1994).
1.2 Adduits de cisplatine-ADN
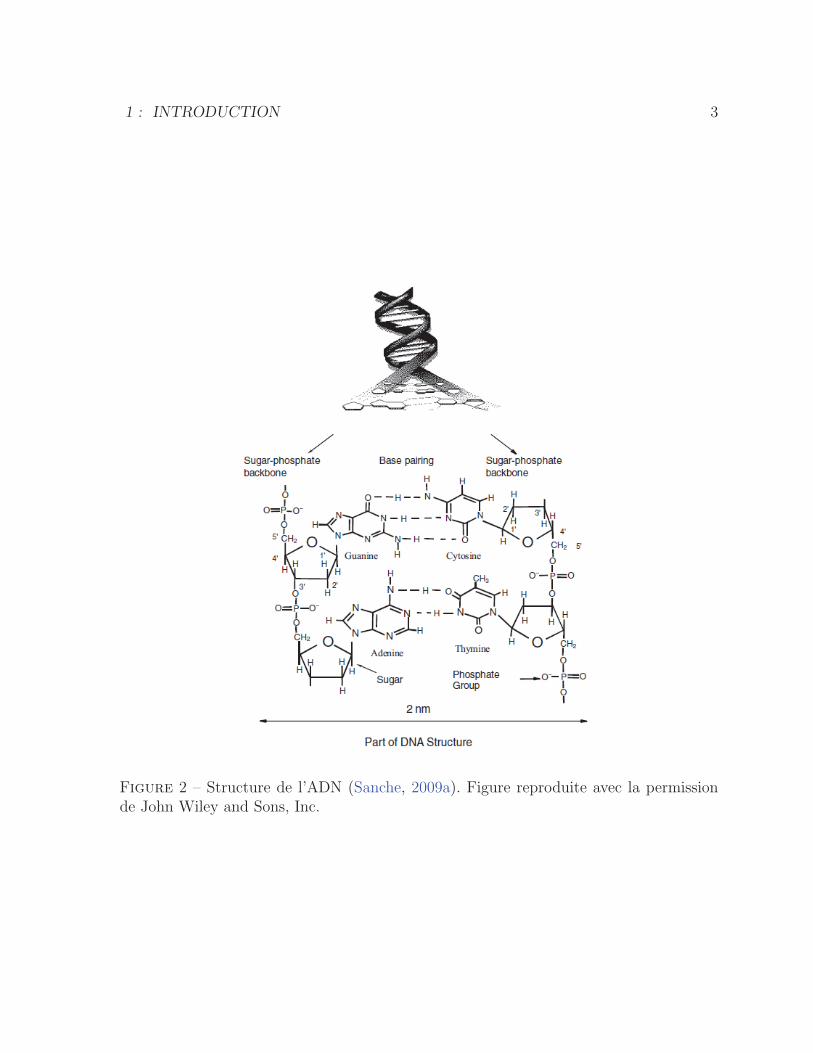
L’ADN est un biopolymere constitue de deux brins, formes d’unites de sucre et de
phosphate, qui sont lies l’un a l’autre par des paires de bases specifiques, soit la thymine
(T) avec l’adenine (A) et la cytosine (C) avec la guanine (G). La liaison entre paires de
bases est assuree par des ponts hydrogene, tel que represente a la figure 2.
Le diamminedichloroplatine (II), communement appele cisplatine, est un agent chi-
1

1 : INTRODUCTION 2
Figure 1 – Avantages theoriques de la radiochimiotherapie
miotherapeutique grandement utilise dans les traitements de differents types de cancer,
soit le cancer des ovaires, des testicules, des poumons, de la tete et du cou. L’activite
antitumorale potentielle du cisplatine a ete decouverte par Rosenberg et al. en 1965 (Ro-
senberg et al., 1965; Rosenberg, 1985). Au cours d’experiences portant sur l’effet d’un
champ electrique sur la croissance des cellules d’Escherichia coli, ce dernier a remarque
que la division cellulaire etait bloquee par la reaction du platine des electrodes avec du
chlorure d’ammonium present dans le tampon.
Le cisplatine entre dans la cellule soit via des transporteurs actifs, plus particuliere-
ment le transporteur cuivre-CTR1 (Kelland, 2007), soit par diffusion passive (Gately and
Howell, 1993). Une fois a l’interieur de la cellule, la faible concentration de chlorure (20
mM) permet l’hydrolyse du cisplatine, ce qui permet la formation du [Pt(NH3)2Cl(OH2)]+et
du [Pt(NH3)2(OH2)2]2+. Sous cette forme, le cisplatine peut alors reagir avec plusieurs
composants cellulaires : proteines, ADN, ARN, filaments d’actine et phospholipides mem-
branaires (Jung and Lippard, 2007; Reedijk, 2009; Jamieson and Lippard, 1999). De tous
ces composes, l’ADN est la cible ayant le plus d’impact sur le fonctionnement de la cellule.
Le cisplatine se lie preferentiellement aux bases d’ADN telles que l’adenine et la
guanine, qui ont de la facilite a donner des charges electroniques. La liaison de l’agent
chimiotherapeutique avec l’ADN peut alors prendre differentes configurations, soient in-
trabrin GG (60-65%), AG (25-30%) et GNG (5-10%) et interbrin (1-3%) (Wozniak and
Blasiak, 2002; Eastman, 1983). Cela engendre des deformations de l’ADN en le pliant et
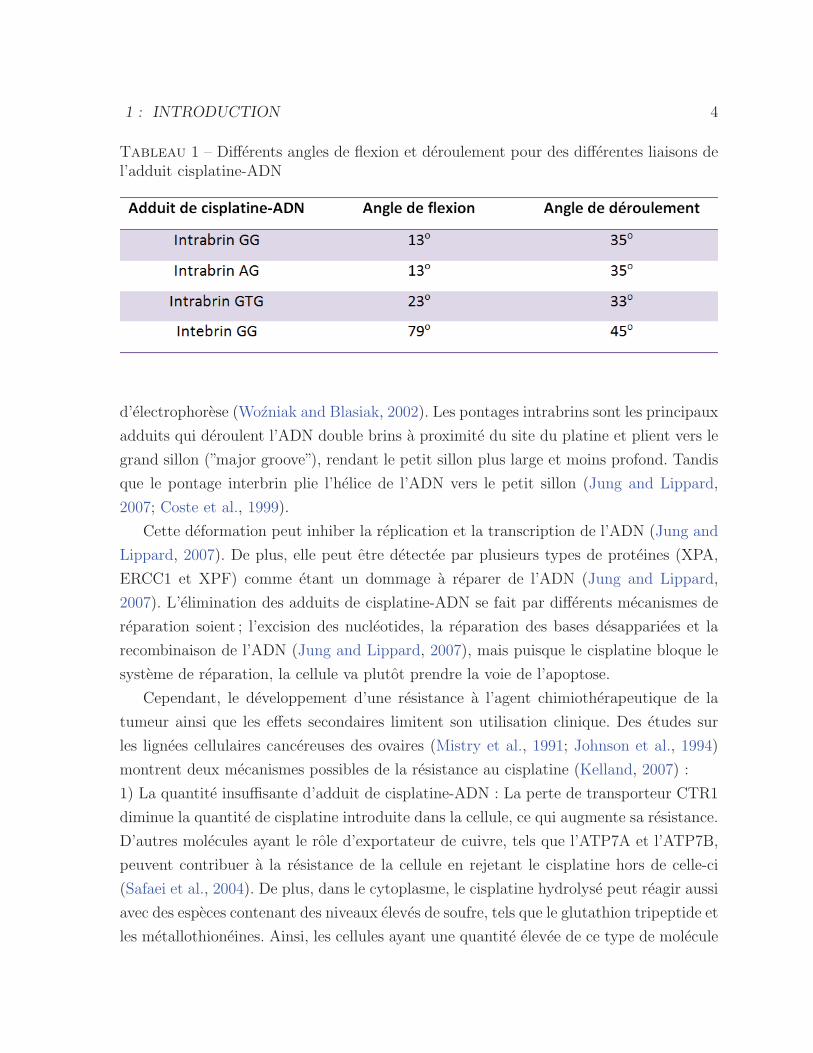
en le deroulant (Wozniak and Blasiak, 2002). Le tableau 1 montre les differents angles de
flexion et de deroulement pour les differentes configurations, mesurees par la technique
1 : INTRODUCTION 3
Figure 2 – Structure de l’ADN (Sanche, 2009a). Figure reproduite avec la permissionde John Wiley and Sons, Inc.
1 : INTRODUCTION 4
Tableau 1 – Differents angles de flexion et deroulement pour des differentes liaisons del’adduit cisplatine-ADN
d’electrophorese (Wozniak and Blasiak, 2002). Les pontages intrabrins sont les principaux
adduits qui deroulent l’ADN double brins a proximite du site du platine et plient vers le
grand sillon (”major groove”), rendant le petit sillon plus large et moins profond. Tandis
que le pontage interbrin plie l’helice de l’ADN vers le petit sillon (Jung and Lippard,
2007; Coste et al., 1999).
Cette deformation peut inhiber la replication et la transcription de l’ADN (Jung and
Lippard, 2007). De plus, elle peut etre detectee par plusieurs types de proteines (XPA,
ERCC1 et XPF) comme etant un dommage a reparer de l’ADN (Jung and Lippard,
2007). L’elimination des adduits de cisplatine-ADN se fait par differents mecanismes de
reparation soient ; l’excision des nucleotides, la reparation des bases desappariees et la
recombinaison de l’ADN (Jung and Lippard, 2007), mais puisque le cisplatine bloque le
systeme de reparation, la cellule va plutot prendre la voie de l’apoptose.
Cependant, le developpement d’une resistance a l’agent chimiotherapeutique de la
tumeur ainsi que les effets secondaires limitent son utilisation clinique. Des etudes sur
les lignees cellulaires cancereuses des ovaires (Mistry et al., 1991; Johnson et al., 1994)
montrent deux mecanismes possibles de la resistance au cisplatine (Kelland, 2007) :
1) La quantite insuffisante d’adduit de cisplatine-ADN : La perte de transporteur CTR1
diminue la quantite de cisplatine introduite dans la cellule, ce qui augmente sa resistance.
D’autres molecules ayant le role d’exportateur de cuivre, tels que l’ATP7A et l’ATP7B,
peuvent contribuer a la resistance de la cellule en rejetant le cisplatine hors de celle-ci
(Safaei et al., 2004). De plus, dans le cytoplasme, le cisplatine hydrolyse peut reagir aussi
avec des especes contenant des niveaux eleves de soufre, tels que le glutathion tripeptide et
les metallothioneines. Ainsi, les cellules ayant une quantite elevee de ce type de molecule

1 : INTRODUCTION 5
sont plus resistantes a l’agent chimiotherapeutique etant donne que celui-ci est intercepte
avant qu’il n’interagisse avec l’ADN. Finalement, une cause importante de la resistance
au cisplatine de la cellule est l’augmentation de sa capacite a se debarrasser de l’agent lie
a l’ADN en surexprimant la proteine ERCC1 impliquee dans la reparation par excision
des nucleotides (NER) (Dabholkar et al., 1992).
2) Augmentation de la tolerance au cisplatine : La resistance peut aussi se produire
par une tolerance accrue a l’adduit cisplatine-ADN, soit par la perte de reparation des
bases desappariees ou par le contournement de l’adduit lors de la replication de l’ADN
(”replicative bypass”) et la diminution de l’apoptose (Kelland, 2007).
1.3 Interaction des rayonnements ionisants avec la
matiere
Les deux processus principaux dans l’interaction de la radiation avec la matiere sont
l’excitation et l’ionisation. Les rayonnements ionisants, soit particulaires ou electroma-
gnetiques, possedent l’energie necessaire pour ioniser des atomes ou des molecules dans
la matiere qu’ils traversent. Des rayonnements sous forme de particules sont utilises en
radiotherapie pour les traitements de cancers, comme des electrons, protons, neutrons,
les ions de carbone ainsi que des rayons α et β. De plus, les rayons γ et les rayons X
sont les deux types de radiation electromagnetique les plus utilises en radiotherapie. En
termes d’energie, ces derniers sont situes vers la fin du spectre electromagnetique (figure
3) et sont suffisamment energetiques pour ejecter un electron d’un atome. L’energie de
liaison typique des orbitales superficielles dans les materiaux biologiques est d’environ
10 eV. Donc, les photons ultraviolets ayant une energie entre 2 et 10 eV sont consideres
non-ionisants et peuvent seulement produire des excitations electroniques, vibrationnelles
et phononiques (Lehnert, 2007).
Les processus d’ionisation impliques dans la radiotherapie electromagnetique sont
principalement l’effet photoelectrique, la creation de paires et l’effet Compton. Ces pro-
cessus creent des electrons primaires, qui a leur tour produisent une grande quantite
d’electrons secondaires, dont la plupart se trouvent a basse energie (≤20 eV) (LaVerne
and Pimblott, 1995; Cobut et al., 1998). Dans la gamme d’energie la plus utilisee en
radiotherapie (100 keV-25 MeV), l’effet Compton est dominant. L’avantage de ce dernier
processus provient du coefficient d’absorption du photon, qui est independant du numero
1 : INTRODUCTION 6
Figure 3 – Spectre electromagnetique
atomique (Z) ; ainsi, la quantite d’energie deposee dans la matiere est la meme que ce soit
dans les tissus mous, dans les muscles ou les os, contrairement a l’effet photoelectrique
(Lehnert, 2007).
1.3.1 Effet direct de la radiation sur l’ADN
En radiotherapie, les dommages crees par la radiation a l’ADN, sont divises en deux
categories, soient les effets directs et indirects. Dans l’effet direct, les particules incidentes
interagissent directement avec l’ADN pour produire des radicaux reactifs, des ions et une
grande quantite d’electrons secondaires (ES). L’effet direct de la radiation ne peut pas etre
reduit par des agents capteurs, puisque les dommages sont produits par le passage direct
de la radiation dans la matiere. Ce processus est dominant pour les rayonnements avec
un transfert d’energie lineaire (TEL) eleve, comme pour les particules α et les neutrons
(Lehnert, 2007).
L’interaction des radiations a haute energie avec la matiere produit un grand nombre
d’ES, soit ∼ 4× 104 electrons par MeV d’energie deposee (Arumainayagam et al., 2010).
La vaste majorite des ES ont une energie en-dessous de 30 eV et peuvent a leur tour pro-
duire une grande quantite de radicaux et d’anions (Sanche, 2009b). Bien que la grande
majorite de ces electrons de basse energie n’ait pas l’energie necessaire pour ioniser les
molecules, ces derniers peuvent tout de meme provoquer des dissociations de molecules
via l’attachement dissociatif, qui est un processus resonnant. Dans ce cas, la resonance se
produit habituellement a de faibles energies (<10 eV) et est caracterisee par la capture
d’un electron incident par une orbitale antiliante non-occupee de la molecule pour for-

1 : INTRODUCTION 7
mer un ion negatif transitoire. Si l’etat negatif transitoire est repulsif et sa demi-vie assez
longue (i.e., de l’ordre ou plus grande qu’une periode de vibration de l’anion), la molecule
se dissocie. La dissociation de l’anion transitoire forme un radical et un anion habituel-
lement stable (Arumainayagam et al., 2010). Par exemple, dans le cas d’une molecule
RH :
e− + RH −→ RH∗− (1.1)
RH∗− −→ R• +H− (1.2)
Bien que les phenomenes d’ionisation et d’excitation soient plus probables a haute
energie (>6 eV) que le phenomene de l’attachement dissociatif, la contribution de ce
dernier dans l’effet direct de la radiation est la plus elevee etant donnee l’augmentation
considerable de l’intensite de la distribution energetique des ES a basse energie (Arumai-
nayagam et al., 2010).
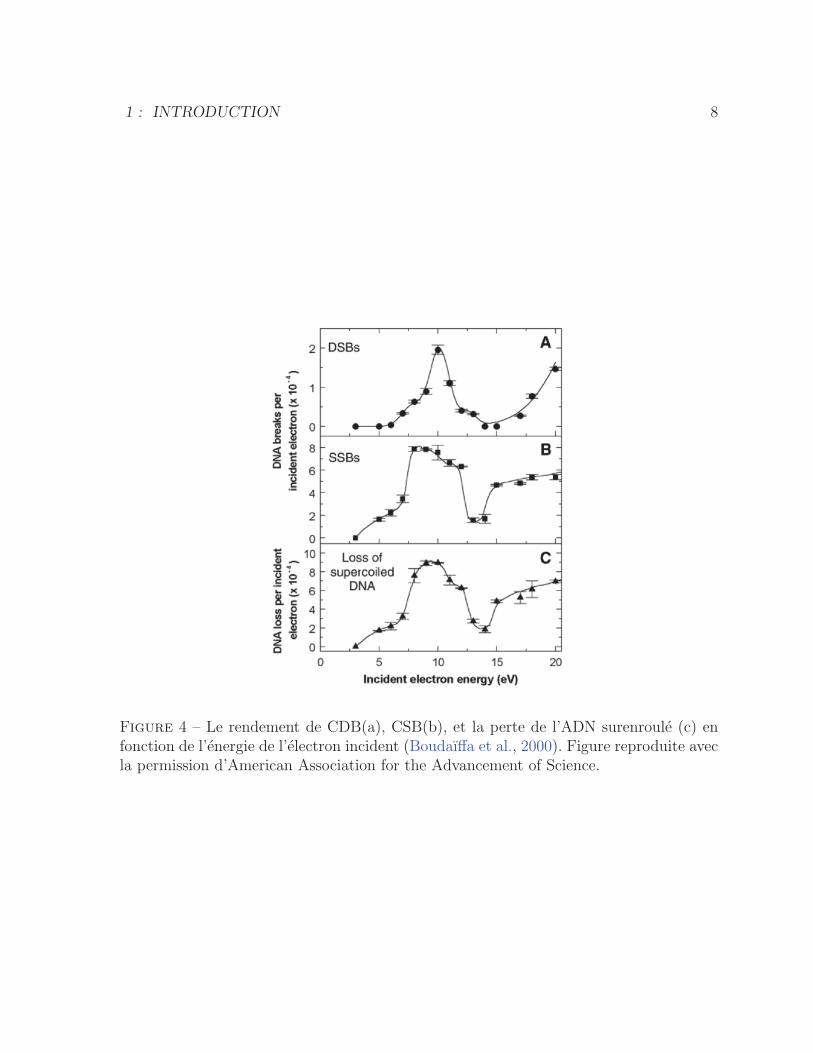
L’effet direct de la radiation a ete etudie par Boudaiffa et al. en bombardant de
l’ADN surenroule en condition seche avec des electrons de basse energie (3-20 eV). Ces
derniers ont mesure la formation de l’ADN circulaire (cassure simple brin (CSB)) et
de l’ADN lineaire (cassure double brins (CDB)). La figure 4 montre une fonction de
rendements tres structuree avec un maximum de CSB et CDB a 10 eV. Ceci indique que
les cassures de brin de l’ADN resultent essentiellement de l’attachement dissociatif avec
une superposition mineure de dissociation par excitation directe (Boudaıffa et al., 2000).
D’autres etudes montrent que les electrons en-dessous du seuil d’excitation electronique
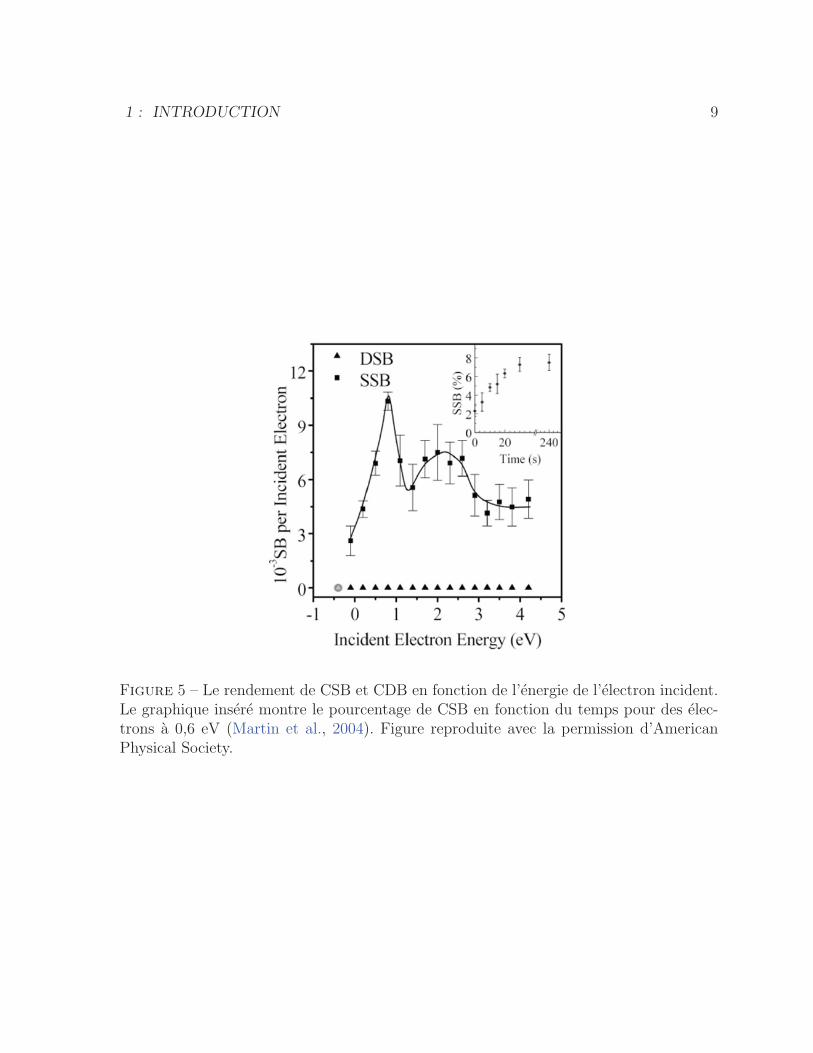
peuvent aussi induire des CSB a l’ADN avec des maxima de rendement a 0,8 et 2,2 eV
(figure 5) (Martin et al., 2004). Par contre, dans la plage d’energie 0-4 eV aucune CDB
n’a ete mesuree (figure 5).
Ray et al. (Ray et al., 2005) ont etudie la dependance de capture de l’electron (<2 eV)
aux sequences de bases de l’ADN. Ils ont montre que la probabilite que ce phenomene
se produise augmente avec le nombre de guanine. La cause de cet effet n’est pas encore
comprise, etant donne que l’affinite electronique de la guanine est comparable a celle des
autres bases. Par contre, la guanine presente un dipole electrique qui est 3 fois superieur
a celui de l’adenine, ce qui peut favoriser la capture electronique.
Des experiences en phase gazeuse montrent que tous les constituants de l’ADN, tels
que le sucre, les groupements phosphates et les nucleobases, subissent l’attachement dis-
1 : INTRODUCTION 8
Figure 4 – Le rendement de CDB(a), CSB(b), et la perte de l’ADN surenroule (c) enfonction de l’energie de l’electron incident (Boudaıffa et al., 2000). Figure reproduite avecla permission d’American Association for the Advancement of Science.
1 : INTRODUCTION 9
Figure 5 – Le rendement de CSB et CDB en fonction de l’energie de l’electron incident.Le graphique insere montre le pourcentage de CSB en fonction du temps pour des elec-trons a 0,6 eV (Martin et al., 2004). Figure reproduite avec la permission d’AmericanPhysical Society.
1 : INTRODUCTION 10
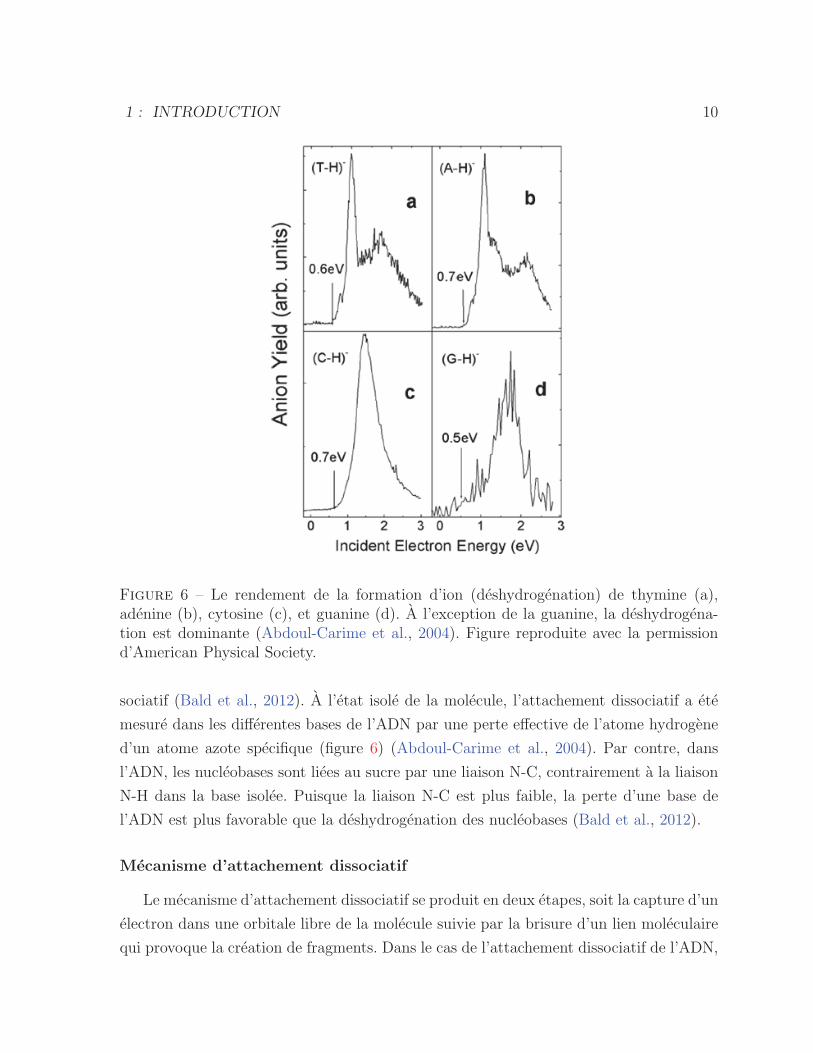
Figure 6 – Le rendement de la formation d’ion (deshydrogenation) de thymine (a),adenine (b), cytosine (c), et guanine (d). A l’exception de la guanine, la deshydrogena-tion est dominante (Abdoul-Carime et al., 2004). Figure reproduite avec la permissiond’American Physical Society.
sociatif (Bald et al., 2012). A l’etat isole de la molecule, l’attachement dissociatif a ete
mesure dans les differentes bases de l’ADN par une perte effective de l’atome hydrogene
d’un atome azote specifique (figure 6) (Abdoul-Carime et al., 2004). Par contre, dans
l’ADN, les nucleobases sont liees au sucre par une liaison N-C, contrairement a la liaison
N-H dans la base isolee. Puisque la liaison N-C est plus faible, la perte d’une base de
l’ADN est plus favorable que la deshydrogenation des nucleobases (Bald et al., 2012).
Mecanisme d’attachement dissociatif
Le mecanisme d’attachement dissociatif se produit en deux etapes, soit la capture d’un
electron dans une orbitale libre de la molecule suivie par la brisure d’un lien moleculaire
qui provoque la creation de fragments. Dans le cas de l’attachement dissociatif de l’ADN,

1 : INTRODUCTION 11
ces deux etapes sont encore aujourd’hui mal comprises. Des experiences de Zheng et al.
(Zheng et al., 2005) avec des oligonucleotides (GCTA et CGTA) ont montre que le clivage
des liaisons phosphodiester (C-O) est preferentiel a celui des liaisons P-O de la chaıne.
Pour ce qui est des liaisons P=O, l’occupation de l’orbitale libre de plus faible energie
necessite une energie plus grande que 2 eV. Il est donc naturel qu’un electron de 0,1-2,0
eV ne puisse cliver cette liaison a moins de faire intervenir l’affinite electronique.
Il a ete predit theoriquement que des electrons de basse energie sont captures par des
bases de l’ADN pour ensuite etre transferes vers des orbitales anti-liantes de groupements
sucre-phosphate, ce qui provoque la rupture de la liaison C-O (Berdys et al., 2004; Simons,
2006; Bald et al., 2012). A l’etat neutre de la molecule, deux electrons occupent l’orbitale
σ de la liaison C-O alors que l’orbitale π∗ est inoccupee. En principe, l’etape de la capture
electronique par l’ADN devrait se produire via cette derniere orbitale et necessite une
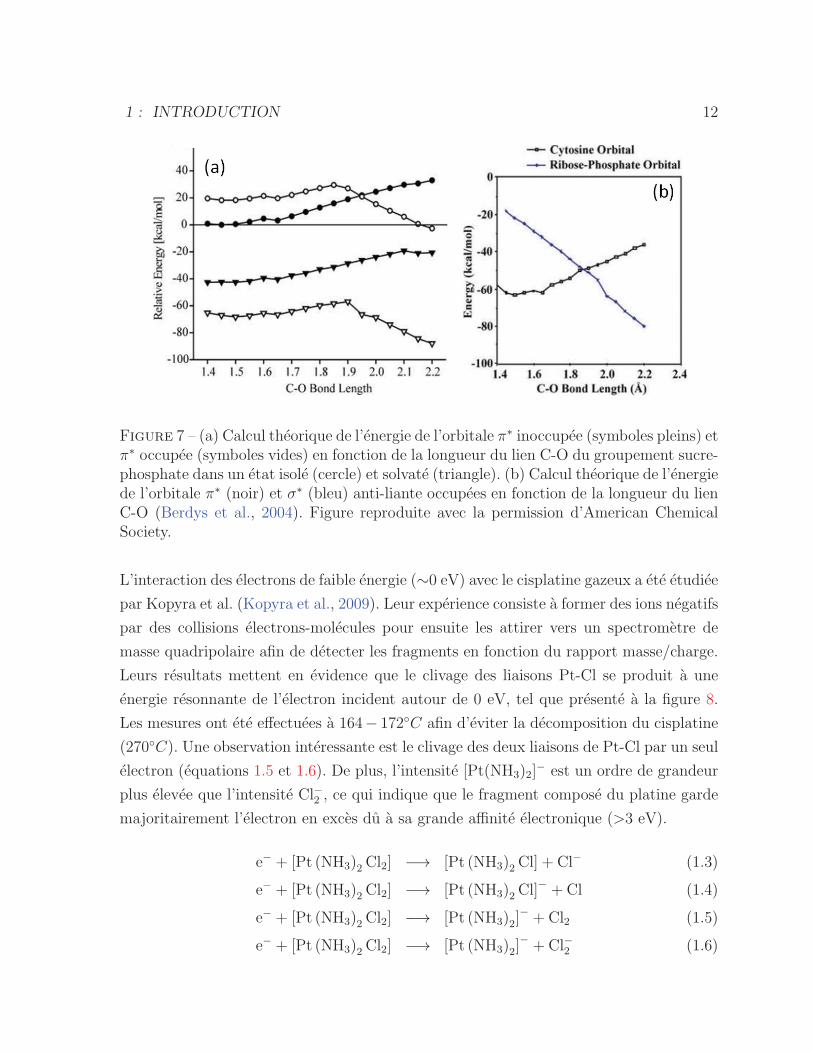
energie minimale de 1 eV pour l’occuper. Des calculs effectues par Berdys et al. (Berdys
et al., 2004) montrent (voir graphique 7 (a)) que la difference entre l’energie de l’etat π∗
inoccupe (•, �) et occupe par un electron en exces (◦, �) change en fonction de la longueur
de la liaison C-O. Les courbes avec des cercles et des triangles presentent respectivement
l’energie de l’orbitale π∗ pour l’ADN isole et dans un environnement solvate. A l’etat
isole de la molecule, il existe une longueur critique de la liaison C-O au-dessus de laquelle
l’energie de l’orbitale π∗ occupee est energetiquement favorable, rendant ainsi la capture
electronique hautement probable, meme a 0 eV. A l’etat solvate, cet etat est toujours
favorable, peu importe la distance interatomique.
L’etape importante pour que le phenomene de dissociation d’un brin se produise est
le transfert de l’electron en exces de l’orbitale π∗ de la base a l’orbitale σ∗ anti-liante
du groupement phosphate. Le graphique 7 (b), issu des calculs de Berdys et al. (Berdys
et al., 2004), montre que cela peut s’effectuer de facon adiabatique lorsque la longueur
de la liaison C-O est 1.9 A, point ou l’energie des deux orbitales sont egales. Au-dessus
de cette distance interatomique, l’orbitale σ∗ anti-liante est meme favorisee. Dans le cas
ou cette orbitale est occupee assez longtemps (10−14 s), le phenomene de dissociation va
se produire. Cela est aussi valable pour les liaisons C-O, N-H ou N1-H.
Attachement dissociatif du cisplatine
Le phenomene d’attachement dissociatif de l’ADN peut etre amplifie par des agents
chimiotherapeutiques tels que le cisplatine. Bien que les mecanismes soient mal compris,
cela pourrait expliquer l’effet synergique de la chimiotherapie et de la radiotherapie.
1 : INTRODUCTION 12
Figure 7 – (a) Calcul theorique de l’energie de l’orbitale π∗ inoccupee (symboles pleins) etπ∗ occupee (symboles vides) en fonction de la longueur du lien C-O du groupement sucre-phosphate dans un etat isole (cercle) et solvate (triangle). (b) Calcul theorique de l’energiede l’orbitale π∗ (noir) et σ∗ (bleu) anti-liante occupees en fonction de la longueur du lienC-O (Berdys et al., 2004). Figure reproduite avec la permission d’American ChemicalSociety.
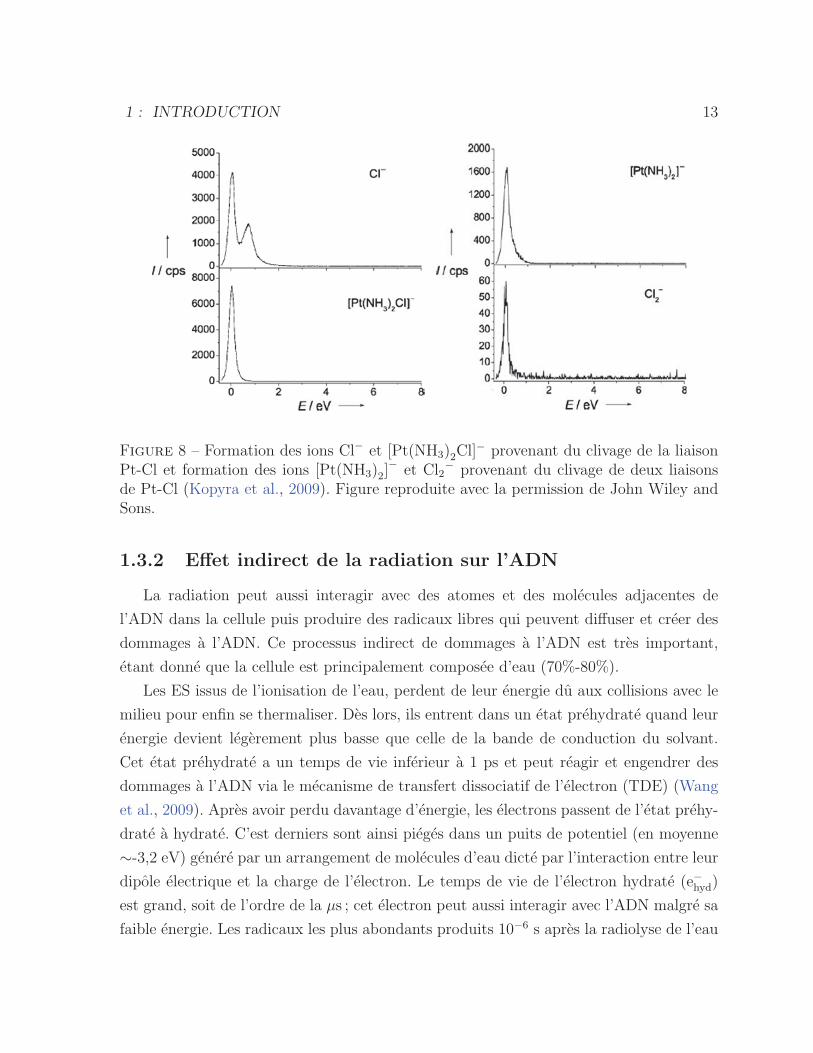
L’interaction des electrons de faible energie (∼0 eV) avec le cisplatine gazeux a ete etudiee
par Kopyra et al. (Kopyra et al., 2009). Leur experience consiste a former des ions negatifs
par des collisions electrons-molecules pour ensuite les attirer vers un spectrometre de
masse quadripolaire afin de detecter les fragments en fonction du rapport masse/charge.
Leurs resultats mettent en evidence que le clivage des liaisons Pt-Cl se produit a une
energie resonnante de l’electron incident autour de 0 eV, tel que presente a la figure 8.
Les mesures ont ete effectuees a 164− 172◦C afin d’eviter la decomposition du cisplatine
(270◦C). Une observation interessante est le clivage des deux liaisons de Pt-Cl par un seul
electron (equations 1.5 et 1.6). De plus, l’intensite [Pt(NH3)2]− est un ordre de grandeur
plus elevee que l’intensite Cl−2 , ce qui indique que le fragment compose du platine garde
majoritairement l’electron en exces du a sa grande affinite electronique (>3 eV).
e− + [Pt (NH3)2 Cl2] −→ [Pt (NH3)2 Cl] + Cl− (1.3)
e− + [Pt (NH3)2 Cl2] −→ [Pt (NH3)2 Cl]− + Cl (1.4)
e− + [Pt (NH3)2 Cl2] −→ [Pt (NH3)2]− + Cl2 (1.5)
e− + [Pt (NH3)2 Cl2] −→ [Pt (NH3)2]− + Cl−2 (1.6)
1 : INTRODUCTION 13
Figure 8 – Formation des ions Cl− et [Pt(NH3)2Cl]− provenant du clivage de la liaison
Pt-Cl et formation des ions [Pt(NH3)2]− et Cl2
− provenant du clivage de deux liaisonsde Pt-Cl (Kopyra et al., 2009). Figure reproduite avec la permission de John Wiley andSons.
1.3.2 Effet indirect de la radiation sur l’ADN
La radiation peut aussi interagir avec des atomes et des molecules adjacentes de
l’ADN dans la cellule puis produire des radicaux libres qui peuvent diffuser et creer des
dommages a l’ADN. Ce processus indirect de dommages a l’ADN est tres important,
etant donne que la cellule est principalement composee d’eau (70%-80%).
Les ES issus de l’ionisation de l’eau, perdent de leur energie du aux collisions avec le
milieu pour enfin se thermaliser. Des lors, ils entrent dans un etat prehydrate quand leur
energie devient legerement plus basse que celle de la bande de conduction du solvant.
Cet etat prehydrate a un temps de vie inferieur a 1 ps et peut reagir et engendrer des
dommages a l’ADN via le mecanisme de transfert dissociatif de l’electron (TDE) (Wang
et al., 2009). Apres avoir perdu davantage d’energie, les electrons passent de l’etat prehy-
drate a hydrate. C’est derniers sont ainsi pieges dans un puits de potentiel (en moyenne
∼-3,2 eV) genere par un arrangement de molecules d’eau dicte par l’interaction entre leur
dipole electrique et la charge de l’electron. Le temps de vie de l’electron hydrate (e−hyd)
est grand, soit de l’ordre de la μs ; cet electron peut aussi interagir avec l’ADN malgre sa
faible energie. Les radicaux les plus abondants produits 10−6 s apres la radiolyse de l’eau

1 : INTRODUCTION 14
sont les hydroxyles (•OH), les electrons hydrates (e−hyd) et les hydrogenes (H•) avec une
valeur G respective de 0,24, 0,28 et 0,06 μmol/J 10−6 s suite a l’irradiation de l’eau.
Radiolyse de l’eau
H2O −→ H2O•+ + e− et H2O
∗ (1.7)
H2O•+ +H2O −→ •OH+H3O
+ (1.8)
H2O∗ −→ •OH+H•,H2 +O• (1.9)
e− +H2O −→ e−hyd (1.10)
O2 et N2O sont des capteurs de l’e−hyd :
e−hyd +O2 −→ O•−2 (1.11)
e−hyd +N2O −→ N2 +• OH+OH− (1.12)
Parmi les radicaux generes par la radiolyse de l’eau, le •OH, un agent oxydant, est celui
qui engendre le plus de dommages a l’ADN. Pour se defendre contre ce type de radicaux,
la cellule contient des composes thioles et des antioxydants qui reagissent comme des
capteurs des radicaux (Lehnert, 2007). Malgre cela, la contribution des radicaux dans les
processus de dommages a l’ADN demeure importante en radiotherapie. Il a ete suggere
qu’entre 30% a 70% des dommages sont dus a la radiolyse de l’eau (DeLara et al., 1995;
Nikjoo et al., 2002). Le radical •OH est tres electrophile et son interaction se produit soit
par l’abstraction de l’electron, par l’ajout d’un OH a une double liaison C-C et C-N ou
bien par l’abstraction d’un H (von Sonntag, 2006). L’action de •OH sur l’ADN produit
des modifications aux bases ainsi que des cassures de brin en interagissant principalement
avec les doubles liaisons des bases et aussi via l’abstraction de H de C4 du sucre. L’e−hydet H•, etant tous deux des reducteurs, generent les memes types de dommages a l’ADN
rendant difficile a distinguer leur effet. Par contre, le potentiel de reduction de H• est plus
petit que e−hyd et dans certains cas, la reaction de reduction d’ion de metal se fait seulement
par e−hyd et non pas par H• (von Sonntag, 2006). De plus, e−hyd reagit avec de nombreux
composes qui sont capables de liberer un anion via le phenomene de dissociation par
capture electronique, ce qui permet la distinction entre e−hyd et H• (Hayon and Allen,
1961; Jortner and Rabani, 1961; von Sonntag, 2006).

1 : INTRODUCTION 15
Tableau 2 – Le rendement des CSB et CDB par •OH, H• et e−hyd pour l’ADN sans etavec l’adduit de cisplatine avec une faible concentration de capteur •OH (5 mM tris) etdans une atmosphere de N2 (Rezaee et al., 2013). Figure reproduite avec la permissionde Radiation Research Society.
En principe, e−hyd ne peut pas produire de cassures de brin de l’ADN (von Sonn-
tag, 1987), puisque ces reactions sont exothermiques et une energie d’activation est donc
necessaire (von Sonntag, 2006). Par contre, les etudes recentes (Rezaee et al., 2013)
montrent que les modifications structurelles et chimiques de l’ADN, en presence de cis-
platine, peuvent diminuer la barriere d’energie de dissociation et favoriser la formation
de cassures de brin de l’ADN. L’attachement de l’electron a une molecule cible avec son
transfert a une liaison specifique et le clivage de cette liaison, est le mecanisme qui ex-
plique la reaction des e−hyd avec l’ADN. En raison de la presence des molecules polaires
de NH3, combinee a la grande affinite electronique du cisplatine et de la modification
structurelle qu’il produit sur l’ADN, la probabilite de capter l’electron (≤ 0 eV) est aug-
mentee. Le tableau 2 montre le rendement des CSB et CDB par differents radicaux issus
de la radiolyse de l’eau, pour le plasmide avec et sans adduit du cisplatine. La presence
de cisplatine augmente les CSB et CDB respectivement d’un facteur 1,5 et 1,2 par le
radical •OH. Cette augmentation en presence de l’adduit de cisplatine est expliquee par
le deroulement de l’ADN, qui rend plus accessible les desoxyriboses aux radicaux.
1.3.3 Effet radiosensibilisateur de l’oxygene
Les radiations ionisantes produisent aussi des radicaux de l’ADN (ADN•). En absence
de l’oxygene, ce type de dommage peut etre chimiquement reparable. Par exemple, par
un ajout d’hydrogene par les molecules qui sont donneur d’hydrogene (Farhataziz and
Rodgers, 1987). La presence de O2 produit le phenomene appele ”fixation de l’oxygene”
qui modifie l’ADN• en generant un radical peroxy (DNA-OO•) (Bertout et al., 2008;
Ferradini and Jay-Gerin, 1999), qui bloque la reparation chimique. En effet, les cellules
1 : INTRODUCTION 16
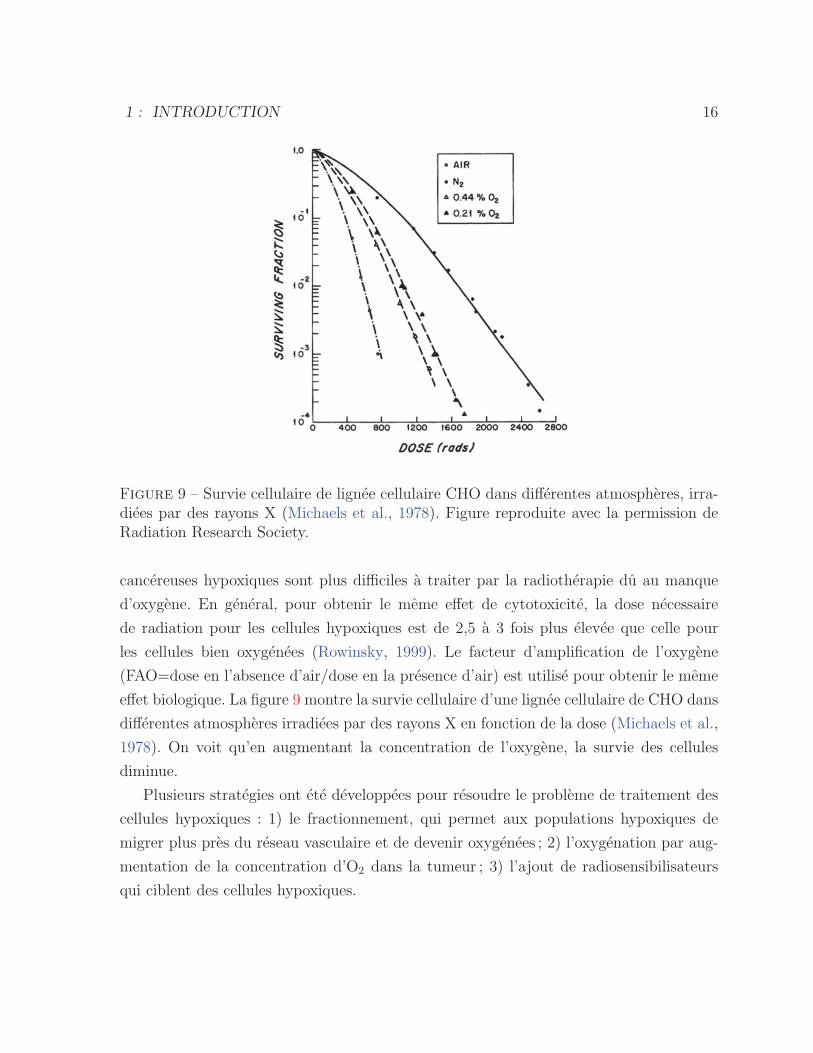
Figure 9 – Survie cellulaire de lignee cellulaire CHO dans differentes atmospheres, irra-diees par des rayons X (Michaels et al., 1978). Figure reproduite avec la permission deRadiation Research Society.
cancereuses hypoxiques sont plus difficiles a traiter par la radiotherapie du au manque
d’oxygene. En general, pour obtenir le meme effet de cytotoxicite, la dose necessaire
de radiation pour les cellules hypoxiques est de 2,5 a 3 fois plus elevee que celle pour
les cellules bien oxygenees (Rowinsky, 1999). Le facteur d’amplification de l’oxygene
(FAO=dose en l’absence d’air/dose en la presence d’air) est utilise pour obtenir le meme
effet biologique. La figure 9 montre la survie cellulaire d’une lignee cellulaire de CHO dans
differentes atmospheres irradiees par des rayons X en fonction de la dose (Michaels et al.,
1978). On voit qu’en augmentant la concentration de l’oxygene, la survie des cellules
diminue.
Plusieurs strategies ont ete developpees pour resoudre le probleme de traitement des
cellules hypoxiques : 1) le fractionnement, qui permet aux populations hypoxiques de
migrer plus pres du reseau vasculaire et de devenir oxygenees ; 2) l’oxygenation par aug-
mentation de la concentration d’O2 dans la tumeur ; 3) l’ajout de radiosensibilisateurs
qui ciblent des cellules hypoxiques.

1 : INTRODUCTION 17
1.4 L’echelle de temps des processus chimiques du
rayonnement
La sequence des reactions qui se produisent tout au long de la trajectoire de la parti-
cule ionisante dans l’environnement cellulaire peut etre divisee en trois etapes (Bald et al.,
2012). La premiere consiste en la ”phase physique”, soit des reactions dans l’intervalle de
temps de 10−15 a 10−12 s apres l’interaction primaire, ce qui inclut des ionisations et ex-
citations electroniques, la formation d’un nombre abondant d’ES et des radicaux comme•OH. La seconde est appelee ”phase chimique”, ou ces reactions se produisent typique-
ment dans l’intervalle de temps de 10−12 a 10−6 s et comprend la relaxation moleculaire, la
reorganisation des molecules et le clivage de liaisons. Finalement, ”la phase biologique”,
ou des modifications se produisent sur une echelle plus longue de plusieurs heures ou
plusieurs annees et inclut une reponse globale du systeme.
1.5 Les dommages induits a l’ADN
La radiation peut produire differents types de modifications a l’ADN, soient les dom-
mages aux bases, des sites abasiques (site AP), des CSB et CDB ainsi que des dommages
multiples (”cluster”) (figure 10). Entre ces dommages, les CDB et les dommages multiples
sont les plus letaux pour la cellule (Ward, 1988). Le tableau 3 montre une comparaison
du nombre de differents types de dommage a l’ADN pour une dose de 1 Gy provenant
d’une source a faible TEL avec leurs effets sur les cellules. Entre autres, le transfert de
charge est un des aspects importants impliques dans la formation des dommages a des
sites particuliers.
1.5.1 Transfert de charge
Le transfert de charge a travers l’ADN est un des phenomenes qui provoque des dom-
mages a l’ADN dans la cellule (Rajski et al., 2000). Les mesures de dommages oxydatifs
a l’ADN double brins a l’aide d’intercalants photo-oxydant montrent qu’une charge peut
etre transferee sur de longues distances, soit de 200 A (Nunez et al., 1999). Une grande
variete d’approches experimentales ont ete utilisees pour mesurer le transport de charge
et ont mene a des conclusions differentes. Certaines etudes montrent que l’ADN a une
conductivite electrique (Okahata, 1967) tandis que d’autres le considerent plus comme

1 : INTRODUCTION 18
Tableau 3 – Differents types de dommage a l’ADN avec une dose de 1 Gy d’une source afaible TEL et l’effet cellulaire (von Sonntag, 2006). Figure reproduite avec la permissionde Springer.

1 : INTRODUCTION 19
Figure 10 – Differents types de dommage a l’ADN par radiation ionisante. (a) Dommagea la base (b) site abasique (c) CSB (d) CDB(e) lesion tandem (f) dommage multiple avecdeux bases endommagees dans deux brins de l’ADN (g) CSB avec dommage a une baseadjacente (h) dommage multiple avec trois bases endommagees (i) dommage multipleavec deux dommages aux bases et CDB (von Sonntag, 2006). Figure reproduite avec lapermission de Springer.
un semi-conducteur, mais avec differentes grandeurs de gap electronique (Fink and Scho-
nenberger, 1999; Porath et al., 2000). Ces variations sont probablement dues a l’integrite
de l’ADN en absence d’eau, a des tensions elevees et aux electrodes utilisees (Boon and
Barton, 2002). Plusieurs etudes ont ete faites sur des oligonucleotides en solution conte-
nant des donneurs et des accepteurs ou le transfert d’electron photo-induit a ete mesure
en fonction de la distance (Murphy et al., 1993; Lewis et al., 1997; Kelley et al., 1997).
Ces resultats sont controverses du au couplage electronique du donneur et de l’accepteur
dans l’ADN (Boon and Barton, 2002). En general, le transfert de charge depend de la
difference d’energie et de la distance entre le donneur et l’accepteur. Quand la distance
entre l’accepteur et le donneur est relativement petite, le transfert de charge se produit
principalement par le mecanisme d’effet tunnel. Ce dernier implique que la vitesse de
transfert de charge diminue exponentiellement avec la distance entre l’accepteur et le
donneur.
Pour de plus grandes distances entre l’accepteur et le donneur, soit au-dela de six
paires de bases, le mecanisme de polaron assisted hopping devient dominant. A ce stade,
le transfert de charge est independant de la distance entre le donneur et l’accepteur (Pa-
thirannehelage, 2011). L’agitation thermique due aux fluctuations structurelles de l’ADN

1 : INTRODUCTION 20
autour d’un polaron est responsable du transfert de charge. Ce phenomene, se produi-
sant entre les bases adjacentes, depend du recouvrement effectif des orbitales moleculaires
(Pathirannehelage, 2011).
Il existe plusieurs etudes theoriques et experimentales qui suggerent que l’oxydation
de l’ADN peut induire une deficience en electrons, donnant lieu a un trou qui se deplace a
travers l’ADN (Ly et al., 1999; Schuster, 2000; Barnett et al., 2001). Par contre, le cas du
transfert d’un electron en exces n’a pas ete beaucoup etudie. Le groupe Sevilla a montre
que le mecanisme en jeu dans le transfert d’electron en exces depend de la concentration
de l’ADN, de sa sequence de bases et de la temperature (Razskazovskii and Swarts, 1997;
Cai and Sevilla, 2000; Cai et al., 2002). Dans cette etude, l’ADN double brins intercale par
mitoxantone a ete utilise et etudie en fonction de la temperature et de la concentration
par la technique Electron Spin Resonance (ESR). Ils ont conclu que le mecanisme d’effet
tunnel est dominant a 77 K et le mecanisme de thermally activated hopping est dominant
a >150 K. De plus, le transfert d’electron est plus grand a plus haute concentration
d’ADN du au transfert de l’electron d’une molecule d’ADN a une autre (Cai and Sevilla,
2004).
Une preference directionnelle dans le transfert de charge a aussi ete observee. Ito et al.
ont montre que le transfert d’electron en exces dans le brin d’ADN complementaire a celui
qui contient le donneur et l’accepteur change d’un facteur 9 si l’orientation du donneur
et de l’accepteur est inversee. Ils ont utilise une photo-excitation selective d’un donneur
(N, N, N’, N’-tetramethyl-1,5-diaminoaphthalene : D), qui initie le transfert d’electron
et produit une reduction subsequente de l’accepteur (5-bromo-2′-desoxyuridine : BrU),
ce qui genere une decomposition de l’ADN (figure 11). Cette decomposition est ensuite
detectee par un gel d’electrophorese et avec un traitement de piperidine qui fragmente
l’ADN la ou il y a modification. Ces resultats montrent que le transfert d’electron de
3’ a 5’ est la direction la plus favorable. D’autres etudes (O’Neill and Barton, 2002)
montrent que la migration de trou est plus favorable dans la direction opposee (de 5’ a
3’). L’asymetrie de recouvrement des Highest Occupied Molecular Orbital (HOMO) des
nucleobases, qui agissent comme les porteurs de charge, a ete proposee pour expliquer
la dependance du transfert de trou en fonction de la direction dans l’ADN (O’Neill and
Barton, 2002). Une proposition equivalente, basee sur le recouvrement Lowest Unoccupied
Molecular Orbital (LUMO) pour le transfert d’electron en exces, a aussi ete suggeree par
Ito et al. (Ito and Rokita, 2004).

1 : INTRODUCTION 21
Figure 11 – Transfert d’electron en exces de N, N, N’, N’-tetramethyl-1,5-diaminoaphthalene (D) a 5-bromo-2’-desoxyuridine (BrU) dans l’ADN. (Ito and Rokita,2004). Figure reproduite avec la permission de John Wiley and Sons.
1.6 Objectifs du projet de recherche
Puisque la grande partie des cellules est composee d’eau, l’effet indirect de la radiation
est un aspect important pour l’etude des dommages a l’ADN. Entre tous les radicaux
produits par la radiolyse de l’eau, le •OH est le plus efficace pour creer des cassures a
l’ADN, alors que l’oxygene (O2) se fixe a l’ADN et inhibe les mecanismes de reparation.
Dans les cellules hypoxiques, cet effet de l’oxygene n’est pas present, rendant le traitement
du cancer plus difficile. Par contre, etant des capteurs d’e−hyd, l’absence d’oxygene rend
importante la contribution de ces electrons aux dommages a l’ADN radio-induit, ce qui
suscite l’interet de ce travail qui consiste a etudier les mecanismes d’interaction entre les
e−hyd et l’adduit cisplatine-ADN.
Une fois lie a l’ADN, l’agent chimiotherapeutique (cisplatine) augmente l’interaction
des electrons avec l’ADN, du a son affinite electronique elevee et son caractere polaire, ce
qui ouvre une voie a des dommages induits supplementaires. Les electrons de basse energie
sont produits en grande quantite par les rayonnements ionisants. Apres avoir perdu leur
energie, ces electrons sont ensuite pieges dans leur environnement polaire forme par l’eau.
Ces e−hyd sont connus inoffensifs pour les cassures de brin a l’ADN, par contre, la presence
du cisplatine deforme le puits de potentiel dans laquelle l’e−hyd est piege et le rend reactif
aux cassures de brin d’ADN.
Dans ce projet, on etudie l’effet des e−hyd sur l’adduit de l’ADN-cisplatine. Pour isoler

1 : INTRODUCTION 22
l’effet des e−hyd et supprimer la contribution des electrons prehydrates, les solutions choisies
sont a faibles concentrations d’ADN (<10−3 M). Cette etude a ete menee sur des sys-
temes simples d’oligonucleotides ayant differentes configurations d’attachement de l’ad-
duit. La presente these consiste a etudier l’effet des e−hyd sur les complexes oligonucleotide-
cisplatine. La technique du gel d’electrophorese combinee a celle d’HPLC vont permettre
de mesurer l’interaction de ces electrons avec l’adduit de cisplatine lie a l’ADN et de quan-
tifier les dommages aux bases. Finalement, la technique de radiolyse pulsee va permettre
de mesurer la constante de vitesse de la reaction de l’e−hyd avec l’adduit de cisplatine-
oligonucleotide et faire la comparaison avec celle d’oligonucleotide seule et le cisplatine
hydrolyse.

ARTICLE 1
Hydrated Electrons React with High Specificity withCisplatin Bound to Single-Stranded DNA
B. Behmand, P. Cloutier, S. Girouard, J. R. Wagner, L. Sanche, and D. J. Hunting J.
Phys. Chem. B, 2013, 117, 15994-15999
Avant-propos : Ma contribution a ete d’effectuer la totalite des experiences et d’ana-
lyser les resultats. J’ai aussi redige l’article sous la supervision de Darel Hunting et Leon
Sanche.
Resume : Dans cet article, l’effet des e−hyd issus de la radiolyse de l’eau a ete mesure sur
des oligonucleotides TTTTTGTGTTT et TTTTTTTGTTT modifies ou non-modifies
par l’adduit de cisplatine. Les echantillons ont ete irradies par des rayons γ avec des
doses de 0 a 2500 Gy et les resultats ont ete analyses par la methode de gel d’electro-
phorese. Ces resultats montrent que les e−hyd interagissent particulierement sur le site de
la liaison de Pt-guanine et il detache l’adduit de cisplatine via le transfert dissociatif de
l’electron. De plus, au contraire de l’effet des radicaux •OH, les e−hyd ne produisent pas
de cassures de brin a oligonucleotide avec ou sans l’adduit de cisplatine.
23

2 : ARTICLE 1 24
ABSTRACT : Short oligonucleotides TTTTTGTGTTT and TTTTTTTGTTT in
solution with and without cisplatin (cisPt) bound to the guanine bases were irradiated
with γ-rays at doses varying from 0 to 2500 Gy. To determine the effect of hydrated
electrons from water radiolysis on the oligonucleotides, we quenched •OH radicals with
ethylenediaminetetraacetic acid (EDTA) and displaced oxygen, which reacts with hy-
drated electrons, by bubbling the solution with wet nitrogen. DNA strand breaks and
platinum detachment were quantified by gel electrophoresis. Our results demonstrate that
hydrated electrons react almost exclusively at the position of the cisPt adduct, where they
induce cisPt detachment from one or both guanines in the oligonucleotide. Given the high
yield of hydrated electrons in irradiated tissues, this reaction may be an important step
in the mechanism of radiosensitization of DNA by cisPt.

2 : ARTICLE 1 25
INTRODUCTION
Cisplatin (cisPt) or cis-diamminedichloroplatinum(II) is a chemotherapeutic agent wi-
dely used in the treatment of testicular, ovary, neck, lung, and head cancers.1−3 cisPt is
composed of a single platinum atom bound to two chlorine atoms and two ammonium
groups. In blood and interstitial fluid, the chloride concentration is high (104 mM), and
thus, cisPt conserves its chlorine atoms and remains intact. In contrast, inside cells, the
chloride concentration is considerably reduced (5 mM), which causes the loss of chlo-
rine atoms by hydrolysis, yielding [Pt(NH3)2Cl(H2O)]+ and [Pt(NH3)2(H2O)2]2+. These
products react with DNA, RNA, and proteins as well as with phospholipids in the cell
membrane.4−6 Of these reactions, the binding of cisPt to DNA is the dominant process.5,7
cisPt binds to the N7 purine sites, preferentially on guanine, yielding 60-65% intrastrand
GG cross-links, 25% intrastrand AG cross-links, with a small percentage of intrastrand
GNG cross-links.8 Such binding results in structural DNA anomalies such as a 23◦ un-
winding and a 33◦ bending angle for the case of GNG intrastrand cross-links. The latter
inhibits DNA replication and transcription, which can prevent cancer cell growth and
lead to apoptosis.9
Clinical studies have shown that concomitant chemotherapy and radiotherapy of can-
cer can enhance the survival rate of patients compared to nonsynchronous treatments.10−12
Two nonexclusive mechanisms, related to the binding of cisPt in vivo with DNA, have
been proposed to explain these results. Studies in cultured cells13 and tumor-bearing
mice14 suggest that the binding of HMG1 to cisPt adducts may inhibit repair of radia-
tion damage.7 There is also evidence that the presence of the chemotherapeutic agent
bound to DNA can increase the initial yield of radiation damage.15
This initial damage to DNA in cells proceeds via two mechanisms referred to as the
direct and the indirect effects.In the direct effect, the incident particle interacts directly
with DNA components (bases, sugar, and phosphate) to produce radical cations and se-
condary electrons (SE). In addition, water molecules tightly bound to the DNA undergo
ionization but rapidly transfer the electron-deficient ”hole” to DNA bases. The vast majo-
rity of SE have energies below 20 eV,16,17 and they in turn can produce large quantities of
highly reactive excited molecules, radicals, cations, and anions.19 The direct effect indu-
ced by low-energy electrons (LEEs) has been elegantly demonstrated in experiments that
measured the yield of DNA single (SSBs) and double (DSBs) strand breaks as a function
of the energy of electrons in the range of 1-20 eV. Under dry conditions, it was found that

2 : ARTICLE 1 26
below 15 eV, the damage results from the formation of transient negative ions.20−22 More
recently, experiments with lyophilized supercoiled plasmid DNA irradiated by LEEs have
shown that the yields of SSB and DSB were considerably enhanced in the presence of
cisPt.23 In the indirect effect, radiation produces reactive species in the vicinity of DNA
that diffuse and react with it.24 Because water is the principal component of cells, most
of these reactive species arise from water radiolysis. Given that at least 50% of the total
damage caused to DNA in irradiated cells arises from the indirect effect,25 this type of da-
mage is important for understanding cisPt radiosensitization. Experiments with aqueous
and dry irradiated plasmids have shown that damages caused by the indirect effect are
at least as important as those resulting from the direct effect.26 The most abundant ra-
dicals at 10−6 s following irradiation of water are •OH, hydrated electrons, and •H with
G values of 0.24, 0.28, and 0.06 μmol/J, respectively.27
•OH radicals are the most reactive species for the induction of DNA damage via the
indirect effect.24 However, before becoming hydrated, LEEs enter in a prehydrated state19
when their energies become lower than that of the conduction level. In the prehydrated
state, those electrons, with lifetimes less than 1 ps, can also react with and damage
DNA18,19 through dissociative electron transfer (DET).28 Time-resolved femtosecond laser
spectroscopy experiments have shown that the purine bases, particularly guanine, are
more sensitive to DET than pyrimidine bases.28 With similar experiments, Lu et al.8,29
have shown a high DET reactivity of prehydrated electrons with cisPt.
Hydrated electrons are trapped in potential energy wells (-3.2 eV), but they still react
with DNA to induce damage on the nucleobases.30,31 However, they do not produce SSBs
or DSBs in normal, unmodified DNA.32 Nevertheless, hydrated electrons have been found
to induce SSBs and interstrand cross-links in double-stranded oligonucleotides having the
radiosensitizer 5-bromodeoxyuridine incorporated into mismatch structures.30,31
DNA damage induced by cisPt in mouse fibroblast cells has been investigated by mass
spectroscopy. Iijima et al. observed the lesion principally at the G-G and A-G sites.33 This
technique has also been used to show the interaction of LEEs with cisPt, which leads to
the loss of chlorine from cisPt in a one- and two-step process via dissociative electron
attachment.34
In the present work, the damages induced by hydrated electrons, generated by wa-
ter radiolysis, on olignucleotide-cisPt complexes are investigated for cisPt mono- and
diadducts.

2 : ARTICLE 1 27
MATERIALS AND METHODS
Reaction of Oligonucleotides with CisPt. The single strand oligonucleotides ODN-
GTG and ODN-TGT were purchased from the DNA synthesis laboratory at the Univer-
sity of Calgary, Alberta, Canada. CisPt was first dissolved in water at a concentration
of 220 μM. The oligonucleotide was then mixed with a solution of 44 μM cisPt to give a
ratio of oligonucleotide/cisPt of 1 :5. The solution was kept at roomtemperature for ∼ 46
h to allow reaction of cisPt with the guanines.
Oligonucleotide-CisPt Purification. The oligonucleotide was purified on a G-25 Se-
phadex microcolumn followed by highperformance liquid chromatography (HPLC) to
separate the different products (oligonucleotide and oligonucleotide-cisPt complex). A
linear gradient (0-15% acetonitrile in ammonium acetate) and a column (5 μm ODS A
250 × 6 mm ; YMC) were used for the separation.
3′ End Labeling of Oligonucleotides. The oligonucleotides were end-labeled with
γ-32P ddATP using terminal deoxynucleotidyl transferase (DTD) and buffer. Afterward,
they were purified with a G-25 Sephadex microcolumn.
Experimental Conditions. The final concentration of the oligonucleotide was 0.03 μM
in phosphate buffer (pH 7.0, 10 mM). •OH radicals were scavenged with ethylenediamine-
tetraacetic acid (25 mM EDTA). N2O was also used to scavenge the hydrated electrons.
To minimize scavenging of hydrated electrons by oxygen, the oligonucleotide solutions
were bubbled with wet nitrogen gas (purity of 99.998%) for 1 min. Considering the short
lifetime of the prehydrated electron (∼10−13 s) and the very low oligonucleotide concen-
tration (∼10−8 M) relative to the high amount of the hydrated electrons generated over
the irradiation time (2.4 ×10−3 M for a dose of 1000 Gy), we expect hydrated electron
reactions to be dominant in our solution, while the action of prehydrated electrons is
considered negligible. Furthermore, if the prehydrated electron has sufficient time to dif-
fuse and react with the DNA, then the •OH should also be able to react with DNA
before being captured by EDTA, and this is not the case, asdemonstrated by the absence
of DNA strand breaks.
Irradiation. Oligonucleotide solutions were irradiated in a cesium-137 Gammacell with
the dose varying between 500 and 2500 Gy (11.6 Gy/min).
Denaturing Gel Electrophoresis. Oligonucleotides were loaded on a 7 M urea dena-
turing 20% polyacrylamide gel. A molecular weight ladder was produced by Maxam and
Gilbert sequencing treatment. An electric field was applied to the electrophoresis gel for

2 : ARTICLE 1 28
2.5 h. The phosphor screen cassette (Molecular Dynamics Inc.) was exposed to the gel
for 16 h and scanned with a Storm fluorescence scanner (Molecular Dynamics Inc.).
RESULTS
Damages Induced to Oligonucleotides, with and without CisPt, by OH Radicals and
Hydrated Electrons. Figure 1 shows the action of •OH radicals and hydrated electrons on
the single-stranded ODN-GTG and the oligonucleotide-cisPt complex (ODN-GTG-cisPt)
in lanes 1-14 and lanes 16-29, respectively. As can be seen, the band corresponding to the
intact ODN-GTG-cisPt (lanes 16-29) migrates slower than the band for the unmodified
oligonucleotide (lanes 1-14). This shift results from a different charge and geometry of
the oligonucleotide containing a cisPt adduct. In lane 1, corresponding to the control (0
Gy) with no cisPt adduct, there are strand breaks (8.8%) that are probably induced by
electron emission from 32P. Strikingly, no such damage was observed for the ODN-GTG-
cisPt at 0 Gy, even though this complex was labeled with 32P at the same specific activity
as the control oligonucleotide.
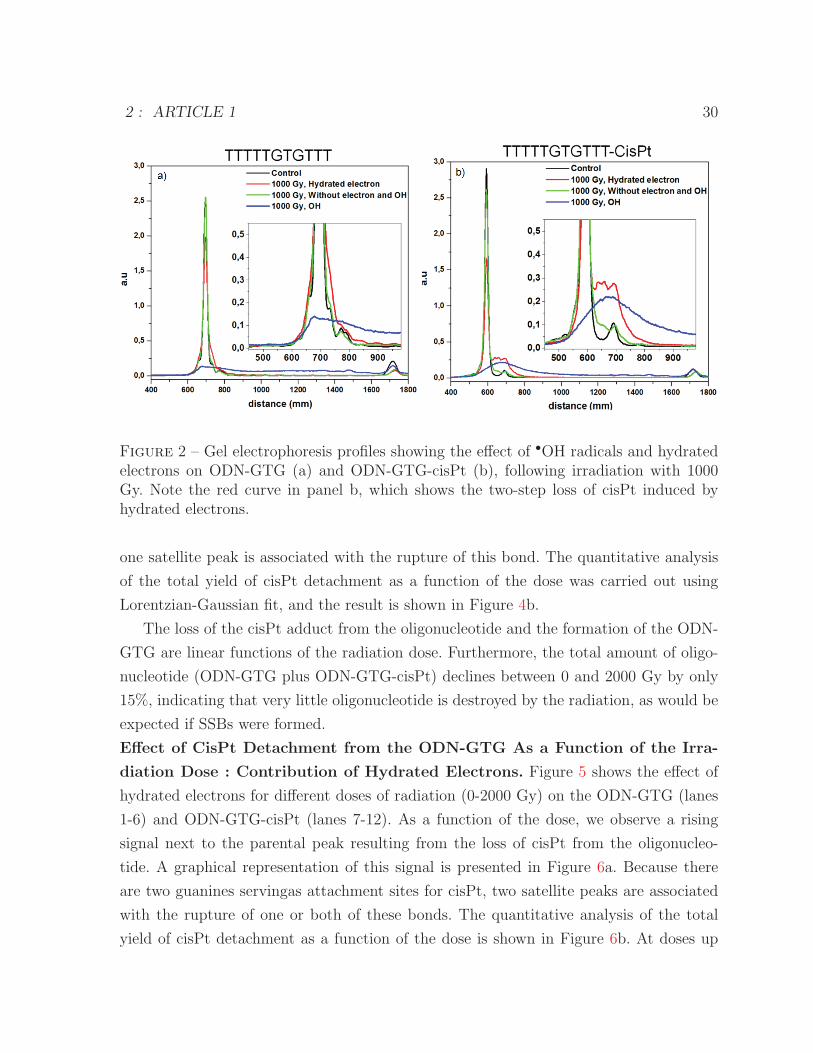
Figure 2 is a graphical representation of the gel in Figure 1 for an irradiation dose of
1000 Gy. The black curves in (a) and (b) show the control oligonucleotide (700 mm of
migration) and ODN-GTG-cisPt (600 mm of migration), respectively, prior to irradiation.
In the latter, a small amount of the oligonucleotide (3.1%) lacking the cisPt adduct is
observed as a satellite peak at 700 mm. The blue curves in (a) and (b) show that extensive
DNA damage is induced by the •OH radical as the principal peak vanishes to give rise to
fragments. This occurs for both the oligonucleotides without and with the cisPt adduct.
In contrast, the red curves show that the oligonucleotide damage induced by the hydrated
electron does not give rise to strand breaks but rather to rupture of the cisPt bond. As
shown in Figure 2b, this effect results in two satellite peaks. Our interpretation is that
the peak at 650 mm of migration corresponds to an oligonucleotide with one cisPt bond
broken and the second peak at 700 mm to the oligonucleotide without cisPt. This suggests
an interaction between the hydrated electron and cisPt leading to bond rupture (one or
two bonds). Presumably, the liberated cisPt then reacts with water in an exothermic
process.34
The green curves, where both •OH radicals and hydrated electrons are captured,
are similar to the control spectra at 0 Gy (black curves), as expected, but also bear
some slight resemblance to the hydrated electron curve, presumably because of the small

2 : ARTICLE 1 29
Figure 1 – Gel electrophoresis showing the effect of •OH and hydrated electrons onoligonucleotides, with and without the cisPt adduct, as a function of irradiation dose.The first lanes (3, 6, 9, 12, 13, 14, 18, 21, 24, 27, 28, 29) for each radiation dose show theeffects of hydrated electrons because the •OH radicals were captured by the scavengerEDTA and O2 was displaced by bubbling the solution with wet nitrogen. In the secondlanes for each dose (4, 7, 10, 19, 22, 25), both •OH radicals and hydrated electrons werescavenged in the presence of EDTA by bubbling the solution with wet N2O. The thirdlanes for each dose (5, 8, 11, 20, 23, 26) show the effect of •OH radicals, with hydratedelectrons being scavenged by O2. Lane 15 is not pertinent.
amount of nonscavenged hydrated electrons. We observe residual breakage of the cisPt
bond, corresponding to about 0.13% for one bond broken and 1.4% for two bonds broken.
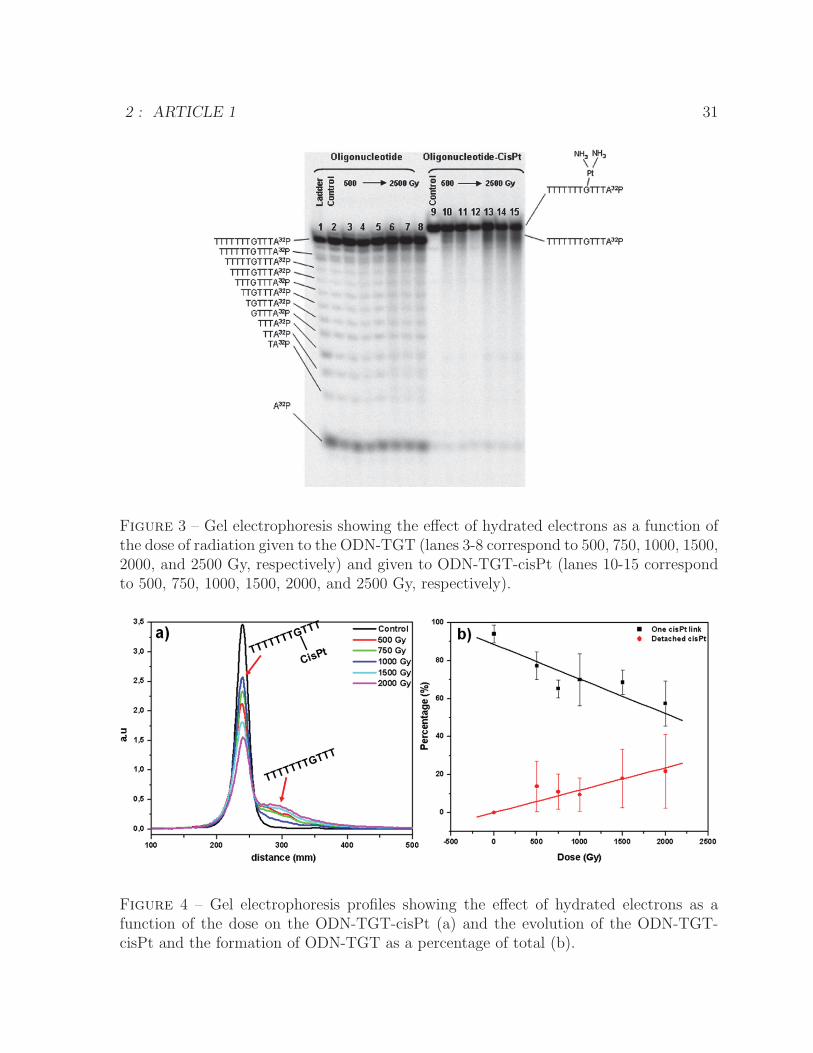
CisPt Monoadduct Detachment from ODN-TGT Induced by Hydrated Elec-
trons. Figure 3 shows the effect of hydrated electrons for different irradiation doses
(0-2500 Gy) on the ODN-TGT (lanes 3-8) and ODN-TGT-cisPt (lanes 10-15). In the
oligonucleotide containing a cisPt adduct, we observe as a function of dose a rising signal
next to the parental oligonucleotide peak, which appears at the same position as the
unmodified oligonucleotide. This signal is therefore attributed to the loss of cisPt from
the oligonucleotide. A graphical representation of this detachment is presented in Figure
4a, where the peak at 300 mm of migration represents the oligonucleotide without cisPt.
Because there is only a single guanine for attachment of cisPt in this oligonucleotide, only
2 : ARTICLE 1 30
Figure 2 – Gel electrophoresis profiles showing the effect of •OH radicals and hydratedelectrons on ODN-GTG (a) and ODN-GTG-cisPt (b), following irradiation with 1000Gy. Note the red curve in panel b, which shows the two-step loss of cisPt induced byhydrated electrons.
one satellite peak is associated with the rupture of this bond. The quantitative analysis
of the total yield of cisPt detachment as a function of the dose was carried out using
Lorentzian-Gaussian fit, and the result is shown in Figure 4b.
The loss of the cisPt adduct from the oligonucleotide and the formation of the ODN-
GTG are linear functions of the radiation dose. Furthermore, the total amount of oligo-
nucleotide (ODN-GTG plus ODN-GTG-cisPt) declines between 0 and 2000 Gy by only
15%, indicating that very little oligonucleotide is destroyed by the radiation, as would be
expected if SSBs were formed.
Effect of CisPt Detachment from the ODN-GTG As a Function of the Irra-
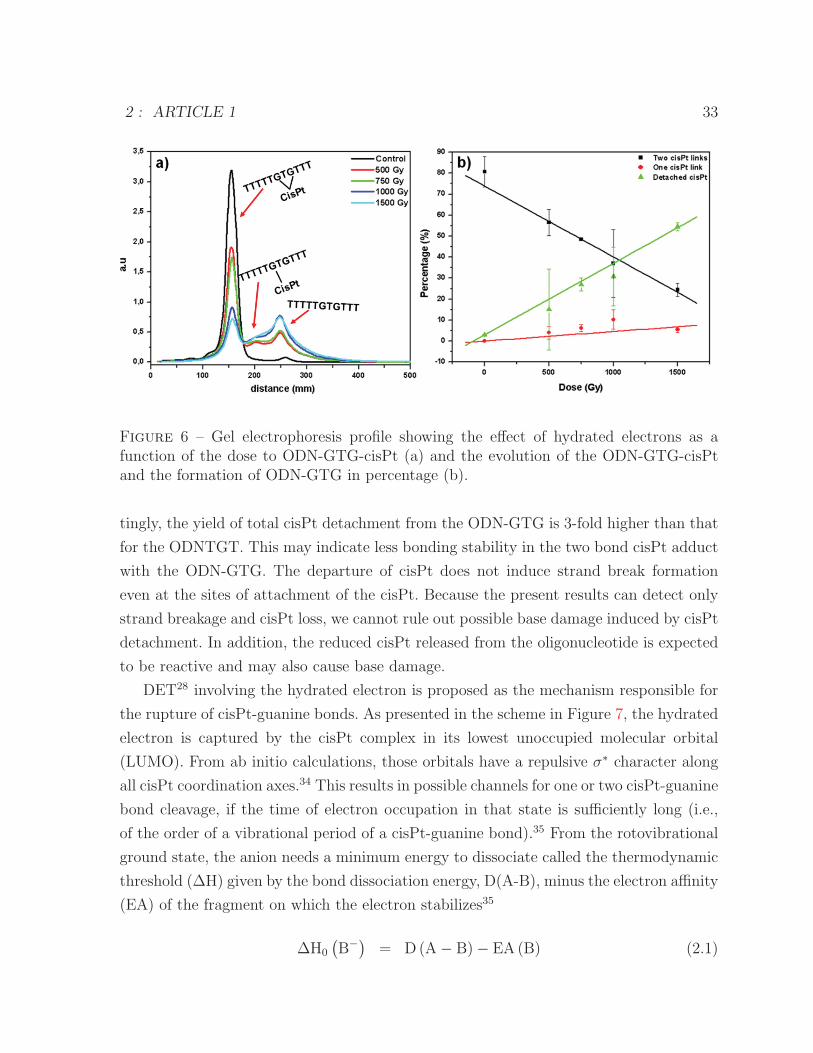
diation Dose : Contribution of Hydrated Electrons. Figure 5 shows the effect of
hydrated electrons for different doses of radiation (0-2000 Gy) on the ODN-GTG (lanes
1-6) and ODN-GTG-cisPt (lanes 7-12). As a function of the dose, we observe a rising
signal next to the parental peak resulting from the loss of cisPt from the oligonucleo-
tide. A graphical representation of this signal is presented in Figure 6a. Because there
are two guanines servingas attachment sites for cisPt, two satellite peaks are associated
with the rupture of one or both of these bonds. The quantitative analysis of the total
yield of cisPt detachment as a function of the dose is shown in Figure 6b. At doses up
2 : ARTICLE 1 31
Figure 3 – Gel electrophoresis showing the effect of hydrated electrons as a function ofthe dose of radiation given to the ODN-TGT (lanes 3-8 correspond to 500, 750, 1000, 1500,2000, and 2500 Gy, respectively) and given to ODN-TGT-cisPt (lanes 10-15 correspondto 500, 750, 1000, 1500, 2000, and 2500 Gy, respectively).
Figure 4 – Gel electrophoresis profiles showing the effect of hydrated electrons as afunction of the dose on the ODN-TGT-cisPt (a) and the evolution of the ODN-TGT-cisPt and the formation of ODN-TGT as a percentage of total (b).

2 : ARTICLE 1 32
Figure 5 – Gel electrophoresis showing the effect of hydrated electrons as a function ofthe dose to ODN-GTG (lanes 2-6 correspond to 500, 750, 1000, 1500, and 2000, respec-tively) and ODN-GTG-cisPt (lanes 8-12 correspond to 500, 750, 1000, 1500, and 2000,respectively).
to 1500 Gy, the parental peak decreases linearly, and product peaks increase linearly.
Very little oligonucleotide containing a single oligonucleotide-cisPt bond was observed.
As shown in Figure 6b, the slope of the dose-response curve for the rupture of one of
the two cisPt-guanine bonds in the ODN-GTG is 7-fold smaller than the slope for the
rupture of both cisPt bonds. Interestingly, the parental peak associated with the complex
ODN-GTG-cisPt decreases faster than the one associated with the ODN-TGT-cisPt in
Figure 4, indicating that it is easier to detach cisPt when it is doubly bonded to the
oligonucleotide than when it is singly bonded.
DISCUSSION
The main effect observed by irradiating the ODN-GTG-cisPt and ODN-TGT-cisPt in
solution is the detachment of the cisPt adduct from the oligonucleotide. In the case of
ODN-GTG-cisPt, this detachment can occur in either a one- or twostep process. Interes-
2 : ARTICLE 1 33
Figure 6 – Gel electrophoresis profile showing the effect of hydrated electrons as afunction of the dose to ODN-GTG-cisPt (a) and the evolution of the ODN-GTG-cisPtand the formation of ODN-GTG in percentage (b).
tingly, the yield of total cisPt detachment from the ODN-GTG is 3-fold higher than that
for the ODNTGT. This may indicate less bonding stability in the two bond cisPt adduct
with the ODN-GTG. The departure of cisPt does not induce strand break formation
even at the sites of attachment of the cisPt. Because the present results can detect only
strand breakage and cisPt loss, we cannot rule out possible base damage induced by cisPt
detachment. In addition, the reduced cisPt released from the oligonucleotide is expected
to be reactive and may also cause base damage.
DET28 involving the hydrated electron is proposed as the mechanism responsible for
the rupture of cisPt-guanine bonds. As presented in the scheme in Figure 7, the hydrated
electron is captured by the cisPt complex in its lowest unoccupied molecular orbital
(LUMO). From ab initio calculations, those orbitals have a repulsive σ∗ character along
all cisPt coordination axes.34 This results in possible channels for one or two cisPt-guanine
bond cleavage, if the time of electron occupation in that state is sufficiently long (i.e.,
of the order of a vibrational period of a cisPt-guanine bond).35 From the rotovibrational
ground state, the anion needs a minimum energy to dissociate called the thermodynamic
threshold (ΔH) given by the bond dissociation energy, D(A-B), minus the electron affinity
(EA) of the fragment on which the electron stabilizes35
ΔH0
(B−) = D(A− B)− EA (B) (2.1)

2 : ARTICLE 1 34
Figure 7 – Dissociative electron-transfer mechanism, involving a hydrated electron, re-sulting in bond cleavage between cisPt and its complex.
Thus, for DET to occur, the potential energy surfaces of the transient anion along
the cisPt-guanine bonds must at least partly lie above •H in the Franck-Condon region.
During the separation of cisPt from guanine, the fragment with the greater EA is ex-
pected to keep the extra electron because it corresponds to the lowest thermodynamic
threshold according to the above equation. In Figure 6b, the conversion of the ODN-
GTG-cisPt to the ODN-GTG occurs as a linear function of dose, which suggests that a
single hydrated electron induces the simultaneous breakage of two cisPt-guanine bonds. A
priori, this double bond break in ODN-GTG-cisPt can occur in two ways : either a cisPt-
guanine bond is first broken and the remaining fragments react with and dissociate the
other cisPt-guanine bond, or the transient anion formed via DET dissociates by simulta-
neously breaking the two cisPt-guanine bonds. The latter mechanism has been observed
by Kopyra et al.34 using a mass spectrometer as a detector, in LEE impact experiments
on gaseous cisPt, containing its original chlorine atoms. They found that a transient
anion formed at ∼0 eV dissociates by simultaneously breaking the bonds between Pt and
the chlorine atoms. The platinated fragment has a much higher probability to keep the
extra electron due to its high electron affinity. This possibility within a ODN-GTG-cisPt
is represented in the scheme of Figure 7. Judging from the huge magnitude of cisPt bond
scission with respect to that of other bonds within the oligonucleotide, observed in our
experiment, hydrated electrons appear to localize preferentially at the site of cisPt, where
they undergo DET. This high affinity for hydrated electrons is probably due to the high
electron affinity of the cisPt adduct.

2 : ARTICLE 1 35
CONCLUSION
The present results using a γ-irradiated oligonucleotide-cisPt solution have demonstra-
ted a strong interaction between hydrated electrons and the cisPt monoadduct at the
guanine base and the intrastrand adduct bonded between two guanine bases separated
by thymine. This interaction leads to hydrated electron capture by the oligonucleotide
almost exclusively at the site of binding of cisPt. The transient anion formed by this
attachment of the hydrated electron dissociates by breaking one or two cisPt-guanine
bonds. In the latter case, cisPt detaches from the oligonucleotide. Although the reaction
of cisPt adducts with hydrated electrons does not lead to strand break formation when
cisPt detaches from DNA, it may induce base damage at adjacent sites and thus be po-
tentially cytotoxic especially in hypoxic tumor cells because oxygen quenches hydrated
electrons.
AUTHOR INFORMATION
Corresponding Author
*E-mail : [email protected]. Address : Universite de Sherbrooke, Facultee
de medecine, Departement de medecine nucleaire et de radiobiologie, 3001, 12e Avenue
Nord, Sherbrooke, QC, Canada, J1H 5N4. Phone : 1 819 820 6868. Fax : 1 819 564-5442.
Notes
The authors declare no competing financial interest.
ACKNOWLEDGMENTS
The authors are grateful to Professor Jean-Paul Jay-Gerin for his helpful comments. Fi-
nancial support for this work was provided by the Canadian Institute of Health Research
(CIHR).
REFERENCES
(1) Kelland, L. The Resurgence of Platinum-Based Cancer Chemotherapy. Nat. Rev.
Cancer 2007, 7, 573-584.
(2) Go, R. S. ; Adjei, A. A. Review of the Comparative Pharmacology and Clinical Ac-
tivity of Cisplatin and Carboplatin. J. Clin. Oncol. 1999, 17, 409-422.

2 : ARTICLE 1 36
(3) Burger, A. M. ; Double, J. A. ; Newell, D. R. Inhibition of Telomerase Activity by
Cisplatin in Human Testicular Cancer Cells. Eur. J. Cancer 1997, 33, 638-644.
(4) Pascoe, J. M. ; Roberts, J. J. Interactions between Mammalian Cell DNA and Inor-
ganic Platinum Compounds-I. Biochem. Pharmacol. 1974, 23, 1345-1357.
(5) Akaboshi, M. ; Kawai, K. ; Maki, H. ; Akuta, K. ; Ujeno, Y. ; Miyahara, T. The Num-
ber of Platinum Atoms Binding to DNA, RNA and Protein Molecules of HeLa Cells
Treated with Cisplatin at Its Mean Lethal Concentration. Jpn. J. Cancer Res. 1992, 83,
522-526.
(6) Speelmans, G. ; Staffhorst, R. W. H. M. ; Versluis, K. ; Reedijk, J. ; Kruijff, B. Cisplatin
Complexes with Phosphatidylserine in Membranes. Biochemistry 1997, 36, 10545-10550.
(7) Jamieson, E. R. ; Lippard, S. J. Structure, Recognition, and Processing of Cisplatin-
DNA Adducts. Chem. Rev. 1999, 99, 2467-2498.
(8) Lu, Q.-B. ; Kalantari, S. ; Wang, C.-R. Electron Transfer Reaction Mechanism of Cis-
platin with DNA at the Molecular Level. Mol. Pharm. 2007, 4, 624-628.
(9) Wozniak, K. ; Blasiak, J. Recognition and Repair of DNA-Cisplatin Adducts. Acta
Biochim. Pol. 2002, 49, 583-596.
(10) Eifel, P. J. Concurrent Chemotherapy and Radiation Therapy as the Standard of
Care for Cervical Cancer. Clin. Pract. Oncol. 2006, 5, 248-255.
(11) Devita, V. T., Jr. ; Hellman, S. ; Resenberg Cancer : Principles and Practice of On-
cology ; Lippcott Williams and Wilkins : New York, 2001.
(12) Brabec, V. In Platinum-Based Drugs in Cancer Therapy ; Kelland, L. R., Farrell,
N., Eds. ; Humana Press, Inc. : Totowa, NJ, 2000.
(13) Zimbrick, J. D. ; Sukrochana, A. ; Richmond, R. C. Studies on Radiosensitization
of Escherichia coli Cells by Cis-Platinum Complexes. Int. J. Radiat. Oncol. Biol. Phys.
1979, 5, 1351-1354.
(14) Lelieveld, P. ; Scoles, M. A. ; Brown, J. M. ; Phil, D. ; Kallman, R. F. The Effect
of Treatment in Fractionatesd Schedules with the Combination of X-Irradiation and Six
Cytotoxic Drugs on the RIF-1 Tumor and Normal Mouse Skin. Int. J. Radiat. Oncol.
Biol. Phys. 1985, 11, 111-121.
(15) Dewit, L. Combined Treatment of Radiation and Cis-Diamminedichloroplatinum
(II) : A Review of Experimental and Clinical Data. Oncol. Biol. Phys. 1987, 13, 403-426.
(16) LaVerne, J. A. ; Pimblott, M. Electron Energy-Loss Distributions in Solid, Dry DNA.
Radiat. Res. 1995, 141, 208-215.
(17) Cobut, V. ; Frongillo, Y. ; Patau, J. P. ; Goulet, T. ; Fraser, M.-J. ; Jay-Gerin, J.-P.

2 : ARTICLE 1 37
Monte Carlo Simulation of Fast Electron and Proton Tracks in Liquid Water-I. Physical
and Physicochemical Aspects. Radiat. Phys. Chem. 1998, 51, 229-243.
(18) Sanche, L. Beyond Radical Thinking. Nature 2009, 461, 358-359.
(19) Alizadeh, E. ; Sanche, L. Precursors of Solvated Electrons in Radiobiological Physics
and Chemistry. Chem. Rev. 2012, 112, 5578-5602.
(20) Martin, F. ; Burrow, P. D. ; Cai, Z. ; Cloutier, P. ; Hunting, D. J. ; Sanche, L. DNA
Strand Breaks Induced by 0-4 eV Electrons : The Role of Shape Resonances. Phys. Rev.
Lett. 2004, 93, 068101/1-068101/4.
(21) Boudaıffa, B. ; Cloutier, P. ; Hunting, D. J. ; Huels, M. A. ; Sanche, L. Resonant
Formation of DNA Strand Breaks by Low-Energy (3 to 20 eV) Electrons. Science 2000,
287, 1658-1660.
(22) Zheng, Y. ; Wagner, J. R. ; Sanche, L. DNA Damage Induced by Low-Energy Elec-
trons : Electron Transfer and Diffraction. Phys. Rev. Lett. 2006, 96, 208101/1-208101/4.
(23) Zheng, Y. ; Hunting, D. J. ; Ayotte, P. ; Sanche, L. Role of Secondary Low-Energy
Electrons in the Concomitant Chemoradiation Therapy of Cancer. Phys. Rev. Lett. 2008,
100, 198101/1-198101/4.
(24) von Sonntag, C. The Chemical Basis for Radiation Biology ; Taylor and Francis :
London, 1987.
(25) deLara, C. M. ; Jenner, T. J. ; Marsden, S. J. ; O’Neill, P. The Effect of Dimethyl
Sulfoxide on the Induction of DNA Double-Strand Breaks in V79-4 Mammalian Cells by
Alpha Particles. Radiat. Res. 1995, 144, 43-49.
(26) Ito, T. ; Baker, S. C. ; Stickley, C. D. ; Peak, J. G. ; Peak, M. J. Dependence of the
Yeild of Strand Breaks Induced by γ-Rays in DNA on the Physical Conditions of Expo-
sure : Water Content and Temperature. Int. J. Radiat. Biol. 1993, 63, 289-296.
(27) Ward, J. F. DNA Damage Produced by Ionizing Radiation in Mammalian Cells :
Identities, Mechanisms of Formation, and Reparability. Prog. Nucl. Acid Res. 1988, 35,
95-125.
(28) Wang, C.-R. ; Nguyen, J. ; Lu, Q.-B. Bond Breaks of Nucleotides by Dissociative
Electron Transfer of Nonequilibrium Prehydrated Electrons : A New Molecular Mecha-
nism for Reductive DNA Damage. J. Am. Chem. Soc. 2009, 131, 11320-11322.
(29) Lu, Q.-B Effects and Applications of Ultrashort-Lived Prehydrated Electrons in Ra-
diation Biology and Radiotherapy of Cancer. Mutat. Res. 2010, 704, 190-199.
(30) Dextraze, M.-E. ; Wagner, J. R. ; Hunting, D. J. 5-Bromodeoxyuridine Radiosensiti-
zation : Conformation-Dependent DNA Damage. Biochemistry 2007, 46, 9089-9097.

2 : ARTICLE 1 38
(31) Cecchini, S. ; Girouard, S. ; Huels, M. A. ; Sanche, L. ; Hunting, D. J. Single-Strand-
Specific Radiosensitization of DNA by Bromodeoxyuridine. Radiat. Res. 2004, 162, 604-
615.
(32) Michael, B. D. ; O’Neill, P. A Sting in the Tail of Electron Tracks. Science 2000, 287,
1603-1604.
(33) Iijima, H. ; Patrzyc, H. B. ; Dawidzik, J. B. ; Budzinski, E. E. ; Cheng, H.-C. ; Freund,
H. G. ; Box, H. C. Measurement of DNA Adducts in Cells Exposed to Cisplatin. Anal.
Biochem. 2004, 333, 65-71.
(34) Kopyra, J. ; Koenig-Lehmann, C. ; Bald, I. ; Illenberger, E. A Single Slow Electron
Triggers the Loss of Both Chlorine Atoms from the Anticancer Drug Cisplatin : Implica-
tions for Chemoradiation Therapy. Chem. Int. Ed. 2009, 48, 7904-7907.
(35) Arumainayagam, C. R. ; Lee, H. L. ; Nelson, R. B. ; Haines, D. R. ; Gunawardane, R.
P. Low-Energy Electron-Induced Reactions in Condensed Matter. Surf. Sci. Rep. 2010,
65, 1-44.

ARTICLE 2
Cisplatin Intrastrand Adducts Sensitize DNA toBase Damage by Hydrated Electrons
B. Behmand, J. R. Wagner, L. Sanche, and D. J. Hunting
J. Phys. Chem. B, 2014, 118, 4803-4808
Avant-propos : Ma contribution a ete d’effectuer la totalite des experiences et d’analy-
ser les resultats. J’ai aussi redige l’article sous la supervision de Darel Hunting, Richard
Wagner et Leon Sanche.
Resume : Les dommages aux bases d’oligonucleotide (TTTTTGTGTTT) avec et sans
l’adduit de cisplatine ont ete quantifies suite a leur interaction avec des e−hyd. De plus, la
structure initiale de l’adduit de cisplatine-oligonucleotide a ete confirmee par spectrome-
trie de masse. Dans cet article, la technique d’HPLC suite a la digestion enzymatique a
ete utilisee pour identifier les dommages aux bases. Ces resultats montrent que les e−hydproduisent des dommages aux guanines, soit les sites d’attachement de cisplatine, et aussi
aux thymines. Le detachement de l’adduit de cisplatine a ete confirme par cette tech-
nique. Par exemple, a 1000 Gy, 2,5 thymines et 1 guanine sont endommagees en moyenne
pour chaque perte de l’adduit de cisplatine.
39

3 : ARTICLE 2 40
ABSTRACT : The oligonucleotide TTTTTGTGTTT with or without a cispla-
tin adduct was reacted with hydrated electrons generated by ionizing radiation. Hy-
droxyl radicals were quenched with ethylenediaminetetraacetic acid (EDTA), and the
solutions were bubbled with wet nitrogen to eliminate oxygen, a scavenger of hydrated
electrons. Prior to irradiation, the structure of the initial cisplatin adduct was identified
by mass spectrometry as G-cisplatin-G. Radiation damage to DNA bases was quantified
by high-performance liquid chromatography (HPLC), after enzymatic digestion of the
TTTTTGTGTTT-cisplatin complex to deoxyribonucleosides. The masses of the plati-
num adducts following digestion and separation by HPLC were measured by mass spec-
trometry. Our results demonstrate that hydrated electrons induce damage to thymines
as well as detachment of the cisplatin moiety from both guanines in the oligonucleotide.
This detachment regenerates both unmodified guanine and damaged guanine, in equi-
molar amounts. At 1000 Gy, a net average of 2.5 thymines and 1 guanine are damaged

3 : ARTICLE 2 41
for each platinum lost from the oligonucleotide. Given the extensive base damage that
occurs for each cisplatin adduct lost, it is clear that, prior to undergoing detachment,
these adducts must catalyze several cycles of reactions of hydrated electrons with DNA
bases. It is likely that a single reaction leads to the loss of the cisplatin adduct and
the damage observed on the guanine base ; however, the damage to the thymine bases
must require the continued presence of the cisplatin adduct, acting as a catalyst. To our
knowledge, this is the first time that platinum-DNA adducts have been shown to have
catalytic activity. We propose two pathways for the interaction of hydrated electrons with
TTTTTGTGTTT-cisplatin : (1) the hydrated electron is initially captured by a thymine
base and transferred by base to base electron hopping to the guanine site, where the
cisplatin moiety detaches from the oligonucleotide via dissociative electron attachment,
and (2) the hydrated electron interacts directly with the platinum-guanine adduct and in-
duces detachment of the cisplatin moiety via dissociative electron attachment. Although
the precise mechanism remains to be elucidated, our results provide important insights
into the radiosensitization of DNA by cisplatin.
INTRODUCTION
Concomitant chemotherapy and radiotherapy have substantially enhanced tumor control
and the survival of patients.1−4 Radiosensitization by chemotherapeutic drugs is pro-
bably due to several factors including an increase in DNA damage, inhibition of DNA
repair, diminution of the apoptosis threshold, and perturbation of the cell cycle such that
cells remain in the sensitive phases for a longer time.5 cis-Diamminedichloroplatinum
(II) or cisplatin is a chemotherapeutic drug used in the treatment of ovary, testicular,
lung, head, and neck cancers.6−8 It binds to the N7 purine sites in DNA, preferentially
on guanine, and creates the structural and chemical anomalies which lead to the effects
listed above.9 When nonthermal electrons interact with this chemotherapeutic agent, a
shape resonance appears near 0 eV in the gas phase, which leads to dissociative electron
attachment (DEA).10 In a polar medium, such as water, this resonance energy is lower by
about 1 eV.11 Cleavage of the bond between the cisplatin moiety (cisPt) and guanine is
also observed on the basis of the fragmentation pattern of Pt-DNA negative ion adducts
during electrospray mass spectrometry.12 Furthermore, electron transfer along the DNA
strand, toward the site of the attachment of cisPt adduct, is expected to increase the
cross section for DEA at the Pt site.13

3 : ARTICLE 2 42
The damage induced by ionizing radiation results from both the direct and indirect
effects. The direct effect is the interaction of either the incident particle or secondary
electrons (SE) generated by the primary radiation with DNA and can produce single and
double DNA strand breaks or base damage. The vast majority of SE have energies of
less than 20 eV.14,15 Zheng et al.16 recently studied the effect of monoenergetic electron
beams (1, 10, 100, 60 000 eV) on thin films of lyophilized DNA with and without cisPt.
They found an enhancement of both single and double strand breaks (SSB and DSB)
in DNA modified by cisPt. Since the major component of cells is water (70-80%), the
indirect effect of radiation is also important and gives rise to about 50% of the total
DNA damage.17,18 In the indirect effect, ionizing radiation interacts with water molecules
and creates the reactive chemical species •OH, H•, and e−hyd that can react with DNA
and create damage. As SE slow down, they start polarizing the electronic orbitals of the
surrounding molecules. As they continue to lose energy below ∼1 eV, they increasingly
polarize the vibrational modes of these molecules, and the phonon modes, to reach a
prehydrated state and become a polaron.19 In this prehydrated state, which lasts for less
than a picosecond, the electron becomes hydrated by surrounding water molecules and
is trapped in an average potential well of ∼-3.2 eV. It has been shown that the hydrated
electron cannot induce strand breaks in unmodified DNA.20,21 However, according to
the results of Behmand et al., based on an electrophoretic assay, hydrated electrons can
induce the detachment of cisplatin, when it is bound to an oligonucleotide.22
Here, we study the effects of the hydrated electrons generated by γ rays on an
oligonucleotide-cisPt complex. Base damage is measured by HPLC following enzyma-
tic digestion of the oligonucleotide. Platinum DNA adducts are measured by HPLC,
mass spectrometry (MS), and electrospray ionization tandem mass spectrometry (ESI-
MS/MS). Our results show that the presence of cisPt adducts greatly increases the da-
mage to guanine and thymine induced by radiation. In addition to confirmation by HPLC
analysis that cisPt detaches from the DNA, our results also indicate that equal amounts
of damaged and undamaged guanine are generated for each cisPt adduct that detaches
from the oligonucleotide.
MATERIALS AND METHODS
The single strand TTTTTGTGTTT (ODN-GTG) was purchased from the DNA synthe-
sis lab (University of Calgary, Alberta, Canada). The details of the reaction of ODN-GTG

3 : ARTICLE 2 43
with cisplatin and purification giving the ODN-GTG-cisPt complex have been reported
elsewhere.22 Although in cells treated with cisplatin, cisPt-GG cross-links outnumber
GTG cross-links, we chose to study the latter adduct because it causes greater DNA
structural deformation.
Experimental Conditions. The concentration of the oligonucleotide was approxima-
tely 14 μM. On the basis of a rate constant calculation, 99.99% of •OH radicals were
expected to be eliminated by 25 mM EDTA as a scavenger. The oligonucleotide solutions
were bubbled for 1 min with wet nitrogen gas (purity of 99.998%) to minimize the oxygen.
Given the low concentration of oligonucleotide in our solutions (14 μM) and the short
half-life of prehydrated electrons, we assume that any reaction of prehydrated electrons
is negligible.
Irradiation. The samples were irradiated with doses ranging from 250 to 1000 Gy using
a cesium-137 irradiator (Best Theratronics, Ottawa, Ontario, Canada).
Digestion. The oligonucleotides (concentration 14 μM) with and without cisplatin were
irradiated and digested at 37 ◦C to deoxyribonucleosides with three enzymes : first, 1 h
with nuclease P1, followed by 1 h with snake venom phosphodiesterase I ; finally, alkaline
phosphatase was added to the sample for 1 h at 37 ◦C. The reaction was stopped with
phosphoric acid and left at 4 ◦C for HPLC analysis, which was performed with a linear
gradient (0-15% acetonitrile in ammonium acetate) and a column (5 μm ODS A 250 ×6 mm ; YMC) for the separation.
Extinction Coefficient. The extinction coefficient of the dG-cisPt-dG adduct (23 050
M−1 cm−1) at 260 nm was calculated on the basis of the extinction coefficients of deoxy-
guanosine (11 500 M−1 cm−1) and cisPt (50 M−1 cm−1).
Mass Spectrometry. The masses of the platinum adducts following digestion were mea-
sured by LC-MS/MS. This machine consists of a Shimadzu HPLC system with an auto
sampler (SIL-HTc), degasser (DGU-14A), column oven (CTO-10ASvp), binary pumps
(LC-10ADvp), UV/vis detector (SPD-20A), and API 3000 tandem mass spectrometer
with a turbo-ion spray source (MDS Sciex, Applied Biosystems). A syringe infusion pump
(Harvard Model 22) was used to optimize the detection conditions, set at a flow rate of 20
μL/min. The cone voltage was 4500 V, and the collision energy was 45 V. Measurements
were made in the negative ion mode.

3 : ARTICLE 2 44
RESULTS
Analysis of the Damage to DNA Bases. In order to investigate the radiosensibili-
zation of a single stranded oligonucleotide by the presence of a single cisplatin adduct,
the oligonucleotide with or without cisPt was irradiated at several doses of ionizing ra-
diation. Analysis of the damage was performed by HPLC following digestion with P1
nuclease and snake venom phosphodiesterase, which cleave phosphodiester bonds to ge-
nerate mononucleotides, and alkaline phosphatase to remove the phosphate from the free
nucleotides. Figure 1a shows the HPLC profile for the ODN-GTG, following enzymatic
digestion to deoxyribonucleosides, without and with cisPt prior to irradiation. The profile
with cisPt shows the absence of dG and an additional peak corresponding to dG-cisPt-dG
as confirmed by MS and discussed below.
Parts b and c of Figure 1 show the HPLC results without and with cisPt, respectively,
as a function of irradiation dose. In the absence of cisPt, the thymidine (dT) peak decrea-
sed gradually with increasing irradiation dose, reflecting the damage to thymine induced
by the hydrated electron, whereas dG remained virtually unchanged. In contrast, with
cisPt, both the thymidine and the dG-cisPt-dG peaks decreased markedly with increa-
sing radiation dose. The dG-cisPt-dG peak disappeared at ∼1000 Gy, while the dG peak,
which was absent at 0 Gy, increased as a function of irradiation dose as a result of the
loss of cisPt from guanine in the oligonucleotide. A quantitative analysis of this process
is shown in Figure 2. In this analysis, we used a value for the extinction coefficient of
dG-cisPt-dG equal to twice that of dG (see Materials and Methods). The black squares
(�) in the figure correspond to the percentage of dG formation from cisPt detachment,
while the red circles (•) correspond to the percentage of the remaining dG attached to
the cisPt. The difference between these two forms of dG indicates the quantity of the
dG which underwent damage as a result of the reaction of hydrated electrons with the
ODN-GTG-cisPt (green triangles (�) on the graph). Thus, the dose-dependent disappea-
rance of the dG-cisPt-dG peak from the HPLC profile corresponds to the generation of
approximately equal amounts of damaged and undamaged dG (Figure 2).
A comparison of the damage induced to dG as a function of the dose with and without
cisPt is shown in Figure 3a, where the potent radiosensitizing effect of the chemothera-
peutic agent is particularly evident. Even at the maximum dose of 1000 Gy, there is no
detectable damage to dG by radiation in the absence of cisPt, whereas, at this dose,
approximately 50% of dG is damaged in the presence of cisPt (Figure 3a). Interestingly,
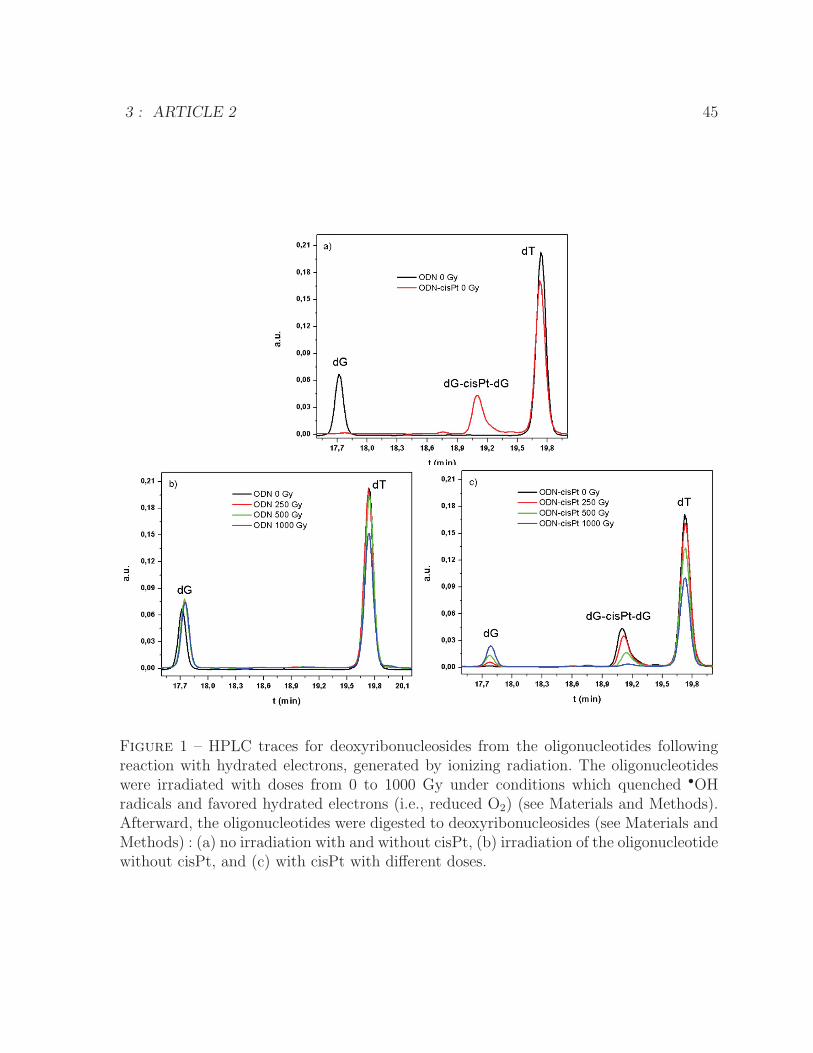
3 : ARTICLE 2 45
Figure 1 – HPLC traces for deoxyribonucleosides from the oligonucleotides followingreaction with hydrated electrons, generated by ionizing radiation. The oligonucleotideswere irradiated with doses from 0 to 1000 Gy under conditions which quenched •OHradicals and favored hydrated electrons (i.e., reduced O2) (see Materials and Methods).Afterward, the oligonucleotides were digested to deoxyribonucleosides (see Materials andMethods) : (a) no irradiation with and without cisPt, (b) irradiation of the oligonucleotidewithout cisPt, and (c) with cisPt with different doses.
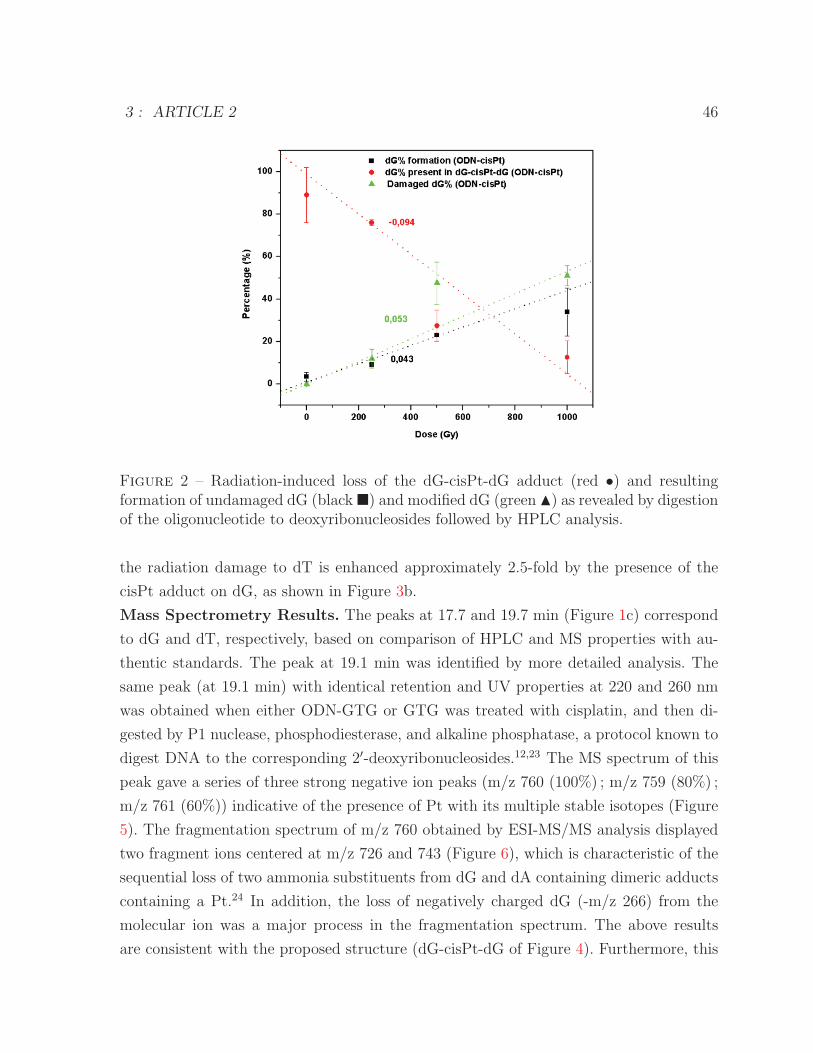
3 : ARTICLE 2 46
Figure 2 – Radiation-induced loss of the dG-cisPt-dG adduct (red •) and resultingformation of undamaged dG (black�) and modified dG (green �) as revealed by digestionof the oligonucleotide to deoxyribonucleosides followed by HPLC analysis.
the radiation damage to dT is enhanced approximately 2.5-fold by the presence of the
cisPt adduct on dG, as shown in Figure 3b.
Mass Spectrometry Results. The peaks at 17.7 and 19.7 min (Figure 1c) correspond
to dG and dT, respectively, based on comparison of HPLC and MS properties with au-
thentic standards. The peak at 19.1 min was identified by more detailed analysis. The
same peak (at 19.1 min) with identical retention and UV properties at 220 and 260 nm
was obtained when either ODN-GTG or GTG was treated with cisplatin, and then di-
gested by P1 nuclease, phosphodiesterase, and alkaline phosphatase, a protocol known to
digest DNA to the corresponding 2′-deoxyribonucleosides.12,23 The MS spectrum of this
peak gave a series of three strong negative ion peaks (m/z 760 (100%) ; m/z 759 (80%) ;
m/z 761 (60%)) indicative of the presence of Pt with its multiple stable isotopes (Figure
5). The fragmentation spectrum of m/z 760 obtained by ESI-MS/MS analysis displayed
two fragment ions centered at m/z 726 and 743 (Figure 6), which is characteristic of the
sequential loss of two ammonia substituents from dG and dA containing dimeric adducts
containing a Pt.24 In addition, the loss of negatively charged dG (-m/z 266) from the
molecular ion was a major process in the fragmentation spectrum. The above results
are consistent with the proposed structure (dG-cisPt-dG of Figure 4). Furthermore, this
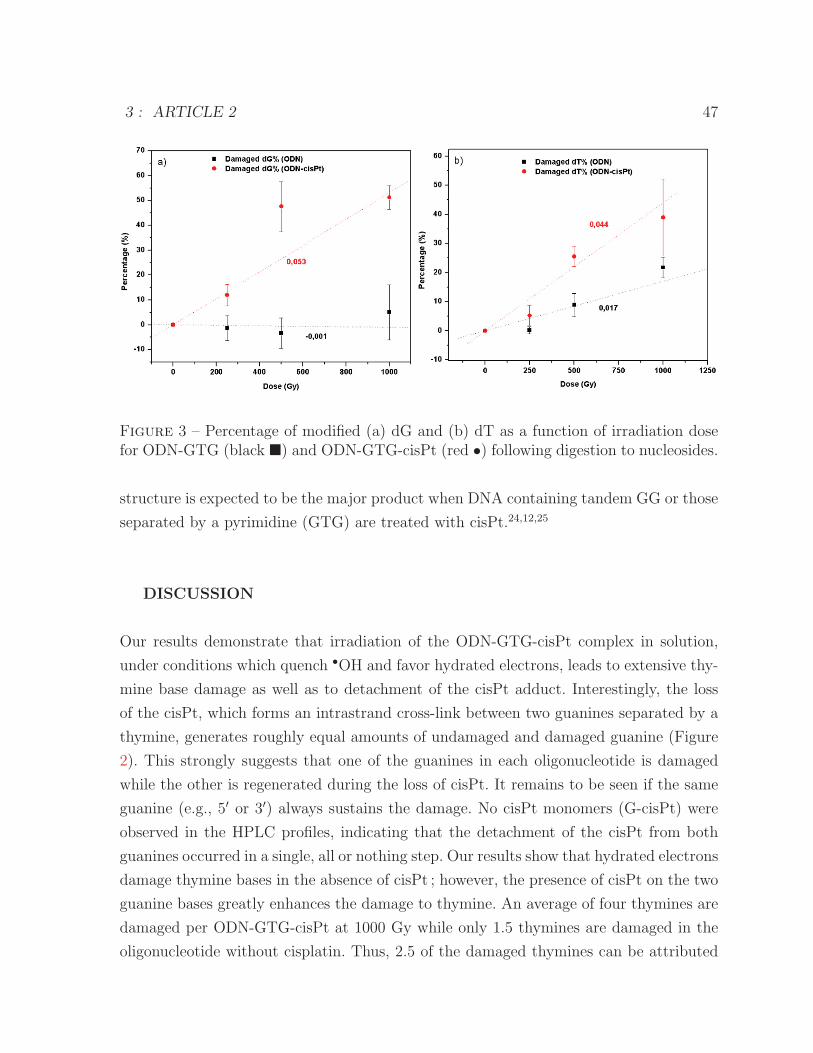
3 : ARTICLE 2 47
Figure 3 – Percentage of modified (a) dG and (b) dT as a function of irradiation dosefor ODN-GTG (black �) and ODN-GTG-cisPt (red •) following digestion to nucleosides.
structure is expected to be the major product when DNA containing tandem GG or those
separated by a pyrimidine (GTG) are treated with cisPt.24,12,25
DISCUSSION
Our results demonstrate that irradiation of the ODN-GTG-cisPt complex in solution,
under conditions which quench •OH and favor hydrated electrons, leads to extensive thy-
mine base damage as well as to detachment of the cisPt adduct. Interestingly, the loss
of the cisPt, which forms an intrastrand cross-link between two guanines separated by a
thymine, generates roughly equal amounts of undamaged and damaged guanine (Figure
2). This strongly suggests that one of the guanines in each oligonucleotide is damaged
while the other is regenerated during the loss of cisPt. It remains to be seen if the same
guanine (e.g., 5′ or 3′) always sustains the damage. No cisPt monomers (G-cisPt) were
observed in the HPLC profiles, indicating that the detachment of the cisPt from both
guanines occurred in a single, all or nothing step. Our results show that hydrated electrons
damage thymine bases in the absence of cisPt ; however, the presence of cisPt on the two
guanine bases greatly enhances the damage to thymine. An average of four thymines are
damaged per ODN-GTG-cisPt at 1000 Gy while only 1.5 thymines are damaged in the
oligonucleotide without cisplatin. Thus, 2.5 of the damaged thymines can be attributed
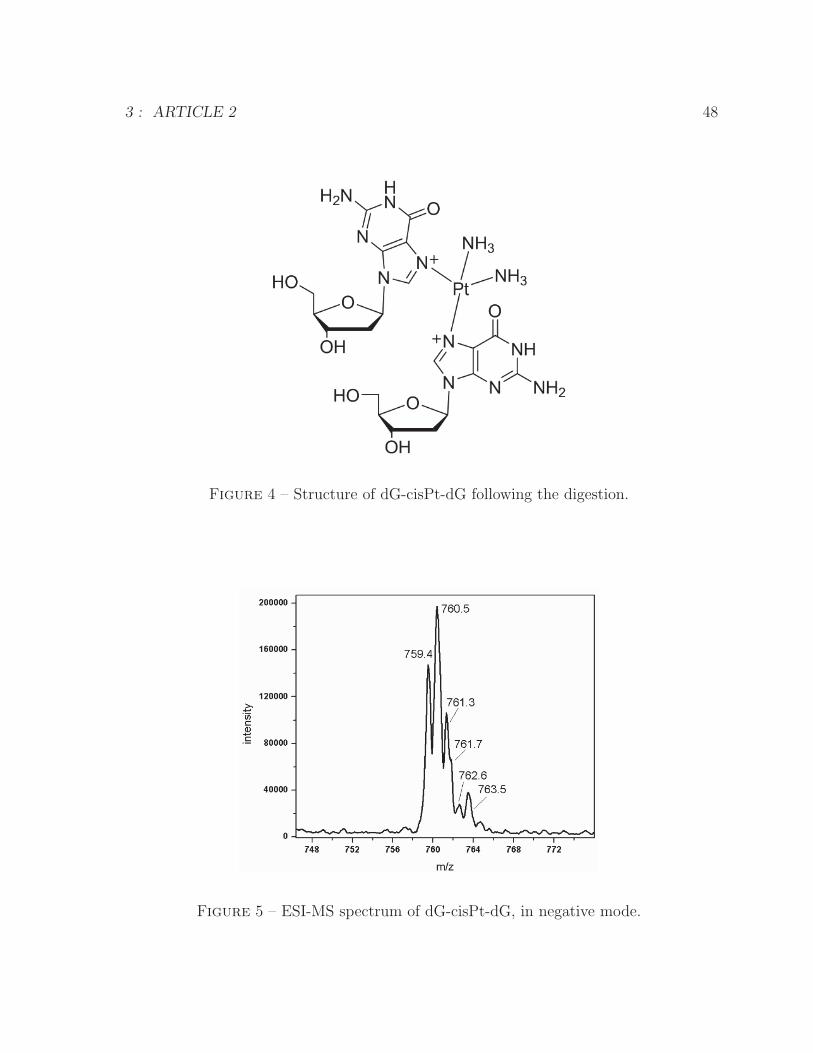
3 : ARTICLE 2 48
Figure 4 – Structure of dG-cisPt-dG following the digestion.
Figure 5 – ESI-MS spectrum of dG-cisPt-dG, in negative mode.
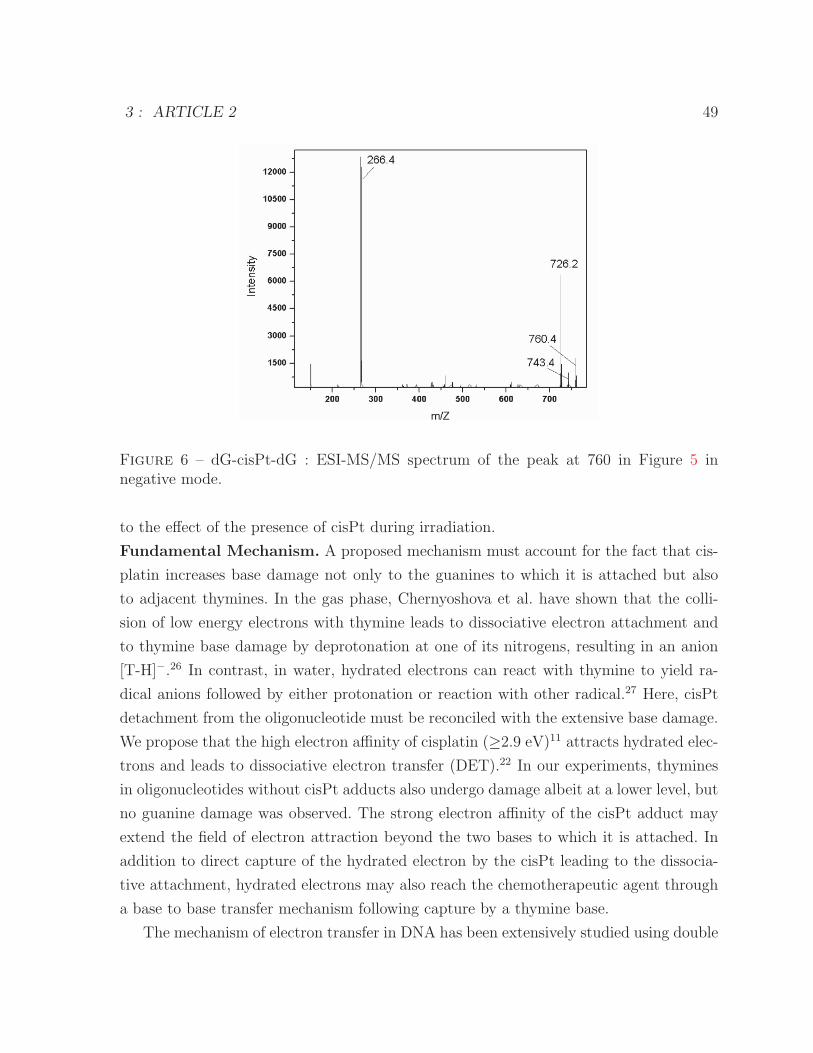
3 : ARTICLE 2 49
Figure 6 – dG-cisPt-dG : ESI-MS/MS spectrum of the peak at 760 in Figure 5 innegative mode.
to the effect of the presence of cisPt during irradiation.
Fundamental Mechanism. A proposed mechanism must account for the fact that cis-
platin increases base damage not only to the guanines to which it is attached but also
to adjacent thymines. In the gas phase, Chernyoshova et al. have shown that the colli-
sion of low energy electrons with thymine leads to dissociative electron attachment and
to thymine base damage by deprotonation at one of its nitrogens, resulting in an anion
[T-H]−.26 In contrast, in water, hydrated electrons can react with thymine to yield ra-
dical anions followed by either protonation or reaction with other radical.27 Here, cisPt
detachment from the oligonucleotide must be reconciled with the extensive base damage.
We propose that the high electron affinity of cisplatin (≥2.9 eV)11 attracts hydrated elec-
trons and leads to dissociative electron transfer (DET).22 In our experiments, thymines
in oligonucleotides without cisPt adducts also undergo damage albeit at a lower level, but
no guanine damage was observed. The strong electron affinity of the cisPt adduct may
extend the field of electron attraction beyond the two bases to which it is attached. In
addition to direct capture of the hydrated electron by the cisPt leading to the dissocia-
tive attachment, hydrated electrons may also reach the chemotherapeutic agent through
a base to base transfer mechanism following capture by a thymine base.
The mechanism of electron transfer in DNA has been extensively studied using double

3 : ARTICLE 2 50
stranded DNA by generating either positive hole transfer or excess electron transfer
between specific sites of DNA base sequences.28−34 In general, electron transfer depends
on the difference in energy and the distance between donor and acceptor molecules,
such that the efficiency of transfer decreases exponentially with distance. In the case
of DNA, several models have been proposed to account for electron transfer.31−33 A
consensus today is that transfer takes place mainly by a hopping process in which charge
is distributed on one or more DNA bases and transfer to other sites is strongly dependent
on base properties and sequence.33,34
Excess electron transfer is base sequence dependent, since some bases are more subject
to protonation (e.g., cytosine) than others (e.g., thymine).29,35 DNA concentration also
has an impact on electron migration distance, since high concentrations provide DNA
to DNA electron transfer.30 In contrast, the latter is independent of concentration when
DNA in water solution is cooled to form ice.36 Electron spin resonance (ESR) has been
used29,30,36 to probe excess electron transfer, following gamma ray irradiation from double
stranded DNA to the interspersed DNA intercalator mitoxantrone (MX), as a function of
temperature and concentration. It was concluded that tunneling operates mainly at low
temperature (<190 K), while, at higher temperatures, thermally activated hopping is the
prevailing process giving a faster migration rate. In contrast to double stranded DNA,
the electron transfer efficiency was only weakly affected by interbase distance in single
stranded DNA.28 To explain this phenomenon, the authors invoked the flexibility of the
single stranded oligonucleotide, which allows changes in conformation to decrease the
distance between the charge donor and acceptor and thus transfer by bypassing several
thymine bases.
In theory, a hydrated electron could be captured by a thymidine base and transferred
between thymidine bases. We suggest that the initial polaron (i.e., the hydrated electron)
interacts with any of the bases to transform into a base polaron. In other words, the
energy of the polarization well of the water around the electron is partly transferred into
structural deformation energy around a base.37 When the charge is thus localized on
one base, two channels may be become available : base damage by protonation at C6 or
migration to the next base by thermally assisted hopping.
Park et al. have shown the existence of hopping through consecutive thymines,38 and
that phenomenon could also explain electron transfer to cisPt. Otherwise, following the
cleavage of the bond between N7 of guanine and the Pt, two pathways are possible : the
bond will break and the extra electron will either go to N7 of guanine (path 1) or to Pt

3 : ARTICLE 2 51
(path2).The first pathway will lead to nonmodified guanine and a reactive cisPt radical.
The second will lead to a potentially reactive Pt(I) species that could react with guanine
and thymine to produce additional damage. It has been demonstrated by Richmond et al.
that Pt(I) reduce the hydroxyl radical adducts of thymine (•TOH).39 This phenomenon
could occur in hypoxic cells, where bases are damaged by hydrated electrons followed by
their reaction with Pt(I). The unexpected observation of damage to bases outside the Pt
attchment sites may provide an important key to understanding the effect of concomi-
tant radiotherapy and chemotherapy treatment, where the cisPt acts as a true catalyst
leading to multiple cycles of long-range DNA damage followed, independently, by detach-
ment of the cisPt adduct and damage to one of the guanine bases to which it was attached.
CONCLUSION
The presence of cisPt greatly increases the damage to DNA bases by hydrated elec-
trons produced by ionizing radiation. This damage is found at the Pt attachment sites
(guanines) but also at other sites (thymine). These results demonstrate the long-range in-
teraction of cisPt with hydrated electrons possibly owing to its high electron affinity. We
propose that cisPt, with its very high electron affinity, attracts electrons leading to DET
and loss of the cisPt. In addition, thymine bases can also serve as entry points for hydra-
ted electrons into the DNA, which subsequently undergo base to base electron hopping
to the cisPt. This electron transfer process has a certain probability of being interrupted
by either DET of the cisPt or protonation of the bases. Moreover, the reactivity of the
detached cisPt one electron reduced may create DNA damage and lead to cell death. The
reactions described here provide a mechanism to explain the radiosensitization of DNA
by cisPt adducts.
AUTHOR INFORMATION
Corresponding Author
*E-mail : [email protected]. Address : Universite de Sherbrooke, Faculte
de medecine, Departement de medecine nucleaire et de radiobiologie, 3001, 12e Avenue
Nord, Sherbrooke, QC, Canada, J1H 5N4. Phone : 1 819 820 6868. Fax : 1 819 564-5442.
Notes
The authors declare no competing financial interest.

3 : ARTICLE 2 52
ACKNOWLEDGMENTS
Financial support for this work was provided by the Canadian Institute of Health Re-
search (CIHR).
REFERENCES
(1) Samant, S. ; Kumar, P. ; Wan, J. ; Hanchett, C. ; Vieira, F. ; Murry, T. ; Wong, F. S.
H. ; Robbins, K. T. Concomitant Radiation Therapy and Targeted Cisplatin Chemothe-
rapy for the Treatment of Advanced Pyriform Sinus Carcinoma : Disease Control and
Preservation of Organ Function. Head Neck 1999, 21, 595-601.
(2) Boscolo-Rizzo, P. ; Gava, A. ; Marchiori, C. ; Baggio, V. ; da Mosto, M. C. Functional
Organ Preservation in Patients with Locoregionally Advanced Head and Neck Squa-
mous Cell Carcinoma Treated by Platinum-Based Multidrug Induction Chemotherapy
and Concurrent Chemoradiotherapy. Ann. Oncol. 2011, 22, 1894-1901.
(3) Peters, W. A., III ; Liu, P. Y. ; Barrett, R. J., II ; Stock, R. J. ; Monk, B. J. ; Berek, J.
S. ; Souhami, L. ; Grigsby, P. ; Gordon, W. ; Alberts, D. S. Concurrent Chemotherapy and
Pelvic Radiation Therapy Compared with Pelvic Radiation Therapy Alone as Adjuvant
Therapy after Radical Surgery in High-Risk Early-Stage Cancer of the Cervix. J. Clin.
Oncol. 2000, 18, 1606-1613.
(4) Candelaria, M. ; Garcia-Arias, A. ; Cetina, L. ; Duenas-Gonzalez, A. Radiosensitizers
in Cervical Cancer. Cisplatin and Beyond. Radiat. Oncol. 2006, 1, 1-17.
(5) Choy, H. ; Mac Rae, R. M. Chemoradiation in Cancer Therapy ; Humana Press : To-
towa, NJ, 2003.
(6) Kelland, L. The Resurgence of Platinum-Based Cancer Chemotherapy. Nat. Rev.
Cancer 2007, 7, 573-584.
(7) Go, R. S. ; Adjei, A. A. Review of the Comparative Pharmacology and Clinical Ac-
tivity of Cisplatin and Carboplatin. J. Clin. Oncol. 1999, 17, 409-422.
(8) Burger, A. M. ; Double, J. A. ; Newell, D. R. Inhibition of Telomerase Activity by
Cisplatin in Human Testicular Cancer Cells. Eur. J. Cancer 1997, 33, 638-644.
(9) Wozniak, K. ; Blasiak, J. Recognition and Repair of DNA-Cisplatin Adducts. Acta
Biochim. Polym. 2002, 49, 583-596.
(10) Kopyra, J. ; Koenig-Lehmann, C. ; Bald, I. ; Illenberger, E. A Single Slow Electron
Triggers the Loss of Both Chlorine Atoms from the Anticancer Drug Cisplatin : Implica-
tions for Chemoradiation Therapy. Angew. Chem., Int. Ed. 2009, 48, 7904-7907.
(11) Arumainayagam, C. R. ; Lee, H. L. ; Nelson, R. B. ; Haines, D. R. ; Gunawardane, R.

3 : ARTICLE 2 53
P. Low-Energy Electron-Induced Reactions in Condensed Matter. Surf. Sci. Rep. 2010,
65, 1-44.
(12) Williams, R. T. ; Nalbandian, J. N. ; Tu, A. ; Wang, Y. Nuclease Digestion and Mass
Spectrometric Characterization of Oligodeoxyribonucleotides Containing 1,2-GpG, 1,2-
ApG, and 1,3-GpXpG Cisplatin Intrastrand Cross-links. Clin. Chim. Acta 2013, 420,
160-170.
(13) For a review, see : Sanche, L. In Radiation Induced Molecular Phenomena in Nucleic
Acid ; Shukla, M. K., Leszczynski, J., Eds. ; Springer : Dordrecht, The Netherlands, 2008 ;
Chapter 19.
(14) LaVerne, J. A. ; Pimblott, M. Electron Energy-Loss Distributions in Solid, Dry DNA.
Radiat. Res. 1995, 141, 208-215.
(15) Cobut, V. ; Frongillo, Y. ; Patau, J. P. ; Goulet, T. ; Fraser, M.-J. ; Jay-Gerin, J.-P.
Monte Carlo Simulation of Fast Electron and Proton Tracks in Liquid Water-I. Physical
and Physicochemical Aspects. Radiat. Phys. Chem. 1998, 51, 229-243.
(16) Zheng, Y. ; Hunting, D. J. ; Ayotte, P. ; Sanche, L. Role of Secondary Low-Energy
Electrons in the Concomitant Chemoradiation Therapy of Cancer. Phys. Rev. Lett. 2008,
100, 198101-1-198101-4.
(17) Nikjoo, H. ; Bolton, C. E. ; Watanabe, R. ; Terrisol, M. ; O’Neill, P. ; Goodhead, D. T.
Modelling of DNA Damage Induced by Energetic Electrons (100 eV to 100 keV). Radiat.
Prot. Dosim. 2002, 99, 77-80.
(18) DeLara, C. M. ; Jenner, T. J. ; Townsend, K. M. S. ; Marsden, S. J. ; O’Neill, P. The
Effect of Dimethyl Sulfoxide on the Induction of DNA Double-Strand Breaks in V79-4
Mammalian Cells by Alpha Particles. Radiat. Res. 1995, 144, 43-49.
(19) Alizadeh, E. ; Sanche, L. Precursors of Solvated Electrons in Radiobiological Physics
and Chemistry. Chem. Rev. 2012, 112, 5578-5602.
(20) Kuipers, G. K. ; Lafleur, M. V. M. Characterization of DNA Damage Induced by
Gamma-Radiation-Derived Water radicals, Using DNA Repair Enzymes. Int. J. Radiat.
Biol. 1998, 74, 511-519.
(21) Kumar, A. ; Sevilla, M. D. Role of Excited States in Low-Energy Electron (LEE)
Induced Strand Breaks in DNA Model Systems : Influence of Aqueous Environment.
ChemPhysChem 2009, 10, 1426-30.
(22) Behmand, B. ; Cloutier, P. ; Girouard, S. ; Wagner, J. R. ; Sanche, L. ; Hunting, D. J.
Hydrated Electrons React with High Specificity with Cisplatin Bound to Single-Stranded
DNA. J. Phys. Chem. B 2013, 117, 15994-15999.

3 : ARTICLE 2 54
(23) Samson-Thibault, F. ; Madugundu, G. S. ; Gao, S. ; Cadet, J. ; Wagner, J. R. Profiling
Cytosine Oxidation in DNA by LC-MS/MS. Chem. Res. Toxicol. 2012, 25, 1902-1911.
(24) Iijima, H. ; Patrzyc, H. B. ; Dawidzik, J. B. ; Budzinski, E. E. ; Cheng, H. C. ; Freund,
H. G. ; Box, H. C. Measurement of DNA Adducts in Cells Exposed to Cisplatin. Anal.
Biochem. 2004, 333, 65-71.
(25) Jamieson, E. R. ; Lippard, S. J. Structure, Recognition, and Processing of Cisplatin-
DNA Adducts. Chem. Rev. 1999, 99, 2467-2498.
(26) Chernyshova, I. V. ; Kontros, J. E. ; Shpenik, O. B. ; Tamuliene, J. ; Balevicius, M.
L. Low-Energy Electron Collisions with Gas-Phase Thymine Molecule. Mater. Sci. 2011,
17, 322-325.
(27) Von Sonntag, C. The Chemical Basis of Radiation Biology ; Taylor and Francis :
London, 1987.
(28) Giese, B. Long-Distance Charge Transport in DNA : The Hopping Mechanism. Acc.
Chem. Res. 2000, 33, 631-636.
(29) Cai, Z. ; Li, X. ; Sevilla, M. D. Excess Electron Transfer in DNA : Effect of Base
Sequence and Proton Transfer. J. Phys. Chem. B 2002, 106, 2755-2762
(30) Cai, Z. ; Sevilla, M. D. Studies of Excess Electron and Hole Transfer in DNA at Low
Temperatures. Top. Curr. Chem. 2004, 237, 103-127.
(31) Ly, D. ; Sanii, L. ; Schuster, G. B. Mechanism of Charge Transport in DNA :
Internally-Linked Anthraquinone Conjugates Support Phonon-Assisted Polaron Hopping.
J. Am. Chem. Soc. 1999, 121, 9400-9410.
(32) Wan, C. ; Fiebig, T. ; Kelley, S. O. ; Treadway, C. R. ; Barton, J. K. ; Zewail, A.
H. Femtosecond Dynamics of DNA-Mediated Electron Transfer. Proc. Natl. Acad. Sci.
U.S.A. 1999, 96, 6014-6019.
(33) Schuster, G. B. Long-Range Charge Transfer in DNA : Transient Structural Distor-
tions Control the Distance Dependence. Acc. Chem. Res. 2000, 33, 253-260.
(34) Jortner, J. ; Bixon, M. ; Langenbacher, T. ; Michel-Beyerle, M. Charge Transfer and
Transport in DNA. Proc. Natl. Acad. Sci. U.S.A. 1998, 95, 12759-12765.
(35) Ito, T. ; Rokita, S. E. Criteria for Efficient Transport of Excess Electrons in DNA.
Angew. Chem. 2004, 116, 1875-1878.
(36) Cai, Z. ; Sevilla, M. D. Electron Spin Resonance Study of Electron Transfer in DNA :
Inter-Double-Strand Tunneling Processes. J. Phys. Chem. B 2000, 104, 6942-6949.
(37) Rezaee, M. ; Sanche, L. ; Hunting, D. J. Cisplatin Enhances the Formation of DNA
Single- and Double-Strand Breaks by Hydrated Electrons and Hydroxyl Radicals. Radiat.

3 : ARTICLE 2 55
Res. 2013, 179, 323-331.
(38) Park, M. J. ; Fujitsuka, M. ; Kawai, K. ; Majima, T. Direct Measurement of the Dy-
namics of Excess Electron Transfer through Consecutive Thymine Sequence in DNA. J.
Am. Chem. Soc. 2011, 133, 15320-15323.
(39) Richmond, R. C. ; Simic, M. G. Effect of Radiation on Cis-Dichlorodiammineplatinum
(II) and DNA in Aqueous Solution. Br. J. Cancer 1978, 37 (Suppl. III), 20-23.

ARTICLE 3
Radiosentization of DNA by Cisplatin AdductsResults from an Increase in the Rate Constant forthe Reaction with Hydrated Electrons andFormation of PtI
B. Behmand, J.-L. Marignier, M. Mostafavi, J. R. Wagner, D. J. Hunting, and L. Sanche
J. Phys. Chem. B, 2015 (DOI : 10.1021/acs.jpcb.5b01752)
Avant-propos : J’ai fait les purifications et la preparation des echantillons. Les ex-
periences ont ete realisees par Jean-Louis Mariginer avec ma participation. J’ai analyse
les donnees et j’ai redige la totalite de l’article sous la supervision de Mehran Mostafavi,
Darel Hunting, Richard Wagner et Leon Sanche.
Resume : Dans cet article, la constante de vitesse de reaction des e−hyd avec oligonucleo-
tide (GTG) lie a l’adduit de cisplatine a ete mesuree par la technique de radiolyse pulsee.
Les resultats montrent que la reaction des e−hyd avec le complexe GTG-cisplatine est 3
fois plus rapide qu’avec GTG sans cisplatine. De plus, pour la premiere fois la formation
de l’intermediaire de PtI a ete mesuree du a l’interaction des e−hyd avec le complexe de
l’adduit de cisplatine-ADN.
56

4 : ARTICLE 3 57
ABSTRACT : Pulse radiolysis measurements of the decay of hydrated electrons
in solutions containing different concentrations of the oligonucleotide GTG with and
without a cisplatin adduct show that the presence of a cisplatin moiety accelerates the
reaction between hydrated electrons and the oligonucleotide. The rate constant of the
reaction is found to be 2.23 × 1010 mol−1 L s−1, which indicates that it is diffusion
controlled. In addition, we show for the first time the formation of a PtI intermediate as
a result of the reaction of hydrated electrons with GTG-cisplatin. A putative reaction
mechanism is proposed, which may form the basis of the radiosensitization of cancer cells
in concomitant chemoradiation therapy with cisplatin.

4 : ARTICLE 3 58
INTRODUCTION
Clinical studies have shown that concomitant chemotherapy and radiotherapy increases
tumor cell killing and patient survival.1−3 Cis-diaminedichloroplatinum(II) (cisplatin) is
a chemotherapy drug used for several types of cancer : lung, testicular, ovary, neck, and
head.4−6 Cisplatin enters cells both by passive diffusion7 and by active transporters.8 Fol-
lowing hydrolysis, activated cisplatin reacts with several cellular components : proteins,
RNA, DNA, membrane phospholipids, and actin filaments,9 but DNA is the most impor-
tant target for cell killing.9,10 Cisplatin adducts (cisPt) specifically bind to the N7 site
of the purine bases, particularly guanine, and distort the DNA conformation by bending
and unwinding the helix.4,9,11 Inhibition of DNA repair and/or increased DNA damage
when cisPt is bound to DNA have been proposed as two possible mechanisms responsible
for radiosensitization by cisPt leading to cancer cell death.9,12−14 However, the precise
mechanisms by which cisplatin adduct increase DNA damage by radiation have not been
elucidated.
Ionizing radiation induces DNA damage by both the direct effect, in which energy
is deposited within the DNA molecule, or by the indirect effect in which molecules sur-
rounding DNA, particularly water, absorb the radiation and generate reactive species,
especially hydroxyl radicals which subsequently induce DNA damage.15,16 Since the cell
is composed of 70% water, it is estimated that at least 50% of total damage to DNA by
γ radiation is due to the indirect effect.17 The reaction of ionizing radiation with water
induces the reactive chemical species •OH, H•, and e−aq that in turn can induce DNA
damage.17 Among these radicals, e−aq does not induce DNA strand breaks.18 However,
Rezaee et al. have shown that e−aq can induce single and double strand breaks in DNA
modified by cisplatin.19 Behmand et al. have proposed that the high electron affinity of Pt
in the oligonucleotide attracts e−aq to the complex leading to dissociative electron transfer
(DET) and detachment of cisPt.20 Richmond et al. have measured the rate constant of
the reaction of e−aq with cisplatin (k = 1.8 × 1010 mol−1 L s−1) and have shown that
the reactive PtI intermediate thus generated reacts with the hydroxyl radical adduct of
thymine (k = 1.2 × 109 mol−1 L s−1).21 It has been demonstrated that cisPt increases
DNA base damage by e−aq not only at the cisPt attachment site (guanine), but also at
other sites.22 Schusser et al. have shown that the rate constant for the reaction of e−aq with
DNA depends on the DNA structure and ionic conditions.23 They observed the same rate
constant over the entire pH-region from 5.0 to 9.0 for double-stranded DNA from calf

4 : ARTICLE 3 59
thymus and thus concluded that the DNA structure is not altered in this pH interval.
Moreover, the interaction of ionizing radiation with DNA produces DNA radicals
(•DNA). The presence of oxygen results in ”fixation” that modifies DNA• generating a
peroxy radical (•DNA-OO),24,25 which leads to DNA lesions. Therefore, hypoxic cancer
cells are more radioresistant. However, oxygen reacts with e−aq and thus the hypoxic
regions of tumor may be susceptible to radiosensitizers, such as DNA-platinum adducts
which react with e−aq. Understanding the mechanism of the reaction of e−aq with the DNA-
cisplatin complex is important to improve the treatment of cancer.
In the present work, we synthesize GTG-cisPt and the rate constant of the reaction
between e−aq and this oligonucleotide, with and without a cisPt adduct, is determined by
pulse radiolysis. In addition, we measure the formation of PtI spectroscopically following
the reaction of e−aq with GTG-cisPt.
MATERIALS AND METHODS
Oligonucleotide-cisPt Complex Preparation. The single-stranded oligonucleotide
(GTG) was purchased from Alpha DNA, Montreal, QC, Canada. Cisplatin was first dis-
solved in water at 4 mmol L−1. Then, the oligonucleotide was mixed with the cisPt
solution (0.4 mmol L−1 GTG) to give a ratio of cisPt/GTG of 10 :1. The solution was
kept at room temperature for 24 h. The product was purified by high performance liquid
chromatography (HPLC) to separate GTG-cisPt from GTG. A column (5 μm ODS A
250×6 mm ; YMC) and a linear gradient (0-15%) of acetonitrile in amonium acetate were
used.
Pulse Radiolysis.We used the transient absorption setup with a streak camera installed
in experimental area EA-3 of the electron pulse facility ELYSE at Paris-Sud University.
The e-beam irradiation was performed using the picosecond pulse radiolysis facility de-
livering a repetition rate of 5 Hz, electron pulses with an energy of 8 MeV. The dose
deposited per pulse (10 Gy) in the samples was deduced from the measurements of the
transient absorbance of e−aq in water and verified just before each series of experiments.
Typically, the electron pulse has a half width (FHWM) of 7 ps and a charge of ∼1 nC.
More details of the system configuration are described elsewhere.26,27 An optical quartz
cell with an optical path length of 1 cm containing the sample is placed as close as possible
to the output window of electron beam to minimize divergence. During the experiment,
the sample is bubbled with argon gas to remove the oxygen, which is a scavenger of

4 : ARTICLE 3 60
e−aq. Furthermore, isopropanol (0.2 mol L−1) or tert-butanol (0.25 mol L−1) was added to
quench the •OH and H• radicals.
RESULTS
The rate constant for the reaction, between, e−aq and the oligonucleotide-cisPt complex was
determined by pulse radiolysis. The solutions (hydrolyzed cisplatin [Pt(NH3)2(OH2)2]2+,
GTG, and GTG-cisPt) were each irradiated with a pulse of 1.4 nC. Approximately 10
transient spectra were recorded and averaged for each solution. The decay of e−aq was
measured at 600 nm for each solution as shown in Figure 1a,b,c, respectively.
Given the very low concentration of the e−aq (3.1×10−6 mol L−1 at 3 ns) compared to
the concentration of the hydrolyzed cisplatin, GTG, and GTG-cisPt, the rate constants
of the reactions were determined from a pseudo-first-order approximation. The observed
rate constant, kobs, is obtained from the e−aq decay on a logarithmic scale of each solution
at different concentrations : hydrolyzed cisplatin, GTG, and GTG-cisPt (Figure 1a,b,c).
Finally, using a linear fit of kobs, the rate constant for each reaction is obtained (Figure
2). The rate constant for the oligonucleotide with a cisPt adducts (2.2×1010 mol−1 L s−1)
is very similar to that for hydrolyzed cisplatin (3.2 ×1010 mol−1 L s−1). However, the
rate constant for the reaction of e−aq with the oligonucleotide-cisPt is 3-fold higher than
the one for the oligonucleotide without cisPt (Table 1).
Pulse radiolysis measurements of a solution containing 2.2×10−4 mol L−1 GTG-cisPt
and 0.25 mol L−1 tert-butanol were performed by observing the entire transient absorp-
tion spectra from 300 to 600 nm. The tert-butanol radical is expected to be present in
solution following the reaction of tert-butanol with •OH and to a lesser extend with H•
atom.29 Due to its weak extinction coefficient in the spectral range under investigation,
this radical has a negligible signature in our transient spectra. Moreover, this radical
is not a reducing species, and thus is not involved in the reduction of the GTG-cisPt
complex. Under these conditions, e−aq, which can react with GTG-cisPt, and the reduced
form of the GTG-cisPt complex are likely the only absorbing species. Figure 3 reports
the kinetics at three selected wavelengths 600, 380, and 310 nm. The decay at 600 nm is
due to the loss of e−aq which has completely decayed after about 700 ns. It is interesting to
note that 380 nm constitutes an isosbestic point showing that the extinction coefficient
of e−aq and the reduced species are the same (ελ=380nm= 2.1×103 mol−1 L cm−1) at this
wavelength. The extinction coefficient of the reduced species at 310 nm is larger than that
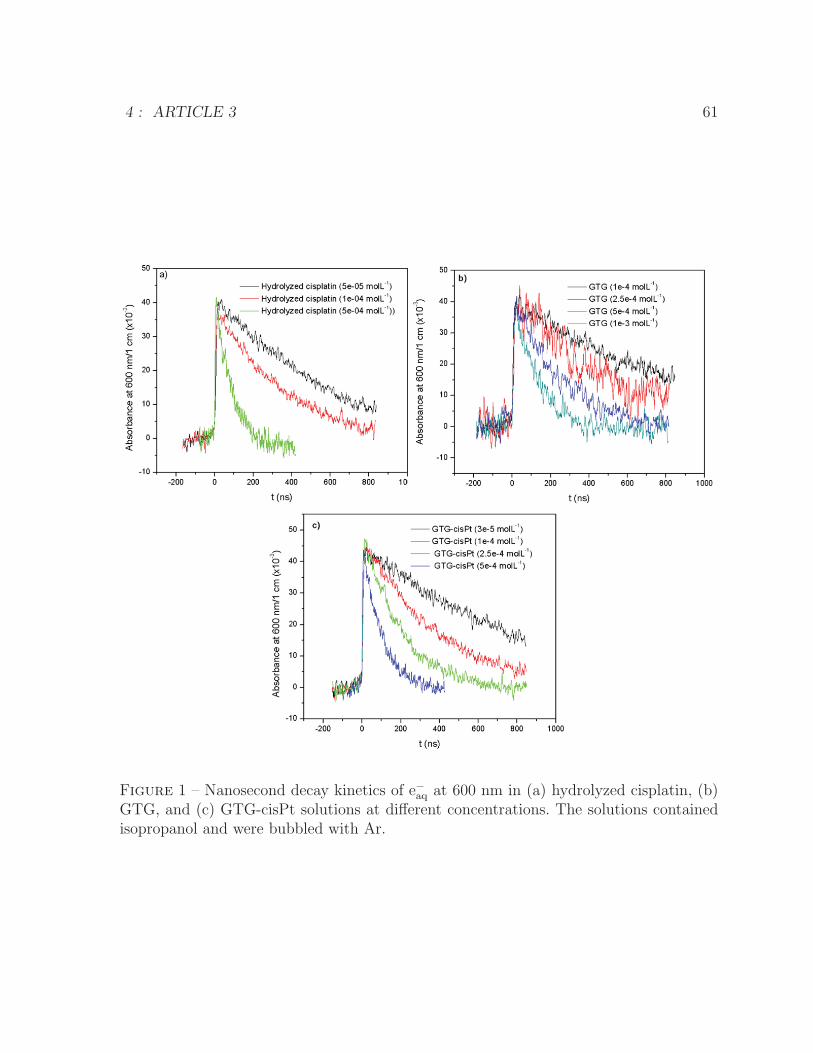
4 : ARTICLE 3 61
Figure 1 – Nanosecond decay kinetics of e−aq at 600 nm in (a) hydrolyzed cisplatin, (b)GTG, and (c) GTG-cisPt solutions at different concentrations. The solutions containedisopropanol and were bubbled with Ar.
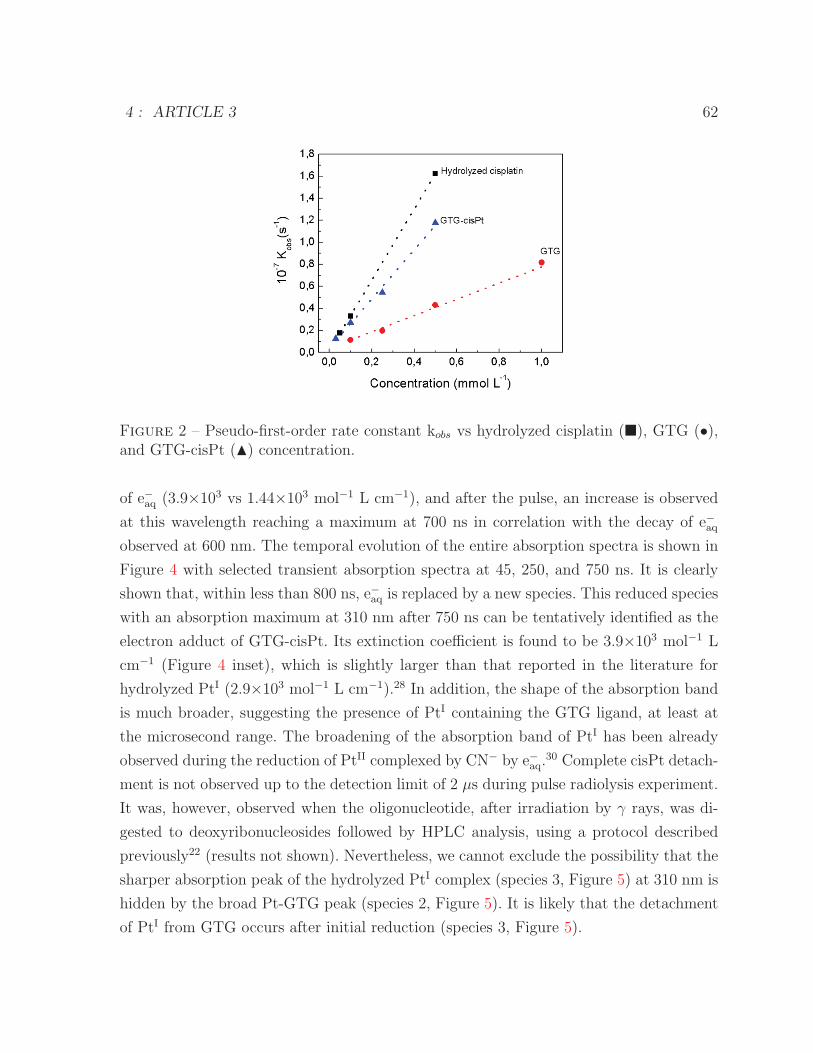
4 : ARTICLE 3 62
Figure 2 – Pseudo-first-order rate constant kobs vs hydrolyzed cisplatin (�), GTG (•),and GTG-cisPt (�) concentration.
of e−aq (3.9×103 vs 1.44×103 mol−1 L cm−1), and after the pulse, an increase is observed
at this wavelength reaching a maximum at 700 ns in correlation with the decay of e−aqobserved at 600 nm. The temporal evolution of the entire absorption spectra is shown in
Figure 4 with selected transient absorption spectra at 45, 250, and 750 ns. It is clearly
shown that, within less than 800 ns, e−aq is replaced by a new species. This reduced species
with an absorption maximum at 310 nm after 750 ns can be tentatively identified as the
electron adduct of GTG-cisPt. Its extinction coefficient is found to be 3.9×103 mol−1 L
cm−1 (Figure 4 inset), which is slightly larger than that reported in the literature for
hydrolyzed PtI (2.9×103 mol−1 L cm−1).28 In addition, the shape of the absorption band
is much broader, suggesting the presence of PtI containing the GTG ligand, at least at
the microsecond range. The broadening of the absorption band of PtI has been already
observed during the reduction of PtII complexed by CN− by e−aq.30 Complete cisPt detach-
ment is not observed up to the detection limit of 2 μs during pulse radiolysis experiment.
It was, however, observed when the oligonucleotide, after irradiation by γ rays, was di-
gested to deoxyribonucleosides followed by HPLC analysis, using a protocol described
previously22 (results not shown). Nevertheless, we cannot exclude the possibility that the
sharper absorption peak of the hydrolyzed PtI complex (species 3, Figure 5) at 310 nm is
hidden by the broad Pt-GTG peak (species 2, Figure 5). It is likely that the detachment
of PtI from GTG occurs after initial reduction (species 3, Figure 5).
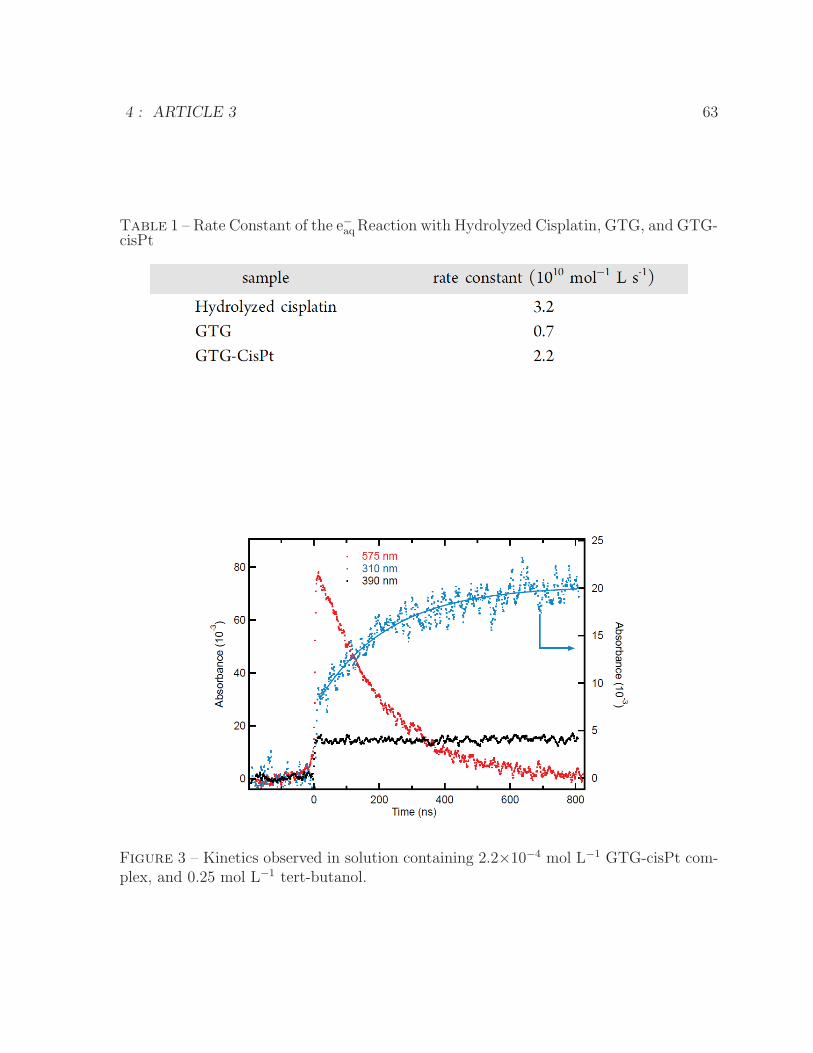
4 : ARTICLE 3 63
Table 1 – Rate Constant of the e−aqReaction with Hydrolyzed Cisplatin, GTG, and GTG-cisPt
Figure 3 – Kinetics observed in solution containing 2.2×10−4 mol L−1 GTG-cisPt com-plex, and 0.25 mol L−1 tert-butanol.
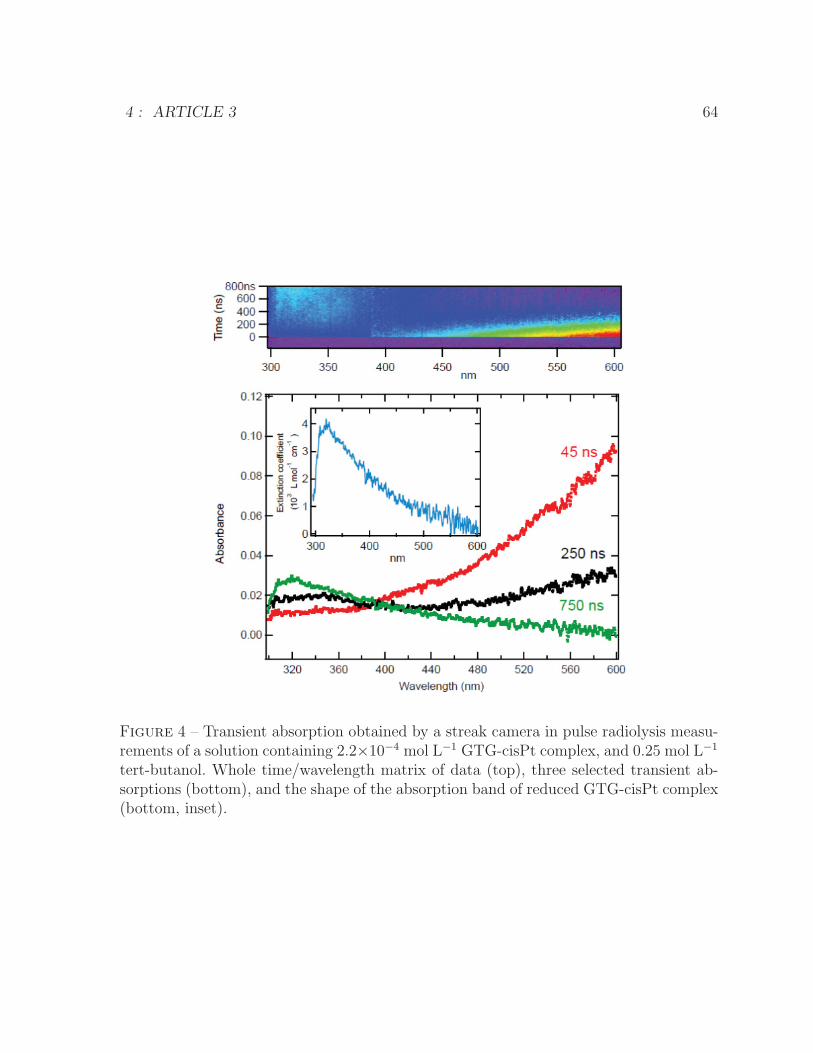
4 : ARTICLE 3 64
Figure 4 – Transient absorption obtained by a streak camera in pulse radiolysis measu-rements of a solution containing 2.2×10−4 mol L−1 GTG-cisPt complex, and 0.25 mol L−1
tert-butanol. Whole time/wavelength matrix of data (top), three selected transient ab-sorptions (bottom), and the shape of the absorption band of reduced GTG-cisPt complex(bottom, inset).

4 : ARTICLE 3 65
Figure 5 – Proposed mechanism of d(GpTpG) release from a GTG-cisPt complex.
DISCUSSION
The radiosensitization of DNA to e−aq by cisplatin adducts has recently been reported,19,22
but the underlying mechanism is not clear. The purpose of this study was to determine
the rate constants for the reaction of e−aq with all components of the system : cispla-
tin, the oligonucleotide, and the oligonucleotide containing a single cisplatin adduct. As
shown in Figure 2, the increase in the decay of e−aq as a function of the substrate concen-
trations indicates that e−aq reacts with all three components listed above. Hydrolyzed
cisplatin alone is the most reactive and the rate constant we determined agrees with
that reported by Richmond et al.21 The oligonucleotide-cisPt was slightly less reactive
than cisplatin alone (∼30%). This latter could be due to screening of the electron-cisPt
potential by the oligonucleotide, thus decreasing the probability of electron attachment
to cisPt. The oligonucleotide-cisPt complex is 3-fold more reactive than the oligonucleo-
tide alone, which is an important enhancement considering that only one cisPt adduct is
present per oligonucleotide. This indicates that the radiosensitization effect of a cispla-
tin adduct may result from its high electron affinity, which promotes electron uptake by
the oligonucleotide-cisPt complex leading to detachment of cisPt. On the basis of HPLC
analysis, the reduction of PtII by e−aq leads to the loss of both G ligands from GTG-cisPt

4 : ARTICLE 3 66
complexes. This may occur through exchange of the ligands with water upon reduction of
PtII to PtI. Although this has been demonstrated for certain Pt complexes, the stability
and propensity for ligand exchange of PtI under our conditions is not known.31 Alterna-
tively, PtI complexes may undergo a redox reaction with the ligand thereby releasing G
radicals. The latter species may undergo further reactions with H2O or radicals in the
reaction mixture.32 Figure 5 shows our proposed mechanism of d(GpTpG) release from a
GTG-cisPt complex following reaction with a single e−aq, by two-step process. Adam et al.
have shown using transient absorption measurement at 310 nm that PtII can be reduced
by e−aq and also by H• radicals (k=1.2×1010 mol−1 L s−1).28
In our experiment the reactive intermediate GTG-PtI may create supplementary da-
mage. The kinetics of appearance of the reduced species and the loss of e−aq at the isosbestic
point at 380 nm, where the extinction coefficient of e−aq and the reduced species are the
same, indicate that one PtI is formed per e−aq lost. The eventual loss of Pt from the tri-
nucleotide is consistent with our previous observation using a 11-mer containing a single
cisplatin adduct, as determined by digestion and HPLC analysis22 and with our results
following irradiation, digestion of the GTG-cisPt in this study (results not shown), and
HPLC analysis. The high electron affinity of Pt attached to the oligonucleotide leads to
Pt detachment and formation of PtI. The presence of polar groups such as NH3 may
also induce modification of the oligonucleotide structure that may favor e−aq attachment
to DNA.19 Moreover, capture of e−aq by the base also creates base damage, such as 5,6-
dihydrothymine, which is much more important in DNA-cisPt complexes than in DNA
without cisPt.22
CONCLUSION
The present pulse radiolysis experiments, which observed the entire absorption spec-
tra from 300 to 600 nm, have clearly demonstrated that the trinucleotide-cisPt complex
is 3-fold more reactive (k = 2.2×1010 mol−1 L s−1) than the trinucleotide without cisPt
(k = 7.4×109 mol−1 L s−1), probably due to the high electron affinity of cisplatin. The
shape of the absorption band of the reduced complex is broad indicating that PtI is
attached to the GTG ligand, at least in the microsecond range. Nevertheless, when the
irradiated oligonucleotide was digested to deoxyribonucleosides followed by HPLC ana-
lysis, the detachment of cisPt by e−aq was observed. These results are consistent with the
previous observation by HPLC and gel sequencing analysis of radiolysis products from

4 : ARTICLE 3 67
longer 11-mer oligonucleotides ;20,22 as well the observation of the strong enhancement
of single and double strand break formation due to e−aq in γ-irradiated plasmid DNA-
cisplatin complexes.19 Taken together, these results strongly suggest that, in cells, the
radiosensitization of DNA by cisplatin adducts probably results, at least in part, from
the increase in the rate constant of the reaction of e−aq with DNA and formation PtI.
AUTHOR INFORMATION
Corresponding Author
*E-mail : [email protected]. Phone : 1 819 820 6868. Fax : 1 819 564-
5442.
Notes
The authors declare no competing financial interest
ACKNOWLEDGMENTS
We would like to thank technical staff of ELYSE accelerator : Jean-Philippe Larbre,
Pierre Jeunesse, Audrey Gayral, Mireille Benoit, and Alexandre Demarque. Financial
support for this work was provided by the Canadian Institute of Health Research (CIHR).
REFERENCES
(1) Boscolo-Rizzo, P. ; Gava, A. ; Marchiori, C. ; Baggio, V. ; da Mosto, M. C. Functional
organ preservation in patients with locoregionally advanced head and neck squamous cell
carcinoma treated by platinum-based multidrug induction chemotherapy and concurrent
chemo-radiotherapy. Ann. Oncol. 2011, 22, 1894-1901.
(2) Samant, S. ; Kumar, P. ; Wan, J. ; Hanchett, C. ; Vieira, F. ; Murry, T. ; Wong, F. S.H. ;
Robbins, K. T. Concomitant radiation therapy and targeted cisplatin chemotherapy for
the treatment of advanced pyriform sinus carcinoma : Disease control and preservation
of organ function. Head Neck 1999, 21, 595-601.
(3) Peters, W. A., III ; Liu, P. Y. ; Barrett, R. J., II ; Stock, R. J. ; Monk, B. J. ; Berek,
J. S. ; Souhami, L. ; Grigsby, P. ; Gordon, W. ; Alberts, D. S. Concurrent chemotherapy
and pelvic radiation therapy compared with pelvic radiation therapy alone as adjuvant
therapy after radical surgery in high-risk early-stage cancer of the cervix. J. Clin. Oncol.

4 : ARTICLE 3 68
2000, 18, 1606-1613.
(4) Kelland, L. The Resurgence of Platinum-Based Cancer Chemotherapy. Nat. Rev.
Cancer. 2007, 7, 573-584.
(5) Go, R. S. ; Adjei, A. A. Review of the Comparative Pharmacology and Clinical Ac-
tivity of Cisplatin and Carboplatin. J. Clin. Oncol. 1999, 17, 409-422.
(6) Burger, A. M. ; Double, J. A. ; Newell, D. R. Inhibition of Telomerase Activity by
Cisplatin in Human Testicular Cancer Cells. Eur. J. Cancer. 1997, 33, 638-644.
(7) Andrews, P. A. ; Mann, S. C. ; Velury, S. ; Howell, S. B. In Platinum and other me-
tal coordination compounds in cancer chemotherapy ; Nicolini, M., Ed. ; MartinusNijhoff
Publishing : Boston, 1988 ; pp 248-254.
(8) Gately, D. P. ; Howell, S. B. Cellular Accumulation of the Anticance Agent Cisplatin :
A review. Br. J. Cancer 1993, 67, 1171-1176.
(9) Jamieson, E. R. ; Lippard, S. J. Structure, recognition, and processing of cisplatin-
DNA adducts. Chem. Rev. 1999, 99, 2467-2498.
(10) Akaboshi, M. ; Kawai, K. ; Maki, H. ; Akuta, K. ; Ujeno, Y. The Number of Plati-
num Atoms Binding to DNA, RNA and Protein Molecules of HeLa Cells Treated with
Cisplatin at its Mean Lethal Concentration. Jpn. J. Cancer Res. 1992, 83, 522-526.
(11) Wozniak, K. ; Blasiak, J. Recognition and repair of DNA-cisplatin adducts. Acta.
Biochim. Polym. 2002, 49, 583-596.
(12) Zimbrick, J. D. ; Sukrochana, A. ; Richmond, R. C. Studies on Radiosensitization
of Escherichia coli Cells by Cis-Platinum Complexes. Int. J. Radiat. Oncol. Biol. Phys.
1979, 5, 1351-1354.
(13) Lelieveld, P. ; Scoles, M. A. ; Brown, J. M. ; Phil, D. ; Kallman, R. F. The Effect
of Treatment in Fractionated Schedules with the Combination of X-Irradiation and Six
Cytotoxic Drugs on the RIF-1 Tumor and Normal Mouse Skin. Int. J. Radiat. Oncol.
Biol. Phys. 1985, 11, 111-121.
(14) Dewit, L. Combined Treatment of Radiation and Cis- Diamminedichloroplatinum
(II) : A Review of Experimental and Clinical Data. Oncol. Biol. Phys. 1987, 13, 403-426.
(15) O’Neill, P. ; Wardman, P. Radiation chemistry comes before radiation biology. Int.
J. Radiat. Biol. 2009, 85, 9-25.
(16) Goodhead, D. T. ; Thacker, J. ; Cox, R. Effects of radiations of different qualities on
cells : Molecular mechanisms of damage and repair. Int. J. Radiat. Biol. 1993, 63, 543-56.
(17) deLara, C. M. ; Jenner, T. J. ; Marsden, S. J. ; O’Neill, P. The Effect of Dimethyl
Sulfoxide on the Induction of DNA Double-Strand Breaks in V79-4 Mammalian Cells by

4 : ARTICLE 3 69
Alpha Particles. Radiat. Res. 1995, 144, 43-49.
(18) Kuipers, G. K. ; Lafleur, M. V. M. Characterization of DNA damage induced by
gamma-radiation-derived water radicals, using DNA repair enzymes. Int. J. Radiat. Biol.
1998, 74, 511-9.
(19) Rezaee, M. ; Sanche, L. ; Hunting, D. J. Cisplatin Enhances the Formation of DNA
Single- and Double-Strand Breaks by Hydrated Electrons and Hydroxyl Radicals. Radiat.
Res. 2013, 179, 323-331.
(20) Behmand, B. ; Cloutier, P. ; Girouard, S. ; Wagner, J. R. ; Sanche, L. ; Hunting, D. J.
Hydrated Electrons React with High Specificity with Cisplatin Bound to Single-Stranded
DNA. J. Phys. Chem. B 2013, 117, 15994-15999.
(21) Richmond, R. C. ; Simic, M. G. Effect of Radiation on Cisdichlorodiammineplatinum
(II) and DNA in Aqueous Solution. Br. J. Cancer 1978, 37 (Suppl. III), 20-23.
(22) Behmand, B. ; Wagner, J. R. ; Sanche, L. ; Hunting, D. J. Cisplatin Intrastrand Ad-
ducts Sensitize DNA to Base Damage by Hydrated Electrons. J. Phys. Chem. B 2014,
118, 4803-4808.
(23) Schusser, H. ; Navaratnam, S. ; Distel, L. Rate Constants for the Reactions of DNA
with Hydrated Electrons and with OH-Radicals. Radiat. Phys. Chem. 2005, 73, 163-168.
(24) Ferradini, C. ; Jay-Gerin, J. P. La radiolyse de l’eau et des solutions aqueuses : his-
torique et actualite. Can. J. Chem. 1999, 77, 1542-1575.
(25) Bertout, J. A. ; Patel, S. A. ; Simon, C. The impact of O2 availability on human
cancer. Nat. Rev. Cancer 2008, 8, 967-975.
(26) Belloni, J. ; Monard, H. ; Larbre, J.-P. ; Demarque, A. ; De Waele, V. ; Lampre, I. ;
Marignier, J.-L. ; Mostafavi, M. ; Bourdon, J. C. ; Bernard, M. ; Borie, H. ; Garvey, T. ;
Jacquemard, B. ; Leblond, B. ; Lepercq, P. ; Omeich, M. ; Roch, M. ; Rodier, J. ; Roux, R.
ELYSE-A Picosecond Electron Accelerator for Pulse Radiolysis Research. Nucl. Instrum.
Methods Phys. Res., Sect. A 2005, 539, 527-539.
(27) Marignier, J.-L. ; de Waele, V. ; Monard, H. ; Gobert, F. ; Larbre, J.-P. ; Demarque,
A. ; Mostafavi, M. ; Belloni, J. Time-resolved Spectroscopy at the Picosecond Laser-
triggered Electron Accelerator ELYSE. Radiat. Phys. Chem. 2006, 75, 1024-1033.
(28) Adams, G. E. ; Broszkiewicz, R. B. ; Michael, B. D. Pulse Radiolysis Studies on
Stable and Transient Complexes of Platinum. Trans. Faraday Soc. 1968, 64, 1256-1264.
(29) Spotheim-Maurizot, M. ; Mostafavi, M. ; Douki, T. ; Belloni, J. Radiation Chemistry
from Basics to Applications in Material and Life Sciences ; EDP Sciences : France, 2008.
(30) Ghosh-Mazumdar, A. S. ; Hart, E. J. Electron Pulse Radiolysis of Aqueous Tetra-

4 : ARTICLE 3 70
chloro and Tetracyano Complexes of PtII. Int. J. Radiat. Phys. Chem. 1969, 1, 165-176.
(31) Waltz, W. L. ; Lilie, J. ; Chandrasekhar, S. ; Woo, D. ; Brown, K. Study of the
Reactions of Platinum Macrocyclic Complex Ions with Free Radicals Formed by Pulse
Radiolysis of Aqueous Media. Inorg.
(32) D’Andelantonio, M. ; Russo, M. ; Kaloudis, P. ; Mulazzani, Q. G. ; Wardman, P. ;
Guerra, M. ; Chatgilialglu, Ch. Reaction of Hydrated Electrons with Guanine Deriva-
tives : Tautomerism of Intermediate Species. J. Phys. Chem. B 2009, 113, 2170-2176.

DISCUSSION
Dans les cellules cancereuses non hypoxiques, l’oxygene agit comme un agent radio-
sensibilisateur en se fixant aux dommages de l’ADN induits par la radiation, inhibant
ainsi les mecanismes de reparation. En contrepartie, l’oxygene agit aussi comme un cap-
teur d’e−hyd, neutralisant ainsi leurs actions sur l’ADN. Les cellules hypoxiques, ou l’effet
de la radiotherapie est moins efficace par manque d’oxygene, se trouvent dans les zones
mal alimentees en sang des tumeurs cancereuses. Par contre, la contribution de l’e−hyd aux
dommages y devient plus importante. Il devient alors important de trouver un moyen
d’amplifier l’effet des dommages a l’ADN par l’e−hyd dans ces cellules resistantes aux trai-
tements. Ainsi, l’utilisation du cisplatine, combinee a la radiotherapie, peut jouer le role
de radiosensibilisateur.
En radiotherapie, les rayonnements ionisants utilises, telles que les rayons γ, generent
une grande quantite d’effet indirect du a leur faible transfert d’energie lineaire (TEL).
La grande majorite de ces effets a pour resultat la production d’electrons prehydrates
(faiblement pieges). Nguyen et al. (Nguyen et al., 2011) ont montre que ces derniers sont
responsables du rendement des CSB et CDB sur l’ADN, a un taux deux fois plus grand
que ceux induits par •OH.
Dans les experiences presentees dans cette these, la faible concentration des echan-
tillons elimine la contribution de la reaction des electrons prehydrates dans le but d’etu-
dier l’effet de l’e−hyd. Pour les rayons γ (TEL≤1 eV/nm), la valeur G des e−hyd est 10 fois
plus grande que pour l’4He (TEL≈100 eV/nm) a 5 MeV (LaVerne, 2004). Malgre la faible
energie de l’e−hyd, le puits de potentiel forme par les molecules d’eau l’entourant peut etre
perturbe par la presence de Pt et des groupements polaires (NH3) du cisplatine, rendant
ainsi l’e−hyd reactif envers l’ADN.
La presente these porte sur l’interaction de l’e−hyd avec des complexes oligonucleotide-
cisplatine, soit un systeme beaucoup plus simple que les plasmides surenroules dont la
71

5 : DISCUSSION 72
sequence de nucleotides n’est pas controlee. Le choix des sequences de nucleotides permet
de choisir les differentes configurations de liaisons du cisplatine afin de mieux etudier leur
impact sur les mecanismes en jeux dans l’effet synergique entre l’agent de chimiotherapie
et la radiotherapie.
5.1 Detachement de l’adduit du cisplatine
Le detachement de l’adduit du cisplatine (cisPt) est l’observation la plus frappante
due a l’interaction entre l’e−hyd et le complexe oligonucleotide-cisPt. Ce phenomene a ete
observe avec trois differentes techniques experimentales et ce, dans differentes conditions
de concentration, de dose, de longueur et de sequence d’oligonucleotides.
Dans le premier article, l’etude de l’effet des differents radicaux sur deux types d’oli-
gonucleotides a ete menee avec des mono- et di-adduit de cisplatine, tel que montre sur la
figure 12. La technique de gel d’electrophorese a permis de mettre en evidence, dans le cas
du mono-adduit TTTTTTTGTTT (ODN-TGT), la croissance d’un pic en fonction de la
dose pres de la bande parentale, ce qui est associe au detachement du cisplatine. Alors
que pour le di-adduit TTTGTGTTTTT (ODN-GTG), un pic supplementaire s’ajoute,
mettant en evidence le detachement d’un et de deux liens N7-Pt. Cet effet n’est visible
qu’en isolant l’effet des e−hyd dans le systeme avec des capteurs appropries. L’effet du•OH a aussi ete etudie et il a ete montre qu’il produit des cassures de brin multiples a
l’ADN avec et sans cisplatine, ce qui contraste avec l’effet des e−hyd, ou aucune cassure
de brins n’a ete observee. De plus, l’interaction de l’e−hyd avec le di-adduit de cisplatine
est plus grande que pour le mono-adduit, generant 3 fois plus de detachement de cisPt
pour ODN-GTG-cisPt qu’avec ODN-TGT-cisPt (article 1). Le comportement lineaire du
detachement de deux liens de cisPt en fonction de la dose montre que ce phenomene se
produit en une etape, soit par un seul electron. Ce resultat corrobore ceux de Kopyra et
al (Kopyra et al., 2009), dont l’experience consistait a irradier par un faisceau d’electrons
monoenergetique du cisplatine en phase gazeuse. Ces derniers ont observe le detachement
des deux Cl du cisplatine par un seul electron possedant une energie proche de 0 eV.
La formation d’un anion transitoire est le mecanisme en jeux dans ce phenomene ;
i.e., le transfert d’e−hyd du systeme aqueux vers une ou un melange d’orbitales libres du
complexe ODN-cisPt. Une fois l’anion transitoire [ODN-cisPt]− forme, l’electron addi-
tionnel pourra se stabiliser sur une base ou sur le cisplatine, s’il reste assez longtemps en
place pour provoquer la relaxation de la polarisation de l’environnement par la charge

5 : DISCUSSION 73
Figure 12 – Complexe ODN-TGT-cisPt et ODN-GTG-cisPt
supplementaire. Par contre, s’il occupe majoritairement une orbitale anti-liante, la mo-
lecule pourra se dissocier avant qu’il soit possible pour l’electron de se stabiliser sur la
molecule.
La figure 13 montre un exemple de ce dernier cas (i.e., le transfert dissociatif de
l’electron (TDE)) avec des courbes de potentiel pour une molecule AB neutre, AB− et
un electron en surplus. L’absorption de cet electron dans une orbitale libre s’effectue
dans la region de Franck-Condon et la dissociation se produit seulement si cette orbitale
est occupee durant au moins la moitie du temps d’une vibration dans la direction de la
dissociation (10−14 s), pour que l’etirement de la liaison AB depasse un seuil critique.
Par exemple, la difference de hauteur de barriere energetique entre AB et AB− neces-
saire pour briser la molecule est dictee par l’affinite electronique (AE) de B. Donc, plus
l’affinite electronique de la molecule est grande, plus la dissociation est favorable energe-
tiquement. Ceci se produit en abaissant l’energie du seuil thermodynamique (ΔH), soit
l’energie minimale necessaire pour dissocier la liaison Pt-G : ΔH(B−)= D(A-B)-AE(B).
Le fragment ayant la plus grande affinite electronique est celui qui a la plus grande proba-
bilite de garder l’electron en surplus, soit le cisplatine dans notre cas. Ceci a ete confirme
par Kopyra et al., dont l’experience du detachement des chlores du cisplatine en phase
gazeuse a montre par spectrometrie de masse un plus grand rendement de fragments
cisPt− et de Cl−2 .
En phase gazeuse, le mecanisme de detachement via l’absorption d’un electron en
surplus est legerement different du cas du TDE en milieu aqueux. Comparativement a la
phase gazeuse, le milieu polaire abaisse l’energie de la courbe de potentiel de la molecule
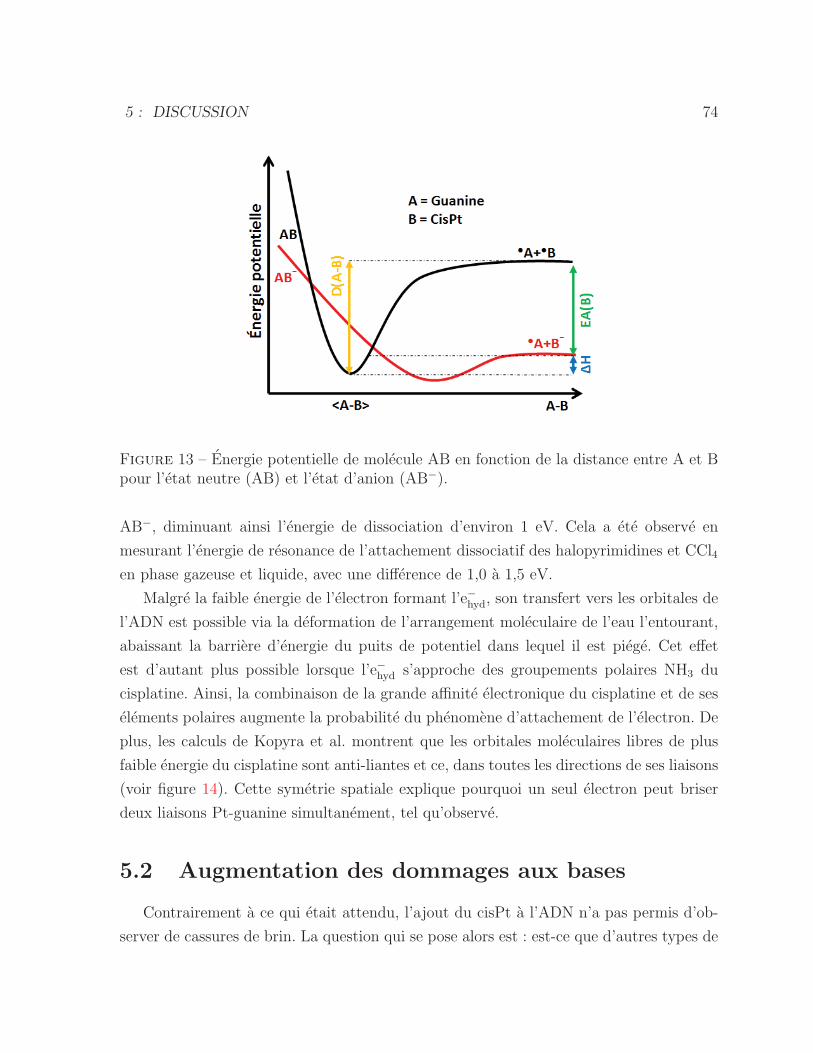
5 : DISCUSSION 74
Figure 13 – Energie potentielle de molecule AB en fonction de la distance entre A et Bpour l’etat neutre (AB) et l’etat d’anion (AB−).
AB−, diminuant ainsi l’energie de dissociation d’environ 1 eV. Cela a ete observe en
mesurant l’energie de resonance de l’attachement dissociatif des halopyrimidines et CCl4
en phase gazeuse et liquide, avec une difference de 1,0 a 1,5 eV.
Malgre la faible energie de l’electron formant l’e−hyd, son transfert vers les orbitales de
l’ADN est possible via la deformation de l’arrangement moleculaire de l’eau l’entourant,
abaissant la barriere d’energie du puits de potentiel dans lequel il est piege. Cet effet
est d’autant plus possible lorsque l’e−hyd s’approche des groupements polaires NH3 du
cisplatine. Ainsi, la combinaison de la grande affinite electronique du cisplatine et de ses
elements polaires augmente la probabilite du phenomene d’attachement de l’electron. De
plus, les calculs de Kopyra et al. montrent que les orbitales moleculaires libres de plus
faible energie du cisplatine sont anti-liantes et ce, dans toutes les directions de ses liaisons
(voir figure 14). Cette symetrie spatiale explique pourquoi un seul electron peut briser
deux liaisons Pt-guanine simultanement, tel qu’observe.
5.2 Augmentation des dommages aux bases
Contrairement a ce qui etait attendu, l’ajout du cisPt a l’ADN n’a pas permis d’ob-
server de cassures de brin. La question qui se pose alors est : est-ce que d’autres types de

5 : DISCUSSION 75
Figure 14 – La plus basse orbitale moleculaire non-occupee de cisplatine (Kopyra et al.,2009). Figure reproduite avec la permission de John Wiley and Sons.
dommage a l’ADN peuvent se produire lors du detachement ou suite au detachement du
cisPt. L’objectif du deuxieme article a donc ete de detecter et de quantifier les dommages
aux bases de l’oligonucleotide ODN-GTG, avec et sans cisPt, induits par l’e−hyd suite a
l’irradiation aux rayons gamma. Il a ete observe par HPLC qu’a 1000 Gy, le detachement
du cisplatine genere 50% de guanines endommagees. Pour cette meme dose, environ 2,5
thymines par detachement de cisPt sont aussi endommagees, ce qui n’etait pas attendu
etant donne que cette base n’est pas le site d’attachement de l’agent chimiotherapeutique.
L’augmentation des dommages aux thymines peut etre due au transfert d’electron
capture entre les thymines. Donc, l’e−hyd serait initialement capture par une thymine et
ensuite, deux voies seraient possibles : 1) le dommage a la base ou 2) le transfert de cet
electron d’une base a une autre par activation thermique (”hopping”) (figure 15) vers le
cisPt. Une autre possibilite est que les dommages observes aux thymines et aux guanines
soient le resultat d’une interaction du cisPt, detache et reactif, avec l’ADN.
5.3 Radiolyse pulsee et la determination de la constante
de vitesse
Puisque la majorite des reactions de nature radicalaire sont generalement tres rapides,
etudier la cinetique des reactions necessite des equipements avec une tres haute resolution
temporelle. Avec la technique de radiolyse pulsee, l’echantillon est excite par un pulse
d’electrons de haute energie, typiquement 3-10 MeV, genere par un accelerateur lineaire

5 : DISCUSSION 76
Figure 15 – Transfert de l’electron d’une base a une autre vers le cisPt
(linac). La duree du pulse d’electrons est de l’ordre de la picoseconde, permettant l’etude
de la cinetique des processus impliquant la radiolyse de l’eau. Ce court pulse d’electrons
est dirige vers le substrat a etudier et un systeme de detection optique (ex. absorption
optique par transmission) permet d’etudier le declin ou l’augmentation des differentes
especes produites.
Le probleme dans les etudes de cinetique des reactions est le changement des concen-
trations des reactifs pendant la reaction. Par exemple, imaginons que nous avons une
reaction entre deux reactifs :
A + B −→ produits (5.1)
r = k[A]n[B]m (5.2)
ou r est la vitesse de reaction, k est la constante de vitesse de reaction, m et n sont
les ordres partiels de reaction puis [A] et [B] sont les concentrations de A et B. Il est
possible d’isoler l’effet du changement de la concentration de A en approximant que
la concentration de B reste constante pour la duree de la reaction. Pour cela, il faut
que la concentration de B soit beaucoup plus grande que celle de A, permettant ainsi
de simplifier l’equation de reaction en considerant une constante de vitesse de reaction

5 : DISCUSSION 77
pseudo-premier ordre qui comprend la concentration de B :
r = (k [B]m0 ) [A]n = kapp [A]
n (5.3)
−d [A]
dt= kapp [A] (5.4)
−∫ [A]t
[A]0
d [A]
[A]=
∫ t
0
kapp dt (5.5)
[A]t = [A]0 e−kapp t (5.6)
Ainsi, si plusieurs experiences sont effectuees avec differentes concentrations de B, il est
alors possible d’extraire la constante de vitesse apparente (kapp) pour chaque concentra-
tion. Finalement le calcul de la pente du graphique de kapp en fonction de [B] donne
directement la constante de vitesse de second ordre (k). Dans la presente etude, [A] cor-
respond a la concentration des e−hyd, qui est induite par la radiolyse de l’eau causee par
le pulse d’electrons.
Avec cette technique, il a ete possible de mesurer la constante de vitesse de reaction
des e−hyd avec le cisplatine seul, le trinucleotide GTG et le complexe GTG-cisPt, tel que
presente dans le troisieme article. Les e−hyd presentent une bande d’absorption optique
maximale a 720 nm dans l’eau pure (Mostafavi and Lampre, 2008). Le maximum de
bande d’absorption est decale a plus haute longueur d’onde si la constante dielectrique
du solvant diminue. Dans notre experience, les mesures de declins des e−hyd ont ete faites
a 600 nm, du a la plus faible quantite de bruit et faible absorption des autres especes a
cette longueur d’onde. Les resultats montrent que la presence du cisPt augmente de 3
fois la vitesse de reaction de l’e−hyd avec le complexe cisPt-GTG en comparaison avec le
GTG sans cisPt. Ceci est une demonstration directe du pouvoir attractif du cisplatine
envers l’e−hyd du a sa grande affinite electronique.
Il a aussi ete possible de mesurer la cinetique de l’interaction entre l’e−hyd et le com-
plexe cisPt-GTG. Ces mesures ont ete effectuees en observant l’ensemble de spectres
d’absorption transitoire de 300 a 600 nm. La constante de vitesse de la formation de
PtI est correlee a celle du declin des e−hyd, montrant l’implication de ces derniers dans le
phenomene. Dans cette etude, les especes reduites presentent un maximum d’absorption
a 310 nm, soit a celle rapportee pour PtI dans la litterature (Adams et al., 1968). Par
contre, la bande d’absorption a 310 nm est elargie par rapport a celle de PtI hydrolyse,
ce qui suggere que cette espece est le radical d’anion de GTG-cisPt. Cependant, nous ne
pouvons pas aussi exclure la formation de PtI hydrolyse sans ligand GTG dans cette re-

5 : DISCUSSION 78
gion spectrale. Cette mesure a ete effectuee a 14 Gy, indiquant que le TDE et la reduction
de cisPt suivie par le detachement cisPt se produit aussi a faible dose.

CONCLUSION ET PERSPECTIVES
Les bienfaits de la combinaison de la radiotherapie et de la chimiotherapie pour la
lutte contre plusieurs cancers sont aujourd’hui reconnus et utilises en traitement clinique.
Bien que les mecanismes de ces traitements individuels soient connus, leurs interactions
synergiques demeurent mal comprises. Dans le cadre de cette these, il a ete demontre
pour la premiere fois que l’irradiation d’un simple systeme d’oligonucleotides-cisplatine
en solution engendre, avec un haut taux de rendement, le detachement de l’agent chimio-
therapeutique via le phenomene de TDE par les e−hyd. Au cours de ce travail, ce phenomene
a ete observe et etudie avec trois differentes techniques experimentales.
La technique de gel electrophorese a permis de mesurer le detachement d’un et de deux
liens de l’adduit du cisplatine avec les guanines des oligonucleotides (ODN-GTG et ODN-
TGT) apres avoir subi differentes doses d’irradiation de rayons gamma en presence de
capteurs des •OH. Il a ete demontre qu’un seul e−hyd peut etre responsable de la dissociation
complete de l’adduit du cisplatine de l’ADN. Ce phenomene implique necessairement la
dissociation par TDE. Il a aussi ete possible d’affirmer qu’aucune cassure de brin d’ADN
n’est generee par TDE.
La technique d’HPLC a permis non seulement de confirmer le detachement de l’ad-
duit du cisplatine apres irradiation, mais aussi d’observer et de quantifier les dommages
engendres aux bases d’ADN. Avec une dose de 1000 Gy par exemple, pour chaque cisPt
detache, il y a 2,5 thymines et 1 guanine d’endommagees de l’oligo (TTTTTGTGTTT).
Ces dommages aux bases, dus au depart du cisPt, donnent des elements de comprehension
de la synergie entre la radiotherapie et la chimiotherapie. Deux voies peuvent expliquer
les dommages aux bases : (1) une fois le cisPtI detache, ce dernier demeure reactif avec
l’ADN ; (2) les dommages aux thymines peuvent se produire pendant le transfert de base
a base de l’electron attache vers le cisPt.
La radiolyse pulsee a permis d’etudier la cinetique du phenomene d’interaction de
79

6 : CONCLUSION ET PERSPECTIVES 80
l’e−hyd avec le complexe cisPt-GTG et de confirmer le role des e−hyd dans le processus. En
mesurant le declin de la population de ces electrons en fonction du temps, il a ete mis en
evidence que la grande affinite electronique de l’agent chimiotherapeutique attire les e−hydvers l’oligonucleotide en solution suite a son irradiation. Les constantes de vitesse extraites
de cette etude ont permis de constater que l’interaction entre l’ADN et les e−hyd est 3 fois
superieure en presence de l’adduit du cisplatine. La technique a aussi permis d’observer
directement l’accroissement du signal provenant de la formation de PtI permettant ainsi
de le correler au declin des e−hyd.
Somme toute, cette these a permis de lever le voile sur une partie de la synergie
entre le traitement de la radiotherapie et de la chimiotherapie. Bien que la grande af-
finite electronique du cisPt etait deja suspectee dans le processus donnant lieu a l’effet
super additif, le phenomene de detachement du cisPt est un phenomene nouveau qui en-
gendre des dommages supplementaires aux bases. Les perspectives d’avenir de ce projet
seraient d’identifier les types de dommages aux bases de l’oligonucleotide. Il sera aussi
interessant de poursuive la meme etude dans l’ADN double brin et comparer avec l’ADN
simple brin. De plus, on peut determiner l’effet de la longueur de l’oligonucleotide sur la
radiosensibilisation par le cisplatine.

REMERCIEMENTS
J’adresse mes sinceres remerciements a mes co-directeurs, Prs Leon Sanche et Darel
Hunting, de m’avoir donne la chance d’entreprendre un projet de doctorat ainsi que pour
leur encadrement.
Je tiens aussi a remercier chaleureusement Pr Richard Wagner pour la generosite de son
temps et pour tous ses bons conseils m’ayant permis de comprendre plus facilement la
partie axee sur la chimie de mon projet.
Je desire de plus remercier vivement nos collaborateurs a l’universite Paris Sud 11, Pr
Mehran Mostafavi et Dr Jean-Louis Marigner, pour la belle opportunite qu’ils m’ont of-
ferte, soit de travailler dans leur labo. Merci pour cette belle experience scientifique et
personnelle !
Merci aussi a vous, Pr Martin Lepage, d’avoir accepte de presider ma soutenance. Merci
aussi aux Prs Christopher J.Wilds et Antonio Conconi d’avoir accepte d’etre membres
de mon jury de these.
Je voudrais aussi remercier Pierre Cloutier et Sonia Girouard de m’avoir initiee aux tech-
niques experimentales. Merci a Pierre Cloutier pour les bons moments qu’on a passes en
jouant au poker.
Merci aussi a Daniel Robillard pour le support technique informatique et pour etre un
bon ami qui a toujours ete la pour moi.
J’aimerai enfin exprimer mes remerciements les plus profonds a ma famille, particuliere-
ment a Jonathan Vermette et a mes parents. Merci de m’avoir soutenue et encouragee
pendant mon doctorat.
81

LISTE DES REFERENCES
Abdoul-Carime, H., Gohlke, S., and Illenberger, E. (2004). Site-specific dissociation ofDNA bases by slow electrons at early stages of irradiation. Physical Review Letters,92(16) :168103–1.
Adams, G. E., Broszkiewicz, R. B., and Michael, B. D. (1968). Pulse radiolysis studieson stable and transient complexes of platinum. Transactions of the Faraday Society,64 :1256.
Arumainayagam, C. R., Lee, H. L., Nelson, R. B., Haines, D. R., and Gunawardane, R. P.(2010). Low-energy electron-induced reactions in condensed matter. Surface ScienceReports, 65(1) :1–44.
Bald, I., Illenberger, E., and Kopyra, J. (2012). Damage of DNA by Low Energy Electrons(< 3 eV). Journal of Physics : Conference Series, 373 :012008.
Barnett, R. N., Cleveland, C. L., Joy, a., Landman, U., and Schuster, G. B. (2001). Chargemigration in DNA : ion-gated transport. Science (New York, N.Y.), 294(5542) :567–571.
Berdys, J., Anusiewicz, I., Skurski, P., and Simons, J. (2004). Damage to model DNAfragments from very low-energy (<1 eV) electrons. Journal of the American ChemicalSociety, 126(20) :6441–6447.
Bertout, J. a., Patel, S. a., and Simon, M. C. (2008). The impact of O2 availability onhuman cancer. Nature reviews. Cancer, 8(12) :967–975.
Boon, E. M. and Barton, J. K. (2002). Charge transport in DNA Elizabeth M Boon andJacqueline K Barton *. pages 320–329.
Boscolo-Rizzo, P., Gava, a., Marchiori, C., Baggio, V., and da Mosto, M. C. (2011).Functional organ preservation in patients with locoregionally advanced head and necksquamous cell carcinoma treated by platinum-based multidrug induction chemotherapyand concurrent chemoradiotherapy. Annals of Oncology, 22(8) :1894–1901.
Boudaıffa, B., Cloutier, P., Hunting, D., Huels, M. a., and Sanche, L. (2000). Resonantformation of DNA strand breaks by low-energy (3 to 20 eV) electrons. Science (NewYork, N.Y.), 287(5458) :1658–1660.
82

LISTE DES REFERENCES 83
Cai, Z., Li, X., and Sevilla, M. D. (2002). Excess Electron Transfer in DNA : Effect of BaseSequence and Proton Transfer. The Journal of Physical Chemistry B, 106(10) :2755–2762.
Cai, Z. and Sevilla, M. D. (2000). Electron Spin Resonance Study of Electron Transferin DNA : Inter-Double-Strand Tunneling Processes. Journal of Physical Chemistry B,104 :6942–6949.
Cai, Z. and Sevilla, M. D. (2004). Studies of Excess Electron and Hole Transfer in DNAat Low Temperatures. pages 103–128.
Candelaria, M., Garcia-Arias, A., Cetina, L., and Duenas Gonzalez, A. (2006). Ra-diosensitizers in cervical cancer. Cisplatin and beyond. Radiation oncology (London,England), 1 :15.
Choy, H. (2003). Chemoradiation in Cancer Therapy. Humana press : Totowa, nj edition.
Cobut, V., Frongillo, Y., Patau, J. P., and Goulet, T. (1998). Monte Carlo Simulation ofFast Electron and. Radiat. Phys. Chem., 3 :229–243.
Coste, F., Malinge, J. M., Serre, L., Shepard, W., Roth, M., Leng, M., and Zelwer, C.(1999). Crystal structure of a double-stranded DNA containing a cisplatin interstrandcross-link at 1.63 Aresolution : Hydration at the platinated site. Nucleic Acids Research,27(8) :1837–1846.
Dabholkar, M., Bostick-Bruton, F., Weber, C., Bohr, V. A., Egwuagu, C., and Reed, E.(1992). Ercc1 and ercc2 expression in malignant tissues from ovarian cancer patients.Journal of the National Cancer Institute, 84(19) :1512–1517.
DeLara, C. M., Jenner, T. J., Townsend, K. M., Marsden, S. J., and O’Neill, P. (1995).The effect of dimethyl sulfoxide on the induction of DNA double-strand breaks in V79-4mammalian cells by alpha particles. Radiation research, 144(1) :43–49.
Eastman, a. (1983). Characterization of the adducts produced in DNA by cis-diamminedichloroplatinum(II) and cis-dichloro(ethylenediamine)platinum(II). Bioche-mistry, 22(16) :3927–3933.
Farhataziz and Rodgers (1987). Radiation chemistry-principles and applications. V ,ew York edition.
Ferradini, C. and Jay-Gerin, J.-P. (1999). La radiolyse de l’eau et des solutions aqueuses :historique et actualite. Canadian Journal of Chemistry, 77(819) :1542–1575.
Fink, H. W. and Schonenberger, C. (1999). Electrical conduction through DNAmolecules.Nature, 398(6726) :407–410.

LISTE DES REFERENCES 84
Gately, D. P. and Howell, S. B. (1993). Cellular accumulation of the anticancer agentcisplatin : a review. British journal of cancer, 67(6) :1171–1176.
Hayon, E. and Allen, O. (1961). Evidence for two kinds of ”h atoms” is the radiationchemistry of water.
Hei, T. O. M. K. and Hall, E. J. (1994). 0 Chemo- and Radiosensitivity. 29(2) :267–271.
Ito, T. and Rokita, S. E. (2004). Criteria for efficient transport of excess electrons inDNA. Angewandte Chemie - International Edition, 43(14) :1839–1842.
Jamieson, E. R. and Lippard, S. J. (1999). Structure , Recognition , and Processing ofCisplatin - DNA Adducts. Medicinal Chemistry.
Johnson, S. W., Swiggard, P. a., Handel, L. M., Johnson, W., Swiggard, A., Brennan,M., Godwin, K., and Owls, R. F. (1994). Relationship between Platinum-DNA AdductFormation and Removal and Cisplatin Cytotoxicity in Cisplatin-sensitive and -resistantHuman Ovarian Cancer Cells Relationship between Platinum-DNA Adduct Formationand Removal and Cisplatin Cytotoxicity in Cisplat. Cancer research, 54 :5911–5916.
Jortner, B. Y. and Rabani, J. (1961). THE DECOMPOSITION OF CHLOROACETICACID I N AQUEOUS SOLUTIONS. pages 2078–2081.
Jung, Y. and Lippard, S. J. (2007). Direct Cellular Responses to Platinum-InducedDNA Damage Direct Cellular Responses to Platinum-Induced DNA Damage. Chemicalreviews, 107(5) :1387–1407.
Kelland, L. (2007). The resurgence of platinum-based cancer chemotherapy. Naturereviews. Cancer, 7(8) :573–584.
Kelley, S. O., Holmlin, R. E., Stemp, E. D. a., and Barton, J. K. (1997). Photoinducedelectron transfer in ethidium-modified DNA duplexes : Dependence on distance andbase stacking. Journal of the American Chemical Society, 119(41) :9861–9870.
Kopyra, J., Koenig-Lehmann, C., Bald, I., and Illenberger, E. (2009). A single slowelectron triggers the loss of both chlorine atoms from the anticancer drug cisplatin :Implications for chemoradiation therapy. Angewandte Chemie - International Edition,48(42) :7904–7907.
LaVerne, J. (2004). Radiation Chemical Effects of Heavy Ions. In Hatano, A. andMozumdar, Y., editors, Charged particle and photon interaction with matter, chapterRadiation chemical effects of heavy ions., page p. 403. Mercel dekker, inc edition.
LaVerne, J. A. and Pimblott, S. M. (1995). Electron energy-loss distributions in solid,dry DNA. Radiation research, 141(2) :208–215.
Lehnert, S. (2007). Biomolecular Action of Ionizing Radiation. Taylor and francis edition.

LISTE DES REFERENCES 85
Lewis, F. D., Wu, T., Zhang, Y., Letsinger, R. L., Greenfield, S. R., and Wasielewski,M. R. (1997). Distance-dependent electron transfer in DNA hairpins. Science (NewYork, N.Y.), 277(5326) :673–676.
Ly, D., Sanii, L., and Schuster, G. B. (1999). Mechanism of charge transport in DNA :Internally-linked anthraquinone conjugates support phonon-assisted polaron hopping.Journal of the American Chemical Society, 121(40) :9400–9410.
Martin, F., Burrow, P. D., Cai, Z., Cloutier, P., Hunting, D., and Sanche, L. (2004). DNAstrand breaks induced by 0-4 eV electrons : The role of shape resonances. PhysicalReview Letters, 93(6) :6–9.
Michaels, H. B., Epp, E. R., Ling, C. C., and Peterson, E. C. (1978). Oxygen sensitizationof CHO cells at ultrahigh dose rates : prelude to oxygen diffusion studies. Radiationresearch, 76(3) :510–521.
Mistry, P., Kelland, L. R., Abel, G., Sidhar, S., and Harrap, K. R. (1991). The rela-tionships between glutathione, glutathione-S-transferase and cytotoxicity of platinumdrugs and melphalan in eight human ovarian carcinoma cell lines. British journal ofcancer, 64(2) :215–220.
Mostafavi, M. and Lampre, I. (2008). The solvated electron : a singular chemical spe-cies. In Spotheim-Maurizot, M., Mostafavi, M., Thierry, D., and Belloni, J., editors,Radiation chemistry : from basics to applications in material and life sciences, chapterRadiation chemical effects of heavy ions., page p. 40. Edp scienc edition.
Murphy, C. J., Arkin, M. R., Jenkins, Y., Ghatlia, N. D., Bossmann, S. H., Turro, N. J.,and Barton, J. K. (1993). Long-range photoinduced electron transfer through a DNAhelix. Science (New York, N.Y.), 262(5136) :1025–1029.
Nguyen, J., Ma, Y., Luo, T., Bristow, R. G., Jaffray, D. a., and Lu, Q.-B. (2011). Directobservation of ultrafast-electron-transfer reactions unravels high effectiveness of reduc-tive DNA damage. Proceedings of the National Academy of Sciences of the UnitedStates of America, 108(29) :11778–11783.
Nikjoo, H., Bolton, C. E., Watanabe, R., Terrissol, M., O’Neill, P., and Goodhead, D. T.(2002). Modelling of DNA damage induced by energetic electrons (100 eV to 100 keV).Radiation protection dosimetry, 99(1-4) :77–80.
Nunez, M. E., Hall, D. B., and Barton, J. K. (1999). Long-range oxidative damage toDNA : Effects of distance and sequence. Chemistry and Biology, 6(2) :85–97.
Okahata (1967). Communications to the editor. Bulletin of the Medical Library Associa-tion, 55(2) :220.

LISTE DES REFERENCES 86
O’Neill, M. a. and Barton, J. K. (2002). Effects of strand and directional asymmetryon base-base coupling and charge transfer in double-helical DNA. Proceedings of theNational Academy of Sciences of the United States of America, 99(26) :16543–16550.
Pathirannehelage, N. S. E. (2011). Charge Transfer in Deoxyribonucleic Acid ( DNA ) :Static Disorder , Dynamic Fluctuations and Complex Kinetic .
Peters, W. a., Liu, P. Y., Barrett, R. J., Stock, R. J., Monk, B. J., Berek, J. S., Souhami,L., Grigsby, P., Gordon, W., and Alberts, D. S. (2000). Concurrent chemotherapyand pelvic radiation therapy compared with pelvic radiation therapy alone as adjuvanttherapy after radical surgery in high-risk early-stage cancer of the cervix. Journal ofClinical Oncology, 18(8) :1606–1613.
Porath, D., Bezryadin, a., de Vries, S., and Dekker, C. (2000). Direct measurement ofelectrical transport through DNA molecules. Nature, 403(6770) :635–638.
Rajski, S. R., Jackson, B. a., and Barton, J. K. (2000). DNA repair : Models for da-mage and mismatch recognition. Mutation Research - Fundamental and MolecularMechanisms of Mutagenesis, 447(1) :49–72.
Ray, S. G., Daube, S. S., and Naaman, R. (2005). On the capturing of low-energyelectrons by DNA. Proceedings of the National Academy of Sciences of the UnitedStates of America, 102(1) :15–19.
Razskazovskii, Y. and Swarts, S. (1997). Competitive electron scavenging by chemicallymodified pyrimidine bases in bromine-doped DNA : Relative efficiencies and relevanceto intrastrand electron migration distances. The Journal of Physical Chemistry B,5647(96) :1460–1467.
Reedijk, J. (2009). Platinum anticancer coordination compounds : Study of DNA bindinginspires new drug design. European Journal of Inorganic Chemistry, (10) :1303–1312.
Rezaee, M., Sanche, L., and Hunting, D. J. (2013). Cisplatin enhances the formation ofDNA single- and double-strand breaks by hydrated electrons and hydroxyl radicals.Radiation research, 179(3) :323–31.
Rosenberg, B. (1985). Fundamental studies with cisplatin. Cancer, 55(10) :2303–l6.
Rosenberg, B., Vancamp, L., and Krigas, T. (1965). Inhibition of Cell Division in Esche-richia Coli By Electrolysis Products From a Platinum Electrode. Nature, 205 :698–699.
Rowinsky, E. K. (1999). Novel radiation sensitizers targeting tissue hypoxia. Oncology(Williston Park, N.Y.), 13(10 Suppl 5) :61–70.
Safaei, R., Holzer, A. K., Katano, K., Samimi, G., and Howell, S. B. (2004). The role ofcopper transporters in the development of resistance to Pt drugs. Journal of InorganicBiochemistry, 98(10 SPEC. ISS.) :1607–1613.

LISTE DES REFERENCES 87
Samant, S., Kumar, P., Wan, J., Hanchett, C., Vieira, F., Murry, T., Wong, F. S. H.,and Robbins, K. T. (1999). Concomitant radiation therapy and targeted cisplatinchemotherapy for the treatment of advanced pyriform sinus carcinoma : Disease controland preservation of organ function. Head and Neck, 21(7) :595–601.
Sanche, L. (2009a). Low-Energy Electron Interaction with DNA : Bond Dissociation andFormation of Transient Anions, Radicals, and Radical Anions, pages 239–293. JohnWiley and Sons, Inc.
Sanche, L. (2009b). Role of secondary low energy electrons in radiobiology and chemo-radiation therapy of cancer. Chemical Physics Letters, 474(1-3) :1–6.
Schuster, G. B. (2000). Long-range charge transfer in DNA : Transient structural distor-tions control the distance dependence. Accounts of Chemical Research, 33(4) :253–260.
Seiwert, T. Y., Salama, J. K., and Vokes, E. E. (2007). The concurrent chemoradiationparadigm–general principles. Nature clinical practice. Oncology, 4(2) :86–100.
Simons, J. (2006). Electrons Cause DNA-Strand Breaks ? 39(10) :772–779.
von Sonntag, C. (1987). The chemical basis of radiation biology. Taylor and francisedition.
von Sonntag, C. (2006). Free-Radical-Induced DNA Damage and Its Repair. Springeredition.
Wang, C. R., Nguyen, J., and Lu, Q. B. (2009). Bond breaks of nucleotides by dissociativeelectron transfer of nonequilibrium prehydrated electrons : A new molecular mechanismfor reductive DNA damage. Journal of the American Chemical Society, 131(32) :11320–11322.
Ward, J. F. (1988). DNA damage produced by ionizing radiation in mammalian cells :identities, mechanisms of formation, and reparability. Progress in nucleic acid researchand molecular biology, 35 :95–125.
Wozniak, K. and Blasiak, J. (2002). Recognition and repair of DNA-cisplatin adducts.Acta Biochimica Polonica, 49(3) :583–596.
Zheng, Y., Cloutier, P., Hunting, D. J., Sanche, L., and Wagner, J. R. (2005). Chemi-cal basis of DNA sugar-phosphate cleavage by low-energy electrons. Journal of theAmerican Chemical Society, 127(47) :16592–16598.













![PLY THICKNESS INFLUENCE IN COMPRESSION AFTER …...tively) using the same modified Hashin failure criterion [34], where the Cohesive Failure Crite-ria [6] was inserted in order to](https://img.pdfslide.fr/doc/110x75/5e92b14d5c415a6d38176d44/ply-thickness-influence-in-compression-after-tively-using-the-same-modiied.jpg)











