Embed Size (px)
Citation preview

N° D’ORDRE: 7385
UNIVERSITE PARIS XI U.F.R SCIENTIFIQUE D’ORSAY
THESE
Présentée
Pour obtenir
Le GRADE de DOCTEUR EN SCIENCES
DE L’UNIVERSITE PARIS XI ORSAY
Par
David LAST
Sujet:
Caractérisation de l’architecture tridimensionnelle du
tissu trabéculaire in vitro et in vivo par IRM
Soutenue le 25 Novembre 2003 devant la Commission d’examen
Monsieur André BRIGUET Madame Geneviève GUILLOT Directeur de thèse Monsieur Alain HERMENT Monsieur Yves MENU Président du jury Monsieur Eric PETIT Rapporteur Madame Françoise PRETEUX Rapporteur


Caractérisation de l’architecture tridimensionnelle du
tissu trabéculaire in vitro et in vivo par IRM
David LAST


Remerciements
Remerciements
Ce travail de thèse s’est déroulé dans l’Unité de Recherche en Résonance Magnétique
Médicale (U2R2M) à Orsay. Un seul « auteur » figure sur la couverture. Ceci est trompeur.
Ce travail n’aurait pu aboutir sous la forme de ce manuscrit si certaines personnes n’y avaient
pas pris part. Je tiens à les en remercier vivement ici.
En premier lieu, je veux saluer la qualité de l’encadrement de ma directrice de thèse, Madame
Geneviève GUILLOT. La richesse de ses connaissances, sa rigueur, son dévouement et sa
disponibilité en font certainement l’un des meilleurs directeurs possibles pour une thèse.
Merci pour tout Geneviève !
Je souhaite remercier le Professeur Jacques BITTOUN de m’avoir accueilli dans son unité
mais aussi pour faire si bien « tourner la boutique ». Les conditions de travail dans lesquelles
je me suis trouvé pendant cette thèse étaient vraiment excellentes: on travaille et on se sent
bien à l’U2R2M !
Je tiens à remercier Monsieur Luc DARRASSE, Monsieur « une idée.s-1 ». C’est grâce à lui
qu’on a pu obtenir de si jolis résultats in vivo. J’ai vraiment apprécié l’intérêt qu’il a eu dans
mon sujet, son dynamisme et l’étendue de ses compétences.
5

Remerciements
Je tiens à remercier mes prédécesseurs Florence REMY et plus particulièrement Theocharis
ANTONIADIS. Je ne l’ai pas connu mais j’ai pu apprécier la quantité et la qualité du travail
qu’il avait accompli pendant sa thèse. L’état dans lequel se trouve mon exemplaire de son
manuscrit en est la preuve…
Je remercie aussi Ludovic De ROCHEFORT. J’ai vraiment aimé travailler avec lui pendant le
stage qu’il a fait dans l’unité. La qualité de son travail et ses capacités à s’adapter à des sujets
nouveaux sont appréciées par l’équipe « hélium » de l’unité depuis qu’il en est doctorant et
cela n’a rien d’étonnant. Je veux aussi remercier les autres stagiaires qui se sont succédés dans
l’unité: Thierry DONADEY, Emilie RONCALI, Françoise RENALDO, et Aurélie HULEUX.
Je tiens à remercier Madame Françoise PEYRIN, Messieurs Jean-Paul JERNOT et Pascal
LAUGIER ainsi que les membres du GDR 2237 « Imagerie et caractérisation tissulaire
appliquées au domaine ostéoarticulaire » pour les longues discussions enthousiastes et
enrichissantes que j’ai pu avoir avec eux.
J’ai aussi beaucoup apprécié la collaboration que nous avons eue avec le Docteur Léon
CHOEL. Ses nombreux allers-retours entre Orsay et Lyon m’ont permis de me rendre compte
de ses compétences, de sa motivation mais aussi de sa grande gentillesse.
Je remercie aussi l’équipe « EP5 » de l’Université de Würzburg pour leur accueil et plus
particulièrement Messieurs Michael SZIMTENINGS, Titus LANZ et Axel HAASE pour la
collaboration très enrichissante que nous avons eue.
Je tiens à remercier les Professeurs Eric PETIT et Françoise PRETEUX d’avoir accepter de
rapporter ma thèse et les Professeurs Yves MENU et André BRIGUET ainsi que Monsieur
Alain HERMENT de m’avoir fait l’honneur de participer à mon jury.
Je souhaite saluer Monsieur Patrick GONORD pour son aide précieuse tant au niveau de la
recherche que de l’enseignement. Je n’ai pas encore trouvé de domaine où on pouvait le
coller… Je vais continuer à chercher !
6

Remerciements
J’ai eu beaucoup de plaisir à travailler avec Jean-Christophe GINEFRI, que cela soit pour la
semaine de « Travaux d’étude » ou au laboratoire. J’ai aussi apprécié sa bonne humeur qui
rend le quotidien plus agréable.
Je tiens aussi à saluer l’encadrement « logistique »: Jean-Pierre RUAUD, Philippe
DOS SANTOS et Catherine BARJOUX. C’est grâce à eux que l’on peut se concentrer sur nos
gros problèmes sans se faire « manger » par les petits.
Je remercie tous les autres collègues de l’unité pour leur soutien: Messieurs Emmanuel
DURAND, Bernard BANDELIER, Xavier MAITRE et Madame Françoise RIOUX ainsi que
Monsieur Guy FISHMAN (de l’IEF) en particulier pour les passionnants trajets dans le
RER B.
J’adresse un grand MERCI à toute l’équipe enseignante EEA et plus particulièrement à ma
tutrice pendant mon monitorat Madame Mireille ROUSSEAU, à Monsieur Robert MEGY, à
Mesdames Claudine FALCETTA et Elisabeth BOUYSSY ainsi qu’à Messieurs Jean-Philippe
DACUNHA et Yves MAIRE.
Je salue et remercie tout autant la petite équipe « électronique numérique » du Magistère de
Physique Fondamentale: son responsable, le Professeur Etienne AUGE ainsi que Messieurs
Madjid ANANE, Ludovic De POUCQUES et Laurent DOLLA avec lesquels se fut un réel
plaisir de faire de l’enseignement.
Je n’oublie pas non plus les compagnons de galère: Lionel CHIRON, Marie
POIRIER-QUINOT, Alexandre VIGNAUD, ainsi que Frédéric ETIENNE, Florian MONSEF
et tous les doctorants de l’équipe « Modèles numériques pour l’IRM ». C’est beaucoup grâce
à eux que j’ai aimé venir travailler chaque matin pendant ces années.
Je remercie aussi tous mes amis et plus particulièrement Vince, Philou, Bruno, Manu,
Caroline, Julien, Lolo, Dali, François, Fred, Michael et Stéphane qui m’ont soutenu et surtout
qui ne m’ont pas oublié malgré mon absence pendant ces (longs) mois de rédaction.
J’adresse aussi un grand MERCI à Monsieur Maurice GABBAY et sa famille qui m’ont
recueilli et aidé quand je n’étais qu’un jeune petit étudiant perdu à Lyon.
7

Remerciements
A Sandra qui m’accompagne maintenant et je l’espère pour très longtemps. Je la remercie
pour son soutien pendant la rédaction de ce manuscrit: il y a des « bouts » d’elle dedans. Je
souhaite aussi remercier sa famille pour leurs encouragements.
Je souhaite enfin remercier ceux grâce auxquels je suis arrivé jusque là, qui ont veillé sur moi
depuis que je suis né, qui ont tout fait pour que je puisse donner le meilleur de moi: mon père
Jacques, ma mère Colette et ma Sœurette Cécile. Je leur en ai fait voir de toutes les couleurs:
ce manuscrit est pour eux.
8

Table des matières
Table des matières
Remerciements.....................................................................................5
Table des matières ...............................................................................9
Introduction .......................................................................................17
Chapitre I: Le tissu osseux et sa caractérisation..............................21
I.1. Description générale de la structure osseuse .......................................... 21
I.2. Régénération osseuse ................................................................................ 23
I.3. L’ostéoporose............................................................................................. 23
I.4. Rôle de la microarchitecture du tissu trabéculaire................................ 24
I.5. Techniques de caractérisation.................................................................. 26
I.5.1. Absorptiométrie.................................................................................................... 26 I.5.2. Histomorphométrie osseuse................................................................................. 28 I.5.3. Ultrasons ............................................................................................................... 29 I.5.4. Tomographie par rayons X ................................................................................. 30
I.6. La technique IRM ..................................................................................... 31
9

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
I.6.1. Phénomène de la RMN ........................................................................................ 32 I.6.1.1. Effet d’un champ magnétique statique sur les protons................................... 32 I.6.1.2. Perturbation radio-fréquence (RF).................................................................. 33 I.6.1.3. Relaxation ....................................................................................................... 34
I.6.2. Le signal de précession libre (FID) ..................................................................... 36 I.6.2.1. Contraste en T1 ............................................................................................... 37 I.6.2.2. Contraste en T2 ............................................................................................... 38 I.6.2.3. Contraste en densité de protons ...................................................................... 39
I.6.3. Séquences d’imagerie........................................................................................... 40 I.6.3.1. Principe du codage de l’espace....................................................................... 40 I.6.3.2. Cas 1D: gradient de lecture............................................................................. 40 I.6.3.3. Formalisme ..................................................................................................... 42 I.6.3.4. Séquence « écho de gradient »........................................................................ 43
I.6.3.4.1. Impulsion radio-fréquence ................................................................. 43 I.6.3.4.2. Gradient de sélection ......................................................................... 44 I.6.3.4.3. Gradient de refocalisation ................................................................. 44 I.6.3.4.4. Gradient de prélecture ....................................................................... 44 I.6.3.4.5. Gradients de codage de phase ........................................................... 45 I.6.3.4.6. Remarques.......................................................................................... 48
I.6.3.5. Séquence « écho de spins » ............................................................................ 48 I.6.3.6. Avantages et inconvénients de ces séquences ................................................ 49
I.6.4. IRM haute résolution........................................................................................... 49 I.6.4.1. Rôle des gradients........................................................................................... 50 I.6.4.2. Rapport Signal sur Bruit (RSB)...................................................................... 51
I.6.4.2.1. Caractéristiques de la sonde .............................................................. 51 I.6.4.2.2. Champ statique................................................................................... 52 I.6.4.2.3. Voxel................................................................................................... 52 I.6.4.2.4. Durée de vie du signal........................................................................ 52
I.6.4.3. Les artéfacts .................................................................................................... 52 I.6.4.3.1. Artéfact de susceptibilité .................................................................... 52 I.6.4.3.2. Décalage chimique............................................................................. 53 I.6.4.3.3. Repliement (aliasing) ......................................................................... 53
I.6.5. Etat de l’art: IRM du tissu trabéculaire in vitro et in vivo .............................. 53
Chapitre II: Quantification de la microarchitecture trabéculaire...57
II.1. Introduction ............................................................................................. 57
II.2. Un préalable: la segmentation................................................................ 58
II.2.1. Histogramme à modes séparés .......................................................................... 58 II.2.2. Histogramme problématique............................................................................. 59 II.2.3. Méthodes dédiées à la segmentation d’images acquises par IRM.................. 60
II.2.3.1. Seuillage ........................................................................................................ 60 II.2.3.2. Autres méthodes ............................................................................................ 61
II.2.4. Caractérisation de la structure osseuse sans segmentation ............................ 62
II.3. Fraction volumique osseuse .................................................................... 62
10

Table des matières
II.4. Paramètres topologiques......................................................................... 63
II.4.1. Notion de topologie: définitions principales ..................................................... 63 II.4.1.1. Connexité....................................................................................................... 63 II.4.1.2. Genre d’une surface....................................................................................... 64
II.4.2. Nombres de Betti ................................................................................................ 65 II.4.2.1. Définition....................................................................................................... 65 II.4.2.2. Propriétés....................................................................................................... 65 II.4.2.3. Illustrations.................................................................................................... 66
II.4.3. Caractéristique d’Euler-Poincaré ..................................................................... 67 II.4.3.1. Formulation de la caractéristique d’Euler-Poincaré...................................... 67 II.4.3.2. Propriété d’additivité..................................................................................... 68 II.4.3.3. Relation entre les nombres de Betti et la caractéristique d’Euler-Poincaré .. 68
II.4.4. Cas des espaces bornés et discrets..................................................................... 69 II.4.4.1. Approche locale............................................................................................. 69 II.4.4.2. Trame de l’image........................................................................................... 69
II.4.4.2.1. Modes de connexité........................................................................... 70 II.4.4.2.2. Phase complémentaire ...................................................................... 71 II.4.4.2.3. Conséquence: effet de la résolution.................................................. 72
II.4.5. Calcul des paramètres ........................................................................................ 73 II.4.5.1. Nombres de Betti d’ordre 0 et 2 .................................................................... 73 II.4.5.2. Contribution à la caractéristique d’Euler-Poincaré ....................................... 74
II.4.6. Tests et mise en œuvre des paramètres sur le tissu trabéculaire.................... 74 II.4.6.1. Amas secondaires .......................................................................................... 75 II.4.6.2. Effet de la résolution sur CN3........................................................................ 76 II.4.6.3. Protocole........................................................................................................ 78
II.4.7. Applications des paramètres topologiques ....................................................... 79
II.5. Paramètres histomorphométriques ....................................................... 80
II.5.1. Surface spécifique ............................................................................................... 81 II.5.1.1. Formulation de la surface d’un ensemble...................................................... 81
II.5.1.1.1. Etude préalable dans R2.................................................................... 81 II.5.1.1.2. Généralisation à R3........................................................................... 82
II.5.1.2. Cas des espaces bornés et discrets................................................................. 82 II.5.1.2.1. Approche locale ................................................................................ 82 II.5.1.2.2. Trame de l’image .............................................................................. 83
II.5.1.3. Méthode de calcul de BS/TV ........................................................................ 85 II.5.1.3.1. Principe de l’algorithme ................................................................... 85 II.5.1.3.2. Images « tests »................................................................................. 85 II.5.1.3.3. Résultats............................................................................................ 86
II.5.2. Epaisseur et espacement des travées................................................................. 88 II.5.2.1. Calcul indirect: hypothèse sur le modèle de tissu trabéculaire ..................... 88 II.5.2.2. Autres approches: méthodes de calcul direct ................................................ 89
II.5.3. Applications des paramètres histomorphométriques...................................... 90
II.6. Paramètres d’anisotropie et d’orientation ............................................ 91
II.6.1. Longueur moyenne d’interception.................................................................... 91 II.6.2. Modélisation par un ellipsoïde........................................................................... 92 II.6.3. Anisotropie de l’ellipsoïde.................................................................................. 93 II.6.4. Orientation de l’ellipsoïde.................................................................................. 94 II.6.5. Applications......................................................................................................... 94
11

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Chapitre III: Validité du rendu par IRM du tissu trabéculaire: confrontation avec une technique de référence, la microtomographie X par rayonnement synchrotron .......................................................97
III.1. Etat de l’art de l’étude in vitro par IRM du tissu trabéculaire et problématique......................................................................................... 97
III.1.1. Caractérisation du tissu trabéculaire in vitro par IRM ................................ 97 III.1.2. Limitations de ces études .................................................................................. 99 III.1.3. Fiabilité de l’IRM pour la caractérisation du tissu trabéculaire.................. 99 III.1.4. Problématique et présentation du travail ..................................................... 101
III.2. Les échantillons utilisés ....................................................................... 102
III.3. Les images ............................................................................................. 103
III.3.1. Images obtenues par IRM .............................................................................. 103 III.3.1.1. Optimisation des paramètres d’acquisition ................................................ 103 III.3.1.2. Paramètres de la séquence.......................................................................... 104 III.3.1.3. Calcul des images....................................................................................... 105 III.3.1.4. Résultats ..................................................................................................... 105
III.3.2. Images obtenues par microtomographie X (rayonnement synchrotron)... 107III.3.2.1. Microtomographie X utilisant le rayonnement synchrotron ...................... 107
III.3.2.1.1. Le rayonnement synchrotron ......................................................... 108 III.3.2.1.2. Instrumentation pour l’imagerie ................................................... 108 III.3.2.1.3. Avantages de cette méthode de tomographie ................................ 109
III.3.2.2. Les acquisitions.......................................................................................... 109 III.3.2.3. Résultats ..................................................................................................... 110
III.4. Recalage des régions d’intérêt ............................................................ 112
III.4.1. Le problème ..................................................................................................... 113 III.4.2. Repositionnement des cylindres..................................................................... 114
III.4.2.1. Axes des cylindres µIRM et SR-µCT ........................................................ 114 III.4.2.2. Alignement des cylindres µIRM et SR-µCT ............................................. 117 III.4.2.3. Bilan du repositionnement ......................................................................... 117
III.4.2.3.1. Enveloppe cylindrique ................................................................... 117 III.4.2.3.2. Plage de linéarité du repositionnement......................................... 118 III.4.2.3.3. Efficacité du repositionnement ...................................................... 119 III.4.2.3.4. Remarque....................................................................................... 120
III.4.3. Ajustement de la résolution............................................................................ 120 III.4.3.1. Problématique ............................................................................................ 120 III.4.3.2. Dégradation des images SR-µCT............................................................... 121 III.4.3.3. Méthode ..................................................................................................... 121
III.4.4. Recherche de la ROI commune ..................................................................... 123 III.4.4.1. Problématique ............................................................................................ 123 III.4.4.2. ROI utilisée pour le calcul ......................................................................... 123 III.4.4.3. Première approche: méthode du volume « OU exclusif » ......................... 124
III.4.4.3.1. Description de la méthode............................................................. 124 III.4.4.3.2. Limitation de cette méthode........................................................... 126
III.4.4.4. Méthode de la longueur moyenne d’interception ...................................... 126
12

Table des matières
III.4.4.4.1. Image paramétrique « MIL2D » ..................................................... 126 III.4.4.4.2. Détermination de la position de la ROI commune ........................ 127 III.4.4.4.3. Avantage de cette méthode ............................................................ 128
III.4.5. Recalage des ROI: effet sur les paramètres de structure ............................ 128 III.4.5.1. Protocole .................................................................................................... 128 III.4.5.2. Résultats ..................................................................................................... 129
III.4.6. Bilan de la méthode de recalage automatique .............................................. 131 III.4.6.1. Efficacité .................................................................................................... 131 III.4.6.2. Etude des deux échantillons problématiques ............................................. 132
III.4.7. Conclusions et perspectives ............................................................................ 134
III.5. Segmentation des images ..................................................................... 135
III.5.1. Images µIRM: méthode d’étiquetage............................................................ 135 III.5.2. Images par rayonnement synchrotron.......................................................... 137
III.5.2.1. Résolution nominale (10 µm) .................................................................... 137 III.5.2.2. Résolution dégradée: conservation de BV/TV0 ......................................... 138
III.6. Confrontation des modalités d’imagerie............................................ 139
III.6.1. ROI choisie et calcul des paramètres ............................................................ 140 III.6.2. Principe de la comparaison ............................................................................ 142 III.6.3. Résultats ........................................................................................................... 143
III.6.3.1. Fraction osseuse ......................................................................................... 145 III.6.3.2. Topologie et orientation de la microstructure ............................................ 146 III.6.3.3. Histomorphométrie .................................................................................... 147 III.6.3.4. Anisotropie................................................................................................. 148
III.6.4. Paramètres pertinents..................................................................................... 150 III.6.4.1. Lien entre paramètres histomorphométriques et fraction osseuse ............. 150
III.6.4.1.1. Loi phénoménologique .................................................................. 150 III.6.4.1.2. Influence sur la confrontation entre les deux modalités................ 151 III.6.4.1.3. Intérêt des paramètres histomorphométriques .............................. 153
III.6.4.2. Lien entre paramètres d’anisotropie et fraction osseuse ............................ 153 III.6.4.3. Caractéristique d’Euler-Poincaré ............................................................... 154
III.6.5. Etude du biais sur la fraction osseuse ........................................................... 155 III.6.5.1. Effet de la résolution .................................................................................. 155 III.6.5.2. Effet de la méthode de segmentation ......................................................... 158 III.6.5.3. RSB des images µIRM et interpolation « zerofill »................................... 159 III.6.5.4. Premiers résultats sur les effets du T2
*....................................................... 162 III.6.5.4.1. Effet de la bande passante ............................................................. 162 III.6.5.4.2. Simulations .................................................................................... 163 III.6.5.4.3. Conclusions ................................................................................... 166
III.7. Recalage des ROI: effet sur la comparaison entre modalités .......... 167
III.7.1. Méthodes .......................................................................................................... 167 III.7.2. Résultats ........................................................................................................... 168
III.7.2.1. Volume analysé.......................................................................................... 168 III.7.2.2. Erreur relative ............................................................................................ 169 III.7.2.3. Confrontation entre les deux modalités ..................................................... 170 III.7.2.4. Conclusion ................................................................................................. 172
III.8. Conclusions ........................................................................................... 172
13

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Chapitre IV: Imagerie 3D in vivo du doigt humain: faisabilité de l’analyse du tissu trabéculaire in vivo.............................................175
IV.1. IRM du tissu trabéculaire in vivo: état de l’art ................................ 175
IV.1.1. Discrimination entre groupes de patients ..................................................... 175 IV.1.2. Quantification des modifications de l’architecture avec l’âge .................... 177 IV.1.3. Suivi de l’efficacité d’un traitement............................................................... 178 IV.1.4. Limitations de ces études ................................................................................ 179 IV.1.5. Problématique et présentation du travail ..................................................... 179
IV.2. Les images ............................................................................................. 180
IV.2.1. Protocole expérimental ................................................................................... 180 IV.2.2. Paramètres d’acquisition ................................................................................ 181 IV.2.3. Calcul des images ............................................................................................ 182 IV.2.4. Premières analyses .......................................................................................... 183
IV.2.4.1. Sur les images ............................................................................................ 183 IV.2.4.2. Sur l’histogramme...................................................................................... 184 IV.2.4.3. Phalange distale ......................................................................................... 184
IV.3. Délimitation de la ROI......................................................................... 186
IV.3.1. Le problème ..................................................................................................... 186 IV.3.2. Première approche .......................................................................................... 186 IV.3.3. Détermination d’une ROI adaptée par ajustement d’une surface
tridimensionnelle ............................................................................................. 187 IV.3.3.1. Récupération de voxels « frontières »........................................................ 188
IV.3.3.1.1. Analyse des bas niveaux de gris .................................................... 188 IV.3.3.1.2. Analyse des hauts niveaux de gris ................................................. 190 IV.3.3.1.3. Voxels « frontières » ...................................................................... 192
IV.3.3.2. Calcul de l’enveloppe tridimensionnelle ................................................... 194 IV.3.3.2.1. Enveloppe convexe......................................................................... 194 IV.3.3.2.2. Solution retenue: α-forme.............................................................. 195
IV.3.3.3. Extraction de la ROI .................................................................................. 197 IV.3.3.4. Résultats et fiabilité de la méthode ............................................................ 200
IV.3.3.4.1. Région trabéculaire ....................................................................... 200 IV.3.3.4.2. Région corticale............................................................................. 201 IV.3.3.4.3. Fiabilité.......................................................................................... 202
IV.4. Segmentation des images ..................................................................... 203
IV.4.1. Caractérisation du bruit de l’image............................................................... 204 IV.4.2. Application de la méthode d’étiquetage ........................................................ 205 IV.4.3. Application de la méthode de réduction du volume partiel......................... 206
IV.4.3.1. Cartographie de la fraction osseuse ........................................................... 206 IV.4.3.2. Algorithme de subdivision......................................................................... 208
IV.4.3.2.1. Première phase .............................................................................. 208 IV.4.3.2.2. Deuxième phase ............................................................................. 209 IV.4.3.2.3. Illustration sur l’image de la phalange distale.............................. 210
IV.5. Premiers résultats................................................................................. 213
14

Table des matières
IV.5.1. Images étudiées ................................................................................................ 213 IV.5.2. Protocole........................................................................................................... 216 IV.5.3. Volume de données analysé ............................................................................ 216 IV.5.4. Fraction osseuse et caractéristique d’Euler-Poincaré.................................. 217
IV.5.4.1. Résultats..................................................................................................... 217 IV.5.4.2. Influence du RSB sur la segmentation....................................................... 218
IV.5.4.2.1. Protocole........................................................................................ 218 IV.5.4.2.2. Résultats......................................................................................... 219 IV.5.4.2.3. Discussion sur les résultats obtenus in vivo .................................. 220
IV.5.5. Anisotropie et orientation............................................................................... 220 IV.5.6. Premières conclusions ..................................................................................... 221
IV.6. Perspectives........................................................................................... 222
Conclusion .......................................................................................225
Bibliographie ...................................................................................229
Annexes............................................................................................243
Annexe 1: Calcul de la caractéristique d’Euler-Poincaré ......................... 245
Annexe 2: Orientation de la microstructure: calcul des trois angles d’Euler (θ,ϕ,ψ).................................................................................................... 249
Annexe 3: Tracé de segments dans un espace discrétisé............................ 253
Annexe 4: Liste des publications liées à ce travail...................................... 259
15


Introduction
Introduction
L’ostéoporose se traduit par une diminution exagérée de la masse osseuse et une détérioration
de la microarchitecture du tissu osseux entraînant une fragilisation de l’os et donc une
augmentation du risque de fracture. En 2000 en France, la maladie touchait environ 30 à 40%
des femmes ménopausées et plus de la moitié de celles de plus de 75 ans pour un coût direct
estimé à 610 millions d’euros par an.
Le tissu osseux constituant le squelette humain se présente sous deux formes: cortical ou
trabéculaire. Le tissu cortical est un tissu compact qui constitue l’enveloppe résistante de l’os.
Il représente 80% de la masse osseuse du squelette. Le tissu trabéculaire est un tissu poreux
constitué de travées osseuses de quelques centaines de microns d’épaisseur, qui forment un
labyrinthe d’espaces intercommunicants occupés par de la moelle osseuse et des vaisseaux.
Cette microstructure a un rôle mécanique puisqu’elle assure la répartition des charges
extérieures dans le volume de l’os. Le tissu trabéculaire ne représente que 20% de la masse du
squelette mais 80% de la surface d’échange entre l’os et la moelle.
Le tissu osseux se renouvelle tout au long de la vie par cycle périodique de dégradation /
régénération d’une durée de deux à trois mois. Ce processus de remodelage osseux se déroule
au niveau de l’interface entre l’os et la moelle. Le tissu trabéculaire présente l’essentiel de la
surface d’échange entre l’os et la moelle et se renouvelle donc cinq fois plus vite que le tissu
cortical. Il a de plus un rôle mécanique important. Il constitue donc un site privilégié pour
détecter l’ostéoporose.
17

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
La densité minérale osseuse (DMO) est le principal déterminant de la résistance mécanique du
tissu osseux trabéculaire. Cependant, des études in vitro ont montré le rôle de la
microarchitecture trabéculaire: des différences de résistance mécanique ont été observées à
DMO équivalente.
En raison de sa capacité à délivrer des images tridimensionnelles de résolution spatiale élevée
et isotrope et de son innocuité totale, l’IRM est une modalité de choix pour évaluer les
paramètres d’architecture tridimensionnelle du tissu trabéculaire. Cependant la résolution
spatiale, limitée par la faible sensibilité de l’IRM, est, au mieux, du même ordre de grandeur
que l’épaisseur des travées osseuses.
Trois objectifs sont visés par ce travail. Le premier est de mettre en œuvre des outils
permettant la caractérisation de la microarchitecture trabéculaire à partir d’images
tridimensionnelles acquises à une résolution de l’ordre de quelques dizaines de microns. Le
deuxième est de les utiliser dans le cadre d’un protocole in vitro afin d’établir la fiabilité de la
représentation de la microstructure trabéculaire par IRM (comparaison à une technique de
référence: la microtomographie X haute résolution par rayonnement synchrotron). Enfin, le
troisième objectif est de se servir des deux premiers afin de développer un protocole
d’imagerie in vivo.
Ce document est composé de quatre chapitres.
Le premier chapitre décrit la structure osseuse et son principe de régénération. Ensuite est
introduit l’intérêt de l’étude du tissu trabéculaire afin de caractériser l’état pathologique de
l’os. Enfin, les principales techniques de caractérisation disponibles à l’heure actuelle sont
succinctement présentées, et plus particulièrement l’IRM.
Le deuxième chapitre présente les paramètres de caractérisation de la microstructure osseuse
que nous avons utilisés dans le cadre de cette étude. Après avoir introduit les problèmes liés à
la segmentation des images, les concepts, les définitions et les méthodes de calcul
tridimensionnel (validées sur des images « test ») de chacune des trois familles de paramètres
envisagées, en plus de la fraction osseuse, sont détaillés: topologie (connectivité, nombre
d’amas déconnectés, nombre de surfaces internes), histomorphométrie (surface sur volume,
espacement et séparation des travées osseuses), et orientation (trois angles d’Euler) et
18

Introduction
anisotropie (deux coeffients caractérisant l’anisotropie maximale et minimale de la
microarchitecture).
Le troisième chapitre correspond à une étude de la validité de la technique IRM pour la
caractérisation de la microstructure trabéculaire. La méthode de référence était la
microtomographie X par rayonnement synchrotron. Cette modalité d’imagerie a déjà
démontré sa fiabilité par rapport à l’histomorphométrie (technique de référence pour
caractériser la structure du tissu trabéculaire) du fait qu’elle permet directement une
évaluation tridimensionnelle de la microarchitecture et de la densité osseuse. Cette étude a été
menée sur 29 échantillons de calcanéum humain (os du talon). Pour l’ensemble des
échantillons, des images 3D acquises par microscopie IRM (images à 66 µm de résolution
isotrope, réalisées sur un microscope RMN à 8.5 T, prototype développé au laboratoire)
étaient disponibles ainsi que des images 3D acquises par microtomographie X haute
résolution par rayonnement synchrotron (images à 10 µm de résolution isotrope, réalisées
avec Madame PEYRIN, en collaboration avec l’ESRF à Grenoble et CREATIS à Lyon). Nous
avons comparé les valeurs des paramètres d’architecture calculés sur les données IRM aux
mêmes paramètres évalués sur les données de microtomographie X. Avant tout, il nous a donc
fallu développer une méthode de recalage automatique des régions d’intérêt entre les deux
modalités afin d’effectuer les comparaisons sur des volumes identiques, à même résolution et
dans la même position.
Le dernier chapitre est une étude de faisabilité de la caractérisation du tissu trabéculaire in
vivo. Nous avons obtenu des images de l’articulation distale du doigt in vivo sur un imageur à
7 T, avec une résolution isotrope de 78 µm. Nous présentons dans ce chapitre, la méthode que
nous avons développée pour isoler automatiquement la région d’intérêt (tissu trabéculaire) des
autres tissus visibles sur ce type d’image (tissu cortical, tendons, peau, vaisseaux
sanguins,…). Nous comparons ensuite deux algorithmes de segmentation à partir des
premières valeurs des paramètres calculés sur une ROI isolée automatiquement.
L’ensemble des algorithmes ont été développés sur une station de travail SUN (400 MHz,
1 Go de RAM, carte graphique 3D), en langage C, sous l’environnement du logiciel
« Advanced Visual Systems » (AVS, Waltham, Ma).
19

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Ce travail de thèse s’inscrit dans le cadre du Groupement de Recherche (GDR) 2237:
« Imagerie et caractérisation tissulaire appliquées au domaine ostéoarticulaire ». Les
collaborations principales au sein de ce GDR ont été menées avec trois équipes lyonnaises:
CREATIS (et en particulier avec Madame Peyrin de l’ESRF de Grenoble), LRMN et LEIBO.
De plus, dans le cadre du programme d’échanges entre laboratoires français et étrangers
proposé par Egide pour le compte du Ministère des affaires étrangères (programme
« Procope »), nous avons pu développer une collaboration avec le laboratoire
« Experimentelle Physik V » (EP5) de l’Université de Würzburg en Allemagne.
20

Chapitre I: Le tissu osseux et sa caractérisation
Chapitre I: Le tissu osseux et sa
caractérisation
Dans les deux premiers paragraphes de ce chapitre, nous décrivons le tissu osseux et les
principes de sa régénération. Au paragraphe 3, nous abordons la principale pathologie
dégénérative de l’os, l’ostéoporose, et l’intérêt de l’étude du tissu trabéculaire pour la
détecter. Le rôle important de la microarchitecture de ce tissu sur les caractéristiques
mécaniques de l’os est ensuite présentée au paragraphe 4. Ces quatre parties ont été rédigées
sur la base de trois références bibliographiques: un dossier de la Fondation pour la Recherche
Médicale ([Dupuy, 2000]), un texte proposé dans INSERM, Expertise collective
([Inserm, 1996]), et le « Handbook » sur l’os de Cowin ([Cowin, 2001]). Enfin, les principales
techniques de caractérisation du tissu osseux sont décrites au paragraphe 5 et plus
particulièrement la technique d’Imagerie par Résonance Magnétique (IRM) au paragraphe 6.
I.1. Description générale de la structure osseuse
L’os est composé pour 65% de petits cristaux minéraux inorganiques (essentiellement
hydroxyapatite de calcium) incorporés dans une matrice organique (35%). Cette matrice (où
se situe le calcium) est composée principalement de collagène de type I (90%) et de protéines
non collagéniques (10%). A l’échelle microscopique, on distingue deux types de tissus osseux
(cf. Figure I.1).
21

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
22
Le tissu cortical, souvent appelé « os cortical », est un tissu compact qui constitue l’enveloppe
résistante de l’os. Il forme un cylindre creux, la cavité médullaire, qui est remplie de moelle
osseuse. Le tissu cortical est constitué d’une juxtaposition de cylindres faits de lamelles
osseuses concentriques dans lesquels les fibres de collagènes sont orientées de manière à lui
donner une résistance mécanique optimale. Au centre et autour de chaque cylindre se trouvent
des capillaires innervés permettant la vascularisation de la moelle osseuse. Ce type de tissu est
situé au niveau de la diaphyse des os longs et entoure les os plats. En terme de masse osseuse,
il constitue 80% du squelette, 50% des vertèbres, 70% du col du fémur.
Le tissu trabéculaire, souvent appelé « os trabéculaire » ou « os spongieux », est un tissu
poreux constitué de travées osseuses de quelques centaines de microns d’épaisseur, qui
forment un labyrinthe d’espaces intercommunicants occupés par de la moelle osseuse et des
vaisseaux. Il est situé au niveau de l’épiphyse des os longs ainsi que dans la partie centrale des
os plats. L’architecture des travées, qui est liée aux forces de pression et de traction
auxquelles est soumis le tissu trabéculaire, permet de distribuer les charges extérieures dans le
volume de l’os. En terme de masse osseuse, il constitue 20% du squelette, 50% des vertèbres,
30% du col du fémur.
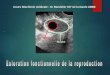
Figure I.1: Coupe d’un os long. La diaphyse est la zone centrale de l’os. Elle est constituée d’un tissu
osseux compact dit « cortical » formant un cylindre creux, la cavité médullaire, qui est remplie de moelle
osseuse. L’épiphyse correspond à l’extrémité de l’os. Elle est principalement constituée d’un tissu
spongieux dit « trabéculaire », dont les pores sont remplis de moelle osseuse (image du polycopié de
DCEM2 du Docteur Lafage-Proust de l’Université de Saint Etienne).

Chapitre I: Le tissu osseux et sa caractérisation
I.2. Régénération osseuse
L’os se régénère tout au long de la vie au cours de cycles périodiques constitués de deux
phases. Dans un premier temps, des cellules appelées « ostéoclastes » dégradent le tissu
osseux. Des facteurs de croissances contenus dans la matrice osseuse se trouvent alors libérés
stimulant des cellules appelées « ostéoblastes » qui synthétisent du tissu osseux. Ce processus
de remodelage se déroule au niveau de l’interface entre l’os et la moelle, au sein d’unités de
remodelage d’environ 100 µm de diamètre indépendantes les unes des autres dans leur
localisation et dans leur activation.
A l'âge adulte, la phase de résorption osseuse dure de deux à trois semaines, celle de
formation environ trois mois. La balance osseuse (différence entre la quantité d’os synthétisé
et résorbé) est équilibrée jusqu'à 30 ans environ: on observe alors un « pic » de masse osseuse
(plus important chez la femme que chez l’homme). Ensuite, la masse osseuse diminue
lentement, aboutissant à un léger déficit de l'ordre de 3 à 5% tous les dix ans.
I.3. L’ostéoporose
La carence de certaines hormones (en particulier chez la femme, à partir de la ménopause)
peut provoquer non seulement un excès de tissu résorbé à chaque cycle, mais aussi une
augmentation de la fréquence du remodelage osseux. Ce processus peut entraîner une perte de
masse osseuse pouvant atteindre, à 80 ans, 25% chez l’homme, 40% chez la femme.
L’ostéoporose est le résultat d’une balance osseuse exagérément négative. Selon
l’Organisation Mondiale pour la Santé (OMS), cette maladie est caractérisée par une
diminution de la masse osseuse et par la détérioration de la microarchitecture du tissu osseux
ce qui fragilise l’os et augmente donc le risque de fracture ([Consensus, 1993]). L'ostéoporose
est une pathologie dégénérative du tissu osseux étroitement liée au cycle de vie du squelette.
L'expression clinique la plus fréquente de l'ostéoporose est la fracture qui, par les
complications qu’elle entraîne (entrée en dépendance, déformations, infirmités,…) fait de
cette pathologie un enjeu majeur de santé publique. En 2000 en France, la maladie touchait
environ 30 à 40 % des femmes ménopausées et plus de la moitié de celles de plus de 75 ans.
Les trois sites les plus affectés sont l’extrémité supérieure du fémur (19% des fractures, 50000
23

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
24
nouveaux cas chaque année), les vertèbres (16% des fractures, 50000 nouveaux cas chaque
année) et l’extrémité distale du radius (14% des fractures, 35000 nouveaux cas chaque année).
Le coût direct en 2000 a été chiffré à environ 610 millions d’euros par an.
L’ostéoporose peut toucher le tissu cortical par son amincissement et par une augmentation de
sa porosité (cf. Figure I.2, A), et le tissu trabéculaire par un amincissement des travées
osseuses entraînant des déconnections de la microstructure (cf. Figure I.2, B).
≈ 5 mm ≈ 3 mm
A B Figure I.2: A: Différences observables sur le tissu cortical entre un cas « jeune » en haut et « âgé » en bas
(coupes d’une côte d’un chien de race « beagle » issues de [Cowin, 2001]). B: Même chose pour le tissu
trabéculaire: le cas « jeune » est à gauche, le cas « âgé » à droite (photographies de tissu trabéculaire
humain issues du polycopié de DCEM2 du Docteur L fage-Proust de l’Université de Saint Etienne). a
Bien que le tissu cortical soit prépondérant en terme de masse osseuse (80% contre 20% pour
le tissu trabéculaire), il ne représente que 20% de la surface d’échange entre l’os et la moelle
(contre 80% pour le tissu trabéculaire). Le tissu trabéculaire se renouvelle cinq fois plus
rapidement que le tissu cortical. Du fait de son rôle mécanique, il constitue donc un site
privilégié pour l’étude de pathologies osseuses de type ostéoporose.
I.4. Rôle de la microarchitecture du tissu trabéculaire
Le squelette étant la charpente du corps humain, les paramètres pertinents pour l’étude du
tissu osseux sont de type mécanique (principalement la résistance et l’élasticité de l’os). On
peut d’ailleurs admettre que le diagnostic de l’ostéoporose se déduit du risque de fracture

Chapitre I: Le tissu osseux et sa caractérisation
donc des indices mécaniques de l’os. De nombreuses études ont cherché à les caractériser soit
directement, soit par l’étude de relations avec d’autres types de paramètres.
Le principal déterminant des caractéristiques mécaniques est la Densité Minérale Osseuse
(DMO). D’ailleurs, l’OMS définit l’état ostéoporotique d’un individu par la valeur de DMO.
A partir d’une statistique faite sur une population de même sexe, jeune et bien portante
(établie pour un site anatomique donné), on définit le « T-score » comme l’écart séparant la
valeur de DMO mesurée à la valeur moyenne de la statistique. Cet écart, exprimé en nombre
d’écart type SD de cette statistique, traduit une perte osseuse (le T-score est négatif). Quatre
états du squelette sont alors définis: « normal » si T-score > -SD, « faible masse osseuse » si
-2.5×SD < T-score < -SD, « ostéoporose », si T-score < -2.5×SD, « ostéoporose sévère » si
T-score < -2.5 et que le patient a déjà eu une fracture.
Cependant, les nombreuses études sur les relations entre la DMO et les caractéristiques
mécaniques de l’os ont montré une dispersion importante des résultats, traduisant le rôle non
négligeable de la microarchitecture trabéculaire. Kleerekoper et al. ([Kleerekoper, 1985]) ont
montré qu’ils ne pouvaient séparer sans ambiguïté un groupe de patients sains d’un groupe de
patients pathologiques en ne considérant que la DMO. Dans une autre étude ([Rice, 1988]),
une loi phénoménologique reliant le module d’Young au carré de la DMO a été établie mais
avec une grande variabilité des données. Plus récemment ([Uchiyama, 1999]), l’analyse de
prélèvements effectués sur des vertèbres (lombaires) a montré que les corrélations entre les
propriétés mécaniques du tissu trabéculaire et la DMO étaient situées entre 0.55 et 0.60. Des
simulations sur modèle 3D de différents scénarios de perte osseuse ([Van Der Linden, 2001],
[Guo, 2002]) ont aussi montré que dans certains cas (en particulier dans le cas
d’amincissement des travées), les effets sur la rigidité de l’os étaient relativement réduits.
Dans une étude comparant différents sites trabéculaires, Morgan et al. ([Morgan, 2003]) ont
pu observer une forte dépendance des relations entre le module d’Young et la densité osseuse
avec le site analysé. Les auteurs ont montré que la cause principale de ces variations était la
différence d’architecture du tissu trabéculaire (les tissus osseux étant soumis à des contraintes
mécaniques qui sont propres au site analysé).
Ces résultats confirment que les caractéristiques de la microstructure du tissu trabéculaire
doivent être prises en compte pour caractériser les attributs mécaniques du tissu osseux. C’est
pourquoi quelques tentatives ont été menées pour trouver un modèle à la structure trabéculaire
25

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
afin de mieux comprendre son comportement et de faciliter son analyse. Par exemple, Gibson
([Gibson, 1985]) a proposé un modèle en fonction de la DMO. La microstructure pourrait être
représentée par un réseau de plaques interconnectées si la DMO est élevée (tissu osseux
soumis à une forte contrainte mécanique). Dans le cas de DMO plus faible (tissu osseux
soumis à une contrainte mécanique moindre), elle serait modélisée par des cylindres
interconnectés. Pour les DMO intermédiaires, le modèle serait une combinaison des deux
précédents. Dernièrement ([Kim, 2002]), un modèle adapté à l’étude du tissu trabéculaire des
vertèbres a aussi été proposé.
I.5. Techniques de caractérisation
Dans cette partie, les principales techniques de caractérisation du tissu trabéculaire sont
décrites, mise à part l’IRM qui fait l’objet du paragraphe suivant (cf. I.6). Nous nous sommes
principalement basés sur un article de 1996 présentant un état de l’art de ces techniques
([Genant, 1996]).
I.5.1. Absorptiométrie
Cette technique est basée sur la mesure de l’atténuation d’un faisceau de photons (émis
généralement par une source de rayons X) résultant de leurs interactions avec la matière.
L’image de projection obtenue correspond à une intégration sur toute l’épaisseur de la zone à
analyser (cf. Figure I.3). Le contraste dans l’image est donné par l’atténuation du faisceau qui
est beaucoup plus sensible dans le tissu osseux que dans les tissus mous: plus la matière
atténue les rayons X, plus le pixel correspondant sera caractérisé par un fort signal. Le tissu
osseux apparaît donc en blanc sur l’image.
26

Chapitre I: Le tissu osseux et sa caractérisation
27
Source (X)
Echantillon
Capteur (film, CCD)
Figure I.3: Principe de l’absorptiométrie.
Du fait de la nature de la mesure, cette technique permet de déterminer la DMO surfacique,
exprimée en g.cm-2.
Il existe deux types de mesures: mono-énergétique (« Single X-ray Absorptiometry »: SXA)
ou bi-énergétique (« Dual X-ray Absorptiometry »: DXA). Cette dernière technique consiste à
acquérir deux projections pour deux longueurs d’onde différentes. Ceci permet de calculer
une image où l’atténuation du faisceau due à la présence d’os (qui constitue l’information
pertinente) est séparée de celle produite lors de la traversée des tissus mous avoisinants. La
DXA est donc particulièrement bien adaptée aux études in vivo où on a souvent une épaisseur
importante de tissus mous sur le tissu osseux à analyser. Elle est d’ailleurs considérée comme
la méthode « Gold Standard » pour l’évaluation de la DMO in vivo et constitue l’examen
principal pour diagnostiquer l’ostéoporose aujourd’hui (cf. I.4).
Un exemple d’examen DXA effectuée sur un échantillon de mandibule humaine ex-vivo est
donné en Figure I.4.
≈ 25
mm
Dent
Tissu cortical
Tissu trabéculaire
Figure I.4: Image DXA d’un échantillon de mandibule humaine. On peut clairement distinguer
l’enveloppe corticale (ce qui est rarement le cas sur les acquisitions in vivo) de la mâchoire et la dent. Par
contre, aucune microarchitecture n’est décelable dans la zone trabéculaire (image: Docteur Choel,
LEIBO - LRMN, Lyon).

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
28
Le principal inconvénient de cette méthode est qu’elle ne permet pas d’obtenir des
informations sur la microarchitecture du tissu trabéculaire (le caractère ionisant de la méthode
limite la résolution à quelques centaines de microns). De plus, la séparation entre les tissus
cortical et trabéculaire est en général impossible. Enfin, la DMO calculée caractérise une
densité de surface puisque le contenu osseux mesuré provient de la projection de tout le
volume. Une étude récente ([Wren, 2000]) sur le tissu trabéculaire du calcanéum (os du talon)
a cependant proposé une formulation permettant d’estimer une fraction osseuse volumique à
partir de la DMO mesurée et de la surface projetée.
I.5.2. Histomorphométrie osseuse
Cette technique consiste en une biopsie de la crête iliaque (partie supérieure de l’os de la
hanche). La Figure I.5 présente la position du patient et l’instrument pour réaliser cet acte qui
est généralement fait sous anesthésie locale. La carotte osseuse prélevée (environ 8 mm de
diamètre) est déshydratée puis incluse dans une résine plastique. Des sections de quelques
microns d’épaisseur sont ensuite réalisées avec un microtome spécial. Un prélèvement
complet présente les deux corticales encadrant le tissu trabéculaire. En fonction de ce que l’on
souhaite observer (cellules, tissu calcifié,…), les coupes sont colorées avec un colorant
adapté.
Figure I.5: Représentation de la biopsie de la crête iliaque. La carotte osseuse que l’on récupère est
d’environ 8 mm de diamètre (image: polycopié de DCEM2 du Docteur Lafage-Proust de l’Université de
Saint Etienne).
L’analyse de ces prélèvements permet d’évaluer avec une excellente résolution spatiale
(échelle du micron) des paramètres reflétant l’architecture du réseau trabéculaire

Chapitre I: Le tissu osseux et sa caractérisation
([Hordon, 2000]). Ce prélèvement donne de plus l’accès à des informations biochimiques très
fines sur la capacité de remodelage du tissu osseux, lié à l’équilibre entre ostéoclastes et
ostéoblastes ([Boivin, 2000]).
Le principal inconvénient de cette technique est son caractère invasif. De plus, l’analyse de
paramètres d’architecture tridimensionnels est difficile à mettre en œuvre car elle nécessite
une reconstruction 3D préalable à partir de coupes qui sont forcément non jointives
([Thomsen, 1996]). Enfin, le site analysé n’est pas particulièrement exposé aux fractures. Les
études d’histomorphométrie osseuse concernant d’autres sites plus pertinents (comme la
vertèbre, cf. [Thomsen, 2002 (a)]) ne peuvent être menées qu’à partir d’échantillons.
I.5.3. Ultrasons
Le principe des techniques ultrasonores est le calcul de paramètres qui dépendent de la densité
osseuse ou de la structure du réseau. Par exemple, la vitesse de propagation du son
(« Ultrasound Bone Velocity », UBV) traversant l’os sera plus élevée dans le tissu cortical
que dans le trabéculaire ou encore l’atténuation des ondes ultrasonores pour une gamme de
fréquence donnée (« Broadband Ultrasound Attenuation » BUA) sera d’autant moins forte
que le tissu est compact (donc dans le tissu cortical). Les images obtenues sont des cartes de
ces paramètres mesurés point par point en déplaçant les transducteurs d’émission et de
réception des ondes ultrasonores. Ce sont donc des images de projections.
Plusieurs études ont déjà montré les liens des paramètres ultrasonores avec la DMO
([Chappard, 1997], [Laugier, 1997]) et avec la microarchitecture ([Nicholson, 2001]).
Actuellement, cette technique devient une alternative intéressante à la DXA (cf. I.5.1):
méthode non ionisante, facilité de mise en œuvre, appareillage peu coûteux pouvant être
portatif, rapidité…
Le principal inconvénient des ultrasons est que seules les extrémités (calcanéum, doigt,
radius) peuvent être étudiées. De plus, l’analyse de la microarchitecture est encore
problématique.
29

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
I.5.4. Tomographie par rayons X
La tomographie par rayons X (« quantitative Computed Tomography », qCT) est basée sur les
mêmes principes physiques que l’absorptiométrie (cf. Figure I.3). Elle consiste à mesurer
l’atténuation d’un faisceau de rayons X monochromatique sur toute une série d’incidences.
Pour chaque incidence, on mesure le nombre N de photons reçus en transmission (les autres
étant absorbés ou diffusés par la matière) pour chaque point du plan de projection à l’aide
d’un capteur CCD. Si N0 est le nombre de photons émis par la source, on montre que le
logarithme du rapport N0/N (qui est fonction de la position dans le plan de projection)
correspond à une intégrale sur l’épaisseur de l’échantillon du coefficient d’atténuation linéaire
du matériau. Radon a montré en 1917 qu’il était possible de reconstruire, à partir des
projections, une distribution spatiale du coefficient d’atténuation linéaire. L’image
reconstruite en tomographie correspond donc à cette distribution. Les algorithmes de
reconstruction proposés sont nombreux. Ils nécessitent généralement une grosse puissance de
calcul.
Il existe une large gamme de tomographes et microtomographes en fonction:
• des propriétés de la source de rayons X utilisée: très focalisée pour un mode point par
point, en éventail ou en cône pour imager une zone plus importante
• de la mécanique pour positionner la zone à imager, le détecteur et la source X
• du type de détecteur: 1D ou 2D, en ligne ou en couronne (ce qui évite les rotations du
capteur puisque à chaque angle correspond un pixel),…
La qualité des images (en terme de résolution spatiale, de rapport signal à bruit et d’artefact)
est limitée principalement par:
• le flux de la source X: s’il est insuffisant, le rapport signal à bruit sera insuffisant ou le
temps de pose deviendra prohibitif
• la cohérence de la source X: la valeur du coefficient d’atténuation dépend de l’énergie de
la source donc de sa longueur d’onde. Si la source est peu cohérente, on obtient une
moyenne des coefficients d’atténuation. La mesure est alors moins précise ce qui peut
causer des artefacts dans l’image
• le détecteur utilisé (CCD): et en particulier sa sensibilité, le bruit qu’il génère (par
exemple: courant d’obscurité), la taille du pixel, l’optique associée,…
De nombreux détails sur les différents algorithmes de reconstruction et les différents types de
tomographe sont donnés dans la Thèse de Pateyron ([Pateyron, 1998]).
30

Chapitre I: Le tissu osseux et sa caractérisation
Les images obtenues par cette méthode d’imagerie donnent une répartition volumique de la
DMO ce qui permet de mesurer très précisément la fraction volumique d’os ([Ding, 1999]),
mais aussi de disposer d’une représentation tridimensionnelle fiable: le réseau trabéculaire,
qui est séparable du tissu cortical, peut alors être analysé (si la résolution spatiale est
suffisante). De nombreuses études ont d’ailleurs montré qu’à partir d’images de
microtomographie X haute résolution de biopsies de crête iliaque ([Müeller, 1998],
[Nuzzo, 2002]) ou de vertèbre ([Banse, 2002]), les évaluations de la DMO et de la structure
du tissu trabéculaire étaient aussi fiables que celles effectuées par histomorphométrie (cf.
I.5.2).
Le principal inconvénient de cette technique d’imagerie est son caractère ionisant ce qui
limite la résolution spatiale accessible in vivo à quelques centaines de microns.
I.6. La technique IRM
En raison de sa capacité à délivrer des images tridimensionnelles de résolution spatiale élevée
et isotrope et de son innocuité totale, l’Imagerie par Résonance Magnétique (IRM) est une
modalité particulièrement intéressante pour étudier la microarchitecture trabéculaire. Dans un
premier temps, nous décrivons les principes physiques de la Résonance Magnétique Nucléaire
(RMN), phénomène lié à des propriétés quantiques de certains noyaux. Ensuite, nous
présentons la technique d’imagerie proprement dite.
La plupart des applications médicales de l’IRM concernent les noyaux d’hydrogène présents
en grande quantité dans le corps humain (eau, molécules organiques hydrogénées comme la
graisse,…). Dans cette partie, nous ne traiterons que de lui. Il faut cependant savoir que
d’autres noyaux sont également étudiés par RMN (principalement en spectroscopie mais aussi
en imagerie pour certains): hélium, carbone, fluor, sodium, phosphore,…
Les ressources bibliographiques sur la présentation de l’IRM sont nombreuses. Pour ce qui est
des concepts, nous nous sommes basés sur quatre livres: [Alaux, 1994], [Farrar, 1971],
[Chen, 1989] et [Haacke, 1999]. De plus, certaines illustrations ont été reprises de la thèse de
31

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Bittoun ([Bittoun, 1987]), d’une documentation de Schering ([Schild, 1990]) et de la thèse de
médecine de Durand ([Durand, 1997]).
I.6.1. Phénomène de la RMN
I.6.1.1. Effet d’un champ magnétique statique sur les protons
Chaque noyau d’hydrogène est constitué d’un proton doté d’un spin ½ à l’origine d’un
moment magnétique. En l’absence de champ magnétique externe, chaque moment
magnétique a une direction propre. Par contre, si un champ magnétique statique 0Br
est
appliqué, deux effets se produisent:
• les spins se regroupent sur deux niveaux d’énergie: le plus faible correspondant à un
moment magnétique parallèle à 0Br
, le plus élevé correspondant à une direction
antiparallèle à 0Br
• les spins précessent: les moments magnétiques des protons subissent un mouvement de
rotation autour de la direction du champ à une pulsation ω0 dite « pulsation de Larmor »
ou pulsation propre. Celle-ci est directement proportionnelle à l’intensité du champ
statique:
ω0 = γB0,
où γ est le rapport gyromagnétique qui caractérise chaque noyau. Pour le proton, on a
γ/2π = 42.5 MHz.T-1.
La séparation d’énergie ∆E entre ces deux niveaux est égale à:
∆E = 2µB0,
où µ est le moment magnétique nucléaire. Dans le cas du proton, on a:
µ = ½γħ,
où ħ = h/2π avec h, la constante de Planck.
A l’équilibre thermique, le système présente un excès de protons dans l’état de faible énergie
dans la proportion P suivante:
kTµBP 4
0= ,
32

Chapitre I: Le tissu osseux et sa caractérisation
où k est la constante de Boltzmann et T la température. A température ambiante (27°) et pour
B0 = 1 T, P est de l’ordre de 10-6: bien que très faible, une aimantation 0Mr
alignée avec 0Br
apparaît au niveau macroscopique:
kTBNµ
PNM 40
2
0
rr
== µ ,
où N est le nombre de protons.
Lorsque cet état d’équilibre est perturbé, l’aimantation Mr
change de direction conformément
à l’équation de mouvement suivante (mécanique semi classique):
BMdtMd rrr
∧=γ Eq. I.1
où Br
est le champ magnétique total appliqué au système. D’après cette équation, si 0BBrr
=
après la perturbation, on observe alors un mouvement de précession autour de 0Br
, à la
pulsation ω0.
I.6.1.2. Perturbation radio-fréquence (RF)
Mr
porte l’information utile: son module est proportionnel à la quantité de protons de
l’échantillon. Il est cependant de très faible intensité. Pour pouvoir détecter Mr
, on cherche à
observer son mouvement de précession ce qui peut être fait à partir de l’analyse de ses
composantes fréquentielles.
Afin d’obtenir un mouvement de précession de l’aimantation, on perturbe le système par un
champ magnétique 1Br
radio-fréquence (« RF ») perpendiculaire à 0Br
. D’après Eq. I.1, on
peut montrer qu’à la fin de l’excitation RF, on obtient un « basculement » de Mr
d’un angle α
(dit « angle de basculement »):
α = γB1τ,
où B1 est l’intensité de la perturbation et τ, son temps d’application.
Les effets de cette perturbation RF peuvent être interprétés au niveau des spins:
• elle donne l’énergie suffisante à certains spins pour passer au niveau d’énergie supérieur
(antiparallèle à 0Br
) entraînant une diminution de Mz, la composante longitudinale de Mr
• elle met en phase les spins ce qui fait apparaître une composante transversale pour
l’aimantation, Mt, qui précesse à ω0.
33

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
34
La Figure I.6 présente la configuration obtenue dans un repère (O,x,y,z) tournant à la
pulsation ω0 autour de (Oz), après application d’un champ RF 1Br
dans la direction (Ox)
pendant un temps τ (l’axe (Oz) correspond à la direction du champ statique 0Br
).
0Br
Mr
zMr
tMr
x
y
z
α
1Br
Figure I.6: Représentation, dans un repère tournant autour de (Oz) à la pulsation ω0, du basculement de
Mr
1Br
obtenu après application d’un champ radio-fréquence selon l’axe (Ox). Deux composantes
apparaissent: une longitudinale Mz (dans la direction de 0Br
) et une transversale Mt qui précesse à ω0.
L’angle de basculement α est directement contrôlé par la perturbation RF. Le signal que l’on cherche à
récupérer est Mt.
C’est donc Mt qui « fournit » le signal RMN.
Remarques:
a) On précise généralement, en même temps que l’angle α, l’axe du repère tournant selon
lequel on applique 1Br
(i.e. la phase de 1Br
: x ou y) afin de connaître le « sens » du
basculement. Dans le cas de la Figure I.6, on écrirait « αx ».
b) Deux valeurs de α sont généralement privilégiées: 90° (pour basculer toute l’aimantation
dans le plan transversal) et 180° (afin d’effectuer un « renversement » de l’aimantation
comme nous le verrons en I.6.2.2).
I.6.1.3. Relaxation
Une fois que le système a été perturbé (i.e. à la fin de l’application de la RF), il revient
progressivement à son état d’équilibre: il y a « relaxation ». Des phénomènes physiques
complexes agissant en parallèle sur les composantes longitudinales et transversales de
l’aimantation en sont à l’origine. Une modélisation simple de ce retour à l’état initial a été

Chapitre I: Le tissu osseux et sa caractérisation
établie, tout à fait acceptable dans de nombreuses applications. L’état d’équilibre est atteint
par deux phénomènes simultanés:
• croissance de la composante longitudinale avec une constate de temps notée T1 (dite temps
de relaxation « longitudinale »). Cette augmentation est liée au retour de spins passés au
niveau d’énergie supérieur vers leur état initial. Ainsi, si on suppose que l’angle de
basculement est de 90°, on aurait:
)1()( 1/0
Ttz eMtM −−=
• décroissance de la composante transversale avec une constate de temps notée T2 (dite
temps de relaxation « transversale »). Cette diminution est liée au déphasage des spins (qui
avaient été mis en phase par la RF). Ainsi, si on suppose que l’angle de basculement est de
90°, on aurait:
2/0)( Tt
t eMtM −=
On peut montrer ([Farrar, 1971]) que T1 (resp. T2) est d’autant plus courte que la densité
spectrale d’énergie (des signaux émis par le système) à ω0 (resp. aux pulsations faibles) est
élevée. Ces constantes de temps de relaxation sont donc fortement liées à l’environnement
électronique et moléculaires des protons: elles permettent donc de caractériser différentes
populations. Par exemple, les valeurs de T1 et T2 associées à des protons situés dans de la
graisse seront inférieures à celles des protons situés dans de l’eau pure. Pratiquement, pour les
tissus mous, T1 varie entre 300 et 2000 ms et T2 entre 30 et 150 ms (pour B0 = 1 T). Pour des
solides, T1 est très grande et T2 très courte.
Les relaxations T1 et T2 agissant simultanément, le retour à l’état d’équilibre de l’aimantation
Mr
se fait par un mouvement spiralé (cf. Figure I.7).
35

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
36
Mr
Figure I.7: Retour à l’état d’équilibre de Mr
. Comme les deux constantes de temps de relaxation T1 et T2
agissent simultanément, le retour (à 0Mr
) se fait par un mouvement spiralé (image: E. Durand, U2R2M,
Orsay).
I.6.2. Le signal de précession libre (FID)
Afin de récupérer la composante transversale Mt, on utilise un circuit résonnant (sonde) placé
dans le plan transversal. On peut ainsi mesurer une tension qui correspond à la force
électromotrice induite par la variation de flux magnétique due au mouvement de . tMr
Le signal recueilli s’appelle « signal de précession libre » plus connu sous le terme anglais de
« Free Induction Decay » (FID). La réception s’effectue sur deux voies correspondant aux
parties réelle et imaginaire du signal (après démodulation en quadrature de phase). Un
exemple de FID est donné sur la Figure I.8.
Figure I.8: Signal de précession libre (« FID »). Ce signal correspond à la force électromotrice induite
dans un circuit résonnant par le mouvement de tMr
. La sinusoïde amortie a une pseudo-période
correspondant à la pulsation propre ω0. La constante de temps de l’enveloppe exponentielle est liée à la
relaxation du système. On l’étudie plus en détail dans la partie I.6.2.2 (image: E. Durand, U2R2M, Orsay).

Chapitre I: Le tissu osseux et sa caractérisation
L’ordre de grandeur de l’amplitude ξ de la FID peut être estimé à partir de l’expression
proposée par Chen et Hoult ([Chen, 1989]) pour un angle de basculement de 90°:
VMIB ∆= 0
10ωξ ,
où IB1 est l’amplitude du champ magnétique RF émis par unité de courant (coefficient
d’induction de la sonde) et ∆V le volume d’échantillon. Pour 1 cm3 d’eau placé dans un
champ statique B0 de 1 T (ω0/2π = 42.5 MHz et M0 ≈ 3.3×10-9 A.m-1) et une boucle
d’émission de 2 cm de diamètre ( IB1 ≈ 2.2×10-5 T.A-1), on a ξ ≈ 20 pV.
L’amplitude de la FID est liée à l’angle de basculement mais aussi à la relaxation du système.
Il est donc important de noter que le contraste obtenu par IRM n’est pas lié uniquement à la
densité de protons: les caractéristiques intrinsèques de chaque tissu (i.e. T1 et T2) peuvent
l’influencer sensiblement. Dans la suite, nous présentons succinctement l’origine du contraste
en IRM à partir de l’exemple d’un échantillon contenant, en proportion identique, deux
populations de protons A et B, caractérisées par des constantes de relaxation différentes: on
supposera T1A < T1
B et T2A < T2
B (la population A pourrait être des protons de la graisse et la
population B de l’eau pure).
I.6.2.1. Contraste en T1
Supposons que l’on envoie, sur l’échantillon présenté ci-dessus, deux RF correspondant à un
angle de basculement de 90°, chacune séparée d’un temps TR. Au moment de la nouvelle RF,
l’aimantation basculée n’est plus M0 mais l’aimantation longitudinale qui est réapparue après
un temps TR soit respectivement:
)1()( 1/0
AR TT
RAz eMTM −−= et )1()( 1/
0B
R TTR
Bz eMTM −−=
pour les familles A et B.
Si TR est court, la différenciation des deux espèces sera facilitée. On dit qu’on obtient un
signal « pondéré T1 » i.e. le paramètre prédominant dans le contraste entre les deux
populations est la valeur de T1. Par contre, si TR est trop long, l’aimantation longitudinale tend
vers M0 pour les deux populations et la distinction devient impossible. Ceci est illustré sur la
Figure I.9.
37

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
38
Intensité
Temps
)1( 1/0
ATteM −−
M0
TR court TR long
avec T B > T A)1( 1/
0BTteM −−
1 1
Figure I.9: Intensité de l’aimantation disponible en fonction de TR. On distinguera d’autant mieux les
protons de la population A de ceux de la B si TR est court. Pour les deux populations, on retrouve, pour TR
assez long, l’aimantation M0 initiale.
I.6.2.2. Contraste en T2
Pour obtenir une pondération en T2, il faut utiliser une RF supplémentaire de 180° avec une
phase en quadrature par rapport à la première perturbation (par exemple 180°y si on avait pris
90°x pour la première RF). Cette perturbation supplémentaire effectue un « retournement »
des spins (cf. Eq. I.1) ce qui permet de les remettre en phase. Une nouvelle FID apparaît: son
amplitude maximale est appelé « écho ». Ce protocole, indiqué en Figure I.10, porte le nom
d’« écho de spins ».
TE/2 RF 90°x RF 180°y TE/2
B D A
Sens de la précession
C Figure I.10: Echo de spins. Au moment de la RF 90°x (A), les spins sont en phase: on observe une première
FID. Une fois la RF coupée, les spins perdent progressivement leur cohérence (B): ceux qui se déphasent
rapidement (en noir) acquièrent une avance de phase plus grande que ceux qui se déphasent lentement (en
gris). TE/2 plus tard, une RF à 180°y est envoyée. Les spins « rapides » se retrouvent en retard de phase
par rapport aux spins « lents » (C). TE/2 plus tard, les spins les plus rapides rattrapent de nouveau les
spins les plus lents (si on suppose que les molécules porteuses de spins ne bougent pas). On obtient une
nouvelle cohérence de phase donc une nouvelle FID (D). En répétant régulièrement les RF à 180°, on peut
créer une série d’échos. Cependant, ceux-ci sont d’int nsité plus faible à cause de l’« effet Te 2 ».

Chapitre I: Le tissu osseux et sa caractérisation
39
M0
Si on suppose que les molécules porteuses de spins ne bougent pas, on compense, par cette
méthode, les inhomogénéités magnétiques statiques (principalement celles de B0) car celles-ci
influencent les spins de la même façon avant et après la RF à 180°y. La décroissance des
échos successifs (que l’on peut obtenir par répétition de la RF à 180°y) se fait avec la
constante de temps « T2 » qui est différente de celle de l’enveloppe des FID que l’on note
« T2* » (T2 > T2
*).
Pour illustrer les conditions d’obtention d’une pondération du signal en T2, on reprend
l’échantillon présenté précédemment (deux populations de protons A et B telles que
T2A < T2
B). Le protocole précédent (Figure I.10) est appliqué, avec une série de RF à 180°y,
chacune étant séparée l’une de l’autre d’un temps TE. L’enveloppe des échos obtenus est donc
respectivement ATteM 2/
0− et BTteM 2/
0−
pour les populations A et B.
La différenciation des deux espèces sera alors facilitée si TE est long (cf. Figure I.11).
Intensité des échos
Temps TE court TE long
avec T2B > T2
A
BTteM 2/0
−ATteM 2/
0−
Figure I.11: Intensité des échos en fonction de TE. On distinguera d’autant mieux les protons de la
population A de ceux de la B si TE est long.
I.6.2.3. Contraste en densité de protons
On obtient un contraste lié à la densité de protons si on a TR long (pour assurer une
récupération totale de l’aimantation longitudinale pour tous les tissus, cf. Figure I.9) et TE
court (pour que la composante transversale ne soit pas influencée par la décroissance T2
propre à chaque tissu, cf. Figure I.11). Dans ces conditions, la seule différence qu’on observe
sur le signal est liée à M0 donc à la quantité de protons.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
I.6.3. Séquences d’imagerie
I.6.3.1. Principe du codage de l’espace
Le codage de l’espace est basé sur l’emploi de gradients de champ magnétique (un sur chaque
axe), superposés au champ statique 0Br
. Ces gradients sont générés par des bobines dans
lesquelles on fait circuler un courant. Les caractéristiques de ce courant (temps d’application,
intensité,…) permettent de contrôler l’amplitude et la durée du gradient. L’emploi des
gradients donne une relation entre la position dans l’espace et le champ magnétique. Or
comme on l’a vu en I.6.1.1, la fréquence de Larmor des protons est proportionnelle au champ
magnétique: on a donc une relation entre la fréquence et la position. L’espace est codé à partir
d’impulsions de gradient sur les trois axes générées à des instants précis. L’ensemble des
impulsions (gradients et RF) forment une « séquence d’imagerie ».
Après avoir détaillé le formalisme couramment utilisé (à partir d’une étude « 1D »), nous
présenterons le rôle des impulsions principales composant une séquence à partir de l’étude
d’une séquence type « écho de gradient ». Enfin, nous aborderons la séquence de type « écho
de spins » basée sur le principe abordé en I.6.2.2.
Remarque:
Il existe de nombreuses autres séquences d’imagerie présentant chacune avantages et
inconvénients en terme de temps d’acquisition, de rapport signal à bruit, d’artéfacts,…. Les
séquences « écho de gradient » et « écho de spins » sont cependant les deux principales
desquelles découlent toutes les autres.
I.6.3.2. Cas 1D: gradient de lecture
Supposons un échantillon parallélépipédique ne contenant que de l’eau placé dans un champ
statique homogène 0Br
. Après émission d’une perturbation RF (par exemple à 90°), on
observe une FID (cf. I.6.2). Si T2* est long, le contenu fréquentiel de la FID (que l’on peut
déterminer par transformée de Fourier) sera une raie à ω0 élargie par l’effet T2* (cf. Figure
I.12, A). Si par contre, après l’émission de la RF, on superpose à 0Br
, un gradient selon l’axe
x, Gx, la pulsation de précession ω va dépendre de la position x des spins suivant la relation:
ω = γ(B0 +Gx×x).
40

Chapitre I: Le tissu osseux et sa caractérisation
41
La FID reçue correspond à la somme des contributions des aimantations transversales
provenant de tout l’échantillon. Son contenu fréquentiel traduit donc la répartition spatiale de
l’aimantation transversale (cf. Figure I.12, B). La transformée de Fourier de la FID permet
donc d’obtenir une cartographie des aimantations transversales donc de la concentration
locale de protons. Ce principe est à la base de la technique d’imagerie. Dans la pratique, la
FID est démodulée (pour ramener le motif en bande de base) et échantillonnée avant de
calculer la transformée de Fourier discrète.
TF
Temps
FID Enveloppe
ωω0
Amplitude
B0
x X2
Protons
X1
A
TF
Temps
FID Enveloppe Amplitude
ω ω1 ω2
X2 X0 ω0
X1
x
xGdxdB=
B0
X1 X2 X0
Protons
B
Figure I.12: Illustration du rôle du gradient de lecture. Un échantillon ne contenant que de l’eau est placé
dans un champ statique B0. Une RF est émise (par exemple à 90°). Si le système reste tel quel (A), on se
trouve dans le cas de la Figure I.8 (l’enveloppe correspondant à l’effet T2*, cf. I.6.2). Si T2
* est long, le
contenu fréquentiel de la FID est une raie à ω0 (légèrement élargie par l’effet T2*). Par contre, si après
l’émission de la RF, on superpose à B0 un gradient Gx (B), les aimantations transversales précessent à une
pulsation de Larmor qui est fonction de leur position. Le contenu fréquentiel de la FID sera donc un
spectre continu traduisant les amplitudes locales de l’aimantation.
Le gradient utilisé pour cette opération est appelé « gradient de lecture » ou « gradient de
codage de la fréquence ». Le profil obtenu correspond à la projection de l’échantillon sur
l’axe du gradient.
Remarque:
Le choix de la direction x est arbitraire: le gradient de lecture peut être appliqué sur n’importe
quel axe.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
I.6.3.3. Formalisme
Pendant l’application du gradient de lecture Gx, la pulsation de précession des aimantations
transversales dépend de leur position sur l’axe x: tMr
xtGxtGBtBt xxx )())(()()( 00 γωγγω +=+== .
Les aimantations transversales se déphasent donc avec une phase instantanée qui est fonction
de leur position x:
duuGxtduutxt
x
t
x )()(),(0
00
∫∫ +== γωωφ .
Si Tech est la période d’échantillonnage de la FID, l’échantillon numérisé n correspond donc à
un état de phase de:
)()(),( 00
0 echx
nT
xech nTxStduuGxtnTxech
γωγωφ +=+= ∫
avec:
duuGnTSechnT
xechx )()(0∫= .
Sx(nTech) est l’aire sous la courbe du gradient de lecture en fonction du temps jusqu’à
l’échantillon n. Si on pose:
kx = γSx(nTech),
on a:
xkttx x+= 0),( ωφ .
Le signal I acquis pour chaque échantillon correspond à la somme, sur tout le volume analysé,
des aimantations transversales mises dans un état de phase qui est fonction de x. On a
donc:
tMr
∫=Volume
xtti
x dxxikzyxMekI o )exp(),,()( ω .
Avant d’être traité, I est démodulé ce qui correspond à une division par . D’après
l’expression précédente, on obtient alors la transformée de Fourier inverse de la projection sur
l’axe x des aimantations transversales. La transformée de Fourier de I donne donc la
cartographie de M
ti oe ω
t selon l’axe x c’est-à-dire la projection du volume sur l’axe x.
L’ensemble des Nx échantillons acquis pour I est généralement appelé l’« espace des k »
(« k-space »).
42

Chapitre I: Le tissu osseux et sa caractérisation
43
Dans la partie suivante, nous détaillons une séquence de type « écho de gradient » ce qui nous
permettra de présenter la généralisation de cette formulation au cas tridimensionnel.
I.6.3.4. Séquence « écho de gradient »
La Figure I.13 présente une séquence de type « écho de gradient ». Nous détaillons dans la
suite chacune des impulsions mise à part celle de gradient de lecture (appliquée selon l’axe x
dans cet exemple) qui a déjà été présentée (cf. I.6.3.2). On suppose que est colinéaire à
l’axe z et que l’effet T
0Br
2* est négligeable.
TR
TE
Gz
t
Gsélection
Grefocalisation
Tz
Nz lignes
Gcodage phase z
Gy
t
Ty
Ny lignes
Gcodage phase y
RF α°RF
t
RF α°
Gx
t
Surfaces égales
Glecture
Gprélecture
FID
t
Echo
Tobs = Nx×Tech
Acquisition
Figure I.13: Représentation d’une séquence d’imagerie de type « écho de gradient ». On suppose que 0Br
est colinéaire à l’axe z. Les impulsions de gradients ne sont en réalité pas rectangulaires mais
trapézoïdales du fait du temps de montée (non nul) des gradients.
I.6.3.4.1. Impulsion radio-fréquence
Cette impulsion est notée « RF » sur la Figure I.13.
Un angle de basculement α de 90° permet de basculer toute l’aimantation longitudinale
(amplitude maximale de la FID). Il peut cependant être intéressant de prendre un angle plus

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
petit pour n’en basculer qu’une partie. On dispose ainsi d’une « réserve » d’aimantation
longitudinale ce qui permet de raccourcir le temps de remontée et donc d’effectuer une
acquisition plus rapide (la conséquence étant que l’aimantation transversale disponible est
d’intensité plus faible).
I.6.3.4.2. Gradient de sélection
Cette impulsion est notée « Gsélection » sur la Figure I.13.
Le gradient de sélection est essentiel dans toutes les applications où la zone à imager est une
partie de l’objet qui est placé dans le champ. C’est évidemment le cas de l’imagerie en 2D
(sélection d’une coupe), mais aussi souvent en 3D (sélection d’un volume appelé « slab »).
Lorsque le patient est placé dans le champ statique 0Br
supposé homogène, tous les protons
ont la même fréquence de Larmor. Une RF à ω0 exciterait donc tout le volume. Pour pouvoir
examiner seulement une zone particulière, on superpose, au moment où on applique la RF, un
gradient de champ Gsélection selon la direction de 0Br
(soit l’axe z dans notre exemple) appelé
« gradient de sélection ». A chaque position z de cet axe, correspond donc un champ B(z):
B(z) = B0 + Gsélection×z.
Les protons ont donc une pulsation de Larmor fonction de leur position sur l’axe z:
zGz sélection×+= γωω 0)( .
Grâce à cette impulsion de gradient, la bande de fréquence excitée et la fréquence centrale de
la RF fixent respectivement l’épaisseur et la position de la zone à étudier.
I.6.3.4.3. Gradient de refocalisation
Cette impulsion est notée « Grefocalisation » sur la Figure I.13.
Du fait de l’impulsion de gradient de sélection pendant l’émission de la RF (cf. I.6.3.4.2), la
phase des aimantations transversales excitées change. Une impulsion de gradient sur le même
axe, dite « gradient de refocalisation », est alors nécessaire pour rétablir l’état de phase initial.
La surface de cette impulsion est optimisée pour chaque type de RF utilisée.
I.6.3.4.4. Gradient de prélecture
Cette impulsion est notée « Gprélecture » sur la Figure I.13.
Du fait d’un certain nombre de délais incompressibles (temps de montée des gradients, temps
morts dans la chaîne d’acquisition,…), on ne peut pas effectuer l’échantillonnage dès la fin de
l’impulsion RF. Pour une FID du type de celle présentée en Figure I.8, cela aurait pour
44

Chapitre I: Le tissu osseux et sa caractérisation
45
Y1 Y2
conséquence que l’on ne pourrait récupérer l’amplitude maximale de signal. Or ces
échantillons correspondent au centre de l’« espace des k » (aimantations transversales en
phase): ils représentent le contraste et ont donc un rôle primordial dans la qualité de l’image.
Afin de les acquérir avec une sensibilité maximale, on cherche à faire apparaître l’écho à un
moment précis de l’impulsion du gradient de lecture. On utilise donc une impulsion de
gradient négative selon l’axe de la lecture, dite impulsion de « gradient de prélecture », qui va
générer un déphasage des aimantations transversales. L’écho ne pourra alors apparaître qu’au
moment où ce déphasage sera compensé par l’impulsion de gradient de lecture. Ainsi, si on
souhaite faire apparaître l’écho au centre de l’impulsion de gradient de lecture (de surface S),
il suffit de placer une impulsion de gradient de prélecture de surface -½S (cf. Figure I.13:
zones grisées): la FID obtenue est alors symétrique.
I.6.3.4.5. Gradients de codage de phase
Ces impulsions sont notées « Gcodage phase » (pour les deux axes y et z) sur la Figure I.13.
Le rôle de ces deux gradients est de placer les aimantations transversales dans un état de
phase dépendant de leurs positions selon ces deux axes. Le principe est très proche de celui du
gradient de lecture (cf. I.6.3.2). Pour l’illustrer, on considère un échantillon (ne contenant que
de l’eau) juste après l’impulsion de refocalisation. Toutes les aimantations transversales du
volume sont donc en phase et précessent à la pulsation ω
tMr
0. La Figure I.14 présente l’état de
phase obtenu pour un plan (y,z).
Y3
Z1
Z2
Z3
Figure I.14: Représentation de 9 aimantations transversales d’une coupe (y,z) d’un échantillon ne
contenant que de l’eau, juste après l’impulsion de refocalisation. Tous les aimantations transversales de
l’échantillon sont en phase et précessent à ω0.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
46
Supposons qu’on applique, pendant une durée Ty, une impulsion de gradient selon l’axe y:
Gcodage phase y(t).
Tant que dure cette impulsion, la pulsation de précession dépend de la position des
aimantations transversales sur l’axe y:
ytGytGBtBt yphasecodageyphasecodagey )())(()()( 00 γωγγω +=+== .
Les aimantations transversales vont donc se déphaser progressivement d’une colonne à l’autre
puisque la pulsation de précession des spins en Y1 est supérieure à celle en Y2 (et ainsi de
suite). Par contre, la cohérence restera au sein de chaque colonne puisque pour un Y donné,
tous les spins voient le même champ magnétique et précessent donc à la même pulsation.
A la fin de cette impulsion de gradient, le champ revient à B0. Tous les spins retrouvent la
même pulsation de précession (ω0) mais conservent leur état de phase (cf. Figure I.15) qui est
donné par:
y
T
yphasecodage yStduuGyttyy
γωγωφ +=+= ∫ 00
0 )(),( ,
où Sy est l’aire sous la courbe du gradient de codage de phase en fonction du temps.
Y1 Y2 Y3
Z1
Z2
Z3
Figure I.15: Etat de la phase des aimantations transversales après l’impulsion de gradient de codage de
phase selon l’axe y.
De la même manière qu’en I.6.3.3, si on pose:
ky = γSy,
on a:
yktty y+= 0),( ωφ .
Le même raisonnement peut être tenu pour le gradient de codage de phase dans la direction z.
A la fin de l’application de l’impulsion de gradient Gz(t) pendant une durée Tz, on a:

Chapitre I: Le tissu osseux et sa caractérisation
47
zkykttzy zy ++= 0),,( ωφ ,
avec:
kz = γSz.
La Figure I.16 donne une représentation de l’état de phase obtenue après cette impulsion, pour
la colonne Y1.
Y1
Z1
Z2
Z3
Y1
Figure I.16: Effet sur la colonne Y1 de l’impulsion de gradient de codage de phase dans la direction z
(l’intensité maximale du champ est en Z1). Pendant l’application du gradient, les aimantations
transversales se déphasent d’une ligne à l’autre (l’écart de phase relatif d’une colonne à l’autre est celui lié
à l’impulsion de gradient de codage de phase dans la direction y). Au moment où ce gradient est coupé,
tous les spins précessent de nouveau à ω0 et l’état de phase est conservé.
On obtient donc une différenciation sur la phase des aimantations transversales selon ces deux
axes, pour chacune des Ny×Nz configurations de ces deux gradients i.e. pour chaque couple
(Sy,Sz) c’est-à-dire (ky,kz). En appliquant le gradient de lecture selon l’axe x dans chaque cas
(cf. I.6.3.2 et I.6.3.3), on a:
zkykxkttzyx zyx +++= 0),,,( ωφ
soit après échantillonnage de la FID (Nx points):
∫∫∫ ++=Volume
zyxtti
zyx dxdydzzkykxkizyxMekkkI o )](exp[),,(),,( ω
L’« espace des k » est donc acquis ligne par ligne (axe de lecture), pour différentes valeurs de
ky et kz, ce qui conduit à un volume de Nx×Ny×Nz échantillons. La transformée de Fourier de I
permet d’accéder à la cartographie Mt(x,y,z), c’est-à-dire l’image 3D du volume
(Nx×Ny×Nz voxels).

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Remarques:
• Les effets des deux impulsions de gradient de codage de phase ont été séparés pour
l’étude. Elles peuvent cependant être simultanées.
• Dans le cas d’imagerie en 2D, on utilise un seul gradient de codage de la phase.
I.6.3.4.6. Remarques
a) Il est à noter que les 3 axes peuvent être interchangés du fait qu’ils jouent un rôle similaire.
b) Afin d’obtenir le TE le plus court possible, toutes les impulsions de gradients peuvent être
simultanées, mis à part le gradient de sélection. Par exemple, les impulsions de gradients de
refocalisation et de prélecture sont souvent fusionnées. Par souci de clarté, les différentes
impulsions ont été volontairement dissociées sur la Figure I.13 et Figure I.17.
I.6.3.5. Séquence « écho de spins »
Dans ce type de séquence (cf. Figure I.17), l’écho est généré par une autre excitation RF à
180° selon le principe décrit en I.6.2.2.
Du fait de l’influence de cette RF, les impulsions de gradients de codage de phase et de
prélecture doivent changer de signe. On peut noter que le gradient de sélection de coupe n’est
pas nécessaire pour la RF à 180°: en effet, celle-ci n’affecte que les aimantations transversales
donc celles qui ont déjà été « basculées » par la première RF.
48

Chapitre I: Le tissu osseux et sa caractérisation
49
TR
½TE ½TE
Gz
t
Gsélection
Grefocalisation
Tz
Nz lignes
Gcodage phase z
Gy
t
Ty
Ny lignes
Gcodage phase y
RF 90°x RF 90°xRF
t
RF 180°y
Gx
t Gprélecture
Surfaces égales
Glecture
FID
t
Echo
Tobs = Nx×Tech
Acquisition
Figure I.17: Représentation d’une séquence d’imagerie de type « écho de spins ». On suppose que 0Br
est
colinéaire à l’axe z. Les impulsions de gradients ne sont en réalité pas rectangulaires mais trapézoïdales du
fait du temps de montée (non nul) des gradients.
I.6.3.6. Avantages et inconvénients de ces séquences
L’avantage principal de la séquence « écho de gradient » est qu’elle permet d’obtenir des
acquisitions relativement rapides. Son principal défaut est qu’il n’y a pas de correction des
inhomogénéités statiques du champ magnétique (cf. I.6.2.2), ce qui peut être gênant dans le
cas de différences de susceptibilité magnétique (cf. I.6.4.3.1).
La séquence « écho de spins » est plus lente (impulsion RF à 180° supplémentaire) mais
permet la correction de l’effet des inhomogénéités statiques.
I.6.4. IRM haute résolution
Les travées osseuses ont une épaisseur de l’ordre de quelques centaines de microns (cf. I.1).
L’analyse de la microarchitecture trabéculaire nécessite donc des images suffisamment

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
résolues, la taille des voxels devant être au moins de l’ordre de la taille caractéristique des
travées. L’intensité des signaux acquis en IRM est très faible (cf. I.6.2) ce qui restreint la
sensibilité de la technique: la résolution spatiale est donc souvent limite pour les études du
tissu trabéculaire. Nous donnons dans ce paragraphe, différents facteurs de l’acquisition qui
l’influencent.
I.6.4.1. Rôle des gradients
La résolution dans les trois directions est fonction de l’aire maximale sous la courbe des
gradients en fonction du temps. On suppose des impulsions de gradients rectangulaires.
La résolution sur les axes de codage de phase (par exemple y) est donnée par:
yyTGyγπδ = ,
où Gy est la valeur maximale du gradient et Ty le temps d’application.
Sur l’axe de lecture (x), on a une formule similaire:
obsxTGxγ
πδ 2= ,
où Gx est la valeur maximale du gradient de lecture et Tobs, la durée d’observation du signal.
Pour améliorer la résolution (pour une valeur de Gx donnée), il faut donc augmenter Tobs. La
durée d’application du gradient de lecture est liée à la période d’échantillonnage Tech par la
relation:
Tobs = Nx×Tech,
où Nx est le nombre d’échantillons acquis (taille de la matrice d’acquisition selon l’axe x).
Pour respecter les conditions de Shannon, le signal doit être filtré de echT21− à
echT21 (filtre
« anti-repliement »). La bande passante du signal (notée BW: « Bandwidth ») est donc donnée
par:
echTBW 1= .
On a donc:
BWNT x
obs= ,
soit:
50

Chapitre I: Le tissu osseux et sa caractérisation
xx NGBWx
γπδ 2= .
Le champ de vue (noté FOV: « Field of View ») s’exprime donc ainsi:
xx G
BWxNFOVγπδ 2== .
BW fixe le champ de vue. Pour améliorer la résolution, on procède de deux façons: en
augmentant Nx à BW fixé, ou en diminuant BW à Nx fixé (ce qui a donc aussi pour effet de
diminuer le champ de vue).
I.6.4.2. Rapport Signal sur Bruit (RSB)
Le principal facteur qui limite la résolution spatiale est le Rapport Signal à Bruit (RSB). En
première approximation, on a:
IB
BWRkTNNNNVMRSB zyxacc 100
4)sin( ×
××≈ δαω ,
où ω0 est la pulsation de Larmor, M0 l’aimantation à l’équilibre, α l’angle de basculement, δV
le volume du voxel (δV = δx×δy×δz), Nacc le nombre d’accumulations des acquisitions, Nx,
Ny et Nz la taille de la matrice d’acquisition, k la constate de Boltzmann, T la température de
la sonde, R la résistance de bruit de la sonde (c’est-à-dire principalement sa résistance de bruit
propre et celle induite par le patient), BW la bande passante et IB1 le coefficient d’induction
de la sonde.
Les facteurs principaux qui influencent le RSB sont donc les suivants.
I.6.4.2.1. Caractéristiques de la sonde
Plus la sonde de réception est petite, plus la détection du signal est bonne. Cependant,
l’homogénéité de la réponse est meilleure pour une sonde de grande dimension. De plus, les
sondes de petite taille sont caractérisées par un bruit dominant lié à leur résistance propre, ce
qui n’est pas la configuration optimale pour le RSB (d’autant plus quand le champ statique B0
est élevé, ce qui est souvent le cas pour les études de microscopie IRM).
Pour utiliser des sondes de petite taille sans affecter le RSB, il faut passer dans la zone où le
bruit dominant est celui du patient: on joue alors sur la température de la sonde. Ainsi, le
développement et l’utilisation de sondes supraconductrices ont déjà montré le grand intérêt de
ce type d’instrumentation pour l’amélioration du RSB ([Ginefri, 2001]).
51

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
I.6.4.2.2. Champ statique
Le champ statique élevé permet d’augmenter ω0 et M0 (cf. I.6.1.1) mais aussi, dans une
moindre mesure, R. Travailler avec des valeurs de B0 élevées permet donc d’améliorer
sensiblement le RSB. Cependant, du fait des limitations technologiques, les champs de vue
accessibles sont restreints (difficulté pour assurer de forts champs statiques homogènes sur un
grand volume): à l’heure actuelle, le champ statique le plus élevé pour un imageur clinique
« corps entier » est de 3 T. De plus, les effets de certains artéfacts sont d’autant plus sensibles
que le champ statique est élevé: l’artefact de susceptibilité magnétique par exemple (cf.
I.6.4.3.1).
I.6.4.2.3. Voxel
Le volume du voxel influence considérablement le RSB: une amélioration de la résolution
d’un facteur 2 dans chaque direction à RSB constant nécessiterait une augmentation du temps
d’acquisition Tacq d’un facteur 64.
Le nombre d’accumulations permet d’améliorer le RSB mais augmente aussi le temps
d’acquisition.
I.6.4.2.4. Durée de vie du signal
Cette expression du RSB simplifiée ne fait pas apparaître certains facteurs, en particulier la
constante de relaxation T2*: plus celle-ci sera courte, plus le RSB sera faible. Celle-ci peut
aussi limiter la résolution. Comme on l’a vu en I.6.4.1, la valeur de Tobs doit être augmentée
pour améliorer la résolution. Ceci ne peut être fait significativement que si la FID n’est pas
noyée dans le bruit donc si T2* est « assez » long.
I.6.4.3. Les artéfacts
Des artéfacts peuvent aussi altérer la qualité de l’image.
I.6.4.3.1. Artéfact de susceptibilité
A s’ajoute un champ magnétique χ×0Br
0Br
où χ est la susceptibilité magnétique du matériau.
Dans le cas de matériau non homogène et plus particulièrement en présence d’interfaces
(comme dans notre cas, entre le tissu calcifié et la moelle), on observe une inhomogénéité
spatiale (statique) du champ magnétique dans le voisinage de l’interface entraînant une
52

Chapitre I: Le tissu osseux et sa caractérisation
diminution plus rapide de l’amplitude de la FID. Cet artéfact peut cependant être compensé
par l’emploi d’une séquence type « écho de spins » (cf. I.6.3.5).
I.6.4.3.2. Décalage chimique
Comme on l’a vu précédemment (cf. I.6.1.3), l’environnement chimique des protons influence
ses caractéristiques. Par exemple, entre des protons d’eau et ceux de molécules de graisse, on
observe un décalage des rapports gyromagnétiques de 3.4 ppm (ce qui correspond à 220 Hz
pour B0 = 1.5 T). Sur l’axe de la lecture, on observe un effet si cet écart de fréquence est
supérieur à la bande passante par voxel: dans ce cas, les protons de la graisse situés à une
position se retrouvent décalés d’un nombre de voxels correspondant à l’écart de fréquence.
On observe alors une superposition de zones différentes sur l’image. Cet artéfact peut aussi
survenir sur l’axe de la sélection de coupe. En effet, des protons de la graisse situés dans une
autre coupe que celle sélectionnée peuvent précesser à ω0 du fait du gradient de sélection:
ceux-ci sont donc aussi excités par la RF. Par contre, il ne peut survenir sur les axes de codage
de phase, puisque le déphasage qu’il génère est constant d’un plan à l’autre.
I.6.4.3.3. Repliement (aliasing)
Cet artefact n’affecte normalement pas l’axe de lecture du fait que la FID est échantillonnée
en respectant les conditions de Shannon (cf. I.6.4.1). Selon les autres axes, il y a aussi
échantillonnage et donc périodisation du champ de vue. Si un signal de protons situés hors du
champ de vue est reçu, il apparaîtra replié et superposé sur l’image. Cet artéfact peut obliger à
prendre un champ de vue plus grand et donc imposer une réduction de la résolution.
I.6.5. Etat de l’art: IRM du tissu trabéculaire in vitro et in vivo
Pour l’imagerie du tissu trabéculaire par IRM in vivo, le signal provient de la moelle osseuse
contenue dans les cavités trabéculaires. Dans la plupart des études in vitro, les tissus mous et
la moelle sont éliminés des prélèvements et remplacés par de l’eau (éventuellement avec un
produit de contraste) qui donne le signal utile. Le tissu solide ne donne aucun signal: il
apparaît sur les images comme du bruit.
Le Tableau I.1 présente les principales publications récentes in vitro (ES: « écho de spins »,
EG: « écho de gradient »). Pour B0 = 1.5 T, la plupart des travaux ont été menées à une
résolution de 156×156 µm2 ou 117×117 µm2 avec une épaisseur de coupe de 300 µm. Les
53

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
analyses, souvent limitées par le faible RSB disponible, sont effectuées sur des voxels
anisotropes. Par contre, elles sont menées dans des conditions très similaires à celles
accessibles in vivo, mis à part les temps de pose qui seraient prohibitifs pour une application
sur un patient. Des études à plus haut champ ont aussi été menées, permettant d’améliorer
sensiblement la résolution et de passer à un voxel isotrope. Pour la plupart de ces travaux, les
auteurs ont utilisé des échantillons dégraissés remplis d’eau dopée avec un produit de
contraste.
Référence Site B0 (T) Résolution (µm) Séquence Temps de pose
[Link, 2003] Radius 1.5 1532×300 ES 3D 1 heure
[Pothuaud, 2002 (a)] Radius 1.5 1562×300 EG 3D ?
[Gardner, 2001] Tibia (souris) 11.7 20×40×60 ES 3D 10 heures
[Beuf, 2001] Vertèbre 1.5 1172×300 ES/EG 3D 30 minutes
[Vieth, 2001] Calcanéum 1.5 1952×300 ES 1 heure
[Borah, 2000] Vertèbre (cochon) 4.7 853 ES 3D 1 heure
[Majumdar, 1998] Fémur,
Calcanéum, Vertèbre
1.5 1172×300 EG 3D 15 minutes
[Hwang, 1997] Radius 9.4 783 ES 3D dédiée 30 minutes
[Hipp, 1996] Humérus,
Fémur, Tibias (bœuf)
8.6 923 ES 3D 1 heure
[Chung, 1995] Vertèbre 9.4 302×150 ES 2D ?
Tableau I.1: Principales images obtenues sur le tissu trabéculaire par IRM in vitro (ES: « écho de spins »,
EG: « écho de gradient »).
Le Tableau I.2 présente les principales publications récentes in vivo. La plupart de ces études
ont été menées sur des imageurs « corps entier » classiques à 1.5 T (disponibles en hôpital).
Les sites concernés sont principalement périphériques (radius, calcanéum) du fait qu’ils sont
faciles d’accès (en particulier pour la sonde de réception).
54

Chapitre I: Le tissu osseux et sa caractérisation
Référence Site Résolution (µm) Séquence Temps de pose
[Beuf, 2002] Genou 1952×1000 EG 3D 12 minutes
[Laib, 2002] Radius 1562×500 EG 3D 12 minutes
[Link, 2002] Calcanéum 1952×1000 ES 2D 8 minutes
[Van Rietbergen, 2002] Calcanéum 1952×500 EG 3D 12 minutes
[Wehrli, 2001] Radius 1372×350 ES 3D dédiée 11 minutes
[Rotter, 2001](*) Calcanéum 2542×2188 EG 3D 11 minutes
Tableau I.2: Principales images obtenues sur le tissu trabéculaire par IRM in vivo (ES: « écho de spins »,
EG: « écho de gradient »). Toutes ces études ont été menées sur des imageurs cliniques (corps entier) à
1.5 T sauf une à 3 T(*).
De nombreuses études sont aussi menées en IRM quantitative pour étudier le tissu
trabéculaire. Comme on l’a vu en I.6.4.3.1, l’interface os/moelle crée une différence de
susceptibilité magnétique qui modifie la valeur de T2*. A partir d’image paramétrique du T2
*,
on peut obtenir une information sur la densité et la structure trabéculaire. Ainsi, Chung et al.
([Chung, 1993]) ont montré qu’il existait des liens étroits entre (T2*)-1 et le module d’Young
d’une part et entre (T2*)-1 et la microstructure d’autre part. Plus récemment, Brismar et al.
([Brismar, 1999]) ont publié une méthode pour déterminer le nombre relatif de travées
osseuses (par mm) dans différentes orientations à partir de mesures du (T2*)-1.
55


Chapitre II: Quantification de la microarchitecture trabéculaire
Chapitre II: Quantification de la
microarchitecture trabéculaire
II.1. Introduction
Du fait de l’importance du tissu trabéculaire dans les propriétés mécaniques de l’os (cf. I.4),
différents outils de caractérisation de la microarchitecture ont été proposés. Ceux-ci ont pour
objectif de rendre compte des propriétés spatiales et géométriques de ce tissu osseux. Dans ce
chapitre, nous présentons plusieurs familles de paramètres pour lesquels nous avons
développé des algorithmes de calcul. Ceux-ci sont directement applicables sur des images
tridimensionnelles dont la résolution est comprise entre une dizaine et quelques centaines de
microns. Pour des acquisitions plus précises (comme celles obtenues par histomorphométrie
osseuse, cf. I.5.2), les méthodes peuvent différer.
Ces paramètres de structure sont employés dans divers buts: i) recherche de corrélations avec
des propriétés mécaniques de l’os, ii) différenciation entre populations saine et pathologique,
iii) étude de l’évolution de la microstructure avec l’age, iv) caractérisation des différences
entre sites osseux… Afin d’illustrer ces différentes applications, nous présenterons, pour
chaque famille de paramètres, les résultats qui ont déjà été obtenus.
Pour des raisons techniques et historiques, les méthodes mathématiques pour caractériser une
microstructure ont d’abord été développées pour traiter des images 2D. Les dispositifs
d’acquisition d’images 3D se généralisant peu à peu, les méthodes de caractérisation ont été
adaptées au cas tridimensionnel. Les paramètres présentés ici sont dédiés à ce type d’étude.
57

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Les symboles utilisés dans ce chapitre sont ceux couramment admis dans la littérature. Ils sont
en général d’origine anglophone. Ils sont définis pour la plupart dans un rapport de 1987
([Parfitt, 1987]) de l’« American Society of Bone and Mineral Research » (ASBMR) qui a
établi une standardisation de la nomenclature des paramètres fréquemment utilisés par cette
communauté scientifique. Cet article est encore aujourd’hui une référence. Nous avons donc
choisi d’adopter les symboles qu’il propose afin de faciliter la lecture d’articles sur ce sujet
(leur signification sera précisée quand chacun d’entre eux sera introduit).
II.2. Un préalable: la segmentation
Les paramètres utilisés dans le cadre de ce travail supposent une représentation binaire du
tissu trabéculaire, c’est-à-dire une image où chaque voxel ne peut être que de type « os » ou
de type « pore ». L’histogramme h de cette image idéale du tissu trabéculaire serait donc de la
forme:
h(x) = a×δ(x-xos) + b×δ(x-xpore),
où δ est l’impulsion de Dirac, x le niveau de gris d’un voxel de l’image (on supposera dans la
suite x∈0,…,255), xos le niveau de gris de la phase osseuse, xpore celui de la phase poreuse
et a et b, les coefficients représentant respectivement la proportion de voxels d’os et de pore
dans l’image (a + b = 1). Dans la pratique, à cause du bruit (propre à chaque type
d’acquisition) et/ou d’une résolution non adaptée à l’imagerie des travées, la distribution des
voxels de chaque phase s’élargit. Il est alors nécessaire de segmenter les images c’est-à-dire
d’attribuer à chaque voxel une des deux phases possibles.
II.2.1. Histogramme à modes séparés
Dans le cas où le Rapport Signal sur Bruit (RSB) est bon et la résolution adaptée aux
structures (c’est-à-dire pour une taille de voxel bien inférieure à celle caractérisant les
travées), les deux modes de l’histogramme sont « suffisamment » séparés (cf. Figure II.1). Un
seuillage simple de l’image avec le seuil σ choisi au niveau de gris équidistant des maxima
des deux modes permet d’obtenir une segmentation « sans ambiguïté » i.e. un nombre
négligeable de voxels changeraient de phase si on faisait varier la valeur du seuil autour de σ.
58

Chapitre II: Quantification de la microarchitecture trabéculaire
59
0
0,005
0 50 100 150 200 250
NG
0,01
0,015
0,02
0,025
Occurrence normalisée NG1 NG2 Seuil σ
Voxels du « Mode 1 »
Voxels du « Mode 2 »½ ½
Figure II.1: Simulation d’histogramme présentant deux modes séparés: les deux phases se distinguent très
nettement. En seuillant l’image avec le seuil σ choisi au milieu de [NG1,NG2] (niveaux de gris
correspondant respectivement aux maxima du premier et du deuxième mode), on obtient une
segmentation non ambiguë car très peu de voxels changeraient de phase si on faisait varier la valeur du
seuil autour de σ.
II.2.2. Histogramme problématique
Dans le cas où le RSB n’est pas suffisant, les modes des voxels de bruit et de signal se
rapprochent sur l’histogramme. De plus, si la résolution n’est pas adaptée, le nombre de
voxels situés pleinement dans l’une des deux phases diminue au profit de voxels contenant
des proportions comparables d’os et de moelle. Cet effet, appelé « volume partiel », se traduit
sur l’histogramme par l’apparition d’une population de voxels caractérisés par des niveaux de
gris intermédiaires.
La Figure II.2 illustre ces deux problèmes (RSB et résolution) qui compliquent
considérablement l’étape de segmentation. Ce type d’histogramme est relativement fréquent
pour des images de tissu trabéculaire obtenues par IRM du fait de la sensibilité réduite de la
technique qui limite la résolution (cf. I.6.4). Pour ce type d’image, la segmentation constitue
une étape cruciale car la précision de l’évaluation des paramètres sera fonction de sa
« qualité ». Elle ne peut donc plus être menée aussi simplement que dans le cas de la Figure
II.1.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
60
0
0,005
0 50 100 150 200 250
NG
0,01
0,015
0,02
0,025
Occurrence normalisée
Phase ?
Volume partiel
Voxels du « Mode 1 »
Voxels du « Mode 2 »
Figure II.2: Simulation d’histogramme bimodal pour un RSB et une résolution limites. Un nombre
important de voxels contiennent des proportions semblables d’os et de pore, pour des niveaux de gris
intermédiaires. Il s’agit du volume partiel qui complique considérablement la segmentation.
De nombreuses méthodes dédiées aux images de tissu trabéculaire acquises par IRM ont déjà
été proposées dans la littérature: dans la partie suivante, nous présentons les principales.
II.2.3. Méthodes dédiées à la segmentation d’images acquises
par IRM
II.2.3.1. Seuillage
De nombreuse techniques de segmentation consistent en un seuillage mais pour un seuil
choisi « intelligemment ».
Chung et al. ([Chung, 1995]) supposent que la distribution des niveaux de gris dans l’image
peut être représentée par la somme d’une distribution de Rayleigh pour le tissu calcifié
(voxels de bruit) et d’une distribution gaussienne pour l’espace des pores (qui sont remplis
d’eau: voxels de signal). Après avoir ajusté ce modèle sur l’histogramme h de l’image, ils
déterminent le seuil optimal σopt d’après l’équation suivante:
Rdxxhopt
=∫σ
0
)( ,
où R est l’amplitude de la distribution de Rayleigh et x le niveau de gris.

Chapitre II: Quantification de la microarchitecture trabéculaire
D’autres techniques utilisent des opérateurs de détection de contour couramment employés en
traitement de l’image (filtrage de Sobel [Hipp, 1996], filtrage de Deriche [Pothuaud, 2000])
pour déterminer le seuil optimal.
Majumdar et al. ([Majumdar, 1996]) ont aussi proposé une technique de seuillage qu’ils ont
utilisée dans de nombreuses autres publications. Celle-ci se base sur trois intensités moyennes
calculées en considérant certaines zones de l’image et en se référant à l’histogramme: IR
caractérisant la zone trabéculaire (contenant donc des voxels d’os et de moelle mais aussi
ceux affectés par l’effet du volume partiel), IB caractérisant le tissu cortical (donc les voxels
d’os) et IL caractérisant la moelle. Le seuil σ choisi est alors celui pour lequel la relation
suivante est la mieux satisfaite:
IR = a(σ)×IB + [1 - a(σ)]×IL,
où a(σ) est la proportion de voxels d’os dans l’image calculée pour le seuil σ. Pour les
auteurs, ce procédé, bien qu’arbitraire, permet une standardisation de la segmentation ce qui
est essentiel pour pouvoir effectuer des comparaisons d’images.
II.2.3.2. Autres méthodes
D’autres méthodes de segmentation plus complexes que les techniques de seuillage ont aussi
été proposées ([Wu, 1994], [Antoniadis, 1998 (a)]). Celles-ci sont basées sur des calculs
statistiques (effectués sur le voisinage de chaque voxel) adaptés à la structure particulière du
tissu osseux.
Enfin des méthodes de réduction de l’effet de volume partiel basées sur la cartographie de la
fraction osseuse peuvent être avantageusement utilisées pour la segmentation (même si ce
n’est pas leur objectif premier). Une technique ([Hwang, 1999]) propose par exemple une
déconvolution itérative de l’histogramme pour se rapprocher au mieux de l’histogramme
idéal. Une autre ([Hwang, 2002]) consiste en une répartition du contenu osseux de chaque
voxel dans huit sous-voxels de manière à assurer une cohérence de la topologie locale.
Les techniques que nous avons utilisées (en particulier [Antoniadis, 1998 (a)] pour l’étude in
vitro et [Hwang, 2002] pour l’étude in vivo) seront décrites plus précisément ultérieurement
(cf. III.5.1 et IV.4.3).
61

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
II.2.4. Caractérisation de la structure osseuse sans segmentation
Afin d’éviter l’étape de segmentation, certaines équipes ont proposé d’autres familles de
paramètres basées sur l’analyse de l’image directement en niveau de gris. Les résultats
obtenus montrent que cette voie d’analyse n’est pas à sous estimer.
En utilisant la probabilité que le voxel soit situé pleinement dans l’os (probabilité égale au
signal de chaque voxel normalisé entre 0 et 1), Hwang et al. ([Hwang, 1997]) ont calculé une
fonction d’autocorrélation spatiale (dans deux directions privilégiées d’un échantillon de tissu
trabéculaire) pour définir une série de paramètres. Ils ont ainsi pu proposer différentes
relations entre le module d’Young et ces paramètres, l’une d’entre elles ayant conduit à un
coefficient de détermination R2 de 0.91.
Plus récemment ([Rotter, 2001]), une étude se basant sur le même type de fonction
d’autocorrélation a montré qu’il était possible de déterminer l’anisotropie de la structure
osseuse.
De nombreux travaux ont aussi montré, sur des images de projection radiographique, l’intérêt
de l’analyse fractale (basée sur la corrélation des niveaux de gris dans des directions données)
pour la caractérisation de la microstrucutre ([Pothuaud, 2000]) ou pour la distinction d’une
population pathologique par rapport à un groupe sain ([Benhamou, 2001]).
Ce type d’attributs n’a cependant pas été envisagé dans le cadre de cette étude. Dans la suite
de ce chapitre, nous supposerons des images segmentées présentant deux ensembles
complémentaires l’un de l’autre: les voxels de la phase osseuse et ceux de la phase poreuse.
II.3. Fraction volumique osseuse
Ce paramètre est égal au nombre de voxels d’os présents dans la région d’intérêt (« Region of
Interest », ROI) sur le nombre de voxels total de la ROI. Ce paramètre est calculable
directement après segmentation. Il est noté « BV/TV » ce qui signifie
« Bone Volume / Tissue Volume ».
La fraction volumique de moelle est évidemment égale à 1-BV/TV.
62

Chapitre II: Quantification de la microarchitecture trabéculaire
II.4. Paramètres topologiques
Parmi les différences observables entre tissu trabéculaire sain et tissu pathologique (cf. Figure
I.2), une des plus flagrantes est certainement la perte de connectivité du réseau trabéculaire.
Les paramètres topologiques permettent d’obtenir une quantification de cette propriété
fondamentale du tissu sans faire aucune hypothèse préalable sur un modèle de la structure.
Ces paramètres devraient donc être d’un grand intérêt pour beaucoup d’études: différenciation
entre populations, lien avec la résistance et l’élasticité de l’os... D’ailleurs, il a déjà été
montré, sur des modèles de structures aléatoires, qu’il était possible de déterminer les
performances mécaniques à partir d’un indice de connectivité ([Feldkamp, 1988]).
A la différence des paramètres dits métriques (associés à une mesure de distance), les
paramètres topologiques sont liés à un dénombrement. Ils ne donnent donc aucune
information de position, de taille…
Les paramètres utilisés pour cette étude sont les trois premiers nombres de Betti et la
caractéristique d’Euler-Poincaré. Nous détaillerons dans un premier temps, les concepts,
définitions et propriétés associés à ces paramètres. Puis nous verrons les modifications liées à
leur application à une image discrète (maillage cubique). Ensuite, nous présenterons les
algorithmes utilisés et leur mise en œuvre particulière que nous avons établie par une étude
préalable sur une série d’images tests. Enfin, les applications récentes de ce type de paramètre
sur le tissu osseux seront discutées.
Dans cette partie, on notera X l’ensemble étudié et XC son complémentaire. Si Ω est l’espace
étudié (R3 dans notre cas), on a donc la relation suivante: X ∪ XC = Ω. Dans le cadre de
l’application sur le tissu trabéculaire, X représentera la phase osseuse et donc XC la phase des
pores.
II.4.1. Notion de topologie: définitions principales
II.4.1.1. Connexité
On dit qu’un ensemble X est connexe si pour tout couple de points (xi,xj) appartenant à X, on
peut au moins associer un chemin reliant xi à xj totalement inclus dans X (cf. Figure II.3).
63

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
64
xi
xj
X
Figure II.3: Illustration de la notion de connexité.
La connexité est la notion mathématique qui permet de définir si un ensemble est composé de
plusieurs amas. Elle est le concept principal de la caractérisation topologique des ensembles.
II.4.1.2. Genre d’une surface
Soit un chemin fermé défini sur une surface S. Si on découpe S le long de ce chemin fermé,
deux configurations sont possibles: la surface se retrouve partagée en deux « sous-surfaces »
connexes ou la surface reste connexe. Ces deux cas sont illustrés sur deux exemples en Figure
II.4.
A B
Figure II.4: Exemples de deux surfaces découpées selon un chemin fermé inclus. Pour une surface
sphérique (A), il n’existe aucun chemin fermé qui laisse la surface connexe: le genre est donc nul. Pour la
surface torique (B), il en existe un. Par contre, tout nouveau découpage sur un chemin fermé définira deux
surfaces connexes. Le genre est donc de 1.
On appelle « genre de la surface S », que l’on note g(S), le nombre maximal de coupures
successives (le long de chemins fermés) que l’on peut appliquer à S de telle façon que la
surface obtenue soit encore connexe ([Coster, 1989]). Ainsi, dans les deux exemples
précédents, on déduit facilement que la surface sphérique a un genre nul (cf. Figure II.4 A), et
la surface torique un genre de 1 (cf. Figure II.4 B). Pour le cas de la surface définie par un
« double tore » (dont une représentation est donnée en Figure II.10), le genre sera égal à 2 et

Chapitre II: Quantification de la microarchitecture trabéculaire
ainsi de suite: le genre donne donc directement une information sur la connectivité d’un
ensemble connexe.
II.4.2. Nombres de Betti
Pour un ensemble X dans un espace de dimension 3, on définit trois nombres de Betti dont
une présentation assez claire est donnée dans un article de Odgaard et Gundersen
([Odgaard, 1993]).
II.4.2.1. Définition
Le nombre de Betti d’ordre 0, β0(X), correspond au nombre de composantes connexes de X.
Le nombre de Betti d’ordre 1, β1(X), correspond au nombre de « connexions redondantes » de
X (nombre d’« anses »). Nous verrons par la suite (cf. II.4.3.3) que β1(X) est directement lié
aux genres des surfaces délimitées par X et donc à la connectivité de X.
Le nombre de Betti d’ordre 2, β2(X), correspond au nombre de cavités de X (ou de « surfaces
internes »). Une cavité de X est un amas connexe de XC isolé dans un amas connexe de X.
Dans le paragraphe suivant, nous donnons les propriétés reliant ces nombres ainsi que des
illustrations afin d’introduire ces notions dans le cadre du tissu trabéculaire.
II.4.2.2. Propriétés
Deux propriétés principales relient les nombres de Betti de X et de XC.
La première relation est la suivante:
1)()( 02 −= CXX ββ Eq. II.1
Celle-ci est un corollaire de la définition de β2. Dans l’exemple suivant où X est une sphère
creuse (cf. Figure II.5), XC est défini par la réunion de deux amas connexes: l’intérieur et
l’extérieur de la sphère creuse (notés respectivement I et E). On a donc β0(X) = 1 et
β0(XC) = 2. X a une cavité puisque I est un amas connexe de XC isolé dans X: β2(X) = 1. Par
contre XC n’en a pas: aucun amas connexe de X n’est isolé dans I ou dans E: β2(XC) = 0.
65

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
66
E
X
Cavité pour X
I
Figure II.5: Illustration d’une surface interne dans le cas où X est une sphère creuse. XC est donc défini
par: XC = I ∪ E où I et E sont respectivement l’intérieur et l’extérieur de la sphère creuse. X a donc une
cavité (du fait de I). Par contre, XC n’en a pas.
A partir de Eq. II.1, on peut exprimer le nombre N de surfaces séparant les deux phases par:
)()()()( 2020 CC XXXXN ββββ +=+= Eq. II.2
La deuxième relation concerne le nombre de Betti d’ordre 1:
)()( 11 XXC ββ = Eq. II.3
Celle-ci signifie que la connectivité n’est pas caractéristique d’un ensemble mais de la surface
séparant les deux phases complémentaires (comme on peut le voir d’après la définition du
genre, cf. II.4.1.2).
II.4.2.3. Illustrations
Pour illustrer les nombres de Betti et les relations précédentes, nous présentons le cas d’une
« mousse » qui est une structure poreuse formée de « cellules » isolées (topologie de type
« sphère creuse », cf. Figure II.6). Si on appelle X l’ensemble des parois et XC le gaz qui est
confiné dans les cellules, N serait très élevé à cause de la grande valeur de β2(X) (nombreuses
cavités). β0(X) serait forcément égal à 1 (aucune paroi ne peut se trouver isolée dans le gaz) et
β1(X) serait faible (très peu d’interconnexions de la structure).

Chapitre II: Quantification de la microarchitecture trabéculaire
67
≈ 100 µm
Figure II.6: Image d’une mousse de polycarbonate obtenue par microscopie électronique. La taille des
cellules varie entre 1 et 10 µm (image issue d’un résumé de D. Wulfson présenté au « 3rd nordic meeting on
materials and mechanics » de Mai 2000).
Par contre, le tissu trabéculaire est composé de travées osseuses formant un labyrinthe de
pores intercommuniquant (cf. I.1 et Figure I.2 B). On ne peut donc théoriquement avoir qu’un
seul amas connexe de tissu calcifié « plongé » dans un seul amas connexe constituant l’espace
des pores (i.e. aucun amas connexe de la phase poreuse ne peut se retrouver isolé dans la
phase osseuse), soit et . D’après Eq. II.2, il n’y a donc
théoriquement qu’une seule surface connexe d’intersection entre les deux phases. De plus, on
doit observer une valeur élevée de β
1)()( 00 == CXX ββ 0)()( 22 == CXX ββ
1(X).
Il existe des algorithmes pour calculer β0(X) (cf. II.4.5.1) à partir duquel on peut déterminer
β2(X) (d’après Eq. II.1). Cependant, on ne peut accéder directement à β1(X) qui est pourtant le
paramètre le plus approprié des trois nombres de Betti pour caractériser la microarchitecture
osseuse. C’est pourquoi on introduit un autre paramètre: la caractéristique d’Euler-Poincaré.
II.4.3. Caractéristique d’Euler-Poincaré
Nous renvoyons aux références suivantes pour plus de détails: [Coster, 1989] et [Serra, 1993].
II.4.3.1. Formulation de la caractéristique d’Euler-Poincaré
Soit X un ensemble qui est délimité par N surfaces connexes Si (i de 1 à N) et g(Si) le genre
associé à chaque surface Si. On définit la caractéristique d’Euler-Poincaré (ou nombre de
connexité) de X par:
∑=
−=N
iiSgXN
13 )](1[)( Eq. II.4

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Le Tableau II.1 présente le nombre de surfaces et la valeur de N3 pour différents exemples.
Ensemble X Nombre de surfaces N3(X)
Sphère creuse (Figure II.5) 2 de genre 0 chacune (1 - 0) + (1 - 0) = 2
Sphère (Figure II.4 A) 1 de genre 0 (1 - 0) = 1
Tore (Figure II.4 B) 1 de genre 1 (1 - 1) = 0
Double Tore (Figure II.10) 1 de genre 2 (1 - 2) = -1
Tableau II.1: Caractéristiques d’Euler-Poincaré pour des formes simples.
On peut remarquer qu’une structure interconnectée a une caractéristique d’Euler-Poincaré
négative dont la valeur absolue est d’autant plus grande que le nombre d’interconnections de
l’ensemble est grand.
II.4.3.2. Propriété d’additivité
La propriété principale de N3 est l’additivité. Si on a X = A ∪ B, alors
)()()()( 3333 BANBNANXN ∩−+= .
Cette propriété n’est pas vérifiée pour les nombres de Betti.
II.4.3.3. Relation entre les nombres de Betti et la caractéristique
d’Euler-Poincaré
La caractéristique d’Euler-Poincaré et les trois nombres de Betti sont reliés par la relation
suivante:
)()()()( 2103 XXXXN βββ +−= ,
ce qui peut s’écrire, d’après Eq. II.2,
)()( 13 XNXN β−= Eq. II.5
si N est le nombre de surfaces d’intersections entre les deux phases.
Comme l’expression de N3(X) (cf. Eq. II.4) ne dépend que des surfaces délimitant X qui ont
la même topologie que celles qui délimitent XC, on a, pour N3, le même type de relation que
pour β1 (cf. Eq. II.3):
)()( 33 CXNXN = Eq. II.6
68

Chapitre II: Quantification de la microarchitecture trabéculaire
En développant l’expression de N3 (cf. Eq. II.4), on peut établir facilement que:
∑=
−=N
iiSgNXN
13 )()( ,
soit, d’après Eq. II.5:
∑=
=N
iiSgX
11 )()(β .
β1(X) représente donc une sorte de « genre global » égal à la somme des genres des surfaces
d’intersections.
Dans le cas du tissu trabéculaire, on a N = 1 (cf. II.4.2.3):
)(1)( 13 XXN β−= Eq. II.7
Il y a donc égalité entre le genre de la surface d’intersection et le nombre de Betti d’ordre 1.
La caractéristique d’Euler-Poincaré évaluée sur le tissu trabéculaire doit donc être négative
(structure interconnectée).
II.4.4. Cas des espaces bornés et discrets
Les images sur lesquelles les paramètres sont calculés sont par définition des espaces bornés
et discrets. Ceci a des conséquences importantes pour pouvoir évaluer les paramètres
topologiques décrits ci-dessus de façon cohérente.
II.4.4.1. Approche locale
Une image est un espace borné et ne représente donc la structure que partiellement. On ne
peut donc évaluer que la contribution du sous-ensemble analysé aux paramètres topologiques
en considérant que celui-ci est représentatif de la structure. Comme on l’a vu en II.4.3.2, seule
la caractéristique d’Euler-Poincaré dispose de la propriété d’additivité. Or celle-ci est
essentielle pour pouvoir estimer la contribution locale de manière non biaisée c’est-à-dire en
prenant en compte les effets de bords. CN3, la contribution locale à N3 de l’image analysée,
est donc le seul paramètre réellement accessible pour estimer la connectivité du réseau
(c’est-à-dire le genre ou le nombre de Betti d’ordre 1).
II.4.4.2. Trame de l’image
La représentation de l’image est forcément de nature discrète. Nous ne discuterons ici que le
cas de la trame carrée en 2D ou cubique en 3D.
69

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
70
II.4.4.2.1. Modes de connexité
Supposons une travée osseuse « réelle » représentée en 2D (Figure II.7 A). Du fait de la
trame, la représentation de cette travée est discrète et donc forcément faussée. Après l’étape
de segmentation (cf. II.2), on obtient une série de pixels qui donne une approximation de la
travée réelle (Figure II.7 B).
Trame de l’image Travée réelle
A
B
Trame de l’image
Travée discrète
Connexions ?
Figure II.7: Travée osseuse « réelle » (A) imagée sur une trame carrée (exemple en 2D). Après
segmentation, la travée est représentée par une série de pixels (B). Suivant l’hypothèse faite pour définir la
connexion entre pixels, la travée qui est réellement d’un seul tenant, peut apparaître sur l’image soit
comme un seul amas connexe, soit comme la réunion ’amas connexes. d
La travée réelle est d’un seul tenant. Par contre, sur l’image discrète, la topologie de la travée
est fonction de l’hypothèse faite pour considérer deux pixels connectés. Sur l’exemple de la
Figure II.7 B, si on suppose que deux pixels sont connectés lorsqu’il existe au moins un
sommet en commun, la travée discrète sera définie par un seul ensemble connexe de 11
pixels. Par contre, si on exige une face commune pour considérer deux pixels connectés, cette
même travée « discrète » sera définie par trois amas connexes de respectivement 5, 2 et 4
pixels.
Cet exemple bidimensionnel simple traduit la nécessaire modification de la définition de la
connexité (cf. II.4.1.1) pour les ensembles discrets. Il faut en effet poser préalablement une loi
définissant la connexion des points entre eux pour pouvoir déterminer la connexité d’un
ensemble discret. Cette loi est appelée « mode de connexité ».
Dans un espace de dimension 2 discrétisé par un maillage carré (pixels), il existe deux modes
de connexité (comme on peut le supposer d’après la Figure II.7 B): 4-connexité (connexion
par une des 4 faces) et la 8-connexité (connexion par une des 4 faces ou un des 4 sommets).
En dimension 3 (voxels), on peut définir trois modes de connexité (cf. Figure II.8):

Chapitre II: Quantification de la microarchitecture trabéculaire
71
6-connexité (connexion par les 6 faces), 18-connexité (connexion par les arêtes) et
26-connexité (connexion par les sommets).
A B C
Figure II.8: Voxels appartenant au voisinage (du voxel central qui n’est pas représenté) au sens de la
6-connexité (A), de la 18-connexité (B) et de la 26-connexité (C). On peut remarquer que deux voxels
voisins au sens de la 6-connexité le sont aussi pour la 18-connexité et la 26-connexité. De même, deux
voxels voisins au sens de la 18-connexité le sont aussi u sens de la 26-connexité. a
On peut noter que le voisinage défini par la 6-connexité est inclus dans celui de la
18-connexité, lui-même inclus dans celui de la 26-connexité.
II.4.4.2.2. Phase complémentaire
Le mode de connexité utilisé pour étudier la phase complémentaire XC doit être cohérent avec
celui choisi pour X. Pour illustrer ce problème, on considère un « anneau » représenté dans un
espace de dimension 2 discrétisé par un maillage carré (cf. Figure II.9).
X XC
Figure II.9: X est un anneau représenté dans un espace de dimension 2 discrétisé par un maillage carré. Si
la 8-connexité est utilisée pour X, alors cet objet apparaît d’un seul tenant (un amas de 14 pixels). Dans ce
cas, XC doit être formée de deux amas connexes. Afin que cela soit respecté, XC doit être étudiée avec le
mode de connexité complémentaire à celui utilisé pour X, i.e. la 4-connexité.
Si on analyse X avec la 8-connexité, X est formé d’un seul amas connexe. Dans ce cas, cet
anneau doit séparer la phase XC en deux amas connexes. Pour que cela soit respecté malgré la
discrétisation de l’image, on ne peut analyser XC avec le même mode de connexité que celui
utilisé pour X: en effet, si on conservait la 8-connexité pour XC, on conclurait que XC est
composé d’un seul amas connecté ce qui irait à l’encontre de l’hypothèse faite sur X. Afin de

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
conserver la cohérence entre X et XC dans le cas d’espace discrets, il faut travailler avec deux
modes de connexité complémentaires, soit pour cet exemple, avec la 4-connexité pour XC.
Dans le cas de l’espace tridimensionnel discret à maillage cubique, les deux modes de
connexité naturellement complémentaires sont la 6-connexité et la 26-connexité: ce sont ces
deux modes qui ont été retenus pour notre étude. Cependant, le choix d’un des deux modes de
connexité pour la phase osseuse (en utilisant son complémentaire pour la phase poreuse) reste
un problème pour la quantification correcte des paramètres topologiques (cf. II.4.4.2.3).
Ainsi, les relations établies pour un espace continu entre β0(XC) et β2(X) (cf. II.4.2.2: Eq. II.1)
et entre CN3(X) et CN3(XC) (cf. II.4.3.3: Eq. II.6) deviennent dans un espace discret:
1)()( 60
262 −= CXX ββ Eq. II.8
)()( 63
263 CXCNXCN = Eq. II.9
En dimension 3, la topologie des surfaces délimitant X en 26-connexité est la même que
celles délimitant XC en 6-connexité.
Remarque:
Il existe d’autres trames d’image (par exemple de type hexagonal en 2D). Elles nécessitent de
même une définition précise du mode de connexité.
II.4.4.2.3. Conséquence: effet de la résolution
La différence entre les deux modes de connexité sera d’autant plus sensible que la taille
caractéristique des objets est proche de la taille des voxels.
La Figure II.10 présente deux objets de type « double tore » (ensemble délimité par une
surface de genre 2): dans un cas (A), le maillage de la trame est adapté à l’objet (i.e. la
résolution de l’image est « suffisante » et l’ensemble apparaît donc « épais »), dans l’autre cas
(B), le maillage est grossier (i.e. la résolution de l’image est limite et l’ensemble apparaît
beaucoup plus fin).
72

Chapitre II: Qu éculaire antification de la microarchitecture trab
73
A: CN3(X6) = CN3(X26) = -1 B: CN3(X6) = 27 ≠ CN3(X26) = -1
Figure II.10: Deux objets de type « double tore » (ensemble défini par une surface de genre 2) imagés avec
un maillage adapté (A) et avec un maillage grossier (B). Dans le cas A, la topologie de la surface délimitant
X est la même que celle délimitant XC quelque soit le mode de connexité envisagé. Par contre dans le cas B,
le mode de connexité influence considérablement le résultat.
Dans le cas de la Figure II.10 A, on trouve CN3(X6) = CN3(X26) = -1. Ceci traduit bien que la
topologie de la surface n’est pas affectée par le choix du mode de connexité: pour les deux
modes envisagés, on trouve une caractéristique d’Euler-Poincaré de -1 (cf. Tableau II.1).
Dans le cas de la Figure II.10 B, on trouve CN3(X6) = 27 ≠ CN3(X26) = -1. Si X est analysé
avec la 6-connexité, il apparaît formé de 27 amas connexes (que l’on peut deviner sur la
figure), chacun ayant un genre nul: CN3(X6) est donc égale à 27. Par contre, si on utilise la
26-connexité, on retrouve le résultat précédent. Cet exemple illustre l’influence considérable
que peut avoir le choix du mode de connexité sur la caractérisation topologique des ensembles
discrets en particulier dans le cas d’un maillage grossier (ce qui est le cas des images de tissu
trabéculaire acquis en limite de résolution).
II.4.5. Calcul des paramètres
II.4.5.1. Nombres de Betti d’ordre 0 et 2
L’algorithme que nous avons mis en œuvre pour calculer β0(X) est inspiré de celui proposé
par Gonzales et Woods ([Gonzalez, 1993]). Les amas connexes sont déterminés à partir
d’opérations de morphologie mathématique. Chaque amas A présent dans l’image est calculé
itérativement selon l’algorithme suivant:
Ak+1 = (Ak ⊕ B) ∩ I,
où ⊕ est l’opérateur de dilatation, B l’élément structurant et I, l’image. La condition d’arrêt
est simplement:
Ak+1 = Ak.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
L’élément structurant choisi permet de sélectionner un mode de connexité: pour un cube
3×3×3, on étudie la 26-connexité, pour une croix 3×3×3, la 6-connexité.
Outre la valeur de β0(X) (pour le mode de connexité choisi), chaque amas connexe de la phase
X est caractérisé par sa fraction volumique.
Pour le calcul de β2(X), il suffit d’appliquer le même algorithme mais sur la phase
complémentaire en veillant bien à prendre le mode de connexité adéquat (cf. II.4.4.2.2: Eq.
II.8).
II.4.5.2. Contribution à la caractéristique d’Euler-Poincaré
Quelques algorithmes ont été proposés dans la littérature pour évaluer, de manière non biaisée
(c’est-à-dire en corrigeant les effets de bords, cf. II.4.4.1), la contribution à la caractéristique
d’Euler-Poincaré CN3. Nous avons utilisé celui proposé par Vogel ([Vogel, 1997]) dont une
présentation succincte est donnée en Annexe 1.
Cet algorithme ne calcule CN3(X) qu’en 6-connexité. Pour évaluer la connectivité de X en
26-connexité, on applique donc ce même algorithme sur XC (cf. II.4.4.2.2: Eq. II.9).
Dans la suite, on considérera la contribution à la caractéristique d’Euler-Poincaré ramenée au
volume analysé afin d’obtenir la connectivité par unité de volume. CN3 sera donc exprimé soit
en voxel-1 soit en mm-3.
II.4.6. Tests et mise en œuvre des paramètres sur le tissu
trabéculaire
Nous avons utilisé 29 images tests provenant d’acquisitions réelles de tissu trabéculaire
obtenues par microtomographie X à très haute résolution (voxel isotrope de 30 µm de coté)
dans des conditions d’imagerie permettant une segmentation par seuillage simple sans
ambiguïté (cf. Figure II.1). A partir de ces données, nous avons pu calculer les dégradations
de ces images pour les tailles de voxels suivantes: 40 µm, 60 µm, 80 µm et 100 µm (la
méthode de dégradation sera détaillée dans le chapitre suivant, cf. III.4.3).
Le but de cette partie n’est pas d’estimer précisément les paramètres mais d’établir la validité
des algorithmes de calculs utilisés dans des conditions proches de celles dans lesquelles ils
seront appliqués.
74

Chapitre II: Quantification de la microarchitecture trabéculaire
On rappelle pour la suite que « X » représente la phase osseuse et donc « XC » la phase
poreuse. Le mode de connexité utilisé sera indiqué par un exposant (6 ou 26) sur X, le mode
de connexité complémentaire étant implicitement utilisé pour XC (cf. II.4.4.2.2).
II.4.6.1. Amas secondaires
Pour la gamme de résolution des images utilisées, les phases X et XC ne devraient
théoriquement être composées que d’un seul amas connexe chacune (cf. II.4.2.3). Ce n’est
cependant pas le cas pratiquement. A cause d’une résolution limite, d’erreurs de segmentation
ou d’artéfacts dans l’image, on observe que chaque phase est composée d’un amas principal et
d’une série d’amas secondaires qui sont, dans la plupart des cas, caractérisés par un genre nul.
L'influence de chaque amas secondaire de X est topologiquement équivalente à celle d’une
sphère ce qui cause l’incrémentation de 1 pour CN3(X) (cf. Tableau II.1). L’influence sur
CN3(X) des amas secondaires de genre nul de XC est la même puisque chacun d'entre eux
définit une surface interne dans l'amas principal de X (« sphère creuse », cf. Tableau II.1). Les
amas secondaires de genre nul conduisent donc à une surestimation de CN3. Pour améliorer
l’évaluation de la contribution à la caractéristique d’Euler-Poincaré, les amas secondaires (qui
doivent correspondre à une fraction volumique négligeable de l’amas principal) sont éliminés
([Last, 2001]). On se trouve alors avec une seule surface séparant les deux phases (cf. II.4.3.3:
Eq. II.7). Le paramètre CN3 peut donc être utilisé directement comme estimateur de la
connectivité du réseau trabéculaire.
A partir du calcul de β0(X) et de β2(X), on peut déterminer les amas principaux et secondaires
(en terme de fraction volumique) des phases X et XC (cf. II.4.5.1). Le Tableau II.2 présente la
valeur moyenne et l’écart type, calculés sur les 29 images, du rapport volume des amas
secondaires VAS par le volume de l’amas principal VAP de la phase osseuse (X) pour chaque
résolution étudiée. Les deux modes de connexité ont été envisagés.
75

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
VAS/VAP (%) 30 µm 40 µm 60 µm 80 µm 100 µm
X6 0.7 ± 0.5 0.9 ± 0.6 3.2 ± 2.6 19.7 ± 32 22.2 ± 23.1
X26 0.3 ± 0.2 0.4 ± 0.2 0.6 ± 0.5 0.9 ± 0.9 1.1 ± 0.9
Tableau II.2: Valeur moyenne et écart type (µ ± σ) du rapport entre le volume des amas secondaires et
celui de l’amas principal d’os pour chaque résolution et pour les deux configurations possibles pour les
modes de connexité (µ et σ ont été calculés sur les 29 images).
On peut observer que plus la résolution est grossière,
• plus les amas secondaires ont un volume important (jusqu’à 22% en 6-connexité)
• plus il y a de différence entre les hypothèses X6 et X26.
Ces résultats illustrent le problème soulevé par la Figure II.10. Plus la résolution se dégrade,
plus il y a de travées qui se retrouvent déconnectées de l’amas principal (ce qui fait augmenter
le volume des amas secondaires) et plus le choix du mode de connexité affecte la
caractérisation de la topologie de la structure. Il est cependant très intéressant de noter que la
proportion relative des amas secondaires en utilisant le mode de connexité X26 reste
négligeable quelle que soit la résolution.
II.4.6.2. Effet de la résolution sur CN3
Pour les 29 images précédentes, on a calculé CN3(X26) et CN3(X6) pour les 5 résolutions
disponibles. La Figure II.11 présente les évolutions obtenues pour trois échantillons
représentatifs.
76

Chapitre II: Quantification de la microarchitecture trabéculaire
77
-18-16-14
30 40 60 80 100
-12-10-8-6-4-2
CN3(X6) (mm-3)
Résolution (µm)
CN3(X
26)(mm-3)
-4
-3
-2
-1
30 40 60 80 100Résolution (µm)
A B Figure II.11: Evolution de CN3(X) en fonction de la résolution pour trois images représentatives. La figure
A correspond à l’hypothèse « X6 », la figure B à « X26 ». Chaque image est représentée par une courbe
avec un motif particulier: cercle, carré et triangle.
La croissance de CN3 quand la résolution se dégrade est quasi systématique pour toutes les
images évaluées avec l’hypothèse « X26 ». Ceci est cohérent avec les observations faites en
II.4.4.2.3: on a une perte d’interconnections de la structure quand la résolution devient
grossière.
Par contre, avec l’hypothèse « X6 », on a une évolution beaucoup plus « chaotique ». On
observe en effet pour beaucoup d’images, une décroissance sensible de CN3 (en particulier au
passage de 40 à 60 µm). Ceci est une particularité de la 6-connexité. Au fur et à mesure que la
résolution se dégrade, les travées osseuses apparaissent de plus en plus « fines » sur l’image.
Beaucoup se trouvent alors composées d’éléments du type de celui présenté en Figure II.12.
Figure II.12: Ensemble composé de 6 voxels formant deux « L » tête bêche. Ces voxels ont un sommet en
commun.
Dans cet exemple, les 6 cubes composant cet amas ont un sommet commun. En 26-connexité,
cet amas est donc de genre nul. Par contre, ce sommet commun n’établit aucune connexion en

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
6-connexité: avec ce mode de connexité, cet amas a la même topologie qu’un tore donc un
genre de 1. Ceci augmente artificiellement le nombre d’interconnections de la structure ce qui
fait diminuer CN3. Pour des tailles de voxels supérieures, on observe souvent une forte
croissance qui est à mettre en parallèle avec les résultats sur la fraction d’amas secondaires
présentés dans le Tableau II.2 (cf. II.4.6.1). Pour les résolutions grossières, l’amas principal
n’est plus représentatif de la structure: on a donc une perte de la connectivité due à
l’élimination des amas secondaires (qui ne sont plus négligeables) ce qui fait augmenter CN3.
Il faut donc bien veiller à ce que la fraction volumique des amas secondaires reste négligeable
pour pouvoir effectuer le filtrage.
Pour chaque image, nous avons calculé l’écart type σ des valeurs de CN3 sur les 5 résolutions
disponibles (pour les deux modes de connexité). La valeur moyenne de σ calculée sur les 29
images, illustrent ce que l’on peut observer sur la Figure II.11. Alors qu’elle est de 2.5 mm-3 si
on applique la 6-connexité sur X, elle n’est que de 0.6 mm-3 avec la 26-connexité: les
évolutions de CN3 avec la 26-connexité appliquée sur la phase osseuse sont donc beaucoup
plus stables qu’avec la 6-connexité.
II.4.6.3. Protocole
Malgré la dégradation de la résolution, nous avons vu que la 26-connexité appliquée sur la
phase osseuse permet de conserver des amas secondaires de volume négligeable (cf. II.4.6.1)
et assure une bien meilleure stabilité de l’évaluation de CN3 que la 6-connexité (cf. II.4.6.2).
Le calcul des paramètres topologiques a donc été mené selon le protocole suivant:
• calcul de β0(X26)
• filtrage des amas secondaires d’os (en ayant vérifié qu’ils ont un volume négligeable par
rapport au volume de l’amas principal)
• calcul de β2(X26) d’après Eq. II.8
• filtrage des amas secondaires de XC isolés dans X (en ayant vérifié qu’ils ont un volume
négligeable par rapport au volume de l’amas principal de X).
• calcul de la contribution à la caractéristique d’Euler-Poincaré en appliquant l’algorithme
sur XC pour obtenir la connectivité de X en 26-connexité (cf. II.4.5.2). La valeur obtenue
est divisée par le volume analysé afin d’obtenir une mesure de la connectivité de la région
d’intérêt CN3 en mm-3 ou voxel-1
78

Chapitre II: Quantification de la microarchitecture trabéculaire
II.4.7. Applications des paramètres topologiques
De nombreuses études topologiques ont déjà été menées sur le réseau trabéculaire. Celles-ci
montrent le grand intérêt de cette famille de paramètres pour la caractérisation de la
microarchitecture du tissu.
A partir d’un modèle à éléments finis, appliqué sur des images de tissu trabéculaire (radius et
vertèbres) obtenues par microtomographie X haute résolution (20 µm isotrope), Kinney et
Ladd ([Kinney, 1998]) ont étudié les relations entre les propriétés mécaniques et la
connectivité du réseau trabéculaire (évaluée avec le même type de paramètres topologiques
que ceux décrits dans cette partie). Ils n’ont trouvé aucun lien direct entre la topologie de la
structure et les propriétés mécaniques. Cependant, en simulant un cycle de remodelage osseux
(cf. I.2) déficitaire, ils ont observé un lien entre la perte d’élasticité résultante et la perte de
connectivité. Les auteurs ont conclu que pour qu’un tissu trabéculaire « pathologique » puisse
conserver ses fonctions mécaniques, il fallait préserver la connectivité du tissu.
D’autres paramètres topologiques ont été définis à partir du calcul du squelette de la structure
trabéculaire. Le squelette est une représentation simplifiée du réseau trabéculaire ayant les
mêmes propriétés topologiques. Par exemple, le squelette d’une sphère est un point, celui
d’un cylindre est son axe de révolution, celui d’une plaque d’épaisseur non nulle, son plan
central.
Saha et al. ([Saha, 2000]) ont proposé une méthode pour associer une classe, parmi 9
possibles, à chaque voxel du squelette afin de caractériser sa topologie locale. Les classes
possibles sont les suivantes: isolé, surface ou courbe intérieure, bord de surface ou extrémité
de courbe, jonctions (courbe-courbe, surface-surface ou courbe-surface) et profil. A partir de
ces considérations, ils ont défini des index topologiques (comme le rapport surface sur
courbe). Les auteurs montrent qu’avec ces index, il est possible de suivre la transformation de
la microstructure d’un modèle en plaque vers un modèle en colonne (cf. I.4), transformation
qui survient souvent quand la densité osseuse diminue, avec l’âge ou dans les cas
d’ostéoporose.
Dans une autre étude ([Pothuaud, 2002 (b)]), il a été montré que des paramètres topologiques
déterminés à partir du squelette peuvent avantageusement compléter des modèles prédictifs
des propriétés mécaniques du tissu trabéculaire basés sur la fraction osseuse.
79

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Sur des biopsies de crête iliaque, Legrand et al. ([Legrand, 2000]) ont aussi montré que la
caractérisation topologique du réseau trabéculaire, calculée à partir du squelette, permettait de
différencier, sur un groupe de 108 individus atteint d’ostéoporose lombaire (T-score < -2.5,
cf. I.4), ceux présentant au moins une fracture vertébrale de ceux n’en présentant aucune.
Enfin, à l’aide d’un modèle tridimensionnel du tissu trabéculaire, des simulations de perte
osseuse ([Guo, 2002]) ont montré qu’à fraction osseuse constante, l’absence de travées avait
des conséquences bien plus importantes sur le module d’Young et la résistance du tissu que
leur amincissement. D’après les auteurs, ceci illustre l’importance du nombre de travées
osseuses et de la connectivité dans les propriétés mécaniques du tissu trabéculaire.
II.5. Paramètres histomorphométriques
Cette famille de paramètres a pour but de caractériser la morphologie de l’architecture du
tissu trabéculaire. Elle a été initialement développée pour l’analyse de coupes histologiques.
Les concepts ont ensuite été adaptés pour les images tridimensionnelles.
Trois paramètres sont présentés dans cette partie. Le premier, surface spécifique, estime le
rapport entre la surface de tissu calcifié et le volume de tissu trabéculaire. Ce facteur est
important car il est directement lié à la capacité de renouvellement du tissu osseux du fait que
le processus de remodelage de l’os se déroule au niveau de l’interface entre l’os et la moelle
(cf. I.2). Les deux autres paramètres sont l’épaisseur et l’espacement des pores. Intuitivement,
on comprend qu’ils peuvent s’avérer pertinents pour caractériser l’état de l’os puisque la
détérioration de l’architecture osseuse peut se manifester par un amincissement des travées et
par un agrandissement de l’espacement entre travées.
La première partie concerne la surface spécifique qui est le paramètre central de cette famille.
Après avoir exprimé la surface d’un ensemble, nous présentons les modifications liées au cas
de l’image discrète. Ensuite, à partir de calculs sur une série de formes simples, nous
proposons un algorithme optimisé pour l’évaluation de ce paramètre. Dans la deuxième partie,
nous indiquons la méthode employée pour déterminer l’épaisseur et l’espacement des travées
à partir de la surface spécifique. Les limites de cette technique sont aussi discutées. Enfin dans
la dernière partie, les résultats de différents travaux sur le tissu trabéculaire menés à l’aide de
cette famille de paramètres sont présentés.
80

Chapitre II: Quantification de la microarchitecture trabéculaire
81
De même que dans la partie II.4, on notera X l’ensemble étudié et XC son complémentaire
(respectivement les phases osseuse et des pores dans notre application).
II.5.1. Surface spécifique
II.5.1.1. Formulation de la surface d’un ensemble
Pour plus de détails sur ce paragraphe, nous renvoyons le lecteur au premier chapitre du livre
de Coster et Chermant ([Coster, 1989]). Afin d’introduire les concepts, nous présentons tout
d’abord le cas de R2 avant de passer à R3.
II.5.1.1.1. Etude préalable dans R2
Supposons un repère orthonormé (O,x,y) dans lequel se trouve un faisceau de droites
parallèles inclinées d’un angle θ par rapport à (Ox) et espacées les unes des autres d’une
distance d. L’intersection de ce faisceau de droites avec un ensemble X placé dans cet espace
est une réunion de segments dont le cardinal N1(X,θ) est égal au nombre de transitions de XC
à X (ou de X à XC). N1(X,θ) est le nombre de connexité de X dans l’espace R1, calculé dans la
direction θ. Une illustration est proposée en Figure II.13.
X
XC
O
y
xθ
TransitionsXC → X
d
Figure II.13: Ensemble X sur lequel est superposé un faisceau de droites parallèles inclinées de θ par
rapport à (Ox) et espacées d’une distance d. Le nombre de segments issus de l’intersection du faisceau de
droites avec X est égal au nombre de transitions de XC à X: N1(X,θ). D(X,θ) = N1(X,θ)×d est la projection
linéaire totale de X dans la direction θ.
L’espacement des droites étant de d, la longueur de la projection orthogonale de X sur une
droite de direction θ + π/2, D(X,θ) est donnée par:

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
dXNXD ×= ),(),( 1 θθ Eq. II.10
D(X,θ) est appelée la projection linéaire totale de X dans la direction θ.
On peut alors déterminer le périmètre L de X en utilisant la relation de Cauchy. On obtient
alors:
dXNEdXDXL ××==∫ ))((),()( 10
πθθπ
,
où E(N1(X)) est l’espérance mathématique de N1(X,θ) sur la direction du faisceau de droites.
II.5.1.1.2. Généralisation à R3
Dans un repère (O,x,y,z), chaque direction du faisceau de droites est déterminée par un couple
d’angles ω = (θ,ϕ) où θ est l’inclinaison par rapport à (Ox) et ϕ celle par rapport à (Oz). On
note aussi d1 et d2 les espacements de deux droites successives du faisceau selon les deux axes
du plan perpendiculaire à la direction ω. On définit alors A(X,ω), la projection surfacique
totale de X dans la direction ω, par:
aXNXA ×= ),(),( 1 ωω Eq. II.11
où a est l’élément de surface correspondant à chaque droite (a = d1×d2).
Selon le théorème de Cauchy, lorsque ω varie, l’espérance mathématique des projections
surfaciques totales est égale au quart de la surface S de X. On a donc:
aXNEdXAXS ××== ∫ ))((4),(1)( 1
4
0
π
ωωπ .
II.5.1.2. Cas des espaces bornés et discrets
De même que pour les paramètres topologiques (cf. II.4.4), on doit tenir compte du fait que
les images sont des espaces bornés et discrets.
II.5.1.2.1. Approche locale
L’espace dans lequel les calculs seront effectués est nécessairement borné. On ne peut donc
accéder entièrement à X mais seulement à l’intersection de X avec le champ de vue analysé.
On considère que l’image est représentative de X. Les paramètres seront calculés par unité de
surface dans R2 ou de volume dans R3.
Dans R2, si AI est l’aire de l’image analysée, on définit la projection linéaire totale de X par
unité de surface dans la direction θ d’après Eq. II.10:
82

Chapitre II: Quantification de la microarchitecture trabéculaire
),(),(),( 1 θθθ XNA
dXNAXD
LII
=×= ,
où NL(X,θ) est le nombre de connexité par unité de longueur des droites du faisceau.
Le périmètre est alors remplacé par le périmètre spécifique LA(X) avec:
))((),()(0
XNEdXDXL LAA ×==∫ πθθπ
.
De même dans R3, si VI est le volume de l’image analysée, on définit la projection surfacique
totale de X par unité de volume dans la direction ω d’après Eq. II.11:
),(),(),( 1 ωωω XNV
aXNVXA
LII
=×= .
On retrouve là aussi le nombre de connexité par unité de longueur des droites du faisceau.
La surface est alors remplacée par la surface spécifique SV(X) avec:
))((4),(1)(4
0
XNEdXNXS LLV ×== ∫π
ωωπ Eq. II.12
C’est donc SV que l’on cherche à calculer sur les images de structures trabéculaires. Nous
utiliserons dans la suite sa notation courante: « BS/TV » qui signifie
« Bone Surface / Tissu Volume ».
Remarques:
a) NL(X,ω) est fonction du nombre de transitions dans la direction ω. On a donc
NL(X,ω) = NL(X,-ω). Pour le calcul de BS/TV, on peut donc se limiter à l’analyse d’un seul
hémisphère: par exemple pour 0 ≤ θ < π et 0 ≤ ϕ < π.
b) En général, les longueurs analysées des droites du faisceau ne sont pas les mêmes d’une
direction à une autre. Par exemple, pour une image parallélépipédique rectangle, la longueur
balayée dans la direction d’une diagonale est plus grande que celle obtenue dans la direction
correspondant à un des côtés. Afin de ne pas favoriser artificiellement certaines directions, il
faut limiter le volume d’analyse à une sphère pour calculer NL(X,ω).
II.5.1.2.2. Trame de l’image
Si on appliquait directement la formulation de BS/TV (cf. II.5.1.2.1: Eq. II.12) au cas d’une
image discrète (limitée à une sphère), il faudrait mener le calcul suivant:
]),(4[lim)(/1
0
1
0,
, ∑∑−
=
−
=∞→∞→
××=M
m
N
nnmLNM
XNNMXTVBS ω ,
83

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
84
où ωm,n est la direction analysée qui est fonction de deux pas angulaires: ωm,n = (θm,ϕn) avec
Mmmπθ ×= et Nnn
πϕ ×= (soit M×N directions étudiées). Pratiquement, ceci signifierait qu’il
faudrait analyser toutes les directions possibles, du moins celles qui seraient définies à partir
du plus petit pas angulaire significatif, et compter toutes les transitions entre la structure
osseuse et le faisceau de droites avec un espacement minimum des droites.
Une telle approche ne serait pas valable pour plusieurs raisons.
En choisissant ωm,n de cette manière, certaines directions seraient privilégiées du fait de la
trame de l’image (en particulier autour des pôles).
De plus, la surface calculée intégrerait toutes les faces des voxels situés en bordure de la
structure osseuse. On se rapprocherait donc de la mesure de la surface spécifique « discrète »
qui est d’autant plus biaisée par rapport à la surface spécifique théorique que la forme
analysée n’est pas adaptée au maillage de l’image. Ceci conduirait alors à une surestimation
de BS/TV. Une illustration de ce problème pour le périmètre spécifique (dimension 2) est
donnée sur la Figure II.14.
Périmètre « discret »
X Périmètre théorique
Figure II.14: Illustration de la surestimation du périmètre discret (trait épais) par rapport au périmètre
théorique (trait pointillé). Une différence du même type est observée sur les surfaces (en dimension 3).
Ensuite, des amas de voxels isolés dans l’image (dus par exemple à des erreurs de
segmentation ou à des artéfacts) seraient pris en compte dans le calcul de la surface et
entraînerait là aussi une surestimation de BS/TV.
Enfin, si le nombre de directions analysées est trop grand, le temps de calcul pourrait devenir
prohibitif.
Dans la partie suivante, nous détaillons les différents facteurs permettant d’optimiser le calcul
de la surface spécifique.

Chapitre II: Quantification de la microarchitecture trabéculaire
II.5.1.3. Méthode de calcul de BS/TV
II.5.1.3.1. Principe de l’algorithme
Avant tout calcul, le volume à analyser est restreint à la plus grande sphère incluse (cf.
remarque b) de II.5.1.2.1).
Pour étudier chaque direction, l’image est tournée (avec une rotation utilisant une
interpolation « par arrondi ») et le calcul du nombre d’intersections est effectué selon le même
axe (nous avons choisi arbitrairement l’axe x). Ainsi, la longueur totale balayée sur les droites
du faisceau est identique quelle que soit la direction analysée puisqu’elle est déterminée à
partir du même axe du voxel. Pour chaque direction, N1 est évalué comme le nombre total
d’intersections (X → XC et XC → X, cf. Figure II.13) divisé par 2. BS/TV est alors calculé à
partir de la valeur moyenne, sur toutes les directions, de N1 divisé par la longueur totale des
droites du faisceau inclus dans la sphère (selon l’expression Eq. II.12).
Les paramètres de l’algorithme sur lesquels on peut agir sont:
• l’espacement des droites du faisceau. On supposera d1 = d2 = d (cf. II.5.1.1.2 pour les
notations). On a étudié dans la suite deux cas: d = 1 et d = 2.
• le nombre de directions analysées. Pour le limiter, une première approche consiste à
analyser une série d’orientations particulières du faisceau de droites. Cependant, il existe
un risque d’erreur si la structure présente une périodicité qui coïncident avec les directions
analysées. Afin d’éviter cela, on choisit les directions aléatoirement.
• le comptage des intersections i.e. le pas de déplacement (noté s) sur les droites du faisceau.
Si s est petit, on risque d’intégrer la contribution de groupes de voxels isolés et donc de
surestimer BS/TV. La technique de filtrage des amas secondaires, développée dans le
cadre de l’étude des paramètres topologiques (cf. II.4.6.1), permet de s’assurer d’une
image débruitée. On peut donc utiliser s minimum: s = 1 voxel.
II.5.1.3.2. Images « tests »
Nous avons testé ces facteurs sur des images représentant des formes géométriques pour
lesquelles la valeur théorique de la surface spécifique est connu. Elles ont été choisies pour
tester les algorithmes dans différentes situations: adaptation plus ou moins bonne de l’objet à
la trame, anisotropie plus ou moins prononcée. Ces formes ont été étudiées pour des tailles
caractéristiques allant de quelques voxels pour simuler une étude en limite de résolution à une
dizaine de voxels pour une étude à haute résolution.
85

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Le Tableau II.3 présente les différentes expressions de la « surface sur volume » (S/V) ainsi
que les tailles des grandeurs caractéristiques de ces formes.
Forme Surface Volume S/V Gamme étudiée
Sphère (R) 4πR2 4πR3/3 3/R 2 ≤ R ≤ 10 voxels
Cube (a) 6a2 a3 6/a 2 ≤ a ≤ 10 voxels
Cylindre (R,h) 2πR2 + 2πRh πR2h 2×(1/R + 1/h) 2 ≤ R,h ≤ 10 voxels
Tableau II.3: Valeurs théoriques du paramètre « surface sur volume » (S/V) pour une sphère de rayon R,
pour un cube de côté a et pour un cylindre de rayon R et de hauteur h. On précise aussi les tailles étudiées
(en voxels).
Le paramètre BS/TV calculé par l’algorithme correspond en fait à S/VI où S est la surface de
la structure et VI le volume de l’image. Pour s’affranchir de la taille de la zone d’étude, la
valeur trouvée est divisée par FV, la fraction volumique de l’objet (FV = V/VI). Ainsi, on peut
directement comparer cette valeur à la valeur théorique de S/V:
V
TOTTOT
TOT FV
S
VV
VS
VS =×= .
II.5.1.3.3. Résultats
Le premier paramètre que nous avons étudié est le nombre de directions. Pour chaque forme
et pour différentes tailles, nous avons calculé S/V pour un nombre de directions variant de 50
à 1000 (par pas de 50). Ces directions ont été choisies aléatoirement. Le rapport entre l’écart
type et la valeur moyenne calculé était inférieur à 1% quelque soit les valeurs de d envisagées.
Nous avons donc choisi d’utiliser pour la suite 100 directions prises aléatoirement.
Pour comparer les paramètres de l’algorithme, nous avons évalué S/V sur chaque forme, pour
différentes tailles. L’erreur relative entre la mesure et la valeur théorique est
systématiquement positive pour la sphère et le cylindre: ceci traduit que BS/TV est surestimé
pour des objets peu adaptés à la représentation discrète (cf. Figure II.14). Pour le cube, dans le
cas où d = 2, l’erreur relative est systématiquement négative: comme la forme est adaptée à la
trame cubique, le fait de ne prendre qu’une droite sur deux dans chaque direction sous-estime
la valeur de la surface. Les erreurs relatives sont généralement plus élevées lorsque la taille de
l’objet est petite. Ceci est principalement dû aux changements de morphologie de la forme
86

Chapitre II: Quantification de la microarchitecture trabéculaire
aux petites tailles: par exemple un sphère de rayon 2 correspond à une croix dans les trois
plans principaux (ce qui est évidemment très différent d’une sphère). Nous avons aussi
calculé la régression linéaire entre S/V théorique et S/V mesuré afin d’établir l’efficacité des
méthodes. Dans le cas idéal, on devrait obtenir un coefficient directeur a de 1 et une ordonnée
à l’origine b de 0. Les résultats sont résumés dans le Tableau II.4.
100 directions aléatoires, d = 1 100 directions aléatoires, d = 2 Forme
a b (voxel-1) R2 a b (voxel-1) R2
Sphère 1.17 0.006 0.99 1.04 0.041 0.99
Cube 0.63 0.29 0.96 0.60 0.29 0.98
Cylindre 1.12 -0.01 0.97 1.10 -3×10-5 0.96
Tableau II.4: Coefficient directeur a, ordonnée à l’origine b et coefficient de détermination R2 des
ajustements entre S/V théorique et mesuré, calculés pour des mesures correspondantes à différentes tailles
des objets.
Les coefficients de détermination sont tous très élevés ce qui traduit que la forme des
expressions analytiques de S/V sont respectées pour chaque objet.
Aucune des deux méthodes ne se distingue nettement. Le cube est une forme problématique
pour les deux algorithmes. Cependant d = 2 semble plus efficace pour la sphère et le cylindre
(ordonnée à l’origine négligeable et coefficient directeur légèrement meilleur que pour le cas
d = 1).
Nous avons donc retenu le protocole de calcul suivant
• restriction du volume d’analyse à la plus grande sphère incluse
• filtrage des amas secondaires
• calcul de BS/TV avec 100 directions choisies aléatoirement, s = 1 et d = 2.
Dans un article de 1997, Simmons et Hipp ([Simmons, 1997]) ont étudié l’influence des
différentes variables de l’algorithme sur l’évaluation de la « densité de travées » (Trabecular
Number ou TbN), paramètre qui correspond, à un facteur 2 près, à la surface spécifique. Leurs
calculs ont été menés sur de véritables acquisitions (échantillons de tissu trabéculaire de bœuf,
imagerie par IRM avec une résolution de 61 µm isotrope): ils ne disposaient donc pas de
valeurs de référence pour déterminer « l’algorithme correct ». Le fait que nous avons tiré des
conclusions similaires de notre étude sur des formes tests semble valider l’algorithme retenu.
87

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
88
II.5.2. Epaisseur et espacement des travées
L’épaisseur et l’espacement des travées osseuses semblent des paramètres pertinents pour la
caractérisation de la microarchitecture trabéculaire. Cependant, ces notions sont
particulièrement difficiles à définir du fait de la forme complexe que la structure trabéculaire
peut prendre. Deux méthodes de calculs sont habituellement admises: nous les présentons ici.
II.5.2.1. Calcul indirect: hypothèse sur le modèle de tissu trabéculaire
Deux modèles sont couramment admis pour la structure trabéculaire (cf. I.4): un modèle en
plaques et un en colonnes. En fonction de l’hypothèse faite sur l’un ou l’autre de ses modèles,
des formulations simples de l’épaisseur et l’espacement des travées ont été développées à
partir de considérations de stéréologie. Elles sont reprises dans le rapport de l’ABSMR déjà
cité ([Parfitt, 1987]).
En dimension 3, on évalue:
• l’épaisseur des travées: notée « Tb.Th » (« Trabecular Thickness ») dans le cas d’un
modèle en plaques ou « Tb.Dm » (« Trabecular Diameter ») dans le cas d’un modèle en
colonnes,
• l’espacement des travées: « Tb.Sp » (« Trabecular spacing ») valables pour les deux
modèles.
Les formulations proposées sont données dans le Tableau II.5 en fonction du modèle de la
structure.
Modèle « Epaisseur » « Espacement »
en plaques TVBSTVBVThTb /
/2. ×= TVBSTVBVSpTb /
/12. −×=
en colonnes TVBSTVBVDmTb /
/4. ×= ]1/1
4[.. −××= TVBVDmTbSpTb π
Tableau II.5: Formulations de l’épaisseur et de l’espacement des travées en fonction du modèle utilisé
(d’après le rapport de l’ABSMR, [Parfitt, 1987]). Dans le cas du modèle en colonnes, l’épaisseur des
travées est appelée « diamètre ». BV/TV est la fraction volumique osseuse (cf. II.3)
Le calcul de l’épaisseur et de l’espacement des travées est donc particulièrement simple avec
l’hypothèse d’un modèle. Par contre celle-ci n’est que très rarement vérifiée dans la pratique,
la structure étant en général beaucoup plus complexe. Ces paramètres sont donc peu

Chapitre II: Quantification de la microarchitecture trabéculaire
significatifs. Pourtant, ils sont couramment évalués ainsi dans la littérature. Nous les avons
donc calculés pour information mais ils seront peu discutés dans la suite.
II.5.2.2. Autres approches: méthodes de calcul direct
Les méthodes indirectes supposent non seulement que la structure soit conforme à un modèle,
mais aussi que l’épaisseur des travées soit relativement constante. Or le simple examen visuel
d’images de tissu trabéculaire montre clairement que les modèles sont très approximatifs et
que la dimension des travées fluctue beaucoup.
C’est pourquoi de nombreux travaux ont été menés pour définir de manière plus précise les
notions d’épaisseur et d’espacement de travées. Ces grandeurs sont en général calculées à
partir de la carte des sphères maximales r(x,y,z) d’un ensemble X ([Hildebrand, 1997]). Pour
chaque voxel (x,y,z) appartenant à X, r(x,y,z) est égal au rayon de la plus grande sphère
incluse dans X et qui contient (x,y,z). En calculant le double de la valeur moyenne de r(x,y,z)
sur tous les voxels (x,y,z) de la phase osseuse, on obtient une estimation de l’épaisseur des
travées. Cette carte, déterminée sur la phase poreuse, permet aussi d’accéder à l’espacement
des travées.
Pateyron ([Pateyron, 1998]) a développé une mesure de l’épaisseur locale des travées à partir
du squelette de la structure trabéculaire (notion que nous avons abordée en II.4.7). En
calculant une carte des distances euclidiennes d(x,y,z) sur chaque voxel du squelette (d(x,y,z)
est égale au rayon de la plus grande sphère centrée en (x,y,z) et incluse dans la phase
osseuse), on obtient la demi épaisseur locale de la travée. A partir de ces mesures, on établit
un histogramme, traduisant la distribution des épaisseurs, qui peut être utilisé pour discriminer
différentes architectures: structures comportant de nombreuses travées d’épaisseur régulières
ou structures plus complexes comportant à la fois des travées épaisses et fines…. La valeur
moyenne de la distribution peut aussi servir d’estimation de l’épaisseur des travées.
Dans une étude assez récente ([Hildebrand, 1999]), des comparaisons ont été faites entre
méthodes directe et indirecte sur des images de microtomographie X de prélèvements de tissu
trabéculaire provenant de quatre sites anatomiques différents. Le calcul de Tb.Th, en
supposant un modèle en plaques, sous-estimait systématiquement l’épaisseur des travées
même pour des sites où ce modèle est couramment admis (comme la tête fémorale:
sous-estimation de 17%). Sur la vertèbre lombaire (généralement associée à un modèle en
colonne), le diamètre, calculée de manière indirecte, était surestimé de 41%. Pour les auteurs,
89

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
ces erreurs montrent qu’une structure ne peut être uniquement associée soit à des plaques soit
à des colonnes d’où l’intérêt d’utiliser les méthodes de mesures directes. Des conclusions
identiques ont été tirées d’une autre étude ([Day, 2000]) sur des prélèvements de tissus
trabéculaires humains et animaux. Les auteurs ont de plus établi un lien entre le biais observé
sur l’évaluation de Tb.Th en utilisant le modèle en plaque et la fraction osseuse.
II.5.3. Applications des paramètres histomorphométriques
Les paramètres histomorphométriques sont certainement les plus fréquemment utilisés pour la
caractérisation de la microarchitecture osseuse.
Par exemple, une étude déjà citée ([Hildebrand, 1999]) a montré une grande variation d’une
série d’indices histomorphométriques en fonction du site osseux analysé. Ainsi, la tête
fémorale a été caractérisée par des travées osseuses épaisses organisées principalement en
plaques. Pour la vertèbre lombaire, les auteurs ont observé une faible fraction osseuse et une
structure plutôt de type colonne. Les indices trouvés pour le calcanéum étaient proches de
ceux de la vertèbre excepté une fraction osseuse légèrement supérieure. Les mesures
effectuées sur la crête iliaque ne se sont pas distinguées des valeurs trouvées pour les autres
sites ce qui semble indiquer que la structure est d’un type intermédiaire à celle des vertèbres
et de la tête fémorale.
Dans une autre étude ([Oleksik, 2000]), les paramètres histomorphométriques calculés sur des
prélèvements de crête iliaque se sont avérés plus discriminants que la fraction osseuse pour
distinguer, dans une population ostéoporotique caractérisée par un T-score < -2.5 (cf. I.4), un
sous-groupe de personnes ayant déjà été touché par des fractures vertébrales, d’un autre
sous-groupe sans fracture.
Plus récemment ([Thomsen, 2002 (b)]), une adaptation du modèle des plaques a été proposée
pour distinguer, sur des coupes histologiques de vertèbres, les travées horizontales des travées
verticales et ainsi pouvoir les caractériser séparément (calcul de Tb.Th, Tb.Sp et BS/TV). Les
auteurs ont étudié les liens de ces paramètres avec le vieillissement. Ils ont établi que
l’épaisseur des travées verticales ne changeait pas avec l’age à la différence de celle des
travées horizontales. Pour les deux types de travées, l’espacement augmente sensiblement
avec l’âge.
90

Chapitre II: Quantification de la microarchitecture trabéculaire
II.6. Paramètres d’anisotropie et d’orientation
Les forces de pression et de traction auxquelles est soumis le tissu trabéculaire influencent
considérablement la morphologie du réseau (cf. I.4). Dans les régions soumises à de fortes
contraintes, le tissu trabéculaire se développe plutôt avec un modèle de plaques alors que dans
les régions où elles sont plus faibles, les travées semblent suivre un modèle en colonnes.
L’orientation de la structure dépend elle aussi des charges appliquées. Ainsi, pour des sites où
la répartition des contraintes est complexe, la structure trabéculaire est plutôt isotrope alors
que pour un tissu trabéculaire soumis principalement à une contrainte, les travées osseuses se
développent avec une symétrie axiale autour de l’axe de la contrainte.
La dernière famille de paramètres que nous avons développée a pour objectif de caractériser
l’anisotropie et l’orientation du réseau trabéculaire. Ces paramètres sont tous déterminés à
partir de la modélisation de la longueur moyenne d’interception par un ellipsoïde. Nous
présenterons donc dans un premier temps cette grandeur et sa modélisation avant de définir
les paramètres mesurés. L’étude de l’architecture trabéculaire étant menée dans un espace de
dimension 3, deux coefficients et trois angles sont nécessaires pour caractériser l’anisotropie
et l’orientation de la structure. Enfin nous indiquerons quelques travaux récents qui ont utilisé
ce type de paramètres.
II.6.1. Longueur moyenne d’interception
La longueur moyenne d’interception (notée MIL: « Mean Intercept Length ») d’un ensemble
X dans la direction ω = (θ,ϕ) est définie à partir du nombre de connexité par unité de longueur
NL(X,ω) que nous avons introduit dans le cadre des paramètres histomorphométriques (cf.
II.5.1.2.1):
),(21),(
ωω
XNXMIL
L×= .
Dans une étude sur des coupes de tissu trabéculaire, Whitehouse ([Whitehouse, 1974]) a
montré expérimentalement que le tracé en coordonnées polaires de la longueur moyenne
d’interception (définie dans R2) suivait une forme proche d’une ellipse. A partir des
caractéristiques de cette ellipse (obtenues par un ajustement), il a proposé une formulation de
l’anisotropie et de l’orientation de la structure trabéculaire. En généralisant ce résultat à R3, on
cherche donc à modéliser MIL(X,ω) par un ellipsoïde.
91

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
II.6.2. Modélisation par un ellipsoïde
Cette modélisation est basée sur celle de Harrigan et Mann ([Harrigan, 1984]) qui ont proposé
une description de l’anisotropie du réseau trabéculaire à l’aide d’un tenseur dérivé de
l’ellipsoïde de MIL(X,ω).
Si on note , le vecteur unitaire portant la direction ω = (θ,ϕ), on peut associer un
vecteur u à MIL(X,ω) avec
),,( zyx nnnnr
r
nXMILu vr×= ),( ω .
Comme MIL(X,ω) décrit un ellipsoïde, la colonne U associée au vecteur u vérifie donc
l’expression matricielle suivante:
r
TU×ME×U = 1,
où ME est la matrice associée à la forme ellipsoïdale:
⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢
⎣
⎡
=
cfe
fbd
eda
ME .
On peut donc écrire:
2),(1ωXMILNMN E
T =×× ,
si N est la colonne associée au vecteur unitaire nr .
ME est un tenseur du deuxième ordre décrivant l’anisotropie. Cette matrice est symétrique et
réelle. Elle est donc diagonalisable dans R3 et la base formée par les vecteurs propres est
orthogonale.
Le principe de calcul des paramètres d’anisotropie et d’orientation est donc basé sur
l’ajustement de l’équation précédente sur les mesures de MIL(X,ω) par la méthode des
moindres carrées ([Press, 1989]) afin d’estimer ME et de calculer sa diagonalisation.
Les valeurs propres (λ1,λ2,λ3) permettent de caractériser l’anisotropie de la structure. A partir
de la position angulaire de la base des vecteurs propres normalisés ),,( 321 VVVrrr
par rapport au
repère de l’image, on peut déterminer les trois angles d’Euler c’est-à-dire l’orientation du
réseau trabéculaire.
On supposera dans la suite (Figure II.15) que:
• λ3 = min(λ1,λ2,λ3)
• les couples (λ1, ) et (λ1Vr
2, ) sont tels que le trièdre 2Vr
),,( 321 VVVrrr
soit direct.
92

Chapitre II: Quantification de la microarchitecture trabéculaire
93
ir
jr
kr
1
1
λVr
2
2
λVr
3
3
λVr
Figure II.15: Illustration de l’ellipsoïde dans le référentiel de l’image ),,( kjirrr
. On indique aussi la base
des vecteurs propres. On suppose que λ3 = min(λ1,λ2,λ3) et que le trièdre ),,( 321 VVVrrr
est direct.
II.6.3. Anisotropie de l’ellipsoïde
On définit à partir des trois valeurs propres (λ1,λ2,λ3), les trois longueurs moyennes
d’interception principales MIL1, MIL2 et MIL3 par:
)1,1min(21
1λλ
=MIL , )1,1max(21
2λλ
=MIL et 3
31λ
=MIL .
Ainsi, sous les hypothèses précédentes (cf. II.6.2), on a donc:
MIL1 ≤ MIL2 ≤ MIL3.
3Vr
porte donc la direction où l’anisotropie du réseau est la plus forte puisque c’est la direction
où on trouve la longueur moyenne d’interception la plus grande donc le nombre
d’intersections par unité de longueur le plus faible. Il correspond donc à l’axe principal de
l’ellipsoïde.
A partir de MIL1, MIL2 et MIL3, on définit deux coefficients caractérisant l’anisotropie de la
structure RMIN et RMAX tels que:
3
1
MILMILRMIN= et ⎥⎦
⎤⎢⎣⎡=
3
2
2
1 ,maxMILMIL
MILMILRMAX .
Ces rapports d’anisotropie sont forcément dans l’intervalle compris entre 0 et 1. Une structure
aura une anisotropie d’autant plus prononcée que ces rapports seront proches de 0. RMIN et
RMAX correspondent respectivement aux rapports minimal et maximal entre les trois longueurs
moyennes d’interception principales. Ils illustrent donc respectivement l’anisotropie la plus

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
forte et la plus faible de la structure. Par exemple, une structure fortement anisotrope dans une
seule direction se traduira par un coefficient RMIN très faible et RMAX proche de 1.
II.6.4. Orientation de l’ellipsoïde
Pour déterminer la position angulaire de la base des vecteurs propres (cf.
hypothèses précédentes présentées en II.6.2) par rapport au référentiel de l’image
),,( 321 VVVrrr
),,( kjirrr
, on
cherche la rotation R telle que:
1)( ViRrr
= , 2)( VjRrr
= , 3)( VkRrr
=
Pour déterminer R, il faut considérer les trois angles d’Euler (θ,ϕ,ψ) dont une définition est
donnée dans le livre de Gourret ([Gourret, 1994]). Nous rappelons en Annexe 2 la définition
de ces trois angles ainsi que la méthode de calcul employée.
II.6.5. Applications
Sugita et al. ([Sugita, 1999]) ont étudié le comportement mécanique d’échantillons prélevés
sur la tête fémorale pour deux types de contraintes, longitudinale et transversale, pour
modéliser l’effort sur le tissu respectivement pendant la marche ou lors d’une chute. Ils ont
établi que lorsque la fraction osseuse diminue, la résistance de l’os diminue dans les deux
axes mais de manière moins flagrante dans la direction transversale. Pour les auteurs, cette
différence de propriétés mécaniques qui est fonction de la direction de la contrainte est causée
par l’anisotropie prononcée de la microarchitecture du tissu trabéculaire fémoral.
A partir d’images de microtomographie X d’échantillons de tissu trabéculaire provenant de
crête iliaque, de vertèbre lombaire et de fémur, Ulrich et al. ([Ulrich, 1999]) ont comparé, en
fonction du site analysé, la capacité des paramètres de microstructure à prédire certaines
constantes d’élasticité du tissu évaluées grâce à un modèle à élément finis. Alors que le
coefficient de détermination R2 n’était que de 0.53 en utilisant un modèle basé uniquement sur
la fraction osseuse, ils ont pu observer une nette amélioration s’ils utilisaient des modèles de
régression multivariables. Le meilleur jeu de paramètres était fonction du site étudié: pour la
tête fémorale, les meilleures prédictions ont été obtenues pour le couple fraction osseuse et
coefficients d’anisotropie (R2=0.82).
94

Chapitre II: Quantification de la microarchitecture trabéculaire
L’analyse de l’orientation de la structure trabéculaire est très peu discutée dans la littérature.
Ceci est principalement du à la difficulté d’effectuer des calculs sur des zones d’intérêt dans la
même position (ce qui est essentiel pour toute comparaison d’orientation de structure).
95


Chapitre III: Validité du rendu par IRM du tissu trabéculaire
Chapitre III: Validité du rendu par IRM du
tissu trabéculaire: confrontation avec une
technique de référence, la
microtomographie X par rayonnement
synchrotron
Les symboles des paramètres utilisés dans ce chapitre sont ceux présentés dans le chapitre II.
Nous renvoyons donc à celui-ci pour toute explication sur leur formulation et leur
signification.
III.1. Etat de l’art de l’étude in vitro par IRM du tissu
trabéculaire et problématique
III.1.1. Caractérisation du tissu trabéculaire in vitro par IRM
L’IRM est une voie d’investigation particulièrement intéressante puisque cette technique est
non invasive et permet d'obtenir des images tridimensionnelles de résolution potentiellement
isotrope (cf. I.6). L’IRM haute résolution in vitro a donc été utilisée dans de nombreuses
97

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
études pour démontrer la possibilité de caractériser le tissu trabéculaire à l’aide de paramètres
de microarchitecture, à des résolutions spatiales proches de celles accessibles in vivo. Nous
résumons les résultats les plus pertinents ci-dessous.
Majumdar et al. ([Majumdar, 1998]) ont étudié les propriétés mécaniques du tissu trabéculaire
à partir d’images IRM (résolution spatiale de 1172×300 µm3 obtenue à 1.5 T) d’échantillons
de calcanéum (os du talon), de fémur et de vertèbre. Sur chaque échantillon, ont été mesurés
la densité minérale osseuse (par tomographie X) ainsi que le module d’Young (par test non
destructif). Après segmentation des images (dont la méthode, détaillée dans
[Majumdar, 1996], est présentée succinctement en II.2.3.1), une série de paramètres
structuraux comme BV/TV, Tb.Th, Tb.Sp, Tb.N (paramètre égal à un facteur près à BS/TV,
cf. II.5.1.3.3) a été évaluée. Des variations significatives entre les différents sites étudiés ont
été observées pour les paramètres de microarchitecture, la densité minérale osseuse et les
propriétés mécaniques. Des modèles prédictifs du module d’Young ont été proposés avec
beaucoup plus d’efficacité quand des paramètres de structure étaient intégrés (par rapport à
l’utilisation de la densité minérale osseuse seule).
La même année ([Link, 1998 (a)]), cette équipe a aussi publié une étude similaire (images
IRM d’échantillons de fémur et de vertèbre, acquises à une résolution spatiale de
117×156×300 µm3 sur le même imageur et segmentées avec le même algorithme) dans
laquelle il a été observé que le tissu trabéculaire fémoral était beaucoup plus hétérogène que
celui prélevé sur les vertèbres. De plus, des corrélations significatives entre les paramètres
histomorphométriques et le module d’Young ont été trouvées, plus élevées dans les vertèbres
que dans le fémur.
Borah et al. ([Borah, 2000]) ont aussi montré qu’il était possible de différencier un groupe de
vertèbres « jeunes » d’un autre de vertèbres « adultes » (prélèvements effectués sur des
cochons) à l’aide de paramètres de microarchitecture calculés sur des images IRM obtenues à
4.7 T avec une résolution spatiale isotrope de 85 µm. Ces auteurs ont aussi remarqué que
l’utilisation de paramètres de microarchitecture (en plus du contenu osseux) améliorait
sensiblement la qualité des modèles de prédictions des propriétés mécaniques.
Plus récemment, Beuf et al. ([Beuf, 2001]) ont utilisé des images IRM de vertèbres lombaires
(obtenues à 1.5 T avec une résolution spatiale de 1172×300 µm3) pour calculer une série de
paramètres: Tb.Th, Tb.Sp, Tb.N, indice de connectivité et MIL. Ces paramètres
morphologiques étaient très étroitement liés à l’âge, au contenu osseux et aux propriétés
98

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
mécaniques, la plus forte corrélation était celle observée entre la contrainte verticale et l’index
de connectivité.
III.1.2. Limitations de ces études
Ces résultats très encourageants montrent le grand intérêt de l’IRM pour étudier le tissu
trabéculaire. On peut cependant remarquer que tous ces travaux ont été menés en limite de
résolution, i.e. la taille de voxel était toujours proche de celle des travées osseuses. Cette
limitation, principalement due à la faible sensibilité de la technique d’imagerie, fait que
l’évaluation de paramètres microarchitecturaux reste discutable. Ainsi, à partir d’échantillons
de tibias et de radius, Majumdar et al. ([Majumdar, 1995]) ont montré l’influence notable
d’une série de facteurs techniques sur l’évaluation de paramètres de structure par IRM: champ
magnétique statique (1.5 T et 4.7 T), séquence d’imagerie (Gradient Echo et Spin Echo) et
leurs paramètres, épaisseur de coupe (variant entre 128 et 1000 µm), résolution dans la coupe
(entre 1282 et 1562 µm2), méthodes de segmentation.
Ceci soulève le problème de la fiabilité des évaluations. En effet, préalablement à toute étude,
il faut estimer le degré de confiance que l’on peut attribuer aux calculs de paramètres sur des
images obtenues par IRM.
III.1.3. Fiabilité de l’IRM pour la caractérisation du tissu
trabéculaire
Pour répondre à cette question, quelques études comparatives ont déjà été menées, dans
lesquelles les mêmes paramètres de structure étaient évalués d’un part par IRM, et d’autre part
par une autre technique d’imagerie prise comme référence.
Ainsi, Hipp et al. ([Hipp, 1996]) ont comparé l’IRM à l’imagerie optique sur 16 échantillons
cubiques de tissu trabéculaire prélevés sur l’humérus, le fémur et la vertèbre de bœuf. Des
paramètres de structure ont été évalués par des méthodes tridimensionnelles sur les images
obtenues par IRM (sur un imageur à 8.6 T, avec une résolution de 92 µm isotrope) et par des
méthodes bidimensionnelles sur les images des 6 faces de chaque échantillon récupérées par
une caméra CCD (résolution de 23×23 µm2). Les évaluations entre les deux modalités étaient
99

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
reliées linéairement, les coefficients de détermination respectivement le plus élevé et le plus
faible étant obtenus pour BV/TV (R2 = 0.81) et pour Tb.N, c’est-à-dire BS/TV (R2 = 0.53).
Dans une autre étude basée sur 7 échantillons cubiques de radius ([Majumdar, 1996]), des
images IRM obtenues à 1.5 T avec une résolution de 1562×300 µm3 ont été comparées à des
acquisitions de microtomographie X avec une résolution de 18 µm isotrope dégradées à la
résolution des images IRM. Mis à part le paramètre de connectivité β1 (qui a été estimé sur
tout le volume de l’échantillon), la valeur de chaque paramètre a été calculée à partir de
mesures bidimensionnelles (valeur moyenne du paramètre sur l’ensemble des coupes
centrales de l’échantillon pour un volume total analysé représentant la moitié de celui-ci).
Pour ces conditions d’acquisition IRM, BV/TV et Tb.Th étaient surestimées d’un facteur 3, β1
d’un facteur 10 tandis que Tb.Sp était sous-estimé d’un facteur 1.6.
Une autre étude comparative sur le même type d’image a aussi été proposée ([Laib, 2001]):
images IRM acquises sur le même imageur à deux résolutions différentes (1562×300 µm3 et
1562×500 µm3) de 15 échantillons cubiques prélevés sur le radius, comparées à la
microtomographie X avec une résolution de 34 µm isotrope. Les deux différences majeures
entre cette étude et la précédente sont que la comparaison entre les deux modalités n’est pas
faite à même résolution et que les paramètres histomorphométriques sont évalués par la
méthode de calcul direct (cf. II.5.2.2). Pour la comparaison entre les deux modalités, aucune
différence notable n’a été trouvée entre les deux résolutions étudiées pour l’IRM. Tout
comme l’étude précédente, les auteurs ont noté la surestimation de BV/TV et la
sous-estimation de Tb.Sp par IRM mais dans des proportions bien inférieures (respectivement
1.5 et 0.8).
La même année, Vieth et al. ([Vieth, 2001]) ont comparé des images IRM de 30 calcanéa
(obtenues à 1.5 T avec deux résolutions: 1952×300 µm3 et 1952×900 µm3) avec la
radiographie X. Afin de s’assurer d’analyser les mêmes zones d’intérêt entre les deux
modalités, ils ont disposé des marqueurs (petits cylindres contenant du produit de contraste)
dans chaque calcanéum. Les coupes radiographiques (900 µm d’épaisseur avec une résolution
dans les coupes de 25 µm isotrope) ont été positionnées dans la partie centrale de chaque
marqueur. Une corrélation élevée a ainsi été observée entre les deux modalités pour certains
paramètres histomorphométriques (en particulier BV/TV et Tb.Sp), cette corrélation étant
d’autant plus forte que les coupes IRM analysées étaient fines. Cependant, une très forte
dépendance de leurs résultats au seuil utilisé pour la segmentation (leurs images présentaient
100

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
un histogramme monomodal d’où une grande difficulté pour déterminer un seuil « correct ») a
aussi été notée.
III.1.4. Problématique et présentation du travail
Le problème de ces études comparatives est que de nombreux facteurs influencent les
résultats et donc les conclusions (qui fluctuent d’une étude à l’autre). On peut citer les zones
analysées qui ne sont pas les mêmes entre les deux modalités, les échantillons qui proviennent
de sites osseux différents, les paramètres évalués en dimension 2 sur une modalité et en
dimension 3 sur l’autre, les modalités qui sont analysées avec des résolutions différentes, la
segmentation (les études citées ci-dessus utilisent une technique de seuillage et la plupart
précisent que les résultats sont fortement dépendant du seuil choisi), l’utilisation de voxels
anisotropes (ce qui peut influencer l’évaluation de certains paramètres en particulier ceux liés
à l’anisotropie et à l’orientation du réseau trabéculaire) ou encore un nombre d’échantillons
limité qui réduit les degrés de signification des résultats obtenus.
Aucune étude de comparaison de l’IRM à une technique de référence n’a donc décorrélé les
différences dues aux phénomènes physiques à l’origine du contraste de celles dues aux
conditions d’imagerie et aux traitements effectués sur les images. Or ceci est nécessaire pour
connaître les précautions avec lesquelles on doit considérer les résultats des évaluations
effectuées sur des images obtenues par IRM.
Ce chapitre a donc pour but d’établir une comparaison quantitative fiable entre l’IRM et une
modalité faisant référence dans la représentation de l’architecture tridimensionnelle du tissu
trabéculaire: la microtomographie X à haute résolution par rayonnement synchrotron. Cette
technique d’imagerie permet d’obtenir une représentation très précise de l’architecture
tridimensionnelle du tissu trabéculaire mais ne serait pas applicable in vivo du fait de son
caractère fortement ionisant (cf. I.5.4).
Nous avons obtenu deux séries d’images tridimensionnelles à haute résolution isotrope (une
par modalité) des mêmes échantillons de tissu trabéculaire prélevés sur le calcanéum humain.
Après avoir identifié pour chaque échantillon, une région d’intérêt commune aux deux
modalités (recalage), une méthode de segmentation robuste adaptée à chaque technique
d’imagerie a été appliquée. Les mêmes paramètres ont ensuite été évalués sur chaque
modalité de la même manière (i.e. en utilisant les mêmes algorithmes de calcul) afin d’établir
une comparaison fiable des deux modalités d’imagerie. De plus, une étude sur la signification
101

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
102
des paramètres calculés a aussi été menée pour essayer d’expliquer certains résultats, ce qui
nous a permis de déterminer les paramètres les plus pertinents de cette étude. Enfin,
l’importance du recalage des régions d’intérêt est aussi discutée.
III.2. Les échantillons utilisés
29 échantillons cylindriques ont été prélevés dans les parties antérieures et postérieures de 15
calcanéa humains dégraissés, à l’aide d’une mèche creuse. Les donneurs étaient composés de
6 hommes et 9 femmes, âgés de 69 à 89 ans. Aucune information n’était disponible sur leur
état pathologique.
Les caractéristiques géométriques de chaque échantillon sont les suivantes: 7 mm de diamètre
et environ 15 mm de hauteur (en fonction du calcanéum) soit un volume d’environ 580 mm3
de tissu trabéculaire. Un exemple est donné en Figure III.1.
A B
Figure III.1: A: photographie de l’os du talon humain, le calcanéum (environ 4 cm sur 3 cm). On peut
observer qu’il est principalement composé de tissu trabéculaire mise à part son enveloppe qui est de type
cortical. B: échantillon cylindrique prélevé, de diamètre 7 mm et de hauteur environ 15 mm.
L’axe du cylindre correspond à l’axe médio-latéral du pied. Cependant, la position exacte de
l’échantillon prélevé dans le calcanéum n’a pas été conservée. Ceci n’est pas gênant compte
tenu de l’objectif de l’étude: le but est d’effectuer une comparaison entre deux modalités et
non d’étudier la position absolue des travées par rapport à un axe donné.
Pour le protocole d’imagerie, chaque échantillon a été placé dans un porte-échantillon adapté
(plexiglas).

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
III.3. Les images
Les 29 échantillons ont été imagés par microscopie RMN dans l’unité (UMR #8081, Orsay) et
par microtomographie X haute résolution par rayonnement synchrotron à l’European
Synchrotron Radiation Facility (ESRF, Grenoble) dans le cadre d’une collaboration avec
Madame Françoise Peyrin de CREATIS (UMR #5515).
III.3.1. Images obtenues par IRM
Comme le tissu osseux a des temps de relaxation très courts (phase solide), il a une
contribution au signal négligeable. En IRM, le réseau trabéculaire est donc imagé en
« négatif »: les pores que l’on remplit préalablement d’eau apparaissent en haut niveau de gris
dans l’image (signal) tandis que le tissu calcifié apparaît en faible niveau de gris (bruit). Pour
limiter la présence de bulles d’air (artéfact qui se présente dans l’image comme du bruit du
fait que l’air ne donne pas de signal par RMN), on place l’échantillon dans un cristallisoir
rempli d’eau sous vide pendant quelques minutes.
L’appareil sur lequel ont été réalisées les images de microscopie RMN est un prototype qui a
été développé dans le laboratoire ([Ruaud, 1991]). Le champ statique est de 8.5 T, soit une
fréquence de Larmor de 360 MHz pour le proton (cf. I.6.1.1). L’ouverture de l’aimant est de
32 mm ce qui permet des champs de vue d’environ 2 cm. Le gradient maximal disponible est
de 220 mT.m-1.
La contrainte de temps d’acquisition n’étant pas critique pour une étude in vitro, la séquence
qui a été utilisée est une « écho de spins » 3D (cf. I.6.3.5) afin de limiter l’effet des artéfacts
de susceptibilité (cf. I.6.4.3.1).
III.3.1.1. Optimisation des paramètres d’acquisition
Compte tenu du temps de montée des gradients, le temps d’écho TE minimum accessible est
de 7 ms. De plus, pour moyenner certains défauts (comme les inhomogénéités de l’impulsion
180°), le nombre d’accumulation NACC doit être au moins de 4.
103

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Les images qui ont été utilisées dans cette étude ont été réalisées par un stagiaire en utilisant
le protocole développé dans la thèse d’Antoniadis ([Antoniadis, 1998 (b)]). Nous détaillons
succinctement dans la suite la méthode utilisée.
L’objectif du protocole d’imagerie est d’obtenir un voxel isotrope ayant une taille de l’ordre
de 100 µm (à cause de la taille caractéristique des travées osseuses) pour un Rapport Signal à
Bruit (RSB) d’environ 10, valeur minimale pour que l’algorithme de segmentation que nous
avons utilisé pour cette étude ([Antoniadis, 1998 (a)], cf. III.5.1) puisse fonctionner
correctement. Le temps d’acquisition doit aussi rester raisonnable (inférieur à une heure).
L’expression du RSB pour une séquence « Spin écho » est donnée par la formule suivante:
ACCTT
TT
NVeeRSBER
0211
−−
⎟⎟
⎠
⎞
⎜⎜
⎝
⎛−∝
où TR est le temps de répétition, T1 et T2 les deux constantes de relaxation et V0 le volume du
voxel. Or le temps d’acquisition TACQ, est relié à NACC par la relation:
TACQ = NACC×Ny×Nz×TR,
où Ny et Nz sont le nombre de voxels selon les deux axes de codage de la phase. On peut alors
montrer que la valeur TR optimisant le RSB (à TACQ constant), maximise l’expression:
R
TT
T
eR
⎟⎟
⎠
⎞
⎜⎜
⎝
⎛−
−11
.
Dans le cas de l’eau pure (T1 = 4 s, T2 = 2 s), la valeur optimale de TR est d’environ 5
secondes. Cette valeur conduirait, pour une matrice de taille 64×64 dans les deux axes de
codage de phase, à un temps d’acquisition d’environ 22 heures ce qui serait prohibitif. Afin de
limiter ce temps, un agent paramagnétique (solution aqueuse de Mn2EDTA) est utilisé pour
réduire T1 ce qui permet de diminuer TR. La concentration utilisée est de 1 mMol.
III.3.1.2. Paramètres de la séquence
Les paramètres de la séquence d’acquisition ont été fixés ainsi:
• TE/TR = 8/200 ms
• une bande passante de 55 kHz
• un champ de vue de 8.4×8.4×16.9 mm3 ce qui permet d’observer l’échantillon entier (sans
repliement)
104

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
• une taille de la matrice d’acquisition de 64×64 selon les deux axes de codage et de 128 sur
celui de lecture, ce qui conduit à un voxel isotrope de 1323 µm3 (2.3×106 µm3), pour un
temps d’acquisition d’environ une heure.
III.3.1.3. Calcul des images
Une interpolation des données est ensuite effectuée par la méthode du « zerofill » d’un facteur
2 dans chaque dimension. Cette technique consiste à ajouter des zéros sur les bords du plan de
Fourier acquis (parties réelle et imaginaire avant le calcul de l’image par transformée de
Fourier) jusqu’à doubler sa taille dans les trois dimensions (la matrice passe de 64×64×128 à
128×128×256 voxels). Il a en effet été montré ([Bernstein, 2001]) que les données situées
dans les coins du plan de Fourier participent utilement à la résolution en appliquant ce type
d’interpolation.
Après calcul de la transformée de Fourier sur ces données, on obtient un voxel isotrope de
66 µm de coté (0.3×106 µm3) et un RSB de l’ordre de 17, ce qui est conforme au cahier des
charges.
III.3.1.4. Résultats
La Figure III.2 présente les trois coupes principales de l’image 3D d’un échantillon.
105

Caractérisation de l’architecture tridim r IRM ensionnelle du tissu trabéculaire pa
106
y
z
16.9 mm
x
z
x
y 8.4 mm
A B C Figure III.2: Trois coupes principales d’une image 3D obtenue par microscopie RMN: longitudinale « X »
(A), longitudinale « Y » (B) et transversale (C). Le champ statique 0Br
est porté par y, la perturbation
radio-fréquence par z (axe de la sonde). Les deux axes de codage de phase (128 voxels chacun) sont x
et y et celui de la lecture, z (256 voxels). On peut remarquer que l’échantillon, légèrement incliné, est
entièrement disponible sur ces images.
1Br
L’histogramme des images (dont un exemple est donné en Figure III.3) est bimodal,
caractéristique d’une image IRM acquise sans volume partiel. Une modélisation précise du
mode associé aux voxels de bruit (tissu calcifié) a été proposée par Gudbjartsson et Patz
([Gudbjartsson, 1995]). En première approximation ([Chung, 1995]), on peut admettre que la
distribution des niveaux de gris dans l’image peut être représentée par la somme d’une
distribution de Rayleigh pour les voxels de bruit et d’une distribution gaussienne pour ceux de
signal (eau, espace des pores).

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
107
0
0,005
0 50 100 150 200 250NG
0,01
0,015
0,02
0,025
Occurrence normalisée
Figure III.3: Exemple d’histogramme calculé sur toute l’image. Cette distribution peut être modélisée par
la somme d’une distribution de Rayleigh pour les faibles niveaux de gris (bruit) et d’une distribution
gaussienne pour les niveaux de gris élevés (signal).
Sur la zone de l’image correspondant à l’échantillon, la sensibilité de la sonde utilisée est
relativement homogène. On peut malgré tout observer des différences le long de son axe (axe
de lecture): plus on s’éloigne du centre de la sonde, plus le niveau de gris correspondant à la
moyenne de la distribution gaussienne sur les histogrammes des coupes transversales
diminue. Par contre, on peut considérer la réponse de la sonde uniforme dans ces plans.
Dans la suite, le symbole utilisé pour référencer les images acquises par microscopie RMN
sera « µIRM ».
III.3.2. Images obtenues par microtomographie X
(rayonnement synchrotron)
III.3.2.1. Microtomographie X utilisant le rayonnement synchrotron
Le principe de la tomographie X a déjà été présenté succinctement au chapitre I (cf. I.5.4). La
microtomographie X par rayonnement synchrotron est une technique particulière qui utilise le
rayonnement généré par un accélérateur à particules comme source de rayons X.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
108
III.3.2.1.1. Le rayonnement synchrotron
La Figure III.4 donne une représentation simplifiée du dispositif disponible à l’ESRF
(Grenoble). Dans un premier temps, les électrons produits par un filament chauffé sont
accélérés dans un accélérateur linéaire qui les amènent à une énergie de 200 MeV. Ensuite ils
subissent une autre accélération dans le synchrotron qui les fait passer à une énergie de
6 GeV. Ils sont enfin injectés dans l’anneau de stockage (dont la circonférence est de 844
mètres) où leur trajectoire est maintenue circulaire par une série d’aimants de courbure
disposés dans l’anneau.
Anneau de stockage
Synchrotron
Accélérateur linéaire
Lignes de lumière
Figure III.4: Représentation simplifiée du synchrotron de l’ESRF (Grenoble). L’anneau de stockage fait
844 m de circonférence. Une trentaine de lignes de lumière sont disposées tangentiellement à celui-ci. Les
acquisitions ont été faites sur la ligne ID19: « Topogr phy and high resolution diffraction beamlight ». a
A chaque passage dans ces aimants, les électrons décélèrent et perdent une partie de leur
énergie sous la forme d’un « rayonnement synchrotron » émis tangentiellement à leur
trajectoire. Ce rayonnement est récupéré au sein de « lignes de lumière » (une trentaine sur le
site de l’ESRF) qui sont positionnées autour de l’anneau de stockage.
III.3.2.1.2. Instrumentation pour l’imagerie
Les expériences d’imagerie se déroulent à l’extrémité d’une de ces lignes, la ligne ID19:
« Topography and high resolution diffraction beamlight ». Sur cette ligne de lumière, se
trouvent successivement un obturateur de faisceau, un monochromateur (pour sélectionner la
longueur d’onde désirée), le support de positionnement des échantillons (selon les trois axes)

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
et la chaîne de détection. Cette dernière comprend notamment un scintillateur, un dispositif
d’agrandissement optique (formé de deux objectifs) et une caméra CCD très rapide de
1024×1024 pixels (19 µm de coté, dynamique de 14 bits). Le scintillateur a pour but de
convertir l’image de rayons X en une image « visible » par le capteur. Cet élément
conditionne la résolution spatiale. Plusieurs scintillateurs sont disponibles: la résolution
spatiale la plus petite que l’on peut obtenir correspond à une taille de pixel de 1.23 µm de coté
(montage encore au stade expérimental). Pour l’imagerie du tissu trabéculaire, la
configuration choisie permet d’obtenir un pixel de 10×10 µm2, soit un champ de vue cubique
d’environ 10 mm de coté.
III.3.2.1.3. Avantages de cette méthode de tomographie
Les principaux avantages du rayonnement synchrotron sont:
• le flux de photons qui est très élevé: de 103 à 104 fois plus grand que celui d’un tube à
rayons X
• le spectre du rayonnement qui est très large et continu (à la différence des sources
classiques): on peut donc sélectionner une longueur d’onde optimale tout en conservant un
flux très important
Pour plus de détails sur cette modalité d’imagerie, nous renvoyons au chapitre 4 de la Thèse
de Pateyron ([Pateyron, 1998]).
Comme on l’a vu en I.5.4, de nombreuses études ont montré l’efficacité de la
microtomographie X pour caractériser le tissu trabéculaire. A partir de biopsies de crête
iliaque, Nuzzo et al. ([Nuzzo, 2002]) ont montré que la microtomographie X par rayonnement
synchrotron donnait une représentation très fiable de la microarchitecture osseuse ainsi qu’une
quantification précise du degré de minéralisation. On peut donc considérer cette technique
comme une référence, d’autant plus que dans les conditions de résolution et de RSB des
acquisitions (cf. III.3.2.2), la segmentation (point crucial pour l’analyse des images, cf. II.2)
peut être réalisée sans ambiguïté.
III.3.2.2. Les acquisitions
La méthode d’acquisition utilisée a été décrite dans un article de Salomé et al.
([Salomé, 1999]).
Les images ont été obtenues en utilisant un faisceau de rayons X monochromatique de 20 keV
extrait du rayonnement synchrotron avec le monochromateur. Pour balayer l’intervalle
109

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
angulaire nécessaire (180°), une projection est acquise tous les 0.2° ce qui conduit à 900
projections pour chaque échantillon. Ce nombre de projections est légèrement supérieur à
celui requis pour pouvoir effectuer correctement la reconstruction. De plus, deux types
d’images ont été aussi récupérés en début et en fin d’acquisition. Le premier correspond à une
image de référence du faisceau (sans objet, obturateur ouvert et même temps d’exposition)
afin d’obtenir une mesure du nombre de photons incidents en fonction de la position dans le
plan de projection (c’est-à-dire N0, cf. I.5.4): cette image permet de corriger les
inhomogénéités du faisceau. Le deuxième correspond à une image du courant d’obscurité du
capteur (acquisition avec l’obturateur fermé pendant le même temps de pose) afin de
quantifier le bruit du capteur.
Le temps de pose total pour l’acquisition des 900 projections est d’environ 1 heure. Le temps
nécessaire et les tailles mémoires utilisées pour reconstruire les images sont très importants.
Afin de les limiter, la reconstruction n’a été faite que sur une partie de la matrice
d’acquisition: 660×660 pixels (au lieu des 1024×1024 disponibles). L’algorithme utilisé est
celui de la rétroprojection filtrée. Grâce à la parallélisation du calcul sur l’ensemble des
serveurs en réseau à l’ESRF, le temps pour reconstruire nos volumes de données a été réduit à
quelques heures par image.
Pour chaque échantillon, on disposait donc d’une image de 660×660×660 voxels, avec une
résolution de 10 µm isotrope (soit un champ de vue de 6.63 mm3) et un RSB d’environ 150.
III.3.2.3. Résultats
La Figure III.5 montre les trois coupes principales d’une image 3D obtenue par cette modalité
d’imagerie.
110

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
111
y
z
x
y 6.6 mm
x
z
A B C
Figure III.5: Trois coupes principales d’une image 3D obtenue par microtomographie X par rayonnement
synchrotron: longitudinale « X » (A), longitudinale « Y » (B) et transversale (C). Le champ de vue
reconstruit (6.63 mm3) ne permet pas de récupérer la totalité de l’échantillon. On peut noter que le
porte-échantillon (plexiglas) est visible.
Compte tenu du champ de vue, l’échantillon n’est pas entièrement imagé: environ la moitié de
sa hauteur est restituée et il est légèrement rogné dans les coupes transversales (cf. Figure
III.5, C). On peut estimer à environ 250 mm3 le volume de tissu trabéculaire disponible sur
chacune de ces images (soit environ la moitié du volume de l’échantillon, cf. III.2).
Du fait de la différence entre les coefficients d’atténuation de l’air (espace des pores) et du
plexiglas, on peut observer le porte-échantillon avec cette modalité d’imagerie: sur chaque
coupe transversale (cf. Figure III.5, C) et sur les coupes longitudinales « X » et « Y » qui sont
situées sur les bords de l’image (ce qui n’est pas le cas sur la Figure III.5, A et B). Cette
particularité n’a aucun effet en IRM puisque l’air, le tissu calcifié et le plexiglas ne produisent
aucun signal (et apparaissent donc comme du bruit sur les images).
L’histogramme calculé sur toute l’image fait apparaître 3 modes (cf. Figure III.6).

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
112
Occurrence normalisée
NG 00 50 100 150 200 250
0,05
0,1
0,15
Figure III.6: Histogramme calculé sur toute l’image. On peut observer trois modes: le premier
caractérisant l’espace des pores, le deuxième le porte-échantillon (plexiglas) et le dernier le tissu calcifié.
Les trois pics correspondent respectivement à l’espace des pores (faibles niveaux de gris i.e.
coefficient d’atténuation faible), au porte-échantillon (niveaux de gris intermédiaires) et au
tissu calcifié (hauts niveaux de gris i.e. coefficient d’atténuation élevé). Les trois modes sont
très nettement séparables. De plus l’histogramme devient bimodal si l’on prend une zone
totalement incluse dans le tissu trabéculaire. Cette distribution traduit le RSB très élevé et
l’adéquation de la taille du voxel par rapport aux dimensions caractéristiques des travées
osseuses. La segmentation de telles images ne pose aucun problème (cf. II.2.1) pour une
région d’intérêt située entièrement dans le tissu trabéculaire.
Dans la suite, le symbole utilisé pour référencer les images acquises par microtomographie X
par rayonnement synchrotron sera « SR-µCT ».
III.4. Recalage des régions d’intérêt
Afin de pouvoir effectuer une comparaison quantitative fiable entre les deux modalités, il faut
s’assurer de mener les calculs des différents paramètres sur les mêmes régions d’intérêt (ROI:
« Region Of Interest ») c’est-à-dire en considérant la même zone de tissu trabéculaire dans
une position identique pour les deux techniques d’imagerie. Pour les paramètres relatifs à
l’orientation de la structure (cf. II.6.4), on comprend facilement que cette étape est même
essentielle. Alors qu’une identification « manuelle » serait envisageable en dimension 2 (bien

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
que relativement fastidieuse), elle serait irréaliste en 3D compte tenu de la complexité de la
microarchitecture. Il faut donc envisager un procédé automatique et non ambigu de recalage
des ROI. Le principe de la méthode proposée consiste à « corriger » les différences observées
sur les images entre les deux modalités.
III.4.1. Le problème
Si on compare les images des coupes principales pour les deux modalités (cf. Figure III.2 et
Figure III.5), on peut constater quatre différences majeures:
• la position de l’échantillon: alors qu’il semble aligné avec l’axe de l’image en SR-µCT, il
apparaît légèrement incliné sur les images µIRM. Ceci est dû au fait que le bobinage du
gradient d’axe z (gradient de lecture) n’est pas exactement colinéaire avec l’axe du
porte-échantillon cylindrique (c’est-à-dire l’axe de la sonde radio-fréquence). Ceci
entraîne une inclinaison de l’image par rapport à cet axe.
• la taille de voxel: pour les deux modalités, on a des voxels isotropes. Cependant la taille du
voxel sur les images SR-µCT est environ six fois plus petite que celle obtenue sur les
images µIRM.
• le champ de vue: l’échantillon est disponible entièrement sur les acquisitions µIRM alors
qu’il n’est que partiellement reconstruit sur les images obtenues par SR-µCT.
• les niveaux de gris: de part la différence des phénomènes physiques à l’origine du
contraste dans l’image (absorption X pour la microtomographie, densité de proton pour
l’IRM), le tissu calcifié apparaît en hauts niveaux de gris et le reste (espace des pores et
fond de l’image) en faibles niveaux de gris sur les images SR-µCT alors que sur les
images µIRM, les faibles niveaux de gris sont à associer au tissu calcifié et au fond de
l’image et les hauts niveaux de gris à l’espace des pores.
Afin de réaliser l’identification des mêmes ROI entre les deux modalités, on a cherché à
« corriger » chacun de ces points. Le traitement s’effectuera en trois parties: repositionnement
des échantillons, ajustement de la résolution et recherche de la ROI commune.
Dans la suite, les images seront repérées par les trois axes (x,y,z) présentés sur la Figure III.2
et la Figure III.5.
113

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
III.4.2. Repositionnement des cylindres
Le but de cette première étape est d’aligner les deux axes des cylindres modélisant
l’échantillon dans les deux modalités d’imagerie (µIRM et SR-µCT).
III.4.2.1. Axes des cylindres µIRM et SR-µCT
La méthode employée pour déterminer les axes des cylindres est basée sur la récupération
pour les deux images de l’enveloppe cylindrique de l’échantillon par ajustement de l’équation
d’un cercle pour chaque coupe transversale. Ce choix est motivé en considérant que l’angle
d’inclinaison de l’échantillon dans l’image est faible (cf. Figure III.2 et Figure III.5). Dans ce
cas, la forme elliptique résultant de l’intersection du plan transversal (x,y) avec le cylindre
incliné peut être approchée par un cercle. Cette approximation sera justifiée dans la partie
III.4.2.3.3.
Sur chaque coupe (x,y) de l’image (pour chaque modalité), les pixels situés en bordure de
l’échantillon sont déterminés en balayant toutes les lignes verticales et horizontales dans un
sens puis dans l’autre. Pour chaque ligne analysée (c’est-à-dire pour un sens de balayage
donné), un pixel au maximum correspondant à une transition du fond de l’image vers
l’échantillon est recherché selon un critère propre à chaque modalité:
• pour les images µIRM, on considère une transition si le niveau de gris passe au-dessus du
seuil correspondant au milieu des deux pics de l’histogramme (cf. Figure III.3)
• pour les images SR-µCT, cette transition est définie entre le porte-échantillon en plexiglas
et l’espace des pores, c’est-à-dire quand le niveau de gris passe sous le seuil correspondant
au milieu des deux premiers pics de l’histogramme (cf. Figure III.6).
En utilisant cette méthode ainsi, deux cas peuvent générer des erreurs.
• Le tissu calcifié se trouve au bord de l’échantillon (cf. Figure III.7). Les pixels
« frontière » erronés sont alors filtrés selon un critère de connectivité (au sens de la
8-connexité, cf. II.4.4.2.1): les amas de pixels « frontière » de taille inférieure à 300 µm
(soit 4 pixels à la résolution des images obtenues par µIRM) sont supprimés. Cette taille
minimale a été déterminée par des tests: elle est de l’ordre de grandeur de l’épaisseur
d’une travée (ce qui revient à supposer que chaque « contact » du tissu calcifié avec le
bord de l’échantillon se fait au niveau d’une seule travée osseuse.
114

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
115
Travée osseuse
Deux amas de pixels « frontière »
Un amas de pixels « frontière » erronés
Figure III.7: Cas où le tissu calcifié se trouve en bordure de l’échantillon (agrandissement d’une image
µIRM). Les droites blanches sont les lignes balayées (dans le sens indiqué par les flèches) afin de
déterminer les pixels « frontière ». Les marques blanches symbolisent des pixels « frontière » trouvés, soit
sur cet exemple, deux amas de pixels « frontière » corrects (rond) mais aussi un amas erroné (carré) dû à
la présence de la travée osseuse. Ces pixels sont filtrés selon un critère de connectivité.
• Dans le cas des images SR-µCT, le cylindre est légèrement rogné dans les coupes
transversales (cf. III.3.2.3): certains pixels situés au bord de ces coupes se retrouvent
directement dans l’espace des pores et sont donc faussement détectés comme des pixels
« frontière ». Pour éviter ces erreurs, on s’assure que les pixels « frontière » successifs
n’ont pas une de leur coordonnées identiques (cf. Figure III.8). Cette correction permet
aussi d’éviter de prendre comme pixel « frontière », les pixels situés aux quatre coins de
chaque coupe transversale. En effet, ceux-ci appartiennent à la même phase que l’espace
des pores car ils sont situés à l’extérieur du porte échantillon (cf. Figure III.5, C).

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
116
Bord de la coupe transversale
Echantillon (espace des pores)
Porte-échantillon (plexiglas)
Pixels frontières ayant même abscisse (cylindre rogné)
Figure III.8: Bord d’une coupe transversale pour une image SR-µCT (agrandissement). Les droites
blanches sont les lignes balayées (dans le sens indiqué par les flèches) afin de déterminer les pixels
« frontière ». Les marques blanches symbolisent des pixels « frontière » trouvés. Du fait que l’échantillon
est incomplet avec cette modalité, certains pixels du bord de la coupe se retrouvent artificiellement de type
« frontière ». Ces pixels successifs (représentés par des carrés) ont une de leurs deux coordonnées égales
(l’abscisse dans cet exemple). Ils sont donc supprimés .
A partir de ces pixels « frontière », la méthode des moindres carrés ([Press, 1989]) est
appliquée afin d’ajuster l’équation d’un cercle pour chaque coupe transversale. Ainsi, pour
chaque modalité, on dispose du rayon Rk et des deux coordonnées du centre (ak,bk) de chaque
cercle associé à chaque coupe transversale k. La Figure III.9 donne une représentation
tridimensionnelle de la forme cylindrique ajustée pour l’image µIRM d’un échantillon.
Figure III.9: Représentation tridimensionnelle de la forme cylindrique d’un échantillon (modalité µIRM).
A part pour quelques coupes problématiques, l’enveloppe de l’échantillon est correctement restituée. On
peut remarquer la légère inclinaison du cylindre (cf. Figure III.2).
L’enveloppe cylindrique est correctement restituée mis à part pour quelques coupes. Nous
reviendrons dans la partie III.4.2.3.1 sur la fiabilité de cette méthode. L’axe (∆) du cylindre

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
117
est calculé, pour chaque modalité, en ajustant (par les moindres carrés) l’équation d’une droite
sur les centres de chaque cercle (ak,bk).
III.4.2.2. Alignement des cylindres µIRM et SR-µCT
Comme cela est représenté sur la Figure III.10, la position angulaire du cylindre peut être
définie à partir de la direction de son axe (∆) qui est inclinée de θ par rapport à x et de ϕ par
rapport à z.
θ
ϕ z
x
y
∆
Figure III.10: Représentation d’un cylindre incliné dans le repère (x,y,z) de l’image. Les deux angles θ et ϕ
permettent de définir la direction de l’axe du cylindre.
A partir de l’expression numérique de (∆) pour chaque modalité que nous avons déterminée
en III.4.2.1, le couple (θ,ϕ) associé à chaque cylindre peut être facilement calculé.
Comme les cylindres SR-µCT semblent alignés avec l’axe z de l’image (ceci sera discuté en
III.4.2.3.3), on choisit d’effectuer une rotation tridimensionnelle (interpolation trilinéaire)
d’angle (-θ,-ϕ) du cylindre µIRM afin de l’aligner avec l’axe z c’est-à-dire l’axe du cylindre
SR-µCT.
Remarque:
Le but de cette partie étant d’aligner les axes des deux cylindres, seul l’angle ϕ est important
pour cette étape (d’autant que, comme les cylindres apparaissent peu inclinés, les valeurs de θ
ne sont pas significatives).
III.4.2.3. Bilan du repositionnement
III.4.2.3.1. Enveloppe cylindrique
Nous avons tout d’abord cherché à établir la robustesse du calcul de l’enveloppe. Le Tableau
III.1 présente la valeur moyenne et l’écart type de Rk, calculés, pour un échantillon, sur

Caractérisation de l’architecture tridimension ire par IRM nelle du tissu trabécula
118
l’image µIRM avant et après repositionnement (156 coupes transversales) et sur l’image
SR-µCT (660 coupes transversales).
µ ± σ µIRM avant µIRM après SR-µCT
Rk (en pixels) 54.1 ± 0.5 53.8 ± 0.7 348.4 ± 3.5
Tableau III.1: Valeur moyenne et écart type (µ ± σ) de Rk pour un échantillon, calculés respectivement sur
les 156 et 660 coupes transversales représentant l’échantillon sur les images µIRM (avant et après
repositionnement) et SR-µCT.
Dans les trois cas, l’écart type de Rk est très faible par rapport à la valeur moyenne.
L’évaluation des rayons d’une coupe à l’autre est donc très stable. Cette constatation permet
de valider la méthode utilisée pour déterminer l’axe du cylindre (cf. III.4.2.1).
III.4.2.3.2. Plage de linéarité du repositionnement
La plage de linéarité du calcul de ϕ a été déterminée comme suit. Nous avons généré une
image test en prélevant arbitrairement, sur une image µIRM, un cylindre colinéaire à l’axe z,
entièrement inclus dans le tissu trabéculaire. Les caractéristiques de ce cylindre (rayon de 50
voxels, hauteur de 150 voxels) ont été choisies pour simuler un échantillon « théorique » (de
dimensions semblables aux échantillons étudiés) dont l’image acquise par µIRM (dans les
mêmes conditions) le présenterait parfaitement colinéaire avec l’axe z. Les trois coupes
principales de ce « cylindre test » sont données en Figure III.11.
A B C Figure III.11: Trois coupes principales (A: longitudinale « X », B: longitudinale « Y » et C: transversale)
d’un échantillon cylindrique « théorique », imagé par µIRM 3D, qui apparaîtrait parfaitement colinéaire
avec l’axe z dans l’image. La hauteur et le rayon du c lindre sont respectivement de 150 et de 50 voxels. y

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
119
Sur ce cylindre, préalablement tourné pour différentes valeurs ϕthéorique de l’angle ϕ, on
effectue le calcul de l’inclinaison de son axe avec la méthode présentée précédemment (cf.
III.4.2.1 et III.4.2.2): ϕmesuré. La Figure III.12 montre ϕmesuré en fonction de ϕthéorique (pour
ϕthéorique ∈ [0.1,30]°).
0,1
1
10
100
0,1 1 10 100
ϕmesuré (°)
ϕthéorique (°)
Plage de linéarité
Figure III.12: ϕmesuré en fonction ϕthéorique pour ϕthéorique ∈ [0.1,30]° (échelle logarithmique). A partir de
cette courbe on peut déterminer une plage de linéarité par comparaison avec la première bissectrice (en
pointillé sur le graphe).
En comparant cette courbe à la première bissectrice, on peut déterminer une zone de linéarité
pour l’alignement du cylindre µIRM: 1°< ϕ <10°. La limite inférieure est due au fait que la
rotation n’est plus significative si l’angle est trop petit. La limite supérieure est liée à
l’approximation de l’ellipse par un cercle (cf. III.4.2.1) qui n’est plus valide pour des
inclinaisons trop fortes.
III.4.2.3.3. Efficacité du repositionnement
Le Tableau III.2 donne la valeur moyenne et l’écart type de l’angle ϕ, calculés sur les 29
échantillons, pour les images µIRM avant et après repositionnement et pour les images
SR-µCT.
µ ± σ µIRM avant µIRM après SR-µCT
ϕ (en °) 2.5 ± 0.3 0.3 ± 0.2 0.5 ± 0.3
Tableau III.2: Valeur moyenne et écart type (µ ± σ) de l’angle ϕ calculés pour les 29 échantillons sur les
images µIRM (avant et après repositionnement) et SR-µCT.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Compte tenu de la gamme de valeurs mesurées pour ϕ sur les 29 échantillons et de la plage de
linéarité de la méthode (cf. III.4.2.3.2), on peut donc conclure que:
• l’approximation de l’enveloppe de l’échantillon par la forme cylindrique peut être utilisée
• les cylindres SR-µCT peuvent être considérés comme alignés avec l’axe z puisque
l’inclinaison mesurée à haute résolution n’est pas significative. L’hypothèse formulée en
III.4.2.2 est donc validée.
• l’alignement des cylindres µIRM est correct puisque la mesure de l’angle ϕ après
repositionnement n’est pas significative.
III.4.2.3.4. Remarque
La méthode présentée pour déterminer l’enveloppe cylindrique est suffisamment précise pour
estimer la direction de l’axe de l’échantillon. Cependant elle serait inadéquate pour définir
rigoureusement la ROI sur laquelle on effectuerait le calcul des paramètres (cf. III.6.1). En
effet, bien que faibles, les fluctuations observées pour Rk (cf. Figure III.9) entraîneraient des
erreurs sur la zone sélectionnée et donc des biais sur les évaluations (fausses déconnections de
travées osseuses,…).
III.4.3. Ajustement de la résolution
III.4.3.1. Problématique
L’objectif de cette étude est d’établir, à partir de l’évaluation de différents paramètres de
microarchitecture, la validité de l’IRM pour caractériser le tissu trabéculaire. De nombreux
travaux ont montré que ces paramètres étaient dépendants de la résolution.
Par exemple, Kothari et al. ([Kothari, 1998]) ont analysé 10 échantillons de tissu trabéculaire
de fémur et de vertèbre à différentes résolutions, à partir d’acquisitions à 40 µm (obtenues par
imagerie optique) qu’ils ont ensuite dégradées pour des épaisseurs de coupe variant de 100 à
1000 µm et une résolution dans la coupe allant jusqu’à 100 µm. Ils ont ainsi pu montrer que
certains paramètres étaient peu dépendants de la résolution (Tb.Sp et Tb.N, c’est-à-dire
BS/TV) mais que d’autres l’étaient fortement (BV/TV, Tb.Th et la famille de paramètres
d’anisotropie et d’orientation). A partir d’acquisitions par microtomographie X (rayonnement
synchrotron) de 10 échantillons de vertèbres à 14, 7 et 2 µm de résolution, Peyrin et al.
120

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
([Peyrin, 1998]) ont obtenu des résultats similaires qui se sont avérés conformes aux
simulations.
L’influence de la résolution est hors du cadre de notre étude. Pour comparer les deux
modalités, on se ramène à la même taille de voxel afin de décorréler au mieux les différences
dues aux techniques d’imagerie de celles dues aux conditions d’acquisition et aux traitements
effectués sur les images. Le but de cette partie est donc de dégrader la résolution des images
SR-µCT afin de s’approcher le plus précisément possible de la taille du voxel obtenue en
µIRM.
III.4.3.2. Dégradation des images SR-µCT
On définit le rayon du cylindre comme la valeur moyenne des rayons obtenus sur chaque
coupe par la méthode présentée en III.4.2.1. On note dans la suite RµIRM, le rayon du cylindre
µIRM calculé après son repositionnement et RSR-µCT celui obtenu pour le cylindre SR-µCT.
La valeur moyenne et l’écart type (µ ± σ) de RµIRM et RSR-µCT, calculés sur les 29 échantillons,
étaient respectivement de 53.2 ± 1.1 voxels et 350.4 ± 4.0 voxels. Les écart types étant très
faibles, on définit le coefficient de dégradation η comme le rapport entre les valeurs
moyennes des rayons de chaque cylindre, calculées sur les 29 échantillons:
><><
=−
µIRM
µCTSR
RR
η .
La résolution de l’image SR-µCT est isotrope, de dimensions N3 (avec N = 660). Pour ajuster
au mieux la taille du voxel SR-µCT à celle du voxel µIRM, la dégradation isotrope de l’image
SR-µCT doit conduire à une image SR-µCTD de dimensions ND3, avec:
)(ηNArrND= ,
où « Arr » est la fonction « arrondie au plus près ». Dans notre cas, l’application numérique
donne η = 6.6 donc ND = 100. Ceci est donc conforme aux résolutions annoncées pour les
deux modalités.
III.4.3.3. Méthode
Les méthodes classiques de dégradation de la résolution (comme le moyennage de voxels) ne
permettent d’obtenir que des rapports η entiers et ne pourraient être appliquées ici. Pour
chaque image SR-µCT, on a donc calculé l’image dégradée associée, SR-µCTD, en trois
étapes illustrées sur la Figure III.13.
121

Caractérisation de l’architecture tridim r IRM ensionnelle du tissu trabéculaire pa
122
• Transformée de Fourier discrète de l’image initiale SR-µCT
• Rognage de cette transformée de Fourier de manière à ne garder que ND voxels du centre
du plan de Fourier dans chaque direction
• Transformée de Fourier discrète inverse du volume précédent
Compte tenu des tailles (N=660, ND=100), les transformées de Fourier Discrète n’ont pu être
évaluées avec un algorithme de calcul rapide (FFT) car ce dernier suppose une image ayant un
nombre de voxels dans chaque direction égale à une puissance de 2. L’algorithme que nous
avons utilisé pour les calculer est celui proposé par Frigo et Johnson du MIT ([Frigo, 1998]):
FFTW (« Fastest Fourier Transform of the West »), disponible en « Freeware » sur leur site
web.
A B C Figure III.13: Les trois étapes de la dégradation. En A, on donne une coupe de l’image SR-µCT originale
(résolution de 10 µm). En B, le module de la transformée de Fourier de A est représenté. On ne conserve
que les ND voxels du centre du plan de Fourier dans chaque direction (carré blanc). La transformée de
Fourier inverse de cette zone est donnée en C: elle correspond à une image dégradée de ND×ND×ND voxels:
SR-µCTD. Tous ces calculs sont menés en dimension 3.
La précision de la dégradation est donc de un voxel dans chaque direction. A titre de
vérification, nous avons déterminé la valeur moyenne et l’écart type (µ ± σ), sur les 29
échantillons, du rayon du cylindre calculé sur les images SR-µCTD: 53.4 ± 1.2 voxels (à
comparer aux 53.2 ± 1.1 voxels trouvés pour les 29 cylindres µIRM repositionnés). Ces
résultats (dont une illustration est donnée en Figure III.14) permettent de valider notre
méthode d’ajustement des tailles de voxels entre les deux modalités.

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
III.4.4. Recherche de la ROI commune
III.4.4.1. Problématique
Le but de cette dernière étape est de trouver la ROI commune entre les cylindres µIRM
repositionné et SR-µCT dégradé, c’est-à-dire entre deux cylindres coaxiaux de même
résolution. Ce problème revient à trouver les positions angulaire θ0 et verticale k0 (car la
hauteur de l’échantillon n’est pas entièrement restituée par la modalité SR-µCT, cf. III.3.2.3)
d’un cylindre par rapport à l’autre. En d’autres termes, si on considère que z est l’axe
commun aux deux cylindres, il faut déterminer:
• la rotation de θ0 du cylindre µIRM autour de z
• la translation de k0 du cylindre µIRM le long de z
pour l’amener dans la même zone et la même position que celui SR-µCTD (nous avons en
effet choisi pour la suite d’effectuer les transformations géométriques sur le volume µIRM
afin de ne pas modifier les images de référence, ce qui pourra faciliter par exemple une future
étude sur les effets de la résolution).
III.4.4.2. ROI utilisée pour le calcul
Pour la recherche de la ROI commune, nous nous sommes basés, pour chaque modalité, sur
une ROI limitée pour chaque coupe, au plus grand cercle inclus dans l’échantillon imagé par
SR-µCT. De cette manière, on s’affranchit du problème lié au fait que l’échantillon apparaît
légèrement « rogné » en SR-µCT (cf. III.3.2.3). Sur la Figure III.14 est donnée une illustration
de la ROI sur laquelle on travaille.
123

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
124
A B
Figure III.14: Illustration de la ROI sur les coupes transversales d’un échantillon pour les images µIRM
après repositionnement (A) et SR-µCT après dégradation (B). Les cercles blancs correspondent à
l’ajustement de l’enveloppe cylindrique pour les deux modalités (par la méthode présentée en III.4.2.1):
les rayons de ces deux cercles sont identiques (ce qui traduit l’égalité des résolutions entre les deux
images). Les cercles gris sont les ROI sur lesquelles est effectuée la recherche de la ROI commune. Ils sont
définis par le même rayon (en voxels): celui du plus grand cercle inclus dans l’échantillon pour l’image
SR-µCTD. Par contre, les hauteurs des cylindres sont propres à chaque modalité du fait de la différence de
champ de vue (cf. III.3.1.4 et III.3.2.3).
Ces deux cylindres (un pour chaque modalité) sont donc coaxiaux et de rayon identique du
fait de l’ajustement des résolutions (cf. III.4.3). Par contre, leur hauteur diffère du fait de la
différence de champ de vue entre les deux modalités (cf. III.3.1.4 et III.3.2.3): NµIRM, le
nombre de coupes transversales disponibles en µIRM, est en moyenne égale à 200 (en
fonction de la taille de l’échantillon, cf. III.2) alors que les images SR-µCTD ne sont
composées que de 100 coupes transversales (cf. III.4.3.2).
III.4.4.3. Première approche: méthode du volume « OU exclusif »
Cette approche consiste à analyser systématiquement toutes les positions possibles du cylindre
µIRM par rapport à celui SR-µCTD en utilisant comme critère de comparaison, le calcul du
volume « OU exclusif » entre les deux modalités.
III.4.4.3.1. Description de la méthode
Pour balayer toutes les positions angulaires possibles, une rotation d’angle θ du cylindre
µIRM autour de son axe (en utilisant l’interpolation trilinéaire) a tout d’abord été calculée,
tous les degrés, pour θ compris entre 0 et 360°. Après segmentation par seuillage simple (cf.
II.2.1), on a pu évaluer un volume « OU exclusif » sur la zone commune entre les deux
modalités pour toutes les translations verticales k du cylindre µIRM laissant le cylindre
SR-µCTD entièrement inclus dans le cylindre µIRM. Le nombre N de translations possibles

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
125
du cylindre µIRM est donné par N = 2×(NµIRM - 100 + 1). En effet, pour un sens donné de
l’échantillon (selon l’axe z), il y a NµIRM - 100 + 1 translations possibles. Or, il faut aussi
envisager le cas où l’échantillon aurait été retourné entre les deux protocoles d’imagerie, d’où
le facteur 2.
Chaque volume « OU exclusif » présente un niveau « 1 logique » pour les voxels où il y a une
différence entre les deux ROI, un niveau « 0 logique » sinon. Quelque soit le couple (k,θ), ces
volumes sont de même taille (celle de la ROI utilisée sur les images SR-µCTD, cf. III.4.4.2):
la somme des voxels au niveau « 1 logique » donne donc l’estimation d’une erreur entre les
deux ROI propre à chaque position étudiée (k,θ): E(k,θ).
Pour chaque échantillon, une image « erreur » E(k,θ) (de dimension N×360) a donc été
calculée. Le recalage (k0,θ0) permettant d’aboutir à une ROI commune aux deux modalités,
est donnée par le couple (k,θ) correspondant au minimum de E(k,θ) (cf. Figure III.15).
θ
k
360 pixels
N pixels
θ0
k0 Figure III.15: Représentation d’une image « erreur » E(k,θ) déterminée à partir du volume « OU
exclusif » calculé entre les deux cylindres (µIRM repositionné et SR-µCTD) pour les N translations
verticales k possibles (N = 2×(NµIRM - 100 + 1), si NµIRM est le nombre de coupes du cylindre µIRM: voir
texte) et pour les 360 rotations d’angle θ (0 ≤ θ < 360°, par pas de 1°) du cylindre µIRM repositionné. Le
couple (k0,θ0) correspondant à l’erreur minimale permet de déterminer la ROI commune aux deux
modalités.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Remarque:
L’étape de segmentation nécessaire au calcul des volumes « OU exclusif » n’est pas critique:
le but ici n’est pas d’avoir la « meilleure » représentation possible du réseau trabéculaire mais
plutôt une approximation permettant la comparaison entre les deux modalités.
III.4.4.3.2. Limitation de cette méthode
Cette technique permet d’obtenir un recalage automatique des ROI. Cependant, le temps de
calcul est rédhibitoire. En effet, pour chaque position angulaire, le calcul du volume « OU
exclusif » doit être fait pour toutes les translations ce qui suppose une analyse de tout le
volume pour chaque couple (k,θ). Ainsi, sur une station SUN 400 MHz, le temps de calcul est
d’environ 5 heures par échantillon.
III.4.4.4. Méthode de la longueur moyenne d’interception
Le principe de cette méthode est basé sur l’analyse de la longueur moyenne d’interception en
dimension 2, MIL2D, évaluée sur chaque coupe transversale k et pour toutes les orientations
possibles θ de chaque cylindre (un par modalité, cf. III.4.4.2). Le calcul de ce paramètre se
déduit très simplement de celui présenté pour un espace de dimension 3 (cf. II.6.1).
MIL2D présente deux avantages:
• ce paramètre reflète l’organisation générale de la structure trabéculaire: comme on l’a vu
en II.6.1, Whitehouse ([Whitehouse, 1974]) a montré que l’orientation et l’anisotropie en
dimension 2 peuvent être estimées à partir de MIL2D
• ce paramètre est relativement peu sensible au contraste: l’étape de segmentation n’est donc
pas critique et peut donc être effectuée par seuillage simple (cf. II.2.1) d’autant plus qu’on
ne cherche pas ici à avoir la « meilleure » estimation possible du paramètre mais une
approximation afin de comparer les deux modalités
III.4.4.4.1. Image paramétrique « MIL2D »
Une image paramétrique « MIL2D » est calculée pour chaque modalité de la manière suivante.
Après rotation du cylindre autour de l’axe z d’un angle θ (en utilisant l’interpolation
trilinéaire), on effectue une segmentation du volume par un seuillage simple (cf. II.2.1).
Ensuite, sur chaque coupe k, on calcule, d’après l’algorithme présenté en II.5.1.3.1 (adapté au
cas 2D), le nombre de transitions N1(k,θ) dans la direction de l’axe x, ainsi que la longueur
des droites du faisceau L, sur la ROI présentée en III.4.4.2. Celle-ci étant circulaire sur chaque
126

Chap du tissu trabéculaire itre III: Validité du rendu par IRM
127
coupe k, on s’assure ainsi de ne pas favoriser artificiellement certaines directions (cf.
remarque b) de II.5.1.2.1).
L’image paramétrique associée à chaque modalité est calculée ainsi:
),(),(
12 θ
θkN
LkMIL D = ,
pour θ compris entre 0 et 180°. Pratiquement, on balaie θ avec un pas angulaire de 1°. La
Figure III.16 présente les deux images paramétriques d’un échantillon.
θ
k
100 pixels
180 pixels
θ
k
NµIRM pixels
180 pixels
A B
Figure III.16: Exemple de deux images paramétriques (une par modalité) de MIL2D(k,θ) calculées pour un
échantillon sur la ROI présentée en III.4.4.2 (A: µIRM repositionnée, B: SR-µCT dégradée). Pour ces
deux images, la dimension de l’axe vertical est identique: 180 pixels qui correspondent à l’intervalle
angulaire de 0 à 180° balayé avec un pas de 1°. La dimension horizontale est différente: elle correspond au
nombre de coupes transversales de l’échantillon disponibles dans chaque modalité (cf. III.4.4.2).
III.4.4.4.2. Détermination de la position de la ROI commune
Pour déterminer le couple (k0,θ0) correspondant au recalage optimal, on recherche le
maximum de la fonction d’intercorrélation entre les deux images paramétriques caractérisant
les deux modalités (cf. Figure III.16). Pour ce faire, on utilise la périodicité de MIL2D:
MIL2D(k,θ) = MIL2D(k,θ + π). De plus, on balaie les deux sens possibles pour les coupes
(miroir d’une des deux images paramétriques) dans le cas où l’échantillon aurait été retourné
entre les deux protocoles d’imagerie.
Du fait de la périodicité de MIL2D, l’angle trouvé par cette méthode, θS, est donné à π près.
Deux couples peuvent donc être solution: (k0,θS) ou (k0,θS + π). Pour lever cette ambiguïté, on
utilise la méthode du volume « OU exclusif » (cf. III.4.4.3): la position angulaire θ0 retenue

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
est celle conduisant à la valeur minimale entre E(k0,θS) et E(k0,θS + π). Cette distinction est
d’ailleurs relativement simple à faire visuellement: une erreur de 180° est en effet très
« frappante » sur les images.
III.4.4.4.3. Avantage de cette méthode
En comparant les deux modalités à l’aide des images paramétriques, le problème du recalage
initialement tridimensionnel devient bidimensionnel: il est donc beaucoup plus simple à
résoudre ce qui permet un gain de temps non négligeable par rapport à la méthode « OU
exclusif » (cf. III.4.4.3).
Ainsi, sur la même station de travail, le temps de calcul n’est que de 15 minutes par
échantillon soit un gain de plusieurs heures.
Nous avons donc retenu cette méthode pour effectuer la recherche de la ROI commune.
III.4.5. Recalage des ROI: effet sur les paramètres de structure
Afin de valider la méthode de recalage des images, il faut vérifier si elle introduit des erreurs
systématiques sur l’évaluation des paramètres d’architecture trabéculaire. En ce qui concerne
la dégradation de résolution des images SR-µCT, nous avons déjà discuté comment le choix
du mode de connexité minimise les variations de topologie selon la résolution (cf. II.4.6.2).
De plus, le critère retenu pour la segmentation de ces images assure la conservation de la
fraction osseuse (cf. III.5.2.2). L’autre opération géométrique irréversible est la rotation 3D
des images µIRM (du fait de l’interpolation). Nous avons suivi le protocole suivant pour
quantifier les effets de cette rotation sur les paramètres d’architecture.
III.4.5.1. Protocole
Un cylindre µIRM préalablement repositionné a été pris comme référence. Différentes
rotations 3D d’angle (θrot,ϕrot) ont été appliquées sur ce cylindre afin de couvrir toute la
gamme des rotations effectuées pour le recalage des 29 échantillons:
• 10 rotations d’angle θrot (ϕrot restant nul) compris entre 0 et 90°: les valeurs de l’angle de
recalage θ0 trouvées étant réparties dans tout l’intervalle [0,360[°, il suffit d’étudier les
effets de l’interpolation sur le premier quadrant [0,90]°, les autres pouvant se déduire de
celui-ci. En effet, les rotations de 90° ou de 180° d’une image discrète sont réversibles car
elles correspondent respectivement à une transposition ou à un miroir de l’image initiale
128

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
• 7 rotations d’angle ϕrot (θrot restant nul) compris entre [0,3.5]° qui correspond à toute la
gamme d’angles ϕ rencontrés dans cette étude (pour l’alignement du cylindre µIRM, cf.
III.4.2)
Après chaque rotation, les images ont été segmentées en utilisant une méthode dédiée à la
microscopie IRM que nous présenterons ultérieurement (cf. III.5.1). Ensuite les paramètres de
microarchitecture décrits au chapitre II ont été calculés.
III.4.5.2. Résultats
Le Tableau III.3 rapporte les valeurs moyennes µ et écarts type σ des erreurs sur les
paramètres de structure relativement à leur évaluation sur le cylindre de référence. µ et σ ont
été calculés sur toute la gamme des angles étudiés. En ce qui concerne les paramètres θ et ϕ
(les deux premiers angles d’Euler), l’erreur a été calculée relativement à la valeur mesurée sur
le cylindre de référence incrémentée de la valeur de l’angle de rotation (respectivement θrot et
ϕrot).
Toutes les erreurs indiquées dans le Tableau III.3 sont donc uniquement dues aux effets de la
rotation 3D discrète.
129

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Erreur relative
(µ ± σ, en %)
Rotations θrot
(ϕrot = 0°)
Rotations ϕrot
(θrot = 0°)
BV/TV 2.4 ± 0.5 2.4 ± 0.5
CN3 4.7 ± 2.2 2.1 ± 0.8
BS/TV 1.1 ± 0.8 2.1 ± 0.4
Tb.Th 1.0 ± 0.5 0.4 ± 0.2
Tb.Sp 1.4 ± 0.2 2.5 ± 0.5
θ(*) 4.4 ± 3.3 1.2 ± 0.7
ϕ(*) 2.9 ± 1.4 3.5 ± 2.5
ψ 1.9 ± 1.4 2.8 ± 0.7
MIL1 1.1 ± 0.8 1.9 ± 0.4
MIL2 0.4 ± 0.3 2.7 ± 0.5
MIL3 3.1 ± 2.0 1.5 ± 0.2
RMIN 2.6 ± 1.9 0.3 ± 0.2
RMAX 0.9 ± 0.7 0.8 ± 0.2
Tableau III.3: Valeurs moyennes et écarts type (µ ± σ) des erreurs sur l’évaluation des paramètres de
structure relativement aux mesures sur le cylindre de référence (c’est-à-dire le cylindre sur lequel aucune
rotation n’a été appliquée) pour les rotations d’angle θrot (10 valeurs comprises entre 0 et 90°, ϕrot restant
nul) et ϕrot (7 valeurs comprises entre 0 et 3.5°, θrot restant nul). Les symboles des paramètres sont
présentés dans le chapitre II. (*)Pour θ et ϕ, les deux premiers angles d’Euler, l’erreur a été calculée
relativement à la valeur mesurée sur le cylindre de référence incrémentée de la valeur de l’angle de
rotation (respectivement θrot et ϕrot).
L’erreur la plus importante, 8.5%, a été trouvée sur θ (le premier angle d’Euler) pour
θrot = 30° (ϕrot restant nul). Pour la fraction osseuse par exemple, les effets de la rotation 3D
étaient particulièrement négligeables (erreur toujours inférieure à 3% quelle que soit la
rotation). Pour les rotations d’angle ϕrot (θrot restant nul), les erreurs étaient systématiquement
inférieures à 3.8% pour tous les paramètres excepté pour ϕ, le deuxième angle d’Euler (erreur
de 7.2% pour ϕrot = 3.5°, θrot restant nul).
130

Chapitre III: Validité du rendu laire par IRM du tissu trabécu
131
On peut donc conclure que le recalage automatique des ROI a très peu d’influence sur les
paramètres de microstructure calculés.
III.4.6. Bilan de la méthode de recalage automatique
III.4.6.1. Efficacité
Avec cette méthode, 27 échantillons (sur les 29) ont pu être recalées automatiquement au
voxel près pour la dégradation et pour la translation, et au degré près pour la position
angulaire. Nous avons d’ailleurs vérifié, pour quelques échantillons, qu’on retrouvait bien les
mêmes couples (k0,θ0) déterminés par la méthode du volume « OU exclusif » (cf. III.4.4.3).
Pour chaque échantillon, on obtient donc un volume cubique par modalité d’imagerie de
6.6 mm de coté (soit 100 voxels à la résolution µIRM) ce qui correspond à un volume
commun d’environ 250 mm3: la ROI commune est en effet imposée par le champ de vue
réduit de l’acquisition SR-µCT (cf. III.3.2.3).
La Figure III.17 présente le résultat obtenu pour un échantillon.
A B C
D E F
Figure III.17: Recalage obtenu pour un échantillon. Chaque figure représente une coupe du champ de vue
cubique: elles sont donc toutes carrées avec un coté de 6.6 mm. Les figures A, B et C sont les images de la
même coupe transversale et les figures D, E et F de la même coupe longitudinale « Y ». Les figures A et D,
B et E, et C et F correspondent respectivement aux images µIRM (100 voxels de côté), SR-µCTD (100
voxels de coté) et SR-µCT (660 voxels de coté).

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
132
MIL2D (en mm)
Du fait que les images SR-µCT dégradées n’ont pas subi de transformations géométriques, on
peut très facilement repérer les coupes identiques d’une résolution à une autre. Visuellement,
la ressemblance est frappante entre les deux modalités d’imagerie.
La Figure III.18 montre, pour un échantillon, les courbes représentant MIL2D calculée pour
chaque modalité sur la ROI commune: en fonction de θ pour une coupe k donnée et en
fonction de la coupe k pour un angle θ donné. On remarque bien que ce paramètre est très
discriminant en terme de position (pour l’angle et la coupe). De plus, ces courbes ont une
forme très similaire pour les deux modalités (même si les valeurs ne sont pas identiques). Ces
deux aspects rendent cette méthode de recalage très fiable et robuste.
012345678
0 50 100
MIL2D (en mm)
kθ (en °)0
1
2
3
4
5
6
7
8
0 50 100 150 A B
Figure III.18: Représentations de MIL2D calculée sur la ROI commune pour un échantillon: en fonction de
θ pour une coupe k donnée (A), en fonction de k pour un angle θ donné (B). Sur ces courbes, les traits noir
et gris correspondent respectivement aux images µIRM et SR-µCTD.
Pour les deux échantillons problématiques, le recalage automatique a pu être obtenu par la
méthode du volume « OU exclusif » (cf. III.4.4.3). Dans la partie suivante, on revient sur ces
deux échantillons afin de comprendre les limites de la méthode de recalage utilisant MIL2D.
III.4.6.2. Etude des deux échantillons problématiques
Seuls deux échantillons (notés dans la suite E1 et E2) ont posé problème. Pour E1, on a observé
une erreur de 90° pour θ0 et de 14 coupes pour k0, et pour E2, une erreur sur k0 de 23 coupes.

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
133
MIL2D (en mm)
Sur la Figure III.19, nous avons représenté deux profils « caractéristiques » de MIL2D (le
premier en fonction de θ pour k fixé, le deuxième en fonction de k pour θ fixé) prélevés sur
les images paramétriques « SR-µCT » (dont un exemple est donné en Figure III.16 B)
associées aux échantillons E1 et E2.
θ (en °)0
1
2
3
4
5
6
7
8
0 50 100 150
MIL2D (en mm)
k0
1
2
3
4
5
6
7
8
0 50 100 A1 B1
MIL2D (en mm)
θ (en °)0
1
2
3
4
5
6
7
8
0 50 100 150
MIL2D (en mm)
k0
1
2
3
4
5
6
7
8
0 50 100 A2 B2
Figure III.19: Profils prélevés sur les images paramétriques « SR-µCT » (dont un exemple est donné en
Figure III.16 B) des deux échantillons problématiques. A1 et B1: E1 (erreur de 90° pour θ0 et de 14 coupes
pour k0). A2 et B2: E2 (erreur sur k0 de 23 coupes). Les figures A1 et A2 correspondent à MIL2D en
fonction de θ (pour une coupe k donnée), les figures B1 et B2 à MIL2D en fonction de k (pour un angle θ
donné).
Pour l’échantillon E1, on peut remarquer que sur la courbe MIL2D en fonction de θ (cf. Figure
III.19 A1), l’écart entre la valeur minimale et maximale est très faible (à comparer avec celui

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
obtenu sur un échantillon pour lequel le recalage a fonctionné, cf. Figure III.18 A). Ceci
illustre la faible anisotropie de la structure trabéculaire de cet échantillon dans les coupes
transversales puisque MIL2D est quasiment indépendant de la direction analysée. La position
angulaire est donc difficile à déterminer. Il est intéressant de noter que l’erreur sur θ0 est de
90° ce qui semble confirmer l’isotropie de la structure: les valeurs maximale et minimale de
MIL2D en fonction de θ ne sont pas significativement différentes.
On peut aussi constater que la courbe MIL2D en fonction de k (cf. Figure III.19 B1) ne fluctue
pas de manière significative (à comparer avec celle de la Figure III.18 B). Le pouvoir
discriminant de MIL2D selon l’axe z est donc réduit ce qui provoque une erreur sur k0.
Pour l’échantillon E2, on retrouve un écart significatif entre les valeur minimale et maximale
de MIL2D en fonction de θ (cf. Figure III.19 A2). Il existe donc des directions privilégiées de
la structure trabéculaire dans les coupes transversales ce qui permet de trouver correctement
θ0. Par contre, comme pour l’échantillon E1, MIL2D ne semble pas discriminant en fonction de
k (cf. Figure III.19 B2) d’où l’erreur sur k0.
Ces deux échantillons illustrent les limites du recalage par la méthode de la longueur
moyenne d’interception. Si la structure ne présente pas d’orientation privilégiée (anisotropie)
dans les coupes transversales, la détermination de θ0 sera problématique. Si, de plus cette
orientation ne change pas « significativement » d’une coupe transversale à l’autre, k0 pourra
être erroné. Dans ces cas de figure, la méthode du volume « OU exclusif » (cf. III.4.4.3) a été
utilisée.
III.4.7. Conclusions et perspectives
La méthode de recalage que nous avons développée est automatique, non ambiguë (il n’y a
pas besoin d’expert), précise et rapide. De plus, dans les conditions d’imagerie de cette étude,
elle n’affecte pas la caractérisation de la microstructure.
Le développement de telles méthodes est essentiel pour de nombreux types d’étude du tissu
trabéculaire. On peut citer par exemple:
• la confrontation de différentes modalités (comme c’est le cas de ce travail). Le recalage est
alors obligatoire en particulier pour les comparaisons de paramètres liés à l’orientation de
la structure (cf. III.6.3.2): il semble en effet très difficile de pouvoir assurer, sur des
protocoles d’imagerie différents, une position identique des échantillons.
134

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
• les applications in vivo comme le suivi de patients dans le temps pour observer l’évolution
de la pathologie (l’ostéoporose étant une pathologie évolutive, cf. I.3), les effets des
traitements,… Compte tenu des contraintes liées à l’imageur ou au patient, il peut s’avérer
difficile d’observer exactement la même ROI sur des images acquises à des instants
différents (position du patient variable à chaque nouvel examen, utilisation d’imageurs
différents ou même de modalités d’imagerie différentes,…). Peu d’améliorations de la
méthode présentée seront nécessaires pour pouvoir l’utiliser dans ce type d’application à
condition que la ROI puisse être détectée précisément. Ceci reste un problème complexe
dans le cas in vivo (où a priori aucune hypothèse ne peut-être faite sur la forme
géométrique de la zone à analyser). Il faudra alors avoir recours à des techniques
d’ajustement de surface 3D. C’est ce que nous détaillerons dans le chapitre IV.
III.5. Segmentation des images
III.5.1. Images µIRM: méthode d’étiquetage
Pour segmenter les images µIRM, nous avons utilisé un algorithme dédié développé dans le
laboratoire: la méthode d’étiquetage ([Antoniadis, 1998 (a)]). Cette technique itérative est
basée sur l’analyse du niveau de gris de chaque voxel et de son voisinage. Nous la présentons
succinctement dans cette partie.
L’initialisation de l’algorithme se fait en attribuant à chaque voxel, une probabilité
d’appartenance à une des deux phases à partir de son intensité (donc de son niveau de gris).
Compte tenu de la dégradation de la sensibilité de la sonde, cette probabilité initiale est établie
à partir de l’analyse de l’histogramme normalisé de chaque coupe transversale (on peut en
effet y supposer une réponse uniforme, cf. III.3.1.4). Comme celui présenté à la Figure III.3,
on peut en première approximation, le modéliser par une fonction h de la forme suivante
([Chung, 1995]):
)2)(
exp(2
)2exp()( 21
21
1
12
2
2G
G
G
G
RR
R xAxxAxh σ
µσπσσ
−−+
−=
où x est le niveau de gris, AR et σR définissent la distribution de Rayleigh qui caractérise les
voxels d’os (voxels de bruit) et AG1, µG1 et σG1 la distribution gaussienne qui caractérise les
voxels d’eau (voxels de signal). En ajustant h (par la méthode des moindres carrés,
135

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
136
[Press, 1989]), on peut obtenir une estimation de σR et µG1 pour chaque coupe transversale.
Pour les voxels de cette coupe qui ont un niveau de gris inférieur (resp. supérieur) à σR (resp.
µG1), la probabilité d’appartenance à la phase osseuse, POS, est fixée à 1 (resp. 0). Pour les
niveaux de gris intermédiaires, cette probabilité est établie à partir de la loi linéaire entre σR et
µG1 comme indiquée en Figure III.20.
0
0,005
0,01
0,015
0 50 100 150 200 250NG
POS Occurrence normalisée
σR µG1
POS = 1
Evolution linéaire de POS
POS = 0
Figure III.20: Evolution de la probabilité d’appartenance à la phase osseuse POS déterminée à partir des
caractéristiques de l’histogramme normalisé de chaque coupe transversale.
Par itérations successives, POS est recalculée pour chaque voxel afin de la faire tendre soit vers
1 soit vers 0 (ce qui permet d’obtenir une segmentation de l’image). Chaque itération se
déroule en trois étapes.
La première consiste à déterminer un segment (ensemble de voxels connectés) auquel le voxel
appartient parmi l’ensemble des segments que l’on peut définir sur un voisinage 3D (pour un
mode de connexité donné): le segment retenu est celui qui conduit à la variance la plus faible.
Deux attributs sont ensuite calculés: l’un lié à la moyenne des valeurs de POS pour les voxels
du segment, l’autre lié au contraste des probabilités entre les voxels du segment et ceux du
reste du voisinage (calculé comme l’écart entre la moyenne des valeur de POS des voxels du
segment et celle des voxels du reste du voisinage).
Enfin, la valeur de POS affectée à chaque voxel est recalculée à partir de ces deux attributs de
manière à la faire tendre vers 0 ou 1. Par exemple, POS augmente (et tend donc vers 1) pour un
voxel ayant ses deux attributs élevés, i.e. le voxel analysé fait partie d’un segment dont les
voxels ont eux-même une forte probabilité d’appartenir à la phase osseuse et qui se détachent
du reste du voisinage. Les voxels les plus problématiques sont ceux pour lesquels la valeur de

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
137
la moyenne des POS sur le segment est intermédiaire et où le contraste de probabilité sur le
voisinage est faible: pour ces voxels, l’itération ne modifie pas significativement POS et
l’évolution suivie par les probabilités de ces voxels (et donc leur convergence vers 0 ou 1)
n’est plus aussi fiable. Ce cas correspond typiquement à des voxels de volume partiel.
Bien qu’il n’y ait pas de preuve formelle de la convergence de cet algorithme, celle-ci a
toujours été obtenue (en général pour un nombre d’itérations inférieur à 20). On peut noter
que le RSB des images µIRM (cf. III.3.1.3) est suffisant pour assurer une très bonne qualité
de la segmentation. Un exemple de cette segmentation pour une coupe transversale d’un
échantillon est donné en Figure III.21.
Figure III.21: Exemple de segmentation d’ ne coupe transversale d’une image µIRM. u
III.5.2. Images par rayonnement synchrotron
III.5.2.1. Résolution nominale (10 µm)
A la résolution nominale, les images SR-µCT peuvent être segmentées directement par
seuillage simple: en effet, comme on l’a vu précédemment (cf. III.3.2.3), les deux modes
correspondant à la phase osseuse et à l’espace des pores sur l’histogramme des images
SR-µCT sont très nettement séparés (cf. Figure III.6). Un exemple de segmentation des
images SR-µCT (à la résolution nominale) est donné en Figure III.22. La convention adoptée
pour la représentation des deux phases est la même que pour les images µIRM (cf. Figure
III.21): les niveaux de gris 0 et 255 sont attribués respectivement au tissu calcifié et à l’espace
des pores.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
138
Figure III.22: Exemple de segmentation d’une coupe transversale d’une image SR-µCT. La convention
choisie pour la représentation est la même que celle des images µIRM: les niveaux de gris 0 et 255 sont
affectés respectivement à la phase calcifiée et à l’espace des pores (cf. Figure III.21).
III.5.2.2. Résolution dégradée: conservation de BV/TV0
A la résolution µIRM, la segmentation par seuillage simple des images SR-µCT dégradées est
plus ambiguë pour certains échantillons du fait de l’effet du volume partiel (cf. histogramme
de la Figure III.23, B).
Occurrence normalisée
NG
0
0,001
0,002
0 50 100 150 200 250
Occurrencenormalisée
NG
0
0,001
0,002
0 50 100 150 200 250
A B Figure III.23: Exemples d’histogramme de deux images SR-µCT dégradées. Dans le cas A, une
segmentation par seuillage simple resterait acceptable. Par contre, dans le cas B, la segmentation n’est
plus aussi simple que dans le cas de la résolution nominale. Sur ces histogrammes, on n’a représenté
volontairement que les occurrences normalisées inférieures à 0.002 afin de visualiser correctement le
mode correspondant aux voxels de signal (phase osseuse).
La technique SR-µCT étant une référence pour la mesure de la fraction osseuse (cf. I.5.4 et
III.3.2.1.3), les images SR-µCT dégradées ont été segmentées par un seuillage assurant la
conservation de ce paramètre mesuré à la résolution nominale. De cette manière, on garantit
que les images SR-µCT dégradées restent encore des références pour la fraction osseuse,

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
139
paramètre le plus pertinent pour la caractérisation de la résistance du tissu osseux (cf. I.4).
Cette valeur de fraction osseuse de référence sera notée dans la suite BV/TV0. Un exemple de
segmentation est donné en Figure III.24.
Figure III.24: Exemple de segmentation d’une image SR-µCT dégradée (conservation de la fraction
osseuse mesurée à haute résolution: BV/TV0). Pour comparaison, nous avons placé la même coupe
transversale que celle représentée pour l’image à haute résolution (cf. Figure III.22).
Remarque:
L’écart relatif entre le seuil conduisant à la conservation de la fraction osseuse et celui
déterminé comme le milieu des deux pics est très faible (environ 3%) pour les images
SR-µCT dégradées sur lesquelles le volume partiel a peu d’effet (image ayant un
histogramme du type de celui présenté à la Figure III.23, A). Il est plus important (environ
26%) pour les autres images (histogramme du type de celui présenté à la Figure III.23, B).
III.6. Confrontation des modalités d’imagerie
Afin d’établir la validité du rendu par IRM du tissu trabéculaire, on effectue une comparaison
des images obtenues par µIRM et par SR-µCT (technique de référence) pour les 29
échantillons, sur chaque paramètre présenté dans le chapitre II. Ces paramètres sont évalués
avec les mêmes algorithmes sur des ROI identiques entre les deux modalités d’imagerie
c’est-à-dire même zone analysée, même position de la ROI et même taille de voxel (cf. III.4).
Les effets de la résolution sur l’analyse d’image de tissu trabéculaire n’étant pas le but de
cette étude, on peut considérer que dans ces conditions, les différences observées ne sont dues
qu’aux techniques d’imagerie: grâce au recalage des ROI, on décorrèle les différences liées
aux paramètres d’acquisition et aux traitements des images de celles qui nous intéressent: les
phénomènes physiques à l’origine du contraste.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
La technique SR-µCT étant une référence pour la fraction osseuse (paramètre le plus pertinent
pour la caractérisation du tissu osseux, cf. I.4) ainsi que pour la microarchitecture (I.5.4 et
III.3.2.1.3), les comparaisons permettent d’obtenir directement le degré de fiabilité de la
µIRM pour chaque paramètre dans des conditions de résolution et de RSB proches de ce que
l’on peut obtenir in vivo (cf. chapitre IV).
III.6.1. ROI choisie et calcul des paramètres
La technique de repérage de l’enveloppe cylindrique utilisée pour le recalage des images (cf.
III.4.2.1) était appropriée pour calculer l’axe du cylindre mais ne serait pas assez rigoureuse
pour définir une zone à analyser (cf. III.4.2.3.4). Nous avons donc choisi, pour les deux
modalités, d’effectuer le calcul des paramètres sur le plus grand cylindre inclus dans
l’échantillon: imposer la ROI de cette façon permet une définition automatique très précise et
parfaitement reproductible.
Les images étant cubiques pour les deux modalités après le recalage des ROI (100 voxels de
côté, cf. III.4.6.1), le diamètre et la hauteur caractérisant les cylindres étudiés dans la suite ont
été choisis égaux à 100 voxels (soit 6.6 mm). Le volume analysé est donc d’environ 225 mm3,
soit 90% du volume total de tissu trabéculaire commun aux deux modalités (qui est limité à
250 mm3 du fait des acquisitions SR-µCT, cf. III.3.2.3). On peut visuellement se rendre
compte de cette restriction en comparant la Figure III.17 et la Figure III.25, où on a représenté
l’extérieur de la ROI analysée par un bord noir.
Sur Figure III.25, on peut observer un épaississement des travées osseuses sur les images
µIRM (A1,3) par comparaison avec celles de SR-µCT (B1,3 ou C1,3).
Par contre, il ne semble pas y avoir de différence entre les deux résolutions de la modalité
SR-µCT (entre B1,3 et C1,3): la dégradation ne paraît donc pas être la cause de cet
épaississement. La segmentation des images ne semble pas non plus en être l’origine puisque
les images segmentées (A4, B4, et C4) semblent traduire correctement la structure
trabéculaire présentées sur les images en niveaux de gris.
140

Chapitre III: Validité du rendu p laire ar IRM du tissu trabécu
141
A B C
1
2
3
4
Figure III.25: Exemple d’une coupe transversale (ligne 1: en niveaux de gris, ligne 2: segmentée) et d’une
coupe longitudinale « Y » (ligne 3: en niveaux de gris, ligne 4: segmentée) sur lesquelles on a limité la ROI
au plus grand cylindre inclus (bordure noire sur les images). Les colonnes A, B et C correspondent
respectivement aux images µIRM, SR-µCT à haute résolution et SR-µCT dégradée. Les volumes étant
recalés, toutes ces images sont carrées et représentent le même champ de vue cubique de 6.6 mm de coté
(soit 100 voxels pour les colonnes A et C et 660 voxels pour la colonne B). La comparaison des modalités
d’imagerie est basée sur le calcul des paramètres effectué sur les volumes (A2,A4) pour la µIRM et
(C2,C4) pour la SR-µCT dégradée (la ROI analysée étant définie par un cylindre de diamètre et de
hauteur égaux à 100 voxels).
Le calcul des paramètres est mené sur les ROI segmentées (présentées en Figure III.25) de
chaque échantillon pour les deux modalités selon le protocole que nous rappelons
succinctement ici (cf. chapitre II).
Sur la ROI cylindrique définie ci-dessus (environ 225 mm3):
• calcul de la fraction osseuse

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
• filtrage des amas secondaires d’os (en 26-connexité) et de moelle (en 6-connexité) (cf.
II.4.6.3)
• calcul de la fraction osseuse après filtrage des amas secondaires (pour vérifier qu’ils
représentent une fraction volumique négligeable) et évaluation de la contribution à la
caractéristique d’Euler-Poincaré (cf. II.4.6.3)
Sur la restriction de la ROI correspondante à la plus grande sphère incluse dans le cylindre
(cf. II.5.1.3.1). Cette sphère (unique) est centrée au centre du cylindre et a un diamètre égal à
la hauteur du cylindre (100 voxels, soit 6.6 mm), ce qui correspond à un volume d’environ
150 mm3:
• filtrage des amas secondaires d’os (en 26-connexité) et de moelle (en 6-connexité) (cf.
II.4.6.3) pouvant apparaître du fait de cette restriction
• calcul des paramètres d’histomorphométrie; les calculs de Tb.Th et Tb.Sp ont été menés
par la méthode indirecte en utilisant le modèle des plaques (cf. II.5.1.3.3 et II.5.2.1)
• calcul des paramètres d’anisotropie et d’orientation (cf. II.6.3 et II.6.4): le but de l’étude
étant de comparer les deux modalités (et non d’étudier l’orientation des travées par rapport
à un axe médical donné), nous avons choisi arbitrairement le repère de l’image comme
référence des trois angles d’Euler.
III.6.2. Principe de la comparaison
Pour chaque paramètre X, on représente chacun des 29 échantillons par un point de
coordonnées (XSR-µCT,XµIRM), où XSR-µCT et XµIRM correspondent respectivement à
l’évaluation du paramètre X sur la ROI « SR-µCT dégradée » et sur la (même) ROI
« µIRM ». Dans le cas idéal, les 29 points se retrouveraient sur la première bissectrice.
On analyse donc pour chaque paramètre le coefficient directeur a, l’ordonnée à l’origine b et
le coefficient de détermination R2 obtenus par le calcul de la régression linéaire effectuée sur
les 29 points.
On peut ainsi étudier la fiabilité de la µIRM pour chaque paramètre: la précision de la mesure
µIRM étant caractérisée par a et R2, et son biais par b. Le degré de signification des résultats,
noté p, est déterminé par test de Student apparié.
142

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
III.6.3. Résultats
Les 29 échantillons analysés couvrent une large gamme de fraction osseuse:
0.03 < BV/TV0 < 0.18. Rappelons que BV/TV0 est la mesure de référence de ce paramètre,
elle est évaluée sur les images SR-µCT à la résolution nominale (cf. III.5.2).
Pour les images analysées, les amas secondaires ne représentent qu’une fraction négligeable
de l’amas principal de la phase calcifiée. La valeur moyenne et l’écart type (µ ± σ) du rapport
VAS/VAP (cf. II.4.6.1) calculés sur les 29 échantillons sont de 0.60 ± 0.63% et de 0.64 ± 0.62%
respectivement pour les modalités µIRM et ST-µCT. Dans ces conditions, on a donc pu
appliquer le principe de filtrage des amas présenté en II.4.6.1.
Les valeurs moyennes et écarts type des différents paramètres, calculés sur les 29 échantillons
sont donnés dans le Tableau III.4. Les évaluations sont cohérentes avec celles déjà obtenues
dans d’autres travaux menés à des résolutions meilleures ou moins bonnes:
• pour les paramètres histomorphométriques dans le calcanéum ([Majumdar, 1998],
[Ulrich, 1999])
• pour la contribution à la caractéristique d’Euler-Poincaré dans le calcanéum
([Majumdar, 1998]) et dans la vertèbre ([Odgaard, 1993])
• pour les indices d’anisotropie dans le calcanéum ([Majumdar, 1998], [Ulrich, 1999])
143

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Paramètre µIRM SR-µCT
BV/TV 0.148 ± 0.047 0.089 ± 0.044
CN3 (mm-3) -2.69 ± 1.03 -2.95 ± 0.79
BS/TV (mm-1) 2.59 ± 0.53 2.10 ± 0.54
Tb.Th (mm) 0.11 ± 0.02 0.08 ± 0.02
Tb.Sp (mm) 0.69 ± 0.19 0.93 ± 0.27
MIL1 (mm) 0.72 ± 0.17 0.89 ± 0.24
MIL2 (mm) 0.81 ± 0.19 1.03 ± 0.28
MIL3 (mm) 0.97 ± 0.20 1.24 ± 0.26
RMIN 0.74 ± 0.06 0.71 ± 0.06
RMAX 0.92 ± 0.05 0.91 ± 0.05
Tableau III.4: Valeurs moyennes et écarts type (µ ± σ) calculés sur les 29 échantillons pour chaque
paramètre.
Le Tableau III.5 dresse le bilan des comparaisons entre les deux modalités pour chaque
paramètre (conformément au paragraphe III.6.2).
144

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
Paramètre a b R2 p
BV/TV 0.88 0.070 0.67 6×10-8
CN3 (mm-3) 0.97 0.16 0.70 2×10-8
BS/TV (mm-1) 0.69 1.14 0.49 2×10-5
Tb.Th (mm) 0.84 0.04 0.74 2×10-9
Tb.Sp (mm) 0.54 0.19 0.58 2×10-6
θ (deg) 0.92 3 0.97 4×10-22
ϕ (deg) 0.99 -2 0.99 2×10-28
ψ (deg) 0.94 -5 0.95 4×10-19
MIL1 (mm) 0.55 0.23 0.62 4×10-7
MIL2 (mm) 0.52 0.27 0.59 1×10-6
MIL3 (mm) 0.54 0.30 0.47 4×10-5
RMIN 0.86 0.13 0.90 5×10-15
RMAX 0.83 0.17 0.74 2×10-9
Tableau III.5: Comparaison des deux modalités pour chaque paramètre. On donne le coefficient directeur
a, l’ordonnée à l’origine b, le coefficient de détermination R2 et le degré de signification p (test de Student
apparié) correspondant à la régression linéaire (calculée sur les 29 échantillons) liant la modalité µIRM à
la modalité de référence (SR-µCT).
Dans la plupart des cas, les corrélations sont fortes (R2 > 0.58) et très significatives
(p < 2×10-6). Pour MIL3 et BS/TV, bien que les degrés de signification soient bons
(p < 4×10-5) , les corrélations sont moyennes (R2 > 0.47). Ces résultats sont analysés pour
chaque paramètre dans les paragraphes suivants.
III.6.3.1. Fraction osseuse
La Figure III.26 montre la confrontation entre les deux modalités pour le paramètre BV/TV.
La corrélation et le degré de signification de la régression sont bons (R2 = 0.67 et p = 6×10-8)
et le coefficient directeur est proche de 1 (0.88): la mesure de BV/TVµIRM est donc plutôt
précise. Cependant, on observe une ordonnée à l’origine de 0.07: ceci signifie que
145

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
146
BV/TVµIRM
BV/TVSR-µCT
l’évaluation de la fraction osseuse par µIRM est systématiquement biaisée de 0.07. Ce biais
correspond à des erreurs de l’ordre de 150% pour les échantillons ayant les plus faibles
fractions osseuses.
0
0,05
0,1
0,15
0,2
0,25
0 0,05 0,1 0,15 0,2 0,25 Figure III.26: Confrontation entre les deux modalités pour le paramètre BV/TV. Le trait plein correspond
à la régression linéaire des données (cf. Tableau III.5 pour les caractéristiques), le trait pointillé à la
première bissectrice.
Ceci semble traduire l’impression visuelle (cf. III.6.1 et Figure III.25). Nous reviendrons plus
en détail sur l’origine de ce biais dans la partie III.6.5.
III.6.3.2. Topologie et orientation de la microstructure
La Figure III.27 montre la confrontation entre les deux modalités pour les paramètres CN3 et
les trois angles d’Euler (θ, ϕ et ψ).
Pour ces deux familles de paramètres, a est proche de 1 et b négligeable. De plus, les
coefficients de détermination sont très proches de 1 et les degrés de signification sont très
bons (p < 2×10-8). L’évaluation de l’organisation générale de la microstructure trabéculaire est
très fiable en µIRM avec un voxel isotrope de taille 66 µm.

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
147
-5-5
-4
-3
-2
-1
CN3µIRM
(mm-3)
θSR-µCT (°)
θµIRM (°)
-4 -3 -2 -1
CN3SR-µCT
(mm-3)
A
-60
-20
20
60
100
-60 -20 20 60 100ϕSR-µCT (°)
ϕµIRM (°)
-150
-100
-50
0
50
100
150
-150 -100 -50 0 50 100 150ψSR-µCT (°)
ψµIRM (°)
-150
-50
50
150
250
-150 -50 50 150 250
B C D Figure III.27: Confrontation entre les deux modalités pour les paramètres CN3 (A), θ (B), ϕ (C) et ψ (D).
Le trait plein correspond à la régression linéaire des données (cf. Tableau III.5 pour les caractéristiques),
le trait pointillé à la première bissectrice.
III.6.3.3. Histomorphométrie
La Figure III.28 montre la confrontation entre les deux modalités pour les paramètres
histomorphométriques: BS/TV, Tb.Th et Tb.Sp.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
148
BS/TVSR-µCT (mm-1)
BS/TVµIRM (mm-1)
1
1,5
2
2,5
3
3,5
1 1,5 2 2,5 3 3,5Tb.ThSR-µCT (mm)
Tb.ThµIRM
(mm)
0,05
0,1
0,15
0,05 0,1 0,15Tb.SpSR-µCT (mm)
Tb.SpµIRM (mm)
0,4
0,6
0,8
1
1,2
1,4
0,4 0,6 0,8 1 1,2 1,4
A B C Figure III.28: Confrontation ente les deux modalités pour les paramètres histomorphométriques: BS/TV
(A), Tb.Th (B) et Tb.Sp (C). Le trait plein correspond à la régression linéaire des données (cf. Tableau
III.5 pour les caractéristiques), le trait pointillé à la première bissectrice.
L’évaluation par µIRM de cette famille de paramètres est très peu performante: les pentes
sont très différentes de 1, les ordonnées à l’origine non négligeables et les corrélations
moyennes. Pour ce qui est de Tb.Th et Tb.Sp, les mauvais résultats obtenus étaient prévisibles
du fait qu’ils dérivent directement de BS/TV et BV/TV.
Comme on peut le voir sur la Figure III.28 (B et C), Tb.Th est systématiquement surestimée et
Tb.Sp sous-estimé. D’après les résultats obtenus pour BV/TV (cf. III.6.3.1) et CN3 (cf.
III.6.3.2), cela semble confirmer l’hypothèse que le biais de fraction osseuse est dû à un
épaississement des travées osseuses et donc à une diminution de l’espacement des travées (cf.
III.6.1 et Figure III.25): « l’apparition » de fausses travées osseuses dans les images µIRM
paraît peu probable du fait que la topologie de la structure osseuse est correctement restituée
par µIRM.
III.6.3.4. Anisotropie
La Figure III.29 montre la confrontation entre les deux modalités pour les paramètres
d’anisotropie: MIL1, MIL2, MIL3 et les rapports RMIN et RMAX.

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
149
MIL1µIRM
(mm)
MIL1SR-µCT (mm)
0,5
0,7
0,9
1,1
1,3
1,5
0,5 0,7 0,9 1,1 1,3 1,5
MIL2SR-µCT (mm)
MIL2µIRM
(mm)
0,5
0,7
0,9
1,1
1,3
1,5
1,7
0,5 0,7 0,9 1,1 1,3
MIL3µIRM
(mm)
1,5 1,7MIL3
SR-µCT (mm)
0,60,6 0,8 1 1,2 1,4
0,8
1
1,2
1,4
1,6
1,8
1,6 1,8
A B C
RMINµIRM
RMINSR-µCT
0,5
0,6
0,7
0,8
0,9
0,5 0,6 0,7 0,8 0,9
RMAXµIRM
RMAXSR-µCT
0,8
0,9
1
0,8 0,9 1
D E
Figure III.29: Confrontation entre les deux modalités pour les paramètres d’anisotropie: MIL1 (A), MIL2
(B) , MIL3 (C) et les deux rapports RMIN (D) et RMAX (E). Le trait plein correspond à la régression linéaire
des données (cf. Tableau III.5 pour les caractéristiques), le trait pointillé à la première bissectrice.
De même que pour les paramètres histomorphométriques (cf. III.6.3.3), l’évaluation des trois
longueurs moyennes d’interception principales est très peu performante en µIRM: les pentes
sont très différentes de 1, les ordonnées à l’origine non négligeables et les corrélations
moyennes. Par contre les deux rapports d’anisotropie sont beaucoup plus fiables: les pentes et
les coefficients de corrélation sont beaucoup plus proches de 1.
MIL1, MIL2 et MIL3 sont systématiquement sous-estimées par µIRM ce qui est encore
cohérent avec l’hypothèse d’un épaississement des travées (cf. III.6.1 et Figure III.25). En
effet, dans ce cas, il y aurait plus d’intersections entre les deux phases par unité de longueur
entraînant une surestimation de NL (cf. II.5.1.2.1) et donc une sous-estimation des longueurs
moyennes d’interception par µIRM.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
150
BS/TV (mm-1)
BV/TV
III.6.4. Paramètres pertinents
Afin de mieux comprendre les résultats de la confrontation entre les deux modalités, nous
avons examiné si les paramètres principaux étaient systématiquement reliés les uns aux autres.
III.6.4.1. Lien entre paramètres histomorphométriques et fraction
osseuse
III.6.4.1.1. Loi phénoménologique
La Figure III.30 présente la surface spécifique, évaluée sur chaque échantillon, en fonction de
la fraction osseuse. Nous nous contenterons de l’analyse de BS/TV puisque c’est le paramètre
principal de la famille « histomorphométrie ».
1
1,5
2
2,5
3
3,5
0 0,05 0,1 0,15 0,2 0,25 Figure III.30: Etude de la relation entre la surface spécifique et la fraction osseuse (les marques en « carré
plein » correspondent à la modalité µIRM, celles en « losange vide » à la SR-µCT). On propose entre ces
deux paramètres, une loi phénoménologique linéaire (trait plein: µIRM, trait pointillé: SR-µCT). Les
caractéristiques de ces ajustements sont données dans au III.6. le Table
Une loi phénoménologique linéaire est proposée pour chaque modalité. Les caractéristiques
de ces ajustements sont précisées dans le Tableau III.6.

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
Modalité Paramètres de l’ajustement R2 p
aµIRM = 10.17 mm-1
µIRM bµIRM = 1.09 mm-1
0.76 7×10-10
aSR-µCT = 12.00 mm-1BS/TV = a× BV/TV + b
SR-µCT bSR-µCT = 1.04 mm-1
0.85 1×10-12
Tableau III.6: Récapitulatif des caractéristiques des deux lois phénoménologiques linéaires (une par
modalité) trouvées entre la surface spécifique et la fraction osseuse.
On peut remarquer une très forte dépendance de la surface spécifique à la fraction osseuse
pour les deux modalités (R2 > 0.76) avec un très bon degré de signification (p < 7×10-10).
Cette loi linéaire présente une ordonnée à l’origine non nulle d’environ 1 mm-1. Ceci semble a
priori paradoxal puisque intuitivement une fraction osseuse qui tend vers 0 semble devoir être
associée à une surface sur volume qui tend aussi vers 0: par exemple, on trouverait une
relation directement proportionnelle entre BS/TV et BV/TV si l’augmentation de la fraction
osseuse était liée à une augmentation du nombre de travées osseuses. Cependant, on a vérifié
que pour certains modèles présentant une anisotropie forte (par exemple pour des structures
générées à partir de formes tests cylindriques ou de plaques), une ordonnée à l’origine non
nulle pouvait apparaître pour la régression linéaire reliant BS/TV à BV/TV. Compte tenu du
caractère anisotrope du calcanéum, une telle loi serait possible dans notre cas. Il faudrait
toutefois d’autres échantillons à très faible fraction osseuse pour pouvoir mesurer avec plus de
précision la limite de BS/TV quand BV/TV tend vers 0 avant de conclure sur un potentiel
modèle de structure pour les échantillons de calcanéum étudiés (et éventuellement envisager
une autre loi phénoménologique que la loi linéaire pour caractériser la relation entre BS/TV et
BV/TV).
III.6.4.1.2. Influence sur la confrontation entre les deux modalités
Dans la suite, on note la loi phénoménologique entre BV/TV évaluée en µIRM et en SR-µCT
(c’est-à-dire BV/TV0): (BV/TV)µIRM = α×(BV/TV)SR-µCT + β. D’après le Tableau III.5, on a
α = 0.88, β = 0.07 (avec R2 = 0.67 et p = 5.8×10-8).
151

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
152
A partir de cette relation ainsi que de celles reliant BS/TV et BV/TV pour chaque modalité
(cf. Tableau III.6), on peut estimer facilement une loi linéaire entre (BS/TV)µIRM et
(BS/TV)SR-µCT:
BTVBSATVBS µCTSRµIRM +×= −)/()/(
avec:
α×= −µCTSR
µIRM
aa
A
µIRMµCTSRµCTSR
µIRMµIRM bb
aa
aB +××−×= −− αβ .
L’application numérique donne une pente A de 0.75 et une ordonnée à l’origine B de
1.02 mm-1. Cette estimation est à comparer avec ce qui a été obtenu expérimentalement (cf.
III.6.3.3 et Tableau III.5): a = 0.69 et b = 1.14 mm-1.
Sur la Figure III.31, on reprend le nuage de points correspondant à l’étude entre les deux
modalités (présenté dans la partie III.6.3.3) sur lequel on a tracé la loi estimée. On remarque
que cette loi est très proche de la loi trouvée expérimentalement.
BS/TVSR-µCT
(mm-1)
BS/TVµIRM (mm-1)
1
1,5
2
2,5
3
3,5
1 1,5 2 2,5 3 3,5 Figure III.31: Confrontation entre les deux modalités pour BS/TV. Par rapport à la Figure III.28 A, on a
rajouté la loi estimée à partir de la relation entre BV/TVµIRM et BV/TVSR-µCT (en trait fin sur la figure). Le
trait épais correspond à la régression linéaire des données (cf. Tableau III.5 pour les caractéristiques), le
trait pointillé à la première bissectrice.
Sur la gamme de fractions osseuses étudiées, la loi « inter-modalités » pour BS/TV peut donc
être très correctement estimée à partir de la relation expérimentale reliant BV/TV évaluée par
µIRM et par SR-µCT (c’est-à-dire BV/TV0). Ceci signifie que le biais sur la mesure de
BV/TV est l’origine principale des erreurs faites sur l’évaluation de la surface spécifique (et

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
153
BV/TV
RMIN
donc des deux autres paramètres histomorphométriques puisque Tb.Th et Tb.Sp dépendent
directement de BS/TV et de BV/TV).
III.6.4.1.3. Intérêt des paramètres histomorphométriques
Les valeurs de R2 étant élevées (cf. Tableau III.6), on peut conclure que les paramètres
histomorphométriques sont principalement déterminés par la valeur de BV/TV pour les
échantillons de calcanéum étudiés. Dans le cadre d’étude de modèles mathématiques
permettant de relier des données « mécaniques » (module d’Young par exemple) à une base
de paramètres de microstructure, ces attributs seront donc d’un intérêt réduit.
III.6.4.2. Lien entre paramètres d’anisotropie et fraction osseuse
La Figure III.32 présente pour chaque échantillon, les deux rapports d’anisotropie en fonction
de la fraction osseuse.
0,5
0,6
0,7
0,8
0,9
0 0,05 0,1 0,15 0,2 0,25BV/TV
RMAX
0,7
0,8
0,9
1
0 0,05 0,1 0,15 0,2 0,25
A B Figure III.32: Etude des relations entre les rapports d’anisotropie et la fraction osseuse: A: RMIN et B:
RMAX. Les marques en « carré plein » correspondent à la modalité µIRM, celles en « losange vide » à la
SR-µCT. On propose, dans chaque cas, une loi phénoménologique linéaire (trait plein: µIRM, trait
pointillé: SR-µCT). Les caractéristiques de ces ajustements sont données dans le Tableau III.7.
Une loi phénoménologique linéaire est proposée pour chaque modalité. Les caractéristiques
de ces ajustements sont précisées dans le Tableau III.7.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Modalité Paramètres de l’ajustement R2 p
cµIRM = -0.69 µIRM
dµIRM = 0.84 0.32 1×10-3
cSR-µCT = -1.19 RMIN = c×BV/TV + d
SR-µCT dSR-µCT = 0.81
0.66 9×10-8
eµIRM = -0.21 µIRM
fµIRM = 0.95 0.04 0.3
eSR-µCT = -0.13 RMAX = e×BV/TV + f
SR-µCT fSR-µCT = 0.92
0.01 0.6
Tableau III.7: Récapitulatif des caractéristiques des différentes lois phénoménologiques linéaires entre les
rapports d’anisotropie et la fraction osseuse, pour chaque modalité.
Les rapports d’anisotropie sont peu dépendants de la fraction osseuse comme cela a déjà été
observé pour des coefficients similaires à ceux-ci, évalués sur des images de
microtomographie X de 15 échantillons de radius ([Laib, 2001]).
On peut aussi noter que RMIN tend à décroître quand BV/TV augmente: la structure
trabéculaire semble d’autant plus anisotrope que la fraction osseuse est grande. Cependant,
pour les deux rapports d’anisotropie, les corrélations sont faibles ce qui confirme que ces
coefficients ne sont pas redondants avec la fraction osseuse. Ces paramètres pourraient donc
être pertinents dans le cadre d’études de modèle de prédiction de caractéristiques mécaniques
du tissu trabéculaire.
III.6.4.3. Caractéristique d’Euler-Poincaré
La Figure III.33 présente pour chaque échantillon, la valeur de la contribution à la
caractéristique d’Euler-Poincaré en fonction de la fraction osseuse et en fonction du rapport
principal caractérisant l’anisotropie de la structure.
154

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
155
BV/TV -5
-4
-3
-2
-1
0,5 0,6 0,7 0,8 0,9
RMIN
CN3 (mm-3)CN3
(mm-3)
-5
-4
0 0,05 0,1 0,15 0,2 0,25
-3
-2
-1
A B
Figure III.33: Etude des relations entre la contribution à la caractéristique d’Euler-Poincaré et la fraction
osseuse (A) et RMIN (B). Les marques en « carré plein » correspondent à la modalité µIRM, celles en
« losange vide » à la SR-µCT. Aucune loi phénoménologique simple n’a pu être trouvée entre ces
paramètres.
Comme on peut le voir, aucune loi phénoménologique ne semble relier CN3 à BV/TV ou à
RMIN. La contribution à la caractéristique d’Euler-Poincaré semble être un paramètre
indépendant qui pourrait certainement améliorer la qualité de modèles prédictifs des
propriétés biomécaniques du tissu trabéculaire. Cela a d’ailleurs déjà été établi sur des
modèles de structure interconnectées ([Feldkamp, 1988]).
III.6.5. Etude du biais sur la fraction osseuse
Le biais sur la mesure de BV/TV par µIRM est particulièrement gênant parce qu’il est la
cause principale de l’erreur commise sur son évaluation (cf. III.6.3.1) mais aussi parce qu’il
est fortement impliqué dans les erreurs observées sur certains paramètres (cf. III.6.4).
Dans cette partie, différentes origines possibles de ce biais sont étudiées individuellement: ce
type d’étude pourrait en effet permettre de déterminer un protocole d’imagerie et de
traitement pour le réduire et/ou le corriger.
III.6.5.1. Effet de la résolution
Pour estimer l’effet de la dégradation de la résolution sur la fraction osseuse, le protocole a été
le suivant:

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
• segmentation des images SR-µCT à 10 µm de résolution. Comme on l’a vu en III.5.2.1,
cette segmentation peut être réalisée sans ambiguïté par seuillage simple (on détermine
d’ailleurs la valeur de référence pour la fraction osseuse, BV/TV0, à partir de ces images)
• dégradation de ces images segmentées pour les ramener à la résolution µIRM (66 µm
isotrope) par la méthode du plan de Fourier (cf. III.4.3)
• segmentation de ces images dégradées. En effet, suite à la dégradation, des niveaux de gris
intermédiaires (autres que 0 ou 255) apparaissent dans l’image. La méthode de seuillage
simple peut encore être utilisée ici car les deux modes de l’histogramme de ces images
dégradées sont clairement séparables (contrairement au cas où la segmentation est
effectuée après la dégradation, cf. III.5.2.2)
• calcul du paramètre BV/TV sur ces images que l’on note dans la suite BV/TV1
La Figure III.34 illustre les différentes étapes de ce protocole. Celui-ci permet de n’observer
que l’effet de résolution sur le paramètre BV/TV puisque la dégradation est effectuée sur le
modèle de référence (image SR-µCT segmentée à résolution nominale) et que les étapes de
segmentation sont effectuées « sans ambiguïté ». La valeur BV/TV1 correspond donc à la
valeur « correcte » de la fraction osseuse de la structure trabéculaire mesurable à la résolution
µIRM.
156

Chapitre III: Validité du rendu laire par IRM du tissu trabécu
157
A B D
Occurrencenormalisée
NG0
0,005
0,01
0,015
0,02
0 50 100 150 200 250 C
Figure III.34: Illustration du protocole permettant d’étudier l’effet de la résolution sur la fraction osseuse.
A: Coupe transversale d’une image segmentée SR-µCT à 10 µm de résolution (660 voxels de coté). B:
Dégradation de l’image haute résolution pour la ramener à la résolution de la modalité µIRM (66 µm soit
100 voxels de coté): des niveaux de gris intermédiaires apparaissent comme on peut le voir sur
l’histogramme normalisé (C) (on n’a représenté volontairement que les occurrences normalisées
inférieures à 0.02 afin de visualiser correctement le mode correspondant aux voxels de la phase osseuse).
Les deux modes restant très nettement séparables, ces images sont segmentées par seuillage simple (D).
La Figure III.35 présente BV/TV1 en fonction de BV/TV0 pour les 29 échantillons ainsi que
l’ajustement linéaire associé (a = 1.00, b = 0.00, R2 = 1.00 et p = 2×10-40). Pour comparaison,
on a tracé sur ce même graphe la première bissectrice. On peut donc clairement conclure que
la dégradation de la résolution n’est pas à l’origine du biais sur la fraction osseuse. Ce résultat
confirme d’ailleurs l’impression visuelle (cf. III.6.1 et Figure III.25).

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
158
0
0,05
0,1
0,15
0,2
0 0,05 0,1 0,15 0,2
BV/TV0
BV/TV1
Figure III.35: BV/TV1 en fonction de BV/TV0 calculées pour les 29 échantillons. Le trait plein correspond
à la régression linéaire des données, le trait pointillé à la première bissectrice.
III.6.5.2. Effet de la méthode de segmentation
Afin de tester l’effet sur l’évaluation de BV/TV de la méthode de segmentation employée
pour les images µIRM, on a comparé les 29 valeurs obtenues pour BV/TVµIRM (c’est-à-dire à
l’aide de la méthode de segmentation par étiquetage, cf. III.5.1) avec celles calculées à partir
des images µIRM segmentées par seuillage simple: BV/TVSEUIL.
La Figure III.36 présente un exemple d’une même coupe transversale de l’image µIRM d’un
échantillon, segmentée par la méthode d’étiquetage (cf. III.5.1) et par seuillage simple.
A B
Figure III.36: Exemple d’une coupe transversale d’une image µIRM segmentée par la méthode
d’étiquetage (cf. III.5.1) (A) et par seuillage simple (B).
La Figure III.37 montre, pour les 29 échantillons, BV/TVSEUIL en fonction de BV/TVµIRM
ainsi que l’ajustement linéaire associé (a = 1.12, b = -0.03, R2 = 0.99 et p = 2×10-28). Pour
comparaison, on a tracé sur ce même graphe la première bissectrice.

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
159
BV/TVµIRM
BV/TVSEUIL
0
0,05
0 0,05 0,1 0,15 0,2 0,25
0,1
0,15
0,2
0,25
Figure III.37: BV/TVµIRM en fonction de BV/TVSEUIL calculées sur les 29 échantillons. Le trait plein
correspond à la régression linéaire des données, le trait pointillé à la première bissectrice.
La pente est légèrement supérieure à 1 mais l’ordonnée à l’origine est légèrement négative:
dans la gamme de fraction osseuse étudiée, il n’y a aucune différence notable entre les deux
méthodes de segmentation (l’ajustement est très proche de la première bissectrice). La
technique de segmentation employée n’est donc pas la cause du biais sur l’évaluation de la
fraction osseuse par µIRM (ce qui confirme là aussi l’impression visuelle, cf. III.6.1 et Figure
III.25).
Ceci illustre aussi le fait que les images µIRM ont un RSB bien supérieur au minimum requis
pour la méthode de segmentation par étiquetage (cf. III.5.1). Les deux modes de
l’histogramme (cf. Figure III.3) sont donc suffisamment séparés pour qu’une segmentation
par seuillage simple soit assez précise.
III.6.5.3. RSB des images µIRM et interpolation « zerofill »
La taille de voxel des images µIRM (66 µm isotrope) a été obtenue à partir d’une acquisition
à une résolution de 132 µm isotrope suivie d’une interpolation par la méthode du « zerofill »
d’un facteur 2 dans chaque direction. Le RSB de ces images était d’environ 17 (cf. III.3.1.3).
Afin de quantifier l’influence de l’interpolation par « zerofill » et du RSB sur l’évaluation de
la fraction osseuse par µIRM, des volumes simulant l’acquisition µIRM ont été générés à
partir des images SR-µCT à haute résolution selon le protocole suivant:
• segmentation des images SR-µCT à 10 µm de résolution par seuillage simple:
l’application de cette méthode n’étant pas ambiguë, la fraction osseuse de référence
BV/TV0 est déterminée sur ces images (cf. III.5.2)

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
• dégradation de ces images segmentées pour obtenir une taille de voxel de 66 µm isotrope
(résolution des images µIRM après l’interpolation par « zerofill ») par la méthode du plan
de Fourier (cf. III.4.3)
• calcul de la transformée de Fourier de ces images et rajout d’un bruit gaussien d’amplitude
réglable
• mise à zéro de la moitié des voxels du plan de Fourier, sur les bords, dans chaque
direction. Le plan de Fourier obtenu simule alors celui correspondant à une image µIRM:
les points centraux représentent les points réellement acquis et les points à zéro sur les
bords (autant que de points centraux) correspondent à ceux rajoutés pour effectuer
l’interpolation d’un facteur 2 par « zerofill »
• calcul de la transformée de Fourier inverse et mesure du RSB: l’amplitude de bruit est
alors ajustée afin d’obtenir un RSB proche de 17 (valeur mesurée sur les images µIRM, cf.
III.3.1.3). L’histogramme obtenu est bien caractéristique d’une image µIRM (cf. Figure
III.3)
• segmentation de ces images: comme la méthode de segmentation n’est pas à l’origine du
biais sur l’évaluation de la fraction osseuse (cf. III.6.5.2), celle-ci est réalisée par seuillage
simple
• calcul du paramètre BV/TV sur ces images que l’on note dans la suite BV/TV2
La Figure III.38 illustre les différentes étapes de ce protocole. BV/TV2, qui est calculée à
partir du modèle de référence (SR-µCT à haute résolution) ne rend compte que des effets de
l’interpolation par « zerofill » et du RSB qui caractérisent les acquisitions µIRM.
160

Chapitre III: Validité du rendu laire par IRM du tissu trabécu
161
A B C
D E G
Occurrencenormalisée
NG0
0,005
0,01
0,015
0,02
0 50 100 150 200 250 F
Figure III.38: Illustration du protocole permettant d’étudier l’effet de l’interpolation par « zerofill » et du
RSB sur la fraction osseuse. A: Coupe transversale d’une image segmentée SR-µCT à 10 µm de résolution
(660 voxels de coté). B: Dégradation de l’image haute résolution pour la ramener à la résolution de la
modalité µIRM (66 µm soit 100 voxels de coté): des niveaux de gris intermédiaires apparaissent. C:
Module de la transformée de Fourier de l’image B. D: Simulation du plan de Fourier d’une acquisition
µIRM: ajout d’un bruit gaussien (pour obtenir un RSB d’environ 17) et mise à zéro de la moitié des voxels
du plan de Fourier, sur les bords, dans chaque direction. E: Image µIRM simulée avec son histogramme
normalisé (F) caractéristique (cf. Figure III.3). L’évaluation de BV/TV2, est effectuée sur l’image G qui
correspond au seuillage simple de l’image E.
La Figure III.39 présente BV/TV2 en fonction de BV/TV0 pour les 29 échantillons ainsi que
l’ajustement linéaire associé (a = 1.09, b = -0.01, R2 = 0.99 et p = 2×10-28). Pour comparaison,
on a tracé sur ce même graphe la première bissectrice.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
162
0
0,05
0 0,05 0,1 0,15 0,2
BV/TV0
0,1
0,15
0,2BV/TV2
Figure III.39: BV/TV2 en fonction de BV/TV0 calculées pour les 29 échantillons. Le trait plein correspond
à la régression linéaire des données, le trait pointillé à la première bissectrice.
La pente légèrement supérieure à 1 est compensée par une ordonnée à l’origine négative: dans
la gamme de fraction osseuse étudiée, la différence entre BV/TV2 et BV/TV0 est négligeable
(l’ajustement est très proche de la première bissectrice). On peut donc clairement conclure
que l’interpolation par « zerofill » ainsi que le niveau du RSB des acquisitions ne sont pas à
l’origine du biais sur l’évaluation de la fraction osseuse.
III.6.5.4. Premiers résultats sur les effets du T2*
Une autre hypothèse pour l’origine du biais sur l’évaluation de la fraction osseuse par µIRM
pourrait être la différence de susceptibilité magnétique au niveau de l’interface entre le tissu
trabéculaire et l’eau (étude in vitro). Cette différence créerait des inhomogénéités locales de
champ magnétique accélérant le déphasage des spins donc diminuant la durée du signal de
précession libre: effets « T2* » (cf. I.6.4.3.1). Cette perte « artificielle » de signal au niveau de
l’interface entre les deux phases pourrait donc être à l’origine de l’épaississement des travées:
les voxels de l’interface tissu calcifié - eau se trouveraient caractérisés par un faible signal
donc comme appartenant à la phase osseuse après segmentation. Ce phénomène pourrait donc
être à l’origine de la surévaluation de la fraction osseuse.
III.6.5.4.1. Effet de la bande passante
Pour modifier l’influence du T2*, on peut agir sur le paramètre de bande passante. En effet,
comme on l’a vu en I.6.4.1, la bande passante est liée à l’inverse du temps d’observation du
signal de précession libre. Pour une image acquise à bande passante élevée (donc à temps

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
163
d’observation court), l’effet de la décroissance T2* est moins sensible que pour une image
acquise à faible bande passante (donc à temps d’observation élevée) et devrait donc permettre
une estimation moins biaisée de la fraction osseuse.
Deux images d’un même échantillon ont été acquises dans les mêmes conditions que
précédemment, pour deux bandes passantes différentes: une proche de celle utilisée pour les
images des 29 échantillons (cf. III.3.1.2): 60 kHz et une autre pour une bande passante moitié:
30 kHz. Ces deux images ont ensuite été segmentées par la méthode d’étiquetage (cf. III.5.1).
La Figure III.40 montre un exemple pour ces deux images.
A B
C D
Figure III.40: Coupe transversale de l’image d’un échantillon acquise avec une bande passante de 60 kHz
(en niveaux de gris: A et segmentée: B) et avec une bande passante de 30 kHz (en niveaux de gris: C et
segmentée: D). La bordure noire sur les images B et D correspond à l’extérieur de la ROI sur laquelle ont
été menés les calculs (plus grand cylindre inclus dans l’échantillon).
Après les mêmes traitements, l’image acquise pour la bande passante de 60 kHz a bien une
fraction osseuse plus faible (0.24) que pour celle acquise à 30 kHz (0.26) mais sans pour
autant atteindre l’écart de 0.07 observé par la confrontation des modalités (cf. III.6.3.1).
III.6.5.4.2. Simulations
Pour compléter cette étude, une simulation de l’effet T2* a aussi été envisagée en utilisant des
opérateurs de morphologie mathématique simples (érosion et dilatation). Différents éléments
structurants ont été testés (de taille variable en fonction de la résolution des images sur
lesquelles ils étaient appliqués):

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
• cube ou croix: pour simuler un effet « isotrope »
• ligne selon un des trois axes pour simuler un effet « anisotrope »: l’axe de lecture est plus
affecté par l’effet T2* que les autres.
Deux études ont été envisagées: diminution de la fraction osseuse des images µIRM par
érosion (pour la comparer à BV/TV0) ou augmentation de la fraction osseuse des images
SR-µCT par dilatation (pour la comparer à BV/TVµIRM). Pour la deuxième approche, la
dilatation a été calculée à différentes résolutions pour permettre un contrôle plus fin des effets
de l’opérateur morphologique sur la fraction osseuse. A partir des images SR-µCT de
référence (images SR-µCT à haute résolution segmentées par seuillage simple, cf. III.5.2.1),
des images SR-µCT à 20 µm et 40 µm de résolution ont pu être calculées à partir de la
méthode présentée en III.6.5.1. Sur ces images, l’opérateur de dilatation a été appliqué avant
de revenir à la résolution µIRM.
Aucune érosion des images µIRM n’a pu être appliquée sans faire chuter très sensiblement la
fraction osseuse et modifier la structure trabéculaire.
La Figure III.41 montre le nuage de points (10 images analysées) correspondant à la relation
entre BV/TV0 et la fraction osseuse, notée BV/TV3, évaluée après dilatation par un cube de
3×3×3 voxels de chaque image SR-µCT à 40 µm, ramenée ensuite à la résolution µIRM.
L’ajustement linéaire donne une pente de 1.35, une ordonnée à l’origine de 0.09 (avec
R2 = 0.92 et p = 1×10-5). Pour comparaison, l’ajustement linéaire qui a été calculé entre
BV/TV0 et BV/TVµIRM (cf. III.6.3.1) est aussi indiqué sur ce graphe.
164

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
165
BV/TV0
BV/TV3
0
0,05
0 0,05 0,1 0,15 0,2 0,25 0,3 0,35
0,1
0,15
0,2
0,25
0,3
0,35
Figure III.41: BV/TV3 en fonction de BV/TV0 calculées pour 10 images SR-µCT (à partir de la résolution
40 µm). Le trait plein correspond à la régression linéaire des données, le trait pointillé à la première
bissectrice et le trait en long pointillé correspond à l’ajustement linéaire effectué sur les 29 échantillons
entre BV/TVµIRM et BV/TVSR-µCT (cf. III.6.3.1).
La Figure III.42 présente aussi le même type de résultats mais entre BV/TV0 et la fraction
osseuse, notée BV/TV4, évaluée après dilatation de chaque image SR-µCT à 40 µm par une
ligne de 3 voxels (orientée dans le sens de l’axe de lecture), ramenée ensuite à la résolution
µIRM.
0
0,05
0,1
0,15
0,2
0,25
0,3
0 0,05 0,1 0,15 0,2 0,25 0,3
BV/TV0
BV/TV4
Figure III.42: BV/TV4 en fonction de BV/TV0 calculées pour 10 images SR-µCT (à partir de la résolution
40 µm). Le trait plein correspond à la régression linéaire des données, le trait pointillé à la première
bissectrice et le trait en long pointillé correspond à l’ajustement linéaire effectué sur les 29 échantillons
entre BV/TVµIRM et BV/TVSR-µCT (cf. III.6.3.1).

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
L’ajustement linéaire donne une pente de 1.22, une ordonnée à l’origine de 0.02 (avec
R2 = 0.99 et p = 3×10-9).
Dans les deux cas présentés en Figure III.41 et Figure III.42, on a bien un biais non nul mais
relativement différent de ce qui a été déterminé entre BV/TV0 et BV/TVµIRM (0.09 ou 0.02 à
comparer avec 0.07, cf. III.6.3.1). De plus, la pente est plus affectée (1.35 ou 1.22 à comparer
avec 0.88, cf. III.6.3.1).
III.6.5.4.3. Conclusions
Afin d’établir la distribution du champ magnétique sur un modèle de tissu trabéculaire, Ford
et al. ([Ford, 1993]) ont utilisé la valeur de -4π×0.3 ppm (unité MKSA) pour la différence de
susceptibilité entre le tissu calcifié et la moelle (du même ordre de grandeur que celle entre le
tissu calcifié et l’eau). Cette valeur numérique a été obtenue en sommant les contributions
élémentaires attendues d’après la composition atomique du tissu osseux. Pour l’imageur sur
lequel les images ont été réalisées (8.5 T), cela correspondrait à une variation de fréquence
maximum de ∆νSUS = 1.3 kHz. De plus, l’inhomogénéité du champ local dépend de façon
complexe de la géométrie des structures et de leur orientation par rapport au champ principal
([Callaghan, 1991]). Il a ainsi été démontré soit théoriquement ([Yablonskiy, 1994]) soit
expérimentalement ([Engelke, 1994], [Selby, 1996]) que la valeur de T2* pour un fluide dans
différents fantômes modélisant la structure osseuse dépendait des paramètres d’architecture
(espacement et épaisseur des travées) et de son orientation par rapport au champ statique 0Br
.
Selon la modélisation numérique de cet effet proposée par Ford et al. ([Ford, 1993]), ∆νSUS
représente la variation maximale de fréquence de résonance sur une échelle de longueur dans
le fluide comparable à l’épaisseur des travées. Dans nos expériences, la bande passante
utilisée est de 55 kHz, soit 400 Hz pour un voxel de taille 132 µm. Cette valeur est trois fois
plus faible que ∆νSUS. Pourtant, nous n’avons pas observé de différence sensible sur la
fraction osseuse en comparant deux images du même échantillon acquises avec une bande
passante de 60 kHz et 30 kHz (cf. III.6.5.4.1). L’intensité de gradient disponible ne permet
pas d’envisager une acquisition à bande passante plus élevée, qui serait de surcroît affectée
d’une perte de sensibilité.
Cette étude est encore incomplète mais il semble particulièrement difficile d’établir l’origine
exacte du biais sur les mesures de la fraction osseuse par µIRM. D’après nos observations,
166

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
l’hypothèse la plus probable serait la présence de microbulles d’air confinées à l’interface
entre le tissu calcifié et l’eau. La différence de susceptibilité magnétique entre l’air et l’eau est
de -4π×0.7 ppm (unité MKSA). Ceci pourrait donc causer une inhomogénéité spatiale de la
différence de susceptibilité magnétique puisque l’interface serait de deux natures: entre le
tissu calcifié et l’eau ou entre l’air et l’eau. Cet effet ne semble cependant pas pouvoir être
simulé de manière simple: il faudrait donc envisager des modèles plus complexes pour
essayer de définir un protocole de correction fiable des images.
III.7. Recalage des ROI: effet sur la comparaison entre
modalités
Dans cette dernière partie, on a essayé de quantifier l’intérêt du recalage des ROI pour la
confrontation entre les deux modalités en menant le même type d’étude mais à partir de ROI
choisies arbitrairement dans les volumes µIRM.
III.7.1. Méthodes
Sur chaque image µIRM, le plus grand parallélépipède rectangle ne contenant que du tissu
trabéculaire a été extrait (cette forme a été choisie pour sa facilité de mise en œuvre). Ces ROI
« en pavé » n’ont été positionnées qu’à partir de considérations géométriques. La Figure
III.43 donne un exemple de ce type de ROI.
Sur ces ROI « en pavé », les paramètres précédents (cf. chapitre II) ont été calculés selon le
protocole qui est rappelé en III.6.1, c’est-à-dire sur toute la ROI pour la fraction osseuse et la
contribution à la caractéristique d’Euler-Poincaré, et sur la restriction à la plus grande sphère
incluse, pour les autres paramètres.
167

Caractérisation de l’architecture tridimension laire par IRM nelle du tissu trabécu
168
A B C
D E F Figure III.43: Exemple de ROI « en pavé » définie sur une image µIRM. La taille et la position de cette
ROI est propre à chaque échantillon. Le volume analysé est en moyenne de 176 mm3. Le rectangle blanc
sur chaque coupe (A: longitudinale « X », B: longitudinale « Y », C: transversale) correspond à
l’empreinte de cette ROI. Les images D, E et F montr nt la segmentation de la ROI présentée. e
III.7.2. Résultats
III.7.2.1. Volume analysé
En moyenne sur les 29 échantillons, le volume de tissu trabéculaire récupéré en utilisant les
ROI « en pavé » était de 176 mm3 soit 78% du volume analysé avec les ROI recalées (cf.
III.6.1). La restriction de la ROI « en pavé » à la plus grande sphère incluse limitait encore
plus la zone de calcul: 47 mm3 en moyenne sur les 29 échantillons soit seulement 31% des
données analysées sur les ROI préalablement recalées (cf. III.6.1). On voit ici le premier
avantage du recalage: on a en effet pu maximiser le volume de l’échantillon sur lequel la
confrontation des deux modalités d’imagerie a été menée. Nous avons aussi observé à
posteriori qu’en moyenne sur les 29 échantillons, 77% des voxels des ROI « en pavé » se
retrouvaient dans les ROI cylindriques communes: en d’autres termes, seulement 23% des
voxels analysés sur les ROI « en pavé » apportaient une « information nouvelle » par rapport
aux ROI cylindriques communes.

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
III.7.2.2. Erreur relative
Le Tableau III.8 détaille la moyenne µ et l’écart type σ de la valeur absolue de l’erreur
relative sur l’évaluation des paramètres par référence aux valeurs obtenues par SR-µCT (sur
la ROI cylindrique commune, cf. Figure III.25) pour les deux types de ROI définies en µIRM:
« en pavé » et cylindrique commune à la modalité SR-µCT (µ et σ ont été calculés sur les 29
échantillons).
Entre les deux types de ROI, deux facteurs agissent simultanément sur l’évaluation des
paramètres:
• la position de la ROI « en pavé » n’est pas la même que celle de la ROI cylindrique
commune. Ceci a évidemment un effet direct sur les paramètres caractérisant l’orientation
de la microstructure. Les évaluations sur les ROI « en pavé » des premier et troisième angles
d’Euler (θ et ψ) présentent les augmentations d’erreur les plus importantes de tous les
paramètres. Elle quintuple pour θ (+71%) et elle double pour ψ (+43%). Bien que l’erreur
augmente de plus du double pour le deuxième angle d’Euler ϕ (5% au lieu de 2% pour
l’évaluation sur les ROI cylindriques communes), elle reste faible ce qui s’explique par la
très faible correction d’inclinaison qui a été nécessaire pour repositionner les images µIRM
(2.5° en moyenne sur les 29 échantillons, cf. Tableau III.2).
• la zone analysée n’est pas de même taille et une partie du volume des ROI « en pavé » est
située à l’extérieur des ROI cylindriques communes. Pour la fraction osseuse (paramètre
insensible aux positions relatives des ROI), on observe une augmentation de l’erreur de
27%. Ceci semble indiquer une dépendance spatiale de ce paramètre, probablement liée à
l’anatomie du site osseux. Par contre, pour les paramètres topologiques,
d’histomorphométrie et d’anisotropie, l’erreur n’augmente que de l’ordre de 10% alors que
pour les deux dernières familles, le volume de données analysées (restriction de la ROI à la
plus grande sphère) représente moins d’un tiers du volume étudié sur les ROI recalées. Il
semblerait donc qu’en ce qui concerne ces paramètres, un « motif » représentatif de la
structure pourrait être extrait.
169

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
Erreur relative à l’évaluation
SR-µCT (en %) µIRM ROI « en pavé »
µIRM ROI cylindrique
commune
BV/TV 111 ± 70 84 ± 57
CN3 18 ± 16 15 ± 12
BS/TV 40 ± 31 27 ± 24
Tb.Th 43 ± 25 42 ± 18
Tb.Sp 31 ± 15 24 ± 14
θ 89 ± 114 18 ± 20
ϕ 5 ± 7 2 ± 3
ψ 74 ± 178 31 ± 85
MIL1 24 ± 14 17 ± 13
MIL2 25 ± 15 19 ± 14
MIL3 28 ± 13 21 ± 13
RMIN 7 ± 4 4 ± 3
RMAX 3 ± 2 2 ± 2
Tableau III.8: Moyenne et l’écart type (µ ± σ) de la valeur absolue de l’erreur relative sur l’évaluation des
paramètres par référence aux valeurs obtenues par SR-µCT sur la ROI cylindrique commune (µ et σ ont
été calculés sur les 29 échantillons).
III.7.2.3. Confrontation entre les deux modalités
Le Tableau III.9 donne les caractéristiques des ajustements linéaires entre les deux modalités
(le coefficient directeur a, l’ordonnée à l’origine b, le coefficient de détermination R2 et le
degré de signification p déterminé par test de Student apparié) qui auraient été trouvés si les
comparaisons avaient été faites entre les ROI « en pavé » pour la µIRM et les ROI
cylindriques pour la SR-µCT.
170

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
Paramètre a b R2 p
BV/TV 0.83 0.090 0.56 3×10-6
CN3 (mm-3) 1.21 0.35 0.66 9×10-8
BS/TV (mm-1) 0.65 1.48 0.34 9×10-4
Tb.Th (mm) 0.80 0.05 0.52 1×10-5
Tb.Sp (mm) 0.42 0.23 0.44 9×10-5
θ (deg) 1.33 -9 0.68 4×10-8
ϕ (deg) 0.96 -2 0.97 4×10-22
ψ (deg) 0.85 -2 0.87 2×10-13
MIL1 (mm) 0.45 0.26 0.52 1×10-5
MIL2 (mm) 0.39 0.34 0.41 2×10-4
MIL3 (mm) 0.40 0.38 0.33 1.1×10-3
RMIN 0.91 0.11 0.79 1×10-10
RMAX 0.62 0.35 0.51 1×10-5
Tableau III.9: Comparaisons entre les deux modalités d’imagerie. Les paramètres sont évalués sur les
ROI « en pavé » pour la µIRM et sur les ROI cylindriques communes pour la SR-µCT. On donne le
coefficient directeur a, l’ordonnée à l’origine b, le coefficient de détermination R2 et le degré de
signification p (test de Student apparié) correspondant à la régression linéaire (calculée sur les 29
échantillons) liant les deux modalités. Ce tableau est à comparer avec celui obtenu pour des ROI recalées:
Tableau III.5 (cf. III.6.3).
On peut constater une dégradation sensible de tous les indices par rapport à ceux obtenus sur
des ROI communes (cf. III.6.3: Tableau III.5), sauf pour le coefficient directeur associé à la
régression linéaire de RMIN. Cependant, compte tenu de l’ordonnée à l’origine trouvée, la
qualité de l’ajustement reste moins bonne pour la gamme de valeurs de RMIN obtenues (i.e.
l’ajustement linéaire est plus « éloigné » de la première bissectrice que dans le cas de la
comparaison sur les ROI communes).
171

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
III.7.2.4. Conclusion
En utilisant des ROI non recalés, les différences observées entre les modalités ne sont pas
dues qu’aux caractéristiques propres aux deux techniques d’imagerie. D’autres sources
d’erreur interviennent avec des effets plus ou moins importants pour les paramètres ce qui
empêche de tirer des conclusions fiables. De plus, le recalage constitue une étape essentielle si
on souhaite effectuer la comparaison des orientations de la structure.
III.8. Conclusions
Nous avons donc pu établir grâce à cette étude in vitro les domaines de fiabilité de l’IRM pour
la caractérisation du tissu trabéculaire.
Les conclusions tirées sont très robustes pour deux raisons principales.
Premièrement, la référence que nous avons utilisée, la microtomographie X à haute résolution
par rayonnement synchrotron, permet d’accéder à un modèle du tissu trabéculaire de très
bonne qualité: les images étudiées étaient caractérisées par un RSB particulièrement élevé et
une résolution parfaitement adaptée à la taille caractéristique des travées osseuses.
Deuxièmement, nous avons bien veillé à restreindre au maximum les sources d’erreur
externes en limitant les degrés de liberté: l’étude a été menée sur les mêmes ROI, à même
taille de voxel (les effets de la résolution étaient hors du cadre de cette étude) et en appliquant
les mêmes algorithmes pour les deux modalités. Ce cadre de travail nous a donc permis de ne
caractériser que les différences liées aux phénomènes physiques à l’origine du contraste:
absorption X pour la microtomographie et densité de protons (eau) pour l’IRM.
Nous avons pu établir que l’organisation générale de la microstructure (c’est-à-dire la
contribution à la caractéristique d’Euler-Poincaré, les trois angles d’Euler et les deux
coefficients d’anisotropie) était restituée avec une grande fiabilité par IRM à 66 µm de
résolution isotrope. L’évaluation de la fraction osseuse par IRM, bien que relativement
précise, était affectée par un biais systématique de 0.07. Les résultats les moins bons ont été
observés pour les paramètres histomorphométriques et les trois longueurs moyennes
d’interception principales: leur très forte dépendance avec la fraction osseuse en était la cause
principale.
172

Chapitre III: Validité du rendu par IRM du tissu trabéculaire
Nous avons essayé d’envisager plusieurs origines possibles au biais sur la fraction osseuse.
Nous avons vérifié qu’il n’était dû ni à la dégradation de résolution, ni à la méthode de
segmentation, ni aux traitements préalables des images µIRM. L’« effet T2* » homogène a
aussi été envisagé et simulé. L’hypothèse la plus probable semblerait être un « effet T2* »
réparti de manière non homogène dans l’espace, causé par exemple par la présence de
microbulles d’air confinées à l’interface entre le tissu calcifié et l’eau. La modélisation d’un
tel effet semble cependant difficile: aucune méthode de correction de ce biais n’a pu être mise
en œuvre pour le moment.
Le recalage des images (qui a permis pour la première fois de comparer l’orientation de la
structure) et l’ajustement des tailles de voxel n’ont jamais été mis en œuvre simultanément
dans les études précédentes et les résultats étaient naturellement moins précis et moins fiables
(cf. III.1).
Nous disposons maintenant d’une série d’outils robustes pour la caractérisation des images de
tissu trabéculaire obtenues par IRM à une résolution accessible in vivo. Le but du chapitre
suivant est de les mettre en œuvre sur des acquisitions obtenues in vivo.
173


Chapitre IV: Imagerie 3D in vivo du doigt humain
Chapitre IV: Imagerie 3D in vivo du doigt
humain: faisabilité de l’analyse du tissu
trabéculaire in vivo
IV.1. IRM du tissu trabéculaire in vivo: état de l’art
De nombreux travaux sur le tissu trabéculaire ont déjà été menés à partir d’images obtenues in
vivo par IRM. Trois axes ont été principalement envisagés: la recherche de paramètres
permettant de discriminer une population saine d’une population pathologique, l’évolution des
caractéristiques du tissu avec l’âge, et le suivi des effets de traitements dans le temps (« étude
longitudinale »). Toutes ces études ont été menées sur des imageurs IRM à 1.5 T, disponibles
couramment en milieu hospitalier
IV.1.1. Discrimination entre groupes de patients
Majumdar et al. ([Majumdar, 1997]) ont étudié trois groupes de patientes (un premier de 10
femmes avant la ménopause, un second de 9 femmes « post ménopause » et un dernier de 11
femmes « post ménopause » atteintes d’ostéoporose) à l’aide de paramètres
histomorphométriques calculés sur des images de radius (60 coupes d’épaisseur 700 µm avec
une résolution dans le plan de 156×156 µm2). L’histogramme présenté étant monomodal, les
images ont été segmentées avec un seuil arbitraire déterminé par la méthode détaillée dans
175

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
[Majumdar, 1996] (que nous avons présentée succinctement en II.2.3.1). Sur des ROI ne
contenant que du tissu trabéculaire (définies manuellement sur chaque coupe), la fraction
osseuse, Tb.Th, Tb.Sp, Tb.N ainsi que la dimension fractale ont été évalués par des méthodes
bidimensionnelles. La fraction osseuse, Tb.Sp et Tb.N sont apparus pertinents pour
discriminer les cas d’ostéoporose chez les patientes ménopausées.
Dans une étude d’images de radius (résolution de 137×137×500 µm3) obtenues sur deux
groupes de patientes (12 femmes en bonne santé et 8 femmes ostéoporotiques), Wehrli et al.
([Wehrli, 1998]) ont calculé, sur des ROI définies manuellement, une série de paramètres
basés sur la fonction d’autocorrélation spatiale. Cette famille de paramètres (que nous avons
abordés dans la partie II.2.4) ne nécessitant pas de segmentation préalable est particulièrement
adaptée pour le cas d’acquisitions en limite de résolution. A l’aide d’une fonction simple
reliant certains de ces paramètres, ils ont pu prédire la « déformation vertébrale » qui est un
indice utilisé pour définir une fracture vertébrale.
A l’aide d’images de calcanéum (résolution dans le plan de 195×195 µm2 pour une épaisseur
de coupe de 500 µm), deux groupes de patients (23 présentant au moins une fracture
ostéoporotique, comparés à 27 cas « sains ») ont été étudiés ([Link, 1998 (b)]). Des
paramètres histomorphométriques ont été calculés en 2D sur des ROI définies manuellement
et segmentées par la méthode de seuillage évoquée précédemment. Des différences
significatives ont pu être observées pour ces paramètres entre les deux groupes.
Une étude similaire ([Majumdar, 1999]) basée sur des images de radius (résolution dans le
plan de 156×156 µm2 pour une épaisseur de coupe de 500 µm) a aussi montré que la
combinaison des paramètres microarchitecturaux avec le contenu minéral osseux mesuré sur
la hanche permettait d’améliorer la distinction, sur des femmes ménopausées, d’un groupe
présentant une fracture du fémur (20 patientes) d’un autre de « contrôle » (19 patientes).
A l’aide d’attributs topologiques calculés sur des ROI positionnées manuellement sur des
images de radius (résolution de 137×137×500 µm3, segmentées par une méthode de réduction
du volume partiel), une discrimination significative a pu être obtenue entre un groupe de 29
femmes présentant une fracture vertébrale d’un autre de contrôle, composé de 50 femmes
([Wehrli, 2001]).
176

Chapitre IV: Imagerie 3D in vivo du doigt humain
Afin d’observer les effets sur le tissu trabéculaire des traitements médicamenteux nécessaires
à la suite d’une transplantation de rein, Link et al. ([Link, 2002]) ont étudié des images de
calcanéum (195×195 µm2 pour une épaisseur de coupe de 1 mm). La segmentation par
seuillage évoquée précédemment ([Majumdar, 1996], cf. II.2.3.1) a été appliquée sur des ROI
positionnées manuellement (comme le plus grand cercle inclus dans le calcanéum). Des
paramètres d’histomorphométrie ont ensuite été évalués. Aucune différence n’a pu être
observée entre un groupe de patients n’ayant pas encore subi la transplantation d’un autre
ayant déjà été opérés. Par contre, une distinction des cas de fractures a pu être obtenue grâce
aux paramètres de microstructure.
Le pouvoir discriminant des paramètres d’histomorphométrie évalués par la méthode directe
(cf. II.5.2.2) a aussi été étudié ([Laib, 2002]) à partir d’images de radius (résolution de
156×156 µm2 pour une épaisseur de coupe de 500 µm) obtenues sur des femmes présentant au
moins une fracture vertébrale par rapport à un groupe de référence.
Des images IRM de radius (similaires à celles utilisées dans le cadre de l’étude précédente)
ont aussi été employées ([Newitt, 2002]) pour effectuer une analyse par éléments finis afin de
déterminer certaines caractéristiques mécaniques du tissu (modules élastiques). En utilisant
ces données ainsi que celles provenant de certains paramètres de structure, il a été possible de
distinguer pour des femmes « post ménopause », un groupe de patientes « saines » d’un autre
ostéoporotiques.
IV.1.2. Quantification des modifications de l’architecture avec
l’âge
Les travaux sur les effets de l’âge sur les caractéristiques du tissu trabéculaire sont moins
nombreux.
Gordon et al. ([Gordon, 1997]) ont étudié les évolutions d’un indice de connectivité et d’une
mesure de l’espacement des travées sur des patients sains âgés de 24 à 65 ans. A partir
d’images de radius (12 coupes d’épaisseur 800 µm avec une résolution dans le plan de
195×195 µm2), ils ont défini automatiquement une ROI sur chaque coupe à partir d’un
algorithme de détection de contour (interface cortical/trabéculaire) qu’ils ont ensuite
177

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
segmentée (par seuillage arbitraire: le seuil étant fixé à 50% de l’amplitude maximum
mesurée dans l’image). Pour chaque image, les paramètres ont été évalués en utilisant une
méthode 2D puis en calculant leur valeur moyenne sur les 6 coupes centrales. Ils ont pu
observer des variations significatives en fonction de l’âge: décroissance de la connectivité et
croissance de l’espacement « inter travées ».
A partir d’images de calcanéum acquises sur 23 femmes en bonne santé âgées de 23 à 74 ans
(28 coupes d’épaisseur 1 mm avec une résolution dans le plan de 195×195 µm2), Ouyang et
al. ([Ouyang, 1997]) ont évalué en 2D des paramètres histomorphométriques (fraction osseuse
et deux indices traduisant l’épaisseur et l’espacement des travées, chacun étant calculé à partir
de leur valeur moyenne sur les 5 coupes centrales). Les ROI (correspondant au plus grand
cercle inclus dans chaque calcanéum) ont été segmentées par seuillage (le seuil étant
déterminé par une méthode similaire à celle évoquée précédemment: [Majumdar, 1996], cf.
II.2.3.1). Des corrélations significatives de ces paramètres avec l’âge ont ainsi pu être
observées et un taux annuel de changement a été proposé pour chacun d’entre eux: perte de
0.52% et de 0.33% respectivement pour la fraction osseuse et l’épaisseur des travées et
accroissement de 0.59% pour l’espacement entre travées.
IV.1.3. Suivi de l’efficacité d’un traitement
Plus récemment ([Van Rietbergen, 2002]), l’IRM a aussi été utilisée dans le cadre d’une étude
longitudinale pour suivre les changements des propriétés mécaniques du tissu trabéculaire
obtenus par un traitement de l’ostéoporose (idoxifène). Pour 3 groupes de femmes (18
femmes ayant reçu un placebo, 23 et 15 femmes traitées respectivement avec 5 mg et 10 mg
par jour d’idoxifène pendant un an), des images de calcanéum ont été acquises avec une
résolution de 195×195×500 µm3, et segmentées par seuillage (le seuil étant déterminé par la
méthode évoquée précédemment: [Majumdar, 1996], cf. II.2.3.1). Sur une ROI positionnée
manuellement, un modèle à éléments finis a été développé afin d’évaluer certains modules
élastiques. Aucune différence significative n’a pu être observée entre les groupes pour la
fraction osseuse. Par contre, pour les groupes traités, les caractéristiques mécaniques ont
considérablement évolué. Cette technique de suivi a donc permis d’établir l’efficacité du
traitement qui ne serait pas apparu si on n’avait analysé que la fraction osseuse.
178

Chapitre IV: Imagerie 3D in vivo du doigt humain
IV.1.4. Limitations de ces études
Les limitations principales des études in vivo qui ont été menées sont:
• la résolution des images utilisées. Souvent proche de la taille caractéristique des travées,
elle rend l’étape de segmentation très problématique. De plus, elle est systématiquement
anisotrope du fait que l’épaisseur de coupe choisie est plus importante que la résolution
dans la coupe. Ceci peut cependant être justifié si la structure est relativement anisotrope
dans cette direction.
• la délimitation de la ROI: du fait des formes complexes que peuvent prendre les zones de
tissu trabéculaire et de la richesse des images in vivo (nombreux autres tissus représentés),
elle est en général obtenue manuellement. Les rares méthodes de délimitation automatique
proposées s’appuyaient sur des particularités anatomiques des images facilitant la
recherche de ROI (tissu cortical très épais par exemple).
• l’évaluation des paramètres de structure est souvent effectuée par des méthodes
bidimensionnelles, du fait de l’anisotropie importante de la résolution (épaisseur de coupe
4 à 5 fois plus élevée que la résolution dans la coupe) due à la sensibilité limite de l’IRM.
Cette approche semble acceptable pour des sites anatomiques sur lesquels les directions
d’orientation principale des travées est bien identifiée (par exemple, la tête du radius).
Mais elle est plus discutable sur le calcanéum (indice d’anisotropie plus faible,
orientations variables des travées)
IV.1.5. Problématique et présentation du travail
Dans ce chapitre, nous avons cherché à établir la faisabilité de l’analyse du tissu trabéculaire
in vivo par IRM à haute résolution. A partir d’images obtenues dans des conditions
d’imagerie proches de celles de l’étude in vitro présentée au chapitre III, nous avons mis en
oeuvre une méthode automatique de récupération de la ROI robuste et employé des
techniques de segmentation fiables du tissu trabéculaire. Les paramètres étudiés au chapitre II
ont donc pu être calculés selon le même protocole qu’au chapitre III.
Les images ont été acquises sur volontaires sains avec un imageur IRM à haut champ (7 T)
« petit animal » (imageur caractérisé par un tunnel d’accès limitant les champs de vue
généralement à l’étude de petits animaux comme la souris). Ceci a imposé le site anatomique
analysé: la phalange distale du majeur. Il n’en reste pas moins un site d’intérêt. Beaucoup de
179

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
recherches sont actuellement menées pour faciliter le suivi des femmes ménopausées. Dans ce
cadre, des dispositifs permettant de mesurer la densité minérale osseuse de la phalange ont été
développés: ce site très facile d’accès permet en effet d’utiliser des doses ionisantes
négligeables ([Michaeli, 1998]). De plus, de nombreux travaux ont déjà été menés sur ce site
([Bouxsein, 1997], [Njeh, 1999], [Drake, 2002]). Mulder et al. ([Mulder, 2000]) ont même
observé des corrélations significatives entre le contenu minéral osseux mesuré sur la phalange
et sur des sites comme la hanche ou la vertèbre, plus classiques pour détecter un état
ostéoporotique.
L’objectif de cette partie (étude de faisabilité) n’est qu’une première étape du développement
d’un protocole basé sur un imageur disponible en routine clinique (imageur « corps entier »)
qui permettra l’analyse de sites anatomiques beaucoup plus nombreux.
Ce travail a été mené dans le cadre d’une collaboration avec l’Université de Würzburg
(Allemagne) soutenue par le programme d’échanges entre laboratoires français et étrangers
proposé par Egide pour le compte du Ministère des affaires étrangères (programme
« Procope »).
IV.2. Les images
IV.2.1. Protocole expérimental
Les images ont été acquises lors de manipulations menées à l’Institut de Physique de
l’Université de Würzburg, avec des membres du groupe « NMR » du laboratoire
« Experimentelle Physik V » (EP5). Nous disposions d’un appareil « petit animal » Bruker
Biospec 70/20 à 7 T (300 MHz), avec un tunnel horizontal de 72 mm de diamètre et un
gradient maximal de 196 mT.m-1 (soit 19.6 G.cm-1).
L’articulation entre les phalanges moyenne et distale du majeur a été placée au centre d’une
sonde en « cage d’oiseau » de 24 mm de diamètre et de 20 mm de long. L’axe principal du
doigt, parallèle au champ statique, a été choisi comme axe de lecture, le plan perpendiculaire
(coupe axiale) étant associé aux deux axes de codage de la phase.
La zone imagée est représentée en Figure IV.1 sur une radiographie X de la main.
180

Chapitre IV: Imagerie 3D in vivo du doigt humain
181
Carpe
Métacarpes
Phalanges proximales
Phalanges moyennes
Phalanges distales
Zone imagée
Figure IV.1: Radiographie X d’une main humaine: vue de dessus (image du site web e-hand.com). Sur
chaque main, il y a 5 métacarpes et 14 phalanges: 5 proximales, 4 moyennes (il n’y en a pas pour le pouce)
et 5 distales. La zone imagée est indiquée par le carré blanc: elle correspond à l’articulation entre les
phalanges moyenne et distale du majeur.
IV.2.2. Paramètres d’acquisition
Outre les optimisations du RSB et de la résolution, on a aussi cherché à minimiser le temps
d’acquisition: en effet, pour une application clinique, le temps passé dans l’imageur doit être
le plus court possible pour ne pas gêner le patient. Ceci nous a imposé le choix d’une
séquence plus rapide que dans le cadre de l’étude in vitro: « écho de gradient 3D » (cf. I.6.3.4)
avec un temps de répétition TR court. Les paramètres de la séquence d’acquisition ont été
fixés ainsi:
• TE/TR = 1/2.6 ms
• une bande passante de 200 kHz
• un angle de basculement optimisé pour obtenir un signal maximum de la moelle
• 16 accumulations
• une matrice d’acquisition de 128×128×128 (taille maximale accessible sur le dispositif
utilisé): pour assurer un voxel isotrope, on a donc utilisé un champ de vue cubique de
20 mm de côté (soit un volume total analysé de 8 cm3).
Les images ont donc été acquises avec une résolution isotrope de 156 µm (soit un volume de
voxel de 3.8×106 µm3) pour un temps d’acquisition de 12 minutes environ.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
182
IV.2.3. Calcul des images
De même que pour l’étude in vitro, une interpolation des données a ensuite été effectuée par
la méthode du « zerofill » d’un facteur 2 dans chaque dimension (cf. III.3.1.3), conduisant à
une matrice 256×256×256. Après le calcul de la transformée de Fourier, la taille de voxel
obtenue est donc de 78 µm isotrope (soit un volume de 0.5×106 µm3) avec un RSB d’environ
10 (le signal et le bruit étant respectivement analysés à partir de voxels situés dans la moelle
et dans le tissu cortical).
Les caractéristiques de ces images sont donc proches de celles des acquisitions in vitro (cf.
III.3.1). La Figure IV.2 montre trois coupes principales d’une image 3D dont l’histogramme
est donné en Figure IV.3.
x
z
Phalange moyenne
Graisse
Phalange distale
20 mm
y
x
Tissu trabéculaire
Tissu cortical
y
z
Phalangedistale
Vaisseauxsanguins
Repliement
Tendons Peau
A B C Figure IV.2: Trois coupes principales d’une image 3D obtenue in vivo par IRM: sagittale (A), coronale (B)
et axiale (C). Les axes x, y et z (256 voxels chacun) sont respectivement l’axe de lecture et les deux axes de
codage de phase. Chaque coupe est carrée (20 mm de coté). On indique sur ces images certains tissus
identifiables ainsi que l’effet du repliement (cf. IV.2.4.1).

Chapitre IV: Imagerie 3D in vivo du doigt humain
183
NG
Occurrence normalisée
0,006
0,008
0,01
0,012
0,014
0,016
0
0,002
0,004
0 50 100 150 200 250
Voxels « Bruit »
Voxels « Volume partiel »
Figure IV.3: Histogramme normalisé de l’image présentée en Figure IV.2. Deux différences majeures
peuvent être notées entre cet histogramme et celui obtenu in vitro (cf. Figure III.3). D’une part, on observe
une population de voxels caractérisés par des niveaux de gris intermédiaires dus à la présence de tissus
mous et de volume partiel (dans la zone trabéculaire). D’autre part, il est très difficile de distinguer la
population des voxels de signal (moelle) sur l’histogra me. m
IV.2.4. Premières analyses
IV.2.4.1. Sur les images
Comme on peut le voir sur la Figure IV.2, de nombreux tissus (autres que trabéculaire)
peuvent être distingués sur ces images: tendons, peau, vaisseaux sanguins, cartilage… On
peut aussi observer l’évolution du type de tissu osseux (cf. I.1). Eloigné de l’articulation
(diaphyse de la phalange), l’os est formé d’une sorte de « cylindre creux » épais empli de
moelle osseuse: le tissu cortical. Plus on se rapproche de l’articulation, plus le tissu cortical
s’amincit et plus les travées osseuses apparaissent dans la moelle. Au niveau de l’articulation
(épiphyse de la phalange), l’os n’est plus formé que de tissu trabéculaire. On peut remarquer
qu’il n’y a pas de séparation précise entre les tissus cortical et trabéculaire (apparition
progressive du tissu trabéculaire au fur et à mesure que l’on se rapproche de l’articulation).
Du fait que le champ de vue a été choisi légèrement inférieur au diamètre de la sonde (cf.
IV.2.1 et IV.2.2), un artefact de repliement apparaît selon les deux axes de codage de phase
(on peut l’observer assez nettement sur la Figure IV.2 C). Celui-ci ne perturbe cependant
l’image qu’au niveau des tissus externes à l’os (qui est situé au centre du doigt donc de la
sonde, cf. Figure IV.2, C). Ce repliement « contrôlé » permet donc d’améliorer la résolution
sans affecter la zone que l’on cherche à analyser.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
La dégradation de la sensibilité de la sonde le long de son axe est perceptible sur les coupes
axiales situées aux extrémités de l’axe de lecture: on peut l’observer aux limites du champ de
vue sur les coupes sagittale et coronale présentées sur la Figure IV.2 (A et B). Comme pour
l’étude in vitro (cf. III.3.1.4 et III.5.1), cette atténuation du signal devra être prise en compte
lors de l’analyse des images.
IV.2.4.2. Sur l’histogramme
A la différence de l’histogramme « classique » des images acquises in vitro (cf. Figure III.3),
il y a ici trois populations de voxels (cf. Figure IV.3). Le tissu cortical, les travées épaisses
(par rapport à la résolution spatiale) et les tendons apparaissent sur l’image en bas niveau de
gris, la moelle et la graisse sous la peau en haut niveau de gris. Les autres tissus mous, les
travées fines (voxels de la zone trabéculaire contenant des proportions semblables de tissu
calcifié et de moelle, i.e. le volume partiel) mais aussi la moelle située dans les coupes axiales
en extrémité de sonde sont caractérisés par des voxels de niveaux de gris intermédiaires. Du
fait de la faible proportion de voxels de niveaux de gris élevés dans l’image, cette population
ne peut être distinguée.
IV.2.4.3. Phalange distale
L’image entière est donc difficilement analysable directement: il faut délimiter, dans un
premier temps, une zone centrée sur les tissus osseux. L’étude s’est principalement axée sur la
phalange distale pour deux raisons: outre la bonne qualité du signal sur cette zone (fort RSB
et bonne homogénéité de la sonde), c’est là que se trouve le plus grand volume de tissu
trabéculaire disponible. La zone correspondant à la phalange distale a donc été extraite de
l’image globale (cf. Figure IV.2) par rognage. La Figure IV.4 présente trois plans principaux
de cette phalange ainsi que les dimensions du volume extrait.
184

Chapitre IV: Imagerie 3D in vivo du doigt humain
185
11.7 mm
8.7 mm
14 mm
8.7 mm 14 mm
11.7 mm
A B C
Figure IV.4: Trois coupes principales correspondant à la phalange distale extraite par rognage de l’image
présentée en Figure IV.2 (A: sagittale, B: coronale, C: axiale). Les dimensions du volume récupéré sont
indiqués sur chaque coupe.
Pour avoir un ordre d’idée du volume de tissu trabéculaire disponible, on peut considérer, en
première approximation, cette phalange comme un tronc de cône avec les caractéristiques
suivantes: un grand et petit diamètres d’environ 10.6 mm et 3 mm et une hauteur d’environ
9.7 mm. Le volume d’intérêt accessible dans cette phalange est donc d’environ 390 mm3.
Sur l’histogramme de ce volume (présenté en Figure IV.5), le mode correspondant aux voxels
de moelle apparaît mais il reste difficile à distinguer de la population des voxels de niveau de
gris intermédiaire. Le mode correspondant au bruit se distingue très nettement.
0
0,002
0,004
0,006
0,008
0,01
0 50 100 150 200 250
Occurrence normalisée
NG
Voxels « Bruit »
Voxels « Volume partiel »
Voxels « Signal »
Figure IV.5: Histogramme de l’image présentée en Figure IV.4. Le mode correspondant au voxels de
moelle (signal) reste difficile à distinguer des autres modes.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
IV.3. Délimitation de la ROI
IV.3.1. Le problème
Le but de l’étude est d’obtenir une caractérisation de la microstructure du tissu trabéculaire.
Afin de pouvoir calculer les paramètres présentés au chapitre II, il faut donc dans un premier
temps, isoler la zone trabéculaire de l’image.
Pour l’étude in vitro, le volume à analyser était relativement simple à isoler puisque sa forme
était parfaitement connue (échantillon cylindre, cf. III.2) et qu’il n’était entouré que de bruit
(fond de l’image « uniforme », cf. III.3.1.4) dont les caractéristiques étaient facilement
accessibles. Ceci facilitait donc considérablement cette étape (cf. III.4.2.1).
Pour les études in vivo, la délimitation de la ROI est beaucoup plus problématique (comme on
peut le voir sur la Figure IV.4). D’une part, la forme de la zone trabéculaire est très complexe
(passage progressif du tissu cortical au tissu trabéculaire au niveau de l’articulation, cf.
IV.2.4.1) et elle est propre à chaque individu: il ne semble donc pas y avoir de modèle simple
que l’on pourrait généraliser. De plus, de nombreux tissus (autres que trabéculaire) sont aussi
représentés sur l’image avec des voxels dont les niveaux de gris se répartissent sur tout
l’histogramme.
IV.3.2. Première approche
L’approche la plus intuitive consiste à « rogner » l’image pour ne garder que des voxels situés
dans la zone trabéculaire (de la même manière que dans la partie III.7). On obtient alors une
ROI « en pavé » dont trois coupes sont montrées sur la Figure IV.6.
186

Chapitre IV: Imagerie 3D in vivo du doigt humain
187
2 m
m
4.3 mm
4.3 mm
5.5
mm
2 m
m
5.5 mm
A B C
Figure IV.6: Exemple de ROI « en pavé » ne contenant que du tissu trabéculaire que l’on peut définir par
« rognage » sur l’image présentée en Figure IV.4. Cette ROI est matérialisée par un rectangle noir sur les
coupes sagittale (A), coronale (B) et axiale (C). Le volume analysable est d’environ 47 mm3.
La zone que l’on peut récupérer de cette manière est d’environ 47 mm3 soit seulement 12% du
volume de tissu trabéculaire disponible estimé (cf. IV.2.4.3). De plus, elle serait encore
fortement réduite pour le calcul de certains paramètres qui nécessitent une restriction
préalable de la ROI à la plus grande sphère incluse (cf. II.5 et II.6). Le volume de cette ROI
est donc peu représentatif.
Elle est d’ailleurs difficile à positionner. Pour cet exemple, celle-ci a été ajustée
« visuellement » mais dans l’optique d’une analyse objective des images (par exemple pour la
comparaison d’examens faits à différents moments ou sur différents patients), il faut prévoir
le minimum d’interventions de l’observateur ce qui nécessite une automatisation du procédé.
Compte tenu des niveaux de gris des autres tissus présents dans l’image, il semble très
difficile de définir des critères pour automatiser ce rognage.
La forme parallélépipédique ne semble donc pas adaptée à ce problème.
IV.3.3. Détermination d’une ROI adaptée par ajustement d’une
surface tridimensionnelle
On cherche à déterminer une forme adaptée à la zone trabéculaire basée sur l’ajustement
d’une surface tridimensionnelle fermée permettant de séparer la zone d’intérêt des autres
tissus présents dans l’image. Nous avons distingué trois étapes:
• détermination des voxels situés sur cette surface

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
• ajustement d’une surface tridimensionnelle fermée à partir de ces voxels
• extraction de l’intérieur de la surface
Comme on l’a vu dans la partie IV.2.4.1, la définition de la zone trabéculaire est assez vague.
Le but n’est donc pas d’en déterminer précisément la frontière mais de pouvoir récupérer le
plus grand volume ne contenant que du tissu trabéculaire. En effet, si d’autres tissus sont
présents dans la zone à analyser, ils risquent d’affecter l’évaluation des paramètres de
microstructure: par exemple, si une partie importante de tissu cortical est incluse dans la ROI,
la fraction osseuse sera surestimée, l’anisotropie de la microstructure moins prononcée,…
Nous détaillons dans la suite, chacune de ces trois étapes.
IV.3.3.1. Récupération de voxels « frontières »
Afin de pouvoir ajuster une surface tridimensionnelle enveloppant « au mieux » la zone
d’intérêt, on doit, dans un premier temps, déterminer des voxels de la phase « trabéculaire »
(i.e. tissu calcifié ou moelle osseuse) qui sont situés « au plus près » de la séparation entre la
zone trabéculaire et les autres tissus présents dans l’image. Il faut donc trouver un critère
permettant de détecter la « frontière » du tissu trabéculaire, sachant que, comme on l’a vu
dans la partie IV.2.4.2, les voxels de cette zone sont caractérisés par des niveaux de gris
élevés (moelle), intermédiaires (travées fines: volume partiel), et bas (travées épaisses).
IV.3.3.1.1. Analyse des bas niveaux de gris
Une première méthode consisterait à associer cette séparation à la surface interne du tissu
cortical. Pour illustrer cette approche, nous avons segmenté le volume (présenté en Figure
IV.4) par seuillage pour un seuil choisi juste au-dessus du mode de l’histogramme
correspondant au bruit (nous avons choisi un niveau de gris de 30, cf. Figure IV.5). Cette
segmentation nous a donc permis d’isoler du reste de l’image, les voxels du tissu cortical, des
travées épaisses et des tendons (cf. IV.2.4.2). La Figure IV.7 montre le résultat de cette
segmentation.
L’analyse des amas (cf. II.4.5.1 et II.4.6.3) montre que le tissu cortical et les travées épaisses
(relativement nombreuses) sont connectés. De plus, comme les tendons et le tissu cortical sont
très proches au niveau des coupes axiales situées au bord de l’image, il y a un risque qu’une
connexion entre ces deux amas apparaisse (c’est d’ailleurs le cas si un nombre de coupes
axiales plus important est conservé lors de la récupération de la phalange distale, cf. IV.2.4.3).
188

Chapitre IV: Imag erie 3D in vivo du doigt humain
189
Enfin, la zone trabéculaire est solidaire du reste de l’image (i.e. « l’intérieur » et
« l’extérieur » du tissu cortical ne forment qu’un seul amas connexe).
A B C
Zone trabéculaire
Tendons Tendons
Tissu cortical
D E
Figure IV.7: Segmentation par seuillage pour un seuil permettant d’isoler les voxels de bruit qui sont
représentés en noir sur les coupes sagittale (A), coronale (B) et axiale (C). On donne aussi deux vues (D:
depuis l’articulation, E: depuis l’extrémité du doigt) de la représentation tridimensionnelle des amas
principaux de ces voxels (chaque amas est associé à un niveau de gris). On peut remarquer que les travées
épaisses sont connectées au tissu cortical et que les tendons sont très proches de ce tissu osseux sur les
coupes axiales situées au bord de l’image.
Il semble donc relativement délicat de déterminer des points « frontières » par cette approche
principalement pour deux raisons. Premièrement, il serait très difficile de différencier les
voxels du tissu cortical, des travées épaisses et des tendons. Deuxièmement, au niveau de
l’articulation, le tissu cortical est de moins en moins présent: le tissu trabéculaire se trouve
alors directement en contact avec des tissus externes à la structure osseuse qui sont
caractérisés sur l’image par des niveaux de gris intermédiaires. Il y a donc une « connexion »

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
entre les voxels qui caractérisent une partie de la zone trabéculaire (volume partiel) et ceux
des tissus externes.
IV.3.3.1.2. Analyse des hauts niveaux de gris
La deuxième technique consiste à associer la « frontière » du tissu trabéculaire à la surface
englobant la moelle osseuse. En effet, comme on l’a vu dans la partie IV.2.4.2, seules la
moelle osseuse (confinée à l’intérieur de l’os) et la graisse (sous la peau) apparaissent en hauts
niveaux de gris dans l’image. Or ces zones sont clairement séparées et forment des amas
déconnectés de voxels de niveaux de gris élevés. Comme l’amas caractérisant la moelle a un
volume bien supérieur à ceux correspondant à la graisse sous la peau, il est très facilement
détectable.
On cherche donc à déterminer un seuil permettant d’isoler les voxels de hauts niveaux de gris.
Pour une image IRM acquise sans volume partiel, l’histogramme peut être représenté en
première approximation, par la somme d’une distribution de Rayleigh et d’une distribution
gaussienne, respectivement pour les modes associés aux voxels de bruit et de signal (cf.
III.3.1.4 et III.5.1). En modélisant l’occurrence des voxels de niveaux de gris intermédiaires
par une gaussienne, l’histogramme présenté sur la Figure IV.5 peut donc, en première
approximation, se mettre sous la forme de la somme d’une distribution de Rayleigh
d’amplitude AR et d’écart type σR (pour les voxels de bruit) et de deux distributions
gaussiennes (la première pour les voxels caractérisés par du volume partiel, la seconde pour
ceux de signal) d’amplitude AG1 (resp. AG2), de moyenne µG1, (resp. µG2) et d’écart type σG1
(resp. σG2):
)2)(
exp(2
)2)(
exp(2
)2exp()( 22
22
2
22
1
21
1
12
2
2G
G
G
G
G
G
G
G
RR
R xAxAxxAxh σ
µσπσ
µσπσσ
−−+
−−+
−= Eq. IV.1
où h est le modèle de l’histogramme réel et x le niveau de gris (x∈0,…,255).
On peut admettre que µG2 serait un seuil acceptable pour isoler les voxels de hauts niveaux de
gris dans l’image. Cependant on ne peut estimer µG2 séparément des sept autres paramètres.
La méthode des moindres carrés ([Press, 1989]) a donc été utilisée pour ajuster la fonction h
aux données expérimentales. La Figure IV.8 montre le résultat de l’ajustement trouvé sur
l’histogramme présenté en Figure IV.5.
190

Chapitre IV: Imagerie 3D in vivo du doigt humain
191
0,004
0,006
0,008
0,01
0
0,002
0 50 100 150 200 250
NG
Occurrence normalisée
Figure IV.8: Ajustement (trait gris) de l’histogramme présenté en Figure IV.5 (en trait noir). Pour
information, les valeurs trouvées pour les huit paramètres de h sont: AR = 0.21, σR = 13.8 niveaux de gris,
AG1 = 0.54, µG1 = 74.1 niveaux de gris, σG1 = 23.8 niveaux de gris, AG2 = 0.25, µG2 = 142.1 niveaux de gris,
σG2 = 33.8 niveaux de gris.
Après avoir effectué le seuillage de l’image présentée en Figure IV.4 (pour un seuil égal à
µG2), on filtre les amas secondaires des voxels de hauts niveaux de gris (cf. II.4.5.1 et
II.4.6.3). L’amas principal correspond alors directement à l’amas représentant la moelle
osseuse. Celui-ci est représenté sur la Figure IV.9.

Caractérisation de l’architecture tridimension laire panelle du tissu trabécu r IRM
192
A B C
D E
Figure IV.9: Amas de moelle osseuse (en blanc) récupéré après seuillage à µG2 de l’image présentée en
Figure IV.4 suivi du filtrage des amas secondaires (A: sagittale, B: coronale, C: axiale). On donne deux
vues (D: depuis l’articulation, E: depuis l’extrémité du doigt) de la représentation tridimensionnelle de cet
amas.
IV.3.3.1.3. Voxels « frontières »
Les voxels « frontière » sont alors déterminés comme les intersections de cet amas avec trois
faisceaux de droites dans les trois directions principales (axes x, y et z représentés sur la
Figure IV.2). Afin d’en limiter le nombre (et donc de réduire le temps de calcul en particulier
pour l’ajustement de la surface, cf. IV.3.3.2), l’espacement des droites dans les directions y et
z est arbitrairement fixé à 5 voxels (soit 390 µm).
Ainsi sur l’amas de moelle de la phalange distale présentée en Figure IV.9, on a déterminé
9899 voxels « frontière » représentés sur la Figure IV.10.

Chapitre IV: Imagerie 3D in vivo du doigt humain
193
A B
Figure IV.10: Représentation tridimensionnelle des voxels « frontière » trouvés (A: vue de l’articulation,
B: vue de l’extrémité du doigt).
Remarques:
a) En considérant uniquement les voxels de niveaux de gris supérieurs à µG2 (cf. Eq. IV.1)
comme représentatifs de la moelle, on surestime considérablement la phase osseuse du fait du
nombre important de voxels de volume partiel dans l’image (cf. histogramme de la Figure
IV.5). Ceci a deux conséquences.
• La segmentation obtenue (cf. Figure IV.9) ne pourrait être utilisée pour effectuer une
caractérisation correcte du tissu trabéculaire. Ce n’est cependant pas l’objectif à ce stade:
ici, la segmentation sert à obtenir une approximation de la zone trabéculaire et non à
caractériser précisément la microstructure (elle est donc beaucoup moins cruciale). Pour le
calcul des paramètres du chapitre II, les algorithmes que nous avons envisagés pour la
segmentation précise des images seront décrits dans la partie IV.4.
• Le volume de moelle ainsi déterminé est sous estimé. En effet, certains voxels ayant un
niveau de gris inférieur à µG2 appartiennent à cette phase et ne sont pas caractérisés
comme tel après segmentation (par exemple, des voxels de moelle appartenant aux coupes
axiales situées à l’extrémité de la sonde). Ce critère permet donc de s’assurer que le
contour de l’amas de moelle sera pleinement situé dans le tissu trabéculaire.
b) Du fait de la méthode employée pour déterminer les voxels « frontière », certains d’entre
eux se trouvent au niveau de travées épaisses donc dans la zone trabéculaire et non à sa
frontière. La surface à ajuster ne devra donc pas passer par tous les voxels « frontière » mais
plutôt les « envelopper ».

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
194
IV.3.3.2. Calcul de l’enveloppe tridimensionnelle
Les concepts associés à la reconstruction de surface tridimensionnelle à partir d’un nuage de
points sont relativement complexes. Leur présentation n’étant pas l’objectif de ce travail, nous
ne développerons que très succinctement les différentes méthodes envisagées: pour de plus
amples informations, nous renvoyons à des références bibliographiques.
En général, ces surfaces sont définies par la réunion de polygones élémentaires, des triangles,
chacun d’entre eux étant déterminé par trois sommets appartenant à l’ensemble des N points
Pi (i de 1 à N) du nuage. Le calcul de base est la triangulation de Delaunay dont une
présentation très claire est donnée dans le chapitre 5 du livre « Computational Geometry in
C » de O’Rourke ([O’Rourke, 2001]).
IV.3.3.2.1. Enveloppe convexe
Dans un premier temps, nous avons envisagé le calcul de l’enveloppe convexe (« convex
hull ») associée au nuage de points Pi. Pour information, on rappelle qu’un ensemble est dit
« convexe » si, pour tout couple de points de cet ensemble, le segment reliant ces deux points
est entièrement inclus dans l’ensemble (cf. Figure IV.11).
A B
A B
A B
Figure IV.11: Exemple d’un ensemble convexe (A) et d’un ensemble non convexe (B).
L’enveloppe convexe est définie comme le plus petit ensemble convexe contenant tout le
nuage de points Pi. De nombreux algorithmes sont disponibles pour ce type de calcul. Nous
avons utilisé celui proposé dans le chapitre 4 du livre cité précédemment ([O’Rourke, 2001]).
Cette approche est cependant apparue insuffisante car la zone à isoler n’est pas convexe a
priori. En utilisant cette méthode, on risquerait donc d’inclure dans la ROI d’autres tissus que
le tissu trabéculaire ou alors il faudrait effectuer un rognage préalable de la phalange pour
n’en garder qu’une partie convexe (ce qui ferait perdre un volume non négligeable de la zone
d’intérêt disponible).

Chapitre IV: Imagerie 3D in vivo du doigt humain
195
IV.3.3.2.2. Solution retenue: α-forme
La solution retenue est basée sur le calcul de l’α-forme (« α-shape ») associée au nuage de
points. Cette technique, développée en 1994 par Edelsbrunner et Mücke
([Edelsbrunner, 1994]), est employée dans de nombreux problèmes de ce type.
Dans un espace de dimension 3, l’α-forme (α > 0) d’un nuage de points Pi est définie
comme l’ensemble des triangles (chacun défini par un triplet de points appartenant à Pi)
pour lesquels il existe une boule de rayon α passant par les trois sommets de chaque triangle
et dont l’intérieur ne contient aucun point Pi.
Pour illustrer la définition de l’α-forme et le rôle du paramètre α, on donne ci-dessous
plusieurs exemples en dimension 2. La formulation de l’α-forme dans cet espace devient
l’ensemble des segments (chacun défini par un couple de points appartenant à Pi) pour
lesquels il existe un disque de rayon α passant par les deux sommets de chaque segment et
dont l’intérieur ne contient aucun point Pi.
La Figure IV.12 montre les disques permettant de définir les segments formant l’α-forme
associée à un nuage de points délimitant un « ruban ». Sur cet exemple, on peut se rendre
compte intuitivement que si α est trop grand, la paroi intérieure du « ruban » ne pourra être
récupérée puisque aucun cercle respectant la définition précédente ne pourra être trouvé pour
les points situés à ce niveau.
Figure IV.12: Illustration (en dimension 2) du calcul d’une α-forme pour un nuage de points délimitant un
« ruban ». Sur cette figure sont représentés l’ensemble des disques définissant les segments composant
l’α-forme (disque de rayon α, ne contenant aucun autre points du nuage que les deux extrémités du
segment). Si α est trop grand, la « paroi intérieure » du « ruban » ne pourra être récupérée (image: cours
de P. Kumar de l’Université de Stony Brook, New Yo k, USA). r

Caractérisation de l’architecture tridim ire par IRM ensionnelle du tissu trabécula
196
Cet exemple illustre que plus la contrainte sur le rayon est forte (c’est-à-dire plus α est grand)
moins l’α-forme est sensible aux détails. Le paramètre α permet donc de régler le degré de
finesse de la forme résultante: quand α tend vers 0, la surface calculée tend vers l’ensemble
vide, quand α tend vers +∞, la surface tend vers l’enveloppe convexe. Ceci est décrit sur la
Figure IV.13 où sont représentées les α-formes associées à un nuage de points en forme de
« α » pour différentes valeurs de α.
A B C
D E F
Figure IV.13: α-formes associées à un nuage de points en forme de « α » (A) pour différentes valeurs
croissantes de α (de B à F). En F, est représenté le cas « α tend vers +∞ », c’est-à-dire l’enveloppe convexe
(images: cours de P. Kumar de l’Université de Stony Brook, New York, USA).
Le choix de α est donc important. S’il est petit, on aura une grande finesse de la
reconstruction mais certains points peuvent se retrouver isolés, entraînant des déconnexions
artificielles de la surface. S’il est grand, on obtiendra une surface fermée mais grossière (on
tend vers l’enveloppe convexe).
L’algorithme proposé par K.L. Clarkson et al. ([Clarkson, 1993]), calcule l’α-forme d’un
nuage de points en estimant préalablement la plus petite valeur du paramètre α telle qu’aucun
point Pi du nuage ne soit isolé (i.e. à l’extérieur de l’α-forme). La surface obtenue est donc
celle présentant le plus haut niveau de détails tout en restant d’un seul tenant.

Chapitre IV: Imagerie 3D in vivo du doigt humain
Cet algorithme est particulièrement bien adapté à notre problème. En effet, la surface à ajuster
ne doit pas passer par tous les voxels « frontière » car certains se trouvent au niveau des
travées épaisses (cf. remarque b) de la partie IV.3.3.1.3): le plus haut niveau de détails ne
serait donc pas souhaitable (outre le problème que poserait une surface qui ne serait pas d’un
seul tenant).
A partir des 9899 voxels « frontière » déterminés sur la région trabéculaire de la phalange
distale étudiée (cf. IV.3.3.1.3), 6524 triangles enveloppant ces points ont ainsi été calculés.
IV.3.3.3. Extraction de la ROI
La dernière étape nécessaire pour sélectionner la zone d’intérêt est de déterminer les voxels
situés à l’intérieur de l’α-forme.
Comme on l’a vu dans la partie précédente (cf. IV.3.3.2.2), la surface est définie par une série
de triangles connectés. Pour pouvoir séparer l’intérieur de l’extérieur de la surface, il suffit
donc de « remplir » la surface délimitée par chaque triangle de façon à ce qu’elle soit
topologiquement d’un seul tenant et « sans trou » c’est-à-dire que :
• les voxels appartenant à la surface de chaque triangle doivent être connectés pour un mode
de connexité donné (nous avons choisi la 26-connexité, cf. II.4.4.2.1).
• la caractéristique d’Euler-Poincaré de chaque surface triangulaire (évaluée selon le même
mode de connexité) doit être égale à 1 (un triangle peut être considéré comme étant
« plein » s’il correspond à un objet de genre nul, cf. II.4.3)
La première étape du remplissage consiste à calculer les trois segments de droite représentant
les trois côtés de chaque triangle. Afin d’assurer la connexion de chaque voxel, la méthode de
tracé proposée par Kaufman ([Kaufman, 1988]) a été utilisée. Celle-ci est basée sur le calcul
de deux projections du segment 3D dans deux plans principaux: le problème revient alors à
celui du tracé de deux segments 2D qui est mené en utilisant l’algorithme de Bresenham
([Bresenham, 1965]). En utilisant cette technique de tracé, on construit les segments voxel par
voxel en respectant la 26-connexité. Plus de détails sont donnés en Annexe 3.
A partir du contour du triangle, on calcule tous les segments de droite reliant un voxel
« sommet » à tous les voxels du côté opposé, en utilisant la même méthode de tracé de
segments 3D. Ceci est illustré sur la Figure IV.14.
197

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
198
Voxel « sommet »
Côté opposé Figure IV.14: Après le tracé des trois cotés, chaque triangle est « rempli » en effectuant le tracé de tous les
segments reliant un voxel « sommet » aux voxels du c té opposé. o
Cette méthode permet d’obtenir une surface triangulaire constituée d’un seul amas connecté
(au sens de la 26-connexité). Cependant, du fait de la nature discrète des images, cela n’assure
pas qu’elle soit « sans trou ». La caractéristique d’Euler-Poincaré est alors calculée pour
déterminer, en 26-connexité, la topologie de la surface obtenue (application de l’algorithme
présenté dans la partie II.4.5.2, sur la phase complémentaire à la surface triangulaire). Tant
que sa valeur n’est pas égale à 1, le remplissage du triangle (présenté sur la Figure IV.14) est
effectué à partir des autres voxels « sommet ».
Le test de cette méthode a cependant montré qu’elle était insuffisante pour finaliser le
remplissage correct de certains triangles. C’est pourquoi on utilise aussi un « filtrage »
morphologique qui consiste en une dilatation suivie d’une érosion ([Gonzalez, 1993]). Cet
opérateur permet de « remplir » des « trous » de petite taille par rapport à celle de l’élément
structurant choisi. Initialement appliqué avec un élément structurant cubique de côté
a = 3 voxels, il est utilisé tant que la caractéristique d’Euler-Poincaré n’est pas égale à 1, en
incrémentant a successivement de 2 voxels.
Cet algorithme nous a permis d’obtenir un triangle « plein » dans tous les cas. Sur la Figure
IV.15, on en rappelle les différentes étapes pour un triangle « test ». On a remarqué que du
fait de la petite taille des triangles qui définissent la surface de la ROI, la première phase (cf.
Figure IV.14) suffisait généralement.

Chapitre IV: Imagerie 3D in vivo du doigt humain
199
A B C
Figure IV.15: Différentes étapes de l’algorithme de remplissage illustrées sur un triangle « test » dont les
sommets sont donnés en A. Le contour est dans un premier temps tracé (B). La condition d’arrêt de la
boucle de remplissage est que la caractéristique d’Euler-Poincaré du triangle soit égale à 1 c’est-à-dire
qu’il ne subsiste aucun « trou » dans la surface pleine (C).
Après application de cet algorithme sur chaque triangle définissant l’α-forme, on obtient donc
une surface d’un seul tenant et sans « trou » au sens de la 26-connexité (cf. Figure IV.16).
A B
Figure IV.16: Représentation tridimensionnelle de la surface trouvée pour la zone trabéculaire de la
phalange distale (A: vue de l’articulation, B: vue de l’extrémité du doigt). Les zones « planes » que l’on
observe en premier plan, correspondent à la restriction de l’image initiale que l’on a effectuée pour
récupérer la phalange distale (cf. Figure IV.4).
L’intérieur et l’extérieur de la surface sont donc associés à deux amas disjoints au sens de la
connexité complémentaire: la 6-connexité (cf. II.4.4.2.1). En utilisant l’algorithme de calcul
de β0 (le nombre de Betti d’ordre 0, cf. II.4.5.1), on a pu extraire l’amas « intérieur »
c’est-à-dire la ROI.

Caractérisation de l’architecture tridi trabéculamensionnelle du tissu ire par IRM
200
Remarque:
D’autres méthodes plus simples ont été envisagées pour récupérer automatiquement la ROI en
particulier en utilisant des opérateurs de morphologie mathématique directement sur l’amas de
moelle (calculé dans la partie IV.3.3.1.2) pour en « remplir » les travées épaisses. Cependant
le choix de la taille de l’élément structurant est très problématique: s’il est trop petit, les
travées épaisses ne sont pas toutes comblées et s’il est trop grand, il modifie sensiblement la
forme de l’amas de moelle.
IV.3.3.4. Résultats et fiabilité de la méthode
IV.3.3.4.1. Région trabéculaire
Nous avons donc pu isoler une ROI située pleinement dans le tissu trabéculaire. La Figure
IV.17 en donne une représentation sur les coupes présentées en Figure IV.4.
A B C Figure IV.17: Représentation de la ROI isolée (en trait blanc) sur les mêmes coupes que celles présentées
en Figure IV.4 (A: sagittale, B: coronale, C: axiale).
Le volume isolé correspond à environ 376 mm3 soit 95% du volume disponible estimé (cf.
IV.2.4.3): cette méthode de détermination de la ROI est donc beaucoup plus efficace que celle
obtenue par rognage (cf. IV.3.2).
Le temps de calcul est d’environ une heure sur une station SUN 400 MHz pour
approximativement 10000 voxels « frontière » analysés. Cependant, si leur nombre n’est pas
limité (comme on l’a fait dans la partie IV.3.3.1.3), la durée du calcul de l’α-forme devient
très vite prohibitive.
Si on peut disposer d’une image où les deux phalanges sont acquises avec un bon RSB, une
analyse globale pourrait alors être envisagée sans avoir à isoler préalablement une phalange

Chapitre IV: Imagerie 3D in vivo du doigt humain
du reste de l’image (cf. IV.2.4.3). En effet, comme notre méthode est basée sur l’analyse de
l’amas de moelle (cf. IV.3.3.1.2), les deux zones trabéculaires pourraient être
automatiquement séparées du reste de l’image puisque la moelle est par définition confinée à
l’intérieur de chaque phalange.
IV.3.3.4.2. Région corticale
En appliquant la même méthode sur les voxels « frontière » du tissu cortical (déterminés à
partir d’un seuillage de l’image présentée en Figure IV.4, pour un seuil égal à σR, cf.
IV.3.3.1.2), la zone enveloppant toute la phalange distale a pu être calculée (8363 voxels
« frontière » conduisant à une α-forme composée de 5763 triangles). En déterminant
l’intersection de cette zone avec le complémentaire de la ROI trabéculaire établie
précédemment, le tissu cortical a pu être extrait du reste de l’image.
La Figure IV.18 montre une représentation sur les coupes présentées en Figure IV.4 ainsi
qu’une visualisation tridimensionnelle.
Un volume de 195 mm3 de tissu cortical a ainsi été isolé. Ce résultat pourrait donc nous
permettre d’étudier aussi le tissu cortical (comme par exemple son épaisseur qui évolue aussi
en cas d’ostéoporose, cf. Figure I.2).
201

Caractérisation de l’architecture tridimension laire panelle du tissu trabécu r IRM
202
A B C
D E
Figure IV.18: Représentation de la ROI « corticale » isolée (en trait blanc) sur les mêmes coupes que celles
présentées en Figure IV.4 (A: sagittale, B: coronale, C: axiale). On donne aussi deux vues (D: depuis
l’articulation, E: depuis l’extrémité du doigt) de la représentation tridimensionnelle de la ROI
« corticale » (en gris) avec la ROI « trabéculaire » (en blanc) déterminée précédemment (cf. Figure IV.16).
IV.3.3.4.3. Fiabilité
Deux autres images (l’une du même patient, l’autre d’un patient différent) de caractéristiques
similaires à celles présentées dans la partie IV.2 ont aussi été testées. Dans les deux cas, nous
avons pu obtenir une délimitation automatique de la ROI. Nous discuterons plus en détail des
résultats obtenus pour l’évaluation des paramètres de structure sur ces ROI dans la partie
IV.5.
Pour ces trois images analysées, l’étape la plus problématique pour la détermination des ROI
s’est avérée être la phase de calcul du seuil pour isoler l’amas de moelle (cf. IV.3.3.1.2). En
effet, pour des images où le mode associé aux niveaux de gris élevés ne se distingue pas
« suffisamment » sur l’histogramme, l’initialisation de l’algorithme des moindres carrés peut
influencer sensiblement le résultat de l’ajustement et donc la valeur trouvée pour le seuil (i.e.

Chapitre IV: Imagerie 3D in vivo du doigt humain
203
µG2). Il faudra donc prévoir une initialisation fiable afin de rendre la méthode la plus
indépendante possible de l’observateur.
Remarque:
La délimitation de la ROI a aussi fonctionné sur les images avant l’interpolation par la
méthode du « zérofill » (c’est-à-dire pour une taille de voxel de 156 µm, cf. IV.2.2).
IV.4. Segmentation des images
Sur la ROI « trabéculaire » déterminée dans la partie précédente (cf. IV.3), on cherche
maintenant à caractériser la phase de chaque voxel (tissu calcifié ou moelle) afin de pouvoir
calculer les paramètres définis au chapitre II.
La Figure IV.19 présente l’histogramme correspondant à la ROI « trabéculaire » déterminée
dans l’étude précédente (cf. IV.3.3.4.1 et Figure IV.17).
Occurrence normalisée
NG 0
0,002
0,004
0,006
0,008
0 50 100 150 200 250 Figure IV.19: Histogramme calculé sur la ROI « trabéculaire » déterminée précédemment (cf. IV.3.3.4.1
et Figure IV.17). Le mode des voxels de bruit est beaucoup moins représenté que sur l’histogramme de la
phalange entière (cf. Figure IV.5).
Malgré l’absence de tout autre tissu au sein de cette ROI, les deux modes « théoriques » de
l’histogramme d’une zone trabéculaire (phase osseuse et moelle) n’apparaissent pas aussi
clairement que dans le cadre de l’étude in vitro (cf. III.3.1.4 et Figure III.3). D’une part, seuls

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
les voxels des travées épaisses sont situés entièrement dans la phase osseuse: le mode
correspondant au bruit est donc beaucoup moins représenté que sur l’histogramme de la
phalange entière (où le tissu cortical et les tendons y contribuaient sensiblement, cf. IV.2.4.2
et Figure IV.5). D’autre part, le volume partiel est plus conséquent que dans le cadre de
l’étude in vitro.
Une segmentation par seuillage simple serait dans ce cas insuffisante. Nous avons donc
envisagé deux solutions que nous présentons dans cette partie. Les conditions d’imagerie étant
proches de celles de l’étude in vitro, nous avons, dans un premier temps, appliqué la méthode
d’étiquetage développée pour la segmentation des images de microscopie IRM (cf. III.5.1).
Dans un deuxième temps, nous avons aussi mis en œuvre une méthode dédiée aux
acquisitions in vivo ayant pour objectif de réduire l’effet du volume partiel (technique
proposée dernièrement par l’équipe de Wehrli, succinctement décrite dans la partie II.2.3.2).
IV.4.1. Caractérisation du bruit de l’image
La phase d’initialisation des deux méthodes de segmentation envisagées est basée sur
l’analyse de l’histogramme de chaque coupe axiale (afin de prendre en compte la dégradation
de la sensibilité de la sonde, cf. IV.2.4.1). Or, sur la ROI, peu de voxels sont situés pleinement
dans le tissu calcifié. Le mode correspondant (voxels de bruit) est donc difficilement
perceptible sur l’histogramme, ce qui pose, dans certains cas, des problèmes de convergence
des algorithmes d’ajustement.
Une caractérisation préalable du bruit a donc été faite pour améliorer l’efficacité des
méthodes. Sur l’image de la phalange distale présentée en Figure IV.4, le modèle h(x) proposé
en IV.3.3.1.2 (cf. Eq. IV.1) a été ajusté sur chaque histogramme des coupes axiales (112 dans
le cas de cette image) par la méthode des moindres carrés afin de calculer, pour chaque coupe,
la valeur de l’écart type de la distribution de Rayleigh: σR.
La moyenne et l’écart type (µ ± σ) de σR sur les 112 coupes étaient de 14.8 ± 3.2 niveaux de
gris. Cette valeur moyenne est d’ailleurs peu différente de la valeur trouvée pour σR à partir
de l’ajustement de l’histogramme 3D (13.8 niveaux de gris, cf. Figure IV.8).
On a donc considéré dans la suite que le bruit n’était pas modifié par la dégradation de la
sensibilité de la sonde. La moyenne de σR sur les 112 coupes a donc été prise comme
204

Chapitre IV: Imag erie 3D in vivo du doigt humain
205
représentative du bruit dans l’image et a été utilisée pour l’initialisation des deux méthodes de
segmentation.
IV.4.2. Application de la méthode d’étiquetage
La méthode d’étiquetage utilisée pour la segmentation des images de tissu trabéculaire
obtenues par microscopie IRM (cf. III.5.1) a été appliquée sur la ROI déterminée
précédemment (cf. IV.3.3.4.1). Seule la phase d’initialisation de l’algorithme a été modifiée:
le couple (σR,µG1) utilisé dans le cadre de l’étude in vitro (histogramme bimodal, cf. Figure
III.20) est remplacé, du fait de la nature de l’histogramme (cf. Eq. IV.1), par (σR,µG2).
La Figure IV.20 présente un exemple de segmentation par cette méthode.
A B C
D E F Figure IV.20: Exemple de segmentation de la ROI trabéculaire (cf. IV.3.3.4.1) par la méthode d’étiquetage
(cf. III.5.1): coupes sagittale (A et D), coronale (B et E) et axiale (C et F).
Visuellement, cette segmentation semble insuffisante: on observe en effet un grand nombre de
travées déconnectées. Ceci est principalement dû au volume partiel: de nombreux voxels de
ces images ont un voisinage contenant majoritairement des voxels affectés par cet artefact, ce
qui correspond à des cas problématiques pour cet algorithme de segmentation (cf. III.5.1) et

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
donc à des erreurs principalement sur les voxels d’os (du fait que les voisinages analysés
contiennent majoritairement des voxels de moelle). La qualité de la segmentation n’est donc
pas aussi bonne que celle obtenue dans le cadre de l’étude in vitro.
IV.4.3. Application de la méthode de réduction du volume
partiel
Dans un article de MRM publié en 2002 ([Hwang, 2002]), l’équipe de Wehrli a proposé une
méthode permettant de réduire l’effet du volume partiel sur des images IRM de tissu
trabéculaire acquises in vivo. Nous la présentons plus en détails ci-dessous.
Le principe de l’algorithme est basé sur la division de chaque voxel en 8 sous-voxels dans
lesquels le contenu osseux est réparti de façon à assurer la conservation de la masse,
c’est-à-dire que la masse osseuse contenue dans chaque voxel doit être égale à la somme des
masses osseuses des 8 sous-voxels. Cette technique ne donne pas accès directement à une
segmentation de l’image mais permet de la faciliter.
IV.4.3.1. Cartographie de la fraction osseuse
Cet algorithme suppose que l’on dispose de la valeur de la fraction osseuse de chaque voxel.
Une cartographie préalable des fractions osseuses (cf. Figure IV.21) est donc établie à partir
de l’intensité de chaque voxel (donc de son niveau de gris) et de la coupe axiale à laquelle il
appartient (pour prendre en compte la dégradation de la sensibilité de la sonde), comme cela a
été fait pour l’initialisation de la méthode d’étiquetage (cf. IV.4.2).
206

Chapitre IV: Imag erie 3D in vivo du doigt humain
207
A B C Figure IV.21: Illustration de la cartographie initiale de la fraction osseuse sur la ROI déterminée en
IV.3.3.4.1: coupes sagittale (A), coronale (B) et axiale (C). L’extérieur de la ROI est en noir sur ces images.
Une valeur de fraction osseuse est attribuée à chaque voxel (0%: noir, 100%: blanc) en fonction de son
niveau de gris et de la coupe axiale à laquelle il appartient (sensibilité de la sonde). Ces images sont donc
représentées selon une échelle de niveau de gris complémentaire de celles présentées précédemment.
La Figure IV.22 montre l’histogramme de la fraction osseuse ainsi calculée. Cette distribution
est à peu de choses près symétrique de celle présentée en Figure IV.19. Les pics aux deux
extrémités proviennent des voxels initialisés respectivement à une fraction osseuse nulle
(voxels de niveau de gris supérieur à µG2) et à une fraction osseuse égale à 1 (voxels de niveau
de gris inférieur à σR). L’allure bimodale du reste de la distribution reflète respectivement les
deux modes gaussiens: voxels de moelle (niveaux de gris élevés) et de volume partiel
(niveaux de gris intermédiaires).

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
208
Fraction osseuse
Occurrence normalisée
0
0,002
0,004
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
0,006
0,008
0,01
0,012
0,014
0,016
Figure IV.22: Histogramme de la cartographie initiale des fractions osseuses. Celui-ci est une sorte de
« symétrique » de celui présenté en Figure IV.19: les voxels de signal correspondent à des fractions
osseuses faibles, ceux de bruit à des fractions osseuse élevées. On n’a représenté volontairement que les
occurrences normalisées inférieures à 0.016 afin de ne pas « écraser » l’histogramme. Pour information,
les proportions de voxels de fractions osseuses inférieures à 0.01 et supérieures à 0.99 sont respectivement
de 0.066 et 0.016.
IV.4.3.2. Algorithme de subdivision
Chaque voxel est alors subdivisé en 8 sous-voxels et son contenu osseux est réparti en deux
phases.
IV.4.3.2.1. Première phase
Pour la première phase, la répartition est basée sur l’hypothèse qu’il n’y a pas d’amas de tissu
calcifié isolé dans la moelle. En d’autres termes, la probabilité pour qu’un sous-voxel
contienne de l’os est d’autant plus forte quand il est entouré de voxels contenant de l’os.
Un poids wi est donc calculé pour chacun des 8 sous-voxels i comme la somme des fractions
osseuses des 7 voxels situés dans le voisinage de i à l’extérieur du voxel où i est défini. Le
contenu osseux du voxel est ensuite réparti dans chaque sous-voxel i proportionnellement à
son poids relatif dans le voxel, wi/wt (où wt, le poids total du voxel, est égal à la somme des 8
wi):
voxelt
iivoxelsous BV
wwBV =− ,
où BVsous-voxel i et BVvoxel sont respectivement les volumes d’os dans le sous-voxel i et dans le
voxel. Ceci assure donc la conservation de la masse osseuse puisque

Chapitre IV: Imagerie 3D in vivo du doigt humain
209
voxeli
ivoxelsous BVBV =∑=
−
8
1.
Ainsi, on peut obtenir une cartographie de la fraction osseuse des sous-voxels de la manière
suivante:
voxelsous
voxelvoxel
t
i
voxelsous
voxel
voxel
voxel
t
i
voxelsous
voxel
t
i
voxelsous
ivoxelsousivoxelsous V
VBVFww
VV
VBV
ww
VBV
ww
VBV
BVF−−−−
−− ××=××=×== ,
où BVFsous-voxel i et BVFvoxel sont respectivement les fractions osseuses du sous-voxel i et du
voxel analysé, Vsous-voxel et Vvoxel, les volumes respectivement d’un sous-voxel et d’un voxel.
Or, comme Vvoxel = 23×Vsous-voxel, on a:
32××=− voxelt
iivoxelsous BVF
wwBVF .
La Figure IV.23 illustre cette première répartition sur un exemple en dimension 2.
d a
c b
0 0.3 0.4
0 0 0.5
0 0.6
0.09 0.38
0 0.33
0 0.3 0.4
0 0 0.5
0 0.6
A B
Figure IV.23: Illustration en dimension 2 du principe de la première répartition du contenu osseux. On
suppose que le pixel étudié (pixel central) a une fraction osseuse de 0.2. Celui-ci est subdivisé en 4
sous-pixels a, b, c et d (A). Chacun de ces sous-pixels i se voit attribuer un poids égal à la somme des
fractions osseuses des 3 pixels situés dans le voisinage de i à l’extérieur du pixel où i est défini:
wa = 0.3 + 0.4 + 0.6 = 1.3, wb = 0.6 + 0.5 = 1.1, wc = 0, et wd = 0.3 (en dimension 3, on aurait 7 voxels voisins
pour chaque sous-voxel). D’après le poids total du pixel wt (égal à la somme des poids de chaque sous-pixel
soit 2.7 dans cet exemple), on peut établir le poids relatif de chaque sous-pixel i: wi/wt. La cartographie de
la fraction osseuse des sous-pixels (donnée en B) est alors déterminée par la formule donnée dans le texte
(« 23 » étant remplacé par « 22 » pour la dimension 2).
IV.4.3.2.2. Deuxième phase
Pour la deuxième phase, l’algorithme utilise le résultat de la première répartition. Chaque
sous-voxel contenant de l’os (c’est-à-dire de fraction osseuse non nulle) est affecté d’une
classe, « centrale » ou « frontière », qui est déterminée à partir de l’analyse de son voisinage
3×3×3. Un sous-voxel est de type « frontière » si les deux critères suivants sont vérifiés:

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
210
• le sous-voxel analysé a au moins un voisin (au sens de la 6-connexité, cf. II.4.4.2.1) qui ne
contient pas d’os
• le sous-voxel analysé n’est pas essentiel dans la topologie des voxels d’os du voisinage
c’est-à-dire que s’il était remplacé par de la moelle, le nombre d’amas connectés de voxels
d’os du voisinage (au sens de la 26-connexité, cf. II.4.4.2.1) ne serait pas modifié.
Si au moins l’un des deux critères n’est pas vérifié, la classe du sous-voxel est « central ». Un
exemple de chaque classe de sous-voxel est donné sur la Figure IV.24,
A B
Figure IV.24: Exemple, en dimension 2, de chaque classe. Le sous-pixel étudié (qui contient de l’os) est
représenté en gris. Les autres sous-pixels contenant de l’os dans son voisinage 3×3 sont indiqués en noir.
En A, le sous-pixel est de type « frontière », en B il est de type « central ».
Le principe de la répartition du contenu osseux du voxel dans les 8 sous-voxels est le même
que lors de la première phase (le calcul des poids se faisant à partir des fractions osseuses des
sept sous-voxels adjacents déterminées lors de la première répartition) sauf que le contenu
osseux est prioritairement distribué sur les sous-voxels de type « central ». La masse osseuse
du voxel qui n’a pas été attribuée est ensuite répartie sur les sous-voxels « frontière ».
La deuxième phase de l’algorithme a donc pour effet de compacter l’os tout en maintenant
l’hypothèse de conservation de la masse osseuse. Ceci se traduit par une augmentation du
nombre de sous-voxels avec une fraction osseuse proche de 0 ou de 1 et une diminution du
volume partiel.
IV.4.3.2.3. Illustration sur l’image de la phalange distale
La Figure IV.25 montre l’évolution de l’histogramme de fraction osseuse au cours des deux
phases. On observe bien une diminution des voxels de fractions osseuses intermédiaires
(volume partiel) qui se retrouvent « rejetées » aux extrémités de l’histogramme (on a pu
chiffrer sur cet exemple que 10% des voxels du volume partiel ont pu être reportés à des
fractions osseuses extrêmes).

Chapitre IV: Imagerie 3D in vivo du doigt humain
211
Fraction osseuse
Occurrencenormalisée
0
0,002
0,004
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
0,006
0,008
0,01
0,012
0,014
0,016
Figure IV.25: Histogrammes des cartographies des fractions osseuses après la première (trait noir épais)
et la deuxième (trait gris épais) phases. On rappelle aussi celui de la cartographie initiale (trait noir fin, cf.
Figure IV.22). On n’a représenté volontairement que les occurrences normalisées inférieures à 0.016 afin
de ne pas « écraser » l’histogramme. Pour information, les proportions de voxels de fractions osseuses
inférieures à 0.01 et supérieures à 0.99 sont respectivement de 0.074 et 0.044 après la première phase et de
0.084 et 0.066 après la deuxième phase (à comparer avec 0.066 et 0.016 pour la cartographie initiale). On
observe bien une diminution du volume partiel au profit des fractions osseuses faibles et fortes.
Après ces deux phases, nous proposons alors d’appliquer un seuillage simple (pour un seuil
correspondant à une fraction osseuse de 0.5) pour obtenir une segmentation de l’image (cf.
Figure IV.26). Visuellement, cette segmentation semble de meilleure qualité que celle
obtenue par la méthode d’étiquetage (cf. IV.4.2 et Figure IV.20).

Caractérisation de l’architecture tridimension laire panelle du tissu trabécu r IRM
212
A B C
D E F Figure IV.26: Exemple de segmentation par la méthode de réduction du volume partiel, sur la ROI
déterminée précédemment (cf. IV.3.3.4.1): coupes sagittale (A et D), coronale (B et E) et axiale (C et F).
On a pour chaque axe de l’image segmentée (D, E et F), le double de voxels par rapport à la Figure IV.20.
Remarque:
a) Ce procédé a des similitudes avec la méthode utilisée pour la segmentation des images
obtenues in vitro par microscopie IRM (cf. III.5.1): ces deux méthodes cherchent en effet à
« étirer » l’histogramme de l’image vers ses extrémités à partir de l’analyse du voisinage de
chaque voxel.
b) L’algorithme tel qu’il est proposé ne donne pas une image segmentée. Nous avons choisi
d’effectuer un seuillage de la cartographie de la fraction osseuse pour l’obtenir. Il pourrait
cependant être intéressant de le remplacer par une méthode de segmentation plus complexe
basée sur cette cartographie: par exemple la méthode d’étiquetage.
c) La conservation de la masse osseuse est assurée au niveau de la cartographie des fractions
osseuses. Or, cette hypothèse n’est plus vérifiée à partir du moment où la segmentation est
effectuée.

Chapitre IV: Imagerie 3D in vivo du doigt humain
IV.5. Premiers résultats
IV.5.1. Images étudiées
Pour cette première étude de faisabilité, deux images du même doigt (d’un même patient) ont
été analysées. Celles-ci ont été acquises d’affilé sans que le patient ne change de position dans
l’imageur. La première image (notée dans la suite « Image 1 ») est celle qui a servi pour
illustrer les différentes étapes de l’étude (cf. IV.2.2, IV.2.3 et Figure IV.2 pour plus de
détails). Les paramètres d’acquisition et les caractéristiques de la deuxième image (notée dans
la suite « Image 2 ») sont très proches:
• TE/TR = 1/2.6 ms
• une bande passante de 200 kHz
• un angle de basculement optimisé pour obtenir un signal maximum de la moelle
• 8 accumulations
• une matrice d’acquisition de 128×128×128 (taille maximale accessible sur le dispositif
utilisé) pour un champ de vue cubique de 20 mm de coté (soit un volume total analysé de
8 cm3).
La même résolution a été obtenue pour cette image (156 µm isotrope) pour un temps
d’acquisition tout a fait acceptable pour l’étude in vivo (6 minutes environ). De même, une
interpolation des données a ensuite été effectuée par la méthode du « zerofill » d’un facteur 2
dans chaque dimension (cf. III.3.1.3), conduisant à une matrice 256×256×256, soit une taille
de voxel après calcul de la transformée de Fourier de 78 µm isotrope avec un RSB mesuré
dans la moelle d’environ 10.
De même qu’en IV.2.4.3, la zone correspondant à la phalange distale a été extraite de l’image
globale par rognage. La Figure IV.27 présente trois plans principaux de cette phalange ainsi
que les dimensions du volume obtenu.
213

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
13.9 mm
9.5 mm
214
12.3 mm
9.5 mm 13.9 mm 12.3 m
m
A B C Figure IV.27: Trois coupes principales correspondant à la phalange distale extraite par rognage de
l’« Image 2 » (A: sagittale, B: coronale, C: axiale). Celle-ci correspond au même doigt du même patient
que celle présentée en Figure IV.4. Les dimensions du volume récupéré sont indiquées sur chaque coupe.
L’histogramme de ce volume est donné en Figure IV.28.
0
0,002
0,004
0,006
0,008
0,01
0 50 100 150 200 250
Occurrence normalisée
NG
Figure IV.28: Histogramme du volume correspondant à la phalange distale de l’« Image 2 ». On peut le
comparer avec celui obtenu pour l’image précédemment étudiée: Figure IV.5.
Sur cette image, la ROI « trabéculaire » a été déterminée de la même manière qu’en IV.3. On
donne en Figure IV.29, trois plans principaux de cette ROI ainsi que son histogramme en
Figure IV.30.

Chapitre IV: Imagerie 3D in vivo du doigt humain
215
A B C
Figure IV.29: Représentation de la ROI isolée (en trait blanc) sur l’« Image 2 » pour les mêmes coupes
que celles présentées en Figure IV.27 (A: sagittale, B: coronale, C: axiale).
0
0,002
0,004
0,006
0,008
0 50 100 150 200 250
Occurrence normalisée
NG
Figure IV.30: Histogramme calculé sur la ROI « trabéculaire » déterminée précédemment sur
l’« Image 2 » (cf. Figure IV.29). On peut le comparer vec celui obtenu pour l’« Image 1 »: Figure IV.19 a
Visuellement, les deux images étudiées semblent relativement proches. Cependant, une
différence importante peut être notée sur l’histogramme de la Figure IV.30. Bien que le RSB
ne soit pas significativement différent, l’effet du volume partiel semble moins sensible sur la
deuxième image (le mode des voxels de signal se distingue plus « facilement » sur cet
histogramme que sur son équivalent pour l’« Image 1 », cf. Figure IV.19).

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
IV.5.2. Protocole
Sur chaque image, après avoir été délimitée (cf. IV.3), chaque ROI a été segmentée. Du fait
que l’application de l’algorithme de réduction du volume partiel (cf. IV.4.3) change la taille
du voxel, deux études ont été menées. Dans le Tableau IV.1, on indique les images utilisées,
le type de segmentation méthod « RVP réduction
du volum ) et le voxels associées. Dans chaque cas, les deux images
présentées en IV.5.1 ont été analysées.
(« E » pour la e d’étiquetage, » pour celle de
e partiel s tailles de
Image source Segmentation Taille de voxel
Etude 1 « zerofill » E 78 µm isotrope
Etude 2 pas de « zerofill » RVP 78 µm isotrope
Tableau IV.1: Récapitulatif des deux études de microarchitecture effectuées sur les deux images
présentées précédemment (cf. IV.5.1). « E » et « RVP » correspondent respectivement à la méthode de
segmentation par étiquetage (cf. IV.4.2) et à la méthode de réduction du volume partiel (cf. IV.4.3).
Les caractéristiques des acquisitions des images étant proches de celles de l’étude in vitro, les
algorithmes de calcul des paramètres (cf. chapitre II) ont été appliqués de la même manière
qu’au chapitre III (cf. III.6.1). La différence principale se situe au niveau du choix des ROI.
Pour la fraction osseuse et la contribution à la caractéristique d’Euler-Poincaré, le calcul est
mené sur la ROI entière (cf. IV.3). Par contre, pour les autres paramètres, le positionnement
e la plus grande sphère incluse dans la ROI (qui a pu être facilement automatisée pour
mettre en œuvre ici. A ce stade, ce positionnement est manuel. Il faudra cependant prévoir
une estriction de ROI.
ties suivantes.
IV.5.3. Volume de données analysé
Le Tableau IV.2 indique les volumes analysés pour chaque image (cf. IV.5.1).
d
l’étude in vitro du fait de la forme cylindrique des échantillons, cf. III.6.1) est plus difficile à
méthode pour déterminer automatiquement cette r
Nous détaillons les résultats obtenus pour les différents paramètres dans les par
216

Chapitre IV: Imagerie 3D in vivo du doigt humain
Image 1 Image 2 Volume
(mm3) ROI entière Restriction sphère ROI entière Restriction sphère
Etudes 1 et 2 376 65 365 65
Tableau IV.2: Volume analysé pour chaque image (cf. IV.5.1).
Les e la
technique de délimitation de la ROI présentée en IV.3.
Pour le re de même rayon a pu être positionnée: les volumes analysés
pour les paramètres histomorphométriques, d’anisotropie et d’orientation sont donc identiques
IV.5.4.1. Résultats
on négligeable de l’amas principal de la phase calcifiée: inférieur à 0.4% pour les
deux images. Dans ces conditions, on a donc pu appliquer le principe de filtrage des amas
présenté en II.4.6.1.
Le Tableau IV.3 indique la fraction osseus à la istique
d’Euler-Poincaré, mesurées sur chaque ima IV.5.1) et les deux ét nvisagées
(cf. IV.5
volumes des ROI entières sont comparables ce qui illustre bien la robustesse d
s deux images, une sphè
entre les deux images.
IV.5.4. Fraction osseuse et caractéristique d’Euler-Poincaré
Pour les deux méthodes de segmentation envisagées, les amas secondaires ne représentaient
qu’une fracti
e et la contribution caractér
ge (cf. pour udes e
.2).
BV/TV CN3 (en mm-3)
Image 1 Image 2 Image 1 Image 2
Etude 1 0.227 0.287 -4.1 -6.7
Etude 2 0.324 0.308 -10.8 -8.8
Tableau IV.3: Fraction osseuse (BV/TV) et contribution à la caractéristique d’Euler-Poincaré (CN3)
mesurées sur chaque image (cf. IV.5.1) et pour les deux études envisagées (cf. IV.5.2).
217

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
La contribution à la caractéristique d’Euler-Poincaré est particulièrement élevée: cela semble
confirmer l’impression visuelle d’un grand nombre de travées interconnectées en particulier
dans les coupes axiales (cf. Figure IV.4 C et Figure IV.27 C).
De même, le nombre important de travées semblant être déconnectées par la segmentation par
étiquetage (cf. IV.4.2: Figure IV.20), paraît se confirmer ici: la valeur absolue de CN3 est plus
faible pour la première étude que pour la deuxième.
Alors que ces deux paramètres varient sensiblement d’une étude à l’autre pour l’« Image 1 »,
ils sont plus stables pour l’« Image 2 ». Il est possible que cela provienne du fait que
l’« Image 2 » soit moins affectée par l’effet du volume partiel que l’« Image 1 » (cf. Figure
IV.30
robustes et cela aboutirait alors à une évaluation plus fiable des deux paramètres. Une autre
hypothèse serait que ces fluctuations sont liées à un RSB limite qui rendrait ces mesures non
IV.5.4.2.1. Protocole
ésente des travées
é
vol
seg
A
gén
typ , pour différentes valeurs de RSB (comprises entre 6 et 40):
par rapport à la Figure IV.19). Les algorithmes de segmentation seraient alors plus
significatives.
Afin d’essayer de lever cette ambiguïté, nous avons étudié, à partir des données de l’étude in
vitro, si le RSB pouvait être à l’origine de ces fluctuations.
IV.5.4.2. Influence du RSB sur la segmentation
Ne disposant d’aucune référence sur le doigt, nous avons utilisé les données in vitro afin de
comparer les deux méthodes de segmentation. Compte tenu de la résolution utilisée dans le
cadre de l’étude in vitro (66 µm) et du site osseux (le calcanéum pr
paisses), on peut considérer que les images ne sont quasiment pas affectées par l’effet de
ume partiel. Nous avons donc pu ainsi n’étudier que l’influence du RSB sur la
mentation pour la fraction osseuse et la contribution à la caractéristique d’Euler-Poincaré.
partir d’une étude similaire à celle présentée en III.6.5.3 (cf. Figure III.38), nous avons
éré, en se basant sur une acquisition haute résolution (rayonnement synchrotron), deux
es d’image
• « acquisition IRM brute »: en effectuant la transformée de Fourier inverse après avoir
ajouté le bruit gaussien, sur le centre du plan de Fourier, c’est-à-dire sur les points qui
n’ont pas été mis à zéro (centre de l’image présentée en Figure III.38 D). La taille de voxel
ainsi obtenue est donc de 132 µm dans chaque direction.
218

Chapitre IV: Imagerie 3D in vivo du doigt humain
219
• ode du « zerofill »: en suivant le même protocole que celui
type d’image (acquisition « brute »), la méthode de segmentation par
réduction du volume partiel a été appliquée; sur le deuxième, celle par étiquetage. Une
comparaison entre les deux méthodes de segmentation a ainsi pu être menée à même taille de
voxel, de la même manière qu’entre les études 1 et 2 présentées en IV.5.2.
IV.5.4.2.2. Résultats
La Figure IV.31 présente l’évolution, pour les images simulées, de la fraction osseuse et de la
contribution à la caractéristique d’Euler-Poincaré en fonction du RSB pour les deux méthodes
de segmentation.
BV/TV
après interpolation par la méth
présenté en Figure III.38. La taille de voxel ainsi obtenue est donc de 66 µm dans chaque
direction.
Pour le premier
RSB0,175 10 15 20 25 30 35 40
RSB
0,18
0,2
CN3 (mm-3)
0,19
-75 10 15 20 25 30 35 40
A B Figure IV.31: Evolution, pour les images simulées, de la fraction osseuse (A) et de la contribution à la
caractéristique
-6
-5
-2
d’Euler-Poincaré en fonction du RSB, pour les méthodes de segmentation par étiquetage
-4
-3
(en noir) et par réduction du volume partiel (en gris). La valeur correspondante à l’image de référence
(SR-µCT à 66 µm) est indiquée par un trait pointillé.
On obtient une estimation sensiblement identique de la fraction osseuse pour les deux
méthodes de segmentation, quel que soit le RSB. Cette valeur est systématiquement
surestimée pour la gamme « classique » de RSB obtenus en IRM (inférieur à 20).
Logiquement, plus le RSB augmente, plus on se rapproche de la valeur mesurée sur l’image
de référence.
Pour la contribution à la caractéristique d’Euler-Poincaré, les deux méthodes permettent
d’accéder à une mesure relativement précise. On peut remarquer que la valeur est surestimée

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
par la métho tude in vitro, cf. Tableau
l’écart des valeurs mesurées pour BV/TV entre les études 1 et 2
Pou soient proches celles de l’étude in vitro:
l’hypothèse d’une dim ages est probable.
ur V
IV.5.5. Anisot t orientation
Le Tableau IV.4 indique les es d ϕ, inés par rapport au repère de
l’im es deu efficient isotropi et RM esurée chaque i (cf.
IV.5.1) et pour les deux études envisagées (cf. IV.5.2).
de d’étiquetage (comportement déjà observé lors de l’é
III.5 et Figure III.27 A: l’ordonnée à l’origine de la régression linéaire entre les modalités
µIRM et SR-µCT est supérieure à 0) et sous-estimée par celle de réduction du volume partiel.
Pour un RSB supérieur à 13, l’écart entre les deux méthodes de segmentation est constant
(environ 1.6 mm-3).
IV.5.4.2.3. Discussion sur les résultats obtenus in vivo
Il est intéressant de noter que
diminue sensiblement sur l’image 2 (d’environ 0.1 pour l’image 1, il passe à environ 0.02
pour l’image 2). Pour CN3, celui-ci diminue aussi: il passe de presque 7 mm-3 à 2 mm-3,
valeur très proche de celle mesurée dans le cadre de l’étude précédente, c’est-à-dire quand
l’effet de volume partiel est négligeable (1.8 mm-3 pour un RSB de 10, cf. IV.5.4.2.2 et Figure
IV.31 B).
r l’image 2, il semble donc que les conditions
inution de l’effet de volume partiel entre les deux im
Dans ce cadre, l’évaluation de la fraction osseuse par l’algorithme de réduction du volume
partiel serait alors plus efficace, puisque la mes e de B /TV par cette méthode est stable
entre les deux images.
ropie e
trois angl ’Euler (θ, ψ, déterm
age) et l x co s d’an e (RMIN AX), m s sur mage
220

Chapitre IV: Imagerie 3D in vivo du doigt humain
θ (en °) ϕ (en °) ψ (en °)
Image 1 Image 2 Image 1 Image 2 Image 1 Image 2
Etude 1 -89 -89 -89 -90 156 157
Etude 2 -90 -89 -90 -90 161 159
RMIN RMAX
Image 1 Image 2 Image 1 Image 2
Etude 1 0.52 0.59 0.80 0.82
Etude 2 0.60 0.63 0.83 0.84
.5.1) et pour les deux études envisagées (cf. IV.5.2).
Tableau IV.4: Angles d’Euler (θ, ϕ, ψ) et coefficients d’anisotropie (RMIN et RMAX) mesurées sur chaque
image (cf. IV
La comparaison de l’orientation de la microstructure entre les deux images est possible ici
sans recalage préalable des ROI du fait que les deux acquisitions ont été menées à la suite et
qu’il a été demandé au patient de garder une position identique.
Les résultats obtenus sur ces paramètres semblent très robustes puisque quelles que soient la
technique et l’image analysée, on trouve une très forte anisotropie de la microstructure selon
axe de lecture (en accord avec l’impression visuelle, cf. Figure IV.4 A,B et Figure IV.27
n utilisée. En ce qui concerne les autres paramètres, il faudrait prévoir des
l’
A,B).
IV.5.6. Premières conclusions
Il est très difficile de conclure sur un jeu de données aussi petit: il faudrait étoffer la base
d’images.
A ce stade, le résultat le plus probant est la forte anisotropie de la structure trabéculaire selon
l’axe du doigt. Ceci apparaît très clairement quelle que soit l’acquisition et la méthode de
segmentatio
confrontations avec d’autres modalités afin d’établir la fiabilité de la mesure. Par exemple, du
221

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
fait que nous pouvons délimiter séparément les zones trabéculaire et corticale, un examen par
DXA (cf. I.5.1) devrait permettre de valider notre étude pour la fraction osseuse.
ion: du fait qu’il a été plus court pour la
ixer le doigt du patient durant l’acquisition (avec un plâtre, par
xemple) pourrait aussi permettre de vérifier notre supposition.
Il apparaît vraisemblable que l’effet du volume partiel soit une des limites principales à la
précision des mesures. L’origine la plus probable de la diminution du volume partiel entre
l’image 1 et l’image 2 serait le temps d’acquisit
deuxième acquisition (6 minutes au lieu de 12, cf. IV.2.2 et IV.5.1), le mouvement
(inévitable) du doigt du patient a été moins important ce qui a certainement atténué l’effet de
volume partiel (images plus « nettes »).
Il pourrait être très intéressant d’effectuer des acquisitions à résolution plus fine quitte à
diminuer dans une certaine mesure le RSB (ce paramètre ne semble en effet ne pas être le plus
critique, cf. Figure IV.31). Ces images pourraient alors nous permettre de valider notre
hypothèse.
Un protocole permettant de f
e
IV.6. Perspectives
Nos premiers résultats montrent la faisabilité de l’étude de la microarchitecture du tissu
abéculaire in vivo. Nous disposons maintenant d’outils fiables et robustes qu’il faudrait
rrait
tre contourné en acquérant le plan de Fourier en plusieurs étapes: par exemple en effectuant
« concaténation » de 8 blocs de 1283 voxels pour en obtenir un seul de 2563.
nsuite, le RSB des images analysées dans cette étude pourrait être encore amélioré:
l’utilisation d’une séquence dite « Gradient Echo refocalisé » permettrait en effet de récupérer
la composante transversale de l’aimantation qui existe encore après les temps de répétition
particulièrement courts de cette étude. Cette « réserve de signal » pourrait ainsi être employée
pour améliorer la résolution.
Les récents développements en instrumentation (en particulier par l’emploi de sonde
supraconductrice, [Ginefri, 2001]) devraient aussi permettre d’obtenir des images de qualité
tr
mettre en œuvre sur d’autres acquisitions pour pouvoir définir le meilleur protocole. Un
certain nombre d’améliorations sont déjà prévues.
Tout d’abord, le problème de la taille limite de la matrice d’acquisition (cf. IV.2.2) pou
ê
la
E
222

Chapitre IV: Imagerie 3D in vivo du doigt humain
comparable sur des imageurs IRM disponibles en milieu hospitalier. Les sites analysés
pourraient ainsi être beaucoup plus nombreux.
Enfin, il faudra aussi prévoir l’adaptation de la méthode de recalage de la ROI développée
dans le cadre de l’étude in vitro (cf. III.4) afin de pouvoir mener des études de comparaisons
in vivo: efficacité de traitements médicamenteux, suivi de l’évolution de la pathologie….
223


Conclusion
Conclusion
La caractérisation
technique est donc utilisée dans de nom
actuelles mettant en jeu l’IRM
(réso
mode,…), soit par les techniques de traitement (influence de la segmentation, comparaison de
régions d’intérêt différentes ou de
contribue à dim
ené cherche à s’affranchir de ces différents facteurs.
s paramètres, les méthodes de
ensionnelles utilisées et développées dans ce travail ont été validées à partir
la validité de la représentation du tissu
agerie proches de ce que l’on peut obtenir in
du tissu trabéculaire par IRM présente de nombreux avantages et cette
breux travaux. Cependant, la plupart des recherches
sont souvent limitées soit par la qualité des acquisitions
lution limite, rapport signal sur bruit faible, histogramme ne présentant qu’un seul
calculs effectués dans des dimensions différentes,…). Ceci
inuer la signification des résultats.
Le travail que nous avons m
Tout d’abord en ce qui concerne les techniques d’évaluation de
calcul tridim
d’études sur des images « test ».
Ensuite, nous avons pu établir par une étude in vitro,
trabéculaire par IRM dans des conditions d’im
vivo. Ce travail a été fait en s’affranchissant d’un nombre important de degrés de liberté ce
qui a permis de ne caractériser que les différences liées aux phénomènes physiques à l’origine
du contraste:
225

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
• les échantillons: 29 échantillons ont été analysés ce qui a assuré des calculs statistiques
significatifs. De plus, les mêmes échantillons ont été utilisés pour les deux modalités
d’imagerie;
menée sur les mêmes régions d’intérêt, dans la même position (ce qui est
essentiel pour comparer l’orientation de la structure) et ce grâce à une méthode efficace de
e sont donc très robustes. Ils ont montré un très bon rendu par IRM
à la contribution à la
c
d’a
éta
plu
obs ation par IRM des
aramètres histomorphométriques. Nous avons aussi émis et vérifié des hypothèses pouvant
ffet semble cependant difficile: aucune méthode de
orrection de ce biais n’a pu être mise en œuvre pour le moment.
nfin, les bases de l’étude du tissu trabéculaire d’images IRM in vivo ont été posées. Des
images très haute résolution du doigt humain à 7 T ont été acquises. Celles-ci ont servi pour
mettre au point une méthode 3D automatique permettant d’isoler séparément les zones
trabéculaire et corticale des autres tissus. En utilisant une technique de segmentation dédiée
• la qualité de la référence: en utilisant la microtomographie X par rayonnement synchotron,
nous disposions d’une série d’image de référence de très bonne qualité (résolution
parfaitement adaptée à la microarchitecture trabéculaire et très haut rapport signal sur
bruit). Cette technique est d’ailleurs couramment admise comme référence pour l’imagerie
trabéculaire;
• le recalage des régions d’intérêt (ROI): la comparaison entre les deux modalités
d’imagerie a été
recalage (rapide, précise et automatique). De plus, les tailles de voxel ont été ajustées et
les mêmes algorithmes de calcul (3D) des paramètres ont été appliqués pour les deux
modalités;
• la segmentation: la méthode utilisée en IRM est parfaitement adaptée aux caractéristiques
des images. Elle n’est pas arbitraire et donne un résultat de très bonne qualité.
Les résultats de cette étud
66 µm de l’organisation générale de la microarchitecture (c’est-à-dire
aractéristique d’Euler-Poincaré, les trois angles d’Euler et les deux coefficients
nisotropie). Par contre l’évaluation de la fraction osseuse, bien que relativement précise,
it affectée par un biais systématique de 0.07 ce qui correspondait pour les échantillons de
s faible fraction osseuse, à des erreurs relatives de l’ordre de 150%. On a d’ailleurs pu
erver que ce biais était la cause principale des erreurs sur l’évalu
p
être à l’origine de ce biais: dégradation de la résolution, méthode de segmentation, traitement
des images IRM (interpolation par la technique du « zerofill »), effet T2*. L’hypothèse la plus
probable semblerait être la présence de microbulles d’air confinées à l’interface entre le tissu
calcifié et l’eau. La modélisation d’un tel e
c
E
226

Conclusion
aux acquisitions IRM in vivo (proposée par l’équipe de Wehrli dans une publication récente
dans le journal Magnetic Resonance in Medicine: 47, 948-957, 2002), nous avons pu effectuer
les premières études des paramè s cependant analysé à ce stade
que deux images acquises sur un même patient.
Améliorations
Des améliorations sont déjà envisagées au niveau de l’instrumentation et de la méthodologie:
• utilisation d’autres séquences d’acquisition: ceci devrait permettre un gain non négligeable
en terme de résolution ou de sensibilité. En particulier, dans le cadre de l’étude in vivo,
nous devrons tester par exemple, la séquence écho de gradient refocalisée;
• emploi de sondes supraconductrices dédiées à l’IRM: les récents travaux menés sur cette
thématique dans l’unité ont montré que cette instrumentation offre une augmentation
Ces sondes devraient perm de qualité similaire sur des appareils
vivo de sites osseux plus fréquemm
vertèbre;
éthode de
nécessaire pour analyser et
doigt.
Perspectives
Les perspectives de ce travail sont nombreuses.
sur les images de doigt, un exam
l’évaluation de la fraction osseuse in vivo.
En ce qui concerne les param t de mettre en œuvre
l direct de l’espacem
semblent beaucoup plus significatives que celle
structure osseuse. De plus, l’ entation
semble une voie intéressante: il est en effet particulièrement intéressant de pouvoir éviter cette
étape surtout lorsque les images sont acquises en limite de résolution, ce qui est souvent le cas
tres de structure. Nous n’avon
substantielle du rapport signal sur bruit et donc une amélioration de la résolution spatiale.
ettre d’obtenir des images
utilisés couramment en routine clinique (imageurs 1.5 T). Ceci faciliterait l’imagerie in
ent affectés par les fractures comme le radius ou la
• adaptation de la m recalage des régions d’intérêt au cas in vivo: ceci est
comparer les autres images que nous avons acquises sur le
A court terme, comme nous pouvons délimiter séparément les zones trabéculaire et corticale
en par DXA de ce site devrait permettre de valider
ètres d’architecture, il pourra être intéressan
les méthodes de calcu ent et de l’épaisseur des travées; ces grandeurs
s basées sur une hypothèse de modèle de la
utilisation de paramètres ne nécessitant aucune segm
227

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
in vivo. Enfin, l’influence
étudiée facilement à partir
Nous allons aussi poursui
avons entamée dans le cadre du GDR. Une première étude, à partir d’images 2D acquises in
déjà permis
trabéculaire et l’âge, le sex
travail a d’ailleurs condui
poursuivre vers une analy dans un premier
o.
A moyen terme, des étude ution de la structure osseuse avec
certaines p
discriminante pour le di
pourront être envisagées
« animal ». Nous disposero nt faciliter considérablement la mise
s projets:
• acquisition par notre un
• recalage des régions d’
effet très difficile de
même zone d’intérêt: i
varie à chaque nouv
érents o
A plus longue échéance, i
de prédiction de grandeu partir d’études par éléments finis ou par tests
échant
des unités travaillant sur ce
de la résolution sur ces paramètres évalués par IRM pourra être
des données in vitro.
vre l’analyse du tissu trabéculaire mandibulaire par IRM que nous
vitro, nous a d’établir un certain nombre de corrélations entre l’état du tissu
e, la présence de dent sur la mandibule et le site mandibulaire (ce
t à la rédaction d’un manuscrit, cf. Annexe 4). Nous souhaiterions
se tridimensionnelle de ce site osseux, in vitro
temps puis in viv
s in vivo comme le suivi de l’évol
l’âge ou avec athologies, afin par exemple, d’établir une base de paramètres
agnostic médical, l’efficacité de traitements médicamenteux,…
sur différents sites osseux, pour des patients ou pour un modèle
ns en effet d’outils qui devraie
en œuvre de ce
ité d’un imageur IRM « petit animal » à 4.7 T
intérêt in vivo ce qui est essentiel dans ce type d’étude. Il semble en
pouvoir s’assurer, expérimentalement, d’analyser exactement la
mages acquises à des moments différents (position du patient qui
el examen), contraintes particulières sur le patient, utilisation
d’imageurs diff u même de modalités d’imagerie différentes,...
l sera aussi possible d’envisager la recherche de modèles efficaces
rs mécaniques à
mécaniques sur des illons ce qui pourrait être mené dans le cadre de collaborations avec
s thématiques.
228

Bibliographie
Bibliographie
[Alaux, 1994] onance magnétique, Paris, Ed: Sauramps
[Antoniadis, 1998 (a)]
8).
[Banse, 2002]
[Bernstein, 2001]
tial resolution and
acquisition strategy, JMRI, 14, 270-280 (2001).
Alaux A., L’image par rés
médical (1994).
Antoniadis T., Scarpelli J-P., Ruaud J-P., Gonord P., Guillot G.,
Bone labelling on micro-magnetic resonance images, Medical
Image Analysis, 3, 119-128 (1998).
[Antoniadis, 1998 (b)] Antoniadis T., Détermination de paramètres structuraux de l’os
trabéculaire par l’analyse d’images RMN à tres haute résolution,
Thèse, Université Paris Sud, Orsay, France (199
Banse X., Devogelaer J.P., Grynpas M., Patient-specific
microarchitecture of vertebral cancellous bone: a peripheral
quantitative computed tomographic and histological study, Bone,
30, 829-835 (2002).
[Benhamou, 2001] Benhamou C.L., Poupon S., Lespessailles, Loiseau S., Jennane R.,
Siroux V., Ohley W., Pothuaud L., Fractal analysis of radiographic
trabecular bone texture and bone mineral density: two
complementary parameters related to osteoporotic fractures, J.
Bone Miner. Res., 16 (4), 697-704 (2001).
Bernstein M.A., Fain S.B., Rieder S.J., Effect of windowing and
zero-filled reconstruction of MRI data on spa
229

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
[Beuf, 2001] Beuf O., Newitt D.C., Mosekilde L., Majumdar S., Trabecular
structure assessment in lumbar vertebrae specimens using
quantitative magnetic resonance imaging and relationship with
[Beuf, 2002]
imaging of normal
[Bittoun, 1987] ar résonance
[Boivin, 2000] ., Santora A.C., Yates J., Meunier
in osteoporotic women,
[Borah, 2000]
R.E., Pierce S.A., Stevens
aging and finite element model, J.
[Bouxsein, 1997] ., Melton
s Int., 7,
[Bresenham, 1965]
Journal, 4, 25-30 (1965).
[Brismar, 1999]
in human vertebrae assessed by MRI, Eur. Radiol.
9 (4), 643-647 (1999).
mechanical competence, J. Bone Miner. Res., 16 (8), 1511-1519
(2001).
Beuf O., Ghosh S., Newitt, D.C., Link T.M., Steinbach L., Ries
M., Lane N., Majumdar S., Magnetic resonance
and osteoarthritic trabecular bone structure in the human knee,
Arthritis & rheumatism, 46 (2), 385-393 (2002).
Bittoun J., Un simulateur numérique d’imagerie p
magnétique nucléaire: conception et application, Thèse, Université
Paris Sud, Orsay, France (1987).
Boivin G.Y., Chavassieux P.M
P.J., Alendronate increases bone strength by increasing the mean
degree of mineralization of bone tissue
Bone, 27 (5), 687-694 (2000).
Borah B., Dufresne T.E., Cockman M.D., Gross G.J., Sod E.W.,
Myers W.R., Combs K.S., Higgins
M.L., Evaluation of changes in trabecular bone architecture and
mechanical properties of minipig verterbrae by three-dimensional
magnetic resonance microim
Bone Mineral Res., 15 (9), 1786-1797 (2000).
Bouxsein M.L., Michaeli D.A., Plass D.B., Schick D.A
M.E., Precision and accuracy of computed digital absorptiometry
for assessment of bone density of the hand, Osteoporosi
444-449 (1997).
Bresenham J.E., Algorithm for computer control of a digital
plotter, IBM System
Brismar T.B., Karlsson M., Li T-Q., Ringertz H., Orientation of
trabecular bone
230

Bibliographie
[Callaghan, 1991]
[Chappard, 1997] Berger G.,
[Chen, 1989] agnetic
[Chung, 1993] s J.L., Kugelmass S.D.,
[Clarkson, 1993]
nstructions, Comp. Geom.: Theory and
re » sur leur site web:
[Consensus, 1993] ce : diagnosis, prophylaxis, and
[Coster, 1989]
is, Ed: Presses du CNRS (1989).
[Day, 2000] ., Hvid I., Weinans H.,
Callaghan P.T., Susceptibility-limited resolution (partie 4.7) dans
Principles of nuclear magnetic resonance microscopy, New York,
Ed: Oxford University Press (1991).
Chappard C., Laugier P., Fournier B., Roux C.,
Assessment of the relationship between BUA and BMD at the
calcaneus using BUA imaging and DXA, Osteoporosis Int., 7,
316-322 (1997).
Chen C.N., Hoult D.I., Signal and noise dans Biomedical m
resonance technology, Bristol, Ed: Adam Hilger (1989).
Chung H-W., Wehrli F.W., William
Relationship between NMR transverse relaxation, trabecular bone
architecture and strength, Proc. Natl. Acad. Sci. USA 90,
10250-10254 (1993).
[Chung, 1995] Chung H-W., Wehrli F.W., Williams J.L., Kugelmass S.D., Wehrli
S.L., Quantitative analysis of trabecular microstructure by 400
MHz nuclear magnetic resonance imaging, J. Bone Miner. Res., 10
(5), 803-811 (1995).
Clarkson K.L., Mehlhorn K., Seidel R., Four results on randomized
incremental co
Applications, 121-185 (1993).
Algorithme disponible en « freewa
cm.bell-labs.com/netlib/voronoi.
Consensus development conferen
treatment of osteoporosis, Am. J. Med., 94, 646-650 (1993).
Coster M., Chermant J.L., Paramètres de bases dans Précis
d’analyse d’images, Par
[Cowin, 2001] Cowin S.C., Section 1: Basic biology dans Bone mechanics
handbook (second edition), Boca Raton, Ed: CRC Press (2001).
Day J.S., Ding M., Odgaard A., Sumner D.R
Parallel plate model for trabecular bone exhibits volume fraction-
dependent bias , Bone, 27 (5), 715-720 (2000).
231

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
[Ding, 1999] Ding M., Odgaard A., Hvid I., Accuracy of cancellous bone
volume fraction measured by micro-CT scanning, J. Biomechanics,
32, 323-326 (1999).
[Drake, 2002] Drake W.M., Brown J.P., Banville C., Kendler D.L., Use of
phalangeal bone mineral density and multi-site speed of sound
conduction to monitor therapy with alendronate in postmenopausal
women, Osteoporosis Int., 13, 249-256 (2002).
Dupuy C[Dupuy, 2000] ., L’ostéoporose ou comment prévenir une maladie du
vieillissement, Dossier réalisé pour le site web de la Fondation
pour la Recherche Médicale (www.frm.org), (2000).
Durand E., Imagerie de la peau humai[Durand, 1997] ne, in vivo, en haute
[Edelsbrunner, 1994]
[Farrar, 1971] concepts in NMR et Relaxation
[Feldkamp, 1988]
media, Phys. Rev. B, 37 (7), 3448-3453 (1988).
[Frigo, 1998] Software
résolution spatiale, par résonance magnétique, Thèse, Université
Paris VI, Paris, France (1997).
Edelsbrunner H., Mucke E. P., Three-dimensional alpha shapes,
ACM Trans. Graphics, 13, 43-72 (1994).
[Engelke, 1994] Engelke K., Majumdar S., Genant H.K., Phantom studies
simulating the impact of trabecular structure on marrow relaxation
time T2’, MRM, 31, 380-387 (1994).
Farrar T.C., Becker E.D., Basic
mechanisms dans Pulse and Fourier transform NMR, Orlando, Ed:
Academic Press (1971).
Feldkamp L.A., David L.C., Topology and elastic properties of
depleted
[Ford, 1993] Ford J.C., Wehrli F.W., Chung H.W., Magnetic field distribution
in models of trabecular bone, MRM, 30, 373-379 (1993).
Frigo M., Johnson S.G., FFTW: An Adaptive
Architecture for the FFT, ICASSP Conference proceedings, 3,
1381-1384 (1998).
Algorithme disponible en « freeware » sur leur site web:
www.fftw.org.
232

Bibliographie
[Gardner, 2001] Gardner J.R., Hess C.P., Webb A.G., Tsika R.W., Dawson M.J.,
Gulani V., Magnetic resonance microscopy of morphological
alterations in mouse trabecular bone structure under conditions of
[Genant, 1996]
d structure: state
[Gibson, 1985]
]
[Gonzalez, 1993]
processing, New York, Ed: Addison Wesley (1993).
7]
4]
[Gudbjartsson, 1995]
10-914 (1995).
[Haacke, 1999] R., Venkatesan R.,
[Harrigan, 1984]
nsor,
Journal of Materials Science, 19, 761-767 (1984).
simulated microgravity, MRM, 45, 1122-1125 (2001).
Genant H.K, Engelke K., Fuerst T., Gluer C.C., Grampp S., Harris
S.T., Jergas M., Lang T., Lu Y., Majumdar S., Mathur A., Takada
M., Non invasive assessment of bone mineral an
of the art, J. Bone Miner. Res., 11, 707-729 (1996).
Gibson L.J., The mechanical behaviour of cancellous bone, J.
Biomechanics, 18 (5), 317-328 (1985).
[Ginefri, 2001 Ginefri J-C., Darrasse L., Crozat P.A., High-temperature
superconducting surface coil for in vivo micro imaging of the
human skin, MRM 45, 376-382 (2001).
Gonzalez R.C., Woods E.R., Representation and description dans
Digital image
[Gordon, 199 Gordon C.L., Webber C.E., Christofou N., Nahmias C., In vivo
assessment of trabecular bone structure at the distal radius from
high resolution magnetic resonance images, Med. Phys., 24 (4),
585-593 (1997).
[Gourret, 199 Gourret J.P., Pipeline graphique dans Modélisation d’images fixes
et animées, Paris, Ed: Masson (1994).
Gudbjartsson H., Patz S., The rician distribution of noisy MRI
data, MRM, 34, 9
[Guo, 2002] Guo X.E., Kim C.H., Mechanical consequence of trabecular bone
loss and its treatment: a three-dimensional model simulation, Bone,
30 (2), 404-411 (2002)
Haacke E.M., Brown R.W., Thompson M.
Magnetic resonance imaging: physical principles and sequence
design, New York, Ed: Wiley-Liss (1999).
Harrigan T.P., Mann R.W., Characterization of microstructural
anisotropy in orthotropic materials using second rank te
233

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
[Hildebrand, 1997]
[Hildebrand, 1999] ger P.,
[Hordon, 2000]
cular architecture in women and men of similar
[Hwang, 1997]
from three-dimensional nuclear magnetic
[Hwang, 1999]
mages
[Hwang, 2002]
bone, MRM, 47, 948-957 (2002).
[Kaufman, 1988]
[Kim, 2002] of vertebral
trabecular bone, Journal of Biomechanics, 35, 1101-1114 (2002).
Hildebrand T., Rüegsegger P., A new method for the
model-independent assessment of thickness in three-dimensional
images, J. Microscopy, 185 (1), 67-75 (1997).
Hildebrand T., Laib A., Müller R., Dequeker J., Rüegseg
Direct three-dimensional morphometric analysis of human
cancellous bone: microstructural data from spine, femur, iliac
crest, and calcaneus, J. Bone Miner. Res., 14 (7), 1167-1174
(1999).
[Hipp, 1996] Hipp J.A., Jansujwicz A., Simmons C.A., Snyder B.D., Trabecular
bone morphology from micro-magnetic resonance imaging, J.
Bone Miner. Res., 11 (2), 286-292 (1996).
Hordon L.D., Raisi M., Aaron J.E., Paxton S.K., Beneton M.,
Kanis J.A., Trabe
bone mass with and without vertebral fractures: 1.
Two-dimensional histology, Bone, 27 (2), 271-276 (2000).
Hwang S.N., Wehrli F.W., Williams J.L., Probability-based
structural parameters
resonance images as predictors of trabecular bone strength, Med.
Phys. 24 (8), 1255-1261 (1997).
Hwang S.N., Wehrli F.W., Estimating voxel volume fractions of
trabecular bone on the basis of magnetic resonance i
acquired in vivo, Int. J. Imaging Syst. Technol., 10, 186-198
(1999).
Hwang S.N., Wehrli F.W., Subvoxel processing: a method for
reducing partial volume blurring with application to in vivo MR
images of trabecular
[Inserm, 1996] Ostéoporose, stratégies de prévention et de traitement, Paris, Les
Editions INSERM (1996).
Kaufman A., Efficient algorithms for scan-converting 3D
polygons, Comp. Graphics, 12, 213-129 (1988).
Kim H.S., Al-Hassani S.T.S., A morphological model
234

Bibliographie
[Kinney, 1998] Kinney J.H., Ladd A.J.C., The relationship between
three-dimensional connectivity and the elastic properties of
trabecular bone, J. Bone Miner. Res., 13 (5), 839-845 (1998).
Kleerekoper M., Villanueva A.R., Stanciu J., Sudhaker Ra[Kleerekoper, 1985] o D.,
[Kothari, 1998] t H.K.,
[Laib, 2001]
m MR images, 37-46
[Last, 2001] stimation de paramètres
[Laugier, 1997] ntet A.M., Berger G., Assessment
[Legrand, 2000]
r bone
Parfitt A.M., The role of three-dimensional trabecular
microstructure in the pathogenesis of vertebral compression
fractures, Calcif. Tissue Int., 37, 594-597 (1985).
Kothari M., Keaveny T.M., Lin J.C., Newitt D.C., Genan
Majumdar S., Impact of spatial resolution on the prediction of
trabecular architecture parameters, Bone, 22 (5), 437-443 (1998).
Laib A., Beuf O., Issever A., Newitt D.C., Majumdar S., Direct
measures of trabecular bone architecture fro
dans Majumdar S., Kay B., Non invasive assessment of trabecular
bone architecture and the competence of bone, Ed: Kluwer Acad.
Press (2001).
[Laib, 2002] Laib A., Newitt D.C., Lu Y., Majumdar S., New
model-independent measures of trabecular bone structure applied
to in vivo high-resolution MR images, Osteoporosis Int., 13 (2),
130-136 (2002).
Last D., Guillot G., Amélioration de l’e
topologiques sur des images 3D d’os trabéculaire obtenues par
microscopie IRM, Récents progrès en génie des procédés, 78,
195-200 (2001).
Laugier P., Droin P., Laval-Jea
of the relationship between acoustic properties and bone mass
density of the calcaneus by comparison of ultrasound parametric
imaging and QCT, Bone, 20, 157-165 (1997).
Legrand E., Chappard D., Pascaretti C., Du Quenne M., Krebs S.,
Rohmer V., M.F.Basle, Audran M., Trabecula
microarchitecture, bone mineral density, and vertebral fractures in
male osteoporosis, J. Bone Miner. Res., 15 (1), 13-19 (2000).
235

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
[Link, 1998 (a)]
2-132 (1998).
[Link, 2002]
S., Changes in calcaneal trabecular
aluation of technical factors
[Majumdar, 1996]
c microscopy and biomechanics, Osteopor. Int.,
[Majumdar, 1997]
d osteoporotic status: in vivo studies
Link T.M., Majumdar S., Lin J.C., Newitt D., Augat P., Ouyang
X., Mathur A., Genant H.K, A comparative study of trabecular
bone properties in the spine and the femur using high resolution
MRI and CT, J. Bone Miner. Res., 13 (1), 12
[Link, 1998 (b)] Link T.M., Majumdar S., Augat P., Lin J.C., Newitt D., Lu Y.,
Lane N.E., Genant H.K., In vivo high resolution MRI of the
calcaneus: differences in trabecular structure in osteoporosis
patients, J. Bone Miner. Res., 13 (7), 1175-1182 (1998).
Link T.M., Saborowski K.K., Kempkes M., Kosch M., Newitt D.,
Lu Y., Waldt S., Majumdar
bone structure assessed with high-resolution MR imaging in
patients with kidney transplantation, Osteoporosis Int., 13 (2),
119-129 (2002).
[Link, 2003] Link T.M., Vieth V., Stehling C., Lotter A., Beer A., Newitt D.,
Majumdar S., High-resolution MRI vs multislice spiral CT: which
technique depicts the trabecular bone structure best ?, Eur. Radiol.,
13, 663-671 (2003).
[Majumdar, 1995] Majumdar S., Newitt D., Jergas M., Gies A., Chiu E., Osman D.,
Keltner J., Keyak J., Genant H., Ev
affecting the quantification of trabecular bone structure using
magnetic resonance imaging, Bone, 17 (4), 417-430 (1995).
Majumdar S., Newitt D.C., Mathur A., Osman D., Gies A., Chiu
E., Lotz J., Kinney J., Genant H., Magnetic resonance imaging of
trabecular bone structure in the distal radius: relationship with
x-ray tomographi
6, 376-385 (1996).
Majumdar S., Genant H.K., Grampp S., Newitt D.C., Truong V.H.,
Lin J.C., Mathur A., Correlation of trabecular bone structure with
age, bone mineral density, an
in the distal radius using high resolution magnetic resonance
imaging, J. Bone Miner. Res., 12 (1), 111-118 (1997).
236

Bibliographie
[Majumdar, 1998]
one, 22 (5), 445-454 (1998).
(3), 231-239 (1999).
[Morgan, 2003] eaveny T.M., Trabecular bone
[Müeller, 1998]
n bone biopsies: a quantitative structural
[Mulder, 2000] J., Michaeli D.A., Siris E., Comparison of bone mineral
ivo assessment of architecture and micro-finite element
[Nicholson, 2001] n P.H.F., Müller R., Cheng X.G., Rüegsegger P., Van Der
Majumdar S., Kothari M., Augat P., Newitt D.C., Link T.M., Lin
J.C., Lang T., Lu Y., Genant H.K., High-resolution magnetic
resonance imaging: three-dimensional trabecular bone architecture
and biomechanical properties, B
[Majumdar, 1999] Majumdar S., Link T.M., Augat P., Lin J.C., Newitt D., Lane N.E.,
Genant H.K., Trabecular bone architecture in the distal radius
using magnetic resonance imaging in subjects with fractures of the
proximal femur, Osteoporosis Int., 10
[Michaeli, 1998] Michaeli D.A., Mirshahi A., Singer J., Rapa F.G., Plass D.B.,
Bouxsein M.L., A new X-ray based osteoporosis screening tool
provides accurate and precise assessment of phalanx bone mineral
content, J. Clin. Densitom., 2, 23-30 (1998).
Morgan E.F., Bayraktar H.H., K
modulus-density relationships depend on anatomic site, J.
Biomechanics., 36, 897-904 (2003).
Müller R., Van Campenhout H., Van Damme B., Van Der Perre
G., Dequeker J., Hildebrand T., Rüegsegger P., Morphometric
analysis of huma
comparison of histological sections and micro-computed
tomography, Bone, 23 (1), 59-66 (1998).
Mulder
density of the phalanges, lumbar spine, hip and forearm for the
assessment of osteoporosis in postmenopausal women, J. Clin.
Densitom., 3, 373-381 (2000).
[Newitt, 2002] Newitt D.C., Majumdar S., Van Rietbergen B., Von Ingersleben
G., Harris S.T., Genant H.K., Chesnut C., Garnero P., MacDonald
B., In v
analysis derived indices of mechanical properties of trabecular
bone in the radius, Osteoporosis Int., 13 (1), 6-17 (2002).
Nicholso
Perre G., Dequeker J., Boonen S., Quantitative Ultrasound and
Trabecular Architecture in the Human Calcaneus, J. Bone Mineral
Res., 16 (10), 1886-1892 (2001).
237

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
[Njeh, 1999] Njeh C.F., Boivin C.M., Gough A., Hans D., Srivastav S.K.,
Bulmer N., Devlin J., Emery P., Evaluation of finger ultrasound in
the assessment of bone status with application of rheumatoid
arthritis, Osteoporosis Int., 9, 82-90 (1999).
Alexandre C., Peyrin F., Synchrotron radiation
of etidronate treatment, J. Bone Miner.
[Odgaard, 1993]
sis on 3-D reconstructions,
[Oleksik, 2000]
al density with
[O’Rourke, 2001] mputational Geometry in C, Cambridge, Ed:
[Ouyang, 1997]
t H. K., High
metry measurements, Calcif. Tissue Int., 60 (2),
[Parfitt, 1987]
enclature, symbol and units, J. Bone Miner.
[Pateyron, 1998]
rotron, Thèse, INSA, Lyon, France
(1998).
[Nuzzo, 2002] Nuzzo S., Lafage-Proust M.H., Martin-Badosa E., Boivin G.,
Thomas T.,
microtomography allows the analysis of three-dimensional
microarchitecture and degree of mineralization of human iliac crest
biopsy specimens: effects
Res., 17, 1372-1382 (2002).
Odgaard A., Gundersen J.G., Quantification of connectivity in
cancellous bone with special empha
Bone, 14, 173-182 (1993).
Oleksik A., Ott S.M., Vedi S., Bravenboer N., Compston J., Lips
P., Bone structure in patients with low bone miner
or without vertebral fractures, J. Bone Miner. Res., 15 (7),
1368-1375 (2000).
O’Rourke J., Co
Cambridge University Press (2001).
Ouyang X., Selby K., Lang P., Engelke K., Klifa C., Fan B.,
Zucconi F., Hottya G., Chen M., Majumdar S., Genan
resolution Magnetic Resonance Imaging of the calcaneus: age-
related changes in trabecular structure and comparison with Dual
X-Ray Absorptio
139-147 (1997).
Parfitt A.M., Drezner M.K., Glorieux F.H., Kanis J.A., Malluche
H., Meunier P.J., Ott S.M., Recker R.R., Bone histomorphometry:
standardization of nom
Res., 2 (6), 595-610 (1987).
Pateyron M., Acquisition et quantification d’images du réseau
trabéculaire osseux en microtomographie tridimensionnelle
utilisant le rayonnement synch
238

Bibliographie
[Peyrin, 1998] Peyrin F., Salome M., Cloetens P., Laval-Jeantet A.M., Ritman E.,
Ruegsegger P., Micro-CT examinations of trabecular bone samples
at different resolutions: 14, 7 and 2 micron leve, Technol. Health
[Pothuaud, 2000]
f trabecular bone projection
[Pothuaud, 2002 (a)]
esolution microcomputed tomography, J. Bone Miner.
[Pothuaud, 2002 (b)]
dicts the
[Press, 1989] .H., Teukolsky S.A., Vetterling W.T., Flannery B.P.,
[Rice, 1988]
apparent density, J.
[Rotter, 2001]
[Ruaud, 1991] t réalisation d’un imageur RMN à très haute
[Saha, 2000] Saha P.K., Gomberg B.R., Wehrli F.W., Three-dimensional digital
topological characterization of cancellous bone architecture, Int. J.
Imaging Syst. Technol., 11, 81-90 (2000).
Care, 6 (5-6), 391-401 (1998).
Pothuaud L., Benhamou C.L., Porion P., Lespessailles E., Harba
R., Levitz P., Fractal dimension o
texture is related to three-dimensional microarchitecture, J. Bone
Miner. Res., 15 (4), 691-699 (2000).
Pothuaud L., Laib A., Levitz P., Benhamou C.L., Majumdar S.,
Three-dimensional-line skeleton graph analysis of high-resolution
magnetic resonance images: a validation study from
34-µm-r
Res., 17 (10), 1883-1895 (2002).
Pothuaud L., Van Rietbergen B., Mosekilde L., Beuf O., Levitz P.,
Benhamou C.L., Majumdar S., Combination of topological
parameters and bone volume fraction better pre
mechanical properties of trabecular bone, J. Biomech., 35,
1091-1099 (2002).
Press W
Numerical recipes in C, Londres, Ed: Cambridge University Press
(1989).
Rice J.C., Cowin S.C., Bowman J.A., On the dependence of the
elasticity and strength of cancellous bone on
Biomechanics, 21 (2), 155-168 (1988).
Rotter M., Berg A., Langenberger H., Grampp S., Imhof H., Moser
E., Autocorrelation analysis of bone structure, JMRI, 14, 87-93
(2001).
Ruaud J-P, Etude e
résolution spatiale, Thèse, Université Paris Sud, Orsay, France
(1991).
239

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
[Salomé, 1999]
]
[Selby, 1996] H.K., Investigation of
[Serra, 1993] rameters and set models dans Image
analysis and mathematical morphology (volume 1) 4ème édition,
Londres, Ed: Academic Press (1993).
[Simmons, 1997] Simmons C.A., Hipp J.A., Method-based differences in the
automated analysis of three-dimensional morphology of trabecular
bone, J. Bone Miner. Res., 12 (6), 942-947 (1997).
[Sugita, 1999] Sugita H., Oka M., Toguchida J., Nakamura T., UEO T., Hayami
T., Anisotropy od osteoporotic cancellous bone, Bone, 24 (5),
513-516 (1999).
[Thomsen, 1996] Thomsen J.S., Barlach J., Mosekilde L., Determination of
connectivity density in human iliac crest bone biopsies assessed by
computerized method, Bone, 18 (5), 459-465 (1996).
[Thomsen, 2002 (a)] Thomsen J.S., Ebbesen N., Mosekilde L., Predicting human bone
strength by vertebral static histomorphometry, Bone, 30 (3),
502-508 (2002).
[Thomsen, 2002 (b)] Thomsen J.S., Ebbesen N., Mosekilde L., Age-related differences
between thinning of horizontal and vertical trabeculae in human
lumbar bone as assessed by a new computerized method, Bone, 31
(1), 136-142 (2002).
[Uchiyama, 1999] Uchiyama T., Tanizawa T., Muramatsu H., Endo N., Takahashi
H.E., Hara T., Three-dimensional microstructural analysis of
human trabecular bone in relation to its mechanical properties,
Bone, 25 (4), 487-491 (1999).
Salomé M., Peyrin F., Cloetens P., Odet C., Laval-Jeantet A.M.,
Baruchel J., Spanne P., A synchrotron radiation microtomography
system for the analysis of trabecular bone samples, Med. Phys., 26
(10), 2194-2204 (1999).
[Schild, 1990 Schild H.H., MRI made easy (…well almost), Berlin, Ed: Schering
AG (1990).
Selby K., Majumdar S., Newitt D.C., Genant
MR decay rates in microphantom models of trabecular bone,
JMRI, 6, 549-559 (1996).
Serra J., Morphological pa
240

Bibliographie
[Ulrich, 1999] Ulrich D., Van Rietbergen B., Laib A., Ruegsegger P., The ability
of three-dimensional structural indices to reflect mechanical
aspects of t e, 25 (1), 55-60 (1999)
[Van Der Linden, 2001] Van Der Linden J.C., Homminga J., Verhaar J.A.N., Weinans H.,
Mechanical consequences of bone loss in cancellous bone, J. Bone
Miner Res., 16 (3), 457-465 (2001).
[Van Rietbergen, 2002] Van Rietbergen B., Majumdar S., Newitt D., MacDonald B.,
High-resolution MRI and micro-FE for the evaluation of changes
in bone mechanical properties during longitudinal clinical trials:
application to calcaneal bone in postmenopausal women after one
year of idoxifene treatment, Clinical Biomechanics, 17, 81-88
(2002).
high-resolution MRI of the calcaneus reflect the true bone
structure?, Investigative Radiology, 36, 210-217 (2001).
(1997).
[Wehrli, 1998] Wehrli F.W., Hwang S.N., Ma J.M., Song H.K., Ford J.C., Haddad
noninvasive assessment with MR microimaging and image
processing, Radiology, 206, 347-357 (1998).
Snyder P.J., Digital topological analysis of in vivo magnetic
resonance microimages of trabecular bone reveals structural
implications of osteoporosis, J. Bone Miner. Res., 16 (8),
1520-1531 (2001).
[Whitehouse, 1974] Whitehouse W.J., The quantitative morphology of anisotropic
trabecular bone, Journal of Microscopy, 101 (2), 153-168 (1974).
rabecular bone, Bon
[Vieth, 2001] Vieth V., Link T.M., Lotter A., Persigehl T., Newitt D., Heindel
W., Majumdar S., Does the trabecular bone structure depicted by
[Vogel, 1997] Vogel H.J., Digital unbiased estimation of the Euler Poincare
characteristic in different dimensions, Acta Stereol., 16/2, 97-104
J.G., Cancellous bone volume and structure in the forearm:
[Wehrli, 2001] Wehrli F.W., Gomberg B.R., Saha P.K., Song H.K., Hwang S.N.,
241

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
[Wren, 2000] Wren T.A.L., Yerby S.A., Beaupre G.S., Carter D.R.,
Interpretation of calcaneus dual-energy X-ray absorptiometry
measurements in the assessment of osteopenia and fracture risk, J.
Bone Miner. Res., 15 (8), 1573-1578 (2000).
[Wu, 1994] Wu Z., Chung H-W., Wehrli F.W., A Bayesian approach to
subvoxel tissue classification in NMR microscopic images of
trabecular bone, MRM, 31, 302-308 (1994).
[Yablonskiy, 1994] Yablonskiy D.A., Haacke E.M., Theory of NMR signal behavior in
magnetically inhomogeneous tissues: the static dephasing regime,
MRM, 32, 749-763 (1994).
242

Annexes
Annexes
rientation de la microstructure: calcul des trois angles
Annexe 3: Tracé de segments dans un espace discrétisé ...............253
Annexe 1: Calcul de la caractéristique d’Euler-Poincaré .............245
Annexe 2: Od’Euler (θ,ϕ,ψ) ................................................................................249
Annexe 4: Liste des publications liées à ce travail .........................259
243


Annexe 1: Calcul de la caractéristique d’Euler-Poincaré
Annexe 1: Calcul de la caractéristique d’Euler-Poincaré
prenant en compte les effets de bord. En effet, les images étant nécessairem
ure et le problème des frontières se pose donc.
Le principe de calcul est basé sur une formule récurrente liant la caractéristique
d’Euler-Poincaré dans un espace de dimension n à celle dans un espace de dimension n-1.
dans la dimension 0 qui correspond à un simple comptage d’éléments d’un
ipe de l’algorithme. Pour plus de détails sur
) Cas « 1D »
L’algorithme de calcul de la caractéristique d’Euler-Poincaré proposé par Vogel
([Vogel, 1997]) permet d’obtenir une estimation non biaisée de ce paramètre, c’est-à-dire en
ent bornées, elles
ne permettent pas d’accéder à toute la struct
Ainsi, quelle que soit la dimension, le calcul revient à celui de la caractéristique
d’Euler-Poincaré
ensemble.
Cette annexe a pour objectif de présenter le princ
sa mise en œuvre nous renvoyons le lecteur à l’article.
A
oit un objet X1 défini par un ensemble de points discrets dans un espace E de dimension 1
(cf. Figure A.1).
S
x0 x10x1 x2 x3 x4 x5 x6 x7 x8 x9
Figure A.1: Exemple d’un objet X1 en dimension 1, défini comme un ensemble de points de N:
X1=x2,x3,x4,x6,x8,x9.
La caractéristique d’Euler-Poincaré de dimension 1, N1(X1) est égale à la différence entre le
nombre de sommets (c’est-à-dire le nombre d’éléments de X1) et le nombre de segments de
(c’est-à-dire le nombre de couple d’éléments consécutifs de X1).
On peut donc écrire:
Eq. A.1
où N0(X) est la caractéristique d’Euler-Poincaré en dimension 0 de l’ensemble X (i.e. son
cardinal), et , l’ensemble défini par:
)ˆ()()( 101011 XNXNXN −=
1X
111 /ˆ XxxxX iii ⊂∪= + .
représente bien le nombre de segments (liens) contenu dans X1. Sur l’exemple de la
Figure A.1, on aurait, N1(X1) = 6 - 3 = 3.
)ˆ( 10 XN
245

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
246
On considère dans la suite une partition de E composée d’ensembles adjacents Ωi tels que la
frontière Fi commune entre Ωi et Ωi+1 soit identique:
Ωi ∩ Ωi+1 = Fi
La Figure A.2 représente trois ensembles adjacents Ω1, Ω2 et Ω3 de cette partition ainsi que
les deux frontières communes Fi-1 et Fi associées.
Ω1 Ω3
Ω2
F1 F2F0 F3
Figure A.2: Exemple de trois ensembles adjacents Ω1, Ω2 et Ω3 ainsi que les frontières communes Fi
associées.
Sur la partition de E, N1(X1) se détermine de la façon suivante:
.
D’après la propriété d’additivité de la caractéristique d’Euler-Poincaré, on peut donc écrire:
Pratiquement, X1 n’est pas accessible entièrement: on ne le connaît que sur un certain
ensemble ΩI. (qui constitue l’« image ») On ne peut alors calculer N1(X1) mais seulement la
contribution de ΩI à N1(X1) que l’on note
)()( 1111 Ui
iXNXN Ω∩=
∑∑ ∩−Ω∩=i
ii
i FXNXNXN )()()( 111111
)(~11 IXN Ω∩
)
. Afin de prendre en compte les effets
de bord, l’auteur propose de calculer (~11 IXN Ω∩ de la manière suivante:
)(21)()(~
111111 III BXNXNXN ∩−Ω∩=Ω∩ Eq. A.2
avec:
,
où BI correspond aux deux frontières de ΩI.
En utilisant Eq. A.1 pour exprimer
III FFB ∪= −1
)(~11 IXN Ω∩ (Eq. A.2) et en remarquant que
est forcément nul (du fait que BI correspond à la réunion de deux points disjoints), on obtient
donc:
)( 10
∧
∩ IBXN
)(21)()()(~
10101011 IIII BXNXNXNXN ∩−Ω∩−Ω∩=Ω∩∧
Eq. A.3

Annexe 1: Calcul de la caractéristique d’Euler-Poincaré
B) Généralisation à la dimension n
un objet défini dans un espace de dimension n. Si on suppose que l’on a accès au
partition Ω de l’espace, l’estimateur proposé
Soit Xn
champ de vue ΩI d’une pour la contribution de
ΩI à la caractéristique d’Euler-Poincaré en dimension n est, d’après Eq. A.3:
)(2 ,1,11 jInnIjnnInnInn −−−
où l’ensemble jX est défini par:
(~ XN 1)()() BXNXNXN ∩−Ω∩−Ω∩=Ω∩∧
XxxxX ij=ˆejii ⊂∪ +/ ,
i étant un vecteur de dimension n, et ej un vecteur unitaire dans la direction j, choisie
ent),
n’importe quelle dimension la contribution de ΩI à la
caractéristique d’Euler-Poinc
(x
arbitrairem
La frontière jIB , du champ de vue ΩI est aussi définie par rapport à la direction ej analysée:
elle correspond à un ensemble de dimension n-1.
On peut ainsi calculer dans
aré.
247


Annexe 2: Orientation de la microstructure: calcul des trois angles d’Euler (θ,ϕ,ψ)
Annexe 2: Orientation de la ic m rostructure: calcul des
,
on en exposant sur les colonnes ou les matrices fait référence à la
base orthonormée directe associée au référentiel de
sera notée B dans la suite.
ls:
La relation matricielle entre un vecteur
trois angles d’Euler (θ,ϕ ψ)
Remarque préalable:
Dans ce qui suit, la notati
base dans laquelle ils s’expriment. Larrr
l’image ),,( kji
Rappe
Xr
exprimé dans une base α, Xα, et ce même vecteur
exprimé dans une base β, Xβ, est basée sur la matrice de changement de base inversible que
l’on notera dans la suite Pα→β:
Xα = Pα→β.Xβ ou Xβ = (Pα→β)-1.Xα.
α β φ autour d’un des
φ e la ro α. On a alors:
(M(φ) ) = (M(φ)α)=M(-φ)α,
où « TM » correspond à la transposée de M.
d’Euler
De plus, si on passe de la base à la base par une rotation d’angle
vecteurs de la base α, on peut facilement montrer que:
Pα→β=M(φ)α,
où M( )α est la matrice d tation exprimée dans la base -1 α T
A) Définition des angles
Dans un espace de dimension 3, on peut passer d’une base orthonormée directe à une autre à
1 2
uivan
r de : on pose
partir d’une rotation R qui peut s’exprimer comme la composition de trois rotations R , R et
R3 définies de la manière s te ([Gourret, 1994]):
• R1, la rotation de θ autou kr
11 )( iiRrr
= et 11 )( jjRrr
=
• R2, la rotation de ϕ autour de : on pose 1ir
212 )( jjRrr
= k et R 22 )( kr r=
3• R , la rotation de ψ autour de kr
2
249

Caractérisation de l’architecture tridim ire par IRM ensionnelle du tissu trabécula
250
(θ,ϕ,ψ) sont les trois angles d’Euler. Sur la Figur
un vecteur et son image par R:
e A.3, on a représenté ces trois rotations pour
Vr
RVr
)(VRVR
rr= . On pose aVVR
rr=)(1 et : on a donc
ba VVRrr
=)(2
Rb VVRrr
=)(3 .
jr
Vr
aVr
kr
θ
ir
1jr
kr
1ir
aVr
bVr
ϕ
2j
r
1
2kir
r
bVr
RVr
ψ
R1(θ) R2(ϕ) R3(ψ)
Figure A.3: Définition des trois rotations R1, R2 et R3 permettant de déterminer les trois angles d’Euler
(θ,ϕ,ψ).
On note dans la suite B1 et B2 les bases ),,( 11 kjirrr
et ),,( 221 kjirrr
.
B) Expression de la matrice de rotation de R
D’après la Figure A.3, on peut facilement établir les expressions matricielles de ces trois
rotations respectivement dans les bases B, B1, et B2:
( )( ) ( )( ) ( )
B
BRM
⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢
⎣
⎡ −
=
100
0cossin
0sincos
1 θθ
θθ
θ
On cherche à exprimer la matrice de la rotation R dans la base B, MRB, telle que:
VRB = MR
B.VB,
(V et VR étant les colonnes associées aux vecteurs
, ( ) ( ) ( )( ) ( )
1
12
cossin0
sincos0
001 B
BRM
⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢
⎣
⎡
−=
ϕϕ
ϕϕϕ , ( )( ) ( )( ) ( )
2
23
100
0cossin
0sincos B
BRM
⎥⎥⎥⎥
⎦
⎤
⎢⎢⎢⎢
⎣
⎡ −
= ψψ
ψψ
ψ .
Vr
et RVr
, cf. Figure A.3).
Il faut donc composer les trois rotations dont les matrices sont données ci-dessus, en veillant
bien à respecter les bases dans lesquelles elles sont exprimées. On peut représenter les
différentes étapes pour passer de VB à VRB de la manière suivante:
,
(Va et Vb étant les colonnes associées aux vecteurs
BR
BBBR
BBBR
RBb
BBBb
RBaBBBa
RB VVVVVVVV ⎯→⎯⎯→⎯⎯→⎯⎯→⎯⎯→⎯⎯→⎯⎯→⎯ →→→→ 11122322112111
aVr
et bVr
, cf. Figure A.3).
On a donc:
VRB = PB→B1.VR
B1 = PB→B1.PB1→B2.VRB2 = PB→B1.PB1→B2.MR3(ψ)B2.Vb
B2

Annexe 2: Orientation de la microstructure: calcul des trois angles d’Euler (θ,ϕ,ψ)
= PB→B1.PB1→B2.MR3(ψ)B2.(PB1→B2)-1.VbB1
= PB→B1.PB1→B2.MR3(ψ) .(PB1→B2) .MR2(ϕ) .(PB→B1) .Va
-1.MR2(ϕ)B1.(PB→B1)-1.MR1(θ)B.VB
ϕ)-1)B1.MR2(ϕ)B1.(MR1(θ)-1)B.MR1(θ)B.VB
VRB = MR1(θ)B.MR2(ϕ)B1.M (ψ)B2.VB
Donc:
MRB = MR1(θ)B.MR2(ϕ)B1.MR3(ψ)B2
= PB→B1.PB1→B2.MR3(ψ)B2.(PB1→B2)-1.MR2(ϕ)B1.VaB1
B2 -1 B1 -1 B
= PB→B1.PB1→B2.MR3(ψ)B2.(PB1→B2)
En remplaçant les matrices de changement de base, on a:
VRB = MR1(θ)B.MR2(ϕ)B1.MR3(ψ)B2.(MR2(
Soit après simplification,
R3
Soit:
( ) ( ) ( ) ( ) ( ) ( ) ( ) ( ) ( ) ( ) ( ) ( )( ) ( ) ( )
B
BM
⎤
⎢⎢⎡
+
−−−
= ψθ
ϕθψϕθψθψϕθψθ
coscos.sin
sin.sincos.cos.sinsin.cossin.cos.sincos.cos
( ) ( ) ( ) ( ) ( ) ( ) ( ) ( ) ( )( ) ( ) (ϕsin ) ( ) ( )
R
⎥⎥⎥⎥
⎦⎢⎢
−+−
ϕψψϕ
ϕθψϕθψθψϕθ
coscos.sin.sin
sin.coscos.cos.cossin.sinsin.cos.
ois angles d’Euler pour la microstructure osseuse
⎣
C) Calcul des tr
Après la diagonalisation de la matrice ME, tenseur du deuxième ordre décrivant l’anisotropie,
on dispose donc, pour les trois valeurs propres (λ1,λ2,λ3), des coordonnées des trois vecteurs
propres normalisés dans la base B:
et .
On suppose de plus que
),,( 321 VVVrrr
B
z
yx
B
VVV
V⎥⎥⎦
⎤
⎢⎢⎣
⎡=
11
1
1 , B
z
yx
B
VVV
V⎥⎥⎦
⎤
⎢⎢⎣
⎡=
22
2
2
B
z
yx
B
VVV
V⎥⎥⎦
⎤
⎢⎢⎣
⎡=
33
3
3
321 VVVrrr
=∧ : ),,( 321 VVVrrr
forment donc une base orthonormée directe.
Pour déterminer la position angulaire de la base des vecteurs propres par rapport
au référentiel de l’image , on cherche à caractériser la rotation R (c’est-à-dire les trois
),,( 321 VVVrrr
),,( kjirrr
angles θ, ϕ, et ψ) telle que:
, 3)( VkRrr
= 1)( ViRrr
= , 2)( VjRrr
=
Soit matriciellement (en posant IB, JB et KB, les colonnes associées respectivement aux
ecteurs et , exprimées dans la base B):
MRB.IB = V1
B, MRB.JB = V2
B, MRB.KB = V3
B
ir
, jr
kr
v
251

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
avec BI ⎥⎦
⎢⎣
=00 , BJ ⎥
⎦⎢⎣
=01 et K ⎥
⎦⎢⎣
=10 .
D’après l’expression de M
B⎤⎡1
B⎤⎡0
B
B⎤⎡0
ment, on trouve facilement: B donnée précédemR
⎟⎠⎞
⎜⎝⎛−= y
x
VV
3
3arctanθ ou πθ +⎟⎠⎞
⎜⎝⎛−= y
x
VV
3
3arctan
( )zV3arccos=ϕ ou ( )zV3arccos−=ϕ
⎟⎠⎞⎜
⎝⎛= z
z
VV
2
1arctanψ ou πψ +⎟⎠⎞⎜
⎝⎛= z
z
VV
2
1arctan
Par contre, on ne peut simplement trouver analytiquement un « triplet solution ». D’après les
expressions précédentes, 8 triplets (θ,ϕ,ψ) seraient possibles. MRB est calculée pour chacun de
ces 8 cas: la matrice retenue est celle permettant de retrouver V1B, V2
B et V3B quand on la
multiplie respectivement par IB, JB et KB. Cependant, du fait des égalités suivantes,
R(θ,ϕ,ψ) = R(θ+π,-ϕ,ψ+π)
R(θ,ϕ,ψ+π) = R(θ+π,-ϕ,ψ)
ϕ,ψ), B
t -½π < θ ≤ ½π.
R(θ,-ϕ,ψ) = R(θ+π,ϕ,ψ+π)
R(θ,-ϕ,ψ+π) = R(θ+π,
il reste une ambiguïté puisque deux triplets valident la condition précédente sur MR . Le
« triplet solution » (θ,ϕ,ψ) est choisi en imposan
252

Annexe 3: Tracé de segments dans un espace discrétisé
253
A) Tracé de segment en dimension 2
Annexe 3: Tracé de segments dans un espace discrétisé
ent de droite reliant les pixels (xOn cherche à tracer le segm 0,y0) et (x1,y1) (x0, y0, x1 et y1
étant des entiers). On supposera x0 < x1.
Pour la suite, on définit la droite qui supporte ce segment:
y = a×x + b avec 01
01
xxyya
−−= et
01
1001
xyxyxb −=
x−.
ette méthode de tracé est la plus intuitive. Elle consiste à effectuer un arrondi pour chaque
où « Arr » est la fonction « arrondie au plus près ».
Sur la Figure A.4, on donne le résultat obtenu en utilisant cette technique pour un exemple:
a = 2.8 et b = 0.
1) Première approche: arrondi simple
C
valeur (entière) xi de x:
yi = Arr(a×xi + b),
x
y
r x de 0 à 3, par la méthode de
l’arrondi simple.
Figure A.4: Représentation du tracé du segment de droite y = 2.8×x pou
Cette méthode de tracé serait totalement insuffisante pour être appliquée dans notre cas.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
254
) Algorithme de Bresenham
e sur la
minimisation de l’« erreur d’approximation ».
A partir du pixel courant Pk, deux pixels sont envisagés pour le pixel suivant Pk+1 en fonction
du coefficient directeur a de la droite: on distingue quatre cas, comme indiqué en Figure A.5.
2
La méthode de Bresenham ([Bresenham, 1965]) est une technique incrémentale basé
x
y
1er octant:0 ≤ a < 1
(x0,y0)8ème octant:
2
-1 ≤ a < 0
ème octant:a ≥ 1
a <
te qui le supporte.
7ème oc ant:t-1
Figure A.5: Présentation des quatre cas possibles pour le tracé d’un segment ayant pour origine (x0,y0) en
fonction du coefficient directeur a de la droi
On résume dans le Tableau A.1, les couples de pixels Pk+1 « candidats » dans chaque
configuration (conformément à la Figure A.6).
Octant 1er 2ème 7ème 8ème
Pk+1 « candidats » NE et E NE et N SE et S SE et E
Tableau A.1: Couple de pixels possibles pour le pixel suivant en fonction du coefficient directeur de la
droite a (cf. Figure A.5). La position des pixels E, N, S, NE et SE est indiquées en Figure A.6.

Annexe 3: Tracé de segments dans un espace discrétisé
255
SE
NE
E Pk
N
S
x
y
Figure A.6: Position des pixels suivants possibles Pk+1 dans le voisinage du pixel courant Pk. Ces pixels sont
référencés à l’aide des « ts cardinaux » (N: nord, E: est, S: sud).
poin
Le pixel Pk+1 retenu (parmi les deux qui sont envisagés) est celui conduisant à la distance
minimum entre la droite réelle et le pixel.
Pour détailler plus précisément le principe de la méthode, on analyse le cas du premier octant
(les autres se déduisant simplement par symétrie). Une illustration est donnée en Figure A.7.
Centres de pixels
vers (x0,y0)
vers (x1,y1) Pk+1 « candidats »
Pixel courantPk
d2
d1
Segment « réel »
y
x NE
E
Figure A.7: Illustration de la méthode de Bresenham pour le cas où a est compris entre 0 et 1 (premier
octant): le pixel suivant est donc soit le pixel situé « est » soit celui situé « nord est ». L’erreur
d’approximation est égale à d2 - d1. Son signe permet de déterminer Pk+1.
Dans ce cas (cf. Tableau A.1), si on pose Pk(xi,yi), les deux pixels Pk+1 « candidats » sont les
pixels situés au « nord est » et à l’« est », c’est-à-dire respectivement (xi+1,yi+1) et (xi+1,yi).
On évalue alors les distances séparant ces deux pixels à la droite réelle (respectivement d1 et
d2 sur la Figure A.7). Le signe de l’erreur d’approximation e(k+1) = d2 - d1, permet de
déterminer Pk+1: si e(k+1) < 0, alors Pk+1 = E, sinon Pk+1 = NE.

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
256
Le résultat du tracé de la A.4) est donnée en Figure A.8. droite précédente (cf. Figure
x
y
Figure A.8: Résultat de la méthode de Bresenham appliquée à la droite présentée en Figure A.4 (cas « 2ème
octant »). Le segment obtenu est donc formé de pixels connectés par les faces ou les sommets (8-connexité).
Cette méthode assure donc la connexion des pixels entre eux au sens de la 8-connexité
(connexion par la face ou le sommet).
B) Tracé de segment en dimension 3
Pour effectuer le tracé du segment de droite reliant les pixels (x0,y0,z0) et (x1,y1,z1) (x0, y0, z0,
x1, y1 et z1 étant des entiers avec x0 < x1), on utilise le tracé des segments 2D correspondant à
deux des trois projections planaires du segment. Ainsi on peut déterminer l’ensemble des
voxels du segment. Cet algorithme qui est illustré en Figure A.9, a été proposé par Kaufman
([Kaufman, 1988]).

Annexe 3: Tracé de segments dans un espace discrétisé
257
x x
y z z0 z1
x1
y1 y0
x0
yi
xi
zi
Projection (2D) dans xOy tracée par la méthode de Bresenham
Projection (2D) dans xOz
A B
1988]). En
), on peut
d
au n xOy et dans le plan xOz. Ces segments 2D
re (x0,z0) et (x1,z1) (B). Chaque voxel du
gment 3D (xi,yi,zi) est calculé de la façon suivante: pour chaque valeur possible de xi (de x0 à x1), on
correspondante de yi et à partir du tracé B, celle qui correspond
tracée par la méthode de Bresenham
Figure A.9: Illustration du tracé de segment 3D par la méthode de Kaufman ([Kaufman,
traçant, par l’algorithme de Bresenham, deux des trois projections planaires (tracé 2D
éterminer l’ensemble des voxels du segment. On a représenté, en A et B, les segments 2D correspondant
x projections du segment 3D respectivement dans le pla
lient respectivement les pixels (x0,y0) et (x1,y1) (A) et les pixels
se
détermine, à partir du tracé A, la valeur
à z . i
D en assurant une connexion des voxels au moins
p s
e cette manière, on construit les segments
ar le sommets (26-connexité).


Annexe 4: Liste des publications liées à ce travail
Annexe 4: Liste des publications liées à ce travail
A) Revue à comité de lecture
AST D., PEYRIN F.,• L GUILLOT G.
Manuscrit en cours de soumission.
agnetic resonance imaging.
iology.
3D microscopy of trabecular bone: accuracy of MR parameters by reference to 3D
synchrotron radiation microtomography in calcaneus samples.
OEUF F., LISSAC M., BRIGUET A., GUILLOT G. • CHOEL L., LAST D., DUB
Trabecular alveolar bone microarchitecture in the human mandible using multislice spin
echo high resolution m
Soumis mi Juillet 2003 à Dentomaxillofacial Rad
B) Conférences avec actes
• LAST D., GUILLOT G., PEYRIN F.
Journal de Radiologie, 83 (10), p. 1474 (2002).
• LAST D., DE ROCHEFORT
r bone images from MR and X-ray
tomography microimages. th
, LAST D., LISSAC M., BRIGUET A., GUILLOT G.
High Resolution MRI of Fresh Mandibular Specimens.
IADR European Section Meeting, Rome 6-8 Septembre 2001.
Journal of Dental Research, 81, p. B267 (2002).
Microscopie IRM de l’os trabéculaire. ème50 Journées Françaises de Radiologie, Paris 19-23 Octobre 2002.
L., PEYRIN F., GUILLOT G.
Automatic match of identical ROIs on trabecula
ESMRMB 19 Annual Meeting, Cannes 22-25 Août 2002.
MAGMA, 15 (1), p. 219 (2002).
• CHOEL L.
259

Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire par IRM
• LAST D., GUILLOT G.
Amélioration de l'estimation de paramètres topologiques sur des images 3D d'os trabéculaire
obtenues par microscopie IRM.
Colloque « Visualisation Image Modélisation », Nancy 15-17 Mai 2001.
Récents Progrès en Génie des Procédés, 15, p. 195-200 (2001).
• GUILLOT G., LAST D., DARRASSE L., SZIMTENINGS M., LANZ T.,HAASE A.
In vivo high-resolution MRI of the finger bone at 7 T.
ISMRM 9th Annual Meeting, Glasgow 21-27 Avril 2001.
Proceedings p. 928.
C) Conférences sans actes
• LAST D., DE ROCHEFORT L., PEYRIN F., GUILLOT G.
3D MR microscopy of trabecular bone: architecture parameters by reference to high
trabéculaire: validité des paramètres d’architecture en IRM 3D
roscopy of trabecular bone: validity of MR parameters by reference to 3D
ography using synchrotron radiation.
-Singapore Biomedical Engineering Symposium, Lyon 11-12 Juin 2002.
resolution microtomography (synchrotron radiation) on matched ROIs.
ICMRM 7th Conference, Snowbird 21-26 Septembre 2003.
• LAST D., DE ROCHEFORT L., PEYRIN F., GUILLOT G.
Microscopie IRM 3D de l’os
par référence à la microtomographie 3D haute résolution (rayonnement synchrotron). ème10 Congrès du GRAMM, Angers 5-7 Février 2003.
• GUILLOT G., LAST D., REMY F., ANTONIADIS T.
The formation factor as a feature of trabecular bone microarchitecture.
6th International Conference on Magnetic Resonance in Porous Media, Ulm 8-12 Septembre
2002.
• LAST D., PEYRIN F., GUILLOT G.
3D mic
microtom
2nd French
260

Annexe 4: Liste des publications liées à ce travail
261
• GUILLOT G., LAST D.
La microscopie IRM de l'os trabéculaire.
Journée GETROA sur la recherche en IRM, 6 Juin 2002.
• LAST D., CHOEL L., LISSAC M., BRIGUET A., DUBOEUF F., BOIVIN G,
GUILLOT G.
Mandibular bone microarchitecture by MRI combined with DEXA measurements.
ICMRM 6P
thP Conference, Nottingham 2-5 Septembre 2001.
UD) Séminaires et autres présentations
• LAST D., GUILLOT G.
Microscopie 3D de l’os trabéculaire: validation de l’IRM par comparaison avec la
microtomographie X.
Séminaire du laboratoire, le 28 Mars 2002.
• LAST D., GUILLOT G.
Le réseau trabéculaire osseux: analyse des données obtenues sur différents sites par
différentes techniques d’imagerie.
Journée annuelle du GDR 2237, le 20 Mars 2001.
• LAST D., GUILLOT G.
Calcul de paramètres topologiques sur le réseau trabéculaire osseux.
Journée annuelle du GDR 2237, le 7 Juillet 2000.
UE) Rédaction d’article en cours
• LAST D., DE ROCHEFORT L., PEYRIN F., GUILLOT G.
Automatic match of ROIs on trabecular bone images from MRI an X-ray microtomography.

Titre: Caractérisation de l’architecture tridimensionnelle du tissu trabéculaire in vitro et in vivo par IRM Résumé: Le tissu osseux se présente sous deux formes, cortical ou trabéculaire. Le tissu cortical est un tissu compact formant l’enveloppe résistante de l’os. Le tissu trabéculaire est un tissu poreux constitué de travées osseuses d’une centaine de microns d’épaisseur, formant un espace interconnecté empli de moelle. Ce tissu est un site privilégié pour détecter l’ostéoporose. En raison de sa capacité à délivrer des images 3D de résolution spatiale élevée et isotrope et de son innocuité, l’IRM est une modalité de choix pour caractériser son architecture. Cependant, la résolution spatiale est limitée, de l’ordre de grandeur de l’épaisseur des travées osseuses. Trois objectifs sont visés par ce travail. Le premier est de mettre en œuvre des outils pour caractériser l’architecture trabéculaire à partir d’images 3D acquises à une résolution de quelques dizaines de microns. Le deuxième est de les utiliser pour établir la validité de la représentation de la structure par microscopie IRM à 8.5 T par comparaison à une technique de référence, la tomographie X par rayonnement synchrotron. Cette étude, menée in vitro sur 29 échantillons de calcanéum, a consisté à comparer les paramètres d’architecture calculés sur des régions d’intérêt recalées automatiquement, communes aux deux modalités d’imagerie. Cette confrontation a montré que l’IRM à (66 µm) P
3P de résolution assurait un très bon rendu de l’organisation générale
de l’architecture. Par contre, l’évaluation de la fraction osseuse, bien que précise, était affectée par un biais important. Des hypothèses sur son origine ont été émises et vérifiées. Enfin, le troisième est d’établir un protocole d’imagerie in vivo. Nous avons obtenu des images 3D à 7 T de l’articulation distale du doigt humain à (78 µm) P
3P de résolution sur lesquelles une méthode automatique pour
isoler les zones trabéculaire et corticale a été développée. Une première évaluation des paramètres calculés sur la région trabéculaire a aussi été effectuée. Title: Three-dimensional characterization of trabecular bone architecture using MRI: in vitro and in vivo studies Abstract: There are two types of bone tissues, cortical and trabecular. Cortical bone is a compact tissue constituting bone envelope. Trabecular bone is a porous tissue consisting in a lattice of trabeculae (around 100 µm width) delimiting an interconnected space filled with marrow. Early detection of osteoporosis is possible in this tissue. As it is a non invasive method able to provide 3D images with isotropic resolution, MRI is of interest for trabecular bone architecture studies. However MRI resolution is at present limited to about 100 µm, i.e. comparable to trabecular thickness. This work had the three following aims. The first one was to develop methods for the assessment of structure parameters in order to characterize trabecular architecture on 3D images with a spatial resolution of a few tens of microns. The second one was to use them to estimate the accuracy of 8.5 T MR microscopy trabecular structure representation by reference to a « Gold Standard » technique: synchrotron radiation X-ray microtomography. This was performed in vitro on 29 calcaneus samples. Architecture parameters computed on common regions of interest (ROI) were compared between the two modalities. These ROIs were previously automatically matched. It was shown that the trabecular structure general organization was reliably represented by MR (66 µm)P
3P
resolution images. On the other hand, bone volume fraction evaluation, although precise, was significantly biased. Several hypothesis were checked to explain this bias. Finally, the third one was to establish an in vivo protocol. (78 µm)P
3P resolution MR images of the human finger
were acquired at 7 T. These images were used to develop an automatic 3D method to delineate trabecular and cortical regions. Computation of architecture parameters was also performed on the trabecular region. Mots-clefs: Microscopie IRM - Tissu osseux trabéculaire - Paramètres de microarchitecture - Recalage de ROI - Délimitation de ROI - Calcanéum - Doigt - Microtomographie X par rayonnement synchrotron
Discipline: Physique Spécialité: Electronique Laboratoire: Unité de Recherche en Résonance Magnétique Médicale U2R2M - CNRS UMR 8081 Université Paris Sud XI - UFR Scientifique d’Orsay F - 91405 ORSAY Cedex