Embed Size (px)
Citation preview

Versatile Cellular Uptake Mediated by Catanionic Vesicles:Simultaneous Spontaneous Membrane Fusion and EndocytosisChloe Mauroy,†,‡,§ Pauline Castagnos,†,§ Julie Orio,‡ Marie-Claire Blache,‡ Isabelle Rico-Lattes,†
Justin Teissie,‡ Marie-Pierre Rols,*,‡ and Muriel Blanzat*,†
†Laboratoire des Interactions Moleculaires et Reactivite Chimique et Photochimique, UMR 5623 Universite Paul Sabatier and CentreNational de la Recherche Scientifique, 118 route de Narbonne, 31062 Toulouse, France‡Institut de Pharmacologie et de Biologie Structurale, UMR 5089 Centre National de la Recherche Scientifique and Universite PaulSabatier, 205 route de Narbonne, 31077 Toulouse, France
*S Supporting Information
ABSTRACT: Lactose-derived catanionic vesicles offer unique opportunities to overcome cellular barriers. These potentialnanovectors, very easy to formulate as drug delivery systems, are able to encapsulate drugs of various hydrophilicity. This articlehighlights versatile interaction mechanisms between these catanionic vesicles, labeled with hydrophilic and amphiphilicfluorescent probes, and a mammalian cell line, Chinese Hamster Ovary. Confocal microscopy and flow cytometry techniquesshow that these vesicles are internalized by cells through cellular energy dependent processes, as endocytosis, but aresimultaneously able to spontaneously fuse with cell plasma membranes and release their hydrophilic content directly inside thecytosol. Such innovative and polyvalent nanovectors, able to deliver their content via different internalization pathways, wouldpositively be a great progress for the coadministration of drugs of complementary efficiency.
KEYWORDS: catanionic vesicles, membrane fusion, drug delivery, endocytosis, glycolipids
■ INTRODUCTIONDuring the last 40 years, a number of drug delivery systemshave been developed to control drug release profile, absorption,and distribution, with a view to improving efficacy andsafety.1−6 The direct release of active molecules inside thecytoplasm is often considered as the most efficient and thesafest mechanism for drug delivery. This promising strategy,which proceeds with drug delivery systems through membranefusion between suitable carriers and the cell membrane, is notstraightforward, due to the competition with endocytoticpathways. It has thus raised important research and develop-ments for the design of synthetic delivery systems. In the earlyseventies, Papahadjopoulos et al. proposed that liposomescomposed of mixtures between phosphatidylethanolamine(PE) and phosphatidylserine (PS) could fuse with plasmamembranes and release their contents into the cytoplasm.7
Indeed, vesicles are attractive systems for drug delivery, due inparticular to their ability to encapsulate either hydrophilic (inthe core) or hydrophobic (in the vesicle membrane) drugs,
controlling their degradation, release, and bioavailability.However, unilamellar vesicles, that are the preferred systemsfor drug delivery,8,9 remain difficult to obtain since multilayeredaggregates are more favorable systems. Therefore, research hasbeen devoted to improve the preparation and stability ofvesicles. In the last 20 years, spontaneous unilamellar vesiclesformation in the absence of applied external forces has beenreported with a new type of surfactants, namely, catanionicsystems.10−14 Catanionic vesicles, obtained by spontaneous self-assemblies of oppositely charged amphiphiles, show goodstability and dispersion properties in aqueous phase15−19 thatmake them potential candidates for drug delivery.5 Aspreviously described,20 we managed to form catanionic vesiclesmade of glycolipids (Figure 1) by a simple and safe preparation
Received: July 1, 2014Revised: September 17, 2014Accepted: October 13, 2014
Article
pubs.acs.org/molecularpharmaceutics
© XXXX American Chemical Society A dx.doi.org/10.1021/mp500458f | Mol. Pharmaceutics XXXX, XXX, XXX−XXX
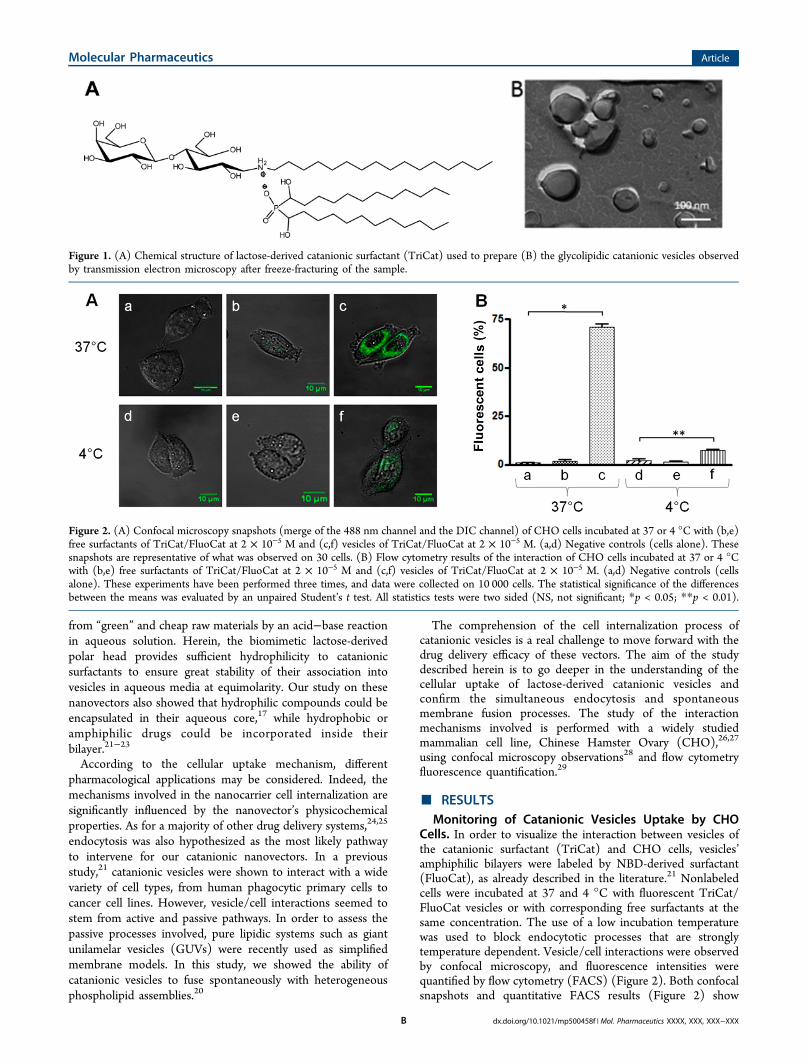
from “green” and cheap raw materials by an acid−base reactionin aqueous solution. Herein, the biomimetic lactose-derivedpolar head provides sufficient hydrophilicity to catanionicsurfactants to ensure great stability of their association intovesicles in aqueous media at equimolarity. Our study on thesenanovectors also showed that hydrophilic compounds could beencapsulated in their aqueous core,17 while hydrophobic oramphiphilic drugs could be incorporated inside theirbilayer.21−23
According to the cellular uptake mechanism, differentpharmacological applications may be considered. Indeed, themechanisms involved in the nanocarrier cell internalization aresignificantly influenced by the nanovector’s physicochemicalproperties. As for a majority of other drug delivery systems,24,25
endocytosis was also hypothesized as the most likely pathwayto intervene for our catanionic nanovectors. In a previousstudy,21 catanionic vesicles were shown to interact with a widevariety of cell types, from human phagocytic primary cells tocancer cell lines. However, vesicle/cell interactions seemed tostem from active and passive pathways. In order to assess thepassive processes involved, pure lipidic systems such as giantunilamelar vesicles (GUVs) were recently used as simplifiedmembrane models. In this study, we showed the ability ofcatanionic vesicles to fuse spontaneously with heterogeneousphospholipid assemblies.20
The comprehension of the cell internalization process ofcatanionic vesicles is a real challenge to move forward with thedrug delivery efficacy of these vectors. The aim of the studydescribed herein is to go deeper in the understanding of thecellular uptake of lactose-derived catanionic vesicles andconfirm the simultaneous endocytosis and spontaneousmembrane fusion processes. The study of the interactionmechanisms involved is performed with a widely studiedmammalian cell line, Chinese Hamster Ovary (CHO),26,27
using confocal microscopy observations28 and flow cytometryfluorescence quantification.29
■ RESULTSMonitoring of Catanionic Vesicles Uptake by CHO
Cells. In order to visualize the interaction between vesicles ofthe catanionic surfactant (TriCat) and CHO cells, vesicles’amphiphilic bilayers were labeled by NBD-derived surfactant(FluoCat), as already described in the literature.21 Nonlabeledcells were incubated at 37 and 4 °C with fluorescent TriCat/FluoCat vesicles or with corresponding free surfactants at thesame concentration. The use of a low incubation temperaturewas used to block endocytotic processes that are stronglytemperature dependent. Vesicle/cell interactions were observedby confocal microscopy, and fluorescence intensities werequantified by flow cytometry (FACS) (Figure 2). Both confocalsnapshots and quantitative FACS results (Figure 2) show
Figure 1. (A) Chemical structure of lactose-derived catanionic surfactant (TriCat) used to prepare (B) the glycolipidic catanionic vesicles observedby transmission electron microscopy after freeze-fracturing of the sample.
Figure 2. (A) Confocal microscopy snapshots (merge of the 488 nm channel and the DIC channel) of CHO cells incubated at 37 or 4 °C with (b,e)free surfactants of TriCat/FluoCat at 2 × 10−5 M and (c,f) vesicles of TriCat/FluoCat at 2 × 10−5 M. (a,d) Negative controls (cells alone). Thesesnapshots are representative of what was observed on 30 cells. (B) Flow cytometry results of the interaction of CHO cells incubated at 37 or 4 °Cwith (b,e) free surfactants of TriCat/FluoCat at 2 × 10−5 M and (c,f) vesicles of TriCat/FluoCat at 2 × 10−5 M. (a,d) Negative controls (cellsalone). These experiments have been performed three times, and data were collected on 10 000 cells. The statistical significance of the differencesbetween the means was evaluated by an unpaired Student’s t test. All statistics tests were two sided (NS, not significant; *p < 0.05; **p < 0.01).
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp500458f | Mol. Pharmaceutics XXXX, XXX, XXX−XXXB
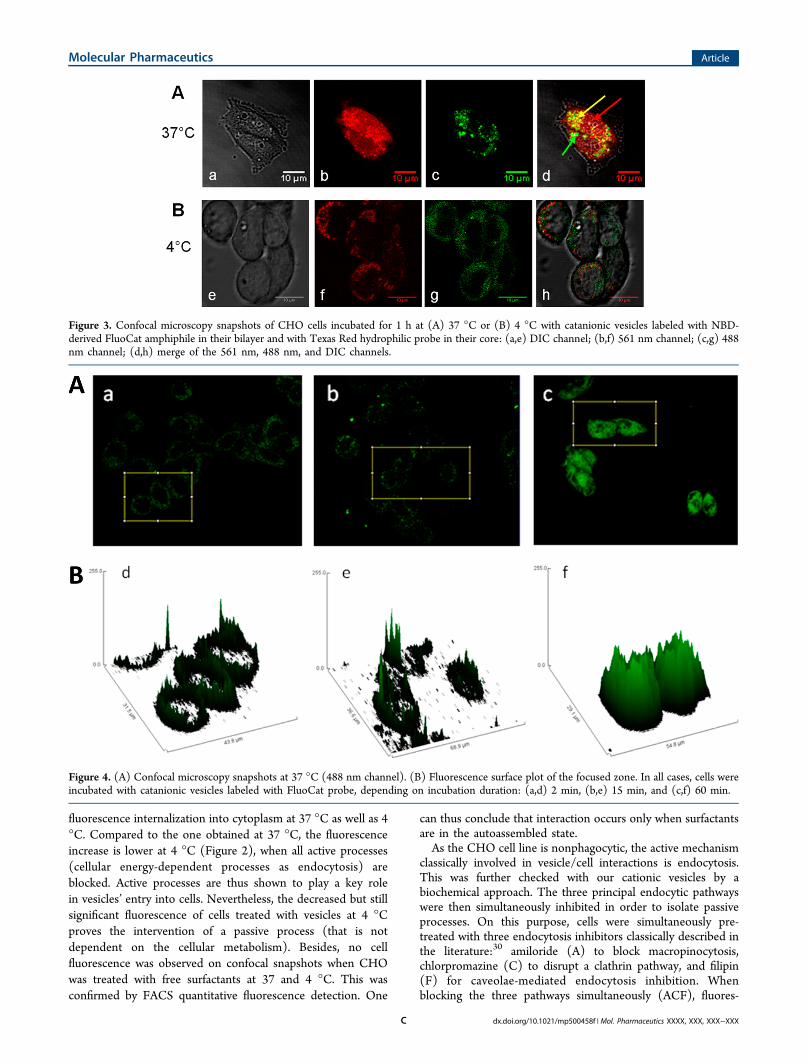
fluorescence internalization into cytoplasm at 37 °C as well as 4°C. Compared to the one obtained at 37 °C, the fluorescenceincrease is lower at 4 °C (Figure 2), when all active processes(cellular energy-dependent processes as endocytosis) areblocked. Active processes are thus shown to play a key rolein vesicles’ entry into cells. Nevertheless, the decreased but stillsignificant fluorescence of cells treated with vesicles at 4 °Cproves the intervention of a passive process (that is notdependent on the cellular metabolism). Besides, no cellfluorescence was observed on confocal snapshots when CHOwas treated with free surfactants at 37 and 4 °C. This wasconfirmed by FACS quantitative fluorescence detection. One
can thus conclude that interaction occurs only when surfactantsare in the autoassembled state.As the CHO cell line is nonphagocytic, the active mechanism
classically involved in vesicle/cell interactions is endocytosis.This was further checked with our cationic vesicles by abiochemical approach. The three principal endocytic pathwayswere then simultaneously inhibited in order to isolate passiveprocesses. On this purpose, cells were simultaneously pre-treated with three endocytosis inhibitors classically described inthe literature:30 amiloride (A) to block macropinocytosis,chlorpromazine (C) to disrupt a clathrin pathway, and filipin(F) for caveolae-mediated endocytosis inhibition. Whenblocking the three pathways simultaneously (ACF), fluores-
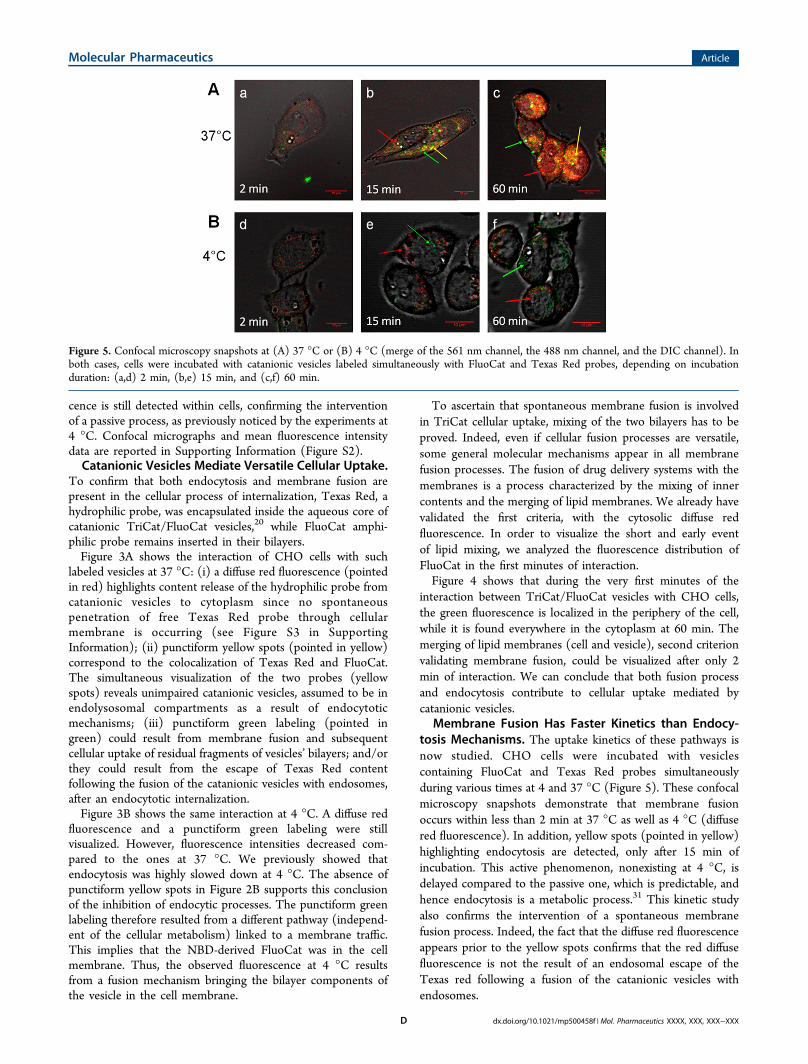
Figure 3. Confocal microscopy snapshots of CHO cells incubated for 1 h at (A) 37 °C or (B) 4 °C with catanionic vesicles labeled with NBD-derived FluoCat amphiphile in their bilayer and with Texas Red hydrophilic probe in their core: (a,e) DIC channel; (b,f) 561 nm channel; (c,g) 488nm channel; (d,h) merge of the 561 nm, 488 nm, and DIC channels.
Figure 4. (A) Confocal microscopy snapshots at 37 °C (488 nm channel). (B) Fluorescence surface plot of the focused zone. In all cases, cells wereincubated with catanionic vesicles labeled with FluoCat probe, depending on incubation duration: (a,d) 2 min, (b,e) 15 min, and (c,f) 60 min.
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp500458f | Mol. Pharmaceutics XXXX, XXX, XXX−XXXC
cence is still detected within cells, confirming the interventionof a passive process, as previously noticed by the experiments at4 °C. Confocal micrographs and mean fluorescence intensitydata are reported in Supporting Information (Figure S2).Catanionic Vesicles Mediate Versatile Cellular Uptake.
To confirm that both endocytosis and membrane fusion arepresent in the cellular process of internalization, Texas Red, ahydrophilic probe, was encapsulated inside the aqueous core ofcatanionic TriCat/FluoCat vesicles,20 while FluoCat amphi-philic probe remains inserted in their bilayers.Figure 3A shows the interaction of CHO cells with such
labeled vesicles at 37 °C: (i) a diffuse red fluorescence (pointedin red) highlights content release of the hydrophilic probe fromcatanionic vesicles to cytoplasm since no spontaneouspenetration of free Texas Red probe through cellularmembrane is occurring (see Figure S3 in SupportingInformation); (ii) punctiform yellow spots (pointed in yellow)correspond to the colocalization of Texas Red and FluoCat.The simultaneous visualization of the two probes (yellowspots) reveals unimpaired catanionic vesicles, assumed to be inendolysosomal compartments as a result of endocytoticmechanisms; (iii) punctiform green labeling (pointed ingreen) could result from membrane fusion and subsequentcellular uptake of residual fragments of vesicles’ bilayers; and/orthey could result from the escape of Texas Red contentfollowing the fusion of the catanionic vesicles with endosomes,after an endocytotic internalization.Figure 3B shows the same interaction at 4 °C. A diffuse red
fluorescence and a punctiform green labeling were stillvisualized. However, fluorescence intensities decreased com-pared to the ones at 37 °C. We previously showed thatendocytosis was highly slowed down at 4 °C. The absence ofpunctiform yellow spots in Figure 2B supports this conclusionof the inhibition of endocytic processes. The punctiform greenlabeling therefore resulted from a different pathway (independ-ent of the cellular metabolism) linked to a membrane traffic.This implies that the NBD-derived FluoCat was in the cellmembrane. Thus, the observed fluorescence at 4 °C resultsfrom a fusion mechanism bringing the bilayer components ofthe vesicle in the cell membrane.
To ascertain that spontaneous membrane fusion is involvedin TriCat cellular uptake, mixing of the two bilayers has to beproved. Indeed, even if cellular fusion processes are versatile,some general molecular mechanisms appear in all membranefusion processes. The fusion of drug delivery systems with themembranes is a process characterized by the mixing of innercontents and the merging of lipid membranes. We already havevalidated the first criteria, with the cytosolic diffuse redfluorescence. In order to visualize the short and early eventof lipid mixing, we analyzed the fluorescence distribution ofFluoCat in the first minutes of interaction.Figure 4 shows that during the very first minutes of the
interaction between TriCat/FluoCat vesicles with CHO cells,the green fluorescence is localized in the periphery of the cell,while it is found everywhere in the cytoplasm at 60 min. Themerging of lipid membranes (cell and vesicle), second criterionvalidating membrane fusion, could be visualized after only 2min of interaction. We can conclude that both fusion processand endocytosis contribute to cellular uptake mediated bycatanionic vesicles.
Membrane Fusion Has Faster Kinetics than Endocy-tosis Mechanisms. The uptake kinetics of these pathways isnow studied. CHO cells were incubated with vesiclescontaining FluoCat and Texas Red probes simultaneouslyduring various times at 4 and 37 °C (Figure 5). These confocalmicroscopy snapshots demonstrate that membrane fusionoccurs within less than 2 min at 37 °C as well as 4 °C (diffusered fluorescence). In addition, yellow spots (pointed in yellow)highlighting endocytosis are detected, only after 15 min ofincubation. This active phenomenon, nonexisting at 4 °C, isdelayed compared to the passive one, which is predictable, andhence endocytosis is a metabolic process.31 This kinetic studyalso confirms the intervention of a spontaneous membranefusion process. Indeed, the fact that the diffuse red fluorescenceappears prior to the yellow spots confirms that the red diffusefluorescence is not the result of an endosomal escape of theTexas red following a fusion of the catanionic vesicles withendosomes.
Figure 5. Confocal microscopy snapshots at (A) 37 °C or (B) 4 °C (merge of the 561 nm channel, the 488 nm channel, and the DIC channel). Inboth cases, cells were incubated with catanionic vesicles labeled simultaneously with FluoCat and Texas Red probes, depending on incubationduration: (a,d) 2 min, (b,e) 15 min, and (c,f) 60 min.
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp500458f | Mol. Pharmaceutics XXXX, XXX, XXX−XXXD

■ DISCUSSION
TriCat catanionic vesicles are easy to formulate sugar-basedsurfactants assemblies constituting “green” alternatives to more“classic” drug delivery systems. This study reports evidence thatcatanionic vesicles are able to deliver hydrophilic andamphiphilic substances into cells following various mechanisms.We thus showed that when the three endocytic pathways are
inhibited (thermally or chemically), the residual fluorescence isstill meaningful and significantly different depending on theinhibition process. Two hypotheses can thus be drawn. First,other active processes than the three ones already mentionedare involved. Second, a passive process is involved.Colocalization experiments (Figure 3) combined to the
results obtained at 4 °C (residual fluorescence at 4 °C) allowsdiscriminating these two hypotheses. At 37 °C, colocalization ofTexas Red and FluoCat probes with a punctiform shape revealsinternalization of TriCat vesicles into cytoplasm, which givesevidence of an endocytic process. However, at 4 °C, such spotswere not visualized, highlighting the absence of any kind ofendocytosis. Nevertheless, the intracytoplasmic detection ofTexas Red demonstrates a spontaneous membrane fusion atlow temperatures.This result validates previous observations obtained with
giant unilamellar vesicles (GUVs) as cellular membrane models.Indeed, we previously proved that heterogeneous structure onGUVs composition is necessary to create a membranedestabilization inducing fusion process.20 Spontaneous mem-brane fusion of catanionic vesicles is thus a passive mechanicalprocess, which requires the presence of phospholipidic domainsinducing interfacial defaults where lipids are less organized. Theresults obtained in this study highlight that these vesicles areable to fuse spontaneously with the membrane of living cells,which possess lipid heterogeneities and notably phaseinterfaces. Indeed, the presence of lipid microdomains rich insphingomyelin and cholesterol induces inhomogeneous lateraldistribution of membranes32−34 enhancing lipid disorganizationand destabilization.Not surprisingly, a kinetics study showed that membrane
fusion is a much faster process than endocytosis pathways,which are complex cellular processes implying intervention ofproteins (indeed Figure 5 shows immediate apparition of greenspots either at 37 or 4 °C, contrary to the yellow ones).35
■ CONCLUSIONS
Very few drug delivery systems have been identified tospontaneously fuse with the plasma membrane. Indeed, thefusion of drug delivery systems with the cell membrane is farfrom being straightforward as different energy barriers have tobe overcome to allow membrane fusion. The cell internalizationmechanism of lactose-derived catanionic vesicle contents pointsout that they are better than “another” drug delivery system.These original nanocarriers have proved to be capable ofencapsulating active principles of various hydrophilicities andusing different cellular uptake pathways to deliver them. Thesecomplementary entry pathways highlight the versatility of suchpotential drug delivery systems. Spontaneous membrane fusionand endocytic processes confer to catanionic vesicles the abilityto deliver compounds either directly into cell cytoplasm or intoendolysosomal compartments. Therefore, perspectives of suchcatanionic vesicles are countless. Indeed, these polyvalent drugdelivery systems offer a wide variety of applications since onecan encapsulate drugs of any hydrophilicity and target any part
of the cell following the cellular uptake pathway used.Coencapsulation of several drugs can even be foreseen,exploiting the difference of release kinetics depending ontheir hydrophilicity. It is worth noticing that the composition ofsuch nano-objects could be also adjusted for specific targeting,by optimizing for instance the nature of surfactant sugar-heador by grafting ligands for specific recognition. Such strategieswould be of great interest for coadministration of several drugsof complementary efficiency, in cancer treatment for instance.
■ MATERIALS AND METHODSMaterials. α-Lactose monohydrate, dodecylamine, meth-
anol, hypophosphorous acid, and dodecylaldehyde used toprepare catanionic surfactants were obtained from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA)12-(N-(7-Nitrobenzo-2-oxa-1, 3-diazol-4-yl)amino)-
dodecanoic acid (NBD) used to synthesize amphiphilicfluorescent probe FluoCat (λex = 488 nm) and hydrophilicfluorescent probe Texas Red (λex = 563 nm) were obtainedfrom Invitrogen Molecular Probes (Carlsbad, CA, USA).
Surfactants Syntheses (TriCat and FluoCat). TriCat. 1-N-Dodecylammonium-1-deoxylactitol-bis(α-hydroxydodecyl)-phosphinate, the tricatenar catanionic surfactant, so-calledTriCat, was obtained following protocols described in theliterature.17,20 It is an ionic pair composed by an in situ acid−base reaction between two surfactants: N-dodecylamino-1-deoxylactitol36 and alpha-bis(hydroxyl) docecylphosphinicacid.37
FluoCat. As previously reported in the literature,21 N-hexadecylammonium-1-deoxylactitol 12-(-N-(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)dodecanoate, so-called FluoCat, isan amphiphilic fluorescent probe synthesized from commer-cially available (molecular probes) 12-(N-(7-nitrobenzo-2-oxa-1,3-diazol-4-yl)amino)dodecanoic acid (NBD), added to N-hexadecyl amino-1-deoxylactitol.
Preparation and Physicochemical Characterization ofCatanionic Vesicles (TriCat, TriCat/FluoCat, TriCat/TexasRed, and TriCat/FluoCat/Texas Red). As already de-scribed,20,21,36 freeze-dried TriCat or TriCat/FluoCat (95:5)was put in water or in Texas Red solution (500 μM in water) atthe concentration of 1 × 10−4 M in TriCat (which is aboveCAC (3 × 10−5 M)), stirred, and then sonicated (Vibra Cell,Bioblock Scientific, titanium probe, pulse rate 30%, intensity×3) for 15 min. Sizes of obtained catanionic vesicles weredetermined using dynamic light scattering (Malvern Instru-ments, Nano ZS ZEN3600, U.K.). The analysis was performedwith a He−Ne laser (633 nm), a scattering angle of 173°, and ata temperature of 25.0 °C ± 0.1 °C. Vesicles’ size andmorphology were checked by transmission electron microscopyusing a JEOL JEM 1011 electron microscope, operating at 120kV. Freeze−fracture experiments were performed with a Balzervacuum chamber BAF (Balzer, Liechtenstein). A small dropletof aqueous solution (5 mM) was sandwiched between twocopper specimen holders and was kept at the desiredtemperature to reach equilibrium. The environment wassaturated with water to avoid evaporation. The sandwich wasthen frozen with liquid propane cooled with liquid nitrogen.The frozen sandwich was additionally fixed to a transport unitunder liquid nitrogen and transferred to the fracture replicationstage in a chamber that was then pumped down to 10−6 mbar at−120 °C. Immediately after fracturing, replication took place byfirst shadowing with platinum/carbon at 45 °C and then withcarbon deposition at 90 °C. The sample was allowed to warm
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp500458f | Mol. Pharmaceutics XXXX, XXX, XXX−XXXE

to room temperature. Replicas were retrieved and cleaned inwater and mounted on 200-mesh copper grids. Observationswere made with a cryo-electron microscope FEI EM120 (120kV), and the images were recorded with an SSCCD 2k Gatancamera. Spectrofluorimetry measurements were performed onvesicles of TriCat/FluoCat (95:5), on a spectrofluorimeter PTI(Photon Technology International) equipped with a EIMACxenon lamp of 175 W. All the slit widths were set at 2 nm. Theexcitation wavelength was set at 488 nm, and the emittedintensity was collected from 500 to 600 nm. The emissionwavelength has been determined at 549 nm.Physicochemical characteristics (shape, size, and stability) of
labeled vesicles with FluoCat and/or Texas Red were checkedto remain identical as “empty” TriCat vesicles without any pHdependence, compatible with flow cytometry or confocal laserscanning microscopy detection. To perform vesicles/cellsinteractions studies, catanionic vesicles were diluted until 2 ×10−5 M. Vesicles of TriCat, labeled or not with FluoCat and/orTexas Red, were checked to be stable under dilution by DLSand tensiometry analyses, even at concentrations inferior to thesurfactant CAC.21
Free surfactants were also prepared at the concentration of 2× 10−5 M.Cell Culture. Chinese Hamster Ovary (CHO) cells (Wild
Type Toronto clone) were grown plated in EMEM (Eurobio)supplemented with 8% (v/v) fetal calf serum (Lonza), 1% (v/v) L-glutamine (2 mM), 1% (v/v) penicillin streptomycin (50IU/mL) (all from Eurobio), 3.5 g·L−1 glucose, 1% (v/v)vitamins, and 2.95 g·L−1 tryptose phosphate (all from Sigma).Culture medium was renewed every 2 days. Flasks were fromNunc. Culture was operated in a 5% CO2 atmosphere at 37 °Cwith 80% humidity.Cytotoxicity Test. A 2,3-bis[2-methoxy-4-nitro-5-sulfo-
phenyl]-2H-tetrazolium-5-arboxanilide (XTT) test was per-formed to check the cytotolerance of catanionic vesiclesdepending on the applied concentration. Cells were seededon 96-well plates for 2 days at 2 × 105 cell/mL. Cells werewashed with phosphate buffer solution (PBS) (Sigma) andincubated for 3 h with solutions of catanionic vesicles rangingfrom 1 × 104 to 2 × 107 M in serum free EMEM. Cells werethen washed with PBS, and 50 μL of a Red Phenol free EMEMsolution containing XTT (0.5 mg/mL) and coenzyme Q (40μL/mL) were added. After 3 h of incubation, 100 μL of asodium dodecyl sulfate solution (10%) was added. The reactionwas then read on a Polarstar Galaxy spectrophotometer (BMGLabtech, France) at a wavelength set at 450 nm.The concentration of vesicles applied on cells was chosen
according to the cytotoxicity results. About 100% viability ofCHO was observed after 3 h of incubation with vesicles at 2 ×10−5 M whatever their composition (TriCat, TriCat/FluoCat,TriCat/Texas Red, and TriCat/FluoCat/Texas Red).Internalization Inhibition. To study the endocytosis
phenomenon, endocytosis inhibiting agents widely describedin the literature30 were applied on CHO cells: amiloride (A),chlorpromazine (C), and filipin (F).Optimal concentration of each inhibitor was determined by a
preliminary study where a range of concentrations was appliedon cells. Chosen concentrations showed maximal inhibitingefficiency and lowest cytotoxicity: (A) 1 mg/mL; (C) 1.3 ×10−2mg/mL; (F) 1.5 × 10−3mg/mL; and (ACF) the mixture ofthe previous amiloride + chlorpromazine + filipin at the sameconcentrations in culture medium. CHO cells were pretreatedwith these agents 1 h in an incubator (37 °C, 5% CO2, in
darkness conditions) before introducing catanionic vesicles,which were then incubated in the same conditions and in thepresence of inhibiting agents for 1 h in the case of FACSanalyses and 15 min in the case of confocal microscopy study.In the case of inhibition of all active processes under cold
temperature, CHO cells were incubated at 4 °C in darknessconditions 15 min before introducing catanionic vesicles, whichwere added while maintaining this temperature throughout thevesicle/cell interaction experiment. Internalization should nothappen. It is indeed well-known that it is a very slow process at4 °C.38
Confocal Laser Scanning Microscopy. Evaluation ofvesicle uptake by cells was performed by confocal laser scanningmicroscopy. CHO cells were seeded on Lab-Tek plates (Nunc)at 8 × 104 cell/mL for 1 day atmosphere at 37 °C with 80%humidity. Subsequently, cells were extensively washed with PBSat room temperature. Catanionic vesicles were diluted in serumfree EMEM at the final concentration of 2 × 10−5 M andincubated for 15 min with the plated CHO cells. Cells werethen washed with cold PBS and then observed in fresh PBS.Live cell imaging was performed with a biphoton microscopeZeiss LSM 710 Inv (obj. ×40 in water), using Argon laser at488 nm and DPSS laser at 561 nm for FluoCat and Texas Redexcitations, respectively. Three images of each conditioncontaining up to 30 individual cells were analyzed with ImageJsoftware (National Institutes of Health) without anyquantification. Images are representative of the original data,corresponding to the fluorescence signal of labeled catanionicvesicles since cells were not labeled. A control without vesicleswas used to set laser intensity in order to limit cellularautofluorescence.
Fluorescence-Activated Cell Sorting (FACS) Analyses.CHO cells were seeded on 24-well plates (Nunc) for 1 day at7.5 × 104 cell/mL in supplemented EMEM in a CO2atmosphere at 37 °C with 80% humidity. Cells were extensivelywashed with PBS. Catanionic vesicles were diluted in serumfree EMEM at the final concentration of 2 × 10−5 M andincubated for 1 h with the plated CHO cells. Cells were thenwashed with cold PBS, harvested using trypsin/EDTA, dilutedin full serum EMEM, and analyzed by FACS (BD FascaliburCell Sorter) by using CellQuest Pro software to quantify thepercentage of fluorescent cells. Laser excitation was at 488 nm.Statistical analysis was performed using Prism 4.0 (GraphPad
Software) by one-way ANOVA and Tukey test. All data wereexpressed as the mean ± SEM of three independent sets ofexperiments on 10,000 cells each. Statistical significance for thisstudy was considered at p < 0.05.
■ ASSOCIATED CONTENT*S Supporting InformationTEM snapshot and size distribution of catanionic vesicles; flowcytometry results; confocal microscopy snapshots; andcytotoxicity of catanionic vesicles. This material is availablefree of charge via the Internet at http://pubs.acs.org.
■ AUTHOR INFORMATIONCorresponding Authors*Tel: +33 5 61 55 73 75. Fax: +33 5 61 55 81 55. E-mail:[email protected].*E-mail: [email protected].
Author Contributions§P.C. and C.M. contributed equally to this work.
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp500458f | Mol. Pharmaceutics XXXX, XXX, XXX−XXXF

NotesThe authors declare no competing financial interest.
■ ACKNOWLEDGMENTSThe imaging platform Toulouse Reseau Imagerie is acknowl-edged for providing the technical support in microscopy andFACS. TRI is funded by the region Midi Pyrenees, the grandToulouse cluster, and the EU FEDER. FONDEREPHAR at thePharmacy Faculty of Toulouse is acknowledged for theirexpertise in cytotoxicity experiments. P.C. and C.M. weresupported by fellowships from the MENESR. We acknowledgefinancial support from the Association Francaise contre lesMyopathies, the ANR contracts Cemirbio and CMIDT,COPOPDT, the Direction Generale de l′Armement, theMinistere des Affaires Etrangeres et Europeennes, and theRegion Midi-Pyrenees (reseau therapie genique).
■ REFERENCES(1) Allen, T. M.; Cullis, P. R. Drug delivery systems: entering themainstream. Science 2004, 303, 1818−1824.(2) Faraji, A. H.; Wipf, P. Nanoparticles in cellular drug delivery.Bioorg. Med. Chem. 2009, 17 (8), 2950−2962.(3) Moses, M. A.; Brem, H.; Langer, R. Advancing the field of drugdelivery: Taking aim at cancer. Cancer Cell 2003, 4 (5), 337−341.(4) Singh, R.; Lillard, J. W., Jr. Nanoparticle-based targeted drugdelivery. Exp. Mol. Pathol. 2009, 86 (3), 215−223.(5) Soussan, E.; Cassel, S.; Blanzat, M.; Rico-Lattes, I. Drug deliveryby soft matter: Matrix and vesicular carriers. Angew. Chem., Int. Ed.2009, 48 (2), 274−288.(6) Torchilin, V. P. Recent advances with liposomes aspharmaceutical carriers. Nat. Rev. Drug Discovery 2005, 4 (2), 145−160.(7) Papahadjopoulos, D.; Poste, G.; Schaeffer, B. E. Fusion ofmammalian cells by unilamellar lipid vesicles: Influence of lipid surfacecharge, fluidity and cholesterol. Biochim. Biophys. Acta, Biomembr.1973, 323, 23−42.(8) Gregoriadis, G.; Allison, A. C. Liposomes in Biological Systems;John Wileys and Sons: New York, 1990.(9) Malmsten, M. Surfactants and Polymers in Drug Delivery. MarcelDekker: New York, 2002.(10) Kaler, E. W.; Murthy, A. K.; Rodriguez, B. E.; Zasadzinski, J. A.Spontaneous vesicle formation in aqueous mixtures of single-tailedsurfactants. Science 1989, 245 (4924), 1371−1374.(11) Marques, E.; Khan, A.; da Graca Miguel, M.; Lindman, B. Self-assembly in mixtures of a cationic and an anionic surfactant: thesodium dodecyl sulfate-didodecyldimethylammonium bromide-watersystem. J. Phys. Chem. 1993, 97 (18), 4729−4736.(12) Menger, F. M.; Binder, W. H.; Keiper, J. S. Cationic surfactantswith counterions of glucuronate glycosides. Langmuir 1997, 13 (12),3247−3250.(13) Blanzat, M.; Perez, E.; Rico-Lattes, I.; Prome, D.; Prome, J. C.;Lattes, A. New catanionic glycolipids. 1. Synthesis, characterization,and biological activity of double-chain and gemini catanionic analoguesof galactosylceramide (gal beta(1)cer). Langmuir 1999, 15 (19),6163−6169.(14) Blanzat, M.; Perez, E.; Rico-Lattes, I.; Lattes, A.; Gulik, A.Correlation between structure, aggregation behaviour and cellulartoxicity of anti-HIV catanionic analogues of galactosylceramide. Chem.Commun. 2003, 2, 244−245.(15) Marques, E. F.; Regev, O.; Khan, A.; Lindman, B. Self-organization of double-chained and pseudodouble-chained surfactants:counterion and geometry effects. Adv. Colloid Interface Sci. 2003, 100−102, 83−104.(16) Vivares, D.; Soussan, E.; Blanzat, M.; Rico-Lattes, I. Sugar-derived tricatenar catanionic surfactant: Self-assembly and aggregationbehavior in the cationic-rich side of the system. Langmuir 2008, 24,9260−9267.
(17) Soussan, E.; Mille, C.; Blanzat, M.; Bordat, P.; Rico-Lattes, I.Sugar-derived tricatenar catanionic surfactant: Synthesis, self-assemblyproperties, and hydrophilic probe encapsulation by vesicles. Langmuir2008, 24 (6), 2326−2330.(18) Ghosh, S.; Ambade, B.; Ray, A. Stable catanionic vesicles as drugdelivery vehicle. Sci. Adv. Mater. 2013, 5 (12), 1837−1846.(19) Qu, W.; Xia, Z.; Guo, H.; Li, X.; Dou, T. Vesicle Formation withan Anionic Surfactant and a Conventional Cationic Surfactant inMixed Systems. J. Dispersion Sci. Technol. 2013, 34 (2), 240−243.(20) Mauroy, C.; Castagnos, P.; Blache, M.-C.; Teissie, J.; Rico-Lattes, I.; Rols, M.-P.; Blanzat, M. Interaction between GUVs andcatanionic nanocontainers: new insight into spontaneous membranefusion. Chem. Commun. 2012, 48, 6648−6650.(21) Boudier, A.; Castagnos, P.; Soussan, E.; Beaune, G.; Belkhelfa,H.; Menager, C.; Cabuil, V.; Haddioui, L.; Roques, C.; Rico-Lattes, I.;Blanzat, M. Polyvalent catanionic vesicles: Exploring the drug deliverymechanisms. Int. J. Pharm. 2011, 403 (1−2), 230−236.(22) Consola, S.; Blanzat, M.; Perez, E.; Garrigues, J.-C.; Bordat, P.;Rico-Lattes, I. Design of original bioactive formulations based onsugar-surfactant/non-steroidal anti-inflammatory catanionic self-as-semblies: A new way of dermal drug delivery. Chem.Eur. J. 2007,13 (11), 3039−3047.(23) Castagnos, P.; Siqueira-Moura, M. P.; Leme Goto, P.; Perez, E.;Franceschi, S.; Rico-Lattes, I.; Tedesco, A. C.; Blanzat, M. Catanionicvesicles charged with chloroaluminium phthalocyanine for topicalphotodynamic therapy. In vitro phototoxicity towards humancarcinoma and melanoma cell lines. RSC Adv. 2014, 4, 39372−39377.(24) Bareford, L. M.; Swaan, P. W. Endocytic mechanisms fortargeted drug delivery. Adv. Drug Delivery Rev. 2007, 59 (8), 748−758.(25) Marianecci, C.; Di Marzio, L.; Rinaldi, F.; Celia, C.; Paolino, D.;Alhaique, F.; Esposito, S.; Carafa, M. Niosomes from 80s to present:the state of the art. Adv. Colloid Interface Sci. 2014, 205, 187−206.(26) Cezanne, L.; Navarro, L.; Tocanne, J.-F. Isolation of the plasmamembrane and organelles from Chinese hamster ovary cells. Biochim.Biophys. Acta, Biomembr. 1992, 1112 (2), 205−214.(27) Gottesman, M. M. Molecular Cell Genetics, 1st ed.; John Wiley &Sons, Inc.: New York, 1985.(28) Torchilin, V. P. Fluorescence microscopy to follow the targetingof liposomes and micelles to cells and their intracellular fate. Adv. DrugDelivery Rev. 2005, 57 (1), 95−109.(29) Huth, U. S.; Schubert, R.; Peschka-Suss, R. Investigating theuptake and intracellular fate of pH-sensitive liposomes by flowcytometry and spectral bio-imaging. J. Controlled Release 2006, 110(3), 490−504.(30) Khalil, I. A.; Kogure, K.; Alkita, H.; Harashima, H. Uptakepathways and subsequent intracellular trafficking in non viral genedelivery. Pharmacol. Rev. 2006, 58, 32−45.(31) Grant, B. D.; Donaldson, J. G. Pathways and mechanisms ofendocytic recycling. Nat. Rev. Mol. Cell Biol. 2009, 10 (9), 597−608.(32) Edidin, M. The state of lipid rafts: From model membranes tocells. Annu. Rev. Biophys. Biomol. Struct. 2003, 32 (1), 257−283.(33) Tocanne, J.-F.; Cezanne, L.; Lopez, A.; Piknova, B.; Schram, V.;Tournier, J.-F.; Welby, M. Lipid domains and lipid/proteininteractions in biological membranes. Chem. Phys. Lipids 1994, 73(1−2), 139−158.(34) Tocanne, J.-F.; Dupou-Cezanne, L.; Lopez, A.; Tournier, J.-F.Lipid lateral diffusion and membrane organization. FEBS Lett. 1989,257 (1), 10−16.(35) Marsh, M.; McMahon, H. T. The structural era of endocytosis.Science 1999, 285 (5425), 215−220.(36) Castagnos, P. Vesicules catanioniques: Design et mecanismes dedelivrance de principes actifs. Ph.D. thesis, Toulouse III, Toulouse,France, 2011.(37) Brun, A.; Etemad-Moghadam, G. New double-chain andaromatic (alpha-hydroxyalkyl)phosphorus amphiphiles. Synthesis2002, 10, 1385−1390.(38) Di Marzio, L.; Marianecci, C.; Cinque, B.; Nazzarri, M.; Cimini,A. M.; Cristiano, L.; Cifone, M. G.; Alhaique, F.; Carafa, M. pH-sensitive non-phospholipid vesicle and macrophage-like cells: Binding,
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp500458f | Mol. Pharmaceutics XXXX, XXX, XXX−XXXG

uptake and endocytotic pathway. Biochim. Biophys. Acta 2008, 1778(12), 2749−2756.
Molecular Pharmaceutics Article
dx.doi.org/10.1021/mp500458f | Mol. Pharmaceutics XXXX, XXX, XXX−XXXH

















![Control of Vascular Sap pH by the Vessel-Associated Cells ... · of Poitiers University. Perfusion Technique This technique (initially used by Van Bel [1974] to study the uptake of](https://img.pdfslide.fr/doc/110x75/5eda89c0febf237c0c3b770d/control-of-vascular-sap-ph-by-the-vessel-associated-cells-of-poitiers-university.jpg)




![Antisenserepression chloroplast inProc. Natl. Acad. Sci. USA90(1993) 6161 centrifugation. The transport of [14C]malate was used to normalize [32P]P, uptake activities of different](https://img.pdfslide.fr/doc/110x75/5ffa530e20ae570318129513/antisenserepression-chloroplast-in-proc-natl-acad-sci-usa901993-6161-centrifugation.jpg)






