
7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 1/40
Cell Structure and FunctionStructure of the Nuclear envelope

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 2/40
Nuclear lamina - A meshwork of lamin filaments providing structuralsupport to the nucleus
-serve as a site of chromatin attachment
Nuclear envelope- composed of:1. inner and outer membranes2. nuclear lamina3. nuclear pore complex
Nuclear pore complex - A large structure forming a transport channelthrough the nuclear envelope .

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 3/40
Lamins . Mostmammalian cells,for example,contain fourdifferent lamins,designated A, B 1,B2, and C. All thelamins are 60- to80-kilodalton (kd)fibrous proteinsthat are related tothe intermediatefilament proteins
of the cytoskeleton

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 4/40
Molecular traffic through nuclear pore complexes

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 5/40
Model of the nuclear pore complex

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 6/40
Selective Transport of Proteins to and from the Nucleus
Proteins are targeted to the nucleus by specific amino acid sequences,called nuclear localization signals
7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 7/40
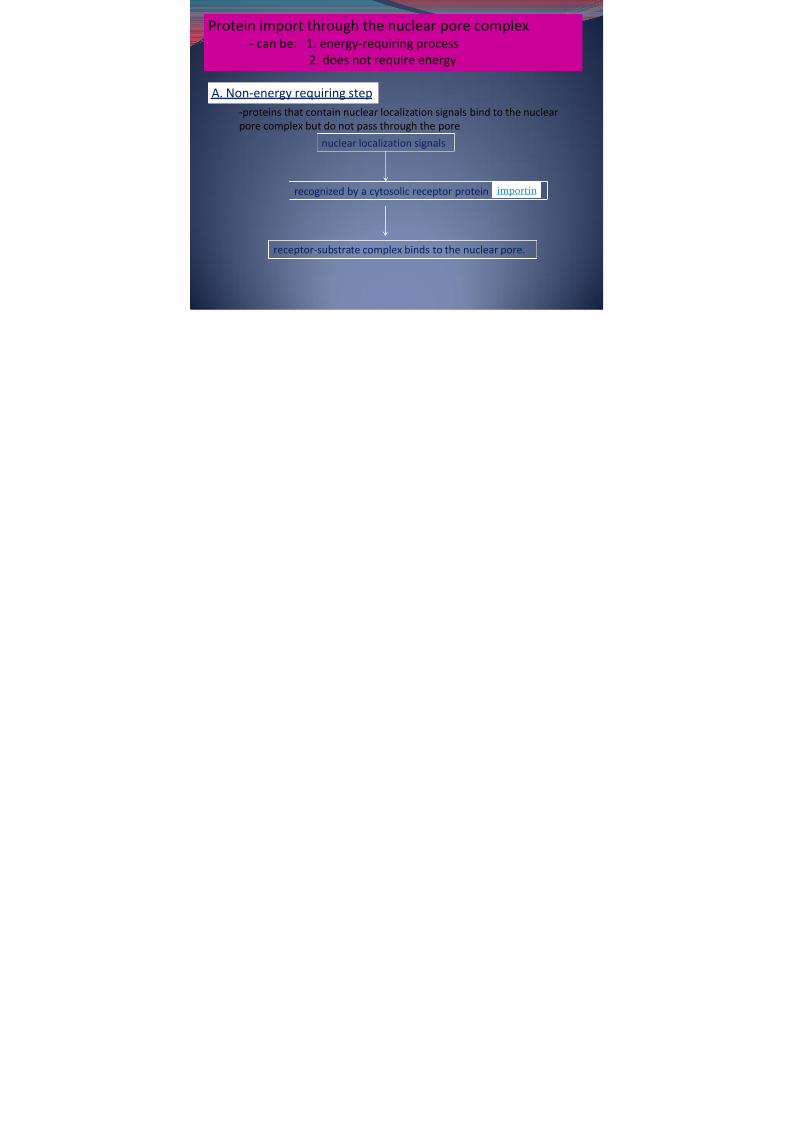
Protein import through the nuclear pore complex- can be: 1. energy-requiring process
2. does not require energy
-proteins that contain nuclear localization signals bind to the nuclearpore complex but do not pass through the pore
A. Non-energy requiring step
nuclear localization signals
receptor-substrate complex binds to the nuclear pore.
recognized by a cytosolic receptor protein importin
7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 8/40
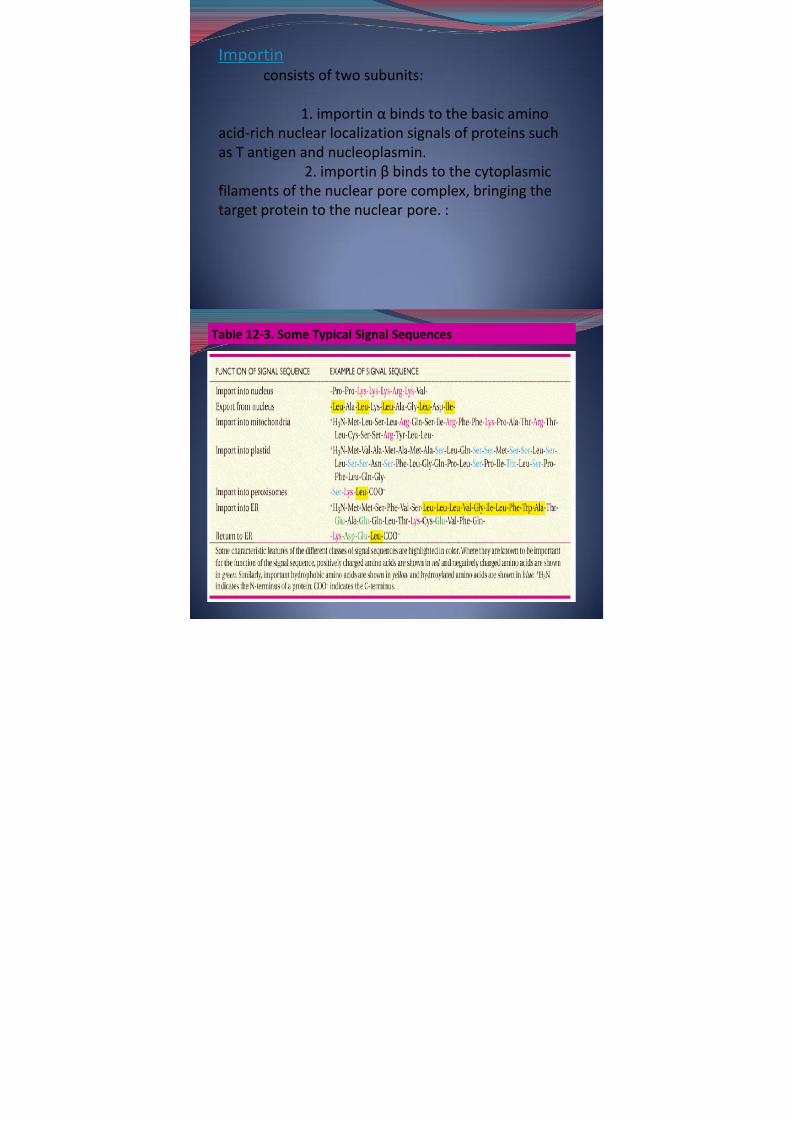
Importin consists of two subunits:
1. importin α binds to the basic aminoacid-rich nuclear localization signals of proteins suchas T antigen and nucleoplasmin.
2. importin β binds to the cytoplasmicfilaments of the nuclear pore complex, bringing the
target protein to the nuclear pore. :

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 9/40
Table 12-3. Some Typical Signal Sequences

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 10/40
B. Energy-requiring step
-requires GTP hydrolysis. A key player in the translocationprocess is a small GTP-binding protein called Ran ,(involved in nuclear import and export)
Enzymes that stimulate GTP binding to Ran are localized to thenuclear side of the nuclear envelope whereas enzymes thatstimulate GTP hydrolysis are localized to the cytoplasmic side.
Role of the Ran protein in nuclear import
1.Transport through the nuclear pore complex is driven by agradient of Ran/GTP,
- a high concentration of Ran/GDP in the cytoplasm- a high concentration of Ran/GTP in the nucleus.
2.Complexes form between target proteins containing nuclearlocalization signals (NLS) importin α, and importin β in thecytoplasm where Ran is in the GDP-bound form.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 11/40
3. Transport through the nuclear pore complex, Ran/GTP binds to
importin β, releasing importin α and the target protein in thenucleus.
4. The Ran/GTP-importin β complex is then transported back to thecytoplasm, where the Ran GTPase-activating protein (Ran GAP)stimulates hydrolysis of the bound GTP to form Ran/GDP.
5. This conversion of Ran/ GTP to Ran/GDP is accompanied by releaseof importin β.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 12/40
Role of the Ran protein in nuclear import

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 13/40
Nuclear export
Proteins are targeted for export from the nucleus by specific
amino acid sequences, called nuclear export signals .

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 14/40
*the nuclear export receptors (called exportins ) are related toimportin β
1. Complexes between target proteins bearing nuclear exportsignals (NES), exportins, and Ran/ GTP form in the nucleus.
2. transport through the nuclear pore complex, Ran GAP
stimulates the hydrolysis of bound GTP, leading to formation of Ran/GDP and release of the target protein and exportin in thecytoplasm.
3. Exportin is then transported back to the nucleus.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 15/40
The Nucleolus
morphologically, nucleoli consist of three distinguishableregions: the fibrillar center, dense fibrillar component, andgranular component
the most prominent substructure within the nucleus
ribosome production factory, designed to fulfill the need forlarge-scale production of rRNAs and assembly of the ribosomalsubunits.
not surrounded by a membrane
organized around the chromosomal regions thatcontain the genes for the 5.8S, 18S, and 28S rRNAs.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 16/40
The Nucleus during Mitosis
1. Dissolution of the Nuclear Envelope -the disassembly of the nuclear envelope marks the end of theprophase of mitosis ( not a universal feature of mitosis and doesnot occur in all cells) a. Closed mitosis
- in which the nuclear envelope remains intact
- the daughter chromosomes migrate to opposite poles of the nucleus, which then divides in two.- occurs in some unicellular eukaryotes (e.g., yeasts)
b. Open mitosis- characterized by breakdown of the nuclear envelope.
- daughter chromosomes then migrate to opposite poles of the mitotic spindle, and new nuclei reassemble aroundthem- occur in cells of higher eukaryotes

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 17/40

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 18/40
Disassembly of the nuclear envelope-involves changes in all three of its components:
1. the nuclear membranes are fragmented into vesicles,2. the nuclear pore complexes dissociate,3. and the nuclear lamina depolymerizes.
At mitosis, Cdc2 and otherprotein kinases phosphorylatethe lamins, causing thefilaments to dissociate into
free lamin dimers.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 19/40
2. Chromosome Condensation
-Recent studies have also identified protein complexes calledcondensins that play a major role in chromosome condensation.
-Condensins are phosphorylated and activated by the Cdc2 proteinkinase, providing a direct link between activation of Cdc2 and mitoticchromosome condensation
3. Re-formation of the Interphase Nucleus-Chromosome decondensation and reassembly of the nuclearenvelope appear to be signaled by inactivation of Cdc2, whichwas responsible for initiating mitosis by phosphorylating cellulartarget proteins, including the lamins, histone H3, and condensins.
-Inactivation of Cdc2 leads to the dephosphorylation of theproteins that were phosphorylated at the initiation of mitosis,resulting in exit from mitosis and the re-formation of interphasenuclei.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 20/40
In concert with dissolution of the nuclear lamina, the nuclearmembrane fragments into vesicles
As the nuclear laminadissociates, the nuclear
membrane fragments intovesicles. The B-type laminsremain bound to thesevesicles, while lamins A andC are released as free
dimers.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 21/40
Re-formation of the nuclearenvelope
1. The first step in reassembly of the nuclear envelope is thebinding of membrane vesiclesto chromosomes, which may
be mediated by both integralmembrane proteins and B-type lamins.
2. The vesicles then fuse,3. the nuclear lamina
reassembles,4. and the chromosomes
decondense
All i C ll h S i S f b

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 22/40
Figure 12-1. The major intracellular compartments of ananimal cell.
All Eucaryotic Cells Have the Same Basic Set of Membrane-enclosed Organelles

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 23/40
Figure 17-1. Overview of sorting of nuclear-encoded proteins in
eukaryotic cells

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 24/40
Endoplasmic reticulum (ER).
2 types: 1. rough ER-ribosomes on its outer surface, functions inprotein processing
- site of synthesis of proteins destined forsecretion
2. smooth ER-not associated with ribosomes and isinvolved in lipid, rather than protein, metabolism
-network of membrane-enclosed tubules and sacs (cisternae) thatextends from the nuclear membrane throughout the cytoplasm
-largest organelle of most eukaryotic cells
-functions as a store for Ca 2+ ions.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 25/40
The Endoplasmic Reticulum and Protein Secretion
The secretory pathway(Pancreatic acinar cells )
These experiments defined apathway taken by secretedproteins, the secretorypathway : rough ER → Golgi →
secretory vesicles → cellexterior.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 26/40
Two types of Protein translocation in cell1. Cotranslational translocation - Proteins can be
translocated into the ER either during their synthesis onmembrane-bound ribosomes (eg. mammalian cells)
2. Posttranslational translocation - Proteins can betranslocated into the ER after their translation has beencompleted on free ribosomes in the cytosol
* both cotranslational and posttranslational pathways are used inyeast.
Signal sequence
A hydrophobic sequence at the amino terminus of a polypeptide chain thattargets it for secretion in bacteria or incorporation into the endoplasmicreticulum in eukaryotic cells .
*signal recognition particle (SRP)

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 27/40
Proteins destined for incorporation into the plasma membrane or themembranes of the ER, Golgi, or lysosomes are initially inserted into the ER
membrane instead of being released into the lumen.
Cotranslational targeting of secretoryproteins to the ER
Posttranslational translocation of proteins into the ER

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 28/40
ER is also the1. site of protein folding,2. assembly of multisubunit proteins,
3. disulfide bond formation,4. the initial stages of glycosylation,5. and the addition of glycolipid anchors to some
plasma membrane proteins.*Indeed, the primary role of lumenal ER proteins is to catalyzethe folding and assembly of newly translocated polypeptides.
The Smooth ER and Lipid Synthesis -major site at which membrane lipids are synthesized in eukaryotic cells.-synthesized lipids are transported from the ER to their ultimatedestinations either in vesicles or by carrier proteins-abundant in cell types that are particularly active in lipid metabolism (eg.steroid-producing cells, such as those in the testis and ovary ; liver, where
it contains enzymes that metabolize various lipid-soluble compounds

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 29/40
flippases – membrane proteinscatalyze the rapidtranslocation
of phospholipidsacross the ERmembrane, resultingin even growth of both halves of thebilayer.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 30/40
Golgi apparatus-consists of organized stacks of disclike compartments called Golgi
cisternae; it receives lipids and proteins from the ER and dispatches them to a
variety of destinations, usually covalently modifying them en route Regions of the Golgi apparatusVesicles from the ER fuse to form theER-Golgi intermediate compartment,and proteins from the ER are thentransported to the cis Golgi network.Resident ER proteins are returnedfrom the ER-Golgi intermediatecompartment and the cis Golginetwork via the recycling pathway.The medial and trans compartmentsof the Golgi stack correspond to thecisternae in the middle of the Golgicomplex and are the sites of mostprotein modifications. Proteins arethen carried to the trans Golginetwork, where they are sorted for
transport to the plasma membrane,secretion, or lysosomes.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 31/40
-In addition to its activities in processing and sorting glycoproteins, the Golgiapparatus functions in lipid metabolism —in particular, in the synthesis of glycolipids and sphingomyelin-In plant cells, the Golgi apparatus has the additional task of serving as the site
where complex polysaccharides of the cell wall are synthesized (eg.hemicellulose and pectins)
Protein Sorting and Export from the Golgi Apparatus-involves the sorting of proteins into different kinds of transport
vesicles, which bud from the trans Golgi network and deliver their contents tothe appropriate cellular locations
Types of secretory pathway:1. constitutive secretory pathway – some proteins are carried from
the Golgi to the plasma membrane-accounts for the incorporation of new proteins and lipids
into the plasma membrane, as well as for the continuous secretion of proteins from the cell.
2. distinct pathway of regulated secretion - Other proteins are
transported to the cell surface

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 32/40
3. specifically targeted to other intracellular destinations - such aslysosomes in animal cells or vacuoles in yeast.
Transport from the Golgi apparatus

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 33/40
In yeasts and plant cells , which lack lysosomes, proteins aretransported from the Golgi apparatus to an additional destination: thevacuole
-assume the functions of lysosomes in these cells as well asperforming a variety of other tasks, such as the storage of nutrientsand the maintenance of turgor pressure and osmotic balance.
Lysosomes -contain digestive enzymes that degrade defunct
intracellular organelles, as well as macromolecules and particlestaken in from outside the cell by endocytosis. On their way tolysosomes, endocytosed material must first pass through aseries of organelles called endosomes .

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 34/40
The intracellular compartments of the eucaryotic cell involved in
the biosynthetic- secretory and endocytic pathways

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 35/40
Vesicular Transport
-a major cellular activity, responsible for molecular traffic between avariety of specific membrane-enclosed compartments.-understanding the molecular mechanisms that control vesiclepackaging, budding, and fusion is being considered
Coat Proteins and Vesicle Budding-first step in vesicular transport is the formation of a vesicle by buddingfrom the membrane.-cytoplasmic surfaces of transport vesicles are coated with proteins, and itappears to be the assembly of these protein coats that drives vesiclebudding by distorting membrane conformation
Three kinds of coated vesicles, which appear to function in different typesof vesicular transport:1. clathrin-coated vesicles
-responsible for the uptake of extracellular moleculesfrom the plasma membrane by endocytosis as well as the transport of molecules from the trans Golgi network to lysosomes

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 36/40
2. nonclathrin-coated or COP-coated vesicles (COP stands for coat protein)- coated vesicles identified as budding from the ER and Golgi complex2 classes:
1. COPII-coated vesicles - bud from the ER and carry their cargoforward along the secretory pathway , to the Golgi apparatus.
2. COPI-coated vesicles - bud from the ER-Golgi intermediatecompartment or the Golgi apparatus and function in the retrievalpathways that serve to retain resident proteins in the Golgi andER.
Incorporation of lysosomal proteins intoclathrin-coated vesicles

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 37/40
Vesicle Fusion-transport vesicle with its target involves two types of events:
1. transport vesicle must specifically recognize the correct
target membrane2. the vesicle and target membranes must fuse, thereby
delivering the contents of the vesicle to the target organelle.
vesicle fusion is mediated by interactions between specific pairs of proteins, calledSNAREs, on the vesicle and target membranes (v-SNAREs and t-SNAREs, respectively)
Rab proteins are a family of small GTP-binding proteins that are related to the Ras
proteins

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 38/40
Vesicle fusion - mediated by binding between specificpairs of v-SNAREs and t-SNAREs on thevesicle and target membranes,respectively. Rab GTP-binding proteinsare required to facilitate formation of v-SNARE/t-SNARE complexes.Following membrane fusion, theNSF/SNAP proteins disassemble theSNARE complex.

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 39/40
Organization of the lysosome
The lysosome system
Endocytosis andlysosomeformation

7/31/2019 Bio 108 Lec 8
http://slidepdf.com/reader/full/bio-108-lec-8 40/40
Peroxisomes- are small vesicular compartments that contain enzymes utilized
in a variety of oxidative reactions.
Mitochondria and (in plants) chloroplasts generate most of the ATP used by cells to drive reactions that require an input of free energy;chloroplasts are a specialized version of plastids, which can also have otherfunctions in plant cells, such as the storage of food or pigment molecules
Recommended

















