Embed Size (px)
Citation preview

BMCP – Cycle cellulaire
04/02/2014FILIPPI Laura L2BMCPA. Touati ([email protected])20 pagesRelecteur 5
Cycle cellulaire
La prof a refusé que l'on prenne le diapo en photo, je n'ai donc pas pu photographier les schémas. Le diapo sur l'ENT n'est pas celui qu'elle nous a projeté c'est pourquoi je n'ai pas pu intégrer tous les schémas, désolée..
Deuxième semestre de BMCP :• Notation indépendante de l'UE indépendante du 1er quad (il faut 10 dans les deux épreuves, elles ne se
compensent pas)• 20h de cours magistraux• + 6h de TP/ED présence obligatoire (du coup, pas de possibilité d'avoir de dette dans cette matière)• 3 ECTS (système européen de transfert et cumul des crédits)
Contrôle des connaissances : – Examen de fin d'année– QR courtes (problème transversal)– 30 minutes– Note sur 14 : + note ED sur 6 (chaque ED compte 2 points)
ENT : Plan et schémas du cours, problème résolus et annales (qui datent des années précédentes et qui correspondent donc à des épreuves de 45 min et pas 30min)
Les objectifs de ED seront mis en ligne sur l'ENT en milieu de semaine prochaine (aux alentours du 12).Certains ED nécessitent une préparation en amont, un prérequis qu'on nous demande de consulter et que l'on doit connaître (mais ce n'est pas le cas du 1er ED)
ED 1 : dosage plasmatiques et urinaires des protéines (pas de travail personnel pour ce TP)ED 2 : Imagerie et microscopie (prérequis à avoir avant ED)ED 3 : Potentiel thérapeutique des cellules souches (prérequis à avoir avant ED + articles scientifiques à lire avant et l’interrogation portera sur un de ces articles)
1/20
Plan
A. Rappel sur la notion de cycle cellulaire : les 4 phases et G0
B. Les points majeurs du cycle cellulaire I. Le point de restriction R en G1 II. Les points de transition en G1/S et en G2/M III.Mise en évidence expérimentale IV. Méthodes d’étude du cycle cellulaire
C. Le système de contrôle du cycle du cellulaire I. Les partenaires moléculaires
D. Conclusion

BMCP – Cycle cellulaire
A. Rappel sur la notion de cycle cellulaire : les 4 phases et G0
Le déroulement du cycle cellulaire va aboutir à l'obtention de deux cellules filles de même patrimoine génétique. Une cellule n'est pas constamment en train de se diviser :
– Une cellule peut être quiescente c'est la phase G0 qui correspond à un état de repos (dans l'incapacité a entrer en division).
– La sénescence est le processus de vieillissement cellulaire. C'est un phénomène ordonné, régulé. Chaque type de cellule peut se diviser un certain nombre de fois. Il y a une marge importante (par exemple) entre la cellule œuf qui est capable de se diviser un grand nombre de fois, et des cellules qui ne peuvent pratiquement pas se diviser comme les neurones
– L'apoptose est la mort cellulaire programmée. C'est un système de barrière et de protection pour ne pas transmettre d'anomalies génétiques aux cellules filles. Elle a également de nombreux rôles importants notamment dans le renouvellement cellulaire, la régénération cellulaire et permet également des gains de fonction.
La cellule doit aussi se différencier, acquérir des caractéristiques morphologiques mais aussi fonctionnelles et cette différenciation s'inscrit aussi dans le cadre du cycle cellulaire.
Au cours du cycle 2 étapes importantes :– Celle qui conduit à la division cellulaire– Celle où la cellule se prépare a cette division cellulaire
Rôles du cycle cellulaire :
• Chez les eucaryotes, il régule : – la croissance cellulaire– le nombre de cellules
• Chez les organismes unicellulaires (levures) : ont pour objectif d'effectuer leur croissance et de se diviser. Les nutriments dont ils ont besoin pour croître sont présents dans son environnement. Si les conditions sont réunies, l'organisme va pouvoir se diviser. Rôle d'adaptation à l'environnement.
• Chez les organismes multicellulaires : La cellule n'est pas isolée, elle est au sein d'un tissu qui, réunis, vont former des organes. L'activation de la division va dépendre des signaux extérieurs : la cellule va recevoir des signaux positifs ou négatifs pour savoir si oui ou non elle doit rentrer en division. Ce phénomène suggère fortement des mécanismes de régulation.Notion d'équilibre des différentes populations cellulaires entre elles car ce cycle cellulaire est important pour la morphogénèse, le renouvellement des tissus, la croissance physiologique, la réparation tissulaire...
Contrôle du cycle cellulaire : à tous les niveaux du cycle
– Le cycle cellulaire est régulé précisément par l'environnement cellulaire par des signaux prolifératifs (+) ou antiprolifératifs (-) :
– Il y a aussi des systèmes de contrôle à différents niveaux du cycle cellulaire. Cela permet de stopper le cycle en cas d'anomalies.
Par exemple, la cellule cancéreuse prolifère de manière indépendante et anarchique ; n'a pas besoin de signaux prolifératifs ou antiproliféraifs. Elle ne vieillit pas, ne rentre pas en appotose et donc devient immortelle. Elle échappe au contrôle social de la cellule.
2/20

BMCP – Cycle cellulaire
Prix nobel pour 3 chercheurs : (note du CR:la prof est passé très rapidement dessus)
– Leland Hartwell (né en 1939), Fred Hutchinson Cancer Research center ? Seattle (USA), est récompensé pour la découverte d'une catégorie de gènes qui commandent le cycle cellulaire. L'un de ces gènes joue un rôle central dans le démarrage de chaque nouveau cycle, d'où son nom « start ». Hartwell a introduit également la notion de « points de contrôle » (checkpoints), conduisant à une approche novatrice du cycle cellulaire.
– Paul Nurse (né en 1949), Imperial Cancer Research Fund, Londres, a identifié par des méthode de génie génétique et de biologie moléculaire l'un des facteurs clés de la régulation du cycle cellulaire, les kinases dépendantes des cyclines (CDK). Il a montré que la fonction des CDK s'est conservée tout au long de l'évolution. Les CDK activent le cycle cellulaire par le biais d'une réaction chimique (phosphorylation) avec d'autres protéines.
– Tim Hunt (né en 1943), Imperial Cancer Research Fund, Londres, est récompensé pour la découverte des cyclines, les protéines qui régulent l'action des CDK. Il a mis en évidence la dégradation des cyclines lors de la division des cellules, un mécanisme qui s'est avéré essentiel pour la régulation du cycle cellulaire.
Les quatre phases du cycle cellulaire et le stade G0
La phase G0 est hors du cycle car il faut que la cellule passe en G1 pour rentrer dans le cycle.
• Phase G1 : – La quantité d'ADN reste fixe. – La transcription et la traduction sont très importantes.– C'est une phase obligatoire mais de durée variable, elle fait la spécificité d'un type cellulaire donné
et elle peut amener à passer en phase G0. – La cellule se prépare à la croissance (phase S). La phase G0 peut être définitive comme dans les neurones ou alors temporaire comme dans les hépatocytes par exemple. C'est pour cela que certaines lésions du foie peuvent être réparées, on va avoir des cellules capables de re-entrer en division et de reconstituer l'organe tel qu'il était auparavant et ce sous l'influence de signaux du milieu.
• Phase S : – Phase de réplication de l'ADN, le matériel génétique est doublé. – Il y a persistance de la traduction et de la transcription
3/20
BMCP – Cycle cellulaire
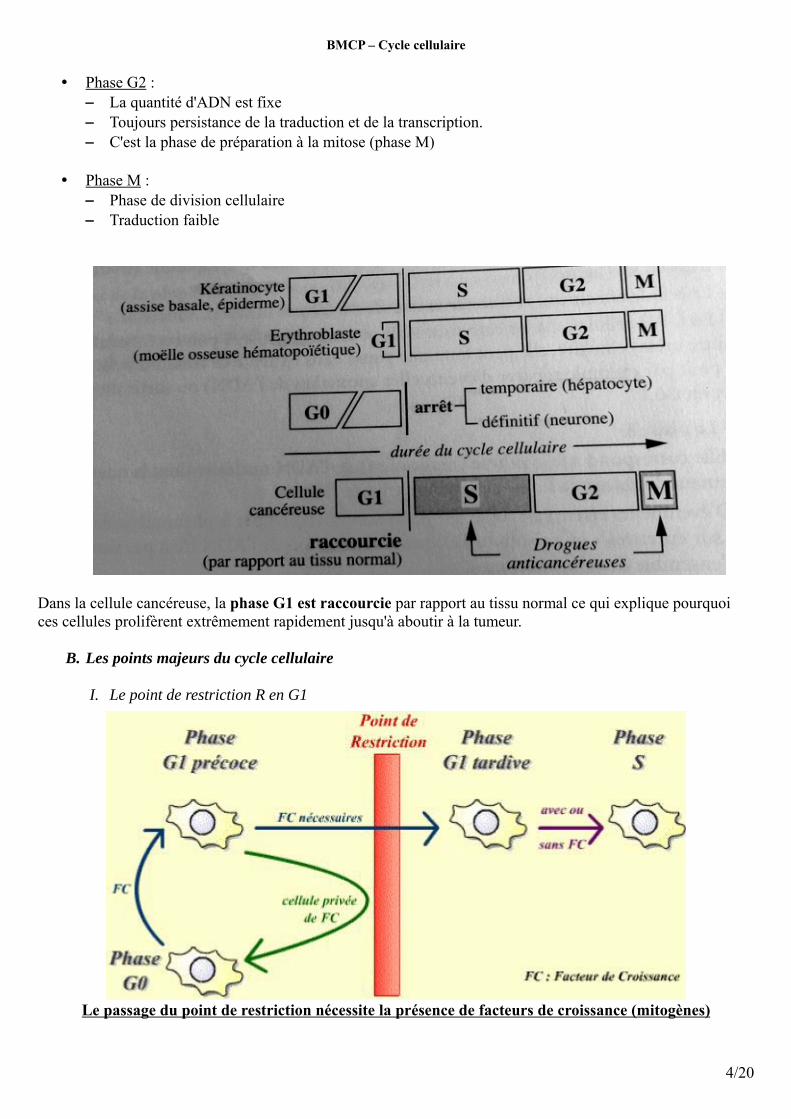
• Phase G2 : – La quantité d'ADN est fixe– Toujours persistance de la traduction et de la transcription. – C'est la phase de préparation à la mitose (phase M)
• Phase M : – Phase de division cellulaire – Traduction faible
Dans la cellule cancéreuse, la phase G1 est raccourcie par rapport au tissu normal ce qui explique pourquoi ces cellules prolifèrent extrêmement rapidement jusqu'à aboutir à la tumeur.
B. Les points majeurs du cycle cellulaire
I. Le point de restriction R en G1
Le passage du point de restriction nécessite la présence de facteurs de croissance (mitogènes)
4/20

BMCP – Cycle cellulaire
NB : Dans ce cours on considère que facteur de croissance = facteur mitogène (bien qu'il y ait quelques différences, les facteurs de croissance agissent plutôt sur la croissance et non la division cellulaire).
Le point R (beaucoup étudié chez la levure) est très important et se situe dans le 1er tiers de G1. Pour arriver jusqu'en G1 la cellule a besoin de signaux positifs de prolifération (donc de facteurs mitogènes ou de croissance). Si l'environnement cellulaire n'est pas permissif, la cellule ne va pas franchir ce point de repère et ne rentrera pas en G1.
Il faut bien différencier dans la phase G1 deux parties • celle avant le point R ou G1 précoce : la cellule a besoin de facteurs de croissance.• celle après le point R ou G1 tardive : la cellule s'est affranchie des facteurs de croissance pour avancer
dans le cycle, elle n'a plus besoin des facteurs pour continuer.
La cellule cancéreuse n'a pas besoin de FC pour proliférer de façon complètement incontrôlée, elles n'ont pas besoin de l'environnement cellulaire.
II. Les points de transition en G1/S et en G2/M
Deux autres points très importants du cycle cellulaire : l'entrée en phase S et l'entrée en phase M.
La succession des phases G1/S/G2/M est immuable et doit être totalement respectée donc il va falloir mettre en place un système de surveillance.Le système de contrôle étudie comment assurer la succession immuable des différentes phases qui caractérisent ce cycle cellulaire. Le système de contrôle comporte un dispositif biochimique qui agit de façon cyclique formé d'un ensemble de protéines interactives, et contrôlée par des « freins ». Le contrôle est assuré par des complexes protéiques CDK-cyclines.
Comment assurer la succession immuable des différentes phases du cycle ?
1 ) Les cyclines et les CDK de la phase G1, de G1/S et S et de M.
Il existe différentes familles de CDK et de cyclines, certaines vont intervenir à des moments du cycle et d'autres à des moments différents. On ne nous demande pas d'apprendre, à quel moment les cyclines vont intervenir, ni quand les CDK1 ou 2 vont intervenir. Il faut bien comprendre que ces complexes se forment de façon spécifique à différents moments du cycle et permettent l'avancée de ce cycle. Globalement on peut dire qu'il y a les complexes CDK-cycline de la phase G1, ceux de la transition G1/S et S et ceux de la phase M.
5/20

BMCP – Cycle cellulaire
2 ) L'anaphase Promoting Complex (APC).
Ce sont des ubiquitine ligases qui agissent au cours de la mitose. L'APC permet de détruire la cohésine qui est une protéine qui retient les chromatides sœurs entre elles, ce qui va donc conduire à leur séparation et donc permettre l'anaphase.
III.Méthodes d’étude du cycle cellulaire
Le contrôle qualité : comment assurer la surveillance du bon déroulement du cycle ?
1) Contrôle des dommages de l'ADN :• Avant l'entrée en phase S (G1)• Au cours de la phase S• Après la réplication de l'ADN (G2)
Si il y dommages de l'ADN, la cellule arrête le cycle et va essayer de réparer les dommages. Soit elle répare et reprend le cycle, soit elle ne peut pas réparer et entre en apoptose. Ce contrôle permanent est indispensable pour éviter de transmettre l'anomalie.
2) Contrôle du fuseau mitotique. Uniquement au cours de la métaphase (mitose). Contrôle que les chromosomes soit rassemblés sur la plaque métaphasique, que le fuseau soit bien constitué pour une bonne répartition des chromosomes.
Les outils pour détecter les anomalies de l'ADN et interrompre le cycle
1) La p53 : elle peut détecter à tout moment une anomalie et arrêter le cycle cellulaire. Elle active alors les systèmes de réparation : soit il y a réparation et reprise du cycle, soit elle induit l'apoptose de la cellule. La p53 est codée par un gène suppresseur de tumeur.
2) Des kinases : ATM (Ataxia Telangiectasia Mutated) et ATR (Ataxia Telangiectasia Related) détectent les anomalies et interviennent dans les phénomènes de réparation. On a également les kinases Chk1 et 2 (Checkpoint Protein Kinases).
3) La protéine MAD (Mitotic Arrest Deficient) : intervient lors de la mitose et bloque l'anaphase.
6/20
BMCP – Cycle cellulaire
IV. Mise en évidence expérimentale
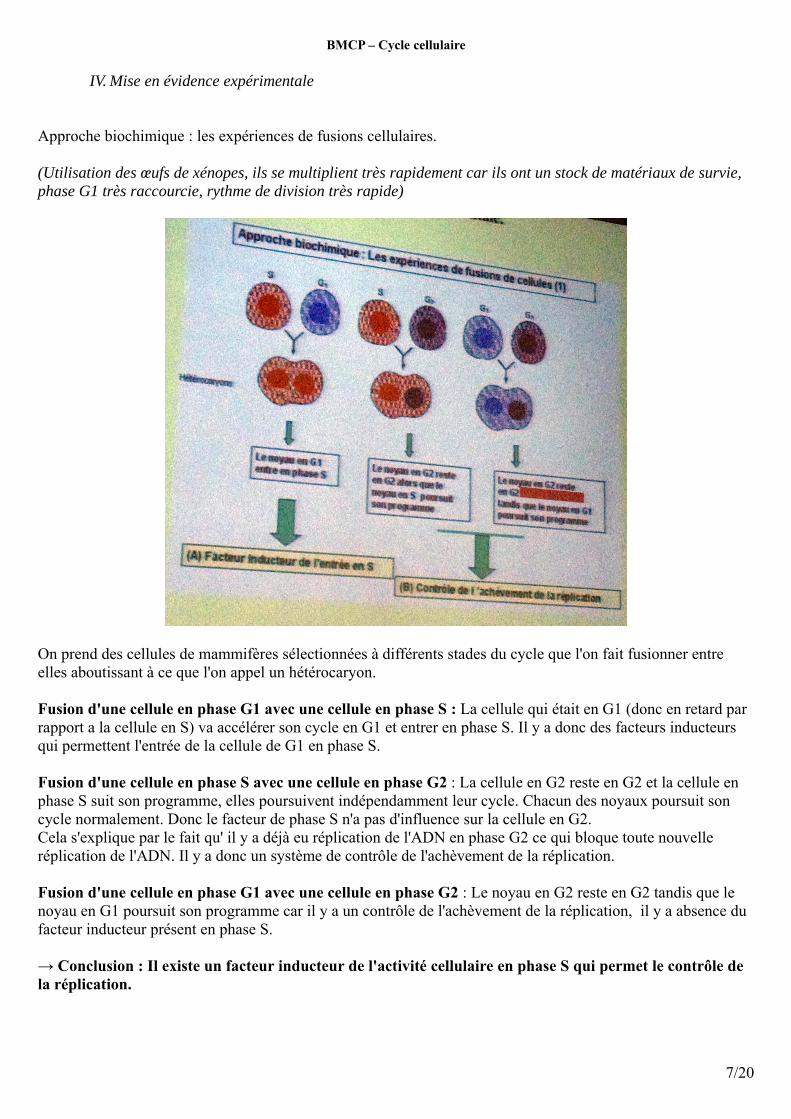
Approche biochimique : les expériences de fusions cellulaires.
(Utilisation des œufs de xénopes, ils se multiplient très rapidement car ils ont un stock de matériaux de survie, phase G1 très raccourcie, rythme de division très rapide)
On prend des cellules de mammifères sélectionnées à différents stades du cycle que l'on fait fusionner entre elles aboutissant à ce que l'on appel un hétérocaryon.
Fusion d'une cellule en phase G1 avec une cellule en phase S : La cellule qui était en G1 (donc en retard par rapport a la cellule en S) va accélérer son cycle en G1 et entrer en phase S. Il y a donc des facteurs inducteurs qui permettent l'entrée de la cellule de G1 en phase S.
Fusion d'une cellule en phase S avec une cellule en phase G2 : La cellule en G2 reste en G2 et la cellule en phase S suit son programme, elles poursuivent indépendamment leur cycle. Chacun des noyaux poursuit son cycle normalement. Donc le facteur de phase S n'a pas d'influence sur la cellule en G2. Cela s'explique par le fait qu' il y a déjà eu réplication de l'ADN en phase G2 ce qui bloque toute nouvelle réplication de l'ADN. Il y a donc un système de contrôle de l'achèvement de la réplication.
Fusion d'une cellule en phase G1 avec une cellule en phase G2 : Le noyau en G2 reste en G2 tandis que le noyau en G1 poursuit son programme car il y a un contrôle de l'achèvement de la réplication, il y a absence du facteur inducteur présent en phase S.
→ Conclusion : Il existe un facteur inducteur de l'activité cellulaire en phase S qui permet le contrôle de la réplication.
7/20

BMCP – Cycle cellulaire
On obtient des résultats similaires in vitro avec des extraits cellulaires prélevés à des stades différents du cycle cellulaire, on ajoute de l'ADN et on regarde si on obtient une réplication ou pas :
On peut obtenir une réplication avec des extraits cellulaires en phase S mais pas en G1 ni en G2. Les cellules en phase S contiennent donc une activité capable d'induire la réplication. On va ensuite caractériser quelle est cette activité et de quoi elle dépend. On s'est rendu compte que l'activité était dépendante d'une kinase CDK (initialement appelée CDC2, mais ce n'est pas le nom a retenir, je l'ai mis parce qu'on peut la retrouver sur certains schémas) activée par sa liaison avec une cycline.
→ Il faut que la réplication soit achevée pour que la cellule rentre en phase G2 et M, donc nécessité de contrôler la réplication et maintenir l'ordre dans l’enchaînement des différentes phases.→ SPF : - Déclenche l'entrée en S d'une cellule en phase G1
- Disparait en fin de S et n'est pas présent dans les cellules en G2 - Pas d'inhibiteur dans les cellules en G2→ signification biochimique : les cellules en phase S contiennent une activité capable d'induire la réplication dans un système in vitro. Cette activité dépend de la protéine cdc2.
Réplication : notion de SPF (S Promoting Factor : facteur inducteur de l'entrée en phase S)• 1 seule réplication par cycle cellulaire• Réplication obligatoire pour l'entrée en mitose• Le SPF ne peut induire la réplication que dans les cellules compétentes (en phase G1)• Les cellules doivent passer par la mitose (M) pour devenir compétentes.
==> Il y a des points de contrôle « checkpoint » : la mitose est retardée jusqu'à ce que le génome soit entièrement répliqué.
8/20
BMCP – Cycle cellulaire
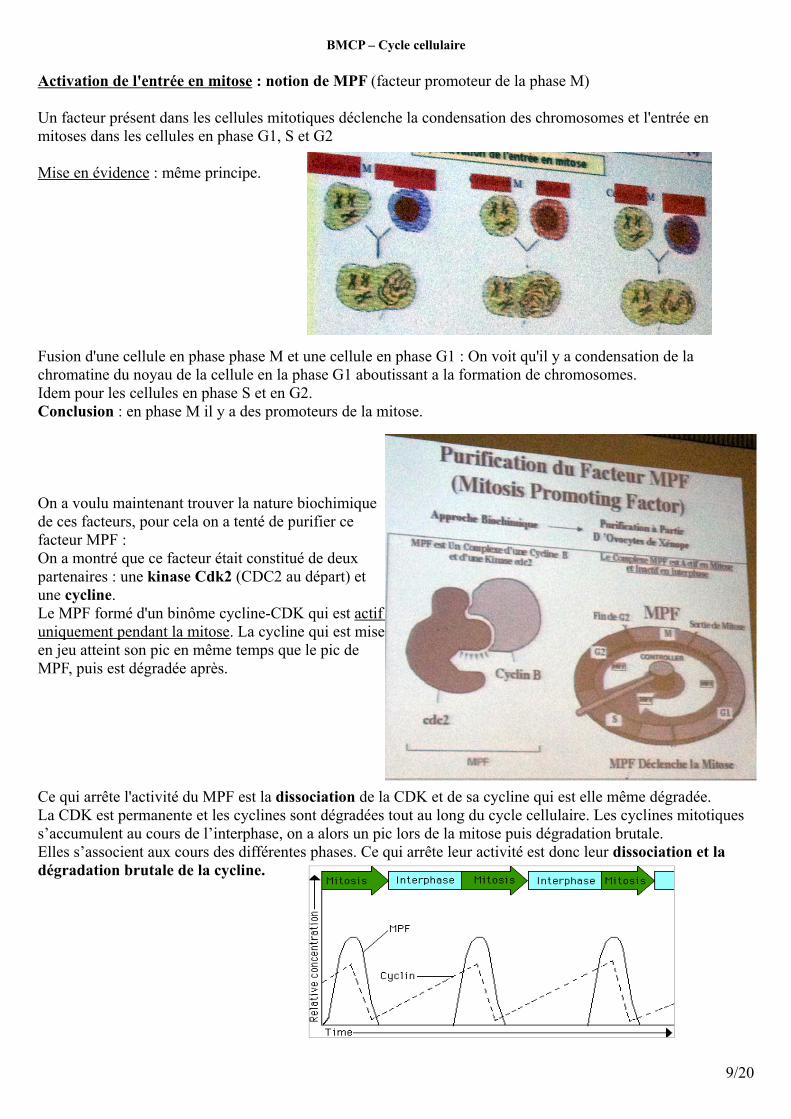
Activation de l'entrée en mitose : notion de MPF (facteur promoteur de la phase M)
Un facteur présent dans les cellules mitotiques déclenche la condensation des chromosomes et l'entrée en mitoses dans les cellules en phase G1, S et G2
Mise en évidence : même principe.
Fusion d'une cellule en phase phase M et une cellule en phase G1 : On voit qu'il y a condensation de la chromatine du noyau de la cellule en la phase G1 aboutissant a la formation de chromosomes.Idem pour les cellules en phase S et en G2.Conclusion : en phase M il y a des promoteurs de la mitose.
On a voulu maintenant trouver la nature biochimique de ces facteurs, pour cela on a tenté de purifier ce facteur MPF :On a montré que ce facteur était constitué de deux partenaires : une kinase Cdk2 (CDC2 au départ) et une cycline.Le MPF formé d'un binôme cycline-CDK qui est actif uniquement pendant la mitose. La cycline qui est mise en jeu atteint son pic en même temps que le pic de MPF, puis est dégradée après.
Ce qui arrête l'activité du MPF est la dissociation de la CDK et de sa cycline qui est elle même dégradée.La CDK est permanente et les cyclines sont dégradées tout au long du cycle cellulaire. Les cyclines mitotiques s’accumulent au cours de l’interphase, on a alors un pic lors de la mitose puis dégradation brutale.Elles s’associent aux cours des différentes phases. Ce qui arrête leur activité est donc leur dissociation et la dégradation brutale de la cycline.
9/20
BMCP – Cycle cellulaire
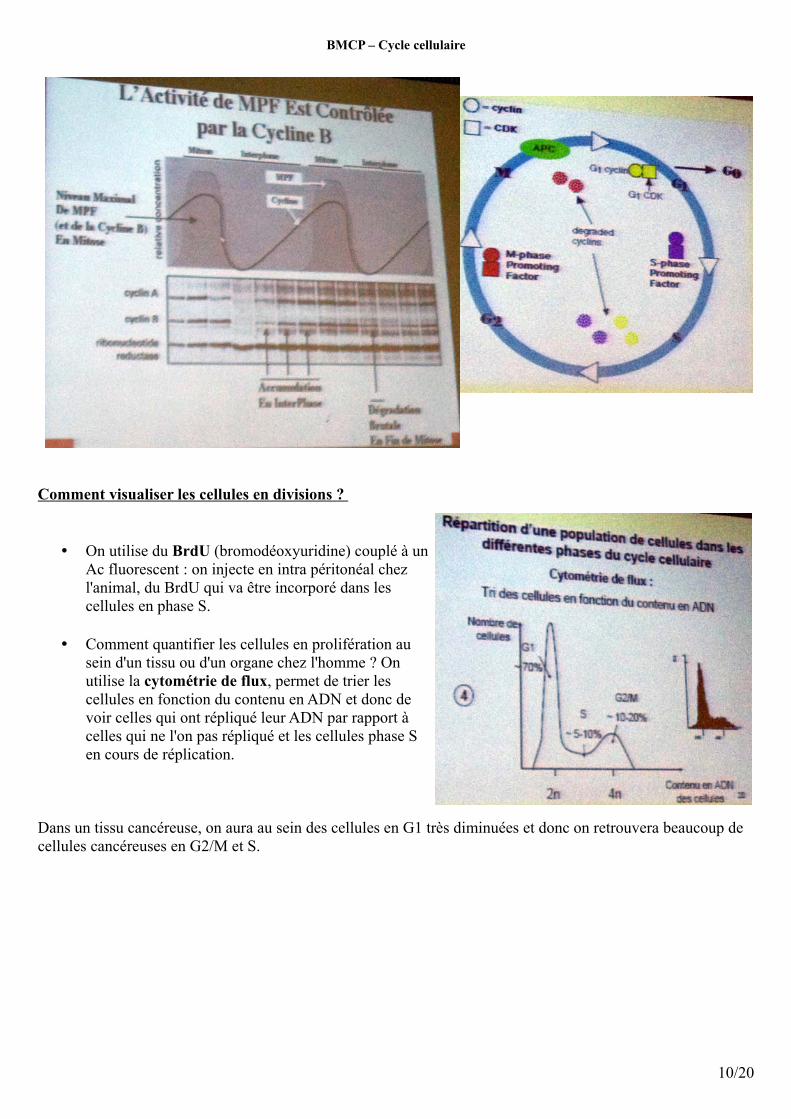
Comment visualiser les cellules en divisions ?
• On utilise du BrdU (bromodéoxyuridine) couplé à un Ac fluorescent : on injecte en intra péritonéal chez l'animal, du BrdU qui va être incorporé dans les cellules en phase S.
• Comment quantifier les cellules en prolifération au sein d'un tissu ou d'un organe chez l'homme ? On utilise la cytométrie de flux, permet de trier les cellules en fonction du contenu en ADN et donc de voir celles qui ont répliqué leur ADN par rapport à celles qui ne l'on pas répliqué et les cellules phase S en cours de réplication.
Dans un tissu cancéreuse, on aura au sein des cellules en G1 très diminuées et donc on retrouvera beaucoup de cellules cancéreuses en G2/M et S.
10/20
BMCP – Cycle cellulaire
C. Le système de contrôle du cycle du cellulaire
I. Les partenaires moléculaires
a) Les cyclines :
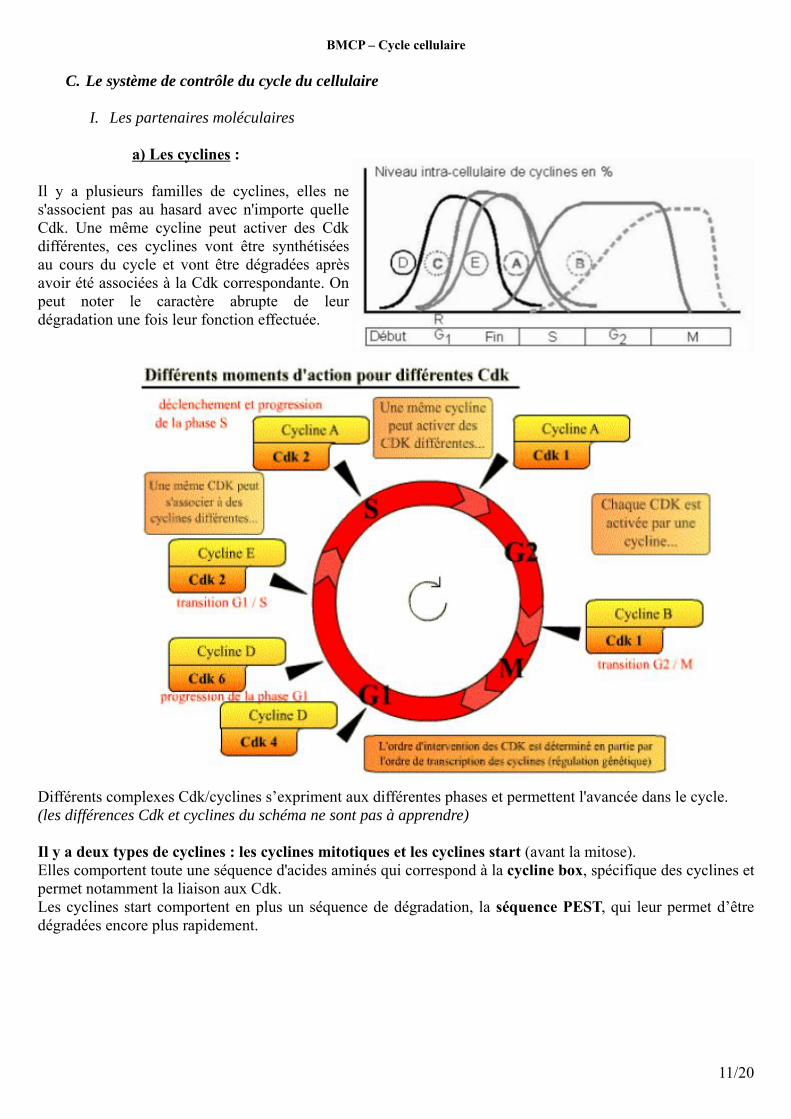
Il y a plusieurs familles de cyclines, elles ne s'associent pas au hasard avec n'importe quelle Cdk. Une même cycline peut activer des Cdk différentes, ces cyclines vont être synthétisées au cours du cycle et vont être dégradées après avoir été associées à la Cdk correspondante. On peut noter le caractère abrupte de leur dégradation une fois leur fonction effectuée.
Différents complexes Cdk/cyclines s’expriment aux différentes phases et permettent l'avancée dans le cycle. (les différences Cdk et cyclines du schéma ne sont pas à apprendre)
Il y a deux types de cyclines : les cyclines mitotiques et les cyclines start (avant la mitose).Elles comportent toute une séquence d'acides aminés qui correspond à la cycline box, spécifique des cyclines et permet notamment la liaison aux Cdk. Les cyclines start comportent en plus un séquence de dégradation, la séquence PEST, qui leur permet d’être dégradées encore plus rapidement.
11/20

BMCP – Cycle cellulaire
• Localisation des cyclines mitotiques :
La cyline B est un cycline mitotique.
Marquage par immunohistochimie avec des AC fluorescents
On voit que la cycline B en G2 est localisée dans le cytoplasme, elle est en cours de synthèse et passe ensuite dans le noyau juste avant la mitose M. Elle est fortement exprimée pendant la prophase (noyau très fluorescent), ensuite pendant la métaphase on va avoir une destruction de cette cycline B et concentration aux deux pôles du fuseau.Donc le premier élément de la régulation des cyclines est la variation de la localisation cytoplasmique et nucléaire de cette cycline au cours du cycle cellulaire. Les cyclines A et B sont fortement synthétisées avant la mitose, on a accumulation au cours de l'interphase avec un pic maximal au cours de la mitose, puis elles sont dégradées brutalement en fin mitose.
• La synthèse de cycline B régule le cycle cellulaire : On cherche à prouver le rôle de la cycline B dans le cycle cellulaire
• Expérience 1
→ A gauche : le taux de cycline est normal
→ Au milieu : on a traité nos cellules avec de la RNase (enzyme dégradant de façon spécifique l'ARN), on s'aperçoit alors que le cycle cellulaire est bloqué. Donc sans cycline B, il n'y a pas d'entrée dans le cycle cellulaire.
→ A droite : on traite les cellules avec une RNase (donc le cycle est arrêté) et on supplée la cellule en cycline B. A ce moment la on observe la reprise de la mitose et du cycle.
• Expérience 2 → On ajoute un ARNm antisens (c'est un ARNm complémentaire (oligonucléotide) de la séquence de l'ARNm d’intérêt, cette liaison détruit l'ARNm cible par dégradation spécifique par une RNase) donc il n'y a pas production de la protéine en question.L'ARNm qui code pour la cycline B est donc détruit, il n'y a pas de synthèse de cycline B et donc pas de mitose, la cellule est bloquée en interphase.Ce sont donc les cyclines B qui permettent l'entrée en mitose, elle régule l'entrée dans le cycle et permettent la division cellulaire.
12/20

BMCP – Cycle cellulaire
Mais la dégradation de la cycline B est requise également pour permettre la sortie de la mitose et l'inactivation du MPF : On évalue l’activité du MPF (CDK+cycline) et on dose la cycline. On retrouve bien l'expression très importante de la cycline au cours de la mitose et dégradation brusque a la fin de la mitose.
→ Au lieu de mettre de l'ARNm antisens avec les ARNm de la cycline B, on met de la cycline Bδ 90 qui est une cycline qui ne peut pas être dégradée par le système de l'ubiquitine, donc la cellule ne sort pas de la mitose. On voit que le MPF au lieu de diminuer reste à un même taux, la cycline continue à augmenter et donc la cellule est arrêtée en mitose, elle ne peut donc pas rentrer en interphase.
Il faut que non seulement il y ait synthèse des cyclines mais aussi dégradation de celle-ci pour que le système fonctionne. Et ceci est valable pour toutes les cyclines du cycle cellulaire, elles fonctionnent toute de la même façon.
b) Les CDK, protéines kinases dépendantes des cyclines
Ce sont des kinases dépendantes des cyclines. Elles sont dépendantes de leur association avec les cyclines pour être actives. Elles ont une structure particulière bilobaire, présente chez toutes les cyclines, avec un domaine en N-ter très conservé qui permet la liaison à la cycline. Les complexe CDK-cycline vont aller phosphoryler des substrats nécessaires à l'avancée du cycle cellulaire.Si la CDK n'est pas associée la cycline, elle est inactive. Lorsque la cycline et sa CDK correspondante s'associent, il y a modification de la conformation de la CDK qui découvre deux poches de fixation de la CDK à l'ATP et au substrat leur permettant de se rapprocher et ainsi de phosphoryler le substrat par transfert d'un groupement phosphate depuis l'ATP sur une serine ou une thréonine.Toutes les CDK sont caractérisées par deux poches de fixation l’une pour la protéine cible l'autre pour l'ATP. C'est la liaison a la cycline qui rend ces deux poches de fixation accessible à l'ATP et au substrat.On note au passage que la CDK est elle même phosphorylée (traitée plus en détail plus loin dans le cours)
13/20

BMCP – Cycle cellulaire
Quels sont les substrats ? • Phase G1 : La CDK-cycline start phosphoryle une protéine cible, la protéine du rétinoblastome (RB), cette phosphorylation permet de franchir le point R de la phase G1. Cette protéine RB est régulatrice du cycle cellulaire, tout comme p53.
Mise en évidence du rôle de la phosphorylation de la protéine RB au cours de la phase G1 :
→ P-RB
→ RB
→ Au Western blot, on utilise deux types d'AC : un qui reconnaît la protéine du RB et un qui reconnaît la forme phosphorylée de cette même protéine (P-RB). On voit que la CDK va phosphoryler la protéine RB qui va se trouver de plus en plus sous forme phosphorylée au western blot, on voit aussi qu'elle migre moins loin car son PM augmente. Donc premier exemple de substrat du CDK-cycline : la protéine du rétinoblastome.
Effet du complexe CDK-cycline :
– Il va phosphoryler des lamines qui vont permettre le désassemblage de la lamina nucléaire donc il y a désassemblage du noyau qui est le premier événement de l'entrée en mitose.
– Fragmentation de l'appareil de Golgi qui intervient au cours de la mitose. Au lieu d'avoir « un gros sac golgien » avec des vésicules empilées les unes sur les autres, et bien il va être fragmenté par le complexe CDK-cycline. Le complexe va aller phosphoryler les protéines G de la famille Rab qui bloque la fusion vésiculaire. → Donc inhibition de la fusion vésiculaire par phosphorylation des protéines G Rab par le complexe.
• Phase M : Autre substrat des complexes CDK-cycline : les microtubules. Il agit sur l'organisation des microtubules, notamment leur longueur.
Visualisation de l'organisation des microtubules :
Effet de la liason cycline B-cdc2 sur la longeur des MT
14/20

BMCP – Cycle cellulaire
On a pris les centrioles de l'interphase, on les met dans un tube à essai on ajoute de la tubuline et le complexe CDKt-cycline et on regarde l’évolution au cours du temps : Il y a modification de la longueur des microtubules ce qui correspond à la réorganisation des microtubules au cours de la mitose. Ce qui est caractéristique c'est qu'au départ on a des MT de longueur variable et au cours du temps, il y a raccourcissement de ceux ci qui correspond à l'avancée dans le cycle cellulaire. → En interphase les MT à proximité du centre cellulaire sont longs, → Prophase : MT courts et dynamiques→ Métaphase : MT très courts et très dynamiquesAu cours de la mitose ils commencent à se raccourcir puis deviennent extrêmement dynamique au cours de la métaphase.
A la fin de la mitose, les complexes sont impliqués aussi dans le mécanisme de cytodiérèse (séparation des cellules filles à la fin de la mitose). La cytodiérèse dépend de la myosine qui va former un anneau contractile qui va séparer la cellule en deux cellules filles. Le complexe CDK-cycline agis en phosphorylant et en déphosphorylant la myosine et permet donc le mécanisme de cytodiérèse.
• Phase S : Complexe CDK-cycline de la phase S vont phosphoryler le complexe de pré réplication favorisant la réplication de l'ADN.
En résumé, les substrats des complexe CDK-cyclines sont :
Pour la phase G1 : La protéine du RBPour la phase M : • les lamines pour le désassemblage du noyau
• les protéines G Rab pour la fragmentation du GolgiPour la fin de la mitose : la myosine pour la cytodiérèse.Pour la phase S : Le complexe de pré réplication
(la liste n'est pas exhaustive)
c) Mode de fonctionnement des complexes CDK - cycline
Ex : cycline B/CDK1 (cycline mitotique)
Première étape très importante : elles doivent s'associer entre elles → modification de la conformation de la CDK1 ce qui permet la liaison du substrat et de ATP et la phosphorylation du substrat.
15/20
BMCP – Cycle cellulaire
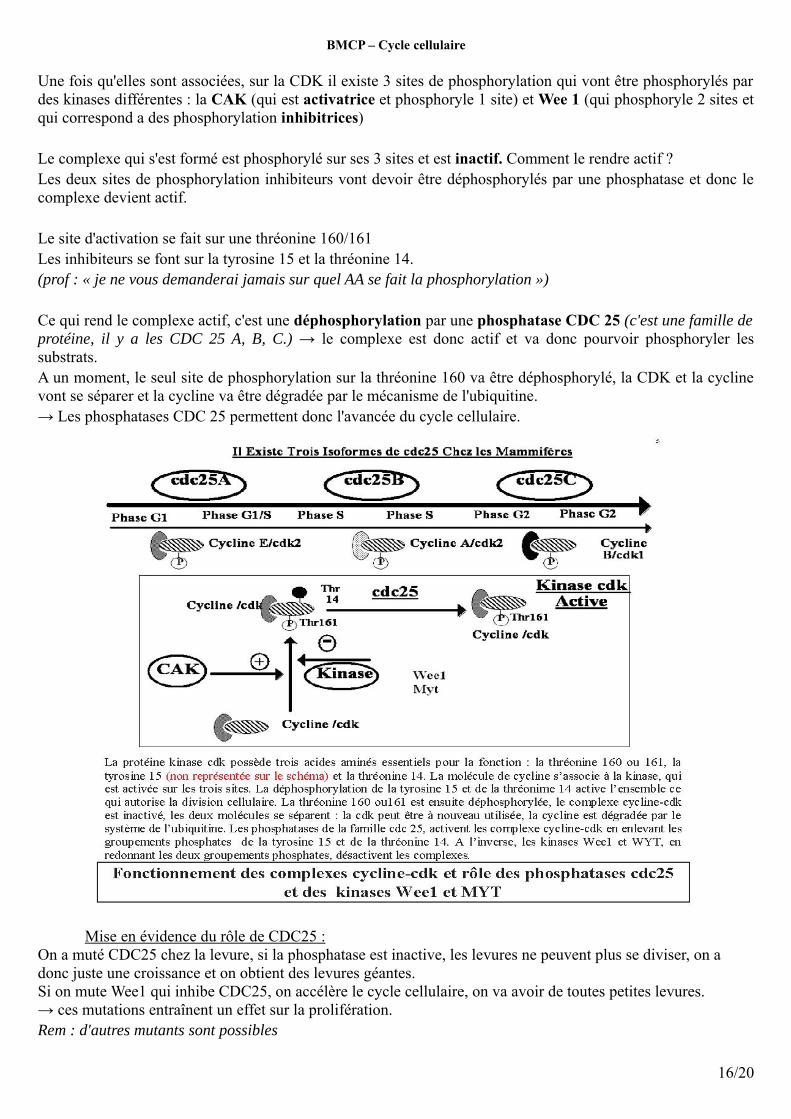
Une fois qu'elles sont associées, sur la CDK il existe 3 sites de phosphorylation qui vont être phosphorylés par des kinases différentes : la CAK (qui est activatrice et phosphoryle 1 site) et Wee 1 (qui phosphoryle 2 sites et qui correspond a des phosphorylation inhibitrices)
Le complexe qui s'est formé est phosphorylé sur ses 3 sites et est inactif. Comment le rendre actif ? Les deux sites de phosphorylation inhibiteurs vont devoir être déphosphorylés par une phosphatase et donc le complexe devient actif.
Le site d'activation se fait sur une thréonine 160/161Les inhibiteurs se font sur la tyrosine 15 et la thréonine 14. (prof : « je ne vous demanderai jamais sur quel AA se fait la phosphorylation »)
Ce qui rend le complexe actif, c'est une déphosphorylation par une phosphatase CDC 25 (c'est une famille de protéine, il y a les CDC 25 A, B, C.) → le complexe est donc actif et va donc pourvoir phosphoryler les substrats.A un moment, le seul site de phosphorylation sur la thréonine 160 va être déphosphorylé, la CDK et la cycline vont se séparer et la cycline va être dégradée par le mécanisme de l'ubiquitine.→ Les phosphatases CDC 25 permettent donc l'avancée du cycle cellulaire.
Mise en évidence du rôle de CDC25 : On a muté CDC25 chez la levure, si la phosphatase est inactive, les levures ne peuvent plus se diviser, on a donc juste une croissance et on obtient des levures géantes.Si on mute Wee1 qui inhibe CDC25, on accélère le cycle cellulaire, on va avoir de toutes petites levures. → ces mutations entraînent un effet sur la prolifération.Rem : d'autres mutants sont possibles
16/20

BMCP – Cycle cellulaire
d) Régulation de l’activité des complexes CDK – cycline
- Pour le site activateur (Thr 160/161) : la CAK (kinase activatrice) et KAP (phosphatase inhibitrice).- Pour les sites inhibiteurs (Thr 14 et Tyr 15) : la Wee1 (kinase inhibitrice) et Cdc25 (phosphatase activatrice).
- Les protéines inhibitrices des CDK :
• La famille CIP/KIP • La famille INK4 (Inhibitor of Kinase 4)
• INK4 est une famille de protéines inhibitrices de la phase G1 dont font partie les protéines p16 et p15. Elles empêchent la formation des complexes cycline-CDK en se fixant à la CDK4. Ce sont des inhibiteurs spécifiques de la phase G1 en agissant sur la CDK 4.A partir du même locus que p16, on peut avoir deux protéines synthétisées, la protéine p19. Les deux protéines interviennent à différents niveaux (traitées dans le prochain cours)
• CIP/KIP : inhibiteurs du cycle cellulaire qui interviennent à tous les niveaux du cycle. Les protéines P21/ P27.Elles bloquent le complexe mais n’empêchent pas sa formation contrairement à p16 et p15, mais elles sont capables d'inhiber tous les complexes CDK-cyclines. → inhibiteur général.
Lésionde l'ADN
17/20

BMCP – Cycle cellulaire
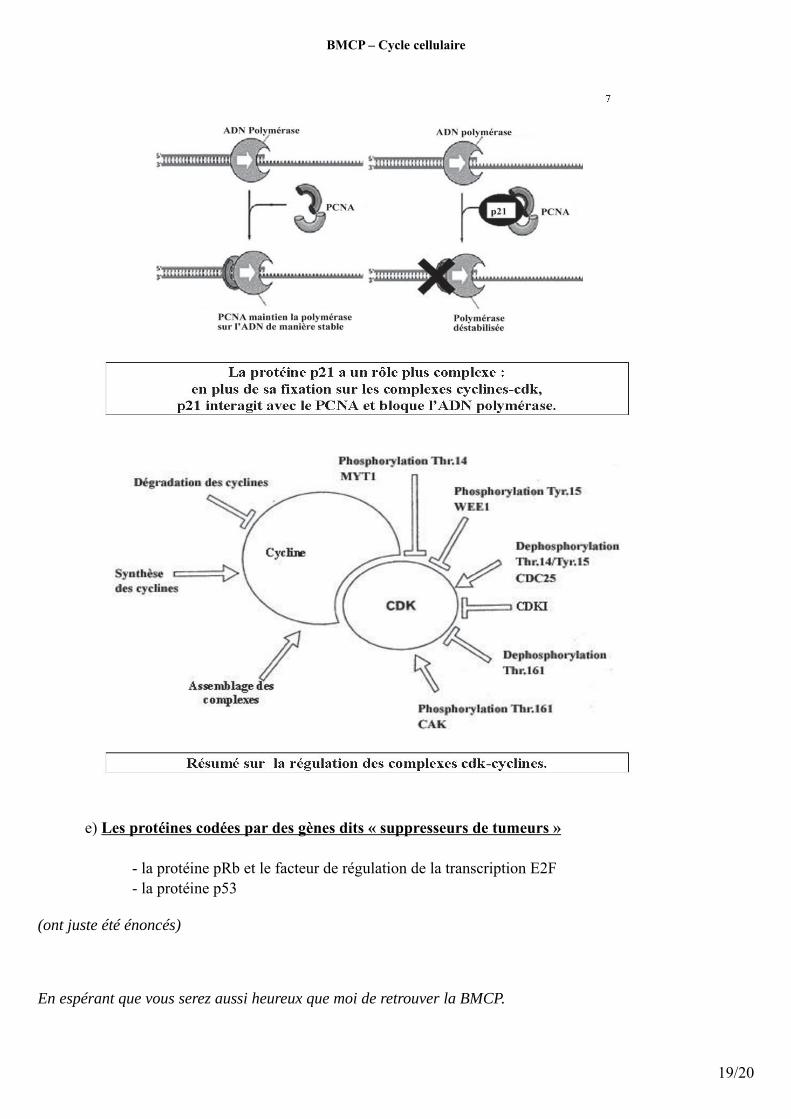
La p21 est un inhibiteur du cycle cellulaire mais a un autre rôle : Elle peut se lier au PCNA (Proliferativ Cell Nuclear Antigen) qui est un élément stabilisateur de la polymérase sur l’ADN, de ce fait la polymérase n'est plus stabilisée car le PCNA ne peut plus se lier sur la polymérase ce qui la rend istable et donc cela bloque la réplication.
18/20
BMCP – Cycle cellulaire
e) Les protéines codées par des gènes dits « suppresseurs de tumeurs »
- la protéine pRb et le facteur de régulation de la transcription E2F - la protéine p53
(ont juste été énoncés)
En espérant que vous serez aussi heureux que moi de retrouver la BMCP.
19/20

BMCP – Cycle cellulaire
20/20





























