Embed Size (px)
Citation preview

de PARIS-SUD ORBAY
labora t oir e de de phytopath ologie
• :'MONTPELLIER ; 1 .... � ,.. ' . ./ . ; . ... '. ' ·� .. ... � " - -·-�\ 111..:�;111.. .. • • ...
.......... � .,., ' .... ,
i ' ' �--! i t 1
·-
' ;
c.
,. ' . . "'
Etude de r agressivité des nouvelles souches de XANTHOMONAS CAMPESTRIS pv. MALVACEARUM (Smith) Oye
virulentes sur les associations de genes majeurs du cotonnier
- les consequences sur la théorie des re lat ions hote-parasite
'
Stéphane CORE E
.. -.....
Beptembr• 1984
'•
\.
, ..
. - . ,
• 1
' .,
1 y
. ) . . .
. ,.
.... .. � .
"\ '

Je t iens à remerc1er tout par t i cu l iêrement Monsi eur FOLLIN mon Maître de S tage , Monsieur GIRARDOT e t Madame ZAPATER pour l eur aide a insi que l ' ensemb l e des se crétaires qui ont permi la r éa l isa t ion de c e documen t .

S 0 M M A I R E
INTRODU CTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
MATERIEL ET METHODES .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
I. - Ma t é r i e l Vég é t a l . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
A) Le s var i é t é s u t i 1 i s é e s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . B) Cul ture d e s p l an t s
I I. Cul ture d e cal s
I I I. X. mal vacearum ( Sm i t h) Dye . . . . . . . . . . . . . . . . . . . . • . . . . . . • . . . . . . . . . .
A ) Le s souche s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
B) Cul tur e s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IV. Inocul a t i on des p l antu l e s
2 2
2 2
3
3 3 3
3 V. Exp l o i ta t ion s t a t i s t i que des donné e s . . . . . . . . . . . . . . . . .. . . . . . . . . . . 4
RES ULTATS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 I. Es s a i prél imina ire : Ino culat ion de R1, R2 , R 1 8 , R1R2 , R 2R 1 8, s ur
l a var i é t é Stonevi l le 20 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 4 II. Es s a i n ° l : Inocul a t ion des nouve l l e s s ouche s sur l a gamme Œima S 4 ,
Mebane BI, 1 0 1 . 1 0 2 B, Gre g g , Ac a l a S J4 ) . . . . . . . . . . . . . . . . . . . . . . .. . .
A ) C l a s s ement des s ouche s . . . . . . . . . . . . . . � . . . . . . . . . . . . . . . . . . . . . . . . . .
B) C l a s sement des var i é t é s
I I I. E s s a i n ° 2 : Ino c u l a t ion d e s s o uches s 3 , T2 , HV 1 , HV2 5 s ur l a même gamme (= e s s ai n ° 1 s imp l if i é ) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
A) C l a s s eme nt des s ouche s . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . .
B) C l a s semen t de s var ié t é s . . . . . . . . . • . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
IV. E s s a i n ° 3 : Ino c u l a t ion d e s nouve l le s s ouche s s ur une gamme de var i ét é s po s s édant toutes l a comb ina i s on de gènes majeur s . . . . . . .
V. Compar a i s on des r é s u l t a t s avec c e ux o b t enus aux E ta t s-Unis . . . . . .
4 4 4
5 5 5
6
6

DIS CUS SION . . . . . . . • . . . . . : . . • . • . . . . . • . . . . . • • . . . . . . • • . . . • • . . . . • . . . .
I. Le s ob serva t i ons de b a s e . . . • • . . • . . . . . . . . . . • . . . . . . . . . . • • • . . . . .
II. Les intera c t ion s d iff é ren t ie l l e s . . . . . . . . . . . . • • . . . . . • . . . . . . • .
III. Adap t a t i on des s ouche s . . . . . . . . . . . . . • • • . • . . . • . . . . . • . . . . . • . . .
IV. L ' e s s a i n ° 3
CONCLUS ION . • . . . . . . • . . • . . • . • • • . . . . . . . . . . . . • • . . . . • • . • • . • . • . . . • • • . .
REFERENCE S BIBLIOGRAPHIQUES • • . . • . • . • . . . • . . . . . • . . • • . . . . • . . • • . • . . .
SOMMAIRE DE S ANNE XE S • • • • • • .• • . • . • • . . . . . • . . . . • . . . . . . . . • . . . • . . • . . . .
7 7
9 1 0 1 1
1 2 1 3
1 6

INTRODUCTION
. La b actér i o s e du cotonni e r causée p a r X. malvace arum ( Sm i th) Dye e s t une des principal e s malad i e s de ce t t e culture , par t i cu l i èreme nt en Af r i que . P rés e nt e s ur t o u t e s l e s p art i e s aér i enn e s de l a p lant e , c e t t e bactér i e p rovoque des symp t ôme s carac téri s t i que s d e nécro s e après une phas e " t ache hui l e u s e " (water soaked l e s i on s ) .
. Les at t aque s p r imaire s déb u t ent à par t i r d e s grain e s ou d e s déche t s végét aux contam inés. La p l uie e t l ' i rrigat i on f acil i t e nt l a d i s s ém ina t i on e t l e s a t t aqu e s s e condair e s ( 1 8) . Le s condi t i o ns opt imal e s d e dév e l oppement s o nt u ne t empérature d e 32- 3 6 ° C jo int e à une f o r t e hygromét ri e et à un e ns o l e i l l eme nt impo r t an t .
. La chut e de rendement e s t t rè s variab l e s e l o n l ' environneme nt e t l e s tade d ' in f e c t i on . E l le a ét é e s t imée à 20 %au Soudan en 1 9 7 0 mai s c e c i r e s t e une app roximat i on , e n rai s on d e s i nt e ract i ons ob s ervée s en tre parc e l l e s s aines e t malad e s ( 1 8) . Le f ai b le développement des re che r ches dans l e domaine d e s t ra i t ements an t ibactériens , l eur coût, l ' ine f f i cac i t é re l at ive d e s méthodes cul t ural e s font q ue l e s e f f o rt s s e son t o r i e nt és ve rs la s éle c tion d e var i ét és rés i s tant e s .
. En e f fe t , l e s pros pecti ons e f fectuée s p ar ��IGHT à part ir de 1 9 34 mon t rent que l e s d i ve r s e s e spèce s de cotonnie r, cul t i vée s ou non , s e comportent de mani ère vari ab l e v i s -à-vi s de l a malad i e . On ob s 2rve :
- La rés i s t ance t o t a l e ( Immuni ty) : aucun s ymn·l6me macros copi que e t réact i on hype r s ens ib l e e n i no cu l a t i on art i f i c i e ll e .
- La rés i s t ance s i mp l e carac tér i s ée par un t r è s f a i bl e déve l oppement d e s l és i on s . Ce t t e rés i s tan ce peut êt re fo r t e e t même donner l i eu à une réact i on HS mai s e l l e n ' e s t p as t o ta le .
- La s en s i b i li t é : dév e l oppeme nt p l us ou mo i ns importan t des tache s ( 4) .
Apr è s hybri d a t i o n suivi e d e p lus ieurs généra t i on s de backro s s , KNIGHT a ob s e rvé l a s égrégat i on de g2n e s "maj eurs " donnant un e rés i s t ance p lus ou moins forte ( an a l y s e mende l li enn e ) àémo ntra•1t p ar l à même 1 'héritab i l i té du caractère "rés i s t ance à l a b ac t é ri o s en ( 3 ) .
. A la s ui t e de ce t te découve r t e , �'H G HT , LAGIERE , B RI:�KE RHOFF , INNES ont m1s en évi dence p l usi eurs gèn e s B de rés i s tance ( cf annexe 2.) .
. Ap rès l e s t ravaux de Vill�DERPL&�K (3 1 - 32 ), ce s gène s majeur s ont été as s imi l és à de s o l i gogènes e t re connus comme l e s c omp o s an t e s de l a rés i s tance ve rt i c a l e ( C HEVAUGEON) .
S ui vant l a même t e rm i no l ogi e , Ja ré s i s t ance hori zont ale e s t déte rminée par l e 'co nt ex t e génét i que " ( ge net i c al background) . BIRD e t H.ADLEY ont a in s i m i s e n évi d ence d e s p o lygène s comp l ex e s appe l és B sm ou Bnm s e l on le génome dont i l s s o nt i s s us ( S tonevi l le ou De l t apine) .
. . 1 ..

2
Ut i l i sé s seu l s , comme ce l a a é t é fai t dans l e p assé (par exemp l e B7 sur S t onevi l le 2 0 ou B2 sur Acala 1 5 1 7 BR2) , l e s gènes B sont rapi d ement surmon t é s , p rovo quant l ' effondrement de l a résistance . L ' hypothèse de la re l a t i on gène pour gène (9 ) pour le coup l e (X. malvace arum , Gossyp i um sp) e st a insi confo rt é e .
. La grande vari ab i l ité du p ouv o i r p athogèn e de c e t t e b a c t é ri e p e rme t c e phénomène . A ce jour 1 8 rac e s o n t é té d é c ri t e s grâce à une gamme d ' hôt e s di fférent i e l le mise au point par HUNTER en 1968 (17) (8 G . hirsutum) e t mod i f i é e par BIRD en 19 80 . En Inde , NAYADU {�2;) é t ab l i t 4 race s se l on la v i ru l ence sur l e s 4 p rinc i p a l e s e spèces d e cotonn i e r . Il dé t e rmine au sei n de chaque race , d e s b i o t yp e s se l on la v i ru l ence sur l es vari é t é s.
Sur m i l i eu art i fi cie l , une p erte d ' agre ssiv i t é a pu être n e t t ement o b se rvé p our c e rt aine s sou ch e s (2- 33) . En revanche l e s vari at i ons de la v i ru l ence son t p lus d i ff i c i l e à me t tre en évidence (7) •
. Pour fai re face aux p rob l èmes que posent l e s int e ra c t i ons entre l ' hôte e t l e pathogène (dépassemen t de l a rési st an c e ) , l e s sé l e c ti onne urs ont ut i l i sé s des comb inai sons de gènes t e l les BzB6 , B 2 B3, B9 L BlOL , Bz B9 K• BzB3B6 , B2B3B7 qui con f è rent une ré sist ance t o t a l e à �· h i rsutum ( B IRD) .
. Ces cotonn i e rs sont ut i l i sé s depu i s 2 0 à 30 an s dans l a p l up art d e s grands p ay s c o t onni e rs , donnant à c e t t e rési st an ce un c arac t ère p art i cu l i è rement stab l e .
. Né anmoins , en 19 80 on a pu o b se rver l ' appari t i on de nouve l l es races v i ru lentes sur t ou t e s ces c omb inaisons en Haute-V o l t a , au Tchad e t au S oudan' ( io) .
. L ' obje t de ce t rava i l est l 'é tude de l ' agre ssivité de ces nouve l le s souches sur d i f fé ren t e s varié t és, ceci débouch an t à un nive au p l us géné ra l sur l 'é tude du type de ré sist anc e qu'offre l e c o t onnier à X. mal vace arum .
. Pour c e c i l a confron t at ion hôt e-p arasi t e a é té é tud i é e sur p l an t u l e (ino cu lat i on d e feu i l l e cotylé dona i re ) e t au n ive au c e l l u l a i re (inocul at i on de cal c e l lu l a i re ) .
MATEJIEL ET METHODE S
1. LE î1ATERIEL VEGETAL
Pima S4 (G. b a rb adense ) Meban e B l ( G. h i rsutum) l 0 l- 1 02 B (" " ) Gre gg (" " ) A c a l a S J4 (" " ) BSO B JA59 2 Jl9 3 MK73
0 Bz + Po lygène s B2B3 l gène in connu 0 B 9LBlO L B2B3 B2B3 B2 B3

3
Le s p l ant s ont été cul t ivés s ur vermi c u l i t e e t al imentés p ar une s o lut i on nut r i t ive ( cf c omp o s i t i on annexe n ° 3 ) .
Les c ond i t i on s de cul t ure s ont l e s s uivante s - 28-2 9°C - 6 5-7 0 %d'hygr omét r i e
1 2 h d e lumi ère p ar jour d'une i ntens ivi t é aporoximat ive d e 2 4 0 0 l ux .
Pendan t l a germination , de fréquen t e s brumi s a t ions ont ét é néces s ai re p our perme t t re un éta l l ement p l us f ac i l e d e s feui l le s c o t y l édonaires .
I I . LA CULTURE DE CAL
A la sui t e d e s t r avaux de RUYACK e t al ( 27 ) c once rnant l ' inoc u l at ion de cal de cotonni e r , il e s t ap p aru intére s s ant de voi r l ' a c t i on d e s nouve l l e s s ouche s s ur 3 vari étés
- Acala 4 4 s an s gène de rés i s t ance - S t onev i l l e 20 ave c le gène B 7 - 1 0 1 - 1 0 2 B p o s s éd an t le c omp lexe B2B 3 .
Les c a l s devai e nt être produ i t s à par t ir de morce aŒde feui l le s o u d e t i g e s d e cotonn i e r . Divers e s méthodes ont été ut i l i s ée s ( 8- 1 6- 1 9- 20-2 1 - 26- 27-29) mai s aucune n ' a p e rmis d ' obt e ni r un mat ér i e l s u f f i s amment ab ondant p our atte indre la phas e vér i tablement i nt ér e s s an t e de l a manipulation : l ' ino c u l a t i on . Aucun p aragrap he ne s e r a d onc consacré à c e t e s s ai dans la p ar t i e r ésu l tat .
III . X. m a l v a c e a r um ( S m i th ) Dy e .
I l s ' ag i t d e nouve l le s s ouch e s ori ginai r e s du Tchad (T), de Hau t e-V o l t a ( HV) et du S o udan ( S ) . Les var i étés dont e l l e s s ont �s sue s s ont rés umée s dans l e t ab l e au s u ivant ( 1 1 12) .,
Souches T l T2 s 1 l s 2 s 3 HV l 1 HV3 HVS 1 HV7
1 l
I s o l ée s d e P ima S4 Re ba Bar ac (Ac 4-4 2 ) e t BJA 59 2 MK73 HC4 75 1 P 279 Gos syp i um barb adeœc
B/ Culture s
Le s souch e s s ont cul t ivée s à l ' obs cur i t é à 2 6°C s ur un mi l i e u PCPA ( P DA modif ié) : c f . la comp o s i t i on annexe n° 5 .
IV . INOCULATION DE P LANTULE
1
Plus i eurs t e chn iques ont été décri t e s ( 1 3- 14- 2 8 ) . C e l l e r e t enue �c� cons i s t e en l ' i no c u l at i o n de p l antule au s t ade coty l éd onaire ( 8 ir s ) à l ' aide d ' une s u s p ensi on dens e de baGtér i e s dan s l ' e au s téri l e ( 1 0 9 - 1o 1 0c/ml ) . Ce t t e s u s pens i on e s t p rép arée à l ' aide de culture bact ér i enne e n phas e de cro i s s ance exp one nt i e l l e ( 48h �Ja:ci ,";um) .
L'i noculat i on se fai t par s cari f i ca t i on de l a face inféri eure de s c o t y l édons à l'ai de d'une p lume à d e s s in t rempée dans l a sus pen s i on .
HV25
?

4
L ' inoculation peut êt re complé té e par une inj e ct ion d an s l e s t is sus foliaire s de s us pens ion bact é rienne à l ' aide d'une s er in gue qui· p e rme t de me t t re mieux en évi dence l a ré action hyp e r s ens ib le (cf. p lanche no 2 annex� ' 1) .
L ' ob s ervation a l ie u 8 à JO jours apr è s l 'inoculat i on e t l a no t a t i on s e fait s e lon une é chelle s imp lifié e compr enant 5 n iveaux
(cf. p lanche n ° 1 annexe 6). V . L ' EXP LOITATION STAT I STIQUE Dn DONNEES
Le s donn é e s r e cu e i llie s par co t �t i on ne p euvent êtr e u t ilis é e s brutes car elle s ne s a t i s font p as a u cri t ère d ' u t ili s a t i on de l ' analy s e de la varian ce . La t r ans formation angulai r e Arcsin -� perme t de s e rapprocher de ce s conditions ( homo généité d e s variance
� �O e t
normalité de la dis t ribution d e s e rreur s ) .
Afin de me t t re en évidence le s int e ract i on s , d e s analy s e s d e variance à 2 voie s o n t é té ré alis é e s . Ce ci néce s s it e u n nombr e de r é p é t i tions con s tan t ( ce la n ' a pu être ob tenu pour l ' e s s ai n ° 3) . Lor s qu ' il y a int e ract i on d e s analy s e s de vari ance à 1 voie s on t fai t e s pour chaque variété (trait ement s ouch e ) et p our chaque souche ( tr aitement varié t é ) .
Ceci p e rm e t d ' évaluer le risque avec le quel l ' hypothè s e nulle d ' égalité de s variance s d e s traitement s peut êt re re j e t é e . Si le r i s que e s t suffi s amment faible , le s t rai t ement s s ont cla s s é s à l ' aide de t e s t d e Ne wman e t Keuls .
I . ES SAI PRELIMINAI RE
RES ULTAT S
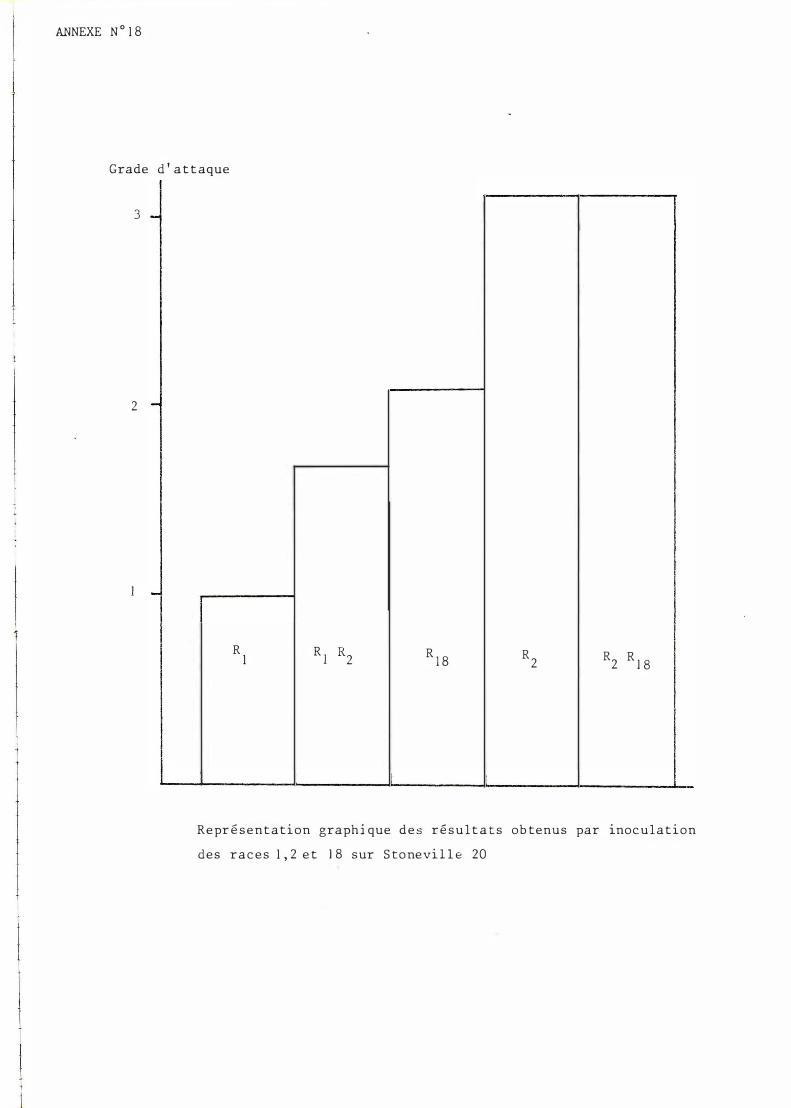
Ino culat i on de RJ, Rz, RJB• RJR2, RzRJ B s ur S t onevi l le 2 0
Ré sulta ts d e s ino cula t i on s : cf . tableau n° l ( annexe 8 ) .
Le s analys e s de variance me t t ent en évidence de s diffé rence s en t re ino culation à un ris que inféri eur à l %o· Le clas s ement ob tenu
e s t s ch émat is é en annexe n° 1 8 .
Comme cela é ta i t prév i s ib le , la race 1 e s t b ien s an s e ffe t face au gène maj eur de r é s i s t ance B 7 . Le s r ace s 2 e t 18 au con t r ai r e cont ourne ce t t e ré s is t ance . L'exp r e s s ion d e s symp t ôme s qu ' elle s engendrent e s t né anmoins n e t t ement iné gale .
En mélange, le s r aces RJRz provoquent d e s symp t ôm e s b e aucoup p lus proche s de ceux obtenus ave c la r ace 1 s eule (nombreu s e s réact i ons hype r
s ens i ble s H .S . ) . En revan che , le mélange RzRJS e s t t o t alement comp arab le à la race 2 seu l2 (les r é s ult a t s ne s ont p as s ignifica t i vement d i ffé rents ) .
I I. E S S AI N° 1 : Ino culat i on des n ouvelle s s ouches sur la g amme P ima S 4 , Meb ane B J , 10 1 - 1 02 B , Gre gg, Acala SJ 4 .
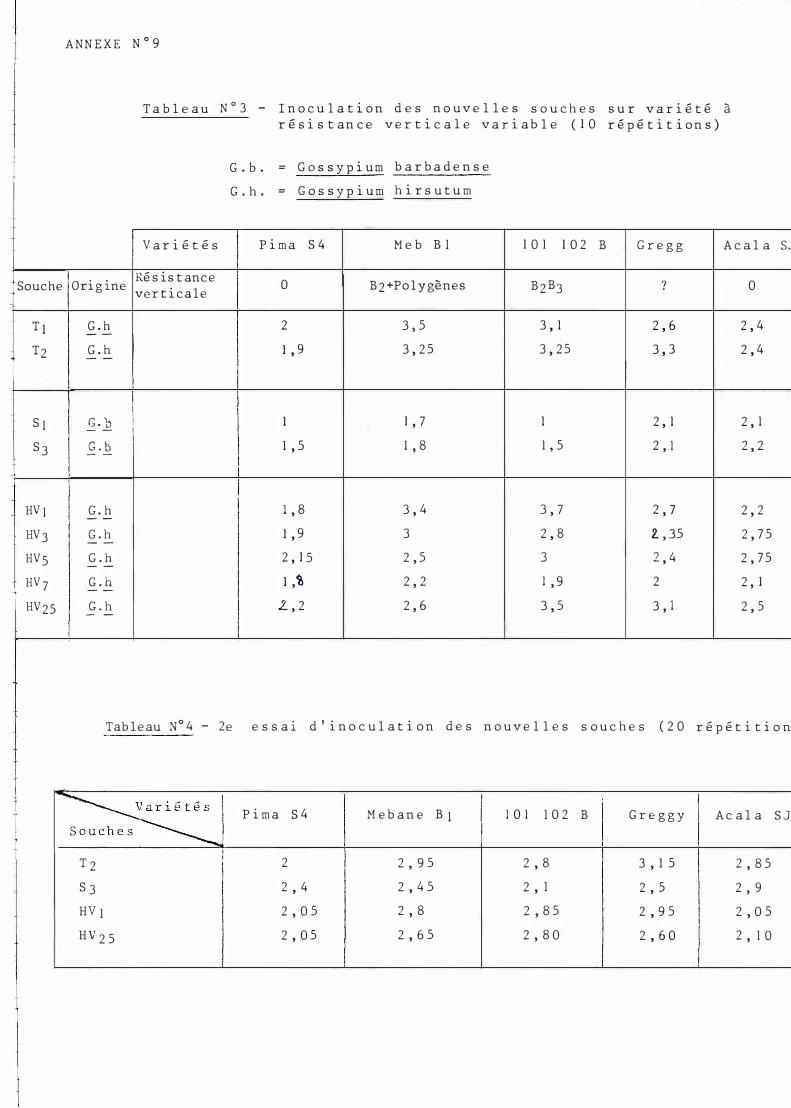
Ré sultats d e s inoculations : cf. t ab le au n ° 3 ( anne xe 9).
P ris e s dans leur ens emb le , le s souch e s d i ffé rencient ne t t ement deux group e s de varié t é s
- P i ma S 4 (�. b arbaden s e ) l ' ens emble d e s autre s varié t é s ( G. hir sutum) .

P ar a l l è l ement , l'ensemb l e des vari é t é s me t en évidence l 'hypoagressivi té de s souches soud anai se s (P ar t i cu l i èrement n e t t e pour 10 1 - 1 02 B ).
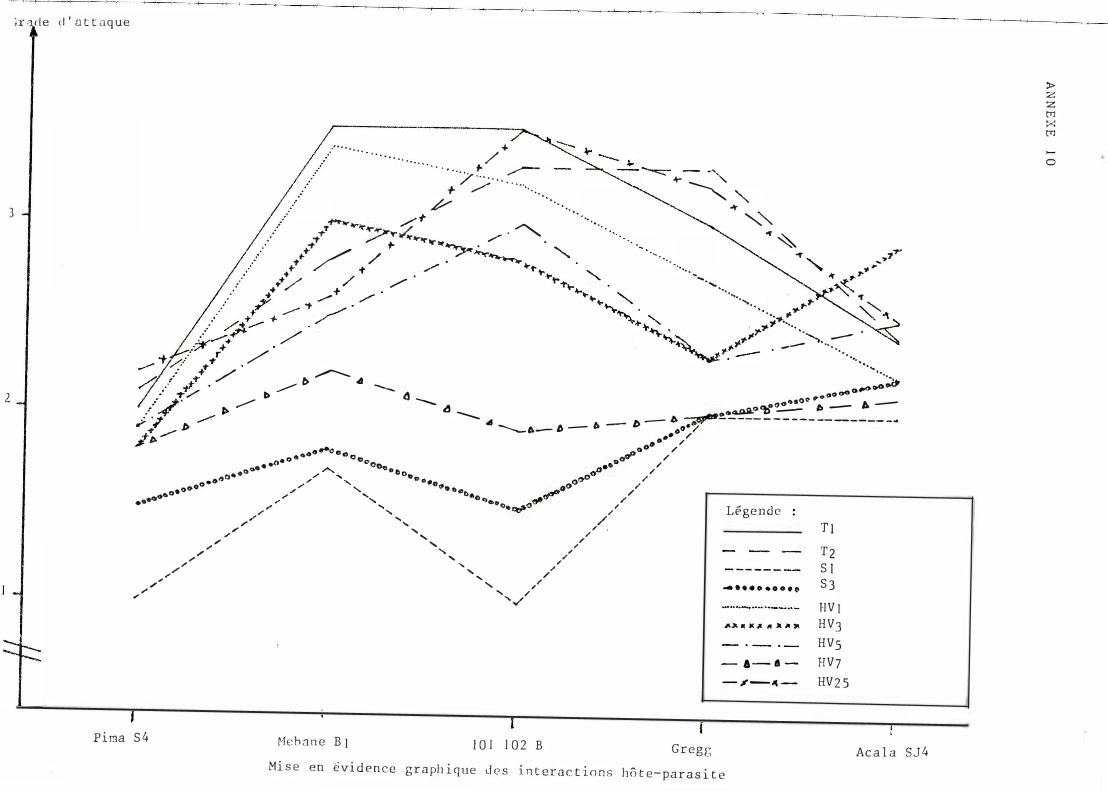
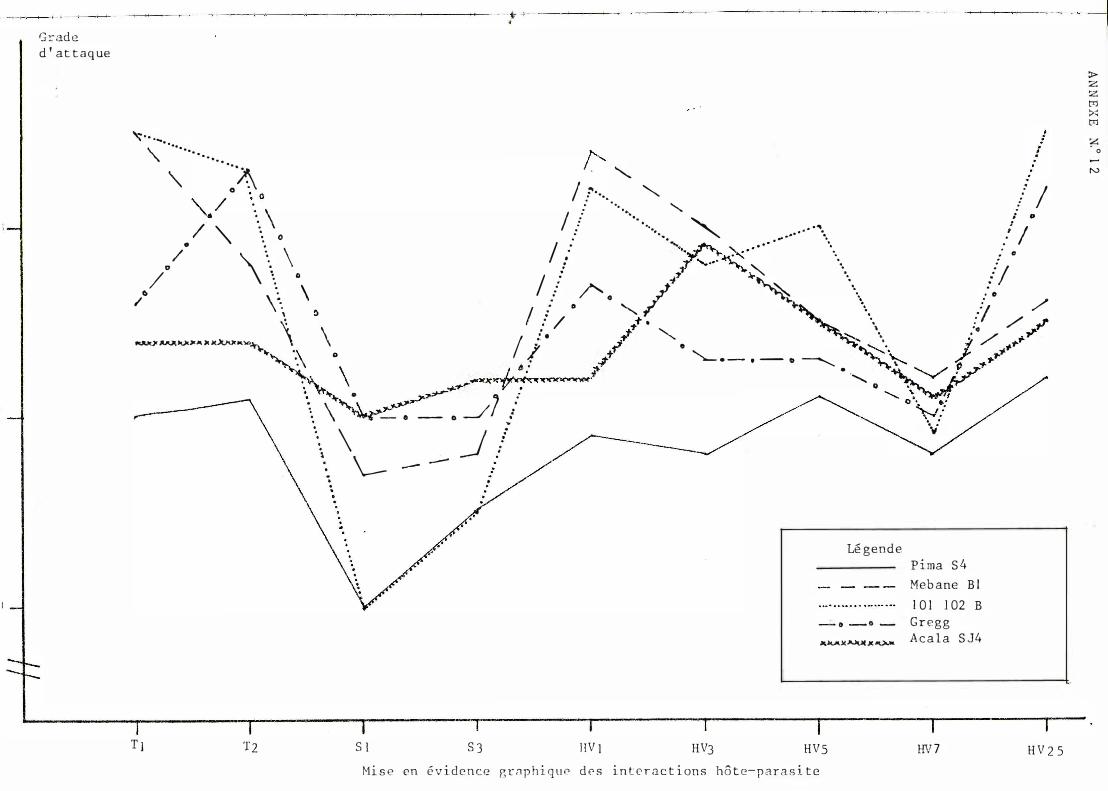
Des d ivergence s de courbe ayant é té déce l é e s à l 'a ide des graph i ques ob t enus à part i r des données brutes (cf . anne xe s n ° l0 et 1 2) , une ana lyse de var i ance à 2 vo i e s (var i é t é s , souche s) a permis de m e t t re en évidence une interact i on i mpo r t ante .
L'int er ac t i on ne perme t t ant pas d'a t t r ibuer la par t de la var i at i on l ié e aux souche s e t ce l le r é su lt ant des var i é t é s , une é tude st a t i st ique à un n iveau plus simple a é t é entrepr i se (é l iminat i on d ' un fact eur de v ar i at i on) .
Toutes l e s ana lyses d e var�ance à l vo�e me t tent en évidence des d i ffé rences sign i f i cat i ve s à un r i sque au plus é g a l à 5 %. Except i on fai t e pour HV7 qui n e pe rme t pas de d i f fé r en c i e r l e s var ié té s. A l a su i t e d e c e s analyse s ( qui r é pondent à l a quest i on : "Y a t- i l d e s d i f fé rences?") des t est s de Newman e t Keu l s ont é té e f f e ctués pour abou t i r au c l assement des souch e s et var i é t é s
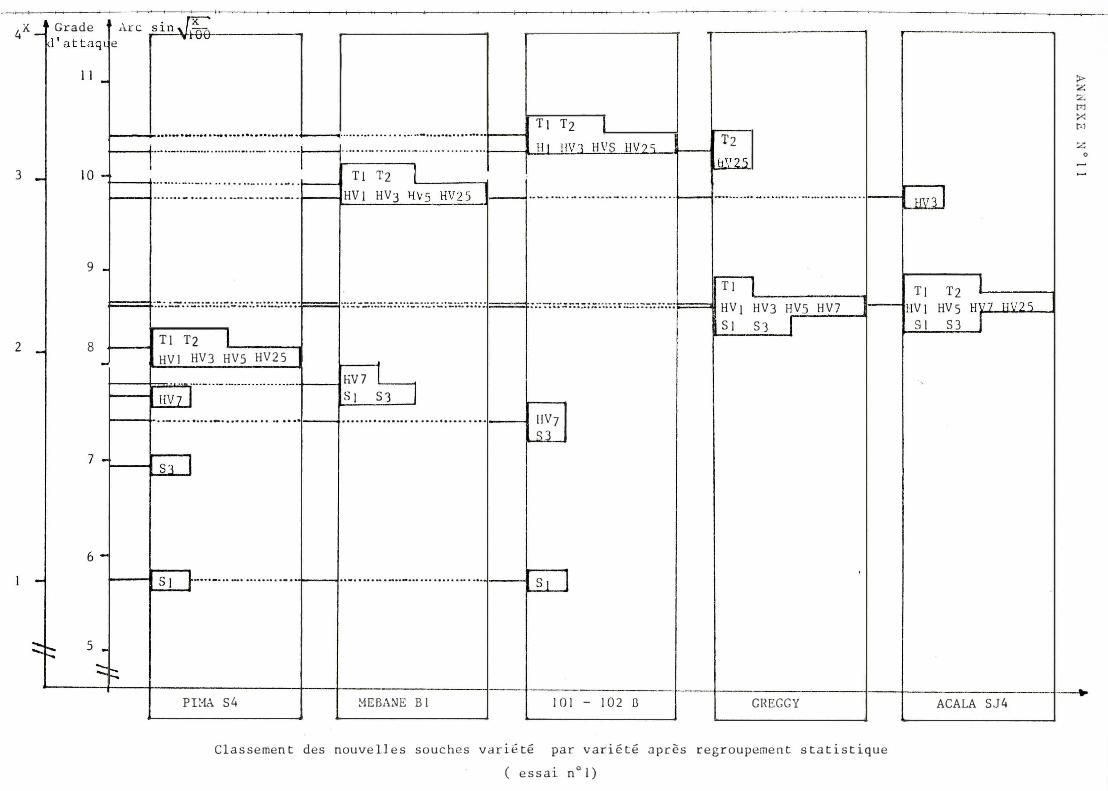
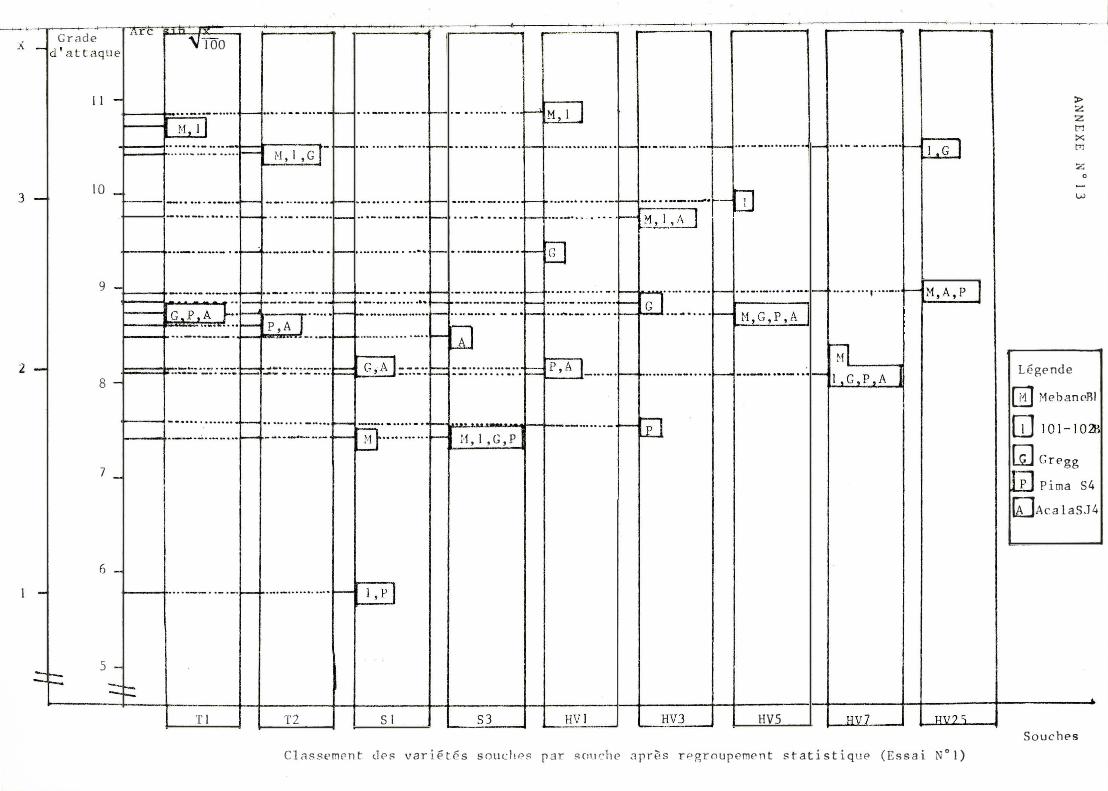
A / Classement des souch e s (cf . annexe n ° ll)
I l con f i rme l a f a i b l e agressivité des souches soudan ai ses. En revan che , l e s souche s T son t toujours parmi l e s plus agressives except i on f a i t e pour l ' Aca l a S J 4 .
L a vari ance d ' agressivité (ampl i tude en tre souches peu e t for t e ment agre ssive ) e st part i cu l i èrement impo r t ante pour l e 1 0 1 - 1 02 B . E l le e st en revanche min imum pour l ' Acala S J 4 pour l e que l , seu l e l a souche HV3 est st at i st i quement di f f é r en t e des aut r e s.
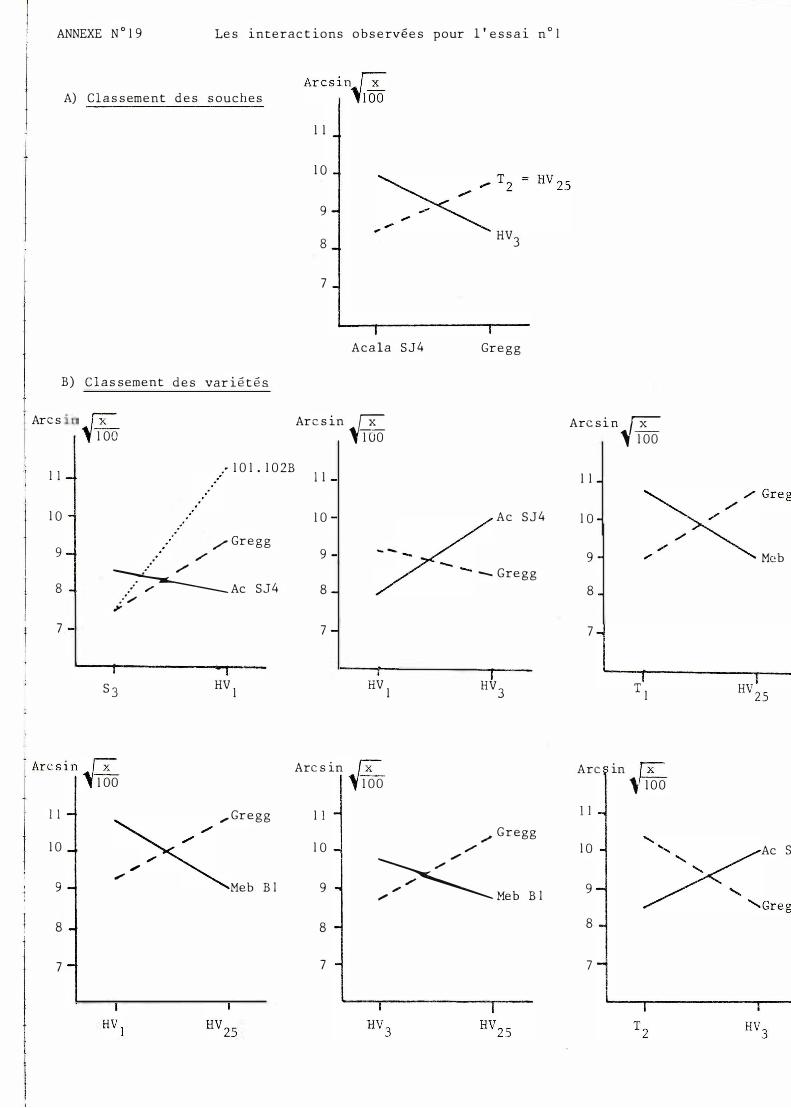
Le cl assement d e s souches n e perm e t pas de m e t t r e en évi dence une interac t i on très n e t t e sinon c e l le r epré sent é e en annexe n ° 19.
G l ob a l emen t , l e c l assement des var i é t é s évo l ue b e aucoup se l on l e s souch e s. Néanmo ins des t endances peuvent êt re obse rvé e s. Ainsi P ima S4 se si tue toujours dans l e groupe de queue e t Acala S J 4 pré sent e une sen si bi l i té re l at i vement const an t e . On n o t e de très nombre use s in terac t i ons. quadrat i ques e t que lques in terac t ions vr aies reprodui t e s en annexe n ° 19.
S i l ' on comb ine l e s c lassement s var i é té e t sou che , l a seu l e i n t e ract i on véri t ab l e e st ce l l e entre l e s souches T2 , HV3 et l e s var i é t é s A c a l a S J 4 e t Gre gg.
Pour chaque souche , l ' ampl i tude entre classe ré si st ante e t sensib le e st re l at ivement const an t e e t faib l e (d e 1 à 3 un i t é s après l a t r ansformat i on Ar cs i n . �).
" loO I I I . E S SAI N'02 : Ino c u l a t i on des sou ches S 3 T? HV1 e t HV25 sur l a même gamme
(= Essai n ° l simp l i fié ) .
Global ement l ' interact i on sou ch e var i é t é est fortement mi se en évidence. Tou t e s l e s analyses de var i an c e à l vo i e (ent r e souch e s e t ent re vari é tés) pe rme t t ent de repousse r l ' hypothèse null e d'é gal i t é de var i ances à un r i squ e toujours infé ri eur à 2,5 %.

6
nette étant
Globalement l'hypovirulence de la souche s 3 n'est pas aussi que dans l'essai précédent. En revanche, Pima S4 est confirmé comme moins sensible que les �· hirsutum.
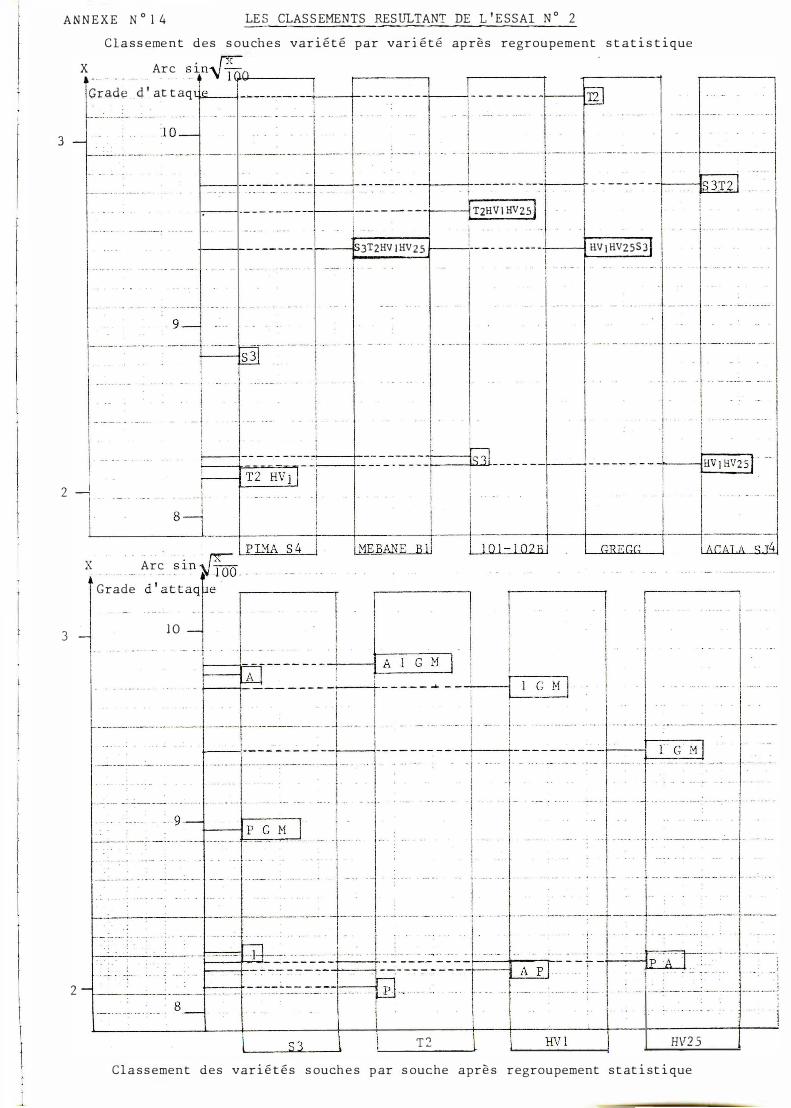
A/ f!������!_9���ch�-��!��-1��-��E��!�� Les résultats observés avec l'essai n°) ne sont que partielle
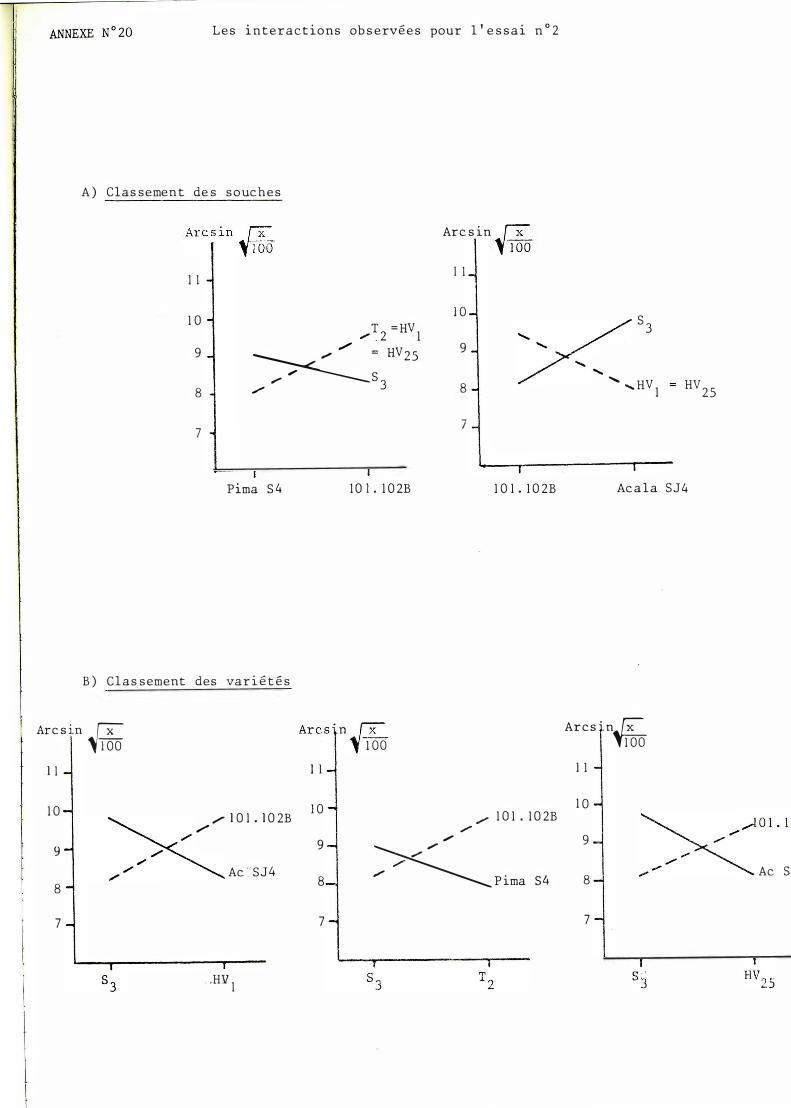
ment retrouvés. Le classement pour 1 01- 1 02 B et Gregg reste inchangé. Celui pour Mebane B I et Acala SJ4 subit des modifications sans présenter d'inversion. En revanche, celui pour Pi�a S� en présente une puisque la souche SJ apparaît plus agressive que les autres. Les interactions observées sont résumées en annexe n° 2 0 .
B/ Classement des variétés selon les souches
On observe pas d'inversion par rapport aux résultats précédents ma�s les groupes ont été refondus. Cela confirme
- la sensibilité particulière de Acala SJ4 vis-à-vis de la souche s 3 - la sensibilité de 1 0 1 -1 02 B pour les autres souches - la bonne tenue de Pima S4 pour toutes les souches.
Les 3 interactions mises en évidence sont résumées en annexe n° 20.
Il est à noter que les différences existant entre les essais 1 et 2 sont délicates à expliquer et font toucher du doigt le problème des artefacts liés aux manipulations en laboratoire. Ainsi, la souche SJ apparaît dans cet essai beaucoup plus agressive que dans l'essai précédent. Ceci a pour conséquence de transformer des interactions quadratiques en interactions vraies.
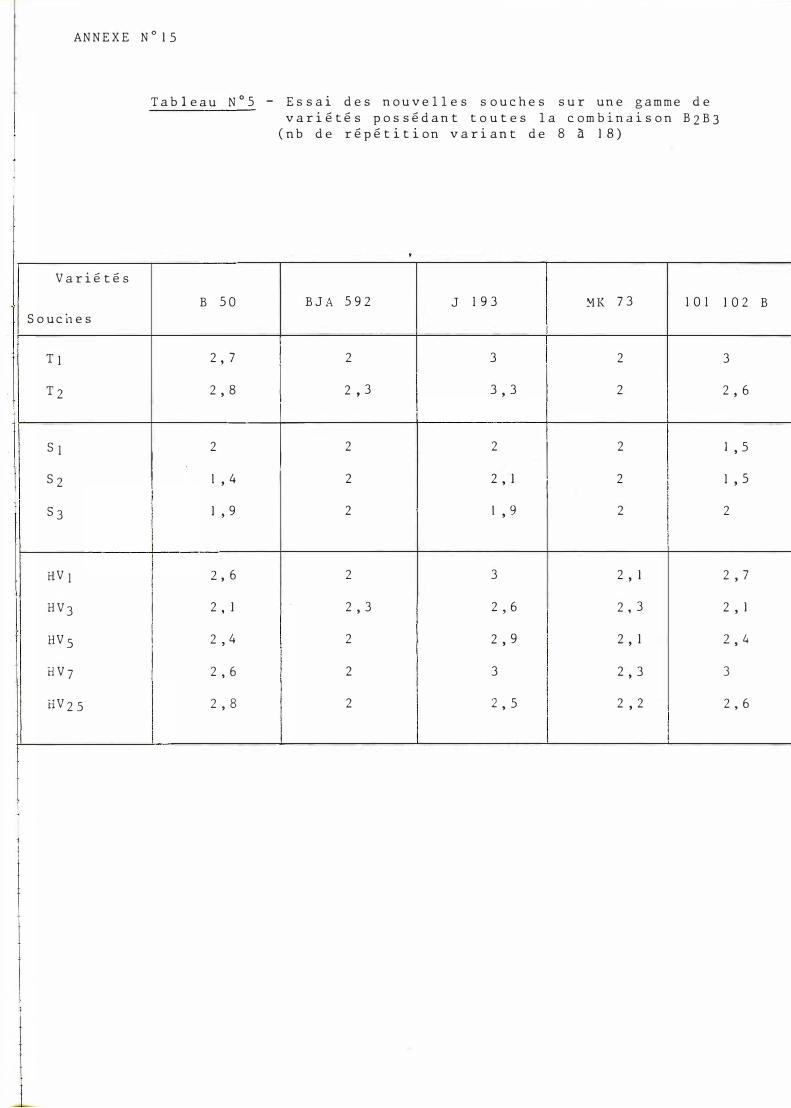
Inoculation des nouvelles souches sur une gamme de variétés possédant tout� la combinaison de gènes B 2B3 (B SO, B J A5 92 , J l 9 3 , MK7 3 et 101- 1 02 B .
Tableau des résultats n°5 (cf. annexe n°lq).
Les résultats n'ont pas été traités de manière statistique étant donné le nombre variable des répétitions.
On note l'hypovirulence généralisée des souches soudanaises et la sensibilité très nette du Jl9 3. La bonne résistance de MK73 et de BJA5 9 2 à l'ensemble des souches est très marquée.
L'observation du graphe (annexe n°16) semble montrer l'absence d'interaction vraie (Parallélisme des courbes).
V. CO�ŒP4�ISON DES RESULTATS AVEC CEUX OBTENUS AUX ETATS UNIS
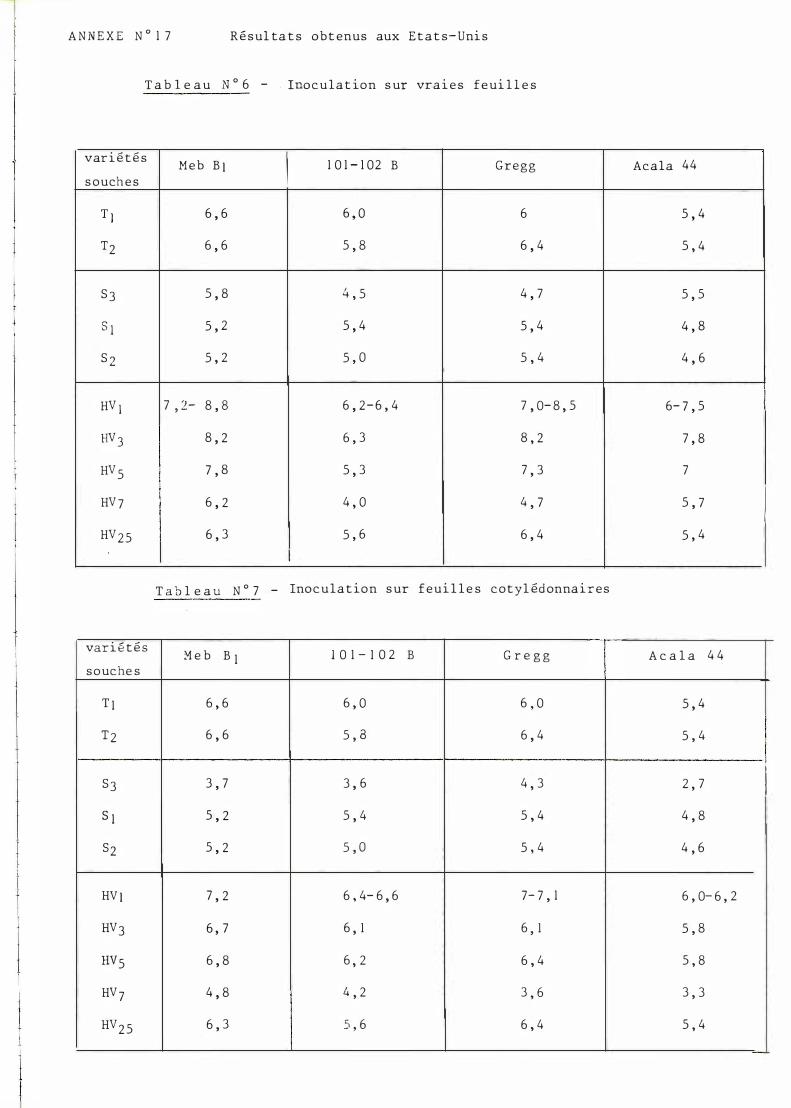
Les nouvelles souches fournies par FOLLIN ont été étudiées par l'équipe de B IRD à College Station (TE XA S ) . Des inoculations ont été faites sur feuilles cotylédonaires et sur feuilles vraies pour lesquelles le niveau d'agressivité semble supérieur (1). Les résultats peuvent être observés dans les tableaux n°6 et 7 cf. annexe no 17.
. . 1 ..

M algré l ' é che l le de n o tation diffé ren te à 10 n�ve aux ( 1 =
ré s is tance t o t ale e t 10 s en s ibil it é m aximum) , le s ré s ult a t s s ont en grande partie re t rouv é s à Montpe l l ie r .
1 ° / Les s ouches s o udanais e s s ont bien caractéris é es p ar leur faible agr e s s ivité.
2 ° / P armi le s s o uche s is s u e s de Hau t e-V o l t a, HV 7 e s t d ' une agre s s ivité n e t t emen t infé rieure.
7
3° / L ' ens emble de ce s nouvelles s ouch e s es t d ' une agr e s s ivité moindre s ur l a varié t é tot alement démunie de gène de rés is t an ce ( Acala SJ4 o u Aca l a 44).
Néanmoin s , cer t ains points en dé s accord peuven t être re l ev é s :
- l' agre s s ivité p ar ticulièrement for t e de l a s ouche HV 1 n'e s t que partiell ement re t rouvé e ( p o ur les varié t é s Me b.B J e t 101-102 B) .
- Au s e in d e s s ouches s oudanais e s , l a d ifférence entre le s s ou ch e s s 1 e t S 3 n'e s t p as t ouj our s re mis e e n évidence . Les s ouches o rigin air e s du Tchad appar ais sent p lus agre s s ive s que dans l es not ations amé r icain e s .
D I S CUSS ION
I. OBSERVATION DE BASE
L ' inocu l at ion du S t onevil le 20 ave c les r aces R1, R2 , R 1 8 et l e s mé l anges RJR2 e t R2R 1 8 aboutit à une doubl e cons tat at ion :
!0/ Le S t onevil le 20 p o s s ède le gène B7 e fficace con t re la race et un comp l exe po l yg énique mal d é f ini. La race 1 8 en cont ournant la résis tance confé rée par B7 provoque d e s s ymp t ômes d ' un niveau moy en. S i 1 'on s ' en t ient aux d é finit ions d u p o uvoir p athogène (9.� 3 2 ) , ce niveau n e devrait p as êt r e dépas s é . Or, avec la r ace 2 le niveau d ' agre s s ivité s e s itue bien p lus haut . Ceci imp lique que la r é s is tance l ié e au comp lexe p o lygénique ait é té dép as s é e e t donc qu ' il y a une certaine adap t ation de l ' agent p athogène au "contexte génét ique g éné ral" de l 'hôt e. Ces o bs e rvations s on t à rapprocher du problème de la r é s is tance du riz à Pyricu l aria Oryz ae grâce au gène Pi K ( 30) .
2 ° / Les obs ervations relative s aux s ouch e s inoculée s en mél anges confirment celle s de F OLLIN et de VERMA e t al. Elles p euvent se r é s umer cormne suit :
Race A (p art ie l l ement + Race B ( agres s ive ) ----1 virulence ident ique à agre s s ive ) la race B (p arfois légère
s ynergie )
Race A (avirulent e : Ré action HS )
+ Race B ( agres s iv e ) ----�Avirul ence et r é action HS il y a prote ction crois é e

1 8
Ceci met directement en cause certaines méthodes de sélections et en particulier celles utilisées pour l'obtention de lignée Multi Adversity Resistant (MAR), le tri des plants se faisant par inoculation au laboratoire de mélange des races : US!, US2, US7 et USI8 ( 1). Il est peu probable que les symptômes observés représentent la moyenne des symptômes issus de l'inoculation de chacune des souches prises séparément (23).
En inoculation au champ le problème des mélanges s.e pose de man�ere moins aiguë puisque l'on peut penser que chaque point infection est indépendant des autres et résulte de la pénétration d'une seule bactérie. Cependant, le niveau de résistance générale exprimé par la plante est fonction de la quantité d'inoculum reçu. Un même mélange n'apportera pas la même quantité d'inoculum actif vis-à-vis de deux hôtes présentant des gênes majeurs différents (23).
L'essai réalisé, par FOLLIN, dont les résultats sont résumés dans le tableau n° 2, met en évidence un phénomène d'interaction entre.deux variétés de coton sensibles sans gènes majeurs de résistance (G. barbadense, G. hirsutum) et deux souches bactériennes. L'hypothèse explicative d'une adaptation des souches à l'espèce est posée.
A partir de ces deux résultats, il a semblé intéressant de vo�r de quelle façon se comportaie�t les nouvelles souches virulentes sur tous les gènes majeurs connus, vis-à-vis d'une gamme de variétés.
Comme cela a été rappelé en introduction, le cotonnier peut réagir de trois façons à la bactériose (4) : résistance totale, résistance partielle, sensibilité. De plus, il apparaît que la résistance partielle peut ou non s'accompagner d'une légère réaction hypersensible (H.S), ce qui fait au total quatre possibilités. Il conviendrait donc de distinguer les résistances avec ou sans hypersensibilité car il semble que l'on ait deux mécanismes différents et indépendants comme le montrent les inoculations en mélange.
Ceci va dans le même sens que les observations de VANDERPLANK (31) qui considère résistance et hypersensibilité de manière distincte puisque "gouvernées séparément tant des points de vue biochimique que génétique".
Ces différentes observations poussent à reconsidérer le problème de la notation des symptômes et leur interprétation. Si les notes 3 , 4 et 5 ne posent pas de problème puisque le niveau d'infection est suffisamment grand pour être correctement apprécié, il convient de s'attarder un peu plus sur les notes l et 2. Théoriquement, la note 1 correspond à l'HS et 2 à une sensibilité trop faible pour que le front d'attaque soit distinct de la blessure d'inoculation.
Dans les essais qui nous intéressent, les souches utilisées sont virulentes sur tous les gènes de résistance connus et donc a priori la note de devait pas être attribuée. En fait, bien qu'elle n'ait jamais été donnée à l'ensemble des répétitions, il est apparu dans plusieurs cas (souches soudannaises) des formes de résistance très proch�de l'hypersensibilité.

9
Le parti qui a été pr1.s est celui de choisir la classe 2, comme une classe "fourre-tout" pour tous les cas ne relevant pas typiquement des notes 1 et 3.
Cette part de subjectivité explique en partie les différences de notation sur des inoculations échelonnées dans le temps. La no. te, 2. peut alors recouvrir aussi bien une r éaction partielle avec hypersenslblllte
ou une forte résistance générale.
II. INTERACTIONS DIFFERENTIELLES (Essaisl et 2)
Les graphiques obtenus (cf annexes 13 et 11) présentent de nombreuses interactions quadratiques (non caractéristiques selon VANDERPLANK (31 et quelques interactions vraies. Les nouvelles souches étant virulentes sur tous les gènes B et sur leur combinaison, les phénomènes de plus ou moins grande sensibilité observés ici ne sont du qu'à "l'arrière plan génétique" des cotonniers, autrement dit à la résistance de type polygénique
Selon VANDERPLANK (32), le type de résistance est caractérisé par l'absence d'interaction différentielle, à savoir qu'il ne peut y avoir variation du pathogène à la suite d'un changement d'hôte (toutes conditions étant égales par ailleurs).
Les principales interactions mises en évidence à l'aide des essais et 2 sont celles que l'on retrouve aussi bien dans le classement des variétés que dans le classement des souches. A savoir :
T2, HV3 et Acala SJ4, Gregg 0 essa1. n
T2, S3 et Pi ma S4, 1 01 102B 0 2 essa1. n
S3, HV1 et Acala SJ4, J 01 l02B 0 2 essa1. n
Dans l'ensemble, ces interactions ne concernent pas toutes les souches, seules : T2, S3, HVl, HV3 et HV25 interviennent.
Les interactions différentielles observées à ce niveau ne peuvent s'expliquer par une action globale polygène-polygène car ainsi, un même hôte ne pourrait agir que d'une seule manière sur l'ensemble des souches pathogènes. Il faut de ce fait pour comprendre, subdiviser l'action des polygènes de l'hôte en une succession d'effets olgogéniques (24).
Ceci peut se représenter schématiquement comme suit
A + B + C + D + E � �ésistance Horizontale
a + b + c + d + e � Agressivité
MODELE ADDITIF
devient
Résistance Horizontale
tA � Ê' D c, D, , •
• t t t t �· b, c, d, ;., y
Agressivité
MODELE �ENE POu�. GENE

10
En subdivisant l a résist ance horizon t a l e en une somme de r ésist ance ve rtica l e , on peut exp l iquer cer tain es r e l ations de type gène p our gène obse rvées à ce niveau puisque , selon VANDERPLANK et FLOR, la mise en évidence de tel l e s rel a tions nécessit e un iquement 2 gènes de résistance e t d eux gènes d'agressivi t é évoluan t de man i ère ind épe nd ant e.
PARLEVLIET et ZADOK dressen t la l iste des conséq uences q u ' ent raîne l a reconnaissance du mo d è l e gène pour gène.
- toutes l es mut ations n ' app ar aissen t pas, ce qui p eut en p ar tie expl iquer l ' aspect durab l e de l a r ésist ance conférée p ar l es p o lygènes
- l a var iance du n iveau de r ésist ance e st moindre
- l e n 1veau g l ob a l de mal adie e st moindre
- l e niveau de sévérité n ' est p as seul emen t fonct ion du n omb re d ' a l l è les positifs d ' a gressi vit é mais aussi de l eur l ocal isation dans l es différents l occi
Enfi n , si cet t e hypoth èse est ret enue, il n ' exist e p l us de différence fondament a l e entre les deux t ypes de résist ance , tous deux ét an t fondés sur l es p r in cip es de l a relation gène pour g ène. C l iffo rd en 1975 (24) reconnaît l 'in t érêt p ratique de subdiviser l a r ésistance en deux classes ma1s ajo ute "Na t ure , 1 am sure, never intended this division".
Ainsi, ce modè le a l ' avan t age de concilie r l es observa t ions scien t ifiques pour d ive r s coup l es hôte- pathogène (dont Gossyprum sp. -
Xan thomonas malvacearum) et un cert ain pragmatisme .
Ill. ADAPTATION DES SOUCHES
S i l ' on admet l ' hypothèse précédente de re l at ions gèn e pour gène au n1veau de l a résist ance géné r a l e, on conçoit imp l icit ement une cer t aine spécial isat ion des souches p uisque l e phénomène perme t t an t le cl assement en race existe égal ement à un deuxième niveau .
Cet t e adap t at ion peut se faire :
- v1s à v1s d ' un cont exte génétique généra l - v1s à v1s d e l ' espèce
Le cho ix de l a gamme d ' hôte pour l es essais 1 e t 2 a ét é fait en fonct ion de ce t te hypothèse déjà avancée à l a suit e des résul t a t s de FOLLIN ( évoqués p récédemment) .
Ainsi , Meb ane B l (B2 + polygènes ) , l 01-102B (B2BJ), Gregg (l gène inconnu) et Aca l a SJ4 (aucun gène majeur) , tous Gossyp i um h i r sut�� ét aien t destinés à vérifier la 1ère hyp o thèse. P ima SJ4 (aucun gène majeur), Gossyp ium barbadense , perme t tait de voir une éventue l l e adap t a t ion à l ' espèce .
Bien en t e ndu , il convient ici d ' être t r è s p r uden t et de n e pas faire d e fina lisme à p ar tir de. quelques obse rvat ions.

JI
Nous nous contenterons donc de noter que les interactions dans l'essai n° 1 restent intra variétales (entre G. hirsut��). On note ainsi une certaine 'préférence" de HV! et T2 pour Gregg par rapport à Acala SJ4, ceci étant à inverser pour les souches S3 et HV3. ReEer ceci à l'origine des souches (cf page 3 ) est très aléatoire car celle-ci n'est pas forcé�ent unique et les filiations entre les variétés utilisées sur le terrain et celles de la gamme sont parfois compliquées.
Dans l'essai n° 2 (essai 1 simplifié) il a été m1s en évidence une interaction intervariétale. Ainsi Pima est plus sensible à S3 que 101 l02B alors que c'est l'inverse pour les souches T2 et HV25· Il est bien entendu tentant de relier ceci au fait que les souches soudannaises ont été isolées de G. barbadense puisque cela va dans le sens de l'hypothèse d'une adaptation du pathogène à son hôte.
L'interaction entre S3, HVI et 10l. I02B, Ac SJ4 est retrouvée dans l'essai 2 de manière amplifiée. En effet, l'ensemble des résultats (interactions vraies et quadratiques) met en évidence une nette "préférence " des souches soudannaises pour AcSJ4 (en terme d'interaction quadratique ceci doit plutôt se traduire en une nette inadaptation des autres souches pour cette variété).
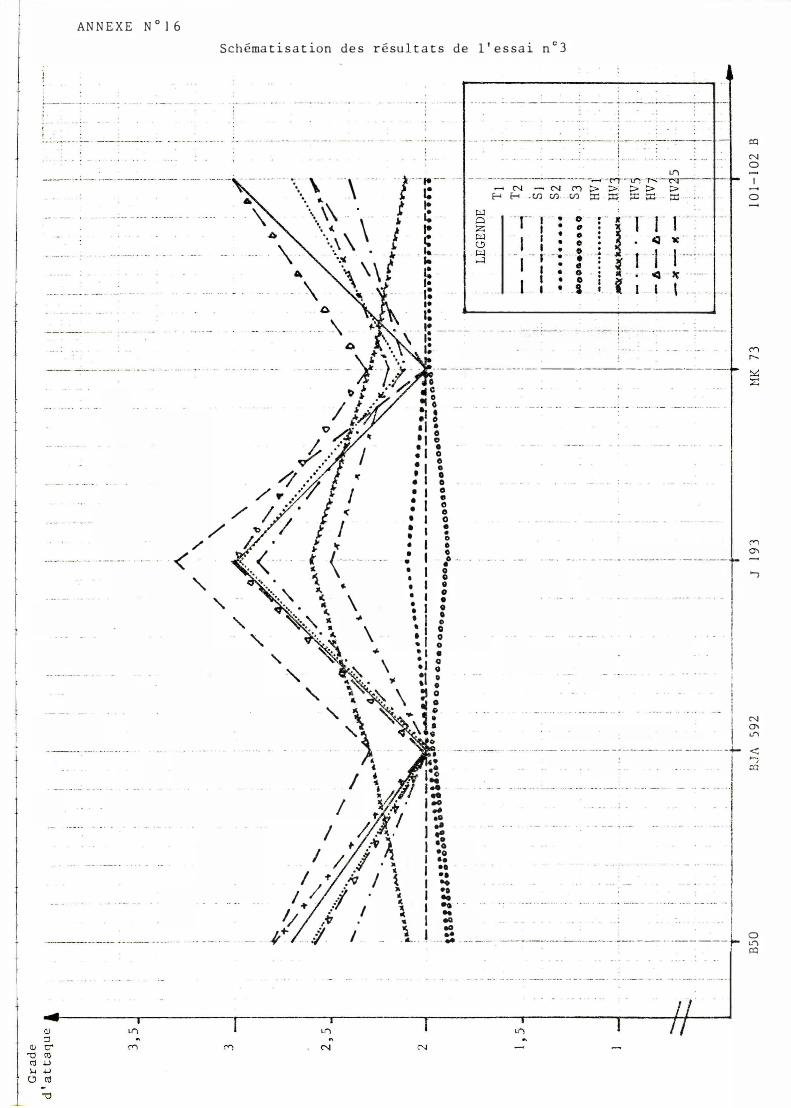
IV- E SSAI No 3
Cet essai a été réalisé afin d'observer l'action des nouvelles souches sur les variétés possédant des gènes majeurs de résistance équivalents (B 2B3 ou B9L BIOL).
Les résultats obtenus correspondent bien à ce qui est observé au champ : c'est à dire principalement une bonne résistance de B JA592 et une sensibilité nette de Jl93.
Pour Jl93 et 101. 102 B on observe une sensibilité plus marquée vis à vis des nouvelles races que des variétés totalement démunies de gènes majeurs de résistance. Ceci peut s'expliquer par la sélection de ces variétés pour une résistance totale aux races existant à ce moment et donc à un effondrement de la résistance de type polygénique que pouvait présenter le noyau de départ (cet aspect de la résistance n'ayant fait l'objet d'aucune sélection).
En dépit de l'aspect parallèle des diverses courbes (cf annexe 16) on peut observer des cas d'interaction quadratique entre les souches soudannaises et l'ensemble des autres. Des cas d'interactions vraies sont observables entre la souche HV3 et les autres souches volta1ques et Tchadiennes pour les variétés BSO, l01 . 102B d'une part et B JA 592, MKï3, d'autre part.
Le faible nombre des interactions reste néanmoins inexpliqué et l'hypothèse d'artefacts de laboratoire n'est pas à exclure.

J2
CONCLUSIONS
Les principales observations à l'etenir des essais entrepris sont
- les indexages erronés qu1.- peuvent résuLter d'inoculat-ion de mélange de souches (14� 23)�
- l'apparente adaptation des souches à la résistance polygénique qui n'est pas sans rappeler les observations de KIYOSAY.4 en 1982 sur le couple pyl'icularia oryzae - riz (30)�
- les difficultés de notation des symptômes liées à la complexité du mode d'expression de la résistance dans le cas de la bactériose du cotonnier�
l'existence d'interaction spécifique sans que les gènes majeurs ne puissent directement être mis en cause� céci étayant l'hypothèse de la généralisation� des relations gène pour gène à tous les n-iveaux de::; la résistance.
Les nouvelles souches étudiées posent le problème de la s�ratégie de sél-ection à mettre en oeuvre dans l'avenir. Il convient ici de constater le faible niveau d'agressivité de ces souches vis à vis des Val'iétés sensibles aux anciennes Paces. Ceci expUque que les gènes B2B3 e-c B9I)310r constituent des coffhinaisons foPtes (FOLLIN)� c'est à dire qu'en leur
D
absence les populations de nouvelles souches régrer-;sent. Ceci sembZ.e nouveau puisque pris un à un� le� gènes majeurs sont tous faibles (exception faite de B? puisque� selon BRINJŒRHOFF et BIRD� la race 2 disparait en son absence).
Eliminer les combinaisons de types B2B3 reviendrait donc à favorisel' le développement des anciennes races et particulièrem2nr. de la l'ace 18 très agressive vis à vis de toutes les autres variétés. I: faut donc cont·inueP à sé lectionnel� des variétés possédant les combinaisonD de gènes majeurs à l'aide de la race 18 inoculée seule puis de sélectionne!:' pour• une meilleure résistance aux nouvelles races en rechePchant Z 'accumulation de gènes mineur•s de résistance. La technique d'inoculation en rrxHange peut ici être envisagée dans la mesure où elle s'applique à des souches d'agressivité proche. En revanche� les tests devPont bien correspondre œ�x conditions d'infections naturelles (préférer les pulvérisations au chanrç).
Cette deuxième pr.ase de la sélection risque d'être longue et déZ1:cate puisque des rr'éthodes simples telles que le back-cPoss ne peuvent plus être envisagées.
Enfin� ayant soulevé le protJL-eme de l'adaptation des souches au contexte génétique général� un point d'interrogation persiste quant au pPoblème de la pérennité de cette nouvelle résistance.
Il faut également rappeler les essais infructueux entrepris pour évaluer l'action de ces nouvelles souches sur cals de cotonnier. La pour.suite de ces essais pourrm:t permettre de savoir .si seule une réaction q.A.al1:tative est observable apl'ès inoculation de cal ou s-i un nivequ moyen d'agPessivüé peut êtr•e mis en év-idence. Dans ce dernier cas� il serait particulièrement intéressant de consta�er si Z.es 1oésultats obtenus sur plantules sont confirmés.

l3
REFE RENCES B IBLIOGRAPHIQUES
1 . BIRD, THAXTON, PERCY, EL-ZIK, HOWELL, POSWAL 1 9 8 4 . Res i s t anc e t o new races of the bac t er ial b l ight pa tho gen and i t s impl i ca t ions w i th in the mul t i-adver sity res i s tance genet i c improvemen t s y s t em fo r c o t to n. A paraître.
2. B RINKERHOFF 1 9 6 3. Var iab i l i ty of Xanthomonas malvacearum : the c o t ton b a c ter ia l b l i ght pa tho gen. Techn i c a l B u l let in o f the Okl ahoma S ta t e Un iver s i ty T 9 8 : 9 5 p.
3. BRINKERHOFF 1 9 70. The var i a t i on �n Xanthomonas malvacearum an t i t s rel a t ion t o contro l . Annua l Rev i ew o f Phytopa t ho l o gy 8 : 85- 1 0 9 .
4. BRINKERHOFF, VERHALEN, JOHNSON , ESSENBERG, RICHARD SON 1 9 84. Devel o pment o f immuni ty t o bac t er ia l b l ight o f c o t t on and i t s impl i c a t ions fo r o ther s d i s eas es . P l an t D i s ea s es 6 8, 2 : 1 68- 1 7 3.
S. CHOWDHURY, VE�� 1 9 80. Mul t i pl i cat i on o f X. malvacearum and a phy l l o pl ane bac t er i um in l eaves of G . h i r s ut i um. lnd ian Phytopatho l o gy 33, 2 : 2 4 5- 2 4 8 .
6 . CASON, R ICHARD SON, E S S ENBERG, B R INKERHOFF, JOHNSON, VENERE 1 9 7 8 . Ul tras truc tural cel l wal l a l t er a t ions in immune c o t ton l eaves ino c u l ated \"i th X . ma lvacear um. Phy t o patho l o gy 6 8 : 1 0 1 5 - 1 0 2 1 .
7 . CROSS 1 9 6 3 . Pathogen ic i ty d i fferenc es i n Tanganyika popul a t i on s o f X . mal vacearurn . Empire Co t ton Gr ower Review 40 : 125 - 1 30 .
8 . DAVIDON IS, HAMILTON 1 9 8 3. P l ant regenerat ion fr om cel l u s t i s s us of Go s sypium h i r s utum L . . P l an t S c ience Let ters 32 : 89- 9 3 .
9 . FLOR 1 9 7 1 . Current s ta tus of the gene-for- gene concept . Annua l Review of Phy t o pa tho l o gy 9 : 2 7 5-2 9 6.
1 0. FOLLIN 1 9 8 2 . Mi s e en évi dence de rac es de Xan thomonas malvacearum virulentes s ur l 'as s o c i a t ion d e gènes B 2B 3 chez G . h i r s utum . Co ton et Fibres Tro picales 36 .
1 1 . FOLLIN 1 9 82. Repo r t o f the bac ter ial b l ight c ommi t ee . Proceed ing of Beltwide C o t ton Res ear ch Conference. San An tonio .
1 2. FOLLIN 1 9 82. Rappor t s ur l ' analyse des s o uches de Xanthomonas ma l vacearum i s o l é s de c o tonnier s a t tein t s de bac t ér i o s es dans l es es s a i s var i é taux de Haute-Vo l ta. Rappor t internes IRCT.

1 3 . FOLL IN 1 9 83 . Rac e s de Xanthomonas campe s t r i s pv . ma l vacearum ( Smi th) Dye en Afri que de l ' Oues t e t en Af r i q ue Centra l e . Co ton e t F ibres Trop i ca l e s 3 8 , 3 : 2 7 4- 2 7 9 .
14
1 4 . FOLLIN 1 9 84 . La s é l e c t ion du c o tonnier ( G . h i rsutum L . ) pour la rés i s tanc e aux malad ies présentes en Afrique du S ud du S ahara ( à par aî t re dans Co ton e t Fibres Tro p i c a le s ) .
1 5 . HAYWARD , WATERS TON 1 9 6 4 . Xan thomonas malvacearum . CMI De s cr i pt ion of pathogenic Fun g i and B a c t e r i a 1 2 .
1 6 . HEINSTE IN , EL SHAGI 1 9 8 1 . Forma t ion o f Gos s yp o l by Go s sy p i um h ir s utum L . ce l ls s us pens ion cul ture s . Journa l o f Natural Produc t s 4 4 , 1 : 1 -6 .
1 7 . HUNTER, BRINKERHOFF , B IRD 1 9 6 8 . The deve l oppement o f a s e t o f upland c o t ton l ines for d i f ferent i a t ing races of Xanthomonas malvacearum . Phyt o pa tho l o gy 5 8 : 830- 8 3 2 .
1 8 . INNES 1 9 83 . Bac t e r i a l b l i gh t of c o t ton . B i o l o g i c a l Review 5 8 1 5 7- 1 7 6 .
1 9 . KATTERMAN , W ILLIAMS , CLAY 1 9 7 7 . The i n f l uence o f a s t r ong s educ ing agent upon t he ini t ia t io n of c a l lus from the germina t ing s e e d l ings of Gos s ypiw� barbadens e . Phy s i o l o gical P l antar um . 40 : 9 8- 1 00 .
20 . . MITCHELL , JOHNS ON , WHITTLE 1 9 80 . Ga l a c to s i dase ac t 1v 1 ty in cul tured c o t ton c e l l s ( Go s sypium h ir s utum L . ) a compari son b e tween c a l l s grow ing on s ucr o se an l ac t o s e . In Vi tro 1 6 , 1 0 : 90 7-9 1 2 .
2 1 . MURASH IBE , SKOOG 1 9 6 2 . A revised med i um for rap i d growth and b io as says w i th tobacco t i s s ue cul tures . Phy s i o l og i c a l P l antarum 1 5 : 4 7 3-49 7 .
2 2 . NAYUDU 1 9 64 . Var iat i on 1n X . ma lvacearum . Ind ian Cot ton Growe r Review 1 8 350-355 .
23 . PARLEVLIET 1 9 83 . Can h o r i zontal res is t ance be r e c o gni s e d in the presence of ve r t i c a l re s i s tance in p l an t s exposed to a mixture o f pathogene races ? Phytopa th o l o gy 3 , 7 3 : 3 7 9 .
24 . PARLEVLIET , ZADOKS 1 9 7 7 . The inte gra t e d concept o f d i s ease res i s t ance ; a new view inc l uding hori zontal and ve r t ical re s i s t ance in p l ant s . Euphyt ica 26 , 5 -2 1 .
25 . PARRY 1 9 8 2 . Le c o tonnier e t s e s produi ts . Ma i sonneuve e t Laros e , Par i s 5 0 2 p .
26 . PR ICE , SMITH , GRUMELES 1 9 7 7 . Cal l us c u l t ures o f s ix s pe c i e s o f c o t t on ( Go s sypium L . ) on def ined me d i a . · P l ant S c ience Le t t e r s 1 0 : 1 1 5- 1 1 9 .

1 '5
2 7 . RUYACK , DOWN ING , S U CHANG, MITCHELL 1 9 79 . Growth o f c a l lus and s u s pens l on cul ture c e l l s from co t ton var i e t ie s ( Go s syp i um h ir s u tum L . ) re s i s t ant and sucep t ib l e t o Xanthomonas ma lvacearum ( E . F . SM . ) Dows . In V i tro 1 5 , 5 : 3 6 8- 37 3 .
2 8 . SAPPENFIELD 1 9 83 . Bac ter i a l b l i ght o f c o t t on : t echnique s and procedur e s f o r b r e e d ing r e s i s tan t or imrnur i e cul t iver s . S o ut hern Cooperat ive S e r i e s Bul l e t in 2 80 .
2 9 . SHENK , H I LDEBRANDT 1 9 7 2 . Med i um and technique s f o r induc t ion and growth o f monoc o ty l e donous and d i co t y l e donous pl ant c e l l c u l t ure s . Canad i an Journal o f B o t any 50 : 1 99-204 .
30 . VALES 1 9 8 3 . Des conna i s s ances s ur l e s r e l a t ions hô te-paras i t e aux s t ratégies de l u t t e c o n t re l a Pyr i c u l ar i o s e du r i z . Thè s e Unive rs i t aire , F ac u l t é d ' Ors ay : 3 1 0 p .
3 1 . VANDERPLANK 1 9 78 . Gene t i c and mo l e cular b a s i s o f p l ant patho gene s i s . Springerve r l a g , Ber l in : 1 6 7 p .
3 2 . VANDERPLA�� 1 9 8 2 . Ho s t- pa thogen interac t i on s . P l ant D i s e a s e Academie Pre s s , New Yo rk : 20 7 p .
3 3 . \�F� , S IN GH 1 9 7 6 . Rac e s o f Xan thomona s ma l vacearurn , l o s s in their virulence and the p ro t e c t ive e f f e c t of avirulent s tra ins heat-h i l l e d c e l l s and phy l lop l ane b a c t e r i a . Journal of P lant D i s e a s e and Pro tec t ion 8 3 , 1 2 : 7 4 8 - 7 5 7 .
34 . VERMA, CHO\vDHURY , S IN GH 1 9 7 9 . Interac t i ons b e tween d i f f e r ent races o f X . ma l vacearum in l e ave s o f G . h i r s u tum . J ourna l o f P l ant Di s e a s e s and Pro t e c t io n 86 , 8 : 460-4 6 4 .

1
l
S OMMAIRE DES ANNE XE S
- F iche t ec hn iquè s ur X . ma l vacearurn ( Sm i t h ) Dye - Les gène s majeurs de r é s i s tance à la Bac t é r i os e - S ol u t i on nut r i t i ve p our l a c u l ture d e s p l an t s sur vermic u l i te - Mi l ieu de c u l ture ut i l i s é pour X . mal vacearum - Le s grade s d ' a t t aq ue de l a gamme de not a t i on de s s ymp tôme s - Mis e en évidence de l a réac t i on hype r s ens ible par inje c t i on - Tabl eaux n °l e t 2 - Tab l eaux n ° 3 e t 4 - Mi s e en évidence gr �phique d e s interac t i ons hôte-para s i t e
(c l a s s ement d e s s ouche s par var i é t é ) - Clas s emen t d e s souche s var i é t é par var i é t é apr ès r e gr oupement
s t a t i s t ique - Mi s e en évidence graph i que des interac t ions hô te -paras i t e
( c l as s emen t d e var i é t é s p a r s ouche ) - Clas s ement de s var i é t é s s ouche par s ouche après regroupement
s t a t i s t ique - Le s c l as s emen t s r é s u l tant de l ' e s s ai n ° 2 - Tab l eau n ° 5 ( ré s u l t a t s d e l ' e s s a i n ° 3) - S ch ématisa t i on des r é s ul t a t s de l ' e s s a i n ° 3 - Le s r é s u l t a t s ob t enus aux E ta t s-Un is - Repr é s entat ion graph ique d es r é s u l t a t s ob tenu s par ino cul a t i on
des rac e s 1 , 2, 1 8 s ur l a var i é t é S t onevil l e 20 - Les intera c t i ons obs ervé e s pour l ' e s s a i n ° l - Le s interac t i ons ob s er vé e s p our l ' e s s ai n ° 2
1 6
Anne xe n °
2 3
5 6
7
8
9
1 0
1 1
1 2
1 3
1 4
1 5 1 6 1 7
1 8 1 9 20
AN N E X E N o l
1� A c
1 l '
1 2 3 4 5 1 6 7 8 9
J O 1 1 1 2 1 3 1 4 1 5 1 6 1 7 1 8
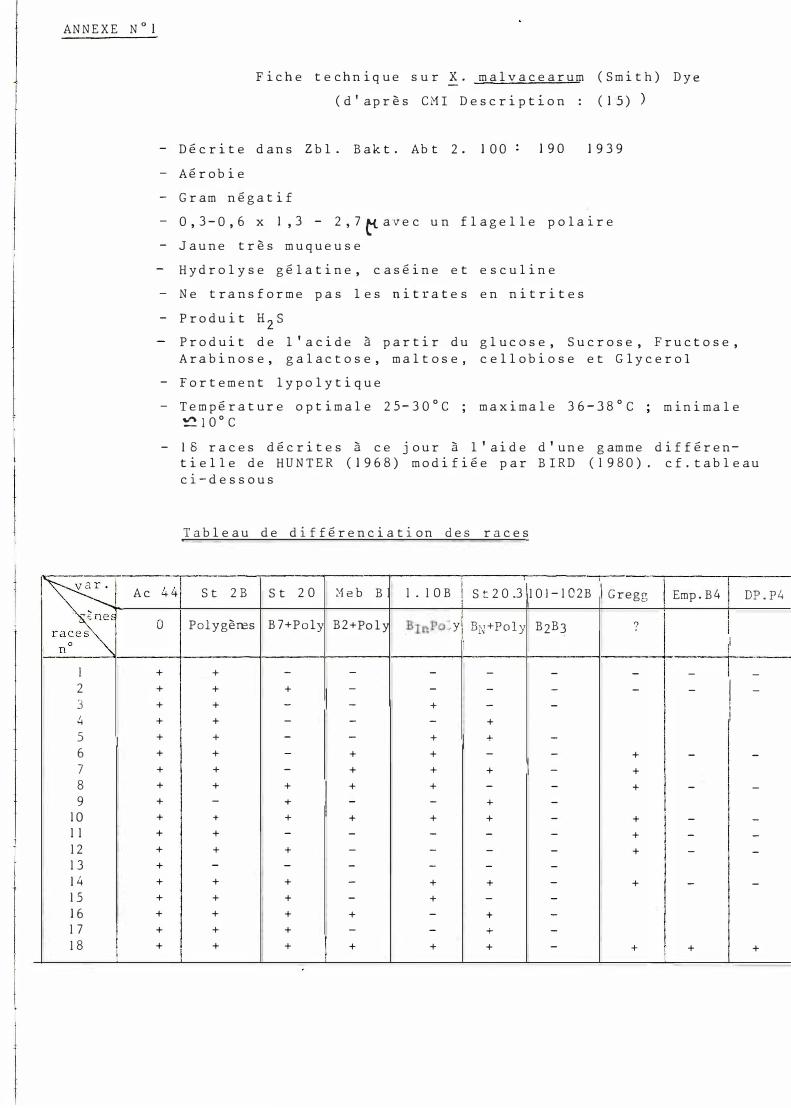
F i c h e t e c hn i q u e s u r ! · m a l v a c e a r u m ( S m i t h ) D y e ( d ' a p r è s C M I D e s c r i p t i o n : ( 1 5 )
- D é c r i t e d a n s Zb l . B a k t . Ab t 2 . 1 0 0 1 9 0 1 9 3 9 - A é r o b i e - G r am n é g a t i f - 0 , 3 - 0 , 6 x 1 , 3 - 2 , 7 t<- a v e c u n f l a g e l l e p o l a ir e - J a u n e t r è s m u q u e u s e
H y d r o l y s e g é l a t i n e , c a s é i n e e t e s c u l i n e - N e t r a n s f o r m e p a s l e s n i t r a t e s e n n i t r i t e s - P r o d u i t H 2 s
P r o d u i t d e l ' a c i d e à p a r t i r d u g l u c o s e , S u c r o s e , F r u c t o s e , A r a b i n o s e , g a l a c t o s e , m a l t o s e , c e l l o b i o s e e t G l y c e r o l
- F o r t e m e n t l y p o l y t i q u e - T e m p é r a t u r e o p t i m a l e 2 5 - 3 0 ° C ; m a x i m a l e 3 6 - 3 8 ° C
� l 0 ° C m i n i m a l e
- 1 8 r a c e s d é c r i t e s à c e j o u r à l ' a i d e d ' u n e g am m e d i f f é r e n t i e l l e d e H U N T E R ( 1 9 6 8 ) m o d i f i é e p a r B I R D ( 1 9 8 0 ) . c f . t a b l e a u c i- d e s s o u s
T a b l e a u d e d i f fé r e n c i a t i o n d e s r a c e s
4 41 1 S t 2 0 .3 \1 0 l - 1 0 2 B 1 1 Emp . B 4 S t 2 B S t 2 0 �1 e b B 1 1 . 1 O B 1 Gregg DP . P 4
i 0 Polygèœs B 7+P o ly B 2 +Po l y B rnPo lyl BN +P o l y B2B3 ? i
1 1 1 1 1
+ + - - - - - - - 1 -+ + + 1 - - - - - - 1 -+ + - - + - - 1
1 + + - - - + 1 + + - - + 1 + -
+ + - + + - - + - -+ + - + + + 1 - + + + + 1 + + - - + - -+ - + - - + -+ + + + + + - + - -
+ + - - - - - + - -+ + + - - - - + - -
+ - - - - - -+ + + - + + - + - -+ + + - + - -+ + + + - + -
+ + + - - + -
+ + + + + + - + + +

A N N E X E N ° 2
G è n e s .
B 1
B 2
B 3
B 4
B 5
B 6
B 7
B 8
B 9 K
!1 9 1.
B l O K
B I O L
B 1 1
B N
B S
B S m
B D m
B ?
L E S G E N E S MA J E U R S
S o u r c e s
O u g a n d a B J I ( h i r s u t u m )
O u g a n d a 6 3 1 1 A l b a r h i r s u t u rn U KB R -----
V a r i é t é s a m é r i c a i n e s
S c h r o e d e r 1 3 0 6 ( h i r s u t u m v . p u n c t a t u m )
N T R / 3 0 ( a r b o r e u m )
G r e n a d i n e w h i t e p o l l e n ( b a r b a d e n s e )
N T I 2 / 3 0 ( a r b o r e u m ) ? U K B R 6 1 / 1 2 (h1 r s u t um )
S t 2 0 e t a u t r e v a r . U S ( h i r s u t u m )
à n o m a l um e t v a r i é t é s d i p l o 1 d e s a f r i c a i n e s n o n c u l t i v é e s
W a y a d 8 ( v a r. c o m m e r c i a l e i n d i e n n e ) ( h e r b a c e um )
A l l e n S I - 2 9 6 ( h i r s u t u m )
K u f r a o a s i s ( h i r s u t urn v ar:püïïët a t u m )
i d il B 9 L
i d à B 9 K
v a r i é t é i n c o n n u e d e s U S A ( h i r s u t u m )
N o r t h ë r n s t a r
S t o r m p r o o f 1
S t . 2 B , E m p ( h i r s u t u m )
D e l t a p i n e ( h i r s u t u rn )
W e s t b u r n 7 0 m u t é p a r i r r a d i a t i o n ( h i. r s u t u m )
D 1 a p r è s I N N E S ( 1 Il ) F O L L I N ( 1 4 )
D e s c r i p t i o n
D o m i n a n t , f a i b l e
D om i n a n t , f o r t
P a r t i e l l e m e n t d o m i n a n t
P a r t i e l l e m e n t d o m i n an t
R é c e s s i f
D o m i n a n c e v a r i a b l e s e l o n l ' a r r i è r e p l a n g é n é t i q u e p a r f o i s e f f e t - s u r B z B 3 ( H u s a i n , S i n g h , S i n g h ,
A g n i h o t r i )
R é c e s s i f
f o r t e m e n t d o m i n a n t
f o r t e m P n t rl om i n � n t
f a i b l e , p a r t i e l l e m e n t d o m i n a n t
f a i b l e
f a i b l e
D om i n a n t
D o m i n an t
D o m i n a n t
P o l y g è n e c o m p l e x e
D o m i n a n t
H é r é d i t é d é t e r m i n é e p a r :
K n ig h t e t C l o u s t o n 1 9 3 9
Kn i g h t C l o u s t o n 1 9 3 9 I n n e s ! 9 6 5 e t 1 9 6 9 B r i n k e r h o f f 1 9 7 0
K n i g h t 1 9 4 4
Kn i g h t 1 9 4 8
Kn i g h t 1 9 5 0
K n i g h t 1· 9 5 3 S a u n d e r s & I n n e s 1 9 6 3 I n n e s 1. 9 6 9
K n i g h t 1' 9 5 3 G r e e n e t B r i n k e r h o f f f l 5 6 I n n e s e t B r o w n 1 9 6 9
K n i g h t 1' 9 5 4
Kn i g h t li 9 6 3 I n n e s 1 9 6 5
T . a e i. è r e � 1 9 6 0 I n n e s 1 9 6 5
K n i g h t 1 9 5 7 I n n e s 1 9 6 5
L a g i è r e s 1· 9 6 0 I n n e s 1 9 6 5
I n n e s 1 9 6 6
G r e e n B r i n k e r h o f f 1 9 6 6 B r i n k e r h o f f 1 9 6 6
G r e e n e t B r i n k e r h o f f 1 9 5 6
G r e e n e t B r i n k e r h o f f t 9 5 6 B i r d e t H a d l e y s 1 9 5 8
B r i n k e r h o f f V e r h a l e n M a m a g h a n & J o h n s o n 1 9 7 8

S o l u t i o n N u t r i t i v e = = � = = = = = = = = = = = = = = =
S o l u t i o n A
K 2 HP 0 4
H g S 0 4 7 H 2 0
H N 0 3 C o n c e n t r é
S o l u t i o n o l i g o é l é m e n t H 2 0 d i s t i l l é e
S o l u t i o n B K N 0 3
N H 4 N 0 3
H a s c o l a t e d e F e r H 2 0 d i s t i l l é e
P a r l i t r e d ' e a u d a n s l ' o r d r e
3 0 g
2 0g
9 7 m l
2 0 m l
9 0 0 m l
5 2 , 5g
4 8 , 0 g
6 m l
9 0 0 m l
S m l d e s o l u t i o n A
l O O O m l d ' e a u S m l d e s o l u t i o n B

A N N E X E N ° 5
M i l i e u d e c u l t u r e u t i l i s � e
p o u r � · m a l v a c e a r um
P o m m e d e t e r r e , C a r o t t e , D e x t r o s e , A g a r . ( P D A m o d i f i é )
P D A d i f c o 3 9 g P e p t o n e 2 , 5 g E x t r a i t d e l e v u r e O , S g M g S 0 4 0 , 3 g C a C o 3 0 , 2 g J u s d e c a r o t t e ! S m l H z O d i s t i l l � e l O O O m l
Aut o c l avé à l 20 ° C pendant 20 minu t e s

A N N E X E N ° 6
2
4
L e s g r a d e s d e l a g a m m e d e n o t a t i on
h y p e r s e n s i b i l i t é
f ront d ' inf e c t i on con fondu ave c l a b l e s s ure d ' inocu l a t i on
• 1 \ ) ' 1,
f r ont d ' infec t i on l argement d i s t inct de la b l e s s u re
3 : fron t d ' in f e c t ion d i s t inct de la b l e s s ure
5 po in t s d ' infe c t i on -di s t an t s de l a zone d ' inocu l a t ion

A N N E XE N ° 7
M i s e e n ê v i d e n c e d e l a r é a c t i o n h y p e r s e n s i b l e p a r i n j e c t i o n
h y p e r s e n s i b i l i t é
s e n s i b i l i t é
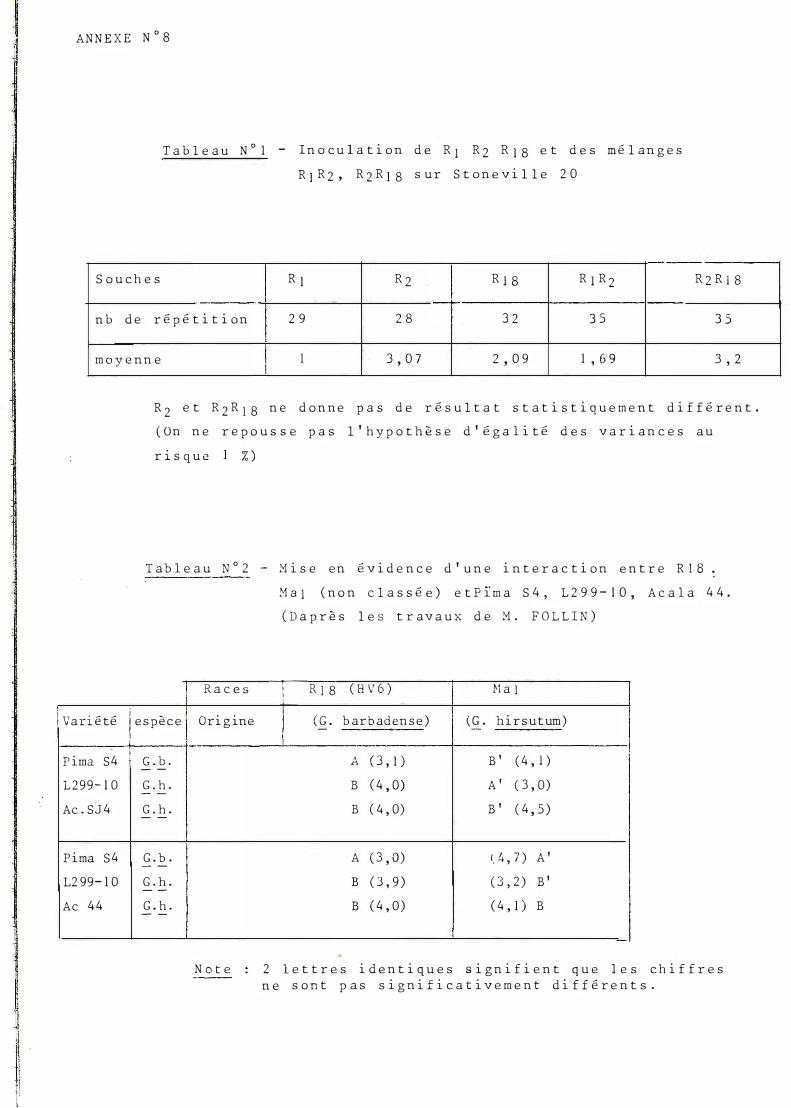
1 A N N E X E N ° 8
T a b l e a u N ° l - I n o c u l a t i o n d e R j R 2 R J S e t d e s m é l a n g e s
R J R 2 , R 2 R 1 8 s u r S t o n e v i l l e 2 0
----S o u c h e s R 1 R 2 -t R J S R ] R 2 R 2 R l 8
n b d e r é p é t i t i o n 2 9 2 8 3 2 3 5 3 5
m o y e n n e 1 3 , 0 7 2 , 0 9 1 ' 6 9 3 ' 2
R 2 e t R 2 R 1 8 n e d o n n e p a s d e r é s u l t a t s t a t i s t i q u e m e n t d i f f é r e n t .
( O n n e r e p o u s s e p a s l ' h y p o t h è s e d ' é g a l i t é d e s v a r i a n c e s a u
r � s q u e %)
T a b l e a u N ° 2 - M i s e e n é v i d e n c e d ' u n e i n t e r a c t i o n e n t r e R I B :
1 R a c e s 1
M a J ( n o n c l a s s é e ) e t P Ï m a S 4 , 1 2 9 9 - 1 0 , A c a l a 4 4 .
( D a p r è s l e s t r a v a u x d e M . F 0 1 L I N )
\R 1 8 ( H V 6 ) 1 �l a 1
1 V a ri é té i ! e s p è c e O r i g ine j ( G . b ar b aà en s e ) 1 ( G . h i r s u t um )
1
r P ima S 4 G . b . - -1 2 9 9 - 1 0 G . h . - -Ac . S J 4 G . h . - -P ima S 4 G . b . - -12 9 9 - 1 0 G . h . - -Ac 4 4 G . h . - -
N o t e
- -\
A ( 3 ' 1 ) B ' ( 4 ' 1 ) i
B ( 4 , O) A ' ( 3 ' 0 )
B ( 4 ' 0) B ' ( 4 , 5 )
A ( 3 ' 0 ) (_ 4 , 7 ) A '
B ( 3 , 9 ) ( 3 , 2 ) B ' B ( 4 ' 0 ) ( 4 ' 1 ) B
1
2 l e t t r e s i d e n t i q u e s s i g n i f i e n t q u e l e s c h i f f r e s n e s o n t p a s s i g n i f i c a t i v e m e n t d i f f é r e n t s .
-
A N N E X E N ° 9
T a b l e a u N ° 3 - I n o c u l a t i o n d e s n o u v e l l e s s o u c h e s s u r v a r i é t é à r é s i s t an c e v e r t i c a l e v a r i a b l e ( 1 0 r é p é t i t i o n s )
G . b . = G o s s y p i u m b a r b a d e n s e
G . h . = G o s s y p i u m h i r s u t u m
V a r i é t é s P i m a S 4 M e b B 1 1 0 l 1 0 2 B G r e g g A c a l a s
�
Souche Ori g ine Ré s is t ance 0 B 2 +Po l y gènes B 2 B3 ? 0 ver t i ca l e
T 1 G . h 2 3 , 5 3 , l 2 , 6 2 , 4 - - l T2 G . h 1 , 9 3 , 2 5 3 , 2 5 3 , 3 2 , 4 - -
1 1 1
S J G . b i 1 1 , 7 l 2 , l 2 , 1 - - 1
S 3 G . b 1 l , 5 l , 8 l ' 5 2 ' l 2 , 2 - -
1
HV ] G . h l , 8 3 , 4 3 , 7 2 , 7 2 , 2 - -
HV3 G . h - -
1 , 9 3 2 , 8 2. , 35 2 , 7 5 HV5 G . h
- -2 , 1 5 2 , 5 3 2 , 4 2 , 7 5
HV 7 G . h 1 ', 2 , 2 l , 9 2 2 , 1 - -
HV25 G . h .2... , 2 2 , 6 3 , 5 3 , 1 2 , 5
1 - -
1
Tab l e au N ° 4 - 2e e s s a J. d ' i n o c u l a t i o n d e s n o u v e l l e s s o u c h e s ( 2 0 r é p é t i t i o n -
�1 1 1 1 P i m a S 4 M e b a n e B 1 1 1 0 1 1 0 2 B G r e g g y A c a l a S J 1 1 1
T 2 1 2 2 ' 9 5 2 , 8 1 3 , 1 5 2 , 8 5 1
S 3 2 , 4 2 , 4 5 2 , 1 2 , 5 2 ' 9 HV 1 2 , 0 5 2 , 8 2 , 8 5 2 , 9 5 2 , 0 5 H v 2 5 2 , 0 5 2 , 6 5 2 , 8 0 2 , 6 0 2 , l 0
;r ade c l ' a t t aq u e
3
2
.... . . . . . . . . . . . . . . . . . / .. .
. . ·
·· · · · · · ,. -�· /
" • - . , ..,..--- -.....::::::-- -+ /
• . . . .... .
� � �
. .. � "' T��- / + � .
�
� ·� � .
++ 7; ..... . / . ""'-.. ·... "" .. ·· / / �,;,....._ . " · . ·- -
. �
•' + / '+
• r
.. / / / .. � ··- � -··
. • + •• � -- " , .
... y / / ·� ., • • •
. r ___.. • _ __. •• .... ··
·. ..._-.;·
• + ..--• .... '·· • "'
_.. /A__.. / "-- . '· • •
. � -�� . .. ......
.-·-----· /
.. ......
· .....
�. /7' ,.
.. . ---- ...
.....--1.}:.-<' •• /
... � ... -· ··-...... -
/} / .. . ./ ....---...
�·�- . - ·· . ·
· ..
. ..._ ...---- . . -
... �- . ---
... ,
,· 7 ...---- •
.. ..
... -:t' - . ---
.. .;..
.- � - -�
.. � ...
• ___.. 1> --- ••••••• ••
/• . . _ . ...... ... - · - · ---..,._ t. - t. - b - -;our- _
_ _ __ _ _ _ _; _ _ _ _
oo / _ _ _ __ _ ..
o o ",..,,. .-., ,. ,. .,
•
•
•
•
/
/
.... . . n "'oo oO
/
..,o• •o r.... Oo •
# /
ooo•o /'" ' <� o., _.,••" / .,o•o ; ' "' o•
.,- / • .,.ao ,.,-"' ' o.,,. ,. 0et0 /
,., " .... , ""•o ... •o /
/ ' ...... tl'(r /
/ .... o • ....,.o /
/ ' ...... "
/ ' /
/ ' /
,., "" .... /
/ ....
"
,.,"" .... /
/ ....
"
, "' .... //
/ .... /
/ .... /
"' ...... /
.... /
' ...//
Lége n d e
- · · · · · · · · �
ICJ< Il • JI. A JI le )1
- 6 - 4 -
- ;Jr --'l -
T I T z S I S 3
HV J HV3
HVs H V 7
HV 2 S
P i ma S 4 �kb a n e B 1 1 0 1 1 0 2 B G r e g g Ac a l a S J 4 Mi s e en � v i d e n c e g r a ph i qu e d e s i n t e r a c t i o n s h ô t e - p a r a s i t e
)> z z tT:1 :x: tT:1
0
4x -f Grade j Ar c s in�------, :d ' a t t a q e
3 -
I l _
1 � ' . . .... . .. .... . . . . ... . .. . . . . .. . ... . . . . ... . . . . . . . .. .......... . . . . .. . 1 ' T t T 2 V
LJ T2
H··· - -- · - - · -.... . . . . . . . . . . , .. _ .. , . . . . . . . . . .. . . . .. . . . . . . . . . .. . . . . . . . . . . . .. . . . . HI J IV3 H s HV25 l � J O �--�--. . . - . . _ . . . . . . . . . . . - . . . . . . � . .
T l T 2 L--.J .. . . .. . - · - · - · · · · --- - - . . · - · · - · · HV J HV3 .. .. . . .. . . .. . . . . . . .. _ .. . . . . . . . . . . �� · - - - - - � - · · · · .. . . . . _ . ... . .. . ... ... . . �
9 -T 1
_mu_]
T 1 T 2 1 J 1 t T 1 T 2 ] J t 1·- · · - · ··-. .. . . . .. . . ... . . . . .. . ... . +===1-.... . . .. .. . .. . . . . . . . . . -. ....... . . . . � t · ··-·- ---- · - . . .. - r .... . _ _ _ , _ _l Il HV 1 HV3 HVS H V 7 . . . - -- · · - · · · . . · · · -- · - . . . .. ..... . . . . . . . _ . . .... .. . . . · - - - -- - � - -... . . . . . . . . . . .... . . .... .. . .. . . .... . . . .. ..... !=='��!
s 1 S
3 r ·
If---tH V 1 HV 5 Hf I-lVZ S 9 1 S 3
2 -
1 -
8 - HV 1 HV 3 HV5 HV 2 5
6 -
HV 7 � ···
·+l---tl S 1 S 3 j , 1 !!Y.Z.J
1 l · ·- · · · · . . . . . - . . . . . . . . . . ·• · · ··1----4 · . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . 1 1 l !V7 ..S.l
1 1 �-· · · -. .. .... .. . . . .. . .. . 1 ! . . . . . .. . .. . -.. --.. . . . . . . . . . .. . ... �
� 5 ..
�
:;t> z z tT1 :x: 1:>1 z
0
----t--+----------t---t---------+--·+- 1-- �_____.. Plt1A S 4 M E BANE B I 1 0 1 - 1 0 2 B Gl{EGGY
C l a s s e men t des nouve l l e s s o u c h e s v a r i ê tê par v a r i ê t ê a p r è s r e g r oupeme n t s t a t i s t i q u e
( e s s a i n ° l )
ACALA SJ 4
Gr-ad e d ' a t t a q ue
"'· "·� .... . . . . . . " · · · · . .
..
! :
.A �
\.. / ·. o
1 ·
'· ·. . '-..
/ " \\ 1 f"··. • .
'-..
/ •
0
1 . .
:
; ' '
.. \ ' \
. .
.
/ .
' •,
'
() \ Il
1 : ··...... ""' ·· · · · ·•. ! 1
/ .
\
. ......:/"';. . - · .
' .
\·. 1 ! ;r .... .. . .<-:·· · · · ..... ! ;
\ ' : /' •• . ... .0 ·.
:
� -�· · ·""'"·� ... \ \ 1 / • • '-.. / ' ·.\.
/:j ..
� '\
. / . ... . .
......... • 0 1 :' · .,. . ,...
: ..
T I Tz
.... . _; '-..
·. '
� \ 1 ... .
\ :/ /
'"' • . -1
"' . . /
·. "' • _ __,.�--�L':._ " '-- •
, ._ :• -•-'
•. ..., , � .� V"":'"'"''lf'""'�_.. - • - o
�·· ;· / .,. r
�. \ �---� J :
� . .......... • •••
•,
- e - o _/ '
'-.. �� '1-1-
• \ .:
0 •
. _} : �, � '
. .
• .. ·'
·, \....- - -'- ·
Vi
•
v
Lé g e n d e
-- o -- �> --
.._><AX-'U\.>C JI( -o-
s 1 8 3 liV I HV3 HVS Mi s e en é v i dence g r D�hi que d e s i n t e r a c t i on s h8 t e- p ar a s i t e
P ima S 4 Meb ane B I
1 0 1 1 02 B G r e g g A c a l a SJ4
HV 7 H V 2 S
> z z M �� M z ' 0 N
Ar·c ,------·--· ---G r a d e :!.. �d ' a t t aque .'J 1 00
3 -
2 -
1 -
.::t::
I l - , 1 � .. . . _
.
... ... . . H ..
. · .. - - ---- ·t-+ ·· --- ·· --· - -H· - . . . . .. .. ;. · ·t-w t·1, 1 1
t , . . . . . . . . . .. . . _ M 1- t--+-· - - · · · ·-·-- . . +-4 · · - · · ·- · · . . · - ·�-+· .. · · .. · · ·· · · ·-- · ·H-· - ... ·· ·- · · . . -4---l . . · · - -· . . . . ..... . +---4 .. · · - - - . . . .. ..
. . 1--. . . . . .. ... . . . . . . . �1 , 1 , G
-·-- · · ·· · · · ·-···w·· -· .. ... . .. ··+--+ -· · · · · · · . . · ·-t--i -···- - · - · · · · · H · .. · · · · · · · -- --�--L__l-(1 -··- -·· · ·· · · · · . . · ·- · . .•.... � .. -- . . · · - · · ---�-i · · · ·- · . . .. · · ·H·· . . •· · · . · · · �.z...!.� ��
1 1----- ------ . -1-4--------- · -- + - +--- - - - - -- . - - · · 1-+ - - --- - - - - - - - -+-kJ -
1 0 -
L.QJ
9 -{
t-·----- -----�---. . . . .. .
·-a--· - ·
- · . ·- -. rE- - -
··-··
···
-·-�- - -- · · .. . . .
·
. .
-·.J$·
.
.. . ..
.
.. . �------... . . . .. . .
.
.
_
. . . . , . . ..... L-JM,A,Pl
• ..
----·-
-·- -· · - . .. . .. . . . . ... --····---
--.. . . . .. . . .. .. . . . . G 1 � t S;:E:;A�}.. .
.
. . . . . . .. . . .. . . . . . . . . . . . . . . . . .. . . . .. .. . . . . ... - -. . . . .. . . .. � . . .. . �1 , G , P , A . ·= . ...:r. . .. p A 1 1 . . . _.__ _ _ _ _ ' - -· ··- ..... . . . . . . . . ... . A
·· ·· ··- ··· · · · H···-·· ···- - -� 8 -
7 -
6 -
5 -
j r-·-·- -· ·- - ·--+-4 .. -. ... ........ . L.J G . A I . . . .. L-J .. . . .. . .. .... .. L..JP,A]
--·--- -· · · - - - r-1 - - - .. · - ·· --� -· ··· ·H - -· · · . . · · · ·· · r� --- · ·· ·1---... . . . . . . . .. . t--i . . ... . .
.
. . .. . .. �.- -.. .. . . . .. ,_ ... i 1 --... ..
... l.J:-1 1 I--·-·-'···· · · ··H- -· ·-- · ·· r-l-!2J.. .. . .. . .. . H , 1 , G , P l �
1 1 - - · ·-- · ·· ·-·� - ------ ·- - . . .. - -1--i �
::t:: .,..___ Tl T2 s 1 ..__ 8 3 - 1 L • ._!!.V ) -·-- '--1:1.V3 HVS HV7 J.IU? '>
C l as semen t d e s var i é té s s ou c h e s p ar. s m.i ch e a p r è s r e groupeme n t s t a t i s t i qu e ( E s s a i N ° 1 )
1 ::.> z z tri ><: tri z
0 -w
L é ge nde
[ill M e b an e..BI
[!] 1 0 1 - 1 0�1
[ÇJ G r e gg tru P i ma S 4
�A c a l a S J 4
S o u c h e s
A N N E X E N ° 1 4 LE S CLAS S EMENTS RESULTANT DE L ' E S SAI N ° 2
3
C l a s s ement des s ouch e s var i é t é par var i é t é ap rès regroupement s t a t i s t ique .
� rx X Arc s �n� �--1G��-d� ��d ' a.t t
.aq � �� -�-----------------=--;-,
- - - --� - - - - -1----11 - - - --
- - - -t-----t _ 1 _ l _
_ m 1 ��� ---�- -; � - - -� ��-�+-+ - - - . . .. • ! - - ··· �-- - - _ _ __ _ _ · __ ·---- ·- ��
-�- - - - · - -- - �· � -=-�----- =--=-=-, . . ,- � - - --- - - -H------ - - --=- - - - - - - - - 3r2J � · · · J �- -- - -----H----- - -- TzHV J HVzs
. � - · - .. �· - � i � . 3-T2HV J HV25 ! . . - �_,_j_--ll RV J HV2SS3 Il. +----+- - -- - - - -- r-- - - - - --� 1 - • · -- - -
. 1 - 1 -� � . . �- ·- . . . . . 1 -- � - .
1 t - . -· J 1 1 1
1 . 1
j · 2 1
3
2
1
X Arc Grade
1 � � � � - -� · ·- · ·
. . .
J O
T 2 HV l HV2 5
C l a s s ement des var i é t é s s ouch e s p ar s ouche aprè s regroupement s t at i st i que
1 - . .
-
AN N E X E N ° 1 5
T a b l e
V a r i é t é s
S o u c h e s
T 1
T 2
s 1 1
s 2 1 S 3 -+--HV 1 1 H V 3 1 1 HV s 1 1 H V 7
HV 2 s l
a u N ° 5 - E s s a i d e s n o u v e l l e s s o u c h e s s u r un e g a m m e d e v a r i é t é s p o s s é d a n t t o u t e s l a c o m b i n a i s o n B 2 B 3
( n b d e r é p é t i t i o n v a r i a n t d e 8 à 1 8 )
B 5 0 B J A 5 9 2 J 1 9 3 1 M K 7 3 1 0 1
1
2 ' 7 1 2 3 1 2
! 1 2 ' 8 2 ' 3 3 , 3 2
2 1 1 2 2 1 2
1 ' 4 2 2 • 1 1
2
1 ' 9 2 1 ' 9 2
2 , 6 2 3 2 ' 1
2 . 1 2 ' 3 2 ' 6 2 ' 3
2 ' 4 2 2 ' 9 2 ' 1
2 ' 6 2 3 2 ' 3
2 ' 8 2 2 • 5 2 ' 2
1 0 2 B
3
2 ' 6
1 ' 5
1 ' 5
2
2 ' 7
2 ' 1
2 ' 4
3
2 • 6
AN N E X E N ° 1 6 S ch éma t i s a t ion de s r é su l t a t s de l ' e s s a i n ° 3
�---·----: -·· · · · ·-· -- · · · · · ·- · --- - --- --- - - - - · - ·-·· : -- - �-----. --·- i---------, . ' . . -- · - - · . - . . - - - .. . - -' . . ' .
.. .. - - - - - - -·-- - - -�-��-- - · -; -�--�--:---�-- ---------:-' : 1.1"'1
- - -- - - - ·· ·· · · \,
, . . -- \· - - � - -;. -- ··
- - - -=-N---�-N-�-;--;r-o;�-.-4 ·. .. . . .. E-< E-< . CJ:l CJ:l . CJ:l ::r: - ::r:: ::r: ::r: - ::r: ....
'" \. \\ · - ! - �= '\ \.-4 \ \ t � •• \ 1- ..
• • Ir . ... _ -
·� \\ f ': \ j � - \ · -
.. - .,: • •• t: . - ; L ·· · -- · 1: -• ·- ··· - - - - -· · - · • • - - - � -
. ---- - -· ---��-�--- · · - · - - · · - � - � ·-·
'• \ 1: . ! . .
0\.l ... ... 1. . . . · - - -
· · - ·- ---· - A: - )j .- · - - ·- - - -- - - -- - ------- - - - -- -·-· -
� � ,.,,:· fi - c ... • .. 0 I l ·' .. � � e1 o
0 � • l • /.J' " . , : ? ...- , . : , -: vr _.· . , : /. ••· ... • 0 // .·· � If e l o
/ .. .. ··1 t 1 : 1 : / 1
.· . 1 < _ : l � / 'D / t ' : • i / h ' . �r • J :
o:1 N 0
1 -0
< ·· .. < . ! \·- - ·- � : · -
r·
' ·,. __ . 1 .. � 1 9
. ·- - - - . · ·-- : - ------ --- -- -- ----r "'
•
•• •• ,, � . \ • 1 : " ·,., . ,._ � 1 : ' '"·. ' " •
• 1 0 ' ··.' • \ - 0
' ··.� � � 1 : �.\ . . , . ' �· . \ :, : '- . ;-;... ·� D " ��· " • 0
' � � \ � : . " � � . , : "\ �l! . .
· -� - .. . 6! • •• •
1 1.: 1 .. ·
, :: 1 t: · ; 1 � 1 ;j , � . 1 ::
t .-\' À 1 •o
/ / ,. " " 1 :: "( 1 :: 1/ 1" :Jq 1 \ 1 :: 1 ! . " 1 � , � ...... 1 � 1 !: / ,i7 . .. 1 ::
__ _ t-+- . .f 1 . t L .. ::
1 1 l 1
- . . . . .. .. i l . 1 N
- ·· · ···-···- - -� -- ---·-·:- - �----�-- - - - -·- - - --- 1 : 1 .1
- 1 - - : · . - - l
l 1 · l . 1 . . i
- - - - · ---:- - -- - - - · - - :-·· --- - - --------t- �
A N N E X E N ° 1 7 R é s ul ta t s obt enus aux E t a t s- Uni s
T a b l e a u N ° 6 - . Ino c u l a t ion s ur vra i e s feui l l e s
var i é t é s 1 · -
M e b B 1 1 0 1 - 1 02 B Gregg Acala 44 s ouch e s
T l 6 , 6 6 , 0 6 5 , 4
T2 6 , 6 5 , 8 6 , 4 5 , 4
S 3 5 , 8 4 , 5 4 , 7 5 , 5
s l 5 , 2 5 , 4 5 , 4 4 , 8
S z 5 , 2 5 , 0 5 , 4 4 , 6 1 1
HV J 7 ? -, - 8 , 8 6 , 2- 6 , 4 7 , 0- 8 , 5 6- 7 ' 5 1 HV 3 8 , 2 6 , 3 8 , 2 7 , 8
HV 5 7 , 8 5 , 3 7 , 3 7
HV 7 6 , 2 4 , 0 4 ' 7 5 , 7 1 HV 25 6 , 3 5 , 6 6 , 4 5 , 4
1
T a b l e a u N ° 7 - Inoculat ion s ur feui l l e s cotyl édonna ires
var i é t é s M e b B 1 1 0 1 - 1 0 2 B G r e g g 1 A c a l a 4 4 souche s
T J 6 , 6 6 , 0 6 , 0 5 , 4
T 2 6 , 6 5 , 8 6 , 4 5 , 4 1 S3 3 , 7 3 , 6 4 , 3 2 , 7 1 S 1 5 , 2 5 , 4 5 , 4 4 , 8
s2 5 , 2 5 , 0 5 , 4 4 , 6
HV 1 7 , 2 6 , 4- 6 , 6 7- 7 ' 1 6 , 0- 6 , 2
HV3 6 , 7 6 ' 1 6 ' 1 5 , 8
nv5 6 , 8 6 , 2 6 , 4 5 , 8
HV 7 4 , 8 4 , 2 3 , 6 3 , 3
HV2 5 6 , 3 5 , 6 6 , 4 5 , 4
ANNEXE N ° 1 8
Grade d ' a t t aque
3 -
2 -
-
R R I R2 R l 8 R2 R2 R I B 1
Repr é s entation graph i que d e s r é s u l t a t s ob tenus par inoculat ion
des races 1 , 2 e t 1 8 sur S to nevi l l e 20
ANNEXE N o 1 9 Le s interac t ions o b s ervée s pour l ' e s s a i n ° l
A) Clas s ement d e s s ouches
B) C l as sement d e s var ié t é s
Arcs in � 1 00
l l .• 1 0 1 . 1 0 2B . .
1 0 . . ·
.·
/ Gre g g 9 /
.. ·· / 8 �Ac SJ4
... ,. ,y
7
S J HV 1
Arc s in � 1 00
I l ,..... Gre g g
1 0 /
/ 9 Meb B I
8
7
Ar cs in_ fx "lOO
1 1
1 0
9 X
T2 = HV 25
' HV 8 3
7
Acala S J 4 Gregg
Arc s in � 1 00
Arc, s i n � 1 00
l 1 1 1
1 0 Ac S J 4 1 0
9 - - 9 - - Gregg
8 8
7 7
HV 1 HV3 i T l
Arc s in � 1 00
Arc in pç; ' 1 00
1 1 l i ., Gregg
1 0 / 1 0 /
� Meb B I 9 9
8 8
7 7
Meb
1 HV 2 5
Ac S
Les interact ions ob s ervé e s pour l ' e s s a i n ° 2
A ) Clas semen t de s s o uches
Arc s in - � li DO
I l
1 0 T = HV ...... - 2 1 9 ........
�= HVzs
,. s 3 8 ........
7
Pima S 4 1 0 l . 1 0 2B
B ) C l as s ement des var i é t é s
Arc s in _ /x llüo
I l
J O .......
9
8
7
1 0 1 . 1 0 2B
Arc s in #a-1 00
Ar c s · n � l OO
1 l J I
1 0 J O _...... 1 0 1 . 1 0 2B / 1 0 1 . 1 0 2B ........ / / 9 /
9 � P ima sq Ac : SJ4 8
8
7 7
. _HV l
s 3
..... ...... -... HV 1 HV2 5
Ac a l a SJ4
Ar cs · n� 1 00
1 1
1 0
9
8 Ac SJ
7