Embed Size (px)
Citation preview

Rapport de stage de recherche de Master 2 Soutenu à Rennes le 14 juin 2010
Analyse comparative des traits d’histoire de vie chez le parasitoïde
Leptopilina heterotoma
Pauline Vuarin M2 Ecologie Fonctionnelle, Comportementale et Evolutive
UFR SVE - Université de Rennes 1
Laboratoire d’accueil : EcoBio - UMR 6553
Responsable scientifique : Joan van Baaren

REMERCIEMENTS
Je remercie chaleureusement Joan van Baaren pour son implication, tant professionnelle que personnelle, dans la bonne réussite de ce stage. Merci pour les conseils méthodologiques, les remarques toujours pertinentes et les propositions nouvelles auxquelles je n’aurais jamais pensé, qui ont grandement contribué à ma progression dans la réalisation des expériences mais aussi dans la rédaction de ce rapport. Merci également pour avoir pris le temps d’aborder avec moi des sujets sans rapport avec le stage, et qui m’ont permis d’en connaître davantage sur le monde de la recherche, sur le recrutement en thèse, ou encore sur les voyages à l’autre bout du monde… Enfin, et c’est le principal à mes yeux, merci de m’avoir encouragée jusqu’au bout et d’avoir fait preuve de compréhension vis-à-vis de mes inquiétudes et de mes préoccupations. Je remercie également l’équipe de choc qui m’a si bien accueillie :
- Cécile pour ses cris de guerre dans les couloirs, dignes des plus beaux hakas, qui m’ont redonné espoir et courage quand ces derniers venaient à manquer !...
- Joffrey pour s’être abstenu de commenter mes erreurs et maladresses, dont il a souvent été le premier témoin ! Et pour ses discussions ornitho !
- Véronique pour ses connaissances passionnantes sur la diversité de couleurs que les moisissures peuvent prendre dans les pots d’élevage !
- Damien pour son aide précieuse dans les innombrables tentatives de réparation du respiromètre !
- Philippe pour m’avoir dévoilé les secrets de l’attirance irrépressible des femelles parasitoïdes pour les larves…
Merci à tous d’avoir répondu à mes questions, d’avoir discuté et partagé vos connaissances avec moi, et ce même sur le temps des pauses, et de m’avoir encouragée jusqu’à la fin. Je tiens surtout, avant tout, envers et contre tout, à remercier Natacha, sans qui les carreaux de chocolat, les discussions personnelles en plein milieu du couloir, les angoisses partagées ou encore les commentaires de messages d’erreur communiqués par R n’auraient pas eu le même intérêt ! Je remercie bien sûr les habitués du RU à 11h30 pile, qui m’ont apporté fous rires, discussions légères mais aussi réflexions scientifiques passionnantes ! Spéciale dédicace à celui qui a pimenté mes pauses thé ! Enfin, je remercie mes parents et ma sœur qui ont contribué à leur manière à l’élaboration de ce rapport via les longues conversations téléphoniques, ainsi que mes amis pour leur soutien, même s’ils n’ont pas vraiment compris le but de mon stage !...

SOMMAIRE
1. INTRODUCTION........................................................................................... 1
2. MATERIELS ET METHODES .................................................................... 4
2.1. MATERIEL BIOLOGIQUE ............................................................................................................................. 4 2.1.1 Biologie des espèces ............................................................................................................................. 4 2.1.2 Origine des espèces .............................................................................................................................. 4 2.1.3. Elevage ................................................................................................................................................ 5
2.2. EXPERIENCES.............................................................................................................................................. 6 2.2.1. Paramètres de développement ............................................................................................................. 6 2.2.2 Mesures biométriques........................................................................................................................... 6 2.2.3. Mesure du taux de métabolisme........................................................................................................... 7 2.2.4. Mesure des taux de lipides................................................................................................................... 7 2.2.5. Mesure de l’activité locomotrice ......................................................................................................... 8 2.2.6. Autres mesures..................................................................................................................................... 8 2.2.7. Motivation des femelles ....................................................................................................................... 8
2.3. ANALYSES STATISTIQUES ......................................................................................................................... 10
3. RESULTATS ................................................................................................. 11
3.1. PARAMETRES DE DEVELOPPEMENT ......................................................................................................... 11 3.2. MESURES BIOMETRIQUES ......................................................................................................................... 11 3.3. TAUX DE METABOLISME ........................................................................................................................... 11 3.4. ACTIVITE LOCOMOTRICE ......................................................................................................................... 12 3.5. TAUX DE LIPIDES ....................................................................................................................................... 13 3.6. AUTRES MESURES..................................................................................................................................... 13 3.7. MOTIVATION DES FEMELLES .................................................................................................................... 14
4. DISCUSSION ................................................................................................ 15
4.1. COMPARAISON DES TRAITS D ’HISTOIRE DE VIE ENTRE POPULATIONS ...................................................15 4.2. REFLEXION SUR LES COMPROMIS EVOLUTIFS AU SEIN DES PO PULATIONS ETUDIEES ............................ 20 4.3. CONCLUSIONS ET PERSPECTIVES............................................................................................................. 23
REFERENCES BIBLIOGRAPHIQUES........................................................ 24

1
1. Introduction
Tout organisme est caractérisé par un ensemble de traits étroitement liés à sa reproduction
et sa survie, appelés traits d’histoire de vie (taille et âge à la maturité sexuelle, taux de
croissance, taux de mortalité, longévité) (Stearns, 2000). L’évolution de ces traits est sous
sélection de facteurs intrinsèques (génétique, physiologie, morphologie) et extrinsèques
(milieu de vie) aux organismes, qui leur imposent ainsi des contraintes. Ces contraintes vont
se traduire par des compromis évolutifs entre les différents traits d’histoire de vie, l’ensemble
des traits ne pouvant être maximisé simultanément (Stearns, 2000 ; Le Lann, 2009). Un
compromis apparaît lorsque l’augmentation de la valeur d’un trait se fait au détriment d’un
autre trait (Roff, 2006). La relation négative entre reproduction et survie (compromis
fécondité-longévité) illustre bien ce concept, l’augmentation des ressources allouées à la
reproduction se faisant au détriment de l’allocation à la survie, comme cela a été démontré
chez l’Hyménoptère parasitoïde Asobara tabida (Ellers et al., 2000). La répartition des
ressources entre les différents traits influence fortement la fitness des organismes, c'est-à-dire
leur succès reproducteur. L’allocation optimale des ressources dans les traits liés par des
compromis dépendant fortement de l’environnement, les organismes vont alors adopter des
stratégies différentes selon le milieu dans lequel ils évoluent, pour optimiser leur fitness
(Askari Seyahooei, 2010 ; Ellers & van Alphen, 1997). Ces stratégies ne se limitent toutefois
pas qu’aux traits d’histoire de vie, mais incluent également des décisions comportementales
ou des adaptations physiologiques par exemple (Stearns, 2000 ; Le Lann, 2009).
Les compromis évolutifs et les modèles d’optimisation ont fait l’objet de nombreuses
études, notamment chez les insectes parasitoïdes. Ces organismes se développent sur ou dans
un autre organisme hôte. Ils en tirent leur subsistance et le tuent comme résultat direct ou
indirect de leur développement. Les adultes mènent une vie libre, au cours de laquelle les
femelles recherchent activement les hôtes, et seuls les stades immatures se développent en
parasites (Eggleton & Gaston, 1990). De par le lien direct qui existe entre leurs décisions de
ponte et leur fitness, les parasitoïdes représentent ainsi un modèle approprié pour l’étude de
ces questions. Ainsi, les conditions abiotiques telles que la température, le régime de
précipitations ou encore l’humidité, et les conditions biotiques telles que la densité des hôtes
ou la compétition peuvent influencer fortement la reproduction et la survie des parasitoïdes.
Certaines études se sont ainsi intéressées à l’évolution des traits d’histoire de vie en fonction

2
des facteurs abiotiques, d’autres en fonction des facteurs biotiques, mais il est possible que
cette évolution soit le fait de la combinaison des deux.
Dans cette optique, la communauté de parasitoïdes de la vallée du Rhône représente un
système d’étude intéressant. Cette communauté est essentiellement composée de deux
espèces de parasitoïdes de larves de drosophiles: Leptopilina heterotoma et Asobara tabida.
Toutefois, elle est progressivement envahie par une autre espèce d’origine méditerranéenne,
Leptopilina boulardi, dont l’aire de répartition progresse vers le nord suite au réchauffement
climatique. Alors que L. heterotoma prédomine au nord de la vallée, son abondance diminue
dans le sud où elle est confrontée à L. boulardi. En effet, cette dernière parasite les mêmes
espèces hôtes et s’avère être un compétiteur supérieur, gagnant systématiquement la
compétition larvaire (Allemand et al., 1999 ; Fleury et al., 1999).
Dans le cadre du projet ANR CLIMEVOL, cette étude consiste à comparer plusieurs traits
d’histoire de vie dans des populations de Leptopilina heterotoma originaires du nord de la
vallée du Rhône où L. boulardi est absente, et des populations du sud où L. boulardi est
installée depuis une trentaine d’années. Toutefois, la vallée du Rhône s’étendant sur plus de
300 kilomètres, il existe un gradient climatique du nord au sud caractérisé par une
augmentation des températures, dont il faut également tenir compte. De par les interactions
compétitives (Allemand et al., 1999) auxquelles s’ajoutent des variations climatiques, cette
communauté est particulièrement adaptée pour investiguer les questions d’évolution des traits
d’histoire de vie chez les parasitoïdes, et a ainsi déjà fait l’objet de plusieurs études. Celle de
Ris (2003), basée sur la comparaison de populations issues du nord et du sud de la vallée, a
montré que les populations du sud présentaient à la fois une fécondité et une longévité plus
élevées que celles du nord, ce qui n’est pas conforme aux prédictions du compromis classique
entre fécondité et longévité. Ce résultat souligne ainsi l’intérêt de mesurer un plus grand
nombre de traits d’histoire de vie, et de considérer d’autres compromis, comme celui entre
reproduction et dispersion.
D’après la littérature et les connaissances acquises par une équipe lyonnaise qui étudie
cette communauté depuis plusieurs années, il est possible d’émettre différentes hypothèses
concernant ces populations, liées soit à la compétition, soit aux variations climatiques.
Au sud de la vallée du Rhône, la compétition devrait sélectionner des femelles présentant
une fécondité élevée, afin de compenser la mortalité de leurs descendants associée à la
présence de L. boulardi. En effet, selon l’hypothèse formulée par Price (1974), les organismes

3
souffrant d’une forte mortalité juvénile devraient effectivement exprimer une forte fécondité.
De plus, ces femelles pourraient disperser leurs œufs à travers plusieurs patchs, adoptant ainsi
une stratégie de bet-hedging qui réduirait encore le risque de perdre tous leurs descendants si
le patch qu’elles ont exploité est ensuite parasité par une femelle L. boulardi. Cette stratégie
conduirait les femelles du sud à passer moins de temps dans chaque patch mais à en visiter un
plus grand nombre au cours de leur vie, nécessitant une activité, un taux de métabolisme et/ou
une capacité de dispersion supérieurs. D’après Waage (1979), la durée de résidence d’une
femelle dans un patch serait en partie déterminée par sa motivation, basée sur la quantité de
kairomones provenant de l’hôte qu’elle perçoit lors de son entrée dans le patch. Une
motivation initiale, et donc une durée de résidence faibles, sont attendues dans les populations
du sud. Ces dernières devraient également être caractérisées par une charge alaire plus faible
que chez les populations du nord pour réaliser cette dispersion entre les patchs.
Concernant l’influence des facteurs abiotiques, il a été montré chez de nombreux
organismes que les individus exposés à des températures basses se développent plus
lentement mais atteignent une taille adulte supérieure que ceux exposés à des températures
plus élevées (Atkinson & Sibly, 1997 ; Angilleta & Dunham, 2003). De ce fait, une durée de
développement et une taille supérieures sont attendues chez les femelles du nord de la vallée
du Rhône. Les milieux froids sélectionneraient une charge alaire réduite chez les drosophiles
(Gilchrist, 2004), ce qui pourrait également être retrouvé chez les parasitoïdes mais qui
s’opposerait alors aux prédictions relatives à la compétition. En revanche, le métabolisme et
l’activité augmentant généralement avec la température chez les ectothermes (May, 1979 ;
Mellanby, 1939), un taux métabolique et une activité locomotrice plus intenses sont donc
attendus chez les femelles du sud.
En prenant en compte les prédictions relatives à la compétition et au climat, les femelles
du sud devraient avoir une fécondité, un taux de métabolisme et une activité plus élevés que
celles du nord. Toutefois, une relation négative entre le nombre d’œufs produits et l’espérance
de vie a été montrée chez une autre espèce de parasitoïde, A. tabida (Ellers et al., 2000), et
l’augmentation du métabolisme et de l’activité accélère la consommation des ressources,
réduisant alors la durée de vie. Les femelles du sud devraient donc exprimer une longévité
plus faible que celles du nord. Elles présenteraient également un taux de lipides inférieur. En
effet, la longévité et les réserves lipidiques semblent liées par une corrélation positive chez les
parasitoïdes (Jervis et al., 2008). De plus, ces réserves lipidiques seraient aussi liées
négativement au nombre d’œufs (Ellers & van Alphen, 1997), renforçant encore les
hypothèses précédemment émises pour ces différents traits.

4
Afin de mettre en évidence les effets de la compétition et du climat sur les traits d’histoire
de vie et les comportements d’exploitation des patchs chez Leptopilina heterotoma, et les
variations dans les compromis évolutifs qui en découleraient, de nombreuses mesures
(biométrie, développement, métabolisme, activité, taux de lipides, fécondité, longévité et
motivation) ont été réalisées sur des femelles issues de populations du nord et du sud de la
vallée du Rhône, en collaboration avec une équipe du laboratoire biométrie et biologie
évolutive à Lyon, également impliquée dans le projet ANR.
2. Matériels et Méthodes 2.1. Matériel biologique
2.1.1 Biologie des espèces
Leptopilina heterotoma est un parasitoïde généraliste largement réparti en Europe, qui
s’attaque aux larves de plusieurs espèces de drosophiles frugivores. Les femelles parasitent
majoritairement les deux premiers stades larvaires de leurs
hôtes, dans lesquels elles déposent généralement un seul
œuf. La larve-hôte poursuit ensuite son développement
jusqu’au stade pupal et la larve du parasitoïde, qui s’était
jusqu’alors développée à l’intérieur de cette larve-hôte,
devient ectoparasite tout en restant à l’intérieur du puparium
de l’hôte. Le parasitoïde en développement consomme les
restes de l’hôte puis se métamorphose à son tour, émergeant
alors du puparium au stade adulte (Prévost et al., 1989 ; Ris
et al., 2004 ; Fleury et al., 2004).
Dans la vallée du Rhône, L. heterotoma se développe
majoritairement dans D. melanogaster et D. simulans, qui
constituent les principales espèces hôtes de cette région.
2.1.2 Origine des espèces
Les populations de parasitoïdes de l’espèce L.
heterotoma utilisées dans nos expériences ont été fondées à
partir d’individus capturés en 2009 sur 4 sites différents Figure 1 : Carte de la vallée du Rhône et des sites de captures
Uchizy Igé
Eyguières
Avignon

5
(figure 1) : Uchizy et Igé au nord de la vallée du Rhône, où L. boulardi est absente, Eyguières
et Avignon au sud, où L. boulardi est présente. Au nord comme au sud, les sites
d’échantillonnage ont été sélectionnés pour leur proximité géographique, permettant ainsi la
répétition des mesures. Chaque population a été constituée à partir d’une trentaine de
femelles.
Les drosophiles de l’espèce D. melanogaster, utilisées comme hôtes au laboratoire, sont
issues d’une population initialement capturée à Sainte Foy-les Lyon, et élevée en laboratoire
depuis 1994.
2.1.3. Elevage
Afin d’obtenir des larves de drosophiles de stade L2 pour les expériences, une
cinquantaine de drosophiles adultes est introduite dans un pot en verre (h = 11,8 cm, ∅ = 5,2
cm) contenant un milieu nutritif eau-agar-sucre-levure-kalmus-nipagine, sur lequel est déposé
une boule de levure vivante qui constitue un milieu de ponte. Le pot est alors placé dans un
incubateur à 25°C sous photopériode LD 16 :8, et les drosophiles sont retirées après 2 heures
de ponte. Les larves sont ensuite récupérées 48 heures plus tard.
Afin d’obtenir des parasitoïdes pour les mesures de traits d’histoire de vie, 100 larves de
D. melanogaster sont déposées dans un tube (h = 9,5 cm, ∅ = 2,3 cm) contenant un milieu
axénique eau-levure-maïs-agar-nipagine, et une femelle L. heterotoma y est également
introduite. Le tube est placé dans un incubateur à 25°C sous photopériode LD 16 :8, et la
femelle est retirée au bout de 24 heures. Les mouches émergent après 7 à 8 jours de
développement, et sont alors prélevées puis dénombrées. L’émergence des parasitoïdes
commence une dizaine de jours plus tard et s’étale davantage dans le temps, elle est alors
vérifiée quotidiennement. Les individus retrouvés en début de matinée ont été conservés à
25°C sous photopériode LD 16:8, dans des pots en verre (h = 8,5 cm, ∅ = 4,5 cm) contenant
un milieu eau-agar-nipagine et du miel. Une partie d’entre eux a été utilisée pour les mesures
de métabolisme 24 heures plus tard, et l’autre pour les mesures d’activité 5 jours plus tard.
Les individus ayant émergé au cours de la matinée ont été congelés à l’azote liquide dans
l’heure qui a suivi leur émergence, puis utilisés ensuite pour les mesures de taux de lipides.
Afin d’obtenir le nombre de femelles de parasitoïdes suffisant pour ces différentes mesures,
une quinzaine de tubes a ainsi été constitué pour chaque population. Toutefois, très peu
d’individus pour la population d’Igé ont été obtenus, et les mesures n’ont finalement pas été

6
réalisées pour cette population à Rennes. La même méthode d’élevage a été utilisée à Lyon
afin de pouvoir comparer et intégrer leurs résultats aux nôtres.
Les mesures comportementales n’étant réalisées qu’à Rennes, il n’était pas nécessaire de
suivre le même protocole d’élevage que pour les traits d’histoire de vie. Une méthode plus
simple et plus rapide a ainsi permis de constituer un élevage de parasitoïdes spécifiquement
pour l’expérience d’exploitation des patchs, en se restreignant à la population d’Uchizy pour
le nord et à celle d’Eyguières pour le sud. Une dizaine de parasitoïdes mâles et femelles sont
placés pendant 24 heures dans un pot en verre (h = 8,5 cm, ∅ = 4,5 cm) dont le milieu eau-
agar-nipagine est recouvert par une fine couche de levure vivante, sur laquelle sont déposées
une centaine de larves. Le pot est alors placé à 25°C sous photopériode LD 16 :8. Les
mouches sont retirées au fur et à mesure de leur émergence, et les parasitoïdes sont ensuite
récupérés et conservés dans des pots avec un milieu eau-agar-nipagine et du miel.
2.2. Expériences
2.2.1. Paramètres de développement
La durée de développement minimale a été mesurée pour chaque tube d’élevage. Elle
correspond ici au nombre de jours écoulés entre le jour de la ponte et le jour où les premiers
individus émergent, et a été calculée séparément pour les mâles et les femelles. Pour chaque
tube, le succès parasitaire a été calculé selon cette formule : (nombre total de parasitoïdes
émergés / (nombre de drosophiles dans tube témoin (T) – nombre de drosophiles émergées).
Le degré d’infestation a quant à lui été déterminé à partir de la formule : ((nombre de
drosophiles T – nombre de drosophiles émergées) / nombre de drosophiles T).
2.2.2 Mesures biométriques
Trois types de mesures ont été réalisées sur au moins 20 femelles pour chaque population.
La masse fraîche a été déterminée chez des femelles congelées à l’émergence, grâce à une
balance Mettler Toledo XP2U de précision ± 0,1 µg. Le tibia postérieur gauche et la surface
de l’aile gauche ont été photographiés à l’aide d’une caméra JVC KY-F reliée à une loupe
binoculaire (0,5x6,3) Olympus SZ-CTV, puis mesurés avec le logiciel Image J. La longueur
du tibia restant constante au cours de la vie des insectes, elle représente ainsi une mesure
fiable pour évaluer leur taille (Nicol, 1999). La surface de l’aile permet quant à elle de
mesurer la charge alaire, qui correspond au rapport de la masse fraîche sur la surface alaire.

7
Les individus présentant une charge alaire faible possèderaient de bonnes capacités de
dispersion (Kolliker-Ott, 2003).
2.2.3. Mesure du taux de métabolisme
Pour chaque population, le taux de métabolisme a été mesuré sur 26 à 30 femelles nourries
et âgées de 24h. Ce taux est assimilé à la quantité de CO2 produite par heure, et est mesuré
grâce à un respiromètre (Sable Systems International). Chaque femelle est placée
individuellement dans un tube en verre (h = 6 cm, ∅ = 2 cm), l’appareillage permettant de
tester jusqu’à 14 individus simultanément. Une pompe capte l’air extérieur qui, après passage
dans une colonne de Drierite-Ascarite pour éliminer le CO2 et l’eau, est envoyé dans les
différents tubes avec un flux constant (Mass Flow Controller MFC-2). L’air de chaque tube
est ensuite expulsé, asséché grâce à du perchlorate de magnésium, puis envoyé dans un
analyseur à infrarouge de CO2 (CA-10A Carbon Dioxide Analyser) pour mesurer la quantité
de CO2 émise par chaque individu. Au cours de l’expérience, la température est maintenue à
25°C et cinq cycles de 1h30 chacun sont enregistrés. Les données issues du premier cycle
n’ont pas été prises en compte dans l’analyse des données. Le logiciel Expedata (Sable
Systems International) permet ensuite de convertir les données en millilitres de CO2 émis par
heure. A la fin de l’expérience, les femelles sont congelées à l’azote liquide, puis pesées à
l’aide d’une balance Mettler Toledo XP2U ± 0,1 µg pour obtenir leur masse fraîche. La masse
étant considérée comme le facteur endogène le plus déterminant pour le taux métabolique,
elle est généralement prise en considération dans les analyses (Nespolo et al., 2008).
2.2.4. Mesure des taux de lipides
Pour chaque population, le taux de lipides a été mesuré sur 22 à 26 femelles congelées à
l’azote liquide une heure maximum après leur émergence, et qui ont ensuite été pesées puis
conservées à -18°C jusqu’à l’extraction. Chaque femelle est broyée, à l’aide d’un pilon, dans
un tube eppendorf contenant 300 µl de méthanol. Après centrifugation pendant 15 minutes, à
1500 tours/min et à 4°C, 150 µl de chloroforme et 60 µl de sulfate de sodium sont ensuite
ajoutés dans chaque tube (le sulfate de sodium permettant la précipitation des protéines qui se
retrouvent dans le culot avec la cuticule et le glycogène). Après une autre centrifugation, le
surnageant de chaque tube est prélevé puis transvasé dans un nouveau tube, qui est conservé
dans la glace (c'est-à-dire le mélange chloroforme-méthanol qui contient les lipides et les
glucides). Les tubes ouverts sont alors placés dans un bain à sec à 90°C, jusqu’à évaporation à
sec des lipides. Après l’ajout de 40 µl d’acide sulfurique, les tubes fermés sont à nouveau

8
placés dans le bain à sec pendant 2 minutes, puis refroidis dans la glace pendant 5 minutes.
Enfin, 960 µl du réactif vanilline sont ajoutés dans les tubes. Le contenu de chaque tube est
agité puis analysé au spectrophotomètre. Les valeurs de densités optiques ainsi obtenues ont
permis de déterminer les quantités de lipides pour chaque femelle, à partir d’une courbe étalon
réalisée préalablement.
2.2.5. Mesure de l’activité locomotrice
Pour chaque population, l’activité locomotrice a été mesurée sur 30 à 31 femelles nourries
et âgées de 5 jours. Chaque femelle est placée individuellement dans une boîte de Pétri (∅ =
3,7 cm), et est filmée pendant 12 heures à l’aide d’une caméra Panasonic® CCTV. Le
dispositif expérimental est placé dans une pièce sans fenêtre, sous lumière blanche artificielle
et à la température de 25±1°C. La caméra permet de filmer jusqu’à 6 individus
simultanément, et est reliée à un ordinateur comportant le logiciel Ethovision (Noldus
Information Technology 1997), qui détecte les individus grâce au contraste entre leur couleur
sombre et le support blanc. Le logiciel enregistre la position de chaque individu 5 fois par
seconde, et permet ainsi de mesurer la distance totale parcourue en cm/s.
2.2.6. Autres mesures
La longueur du tibia, la durée de développement, le succès parasitaire et le degré
d’infestations ont également été mesurés à Lyon. De plus, des mesures de fécondité, de
longévité et de taille d’œufs y ont été réalisées afin de compléter les données (tableau 1).
L’ensemble de ces mesures a été effectué sur des femelles issues des mêmes populations que
celles de Rennes, mais aussi sur des femelles originaires d’Igé.
Tableau 1: Détail des mesures réalisées à Lyon.
Fécondité Longévité Biométrie Développement à l'émergence avec nourriture avec ponte longueur du tibia durée de développement
totale taille des œufs succès parasitaire degré d'infestation
2.2.7. Motivation des femelles
La motivation initiale a été testée chez 15 femelles nourries, accouplées et âgées de 5 jours
par population et par modalité. Pour s’assurer que les femelles soient expérimentées, chacune
d’entre elles a été en contact pendant 2 heures avec 16 larves de D. melanogaster de stade L2,

9
Figure 2 : Schéma du dispositif expérimental. A gauche : la veille de l’expérience, à droite : le jour de l’expérience après retrait de l’anneau et des larves.
la veille de l’expérience. Elles ont ensuite été isolées dans une boîte de Pétri (∅ = 5,5 cm)
avec un coton imbibé de miel jusqu’au lendemain matin.
Le dispositif expérimental est constitué d’une boîte de Pétri (∅ = 5,5 cm) dont le fond est
recouvert par une fine couche d’un milieu eau-agar-nipagine. La veille d’une observation, en
début d’après-midi, un anneau en caoutchouc (∅ = 2 cm) est placé au centre de cette boîte, et
200 ml de levure vivante liquide sont déposés sur la zone délimitée par l’anneau,
correspondant à un patch. Selon la modalité testée, 4 (patch pauvre) ou 16 (patch riche) larves
de D. melanogaster de stade L2 sont ensuite déposées sur la levure, et le dispositif est placé à
25°C. Le lendemain matin, soit 18 heures plus tard, l’anneau et les larves sont alors retirés
afin qu’il ne reste que les kairomones larvaires dans le patch.
L’ensemble des observations a été réalisé le matin, les femelles étant davantage motivées
en début de journée (observation personnelle). L’expérience débute lorsqu’une femelle est
introduite dans le dispositif expérimental, c'est-à-dire lorsque le couvercle de la boîte de Pétri
sous lequel elle se trouve est posé sur le dispositif expérimental, rendant ainsi le patch
accessible pour la femelle. Les entrées et sorties du patch, les phases de déplacements, de
déplacements couplés à du sondage par l’ovipositeur, d’immobilité, et les comportements de
nettoyage sont enregistrés au fur et à mesure grâce au logiciel The Observer. L’expérience se
termine lorsque la femelle passe plus de 60 secondes en dehors du patch sur l’agar, ou
lorsqu’elle monte sur le bord de la boîte de Pétri ou sur le couvercle. L’expérience est
également terminée si la femelle n’est pas entrée dans le patch au bout de 600 secondes.

10
2.3. Analyses statistiques
Pour les paramètres de développement et les mesures biométriques, la normalité et
l’homoscédasticité des données ont préalablement été vérifiées (test de Shapiro-Wallis et test
de Bartlett), afin de déterminer la catégorie de test à appliquer (paramétrique ou non
paramétrique). La durée de développement minimale et le succès parasitaire ont été comparés
entre populations par des tests de Kruskal-Wallis, puis les populations ont ensuite été
comparées deux à deux par des tests de Wilcoxon lorsque le premier test était significatif. Le
degré d’infestation a quant à lui été analysé grâce à une ANOVA. La masse fraîche, la
longueur du tibia gauche et la charge alaire des femelles ont été comparées entre populations à
l’aide d’une ANOVA. Dans les cas d’un test significatif, un test post-hoc de Tukey a été
réalisé ensuite afin de déterminer le sens de la différence.
Des modèles linéaires généraux ont permis d’analyser l’effet de la population sur le taux
de lipides et sur le taux de métabolisme des femelles, avec la masse en covariable et après
avoir appliqué les tests de Shapiro et Bartlett. La vérification de la normalité et de
l’homoscédasticité des résidus a ensuite permis de valider les modèles, et la valeur du critère
d’Akaike de déterminer le modèle le plus parcimonieux parmi les différents modèles testés
(additifs ou interactifs). Un test de corrélation de Pearson a également été réalisé entre la
variable testée et la masse lorsque cette dernière avait un effet significatif sur la variable, afin
de déterminer le sens de la relation. L’activité locomotrice a quant à elle été comparée entre
les populations à l’aide d’un test de Kruskal-Wallis.
Enfin, les données issues de l’expérience comportementale ont été analysées grâce à un
modèle de risques proportionnels, aussi appelé modèle de régression de Cox. Ce modèle
exprime le temps passé dans le patch en terme de probabilité par unité de temps que la femelle
quitte le patch, sachant qu’elle y est toujours (Wajnberg, 2006 ; Martel et al., 2008). Il a pour
équation :
où h(t) correspond à la fonction du taux de risque qui décrit la tendance de la femelle à quitter
le patch, h0(t) à la fonction de risque de base, t au temps écoulé depuis que la femelle est
entrée dans le patch, et βi aux coefficients donnant la contribution relative des p variables.
L’ensemble des analyses statistiques a été réalisé avec le logiciel R 2.8.0.
11
Avignon n = 24
Eyguières n = 22
Uchizy n = 30
NS NS *
B
0.0
0.1
0.2
0.3
0.4
0.5
0.6
Cha
rge
alai
re (m
g/m
m²)
Avignon n = 24
Eyguières n = 22
Uchizy n = 30
NS
A
0.0
0.1
0.2
0.3
0.4
0.5
0.6
Mas
se fr
aîch
e (m
g)
3. Résultats
3.1. Paramètres de développement
L’analyse des résultats obtenus à Rennes ne montre pas de différence entre populations
pour la durée de développement minimale (mâles : Kruskal-Wallis χ² = 4,640 ; ddl = 2 ; p =
0,098 / femelles : Kruskal-Wallis χ² = 0,950 ; ddl = 2 ; p = 0,622), ni pour le degré
d’infestation (ANOVA ; F = 1,309 ; ddl = 2 ; p = 0,284). En revanche, il existe une différence
pour le succès parasitaire (Kruskal-Wallis χ² = 6,532 ; ddl = 2 ; p = 0,038), le succès
parasitaire d’Avignon étant significativement supérieur à celui d’Eyguières (Wilcoxon ; w =
107 ; p = 0,002).
3.2. Mesures biométriques
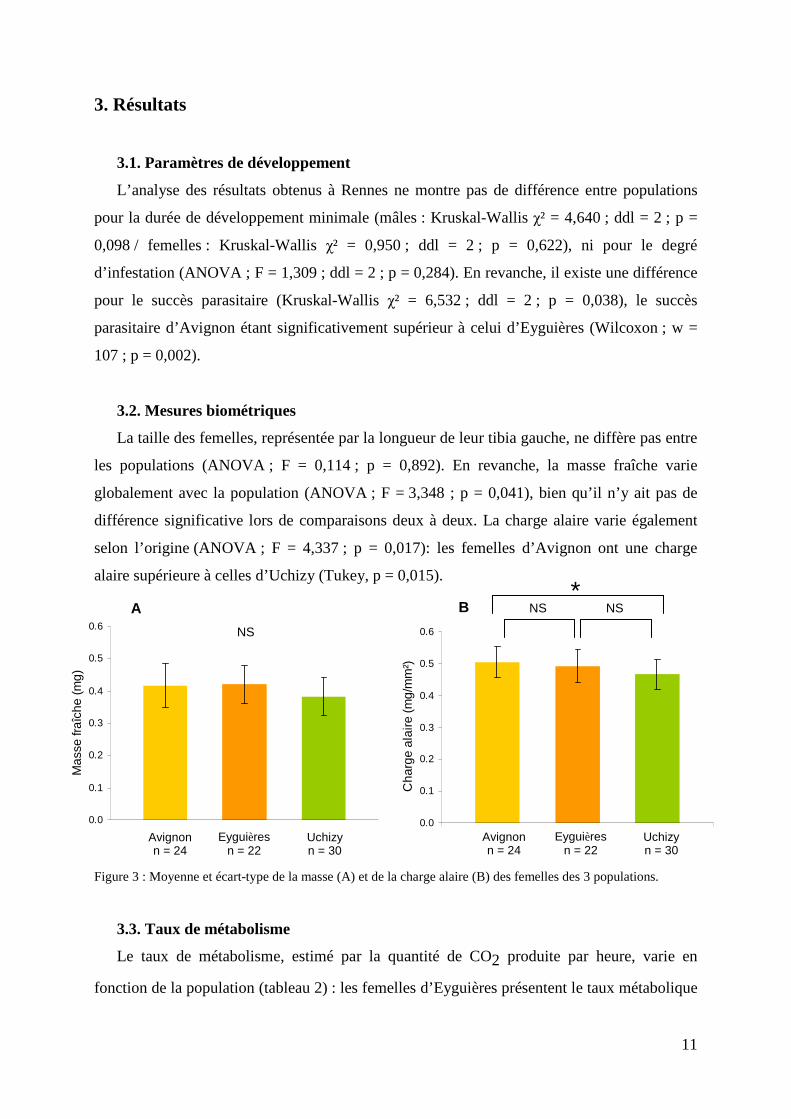
La taille des femelles, représentée par la longueur de leur tibia gauche, ne diffère pas entre
les populations (ANOVA ; F = 0,114 ; p = 0,892). En revanche, la masse fraîche varie
globalement avec la population (ANOVA ; F = 3,348 ; p = 0,041), bien qu’il n’y ait pas de
différence significative lors de comparaisons deux à deux. La charge alaire varie également
selon l’origine (ANOVA ; F = 4,337 ; p = 0,017): les femelles d’Avignon ont une charge
alaire supérieure à celles d’Uchizy (Tukey, p = 0,015).
Figure 3 : Moyenne et écart-type de la masse (A) et de la charge alaire (B) des femelles des 3 populations.
3.3. Taux de métabolisme
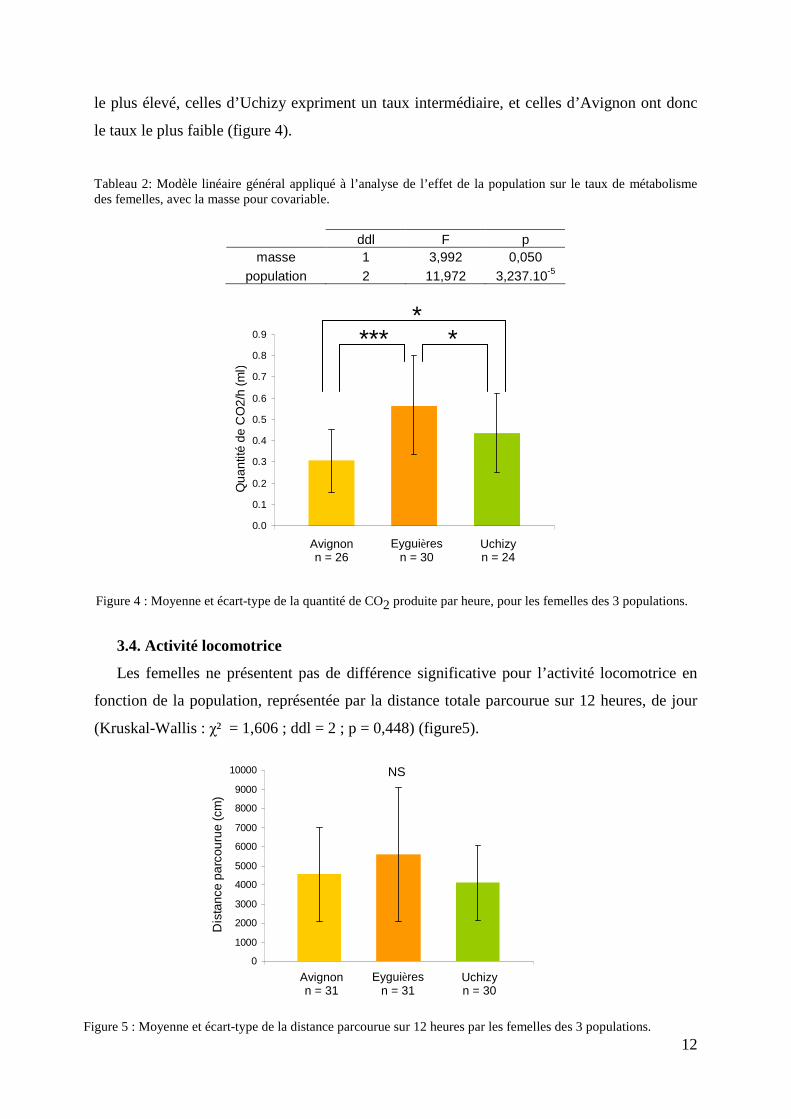
Le taux de métabolisme, estimé par la quantité de CO2 produite par heure, varie en
fonction de la population (tableau 2) : les femelles d’Eyguières présentent le taux métabolique
12
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
Dis
tanc
e pa
rcou
rue
(cm
)
Avignon n = 31
Eyguières n = 31
Uchizy n = 30
NS
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Qua
ntité
de
CO
2/h
(ml)
Avignon n = 26
Eyguières n = 30
Uchizy n = 24
*** * *
le plus élevé, celles d’Uchizy expriment un taux intermédiaire, et celles d’Avignon ont donc
le taux le plus faible (figure 4).
Tableau 2: Modèle linéaire général appliqué à l’analyse de l’effet de la population sur le taux de métabolisme des femelles, avec la masse pour covariable.
3.4. Activité locomotrice
Les femelles ne présentent pas de différence significative pour l’activité locomotrice en
fonction de la population, représentée par la distance totale parcourue sur 12 heures, de jour
(Kruskal-Wallis : χ² = 1,606 ; ddl = 2 ; p = 0,448) (figure5).
ddl F p masse 1 3,992 0,050
population 2 11,972 3,237.10-5
Figure 4 : Moyenne et écart-type de la quantité de CO2 produite par heure, pour les femelles des 3 populations.
Figure 5 : Moyenne et écart-type de la distance parcourue sur 12 heures par les femelles des 3 populations.
13
0
2
4
6
8
10
12
14
Tau
x de
lipi
des
(µl)
Avignon n = 23
Eyguières n = 22
Uchizy n = 26
NS
3.5. Taux de lipides
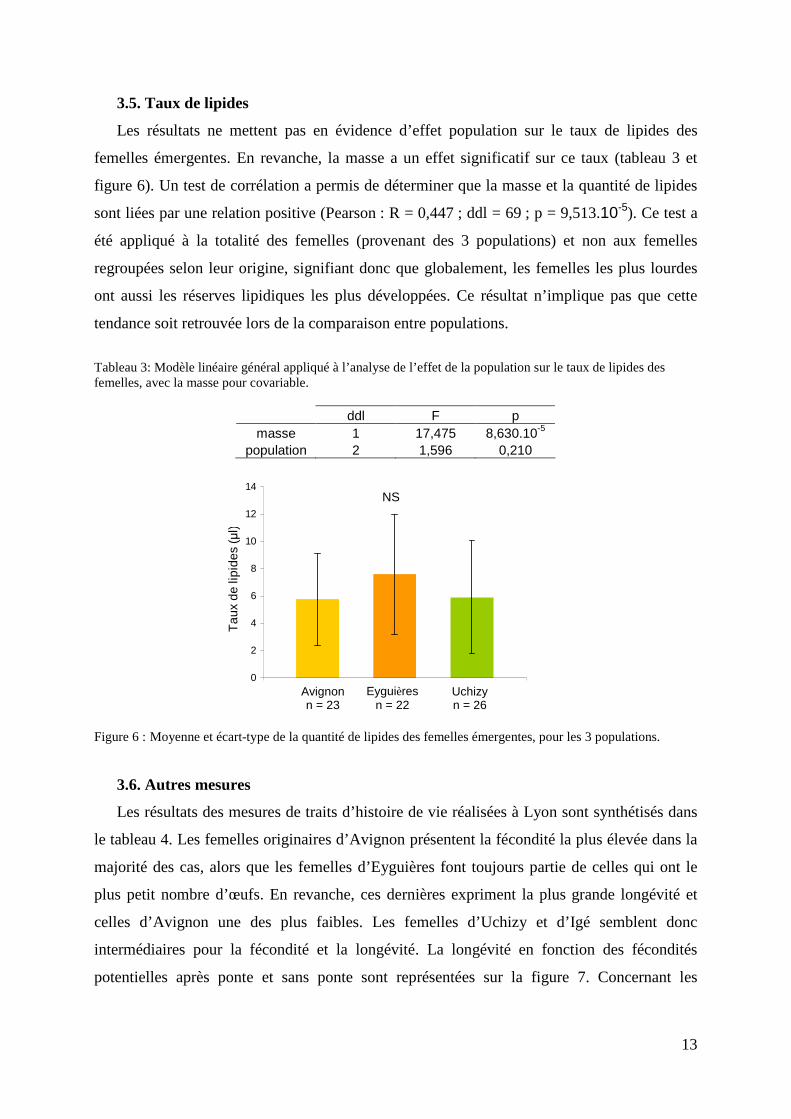
Les résultats ne mettent pas en évidence d’effet population sur le taux de lipides des
femelles émergentes. En revanche, la masse a un effet significatif sur ce taux (tableau 3 et
figure 6). Un test de corrélation a permis de déterminer que la masse et la quantité de lipides
sont liées par une relation positive (Pearson : R = 0,447 ; ddl = 69 ; p = 9,513.10-5). Ce test a
été appliqué à la totalité des femelles (provenant des 3 populations) et non aux femelles
regroupées selon leur origine, signifiant donc que globalement, les femelles les plus lourdes
ont aussi les réserves lipidiques les plus développées. Ce résultat n’implique pas que cette
tendance soit retrouvée lors de la comparaison entre populations.
Tableau 3: Modèle linéaire général appliqué à l’analyse de l’effet de la population sur le taux de lipides des femelles, avec la masse pour covariable.
ddl F p masse 1 17,475 8,630.10-5
population 2 1,596 0,210
Figure 6 : Moyenne et écart-type de la quantité de lipides des femelles émergentes, pour les 3 populations.
3.6. Autres mesures
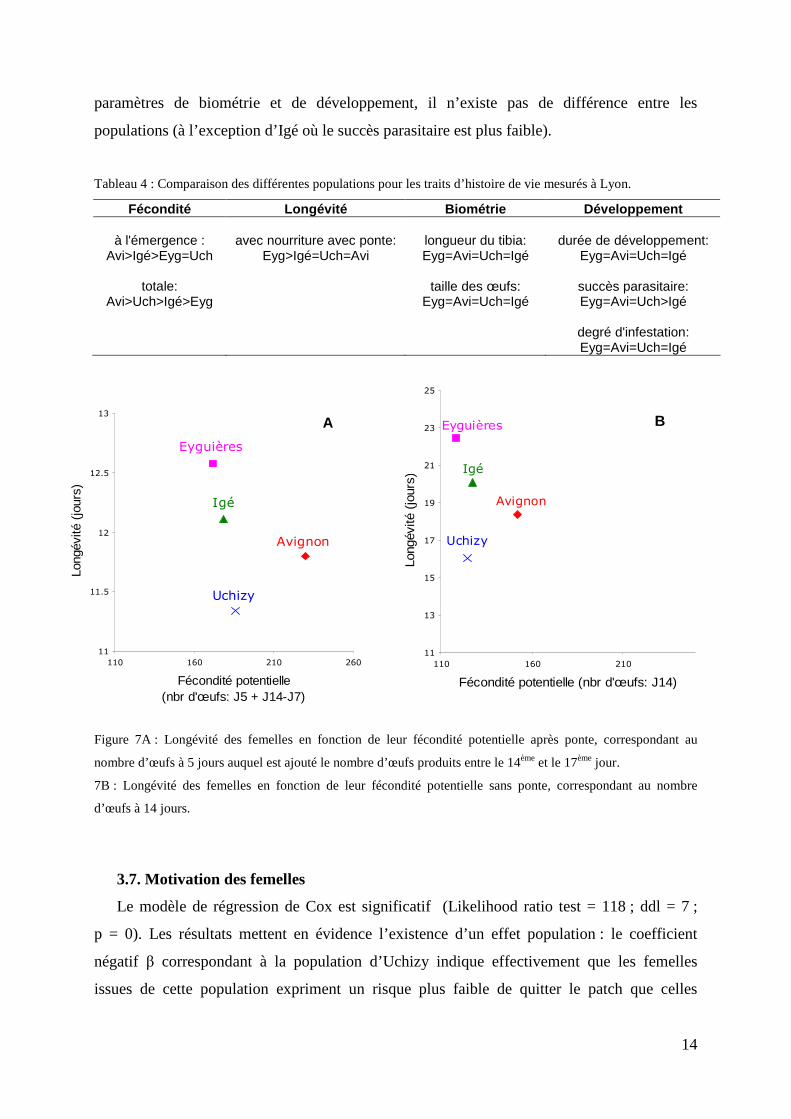
Les résultats des mesures de traits d’histoire de vie réalisées à Lyon sont synthétisés dans
le tableau 4. Les femelles originaires d’Avignon présentent la fécondité la plus élevée dans la
majorité des cas, alors que les femelles d’Eyguières font toujours partie de celles qui ont le
plus petit nombre d’œufs. En revanche, ces dernières expriment la plus grande longévité et
celles d’Avignon une des plus faibles. Les femelles d’Uchizy et d’Igé semblent donc
intermédiaires pour la fécondité et la longévité. La longévité en fonction des fécondités
potentielles après ponte et sans ponte sont représentées sur la figure 7. Concernant les
14
11
11.5
12
12.5
13
110 160 210 260
Fécondité potentielle (nbr d'œufs: J5 + J14-J7)
Long
évité
(jo
urs)
Eyguières
Igé
Uchizy
Avignon
A
11
13
15
17
19
21
23
25
110 160 210
Fécondité potentielle (nbr d'œufs: J14)
Long
évité
(jo
urs)
Uchizy
Eyguières
Igé
Avignon
B
paramètres de biométrie et de développement, il n’existe pas de différence entre les
populations (à l’exception d’Igé où le succès parasitaire est plus faible).
Tableau 4 : Comparaison des différentes populations pour les traits d’histoire de vie mesurés à Lyon.
Fécondité Longévité Biométrie Développement
à l'émergence : Avi>Igé>Eyg=Uch
avec nourriture avec ponte: Eyg>Igé=Uch=Avi
longueur du tibia: Eyg=Avi=Uch=Igé
durée de développement: Eyg=Avi=Uch=Igé
totale:
Avi>Uch>Igé>Eyg taille des œufs:
Eyg=Avi=Uch=Igé succès parasitaire: Eyg=Avi=Uch>Igé
degré d'infestation: Eyg=Avi=Uch=Igé
Figure 7A : Longévité des femelles en fonction de leur fécondité potentielle après ponte, correspondant au
nombre d’œufs à 5 jours auquel est ajouté le nombre d’œufs produits entre le 14ème et le 17ème jour.
7B : Longévité des femelles en fonction de leur fécondité potentielle sans ponte, correspondant au nombre
d’œufs à 14 jours.
3.7. Motivation des femelles
Le modèle de régression de Cox est significatif (Likelihood ratio test = 118 ; ddl = 7 ;
p = 0). Les résultats mettent en évidence l’existence d’un effet population : le coefficient
négatif β correspondant à la population d’Uchizy indique effectivement que les femelles
issues de cette population expriment un risque plus faible de quitter le patch que celles

15
provenant de la population d’Eyguières. De par le signe positif de son coefficient, le
mouvement qui correspond aux phases de déplacement sans sondage constitue le seul facteur
qui augmente le risque de quitter le patch (tableau 4).
Tableau 4: Coefficients de régression estimés (β), taux de risque (expβ), erreur standard (ES), et degré de significativité (p) pour la tendance des femelles à quitter le patch.
4. Discussion
Les résultats obtenus ne vont pas tous dans le sens des prédictions relatives aux effets de la
compétition sur l’évolution des traits d’histoire de vie chez Leptopilina heterotoma. En effet,
l’influence de cette compétition n’a pas pu être totalement séparée de celle exercée par le
climat, et les populations ne se différencient pas clairement selon l’opposition nord-sud. Dans
un premier temps, il est donc nécessaire de reprendre les traits d’histoire de vie un par un,
pour essayer de comprendre quels sont les facteurs et les pressions évolutives responsables de
la direction dans laquelle ils ont évolué au sein de chaque population. Ensuite, il est essentiel
de mettre ces traits d’histoire de vie en relation les uns avec les autres, pour faire ressortir les
compromis évolutifs qui pourraient les lier, et ainsi mettre en évidence les stratégies adoptées
par les différentes populations. Enfin, cette étude apporte de nouvelles connaissances sur la
communauté de parasitoïdes de la vallée du Rhône, et pourrait ainsi contribuer à la réflexion
concernant la pertinence du choix des traits d’histoire de vie à intégrer dans les études
comparatives de ce genre.
4.1. Comparaison des traits d’histoire de vie entre populations
Les résultats concernant la taille et la durée de développement ne correspondent pas aux
hypothèses de départ, puisqu’il n’existe pas de différences entre les populations pour ces deux
paramètres. Il a pourtant été montré que de nombreux ectothermes atteignent des tailles
β exp(β) ES z p Population Uchizy -0,792 0,453 0,298 -2,66 7,8.10-3
Kairomones 0,046 1,047 0,025 1,85 6,5.10-2 Mouvement 2,182 8,864 0,249 8,77 0
Sondage -0,163 0,850 0,047 -3,45 5,6.10-4 Immobilité -1,901 0,149 0,236 -8,04 8,9.10-16 Nettoyage -0,444 0,641 0,137 -3,24 1,2.10-3
Sortie du patch -1,806 0,164 0,257 -7,03 2,0.10-12

16
adultes supérieures lorsqu’ils sont élevés à des températures basses. De plus, une taille
supérieure a également été retrouvée chez les individus vivant à de hautes altitudes et
latitudes, où le climat est globalement plus froid (Atkinson & Sibly, 1997). Les écarts de
température entre le nord et le sud de la vallée du Rhône ne sont peut être pas suffisamment
importants pour induire de telles variations dans les populations de L. heterotoma étudiées ici.
La charge alaire des femelles d’Avignon n’est pas significativement différente de celle des
femelles d’Eyguières, mais est toutefois supérieure à celle des femelles d’Uchizy, qui auraient
donc la meilleure capacité de dispersion. Ce résultat contredit ainsi l’hypothèse d’une
meilleure capacité de dispersion des femelles du sud par rapport à celles du nord, hypothèse
reliée à la compétition avec L. boulardi. Néanmoins, une étude sur Drosophila subobscura a
mis en évidence que les individus vivant en milieu froid ont une charge alaire inférieure à
ceux vivant dans des milieux plus chauds (Gilchrist, 2004). Un phénomène similaire pourrait
alors se produire chez les parasitoïdes, et expliquer ainsi le sens des résultats.
A Rennes comme à Lyon, le degré d’infestation ne varie pas selon la population. Les
populations du nord et du sud exploitant les mêmes espèces hôtes, les caractéristiques de
développement et de résistance au parasitisme de ces dernières pourraient être similaires au
nord et au sud de la vallée, et expliquer ainsi l’absence de variation pour ce paramètre. En
revanche, le succès parasitaire d’Avignon est supérieur à celui des autres populations d’après
les résultats de Rennes, mais la comparaison avec ceux de Lyon n’est pas possible puisque les
analyses n’ont pas été réalisées sur le même nombre de populations. Une virulence plus forte
chez les individus d’Avignon pourrait expliquer le résultat obtenu à Rennes.
Concernant le taux de métabolisme, les résultats ne suivent qu’en partie les hypothèses. Il
était attendu que les femelles du sud aient un métabolisme plus élevé que celles du nord, ce
qui est effectivement retrouvé dans la population d’Eyguières, mais pas dans celle d’Avignon
dont le taux métabolique moyen est inférieur à celui de la population d’Uchizy. Le taux de
métabolisme augmentant avec la température chez les ectothermes (May, 1979), il est donc
surprenant que la population d’Eyguières ne suive pas la même tendance que celle d’Avignon.
Tout comme pour le métabolisme, il était attendu que les femelles du sud soient plus
actives que celles du nord. Cependant, aucune différence entre populations n’a été mise en
évidence pour l’activité locomotrice. L’hypothèse selon laquelle les femelles du sud auraient
recours à une stratégie de bet-hedging amène pourtant à penser qu’une dispersion des pontes à
travers plusieurs patchs nécessiterait une activité locomotrice plus intense, ou une charge
alaire réduite. De plus, ce résultat va également à l’encontre d’une précédente étude réalisée

17
chez la même espèce, où l’enregistrement du rythme d’activité avait permis de montrer que
des femelles originaires de Lyon exprimaient un taux d’activité plus faible que celles issues
d’Antibes, au sud de la vallée (Fleury et al., 1995). Mesurer l’évolution de l’activité
locomotrice au cours de la journée (heure par heure par exemple), plutôt que la mesurer
globalement sur 12 heures, permettrait peut-être de mettre en évidence des différences entre
populations que le protocole utilisé ici n’a pu faire ressortir.
Les populations étudiées ne se différencient pas non plus par leur taux de lipides. Ce
résultat impliquerait que les différentes populations extraient la même quantité de ressources
lipidiques de leurs hôtes, ce qui n’est pas surprenant puisqu’elles exploitent les mêmes
espèces hôtes. Il faut cependant noter que le taux lipidique mesuré ici correspond à la quantité
de lipides contenue dans la totalité du corps de l’insecte. Les lipides sont impliqués dans
différentes fonctions chez les insectes, pouvant jouer un rôle dans la survie, la dispersion ou
encore constituer une source d’énergie cruciale pour la production des œufs (Pélosse, 2008 ;
Ellers & van Alphen, 1997). L’idéal serait donc de distinguer la quantité de lipides contenue
dans les œufs de celle contenue dans le reste du corps, mais il est difficile de séparer les
lipides de ces deux compartiments sur un seul individu, les œufs constituant un volume trop
faible pour les analyses biochimiques. Une solution consisterait alors à combiner les œufs de
plusieurs femelles pour pouvoir évaluer la quantité de lipides moyenne allouée à la
reproduction par les individus de chaque population, mais l’analyse serait moins précise.
Les prédictions relatives aux impacts de la compétition et de la température sur la
fécondité entrent en opposition pour certains traits d’histoire de vie. En effet, alors que la
compétition liée à la présence de L. boulardi pourrait sélectionner une fécondité élevée chez
les femelles du sud, une grande taille est généralement synonyme d’une fécondité élevée chez
les insectes (Nylin & Gotthard, 1998). Cette dernière hypothèse inclurait donc que les
femelles du nord aient un nombre d’œufs plus important puisqu’elles seraient également plus
grandes à cause du climat. Les résultats n’ayant pas montré de différence de taille entre
populations, la relation entre taille et fécondité ne peut donc pas être appliquée aux
populations étudiées. Les femelles d’Uchizy présentent en réalité la fécondité à l’émergence
la plus faible, allant ainsi à l’encontre de l’hypothèse relative au climat, mais dans le sens de
celle relative à la compétition. La fécondité élevée des femelles d’Avignon (fécondité à
l’émergence comme fécondité totale) suggère quant à elle que la compétition puisse
influencer ce trait, mais la faible fécondité à l’émergence des femelles d’Eyguières va à
l’encontre de cette hypothèse. Les effets de la compétition et de la température sur la

18
fécondité ne semblent donc pas être aussi facilement identifiables, et pourraient être
partiellement masqués par la relation qui lie le nombre d’œufs à leur taille. En effet, un
nombre d’œufs plus important peut avoir pour conséquence un investissement par œuf plus
faible, avec pour conséquence des œufs plus petits. Cependant, l’absence de différence dans la
taille des œufs ne permet pas de vérifier l’existence de cette relation chez les populations de L.
heterotoma étudiées.
La longévité d’une femelle parasitoïde peut être influencée par les caractéristiques de ses
traits d’histoire de vie. Par exemple, une fécondité élevée, un important taux de métabolisme
et une forte activité sont généralement associés à une longévité courte (Ellers et al., 2000).
D’après cette relation, les femelles du sud devraient donc vivre moins longtemps que celles du
nord. Encore une fois, les résultats ne correspondent pas parfaitement aux prédictions. Les
femelles d’Eyguières présentent bien la longévité la plus forte, mais ce n’est pas le cas des
femelles d’Avignon, dont la longévité n’est pas différente de celle des femelles des
populations du nord. Considérer la relation entre la longévité et les autres traits pourrait
permettre de mieux comprendre ce phénomène, les traits d’histoire de vie n’étant pas
indépendants les uns des autres.
Les résultats de l’expérience sur l’exploitation des patchs sont relativement conformes aux
prédictions émises au départ. En effet, en exprimant un risque plus faible de quitter le patch
que les femelles d’Eyguières, les femelles d’Uchizy présenteraient une durée de résidence
dans le patch plus élevée. Lors d’une étude sur l’approvisionnement en hôtes chez le
parasitoïde Nemeritis canescens (1979), Waage a suggéré qu’une femelle entre dans un patch
avec un certain niveau de motivation pour rester dans ce patch, qui serait fonction de la
concentration en kairomones corrélée à la densité des hôtes. Suite à l’habituation progressive
de la femelle à cette concentration, le départ du patch serait alors provoqué par la diminution
de la motivation jusqu’à l’atteinte d’un seuil. La durée de résidence dans le patch serait donc
plus importante pour une concentration en kairomones plus élevée. Cependant, la motivation
initiale pourrait également être sous le contrôle d’autres facteurs, et varier entre populations
selon l’environnement, la compétition, les paramètres physiologiques ou encore l’histoire
évolutive des parasitoïdes. En reliant la durée de résidence au niveau de motivation, la
compétition pourrait avoir constitué une pression de sélection favorisant une faible motivation
initiale chez les femelles du sud, et être ainsi à l’origine de la tendance observée.
19
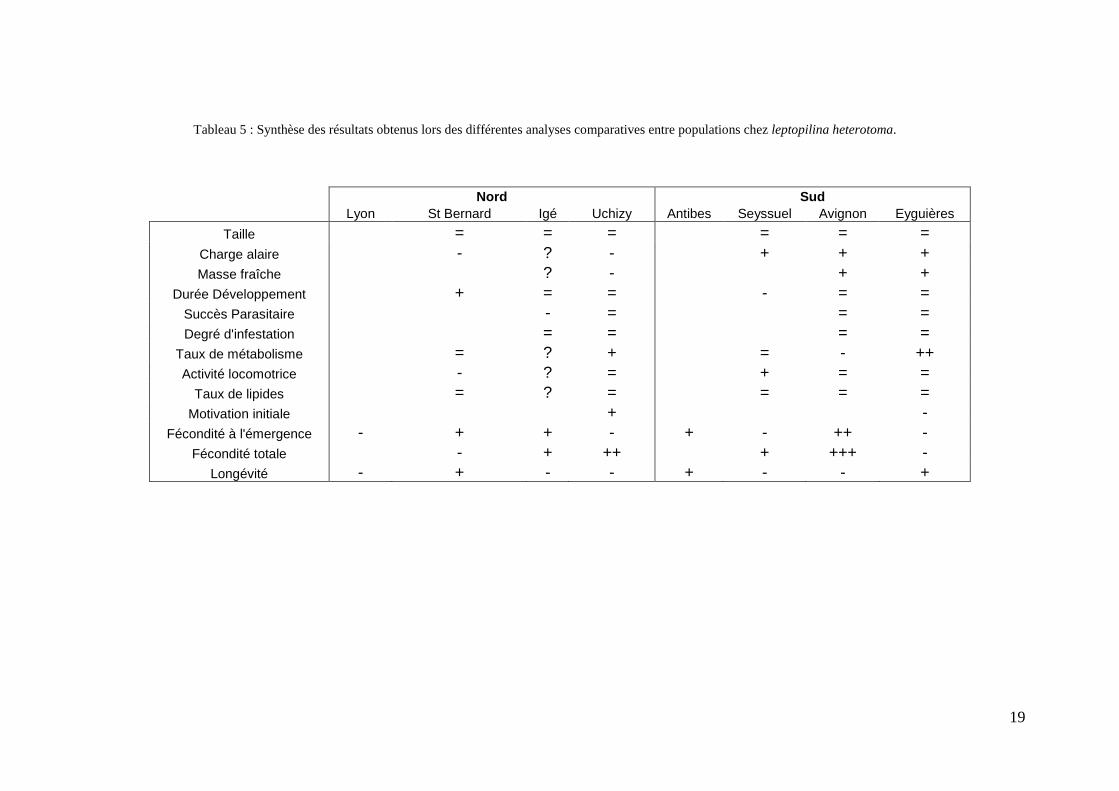
Tableau 5 : Synthèse des résultats obtenus lors des différentes analyses comparatives entre populations chez leptopilina heterotoma.
Nord Sud Lyon St Bernard Igé Uchizy Antibes Seyssuel Avignon Eyguières
Taille = = = = = = Charge alaire - ? - + + + Masse fraîche ? - + +
Durée Développement + = = - = = Succès Parasitaire - = = = Degré d'infestation = = = =
Taux de métabolisme = ? + = - ++ Activité locomotrice - ? = + = =
Taux de lipides = ? = = = = Motivation initiale + -
Fécondité à l'émergence - + + - + - ++ - Fécondité totale - + ++ + +++ -
Longévité - + - - + - - +

20
4.2. Réflexion sur les compromis évolutifs au sein des populations étudiées
Les traits d’histoire de vie n’étant pas indépendants les uns des autres, il est très probable
que des compromis évolutifs s’opèrent au sein des populations de Leptopilina heterotoma,
influencés par la compétition mais aussi par le climat. Etablir les relations existant entre traits
pourrait permettre de mieux comprendre les résultats obtenus.
L’analyse de ces résultats met en avant un contraste inattendu entre les deux populations
du sud, qui s’opposent pour plusieurs traits dont la fécondité et la longévité. Le compromis
liant la reproduction et la survie est aujourd’hui largement admis (Jervis et al., 2008 ; Roff,
1996 ; Stearns, 1989), et a été démontré chez de nombreux organismes. Chez le parasitoïde
Asobara tabida par exemple, ce compromis résulte du fait que le métabolisme somatique et la
production des œufs dépendent des mêmes réserves énergétiques, chaque œuf produit
diminuant ainsi la quantité d’énergie disponible pour la survie. Ainsi, les femelles qui
produisent un plus grand nombre d’œufs vivent moins longtemps (Ellers et al., 2000).
Malgré les contrastes observés dans cette étude, il est toutefois possible d’identifier les
stratégies adoptées par chaque population, et de mettre ainsi en évidence les éventuels
compromis évolutifs qui les sous-tendent. Alors que la population d’Avignon présente une
fécondité élevée au détriment de la longévité, celle d’Eyguières exprime le patron inverse.
Ces résultats sont particulièrement étonnants puisque des populations originaires de la même
zone géographique, et donc potentiellement soumises à des pressions de compétition et des
conditions climatiques similaires, présentent des tendances inversées. De plus, ces résultats
s’opposent à ceux de Ris (2003), qui avait montré une fécondité et une longévité supérieures
dans des populations de L. heterotoma d’Antibes, située au sud de la vallée où L. boulardi est
présente, par rapport à des populations de Lyon. Cette observation souligne encore le
caractère paradoxal des stratégies mises en évidence dans les populations étudiées. Toutefois,
les résultats de Ris sont eux-mêmes surprenants car les populations d’Antibes investissent
simultanément dans la reproduction et la survie, ne réalisant pas de compromis entre ces
traits. Au nord de la vallée, les deux populations ont une fécondité similaire (à l’émergence et
totale), mais la longévité de la population d’Igé est supérieure à celle d’Uchizy. Les femelles
d’Igé semblent avoir recours à une stratégie intermédiaire, avec une fécondité et une longévité
moyennes, alors que la population d’Uchizy semble être la seule à ne pas exprimer ce
compromis puisqu’elle est caractérisée par des valeurs faibles pour ces deux traits.
Bien que fécondité et longévité soient intimement liées, le coût de la reproduction ne se
répercute pas uniquement sur la survie mais affecte également d’autres traits. Les compromis

21
qui en découlent vont tout autant jouer un rôle dans l’élaboration des stratégies que les
organismes mettent en place pour optimiser leur fitness en fonction de l’environnement. Il
existe par exemple des compromis entre la reproduction actuelle et la reproduction future
(Stearns, 1989), entre la reproduction et la dispersion, ou encore entre le nombre et le volume
des œufs.
Les femelles d’Avignon présentent à la fois la fécondité la plus élevée mais aussi la charge
alaire la plus importante. De ce fait, cette population semble exprimer un compromis entre
reproduction et dispersion, l’investissement dans la fécondité se faisant au détriment des
capacités de dispersion. Le sens de ce compromis est ainsi conforme aux hypothèses relatives
au climat pour les populations du sud. Il ne l’est donc pas vis-à-vis des hypothèses relatives à
la compétition, qui suggèreraient une forte fécondité couplée à une bonne capacité à disperser.
Des contraintes intrinsèques aux parasitoïdes pourraient empêcher la maximisation simultanée
de ces deux traits, et être responsables du compromis justement mis en évidence. En revanche,
bien que les femelles d’Eyguières aient une fécondité plus faible que celles d’Avignon, leur
charge alaire n’est pas différente. Encore une fois, cette population se distingue en ne suivant
pas les prédictions, et aucun compromis ne semble se dégager entre ses traits. L’analyse
s’avère encore plus difficile pour les populations du nord de la vallée. En effet, l’absence de
données pour la majorité des traits d’histoire de vie chez les femelles d’Igé rend délicate la
comparaison entre les deux populations du nord. Les femelles d’Uchizy présentent toutefois
une fécondité et une charge alaire plus faibles que les femelles d’Avignon, suggérant qu’elles
réalisent aussi un compromis entre reproduction et dispersion, mais que ce compromis est
inversé. Tout comme pour Avignon, le compromis exprimé dans la population d’Uchizy
respecte les hypothèses relatives au climat, mais pas celles liées à l’absence de compétition au
nord. Les résultats obtenus pourraient aussi s’expliquer par la répartition des ressources. Les
vergers étant moins nombreux et plus distants au nord qu’au sud, les femelles avec une charge
alaire faible (et donc une bonne capacité à disperser) seraient sélectionnées au nord de la
vallée. Les femelles d’Uchizy seraient davantage caractérisées par un compromis entre
reproduction et dispersion, plutôt qu’entre reproduction et survie. Enfin, il n’est pas possible
de conclure pour Igé par manque d’information concernant la capacité de dispersion.
Bien qu’il existe d’importantes différences dans le nombre d’œufs entre les populations,
les résultats de cette étude ne permettent pas de déterminer si un compromis opère entre le
nombre et le volume des œufs, puisque les mesures de taille d’œufs réalisée à Lyon sont
semblables d’une population à l’autre. Une précédente étude réalisée sur la même espèce et
sur un gradient de 50 kilomètres seulement avait pourtant montré que le volume des œufs

22
d’une population du sud était supérieur à celui d’une population du nord (Vayssade, 2009). La
variation dans la taille des œufs semble donc possible chez cette espèce, mais ne s’applique
pas à toutes les populations étudiées.
Différentes explications pourraient être à l’origine des contrastes et des « incohérences »
retrouvées dans les populations de Leptopilina heterotoma utilisées pour cette étude. Les deux
populations du sud s’opposent fortement malgré leur proximité géographique. Deux
hypothèses peuvent être formulées concernant ce phénomène. Premièrement, il est tout à fait
possible que les microhabitats d’où proviennent les individus utilisés pour fonder les élevages
soient différents. En effet, il n’est pas possible d’exclure l’influence de facteurs abiotiques tels
que le traitement chimique des vergers ou la composition du paysage par exemple. De même,
les conditions biotiques peuvent varier d’un site d’échantillonnage à un autre, comme la
densité et la répartition des fruits qui conditionnent la répartition des hôtes, ou encore la
densité des compétiteurs. Les populations naturelles d’Avignon et d’Eyguières pourraient
donc avoir été soumises à des conditions biotiques et abiotiques différentes, et s’être ainsi
adaptées localement à leurs environnements respectifs. Cette adaptation locale pourrait avoir
conduit à une différenciation progressive des populations, qui se manifesterait alors par des
traits d’histoire de vie différents, et donc des stratégies différentes. La stratégie optimale peut
effectivement changer radicalement selon l’habitat et les patrons de distribution de la
ressource (Askari Seyahooei, 2010). Une deuxième hypothèse conduirait à penser que la
population de laboratoire pour Eyguières ne serait pas représentative de la population
naturelle. En effet, elle se conforme peu aux prédictions, et se distingue fortement d’Avignon
ainsi que de d’autres populations étudiées précédemment (voir Ris, 2003 ; Vayssade, 2009),
pour plusieurs traits. Cette opposition de stratégies pourrait directement résulter d’un
problème d’échantillonnage ou d’élevage, qui aurait créé un goulot d’étranglement (Simoes et
al., 2008). En effet, si les individus collectés sur le terrain proviennent d’un site très
particulier, ils pourraient s’y être adaptés en développant des caractéristiques singulières que
ne possèderaient pas les individus d’autres sites. Une autre possibilité réside dans le fait
qu’une partie seulement des femelles collectées pourraient s’être effectivement reproduites en
laboratoire, ayant pour conséquence une faible variabilité génétique au sein des individus de
l’élevage, qui descendraient tous d’un nombre très réduit de femelles. Là encore, si ces
femelles présentent des caractéristiques particulières, elles seront vite transmises à l’ensemble
de la population. Dans les deux cas, l’élevage aurait été constitué à partir d’individus

23
exceptionnels non représentatifs de la population d’origine, ce qui pourrait expliquer la
singularité des résultats trouvés pour Eyguières.
4.3. Conclusions et perspectives
Cette étude avait pour vocation de comparer l’évolution des traits d’histoire de vie chez
des populations de parasitoïdes originaires du nord et du sud de la vallée du Rhône, qui sont
soumises à des pressions de compétition et des conditions climatiques différentes, afin de
confirmer les résultats précédemment obtenus sur d’autres populations. Les résultats obtenus
ont montré que des variations de traits et de comportements peuvent exister entre les individus
des populations étudiées. De possibles compromis évolutifs ont même pu être identifiés dans
certains cas, notamment entre reproduction et survie, mais dans des sens parfois opposés chez
des populations pourtant originaires de la même zone géographique. Des stratégies
contrastées ont ainsi été mises en évidence, tout particulièrement entre les deux populations
du sud, Avignon et Eyguières. Les compromis évolutifs semblent donc pouvoir être
influencés à la fois par la compétition mais aussi par le climat, et très probablement par de
nombreux autres facteurs relatifs à l’environnement.
Les résultats de cette étude viennent compléter ceux de précédentes études qui s’étaient
déjà intéressées aux traits d’histoire de vie chez Leptopilina heterotoma. Bien que ce travail
apporte de nouveaux éléments de réflexion sur l’évolution de la communauté de parasitoïdes
de cette vallée, il souligne également la complexité et la multiplicité des mécanismes qui
sous-tendent les compromis, ainsi que la difficulté liée à leur analyse. Le recoupement de
certains résultats au travers des différentes études amène à penser que certains traits sont plus
pertinents que d’autres pour étudier l’évolution des histoires de vie, tout au moins dans la
communauté de parasitoïdes de la vallée du Rhône. Ainsi, il serait peut être plus efficace de se
concentrer sur les traits qui varient fortement entre populations, tels que la fécondité ou la
longévité, plutôt que sur des traits qui sont similaires sur l’ensemble de la vallée, comme la
taille ou le taux de lipides. De plus, il sera également nécessaire de multiplier les répétitions et
les populations étudiées dans les analyses futures, pour espérer distinguer des tendances
claires.

24
REFERENCES BIBLIOGRAPHIQUES
Allemand R., Fleury F., Lemaitre Claire & Boulétreau M., 1999. Dynamique des populations et
interactions compétitives chez deux espèces de Leptopilina, parasitoïdes de drosophiles, dans la vallée du Rhône (Hymenoptera : Figitidae). Annales de la Société Entomologique de France 35 : 97-103.
Angilleta M.J. & Dunham A.E., 2003. The temperature-size rule in ectotherms : simple evolutionary
explanations may not be general. The American Naturalist 162 : 332-342. Askari Seyahooei M., 2010. Life history evolution in Hymenopteran parasitoids: the role of host and
climate. Doctoral thesis, Leiden university. Atkinson D. & Sibly R.M., 1997. why are organisms usually bigger in colder environments ? Making
sense of a life history puzzle. Trends in Ecology and Evolution 12 : 235-239. Eggleton P. & Gaston K.J., 1990. Parasitoids species and assemblages – Convenient definitions or
misleading compromises. Oikos 59: 417-421. Ellers J., Driessen G. & Sevenster J.G., 2000. The shape of yhe trade-off betwenn egg production
and life span in the parasitoid Asobara tabida. Netherlands Journal of Zoology 50 : 29-36. Ellers J. & van Alphen J.J.M., 1997. Life history evolution in Asobara tabida: plasticity in allocation
of fat reserves to survival and reproduction. Journal of Evolutionary Ecology 10: 771-785. Fleury F, Allemand R., Fouillet P. & Boulétreau M., 1994. Geographic variation in locomotor
activity rhythms of Leptopilina heterotoma : inheritance and role in species richness of the Drosophila parasitoid community. Norwegian Journal of Agricultural Sciences 16 : 191-197.
Fleury F, Allemand R., Fouillet P. & Boulétreau M., 1995. Genetic variation in locomotor activity
rhythm among populations of Leptopilina heterotoma (Hymenoptera : Eucoilidae), a larval parasitoid of Drosophila species. Behavior Genetics 25 : 81-89.
Fleury F, Allemand R., Vavre F., Fouillet P. & Boulétreau M., 2009. Adaptive significance of a
circadian clock : temporal segregation of activities reduces intrinsic competitive inferiority in Drosophila parasitoids. Proceedings of the Royal Society of London B 267 : 1005-1010.
Fleury F., Ris N., Allemand R., Fouillet P., Carton Y. & Boulétreau M., 2004. Ecological and
genetic interactions in Drosophila-parasitoids communities : a case study with D. melanogaster, D. simulans and their common Leptopilina parasitoids in south-eastern France. Genetica 120 : 181-194.
Gilchrist G.W. & Huey R.B., 2004. Plastic and genetic variation in wing loading as a function of
temperature within and among parallel clines in Drosophila subobscura. Integrative Compilation Biology 44: 461-470.
Jervis M.A, Eller J. & Harvey J.A., 2008. Resource acquisition, allocation, and utilisation in parasitoid reproductive strategies. Annual Review of Entomology 53: 361-385.
Kolliker-Ott U.M., Blows M.W. & Hoffmann A.A., 2003. Are wing size, wing shape and
asymmetry related to field fitness of Trichogramma egg parasitoids, Oikos 100: 563-573. Le Lann C., 2009. Thèse de doctorat, université de Rennes 1, p.

25
Martel V., Wajnberg E. & Boivin G., 2008. Patch time allocation in male parasitoids. Ecological Entomology 33: 608-613.
May M.L., 1979. Insect thermoregulation. Annual Review of Entomology 24: 313-349. Mellanby K., 1939. Low temperature and insect activity. Nespolo R.F., Roff D.A. & Fairbairn D.J., 2008. Energetic trade-off between maintenance costs and
flight capacity in the sand cricket (Gryllus firmus). Functional ecology 22 : 624-631. Nicol C.M.Y. & Mackauer M., 1999. The scaling of body size and mass in a host-parasitoid
association: influence of host species and stage. Entomologia Experimentalis Et Applicata, 90: 83-92.
Nylin S. & Gotthard K., 1998. Plasticity in life history traits. Annual Review of Entomology 43: 63-83.
Pelosse P., 2008. Rôle des compromis évolutifs dans la spécialisation et la coexistence d’espèces
compétitrices. Développement théoriques et expérimentaux chez les insectes parasitoïdes. Thèse de doctorat, université de Lyon 1, 186 p.
Prévost G., Boulétreau M. & Wajnberg E., 1989. Effets de la surpopulation larvaire sur l’évolution
des larves de Drosophiles parasitées par les Hyménoptères Cynipides du genre Leptopilina. Bulletin de la Société Ecophysiologique 14 : 109-113.
Price P.W., 1974. Strategies for egg production. Evolution 28 : 76-84. Ris N., 1993. Hétérogéneité spatiale, plasticité phénotypique et trade-off environnementaux : rôle de
l’espèce hôte et de la température dans la différenciation génétiques des populations du parasitoïde Leptopilina heterotoma (Hymenoptera). Thèse de doctorat, université de Lyon 1, 159 p.
Roff D.A. & Fairbairn D.J., 2006. The evolution of trade-offs : where are we? Journal compilation
20 : 433-447. Stearns S.C., 2000. Life history evolution : successes, limitations and prospects. Naturwissenschaften
87 : 476-486. Vayssade C., 2009. Influence de la distribution des hôtes et de la compétition sur les traits d’histoire
de vie de deux populations de parasitoïdes de drosophiles. Mémoire de recherche de master 2, université de Rennes 1, 25p.
Waage J.K., 1979. Foraging for patchily-distributed hosts by the parasitoid, Nemeritis canescens.
Journal of Animal Ecology 48: 353-371. Wajnberg E., 2006. Time allocation strategies in insect parasitoid: from ultimate predictions to
proximate behavioral mechanisms. Behavioral Ecology and Sociobiology 60 : 589-611.

26
RESUME
L’évolution des histoires de vie est façonnée par de nombreux facteurs internes et externes aux organismes. Cette étude a consisté à comparer les traits d’histoire de vie de plusieurs populations du parasitoïde Leptopilina heterotoma, sur un gradient nord-sud dans la vallée du Rhône. Ces populations sont soumises à des pressions de compétition et à des conditions climatiques différentes, influençant probablement l’allocation énergétique dans les différents traits, et donc les éventuels compromis évolutifs qui s’opèrent entre ces traits. Les résultats ont mis en évidence l’existence de variations entre populations du nord et du sud pour certains traits, comme la longévité, la fécondité, la charge alaire et le métabolisme. Par contre, d’autres sont similaires entre les populations, confortant des résultats précédents. Cette étude permet d’apporter de nouveaux éléments de réflexion quant à la pertinence du choix des traits d’histoire de vie dans les analyses comparatives s’intéressant aux facteurs impliqués dans l’évolution de ces traits. Cependant, deux populations proches géographiquement expriment des stratégies contrastées, et parfois même opposées bien qu’elles subissent apparemment les mêmes pressions compétitives et le même climat. Cette opposition pourrait s’expliquer par des adaptations locales conduisant à un investissement différent de la ressource énergétique ou par un goulot d’étranglement accidentel lors de la mise en place des élevages. Ce contraste entre ces deux populations permet de mettre en évidence la plasticité de l’allocation des ressources entre les différents traits. Mots clés : parasitoïdes, traits d’histoire de vie, compromis évolutifs, compétition, climat
ABSTRACT Life history evolution of organisms is shaped by several intrinsic and extrinsic factors. The present study aims to compare life history traits of different populations of the parasitoid Leptopilina heterotoma, along a north to south cline in the Rhône valley. These populations suffer from different competitive pressions as well as climatic conditions, probably mediating resource allocation to different traits, and thus the potential trade-offs which occur between traits. Results have highlighted variations between populations for some of the traits, like fecundity, longevity, wing loading and metabolism. By contrast, others traits are similar between populations, consistent with precedent results. This study brings new insights concerning the relevance of the life history traits chosen in comparative analysis which focus on the factors implicated in traits evolution. However, two populations from the same geographic origin show contrasted strategies, even opposite ones despite the fact that they suffer similar competitive and climatic pressions. This opposition may results from local adaptations leading to differential resource allocation or from a bottleneck at the beginning of the rearing. These contrast between populations put in evidence the plasticity of resource allocation to different traits. Key words : parasitoids, life history traits, trade-off, competition, climate