Embed Size (px)
Citation preview

BACTERIOLOGIE M # d e c i n e e t M a l a d i e s I n f e c t i e u s e s - 1986 - 11 b is - 644 ~ 654
Beta-lactamases :identification, aspects pidemiologiques*
par A, PHILIPPON** et G. PAUL**
rSle,
RESUME Historiquement, les Mactamases ont ~t~ le premier m~canisme de r~sistance individualis~ dans le cadre de la r~sistance naturelle (cdphalosporinase de bacilles & Gram-n~gatif) et
acquise (pdnicillinase de S. aureus). Avec I'av~nement des ~-Iactamines ~ large spectre comme I'ampicilline et les cdphalosporines de premiere gdn~ration (cdphalothine par exemple) sont apparues les p~nicillinases plasmidiques des bacilles & Gram-n~gatif, la principale d'entre elles ~tant la ~-Iactamase de type TEM. Le r61e ~piddmique de ces p~nicillinases est d'autant plus efficace qu'elles sont transposables, d'o~ I'extension plus r~cente ~ des esp~.ces jusque-I~ dpargndes comme N. #onorrhoeae, H. influenzae, H. ducreyi, voire N. meningitidis. La d~couverte plus r~cente de ~-Iactamines peu inductrices, r~putdes non hydrolysables a modifid I'image de la rdsistance acquise. La predisposition de certains esp~ces (phdnotype cdphalosporinase- inductible) aptes ~ produire constitutivement (mutation) des quantitds ~lev~es de ~-Iactamase, leur donne les moyens de s'adapter en relation avec I'inactivation de la plupart des Mactamines pr~sentes. La derni~re dtape enzymatique connue dans la r~sistance aux nouvelles ~-Iactamines s'oriente vers les ~-Iactamases large spectre produites en grande quantit~ et rencontr~es jusqu'~ present chez certaines ent~robact~ries comme les Klebsiella (K. oxytoca, K. pneumoniae). Une de ces enzymes est encore un exemple unique de Mactamase adapt~e par mutation (SHV-2) aux nouvelles Mactamines. Ainsi, le monde bactdrien poss~de en son sein de nombreuses potentialit~s d'adaptation aux nouvelles thdrapeutiques. Seule la pression de sd- lection, suite souvent ~ des abus de prescription, et des r~gles d'hygi~ne ddfectueuses permettront leur ex- pression et leur extension. Aussi, conviendra-t-il de surveiller toute dmergence de souches anormalement rdsistantes et d'appr~cier son incidence dans le temps, le m~canisme enzymatique restant prddominant jusqu't~ prdsent.
Mots-cl~s : Mactamases- Identification - Classification - R61e- R~partition
La d~couverte d'enzymes inactivant la benzylp~nici l l i - ne en presence d'extraits bact~riens, par exemple d'E. c o / i ,
date de 1940. Ceci d~montrai t pr~coc~ment I'existence d'un m~canisme enzymatique de la r~sistance, en I'occur- rence, naturelle. L'enzyme fu t d~nomm~e b tor t <<p~nicilli- nase>), le terme de ~-Iactamase n'ayant (~t~ propos~ qu'en 1960 par Pollock. Quelques ann~es plus tard fu t rapide- ment identifi~e la p6nicil l inase de S. a u r e u s (r~sistance acquise). Cette ~mergence de la r~sistance bact~rienne en relation avec les /~-Iactamases ne devait plus s'arr~ter.
Celles-ci se definissent par rapport au cycle #-Iactame dont I 'hydrolyse entraine la perte d'activit~ antibact~rienne de la ~-Iactamine. Avec certains substrats, I 'hydrolyse est di f f ic i le b visualiser par les techniques usueltes, laissant pr~- juger, ~ tort , d'une bonne stabil it~ telle celle des nouvelles fl-lactamines, c~phalosporines de troisibme g~n~ration, par exemple. II s'agit, en fait d 'une hydrolyse faible et lente
(66), le m~canisme de <(trapping)> ayant ~t~ ini t ialement propos~ (63).
Deux autres m~canismes de r~sistance ont ~t~ identi- fies. II s'agit soit d 'une d iminut ion de la di f fusion des an- t ibiot iques ~ travers la paroi bact~rienne des germes Gram-n~gatif, essentiellement au niveau de prot~ines tri- m~res (porines), soit de modif ications d'un ou plusieurs si- tes d'action de ces antibiot iques dans la membrane cyto- plasmique,d~nomm6s prot~ines de liaison ~ la p6nici l l ine (PLP) (2, 9, 10, 27).
*Communication pr~sent~e b la r~union de la Soci~t~ de Pathologie infectieuse de Langue franc;aise, tenue b Saint-Mand6 (H6pital d'ins- truction des arrn6es ~6gin), les 5 et 6 d~cembre 1986 sur le th~me : ¢{ Les nouvelles betalactamines en 1986>) **Service de Bact~riologie, Centre Hospitalier Universitaire Cochin, 27 rue du Fg Saint-Jacques, F-75014 PARIS
644

La responsabilit~ des /3-1actamases (incidence nulle d'autres enzymes d' inactivation, amidases et est~rases) est mentionn~e comparativement aux autres facteurs (tableau I).
TABLEAU I Facteurs de r~sistance aux/~-Iactamines selon les bact~ries
Facteur Gram-positif a Gram-n~gatif
Enzymes (~-Iactamases) -}- +J r '+ Paroi (porines) -I- "+ Membrane (PLP) -t-~- -I- -I-
a : la tolerance reste ~ ~valuer en clinique
L'apparit ion r~cente de nouvelles molecules d'h~misyn- th~se ou de synth~se au sein de la famitle des ~-Iactamines permit d' individualiser de nouveaux groupes tels les inhibi- teurs ezymatiques, les monobactames, les carbap~n~mes, reftets d'une recherche efficace en relation directe avec les ~-Iactamases, dont les axes ~taient :
-- inductibi l i t~ diminu~e (c~phalosporinases chromoso- miques)
- aff init~ faible (c~phalosporinases chromosomiques) - stabilit~ augment~e (p~nicillinases et c~phalospori-
Bases) -- effet inhibiteur irreversible (p~nicillinases)
La vitesse de p~n~tration am~lior~e ~ travers la paroi, et enfin le meilleur tropisme possible pour les cibles sont les axes de recherche li~s aux autres m~canismes de r~sis- tance. La mult ipl ici t~ actuelle des antibiol~iques de cette famil le illustre bien les progr~s accomplis mais ne rend que plus perplexe le choix th~rapeutique. Le tableau II retrace les principales ~tapes relatives ~ la d~couverte de nouvelles (~-Iactamines et/~-tactamases.
TABLEAU II Historique des ~-Iactamines et des/3-tactamases
1940 1944 1948 1955 1955 1960 1961 1962 1965 1967 1970 1972 1973 1974
1976
1977
1978 1979 1980 1981 1982 1983
1984
1986
Benzylp~ntcilline
P h~noxym~thylp~ nicilline C~phalosporine C 6-APA (amidase) M~thicilline Ampicilline, oxacilline C~phalothine
Carb(~nicilline
C(~phamycines C~foxitine
(¢P~niciltinase)> P~nicillinase (S. aureus)
/~-Iactamase, Inhibiteurs
~-Iactamase TE M (Temoniera)
Carb~nicillinase Stabilit~ aux ~-Iactamases
TEM-1 H. influenzae Transposon TnA
Thi~namycine, Nocardicine TEM-1 N. gonorrhmae A M ~tho×yiminoc~phalospo- tines Ac.clavulan ique, Sulbactam Latamoxef, Ceftazidirne Irnipenem M onobactames
~-Iactamase PSE
C~phalosporinase d~r~prim~e ~-Iactamases Klebsiella TEM-1 N. meningitidis BRO-1 B. catarrhalis LC R 1 P. aeruginosa Pase S. faecalis OXA4,OXA5,OXA6,OXA7 Pase F. nucleatum Pase P. multocida, ROB-1 NPS-1, SHY-2, CAR-4, SAR-1, OHIO
L'appari t ion continuel le des ~-Iactamases chez des esp~ces bact~riennes jusqu'ators sensibles ainsi que I'~mer- gence d'autres ~-Iactamases chez celles d~j~ r~sistantes pose le probl~me de I'origine, de la phylog~n~se et aussi de I '~pid~miologie de ces enzymes. Aussi, I ' ident i f icat ion devra se fonder sur une approche m~thodologique precise de plusieurs crit~res (3), justif iant r~ellement alors la d~nomi- nation d'enzymes nouvelles ou d~montrant I '~volution de celles d~j& conr)ues.
Le tableau III indique les principaux crit~res utilis~s dans I ' ident i f icat ion de quelques unes de ces enzymes com- mes les p~nicillinases plasmidiques de S. aureus, celles de bacilles ~ Gram-n~gatif (TEM, OXA, CARB), celles d'une c~phalosporinase inductible (I) ou d~r~prim~e (C) d'ent~- robact~ries et enfin de ~-Iactamases & large spectre de Klebs ie l la (7, 11, 13, 21, 22, 29, 30, 31, 51, 60, 64).
LOCALISATION - BIOGENESE
La Iocalisation de ces enzymes est exocellulaire chez les esp~ces ~ Gram-posit i f (S. aureus, Baci l lus) ou p~riplasmi- que (li~e ~ la paroi) chez celles ~ Gram-n~gatif. Chez les ent~robact~ries, une barri~re de perm~abilit~ y est, souvent, associ~e en raison de la diffusion sp~cifique des molecules selon leur hydrophobici t~ par les porines.
Le d~terminisme de leur synth~se, encore incompl~- tement ~lucid~, diff~rencie les (3-1actamases inductibles (p~nicillinases de germes ~ Gram-positif, S. aureus, Bac i l lus et c~phalosporinases de certaines esp~ces de bacilles ~ Gram n~gatif) de celles constitutives, synth~tis~es en I'absence d' inducteur (~-Iactamine). Ce sont les p~nicillinases plasmi- diques des bact~ries ~ Gram-n~gatif comme TEM, 0 X A , PSE mais aussi les ~-Iactamases chromosomiques des Kleb- siella.
Un des axes de recherche f~cond a ~t~ de s~lectionner des antibiotiques ayant pas ou peu d'effet inducteur, en particulier ~ I;~gard des c~phalosporinases sp~cifiques d'en- t~robact~ries (33). Les premieres ~-tactamines ~ large spec- tre (ampicill ine, c~phalosporines de premiere g~n~ration), boBs inducteurs, d~clenchaient, en effet, la product ion de I'enzyme chez les esp~ces <(c~phalosporinase inductible)) vis-a-vis de ces antibiot iques et donc la r~sistance naturelle. L'un des progr~s r~cents a ~t~ I 'obtent ion de p~nicillines (acyl-ur~idop~nicillines) ou de c~phalosporines peu ou pas inductrices, permettant une extension du spectre des #-Iactamines aux esp~ces de ph~notype ((c~phalosporinase)) (45).
L'effet hautement inducteur de certains ~-Iactamines comme la c~foxit ine ou I ' imipenem avec E. cloacae montre une apti tude g~n~tique de certaines esp~ces ~ produire des quantit~s d'enzyme cent ~ mil le fois sup~rieures ~ la nor- male (54). Le ou les m~canismes qui r~gulent cette hyper- product ion sont encore mal connus (8, 35). L'~mergence de souches d~r~prim~es (constitutives) montrant une biosyn-
645

TABLEAU III Param~tres d'identification des/~-Iactamases
S. aureus TEM OXA CARB CASE I CASE C Large spectre
Localisation Exocellulaire ÷ -- -- ÷ + Periplasmique
Biog~n~se lnductible ÷ ÷ ÷ Constitutive -- ~- ~- ~- -- ~- ÷
Activit~ enzymatique 1 --10 (mU/mg) ÷ ÷a ÷ ÷ > oo + + + - +
Profil enzymatique c (Vmax) Pase Pase Pase Pase Case Case Large
Inhibition d Ac. Clavulanique ÷ ÷ ÷ ÷ ÷ Coxacilline ÷ ÷ ÷ ÷ ÷-~-÷ ÷ - ~ ÷ ÷ Chlorures -- -- ÷ . . . .
Point iso~lectrique e 8,9 5,4-5,6 7-8 4,3-5,75 > 7 > 7 5-8 Poids mol~culaire (X103) 29 25 23-44 15-28 23-42 G~n~tique
Chromos0ne f -~- ÷ Jr Plasmide ~- -~ -~- -~- -- --g --g Transposon -- ÷ ÷ -'}- -- --
a TEM-1 et H. inf luenzae b : activitY augment6e jusqu'~ 5000 lois (mutants d~r~prim~s) c distinguant les p~nicillinases (Pase), sub-divis~es en TEM, oxacillinases, carb~nicillinases . . . . des c~phalosporinases (Case), de celles ~ large spec-
tre avec hydrolyse nette de/~-Iactamines dites non hydrolysables. d autres inhibiteurs utilis~s : parachtoromercuribenzoate, anticorps... e crit~re discriminant de nombreux types : TEM-1 et TEM-2, OXA-1 ~ OXA-7. . . f : sp~cifiques d'esp~ces, quelquefois transposables g : saul exception : pLQ3 (24) et pBP 60 (20)
th~se d~r~gul~e de la c~phalosporinase constitue un exem- pie r~cent d'adaptabil it~ bact~rienne dont les consequences en termes de r~sistance crois~e seront examinees plus loin. L'activit~ enzymatique est, souvent, en relation avec la CMI. Selon les souches (mutants d~r~prim~s ou constitu- tifs, plasmides multicopies), I 'activit~ #-Iactamase varie de quelques mil l iunit~s ~ 10 000 mil l iunit~s par mg prot~ines.
CONSTANTES CINETIQUES
Le crit~re principal d ' ident i f icat ion est d~fini par la vi- tesse d' inact ivat ion ou d'hydrolyse de l 'ant ib iot ique pris en compte (V max). Diff~rents profi ls de substrats sont ainsi individualis~s selon les #-Iactamases d6finissant des types enzymatiques (7, 11, 13, 21, 22, 24, 29, 30, 31,34, 42, 43, 44, 46, 51, 60, 64) corral,s avec la r~sistance de la bact~rie. Init ialement, I'existence d'une correlation directe entre la vitesse d'hydrolyse mesur~e et la r~sistance, appr~- cite en CMI ~tait admise. Ne tenir compte que de vitesses d'hydrolyse 61ev~es s'est av~r~ insuffisant pour expl iquer I'exemple r~cent des interactions entre les c~phalosporina- ses et les #-lactamines dites non hydrolysables (Vmax < 1) (66). Une meilleure appr6ciation et donc une s61ection de nouveaux antibiotiques prend aussi en compte leur aff init~ (constante de Michaelis-Menten ou Km) pour I 'enzyme examinee. Cette valeur est plus di f f ic i le ~ mesurer mais essentielle dans la r~sistance ou la sensibilit~ ~ un antibio- t ique de cette famille. La modif icat ion d'aff ini t~, vraisem-
blablement par mutat ion, peut ~largir consid~rablement le spectre d'une enzyme (tableau VI I ) (20).
De nombreux types enzymatiques ont ~t~ r~pertori~s et distingu~s en :
- C # p h a l o s p o r i n a s e s (Case) - P # n i c i l l i n a s e s (Pase) : TEM, OXA (oxacillinases),
CARB (carb~nicillinases) . . . - # - I a c t a m a s e s & l a rge s p e c t r e
- C ~ f u r o x i m a s e s
- CXases (oxyiminocephalosporin #-Iactamases).
Le terme p~nicill inase d~signe une hydrolyse pr~f~- rentielle mais ~ des degr~s divers des p~nicill ines, contrai- rement ~ celui de c~phalosporinase inactivant surtout les c~phalosporines. Les CXases montrent, en particul ier, I 'hy- drolyse des oxyimino-c~phalosporines (53) et quelquefois de I' imipenem (56). Dans l 'optique de la cl inique, il n'est pas n~cessaire de retenir les subtilit~s d ' ident i f icat ion des #-Iactamases r~pertori~es dans la l i tt~rature comme TEM-1, TEM-2, OXA-1, OXA-2, OXA-3, HMS, PIT-2, PSE-1, PSE-2, PSE-3, LCR-1, ROB-l , TLE-1, OXA-4, OXA-5, OXA-6, OXA-7, BRO-1, AER-1, CARB-4, OHIO, SAR-1, d'autant qu' i l s'agit quelquefois d'un seul exemplaire rap- port~ dans ,la l i t t~rature c0mme HMS, TLE-1, AER-1, CARB-4 (13, 29 ,30 , 46, 59). En revanche, la signif ication d 'un type donn~ en termes de r~sistance (TEM, CARB, OXA, Case inductible, Case d~r~prim~e, #-Iactamase large spectre), sa fr6quence par esp~ce bact~rienne et pal pays
646

est d'une importance clinique extreme en raison du choix th~rapeutique.
A U T R E S CRITERES
Les inhibiteurs de/~-Iactamases, utilis~s d~s la d~couver- te de celles-ci ont un int~r~t non seulement th~rapeutique mais d'identification. Un profi l d' inhibit ion par type d'en- zyme est alors bas~ sur le comportement vis-a-vis de d i f f , - rents inhibiteurs de type ~-Iactamine, analogues structuraux donc comp~titifs, chimiques (EDTA, chlorures, pCMB) ou immunologiques (imrnuns6rurns anti-TEM, anti-SHV-1, an t i -CARB. . . ) .
-- Enfin, pour deux ~-Iactamases ayant m~me profil de substrat et m~me profil d' inhibit ion, la d~termination comparative du point iso~lectrique (pl) peut permettre de les diff~rencier (tableau IV). Cette distinction fine par le pl a un grand int~r~t dans I'inventaire de la r~sistance en- zymatique et Iors d'enqu~tes ~pid~miologiques ~tablissant la distribution des/~-Iactamases par esp~ces et par pays (29, 48, 58, 67). Elle est peu utile au clinicien. Ainsi, les enzy- mes de type TEM sont ais~ment distinguables par te pl (5,4 et 5,6) mais ne different que par un seul acide amin~, torsque I'on examine leur s~quence primaire (1). Parmi les autres crit~res, le poids mol~culaire est un ~l~ment de clas- sification de moindre int~r~t en raison des faibles variations observ~es pour un type donn~. L'avenir des sondes nucl~i- ques apparait prometteur, car elles permettront des ~tudes d'homologies g~n~tiques plus rapides que la d~termination de s~quences primaires (1,39).
TABLEAU IV Points iso(~lectriques d'enzymes plasmidiques
Type TEM Type CA R B
Type OXA
TEM-1 (5, 4) TEM-2 (5, 6) CARB-1 (5, 3), CARB-2 (5, 7), CARB-3 (5,75), CARB-4 (4,3) OXA-1 (7,4), OXA-2 (7,7 et 7,4), OXA-3 (7,0), OXA-4 (7,45-7,5), OXA-5 (7,6), OXA-6 (7,8), OXA-7 (7,65)
D E T E R M I N I S M E G E N E T I Q U E
Ce crit~re d'individualisation d'une prot~ine est essen- tiel dans I'histoire de ces enzymes et leur ~pid~miologie. Celles-ci sont soit chromosomiques et sp~cifiques d'esp~ce relies les c~phalosporinases d'E. co/i, d'E. cloacae, de B. fragilis, les p~nicillinases de Klebsiella, soit plasmidiques comme celles de bact~ries ~ Gram-n~gatif (p~nicillinases de type TEM, OXA, PSE . . . . ). Les/~-Iactamases plasmidiques comme celle de type TEM d~couverte en 1965 jouent un grand r~le ~pid~miologique, en raison de leur extension des esp~ces taxonomiquement tr~s diff~rentes. Le transfert du g~ne/~la (TEM) sur de nombreux plasmides d'incompa- tibilit~s diff~rentes a ~t~ ~ I'origine, en 1974, de la d~cou- verte ((physique)) du transposon (12). Une enzyme chromo-
somique chez certaines esp~ces comme SHV-1 (PIT-2) ou PSE-4 peut ~tre plasmidique chez d'autres. Les transposons ~-Iactamases TEM-2 et TEM-1 tels Tn 1, Tn 2 et Tn 3 sont responsables d'~pid~mies et de l'extension assez r~cente de la r~sistance aux p~nicitlines ~ large spectre ~ des esp~ces jusque I~ ~pargn~es : H. influenzae, N. gonorrhoeae, H. ducreyL L'avenir devrail~ montrer d'autres exemples d'~chan- ges interbact~riens (6). Un aspect encore incompl~tement r~solu du d~terminisme g~n~tique concerne la nature du ou des syst~mes de r~gulation des c~phalosporinases chro- . mosomiques des bacilles ~ Gram-n~gatif comme E. cloacae ou C. freundii (8, 35, 54). Un autre aspect m~rite d'etre rapport~ : il s'agit de la possibilit~ d'une amplification du g~ne ~la (7).
CLASSIFICATION
Diff~rentes classifications des /~-Iactamases, dans les- quelles devraient pr~valoir les termes de c~phalosporinase ou de p~nicillinase, ont 6t~ propos~es par diff~rents auteurs (7, 29, 34, 51, 60). L'absence de r~gles pr~cises et admises au niveau international, en ce qui concerne les crit~res d'identif ication ~ utiliser obligatoirement et d'autre part, la valeur excessive ~ donner ~ d'autres comme actuellement le point iso~lectrique entretiennent la confusion. Ces pro- blames de sp~cialistes devraient 6tre r~solus par I'adoption de crit~res rationnels, rendus n~cessaires par la d~couverte continuelle de ((nouvelles enzymes)). Aussi, avant de propo- ser une nouvelle d~nomination, a-t-on ~tudi~ comparative- ment I'enzyme ~ identifier avec celles d~j~ identifi~es. Aussi en I'bsence de r~gles pr~cises, n'est-il gu~re surprenant que pour un m~me type enzymatique, il puisse y avoir plusieurs appellations, variables selon le pays comme.-]e montrent les exemples de ~-Iactamases de bacilles ~ Gram- n~gatif (Ent~robact~ries et Pseudomonas) (tableau V) (7, 30, 34, 40, 51). II conviendra dans I'avenir de suivre des r~gles plus strictes de caract~risation et de d~nomination afin d'~viter les confusions et d'arriver ~ un effort de clari- fication pour le non sp~cialiste.
TABLEAU V Synonymes de quelques ~-Iactamases de bacilles & Gram n~gatif
(3O, 4O)
Richmond Mitshuhashi Matthew Pitton Labia Philippon
Classe Ilia Type la TEM-1 PIT-1 Ilia Ib TEM-2 PIT-1 IV SHY-1 PIT-2 Va II OXA-1 Vb III OXA-2 V OXA-3
V IV PSE-1 CARB-2 PSE-2 OXA-4* PSE-3 PSE-4 CA R B-1
*appellation donn~e & un autre type d'enzyme tr~s proche de OXA- 1 (31)
647

C O M P O R T E M E N T DES # - L A C T A M I N E S S E L O N LES TYPES D ' E N Z Y M E
Les probl6mes pos6s par I ' ident i f icat ion et la classifica- t ion d'une fl-lactamase ou encore celui de tenter de pr6ciser le d6terminisme g~n6tique de la r6sistance (chromosomique, plasmidique ou transposable) lors de la d#couverte d'un nouveau type d'enzyme peuvent apparaitre de la s~manti- que. Pour le clinicien, seul importe le meilleur choix th6ra- peutique orient6 souvent en fonct ion des connaissances fondamentales et bact6riologiques les plus r6centes,
L'un des aspects diff ici les de ce probl~me de la r6sistan- ce concerne le comportement exact de telie ou telle/~-Iac- tamine vis-a-vis des diff6rentes ~-Iactamases connues, non seulement in vitro, ce qui est assez ais6ment r6alisable mais surtout in vivo, afin de d61imiter les condit ions de la r6sis- tance cl inique, c'esto~-dire de l'~chec clinique.
Au laboratoire, ce r61e de l 'enzyme seule est quelque- fois malais# ~ d~terminer pour une souche cl inique posse- dant un autre type enzymatique ou un autre m6canisme de r6sistance (imperm6abilit6, modif icat ion d 'af f in i t6 d'une ou plusieurs PLP) surajout6 au premier.
Si le type enzymatique identif i6 importe, d'autres param~tres sont ~ prendre en consid6ration comme la quantit6 d'enzyme produite pouvant varier de quelques mU ~ plusieurs mill iers de mU/ml par modif icat ion d'acti- vit~ du g~ne ( induct ion, mutants constitut i fs) ou en pre- sence d'une populat ion bact6rienne dense (effet inocu- lure).
Plusieurs approches m~thodologiques permettent de connaitre le comportement pr6cis des/~-Iactamines en fonc- t ion de la ~-Iactamase. Parmi celles-ci, la comparaison des sensibilit6s, d6termin6es par les CMI, est 6tablie apr6s transfert du g~ne (~4actamase (/~la) obtenu par transforma- t ion, conjugaison ou transduction ~ la m6me souche sensi- ble (isog6nique) (14, 15). Une autre modalit6 g6n6tique, plus comptexe, concerne la transposition du m~me g6ne ~la sur des plasmides multicopies ou encore en I'absence d'excision naturelle de cloner le g~ne chromosomique (15).
Le r61e de la fl-lactamase chromosomique (c6phalo- sporinase non transf6rable) est aussi appr6ci6 par la s~lec- t ion in vitro ou in vivo de mutants consti tut i fs ou d6r6- gul6s (4, 8, 54, 62). La figure 1 il lustre les CMI de diff6- rentes fl-lactamines, certaines non hydrolysables en termes de Vmax, pour une souche de S. marcescens c6phalospo- rinase inductible et son variant d6r6prim6 (8).
Une autre approche m6thodologique est celle de la surveillance de la sensibilit6 des souches cliniclues. Les 6tu- des mult icentriques sur des 6chantillonnages comportant plusieurs mitliers de souches sont encore trop rares, mais donnent des indications primordiales sur la concordance du comportement des /~-Iactamines par ph~notype de
100
CMI (mg/I)
60
20
0
-lactamines
TIC
Souche inductible
[ ] Souche d6r6prirn~e
JJ m°! "PIP "CFM "CTX MOX
MEZ MEC CPZ CAZ AZT IMI'
a : ~-Iactamine, TIC, Ticarcilline ; MEZ, Mezlocilline ; PIP, Pip~racil- line ; MEC, M6cillinarn ; CFM, C~famandole ; CPZ, C6fopSrazone ; CTX, C6fotaxime ; CAZ, Ceftazidime ; MOX, Latamoxef ; AZT, Az- tr~onam ; IMI, Imipenem
FIGURE 1 Comportement des/3-1actamines (CMI) chez S. m a r c e s c e n s
et son variant d(~r~prim# (8)
r~sistance mais ~galement sur la prevalence de tel ou tel m6- canisme comme cela a ~t6 r6cemment mis en oeuvre pour P. aeruginosa (40, 61, 67). Enfin, route souche de sensibili- t~ anormale ou inconnue pour une esp~ce donn~e sera soi- gneusement examinee en fonct ion des m6canismes de r6sis- tance connus, type de #-Iactamases par exemple.
II faut ainsi indiquer le r61e r6cent (1983) observ6 avec les ~-Iactamases ~ large spectre de Klebsiella, en particul ier celles de K. oxytoca, qui pourrait constituer la menace des prochaines ann~es d'autant que plusieurs pays dont la France signalent maintenant leur apparit ion (11, 22, 64). Parmi celles-ci, I 'enzyme plasmidique SHV,2, observ6e chez plusieurs esp~ces & Gram n6gatif comme K, ozaenae, cons- t i tue un exemple unique d'adaptation par mutat ion de I'en- zyme SHV-1, pr6c6demment ddnomm~e PIT-2 (tableau Vl) . II y a augmentation significative des CMI de diff6rentes c6- phalosporines non hydrolysables comme le c#fotaxime, la ceftr iaxone ou la ceftazidime mais aussi du monobactam examine, I'aztr6onam.
TABLEAU Vl Sensibilit6 des/~-Iactamines aux/~-Iactamases SHV-1 et SHV-2 (12)
/~-factamine K. ozenae a E. coli S SHV1 SHV 2 a
P6nicilline G >2048 8 512 >2048 Ampicilline > 2048 2 1024 > 2048 C~falotine 128 8 32 128 C~furoxime 8 2 8 8 C6fotetan 0,125 0,06 0,06 0,25 C~fotaxime 4 0,03 0,03 4 Ceftriaxone 4 0,03 0,03 4 Ceftazidime 2 0,125 0,125 4 HR 810 1 0,015 0,03 1 Aztr~onam 1 0,06 0,03 1 I mipenem 0,5 0,5 0,5 0,5
a : plasmide pBP 60
648
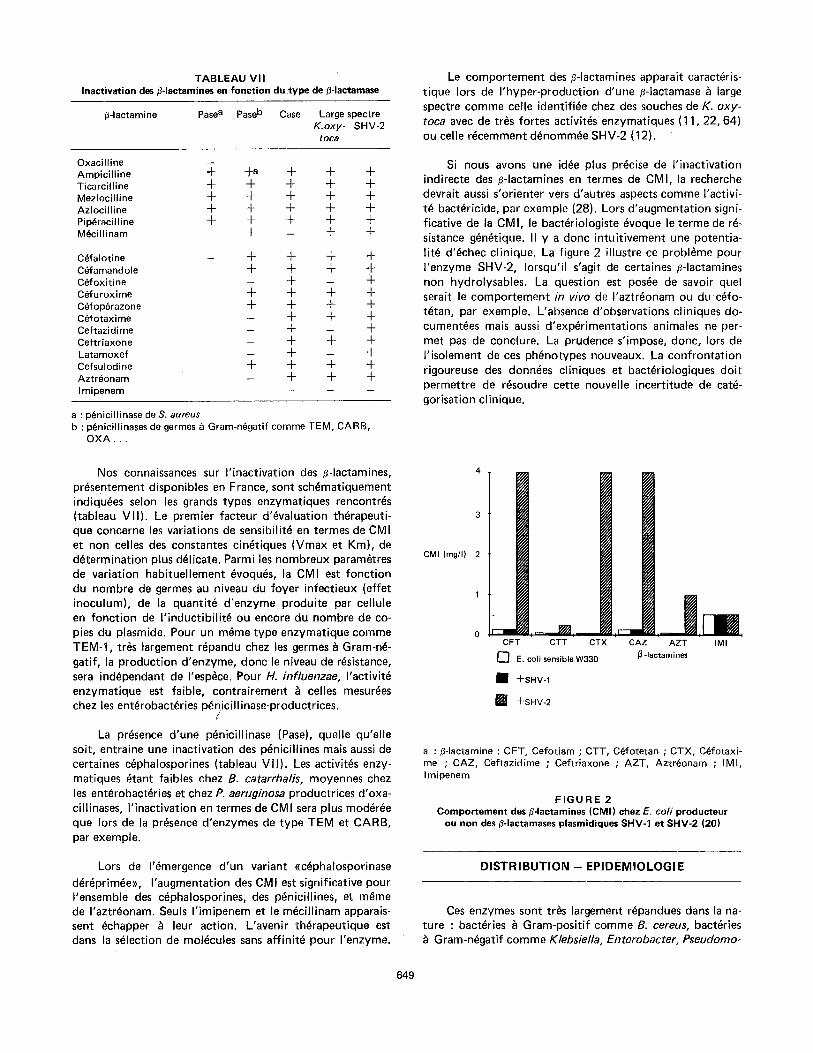
T A B L E A U V I I Inactivation des #-Iactamines en fonction du .type de (~-Iactamase
fl-lactamine Pase a Pase b Case Large spectre K.oxy- SHV-2
toc~
Oxacilline Ampicilline ~- Ticarcilline 4- Mezlocilline 4- Azlociltine 4- Pip~racilline 4- M~cillinam
C~falotine C~famandole Cefoxitine C~furoxime C~fop~razone Cefotaxime Ceftazidi me Ceftriaxone Latamoxef Cefsulodine Aztr~onam I mipenem
+a + + + + + + + + + + + + + + + + + + + + - + +
+ + + + + + + +
+ + + ~ +
+ + + + _ + + + _ + _ + - + + + - + - + + + + + - + + +
a : p6nicillinase de S. aureus b : p6nicillinases de germes ~ Gram-n~gatif comme TEM,
OXA...
CARB,
Nos connaissances sur I'inactivation des fl-lactamines, pr~sentement disponibles en France, sont sch~matiquement indiqu~es selon les grands types enzymatiques rencontres (tableau V I I). Le premier facteur d'~valuation th~rapeuti- que concerne les variations de sensibilit~ en termes de CMI et non celles des constantes cin~tiques (Vmax et Km), de d~termination plus d~licate. Parmi les nombreux param~tres de variation habituellement ~voqu~s, la CMI est fonction du nombre de germes au niveau du foyer infectieux (effet inoculum), de la quantit~ d'enzyme produite par cellule en fonction de I ' inductibil i t~ ou encore du nombre de co- pies du plasmide. Pour un m~me type enzymatique comme TEM-1, tr6s largement r~pandu chez les germes b Gram-n~- gatif, la production d'enzyme, donc le niveau de r~sistance, sera ind~pendant de I'esp~ce. Pour H. influenzae, I'activit~ enzymatique est faible, contrairement ~ celles mesur~es chez les ent~robact~ries p~nicillinase-productrices.
/
La presence d'une p~nicillinase (Pase), quelle qu'elle soit, entraine une inactivation des p~nicillines mais aussi de certaines c~phalosporines (tableau VII). Les activit~s enzy- matiques ~tant faibles chez B. catarrhalis, moyennes chez les ent~robact~ries et chez P. aeruginosa productrices d'oxa- cillinases, I'inactivation en termes de CMI sera plus mod~r~e que Iors de la presence d'enzymes de type TEM et CARB, par exemple.
Lors de I'~mergence d'un variant ((c~phalosporinase d~r~prim~e)), I'augmentation des CMI est significative pour I'ensemble des c~phalosporines, des p~nicillines, et m~me de I'aztr~onam. Seuls I'imipenem et le m~cillinam apparais- sent ~chapper ~ leur action. L'avenir th~rapeutique est dans la s~lection de molecules sans affinit~ pour I'enzyme.
Le comportement des fl-lactamines apparait caract~ris- tique Iors de I'hyper-production d'une ~-Iactamase & large spectre comme celle identifi~e chez des souches de K. oxy- toca avec de tr~s fortes activit~s enzymatiques (11,22, 64) ou celle r~cemment d~nomm~e SHV-2 (t2).
Si nous avons une idle plus precise de l' inactivation indirecte des 13-1actamines en termes de CMI, la recherche devrait aussi s'orienter vers d'autres aspects comme I'activi- t~ bactericide, par exemple (28). Lors d'augmentation signi- ficative de la CMI, le bact~riologiste ~voque le terme de r~- sistance g~n6tique. II y a donc intuitivement une potentia- lit~ d'~chec clinique. La figure 2 illustre ce probl6me pour I'enzyme SHV-2, Iorsqu'il s'agit de certaines fl-lactamines non hydrolysables. La question est posse de savoir quel serait le comportement in vivo de I'aztr~onam ou du c~fo- t~tan, par exemple. L'absence d'observations cliniques do- cument~es mais aussi d'exp~rimentations animales ne per- met pas de conclure. La prudence s'impose, donc, Iors de l'isolement de ces ph~notypes nouveaux. La confrontation rigoureuse des donn~es cliniques et bact6riologiques doit permettre de r~soudre cette nouvelle incertitude de cat~- gorisation clinique.
CMI (mg/I) 2
m CFT CTT CTX CAZ AZT IMI
[ ] E. coli sensible W 3 3 0 ~-Iactamines
1B +SHV-1
[ ] -FSHV-2
a : ~-Iactamine : CFT, Cefotiam ; CTT, C6fotetan ; CTX, C~fotaxi- me ; CAZ, Ceftazidime ; Ceftriaxone ; AZT, Aztr~onam ; IMI, Imipenem
F I G U R E 2 Comportement des/3-1actamines (CMI) chez E. coli producteur
ou non des/~-Iactamases plasmidiques SHV-1 et SHV-2 (20)
D I S T R I B U T I O N - E P I D E M I O L O G I E
Ces enzymes sont trbs largement r~pandues dans la na- ture : bact~ries b Gram-positif comme B. cereus, bact~ries
Gram-n~gatif comme Klebsiella, Enterobacter, Pseudomoo
649
nas, bact~ries ~ Gram-negatif ana~robies strictes tel(es B. fragilis, streptomyc~tes, actinomyc~tes (N. asteroides, Mycobacterium), levures, algues bleues et m~me tissus de certains mammif~res comme en t~moigne chez I'hom- me et les primates, I ' inactivation de I ' imipenem par une dihydropeptidase rdnale (19). Selon te type enzymati- que habituel lement trouv~ et malgr~ I'association possi- ble d'un autre m~canisme de resistance, I'expression pheno- typique ((naturelle)) de nombreuses esp~ces de bacilles Gram-n~gatif est maintenant connue des bact~riologistes medicaux (16, 38, 45).
Un aspect plus nouveau concerne les types d'enzy- mes responsables de la resistance acquise et leur inciden- ce, plus particuli~rement en France.
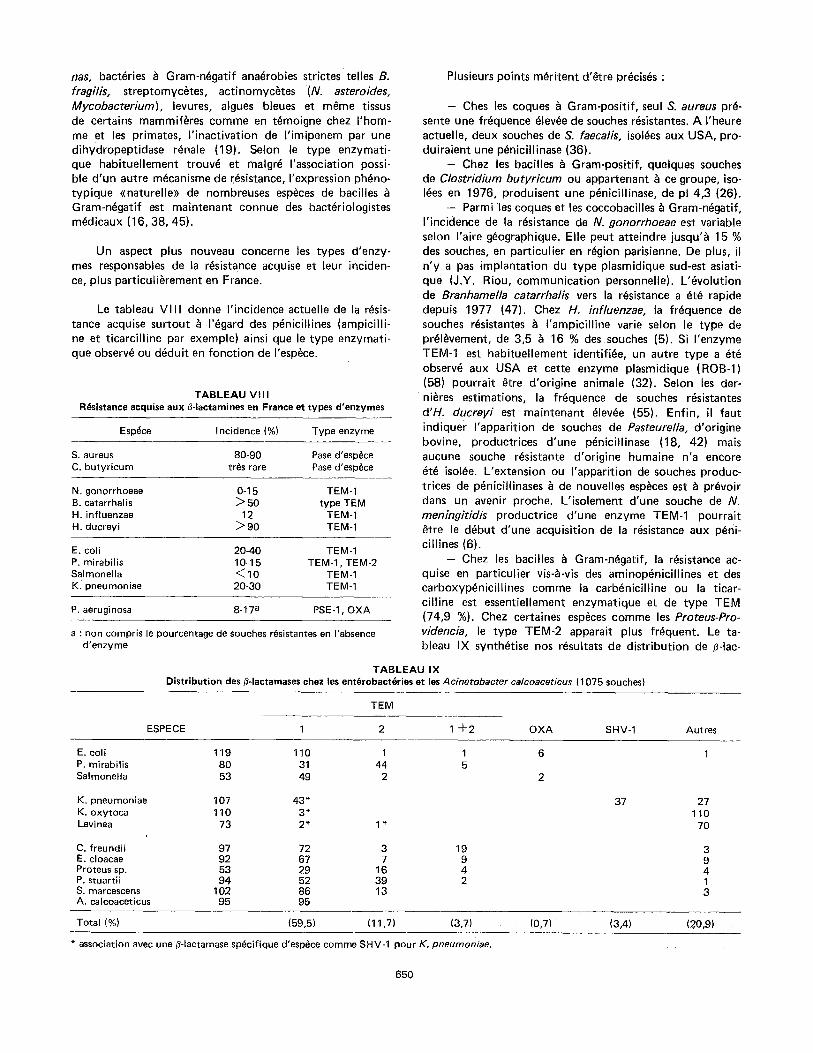
Le tableau V I I I donne I'incidence actuelle de la resis- tance acquise surtout ~ I'egard des p~nicillines (ampicil l i- ne et t icarcil l ine par exemple) ainsi que le type enzymati- que observe ou deduit en fonct ion de I'esp~ce.
T A B L E A U VIII R(~sistance acquise aux fl-lactamines en France et types d'enzymes
Esp~ce I ncidence (%) Type enzyme
S. aureus 80-90 Pase d'esp~ce C. butyricum tr~s rare Pase d'esp~ce
N. gonorrhoeae 0-15 TEM-1 B. catarrhalis ~ 50 type TEM H. influenzae 12 TEM-1 H. ducreyi > 90 TEM-1
E. coil 20-40 TEM-1 P. mirabilis 10-15 TEM-1, TEM-2 Salmonella < 10 TEM-1 K. pneumoniae 20-30 TEM-1
P. aeruginosa 8-17 a PSE-1, OXA
a : non compris le pourcentage de souches r~sistantes en I'absence d 'enzyme
Plusieurs points m~ritent d'etre pr~cis~s :
- Ches les coques ~ Gram-positif, seul S. aureus pr~- sente une fr~quence ~lev~e de souches r~sistantes. A I'heure actuelle, deux souches de S. faecalis, isol~es aux USA, pro- duiraient une p~nicillinase (36).
- Chez les bacilles ~ Gram-positif, quelques souches de Clostridium butyricum ou appartenant ~ ce groupe, iso- I~es en 1976, produisent une p~nicillinase, de pl 4,3 (26).
- Parmi "les coques et les coccobacilles ~ Gram-n~gatif, I ' incidence de la r~sistance de N. gonorrhoeae est variable selon I'aire g~ographique. Elle peut atteindre jusqu'& 15 % des souches, en particulier en r~gion parisienne. De plus, il n 'y a pas implantat ion du type plasmidique sud-est asiati- que (J.Y. Riou, communicat ion personnelle). L'~volut ion de Branhamella catarrhalis vers la r~sistance a ~t~ rapide depuis 1977 (47). Chez H. influenzae, la fr6quence de souches r~sistantes & I 'ampicil l ine varie selon le type de pr~l~vement, de 3,5 ~ 16 % des souches (5). Si I'enzyme TEM-1 est habituel lement identifi~e, un autre type a ~t~ observe aux USA et cette enzyme plasmidique (ROB-l) (58) pourrait ~tre d'origine animale (32). Selon les der-
n i t r es estimations, la frequence de souches resistantes d'H. ducreyi est maintenant ~lev~e (55). Enfin, il faut indiquer I 'apparit ion de souches de Pasteurella, d'origine bovine, productrices d'une penicillinase (18, 42) mais aucune souche r~sistante d'origine humaine n'a encore ~t~ isol~e. L'extension ou I 'apparit ion de souches produc- trices de p~nicitlinases ~ de nouvelles esp~ces est ~ pr~voir dans un avenir proche. L' isolement d'une souche de N. meningitidis productrice d'une enzyme TEM-1 pourrait #}tre le d~but d'une acquisition de la r~sistance aux p~ni- cillines (6).
- Chez les bacilles ~ Gram-n~gatif, la resistance ac- quise en particulier vis-a-vis des aminop~nicil l ines et des carboxyp~nicitl ines comme la carb~nicill ine ou la ticar- cil l ine est essentiellement enzymatique et de type TEM (74,9 %). Chez certaines esp6ces comme les Proteus-Pro- videncia, le type TEM-2 apparait plus frequent. Le ta- bleau IX synth~tise nos r~sultats de distr ibution de /]-Iac-
TABLEAU IX Distribution des ~-Iactamases chez les ent6robact6ries et les Acinetobacter calcoaceticus (1075 souches)
TEM
ESPECE 1 2 1 + 2 OXA SHV-1 Autres
E. coil 119 110 1 1 6 1 P. mirabilis 80 31 44 5 Salmonella 53 49 2 2
K. pneumoniae 107 43* 37 27 K. oxytoca 110 3* 110 Levi nea 73 2" 1 * 70
C. freundii 97 72 3 19 3 E. cloacae 92 67 7 9 9 Proteus sp. 53 29 16 4 4 P. stuartii 94 52 39 2 1 S. marcescens 102 86 13 3 A. calcoaceticus 95 95
Total (%) (59,5) (11,7) (3,7) (0,7) (3,4) . . . . . (?0,9)
* association avec une fl-lactamase sp~cifique d'esp~ce comme SHY-1 pour K. pneumoniae.
650
tamases pour des souches resistantes ~ la carb~nicil l ine iso- lees en 1977 et en 1978 au CHU Cochin, ~ I 'exception des Salmonella (41). Aucune souche ~ cette p~riode ne mon- trai t une activite c~phalosporinase excessive. Pour I'esp~ce P. aeruginosa, predominent d'autres types tels PSE-1 ou CARB-2 et differentes oxaciltinases (OXA) (44, 48). Le meme type de dist r ibut ion a et~ observe en Grande-Bre- tagne (67). On remarquera I 'apparit ion de nouveaux types, encore limitee ~ quelques souches au-cours de ces derni~- res annees (25, 31, 44, 46, 57, 59). Ces differentes enzy- mes de type penicill inase (LCR-1, NPS-1, CARB-4, OXA- 4), bien que distinguables par certains crit~res d' identi f i - cation precedemment indiqu~s permettant une epidemio- Iogie fine, ont un r61e apparemment semblable vis-a-vis des fl-lactamines (tableau VI I I ) . Nous n'avons pas assez de renseignements en ce qui concerne les bacilles ~ Gram- n~gatif anaerobies stricts, du moins en France. Des peni- ciliinases ont ete recemment decrites chez F. nucleatum (65), de meme qu'une fl-lactamase hydrolysant la c~foxi- t ine et transferable chez B. fragilis.
- L'aspect recent de la r~sistance acquise aux ~-Iac- tamines non hydrolysables comme les cephalosporines de 3~me g~neration concerne I'emergence de souches der~- primees pour la product ion d'une c~phalosporinase (8, 17, 37, 54). Les caract6ristiques de cette resistance cons- t i tut ive sont les suivantes :
• au cours ou apr6s prescription de fl-lactamines large spectre
• quelquefois au cours d'une association avec un autre ant ibiot ique
• emergence chez le malade en quelques jours • bacilles ~ Gram-negatif de phenotype cephalospo-
rinase inductible (E. cloacae, S. marcescens, C. freundii, Proteus indol 4-, P. aeruginosa)
• selection de mutants consti tut i fs (taux de muta- t ion 10 "s ~ 10 -7 )
• biosynth~se de fl-lactamase augmentee de 100- 1000 fois
• hydrolyse faible, di f f ic i le & visualiser de differen- tes fl-tactamines
• resistance crois~e entre /~-Iactamines, sauf excep- t ions (tableau V I I I )
• incidence non negligeable en France, du moins dans certains h6pi taux (16, 38, 48, 61).
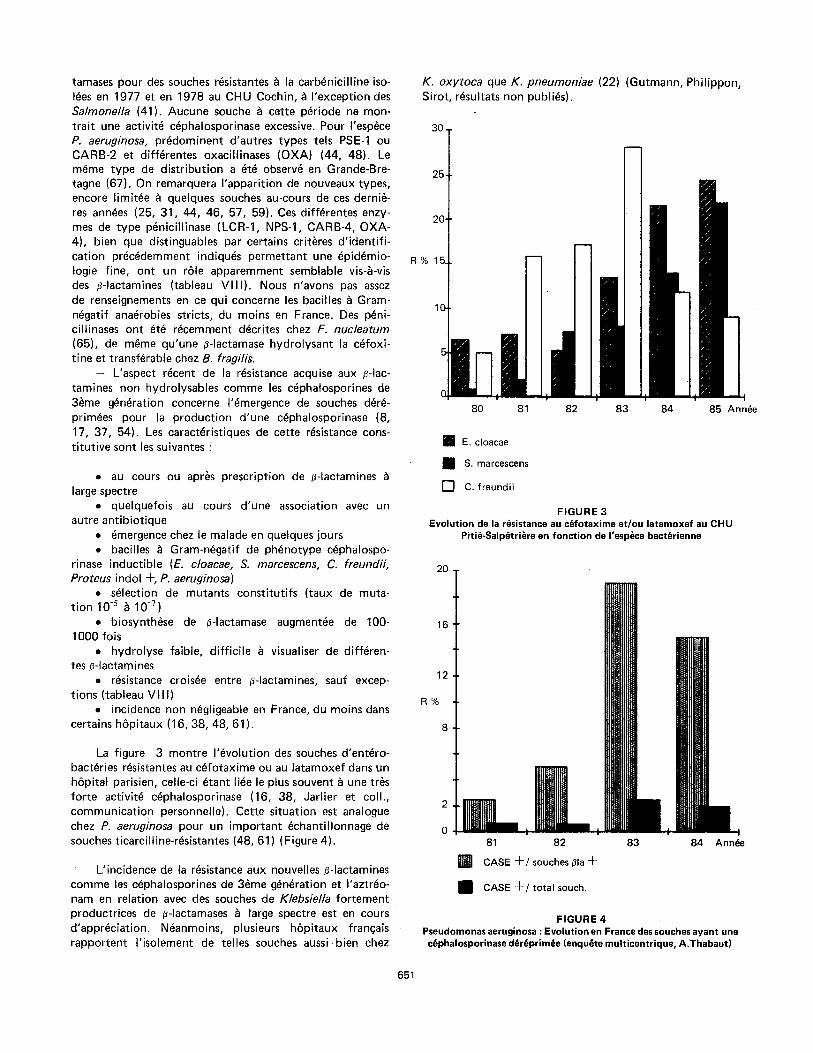
La figure 3 montre I 'evolution des souches d'entero- bact~ries r~sistantes au cefotaxime ou au latamoxef dans un h6pital parisien, celle-ci etant li~e le plus souvent ~ une tr~s forte activite cephalosporinase (16, 38, Jarlier et coll., communicat ion personnetle). Cette situation est analogue chez P. aeruginosa pour un important ~chantil lonnage de souches ticarciltine-resistantes (48, 61) (Figure 4).
L'incidence de la r~sistance aux nouvelles #-Iactamines comme les cephalosporines de 3~me generation et I'aztreo- nam en relation avec des souches de Klebsiella fortement productrices de ~-Iactamases & large spectre est en cours d'appr~ciation. Neanmoins, plusieurs h6pi taux francais rapportent I' isolement de telles souches aussi b i en chez
K. oxytoca que K. pneumoniae (22) (Gutmann, Phil ippon, Sirot, r~sultats non publi~s).
30.
25.
20"
R%15.
t iJ d L
80 81 82 83 84 85 Annie
IN E. cloacae
[ ] S. marcescens
[ ] C. freundii
FIGURE 3 Evolution de la r~sistance au c~fotaxime et/ou latarnoxef au CHU
Piti~-Salp~tri~re en fonction de l'esp~ce bact~rienne
20
16
12
R%
8
81 82
~ l CASE Jr-/souches/~la 4-
i CASE 4-/total souch.
83 84 Annie
F IGURE 4 Pseudomonas aeruginosa :Evolution en France des souches ayant une
c~phalosporinase d6r~prim~e (enqu~te multicentrique, A.Thabaut)
651

CONCLUSIONS - PERSPECTIVES
Le premier m~canisme de r~sistance individualis~, tant dans le cadre de la r~sistance naturelle qu'acquise, a montr~ I ' intervent ion de fl-lactamases. Par la suite, le rble des p~ni- cillinases plasmidiques ou transposables chez les bacilles Gram-n~gatif (TEM-1 par exemple) a r~v~l~ leur pouvoir ~pid~mique. La d~couverte de nouvelles ~-Iactamines r~- put~es non hydrolysables est suivie actuel lement de I'~mer- gence de souches r~sistantes &ces antibiot iques. Le m~ca- nisme de r~sistance individualis# est en relation avec la synth~se d~r~gul~e de c~phalosporinase. Cette r~sistance const i tut ive est caract~ris~e par un spectre d ' inact ivat ion
plus large que celui des penicillinases plasmidiques. Serait-ce une adaptat ion ~ une pression de s~lection dif f~rente ? L' in- dividual isat ion d'autres enzymes de type p~nicill inase, de ~-Iactamases ~ large spectre, en part icul ier par mutat ion, de CXases ou encore leur extension aux bact~ries ana~robies strictes montre les ressources 6volutives de la r~sistance en- zymat ique aux fl-lactamines dans le monde bact~rien, en par- t icul ier chez les esp~ces ~ Gram-n~gatif, bien que les autres m~canismes connus pourraient ~tre plus importants & I'avenir comme peut-~tre pour la r~sistance ~ I ' imipenem (49).
REMERCIEMENTS Les auteurs de cette revue adressent leurs remercie-
ments ~ Pierre N6vot pour les critiques et l'aide apport6e /l la r6alisation de ce travail.
SUMMARY B-lactamases : identification, incidence, epidemiology
Originally, the ~-Iactamases were the first mechanism of ~-Iactam resistance identified for nature/resistance such as inducible cephalosporinase produced by gram-negative rods and
lately, in acquired resistance mediated by penicillinase of S. aureus. With the discovery of broad-spectum ~-Iactams such as ampicillin and cephalosporins (cephalothin for example) the plasmid-mediated penicilli- nases of gram-negative rods had emerged, mostly due to the ~-Iactamase TEM-type. The epidemiological role of these enzymes was explained because most of them were transposable. This genetic determinism is implicated in the recent spread to other susceptible species, including N. gonnorheae, H. influenzae, H. ducreyi, and probably N. meningitidis. Comparatively, with the advent of poor inducers and unhydrolyzable ~-Iactam antibiotics, the novel resis- tance status was correlated to the predisposition of several cephalosporinase-inducible species to emerge by selection of derepressed or constitutive variants producing high levels of this ~-/actamase. The pattern of susceptibility was deeply modified towards the unhydrolyzable ~-Iactam antibiotics such as third-genera- tion cephalosporins and monobactam. Very recently, some reports in several countries focused on a diffe- rent modality of resistance related to the broad-spectrum ~-Iactamases already identified in several species but high/y produced by a few strains of Enterobacteriaceae such as Klebsiella (K. oxytoca, K. pneumoniae). Another characteristic of this type of enzyme was the acquisition of an extented spectrum to new ~-Iactam antibiotics, probably by mutation in one case (SHV-2). Among bacteria, some of them may show a poten- tial capacity to adapt to newer and abusively prescribed antibiotics by selection pressure, and the resistance is perpetuated because of inefficient rules of hygiene. We must always survey the emergence of unusual resistant isolates due to a different mechanism as previously identified but the enzymatic inactivation ap- pears always predominant.
Key-words : Hactamases- Identification - Classification - Incidence - Distribution
1.
2.
3,
4.
5.
B I B L I O G R A P H I E
AMBLER R.P. -- The structure of ~-Iactamases. Trans. R. Soc. Lond. (Biol.), 1980, 289, 321-331
BRYAN L.E. -- Antimicrobial drug resistance. Academic Press Inc., Harcourt Brace Jovanich, Publishers, 1984
BUSH K., SYKES R.B. -- Methodology for the study of j3-1ac- tamases. Antimicrob. Agents Chemother., 1986, 30, 6-10
CURTIS N.A.C., EISENSTADT R.L., RUDD C., WHITE A.J.-- Inducible type I ~-Iactamases ef Gram-negative bacteria and re- sistance to /~-Iactam antibiotics. J. Antimicrob. Chemother., 1986, 17, 51-61
DABERNAT H., DELMAS C., LARENG M.B. -- Prevalence de la r~sistance aux antibiotiques des Haemophilus influenzae
6.
7.
8 .
isol~s en France. Un an d'activit~ du r~seau de surveillance des infections & Haernophilus influenzae. Path. Biol.; 1986, 34, 297-540
DILLON J.R., PAUZE M., YEUNG K.M. -- Spread of penicilli- nase-producing and transfert plasmids from the gonococcus to Neisseria meningitidis. Lancet, 1983, 1, 532-535
FOSTER T . J . - - Plasmid-determined resistance to antimicrobial drugs and toxic metal ions in bacteria. Microbiol. Rev., 1983, 47, 361-409
GOERING R.V., SANDERS C.C., SANDERS F.W. -- Inducible /3-1actamases and /~-Iactam resistance. Proc. IX Inter. Cong. Inf. Parasit. Dis., 130-134, Munich, 1986
652

9 . G U T M A N N L . - - M~canismes d'action des #-Iactamines. J. Enseign. Post-Univ. Piti#-Salp#tri#re, 1985, pp. 87-99
10. GUTMANN L., WILLIAMSON R., COLLATZ E . - A possible role of porins in bacterial ant ibiot ic resistance. Ann. Intern. Med., 1984, 101, 554-557
11. HART C.A., PERCIVAL A. -- Resistance to cephalosporins among gentamici~ resistant Klebsiellae. J. Antimicrob. Chemo- thor., 1982, 9, 275-286
12. HEDGES R.W., JACOB A.E. -- Transposition of ampicillin re- sistance from RP4 from RP4 to other replicons. Mol. Gen. Genet., 1974, 132, 31-40
13. HEDGES R.W., MEDEIROS A.A., COHENFORD M., JACO- BY G . A . - - Genetic and biochemical properties of AER-1, a novel carbenicil l in-hydrolyzing #4actamase from Aeromonas hydrophila, Antimicrob. Agents Chemother., 1985, 27, 479- 484
14. JACOBY G.A., SUTTON L. -- Act ivi ty of #-Iactam antibiotics against Pseudomonas aeruginosa carrying R plasmids determi- ning dif ferent #-Iactamases. Antimicrob. Agents Chemother., 1979, 16, 243-245
15. JACOBY G.A., SUTTON L. -- #-Iactamases and #-Iactam resis- tance in Escherichia coll. Antimicrob. Agents Chemother., 1985, 28, 703-705
16. JARLIER V., BISMUTH R., NICOLAS M.H., N'GUYEN J., TRUFFOT C., GROSSET J. -- Survey of the phenotypes of susceptibility to #-Iactams in Enterobacteriaceae of the Piti~- Salp~tri6re hospital. J. Antimicrob. Chemother., 1984, 14, sup- pl. B 59-65
17. JARLIER V., PHILIPPON A., NICOLAS M.H., BISMUTH R., PAUL G., FUSCIARDI J. -- Enterobactercloacae : Emergence in vivo d'un variant r6sistant aux nouvelles #-Iactamines Iors d'un traitement lamoxactam-gentamicine. Path. Biol., 1984, 32, 399-403
18. JOLY B., MARTEL J.L., MICHEL R., REYNAUD A . ,CLU- ZEL R . - - Sensibilit~ aux antibiotiques et production de #-Iac- tamase chez les souches de Pasteurella d'origine bovine isolees en France. M6d. Mal. Inf., 1986, Sp6cial Mars, 52-56.
19. KAHAN F.M., KROPP H., SUNDELOF J.G., BIRNBAUM J. Thienamycin : development of imipenem6ilastin. J. Antimi- crobiol. Chemother., 1983, 12, suppl. D, 1-35
20. KLIEBE C., NIES B.A., MEYER J.F., TOLSDORFF-NEUTZ LING R.M., WIEDEMANN B. -- Evolution of plasmid-coded resistance to broad-spectrum cephalosporins. Antimicrob. Agents. Chemother., 1985, 28, 302-307
21. LABIA R., GUIONIE M., BARTHELEMY M., PHILIPPON A. -- Properties of three carbenicill in-hydrolysing #-Iactamases (CARB) from Pseudomonas aeruginosa : identif ication of a new enzyme. J. Antimicrob. Chemother., 1981,7, 49-56
22. LABIA R., MORAND A., GUIONIE M., HEITZ M., PITTON J.S. - Betalactamases d~ Klebsiella oxytoca : Etude de leur action sur les c~phalosporines de troisi~me g~n6ration. Path. Biol., 1986, 34, 611-615
23. LEVESQUE R.P., ROY P . H . - Mapping of a plasmid (pLQ3) from Achromobacter and cloning of its cephalosporinase gene in Escherichia coil Gene., 1982, 18, 69-75
24. LEVESQUE R., ROY P., LETARTE R., PECHERE J . C . - A plasmid-mediated cephalosporinase f rom Achromobacter species. J, Inf. Dis., 1982, 145,753-761
25. LIVERMORE D.M., JONES C.S. - Characterization of NPS-1, a novel plasmid-mediated/3-1actamase, from two Pseudomonas aeruginosa isolates. Antimicrob. Agents Chemother., 1986, 29, 99-103
26. MAGOT M. -- Some properties of the Clostridium butyricum group #-Iactamase. J. Gen. Microbiol., 1981, 127, 99-103
27. MALOUlN F., BRYAN L.E. -- Modif ication of penicill in-bin- ding proteins as mechanisms of #-Iactam resistance. Antimicrob. Agents Chemother., 1986, 30, 1-5
28. MANIAN F.A., ALFORD R.H. - Discrepancies between disk diffusion and broth susceptibility studies of the activity of ticarcillin plus clavulanic acid against ticarcillin-resistant Pseu- domonas aeruginosa. Antimicrob. Agents Chemother., 1986, 30, 35-38
29. MATTHEW M. -- Plasmid-mediated fl-lactamases of gram nega- tive bacteria : properties and distribution. J. Antimicrob. Che- mother., 1979, 5, 349-358
30. MEDEIROS A.A. -- /3-1actamases. Brit. Med. Bull., 1984, 40, 18-27
31. MEDEIROS A.A., COHENFORD M., JACOBY G.A. -- Five novel plasmid determined #-Iactamases. Antimicrob. Agents Chemother., 1985, 2, 715-719
32. MEDEIROS A.A., LEVESQUE R., JACOBY G.A. -- An ani- mal source for the ROB-1 #-Iactamase of Haemophilus influen- zae type b. Antimicrob. Agents Chemother., 1986, 29, 212- 215
33. MINAMI S., YOTSUJI A., INOUE M., MITSUHASHI S . - Induction of #-Iactamase by various #-Iactam antibiotics in Enterobacter cloacae. Antimicrob. Agents Chemother., 1980 18, 382-385
34. MITSUHASHI S., YAMAGISHI S., SAWAI T., KAWABE H. Biochemical mechanisms of plasmid-mediated resistance.In R factor drug resistance plasmid ; edited by S. Mitsuhashi, University of Tokyo Press, 1977, 195-251
35. M U R A Y A M A S.Y., YAMAMOTO T., SUZUKI I., SAWAI T. Mutation of Escherichia coil capable of expressing gene(s) for #-Iactamase production of Citrobacter freundiL Antimicrob. Agents Chemother., 1986, 29, 707-709
36. MURRAY B.E., MEDERSKI-SAMAROJ B. -- Transferable #-Iactamase : A new mechanism for in vitro penicillin resistan- ce in Streptococcus faecalis. J. Clin. Invest., 1983, 72, 1168- 1171
37. NAUCIEL C., PHILIPPON A., RONCO E., PILLIOT J., GUE- NOUNOU M., PAUL G., BRUNEL D., OUTIN H.D. -- Septi- c~mies ~ Enterobacter cloacae et E. aerogenes : Emergence de variants r~sistants (c6phalosporinase d~r~prim~e) en cours de traitement par des c6phalosporines de troisi~me g6n~ration. Presse M#d., 1985, 14, 673-676
38. NICOLAS M.H., JARLIER V., FIEVET M.H., N'GUYEN J., TRUFFOT C . - - Situation de la r6sistance des ent~robact~ries au c~fotaxime et lamoxactam au groupe hospitalier Piti~-Sal- p~tri6re. Pat& Biol., 1984, 32, 393-395
39. OUELLETTE M., ROY P.H. -- Analysis by using ONA probes of the OXA-1 #-Iactamase gene and its transposon. Antimicrob. Agents Chemother., 1986, 30, 46-51
40. PHILIPPON A. -- Les #-Iactamases. Techn. Bio1.,1983, 2, 80-84
653

41. PHILIPPON A., FOURNIER G., CORNEL E., PAUL G., LE. MINOR L., NEVOT P. - - Les/3-tactamases de Salmonella r~sis-. tantes ~ I'ampicilline. Ann. Microbiol. (Inst. Pasteur), 1984, 135A, 229-238
42. PHILIPPON A., JOLY B., REYNAUD D., PAUL G., MAR- TELL J.L., SIROT D., CLUZEL R., NEVOT P. - Characteri- zation of a ~3-1actamase f rom Pasteurella multocida. Ann. Microbiol. (Inst. Pasteur), 1986, 137A, 153-158
43. PHILIPPON A., PAUL G., JACOBY G.A. -- Properties of PSE-2 /3-1actamase and genetic for its production in Pseudo- monas aeruginosa. Antimicrob. Agents Chemother., 1983, 24, 362-369
44. PHILIPPON A.M., PAUL G.C., JACOBY G.A. - New plas- mid mediated oxacil l in hydrolyzing /3-1actamase in Pseudo- monas aeruginosa. J. Ant imicrob. Chemother., 1986, 17, 415-422
45. PHILIPPON A., PAUL G., NEVOT P. - M~canisme de r6sis- tance enzymatique aux ~-Iactamines. Presse M~d., 1986, sous presse
46. PHILIPPON AI, PAUL G., THABAUT A.,-JACOBY G.A. -- Properties of a novel carbenicil l in-hydrolyzing /3-1actamase (CARB-4) specified by a P2-plasmid from Pseudomonas aeru- ginosa. Antimicrob. Agents Chemother., 1986, 29, 519-520
47. PHILIPPON A., RIOU J.Y., GUIBOUDENCHE M., SOTO- LONGO F. -- ~-Iactamases of Branhametla catarrbalis : Detec- t ion -- Frequency -- Inhibit ion. Drugs, 1986, 31, suppl. 3, 64-69
48. PHILIPPON A., THABAUT A., MEYRAN M., NEVOT P. -- Distribution des ~-Iactamases constitutives chez P. aeruginosa. Presse M~d., 1984, 13, 772-776
49. QUINN J.P., DUDEK E.J., DIVINCENZO C.A., LUCKS D.A., LERNER S.A . - - Emergence of resistance to imipenem during therapy for Pseudomonas aeruginosa infections. J. Inf. Dis., 1986, 154, 289-294
50. REID A.J., AMYES G.B. - Plasmid penicillin resistance in Vibrio cholerae : Identif ication of new ~-Iactamase SAR-I. Antimicrob. Agents Chemother., 1986, 30, 245-247
51. RICHMOND M.H., SYKES R.B. -- The/3-1actamases of gram- negative bacteria and their possible phy~ological role. In Rose A.H., Tempest D.W., eds. Adv. Microb. Physiol., 1973, 9, 31-88
52. RUBIN L.G., MEDEIROS A.A., YOLKEN R.H., MOXON E.R. Ampicil l in treatment failure of apparently/3-1actamase-negative Haemophilus influenzae type b meningitis due to novel/3-1acta- mase. Lancet, 1981, i i, 1008-1010
53. SAINO Y., INOUE M., MITSUHASHI S. - Purification and properties of an inducible cephalosporinase from Pseudomo- nas maltophil ia. Ant imicrob. Agents "Chemother., 1964, 25, 362-365
54. SANDERS C.C., SANDERS F.W. -- Emergence of resistance during therapy with the newer /~-Iactam antibiotics : role of inducible ~-Iactamases and implications for the future. Rev. Infect. Dis., 1983, 5, 639-648
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
SANSON-LE PORS M.J., CASIN I., THEBAULT M.C., PEROI- Y. -- Evolution de la r~sistance aux antibiotiques de 192 sou- ches d'Haemophilus ducrevi isol~es ~ Paris entre 1976 et 1984 et d~termination du contenu plasmidique de 15 soL~ches. 2~me Cong. Mond. MST, Paris, 1986
SATO K., FUJII T., OKAMOTO R., INOUE M., MITSUHASHI S. -- Biochemical properties of/3-1actamase produced by Flavo- bacterium odoratum. Antimicrob. Agents Chemother., 1985, 27, 612-614
SHLAES D.M., MEDEIROS A.A., KRON M.A., CURRIE-Mc CUMBER, PAPA E., VARTIAN C . V . - Novel plasmid-media- ted /~-Iactamase in members of the family Enterobactericeae f rom Ohio. A'ntimicrob. Agents Chemother., 1986, 30, 220- 224
SIMPSON I.N., HARPER P.B., O 'CALLAGHAN C.H. -- Princi- pal /~-Iactamases responsible for resistance to ~-Iactam antibio- tics in urinary tract infections. Antimicrob. Agents Chemother. 1980, 17, 929-936
SIMPSON N., PLESTED S.J., BUDIN-JONES M.J., JEES J., HEDGES R.W., JACOBY G.A. -- Characterization o:f a novel plasmid-mediated beta-lactam resistance in Pseudomonas aeru- ginosa. FEMS Microbiol. Lett., 1983, 19, 23-27
SYKES R.B., MATTHEW M. -- The /3-1actamases of Gram-ne- gative bacteria and their role in resistance to /3-1actam anti- biotics. J. Antimicrob. Chemother., 1977, 2, 115-157
THABAUT A., PHILIPPON A., MEYRAN A. -- Activit~ com- par~e des /3-!actamines actives sur Pseudomonas aeruginosa en fonct ion des ph~notypes de r~sistance. Presse M~d., 1984, 13, 768-771
THABAUT A., MEYRAN A., PHI LIPPON A. --Pseudomonas aeruginosa - In vitro emergence of resistance to/3-1actams (car- boxy, ureido-penicillins and third generation cephalosporins), 13th Inter. Cong. Chemother., 1983, Proc. 90, 35-42
THEN L.R., ANGHER P. -- Trapping of non hydrolysable ce- phalosporins by cephalosporinase in Enterobacter cloacae and Pseudomonas aeruginosa as a possible resistance mechanism. Antimicrob. Agents Chemother., 1982, 21, 711-717
THEN L.R., GLAUSER M.P., ANGHER P., ARISAWA M -- Cephalosporin resistance in strains of Klebsiella oxytoca iso- lated during antibiot ic therapy. Zbl. Bakt. Hyg., I. Abt. Orig. A, 1983, 254, 469-479
TUNER K., LINDQVIST L., NORD C.E.,Characterization of a new ~-Iactamase f rom Fusobacterium nucleatum by substrate profiles and chromatofocusing patterns. J. Antimicrob. Chemo- then, 1985, 1G, 23-30
VU H., NIKAIDO H. -- Role of/3-1actam hydrolysis in the me- chanism of resistance of a f3-1actamase constitutive Enterobac- ter cloacae strain to expanded-spectrum/~-Iactams. Antimicrob. Agents Chemother., 1985, 27, 393-398
WILLIAMS R.J., LIVERMORE D.M., LINDRIDGE M.A., SAID A.A., WILLIAMS J.D. -- Mechanisms of beta-lactam re- sistance in British isolates of Pseudomonas aeruginosa, J. Med. Microbiol., 1984, 17, 283-293
654