Embed Size (px)
Citation preview

4/5/2018
1
BIOLOGY AND ECOLOGY OF
CIGUATOXINS PRODUCERS
Chui Pin LEAW PHD
IOES, University of Malaya
Harmful benthic dinoflagellates
Coolia
Gambierdiscus
Ostreopsis
Prorocentrum
Fukuyoa
Amphidinium

4/5/2018
2
e.g. Yasumoto et al. 1979, 1987, 2005; Chambell et al. 1987; Chinain et al. 1999 and many more…
O u t l i n e
Taxonomy and biodiversity
Eco-physiology

4/5/2018
3
Gambierdiscus toxicus Adachi et Fukuyo 1979
Adopted from Adachi & Fukuyo 1979
Courtesy of J. Larsen Sousa & Silva 1956
4/5/2018
4
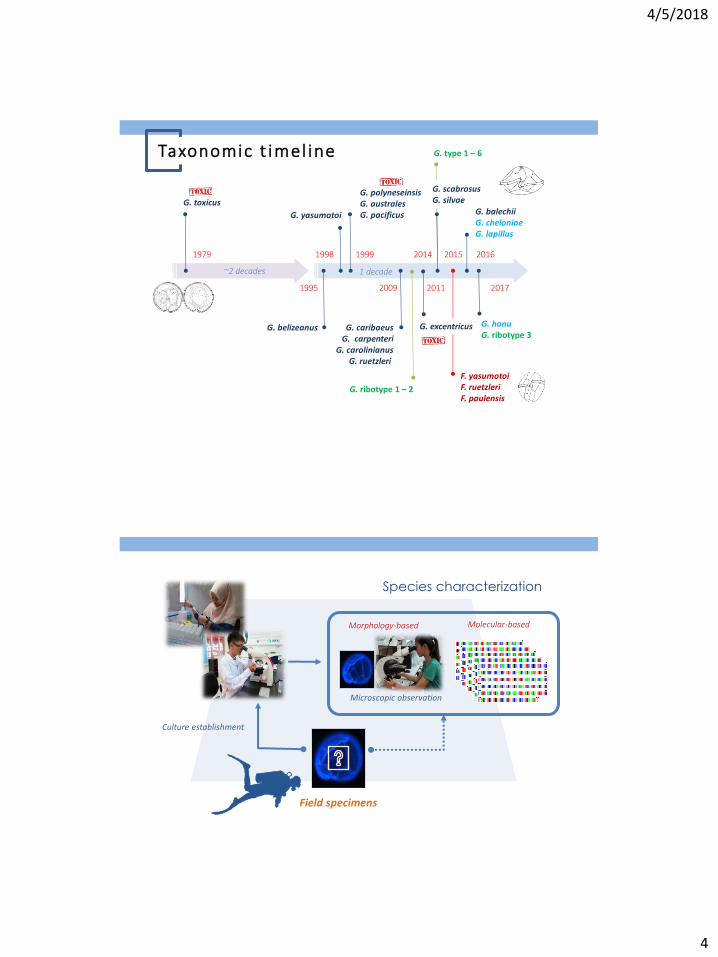
Taxonomic t imel ine
G. toxicus
1995
G. belizeanus
1979 1998 1999
G. polyneseinsis G. australes G. pacificus G. yasumotoi
1 decade ~2 decades
2009
G. caribaeus G. carpenteri
G. carolinianus G. ruetzleri
2011
2015 2016
2017
G. balechii G. cheloniae G. lapillus
G. honu G. ribotype 3
G. excentricus
2014
F. yasumotoi F. ruetzleri F. paulensis
G. scabrosus G. silvae
G. type 1 – 6
G. ribotype 1 – 2
Morphology-based Molecular-based
Species characterization
Culture establishment
Field specimens
Microscopic observation
4/5/2018
5
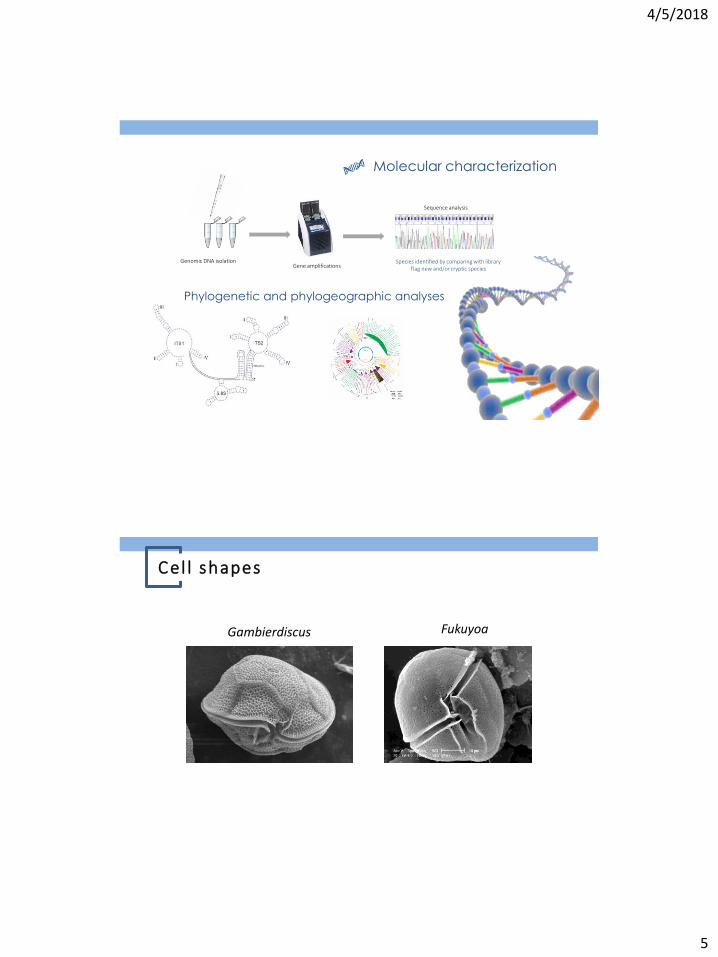
Molecular characterization
Sequence analysis
Species identified by comparing with library Flag new and/or cryptic species Gene amplifications
Phylogenetic and phylogeographic analyses
Genomic DNA isolation
Gambierdiscus Fukuyoa
C el l shapes
4/5/2018
6
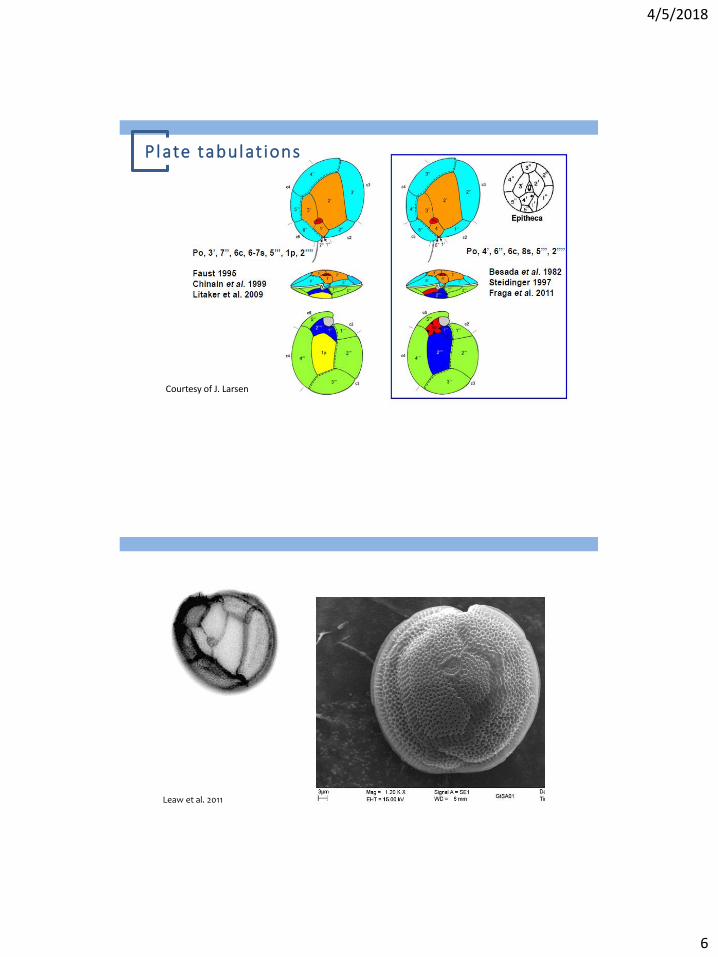
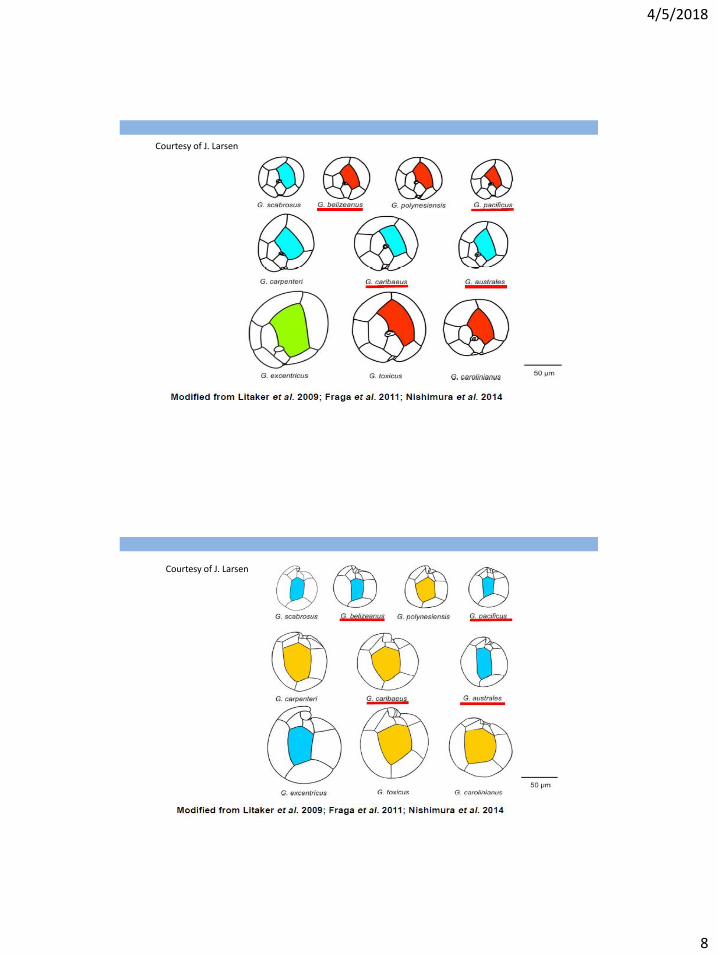
Courtesy of J. Larsen
Plate tabulat ions
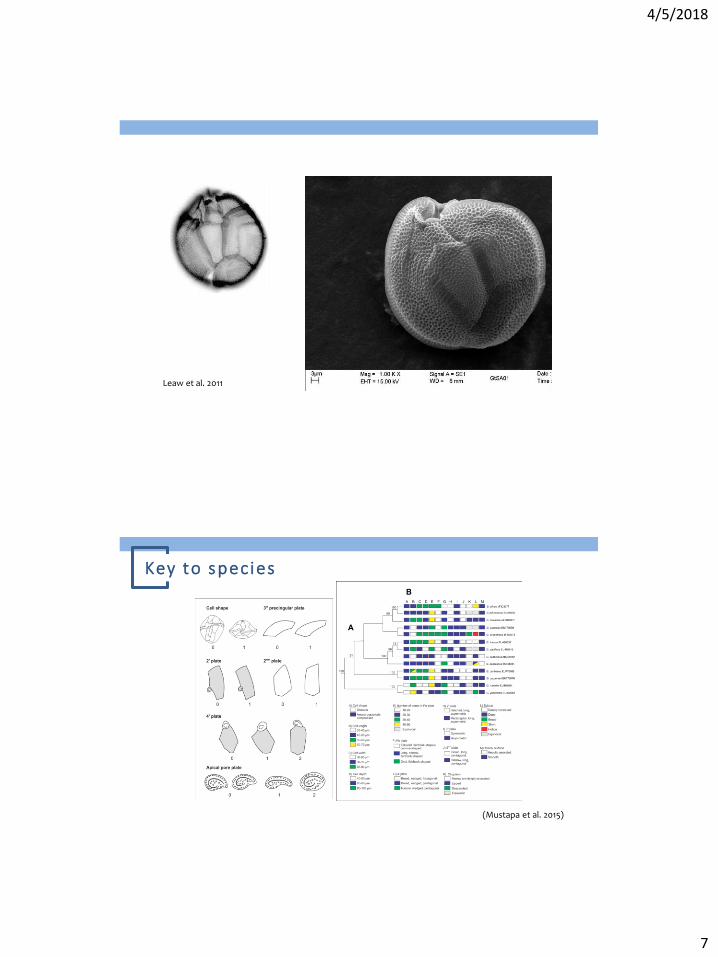
Leaw et al. 2011
4/5/2018
7
Leaw et al. 2011
(Mustapa et al. 2015)
Key to species
4/5/2018
8
Courtesy of J. Larsen
Courtesy of J. Larsen
4/5/2018
9
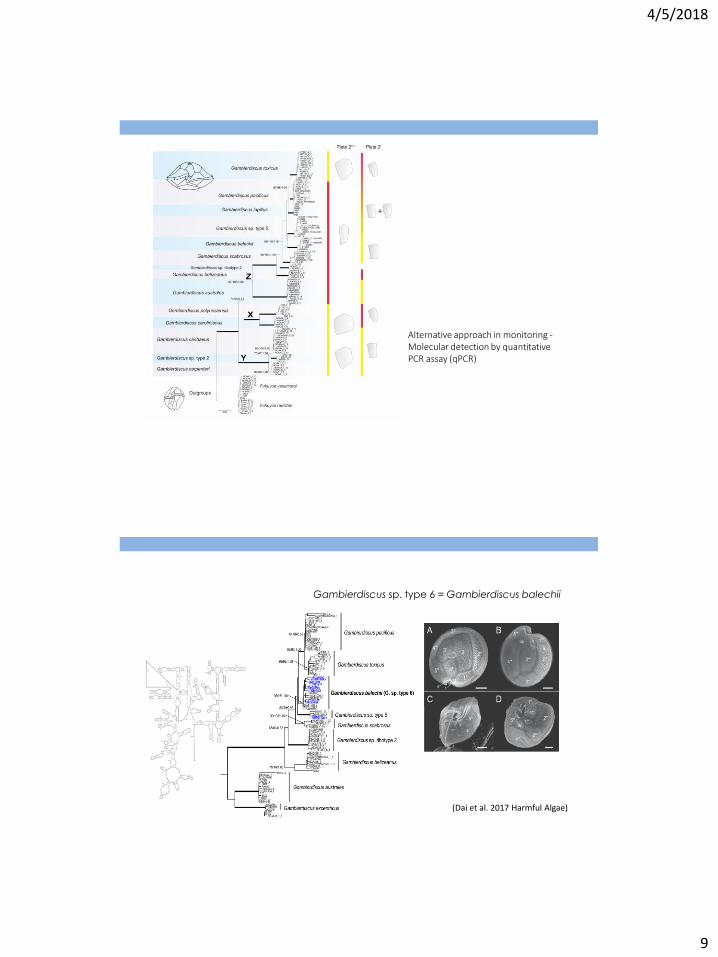
Alternative approach in monitoring - Molecular detection by quantitative PCR assay (qPCR)
Gambierdiscus sp. type 6 = Gambierdiscus balechii
(Dai et al. 2017 Harmful Algae)

4/5/2018
10
L i fe h istory
Bravo et al. 2014
Dividing cells were mostly in the cluster comprising the largest cells, indicating that mitosis takes place in diploid cells.
PHYSIOLOGICAL AND ECOLOGICAL ADAPTIONS OF CIGUATOXINS PRODUCERS

4/5/2018
11
Distr ibut ion
35 oN – 37 oS (Parson et al. 2012)
Adopted from Rhodes et al. 2017
4/5/2018
12
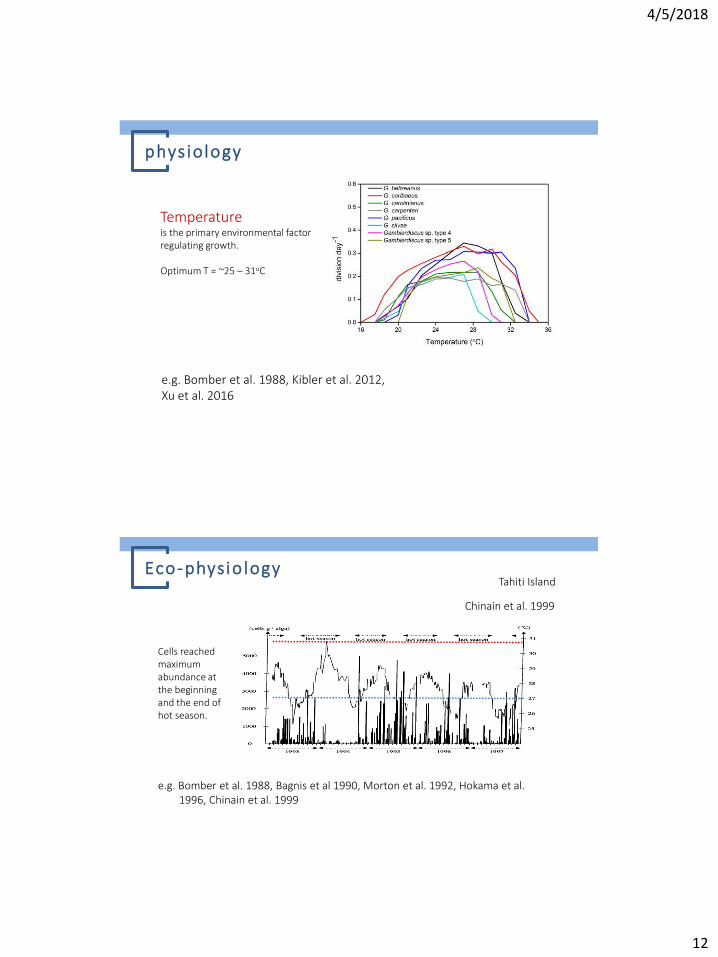
Temperature is the primary environmental factor regulating growth. Optimum T = ~25 – 31oC
phys iology
e.g. Bomber et al. 1988, Kibler et al. 2012, Xu et al. 2016
Eco-physio logy
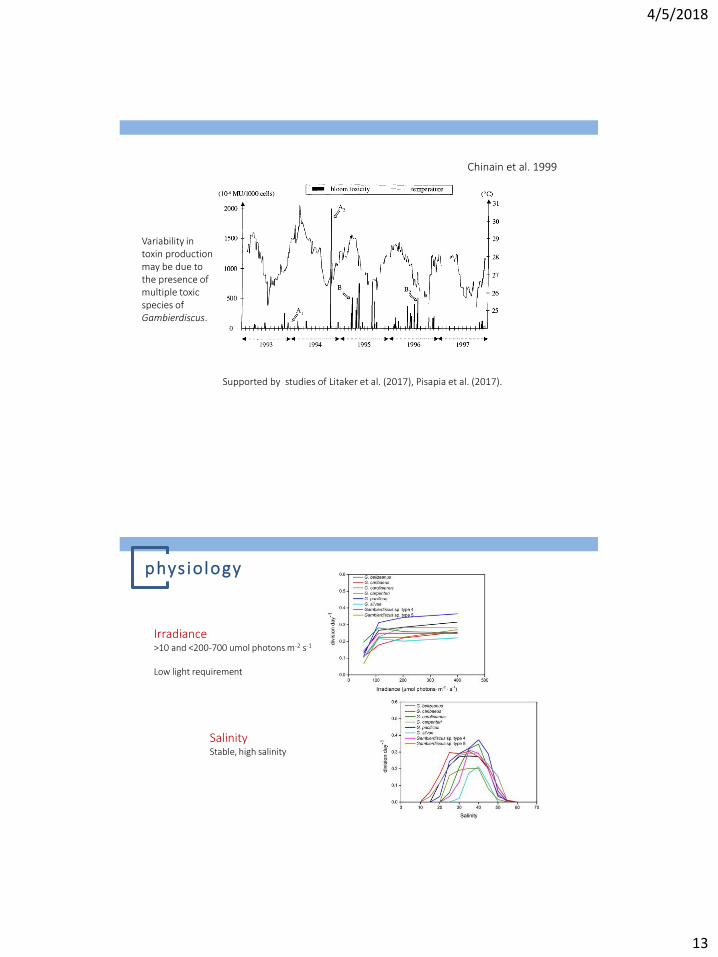
Chinain et al. 1999
Cells reached maximum abundance at the beginning and the end of hot season.
e.g. Bomber et al. 1988, Bagnis et al 1990, Morton et al. 1992, Hokama et al. 1996, Chinain et al. 1999
Tahiti Island
4/5/2018
13
Chinain et al. 1999
Variability in toxin production may be due to the presence of multiple toxic species of Gambierdiscus.
Supported by studies of Litaker et al. (2017), Pisapia et al. (2017).
Irradiance >10 and <200-700 umol photons m-2 s-1
Low light requirement
phys iology
Salinity Stable, high salinity

4/5/2018
14
BHAB species
Species diversity?
Healthy coral reef
Degraded coral reef
Alteration of trophic transfer
Local stresses
Climate changes
Disturbance Frequency HIGH
Reef Recovery LOW
Algal palatability HIGH
CFP vector shift
Degraded reefs
4/5/2018
15
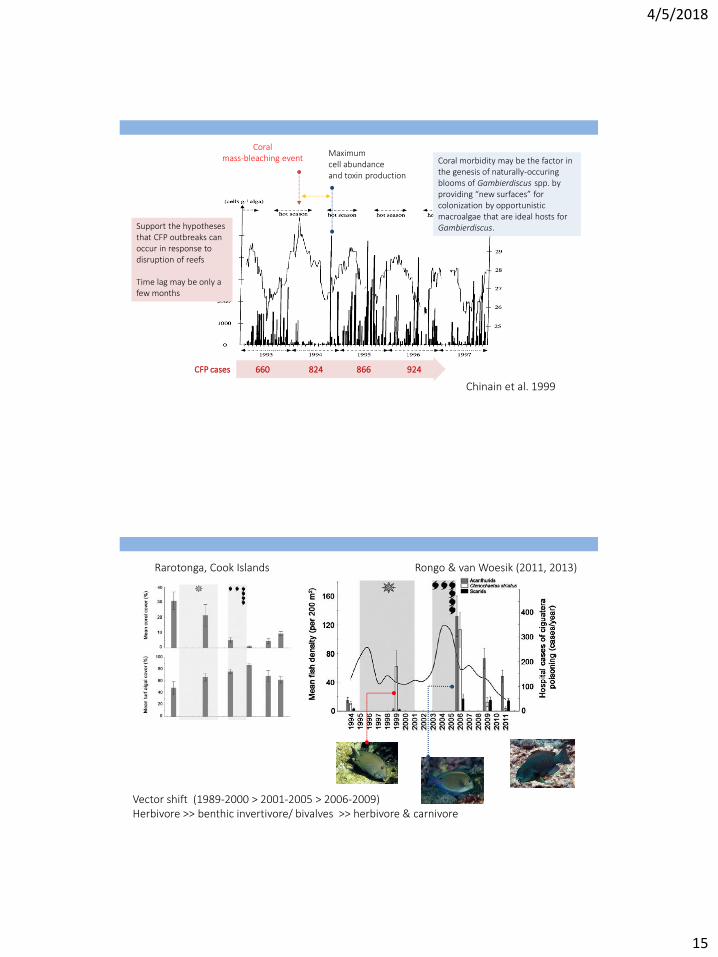
Chinain et al. 1999
Coral mass-bleaching event
660 824 866 924 CFP cases
Maximum cell abundance and toxin production
Coral morbidity may be the factor in the genesis of naturally-occuring blooms of Gambierdiscus spp. by providing “new surfaces” for colonization by opportunistic macroalgae that are ideal hosts for Gambierdiscus. Support the hypotheses
that CFP outbreaks can occur in response to disruption of reefs Time lag may be only a few months
Rongo & van Woesik (2011, 2013) Rarotonga, Cook Islands
Vector shift (1989-2000 > 2001-2005 > 2006-2009) Herbivore >> benthic invertivore/ bivalves >> herbivore & carnivore

4/5/2018
16
Non-destructive sampling approach
(Tester et al. 2012, 2014; Tan et al. 2013)
Reef surveys and sample collection
Underwater photoquadrat survey was performed at the sampling sites to record the percent coral coverage, variety of benthic substratum and the biotic inhabitants (e.g. fishes, invertebrates).
4/5/2018
17
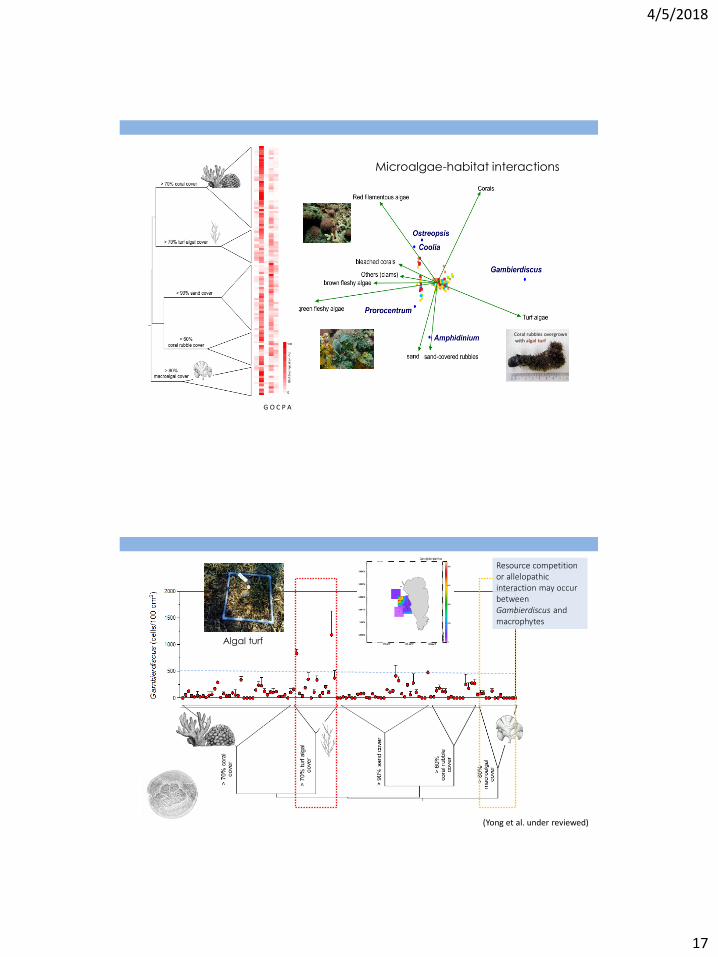
Microalgae-habitat interactions
Coral rubbles overgrown with algal turf
G O C P A
(Yong et al. under reviewed)
Resource competition or allelopathic interaction may occur between Gambierdiscus and macrophytes
Algal turf
4/5/2018
18
(Yong et al. under reviewed)
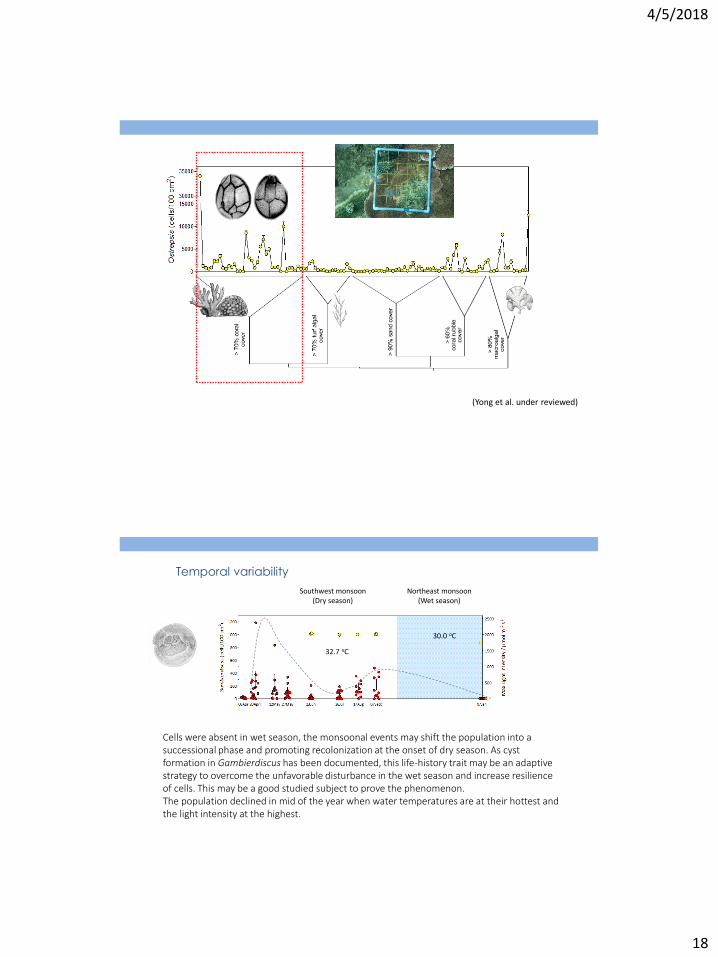
Northeast monsoon (Wet season)
Southwest monsoon (Dry season)
Temporal variability
32.7 oC
30.0 oC
Cells were absent in wet season, the monsoonal events may shift the population into a successional phase and promoting recolonization at the onset of dry season. As cyst formation in Gambierdiscus has been documented, this life-history trait may be an adaptive strategy to overcome the unfavorable disturbance in the wet season and increase resilience of cells. This may be a good studied subject to prove the phenomenon. The population declined in mid of the year when water temperatures are at their hottest and the light intensity at the highest.

4/5/2018
19
Northeast monsoon (Wet season)
Southwest monsoon (Dry season)
Temporal variability
Ostreopsis temporal distribution displayed a manner opposite to that of Gambierdiscus. It reach maximum abundance in Jun, which is the hottest time of the year. It seems to withstand the harsh weather in the wet season, and perhaps the trait of ability to form massive mat is the advantage here.
Experiment on substrate/host preference
Gambierdiscus cultures
Collecting macroalgae
Experimental setup
Cell count
e.g. Parson et al. 2011, 2015
4/5/2018
20
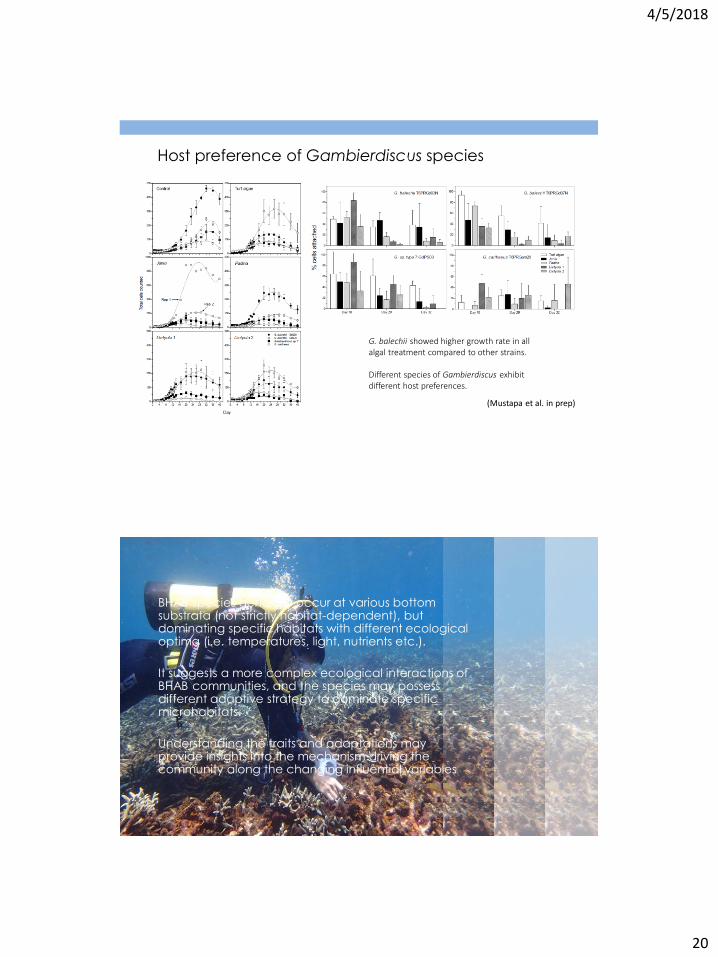
Host preference of Gambierdiscus species
G. balechii showed higher growth rate in all algal treatment compared to other strains. Different species of Gambierdiscus exhibit different host preferences.
(Mustapa et al. in prep)
BHAB species generally occur at various bottom substrata (not strictly habitat-dependent), but dominating specific habitats with different ecological optima (i.e. temperatures, light, nutrients etc.).
It suggests a more complex ecological interactions of BHAB communities, and the species may possess different adaptive strategy to dominate specific microhabitats.
Understanding the traits and adaptations may provide insights into the mechanism driving the community along the changing influential variables

4/5/2018
21

4/5/2018
22

![Guide to Biology - WordPress.com · Significant contribution Nelson Thornes AQA Science [GCSE Biology] 3 Active Transport Two of the main ways in which diffused substances are transported](https://img.pdfslide.fr/doc/110x75/5e8a088f531a680df900aee2/guide-to-biology-significant-contribution-nelson-thornes-aqa-science-gcse-biology.jpg)