Embed Size (px)
Citation preview

Ronéo 1, Cours 2 UE1 Page 1 sur 18
UE1 Bases moléculaires et cellulaires des pathologies
Pr Hélène Cavé
Mercredi 27/09/2017 de 15h30 à 17h30
Ronéotypeur : Ji-Yun PARK
Ronéoficheur : Thomas POUCHEPARADJ
Cours 2 – UE1
Mécanismes moléculaires de la
transduction des signaux
Le métabolisme de l’information partie 2

Ronéo 1, Cours 2 UE1 Page 2 sur 18
Sommaire :
I. Récepteurs-enzymes
A) Activité enzymatique associée : voie JAK/STAT
1. JAK
2. STAT
3. SOCS
4. Etapes de signalisation
5. Combinatoires
B) Activité enzymatique intrinsèque (Récepteurs Tyrosine Kinase)
1. Ligands
2. Récepteurs
3. Etapes de la signalisation
4. Cascades des MAPKs
5. Voie PI3K
II. Réseau de signalisation : dialogue ou « cross-talk » entre voies de signalisation
A) Les Interactions fonctionnelles
B) Les Interférences
C) Métabolisme de l’information/ Métabolisme énergétique
III. Pathologies touchant les circuits de signalisation
A) Neurofibromatose de type 1
B) Mutation de gènes de la voie Ras dans les tumeurs
C) La lèpre

Ronéo 1, Cours 2 UE1 Page 3 sur 18
I. Récepteurs enzymes
Il existe 2 types de récepteurs enzymes :
• les récepteurs à activité enzymatique associée : voie JAK/STAT
• les récepteurs à activité enzymatique intrinsèque (l’activité enzymatique est portée par le
récepteur lui-même) : récepteurs Tyrosine kinase, récepteurs Ser/Thr kinase, et récepteurs guanylate-
cyclase
A) Activité enzymatique associée : voie JAK/STAT
Cette voie fait intervenir 3 types d'acteurs : JAK, STAT et SOCS
1. JAK (Janus Kinase)
C'est une famille de 4 protéines (JAK1, JAK2, JAK3, TYK2). Elles sont constituées de 4 domaines :
• pseudo-kinase : c'est un homologue du domaine kinase qui a subi des mutations au niveau des
acides-aminés indispensables à la fonction enzymatique. Il est dons inactif en terme enzymatique et
permet la régulation de l'activité du domaine kinase
• kinase : possède l'activité catalytique de phosphorylation
• FERM : permet la liaison avec le récepteur membranaire
• SH2 : c'est un motif récurrent des protéines ; il constitue un site de reconnaissance des
protéines au niveau des tyrosines phosphorylées
→ Quelques exemples de domaines protéiques récurrents :
Les domaines SH sont les domaines récurrents les plus représentés qui ont été définis par homologie à
une protéine oncogénique, la protéine Src (d'où leur nom« Src Homology »).
La protéine Src comporte 3 motifs :
• SH1 : domaine enzymatique à activité Tyr kinase
• SH2 : domaine qui ne possède pas d'activité enzymatique mais qui permet la liaison à d'autres
protéines au niveau des Tyrosines phosphorylées avec des spécificités variables. Globalement, on a
une affinité 1000 fois plus importante que celle de la liaison substrat/ligand de la Tyr kinase.
• SH3 : site de liaison à des domaines riches en Proline
→les protéines JAK sont capables de se fixer à des tyrosines phosphorylées grâce à leur domaine
SH2.
Sur le même principe, il existe également des domaines d'homologie à la Pleckstrine (PH) qui sont des
motifs de liaison aux phospho-inositides.

Ronéo 1, Cours 2 UE1 Page 4 sur 18
2. STAT (Signal Transducer and Activator of Transcription)
C'est une famille de 7 facteurs de transcription (STAT1 à STAT7) constitués de 4 domaines :
• domaine de liaison à l'ADN (comme tout facteur de transcription)
• domaine de dimérisation
• domaine SH2 qui permet de se fixer à une Tyrosine phosphorylée
• tyrosine conservée (en C-ter) qui peut être phosphorylée
STAT est capable de former des homodimères via le domaine SH2 lorsque la tyrosine conservée est
phosphorylée. « jeu d'emboîtement » avec le domaine SH2 d'un monomère qui s'accroche sur la
tyrosine phosphorylée de l'autre monomère et réciproquement)
3. SOCS (Suppressor of Cytokine Signaling)
C'est une famille de 7 protéines (SOCS1 à SOCS7) constitués de 3 domaines d'interaction protéine-
protéine :
• domaine KIR (Kinase Inhibitory Region) capable d'inhiber directement la fonction kinase de
JAK
• domaine SH2 qui permet de se fixer à une Tyrosine phosphorylée
• Boîte SOCS capable de recruter le système d'ubiquitine transférase et induire ainsi la
dégradation par le protéasome, du complexe Récepteur+JAK+SOCS
4. Etapes de la signalisation
Contrairement aux récepteurs à 7 domaines transmembranaires, la voie JAK/STAT est une voie de
signalisation qui concerne un nombre très restreint de molécules.
→Elle est le plus souvent utilisée par : • les facteurs de croissance
• les cytokines
• l'hormone de croissance (GH)
• la leptine
• l'Interféron...

Ronéo 1, Cours 2 UE1 Page 5 sur 18
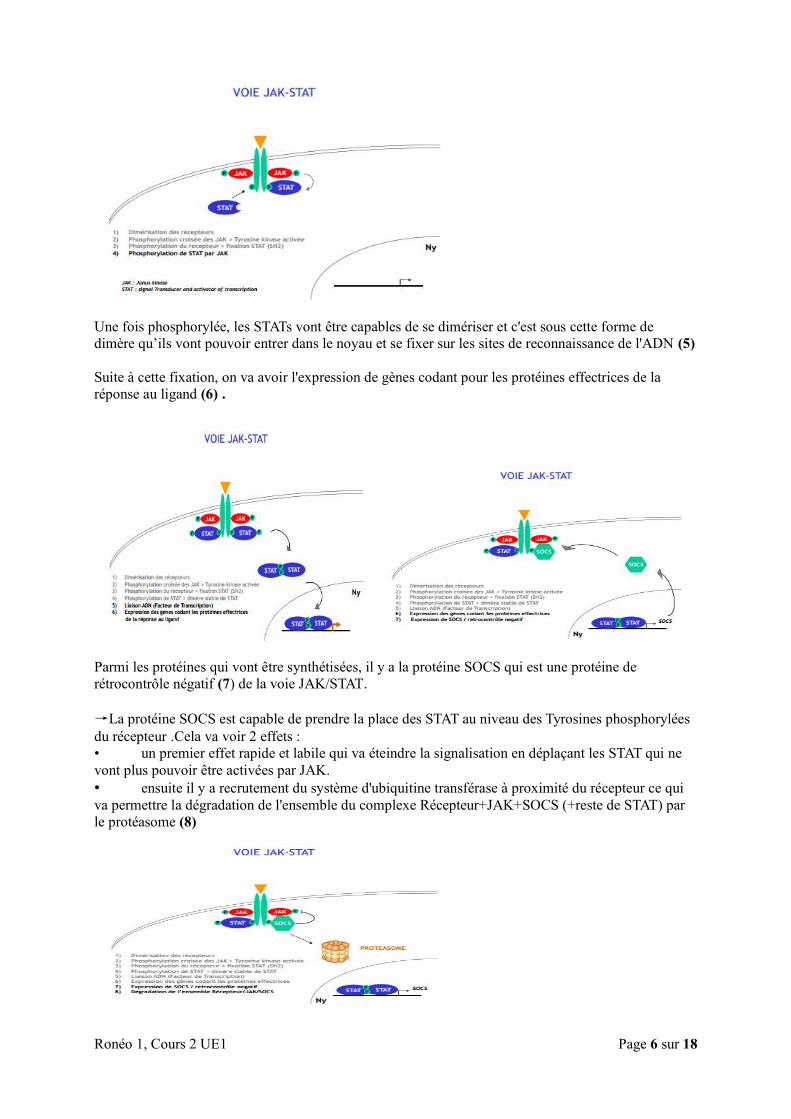
→La voie JAK/ STAT se fait en 8 étapes : 1. Dimérisation des récepteurs
2. Phosphorylation croisée des JAK ce qui va activer la Tyrosine Kinase
3. Phosphorylation du récepteur permettant la fixation de STAT via le domaine SH2
4. Phosphorylation de STAT
5. Liaison à l'ADN (facteur de transcription)
6. Expression des gènes codant pour les protéines effectrices
7. Expression de SOCS (rétrocontrôle négatif)
8. Dégradation de l’ensemble Récepteur/JAK/SOCS
→ Explication des étapes
• Au repos :
le récepteur est fiché dans la membrane. Sur ce récepteur, est accrochée une protéine JAK.
Le récepteur lui-même n'a pas d'activité enzymatique mais il est étroitement associé à la protéine JAK,
qui elle, est porteuse d'une activité enzymatique.
• Fixation du ligand :
En présence du ligand, les deux monomères du récepteur vont s'associer entre eux pour former un
dimère (1).
Cela va entraîner la juxtaposition des protéines JAK.
La protéine JAK a une activité très faible lorsqu' elle est isolée. Cependant, avec la dimérisation des
récepteurs, les 2 protéines JAK vont se phosphoryler mutuellement ce qui va activer leur fonction
kinase (2).
Ensuite, les protéines JAK vont phosphoryler le récepteur au niveau des tyrosines permettant
l'apparition d'un site d'ancrage par les protéines STAT (via le domaine SH2) (3).
JAK va alors phosphoryler STAT au niveau de la tyrosine conservée en C-ter (4) .
Ronéo 1, Cours 2 UE1 Page 6 sur 18
Une fois phosphorylée, les STATs vont être capables de se dimériser et c'est sous cette forme de
dimère qu’ils vont pouvoir entrer dans le noyau et se fixer sur les sites de reconnaissance de l'ADN (5)
Suite à cette fixation, on va avoir l'expression de gènes codant pour les protéines effectrices de la
réponse au ligand (6) .
Parmi les protéines qui vont être synthétisées, il y a la protéine SOCS qui est une protéine de
rétrocontrôle négatif (7) de la voie JAK/STAT.
→La protéine SOCS est capable de prendre la place des STAT au niveau des Tyrosines phosphorylées
du récepteur .Cela va voir 2 effets : • un premier effet rapide et labile qui va éteindre la signalisation en déplaçant les STAT qui ne
vont plus pouvoir être activées par JAK.
• ensuite il y a recrutement du système d'ubiquitine transférase à proximité du récepteur ce qui
va permettre la dégradation de l'ensemble du complexe Récepteur+JAK+SOCS (+reste de STAT) par
le protéasome (8)

Ronéo 1, Cours 2 UE1 Page 7 sur 18
5. Combinatoires
Pour chaque récepteur, il existe un usage préférentiel d'1 ou 2 types de JAK.
Par exemple, pour JAK1, on a le récepteur à l'IL (interleukine) 10 et 6 et pour JAK2,le
récepteur à l'EPO. (les récepteurs médians des signaux de développement et de prolifération
des cellules hématopoïétiques utilisent préférentiellement JAK2).
Certains récepteurs peuvent s'associer à deux types de JAK différents comme les récepteurs à
l'INF-γ (JAK1 et JAK2).
De même, il existe des combinatoires JAK/STAT préférentielles :
La protéine STAT induit l'expression d'un groupe déterminé de gènes pouvant varier selon le
type de cellule. Par exemple dans le cas du récepteur à l'IL-4, avec une même STAT6, selon qu'on est
dans une cellule lymphoïde ou un macrophage, on va avoir des gènes exprimés qui vont être
différents.
Exemple de l'INF-γ :
C'est une cytokine immunostimulante produite par les lymphocytes T et les cellulules NK pour
répondre aux infections virales. Elle peut être surproduite dans certaines maladies autoimmunes et
est utilisée comme médicament pour traiter certains cancers. →La fixation de l'IFN sur son récepteur entraîne : • la fixation de JAK1 et 2 au récepteur
• activation puis dimérisation de STAT1 suite à sa phosphorylation
• transcription de séquences spécifiques de réponse à l'INF

Ronéo 1, Cours 2 UE1 Page 8 sur 18
D'après le schéma précédent, on s'aperçoit que :
• avec ses 3 composantes (récepteur, JAK, STAT), la voie JAK/STAT est relativement
simple et très rapides avec des réactions linéaires
• par contre, les voies comprenant des composants plus nombreux présentent une capacité
supérieure d'amplification du signal.
B) Activité enzymatique intrinsèque (Récepteurs Tyrosine Kinase)
1. Ligands
Les récepteurs à activité Tyrosine Kinase sont les récepteurs des facteurs de croissance
NB : Tous les facteurs de croissance n'ont pas un récepteur à activité intrinsèque : certains facteurs de
croissance ont un récepteur JAK/STAT
Les facteurs de croissance permettent essentiellement :
• la prolifération
• la survie cellulaire (en protégeant la cellule de l'apoptose)
2. Structure des récepteurs
Il existe environ 60 types de récepteurs répartis en 20 sous-familles mais leur structure générale reste
commune les uns avec les autres :
• Domaine extracellulaire très glycosylé et très riches en Cystéine, « en pseudo-globulin » ou
« immunoglobulin-like » • Domaine transmembranaire
• Domaine intracytoplasmique qui porte la fonction kinase

Ronéo 1, Cours 2 UE1 Page 9 sur 18
3. Etapes de la signalisation
Activation des récepteurs Tyr Kinase (TK)
Au repos, les récepteurs TK sont fichés dans la membrane avec une fonction kinase qui n'est pas
active.
Lorsque le ligand se fixe sur les récepteurs, on a la dimérisation de ces derniers avec une mise à
proximité des fonctions kinases de chacun des monomères. De même que les protéines JAK, les TK intrinsèques de ce type de récepteur sont inactives
lorsqu’elles ont isolées. Cependant, la dimérisation des récepteurs permet une mise à proximité des
domaines TK qui peuvent alors se phosphoryler mutuellement (=auto-phosphorylation) au niveau
d’une tyrosine intracellulaire du récepteur →activation des TK→phophorylation sur plusieurs
sites du récepteur
Les tyrosines phosphorylées du récepteur vont attirer les protéines à domaine SH2.(et également des
protéines cytoplasmiques)
La phosphorylation des Tyr du récepteur a 2 effets :
• Activation d'une fonction enzymatique (ex : phospholipases, lipides kinases) suivie de la
production d'une petite molécule (ex : PIP3)
• Activation d'une cascade (interactions en série entre macromolécules) (ex : protéines
kinase)

Ronéo 1, Cours 2 UE1 Page 10 sur 18
Il y a donc une forte amplification du signal à chaque étape avec parfois des branchements (ou des
interactions entre différentes voies).
Enfin, ce signal est inactivé d’une part, par des phosphatases qui vont déphosphoryler les tyrosines du
récepteur et d’autre part, par l'endocytose des récepteurs.
Exemple du récepteur à l'EGF (EGFR)
Un récepteur à activité TK peut contenir plusieurs sites de liaison au SH2 et activer plusieurs
effecteurs :
La fixation du ligand (EGF) induit une dimérisation des récepteurs. Suite à cette dimérisation, on va
avoir la phosphorylation des tyrosines dans la partie intracellulaire du récepteur activant ainsi son
activité TK.
Cette phosphorylation des tyrosines va également permettre l'apparition, sur le récepteur, de sites
d'ancrage aux protéines porteuses de domaine SH2 en particuliers, Grb2.
Grb2 est une protéine possédant 3 domaines récurrents :
• un domaine SH2
• et 2 domaines SH3
Sa fonction est de se lier aux protéines & établir des liaisons protéine-protéine.
Il est donc le prototype d'un adaptateur moléculaire (=protéine se liant à d'autres protéines
sans activité catalytique propre)
Il se lie au récepteur (via le domaine SH2) et attire vers lui, la protéine Sos.
Sos est un facteur d'échange (=GEF) et agit sur la protéine Ras (=un commutateur
moléculaire) par contact protéine/protéine. En effet, Sos va modifier la conformation de Ras
qui va ouvrir sa poche à nucléotide et laisser partir le GDP laissant alors place au GTP qui, en
se fixant sur Ras, va l'activer.
Ras est une « petite protéine G ». C'est un monomère très similaire à la SU α de la protéine G.
C'est un commutateur moléculaire et possède donc la fonction GTPase qui lui confère la
capacité à hydroliser le GTP. Cette fonction a besoin d'être activée par une protéine GAP, la
plus connue étant NF1 qui code pour la neurofibromine.
Ronéo 1, Cours 2 UE1 Page 11 sur 18
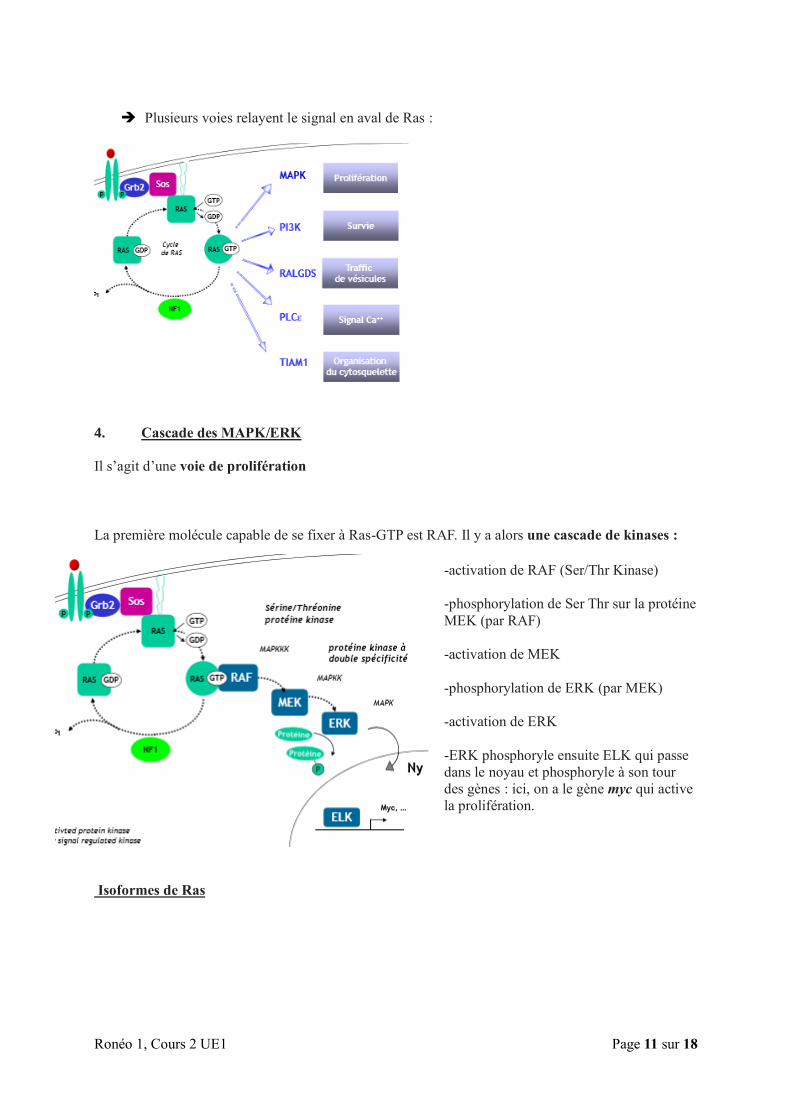
Plusieurs voies relayent le signal en aval de Ras :
4. Cascade des MAPK/ERK
Il s’agit d’une voie de prolifération
La première molécule capable de se fixer à Ras-GTP est RAF. Il y a alors une cascade de kinases :
-activation de RAF (Ser/Thr Kinase)
-phosphorylation de Ser Thr sur la protéine
MEK (par RAF)
-activation de MEK
-phosphorylation de ERK (par MEK)
-activation de ERK
-ERK phosphoryle ensuite ELK qui passe
dans le noyau et phosphoryle à son tour
des gènes : ici, on a le gène myc qui active
la prolifération.
Isoformes de Ras

Ronéo 1, Cours 2 UE1 Page 12 sur 18
La famille des protéines Ras (=sous-famille de la protéine G) comprend 4 protéines codées par
seulement 3 gènes très homologues (effet de l’épissage alternatif) : Hras, Nras et Kras. Ce dernier
code pour 2 protéines, Kras A et Kras B.
Au niveau de l’extrémité C-ter, on a une région hypervariable composée de résidus spécifiant les
modifications post traductionnelles des protéines.
Modifications post-traductionnelles de Ras
Ras est une protéine hydrophile ; elle subit 2 types de modifications :
-la palmitoylation : ajout d’acides gras par des Palmitoyl-transférases -la prénylation : ajout d’acides gras isoprénoides sur une cystéine en C-ter ou appartenant
à un motif CAAX . Elle est réalisée par les farnesyl-transférase ou granyl/geranyl-
transférase
Ces modifications confèrent à Ras la capacité de s’ancrer à la membrane et il existe des
différences selon les types de protéine RAS. (Ras est une protéine hydrophile, et par conséquent cytoplasmique. La palmitoylation et la prénylation
consistent à ajouter des acides gras qui sont des molécules hydrophobes →ancrage de Ras à la
membrane)

Ronéo 1, Cours 2 UE1 Page 13 sur 18
5. Voie PI3K(Phosphatidyl Inositol 3 Kinase) / AKT
Il s’agit d’une voie de prolifération et de survie cellulaire.
/ ! /PI3K est une LIPIDE Kinase →permet la phosphorylation de lipides
PIP2 (Phosphatidyl inositol (4,5) Phophate 2) est phosphorylé par PI3K, grâce à un ATP, et devient
PIP3 (Phophatidyl Inositol (3, 4,5) Phosphate 3). → PIP2 est utilisé comme précurseur de seconds
messagers (=PIP3)
Cette réaction est inversée par PTEN .
Structure de PI3K
PI3K est composé de 2 sous unités :
-P110 = SU catalytique
Elle contient :
-un domaine kinase
-un domaine de liaison à Ras
-un domaine d’ancrage membranaire C2
-un domaine de liaison à P85
-P85 = SU régulatrice
Elle contient :
-un domaine de liaison à P110
-2 domaines SH2
C’est donc une protéine adaptatrice → permet la liaison protéine/protéine
Il existe 3 modes d’activation de PI3K :
→directement, en se fixant sur le récepteur à, activité TK via les domaines SH2 de sa sous-unité
P85 →via Ras →via IRS = Substrat du Récepteur à l’Insuline
Description de la voie PI3K/AKT
/ !/PIP2 et PIP3 étant lipophiles, ils sont ancrés dans la membrane
Lorsque la SU P110 de PI3K se fixe à Ras, il y a :

Ronéo 1, Cours 2 UE1 Page 14 sur 18
→activation de PI3K
→transformation de PIP2 en PIP3
→augmentation de la concentration de PIP3
PIP3 est reconnu par les domaines PH portés par les protéines AKT (=PKB), PDK1 et
PDK2→activation de AKT
Une fois activé, AKT phosphoryle des protéines ce qui va modifier l’activité cellulaire avec une
activation ou une inhibition des cibles cellulaires :
Un récepteur à activité TK peut contenir plusieurs sites de liaison aux SH2 et activer plusieurs
effecteurs : Par exemple, les Tyr phophorylées du récepteur peuvent recruter la protéine Cbl qui est
associée à une Ubiquitine Ligase → voie de dégradation des protéines et du récepeteur
Exemple : Listeria monocytogenes
-Certaines bactéries pathogènes utilisent le métabolisme des PI pour augmenter leur virulence.
-C’est le cas de Listeria monocytogenes qui est une infection dangereuse surtout chez la femme
enceinte et les personnes immuno-déprimées .
-Cette bactérie reconnait un récepteur TK et active la voie PI3K. → Internalisation de la bactérie et
modification du cytosquelette -Il y a un détournement de la machinerie cellulaire de signalisation pour permettre l’intégration de la
bactérie qui a une action intracellulaire.

Ronéo 1, Cours 2 UE1 Page 15 sur 18
II. Réseau de signalisation : dialogue ou « cross-talk » entre voies de signalisation
Les voies de signalisation peuvent être mises en œuvre simultanément dans une cellule, elles sont très
polyvalentes et également partagées par différentes cellules, facteurs de croissance, cytokines…
Le schéma ci-contre illustre la complexité du réseau
de signalisation : activation de plusieurs voies dont
JAK/STAT, Phospholipase C (PLC), Ras MAPK et
beaucoup d’autres.
Les signaux d’entrées et les sorties (conséquences
moléculaires) sont extrêmement variables mais le
noyau de processus central est lui conservé, il
existe en effet un nombre limité de
possibilités/circuits.
En fait la spécificité de la réponse est permise par de nombreuses molécules gravitant autour
du système, elles correspondent au contrôleur du système. Il a ainsi des contrôles au niveau des
entrées et des sorties mais également des rétrocontrôles négatif ou positif variables d’une cellule à
l’autre.
A un même signal la cellule répond différemment. Le contexte cellulaire, les molécules de régulation
synthétisées et les rétrocontrôles font qu’un même signal interprété dans une cellule va être interprété
différemment dans une autre cellule. De même la réponse sera différente selon son niveau de
différenciation.
Par ailleurs, pour un même type de cancer, les mutations seront différentes selon qu’il soit présent chez
un enfant ou chez un adulte.
Ex : Jak2 impliqué dans les syndromes myéloprolifératif, les mutations sont différentes :
Elles entrainent chez l’adulte à une réponse transcriptionnelle conduisant à une myoprolifération alors
que chez l’enfant il n’y a pas de réponse transcriptionnelle.)
Le contexte cellulaire est donc très important et permet avec peu de mécanismes une grande diversité.
→ Plusieurs voies peuvent être activées en aval d’un stimulus, peuvent interagir entre elles, on
distingue plusieurs types d’interaction :
Des Interactions fonctionnelles
Des Interférences
Une compartimentation
A) Les Interactions fonctionnelles
Ce sont des interactions mises à profit pour la cellule assurant ainsi un dialogue positif entre les
différentes voies.
Interactions de convergence
Convergence additive : A ou B nécessaire
Exemple : l’IFN qui active STAT1 grâce à la voie JAK mais cette dernière peut également activer PI3k
qui peut phosphoryler STAT1 qui devient alors plus actif. (la voie PI3k n’est pas indispensable mais
permet une meilleure activation). Il y a un effet additif.

Ronéo 1, Cours 2 UE1 Page 16 sur 18
Convergence intégrative : A et B nécessaire
Exemple avec le développement de la drosophile : 2 signaux (bleu/rouge) sont
produits touchant des bandes de cellules différentes, avec une partie recevant
les 2 signaux (violet). Seules les cellules ayant reçu les 2 signaux peuvent
produire le 3ème signal.
Ainsi les 3 signaux sont indispensables. (Important pour la spéciation et la
différenciation cellulaire)
Le système reçoit différents types de signaux et doit être capable de donner une réponse unifiée aux
besoins de l’organisme.
Interactions de régulation
B) Les Interférences
Certaines interactions sont non voulues, les interférences. On souhaite que les signaux se produisent de
façon distincte les uns des autres.
La cellule s’est prémunie contre ces interférences :
Différenciel d’affinité des kinases pour leurs cibles
Les kinases appartiennent à des familles et il existe une diversité des kinases produites à chaque fois.
Le signal peut être le même en terme de modalité général de fonctionnement mais va activer des
kinases et des cibles différente. Ceci participe à la spécificité des voies de signalisation.
Des phosphatases suppriment les phosphorylations inappropriées des kinases par déphosphorylation.
L’isolement spatial : la cellule est organisée en compartiment ce qui permet d’isolé des secteurs dans
lesquels se produisent les signalisations différentes.
Ce système permet une optimisation du fonctionnement des voies de signalisation et de minimiser les
interférences.
Exemple des PKA :
Découverte des AKAP, protéines d’assemblage de
complexes macromoléculaire sur lesquels se fixe PKA,
les molécules de régulation dans un domaine
subcellulaire.
Ceci permet une activation coordonnée des protéines kinases car les composants sont
regroupés aboutissant à une action optimisée, au maintien de la spécificité des signaux et par
conséquent à une efficacité plus importante.
L’AMPc va réguler la réponse à des stimuli externe dans une région de la cellule délimitée et pour un
temps limité : Création d’un gradient spatio-temporel.

Ronéo 1, Cours 2 UE1 Page 17 sur 18
C) Métabolisme de l’information/ Métabolisme énergétique
Ces deux mécanismes sont assez proches mais diffèrent au niveau de la quantité de molécules :
La signalisation correspond à un
transfert d’information de faibles
quantités.
Le métabolisme énergétique permet le
transfert de grandes quantités de
masse. De plus les quantités en enzyme
sont beaucoup plus faibles que celles en
substrat, l’enzyme est limitante.
Dans la signalisation il y a un équilibre entre les quantités de substrat et d’enzyme permettant une
régulation de la signalisation.
III. Pathologies touchant les circuits de signalisation
A) Neurofibromatose de type 1
Maladie du développement, autosomique dominante due à une
haploinsuffisance de NF1 (GAP protéine, inhibiteur de Ras), il y a perte d’un
allèle du gène.
L’absence de NF1 conduit à une activation anormalement prolongée de Ras.
Ceci crée une fragilité cellulaire avec un risque important de développer des
tumeurs maligne spécifique.
B) Mutation de gènes de la voie Ras dans les tumeurs
Ras est souvent impliqué dans le développement de cancer, en effet les mutations activatrice de Ras
sont très fréquente dans ces derniers et les conséquences de ces mutations sont très variées.
Mutations de Ras fréquentes : (valeurs à ne pas connaitre)
_85% cancers du pancréas
_55% cancers de la thyroïde
_44% cancers du colon
A noter que KRas et NRas sont plus impliqués dans les tumeurs que HRas (or de nombreuses études
portait sur le rôle de HRas)
Différentes anomalies dans la voie de signalisation représente des cibles thérapeutiques potentielles :
_récepteur surexprimé (EGFR)
_récepteur muté (EGFR)
_Intermédiaires surexprimés (Ras, Raf, Braf,…)
_ligand surproduit

Ronéo 1, Cours 2 UE1 Page 18 sur 18
De nombreuses thérapeutiques ont été développées :
_Inhibiteurs de farnesyl transferase touchant
la capacité de Ras à fixer la membrane.
_Statines
_L’Herceptine particulièrement importante
dans le cancer du sein.
C) La lèpre
Problème de santé publique majeur dans certains pays, la lèpre est une atteinte provenant d’une
mycobactérie, la Mycobacteriumleprae affectant le système nerveux périphérique.
Elle est capable de lier à la surface des cellules de Schwann (présentes au niveau des gaines nerveuses)
les récepteurs ErbB2 (récepteurs à Tyrosine kinase).
La voie MAPK est anormalement activée ce qui a pour conséquence la démyélinisation de ces
cellules.
La progression de la lèpre peut être bloquée :
Au niveau du récepteur par l’Herceptine (utilisé dans le cancer du sein) ou PK166
Au niveau de la phosphorylation de Erk1/2 par l’inhibiteur de Mek UO126
(A noter que la lèpre est soignée par antibiotiques, en effet Herceptine représente un cout trop
important que ne peuvent se permettre les pays touchés)
Les voies de signalisations sont extrêmement polyvalentes et peuvent être communes entre le
cancer du sein et la lèpre.





























