Embed Size (px)
Citation preview

UE8 – Système neurosensorielLundi 25 février 2013 à 13h30Pr LozeronRonéotypeur : Romain BourdoncleRonéolectrice : Ariane Corret
Cours n°17 : Somesthésie, Généralités, Système Lemniscal - 1 ère partie
Plan :
I - Définition
II - Généralités sur la sensibilité A- Physiologie de la sensibilité B- Détails sur les 5 étapes
III - Système lemniscal A- Réception B- Transduction C- Modulation D- Transmission E- Intégration
Avant Propos : Oui, je sais, ce cours fait 24 pages… Mais ne vous inquiétez pas, c’est surtout pour plus de clarté. Cette ronéo contient l’intégralité des propos du Prof en cours, ainsi qu’une grande majorité des diapos. Je vous invite d’ailleurs à aller les regarder sur weebly car je ne suis pas sur qu’elles passent toutes. Il y a aussi des images d’IRM que je n’ai pas mise car en ayant tenté une photocopie, je me suis vite rendu compte des dégâts sur la ronéo finale. (J’ai tenté de refaire un cerveau en paint). Le cours sur la Somesthésie est en deux parties. Il y a beaucoup de points que le prof développe dans la seconde partie. Il n’y avait pas vraiment de plan du coup, j’ai essayé d’organiser le tout. Soyez prudent sur la partie Modulation, ça me paraissait logique, et le prof ne caractérise pas vraiment la modulation sinon.. Bon courage à tous !

I - Définition
Etymologie grecque de somesthésie.
SOMA : corps AISTHESIS : sensation
=> Sensibilité du corps
II - Généralités sur la sensibilité
Il y a deux grands types de sensibilité :
Sensibilité viscérale: qui est inconsciente Récepteurs viscéraux (chémorécepteurs, mécanorécepteurs...). Sensibilité inconsciente.- récepteurs au pH ou P02 : inconscient - sensibilité consciente pour un certain nombre d’organes : une douleur dans la poitrine peut traduire un infarctus du myocarde. Donc un viscère, un organe peut entrainer une douleur.
Sensibilité somatique (donc du corps) : qui est conscienteIl y a différent type de sensibilités somatiques. On les différencie parce qu’elles passent par des voies différentes, par des mécanismes différents et mettent en jeu des reflexes différents.
- Tact discriminatif (sensibilité de la peau) : qui permet de discriminer, de séparer deux objets assez précisément. Il y a une deuxième partie du tact, dont on entend moins parler : le tact protopathique, on sait qu’on nous touche mais c’est beaucoup plus grossier et c’est véhiculé par un autre type de fibre.
- Proprioception « Réception du soi » (position des membres et corps) : c’est le sens de position des membres. Pour avoir de l’équilibre il faut trois choses : la vue, le système vestibulaire et le cervelet. Si on n’a plus de proprioception, on est obligé d’avoir une bonne vue.
EX : Si tu lances une balle en l’air, sans voir ta main, tu pourras la rattraper (et oui !) parce que tu sais où est ton membre. Il y a des capteurs de sensibilité pour connaitre le sens de position des membres. Ces capteurs font intervenir certaines fibres nerveuses dans les nerfs périphériques et une certaine voie de sensibilité qui est, ici, la voie LEMNISCALE au niveau du SNC.
En clinique, il y a des maladies qui atteignent spécifiquement ces fibres. Quand on a un patient qui a un trouble de l’équilibre et une atteinte proprioceptive, on aura une orientation diagnostique.
- Douleur : il existe différents types de douleur (ce sera vu dans la partie II)- Température - Sensibilité sensorielle spécialisée (vision, goût, odeurs, audition…)

A- Physiologie de la sensibilité
Pour sentir il faut ces 5 étapes :
1 - Réception : Récupérer l’énergie d’un stimulus via… des récepteurs.
2- Transduction : Transformer ce stimulus en changement de potentiel de membrane. Et ce n’est pas si simple : par exemple pour le toucher il va falloir transformer un message mécanique en signal électrique.
3- Transmission : Transfert du message jusqu’au système nerveux central. Cet influx nerveux pourra être modulé.
4- Modulation : Variations appliquées au message. Il y a des mécanismes inhibiteurs, des mécanismes activateurs qui pourront intervenir dans des mécanismes de défense en particulier.
5- Intégration : Interprétation des données. Pour bouger ta main il faut aussi que tu ais de la sensibilité en plus de l’activité motrice. Il est impératif de savoir où est notre main. C’est la même chose que pour envoyer la balle. Si tu veux faire du moteur, il faut que tu ais intégré le sensitif pour prévenir les voies motrices, les centres moteurs, que tu as bougé. Et ça se passe au niveau du SNC.
1 - Organisation du système sensitif +++
La sensibilité se fait avec trois neurones. (« Sauf quand elle se fait avec un peu plus… » Cf. Partie II)
- Neurone de premier ordre (1er neurone)Délivre l’information sensitive
- Neurone de deuxième ordre (2ème neurone) ou deutoneurone Dans la moelle ou le tronc cérébral
- Neurone de troisième ordre (3ème neurone)
A chaque fois, il y a un relais.
(Pour la motricité, il faut 2 neurones : un du cortex jusqu’à la moelle, et un qui va de la moelle jusqu’au muscle)
Une atteinte à ces trois niveaux se traduira cliniquement différemment. C’est important de connaitre ces trois neurones, car on n’aura pas les mêmes symptômes cliniques. Il est IMPORTANT de savoir qu’il y a trois neurones (apprenez aussi les détails sur la localisation dans la suite du cours)

2- Types de sensibilité
a. La sensibilité tactile :
- Le tact fin (ou sensibilité tactile épicritique) discriminative. Il faut connaitre ces trois termes +++.
Par exemple, la main est très précise elle peut discriminer deux petites piqures à l’aiguille à quelques mm l’une de l’autre, alors qu’au milieu du dos… Donc, il permet de distinguer deux points de pression sur la peau (la plus importante au niveau de lèvres et de la pulpe des doigts). Elle est liée au nombre de récepteurs par surface.
- Le tact grossier ou sensibilité protopatique (pas important). On sait qu’on touche mais on n’a pas de localisation précise.
b. La sensibilité proprioceptive (récepteurs articulaires, récepteurs golgiens, etc.) : sens de position des segments de membres dans l’espace et les uns par rapport aux autres.
c. La sensibilité thermique.
d. La sensibilité algique (douloureuse) ou nociception.
3- Schéma commun de la sensibilité.
Que ce soit pour le système lemniscale ou extralemniscale, on aura toujours 3 neurones.
Un premier neurone, dit neurone afférent. Le corps cellulaire de ce premier neurone est dans le ganglion rachidien postérieur. L’autre partie est périphérique, en dehors de la moelle.
Puis au niveau du deuxième neurone, (juste avant), on entre dans la moelle. (Différent du système extraL)
Puis un troisième neurone qui va jusqu’au cortex CONTROLATERAL.
La sensibilité de l’hémicorps gauche est perçue et integrée au niveau du cortex droit. C’est comme pour la motricité, ca croise.
Le premier relais est dans la moelle.Le deuxième relais se fait dans le thalamus.
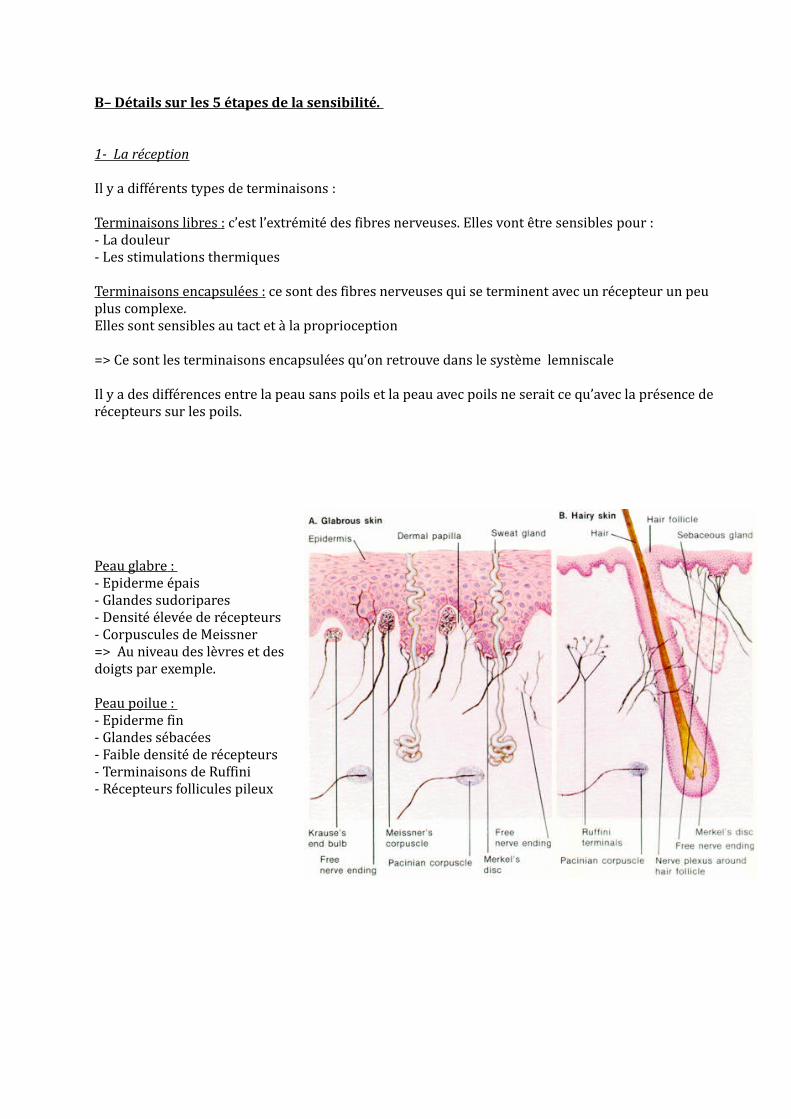
B– Détails sur les 5 étapes de la sensibilité.
1- La réception
Il y a différents types de terminaisons :
Terminaisons libres : c’est l’extrémité des fibres nerveuses. Elles vont être sensibles pour : - La douleur- Les stimulations thermiques
Terminaisons encapsulées : ce sont des fibres nerveuses qui se terminent avec un récepteur un peu plus complexe. Elles sont sensibles au tact et à la proprioception
=> Ce sont les terminaisons encapsulées qu’on retrouve dans le système lemniscale
Il y a des différences entre la peau sans poils et la peau avec poils ne serait ce qu’avec la présence de récepteurs sur les poils.
Peau glabre : - Epiderme épais- Glandes sudoripares- Densité élevée de récepteurs- Corpuscules de Meissner=> Au niveau des lèvres et des doigts par exemple.
Peau poilue : - Epiderme fin- Glandes sébacées- Faible densité de récepteurs- Terminaisons de Ruffini- Récepteurs follicules pileux

2- Transduction
Le récepteur transforme un signal extérieur à travers une modification de potentiel de membrane. Le récepteur, la terminaison nerveuse, qu’elle soit encapsulée ou libre, entraine la création d’un potentiel d’action PA qui va pouvoir être transmis. Cela va être possible grâce à un potentiel de récepteur (PR).
Gentil rappel du prof sur nos cours de Mercadier de la P1 : Le potentiel gradué, qui augmente progressivement en fonction de l’intensité du stimulus et de la distance à laquelle on est du point du stimulus, va être plus ou moins grand. S’il dépasse un certain seuil de repos de la membrane (= le seuil de dépolarisation de la membrane), il va permettre d’ouvrir certains canaux ioniques. Et donc si le potentiel gradué ouvre les canaux ioniques, il y a un potentiel d’action et à partir de ce moment là, c’est la loi du tout ou rien : on décharge un PA ou on n’en décharge pas.
C’est donc la transduction : transformer un message chimique ou mécanique (ou thermique… ou douloureux) en un Potentiel d’Action qui lui va être transmis dans le système nerveux.
On va revoir tout ça dans la suite du cours.
3 - Transmission
On va donc transmettre cet influx nerveux. Initialement dans le SNP (dans les nerfs) dans les bras et dans les jambes et on va le transmettre avec des fibres particulières. Il y a différents types de fibres, on différencie essentiellement les fibres Aβ, Aδ, et les fibres C.
Les fibres Aβ vont faire la sensibilité discriminative, le tact épicritique, tact fin, Les fibres Aδ vont faire le tact non discriminatif, la température, la douleur.

Les fibres C vont faire la température et la douleur. Le tact discriminatif c’est la voie lemniscale => la transmission se fait par les fibres Aβ L’autre c’est la voie extralemniscale (la partie II) => Aδ et C
Grace à ces fibres nerveuses (au niveau du doigt) « coucou c’est un doigt »Les fibres nerveuses passent dans le bras, le corps cellulaire est dans le ganglion rachidien postérieur, et il va faire relais avec un deuxième neurone qui sera situé dans la moelle ou dans la moelle allongée (moelle allongée = anciennement bulbe) pour rejoindre le SNC.
Entre chaque vertèbre sort ce qu’on appelle une RACINE. Ces racines vont mélanger leurs fibres entre elles au niveau des PLEXUS.Et ça va ressortir à l’autre bout sous forme d’un nerf.
Donc en gros : Bras nerfs => PLEXUS où ils se mélangent => et ca donne une racine qui sort de la vertèbre.
Ces racines apportent la sensibilité de zones très précises (on reverra ça dans la partie II) mais ca forme des DERMATOMES.
Ex : La racine T4 passe par le mamelon. Si on coupe T4, on aura une perte de sensibilité au niveau du mamelon. Cliniquement c’est capital. Le pli de l’aine c’est T12-L1L’ombilic c’est T10Xiphoïde c’est T6
C’est très utile quand on a des pathologies de la moelle épinière. On peut dire « ah il ne sent pas de l’ombilic et bien l’ombilic c’est T10 » Donc on sait que c’est une liaison au niveau de T10.
En neurologie, il faut localiser la lésion. Les examens ne suffisent pas toujours comme dans certaines spécialités…
Le transfert de l’information des récepteurs jusqu’au cortex se fait par deux types de voies :
- La voie lemniscale qui passe par une zone du Tronc Cérébral qui s’appelle le Lemnisque MédianElle est DISCRIMINATIVE (reconnaissance)C’est pour la sensibilité tactile épicritique.Et la sensibilité proprioceptive.
- La voie extra-lemniscale ou faisceau spinothalamique:Elle caractérise l’ URGENCE (douleur, alerte T°..)La sensibilité thermique.La nociception.La douleur c’est important, cela apporte un message.
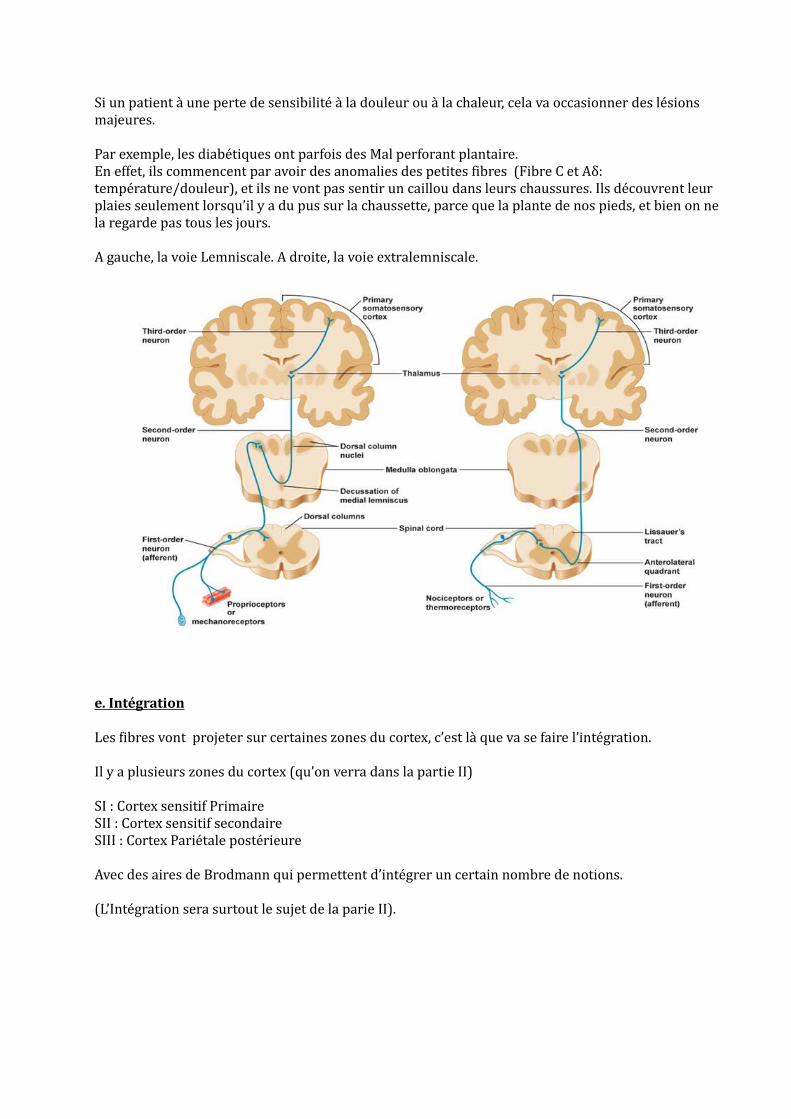
Si un patient à une perte de sensibilité à la douleur ou à la chaleur, cela va occasionner des lésions majeures.
Par exemple, les diabétiques ont parfois des Mal perforant plantaire. En effet, ils commencent par avoir des anomalies des petites fibres (Fibre C et Aδ: température/douleur), et ils ne vont pas sentir un caillou dans leurs chaussures. Ils découvrent leur plaies seulement lorsqu’il y a du pus sur la chaussette, parce que la plante de nos pieds, et bien on ne la regarde pas tous les jours.
A gauche, la voie Lemniscale. A droite, la voie extralemniscale.
e. Intégration
Les fibres vont projeter sur certaines zones du cortex, c’est là que va se faire l’intégration.
Il y a plusieurs zones du cortex (qu’on verra dans la partie II)
SI : Cortex sensitif PrimaireSII : Cortex sensitif secondaireSIII : Cortex Pariétale postérieure
Avec des aires de Brodmann qui permettent d’intégrer un certain nombre de notions.
(L’Intégration sera surtout le sujet de la parie II).

III - Système Lemniscal
Le système Lemniscal est en rapport avec différentes sensibilités :
-Tact discriminant-Pression-Proprioception
Le système lemniscal est là pour transmettre ces informations grâce à trois neurones.
- Neurone afférent primaire
C’est celui qui est dans le nerf périphérique. Et qui a son corps cellulaire dans le ganglion rachidien postérieur (!!).
Il se projette au niveau de la moelle longue (donc le tronc cérébrale) très haut. Il va de l’extrémité de ton orteil jusqu’au tronc cérébral.
Il remonte sur la moelle dans le cordon postérieur HOMOLATERAL.
- Deuxième neurone ou deutoneurone :
Au niveau de la moelle longue, dans les noyaux graciles et cunéiformes (= Noyaux de Goll et Burdach), il y a un relais vers le deuxième neurone
Ce deuxième neurone va se projeter de façon CONTROLATERALE sur le noyau Ventro-Postero- Latéral (VPL) du Thalamus ou sur le Ventro-Médian du thalamus.
(Il y a une erreur sur le schéma, il y a une fille dans l’amphi qui a gentiment signalé « mais Monsieur, le Thalamus est Controlatéral », il a avoué s’être trompé sur le schéma… )
Le thalamus est un noyau gris central. (C’est de la substance grise au centre du cerveau)Au final ça croise la ligne médiane, ça décusse.
- Troisième neurone : au niveau du noyau VPL du thalamus et se projette au niveau du cortex pariétal toujours en CONROLATERAL.
IL y a une SOMATOTOPIE
Le corps – La localisation.
Il y aura une discrimination spatiale des différentes parties du corps. Ce qui est beaucoup moins vrai pour l’extralemniscal. Cela veut dire, qu’il y aura tout le long de la voie une représentation des différentes parties du corps. Une cartographie corporelle. On le verra plus tard dans le cours, donc gardez l’idée à l’esprit.

A – La Réception
Il y a :
- 6 Récepteurs tactiles de la peau
Terminaisons libres (il y en a peu)Follicules pileuxDisques de MerkelCorpuscules de MeissnerCorpuscules de PaciniCorpuscules de Ruffini
- 3 Récepteurs proprioceptifs
Fuseau neuromusculaireOrgane de GolgiRécepteurs articulaires
=> Plusieurs récepteurs différents qui ont des fonctions légèrement ou très différentes…
Les Récepteurs tactiles :
Les récepteurs tactiles sont soit à : • Terminaisons libres:– Toucher et pression (tact grossier)(– Entre les cellules épiderme)
Ou sont des : • Récepteurs pileux :– distorsion du poil et mouvements de la peau(– Adaptation rapide– contact initial et mouvements suivants)
1- Cellules de MerkelCouches profondes de l’épidermeTact fin et pression
2- Corpuscules de Meissner: « retenez Meissner, il est important Meissner, regardez sa localisation Meissner, il est important Meissner » … Perso, je vais l’apprendre.– Jonction derme épiderme– Tact fin, pression, vibration basse fréquence – Paupière, lèvres, bout des doigts, mamelons, organes génitaux externes
« Meissner retenez qu’il est important »
3- Corpuscules de Paccini:– Sensible pression profonde, quelque chose de dynamique, c’est au mouvement qu’il est sensible– Adaptation rapide (mouvement)
4- Corpuscules de Ruffini : également sensible à la pression, sensible à la pression continu – (Derme profond– located in the reticular dermis(deep))

« Vous avez une ronéo Hein… bah si vous voulez recopier tout ça bon… bof »
Cellules de Merkel : terminaisons libres (cellules épithéliales modifiées)Couches profondes de l'épiderme (paumes et plantes)mécano-réception.
Corpuscules de PacciniDerme et autres tissus sous-cutanés. doigts, des organes génitaux et des seins.Ils ont de 1 à 4 mm de long et de 0.5 à 1.0 mm de diamètre. La fibre nerveuse centrale est démyélinisés et est entourée de cellules aplaties et disposées de façon concentrique.Mécanorécepteurs pressions, vibration, déformation (articulation vessie)Adaptation rapide (mouvement).
Corpuscules de RuffiniDerme profondPetits et allongés, ils font environ 1 mm de long par 0.1 mm de diamètre.Etat d'étirement de la peau (statique).Bulbes de Krause sont de petits corpuscules situés dans le derme de tissus muqueux (blanc de l'oeil, les paupières, la langue et les muqueuses génitales externes).Mécanorécepteurs.
Corpuscules de MeissnerJonction derme épidermeZones Glabre de grande sensibilité tactile (surface palmaire des doigts, la plante des pieds, les paupières, les lèvres, les régions génitales externes et le mamelon des seins).Textures
=> Pas très important.
Schéma récapitulatif :
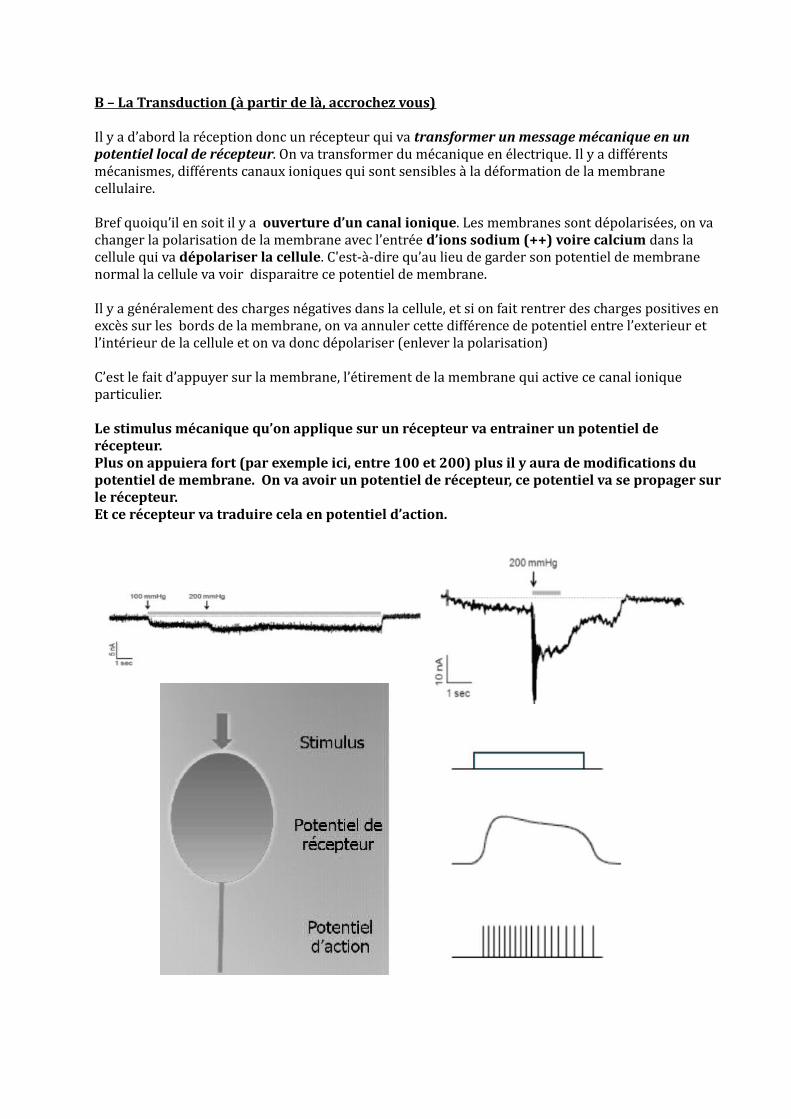
B – La Transduction (à partir de là, accrochez vous)
Il y a d’abord la réception donc un récepteur qui va transformer un message mécanique en un potentiel local de récepteur. On va transformer du mécanique en électrique. Il y a différents mécanismes, différents canaux ioniques qui sont sensibles à la déformation de la membrane cellulaire.
Bref quoiqu’il en soit il y a ouverture d’un canal ionique. Les membranes sont dépolarisées, on va changer la polarisation de la membrane avec l’entrée d’ions sodium (++) voire calcium dans la cellule qui va dépolariser la cellule. C'est-à-dire qu’au lieu de garder son potentiel de membrane normal la cellule va voir disparaitre ce potentiel de membrane.
Il y a généralement des charges négatives dans la cellule, et si on fait rentrer des charges positives en excès sur les bords de la membrane, on va annuler cette différence de potentiel entre l’exterieur et l’intérieur de la cellule et on va donc dépolariser (enlever la polarisation)
C’est le fait d’appuyer sur la membrane, l’étirement de la membrane qui active ce canal ionique particulier.
Le stimulus mécanique qu’on applique sur un récepteur va entrainer un potentiel de récepteur. Plus on appuiera fort (par exemple ici, entre 100 et 200) plus il y aura de modifications du potentiel de membrane. On va avoir un potentiel de récepteur, ce potentiel va se propager sur le récepteur. Et ce récepteur va traduire cela en potentiel d’action.

Exemple +++: on appuyer sur la peau. On a fait rentrer des charges positives dans la cellule. Le potentiel de membrane de cette cellule était de -70mv, elle est chargée négativement à l’intérieur. Et on lui fait rentrer des charges positives. Elle est donc de moins en moins négative et elle va passer ou elle peut passer un certain seuil.
La particularité de ce potentiel de récepteur est qu’il se perd avec la distance. Plus on s’éloigne, plus on se rapproche de l’extrémité du récepteur, plus on perd l’intensité du potentiel. La propagation est decrémentielle et locale
Ce PR est donc dépendant de :- la distance : le potentiel est decrémentiel, c’est de moins en moins avec la distance du point de stimulation- l’intensité : plus j’appuie fort, plus I est grande. L’amplitude du PR est proportionnelle avec l’intensité.
Ce potentiel de récepteur va donner lieu à un potentiel d’action si et seulement si on a appuyé suffisamment pour dépasser le seuil. Il ne faut pas oublier que la génération du PA se fait à l’extrémité du récepteur : « TRIGGER ZONE »
Il faut donc que le PR gradué réussisse à traverser tout le récepteur pour arriver à la sortie de l’axone, avec une amplitude suffisamment grande, pour générer un PA.
Ici, on n’en a pas, on n’a pas appuyé assez fort. Et d’ailleurs c’est vrai, on peut le démontrer, on peut toucher tout doucement quelqu’un sans qu’il ne le sente.
Ensuite il y a transduction, au moment où, à l’extrémité de l’axone, on a passé le seuil. On passe le seuil, et on déclenche un nombre de potentiel d’action. Et à partir de ce moment là, c’est la loi du tout ou rien, le PA se propage, de façon saltatoire, sur les fibres myélinisées du système lemniscal (de nœud de Ranvier en nœud de Ranvier).
Grâce à cette conduction saltatoire, l’influx nerveux ne s’arrête pas. On passe dans le numérique, le potentiel générateur (= gradué= récepteur) c’était de l’analogique.
Et on transmet cet influx nerveux sur le premier neurone, puis grâce à des synapses, sur le deuxième et le troisième.

C- Modulation :
1- Champ récepteur :
C’est une zone innervée par un récepteur plus ou moins grande suivant les zones de l’organisme. L’intensité du message concerné va être différente en fonction de la distance (Si on est proche ou loin du centre du champ récepteur)
L’extrémité de la fibre nerveuse se divise en petites parties au niveau de la peau.
- Chaque neurone a sa propre zone de réception qui chevauche la zone de réception des neurones voisins
- Chaque neurone a une sensibilité différente au stimulus en fonction de la zone du champ récepteur stimulée
La fibre colorée à un champ récepteur plutôt petit. Une petite zone sous sa dépendance qui se chevauche légèrement avec la suivante.
Si on stimule une zone de la main avec une aiguille, on enregistre l’activité du nerf médian par exemple. Quand je stimule là, j’active ce neurone là. Quand je stimule ici, j’active ce neurone ci…
Ce champ récepteur est donc plus ou moins grand, et cela en fonction des récepteurs.
On rappelle que Meissner c’est fin, par sa localisation et par son champs récepteur.De même pour Meckel.Pacini et Ruffini ce n’est pas fin. On peut activer le même récepteur sur une grande distance.
Sur le schéma, on voit qu’il y a des zones plus ou moins fines.

Par ailleurs, les récepteurs ont une caractéristique principale.C’est soit de s’adapter rapidement, soit de s’adapter lentement. Et on appelle ça une réponse phasique ou une réponse tonique.
Réponse phasique : exemple du corpuscule de Meissner qui est sensible au changement. Des potentiels d’action sont transmis au moment où on appuie et au moment où on relâche. Il y a une adaptation rapide.
Réponse tonique : Merckel, lui, continue à décharger pendant toute la durée de la pression. Mais il décharge de moins en moins avec le temps.
Il y a des récepteurs phasiques, sensibles au changement (on appuie ou non) et des récepteurs toniques qui déclenchent des potentiels d’action permanents tant que dure la stimulation (on appuie combien de temps)
2 - Intensité de la stimulation
Comment code-t-on l’intensité de la stimulation?
Par deux mécanismes différents :
-soit le nombre de terminaisons nerveuses activées : si on a une pression plus forte on va le sentir parce qu’on aura plus de fibres activées.
- Et ca va se traduire par la fréquence de stimulation : Plus ca va être intense, plus le potentiel gradué (ou PR) sera important. Cette intensité de stimulation de PR va être traduite en fréquence de stimulation de PA. C’est une façon de traduire l’intensité de la stimulation.L’intensité de stimulation est transmise par la fréquence de PA
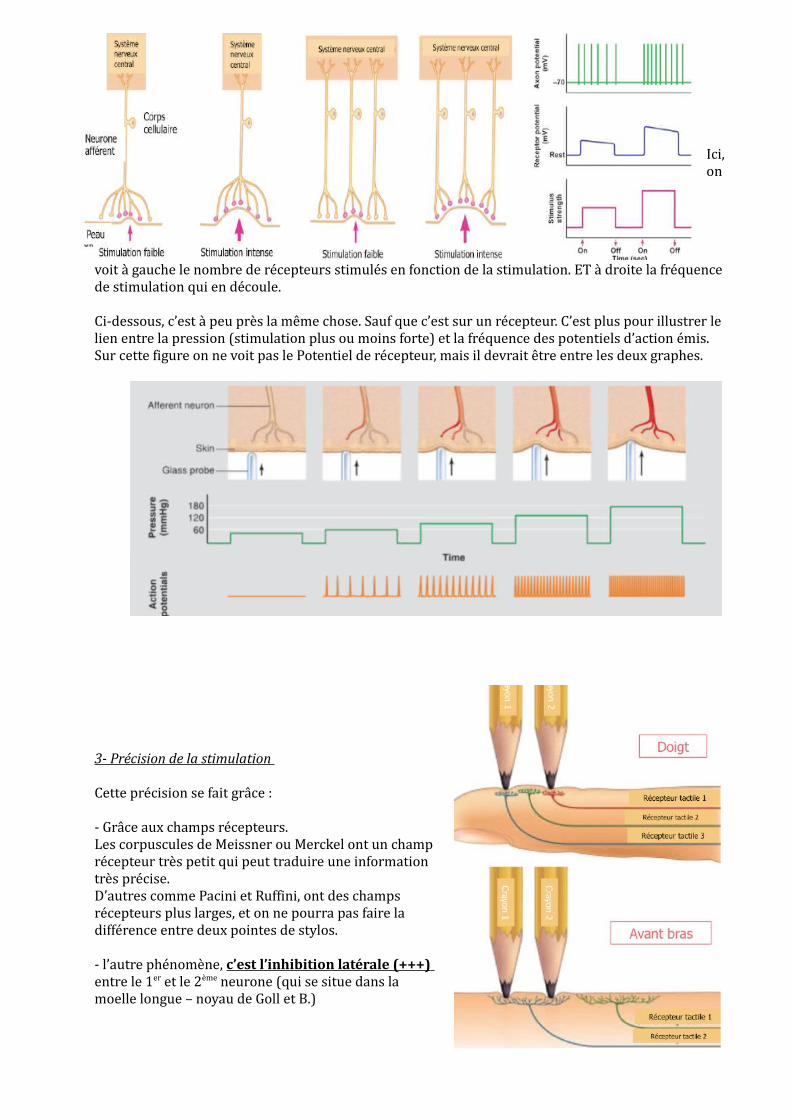
Ici, on
voit à gauche le nombre de récepteurs stimulés en fonction de la stimulation. ET à droite la fréquence de stimulation qui en découle.
Ci-dessous, c’est à peu près la même chose. Sauf que c’est sur un récepteur. C’est plus pour illustrer le lien entre la pression (stimulation plus ou moins forte) et la fréquence des potentiels d’action émis. Sur cette figure on ne voit pas le Potentiel de récepteur, mais il devrait être entre les deux graphes.
3- Précision de la stimulation
Cette précision se fait grâce :
- Grâce aux champs récepteurs. Les corpuscules de Meissner ou Merckel ont un champ récepteur très petit qui peut traduire une information très précise. D’autres comme Pacini et Ruffini, ont des champs récepteurs plus larges, et on ne pourra pas faire la différence entre deux pointes de stylos.
- l’autre phénomène, c’est l’inhibition latérale (+++) entre le 1er et le 2ème neurone (qui se situe dans la moelle longue – noyau de Goll et B.)
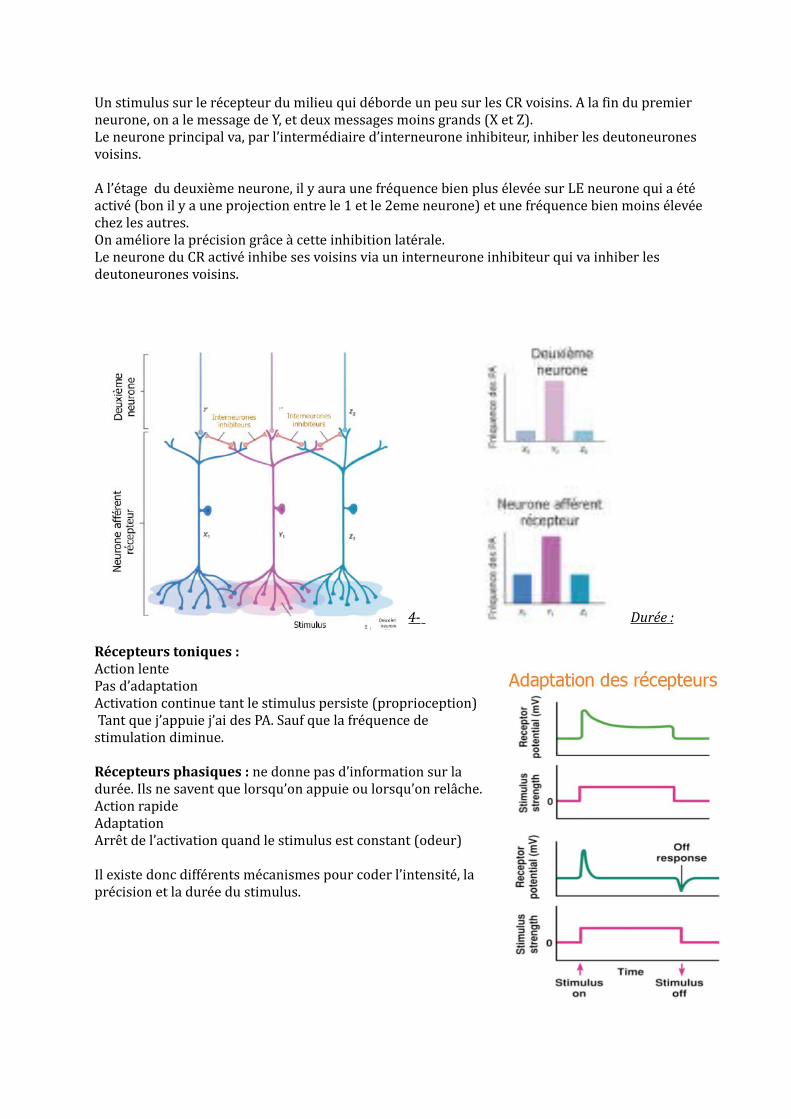
Un stimulus sur le récepteur du milieu qui déborde un peu sur les CR voisins. A la fin du premier neurone, on a le message de Y, et deux messages moins grands (X et Z). Le neurone principal va, par l’intermédiaire d’interneurone inhibiteur, inhiber les deutoneurones voisins.
A l’étage du deuxième neurone, il y aura une fréquence bien plus élevée sur LE neurone qui a été activé (bon il y a une projection entre le 1 et le 2eme neurone) et une fréquence bien moins élevée chez les autres.On améliore la précision grâce à cette inhibition latérale. Le neurone du CR activé inhibe ses voisins via un interneurone inhibiteur qui va inhiber les deutoneurones voisins.
4- Durée :
Récepteurs toniques : Action lentePas d’adaptationActivation continue tant le stimulus persiste (proprioception) Tant que j’appuie j’ai des PA. Sauf que la fréquence de stimulation diminue.
Récepteurs phasiques : ne donne pas d’information sur la durée. Ils ne savent que lorsqu’on appuie ou lorsqu’on relâche. Action rapideAdaptationArrêt de l’activation quand le stimulus est constant (odeur)
Il existe donc différents mécanismes pour coder l’intensité, la précision et la durée du stimulus.
D – Transmission
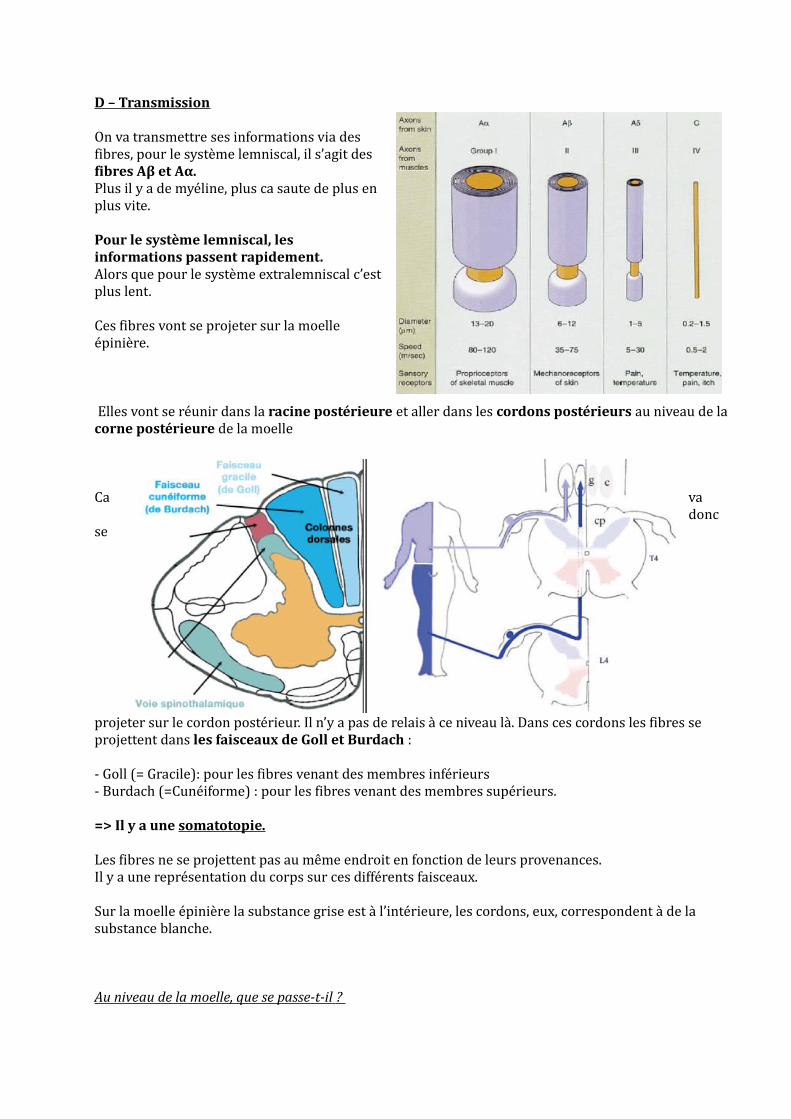
On va transmettre ses informations via des fibres, pour le système lemniscal, il s’agit des fibres Aβ et Aα. Plus il y a de myéline, plus ca saute de plus en plus vite.
Pour le système lemniscal, les informations passent rapidement.Alors que pour le système extralemniscal c’est plus lent.
Ces fibres vont se projeter sur la moelle épinière.
Elles vont se réunir dans la racine postérieure et aller dans les cordons postérieurs au niveau de la corne postérieure de la moelle
Ca va donc
se
projeter sur le cordon postérieur. Il n’y a pas de relais à ce niveau là. Dans ces cordons les fibres se projettent dans les faisceaux de Goll et Burdach :
- Goll (= Gracile): pour les fibres venant des membres inférieurs- Burdach (=Cunéiforme) : pour les fibres venant des membres supérieurs.
=> Il y a une somatotopie.
Les fibres ne se projettent pas au même endroit en fonction de leurs provenances. Il y a une représentation du corps sur ces différents faisceaux.
Sur la moelle épinière la substance grise est à l’intérieure, les cordons, eux, correspondent à de la substance blanche.
Au niveau de la moelle, que se passe-t-il ?

On monte par les faisceaux de Goll et Burdach mais également, il y a un relais local sur un motoneurone et des interneurones (partie II).
On remarque dans les faisceaux graciles et cunéiformes les différentes parties du corps. A l’intérieur, la région sacrée. Intermédiaire, les régions thoracique et lombaire.En périphérie, les membres supérieures.
A l’exterieur, c’est le faisceau cunéiforme/BurdachA l’intérieur, c’est le faisceau gracile/Goll
Il y a encore une fois une représentation du corps humain
A quoi sert le motoneurone ? Il sert à un reflexe. - Il y a la voie afférente, sensitive, qu’on vient de voir.- Un relais central ou pas- Et une voie efférente, vers l’effecteur.
Donc trois parties : Afférente – Centrale – Efférente
Le reflexe du motoneurone, c’est le reflexe myotatique.
A l’étirement d’un muscle, au lieu d’avoir une force proportionnelle à l’étirement de mon muscle, j’ai une force bien plus importante, il y a un phénomène actif.
L’allongement, n’explique pas à lui seul l’augmentation de force, qui se limitera à ça (E)
Il y a beaucoup plus de force qui se dégage, il y a un phénomène actif, c’est le reflexe myotatique qui vient du fuseau neuromusculaire.
Il y a des fibres sensitives au niveau du muscle qui sont sensible à l’étirement. Quand ces fibres sont activées, elles vont faire relais avec le motoneurone. Donc au lieu d’aller uniquement dans les faisceaux de Goll et Burdach, elles vont aussi aller vers le motoneurone alpha qui lui va contracter les fibres du muscle en question.

Ex : je me pense en avant. J’étends mon triceps sural (derrière au niveau du mollet). Deux possibilités : - je me casse la gueule. - j’active mon fuseau neuromusculaire, j’étends mon fuseau neuromusculaire.
Je vais par reflexe contracter mon triceps sural et je me redresse.
Si je te lance une balle, et que tu essaies de la rattraper. Ton bras ne va pas tomber sous le poids. Parce que, par reflexe, tu vas contracter ton bras.
L’étirement du muscle, va entrainer la contraction.
De même, si je prends un verre d’eau et que je remplis rapidement d’eau, il est peu probable que je le lache. Grace au fuseau neuromusculaire, je vais augmenter la force, parce que je vais voir que mon muscle s’étend, pour pouvoir continuer à porter le verre. Après ca devient actif, mais il y a d’abord une partie reflexe.
On a pu prouver que c’était un seul neurone, parce qu’on sait qu’il faut moins d’une milliseconde pour traverser la moelle épinière. Quand on stimule un neurone afférent, qu’on enregistre l’activité à l’entrée de la moelle (1) et celle du motoneurone de l’autre côté (2) on obtient la différence de temps entre l’arrivée du potentiel en (1) et l’arrivé du potentiel en (2).
Et on a calculé que c’était la durée d’une seul synapse, de l’ordre d’une milliseconde. On a pu dire que c’était un reflexe. Le reflexe myotatique, qui est un reflexe monosynaptique. Il n’y a que le passage du neurone afférent au neurone efférent.
Au niveau articulaire, il y a d’autres types de récepteurs. Les articulations peuvent faire mal, on peut sentir la position du l’articulation. Il y a des organes de Golgi (partie II) et surtout des corpuscules de Ruffini qui permette de sentir l’étirement articulaire. On peut voir les modifications au niveau articulaire et ca permet la proprioception. On connait le sens de position des membres parce qu’on connait le sens de l’étirement des articulations.
Si je connais la position de mon membre droit, sans regarder ma main droite je peux faire la même chose avec l’autre main. Allez viens ! Regarde tout ce qu’on peut faire ! C’est génial nan ?

ET enfin, on monte dans la moelle :
Certaines fibres ont fait le reflexe myotatique, d’autres montent dans la moelle par la région cordonale postérieure (donc par les faisceaux de Goll et Burdach).On rappelle, de l’intérieure vers l’extérieure : Sacré, lombaire, thoracique cervical.
Bref, on remonte dans la moelle spinale (=moelle épinière) pour croiser au niveau de la moelle allongée basse. Ca décusse, ça croise à ce niveau là.
Moralité, on pourra s’avoir si un patient à une atteinte de la sensibilité au tact ou à la proprioception de l’autre coté que les lésions. On pourra localiser la lésion.
Donc ça croise, avec relais dans les noyaux de Goll et Burdach (même noms que les faisceaux)Le deuxième neurone nait de la moelle allongée basse. C’est là que ce fait le relais. Et là je croise. Et je passe de l’autre côté.
Au niveau de la protubérance (=le pont) :
Un peu plus haut, on est au niveau de la zone beaucoup plus antérieure : le LEMNISQUE MEDIAN (qui donne son nom à la voie, la voie lemniscale qui passe par le lemnisque médian).
Encore une fois, on a une représentation des fibres selon une cartographie prédéfinie : sacré, lombaire, thoracique, cervicale.
« Je ne vous demande pas de connaitre si le sacré est antérieur, latéral, peut importe, juste de savoir qu’il y a une représentation, qu’il y a une somatotopie. Que le corps est représenté, et que toutes les
fibres ne sont pas mélangées. Il y a les fibres sacrées qui sont différenciées des fibres lombaires, thoracique et cervicale. De la même façon que dans les faisceaux de Goll et Burdach. Il ne faut
certainement pas l’apprendre par cœur. Quoique, comme je suis vache, et que moi je ne le connais pas par cœur, je vais vous demander à vous de l’apprendre par cœur, c’est ce qu’on appelle le
syndrome du petit chef : Il faut en chier ! »
Vous ferez donc en fonction de votre envie/interprétation hein !
A Gauche, la Moelle allongée basse avec la décussation et le relais avec le 2ème neurone.

A droite, la Moelle allongée, avec le Lemnisque Médian.Au niveau du tronc cérébral :
- On est vraiment au niveau de la partie médiane du pont, on le voit bien avec le faisceau pyramidal qui passe devant et le lemnisque médian. Avec encore la somatotopie.
- Un peu plus haut, au niveau des pédoncules cérébraux, il y a encore le lemnisque médian qui passe à côté d’un noyau rouge (parce qu’il apparait rouge quand on coupe un cerveau en tranche)
- Le deuxième neurone est parti du TC, de la moelle allongée basse, jusqu’au Thalamus. Au niveau du Thalamus, il fait relais avec le troisième neurone. Mais en controlatéral. On a changé de côté.
- Dans le thalamus, il y a plein de noyaux. Il y en a deux qui nous intéressent particulièrement. Ils sont relativement externes, c’est le Ventro-postéro-Latéral (VPL) et le ventro-postéro-médian (VPM).
C’est sur ces deux noyaux que se projettent les voies lemniscales. Et là encore, sur le schéma agrandi en forme de berlingot, la partie VPL dans laquelle passent les fibres sacrées et du Tronc. Et le VPM dans lequel passent les fibres qui véhiculent la sensibilité extralemniscale proprioceptive/tact fin de la face.

Il y a une somatotopie thalamique. Là encore, le schéma corporel est conservé pour les fibres.
Au niveau du VPM :
Il a fallu récupérer les fibres venant de la face. Les fibres du tact fin/proprioceptive au niveau de la face ne passent pas par le faisceau de Goll et Burdach, mais grâce au nerf V, le trijumeau.
Ca passe par le noyau de la 5ème paire crânienne dans le TC et ça rejoint le thalamus au niveau du VPM.
Et ça va se projeter sur le cortex avec la encore une somatotopie.
Entre les deux hémisphères, les membres inférieurs sont représentés. Puis sur chaque hémisphère, le Thorax sur la partie supérieure, puis la main très largement représentée, puis la figure et les lèvres, le larynx et puis la langue.
Toute cette zone là (en hachurée, en arrière de la scissure de Rolando) est vascularisée par l’artère cérébrale moyenne. C’est l’artère qui se bouche le plus facilement dans les AVC. Dans les AVC l’homoculus moteur est similaire au sensitif et cela entraine un déficit brachio-facial.
Cortex sensitif est localisé en arrière de la scissure de Rolando, dans le lobe pariétal.
L’homoculus sensitif : les mains sont très grosses, comme les lèvres car elles sont très sensibles. Les bras et le tronc, au contraire, sont tout petits car ils ont un faible pouvoir discriminatif.

E. Intégration
Organisation en Colonne du Cortex :
En arrière de Rolando, on a le cortex sensitif primaire (SI) avec la représentation des différentes parties du corps. La circonvolution pariétale antérieure coupée en tranche à plusieurs parties : 4 en particuliers (partie II)
Aux pieds du SI, on a le cortex somatosensitif secondaire (SII).En arrière du SI, on a le cortex pariétal postérieur (SIII) avec les aires 5 et 7 de Brodmann
Il y a d’abord une aire de projection de la proprioception, c’est l’aire 3a. Sur l’aire 3b, qui est une aire d’entrée principale, il y a la sensibilité tactile et vibratoire qui elle-même va aller se projeter (après ses fibres vont faire) par ses fibres qui vont faire des communications avec l’aire 1 et 2 pour la texture (A1) pour la taille et les formes (A2)
Proprio récepteurs vont sur 3a du SI, puis ça va se projeter à nouveau sur l’aire 2 puis sur le Cortex pariétal postérieur sur l’aire 5 et 7 (Partie II)
Sur l’aire 3b, les fibres se projettent, puis sur l’Aire 1 du SI, puis SII.
Et vous aurez la suite dans la fameuse partie II.






![brirsa.files.wordpress.com · Web view2020. 4. 1. · Exercice de compréhension sur le CORROYAGE – professeur M. MERCADIER – IRSA menuiserie [Tapez le texte]](https://img.pdfslide.fr/doc/110x75/60128292864677645a7c496e/web-view-2020-4-1-exercice-de-comprhension-sur-le-corroyage-a-professeur.jpg)






















