Embed Size (px)
Citation preview

Diversité des pollinisateurs et de la ressource florale dans les vergers de Martinique
Clara SINGH
Master 2 Ecologie et Biosciences de l’Environnement
Stage encadré par Eddy Dumbardon-Martial (FREDON)
En partenariat avec François Meurgey (SHNLH )
(Mars – Août 2015)
Tuteur de stage : Annie OUIN
Correcteur : Aude VIALATTE

Remerciements
Mes premiers remerciements s’adressent à Eddy Dumbardon-Martial pour m’avoir encadrée et guidée tout au long de ce stage. La confiance qu’il m’a accordée m’a permis de réaliser cette mission en toute autonomie. Sa gentillesse et son calme m’ont aidée à gérer les imprévus et à surmonter les difficultés du stage. Je le remercie aussi d’avoir facilité mon acclimatation à la Martinique.
Je remercie Annie Ouin, qui depuis la métropole m’a confortée dans le bon déroulement de ma mission. Cela a permis de maintenir un contact rassurant avec la France et ce, malgré la distance.
Je remercie Denise Duféal pour m’avoir accueillie à la FREDON, me permettant de réaliser ce stage qui a parfaitement répondu à mes attentes.
Merci à Chloé Pierre, les échanges que nous avons eus dans le cadre de ce travail ont été enrichissants. Elle m’a apporté son aide et son soutien jusqu’à la fin de cette mission. .
Un grand merci à Alain Pauly pour son travail d’identification et pour m’avoir formée à distance à identifier certains spécimens. Merci a François Meurgey pour l’identification des abeilles sauvages de référence ainsi que pour ses conseils.
Je remercie les stagiaires, Lucie Abolivier et Eddy Barthélémy qui ont rendu ce stage convivial. Un grand merci également à l’ensemble du personnel. Merci à Pierre-Damien, Hilaire, Sébastien et Jean-Claude pour m’avoir véhiculée de mon domicile à la FREDON. Merci à Alain, Armand, Caroline, Synthia, Denis, Jessica, Rémi, Teddy, Valérie et Jacqueline pour leur accueil et leur disponibilité.
Un grand merci à tous les agriculteurs que j’ai rencontrés dans le cadre de cette mission. Ils m’ont tous merveilleusement bien accueillie, me faisant découvrir les spécialités de leur terroir. La campagne de terrain a ainsi pu se réaliser dans de bonnes conditions.
Merci à l’association AVJT (Association des Vergers et Jardins Tropicaux) grâce à laquelle j’ai pu entrer en contact avec de nombreux agriculteurs.
Merci à ma famille, toujours présente pour me soutenir dans mes études. Merci à Pedro de ne jamais être très loin et de m’épauler en toutes circonstances. Merci à Alison et Tricia mes camarades de l’ENSAT avec qui j’ai partagé mes aventures à distance. Cette année à été top grâce à vous.
Merci encore à Annie Ouin et Romain Carrié de l’UMR Dynafor (INRA) de m’avoir poussée vers cette expérience originale et formatrice.
Merci à l’équipe d’internes en ophtalmologie du CHU La Meynarde d’avoir assuré mon suivi pendant plusieurs mois et sans qui mon stage n’aurait pas pu se poursuivre.
Un grand merci à toutes les personnes que j’ai rencontrées en Martinique, grâce auxquelles, ce séjour restera inoubliable.

Table des matières Remerciements Table des matières Tables des figures
Résumé .................................................................................................................................................... 6
Introduction ............................................................................................................................................. 1
1. Contexte scientifique de l’étude .......................................................................................................... 2
2.1 Importance de la biodiversité dans l’établissement des services écosystémiques ........................ 2 2.2 La pollinisation entomophile : un service écosystémique indispensable à la production agricole 2
2.3Les Associations ruches-vergers dans le monde ............................................................................ 3 2.3 Etat des lieux sur l’association ruches-vergers dans les Antilles .................................................. 4 2.4 Prise en considération des pollinisateurs sauvages dans ces systèmes agricole............................ 4 2.5 Les pollinisateurs sauvages sont étroitement liés aux conditions du milieu ................................. 5 2.6 La FREDON un syndicat professionnel qui promotionne les principes agro-écologiques. .......... 6
2.6 Les missions du stage .................................................................................................................... 7
2. Matériels et Méthodes ......................................................................................................................... 9
2.1 Le site d’étude .............................................................................................................................. 9
2.1.1. Eléments de géographie ........................................................................................................ 9
2.1.2. Particularités climatiques ...................................................................................................... 9
2.1.3. Choix et localisation des parcelles étudiées .......................................................................... 9 2.2 Les cultures fruitières étudiées ................................................................................................... 11
2.3 Les insectes pollinisateurs : modèle biologique .......................................................................... 12 2.3.1. Choix des pollinisateurs étudiés ......................................................................................... 12 2.3.2. Les Apoïdes ........................................................................................................................ 12
2.3.3. Les syrphes ......................................................................................................................... 13
2.4 Le protocole de terrain ............................................................................................................... 14
2.4.2. Capture des pollinisateurs ................................................................................................... 14
2.4.3 Les relevés floristiques ........................................................................................................ 17
2.4.4Estimation des éléments constitutifs de l’environnement proche ......................................... 17 2.5 Le protocole de laboratoire ........................................................................................................ 18
2.6 Enquête auprès des arboriculteurs au sujet de leurs techniques et pratiques de désherbage ...... 18
2.7 Analyses statistiques ................................................................................................................... 18
2.7.1. Estimation de l’effort d’échantillonnage avec des courbes d’accumulation....................... 19
2.7.2. Etude de l’abondance et de la diversité des pollinisateurs selon les sites ........................... 19 2.7.3. Analyse de l’effet de l’heure de capture sur l’abondance et la diversité des pollinisateurs 20
2.7.4 Effet des adventices sur l’abondance et la diversité des pollinisateurs ............................... 20 2.7.5. Etudes des interactions plantes pollinisateurs ..................................................................... 20 2.7.6. Effet de l’hétérogénéité à l’échelle de la parcelle et effet de l’environnement proche sur les communautés de pollinisateurs. .................................................................................................... 21
2.7.7. Effet des pratiques .............................................................................................................. 21
3. Résultats........................................................................................................................................ 22
3.1 Le matériel collecté ..................................................................................................................... 22
3.1.1 Représentativité des groupes collectés .......................................................................... 22 3.1.2 Evaluation de l’effort d’échantillonnage ............................................................................. 23
3.2 Diversité et abondance des pollinisateurs .................................................................................. 24 3.2.1 Les pollinisateurs recensés .................................................................................................. 24
3.2.2 Abondance et diversité des pollinisateurs selon les sites ..................................................... 26 3.2.3 Effet de l’heure de capture sur l’abondance et la diversité des pollinisateurs ..................... 27
3.3 Effet des adventices sur les communautés de pollinisateurs ....................................................... 28 3.3.1 Bilan sur les relevés floristiques .......................................................................................... 28
3.3.2 Effet de la diversité des adventices sur l’abondance et la diversité des pollinisateurs ........ 28
3.3.3 Effet du nombre d’inflorescences et du recouvrement floral sur l’abondance et la diversité des pollinisateurs .......................................................................................................................... 29
3.3.4 Effet de la diversité des plantes en fleurs sur l’abondance et la diversité des pollinisateurs 31

3.3.5 Description de l’interaction plantes-pollinisateurs .............................................................. 33 3.4 Effet de l’hétérogénéité à l’échelle intra-parcellaire et effet de l’environnement proche sur l’abondance et la diversité des pollinisateurs .................................................................................... 34
3.4.1 Effet de la bordure de parcelle sur l’abondance et la diversité des pollinisateurs. .............. 34
3.4.2 Effet des éléments constitutifs du périmètre de la parcelle sur l’abondance et la diversité des pollinisateurs .......................................................................................................................... 35
3.5 Effet des techniques et pratiques de gestion des adventices sur les communautés d’adventices et sur les communautés de pollinisateurs. ............................................................................................. 36
4. Discussion ......................................................................................................................................... 37
4.1Méthodologie et effort de prospection ......................................................................................... 37
4.1.1 Les trois techniques de captures sont complémentaires ...................................................... 37 4.1.2 Un effort d’échantillonnage suffisant mais des localités sur l’île restent à étudier ............ 37
4.1.3 Les pollinisateurs ont une activité de butinage plus importante le matin ............................ 38 4.2 Les Apoïdes et les syrphes capturés dans les couverts des vergers ............................................ 38
4.2.1. Une faible diversité des pollinisateurs recensés en raison des particularités du milieu insulaire ........................................................................................................................................ 38
4.2.2. Des communautés de pollinisateurs non équitables .......................................................... 39 4.2.3. La diversité et l’abondance des pollinisateurs diffère selon les parcelles mais les espèces sont représentées de manières homogènes à l’échelle globale ..................................................... 39
4.3 La ressource florale explique en partie la présence des pollinisateurs dans les vergers ............. 40 4.3.1 La ressource florale a un effet positif sur l’abondance des pollinisateurs mais aucun effet sur la diversité n’a été démontré. Il y a un effet de l’échelle et/ou de la technique de capture sur ce résultat. ..................................................................................................................................... 40
4.3.2 Le caractère plus ou moins attractif des espèces de fleurs atténue les effets de l’abondance et de la diversité de la ressource florale sur les communautés de pollinisateurs .......................... 41 4.3.3 Une étude sur une échelle de temps plus longue permettrait peut-être de voir l’effet de la richesse spécifique des adventices (en fleurs et non en fleurs) sur les communautés de pollinisateurs. ................................................................................................................................ 41
4.3.4 Il existe des préférences, certaines fleurs des couverts sont plus butinées que d’autres ...... 42
4.3.5 Ingénierie et contribution à la mise en place du système agro-écologique ......................... 43 Conclusion ............................................................................................................................................. 46
Bibliographie ......................................................................................................................................... 47
Annexes
Annexe 1 : Abondance des différentes familles d’insectes capturés au fauchage
Annexe 2 : Abondance des différentes familles d’insectes capturés à l’aide des bols colorés
Annexe 3 : Extrait de la collection entomologique regroupant les pollinisateurs recensé
Annexe 4 : Planning d’une journée de terrain pour la mise en place du dispositif 1
Annexe 5 : Etude de la diversité des pollinisateurs à l’aide des bols colorés

Table des figures
Figure 1 : Schéma récapitulatif des différentes hypothèses posées……………………. 8
Figure 2 : Carte de localisation des parcelles échantillonnées pour l’étude………….. 10
Figure 3 : Représentation du dispositif de terrain n°1………………………………… 15
Figure 4 : Représentation du dispositif de terrain n°2………………………………… 16
Figure 5 : Abondance de chaque groupe de pollinisateurs (abeilles sauvages, abeilles domestiques et syrphes) selon la technique de capture (piégeage par bols colorés, fauchage et chasse à vue)……………………………………………………………….
22
Figure 6 : Courbe d’accumulation de la richesse spécifique cumulée des pollinisateurs capturés au fauchage. 60 sites ont été échantillonés (1site=1 transect)....
23
Figure 7 : Courbe d’accumulation de la richesse spécifique cumulée des pollinisateurs capturés à vue. 20 sites ont été échantillonnés (1 site =1 parcelle)……...
23
Figure 8 : Boîtes à moustaches des abondances de pollinisateurs selon la parcelle échantillonnées…………………………………………………………………….........
26
Figure 9 : Boîtes à moustaches des abondances de pollinisateurs selon l’heure de capture par fauchage……………………………………………………………………
27
Figure 10 : Abondance des pollinisateurs capturés à vue en fonction du nombre d’inflorescences à l’échelle de la parcelle……………………………………………....
30
Figure 11 : Abondances des pollinisateurs en fonction de la diversité des plantes en fleurs……………………………………………………………………………………..
31
Figure 12 : Courbe d’accumulation de la richesse spécifique cumulée des pollinisateurs capturés à vue sur 35 espèces végétales en fleurs
32
Figure 13 : Indice Trophic Niche Breadth selon l’espèce de pollinisateur capturé en chasse à vue……………………………………………………………………………...
33
Figure 14 : Nombre de visites (toutes espèces de pollinisateurs confondues) selon les familles des fleurs butinées Les données sont issues des captures à vue………………..
33
Figure 15 : Nombre de visites (toutes espèces de pollinisateurs confondues) pour chaque espèce en fleurs à partir des captures de la chasse à vue………………………..
34
Figure 16: Boîtes à moustache de la richesse spécifique des adventices selon la position du transect……………………………………………………………………...
35

Résumé L’étude porte sur la diversité des pollinisateurs dans les vergers de Martinique et se concentre
surtout sur l’effet de la flore adventice sur les communautés de pollinisateurs. En effet, l’un
des objectifs du projet Agriculture Apiculture Environnement est de faire un recensement des
pollinisateurs dans les vergers, puis, de détecter les leviers d’action favorisant ces
pollinisateurs dans les exploitations. L’étude se concentre surtout sur l’effet de la flore
adventice des couverts herbacés sur les pollinisateurs. Les groupes étudiés ici sont les
Apoïdes et les Syrphes. 20 parcelles de goyaves et d’agrumes ont été sélectionnées sur l’île et
des relevés au filet entomologiques (fauchage et chasse à vue) ont été réalisés. Un premier
essai de piégeage à l’aide de « bols colorés » est aussi intégré à l’étude. Des relevés
floristiques dans les couverts sont effectués et sont mis en relation avec les communautés de
pollinisateurs. Les effets des éléments semi-naturels et de la gestion de l’enherbement sur les
pollinisateurs sont abordés. L’étude montre que l’abondance et la diversité de la flore
adventice explique en partie les abondances et diversités de pollinisateurs observées.
Certaines espèces de fleurs sont plus attractives pour les pollinisateurs et ont un intérêt
agronomique plus prononcé. Le couvert, selon les cortèges floristiques qui le composent peut
être utilisé pour attirer les pollinisateurs et favoriser la pollinisation de la plante cultivée.
Aucun effet des éléments semi-naturels n’a été démontré et une étude plus précise à plus
grande échelle est nécessaire. L’effet des techniques de désherbage n’a pas été analysé car
tous les agriculteurs procèdent à un désherbage mécanique. Cette étude a permis de faire un
état des lieux des pollinisateurs dans les vergers et la méthodologie devra par la suite être
affinée pour répondre aux nouveaux questionnements que ce stage aura permis de soulever.
Mots clefs : agroécologie, pollinisateurs, flore adventice, vergers

Abstract The study focuses on pollinator diversity in the orchards of Martinique and concentrates
primarily on the effect of the weeds on pollinator communities. Indeed, one of the objectives
of Agriculture Apiculture and Environnement project is to make a census of pollinators in
orchards, then detect levers supporting these pollinators on farms. The groups studied here are
the Apoidea and Hoverflies. Twenty fields of guavas and citrus were selected on the island
and entomological catch with a net (mowing and hunting view) have been achieved. A first
trapping test using "colored bowl" is also integrated into the study. Floristic sample are
carried in vegetation covers and are linked with the pollinator communities. The effects of
semi-natural elements and the management of weeds on pollinators are addressed. The study
shows that the abundance and diversity of weeds flora partly explains the abundance and
diversity of pollinators observed. Some species of flowers are most attractive to pollinators
and have a more pronounced agronomic interest. The vegetation cover, according to the
processions that make up the flora can be used to attract pollinators and promote pollination
of the crop. No effect of semi-natural elements has been demonstrated and a more precise
study at a larger scale is required. The effect of weeding techniques was not analyzed because
all the farmers carry out mechanical weeding. This study enabled to do an inventory of
pollinators in orchards and methodology will then be refined to answer new questions that this
internship raised.
Key Words : Agroecology, polliniators, weeds flora, orchards.

1
Introduction
Après la seconde guerre mondiale, la modernisation de l’agriculture est une priorité, et
entraîne le passage d’une agriculture traditionnelle à une culture moderne lors de la
« Révolution verte » basée sur de nouveaux principes agronomiques. L’Europe, à travers la
Politique Agricole Commune soutient financièrement la production agricole et encourage les
innovations techniques et technologiques contribuant à augmenter les productions. Cette
période se traduit par un bouleversement des pratiques agricoles parmi lesquelles la
mécanisation, l’usage d’intrants (engrais et produits phytosanitaires), la sélection de variétés
plus productives occupent une place importante (FAO, 1996).
Si ce mode de production a permis de garantir la sécurité alimentaire à de nombreux pays et à
rétablir une bonne croissance démographique, nombreuses ont été les répercussions négatives
d’ordre socio-économique (augmentation des crédits agricoles, fragilisation financière des
petites exploitations, pertes des savoirs traditionnels…) et environnementales (pollution des
milieux, diminution de la biodiversité…), qui à ce jour, sont toujours d’actualité. Il existe un
consensus scientifique montrant d’une part que l’agriculture est l’une des premières cause du
déclin de la faune et de la flore sauvage, mais aussi, dans d’autres contextes, qu’elle est le
garant du maintien d’une certaines biodiversité. L’agriculture d’aujourd’hui se veut donc
conservatrice tout en valorisant et tirant profit des services écosystémiques qu’offre la
biodiversité (Le Roux, 2008).
A partir de 2003, la Politique Agricole Commune diminue sa part d’aide à la production
conditionnant ses aides à plusieurs mesures permettant la mise en place de pratiques agricoles
respectueuses de l’environnement : verdissement de la PAC (Heidseck & Allier, 2013). Inscrit
au cœur du Grenelle de l’environnement, le Plan Ecophyto a pour objectif de réduire de façon
significative l’usage des produits phytosanitaires tout en développant des pratiques
alternatives (www.agriculture.gouv.fr/ecophyto). Au cours d’une conférence nationale en
janvier 2015, leMinistère de l’agriculture, encourage l’agriculture française vers une transition
agroécologique (www.Agriculture.gouv.fr). Le projet Agroécologie proposé par le Ministère
qui se décline en plusieurs programmes nationaux (Plan Ecophyto, Plan Apiculture, Plan
Agriculture biologique…), a pour ambition d’inscrire l’agriculture française dans la
performance économique et environnementale(www.actu-environnement.com).
La Martinique qui subit, au même titre que les autres territoires européens, la crise alimentaire
et environnementale (pollution à la chlordécone(www.daaf972.agriculture.gouv.fr), tente
d’adapter ses systèmes de production tropicaux aux grandes orientations gouvernementales.

2
Fernandes (2009) met cependant en évidence que l’absence de référentiels et le faible niveau
d’appuis techniques adaptés aux conditions locales constituent quelques-uns des freins à
l’innovation technique de l’agroécologie en Martinique.La FREDON (Fédération Régionale
de défense contre les organismes nuisibles), un syndicat mixte professionnel participe à
l’établissement de ces référentiels techniques et scientifiques et met en place différentes
actions qui s’inscrivent toutes dans une démarche agro-écologique.Plusieurs actions portent
sur les auxiliaires des cultures et sur les services écosystémiques associées. Depuis 2014, le
projet Agriculture Apiculture Environnement aspire à mettre en place un système agro-
écologique associant les productions végétales et l’activité apicoles tout en favorisant les
pollinisateurs sauvages dans les milieux cultivés. C’est dans ce contexte que s’inscrit ce stage
de fin d’étude qui a pour objectif , dans un premier temps de faire un recensement des
pollinisateurs domestiques et sauvages dans les vergers. Dans un second temps, cette mission
consistera à détecter les potentiels leviers d’actions pouvant être utilisés afin de favoriser ces
pollinisateurs dans les exploitations des arboriculteurs et à terme d’améliorer la pollinisation
des productions fruitières.
1. Contexte scientifique de l’étude
2.1 Importance de la biodiversité dans l’établissement des services écosystémiques
Au sein d’un agroécosystème la biodiversité assure l’approvisionnement continu en biens et
services (Altieri, 1999). Dans ce contexte, l’agroécologie intègre aussi la notion de service
écosystémique, c’est à dire un service fourni parla nature qui améliore le bien-être humain
(Daily, 1997)et dans lequel la diversité et l’abondance des organismes assurent le bon
fonctionnement de l’agroécosystème(FAO, 2008 ; Naeem et al, 1995).Plus le système
agricole abrite une grande diversité d’organismes, plus il sera à même de résister aux
agressions et de se remettre après une perturbation (Tilman et al, 1997 ; Walker et al, 1992).
Cette théorie peut s’appliquer à différents services écosystémiques tels que la décomposition
de la matière organique, le contrôle biologique des ravageurs ou la pollinisation (Winfree &
Kremen, 2009).
2.2La pollinisation entomophile : un service écosystémique indispensable à la production agricole
Actuellement, la pollinisation retient l’attention du fait du déclin massif de pollinisateurs
observé à l’échelle mondiale qui inquiète en raison de leur intervention cruciale dans la
production agricole (Rasmont, 1988 ; Williams, 1994).La pollinisation, étape incontournable

3
de la reproduction des plantes à fleurs sauvages et cultivées (Pesson & Louveau,
1995 ;Albrecht, 2007), se définit par le transport de grains de pollen, depuis les étamines
d’une fleur, jusqu’au pistil d’une autre fleur de la même espèce pour conduire à la
fécondation(Pesson & Louveau, 1995 ; O’Tool& Raw, 1999). Le transport du pollen peut se
faire par des agents abiotiques (anémophilie, hygrophylie) ou par des agents biotiques
(insectes, mammifères, oiseaux).Dans ce dernier cas, le mode de pollinisation le plus répandu
est la pollinisation entomophile basée sur une relation mutualiste entre l’insecte et l’espèce
végétale. L’insecte floricole se nourrit du nectar ou du pollen de la fleur et assure en échange
la reproduction et la pérennité de l’espèce végétale (Pesson & Louveau, 1995 ; O’Tool& Raw,
1999). De nombreux groupes taxonomiques tels que les diptères, coléoptères, lépidoptères
sont connus pour leur action de pollinisation. Cependant, l’abeille domestique présente une
activité pollinisatrice globalement plus élevée que celle des autresinsectes floricoles du fait de
la taille de ses colonies autant que ses caractères morphologiques, de son régime alimentaire
et de son comportement de butinage (Bourgeois, 2006 ; Le Féon, 2010 ; Winston, 1993).Du
fait de l’importance de la pollinisation dans la production agricole et la reproduction des fleur
sauvage, se service prend une valeur économique. La valeur de l’activité pollinisatrice des
insectes s’éleverait à153 milliards d’euros (Gallais et al, 2009). En effet, les insectes
pollinisateurs participeraient à 35% de la production mondiale de nourriture (Gallais, 2009) et
ont un effet direct sur les rendements (Corbet, 1992 ; Fohouo, 2002) en assurant la
reproduction de 70% des plantes cultivées (Klein, 2007).
2.3Les Associations ruches-vergers dans le monde Les performances de l’abeille domestique sont utilisées par certains agriculteurs dans le but
d’améliorer de manière qualitative et quantitative les rendements de diverses cultures. On
parle alors de « pollinisation dirigée » (Louveaux & Pesson, 1995).En Californie,
l’association entre les producteurs d’amandiers et les apiculteurs en est un parfait exemple et
constitue le système d’association le plus grand à l’échelle mondiale. En effet, la culture
d’amande nécessite aujourd’hui60 % des abeilles domestiques des Etats-Unis ce qui équivaut
à 2.5 ruches/ha lorsque les pollinisateurs sauvages sont en diapause (Summer & Boriss,
2006). En France, plusieurs études sont menées sur la pollinisation et les pollinisateurs en vue
d’une amélioration des productions. Le projet POLAPIS par exemple, vise à obtenir des
références pour une meilleure maîtrise de la pollinisation des cultures de colza en passant par
une gestion raisonnée des colonies d’abeilles domestiques et la favorisation de l’activité des
pollinisateurs sauvages (http://www.itsap.asso.fr).

4
2.3 Etat des lieux sur l’association ruches-vergers dans les Antilles Dans le souci d’une amélioration quantitative et qualitative de leur production, les
arboriculteurs de Martinique tententaussi de valoriser l’activité pollinisatrice des abeilles dans
leurs vergers en y installant des colonies d’abeilles domestiques sur la base d’une
collaboration avec les apiculteurs, eux-mêmes souvent en recherche de sites de pollinisation
durant une partie de l’année. Cependant, dans les Antilles, les références scientifiques,
techniques et économiques sur ce type d’association sont inexistantes localement et seules des
expériences empiriques au sein de certaines exploitations font état de ces systèmes sans
vraiment qu’il y ait un retour objectif des modalités de leur mise en place ni de leur impact
pour chacun des partenaires (Comm. E.Dumbardon-Martial). Afin d’accompagner les
agriculteurs dans cette démarche et de mettre en place une collaboration efficace et durable
entre les apiculteurs et les arboriculteurs,il est nécessaire d’acquérir des données sur les
pollinisateurs des vergers en Martinique. C’est dans ce contexte qu’intervient la FREDON par
son projet « Agriculture Apiculture et Environnement »qui vise à acquérir localement des
références afin de mettre en place un système agro-écologique associant arboriculture et
apiculture tout en favorisant les pollinisateurs sauvages.
2.4 Prise en considération des pollinisateurs sauvages dans ces systèmes agricoles en région tropicales
Toutes les productions végétales ne dépendent pas exclusivement de l’abeille mellifère.Bien
que l’efficacité de l’abeille domestique ait été démontrée à plusieurs reprises, des études plus
récentes ont mise en évidence l’importante contribution des pollinisateurs sauvages à la
production agricole (Bosh & Blas, 1993 ; Brittain et al, 2014 ; Corbet et al, 1992 ; Fohouo et
al, 2002 ;Klein et al, 2003;Kremen et al, 2002 ; Pérez-Balam et al, 2012 ; Winfree, 2007). Ces
derniers peuvent assurer un véritable soutient aux abeilles domestiques voire atteindre une
efficacité équivalente, si une diversité et une abondance suffisante sont maintenues (Brittain et
al, 2014 ; Klein, 2003 ; Winfree, 2007). Les pollinisateurs sauvages sont très diversifiés et
présentent des traits fonctionnels variés. Cette diversité fonctionnelle agit positivement sur
l’efficacité de la pollinisation et peut être utilisée (Fontaine et al, 2006 ; Hoehn et al, 2008).
La diversité et l’abondance des pollinisateurs ont un effet positif sur les rendements et
deviennent alors des facteurs de production (Kremen, 2002 ;Vaissière, 2005). Les abeilles
sauvages et les syrphes par exemple participent de façon significative à la fructification des
cultures indépendamment de l’abondance de l’abeille mellifère. Winfree (2007) a montré que
la pollinisation des fleurs de pastèques par les abeilles sauvages est suffisante pour assurer
une bonne production. En milieu tropical, la dépendance de certaines cultures à la
pollinisation par des insectes sauvages est forte. Roubick (1995) estime que 70 % des 1330

5
cultures tropicales sont dépendantes de la pollinisation par les insectes sauvages. En effet,
avant que l’abeille domestique ne soit importée lors de la colonisation, de nombreuses
cultures tropicales dépendaient des pollinisateurs sauvages qui jouaient donc un rôle central
dans la production agricole.La culture de café, par exemple, dépend de la diversité des
abeilles sauvages(Klein et al, 2003 ; Ricketts, 2004). Les fleurs de manguiers sont butinée par
l’abeille domestique, cependant, ce sont les syrphes et les abeilles sauvages qui en assurent la
pollinisation. Les fleurs d’agrumes sontaussi pollinisées par une importante diversité de
pollinisateurs sauvages (Louveaux & Pesson, 1995).
Dans les Antilles, aucune référence en milieu agricole ne témoigne de la participation des
pollinisateurs domestiques et sauvages à la pollinisation des plantes cultivées. Une étude
portant sur la diversité des abeilles sauvages et sur les fleurs butinées en Guadeloupe,permet
d’estimer la diversité des abeilles sauvages pouvant être observée dans les îles des Petites
Antilles (Meurgey, 2014). Cette étude a aussi mis en évidence des interactions entre les
espèces de fleurs et les pollinisateurs eta aussi été menée en Martinique (Meurgey &
Dumbardon, sous presse).
2.5 Les pollinisateurs sauvages sont étroitement liés aux conditions du milieu
Si les pollinisateurs sauvages et domestiques doivent être pris en considération dans la mise
en place du système agroécologique, les facteurs qui conditionnent leur présence doivent être
inclus dans l’étude. Pour se maintenir, les abeilles ont besoins de trois ressources différentes :
du pollen pour l’apport de protéines, du nectar pour les sucres et les acides aminés, et des sites
de nidification pour la reproduction et l’hivernage (Rollin, 2013). Des études ont montré la
relation étroite existante entrel’abondance et la diversité de la ressource florale présente dans
ou / à proximité des exploitations et l’abondance et la diversité des pollinisateurs sauvages en
milieu agricole (Ghazoul, 2006 ;Haenke, 2009 ; Hine & Hendrix, 2005 ; Holzchuh, 2007 ;
Kwaiser & Hendrix, 2007) Les prairies permanentes et la flore dans les parcelles cultivées
favorisent la présence des pollinisateursen milieu agricole.La proximité des zones naturelles
ou des éléments-semi naturels tels que les haies, les bosquets,les bordures de parcelles, a un
effet positif sur les communautés de pollinisateurs de part les sites de nidification et les
ressources florales qu’elles peuvent offrir aux pollinisateurs (Heidseck & Allier, 2013). La
distance aux éléments-semi naturels a un effet positif sur la diversité des pollinisateurs mais
aussi sur les rendements (Klein, 2008 ; Rickett, 2008) L’hétérogénéité et la complexité des
paysages favorisent la présence de pollinisateurs sauvages dans le milieu en comparaison aux
milieux simples et homogènes (Holzchuh, 2007, Tscharntke, 2005, Steffan-Deweneter, 2002).
Les pratiques culturales ont aussi un effet. Par exemple, certaines pratiques culturales telles

6
l’utilisation de produits phytosanitaires ont un effet négatif sur la ressource florale au sein de
la mosaïque agricole etagissent par conséquent négativement sur les communautés de
pollinisateurs (Holzschuh, 2010).
2.6 La FREDON un syndicat professionnel qui promotionne les principes agro-écologiques.
La FREDON (Fédération Régionale de Défense contre les Organismes Nuisibles), syndicat
professionnel agricole créé en 1958 dont les principales actions sont inscrites dans le code
rural aspire à développer et à mettre en application, dans le cadre de la protection des cultures,
des méthodes qui s’inscrivent dans une démarche agroécologique. En plus de ses actions
relatives à la lutte collective contre les rongeurs, au diagnostic sanitaire des végétaux, à la
surveillance du territoire (http://www.fredon972.fr), elle a plusieurs projets s’inscrivant
pleinement dans l’agroécologie. Tel est le cas de l’action PBI (Protection Biologique
Intégrée) qui a pour but de mettre en place des moyens de lutte intégrée en arboriculture
fruitière et en culture maraîchère. La FREDON assure un accompagnement technique des
agriculteurs afin de maintenir et valoriser la biodiversité des milieux agricoles martiniquais..
L’action PBI consiste alors aussi à décrire et inventorier la biodiversité dans les exploitations
agricoles et à identifier les potentielles espèces d’intérêt agronomique pour leur valorisation
dans la lutte biologique inondative et de conservation.
Depuis 2014 la FREDON développe une nouvelle action Agriculture Apiculture
Environnement dont la finalité est l’acquisition de références pour le développement d’un
système de culture agroécologique, associant productions végétales et activité apicole.
L’objectif de ce projet est de favoriser l’action pollinisatrice des abeilles domestiques tout en
conservant la diversité des pollinisateurs sauvages. Les arboriculteurs sont à cette occasion
sensibilisés à l’action pollinisatrice des pollinisateurs sauvages et domestiques et des relevés
sont réalisés dans leurs exploitations. Cette action est divisée en deux axes. Le premier
consiste à étudier les pollinisateurs des fleurs de goyaviers et de mesurer leur contribution à la
production de goyaves. Le deuxième axe consiste à identifier les pollinisateurs présents dans
les vergers et de d’identifier les facteurs qui expliquent leur présence ou non dans ces milieux
cultivés. C’est sur ce deuxième axe que se concentre ce stage.

7
2.6 Les missions du stage Ce stage consiste à répondre à la problématique suivante :
Dans le contexte insulaire tropical, quels sont les leviers d’action à prendre en compte pour l’établissement d’un système agroécologique associant verger et apiculture qui ne soit pas défavorable aux pollinisateurs?
Pour répondre à cette problématique, j’ai eu pour mission de réaliser un inventaire des
pollinisateurs présents dans les couverts des vergers et de caractériser leur communauté
(abondance, diversité des espèces, période d’activité). Dans un second temps, j’ai procédé à
des relevés floristiques des adventices afin de mettre en relation les communautés de
pollinisateurs et la ressource florale des couverts. L’étude des pollinisateurs des fleurs des
plantes cultivées (notamment les fleurs de goyaviers) fait partie d’une étude menée en
parallèle par la FREDON et ne fait pas parti des missions du stage. Cependant, du fait de la
complémentarité des deux études, certaines observations peuvent aider à l’interprétation des
résultats. Une description précise des interactions entre les fleurs adventices et les
pollinisateurs devra me permettre de mettre en évidence desfleurs du couvert d’intérêt qui
pourront être utilisées dans des aménagements agro-écologique pour favoriser les abeilles et
les syrphes dans les vergers. Une première approche des effets des infrastructures agro-
écologique au sein des exploitations sera réalisée pour apporter des premières informations
sur l’effet des éléments semi-naturels sur les communautés de pollinisateurs. Cette partie du
stage ne sera pas très approfondie et devra faire l’objet d’une étude à part entière
ultérieurement. L’étude est réalisée en milieu non contrôlé chez des arboriculteurs volontaires.
Le stage comprend alors un travail de démarchage auprès des agriculteurs et l’organisation
d’entretiens au cours duquel ils sont interrogés sur leurs techniques de gestion de
l’enherbement et sur les traitements utilisés sur les parcelles visitées. Un rendu des résultats
aux agriculteurs est ensuite effectué.

8
Hypothèses :
� H1 : Il y a un effet positif de la ressource floral (recouvrement, nombre
d’inflorescences, diversité des adventices, diversité des plantes en fleur) dans les
vergers sur l’abondance et la diversité de pollinisateurs.
� H2 : Les différents pollinisateurs utilisent une diversité variable de ressources florale
dans les couverts et il existe des préférences.
� H3 : Il y a un effet positif de l’environnement proche (éléments semi-naturels à
proximité des parcelles étudiées tels que les haies et les bordures de parcelle)sur les
communautés de pollinisateurs. La bordure de parcelle est attractive pour les
pollinisateurs et a un effet positif sur l’abondance et la diversité des pollinisateurs
� H4 : Il y a un effet positif ou négatif des techniquesde désherbage sur les adventices
des vergers et un effet indirect sur les communautés de pollinisateurs.
Figure 1 : Schéma récapitulatif des différentes hypothèses posées

9
2. Matériels et Méthodes 2.1 Le site d’étude
2.1.1. Eléments de géographie
L’étude est réalisée en Martinique. Elle est un des départements français d’Outre-Mer situé
dans l’archipel des Petites Antilles. Ces dernières séparent la mer des Antilles (communément
appelée mer des Caraïbes) de l’océan Atlantique. La Martinique est une île volcanique unique
de 1100 km2 avec seulement quelques dépendances. Le relief dans la moitié nord y est très
accidenté en raison de la montagne Pelée (1397m) toujours en activité, isolée à l’extrémité
nord et du Piton du Carbet (1196m). L’unique plaine du Lamentin recouvre le centre de l’île
et une alternance de vallées et de collines (Mornes) caractérise la moitié sud de la Martinique
(Sastre & Breuil, 2007).
2.1.2. Particularités climatiques
La Martinique bénéficie d’un climat tropical maritime et d’un fort ensoleillement. Les
températures montrent de faibles amplitudes saisonnières et peu de variations à l’échelle de
l’île (DIREN, 2008). Il est cependant possible de distinguer deux saisons sur une année. La
saison sèche ou carême débute en février et se termine en avril. C’est une période de
sècheresse durant laquelle peuvent se succéder plusieurs semaines sans aucun épisode
pluvieux. La saison humide ou hivernage s’étend de mai-juin à novembre et des perturbations
climatiques importantes peuvent avoir lieu avec des risques cycloniques en début de saison
(DIREN, 2008 ; Sastre & Breuil, 2007). Les températures minimales (17 à 25 °C) sont
ressenties pendant l’hivernage dans les reliefs du nord de l’île tandis que les températures
maximales (28 à 32 °C) sont ressenties pendant le carême le long des côtes. C’est à la fin du
carême et au début de l’hivernage qu’il est possible d’observer le maximum d’espèces
végétales en floraison (DIREN, 2008 ; Sastre & Breuil, 2007)et qu’il est alors stratégique
d’étudier les pollinisateurs.
2.1.3. Choix et localisation des parcelles étudiées
L’étude est réalisée sur l’ensemble du territoire dans les localités propices à l’arboriculture
fruitière. Tous les agriculteurs sélectionnés ont déjà eu recours aux services de la FREDON et
ont participé à diverses études. Certains arboriculteurs ont été sélectionnés dans la base de
données de l’Association des Vergers et Jardins Tropicaux Organisation de producteurs de
Martiniquequi travaille en collaboration avec la FREDON.
Les relevés ont lieu dans des parcelles de goyaviers et d’agrumes. En effet, ce sont les
cultures fruitières les plus importantes sur l’île et représentent 90 % de la surface fruitière
10
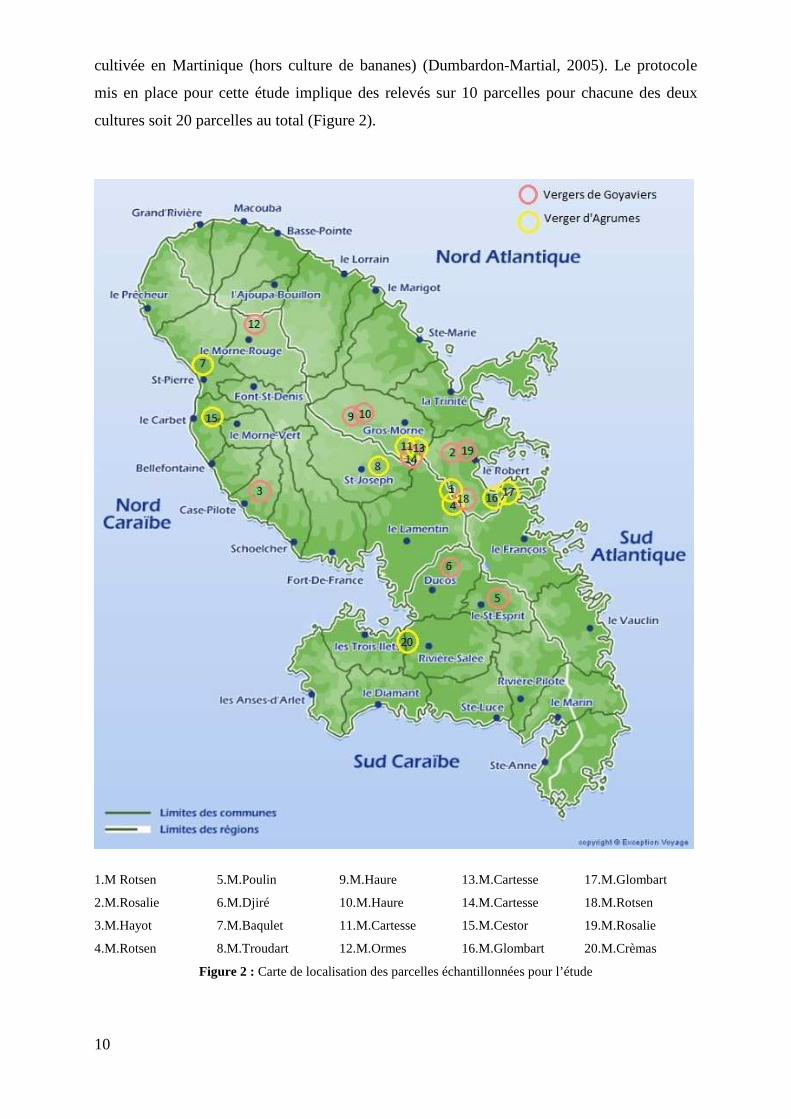
cultivée en Martinique (hors culture de bananes) (Dumbardon-Martial, 2005). Le protocole
mis en place pour cette étude implique des relevés sur 10 parcelles pour chacune des deux
cultures soit 20 parcelles au total (Figure 2).
1.M Rotsen 5.M.Poulin 9.M.Haure 13.M.Cartesse 17.M.Glombart
2.M.Rosalie 6.M.Djiré 10.M.Haure 14.M.Cartesse 18.M.Rotsen
3.M.Hayot 7.M.Baqulet 11.M.Cartesse 15.M.Cestor 19.M.Rosalie
4.M.Rotsen 8.M.Troudart 12.M.Ormes 16.M.Glombart 20.M.Crèmas
Figure 2 : Carte de localisation des parcelles échantillonnées pour l’étude

11
2.2 Les cultures fruitières étudiées
2.2.1. Le Goyavier
La culture de goyave s’inscrit actuellement dans une stratégie de diversification fruitière
comme alternative aux productions actuellement en crise sur l’île. Appartenant à la famille
des Myrtacea, le goyavier est reconnaissable par sa taille (entre 2 et 8 mètres de hauteur), et
son tronc aux motifs atypiques du fait de la desquamation de fines plaques d’écorces (Le
Bellec & Lavigne, 2006). La fleur de goyavier se caractérise par des étamines externes à filets
très longs. Les pétales de la corolle blanche, bien écartés rendent les étamines bien visibles et
accessibles aux pollinisateurs. La fleur fécondée produit un fruit, une baie globuleuse dont la
taille varie selon les variétés (Le Bellec & Lavigne, 2006).
Crédit photo : Clara SINGHCrédit photo Emilie LAFONT
L’espèce Psidium guayava, est originaire d’Amérique tropicale. On la retrouve à la fois en
tant que plante cultivée dans les vergers et arbre sauvage très répandu sur l’île. La poussée
végétative du goyavier est régulière et l’arbre présente deux périodes de fructification (Le
Bellec & Lavigne, 2006). La première a lieu début février (30% de la production annuelle) et
la seconde en juillet (70% de la production annuelle). Psidium guayava présente une
importante diversité variétale (150 variétés dans le monde). La Martinique a procédé à une
sélection afin de ne conserver que celle d’une qualité supérieure propice à la transformation.
La production de goyave est étroitement liée aux possibilités de transformation sur place
(sorbets, nectars, jus, confitures…). Les variétés se distinguent en 3 groupes : fruit de bouche,
fruit d’usine, fruit mixte. Les variétés les plus représentées à la Martinique sont : Beaumont,
Centeno Profilic, Red hybride, et Red suprême Ruby.

12
2.2.2. Les agrumes
Les agrumes sont les principales cultures fruitières de Martinique et forment une production
tournée essentiellement vers le marché local. Les cultures d’agrumes occupent à elles seules
75% des surfaces fruitières cultivées. Le terme « agrume » désigne à la fois les espèces du
genre Citrus, Fortunella et Poncirus. Ces arbres fruitiers, originaires du Sud-est-Asiatique
appartiennent à la famille des Rutaceae et comptent de nombreuses espèces. Les espèces du
genre Citrus sont : l’Oranger : C.sinensis, le Mandarinier :C.reticulata, leLimetier :
C.aurantifolia et le Pomelos : C.paradisi. Au cours des années 80, quelques tentatives de
développement de la filière arboricole vers l’exportation ont été initiées. Cependant, les
problèmes de commercialisation et de compétitivité on freiné ces démarches.
2.3 Les insectes pollinisateurs : modèle biologique
2.3.1. Choix des pollinisateurs étudiés
Beaucoup d’insectes (diptères, les lépidoptères hétérocères, les coléoptères, les hyménoptères
ou encore les lépidoptères) présentent une activité floricole (Fenster, 2004). Cependant,
l’activité floricole ne justifie pas l’action de pollinisation (Couplan et al, 2014, Vaissière,
2005). Seulement une portion restreinte des insectes floricoles contribue activement à la
reproduction sexuée des fleurs de plantes sauvages et cultivées. En Europe, plusieurs études
ont démontré que les pollinisateurs les plus efficaces sont des hyménoptères appartenant à la
super-famille des Apoïdes Anthophiles (nom scientifique : Apoidae) c'est-à-dire les abeilles
domestiques et sauvageset cela a été confirmé à l’échelle mondiale(Klein et al, 2007 ; Le
Feon, 2010, Roubick et al, 1995). Les syrphes (nom scientifique : Syrphidae) peuvent être
considérés comme importants pour la pollinisation des cultures bien que leur efficacité soit
moindre que celle des abeilles (Le Féon, 2010). A partir de ces informations et dans le cadre
de cette étude, le choix des pollinisateurs étudiés se restreint donc aux Apoïdes (abeilles
sauvages et domestiques) et aux syrphes.
2.3.2. Les Apoïdes
� Généralités
Les Apoïdes Anthophiles comprennent neuf familles dont celle des Apidae qui regroupe les
pollinisateurs les plus connus, notamment l’abeille domestique. Les 8 autres familles sont : les
Andrenidae, les Halictidae, les Colletidae, les Mellitidae, les Megachilidae et les
Anthophoridae (Terron, 1980). A l’échelle mondiale, environ 25 000 espèces seraient
recensées (Millenium Ecosystem Assesment). Toutes les familles ne sont pas toujours toutes
représentées sur un territoire donné ; par exemple, 7 familles et 865 espèces sont recensées en
France (Rasmont et al, 1995). En Martinique et plus largement dans les petites Antilles, peu

13
d’études ont été réalisées sur les insectes pollinisateurs et la diversité des Apoïdes sur l’île est
encore très peu connue. Un recensement récent permet de dire qu’il y aurait 14 espèces
d’abeilles (Meugrey & Dumbardon-Martial, 2015 sous presse).
� Abeilles domestiques
L’abeille domestique Apis melliferaconstitue le pollinisateur le plus performant du fait de sa
morphologie et de la taille de ses colonies. Elle possède des traits morphologiques
fonctionnels spécifiques à la récolte du pollen (O’Tool& Raw,1999). Sa pilosité lui permet de
retenir le pollen lorsqu’elle butine les fleurs etle transfère à l’aide du « peigne » sur des
organes spécifiques localisés au niveau de ses pattes postérieures. Elle fabrique ensuitedes
pelotes de pollen qu’elle stocke dans les « corbeilles à pollen » (Couplan et al, 2014, O’Tool&
Raw, 1999).De plus, L’abeille domestique se caractérise par une activité très élevée en
comparaison à d’autres insectes floricoles (Bourgeois, 2006). Elle se distingue des autres
insectes pollinisateurs parson comportement plutôt généraliste (polylectiques)qui lui permet
d’assurer la pérennité d’une importante diversité d’espèces végétales (Fontaine, 2006).
� Abeilles sauvages
Les abeilles sauvages sont le plus souvent des abeilles solitaires. Les femelles édifient toutes
leur propre nid et le type de nidification varie d’une famille à l’autre (Villemant, 2005). La
majorité des abeilles sauvages creusent leurs nids dans le sol et sont dites terricoles. D’autres
édifient leur nid dans du bois creux ou réutilisent des galeries dans du bois mort et sont
qualifiées d’abeilles xylicoles. Les autres, dites à nid libre fabriquent leur nids avec de la
résine ou de la boue mélangée à des cailloux (Villemant, 2005). Les abeilles sauvages doivent
pouvoir trouver la ressource floricole (pollen et/ou nectar), nécessaire à leurs besoins nutritifs
et à ceux des larves, et ce, à proximité de leur nid. Le régime alimentaire des abeilles
sauvages est variable et plus ou moins spécifique selon les familles ou les espèces.
2.3.3. Les syrphes
Les syrphes correspondent à une famille de diptères. Plus de 6000espèces sont recensées en
dans le monde (Sommagio, 1999) et 500 ont été identifiées en France (Sarthou & Speight,
2005). Les adultes mesurent entre 4 et 25 mm et sont reconnaissables à leur vol stationnaire et
leurs mouvements latéraux très rapides (Sommagio, 1999). Les syrphes constituent un
auxiliaire des cultures de double intérêt pour les agriculteurs. En effet, les larves à action de
prédation peuvent assurer la régulation de certains ravageurs notamment les puceronsLa
majorité des larves sont entomophages, d’autres sont macrophages ou phytophage. De plus,
sauf exception, tous les syrphes adultes sont floricoles et récoltent le nectar ou le pollen

14
nécessaire à leurs besoins et peuvent participer à la pollinisation. Les syrphes sont capables
d’occuper une importante diversité d’habitats. En effet, il est possible d’en retrouver en milieu
ouvert comme en milieu fermé (Sarthou & Speight, 2005).
2.4 Le protocole de terrain
2.4.1. Description du dispositif
Afin de pouvoir estimer la relation entre les communautés de pollinisateurs et les cortèges
floristiques, le dispositif est mis en place sur la base de plusieurs transects (Le Feon,
2010 ;Mandelik et Roll, 2009). Sur chacune des parcelles sont définies 3 transects de 30mde
longueur (Bartomeus et al, 2010) (Figure 3& 4). Un premier transect est positionné en
bordure de champ, un deuxième est localisé entre le 2ème et le 3ème rang et le troisième est mis
en place entre le 4ème et le 5ème rang.
2.4.2. Capture des pollinisateurs
Au filet entomologique
1. Le fauchage
Les captures sont réalisées au filet entomologique le long des trois transects mis en place dans
le champ (Figure 3). Sur chaque transect, des relevés non sélectifs de type fauchage sont
effectués au filet (« chasse à l’aveugle ») afin de prélever divers arthropodes dont les
pollinisateurs et d’apporter des données d’abondance et de diversité. Le récolteur avance d’un
pas lent le long du transect et effectue des mouvements rapides latéraux de va-et-vient avec le
filet à hauteur de la strate herbacée. Ces relevés sont réalisés à 8h puis sont répétés à 11h puis
à 15h afin que les relevés soient représentatifs de la diversité de l’entomofaune car tous les
pollinisateurs n’ont pas une activité synchrone au cours d’une journée afin d’éviter la
concurrence pour l’accès à la ressource (Fouhouo, 2002). De plus, de savoir s’il existe une
heure préférentielle d’activité floricole et d’ajuster le protocole pour les futures études sur les
pollinisateurs à la FREDON.

15
Figure 3 : Dispositif de terrain n°1. Le long des transects rouges sont réalisés les relevés de type fauchages ( à 8h00, 11h00 et 15h00). Le long des transects bleus est effectuée la chasse à vue de 8h30 à 10h (30 min/transect). Les relevés floristiques sont faits sur la zone d’échantillonnage en rouge sur une largeur d’un mètre).
Les insectes collectés sont transférés dans des bocaux contenant du papier absorbant imbibé
d’alcool à 90°. Une fois les insectes collectés immobiles, le contenu des bocaux est transvasé
dans un petit pilulier. Les bocaux peuvent alors être réutilisés à 11h et à 15h. Les échantillons
sont ensuite transportés dans ces piluliers jusqu’au laboratoire, sont vidés de leur alcool pour
être placés au congélateur.
2. La chasse à vue
Le long des trois transects (figure 3), les pollinisateurs sont capturés de façon sélective en
activité de butinage (Payette, 2013) pendant 30 minutes sur chaque transect (Bartomeus,
2010). Pour chaque capture, le pollinisateur et la plante butinée sont identifiés. La chasse à
vue a pour objectif de décrire les interactions entre les pollinisateurs et les fleurs butinées et
de regarder s’il existe des préférences selon les espèces.Afin de caractériser la diversité des
fleurs butinées par chaque espèce de pollinisateurs, l’indice Trophic Niche Breadth est calculé
(Wolfgang, 1996).
Par piégeage « bols colorés »
Le long de ces mêmes transects sont installés linéairement des pièges colorés (bols) (Figure
4). L’utilisation de cette technique constitue un premier essai de piégeage des pollinisateurs
en Martinique.Si les pièges colorés sont souvent utilisés pour évaluer la diversité des

16
pollinisateurs dans de nombreuses études, ils n’ont jamais été utilisé à notre connaissance en
Martinique, ni sur une île océanique dans les tropiques (Le Féon, 2010 ; Kwaiser & Hendrix,
2007 ; Mandelik & Roll, 2009). A cette occasion, il s’agira de vérifier que le fauchage
n’entraîne pas une sous-estimation de la biodiversité auquel cas la capture par pièges
apporterait des données complémentaires à l’étude (Payette, 2013). L’utilisation de pièges
permet également de compléter les relevés par une technique passive dont la qualité ne
dépend pas du récolteur. Pour se faire sur chaque transects, 3 bols colorés sont alignés et
séparés les uns des autres par une distance de 15m. Il ya a alors en tout 9 pièges par parcelle.
Les bols utilisés sont jaunes, bleus et blancs afin d’attirer une diversité maximale de
pollinisateurs. En effet, tous les pollinisateurs ne sont pas attirés par les mêmes couleurs et il
existe des préférences (Leong & Thorp, 1999). Il y a un bol de chaque couleur par transect
alignés dans un ordre précis, et, cet ordre est conservé sur tous les transects des parcelles
étudiées.
Figure 4 : Dispositif de terrain sur lequel seront effectués les relevés entomologiques par piégeage à l’aide des
bols colorés. Les ronds représentent chacun des pièges.
Ces bols contiennent de l’eau savonneuse dans laquelle les pollinisateurs glissent et se noient.
Chaque piège est placé à hauteur de la strate herbacée. Les bols posés sur les parcelles au jour
J sont retirés au jour J+3. Lors de la récupération des échantillons à J+3, les pièges sont vidés

17
et les insectes sont récupérés dans une passoire puis transférés dans des piluliers remplis
d’alcool à 90°. Les échantillons sont ensuite transportés ainsi jusqu’au laboratoire où ils
seront traités.
2.4.3 Les relevés floristiques
Les transects utilisés pour les relevés floristiques sont ceux sur lesquels sont réalisés les
relevés entomologiques par fauchage (figure 1). Sur une largeur de 0,5m de chaque côté du
transect, des relevés qualitatifs des plantes en fleurs et non en fleurs sont effectués afin
d’obtenir la richesse spécifique des adventices. Une liste d’espèces est alors attribuée à
chaque transect et informe sur la composition floristique. Les espèces adventices sont
identifiées à l’aide des planches photographiques du guide Adventille Guadeloupe et
Martinique et des clefs de la flore de phanérogames de Guadeloupe et de Martinique (Fournet,
2012). Pour chaque espèce en fleurs recensée à l’exception des Poacées (Bartomeus, 2010), le
nombre d’inflorescences ou d’unités florales est compté. L’unité florale est définie dans
l’étude comme une inflorescence accessible aux pollinisateurs par le vol. On compte deux
unités florales si le pollinisateur doit voler pour passer de l’une à l’autre (Bartomeus, 2010 ;
Dick, Corbet & Pywell, 2002). Une somme de toutes les inflorescences (toutes espèces
confondues) est aussi calculée. A chaque espèce en fleurs est aussi attribuée une valeur de
recouvrement qui repose sur une estimation visuelle (Steffan-Dewenter, 2001). En effet, la
taille de la corolle pouvant être très variable selon les espèces, la relation pollinisateurs-
recouvrement floral peut différer de la relation pollinisateurs-nombre d’inflorescences. Une
somme des recouvrements, toutes espèces confondue, est également calculée pour chaque
transect.
La sélection des parcelles est faite de sorte que les relevés soient réalisés sur des parcelles
variées du point de vue du couvert végétal. Le dispositif est alors mis en place sur un nombre
équivalent de parcelles fleuries, moyennement fleuries et peu ou pas fleuries.
2.4.4Estimation des éléments constitutifs de l’environnement proche
Sur chaque parcelle est réalisée une description de l’environnement proche. En effet, une
étude paysagère et une estimation des surfaces des éléments semi-naturels sur le terrain est
difficilement réalisable compte-tenudes moyens dont dispose la FREDON, du temps
nécessaire à cartographier chaque élément du paysage en raison de la complexité et de
l’hétérogénéité de la mosaïque des vergers caractéristiques des exploitations en milieu
tropical. Le périmètre de chaque parcelle est alors mesuré. Tous les éléments constitutifs de ce
périmètre sont mesurés. Les rapports entre la longueur de chacun des éléments sur la longueur
totale du périmètre de la parcelle sont calculés. Ces rapports expriment la proportion de

18
chaque élément de l’environnement proche de la parcelle étudiée. Les éléments de
l’environnement proche observés sur le terrain sont : les haies (regroupant les haies et les
haies vives), les bordures et les parcelles adjacentes. D’autres éléments tels que les cours
d’eau et les lisières de bosquets ont été mesurés mais sont trop peu représentés pour les
inclure dans les analyses de données. Du fait de la disponibilité des parcelles, la sélection des
différentes parcelles de l’étude n’ont pas pu être choisies en fonction de la proportion des
éléments constitutifs du périmètre. Il s’agit donc d’une première approche avec les données
disponibles obtenues sur les parcelles échantillonnées.
2.5 Le protocole de laboratoire Les pollinisateurs servant de modèle pour l’étude (abeilles domestiques, syrphes et abeilles
sauvages) sont identifiés au rang de l’espèce et mis en collection. Les Anthophiles sont
identifiés à l’aide d’une boîte de référence dont les spécimens ont été identifiés par François
Meurgey de la société de l’Herminier. Alain Pauly de l’Institut royal des Sciences Naturelles
de Belgique, a permis l’identification des abeilles sauvages appartenant à la famille des
Halictidae, et nous a transmis les critères d’identification pour que nous puissions les
identifier directement au laboratoire. Il a cependant continué de vérifier les identifications au
cours de l’étude. Le reste des insectes collectés sont identifiés au rang de la famille par Eddy
Dumbardon-Martial et Chloé Pierre (FREDON).
2.6 Enquête sur les pratiques de désherbage Un court questionnaire est établi et complété lors d’un entretien avec l’agriculteur. Ce dernier
est ainsi interrogé sur ses techniques et les pratiques qu’il emploie sur la-parcelle étudiée :
- Quelles technique de désherbage utilise-t-il (mécanique, chimique, manuelle,
autres…) sur une année ?
- S’il utilise plusieurs techniques, les utilise-t-il de façon équivalente ou utilise-t-il
préférentiellement une technique par rapport aux autres ?
- A quelle fréquence désherbe-t-il ?
- Quels traitements ont été effectués sur la parcelle étudiée pendant les six mois
précédents les relevés ?
- La parcelle étudiée est-elle irriguée ?
- Intervient-il dans la gestion des éléments de l’environnement proche (haies, lisières…)
- Sait-il s’il y a des ruches à proximité de l’exploitation ?
2.7 Analyses statistiques L’échelle à laquelle sont réalisées les analyses de données est variable selon les hypothèses
testées. Ce sont exclusivement les données de fauchage et de chasse à vue qui servent au

19
traitement de données. En effet, tous les individus capturés à l’aide des « bols colorés » ne
sont pas tous identifiésen raison de la grande quantité d’individus piégés et des temps
d’identification.
Les valeurs de diversité des pollinisateurs utilisées pour toutes les analyses statistiques de
cette étude résultent d’un calcul d’indice de Shanon:
H’= -∑ pilnpi
(i=une espèce de pollinisateur, pi = proportion du pollinisateurs i parmi tous les pollinisateurs
capturés). Cet indice inclue tous les pollinisateurs capturés (abeilles domestiques, sauvages et
syrphes).
2.7.1. Estimation de l’effort d’échantillonnage avec des courbes d’accumulation
Pour le fauchage et la chasse à vue, des courbes d’accumulation représentant la richesse
spécifique cumulée des pollinisateurs capturés en fonction du nombre de sites échantillonnées
ont été tracées. Les courbes d’accumulation témoignent de la qualité de l’effort
d’échantillonnage et permettent de savoir si l’ensemble de la diversité des pollinisateurs des
milieux échantillonnés a été capturée. Pour le fauchage, la courbe est réalisée à l’échelle du
transect (1 site = 1 transect et il y a 60 sites au total). Pour la chasse à vue, l’échelle est celle
de la parcelle (1 site =1 parcelle =20 parcelles et il y a 20 sites au total).
2.7.2. Etude de l’abondance et de la diversité des pollinisateurs selon les sites
Pour cette analyse,le transect est utilisé comme unité d’étude et ce sont les pollinisateurs
capturés au fauchage sur les trois transects (toutes heures et tous transects confondus) des 20
parcelles qui sont utilisés comme données. Les abondances de pollinisateurs ne suivent pas
une distribution normale. Une comparaison des moyennes de plusieurs échantillons via une
ANOVA n’est pas possible car les conditions d’application nécessaires ne sont pas réunies.
Un test alternatif non paramétrique de Kruskal-Wallis est utilisé. Il s’agit d’un test de
comparaison des médianes de plusieurs échantillons. Il y a donc 20 échantillons avec 3
répétitions par échantillon (1 transect = 1 répétition). Le même test est utilisé pour comparer
la diversité des pollinisateurs selon les parcelles échantillonnées. En effet, les indices de
diversité de pollinisateurs ne suivent pas une loi normale. Une ANOVA ne peut pas être
utilisée.

20
2.7.3. Analyse de l’effet de l’heure de capture sur l’abondance et la diversité des pollinisateurs
Les pollinisateurs capturés au fauchage à 8h00, 11h00 et 15h00 sur chacun des 3 transects
(tous transects confondus) sont utilisés pourcette analyse. L’unité d’étude pour ce test est
donc le transect. Les données d’abondance et de diversité des pollinisateurs ne permettent pas
la réalisation d’une ANOVA. Un test de Kruskal-Wallis de comparaison des abondancesde
trois échantillons (8h00, 11h00 et 15h00) est utilisé. Ce même test est utilisé afin de comparer
la diversité des pollinisateurs entre les différentes heures de capture.
2.7.4Effet des adventices sur l’abondance et la diversité des pollinisateurs
Plusieurs variables sont utilisées pour tester l’effet des adventices sur les communautés de pollinisateurs :
- Richesse spécifique des adventices totale (en fleurs et non en fleurs)
- Recouvrement des plantes en fleurs : « recouvrement florale »
-Nombre d’inflorescences
- Diversité des plantes en fleurs calculée à partir d’un indice de Shanon : H’= -∑ pilnpi (i=espèce végétale en fleurs, pi=nombre d’inflorescence de l’espèce 1 par rapport aux autres espèces en fleurs)
Dans un premier temps, les données utilisées pour cette analyse sont les pollinisateurs
capturés au fauchage. L’analyse se fait donc à l’échelle du transect. Afin d’étudier une
potentielle relation entre les variables explicatives ci-dessus et les communautés de
pollinisateurs, un Modèle linéaire Généralisé est utilisé pour chacune des variables pour
expliquer l’abondance des pollinisateurs. Ces modèles suivent une loi de Poisson. Ces mêmes
modèles sont réutilisés pour regarder l’effet sur la diversité des pollinisateurs. Dans ce dernier
cas, ces Modèles Linéaires Généralisés suivent une loi de Gauss.
Dans un second temps, ces modèles linéaires généralisés sont reproduis à l’échelle de la
parcelle avec les données de capture à vue. Les données floristiques utilisées sont alors issue
de calculs de moyennes des relevés floristiques des 3 transects de chaque parcelle.
2.7.5. Etudes des interactions plantes pollinisateurs
Calcul de la diversité des fleurs du couvert butinées par chaque espèce de pollinisateur
L’indice Trophic Niche Breadth NB (Wolfgang, 1996) est calculé à partir des données
obtenues lors de la chasse à vue. Il s’agit d’un calcul d’indice de Shanon : H’= -∑ pilnpi
(i=une espèce de pollinisateurs, pi = proportion de fleurs butinées par le pollinisateur i par
rapport à toutes les autres fleurs).

21
Plus l’indice NB est élevé, plus le pollinisateur utilise une ressource florale diversifiée. Un
indice par espèce de pollinisateurs (toutes parcelles confondues) est calculé.
Estimation du nombre d’espèce en fleurs nécessaire pour capturer l’ensemble des espèces
du milieu à vue.
Une courbe d’accumulation représentant la richesse spécifique cumulée des pollinisateurs
capturés à vue, en fonction de la richesse spécifique des plantes en fleurs sur lesquelles ont été
capturés les pollinisateurs, a été tracée. Cette courbe rassemble alors les données de chasse à
vue des 20 parcelles. Elle a pour objectif de regarder à partir de combien d’espèces en fleurs il
est possible de recenser l’ensemble des pollinisateurs capturés à vue dans cette étude.
2.7.6. Effet de l’hétérogénéité à l’échelle de la parcelle et effet de l’environnement proche sur les communautés de pollinisateurs.
Effet de l’hétérogénéité intra-parcellaire : effet transect
Afin de tester l’effet de la position du transect sur les communautés d’adventices, un test de
comparaison de 3 échantillons (1 échantillon par transect) est utilisé. Les conditions
d’application de l’ANOVA ne sont pas réunies et le test de Kruskal-Wallis est utilisé pour
comparer la richesse spécifique des adventices, le recouvrement floral et le nombre
d’inflorescences entre les 3 transects.
Un test de comparaison de trois échantillons de Kruskal-Wallis est choisi pour comparer
l’abondance des pollinisateurs entre les trois transects. Ce même test est réutilisé pour tester
l’effet de la position du transect sur la diversité des pollinisateurs.
Effet des éléments constitutifs du périmètre sur l’abondance et la diversité despollinisateurs
Afin d’étudier les effets des éléments constitutifs du périmètre des parcelles sur les
communautés de pollinisateurs, un Modèle Linéaire Généralisé additif incluant les
proportions de bordure, de haies et de parcelle adjacente a été utilisés. Ce modèle suivant une
loi de poisson a été réalisé pour regarder l’effet sur l’abondance des pollinisateurs. Ce même
modèle suivant une loi de Gauss a été utilisé pour étudier l’effet des différentes variables sur
la diversité.
2.7.7. Effet des pratiques
Aucune analyse statistique n’a pu être réalisée. En effet, les techniques de désherbages
utilisées ne sont pas assez variées pour effectuer des comparaisons. Il s’agit donc simplement
d’une analyse descriptive.
22
3. Résultats
Les analyses ont essentiellement été réalisées avec les données obtenues à l’aide du fauchage
et de la chasse à vue. Les pollinisateurs capturés à l’aide des bols colorés ne sont pas encore
tous identifiés au rang de l’espèce et seules les données d’abondances sont disponibles
3.1 Le matériel collecté
3.1.1 Représentativité des groupes collectés
Au total 3785 spécimens ont été collectés et sont répartis dans 96 Familles (Annexe x).
Certains ordres tels que les diptères (autres) sont bien représentés et présentent une abondance
notable (toutes familles confondues). Les taxons appartenant aux groupes étudiés (abeilles
domestiques, abeilles sauvages et syrphes) représentent 34% de l’ensemble des spécimens
collectés.
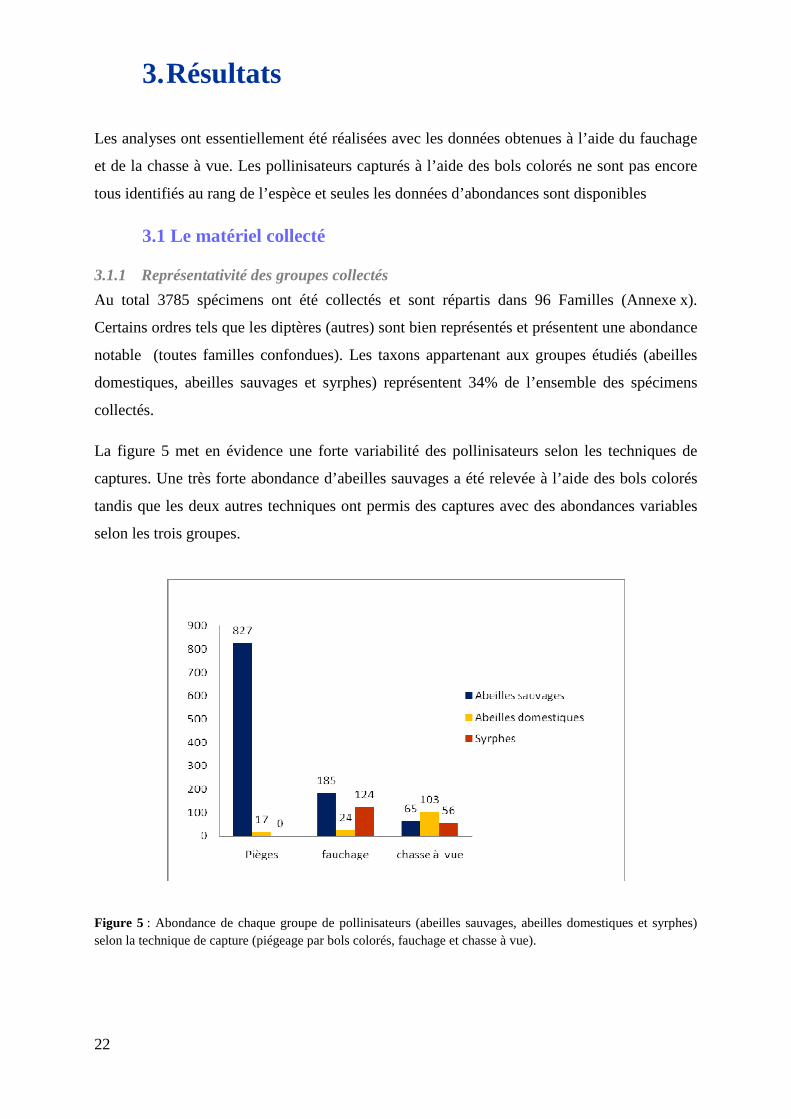
La figure 5 met en évidence une forte variabilité des pollinisateurs selon les techniques de
captures. Une très forte abondance d’abeilles sauvages a été relevée à l’aide des bols colorés
tandis que les deux autres techniques ont permis des captures avec des abondances variables
selon les trois groupes.
Figure 5 : Abondance de chaque groupe de pollinisateurs (abeilles sauvages, abeilles domestiques et syrphes) selon la technique de capture (piégeage par bols colorés, fauchage et chasse à vue).
23
0 10 20 30 40 50
05
1015
2025
Sites
Ric
hess
e sp
écifi
que
cum
ulée
5 10 15
05
1015
2025
Sites
Ric
hess
e sp
écifi
que
cum
ulée
3.1.2 Evaluation de l’effort d’échantillonnage
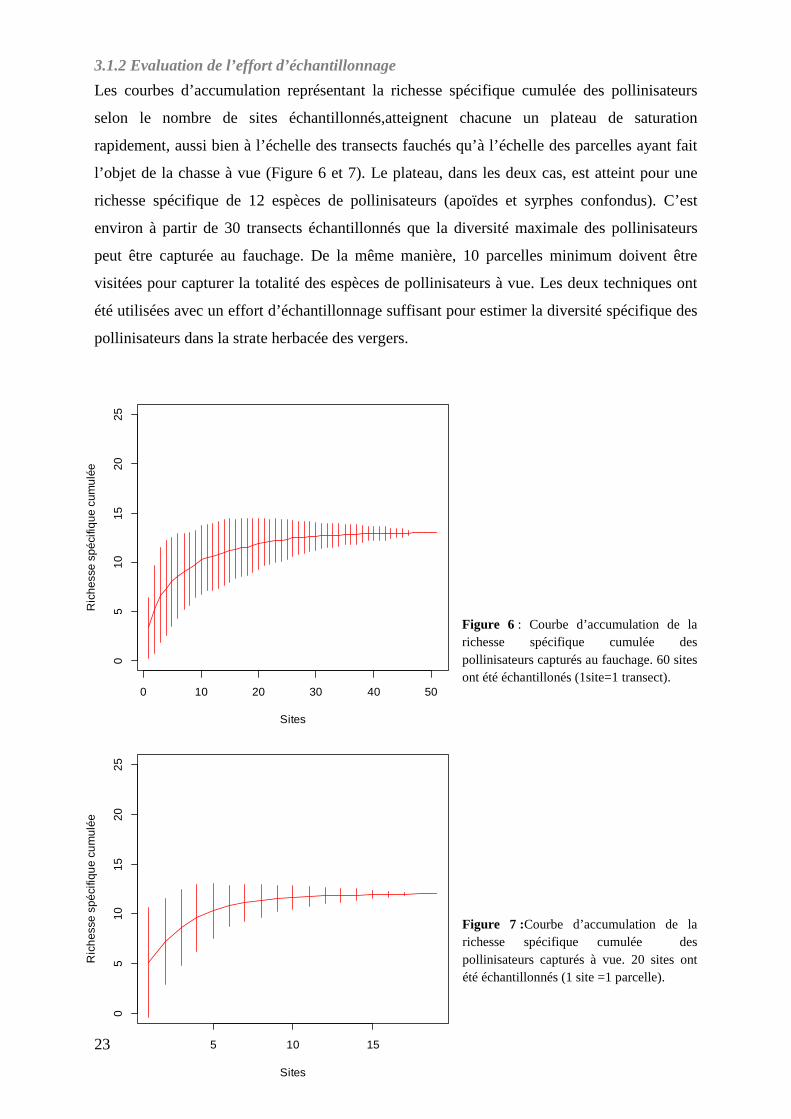
Les courbes d’accumulation représentant la richesse spécifique cumulée des pollinisateurs
selon le nombre de sites échantillonnés,atteignent chacune un plateau de saturation
rapidement, aussi bien à l’échelle des transects fauchés qu’à l’échelle des parcelles ayant fait
l’objet de la chasse à vue (Figure 6 et 7). Le plateau, dans les deux cas, est atteint pour une
richesse spécifique de 12 espèces de pollinisateurs (apoïdes et syrphes confondus). C’est
environ à partir de 30 transects échantillonnés que la diversité maximale des pollinisateurs
peut être capturée au fauchage. De la même manière, 10 parcelles minimum doivent être
visitées pour capturer la totalité des espèces de pollinisateurs à vue. Les deux techniques ont
été utilisées avec un effort d’échantillonnage suffisant pour estimer la diversité spécifique des
pollinisateurs dans la strate herbacée des vergers.
Figure 6 : Courbe d’accumulation de la richesse spécifique cumulée des pollinisateurs capturés au fauchage. 60 sites ont été échantillonés (1site=1 transect).
Figure 7 :Courbe d’accumulation de la richesse spécifique cumulée des pollinisateurs capturés à vue. 20 sites ont été échantillonnés (1 site =1 parcelle).
24
Tous les individus capturés par les pièges « bols colorés » n’ont pas tous pu être identifiés
dans les temps. La courbe d’accumulation des espèces capturées sera donc réalisée
ultérieurement à la FREDON.
3.2 Diversité et abondance des pollinisateurs
3.2.1 Les pollinisateurs recensés
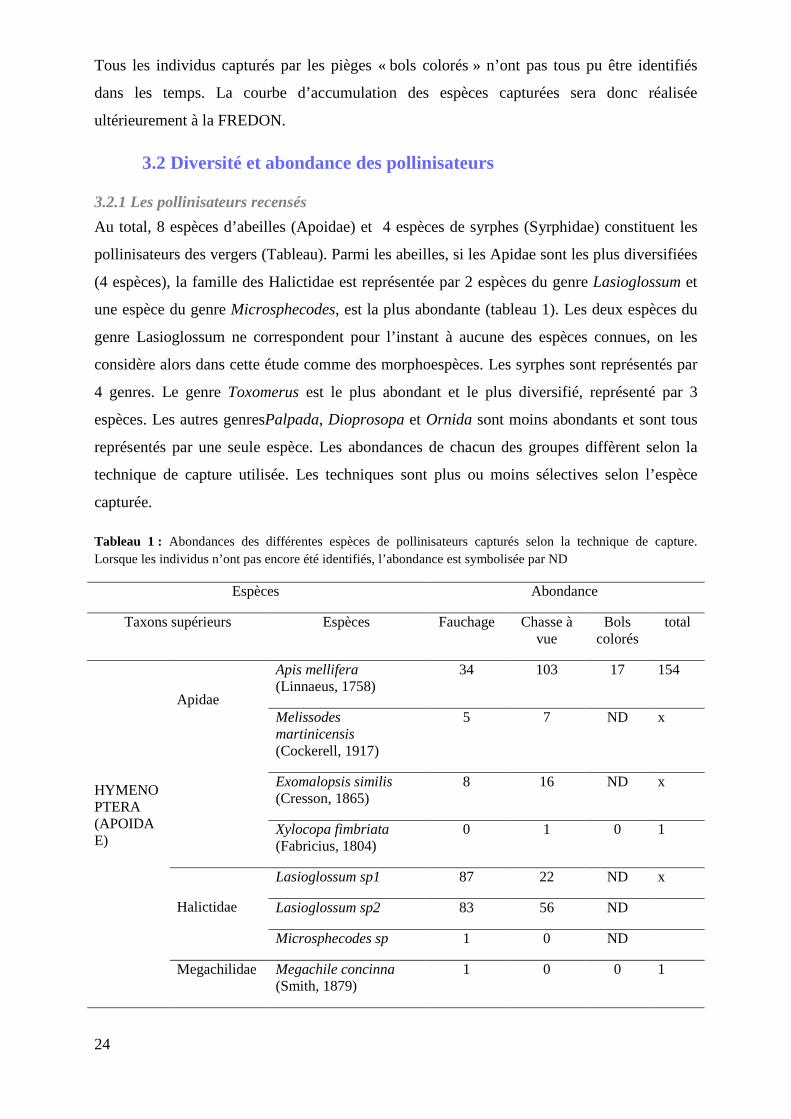
Au total, 8 espèces d’abeilles (Apoidae) et 4 espèces de syrphes (Syrphidae) constituent les
pollinisateurs des vergers (Tableau). Parmi les abeilles, si les Apidae sont les plus diversifiées
(4 espèces), la famille des Halictidae est représentée par 2 espèces du genre Lasioglossum et
une espèce du genre Microsphecodes, est la plus abondante (tableau 1). Les deux espèces du
genre Lasioglossum ne correspondent pour l’instant à aucune des espèces connues, on les
considère alors dans cette étude comme des morphoespèces. Les syrphes sont représentés par
4 genres. Le genre Toxomerus est le plus abondant et le plus diversifié, représenté par 3
espèces. Les autres genresPalpada, Dioprosopa et Ornida sont moins abondants et sont tous
représentés par une seule espèce. Les abondances de chacun des groupes diffèrent selon la
technique de capture utilisée. Les techniques sont plus ou moins sélectives selon l’espèce
capturée.
Tableau 1 : Abondances des différentes espèces de pollinisateurs capturés selon la technique de capture. Lorsque les individus n’ont pas encore été identifiés, l’abondance est symbolisée par ND
Espèces Abondance
Taxons supérieurs Espèces Fauchage Chasse à vue
Bols colorés
total
HYMENOPTERA (APOIDAE)
Apidae
Apis mellifera (Linnaeus, 1758)
34 103 17 154
Melissodes martinicensis (Cockerell, 1917)
5 7 ND x
Exomalopsis similis (Cresson, 1865)
8 16 ND x
Xylocopa fimbriata (Fabricius, 1804)
0 1 0 1
Halictidae
Lasioglossum sp1 87 22 ND x
Lasioglossum sp2 83 56 ND
Microsphecodes sp 1 0 ND
Megachilidae Megachile concinna (Smith, 1879)
1 0 0 1

25
Quelques pollinisateurs recensés
a) b
c) d)
e) f)
g) h)
26
Photographies des polinisateurs recensés : Apis mellifera (a), Xylocopa fimbriata (b), Exomalopsis simlis (c), Lasioglossum spp (d), Ornida obesa €, Palpada vinetorum (f), Melissodes martinicensis (g), Toxomerus spp (h).
Crédit Photo : Chloé PIERRE
DIPTERA
Syrphidae
Toxomerus floralis Fabricius (1798)
87 7 0 94
Toxomerus dispar (Fabricius, 1794)
18 7 0 25
Toxomerus pulchellus (Macquart, 1846)
1 0 0 1
Dioprosopa clavata (Fabricius, 1974)
15 13 0 28
Palpada vinetorum (Fabricius, 1798)
3 24 0 27
Ornida obesa e) (Fabricius, 1775)
0 2 0 2
3.2.2 Abondance et diversité des pollinisateurs selon les sites
1 3 5 7 9 11 13 15 17 19
05
1015
2025
parcelles
Abo
ndan
ce d
es p
ollin
isat
eurs
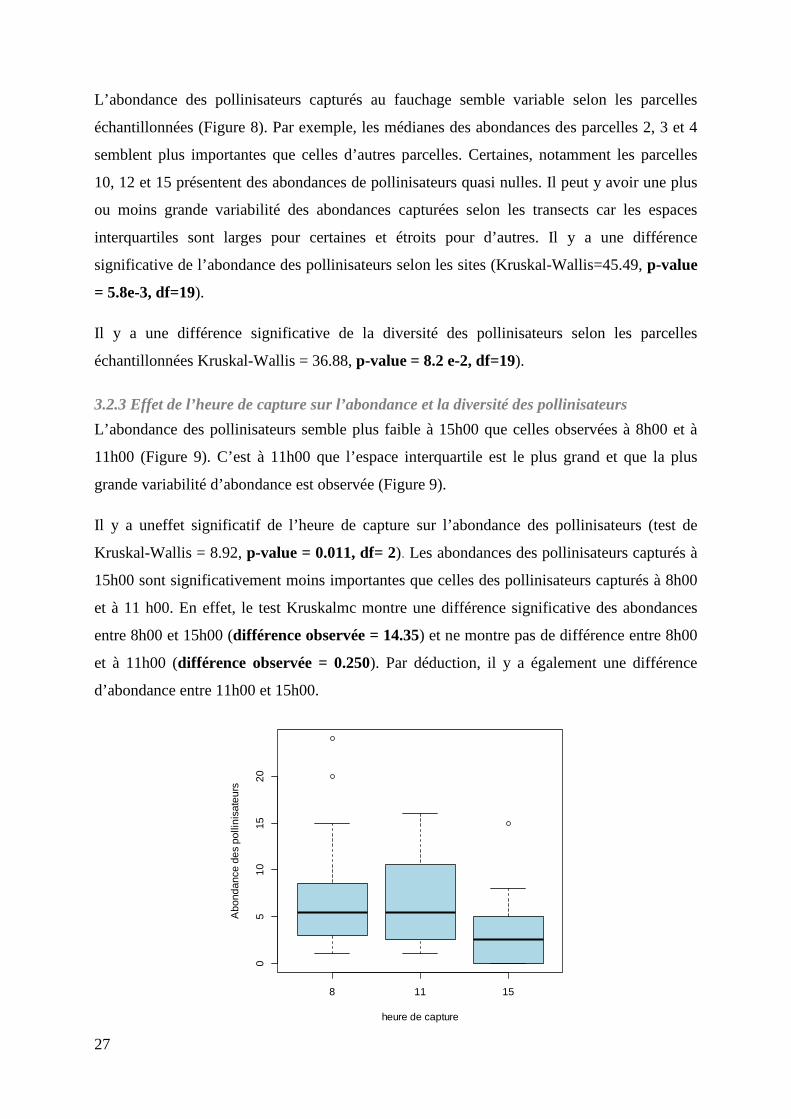
Figure 8 : Boîtes à moustaches des abondances de pollinisateurs selon la parcelle échantillonnées. La médiane est représentée par le trait noir épais, le rectangle orange correspond à l’écart entre le 1er et le 3ème quartile, les segments fins représentent les valeurs extrêmes.
27
L’abondance des pollinisateurs capturés au fauchage semble variable selon les parcelles
échantillonnées (Figure 8). Par exemple, les médianes des abondances des parcelles 2, 3 et 4
semblent plus importantes que celles d’autres parcelles. Certaines, notamment les parcelles
10, 12 et 15 présentent des abondances de pollinisateurs quasi nulles. Il peut y avoir une plus
ou moins grande variabilité des abondances capturées selon les transects car les espaces
interquartiles sont larges pour certaines et étroits pour d’autres. Il y a une différence
significative de l’abondance des pollinisateurs selon les sites (Kruskal-Wallis=45.49, p-value
= 5.8e-3, df=19).
Il y a une différence significative de la diversité des pollinisateurs selon les parcelles
échantillonnées Kruskal-Wallis = 36.88, p-value = 8.2 e-2, df=19).
3.2.3 Effet de l’heure de capture sur l’abondance et la diversité des pollinisateurs
L’abondance des pollinisateurs semble plus faible à 15h00 que celles observées à 8h00 et à
11h00 (Figure 9). C’est à 11h00 que l’espace interquartile est le plus grand et que la plus
grande variabilité d’abondance est observée (Figure 9).
Il y a uneffet significatif de l’heure de capture sur l’abondance des pollinisateurs (test de
Kruskal-Wallis = 8.92, p-value = 0.011, df= 2). Les abondances des pollinisateurs capturés à
15h00 sont significativement moins importantes que celles des pollinisateurs capturés à 8h00
et à 11 h00. En effet, le test Kruskalmc montre une différence significative des abondances
entre 8h00 et 15h00 (différence observée = 14.35) et ne montre pas de différence entre 8h00
et à 11h00 (différence observée = 0.250). Par déduction, il y a également une différence
d’abondance entre 11h00 et 15h00.
8 11 15
05
1015
20
heure de capture
Abo
ndan
ce d
es p
ollin
isat
eurs

28
Figure 9 : Boîtes à moustaches des abondances de pollinisateurs selon l’heure de capture par fauchage
Même si l’abondance des pollinisateurs diminue l’après-midi, les abeilles et les syrphes
recensés à 15h00 ne sont pas moins diversifiés. En effet, l’heure de capture n’agit pas
significativement sur la diversité des pollinisateurs (Test de Kruskal-Wallis = 0.44, p-value =
0.80, df = 2).
3.3 Effet des adventices sur les communautés de pollinisateurs
3.3.1 Bilan sur les relevés floristiques
Au total, 118espèces d’adventices réparties dans 28 familles ont été recensées. Parmi toutes
les espèces d’adventices inventoriées, 71 ont été observées en fleurset ont contribué aux
estimations de recouvrement floral et de nombre d’inflorescences. Les pollinisateurs ont été
observés en activité de butinage sur 35 espèces en fleurs soit 50% des fleurs que l’on peut
rencontrer dans les couverts.
3.3.2 Effet de la diversité des adventices sur l’abondance et la diversité des pollinisateurs
Il n’y a pas d’effet significatif de la richesse spécifique totale des adventices (en fleurs et non
en fleurs) sur l’abondance et sur la diversité des pollinisateurs (Tableau 2). De plus, les
Pseudos R2 sont très faibles et les deux modèles ne permettent pas d’expliquer l’abondance ni
la diversité des pollinisateurs
Tableau 2 : Effet de la richesse spécifique des adventices : S adventives (en fleur et non en fleurs) sur les communautés de pollinisateurs sur les 3 transects des 20 parcelles étudiées. Deux modèles linéaires ont été réalisés. Le modèle qui porte sur l’abondance des pollinisateurs suit une loi de Poisson (1) tandis que celui qui porte sur la diversité des pollinisateurs (indice de Shanon) suit une loi de Gauss (2).
Modèles Variable explicative Coefficient z-value
p-value Pseudo R2
Abondance pollinisateurs GLM (1)
Intercept 1.76+/-0.18 9.48 *** 0.05 S adventices
(en fleurs et non en fleurs)
-0.002+/-0.001 -0.205 NS
Diversité pollinisateurs GLM (2)
Intercept -1.5+/-0.59 -2.53 * 0.06 S adventices
(en fleurs et non en fleurs)
0.05+/-0.03 1.660 NS
*p<0.05 ; **p<0.01 ; ***p<0.001 NS=non significatif
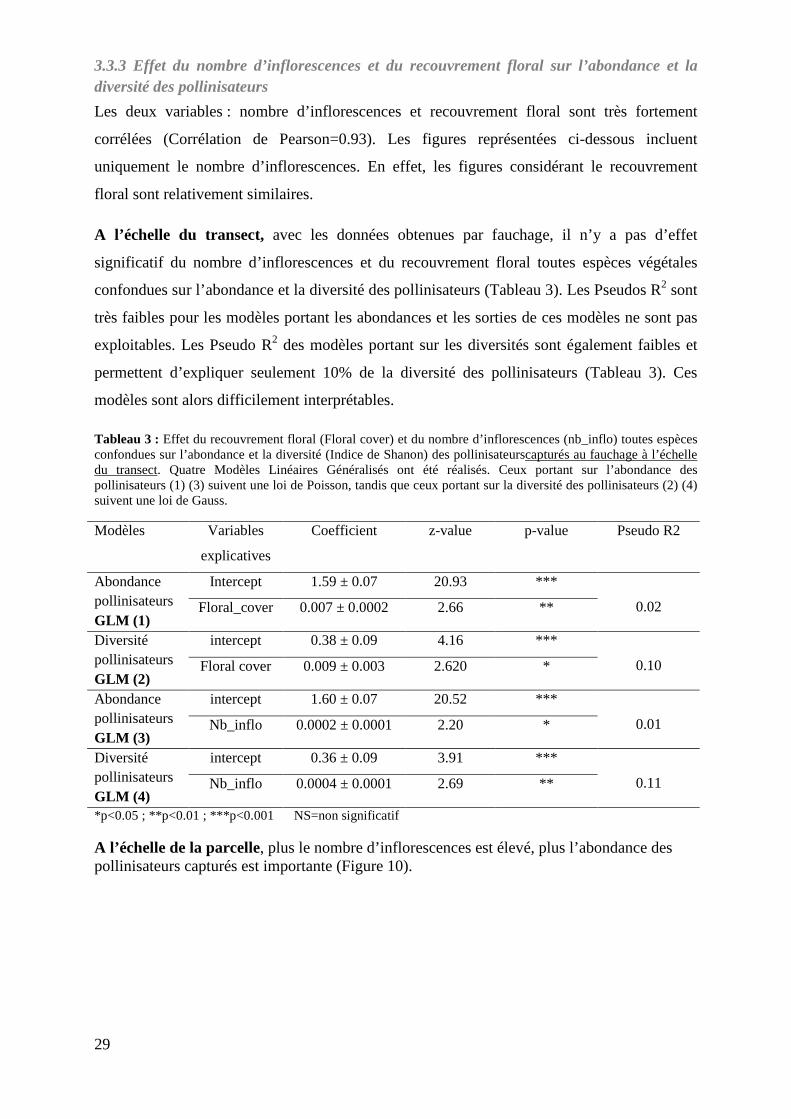
29
3.3.3 Effet du nombre d’inflorescences et du recouvrement floral sur l’abondance et la diversité des pollinisateurs
Les deux variables : nombre d’inflorescences et recouvrement floral sont très fortement
corrélées (Corrélation de Pearson=0.93). Les figures représentées ci-dessous incluent
uniquement le nombre d’inflorescences. En effet, les figures considérant le recouvrement
floral sont relativement similaires.
A l’échelle du transect, avec les données obtenues par fauchage, il n’y a pas d’effet
significatif du nombre d’inflorescences et du recouvrement floral toutes espèces végétales
confondues sur l’abondance et la diversité des pollinisateurs (Tableau 3). Les Pseudos R2 sont
très faibles pour les modèles portant les abondances et les sorties de ces modèles ne sont pas
exploitables. Les Pseudo R2 des modèles portant sur les diversités sont également faibles et
permettent d’expliquer seulement 10% de la diversité des pollinisateurs (Tableau 3). Ces
modèles sont alors difficilement interprétables.
Tableau 3 : Effet du recouvrement floral (Floral cover) et du nombre d’inflorescences (nb_inflo) toutes espèces confondues sur l’abondance et la diversité (Indice de Shanon) des pollinisateurscapturés au fauchage à l’échelle du transect. Quatre Modèles Linéaires Généralisés ont été réalisés. Ceux portant sur l’abondance des pollinisateurs (1) (3) suivent une loi de Poisson, tandis que ceux portant sur la diversité des pollinisateurs (2) (4) suivent une loi de Gauss.
Modèles Variables
explicatives
Coefficient z-value p-value Pseudo R2
Abondance pollinisateurs GLM (1)
Intercept 1.59 ± 0.07 20.93 ***
0.02 Floral_cover 0.007 ± 0.0002 2.66 **
Diversité pollinisateurs GLM (2)
intercept 0.38 ± 0.09 4.16 ***
0.10 Floral cover 0.009 ± 0.003 2.620 *
Abondance pollinisateurs GLM (3)
intercept 1.60 ± 0.07 20.52 ***
0.01 Nb_inflo 0.0002 ± 0.0001 2.20 *
Diversité pollinisateurs GLM (4)
intercept 0.36 ± 0.09 3.91 ***
0.11 Nb_inflo 0.0004 ± 0.0001 2.69 **
*p<0.05 ; **p<0.01 ; ***p<0.001 NS=non significatif A l’échelle de la parcelle, plus le nombre d’inflorescences est élevé, plus l’abondance des pollinisateurs capturés est importante (Figure 10).
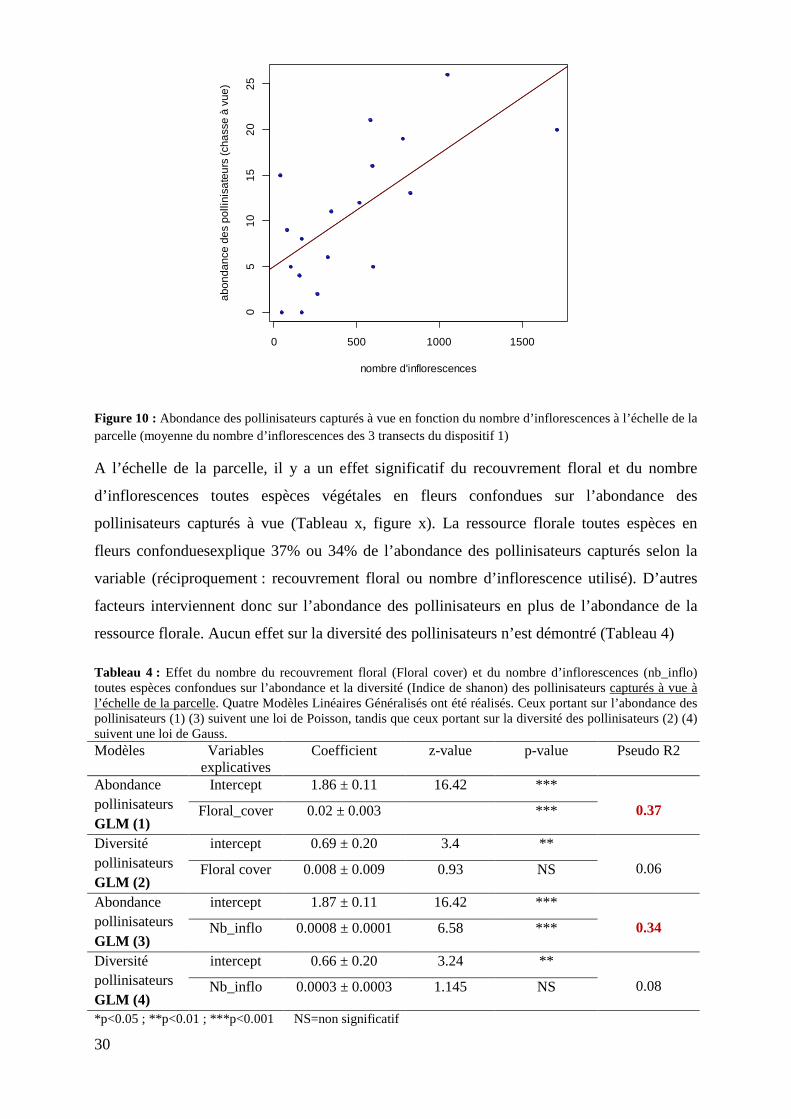
30
0 500 1000 1500
05
1015
2025
nombre d'inflorescences
abon
danc
e de
s po
llini
sate
urs
(cha
sse
à vu
e)
Figure 10 : Abondance des pollinisateurs capturés à vue en fonction du nombre d’inflorescences à l’échelle de la parcelle (moyenne du nombre d’inflorescences des 3 transects du dispositif 1)
A l’échelle de la parcelle, il y a un effet significatif du recouvrement floral et du nombre
d’inflorescences toutes espèces végétales en fleurs confondues sur l’abondance des
pollinisateurs capturés à vue (Tableau x, figure x). La ressource florale toutes espèces en
fleurs confonduesexplique 37% ou 34% de l’abondance des pollinisateurs capturés selon la
variable (réciproquement : recouvrement floral ou nombre d’inflorescence utilisé). D’autres
facteurs interviennent donc sur l’abondance des pollinisateurs en plus de l’abondance de la
ressource florale. Aucun effet sur la diversité des pollinisateurs n’est démontré (Tableau 4)
Tableau 4 : Effet du nombre du recouvrement floral (Floral cover) et du nombre d’inflorescences (nb_inflo) toutes espèces confondues sur l’abondance et la diversité (Indice de shanon) des pollinisateurs capturés à vue à l’échelle de la parcelle. Quatre Modèles Linéaires Généralisés ont été réalisés. Ceux portant sur l’abondance des pollinisateurs (1) (3) suivent une loi de Poisson, tandis que ceux portant sur la diversité des pollinisateurs (2) (4) suivent une loi de Gauss. Modèles Variables
explicatives Coefficient z-value p-value Pseudo R2
Abondance pollinisateurs GLM (1)
Intercept 1.86 ± 0.11 16.42 ***
0.37 Floral_cover 0.02 ± 0.003 ***
Diversité pollinisateurs GLM (2)
intercept 0.69 ± 0.20 3.4 **
0.06 Floral cover 0.008 ± 0.009 0.93 NS
Abondance pollinisateurs GLM (3)
intercept 1.87 ± 0.11 16.42 ***
0.34 Nb_inflo 0.0008 ± 0.0001 6.58 ***
Diversité pollinisateurs GLM (4)
intercept 0.66 ± 0.20 3.24 **
0.08 Nb_inflo 0.0003 ± 0.0003 1.145 NS
*p<0.05 ; **p<0.01 ; ***p<0.001 NS=non significatif
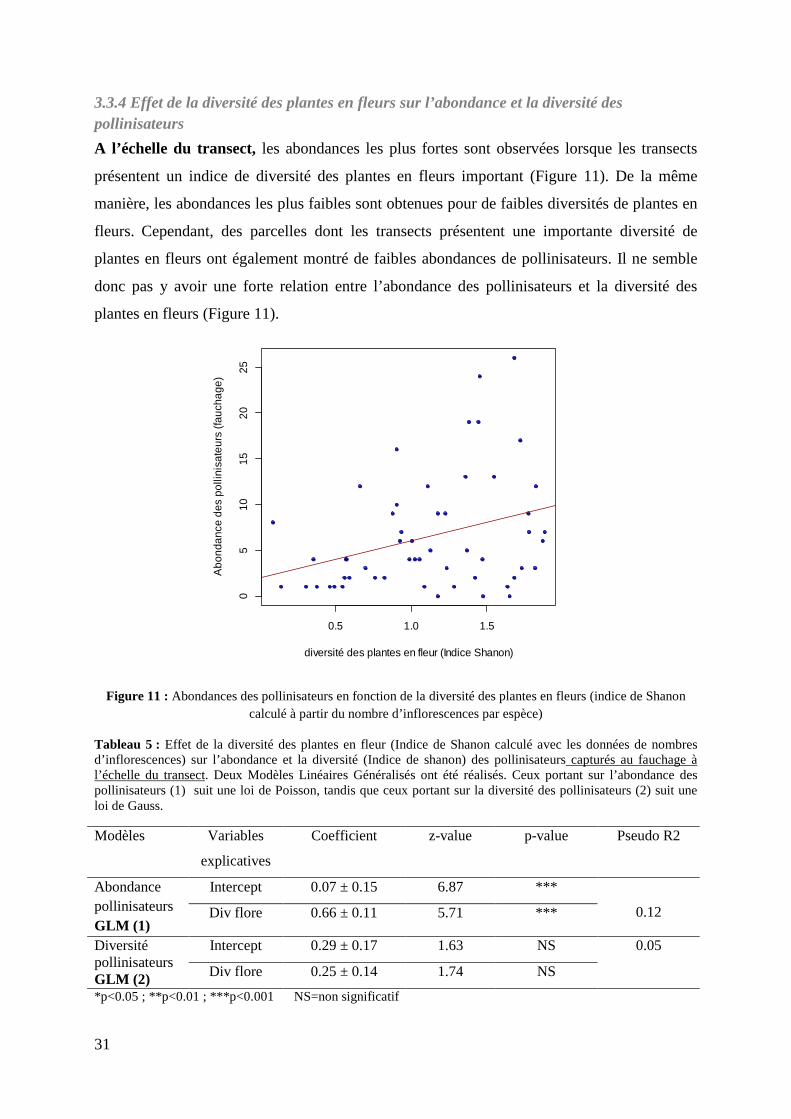
31
3.3.4 Effet de la diversité des plantes en fleurs sur l’abondance et la diversité des pollinisateurs
A l’échelle du transect, les abondances les plus fortes sont observées lorsque les transects
présentent un indice de diversité des plantes en fleurs important (Figure 11). De la même
manière, les abondances les plus faibles sont obtenues pour de faibles diversités de plantes en
fleurs. Cependant, des parcelles dont les transects présentent une importante diversité de
plantes en fleurs ont également montré de faibles abondances de pollinisateurs. Il ne semble
donc pas y avoir une forte relation entre l’abondance des pollinisateurs et la diversité des
plantes en fleurs (Figure 11).
0.5 1.0 1.5
05
1015
2025
diversité des plantes en fleur (Indice Shanon)
Abo
ndan
ce d
es p
ollin
isat
eurs
(fau
chag
e)
Figure 11 : Abondances des pollinisateurs en fonction de la diversité des plantes en fleurs (indice de Shanon calculé à partir du nombre d’inflorescences par espèce)
Tableau 5 : Effet de la diversité des plantes en fleur (Indice de Shanon calculé avec les données de nombres d’inflorescences) sur l’abondance et la diversité (Indice de shanon) des pollinisateurs capturés au fauchage à l’échelle du transect. Deux Modèles Linéaires Généralisés ont été réalisés. Ceux portant sur l’abondance des pollinisateurs (1) suit une loi de Poisson, tandis que ceux portant sur la diversité des pollinisateurs (2) suit une loi de Gauss.
Modèles Variables
explicatives
Coefficient z-value p-value Pseudo R2
Abondance pollinisateurs GLM (1)
Intercept 0.07 ± 0.15 6.87 ***
0.12 Div flore 0.66 ± 0.11 5.71 ***
Diversité pollinisateurs GLM (2)
Intercept 0.29 ± 0.17 1.63 NS 0.05
Div flore 0.25 ± 0.14 1.74 NS
*p<0.05 ; **p<0.01 ; ***p<0.001 NS=non significatif
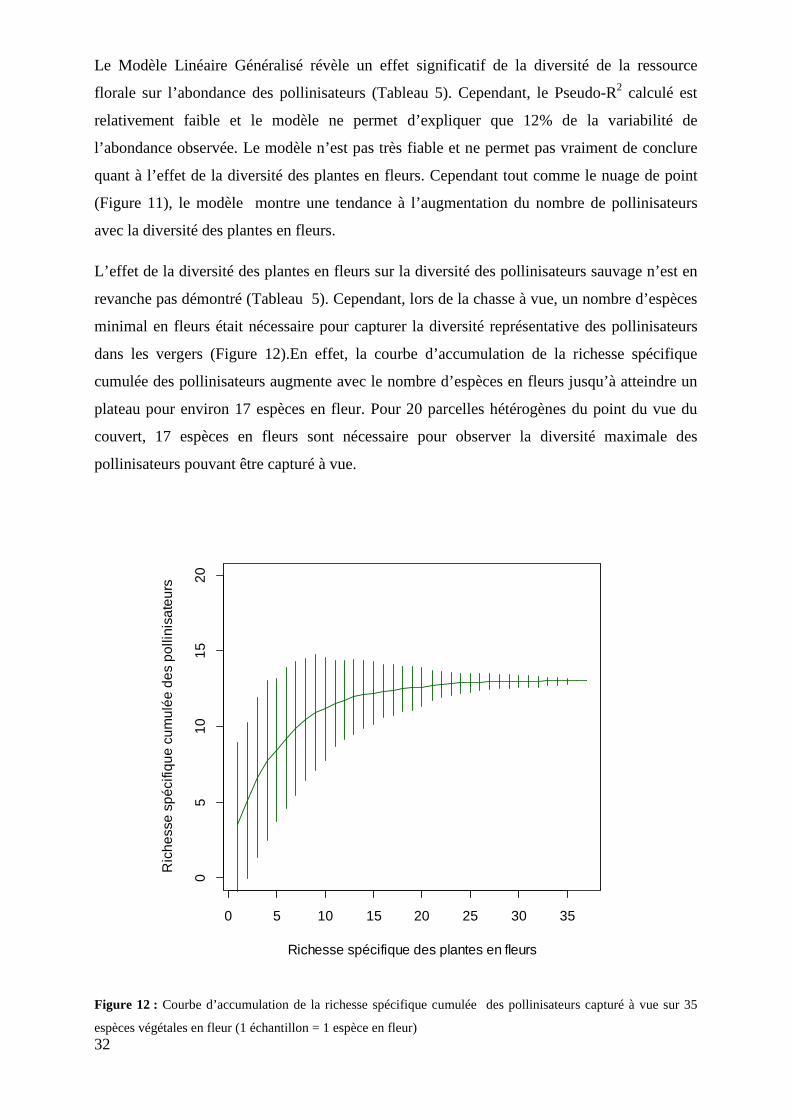
32
Le Modèle Linéaire Généralisé révèle un effet significatif de la diversité de la ressource
florale sur l’abondance des pollinisateurs (Tableau 5). Cependant, le Pseudo-R2 calculé est
relativement faible et le modèle ne permet d’expliquer que 12% de la variabilité de
l’abondance observée. Le modèle n’est pas très fiable et ne permet pas vraiment de conclure
quant à l’effet de la diversité des plantes en fleurs. Cependant tout comme le nuage de point
(Figure 11), le modèle montre une tendance à l’augmentation du nombre de pollinisateurs
avec la diversité des plantes en fleurs.
L’effet de la diversité des plantes en fleurs sur la diversité des pollinisateurs sauvage n’est en
revanche pas démontré (Tableau 5). Cependant, lors de la chasse à vue, un nombre d’espèces
minimal en fleurs était nécessaire pour capturer la diversité représentative des pollinisateurs
dans les vergers (Figure 12).En effet, la courbe d’accumulation de la richesse spécifique
cumulée des pollinisateurs augmente avec le nombre d’espèces en fleurs jusqu’à atteindre un
plateau pour environ 17 espèces en fleur. Pour 20 parcelles hétérogènes du point du vue du
couvert, 17 espèces en fleurs sont nécessaire pour observer la diversité maximale des
pollinisateurs pouvant être capturé à vue.
0 5 10 15 20 25 30 35
05
1015
20
Richesse spécifique des plantes en fleurs
Ric
hess
e sp
écifi
que
cum
ulée
des
pol
linis
ateu
rs
Figure 12 : Courbe d’accumulation de la richesse spécifique cumulée des pollinisateurs capturé à vue sur 35
espèces végétales en fleur (1 échantillon = 1 espèce en fleur)
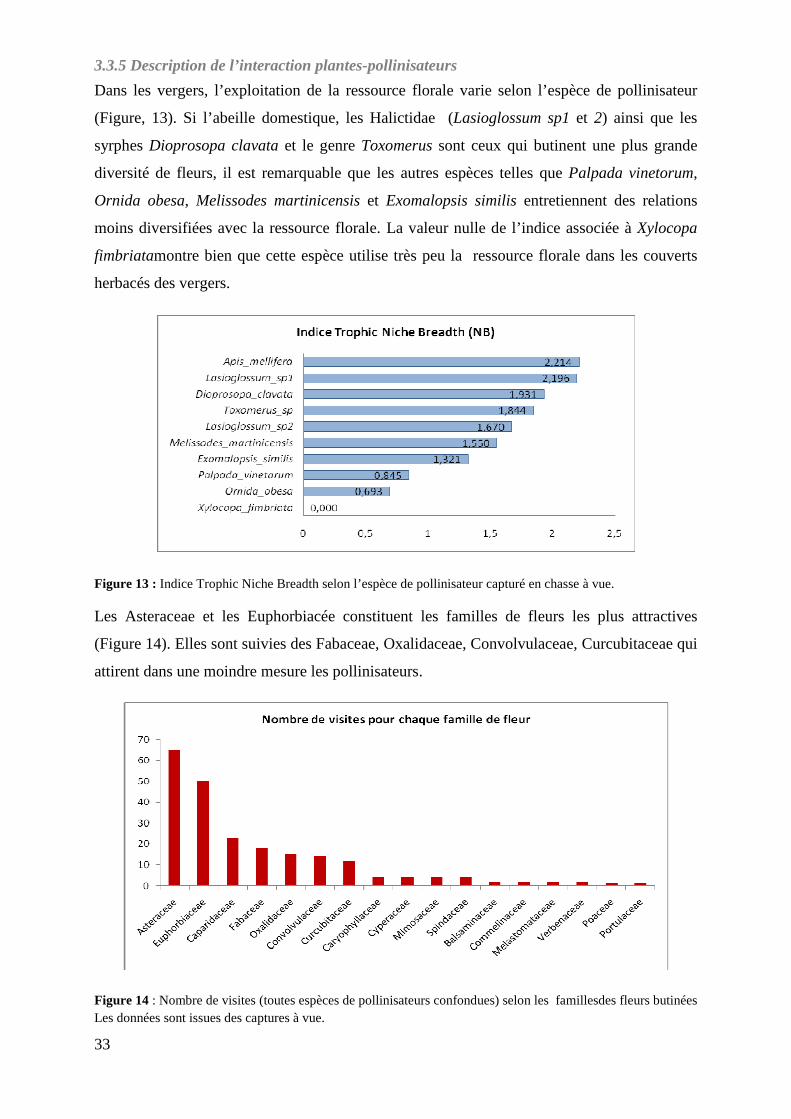
33
3.3.5 Description de l’interaction plantes-pollinisateurs
Dans les vergers, l’exploitation de la ressource florale varie selon l’espèce de pollinisateur
(Figure, 13). Si l’abeille domestique, les Halictidae (Lasioglossum sp1 et 2) ainsi que les
syrphes Dioprosopa clavata et le genre Toxomerus sont ceux qui butinent une plus grande
diversité de fleurs, il est remarquable que les autres espèces telles que Palpada vinetorum,
Ornida obesa, Melissodes martinicensis et Exomalopsis similis entretiennent des relations
moins diversifiées avec la ressource florale. La valeur nulle de l’indice associée à Xylocopa
fimbriatamontre bien que cette espèce utilise très peu la ressource florale dans les couverts
herbacés des vergers.
Figure 13 : Indice Trophic Niche Breadth selon l’espèce de pollinisateur capturé en chasse à vue.
Les Asteraceae et les Euphorbiacée constituent les familles de fleurs les plus attractives
(Figure 14). Elles sont suivies des Fabaceae, Oxalidaceae, Convolvulaceae, Curcubitaceae qui
attirent dans une moindre mesure les pollinisateurs.
Figure 14 : Nombre de visites (toutes espèces de pollinisateurs confondues) selon les famillesdes fleurs butinées Les données sont issues des captures à vue.

34
Le nombre de visites des pollinisateurs sur les fleurs du couvert dans les parcelles étudiées
varie selon les espèces végétales visitées (Figure 15). La chasse à vue permet alors de mettre
en évidence des préférences. Les fleurs les plus butinées sont Tridax procumbens et
Euphorbia heterophylla. Cleome rutidosperma et Oxalis barrelieri sont des fleurs
régulièrement butinées par les pollinisateurs dans les vergers.
Figure 15 : Nombre de visites (toutes espèces de pollinisateurs confondues) pour chaque espèce en fleurs à partir des captures de la chasse à vue.
3.4Effet de l’hétérogénéité à l’échelle intra-parcellaire et effet de l’environnement proche sur l’abondance et la diversité des pollinisateurs
3.4.1Effet de la bordure de parcelle sur l’abondance et la diversité des pollinisateurs.
Il n’y a pas de différence significative du recouvrement floral des adventices selon la position
du transect décrite dans le dispositif 1(Kruskal-Wallis = 0.58, p-value= 0.74, df =2). Il n’y a
pas non plus d’effet significatif de la position du transect sur le nombre d’inflorescences
(Kruskal-Wallis = 0.90, p-value = 0.63, df=2). L’abondance de la ressource florale est donc
relativement homogène à l’échelle de la parcelle et les bordures ne présentent pas plus de
ressources que les inter-rangs à l’intérieur des parcelles.
Les bordures de parcelles semblent présenter une richesse spécifique totale des adventices (en
fleurs et non en fleurs) plus élevée que celles recensées sur les transects 2 et 3 (Figure 16). Il y
a un effet significatif de la position du transect sur la richesse spécifique des adventices
(Kruskal-Wallis= 13.29, p-value = 0.001, df=2). Les transects en bordure de parcelle
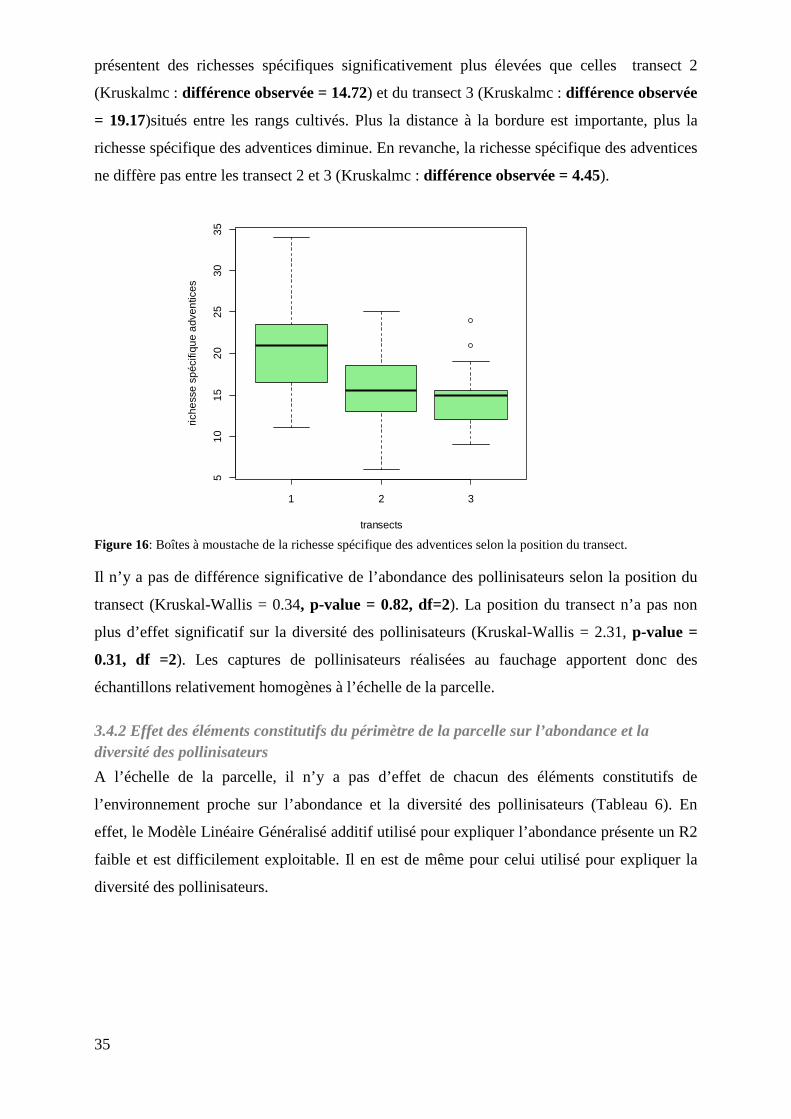
35
1 2 3
510
1520
2530
35
transects
riche
sse
spéc
ifiqu
e ad
vent
ices
présentent des richesses spécifiques significativement plus élevées que celles transect 2
(Kruskalmc : différence observée = 14.72) et du transect 3 (Kruskalmc : différence observée
= 19.17)situés entre les rangs cultivés. Plus la distance à la bordure est importante, plus la
richesse spécifique des adventices diminue. En revanche, la richesse spécifique des adventices
ne diffère pas entre les transect 2 et 3 (Kruskalmc : différence observée = 4.45).
Figure 16: Boîtes à moustache de la richesse spécifique des adventices selon la position du transect.
Il n’y a pas de différence significative de l’abondance des pollinisateurs selon la position du
transect (Kruskal-Wallis = 0.34, p-value = 0.82, df=2). La position du transect n’a pas non
plus d’effet significatif sur la diversité des pollinisateurs (Kruskal-Wallis = 2.31, p-value =
0.31, df =2). Les captures de pollinisateurs réalisées au fauchage apportent donc des
échantillons relativement homogènes à l’échelle de la parcelle.
3.4.2 Effet des éléments constitutifs du périmètre de la parcelle sur l’abondance et la diversité des pollinisateurs
A l’échelle de la parcelle, il n’y a pas d’effet de chacun des éléments constitutifs de
l’environnement proche sur l’abondance et la diversité des pollinisateurs (Tableau 6). En
effet, le Modèle Linéaire Généralisé additif utilisé pour expliquer l’abondance présente un R2
faible et est difficilement exploitable. Il en est de même pour celui utilisé pour expliquer la
diversité des pollinisateurs.
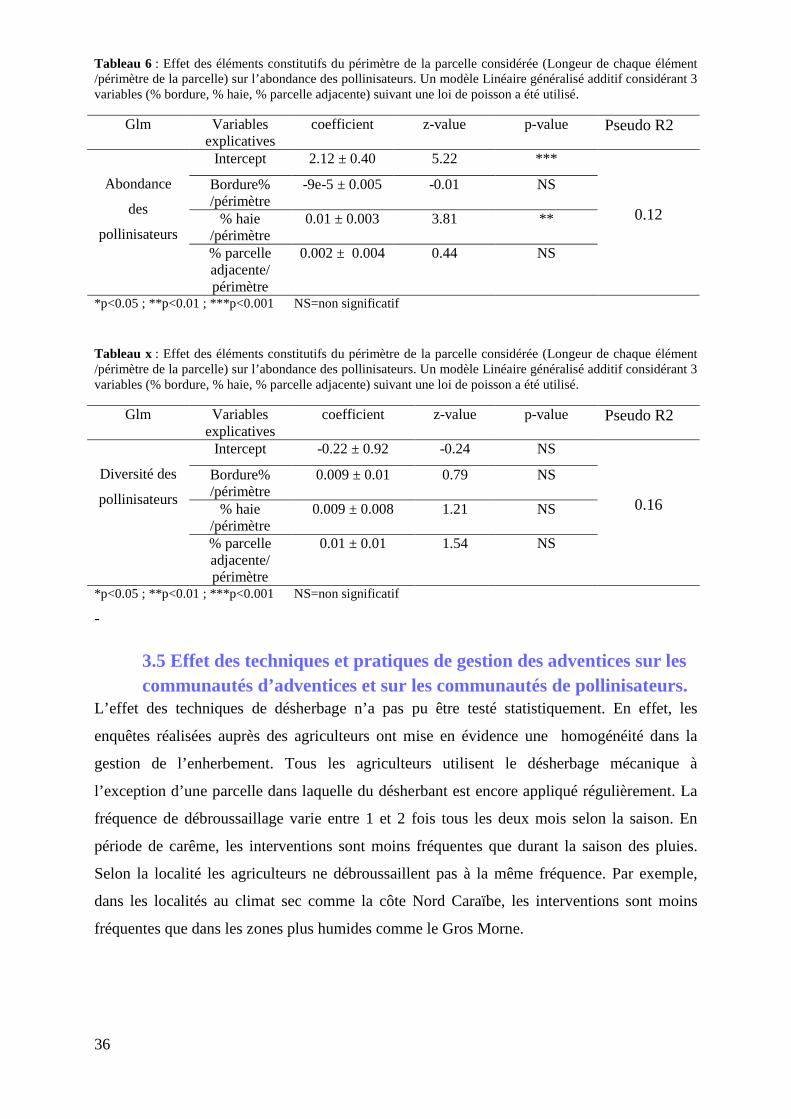
36
Tableau 6 : Effet des éléments constitutifs du périmètre de la parcelle considérée (Longeur de chaque élément /périmètre de la parcelle) sur l’abondance des pollinisateurs. Un modèle Linéaire généralisé additif considérant 3 variables (% bordure, % haie, % parcelle adjacente) suivant une loi de poisson a été utilisé.
Glm Variables explicatives
coefficient z-value p-value Pseudo R2
Abondance
des
pollinisateurs
Intercept 2.12 ± 0.40 5.22 ***
0.12
Bordure% /périmètre
-9e-5 ± 0.005 -0.01 NS
% haie /périmètre
0.01 ± 0.003 3.81 **
% parcelle adjacente/ périmètre
0.002 ± 0.004 0.44 NS
*p<0.05 ; **p<0.01 ; ***p<0.001 NS=non significatif
Tableau x : Effet des éléments constitutifs du périmètre de la parcelle considérée (Longeur de chaque élément /périmètre de la parcelle) sur l’abondance des pollinisateurs. Un modèle Linéaire généralisé additif considérant 3 variables (% bordure, % haie, % parcelle adjacente) suivant une loi de poisson a été utilisé.
Glm Variables explicatives
coefficient z-value p-value Pseudo R2
Diversité des
pollinisateurs
Intercept -0.22 ± 0.92 -0.24 NS
0.16
Bordure% /périmètre
0.009 ± 0.01 0.79 NS
% haie /périmètre
0.009 ± 0.008 1.21 NS
% parcelle adjacente/ périmètre
0.01 ± 0.01 1.54 NS
*p<0.05 ; **p<0.01 ; ***p<0.001 NS=non significatif
-
3.5 Effet des techniques et pratiques de gestion des adventices sur les communautés d’adventices et sur les communautés de pollinisateurs.
L’effet des techniques de désherbage n’a pas pu être testé statistiquement. En effet, les
enquêtes réalisées auprès des agriculteurs ont mise en évidence une homogénéité dans la
gestion de l’enherbement. Tous les agriculteurs utilisent le désherbage mécanique à
l’exception d’une parcelle dans laquelle du désherbant est encore appliqué régulièrement. La
fréquence de débroussaillage varie entre 1 et 2 fois tous les deux mois selon la saison. En
période de carême, les interventions sont moins fréquentes que durant la saison des pluies.
Selon la localité les agriculteurs ne débroussaillent pas à la même fréquence. Par exemple,
dans les localités au climat sec comme la côte Nord Caraïbe, les interventions sont moins
fréquentes que dans les zones plus humides comme le Gros Morne.

37
4. Discussion 4.1Méthodologie et effort de prospection
4.1.1 Les trois techniques de captures sont complémentaires
Cette étude fournit pour la première fois des données sur la diversité et l’abondance des
insectes pollinisateurs associés aux cultures fruitières en Martinique. Les différentes
techniques de capture ont permis de recenser une diversité des pollinisateurs représentative de
celle du milieu étudié et se sont montrées complémentaires. En effet, selon la technique de
captures et l’échelle utilisée comme unité de référence, les effets des différentes variables
environnementales étaient plus ou moins visibles et démontrés. La méthodologie, de part la
diversité techniques de capture, des échelles utilisées et des différentes variables testées peut
servir de support pour les études ultérieures et pourra alors être améliorée et affinée.
Les pièges « bols colorés » constituent une méthode de capture plus sélective que le fauchage
et conformément à d’autres études, dans d’autres régions, cela permet d’attraper une très
grande quantité d’abeilles sauvages, notamment des Halictidae (Le Feon, 2010 ; Mandelick et
Roll, 2009 ; Roultson et al, 2007). Il a été noté que les bols colorées sous-estiment les abeilles
de grandes tailles (Le Feon, 2010 ; Westphal, 2008). Les abeilles domestiques et autres
Apidae (Exomalopsis similis et Melissodes martinicensis) sont alors peut- être sous estimées
par rapport aux Halictidae (Annexe).En Martinique cette méthode exclue catégoriquement la
capture des syrphes tandis que dans ces autres études, certains syrphes, certes moins
abondants que les Apoïdes, avaient tout de même été piégés dans les bols colorés (Le Féon,
2010). L’utilisation des bols colorés est complémentaire des captures au filet (Westphal,
2008) et serait adaptée pour une étude des abeilles sauvages à une échelle plus globales ainsi
que pour effectuer des comparaisons entre les parcelles étudiées.
4.1.2 Un effort d’échantillonnage suffisant mais des localités sur l’île restent à étudier
Malgré l’effort d’échantillonnage suffisant et même si les principales zones de production
fruitières ont été visitées et apportent l’essentiel des informations, d’autres localités sur l’île
pourraient être échantillonnées. Cela assurerait que la diversité recensée est représentative de
la diversité réelle des couverts des vergers à l’échelle du territoire. Certaines localités comme
la côte Atlantique (Sainte-Marie, le François, le Vauclin), le nord de l’île (le Prêcheur) ou
encore l’extrémité sud n’ont pas été échantillonnées en raison de l’état du couvert des
parcelles au moment de la campagne de terrain et de la disponibilité des agriculteurs.

38
4.1.3 Les pollinisateurs ont une activité de butinage plus importante le matin
Les plus faibles abondances de pollinisateurs observées l’après-midi suggèrent une activité
pollinisatrice plus importante le matin. Cela a déjà été constaté dans des études portant sur
l’activité journalière des pollinisateurs en milieu tropical (Fohouo, 2002 ;Roubick, 1989).
Certains agriculteurs ont témoigné d’une activité très matinale des abeilles domestiques dans
leurs vergers à partir de 6h00 (Commentaire : Poulin, Glombart, Troudart). En milieu
tempéré, l’activité de butinage a lieu préférentiellement le matin et en milieu de journée. La
méthodologie de certaines études implique d’ailleurs des relevés entre 10h00 et14h00.En
Martinique, le soleil se lève et se couche plus tôt et les pollinisateurs seraient actifs dès le
lever du jour. Les abeilles sauvages iraient jusqu’à butiner des Graminées avant même le lever
du jour. En effet, la rosée facilite la déhiscence des grains de pollen qui sèchent ensuite
rapidement au cours de la matinée (Jaeger, 1954 ; Roubick, 1989).Sur le terrain, pendant la
période de carême, il a été constaté un flétrissement rapide de certaines fleurs butinées en fin
de matin du fait de la chaleur. Les pollinisateurs seraient alors actifs le matin pour optimiser
leur récolte en pollen et en nectar. Les abeilles sauvages récoltent la ressource pour ensuite la
transporter jusqu’à leur nid pour approvisionner leurs larves. Les abeilles conditionneraient
leur énergie le matin à la recherche de ressources, et l’après midi serait davantage consacré a
nourrir les larves au nid. Effectuer des relevés plus tôt avant 8h00 enrichirait peut-être les
données concernant la présence des pollinisateurs domestiques et sauvages ainsi que les fleurs
butinées dans les couverts. Bien que la diversité des pollinisateurs ne soit pas différente selon
l’heure de capture, la période d’activité floricole peut varier sensiblement dans la matinée
entre les différentes espèces (Njoroge et al, 2010). Les syrphes et les abeilles n’ont pas le
même comportement alimentaire et ne sont pas dissociés dans cette étude.
4.2 Les Apoïdes et les syrphes capturés dans les couverts des vergers
4.2.1. Une faible diversité des pollinisateurs recensés en raison des particularités du milieu
insulaire
Au total, 14 espèces de pollinisateurs (abeilles et syrphes confondus) on été recensées. Cela
peut paraître étonnant lorsque des études en Europe collectent plus de 50espèces (Kwaiser &
Hendrix, 2007 ; Le Féon, 2010). L’isolement au continent américain a un effet sur la diversité
des espèces et conduit à de l’endémisme. Dans les Antilles Française, l’entomofaune
représenterait moins de 20% de ce qui est observé sur le continent en Guyane (Touroult,
2011). La richesse spécifique d’une île est directement liée à la diversité des milieux, à la
superficie de l’île et à la distance au continent (Mac Arthur & Wilson, 1967 ; Touroult, 2011).

39
La Martinique est une île dont la superficie est réduite (1128) (Sastre & Breuil, 2007) et
répond à cette théorie de l’équilibre insulaire.
4.2.2. Des communautés de pollinisateurs non équitables
Parmi les pollinisateurs recensés, des Apidae, une Megachilidae, desHalictidae et des
Syrphidae ont été identifiés. La Martinique compte 14 espèces d’abeilles (Apoidae :
Anthophila) ;(Meurgey & Dumbardon-Martial, sous presse). Ainsi, 46% de la faune
apidologique de l’île sont présents dans les vergers. La majorité des espèces ont une
répartition géographique large (répartition insulaire et continentale) et seule Melissodes
martinicensis est endémique stricte à l’île. Nos résultats d’abondance montrentaussi la
présence de cette espèce dans les parcelles agricoles. En effet, nous la croyions peu présente
en milieu cultivé et les abondances de cette espèce révélées par les bols colorés corrigent cette
sous-estimation. Chez les Halictidae, les deux espèces du genre Lasioglossum (Dialictus)
sont de loin les plus abondantes et dominent constamment les communautés de pollinisateurs.
Les premières analyses taxonomiques de ces deux espèces prétendent qu’il s’agit d’espèces
nouvelles pour la science. Les Halictidae et les Apidés du genre Melissodes sont des nicheurs
du sol et nidifient dans les chemins terreux, les sols nus et parfois même au sein des parcelles
cultivées (Normandin et al, 2014 ; Villemant, 2005). Les exploitations agricoles et leurs
environs peuvent offrir des terrains propices a la nidification de certains pollinisateurs qui
sont alors plus abondants. La présence d’Exomalopsis similis dans les parcelles est très
variables, nous avons constaté sur le terrain qu’elle peut être aussi bien absente qu’abondante
très localement à proximité d’un patch de fleur attractive pour elle. L’abeille domestique a été
retrouvée dans presque toutes les parcelles en abondance variable. Les agriculteurs
témoignaient de la présence ou non de ruches ou d’essaims à proximité ou dans les
exploitations qui peuvent fortement influencer les observations. Les syrphes sont
principalement représentés par les espèces Toxomerus floralis et Toxomerus dispar. Ces
dernières, cosmopolites font parties des espèces les plus abondantes dans toutes les Antilles et
l’Amérique(Thompson, 1981).Ces syrphes sont particulièrement associés à la strate herbacée
des couverts et sont des prédateurs de cicadelles elles aussi abondantes dans ce milieu.
4.2.3. La diversité et l’abondance des pollinisateurs diffère selon les parcelles mais les espèces sont représentées de manière homogène à l’échelle globale
Les analyses ont montré que la diversité et l’abondance des pollinisateurs varient selon les
parcelles échantillonnées. Cependant, en général, l’ensemble des espèces recensées ont toutes
été observées dans les différentes localités à l’exception du Morne Rouge pour laquelle une
seule parcelle a été étudiée et peu de données ont pu être obtenues en raison des conditions

40
météorologiques et de la disponibilité des agriculteurs. On suppose donc que l’effet de la
localité sur l’île (Nord, Sud, plaine du centre côte Atlantique et côte Caraïbe) qui n’a pas pu
être incluse dans l’étude n’a pas eu d’impact sur l’estimation de larichesse spécifique des
communautés de pollinisateurs dans les vergers. La variabilité observée entre les parcelles est
donc due à des facteurs liés à l’environnement, aux paysages, aux ressources ainsi qu’aux
pratiques culturales. Parmi ces facteurs la ressource florale comme facteur structurant les
communautés de pollinisateurs a pu être étudiée.
4.3 La ressource florale explique en partie la présence des pollinisateurs dans les vergers
4.3.1 La ressource florale a un effet positif sur l’abondance des pollinisateurs mais aucun effet sur la diversité n’a été démontré. Il y a un effet de l’échelle et/ou de la technique de capture sur ce résultat.
Le nombre d’inflorescences et le recouvrement floral toutes espèces végétales confondues
expliquent en partie l’abondance et n’expliquent pas la diversité des pollinisateurs.Cependant,
cela n’est visible qu’à l’échelle de la parcelle avec les données de chasse à vue. La diversité
de la ressource florale a peu d’effet sur l’abondance et aucun effet sur la diversité des
pollinisateurs. Le fauchage est sans doute un peu aléatoire et la chasse à vue plus précise pour
mettre en évidence de telles corrélations. De plus, l’échelle des transects n’est peut être pas
adaptée du fait de la petite taille des parcelles car les pollinisateurs peuvent aisément se
disperser dans les parcelles qui doivent alors être prises comme unité d’étude. Les résultats
obtenus sont plus ou moins cohérents avec d’autres études réalisées qui ont montré des effets
significatifs positifs de l’abondance et de la diversité de la ressource florale sur l’abondance
et la diversité des abeilles sauvages (Hine & Hendrix, 2005, Holzchuh, 2007, Potts et al,
2003). Kaiser & Hendrix (2007) ont ainsi montré un effet positif de la diversité des espèces
en fleurs (à abondance constante) sur l’abondance des pollinisateurs dans l’Iowa. Les
pollinisateurs recensés dans cette étude sont aussi essentiellement des Halictidae et des
Anthophoridae du genre Melissodes. En Allemagne, il a été démontré que la diversité des
syrphes est étroitement corrélée positivement à la densité et à la diversité de la ressource
florale (Haenke, 2009). En revanche, parmi d’autres études Steffan-Dewenter & Tscharntke
(2001)montré que le recouvrement floral agit positivement plus sur l’abondance que sur la
diversité des pollinisateurs.Cependant, ils ont aussi démontré que la diversité des plantes en
fleur agit surtout sur la diversité des pollinisateurs, relation que nous n’avons pas exactement
observée.D’autres facteurs interviennent sans doute dans la structuration des communautés de
pollinisateurs notamment les éléments du paysage ou la conduite des cultures notamment les
pratiques portant atteinte au couvert et à la ressource florale. En effet, une étude a montré que

41
l’effet de la ressource florale varie selon le système de culture (conventionnel ou biologique)
(Holzchuh, 2007).
4.3.2 Le caractère plus ou moins attractif des espèces de fleurs atténue les effets de l’abondance et de la diversité de la ressource florale sur les communautés de pollinisateurs
L’analyse faite sur les effets de l’abondance et de la diversité de la ressource florale
(recouvrement et nombre d’inflorescences) sur l’abondance et la diversité des pollinisateurs
montre qu’il existe une différence d’attractivité des plantes, les pollinisateurs butinant une
fraction restreinte des espèces disponibles. L’attractivité des fleursn’a pas été prise en
considération dans l’étude et une parcelle peut avoir un taux de recouvrement important et ne
pas attirer de pollinisateurs. Cela a été observé au Gros Morne dans une parcelle dans
laquelle a été mise en place une plante de couverture. L’espèce plantée Arachis pintoi
présentait alors un fort recouvrement floral mais l’abondance et la diversité des pollinisateurs
étaient quasi-nulle. Ce phénomène atténue alors peut être l’effet de la ressource florale sur les
pollinisateurs. A L’inverse, sur des parcelles à fort recouvrement à raison d’une seule espèce
en fleur ou presque, de fortes abondances et diversités de pollinisateurs qui se partagent la
ressource ont été observées du fait de la très forte attractivité de l’espèce de fleur présente
(parcelle de M. Poulin à Saint esprit : Tridax procumbens, parcelle de M. Rotsen au
Lamentin : Cleome rutidosperma parcelle de M. Glombart au Robert : Momordica charantia,
Parcelle de M. Baqluet à Saint-Pierre : Euphorbia heterophylla). Ce phénomène peut en partie
expliquer pourquoi, dans certaines situations, malgré de forts indices de diversité des plantes
en fleurs, de faibles abondances de pollinisateurs ont été relevées. Toutes les espèces en fleurs
ne se valent pas. Une relation étroite entre la richesse spécifique des abeilles et la qualité du
nectar des fleurs a d’ailleurs été démontré (Potts et al, 2003) et, selon la composition du
cortège les effets sont donc plus ou moins marqués. Afin de mieux évaluer l’importance de la
ressource florale sur la présence des pollinisateurs dans les vergers, une mesure de
l’attractivité individuelle de l’espèce et de l’attractivité à l’échelle de la communauté
(attractivité d’une espèce par rapport aux autres espèces présentes) fournirait des indications
plus précises quant aux potentielles mesures à mettre en place pour favoriser les pollinisateurs
dans ces milieux.
4.3.3 Une étude sur une échelle de temps plus longue permettrait peut-être de voir l’effet de la richesse spécifique des adventices (en fleurs et non en fleurs) sur les communautés de pollinisateurs.
La richesse spécifique totale des adventices n’a pas d’effet visible sur l’abondance et la
diversité des pollinisateurs au moment où sont effectués les relevés. Parmi les plantes
recensées, peu sont en fleurs et toutes ne sont pas attractives. L’effet des plantes en

42
fleursaurait un effet plus visible dans l’immédiat que la richesse spécifique totale qui elle,
agirait sur un temps plus long. L’effet porterait alors plutôt sur la stabilité de l’activité de
butinage des pollinisateurs dans le temps cartoutes les adventices n’ont pas une floraison
synchrone. La succession des floraisons des différentes espèces dans le temps assurerait
alors le maintien et la redondance de la ressource florale disponible pour les pollinisateurs.
Ceci est d’autant plus vérifié en milieu tropical car les saisons ne sont pas aussi marquées
qu’en milieu tempéré et il y a un étalement de la floraison sur toute l’année.
4.3.4 Toutes les espèces en fleurs n’ont pas la même attractivité pour les pollinisateurs
Certaines espèces de fleurs sont plus butinées que d’autres dans les couverts Les captures à vue démontrent que toutes les espèces végétales n’ont pas une attractivité
équivalente pour les pollinisateurs. Sur toutes les fleurs butinées, cinq espèces se distinguent
et sont plus butinées que les autres : Tridax procumbens, Euphorbia heterophylla, Cleome
rutidosperma, Oxalis barrelieri et Momordica charantia. Parmi elles, certaines ont parfois
peut-être causé le biais dans l’étude de l’effet de la ressource florale sur les pollinisateurs
(§4.3.2). Les espèces les plus attractives appartiennent toutes à des familles différentes et
offrent peut être des ressources complémentaires. Conformément à des observations réalisées
dans d’autres études, les Asteraceae sont les plus attractivesmais sont aussi les plus
diversifiées en espèces butinées que ce soit par les abeilles ou par les syrphes. Certaines
familles de plantes regroupent différentes espèces attractives. C’est le cas des Fabaceae, et
Convolvulaceae dont différentes espèces ont attiré les pollinisateurs. En revanche, bien que
les Capparidaceae et les Oxalidaces ont aussi attiré les pollinisateurs, elles le doivent chacune
qu’à une seule espèce. Les différentes familles de fleurs les plus butinées regroupent des
morphologies, des tailles et des couleurs de fleurs variées et permettent alors de satisfaire
divers pollinisateurs. Les ressources nutritives des différentes familles sont peut-être
complémentaires et assurent la santé des pollinisateurs (ITSAP). Le nombre de visites par
espèce et par famille de fleur confond les syrphes et les abeilles sauvages, or leur régime et
comportement alimentaire sont différentsEn effet, les syrphes butinent par exemple des
plantes anémogames telles que les Poaceae et les Cyperaceaequi ne sont pas toujours butinées
par les Apoïdes par exemple (Le Feon ; 2010).
Il est difficile de dissocier l’effet attractif d’une fleur de l’effet de l’abondance de cette
même fleur
Les espèces les plus butinées étaient aussi souvent les plus abondantes. Il est donc difficile de
dissocier l’attractivité de l’espèce en fleur de l’abondance de cette ressource. En effet, une
espèce de fleurs peu butinée dans des couverts diversifiées, peut devenir très attractive si c’est
l’espèce en fleur prépondérante dans une parcelle. Cela a été observé par exemple pour

43
l’Asteraceae Vernonia cinerea et pour la Curcubitaceae Momrordica charantia, très
communes dans les vergers. Les syrphes par exemple, butinent les fleurs les plus abondantes,
celles qu’il pourra le plus probablement rencontrer (Branquart & Hemptinne, 2000 ) afin de
réduire le temps et l’énergie consacré à la recherche des ressources florales (Branquart
&Hemptinne, 2000 ; Cowgill et al, 1993a).Il existe donc des préférences selon la disponibilité
des ressources et, des fleurs présentes en grande quantité sont peut-être butinées à défaut de
rien ou de meilleures ressources.
Les différents pollinisateurs utilisent des ressources du couvert plus ou moins diversifiées
Les pollinisateurs, selon l’espèce à laquelle ils appartiennent utilisent une ressource plus ou
moins diversifiée dans la strate herbacée des couverts. L’abeille domestique déjà décrite
comme un pollinisateur généraliste (O’Toole & Raw, 1999) a été observée en activité sur la
plus grande diversité des plantes en fleurs par rapport aux autres espèces d’abeilles et de
syrphes conformément à d’autres études (Reader et al, 2005). Les abeilles sauvages
considérées dans la littérature comme des espèces plus spécialistes montrent des
comportements différents selon les espèces.Le régime alimentaire des Halictidae a peu été
étudié et il est difficile de juger de sa spécificité. Cependant, ce sont des abeilles ayant des
distances de vol réduites du fait de leur petite taille. Si les Halictidaes nichent au sein de
l’exploitation, elles iront préférentiellement chercher la ressource florale à proximité de leur
nid dans la strate herbacée des couverts et butinerait alors une plus importante diversité de
fleurs en comparaison à d’autres (Melissodes martinicensis, Xylocopa fimbriata) pouvant
rechercher leur ressource favorite sur de plus grande distance. L’espèce Xylocopa fimbriata
est connue pour butiner une plus faible diversité de fleurs comme certaines passiflores et
légumineuses parmi quelques autres fleurs. Les syrphes du genre Toxomerus sont associés
aux couverts herbacés des milieux ouverts et butinent donc une importante diversité de fleurs.
4.3.5 Ingénierie et contribution à la mise en place du systèmeagro-écologique
Les fleurs adventices peuvent constituer un levier d’action pour favoriser les pollinisateurs
qui butinent à la fois les fleurs du couvert et celles de la plante cultivée.
Les pollinisateurs dont le régime alimentaire est le moins diversifié dans les couverts (Ornida
obesa, Palpada vinetorum et Exomalopsis similis) ont été observés sur les fleurs de goyavier
lors de l’étude menée en parallèle par la FREDON sur les pollinisateurs du goyavier. Il est
possible que ces espèces butinent préférentiellement les fleurs de goyavier et utilisent les
ressources du couvert en complémentation. L’abeille domestique et les Halictidaes sont
autant retrouvées dans les couverts que sur les fleurs de goyaviers. Il est alors envisageable de
favoriser tous ces pollinisateurs en utilisant les fleurs butinées dans les couvert. En effet, le

44
couvert dans les vergers n’entre pas en concurrence avec la culture fruitière et peut être
valorisé. De plus, en arboriculture fruitière, la flore adventice et donc les fleurs butinées sont
naturellement favorisées par rapport à d’autres systèmes de cultures qui nécessitent un
retournement du sol régulier et dans lesquels le couvert ne peut pas s’installer durablement.
En milieu tropical, les plantes adventices sont abondantes toute l’année et constituent une
forte contrainte pour les agriculteurs qui doivent constamment gérer l’enherbement. La
valorisation du couvert aurait donc un double avantage par une nouvelle gestion des
adventices et par l’attraction des pollinisateurs qui peuvent à terme avoir une incidence sur la
pollinisation de leur culture.
Le Xylocope et l’abeille Centris Versicolor(Commentaire, Dumardon-Martial) sont des
pollinisateurs du goyavier et des agrumes, mais, étant quasi absents dans les couverts, la flore
adventice ne constitue pas un outil utilisable pour les attirer dans les vergers. D’autres leviers
d’action tels que les éléments semi-naturels sont envisageables.
Même si certaines espèces n’ont pas été observées sur les fleurs de goyaviers (notamment les
syrphes du genre Toxomerusou l’espèce Melissodes martinicensis), ilsne sont pas à exclure du
fait des autres services qu’ils peuvent rendre à l’agriculture. Les pollinisateurs des fleurs
sauvages peuvent entretenir le couvert dans les vergers et certains syrphes peuvent assurer le
contrôle biologique des ravageurs au sein de l’exploitation(White et al, 1995).
La diversité et l’abondance des fleurs dans les couvert doit être conservées pour favoriser
les pollinisateurs dans les parcelles cultivées
La construction de la courbe d’accumulation a montré que malgré la sélection des ressources
florales dans les couverts par les pollinisateurs, une richesse spécifique minimale de plantes
en fleursest nécessaire pour que les parcelles présentent la diversité maximale des
pollinisateurs ayant pu être observéeà vue dans cette étude. Cetteanalyse justifie que malgré
les effets moyennement démontrés de la diversité de la ressource florale sur les pollinisateurs,
la diversité des plantes en fleurs structure en partie les communautés de pollinisateurs. Plus le
couvert est diversifié, plus il y aura une redondance et une stabilité de la ressource florale et
plus la diversité des pollinisateurs sera favorisée. De plus, du fait du peu d’espèces de
pollinisateurs recensées, une utilisation fine des espèces butinées par chaque pollinisateur est
possible par la constitution d’assemblages floristiques avec les fleurs les plus attractives au
sein des parcelles ou sur leur pourtour. Il serait aussi judicieux de choisir des espèces dont les
floraisons se succèdent dans le temps pour assurer durablement la disponibilité des ressources
pour les pollinisateurs.

45
A l’échelle du paysage, le maintien des éléments semi-naturels est un levier d’action
Une étude à l’échelle du paysage apporterait sans doute plus d’informations en cohérence
avec d’autres études qui ont montré l’effet positif de la complexité et de l’hétérogénéité du
paysage sur la qualité des services écosystémiques dont la pollinisation. De plus, la ressource
florale et relativement homogène à l’échelle de la parcelle et c’est à une échelle plus large
qu’il faut agir. L’étude préliminaire des éléments constitutifs du périmètre n’a pas apporté de
résultats permettant de qualifier les haies, bordures et parcelles adjacentesde leviers d’action
pour la mise en place du système agro-écologique. Cependant, une étude à montré que la
distance aux éléments semi-naturels n’affecte pas les communautés de pollinisateurs si le
paysage contient une proportion importante de praire semi-naturelle importante(Jaucker et
al,2009). Les différents paramètres tels que les éléments semi-naturels et la ressource florale
doivent alors peut-être être considérés ensemble dans le système. De plus, les éléments semi-
naturels n’ont pas toujours le même effet selon les groupes considérés : syrphes ou apoïdes
(Jauker et al, 2009). Bien que l’étude ne permette pas de mettre en évidence l’effet positif des
éléments semi-naturels, il est acquis qu’ils offrent les ressources nécessaire au maintien des
pollinisateurs telles que la ressource florale et les sites de nidifications. Ils doivent alors être
tout comme les fleurs butinées être conservé au sein et à proximité des exploitations.
Réflexion sur une application des MAE en Martinique
En 2013, lors du verdissement de la PAC, des Mesures Agri-environnementales (MAE) ont
été développées pour notamment favoriser la biodiversité dans les milieux agricoles. Diverses
mesures ont été instaurées pour les pollinisateurs sauvages et domestiques. Ce n’est pas
encore le cas en France mais certaine mesure sont favorables aux pollinisateurs dans le cadre
des MAE, qui favorisent notamment la restauration des habitats favorables,
l’approvisionnement en abondance de ressources le plus souvent florales et limitent des effets
des intrants chimiques (Heidseck & Allier, 2013).
Les mosaïques agricoles en milieu tropical se caractérisent par de petites parcelles du fait de
la superficie disponible et par des milieux très hétérogènes propices à la présence des
pollinisateurs. L’entomofaune dans les îles tropicales est fortement liée à la forêt primaire qui
recouvrait l’île autrefois car, elle n’a pas eu le temps de s’adapter aux milieux ouverts
résultants du changement rapide d’utilisation des terres, pour la mise en place d’une
agriculture plus productive (Touroult, 2011). Cette complexité des milieux et la topographie
se sont imposés aux agriculteurs qui ont adapté leurs exploitations aux contraintes naturelles
du milieu. L’aménagement d’ « éléments fixes du paysages à enjeux de biodiversité » (haies,
bandes fleuries) n’est peut-être pas approprié dans les vergers visités car les ressources
nécessaires aux pollinisateurs associées aux éléments semi-naturels et naturels sont

46
finalementnaturellement présentes et nécessitent d’avantage un entretien plutôt qu’une
restauration comme c’est le cas en Europe par exemple.Cependant, cette observation n’est
pas une généralité et ne s’applique pas à toutes les cultures ni à tous les agriculteurs en
Martinique, mais uniquement aux exploitations visitées pour l’étude. La réflexion au sujet
d’autres cultures, par exemple amènerait à des perspectives différentes.
Les arboriculteurs ayant contribué à l’étude soutiennent tous ou presque une agriculture
durable et la grande majorité n’appliquent pas ou peu d’intrants chimiques (produits
phytosanitaires) dans leur verger. Le désherbage mécanique est le mode de gestion de
l’enherbement le plus fréquemment observé dans les parcelles étudiées. Rencontrer une plus
grande diversité d’arboriculteurs aux modes de gestion et conduites de cultures différents
permettrait d’élargirla réflexion quant aux mesures à mettre en place en Martinique selon les
différents contextes pour favoriser les pollinisateurs dans les vergers.
Conclusion L’objectif de cette mission était dans un premier temps, de caractériser les communautés de
pollinisateurs présentes dans les vergers d’agrumes et de goyaviers en Martinique. Dans un
second temps, il s’agissait d’identifier les leviers d’action utilisables pour la mise en place
d’un système agroécologique associant la production fruitière et l’activité apicole, tout en
favorisant les pollinisateurs sauvages. La ressource florale des couverts des vergers est le
principal levier d’action qui a pu être mis en évidence. En effet, ce stage a permis de
démontrer l’effet positif de l’abondance et de la diversité de la ressource florale sur les
communautés de pollinisateurs. Des espèces de fleurs adventices attractives ont été
identifiées et sont à conserver dans les couverts des parcelles pour favoriser les pollinisateurs
dans les exploitations et les amener à polliniser les fleurs de la plante cultivée. Au vu de ces
résultats, il est possible d’orienter des propositions de gestion vers la conservation de la flore
adventice d’intérêt pour les pollinisateurs dans les couverts plutôt que la mise en place
d’aménagements tels que les bandes fleuries. Il serait en effet plus stratégique d’utiliser le
capital floristique que les parcelles offrent naturellement.
Une étude plus précise à l’échelle de l’exploitation ou du paysage conduirait à identifier les
éléments semi-naturels propices à la présence des pollinisateurs en milieu agricole. Un travail
de cartographie estimant les surfaces de haies, de bosquets et de bordures pourrait ainsi être
intégré à l’étude. Une estimation des surfaces favorables à la nidification des abeilles
sauvages dans le sol complèterait ce travail. De plus, un suivi à l’échelle de l’exploitation sur
un temps plus long permettrait d’étudier l’effet de la gestion de l’enherbement sur les
communautés de pollinisateurs. La gestion des pollinisateurs est une gestion intégrée et doit

47
se faire à plusieurs échelles tout en faisant intervenir divers facteurs qu’ils soient
environnementaux ou liés à la conduite de culture des agriculteurs.
Bibliographie
Albrecht, M., Duelli, P., Müller, C., Kleijn, D., Schmid, D,2007. The Swiss agri- environment scheme enhances polliniators diversity and plant reproductive success in nearby intensively managed farmland, J. Appl. Ecol, 44 : 813-822
Altieri, M.A., 1999. The Ecological role of biodiversity in agroecosystems. Agric.Ecosyst.Environ, 74 :19-31.
Barbier, E,1964. Pollinisation et fructification du clémentinier. Ann Abeilles,7 : 63-80. Bartomeus, I., Montserrat, V., Steffan-Dewenter, I, 2010. Combined effect of impatiens
glandulifera invasion and landscape structure on native plant pollination.J. Ecol,98 : 440-450.
Benton, T.G., Vickery, J.A. Wilson. J.D, 2003. Farmland biodiversity : is habitat heterogeneity the key ? Trends in Ecology and Evolution, 18 :182-188.
Bourgeois, G, 2006. Différentes abeilles butineuses pour la pollinisation du bleuet nain. Séminiare en phytologie, 11p.
Branquart, E., Hemptinne, J.L, 2000. Selectivity in the exploitation of floral ressources by hoverflies (Diptera : Syrphinae). Ecography,23 : 732-742.
Brittain, C., William, N., Kremen, C., Klein, A-M , 2013. Synergistics effects of non-Apis bees and honney bees for pollination services. Pro .R. Soc. B Biol. Sci.280 :1-7.
Bosh, J., Blas, M, 1993. Foraging Behavior and Pollinating Efficiency of Osmia cornuta and Apis mellifera on Almond (Hymenoptera, Megachilidae and Apidae) Appl. Entomol. Zool. 29 : 1-9.
Corbet, S., Willimas, I., Osborne, J, 1992. Bees and the pollination of crops and wilds flowers in the European Community. Apiacta, 2p.
Couplan, F., Lazarin A., Lazarin G, 2014. Abeilles et pollinisation en danger, Pontarlier, Edition du Belvédère, 173pages.
Cowgill, S.E., Wratten, S.D., Sotherton, N.W, 1993a. The selective use of floral ressources by hoverfly Episyrphus balteatus (Diptera : Syrphidae) on farmland. Ann.Appl.Biol.122 :223-231.
Daily, G.C, 1997.Nature’s services.Societal dependance on natural ecosystems.Island Press, Wahsington, 392 pp.
Direction Régionale de l’ENvironnement en Martinique, 2008. Profil environnemental. www.side.developpementdurable.gouv.fr/userfiles/file/P18/Publications/Profil_Environemental_2009.pdf, consulté le 19/03/2015.
Dicks, L.V., Corbet, S.A.& Pywell, R.F. 2002. Compartmentalization in plant-insect flower visitor webs.J. Anim. Ecol. 71 : 32-43.
Dumardon-Martial, E , 2005. L’inventaire faunistique des vergers de goyaves : un premier pas vers une protection intégrée.
Faohouo, F-N., Messi, J., Pauly, A. 2002. L’activité de butinage des Apoïdes sauvages

48
(Hymenoptera Apoïdea) sur les fleurs de maïs à Yaoundé (Cameroun) et reflexion sur la pollinisation des graminnées tropiclales. Biotechnol. Agron. Soc. Environ. 6 : 87-98
FAO, 1996. Sommet mondial d’alimentation, 6.Les leçons de la révolution verte-vers une nouvelle Révolution verte. Document d’information technique. www.fao.org/docrep/003/w2612f/w2612f06.htm consulté le 23/03/2015.
FAO, 2008.L’agriculture pour la biodiversité, Introduction / Introduction /Biodiversité agricole à la FAO. http://www.fao.org/docrep/010/i0112f/i0112f00.htm.
FAO, 2008. L’agriculture pour la biodiversité, Agroécosystèmes, services et personnes, La biodiversité pour la préservation des fonctions des agroécosystèmes.http://www.fao.org/docrep/010/i0112f/i0112f00.htm.
Fenster, C.B., Armbruster, W.S., Wilson, P., Dudash, M.R., Thompson, J.D, 2004. Pollination Syndrome and floral specialization.Annu. Rev. Ecol. Syst. , 35 : 375 – 403.
Fernandes, P., Ludovic, T., Crance, J., Minatchi, S. 2009. Innovations agroécologiques en Martinique : freins et leviers organisationnels, institutionnels techniques et économiques. Innovations Agronomiques, 4 :457-466.
Fluri, P.,Pickhardt, A., Cottier, V., Charriere, J- D. 2001. La pollinisation des plantes à fleurs par les abeilles : Biologie, Ecologie, Economie. Agroscope Liebefelf-Poiseux, Centre de recherche Apicole, 27p.
Fontaine, C., Dajoz, I., Meriguet, J., Loreau, M. 2006. Functional Diversity of Plant- Polliniator Interaction Webs Enhance the Persistance of Plant Communities. PloS Biology, 4 : 0129-0135.
Gallais, N., Salles, J.M., Seetle, J., Vaissière , B.2009. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline, Ecol. Econom. 68 : 810-821
Ghazoul, J. 2006. Floral diversity and the facilitation of pollination.J. Ecol. 94 : 295-304.
Gliessman, S.R., 2007. Agroecology : the ecology of sustainable food systems. CRC Press, Taylor & Francis, 384 p
Goulson, D., Nicholls, E., Botias, C., Rotheray, E. 2018. Bee declines driven by combines stress from parasite, pesticides and lack of flowers. Sciencesmag, 347 : 1-9.
Haenke, S., Scheid, B., Schaefer, M., Tscharntke, T., Cartsen, T. 2009. Increasing syrphid fly diversity and density in sown flower strips within simple vs complex landscape.J. Appl. Ecol. 46 : 1106-1114.
Heidsieck, H., Allier, F., 2013. Proposition pour une prise en compte des insectes pollinisateurs dans les politiques agricoles nationales et européennes. Cahier technique de l’ITSAP (Institut de l’abeille).
Hine, H.M., Hendrix, S. 2005. Bumble Bee (Hymenoptera :Apidae) Diversity and Abundance in Tallgrass Prairies Patches : Effect of local Landscape Floral Ressources. Environ. Entomol.34 : 1477-1484.
Hoehn, P., Tscharntke, T., Tylianakis, J.M., Steffan-Dewenter, I. 2008. Functional group diversity of bee pollinators increase crop yield. Proc. R. Soc. B Biol Sci. 275 : 2283-2291.
Holzschuh, A., Steffan-Dewenter, I., Kleijn, D., Tscharntke, T. 2007. Diversity of flower- visiting bees in cereal fields : effect of farming system, landscape composition and regional contexte.J. Appl. Ecol.44 : 41-49.
Holzchuh, A., Steffan-Dewenter, I., Tscharntke, T. 2.10. How do landscape composition and configuration, organix farming and fallow strip affect the diversity of bees, wasps,and their parasitoids.J. Appl. Ecol.79 : 491-500.

49
Jaeger, P. 1954. Observation de quelques cas d’entomophilie chez les Graminées Tropicales. C.R. Hebd. Séance Acad.Sci. 239p.
Jauker, F., Diekötter, T., Schwarzback, F., Wolters, V. 2009.Pollinators dispersal in an agricultural matrix : opposing responses of wild bees and hoverflies to lansdcape structure and distance from main habitat. Landscape Ecol24 : 547-555.
Kwaiser, K.S., Hendrix, S.D., 2007.Diversity and abundance of bees (Hymenoptera : Apiforme)in native and ruderal grassland og agriculturaly dominated landscape. Agricul.t Ecosys. Environ.124, :200-204
Klein, A-M., Vaissière, B., Cane, J., Steffan6Dewenter, I., Cunningham, S., Kremen, C., Tscharntke, T. 2007. Importance of pollinators in changing landscape for world crop.Pro. R. Soc. B. Biol. Sci.274 : 303-3013
Klein, A.M., Cunnigham, S.A., Bos, M., Steffan-Dewenter, I. 2008. Advance in pollination ecology from tropical plantation crops. Ecology.89 : 935-943.
Krebs, J.R., Wilson, J.D., Bradbury, R.B. & Siriwaerdena, G. M, 1999The second silence spring ? Nature.400 : 6111-612
Kremen, C., Wiliams, N.M., Thorps, R.W. 2002. Crop poliniation from native bees at risk from agricultural intensification. Proc.Natl.Acad.Sci.99 :16812-16816.
Lavigne, C., Lesueur-Jannoyer, M., de Lacroix, S., Chauvet, G., Lavigne, A., Dufeal, D. 2011. De la production fruitière intégrée à la gestion écologique des vergers aux Antilles. Innovation Agronomique, 16 :53-62.
Le Bellec, F., Lavigne, C, 2006. La goyave aux Antilles : intérêt pour la diversification Fruitière[en ligne], 2006, dernière mise à jour en 2008, consulté le 24/03/2015, disponiblesur :http://caribfruits.cirad.fr/production_fruitiere_integree/conduite_de_son_verger/les_cultures/Goyave.
Le Féon, V., 2010. Insectes pollinisateurs dans les paysages agricoles : approche pluri-échelle du rôle des habitats semi-naturels, des pratiques agricoles et des cultures entomophiles. 257p.
Le Roux, X., Barbault, R., Baudry, J., Burel, F., Doussan, I., Garnier, E., Herzog, F., Lavorel, S., Lifran, R., Estrade, J-R., Sarthou, J-P., Trommetter, M. 2008. Agriculture et Biodiversité, Valoriser les synergies, Synthèse du rapport d’expertise.9p.
Leong, J., Throp, R-W. 1999. Colour-coded sampling : the pan trap colour preferences of oligolectic and nonologolectic bees associated with a vernal pool plant.Ecol Entomol.24 : 329-335.
MacArthur, R., Wilson, E.O. 1967. The Theory of Island Biogeography. Princeton University Press. 203p.
Mandelik, Y., Roll, U., 2009. Diversity patterns of wild bees in almond orchards and their surrounding landscape, Isr. J. Plant. Sci. 57 : 185-19
Meurgey, 2014. Diversité des abeilles sauvages en Guadeloupe et leur contribution la flore butinée. (Hymenoptera, Athophila, Apidae et Megachilidae). 63p
Naeem, Shahid, Lindsey, J., Thompson, Sharon, P., Lawler, John, H., Richard, M., Woodfin.1995. Empirical Evidence that Declining Species Diversity May Alter the
Performance of Terrestrial Ecosystems.Proc. R. Soc. B Biol.Sci.347(1321) : 249-262. Njoroge, G.N., Gemmill, B., Bussmann, R., Newton, L.E., Ngumi, V.M. 2010. Diversity
and efficiency of wild pollinators of watermelon (Citrullus, lanatus (Thunb.) Mansf.) at Yatta (Kenya). Journal of Applied Horticulture. 12 : 35-41.

50
Normandin, E., Fournier, V.,Rivard, N. 2014.Les abeilles sauvages : une étude de leur diversité. .Bulletin de conservation du ParcNational des ïles-de-Boucherville. 4p.
O’Toole, C., Raw, A., 1999. Bees of the world.Blandford. New York. 192p. Payette, A., 2013. Contribution à l’inventaire des espèces d’abeilles (Hymenoptera :
Apoidea) dans des cultures de la Canneberge à gros fruits, Vaccinium macrocarpon, de la province du Québec. Rapport d’inventaire pour le MAPAQ. 12 p.
Potts, S.G., Vulliamy, B., Dafni, A., Ne’Eman, G., Willmer, P, 2003. Linking bees and flowers : How do floral communities structure pollinator communities ? Ecology.84 : 2628-2642.
Perez-Balam, J., Quezada-Euàn, J.J.G., Alfaro-Bates, R., Medina, S., McKendrick, L., Soro, A., Paxton, R.J, 2012,The contribution of honney bees,flies ans wasps to avocado (Persea americana) pollination in souther Mexico. J. Poll. Ecol, 8 : 42-47.
Pesson, P., Louveau, J. 1994. Pollinisation et production végétale, INRA, Paris, 663 pp. Rasmont, P., Ebmer, A., Banaszak, J., Van Der Zanden, G. 1995. Hymenoptera Apoidea
Gallica – Liste taxonomique des abeilles de France, de Belgique, de Suisse et du Grand-duché du Luxembourg.Bul.l Soc. Entomol. Fr.100 : 1-98
Reader, T., MacLeod I., T.Elliott, P., J.Robinson, O., Manical, A. 2005. Inter-Order Interaction Flower-Visisting Insect : Foraging bees Avoid Flowers Previously Visisted by Hoverflies. J. Insect. Behav.18 : 51-57.
Roubick, D-W. 1989. Ecology and Natural History of tropical bees. University Press Cambridge. 514 p.
Rollin, O. 2013. Etude multi-échelle du patron de diversité des abeilles et utilisation des ressources fleuries dans un agrosystème intensif.184p
Roubick, D.W.1995. Polliniation of cultivated plants in the tropics. Foof & Agriculture org, Rome. 196p.
Roulston, T’ai H., Smith S.A., Brewster, A.L. 2007. A comparison of Pan Trap and Intensive Net Sampling Techniques for Documenting a Bee (Hymenoptera : Apiforme) Fauna. Journal of the Kansas Entomological Society 80 : 179-181.
Sarthou, J-P., Speight M.C.D. 2005.Les Diptères Syrphidés, peuples de tous les espaces. Insectes. 137 : 3-8.
Sastre, C., Breuil, A. 2007. Eléments de géographie antillaise. Dans : Biotope, (Ed.) : Plantes, Milieux et Paysages des Antilles françaises. Ecologie, biologie, identification, protection et usages. Mèze, (Collection Parthénope), 22-31.
Sommagio, D. 1999. Syrphidae/can they be used as environmental bioindicators ? Agricult.Ecosys. Environ. 74 : 343-356
Steffan-Dewenter, I. 2002. Landscape context affects trap nesting bees, waps and their natural Enemies. Eco.l Entomol. 27 : 631-637.
Summer, D-A., Boriss, H. 2006.Bee-conomics and the Leap in Pollination Fees. Giannini Foundation of Agricultural Economics.9-11.
Tscharntke, T., Klein, A.M., Steffan-Dewenter, I., Thies, C. 2005. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service managment. Ecol. Lett.8 : 857-874.
Thies,C., Tscharntke, T. 1999. Landscape Structure Biological Control in Agroecosystems. Sciences285 : 893-895
Tilman, D., Knops, J., Wedin, D. 1997. The Influence of Functional Diversity and composition on Ecosystems processes. Sciences277 : 1300-1302.
Thompson, FC. 1981. The flower flies of West Indies (Diptera:Syrphidae). Mem EntomolSoc Wash 9:1-20

51
Touroult, J. Insectes des Antilles et de la Gyanne : La forêt avant tout. Insectes 162 : 43-46 Vaissière B. 2005. Abeilles et Pollinisation. Académie d’Agriculture de France, 4p. Villemant, C., 2005. Les nids d’abeilles solitaires et sociales, Revue insectes 137 :13-17 Walker, Brian, H. 1992. Biodiversity and Ecological Redundancy.Conserv. Biol.6 :18-23. Westphal, C., Bommarco, R., Carré, G., Lamborn, E., Morison, N., Petanidou, T.,
Potts,S.G., Roberts, S.P.M., Szentgyörgyi, H., Tscheulin, T., Vaissière, B., Woyciecowski., Biesmeijer, J.C., Kunning, W.E., Settele, J., Steffan-Dewenter, I. 2008. Measuring Bee Diversity in Different European Habitat and Biogeographical Regions. Ecol. Mono.78 : 653-671.
William, I-H. 1994. The dependence of crop polliniation within the European Union on polliniation by honey bees.Agricut. Zool. Rev.6 :229-257. Winfree, R., Williams, N-M., Dushoff, J., Kremen, C.2007. Native bees provide insurance
against on going honney bee losses ecology letter 10 : 1105-1113. Winfree, R., Kremen, C. 2009. Are ecosystem services stabilized by differences among
species ?A test using crop pollinsation.Proc. R. Soc. B Biol. Sci.276 : 229-237 Winston, M. 1993. La biologie de l’abeille, Nauweleart et Frisson-Roche, Beauvechain et
Paris. 276 pp. White, A-J., Wratten, S., A.Berry,N., Weigmanni,U. 1195. Habitat Manipulation to
enhance Biological Control of Brassica Pest by Hover Flies (Diptera :Syrphidae). J. Econ. Entomol.88 : 1171-1176
Wolfgang, W., Imperatriz-Fonsesca,V-L., Engels, W. 1996. Ressource partitionning between Highly Eusocial Bees and Possible Impact of the Introduced Africanized Honney Bee on Native Stingless Bees in the Brazilian Atlantic Rainforest. Stud Neotrop Fauna & Environm. 31 : 137-151.
52
ANNEXES
Annexe 1 Abondance des différents groupes d’insectes capturés au fauchage
Groupes Abondance Groupes Abondance Achilidae 1 Heteroptera 1 Agromyzidae 30 Icheumonidae 2 Agronidae 1 Issidae 2 Airaignée 1 Lampyridae 5 Anthocoridae 1 lauxaniidae 3 Aphididae 13 Lepidoptera 55 Araignée 74 Lonchaeidae 2 Berytidae 18 Lygaelidae 54 Braconidae 26 Michichiidae 1 Bupestidae 5 Micropezidae 4 Caelifera 20 Milichiidae 6 Calliphoridae 3 Miridae 78 Calyptratae 1 Muscoidae 9 Chalcididae 17 Myrmycinae 1 Chalcidoidae 39 Nematocera 21 Chloropidae 74 Odonata 1 Chrisopidae 1 Orthoptera 3 Chrysomelidae 20 Otitidae 43 Chrysopidae 4 Pentatomidae 12 Cicadellidae 93 Phoridae 8 Cixiidae 8 Pipunculidae 4 Coccinellidae 30 Rhopalidae 5 Coreidae 11 Sarcophagidae 3 Curculionidae 8 Scelionidae 1 Delphacidae 26 Scoliidae 2 Diptera (ND) 32 Sespidae 11 Dolichopodidae 40 Sphecidae 1 Drosophilidae 5 Stratomydae 1 Dryinidae 3 Tachinoidae 55 Ensifera 6 Tenebrionidae 5 Ephydridae 33 Tephritidae 19 Figitidae 3 Tettigoniidae 28 Formicidae 72 Thysanoptera 1 Hemiptera 2 Tiphiidae 5 Hemiptera (ND) 14 Vespidae 2
Abondance totale : 1206 individus

53
Annexe 2 Abondance des différents groupes d’insects capturés à l’aide des « bols colorés »
Abondance Groupe Abondance
Agromyzidae 41 Issidae 9
Anthocoridae 4 Lampyridae 7
Aphidiidae 21 Lauxaniidae 4
Araignée 20 Lepidoptera 44
Asteiidae 1 Lygaelidae 15
Blattoptera 2 Membracidae 1
Bostrichidae 1 Micropezidae 19
Braconidae 7 Milichiidae 16
Caelifera 1 Miridae 16
Calliphoridae 1 Muscoidae 13
Carabidae 2 Nematocera 52
Carnidiae 6 Nitidulidae 2
Chalcididae 10 Oedenidae 1
Chalcidoidae 44 Otitidae 24
Chloropidae 65 Pentatomidae 4
Chrysomelidae 9 Phoridae 34
Chrysopidae 1 Pipunculidae 1
Cicadellidae 51 Pompilidae 1
Cixiidae 27 Psocoptera 8
Coccinellidae 11 Rhopalidae 1
ColeopteraND 3 Scarabeidae 3
Curculionidae 20 Sespidae 3
Cynipidae 5 Sphaeroceridae 3
Delphacidae 17 Sphecidae 7
Diptera ND 38 Staphyliridae 4
Dolichopodidae 70 Stratyomidae 6
Drosophilidae 86 Tachinoidae 77
Elateridae 1 Tenebrionidae 23
Empididae 5 Tephritidae 4
Ephydridae 34 Termitidae 2
Formicidae 109 Tettigoniidae 9
Grallipeza 1 Thysanoptera 16
Hemiptera ND 4 Tingidae 6
Heteroptera ND 6 Tiphiidae 1
Hymenoptera ND 5 Tropiduchidae 1 Abondance totale: 1178

54
Annexe 3 : Extrait de ka collection entomologique de la FREDON.
55
Annexe 4 : Planning d’une journée de terrain pour la mise en place du dispositif 1
Planning d’une journée de terrain : Dispositif 1
Le dispositif 1 peut être mis en place le lundi, mardi et jeudi (journées complètes). Le dispositif 1 est mis en place sur 20 parcelles et nécessite donc 20 journées complètes de terrain (à partir de mi-avril jusqu’à fin juin). Le mercredi et le vendredi sont consacrés au tri et à l’identification des échantillons ainsi qu’à la saisie et la mise en forme des données.
Horloge Taches
7h00 Arrivée sur la parcelle + analyse de la parelle et mise en place des 3
transects
8h00 Premier fauchage sur les 3 transects
8h30 à 9h00 Chasse à vue sur le transect 1
9h00 à 9h30 Chasse à vue sur le transect 2
9h30 à 10h00 Chasse à vue sur le transect 3
10h à 10h30 Moment de battement (photos + dessins de la parcelle)
10h30 à 11h00 Mesure du périmètre de la parcelle et des éléments constitutifs de
l’environnement proche
11h00 Deuxième fauchage sur les 3 transects
11h30 à 12h00 Petite pause
12h00 à 15h00 Relevés floristiques qualitatifs, recouvrement par espèce des plantes en
fleurs, abondance fleurs ouvertes
15h00 Troisième fauchage sur les 3 transects
15h30 Départ de la parcelle
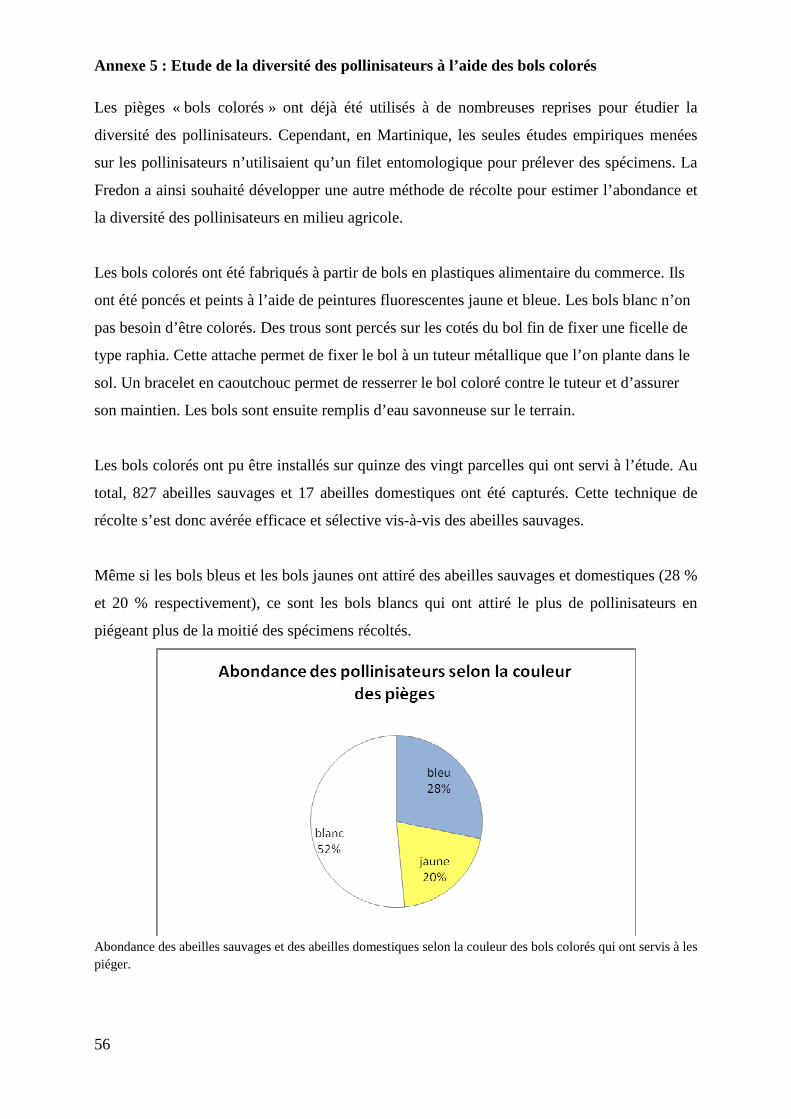
56
Annexe 5 : Etude de la diversité des pollinisateurs à l’aide des bols colorés Les pièges « bols colorés » ont déjà été utilisés à de nombreuses reprises pour étudier la
diversité des pollinisateurs. Cependant, en Martinique, les seules études empiriques menées
sur les pollinisateurs n’utilisaient qu’un filet entomologique pour prélever des spécimens. La
Fredon a ainsi souhaité développer une autre méthode de récolte pour estimer l’abondance et
la diversité des pollinisateurs en milieu agricole.
Les bols colorés ont été fabriqués à partir de bols en plastiques alimentaire du commerce. Ils
ont été poncés et peints à l’aide de peintures fluorescentes jaune et bleue. Les bols blanc n’on
pas besoin d’être colorés. Des trous sont percés sur les cotés du bol fin de fixer une ficelle de
type raphia. Cette attache permet de fixer le bol à un tuteur métallique que l’on plante dans le
sol. Un bracelet en caoutchouc permet de resserrer le bol coloré contre le tuteur et d’assurer
son maintien. Les bols sont ensuite remplis d’eau savonneuse sur le terrain.
Les bols colorés ont pu être installés sur quinze des vingt parcelles qui ont servi à l’étude. Au
total, 827 abeilles sauvages et 17 abeilles domestiques ont été capturés. Cette technique de
récolte s’est donc avérée efficace et sélective vis-à-vis des abeilles sauvages.
Même si les bols bleus et les bols jaunes ont attiré des abeilles sauvages et domestiques (28 %
et 20 % respectivement), ce sont les bols blancs qui ont attiré le plus de pollinisateurs en
piégeant plus de la moitié des spécimens récoltés.
Abondance des abeilles sauvages et des abeilles domestiques selon la couleur des bols colorés qui ont servis à les piéger.
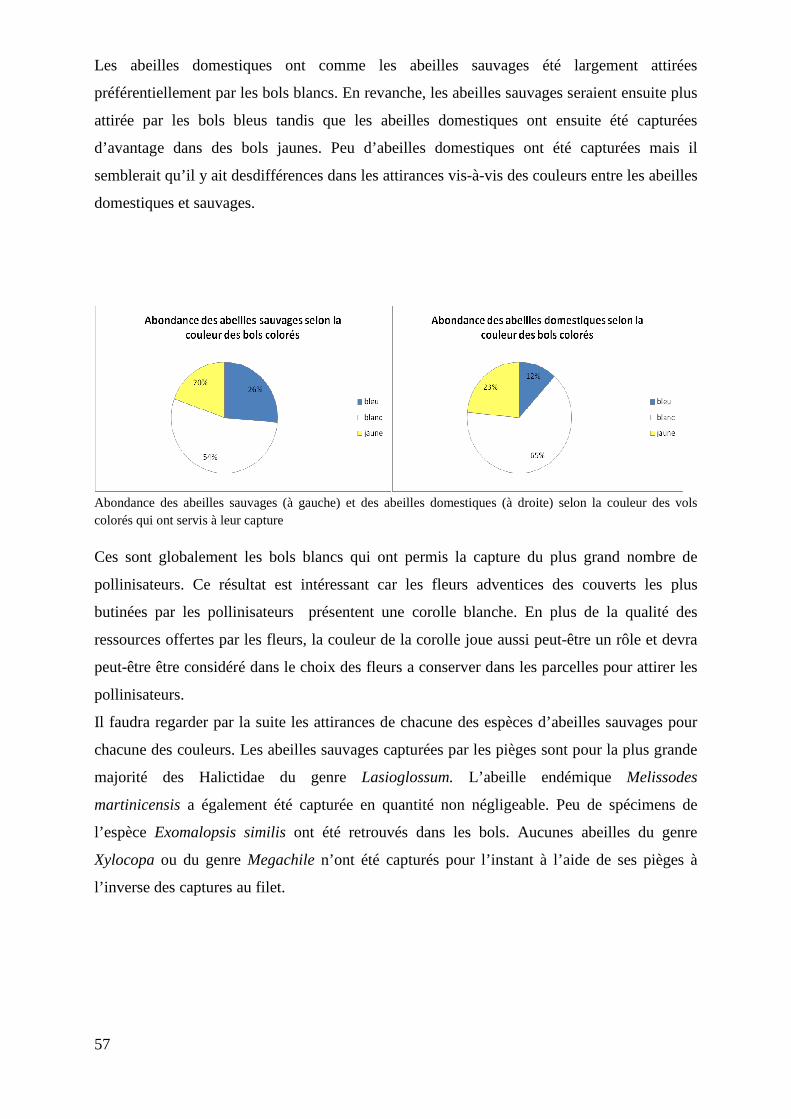
57
Les abeilles domestiques ont comme les abeilles sauvages été largement attirées
préférentiellement par les bols blancs. En revanche, les abeilles sauvages seraient ensuite plus
attirée par les bols bleus tandis que les abeilles domestiques ont ensuite été capturées
d’avantage dans des bols jaunes. Peu d’abeilles domestiques ont été capturées mais il
semblerait qu’il y ait desdifférences dans les attirances vis-à-vis des couleurs entre les abeilles
domestiques et sauvages.
Abondance des abeilles sauvages (à gauche) et des abeilles domestiques (à droite) selon la couleur des vols colorés qui ont servis à leur capture
Ces sont globalement les bols blancs qui ont permis la capture du plus grand nombre de
pollinisateurs. Ce résultat est intéressant car les fleurs adventices des couverts les plus
butinées par les pollinisateurs présentent une corolle blanche. En plus de la qualité des
ressources offertes par les fleurs, la couleur de la corolle joue aussi peut-être un rôle et devra
peut-être être considéré dans le choix des fleurs a conserver dans les parcelles pour attirer les
pollinisateurs.
Il faudra regarder par la suite les attirances de chacune des espèces d’abeilles sauvages pour
chacune des couleurs. Les abeilles sauvages capturées par les pièges sont pour la plus grande
majorité des Halictidae du genre Lasioglossum. L’abeille endémique Melissodes
martinicensis a également été capturée en quantité non négligeable. Peu de spécimens de
l’espèce Exomalopsis similis ont été retrouvés dans les bols. Aucunes abeilles du genre
Xylocopa ou du genre Megachile n’ont été capturés pour l’instant à l’aide de ses pièges à
l’inverse des captures au filet.