Embed Size (px)
Citation preview

LABORATOIRE DE BIOLOGIE DU DEVELOPPEMENTET DE LA DIFFERENCIATION
Option : Cancer & environnement
Par
Mademoiselle YAHIA RADIA
Sujet du mémoire :
Recherche des marqueurs sérologiques du Virus Epstein-Barr (EBV)
chez des femmes ayant un cancer du sein et issues de l’Ouest Algérien
Soutenu le :
Devant le jury composé de :
Président : Professeur F.Z. EL KEBIR Université d’Oran
Encadreur : Professeur T. SAHAOUI Université d’Oran
Co-encadreur : Docteur D.K.DERKAOUI Université d’Oran
Examinateur : Professeur A. AOUES Université d’Oran
Examinateur : Professeur H.BOUCHERIT Université d’Oran
Invité d’honneur : Docteur H. MOULAY Université d’Oran
2010-2011
Faculté des Sciences
Département de Biologie
Mémoire présenté en vue de l’obtention du diplôme de magister en biologie

DEDICACES
A mes chers parents, qui m’ont toujours soutenue durant mon parcours
scolaire. Merci pour votre confiance et votre affection. Voici le résultat de tous vos efforts.
A mom frère Lotfi et sa femme Sarah, à ma sœur Myriame et son mari Farouk
à ma petite jolie sœur Karima, qui m’ont toujours encouragée. Je suis heureuse et fière de
vous avoir à mes côtés.
A mon fiancé Amine pour ton soutien durant mon mémoire avec ses moments
difficiles, ses hauts et ses bas, pour ta présence auprès de moi.
A mes futurs beaux‐parents, Merci pour votre gentillesse.
A mon joli cœur princesse Lyna Syrine qui m’a enchantée et égaillée depuis sa
naissance.
A ma très chère cousine Anissa que je n’oublierai jamais.
A toutes mes amies de longue date, Amina, Fouzia et Assia.
Aux familles Daoudi, Yahia, Merzouk, Boussiala, Dehmani.

REMERCIEMENTS
Je remercie dieu le tout puissant qui m’a donné la volonté, le courage d’avoir achevé ce
travail.
Mes remerciements les plus profonds vont au PR ELKEBIR FATIMA ZOHRA qui
m’a accueillie chaleureusement dans son laboratoire me permettant ainsi d’accéder
directement à la concrétisation de ce travail
Je tiens à la remercier également pour sa grandeur d’âme et son ouverture d’esprit, pour
sa patience ainsi que pour le grand intérêt qu’elle a consacré à la lecture de ce manuscrit.
Je tiendrai compte de ses précieuses critiques scientifiques qui ne font qu’améliorer ce
travail.
Je remercie vivement le PR BOUCHERIT ELHASSEN d’avoir examiné ce mémoire
en consacrant un temps précieux. Ses remarques scientifiques ainsi que ses critiques m’ont
beaucoup servies à l’amélioration de ce travail.
Je le remercie de m’avoir permis l’accès au service et de m’avoir autorisée d’obtenir les
prélèvements biologiques nécessaires sans lesquels ce travail n’aurait pas eu lieu.
Merci professeur pour votre bonne humeur.
Je remercie Mr LE PR AOUAS ABK d’avoir accepté d’être examinateur de mon
mémoire. Encore merci pour sa disponibilité, sa patience et ses critiques .C’est pour moi un
honneur et un privilège de vous voir parmi mon jury. Je vous exprime ma plus grande
gratitude et mon plus profond respect.
Un grand merci au Dr DERKAOUI DALIA, merci de m’avoir co-encadrée. Je vous
remercie d’avoir été patiente à mon égard .D’avoir su me guider dans ce travail, d’avoir été
présente chaque fois que j’en avais besoin. Ce fut très instructif aussi bien scientifiquement
que personnellement. Ce fut un plaisir de travailler à vos côtés.

Je remercie le PR SAHRAOUI TEWFIK pour ses précieuses connaissances
scientifiques pour sa gentillesse et sa bonne humeur ainsi que pour sa patience et ses conseils
qui n’ont fait qu’apporter un précieux plus à mon travail. Enfin, je le remercie pour sa
disponibilité dans les grands moments de mon cursus.
Je tiens à remercier vivement le Dr MELOULI HAMID pour m’avoir accueillie
chaleureusement dans son laboratoire (institut Pasteur à Alger). Ce travail n’aurait pas pu
se faire sans son aide précieuse, sa patience, sa gentillesse, ses compétences et son
professionnalisme. Ses encouragements, associés aux moyens matériels dont je me suis servie,
m’ont permis de mener à bien la recherche présentée dans ce mémoire. C’est pourquoi je lui
exprime ma plus grande gratitude.
Je remercie vivement le Dr MOULAY pour son aide précieuse, sa gentillesse et ses
conseils.
Je tiens à remercier l’ingénieur FOUZIA de l’institut Pasteur. J’ai beaucoup appris
à ses côtés, elle m’a transmis sa passion et sa volonté, merci pour les nombreuses discussions
que nous avons eues à la paillasse, pour tes conseils techniques, ton ouverture d’esprit. Merci
de m’avoir aidée, ses conseils ont été très précieux.
Je remercie Pr SENHADJI RACHID pour sa disponibilité et son encouragement.
Je tiens personnellement à remercier Dr FARIDA MESLI, une autre personne qui a
beaucoup compté durant mon mémoire. Je vous remercie pour votre confiance et votre grande
ouverture d’esprit.
Je remercie les membres permanents de l’hôpital universitaire d’Oran, pour leur
disponibilité, leur soutien, leur professionnalisme, leur extrême gentillesse. Les conversations
que nous avons eues, furent extrêmement utiles ainsi qu’un plaisir.

Je présente enfin mes amitiés à tous les étudiants qui ont travaillé dans le laboratoire
du développement et de la différenciation, durant cette période, merci au Dr Zaoui
Chahinez (2ème année), que j’ai eu le plaisir de connaitre. A toute la promotion cancer et
environnement à mes grandes sœurs Hayette, Amina, Souhila, Soria, Houria à mon ami
Youcef et un très grand merci à Fatiha. Ils ont fait de ce mémoire un travail très
intéressant.
A tous et à ceux que j’ai oubliés, merci pour les belles années passées avec vous.
Grand merci à tous

RÉSUMÉ
Le virus d’Epstein-Barr (EBV) est un virus oncogène ubiquitaire, associé à différents
types de carcinomes tels que le carcinome du nasopharynx et le carcinome gastrique. Des
études ont rapporté la présence du génome de l’EBV dans les cellules tumorales de cancer du
sein (Labrecque et al, 1995), principalement dans les types infiltrants (Bonnet et al, 1999).
Ceci suggère un rôle potentiel de l’EBV dans la progression tumorale (Arbach et al, 2006).
Afin de mieux cerner l’effet de l’EBV dans les carcinomes mammaires. Nous avons essayé
d’apporter un éclaircissement quant à la présence des marqueurs sérologiques tumoraux
impliqués dans le cancer du sein, notre étude a portait sur 24 patientes toutes atteintes de
cancer du sein et issues de l’Ouest Algérien. Les résultats obtenus a l’issu de l’étude
histologique font ressortir une atteinte prédominante du sein gauche et type histologique le
plus répandu dans notre étude est le CCI. La partie la plus importante de ce mémoire c’est
concentre sur l’étude sérologique. Trois techniques différentes ont été nécessaires à la
réalisation de cette partie, toutes exprimaient les mêmes résultats .L’association entre le
cancer du sein et le virus Epstein-Barr reste toujours inconnue.
Mots clés : cancer mammaire, virus Epstein-Barr, marqueurs sérologiques.

Abstract
The Epstein-Barr virus (EBV) is a ubiquitous virus oncogene, associated with different
types of carcinomas such as nasopharyngeal carcinoma and gastric carcinoma. Studies
have reported the presence of EBV genome in tumor cells of breast cancer (Labrecque et
al., 1995), mainly in the infiltrating type (Bonnet et al., 1999). This suggests a potential
role of EBV in tumor progression (Arbach et al., 2006). To better understand the effect of
EBV in breast carcinomas. We tried to make a clarification regarding the presence of
serological markers involved in tumor breast cancer, our study included 24 patients all
suffering from breast cancer from western Algeria. At the end the results of histological
study show an impairment of the left breast and the most common histological type in our
study is the ICC. The most important part of this thesis is focused on the serological study.
Three different techniques were needed to carry out this part, all expressed the same
results. The association between breast cancer and the Epstein-Barr virus remains
unknown.
Keywords
Breast cancer, Epstein-Barr virus, serological markers.

Sommaire
Liste des abréviations
Liste des figures
Liste des tableaux
Résumé
Abstract
Introduction……..……………………………………………….....................................1
Chapitre I : Anatomie, histologie et physiologie du sein
1. Anatomie structure de la glande mammaire…………………………………..……2
2. Histologie …………………………………………………………………….…..........3
3. origine embryonnaire du tissu mammaire
4. Vascularisation
4.1. La vascularisation sanguine
4.2. La vascularisation lymphatique…………………………………….…...........4
5. Innervation
6. La physiologie de la glande mammaire
6.1. Facteurs hormonaux………………………………………………….……………5
6.1.1. Les estrogènes
6.1.2. La progestérone
6.1.3. La prolactine
6.1.4. Autres hormones…………………………………………………………….6
6.2. Facteurs de croissance
7. Développement de la glande mammaire…………………………………………....7

Chapitre II : Anatomie pathologique du sein
1. Les pathologies bénignes de la glande mammaire……………………………........9
1.1. L’adénofibrome
1.1.1. Adénofibrome-unique
1.1.2. L’adénofibromatose
1.1.3. L’adénofibrome juvénile………………………...………………………………10
1.2. Le kyste rétro-aréolaire de l’adolescente
1.3. L’ectasie galactophorique
1.4. Les abcès du sein………………………………………………………………………11
1.5. La mastopathie fibrokystique
1.6. La papillomatose juvénile……………………………………………………………...12
1.7. La tumeur phyllode
2. Pathologies malignes du sein…………………………...……………………………......13
2.1. Les carcinomes mammaires ou épithéliomas
2.1.1. Les carcinomes lobulaires
2.1.2. Les carcinomes canalaires
2.2. Les sarcomes mammaires………………………………………………………………14
3. Mécanisme de carcinogenèse mammaire………………………………………………..15
3.1 Initiation de la progression tumorale………………………………………….....16
3.2. Promotion tumorale

3.3. Métastases des carcinomes mammaires………………………………………...17
4. Classification des tumeurs mammaires…………………………………………………19
4.1. Classifications histologiques des carcinomes infiltrants
4.1.1 Classification de l’OMS (1981)
4.1.2. Classification de l’OMS modifiée (2002-2003)
4.2. Classification TNM……………………………………………………………...20
4.3. Classification SBR…………………………………………………………….....23
Chapitre III : Epidémiologie du cancer du sein
1. Epidémiologie du cancer du sein……………………………………………………......24
2. Epidémiologie du cancer du sein en Algérie…………………………………….......26
3. Etude épidémiologique du cancer du sein à Oran
4. Aspect histologique du cancer du sein………………………………………………......27
5. Facteurs de risques de cancer du sein…………………………………………………...29
5.1. Facteurs génétiques
5.2. Facteurs environnementaux…………………………………………………......30
5.3. Facteurs hormonaux
5.3.1. Hormones endogènes………………………………………………………..31
5.3.2. Hormones exogènes……………………………………………………….....32
5.4. Facteurs pronostiques et survie…………………………………………….........33
Chapitre IV : Virus et cancer du sein1. Virus et cancer du sein…………………………………………………………………...34
1.1. MMTV (Mouse Mammary Tumor Virus)

1.2. HPV (Les papillomavirus humains)
1.3. EBV (Epstein-Barr)…………………………………………………………………….35
1.3.1. La famille des herpesviridae et le virus d’Epstein-Barr
1.3.1.1. La famille des herpesviridae
1.3.1.2 Historique…………………………………………………………………………36
1.3.1.3. Structure du virus………………………………………………………………...37
1.3.1.4. Génome viral de l’EBV
1.3.1.5. Polymorphisme…………………………………………………………………....39
1.3.1.6. Les voies de transmission
1.4. Tropisme cellulaire
1 .4.1 In vivo
1.4.2. In vitro……………………………………………………………………………40
1.5.. Cycle biologique du virus d’Epstein-Barr……………………………………...........41
1.5.1. Entrée du virus dans les cellules cibles
1.5.2. Latence et maintien du génome………………………………………………...........42
1.5.2.1. Les différents types de latences virales
1.5.2.2. Les protéines de latence : description, régulation et fonction des protéines
de latence…………………………………………………………………………………….43
A. Les promoteurs des protéines de latence
1.5.2.3. La famille des Antigènes Nucléaires de l’EBV, les protéines EBNAs….....44
A .EBNA-LP
B. EBNA2……………………………………………………………………………...45
C. EBNA-3 (-3A, -3B et -3C)

D. EBNA1……………………………………………………………………………...47
1.5.2.4. Les protéines membranaires de latence………………………………….....48
A. LPM-1 (protéine membranaire de latence-1)
B. LMP-2 (protéine membranaire de latence-2)
1.5.2.5. Les transcrits de la région BamHI A (BART, BamHI A Rightward
Transcripts)………………………………………………………………………………….51
1.5.3. L’infection lytique…………………………………………………………………….52
1.5.4. Réactivation……………………………………………………………………….......53
1.6. Pathologies associées au virus d’Epstein-Barr
1.6.1. Infection primaire et pathologies
A. La mononucléose infectieuse
1.6.2. Infection persistante et pathologies tumorales
1.6.2.1. Pathologies de l’hôte immunodéprimé……………………………………...55
1. Désordres Lymphoprolifératifs Post-Transplantation (PTLD)
2. Lymphomes associés à l’EBV chez le malade du SIDA……………………….....57
A. Léiomyosarcomes liés à l’Immunodéficience……………………………........58
B. Leucoplasie orale chevelue
1.6.2.2. Pathologies de l’hôte immunocompétent…………………………………………59
1. Lymphome de Burkitt (LB)
2. Le lymphome de Hodgkin………………………………………………………….61
3. Le carcinome du rhinopharynx
4 .Maladies associées à l’EBV dans les cibles cellulaires non-conventionnelles…...63
A. Carcinome gastrique et carcinome de type lymphoépithéliome……………..64
B. Maladies lymphoprolifératives à cellules T

C. Pseudotumeurs inflammatoires de type Tumeurs des cellules dendritiquesfolliculaires (CDF)…………………………………………………………………...65
D. Le cancer du sein………………………………………………………………..66
Premières études……………………………………………………………..67
1. Etudes sérologiques
2. Etudes de la tumeur
II. Matériels et méthodes
1. Population étudiée ………………………………………………………………………..68
2. Prélèvements
2.1. Prélèvement de Sérums
2.2. Prélèvement de biopsie
3. Etude sérologique
3.1. La technique Elisa (enzyme-lynked immunoessay)………………………………….69
3.1.1. Le principe de la technique
3.1.2. Protocol expérimental
3.2. La technique IFI (immunofluorescence indirecte)…………………………………...70
3.2.1. Le principe de la technique
3.2.2. Protocol expérimental
3.3. La technique de Western Blot
3.3.1. Le principe de la technique
3.3.2. Protocol expérimental………………………………………………………………...71
4. Etude histologique du tissu tumorale
III. Résultats de l’étude histologique
1. Résultats de l’étude histologique………………………………………………………..74

1.1. La localisation anatomique
1.2. La contraception
1.3. L’âge des patientes……………………………………………………………………...75
1.4. Les carcinomes cibles
1.5. Le grade…………………………………………………………………………………76
2. Résultats de l’étude sérologique
IV- Discussion et conclusion et perspective
-Discussion et conclusion……………………………………………………………………82
-Perspectives…………………………………………………………………………………86
- Annexe
- Références

ABREVIATIONSADNc : Acide désoxyribonucléique complémentaire
ARNm : Acide ribonucléique messager
BZLF1 : Z Epstein Barr Virus Replication Activator
CD : Cluster de Différentiation
CMH : Complexe Majeur d’Histocompatibilité
CMV : Cytomégalovirus
CAEBV: Chronic Active Epstein-Barr Virus infection
EA: Early Antigen
EBER : Epstein-Barr Encoded RNA
EBV: Epstein-Barr virus
EBNA : Epstein-Barr Nuclear Antigen
EBV : Epstein-Barr Virus
EGF: Epithelial Growth Factor
ELISA: Enzyme Linking Immunosorbent Assay
EMMP: Extracellular Matrix Metalloproteinase
FGF: Fibroblast Growth Factor
GAPDH : Glycéraldéhyde 3-Phosphate Déshydrogénase
GFP: Green Fluorescent Protein
gp : Glycoprotéine
HSV1 : Herpes Simplex Virus de types 1
HIS: Hybridation In Situ
HPV: Human PapillomaVirus
Ig: Immunogobuline
IHC: ImmunoHistoChimie

IGF: Insulin Growth Factor
IB: Inhibitor of NF B
IARC: International Association for Research on Cancer
IFI : immunofluorescent assays
IR : Internal Repeat
KDR : Kinase insert domain-containing receptor
LB : Lymphocytes B
LCL : Lignées Cellulaires Lymphoblastoïdes
LMP : Latent Membrane Protein
LT : Lymphocytes T
MEC : Matrice extracellulaire
MH : Maladie de Hodgkin
miARN : microARN
MMTV: Mouse Mammary Tumor Virus
MNI: Mononucléose infectieuse
NPC : Carcinome du nasopharynx
ORF : Cadre de lecture ouvert
Ori lyt : Origine de réplication lytique
Ori P : Origine de réplication latente
PBS: Phosphate Buffer Saline
PCR: Polymérase Chain Reaction
PDGF: Platelet-derived Growth Factor
PTLD: Syndrome lymphoprolifératif post-transplantation
PKC : activateur de la protéine kinase C
RS: Reed-Sternberg

SCID: Severe Combined immunodeficiency
SIDA : Syndrome de l’immunodéficience humaine
TBP : TATA Box Binding Protein
TEM: Transition épithélio-mésenchymateuse
TGF: Transforming Growth Factor
TK: Thymidine Kinase
TSP1: Trombospondine 1
UL: Unique long
US: Unique short
VCA: Viral Capsid Antigen
VEGF: Vascular Endothelial Growth Factor
VIH : Virus de l’immunodéficience humaine
ZEBRA : BamH1 Z Epstein-Barr virus Replication Activator

Liste des figuresFigure 1 : Anatomie et sur la glande mammaire structure de la glande mammaire………………………………2
Figure 2 : Schéma de l’action hormonale…………………………………………………………………………6
Figure 3 : Evolution de la glande mammaire féminine au cours de la vie………………………………………..7
Figure 4 : Evolution des différents types de carcinomes mammaires…………………………………...……….15
Figure 5 : Mécanismes de la cancérogénèse……………………………………………………………………..18
Figure 6 : Incidence du cancer du sein dans le monde tout âge confondu……………………………………….25
Figure 7 : Incidence du cancer du sein dans le monde…………………………………………………………..26
Figure 8: Représentation de pourcentage de cancer du sein à Oran 1999-2008…………………………………27
Figure 9 : Représentation de pourcentages de quelques types de cancer du sein.
Figure 10 : Pourcentage de la morphologie de cancer du sein…………………………………………………...28
Figure 11: pourcentage des sources à Oran.
Figure12 : Représentation schématique linéaire du génome d’EBV (souche B95-8)……………………………38
Figure 13 : Génome épisomal de l’EBV et transcrits des gènes de latence.
Figure 14 : Représentation en fonction de la localisation de la tumeur………………………………………….74
Figure 15 : Représentation graphique en fonction de la contraception.
Figure 16: Représentation graphique de la fréquence de cancer du sein selon l’âge…………………………….75
Figure 17 : Représentation graphique en fonction des carcinomes.
Figure 18 : Représentation des grades apparus dans la population cible………………………………………...76
Figure 19 : Résultat IgG–VCA…………………………………………………………………………………..77
Figure 20 : Résultat -IgM VCA.
Figure 21 : Résultat IgA VC.………………………………………………………………………………….....78
Figure 22 : Résultat IgG-EA
Figure 23 : Résultat IgA-EA…………………………………………………………………………………..…79
Figue 24 : Représentation des trois Blots regroupant toutes les patientes EBNA…………………………….....80
Figure 25 : Marqueur de taille pour la technique en Western Blot………………………………………………81

Liste des tableaux
Tableau 1: Classification TNM de l‘UICC……………………………………………………21
Tableau II : Classification pTNM…………………………………………………………..…22

Introduction
1
I. IntroductionLe cancer du sein est dans la plupart des pays, le cancer le plus fréquent chez la
femme. Chaque année dans le monde, plus d’un million de nouveaux cas apparaissent, soit
30% des cas de cancer féminins dans les pays industrialisés et 14% dans les pays en voie
de développement. C’est aussi la 1ère cause de mortalité par cancer chez la femme avec 410
000 décès annuels (Rochefort et Rouessé, 2008). Plus de 9000 nouveaux cas de cancer du
sein sont enregistrés chaque année en Algérie, il reste le motif principal des consultations
en oncologie. La mortalité par cancer du sein est d’environ 3500 cas par an (Abid, 2009).
Le cancer est la première cause de mortalité dans le monde, il reste une maladie
multifactorielle où plusieurs événements peuvent être à l’origine de cette pathologie, on
notera à titre d’exemple les facteurs génétiques tels que les gènes BRCA1 et BRCA2 et
héréditaires, les facteurs environnementaux, à savoir la pollution, l’alimentation, les
radiations (les rayons UV, les rayons X …) et les infections par certains germes. Le tabac
et l’alcool à leur tour représentent un facteur de risque extrêmement important pour
certains cancers tel que les cancers bronchiques et les cirrhoses hépatiques, le cancer de la
bouche, de larynx, la gorge, les reins, le col, de l’utérus et le cancer du colon.
Le cancer est donc une maladie mal connue d’origines diverses, aussi et tout
récemment, la notion de cancérisation par virus revient en force dans le domaine de la
recherche scientifique puisque les cancers viro-induits sont de plus en plus étudiés.
Environ 15 à 20% des cancers sont induits par des infections virales, ce pourcentage
est encore plus élevé dans les pays à faibles ressources où les conditions d’infection sont
plus répandus. En effet, au cours de ces dernières années l’origine virale du cancer s’est
imposée dans la compréhension du processus tumoral, et de multiples investigations sont
proposées en réponse à des hypothèses qui ont été émises.
Des recherches sont menées dans le cadre des études épidémiologiques et
biologiques pour pouvoir bien comprendre le lien qui peut exister entre le virus et le
développement du cancer. Plusieurs familles de virus sont susceptibles d’induire le cancer
comme le virus d’Epstein-Barr (EBV), le virus de l’hépatite B (VHB), le virus de l’hépatite
C (VHC), le virus du lymphome humain à cellules T de type I (HTLV-I), le virus du
papillome humain (VPH) et quelques rétrovirus tel que le MMTV et autres.
Ainsi dans ce mémoire de Magister, nous avons essayé d’explorer la voie virale,
l’objectif est de rechercher les marqueurs sérologiques du virus Epstein-Barr (EBV) chez
des femmes atteintes de cancer du sein dans la région de l’Ouest Algérien.
Chapitre I : Anatomie, histologie et physiologie du sein
2
1. Anatomie et structure de la glande mammaireLes glandes mammaires sont responsables de la lactation (production du lait),
fonction primordiale dans la nutrition (et donc la survie) du nouveau né. Dans le lait
maternel, le nourrisson puise tous les éléments nécessaires pour son développement :
minéraux, vitamines, glucides, lipides et protéines. Parmi les protéines, on trouve les
anticorps, dont les IgA, très actives dans la défense immunitaire du tube digestif du
nouveau-né. Ce lait essentiel pour le nourrisson est fabriqué au niveau des lobes dans les
glandes mammaires, un tissu complexe dont le développement et fonctionnement est sous
l’influence d’hormones et de facteurs de croissance.
Le sein se compose de la glande mammaire, entourée par du tissu conjonctif et du
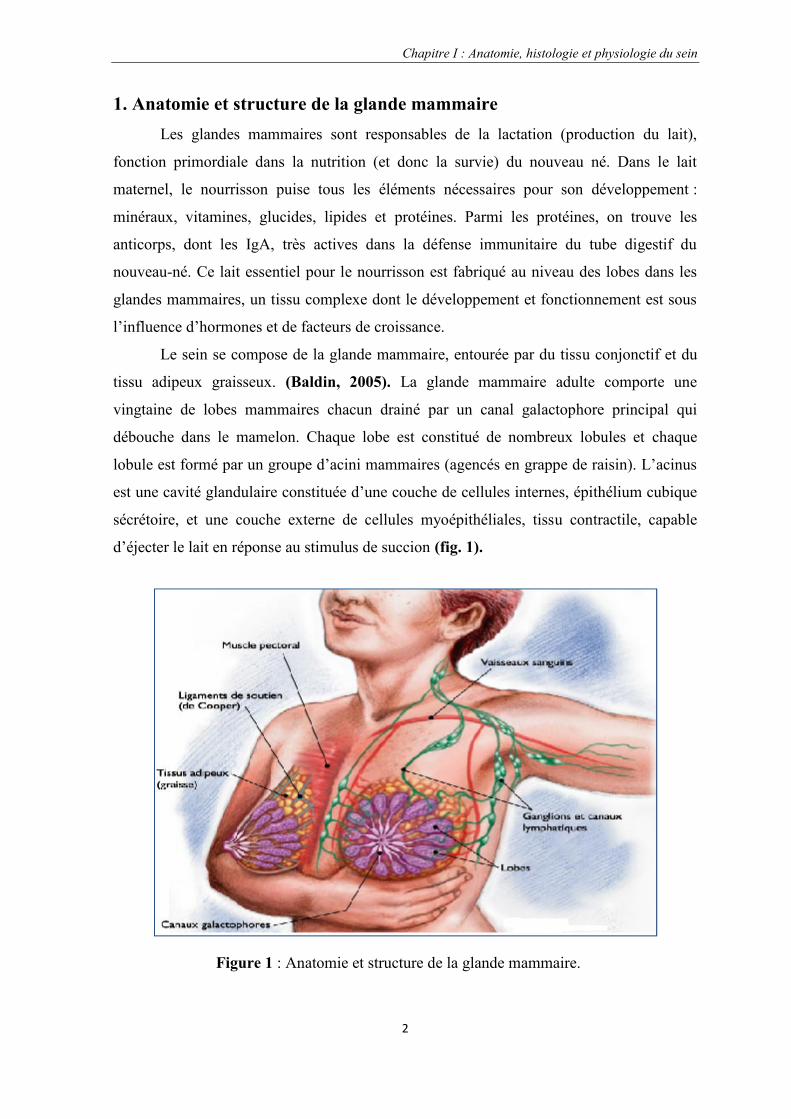
tissu adipeux graisseux. (Baldin, 2005). La glande mammaire adulte comporte une
vingtaine de lobes mammaires chacun drainé par un canal galactophore principal qui
débouche dans le mamelon. Chaque lobe est constitué de nombreux lobules et chaque
lobule est formé par un groupe d’acini mammaires (agencés en grappe de raisin). L’acinus
est une cavité glandulaire constituée d’une couche de cellules internes, épithélium cubique
sécrétoire, et une couche externe de cellules myoépithéliales, tissu contractile, capable
d’éjecter le lait en réponse au stimulus de succion (fig. 1).
Figure 1 : Anatomie et structure de la glande mammaire.

Chapitre I : Anatomie, histologie et physiologie du sein
3
2. HistologieLa glande mammaire est formée d'une part de tubes glandulaires ou galactophores,
bordés d'une double couche cellulaire : une interne formée de cellules épithéliales
cylindriques ou cylindro-cubiques, l’autre externe faite de cellules myoépithéliales. Le tout
est entouré de la membrane basale. Et d’autre part, d’un tissu conjonctif, qui au moment de
la puberté se différencie en deux parties : le tissu interstitiel banal qui constitue le support
des lobes comportant de nombreux adipocytes et la plaque aréolomamelonnaire qui est
tapissée par un épiderme plus ou moins pigmenté. Le derme sous-jacent comporte des
annexes cutanées, un réseau lymphatique bien développé et de nombreux faisceaux
musculaires lisses (Gosset, 2007).
3. Origine embryonnaire du tissu mammaireEntre la partie craniale et caudale, il y a apparition d’une bande mammaire qui
deviendra une crête mammaire. C’est à partir de celle-ci, que les ébauches mammaires
vont apparaître et que la crête mammaire existe virtuellement à l’âge adulte. Chez
l’homme, l’ébauche est unique et située au niveau du 4ème espace intercostal. On peut avoir
des ébauches mammaires ectopiques, exemple de sein surnuméraire, qui peut être l’objet
d’un cancer du sein (Amice, 2006).
4. Vascularisation
4.1. La vascularisation sanguineElle est assurée par deux artères, branches de l'artère sous-clavière :
l'artère mammaire externe, superficielle ;
l'artère mammaire interne (ou thoracique interne), profonde, a un trajet sous-
costal à proximité du bord du sternum.
Ces artères se divisent dans le conjonctif pour alimenter un réseau capillaire dense
péri-alvéolaire. Le sang veineux rejoint des troncs veineux qui ont un trajet parallèle aux
artères. La veine mammaire externe se jette dans la veine sous-clavière. La veine
mammaire interne se jette dans le tronc veineux brachio-céphalique. Par ailleurs un réseau
veineux sous-cutané, bien apparent au cours de la grossesse, se draine dans la veine
axillaire.

Chapitre I : Anatomie, histologie et physiologie du sein
4
4.2. La vascularisation lymphatiqueDans la glande, le réseau lymphatique est périlobulaire. Ce réseau alimente des
troncs lymphatiques présentant la même distribution que les vaisseaux sanguins. Le réseau
externe est le plus important : il draine les lymphatiques des 3/4 inférieurs et externes de la
glande. Il rejoint les lymphatiques axillaires. Le réseau interne, rejoint la chaîne
lymphatique parasternale, le long du bord externe du sternum. La distribution des
lymphatiques a des conséquences importantes en pathologie. Les cancers mammaires
donnent rapidement des métastases ganglionnaires dont la situation est fonction du
territoire de la tumeur primitive.
5. InnervationElle est assez riche, associant des fibres végétatives et des fibres sensitives. Les
fibres végétatives forment des plexus autour des vaisseaux, des canaux et des alvéoles.
L'innervation sensitive est très développée au niveau de la peau du mamelon. A ce niveau,
les corpuscules sensoriels sont nombreux. Ils ont un rôle fonctionnel au moment de la tétée
(Amice, 2006).
6. La physiologie de la glande mammaireLe sein humain dépend essentiellement de trois hormones : l'œstradiol, la
progestérone, la prolactine. Hormis la période pubertaire, où la croissance mammaire
dépend de l'œstradiol, seul sécrété par l'ovaire pendant plusieurs années, le sein est soumis,
dès que commencent les cycles ovulatoires, à l'alternance des sécrétions d'œstradiols et de
progestérone. La prolactine intervient au moment de la grossesse pour stimuler la
« mammogénèse », et lors de la lactation pour permettre la « lactogènes ». Ce sont les
actions synergiques et antagonistes de l'œstradiol et de la progestérone sur la prolifération
et la différenciation mammaires. Il s'agit d'une question essentielle, dans la mesure où de
l'équilibre de ces deux hormones dépend l'eutrophie du tissu mammaire au cours des
différentes périodes de la vie génitale, lors des cycles menstruels spontanés, mais aussi lors
de divers traitements hormonaux (inducteurs de l'ovulation, contraceptifs, ou substitutifs de
la ménopause) et aussi dans l'élaboration de stratégies en matière de prévention du cancer
du sein.

Chapitre I : Anatomie, histologie et physiologie du sein
5
6.1. Facteurs hormonaux
6.1.1. Les ŒstrogènesIls ont un rôle essentiel dans le développement mammaire, de nombreuses études in
vitro ont montré que les œstrogènes stimulaient la prolifération des cellules épithéliales. En
effet, in vivo, ils n'ont pas d'effet prolifératif direct mais stimulent la production d'un
certain nombre de facteurs de croissance (TGFα, IGF1, PDGF) par les éléments de la
matrice extracellulaire. Cependant, cette matrice extracellulaire, abondante sur le sein au
repos est très réduite autour des bourgeons en croissance ; ceci suggère un autre
mécanisme d'action sans doute prépondérant des œstrogènes. La matrice extracellulaire a
un rôle inhibiteur sur la croissance du sein, les œstrogènes agiraient en favorisant sa
destruction locale permettant aux bourgeons mammaires de proliférer.
6.1.2. La progestéroneElle est nécessaire à la différentiation lobulo-alvéolaire du sein. Ses effets sur la
prolifération épithéliale restent controversés : pour certains, elle inhibe la prolifération
cellulaire, pour d'autres elle est plutôt mitogène. In vivo et contrairement à ce qui se passe
au niveau de l'endomètre, l'index mitotique des cellules épithéliales est maximal en phase
lutéale. L’action de la progestérone n'est probablement pas univoque :
effet mitogène sur les cellules dont la prolifération dépend de l'EGF, effet
antimitogène sur celles dont la prolifération ne dépend pas de l'EGF ;
action antiproliférative sur l'épithélium canalaire et acineux, proliférative sur les
terminaisons ductulolobulaires ;
effet biphasique dans le temps : prolifératif en phase lutéale précoce puis
antiprolifératif.
Quoiqu'il en soit, les deux hormones, œstradiol et progestérone, agissent en
synergie et sont nécessaires au développement d'une glande apte à la lactation.
6.1.3. La prolactineLa prolactine est l'hormone lactogène : après l'accouchement, sa sécrétion intense
provoque la montée laiteuse. Elle a aussi un rôle dans le développement de la glande
mammaire, elle contribue à la différentiation des alvéoles au cours de la grossesse.
L'entretien de la lactation ou galactopoïèse se fait sous son influence.
Chapitre I : Anatomie, histologie et physiologie du sein
6
La sécrétion de prolactine va être entretenue pendant toute la durée de l'allaitement,
en réaction à la succion du mamelon par le bébé. En dehors de cette période, la prolactine
n'intervient pas dans la fonction mammaire.
6.1.4. Autres hormonesLes autres hormones (hormone de croissance, l'hormone lactogène placentaire,
cortisol) interviennent également dans le développement de la glande mammaire
(Ponsy, 1995).
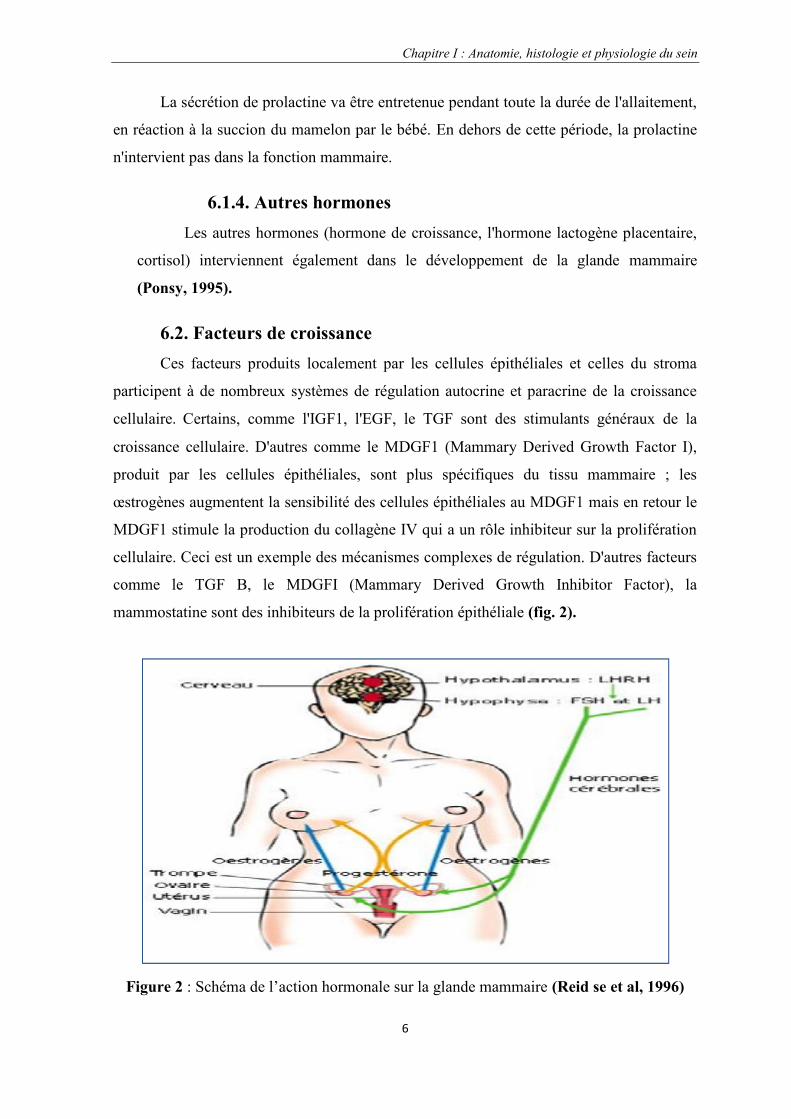
6.2. Facteurs de croissanceCes facteurs produits localement par les cellules épithéliales et celles du stroma
participent à de nombreux systèmes de régulation autocrine et paracrine de la croissance
cellulaire. Certains, comme l'IGF1, l'EGF, le TGF sont des stimulants généraux de la
croissance cellulaire. D'autres comme le MDGF1 (Mammary Derived Growth Factor I),
produit par les cellules épithéliales, sont plus spécifiques du tissu mammaire ; les
œstrogènes augmentent la sensibilité des cellules épithéliales au MDGF1 mais en retour le
MDGF1 stimule la production du collagène IV qui a un rôle inhibiteur sur la prolifération
cellulaire. Ceci est un exemple des mécanismes complexes de régulation. D'autres facteurs
comme le TGF B, le MDGFI (Mammary Derived Growth Inhibitor Factor), la
mammostatine sont des inhibiteurs de la prolifération épithéliale (fig. 2).
Figure 2 : Schéma de l’action hormonale sur la glande mammaire (Reid se et al, 1996)

Chapitre I : Anatomie, histologie et physiologie du sein
7
7. Développement de la glande mammaireL'organisation tissulaire et l'activité épithéliale sont le résultat d'un équilibre entre
les influences des hormones sexuelles œstrogènes et progestérone, de la prolactine, de
l’hormone de croissance et de facteurs de croissance divers (EGF, insuline, IGF-1). A un
moindre degré, les glucocorticoïdes et les minéralocorticoïdes peuvent également
intervenir. Au stade embryonnaire, un motif de base de la glande mammaire, commun aux
deux sexes, est mis en place. A la différence des autres organes, la glande mammaire reste
très peu présente jusqu’à la puberté. Son développement est étroitement contrôlé par des
hormones stéroïdiennes (œstrogène, progestérone, corticoïdes) et non-stéroïdiennes
(prolactine et ocytocine) agissant en synergie avec le microenvironnement conjonctif des
acini (facteurs de croissance locaux). (fig. 3).
De la naissance à la puberté, la croissance et la ramification des canaux
galactophores sont sous l’influence des hormones œstrogènes (ovaire en développement) et
corticoïdes (stéroïdes de la glande corticosurrénale) soutenues par des facteurs de
croissance produits par le tissu conjonctif de la glande elle-même, tels que HGF, IGF-1,
activine et épimorphine.
Durant la première grossesse les œstrogènes, la progestérone, la prolactine et les
corticoïdes, encore soutenues par la production locale de facteurs de croissance tels que
neuréguline, activine, FGF et épimorphine, induisent une forte activité proliférative des
cellules épithéliales lobulaires (développement lobulo-acineux).
Figure 3 : Evolution de la glande mammaire féminine au cours de la vie.

Chapitre I : Anatomie, histologie et physiologie du sein
8
Ces facteurs provoquent également l’accumulation de matériel sécrétoire dans ces
mêmes cellules (entrainant ainsi l’augmentation transitoire du volume des glandes
mammaires).
Durant la lactation les cellules épithéliales lobulaires sont gorgées des composants
du lait. La sécrétion vers la lumière de l’acinus est provoquée par l’action de la prolactine
et soutenue par les corticoïdes. Dans ce processus la partie supérieure (apicale) de la
cellule se détache pour former la sécrétion lactée (phénomène d’apocytose).
A l’arrêt de lactation causé par le sevrage, la glande mammaire régresse. Le
processus dit d’involution implique l’arrêt d’expression des gènes codant les protéines du
lait (caséine et lactalbumine et lactoglobuline) et la disparition progressive des cellules
épithéliales, remplacées par du tissu adipeux.
Durant la ménopause les cellules épithéliales s’atrophient davantage par manque de
soutien hormonal. Le tissu adipeux devient dense et le tissu conjonctif perd sa fonction car
les fibres de collagène et d’élastine sont altérées.
L’ensemble de ces processus illustre bien l’étroite hormonodépendance de la
glande mammaire aussi bien au niveau anatomique que fonctionnel (Coudert et al, 2007).

Chapitre II : Anatomie pathologique du sein
9
1. Pathologies bénignes de la glande mammaireLa pathologie bénigne est peu connue et a été beaucoup moins étudiée que le cancer
du sein, bien qu’elle soit encore plus fréquente, les pathologies mammaires humaines
bénignes regroupent un large éventail de lésions physiopathologiques des divers
composant du sein. En 1985, sur la base des travaux de Dupont et Paget, un consensus
national de classification histologique a été adopté (Dupont et al., 1985). Le concept de
maladie proliférative épithéliale bénigne identifie les caractéristiques histologiques des
maladies bénignes mammaires ayant un potentiel d’évolution vers le cancer du sein.
1.1. L’adénofibrome
1.1.1. Adénofibrome uniqueL’adénofibrome nommé par Charles Gros « la tumeur de la fiancée » est la tumeur
bénigne la plus fréquente chez les jeunes filles de moins de 25 ans (Morrow M et al.,
1998). Il s’observe plutôt chez les jeunes filles pubères qu’en période prépubertaire.
Histologiquement, la prolifération intéresse le tissu épithélial canaliculaire et stroma
fibreux lobulaire. Du point de vue évolutif, l’adénofibrome peut soit resté stable, soit
augmenter de taille (notamment lors des grossesses), soit régresser spontanément.
En échographie, l’adénofibrome correspond à un nodule hypo-échogène avec un
renforcement postérieur. (Ce dernier n’est présent que dans 40% des cas). Les contours
sont réguliers. Cette masse bénigne présente un grand axe parallèle au plan cutané avec un
rapport L sur e supérieur à 1,4 dans 86% des cas (Fornage et al., 1989). Des zones
d’infarctus intra-tumoraux peuvent survenir et donner des calcifications séquellaires (mais
cet aspect est rare chez l’adolescente). Actuellement, une chirurgie d’exérèse est conseillée
par certaines équipes chirurgicales vers 25 ans ou 30 ans, voire plus tôt en cas de masse
volumineuse (Stehr et al., 2004).
1.1.2. L’adénofibromatoseCliniquement, l’adolescente se présente avec des nodules multiples des deux seins,
fermes, mobiles, indolores. Les adénofibrome peuvent être multiples et bilatéraux dans 10
à 20% des cas, ils sont plus fréquents dans la race noire. Une surveillance clinique et
échographique est instaurée.

Chapitre II : Anatomie pathologique du sein
10
1.1.3. L’adénofibrome juvénileL’adénofibrome juvénile survient à l’adolescence, généralement entre 14 et 19 ans.
Sa fréquence est estimée à 4% de l’ensemble des adénofibrome. Cliniquement, il existe
une augmentation rapide et douloureuse du volume mammaire. Par palpation d’un nodule
ferme, le plus souvent unique, moins volontiers multiple. L’aspect échographique peut être
celui d’un adénofibrome classique ou celui d’un nodule hétérogène.
1.2. Le kyste rétro-aréolaire de l’adolescenteLes kystes rétro-aréolaires de l’adolescente sont réputés rares, mais un certain
nombre est méconnu, car asymptomatique. Sur le plan anatomique, il existe des glandes
sébacées aréolaires centrales qui ne sont pas associées à un follicule pileux mais à de
petites glandes mammaires accessoires. Le sébum est ainsi libéré dans un petit canal
galactophorique provenant de lobules mammaires accessoires plus profondément situés
sous l’aréole. Ce canal commun s’ouvre à la surface de l’aréole au centre d’un tubercule de
Montgomery (Huneeus et al., 2003). Ils peuvent être silencieux et souvent on découvre à
l’échographie un kyste contro-latéral asymptomatique. Echographiquement, ces kystes
correspondent à une image liquidienne arrondie ou ovalaire ou de type lobulaire. Le
caractère douloureux et la paroi épaissie traduisent le caractère inflammatoire. L’évolution
spontanée se fait vers la disparition ; il existe rarement une récidive. Un traitement
antibiotique anti-inflammatoire peut être proposé.
1.3. L’ectasie galactophoriqueContrairement à des notions anciennes, l’ectasie galactophorique peut s’observer
chez la fillette et l’adolescente. L’hypothèse actuelle est celle d’une maladie auto-immune
à médiation cellulaire. Sur le plan anatomopathologique, il existe au début une dilatation
simple des canaux sans altération des parois, ni de l’épithélium (phase asymptomatique).
Cliniquement, elle peut soit être asymptomatique, soit se révéler par un écoulement séreux
ou verdâtre, ou par une mastite. Des cas de régression spontanée ont été reportés (Stringel
et al., 1986). A l’échographie, il existe une dilatation des canaux galactophoriques
supérieure à 3mm. L’évolution peut se faire vers la mastite à plasmocytes avec
d’éventuelles poussées inflammatoires infectieuses (abcès avec pus stérile ou microbien)
ou bien vers la régression.

Chapitre II : Anatomie pathologique du sein
11
1.4. Les abcès du seinChez le nouveau-né, l’abcès mammaire fait souvent suite à la crise génito-
mammaire néonatale et survient dans les trois premières semaines de la vie. Les
manipulations de la région mammaire favorisant ces abcès. Chez la fillette plus âgée et
l’adolescente, l’abcès survient par surinfection d’une ectasie galactophorique ou d’un kyste
rétro-aréolaire. Une éventuelle fistule peut favoriser les récidives. L’abcès du sein peut
correspondre plus rarement à un embole septique à partir d’un foyer lointain et à
l’extension d’une infection cutanée. Cliniquement, le sein est rouge, chaud, douloureux.
Quand l’abcès est collecté, on peut palper une masse fluctuante. Les signes généraux et
l’écoulement purulent sont inconstants. Lorsque ce dernier est présent, un examen
cytobactériologique peut être réalisé. Au stade collecté, l’image est celle d’un kyste à
contours épais ou à paroi épaissie avec un renforcement postérieur.
Il est possible de visualiser des amas hyper-échogènes. On peut également observer
des collections hypo-échogènes (Boisserie-La et al., 1993). Le risque est l’évolution vers
la cellulite qui sera prévenue par une antibiothérapie systématique adaptée à un
antibiogramme réalisé sur le liquide d’écoulement ou sur le liquide de drainage. Certains
auteurs préconisent un drainage chirurgical par une incision radiée ou périaréolaire. Lors
d’abcès récidivants, la recherche d’une fistule et son excision seront réalisées.
1.5. La mastopathie fibrokystiqueLa maladie fibrokystique est un terme contesté. Elle associe des lésions kystiques,
de fibrose, d’hyperplasie épithéliale lobulaire, d’adénose sclérosante. Les kystes
mammaires se développent à partir d’une dilatation des canaux intra-lobulaires. Sa
fréquence est d’environ 10% des tuméfactions mammaires avant 20 ans. Cliniquement,
l’adolescente ressent des mastodynies prémenstruelles à type de pesanteur. Des kystes
peuvent être palpés ; leur tension est parfois telle qu’ils peuvent être confondus avec des
masses solides et l’échographie est indispensable pour le diagnostic. Si un doute persiste,
une aspiration à l’aiguille sera effectuée. Au cours du cycle et d’un cycle à l’autre, la taille
et le nombre des kystes sont variables. A l’échographie, Il existe de nombreux nodules
ovalaires transsoniques, aux contours réguliers et présentant un renforcement postérieur
dont l’aspect est typique de kystes. La majorité de ces jeunes filles présente une
insuffisance lutéale. Un traitement par une pilule œstro-progestative à climat
progestéronique peut être proposé (McDivitt et al., 1996).

Chapitre II : Anatomie pathologique du sein
12
1.6. La papillomatose juvénileLa papillomatose juvénile est une entité anatomo-clinique particulière, décrite par
Rosen pour la première fois en 1980 ; elle touche dans 70% des cas des jeunes filles de
moins de 25 ans présentant un pourcentage élevé (environ 30%) d’antécédents familiaux
de cancer du sein. Cette affection rare constitue un marqueur de risque familial ou
individuel de cancer du sein (Bazzocchi et al., 1986). Macroscopiquement, la tumeur se
compose de nombreux kystes avec une lumière encombrée de sécrétions au sein d’un tissu
dense. Microscopiquement, on observe des kystes bordés d’un épithélium aplati avec une
métaplasie papillaire apocrine et une hyperplasie floride de l’épithélium canalaire.
Cliniquement, on palpe une masse douloureuse, ferme du sein, qui peut
s’accompagner parfois d’un écoulement mamelonnaire et qui est unilatéral. L’échographie
retrouve une lésion hypoéchogène avec des zones kystiques périphériques. Le traitement
est fonction de plusieurs paramètres : s’il s’agit d’une lésion localisée, qu’il n’existe pas
d’antécédent familial de cancer du sein, une segmentectomie large est réalisée avec une
surveillance attentive. Si l’histologie montre des lésions diffuses et atypiques, que
l’ensemble de la plage mammaire est ponctué de microcalcifications, il faut discuter
l’opportunité d’une mastectomie sous-cutanée avec prothèse d’emblée. Etant donné
l’incertitude du pronostic, une surveillance des deux seins tous les 2 ans est instaurée. Un
dépistage chez les parents proches devra être institué.
1.7. La tumeur phyllodeC’est une tumeur rare survenant le plus souvent dans la quatrième décennie. Seule
une cinquantaine de cas ont été publiés chez l’adolescente. Il n’existe pas de critère absolu
pour juger de la bénignité ou de la malignité des tumeurs phyllodes, le seul critère de
malignité absolue demeurant la présence de métastases (ganglionnaire, pulmonaire et
hépatique). Cliniquement, c’est une tumeur qui augmente rapidement de taille, Polylobée,
mais qui peut également avoir les caractères d’un adénofibrome banal (Buchberger,
1991). A l’échographie, l’aspect peut être similaire à celui décrit dans l’adénofibrome
juvénile, mais il faut noter la possibilité de zones liquidienne intra tumorales. L’examen
anatomo-pathologique affirme le diagnostic, une tumorectomie élargie est nécessaire pour
éviter les récidives et semble suffisantes chez l’adolescente, la survenue de métastases
étant exceptionnelle à cet âge.

Chapitre II : Anatomie pathologique du sein
13
2. Pathologies malignes du seinLa distinction entre les différents types de cancer est basée sur leur origine
tissulaire et segmentaire, sur le degré d'infiltration en dehors du système galactophorique et
sur la différenciation de la tumeur par rapport à la glande mammaire normale (Chevalier,
2000).
2.1. Les carcinomes mammaires ou épithéliomasNées de l'épithélium des lobules ou canaux, ces tumeurs malignes représentent à
elles seules 98% des tumeurs malignes du sein.
2.1.1. Les carcinomes lobulairesLes carcinomes lobulaires naissent dans les canalicules terminaux intralobulaires.
Les carcinomes lobulaires in situ représentent environ 1% des cancers du sein, surviennent
dans 90% des cas en période d'activité génitale et sont volontiers multicentriques et
bilatéraux ; ils n'ont pas de traduction radiographique directe. Seuls 20% des carcinomes
lobulaires in situ deviennent invasifs à 5 ans. Les carcinomes lobulaires infiltrants ou
carcinomes à petites cellules de Haagensen représentent environ 5% des cancers du sein ;
la recherche des récepteurs en œstrogènes est positive dans 90% des cas, alors qu'elle ne
l'est que dans 55% des cas pour les autres carcinomes mammaires invasifs ; lors de
l'examen anatomo-pathologique on trouve souvent des foyers de carcinomes lobulaires in
situ associés.
2.1.2. Les carcinomes canalairesCes tumeurs naissant dans les canaux galactophores de préférence de 3ème ordre,
représentent la grande majorité des carcinomes.
Les carcinomes canalaires in situ représentent environ 2% des cancers du sein et
comportent plusieurs variétés architecturales, allant du carcinome papillaire au
comédocarcinome progressant lentement de proche en proche au sein du système
galactophorique ; ce qui lui donne un pronostic favorable ; la bilatéralité
concomitante ou secondaire existe cependant dans 15 à 20% des cas ;
Les carcinomes canalaires invasifs représentent le groupe le plus important des
cancers du sein (environ 75%) ; différents sous-groupes ont été individualisés,
tenant compte essentiellement du degré de différenciation ; il existe d'autre part des
sous-groupes particuliers, caractérisés par des spécificités histologiques propres.

Chapitre II : Anatomie pathologique du sein
14
Le carcinome colloïde ou mucineux : appelé ainsi en raison de la production d'une
grande quantité de mucus extra-cellulaire. Les éléments carcinomateux sont en
quantité souvent faible par rapport à la substance colloïde ;
Le carcinome médullaire : constitué de cellules peu différenciées, atypiques, dans
un stroma peu abondant avec intense infiltration lymphoïde. Les limites de cette
tumeur apparaissent cependant bien circonscrites avec fréquemment présence de
foyers de nécrose. Son évolution serait plus favorable que ne laisseraient prévoir
les importantes anomalies cytonucléaires ;
Le carcinome tubulaire : représente une variété de carcinome très différencié, dont
les cellules sont régulières et disposées en tubules. Le pronostic est habituellement
favorable ;
Le carcinome cylindromateux ou adénoïde kystique : se présente histologiquement
comme les cylindromes des glandes salivaires. L'évolution de ces formes est lente
et leur pronostic assez favorable ;
Le carcinome apocrine est formé de cellules à abondant cytoplasme éosinophile
analogue à celui des cellules apocrines métaplasiques ;
Les carcinomes métaplasiques : forme spinocellulaire ou épidermoïde ; il s'agit le
plus souvent de formes induites par des remaniements nécrotiques. Fréquemment
accompagnées d'une stroma-réaction riche en fibroblastes, elles sont souvent
interprétées à tort comme carcinosarcomes ;
Le carcinome riche en lipides est exceptionnel mais d'un pronostic
particulièrement péjoratif ;
La maladie de Paget du mamelon et la mastite carcinomateuse sont deux entités
qui ne doivent pas être considérées comme des types histologiques particuliers ;
mais représentent en fait des modes d'extension possibles de n'importe quel cancer
mammaire.
2.2. Les sarcomes mammairesLes sarcomes mammaires constituent la seconde variété de tumeurs malignes du
sein, mais sont rares (1%). Ils peuvent naître à partir du contingent mésenchymateux d'une
tumeur bénigne préexistante, cette composante prenant le pas sur la composante
épithéliale, qui s'efface. Ceci est le cas des tumeurs phyllodes, classées en 4 catégories ou 3
grades dont seul le stade IV ou grade 3 est véritablement malin.
Chapitre II : Anatomie pathologique du sein
15
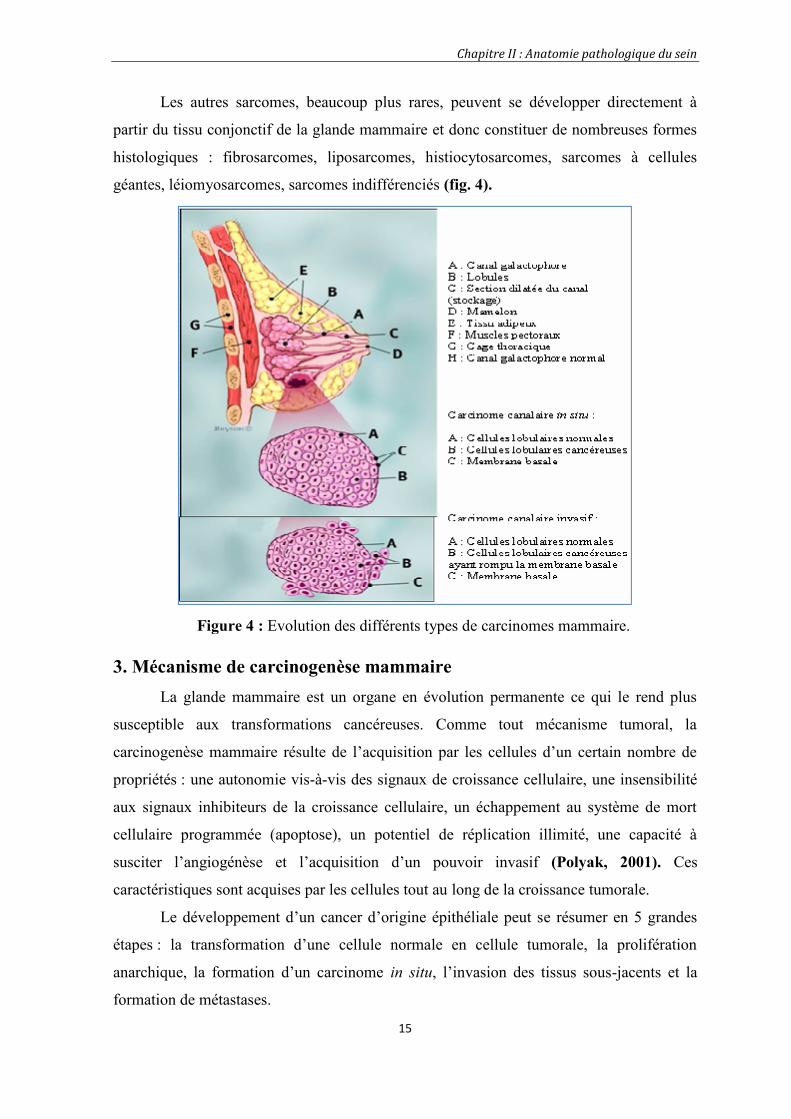
Les autres sarcomes, beaucoup plus rares, peuvent se développer directement à
partir du tissu conjonctif de la glande mammaire et donc constituer de nombreuses formes
histologiques : fibrosarcomes, liposarcomes, histiocytosarcomes, sarcomes à cellules
géantes, léiomyosarcomes, sarcomes indifférenciés (fig. 4).
3. Mécanisme de carcinogenèse mammaireLa glande mammaire est un organe en évolution permanente ce qui le rend plus
susceptible aux transformations cancéreuses. Comme tout mécanisme tumoral, la
carcinogenèse mammaire résulte de l’acquisition par les cellules d’un certain nombre de
propriétés : une autonomie vis-à-vis des signaux de croissance cellulaire, une insensibilité
aux signaux inhibiteurs de la croissance cellulaire, un échappement au système de mort
cellulaire programmée (apoptose), un potentiel de réplication illimité, une capacité à
susciter l’angiogénèse et l’acquisition d’un pouvoir invasif (Polyak, 2001). Ces
caractéristiques sont acquises par les cellules tout au long de la croissance tumorale.
Le développement d’un cancer d’origine épithéliale peut se résumer en 5 grandes
étapes : la transformation d’une cellule normale en cellule tumorale, la prolifération
anarchique, la formation d’un carcinome in situ, l’invasion des tissus sous-jacents et la
formation de métastases.
Figure 4 : Evolution des différents types de carcinomes mammaire.

Chapitre II : Anatomie pathologique du sein
16
3.1 Initiation de la progression tumoraleLa première étape du processus de cancérisation mammaire est une phase
d’initiation. Elle se caractérise par une accumulation d’altérations géniques que les
systèmes de réparation, débordés ou défectueux, ne peuvent plus compenser. De telles
transformations cellulaires ont pour conséquence une surexpression des facteurs pro-
oncogéniques. La cellule reste tout de même contrôlée par l’environnement cellulaire grâce
aux jonctions « gap ». Les facteurs pro-oncogéniques sont généralement des facteurs de
croissance secrétés par les cellules de carcinomes mammaires. Une fois synthétisés, ces
facteurs stimulent les cellules cancéreuses et les cellules du stroma via des mécanismes
auto- et paracrines. Les facteurs de croissance identifiés inclus l’EGF (Epithelial Growth
Factor), le TGF-b, le VEGF (Vascular Endothelial Growth Factor), l’IGF-2, le PDGF
(Platelet-derived Growth Factor) et le FGF (Fibroblast Growth Factor). L’EGF, le TGF-b
l’IGF-1 et l’EGF-2 sont exprimés et sécrétés par des lignées cellulaires et des tissus de
carcinomes du sein et représentent des agents mitogènes pour la composante épithéliales de
ces tumeurs. Le PDGF et le FGF sont également produit par les cellules tumorales et sont
responsables de la prolifération stromale retrouvée dans beaucoup de carcinomes
mammaires.
Les autres oncogènes fréquemment retrouvés surexprimés dans les carcinomes
mammaires regroupent les membres de la famille de récepteurs à l’EGF (EGFR) incluant
notamment erbB-2, HER-3 et HER-4, ainsi que des membres des familles ras et myc
(Eccles, 2001; Neve et al. 2001). Les mécanismes de cancérisation sont aussi provoqués
par la perte des fonctionnalités de gènes suppresseurs de tumeur tels que p53, RB1 ou
BRCA1 et BRCA2.
3.2. Promotion tumoraleLes cellules mammaires génétiquement anormales entrent ensuite dans la seconde
étape dite de promotion où elles échappent aux contrôles de leur multiplication cellulaire et
de leur apoptose, suite à la perte de fonctionnalité d’anti-oncogènes. Elles perdent
également leur capacité de communication intercellulaire. La prolifération anarchique
d’une cellule transformée peut aboutir à la formation d’une tumeur qui ne franchit pas la
membrane basale et reste localisée au niveau de l’épithélium : on parle alors de carcinome
in situ. Cette première étape de transition d’un épithélium normal à un carcinome in situ est
caractérisée par un indice mitotique élevé et une absence de différenciation cellulaire.

Chapitre II : Anatomie pathologique du sein
17
Durant plusieurs mois voire plusieurs années, l’apoptose et la prolifération des
cellules tumorales peuvent s’équilibrer et le carcinome rester circonscrit à l’épithélium
sans franchissement de la membrane basale.
3.3. Métastases des carcinomes mammairesAu sein du foyer tumoral primaire, certaines cellules peuvent acquérir la capacité
d’envahir le tissu sous-jacent et de former des métastases à distance de la tumeur d’origine.
Cette capacité d’invasion nécessite l’acquisition de nouvelles propriétés biologiques par les
cellules tumorales. Elles subissent notamment une désorganisation des jonctions
intercellulaires favorisant ainsi la désolidarisation des cellules de l’épithélium et leur
progression dans le tissu sous-jacent. Les cellules acquièrent un phénotype proche des
cellules mésenchymateuses ainsi que la capacité à migrer et dégrader les constituants de la
membrane basale et de la matrice extracellulaire (MEC).
Ceci est rendu possible grâce à l’expression de diverses protéases, parmi lesquelles
on retrouve les métalloprotéases matricielles (MMPs), qui ont la capacité de dégrader la
majorité des éléments de la MEC et la membrane basale. Les cellules tumorales s’infiltrent
alors dans le stroma sous-jacent et se divisent activement. Lorsque la tumeur grossit,
l’apport en oxygène et en nutriments ne peut plus alors se faire par simple diffusion et cet
état entraîne une hypoxie intratumorale. Cette hypoxie induit une surexpression de gènes
angiogéniques et va déclencher un processus de néo-vascularisation qui va favoriser la
progression tumorale (Kerbel et Folkman, 2002 ; Pugh et Ratcliffe, 2003). En effet, du
fait de l’angiogenèse qu’elles ont induite, les cellules tumorales se trouvent fréquemment à
proximité de néovaisseaux. Certaines d’entre elles vont digérer la membrane basale des
néovaisseaux ou la paroi lymphatique et pénétrer lors d’un processus d’intravasation dans
la circulation sanguine ou lymphatique. Quelques cellules se fixeront à la paroi des
capillaires où le flux sanguin est faible et formeront des agrégats ou emboles cellulaires.
Les cellules tumorales quittent ensuite ces vaisseaux au cours de l’extravasation pour
envahir d’autres organes où elles prolifèrent et créent des métastases. Les organes les plus
fréquemment atteints lors des métastases des cancers du sein sont les poumons, les os et le
foie ; souvent, les patientes développent également des métastases multiples aux
organes.(fig. 5).

Chapitre II : Anatomie pathologique du sein
18
Figure 5 : Mécanismes de la cancérogénèse (FNCLCC, 2009).

Chapitre II : Anatomie pathologique du sein
19
4. Classification des tumeurs mammairesDeux types de classifications sont le plus couramment utilisés : La classification
histologique et la classification clinique.
4.1. Classifications histologiques des carcinomes infiltrants
4.1.1. Classification de l’OMS (1981) : la classification de
l’OMS des carcinomes (C) infiltrantsDatant de 1981, définit sur le plan morphologique 14 entités différentes elle ne
tenait pas compte des problèmes rencontrés par le pathologiste en pratique quotidienne.
Elle avait le mérite de classifier, selon leur aspect morphologique, différents types de
carcinomes infiltrants à pronostic inégal :
Carcinome canalaire infiltrant ;
Carcinome canalaire infiltrant avec composante intra canalaire prédominante ;
Carcinome lobulaire infiltrant ;
Carcinome mucineux ;
Carcinome médullaire ;
Carcinome papillaire ;
Carcinome tubuleux ;
Carcinome adénoïde kystique ;
Carcinome sécrétant (juvénile) ;
Carcinome apocrine ;
Carcinome métaplasiques ;
Autres : Carcinome à cellules riche en lipides; Carcinome à petites cellules ;
Carcinome à cellules en bague de chaton.
4.1.2. Classification de l’OMS modifiée (2002-2003)Cette classification tient compte des données morphologiques et
immunohistochimiques et elle a le mérite d’être exhaustive. Au total 21 entités de
carcinomes mammaires infiltrants sont définies :
Carcinome canalaire infiltrant de type non spécifique (SAI) ;
Carcinome lobulaire infiltrant ;
Carcinome tubuleux ;
Carcinome médullaire ;

Chapitre II : Anatomie pathologique du sein
20
Carcinome produisant de la mucine ;
Carcinome cribriforme infiltrant ;
Carcinome endocrine du sein ;
Carcinome métaplasique ;
Carcinome apocrine ;
Carcinome à cellules riches en lipides ;
Carcinome sécrétant (juvénile) ;
Carcinome adénoïde kystique ;
Carcinome à cellules acineuse ;
Carcinome à cellules claires (riche en glycogène) ;
Carcinome mammaire à cellules géantes ostéoclastiques ;
Carcinome mammaire avec faits choriocarcinomateux ;
Carcinome oncocytique ;
Tumeur mélanocytaire ;
Carcinome sébacé ;
Carcinome micropapillaires ;
Carcinome muco-épidermoïde.
4.2. Classification TNMLa classification TNM proposée par Pierre Denaix a le mérite de répondre à ces
exigences. Elle a été retenue comme base de classification par le comité de nomenclature et
de statistique de l’UICC. Elle est basée sur le principe de l’extension anatomique déterminé
par la clinique et l’histopathologie (tableau I).
A côté de la classification TNM, il existe une deuxième classification post-
chirurgicale nommée pTNM. Cette classification post-chirurgicale diffère de la première
TNM surtout en ce qui concerne les adénopathies (pN). De plus, elle permet de préciser
de façon rigoureuse la taille réelle de la tumeur appelée pT après prélèvement (tableau II).

Chapitre II : Anatomie pathologique du sein
21
Tableau I : Classification TNM de l‘UICC (Singletary et Allred, 2002).
T : Tumeur
Tx Détermination de la tumeur primitive impossible.
Tis Carcinome in situ ; Carcinome intracanalaire ou canalaire lobulaire in situ ; Maladie de Paget du mamelon sans tumeur décelable.
T0 Pas de signes de tumeur primitive.
T1
T1 Tumeur de taille ≤ 2 cm.T1a > 0,1cm et strictement ≤ 0,5cm.T1b > 0,5cm et strictement <1cm.T1c > 1cm et strictement < 2cm.
T2 Tumeur > 2cm et <5cm.T3 Tumeur > 5cm.
T4
Tumeur de toute taille avec l’extension directe à la paroi thoracique ou à la peau.T4a L’extension à la paroi thoracique.
T4b Œdème (y compris la peau d’orange) ; Ou ulcération du sein ; Ou Nodule de perméation au même sein.
T4c A la fois T4a et T4b.T4d Carcinome inflammatoire.
N : adénopathies régionalesNx Appréciation impossible de l’attente ganglionnaire.N0 Absence de signe d’envahissement ganglionnaire.N1 Ganglions axillaires homo-latéraux mobiles.
N2 Ganglions axillaires homo-latéraux fixés entre eux ou à d’autres structures.
N3 Ganglions mammaires internes homolatéraux.
M : Métastases à distanceMx Détermination impossible de l’extension métastatique.M0 Absence de métastase à distance.M1 Présence de métastases à distance.
Chapitre II : Anatomie pathologique du sein
22
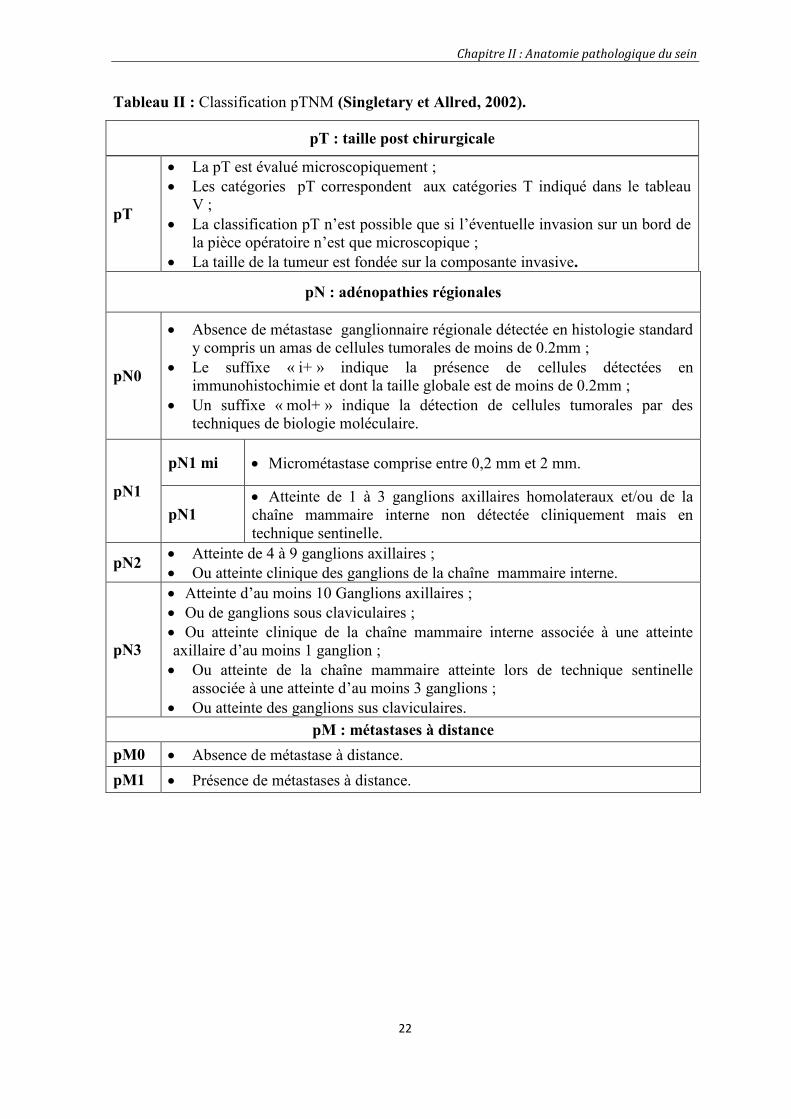
Tableau II : Classification pTNM (Singletary et Allred, 2002).
pT : taille post chirurgicale
pT
La pT est évalué microscopiquement ; Les catégories pT correspondent aux catégories T indiqué dans le tableau
V ; La classification pT n’est possible que si l’éventuelle invasion sur un bord de
la pièce opératoire n’est que microscopique ; La taille de la tumeur est fondée sur la composante invasive.
pN : adénopathies régionales
pN0
Absence de métastase ganglionnaire régionale détectée en histologie standardy compris un amas de cellules tumorales de moins de 0.2mm ;
Le suffixe « i+ » indique la présence de cellules détectées enimmunohistochimie et dont la taille globale est de moins de 0.2mm ;
Un suffixe « mol+ » indique la détection de cellules tumorales par destechniques de biologie moléculaire.
pN1
pN1 mi Micrométastase comprise entre 0,2 mm et 2 mm.
pN1 Atteinte de 1 à 3 ganglions axillaires homolateraux et/ou de lachaîne mammaire interne non détectée cliniquement mais entechnique sentinelle.
pN2 Atteinte de 4 à 9 ganglions axillaires ; Ou atteinte clinique des ganglions de la chaîne mammaire interne.
pN3
Atteinte d’au moins 10 Ganglions axillaires ; Ou de ganglions sous claviculaires ; Ou atteinte clinique de la chaîne mammaire interne associée à une atteinteaxillaire d’au moins 1 ganglion ; Ou atteinte de la chaîne mammaire atteinte lors de technique sentinelle
associée à une atteinte d’au moins 3 ganglions ; Ou atteinte des ganglions sus claviculaires.
pM : métastases à distancepM0 Absence de métastase à distance.pM1 Présence de métastases à distance.

Chapitre II : Anatomie pathologique du sein
23
4.3. Classification SBRLe type histologique d'un cancer et son degré de différenciation sont les facteurs les
plus anciennement connus : plus une tumeur est différenciée, plus l'évolution est favorable.
Les signes inflammatoires sont par contre un facteur très défavorable. La forme commune
canalaire infiltrante représentant 65% des carcinomes mammaires, d'autres facteurs ont été
définis. Le grade SBR ou le "grading" de Scarff-Bloom et Richardson est le plus souvent
utilisé.
Cette méthode appliquée aux carcinomes infiltrants consiste à noter trois
paramètres : la formation de tubes (degré de différenciation), l'anisonucléose et le taux des
mitoses. Trois niveaux sont définis : du grade I, de pronostic plus favorable, au grade III de
mauvais pronostic. Le grade SBR est le meilleur facteur pronostique reconnu à ce jour.
Pourtant, il n'est déterminé systématiquement que dans certains pays. Les facteurs
histologiques et le grade SBR sont encore aujourd'hui les plus utilisés comme facteurs
pronostiques mais ils sont difficiles à standardiser d'une équipe à l'autre (Rouëssé et al.,
1997) De plus, le taux de survie des femmes présentant un cancer du sein N- n'est que de
80%. Ainsi des facteurs biologiques propres à la tumeur ont été développés.

Chapitre III : Epidémiologie du cancer du sein
24
1. Epidémiologie de cancer du sein dans le mondeLe cancer du sein est le plus fréquent des cancers dans les pays occidentaux et
constitue le premier cancer féminin dans le monde avec environ 1 million de cas par an
(Dumistrescu et al, 2005). Le cancer du sein reste rare chez les hommes puisque 1%
d’entre eux présentent cette pathologie. En 2005, sur 60 000 femmes qui sont décédées
d’un cancer, environ 11 000 l’ont été à la suite d'un cancer du sein, ce qui fait de cette
maladie la première cause de mortalité par cancer chez la femme. Avec près de 42 000
nouveaux cas estimés chaque année, cela représente 36,7 % de l’ensemble des nouveaux
cas de cancers chez la femme. Une femme sur 10 développera un cancer du sein au cours
de sa vie. Cette pathologie est rare avant 30 ans, 95% des cas surviennent après 40 ans dont
près de la moitié sont diagnostiqués entre 50 et 69 ans. Par ailleurs, son incidence s’accroît
régulièrement, + 60% en 20 ans, ce qui fait de cette pathologie un problème majeur de
santé publique (Trétarre et al, 2004 ; Hill & Doyon, 2008). La mortalité par cancer du sein
est d’environ 3500 cas annuels (ABID 2009).
Globocan Le cancer du sein est le plus fréquent des cancers féminins, il frappe
nettement plus les pays industrialisés que les pays en voie de développement ou sous-
développés. (Globocan, 2008, IARC). (Figure 6)

Chapitre III : Epidémiologie du cancer du sein
25
Figure 6 : Incidence du cancer du sein dans le monde tout âge confondu.
(Globocan, 2008)
À l’échelle du globe, on estime qu’il y a eu 1,4 million de nouveaux cas de cancer
du sein estimés en 2008, représentant 1/5 de tous les nouveaux cas de cancer. C’est aussi
la forme la plus fréquente de décès par cancer chez les femmes dans le monde, représentant
1/8 de tous les décès par cancer (Globocan 2008). Le Japon qui présente un développement
socio-économique analogue à celui des pays industrialisés montre une incidence faible
28,61/100 000 (données de l'IARC) (fig. 6)
Chapitre III : Epidémiologie du cancer du sein
26
2. Epidémiologie du cancer du sein en AlgérieSelon des auteurs Algériens, une étude multicentrique rapportée en 2005 portant sur
un échantillon de 4892 patientes a révélée que les deux tiers des femmes atteintes du
cancer du sein ont moins de 50 ans et 22% ont moins de 40 ans. L’âge moyen des patientes
se situe autour de 48,3. Cette pathologie semble, en revanche, épargner les femmes
ménopausées.les 2/3 des femmes atteintes de cancer du sein sont en activité génitale et
49,10% pratiquent la contraception. La moitié des femmes (50%), révèle la même étude,
elles consultent dans les premiers mois de l’apparition du nodule alors que l’autre moitié
ne le fait que trois mois plus tard.
S’agissant de l’épidémiologie de la pathologie, le nord du pays vient en tête avec
71%, avec 44,36% dans la capitale, l’Ouest (26%), l’Est (15,96%) et le Sud (8,96%)
(Registre Oran, Alger, Sétif, 2006).
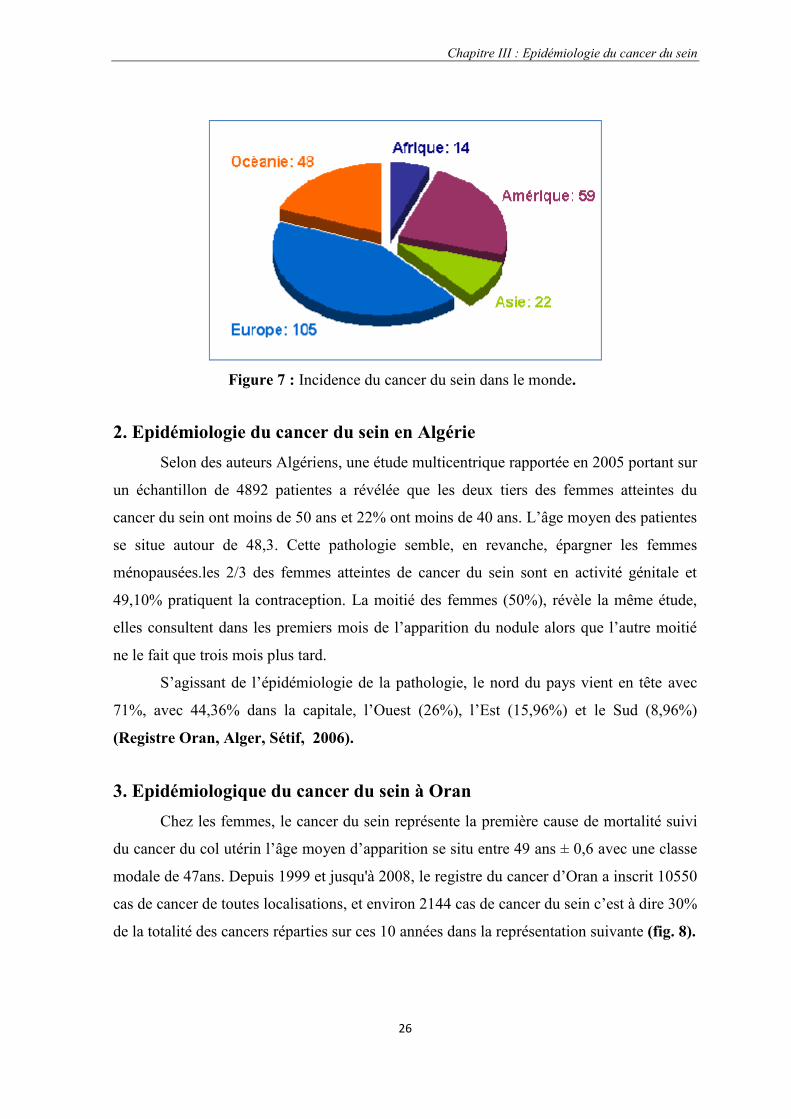
3. Epidémiologique du cancer du sein à OranChez les femmes, le cancer du sein représente la première cause de mortalité suivi
du cancer du col utérin l’âge moyen d’apparition se situ entre 49 ans ± 0,6 avec une classe
modale de 47ans. Depuis 1999 et jusqu'à 2008, le registre du cancer d’Oran a inscrit 10550
cas de cancer de toutes localisations, et environ 2144 cas de cancer du sein c’est à dire 30%
de la totalité des cancers réparties sur ces 10 années dans la représentation suivante (fig. 8).
Figure 7 : Incidence du cancer du sein dans le monde.
Chapitre III : Epidémiologie du cancer du sein
27
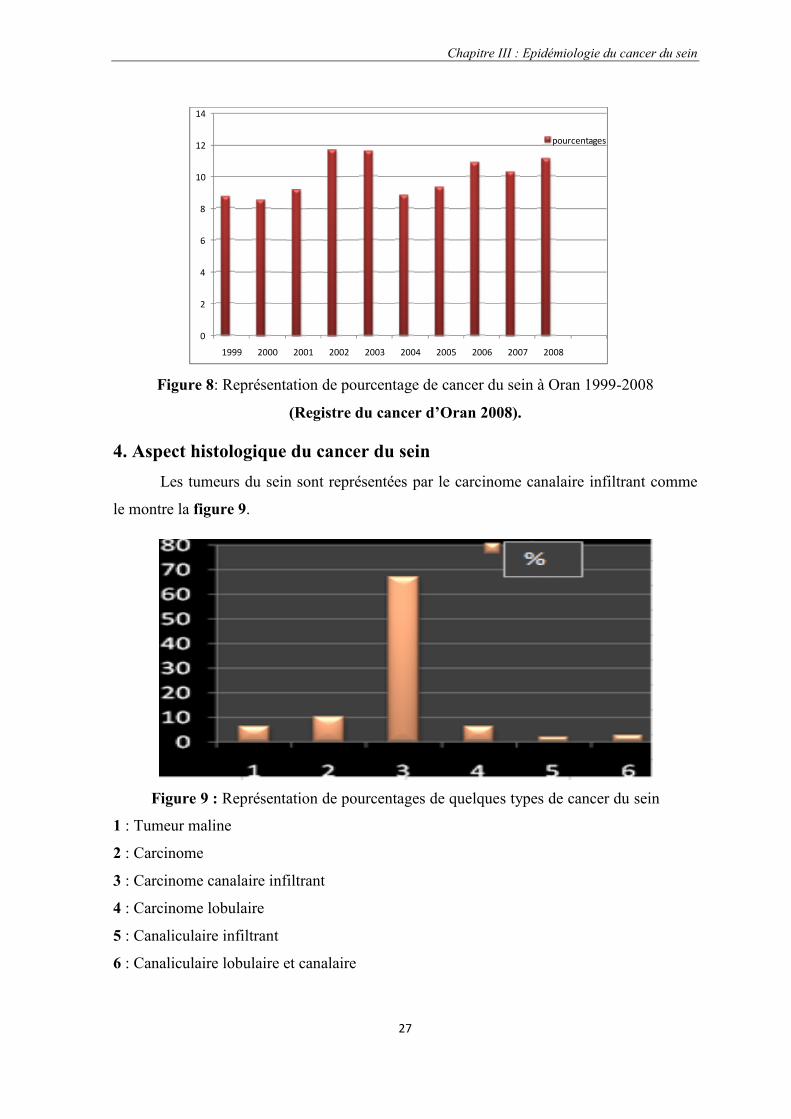
4. Aspect histologique du cancer du seinLes tumeurs du sein sont représentées par le carcinome canalaire infiltrant comme
le montre la figure 9.
1 : Tumeur maline
2 : Carcinome
3 : Carcinome canalaire infiltrant
4 : Carcinome lobulaire
5 : Canaliculaire infiltrant
6 : Canaliculaire lobulaire et canalaire
0
2
4
6
8
10
12
14
1999 2000 2001 2002 2003 2004 2005 2006 2007 2008
pourcentages
Figure 8: Représentation de pourcentage de cancer du sein à Oran 1999-2008
(Registre du cancer d’Oran 2008).
Figure 9 : Représentation de pourcentages de quelques types de cancer du sein
sein.
Chapitre III : Epidémiologie du cancer du sein
28
Le carcinome canalaire infiltrant domine les autres carcinomes avec un pourcentage
de 67,1% c’est à dire que environ 1438,1 des cas de cancer du sein sont des « carcinomes
canalaires infiltrants » (1999-2008), suivit par « les carcinomes» avec pourcentage de
10,1% environ 216,5 des cas, le carcinome lobulaire est de 6,2%.
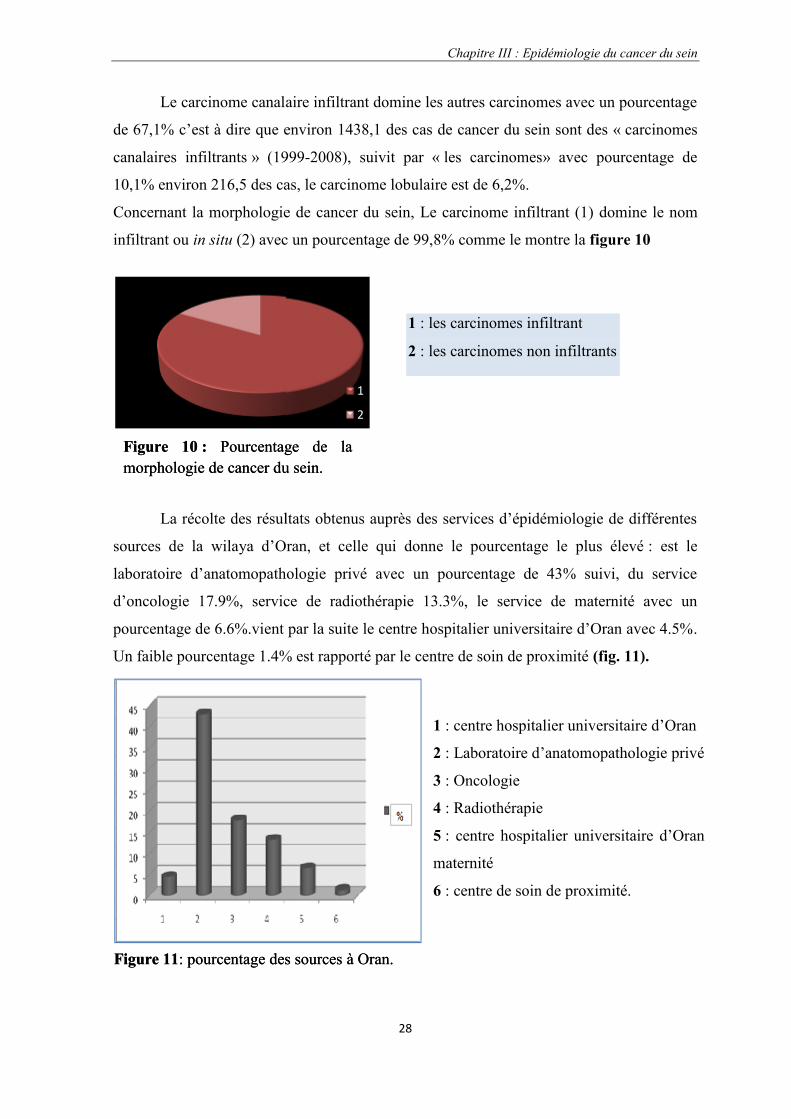
Concernant la morphologie de cancer du sein, Le carcinome infiltrant (1) domine le nom
infiltrant ou in situ (2) avec un pourcentage de 99,8% comme le montre la figure 10
La récolte des résultats obtenus auprès des services d’épidémiologie de différentes
sources de la wilaya d’Oran, et celle qui donne le pourcentage le plus élevé : est le
laboratoire d’anatomopathologie privé avec un pourcentage de 43% suivi, du service
d’oncologie 17.9%, service de radiothérapie 13.3%, le service de maternité avec un
pourcentage de 6.6%.vient par la suite le centre hospitalier universitaire d’Oran avec 4.5%.
Un faible pourcentage 1.4% est rapporté par le centre de soin de proximité (fig. 11).
1 : les carcinomes infiltrant
2 : les carcinomes non infiltrants
Figure 10 : Pourcentage de lamorphologie de cancer du sein.
1 : centre hospitalier universitaire d’Oran
2 : Laboratoire d’anatomopathologie privé
3 : Oncologie
4 : Radiothérapie
5 : centre hospitalier universitaire d’Oran
maternité
6 : centre de soin de proximité.
Figure 11: pourcentage des sources à Oran.
Chapitre III : Epidémiologie du cancer du sein
28
Le carcinome canalaire infiltrant domine les autres carcinomes avec un pourcentage
de 67,1% c’est à dire que environ 1438,1 des cas de cancer du sein sont des « carcinomes
canalaires infiltrants » (1999-2008), suivit par « les carcinomes» avec pourcentage de
10,1% environ 216,5 des cas, le carcinome lobulaire est de 6,2%.
Concernant la morphologie de cancer du sein, Le carcinome infiltrant (1) domine le nom
infiltrant ou in situ (2) avec un pourcentage de 99,8% comme le montre la figure 10
La récolte des résultats obtenus auprès des services d’épidémiologie de différentes
sources de la wilaya d’Oran, et celle qui donne le pourcentage le plus élevé : est le
laboratoire d’anatomopathologie privé avec un pourcentage de 43% suivi, du service
d’oncologie 17.9%, service de radiothérapie 13.3%, le service de maternité avec un
pourcentage de 6.6%.vient par la suite le centre hospitalier universitaire d’Oran avec 4.5%.
Un faible pourcentage 1.4% est rapporté par le centre de soin de proximité (fig. 11).
1
2
1 : les carcinomes infiltrant
2 : les carcinomes non infiltrants
Figure 10 : Pourcentage de lamorphologie de cancer du sein.
1 : centre hospitalier universitaire d’Oran
2 : Laboratoire d’anatomopathologie privé
3 : Oncologie
4 : Radiothérapie
5 : centre hospitalier universitaire d’Oran
maternité
6 : centre de soin de proximité.
Figure 11: pourcentage des sources à Oran.
Chapitre III : Epidémiologie du cancer du sein
28
Le carcinome canalaire infiltrant domine les autres carcinomes avec un pourcentage
de 67,1% c’est à dire que environ 1438,1 des cas de cancer du sein sont des « carcinomes
canalaires infiltrants » (1999-2008), suivit par « les carcinomes» avec pourcentage de
10,1% environ 216,5 des cas, le carcinome lobulaire est de 6,2%.
Concernant la morphologie de cancer du sein, Le carcinome infiltrant (1) domine le nom
infiltrant ou in situ (2) avec un pourcentage de 99,8% comme le montre la figure 10
La récolte des résultats obtenus auprès des services d’épidémiologie de différentes
sources de la wilaya d’Oran, et celle qui donne le pourcentage le plus élevé : est le
laboratoire d’anatomopathologie privé avec un pourcentage de 43% suivi, du service
d’oncologie 17.9%, service de radiothérapie 13.3%, le service de maternité avec un
pourcentage de 6.6%.vient par la suite le centre hospitalier universitaire d’Oran avec 4.5%.
Un faible pourcentage 1.4% est rapporté par le centre de soin de proximité (fig. 11).
1 : les carcinomes infiltrant
2 : les carcinomes non infiltrants
Figure 10 : Pourcentage de lamorphologie de cancer du sein.
1 : centre hospitalier universitaire d’Oran
2 : Laboratoire d’anatomopathologie privé
3 : Oncologie
4 : Radiothérapie
5 : centre hospitalier universitaire d’Oran
maternité
6 : centre de soin de proximité.
Figure 11: pourcentage des sources à Oran.

Chapitre III : Epidémiologie du cancer du sein
29
5. Facteurs de risques
Il n’existerait pas de facteur unique responsable de l’apparition d’un cancer du sein.
En réalité, plusieurs facteurs de risque susceptibles d’augmenter le développement de ce
type de cancer ont été mis en évidence : facteurs génétiques, environnementaux et
hormonaux….etc.
5.1. Facteurs génétiques
Seuls 5 à 10% des cas de cancesr sont liés à des anomalies génétiques
héréditaires. La probabilité qu’une femme de 30 ans, ayant une mère ou une sœur atteinte
d’un cancer du sein, développe cette pathologie avant l’âge de 70 ans est compris entre 7%
et 18% (Sakorafas et al, 2002). Plusieurs gènes impliqués dans la carcinogenèse
mammaire héréditaire ont été identifiés. Les principaux sont BRCA1, BRCA2, p53, PTEN
et ataxia-telangiectasia (AT). La grande majorité des cancers du sein héréditaires peuvent
être attribués aux gènes BRCA1 et BRCA2 (Dumitrescu et al, 2005). BRCA1 est localisé
sur le chromosome 17 et code une protéine nucléaire impliquée notamment dans le
contrôle de la recombinaison mitotique, dans la ségrégation des chromosomes, la
régulation transcriptionnelle ainsi que dans la réparation de l’ADN. Cette protéine agit
comme une ubiquitine ligase de type E3 dont les substrats sont certaines histones, la
tubuline γ, ERα, NPM1, la sous unité d’ARN polymérase II, l’isoforme A du PR et le
facteur TFIIE (Wu et al, 2008). BRCA2 est localisé sur le chromosome 13 et code une
histone acétyle transférase impliquée dans la régulation de la transcription avec une
fonction suppresseur de tumeur. Elle est impliquée dans la réparation de l’ADN et interagit
avec BRCA1 dans les voies d’activation de la protéine p53. Les femmes portant des
mutations de type délétion au niveau de BRCA1 ou BRCA2, présentent un risque
considérable de développer un cancer du sein (80%). Ce risque est environ dix fois
supérieur à celui de la population générale (Daly et al, 2005). Le gène p53 localisé sur le
chromosome 17 est un des gènes les plus communément mutés dans les cancers humains
(environs 50% des cancers). Les femmes affectées par une mutation de p53 présentent un
risque plus élevé de développer un cancer du sein avant l’âge de 45 ans. Enfin, les
mutations des gènes PTEN et AT, observées chez des patientes atteintes respectivement du
syndrome de Cowden et de l’ataxie télangiectasie, augmentent de 25 à 50% le risque de
cancérogenèse mammaire.

Chapitre III : Epidémiologie du cancer du sein
30
5.2. Facteurs environnementaux
Les variations d’incidence observées entre différents pays et zones
géographiques amènent à parler de risques environnementaux. Les radiations ionisantes
constituent le facteur de risque environnemental le mieux décrit. Les animaux de
laboratoire et les populations humaines ayant été exposés à de fortes doses de radiations
ionisantes présentent un taux de cancer du sein relativement élevé (Ronckers et al, 2004).
Un certain nombre de produits chimiques environnementaux peuvent augmenter le risque
de cancer du sein. C’est notamment le cas des organochlorés tels que le
Dichlorodiphényltrichloroéthane (DDT) ou les Polychlorobiphényls (PCB) qui ont
longtemps été utilisés comme pesticides. Les dioxines ou encore les solvants organiques
utilisés dans l’industrie sont également incriminés (Coyle et al, 2004). Le régime
alimentaire semble aussi avoir une importance dans l’apparition des cancers mammaires.
La surconsommation de protéines et de graisses animales a longtemps été incriminée or il
apparaît qu’un régime alimentaire riche en acide gras oléique trouvé dans l’huile d’olive
par exemple protège des cancers du sein (Wang et al, 2008). Par ailleurs, le taux de cancer
mammaire est plus élevé dans les contrées occidentales qu’au Japon ou autres pays d’Asie
moins consommateurs de graisses animales. Il est clairement démontré que les femmes
obèses post-ménopausées présentent un risque supérieur de développer un cancer
mammaire (Carmichael et al, 2004 ; Dumitrescu et al, 2005). La consommation d’alcool
ainsi qu’une consommation active ou passive de tabac augmente également ce risque
(Morabia et al, 1996 ; Johnson et al, 2000 ; Thygesen et al. 2008). Enfin, la pratique
d’une activité physique diminue le risque de développer un cancer du sein chez les femmes
pré et post-ménopausées (Maruti et al, 2008).
5.3. Facteurs hormonaux
Certaines hormones stéroïdes jouent un rôle important dans le développement
de la glande mammaire. Elles jouent aussi un rôle important en tant que facteur de risque
du cancer du sein. Les carcinomes mammaires hormono-dépendants s'observent chez
approximativement 60% des patientes pré-ménopausées et chez 75% des patientes
ménopausées. On distingue les hormones endogènes (œstrogènes et progestérone) des
hormones exogènes pouvant être apportées par les contraceptifs oraux, les traitements
hormonaux substitutifs ou encore les xénooestrogènes.

Chapitre III : Epidémiologie du cancer du sein
31
5.3.1. Hormones endogènes
Le risque semble dépendre essentiellement du temps d'exposition de
l'épithélium mammaire à ces hormones. De nombreuses variables, incluant l'âge des
premières règles, celui de la première grossesse et de la ménopause ainsi que le nombre de
grossesses, sont à prendre en compte. Chez les femmes pré-ménopausées, les œstrogènes
sous forme d’œstradiol sont majoritairement produits par les ovaires. Après la ménopause,
les œstrogènes continuent à être produits au niveau du tissu adipeux notamment dans la
glande mammaire. Une ménarche précoce (avant 12 ans) augmente de 10 à 30 % le risque
de cancer mammaire. De même, une ménopause tardive élève le risque de 3% pour chaque
année dépassant l'âge moyen de la ménopause. Ces résultats s'expliquent par
l'augmentation, chez ces femmes, du nombre de cycles ovulatoires. A l'inverse, des
grossesses précoces ou nombreuses et des lactations prolongées, diminuent le risque de
cancer du sein (Dumitrescu et al, 2005; Sakorafas et al, 2002). D'une manière générale,
la concentration et le type d'hormones présentes dans le sérum influencent le risque de
carcinogenèse mammaire. Ainsi, une augmentation du taux d'œstrone, d'œstradiol et/ou de
testostérone est corrélée à une augmentation du risque de cancer du sein (Feigelson et al,
1996; Key et al, 2002).
La progestérone agit par l’intermédiaire de son récepteur (PR), dont il existe
deux isoformes (PR A et PR B), transcrites à partir de deux promoteurs différents d’un
même gène. Le rôle des isoformes est mal connu dans la tumeur mammaire mais il apparaît
que l’augmentation du rapport A/B induit des gènes impliqués dans l’adhésion cellulaire
aboutissant à la migration des cellules cancéreuses vers la moelle osseuse ce qui
correspond à un phénotype plus agressif (Jacobsen et al., 2002) Mais une étude
contradictoire a rapporté que la progestérone n'augmenterait pas ce risque et pourrait avoir
un effet préventif (Campagnoli et al., 2005).

Chapitre III : Epidémiologie du cancer du sein
32
5.3.2. Hormones exogènes
Les traitements hormonaux substitutifs et la contraception orale sont les deux
sources d'hormones exogènes les plus étudiées.
Les contraceptifs oraux contiennent généralement un œstrogène et de fortes
doses de progestérone. Les résultats, même s'ils sont pour la plupart controversés, montrent
qu'ils n'augmentent que modestement le risque de cancer du sein. Ce risque varie selon
l'âge d'ingestion des premiers contraceptifs et devient nul environ 10 ans après l'arrêt du
traitement (Henderson et al, 2000; Dumitrescu et al, 2005).
En France, les traitements hormonaux substitutifs les plus couramment
prescrits combinent des œstrogènes, généralement administrés sous forme de patchs ou de
gels, à de la progestérone micronisée ou à des progestatifs de synthèse. Par ailleurs, les
autorités sanitaires recommandent désormais un traitement hormonal substitutif de courte
durée. L'examen des taux de cancer selon le type d'hormones suggère qu'avec l'œstrogène
utilisé seul (traitement réservé aux femmes hystérectomisées), le sur-risque de développer
un cancer du sein est minime voire inexistant. En revanche, lorsqu'il est associé à un
progestatif de synthèse, le risque de cancer du sein est augmenté de 40 % et ce,
indépendamment de la voie d'administration de l'œstrogène. La combinaison œstrogène et
progestérone micronisée semble dépourvue d'effet cancérigène, tout du moins à court
terme (Fournier et al, 2005). Les xénooestrogènes sont des substances chimiques,
d'origine naturelle ou synthétique, capables de se lier aux récepteurs des œstrogènes (ER)
et d'induire ou moduler une réponse relayée par ces récepteurs. Ces xénooestrogènes ou
œstrogène-mimétiques proviennent essentiellement de l'industrie, de l'agriculture et des
rejets urbains. Les lignées de tumeurs mammaires MCF-7 et T47D, sensibles à l’œstradiol,
se multiplient davantage en présence de ces molécules (Soto et al, 1995; Lascombes et al,
2000). Ces résultats suggèrent que ces molécules peuvent avoir un impact non négligeable
sur l'incidence des cancers mammaires hormono-dépendants.

Chapitre III : Epidémiologie du cancer du sein
33
5.4. Facteurs pronostiques et survie
Un grand nombre de facteurs interviennent dans l'établissement d'un pronostic
de survie. L'âge joue un rôle très important : en absence de métastases, la survie à 5 et 10
ans d’une patiente est respectivement de 67% et 52% lorsqu’elle a moins de 35 ans, de
79% et 63% entre 35 et 65 ans ; pour les femmes de plus de 75 ans la survie passe à 51% et
25%. L'évolution de la maladie est d'autant plus sévère que la taille de la tumeur est
volumineuse. Le taux de survie à 5 ans est de 99 % si la tumeur est de 1 cm ou moins, alors
qu'il n'est plus que de 86 % pour une taille comprise entre 3 et 5 cm (Donegan et al, 1997).
L'envahissement ganglionnaire axillaire est le meilleur indicateur de survie des patientes
atteintes d’un cancer du sein. Le nombre de ganglions infectés et leur niveau
d'envahissement suffisent à prédirent le taux de survie. En effet, le taux de survie à 5 ans
est de 73 % pour des patientes ayant de 1 à 3 ganglions infectés, de 45,7 % pour 4 à 12
ganglions et de 28,4 % pour 13 ganglions ou plus. Les formes d'emblée métastatiques ont
un très mauvais pronostique à 5 et 10 ans : respectivement 15 % et 5 % (Cianfrocca et al,
2004).
Enfin, la présence des récepteurs hormonaux a aussi valeur de facteur
pronostique. La survie à 5 ans pour un cancer du sein non métastatique est de 74 % en
présence de récepteurs des œstrogènes α et de 66 % en leur absence. Pour les récepteurs à
la progestérone, la survie à 5 ans est de 81 % en leur présence et de 73 % en leur absence.
Ces facteurs sont plutôt favorables, parce que la présence de ces récepteurs est indicateur
de la sensibilité des cellules tumorales à l'hormonothérapie d'une part, et car de
nombreuses études montrent que les récepteurs des œstrogènes diminuent les capacités
invasives des cellules tumorales d'autre part. Le grade histologique est aussi un facteur
pronostic important. Il est basé sur la différenciation glandulaire, le pléïomorphisme
nucléaire et le nombre de mitoses par champ.

Chapitre IV : Virus et cancer du sein
34
Récemment, des virus ont été évoqués comme étant des facteurs de risque dans la
progression tumorale.
1. Virus et cancer du sein
1.1. MMTV (Mouse Mammary Tumor Virus)Ce virus est associé à une forte incidence de cancers du sein chez certaines races de
souris et agit par mutation insertionnelle. Le rôle d'un rétrovirus dans le cancer du sein, a
donc été soupçonné mais les séquences de rétrovirus endogènes déjà insérées dans le
génome humain peuvent interférer dans la recherche de séquences d'un autre virus. Wang
et al. 1995 ont mis au point une PCR qui détecte une séquence codant pour les protéines de
l'enveloppe qui possède de fortes homologies de séquence avec le MMTV murin mais pas
avec les rétrovirus endogènes. Cette séquence a été détectée par PCR dans environ 40%
des cancers du sein testés (Wang et al., 1995). L'expression de cette séquence a été mise
en évidence par RT-PCR dans 66% des tumeurs « hMMTV » positives (Wang et al.,
1998).
1.2. HPV (Les papillomavirus humains)De type 16 ou 18 sont associés au développement des cancers de l'utérus et peuvent
induire l'immortalisation de cellules épithéliales mammaires normales en culture (Band et
al., 1990). La présence de séquence HPV 16 et 18 a été recherchée par PCR dans des
tumeurs du sein inclues en paraffine (Di lonardo et al., 1992 ; Bratthauer et al., 1992 ;
Wrede et al., 1992). Une seule étude a retrouvé la présence de HPV dans environ 30% des
cas mais l'ADN viral n'a pu être détecté par HIS (Di Lonardo et al. 1992). Plus récemment
des séquences de HPV 16 ont été mises en évidence par PCR dans des tumeurs du sein de
femmes traitées pour des cancers de l'utérus de haut grade (CIN III). Il a été montré que le
CIN III devait précéder le cancer du sein pour que l'on puisse détecter des séquences HPV
dans les carcinomes mammaires (Hennig et al., 1999). A l'heure actuelle, aucun de ces
virus n'est clairement associé au cancer du sein. Pour HPV, les résultats de PCR divergent
et la présence du virus in situ n'a été montrée que dans un seul cas. Pour le « MMTV
humain », peu d'études ont été publiées pour permettre une conclusion précise.

Chapitre IV : Virus et cancer du sein
35
1.3. EBV (Epstein-Barr)L’EBV a été détecté dans le lait maternel en 1991 (Junker et al, 1991). Par ailleurs,
l'EBV a été mis en évidence par des techniques de PCR conventionnelle dans 20% à 50%
des cancers du sein (Labrecque et al, 1995 ; Luqmani et al, 1995; Bonnet et al, 1999;
Fina et al, 2001). La charge virale a été quantifiée par Q-PCR dans des biopsies de cancer
du sein et des cellules tumorales isolées par microdissection (Arbach et al., 2006). Les
résultats obtenus révèlent un faible nombre de copies, ce qui explique la difficulté à mettre
en évidence la présence du virus. De plus, la distribution du génome viral est très
hétérogène au sein de la même tumeur et d’une tumeur à l’autre. D’autres parts, il a été
détecté par RT-PCR dans les tumeurs EBV positives, des transcrits de gènes latents :
EBNA1 a été retrouvé dans 80% des cas et les transcrits de deux oncogènes viraux LMP1
et BARF1 ont été détectés respectivement dans 20% et 57% des cas. Les résultats obtenus
en utilisant les microdissections confirment ainsi la présence de l’EBV dans les cellules
tumorales mammaires et la détection des transcrits de gènes latents montre que le génome
viral est sous forme latente. Par ailleurs, la possibilité selon laquelle le virus contribuerait à
la progression de cette tumeur a été suggérée (Arbach et al., 2006).
1.3.1. La famille des herpesviridae et le virus d’Epstein-Barr
1.3.1.1. La famille des herpesviridaeLa famille des Herpesviridae est divisée en trois grands sous-groupes (α, β et γ) en
fonction de critères basés sur leur tropisme in vivo, et sur la durée de leur cycle lytique. Les
herpèsvirus qui possèdent une étroite spécificité d’espèce, sont très largement répandus. En
effet, la majorité des espèces animales peuvent être infectées au moins par l’un de ces
virus. Une des particularités de ces virus est qu’ils sont responsables d’infections dites
latentes et persistent tout au long de la vie de l’individu infecté. Si les primo-infections
sont souvent asymptomatiques, les herpèsvirus sont cependant associés à de graves
pathologies, notamment chez les sujets immunodéprimés. Actuellement, huit virus herpès
humains (HHV) sont identifiés et associés à différentes pathologies. Les virus de cette
famille ont la propriété d’établir une infection latente leur permettant de persister à long
terme dans les cellules « réservoirs » de leur hôte. La phase de latence peut être
interrompue (phénomène de réactivation) pour produire de nouveaux virus infectieux,
entrée dans le cycle lytique. Le cycle lytique est composé d’une succession ordonnée de
phases de transcription/traduction. Les phases précoce et tardive sont respectivement

Chapitre IV : Virus et cancer du sein
36
définies comme les phases de transcription/traduction avant et après la réplication du
génome viral (Kieff, 1996).
1.3.1.2 Historique
En 1958, Denis Burkitt, un chirurgien anglais travaillant en Ouganda, a décrit un
Cancer commun affectant des enfants en Afrique équatoriale (Burkitt, 1958). La
distribution géographique du lymphome de Burkitt (LB) et les conditions climatiques
associées suggèrent qu'un agent infectieux pourrait en être responsable (Burkitt, 1962). En
1964, Epstein, Achong et Barr ont identifié des particules virales par microscopie
électronique dans des lignées cellulaires provenant d’une biopsie de lymphome de Burkitt.
Ces particules virales, faisant partie de la famille des herpèsvirus, ont été dénommées virus
d’Epstein-Barr (EBV) (Epstein et al., 1964). Au milieu des années 60, des études séro-
épidémiologiques ont montré que les sérums des patients atteints de lymphome de Burkitt
avaient des taux sériques plus élevés d'anticorps dirigés contre les antigènes de l’EBV que
des sérums contrôles (Henle, 1966). Ces analyses sérologiques ont également identifié
l’EBV comme agent étiologique de la mononucléose infectieuse (MNI). Ces analyses ont
démontré que l'infection par l'EBV était associée à une autre maladie maligne, le
carcinome indifférencié du nasopharynx (CNP) (Henle et al, 1968; zur Hausenw et al,
1970), confirmant la relation de ce virus à des cancers chez l’homme (Henle et Henle,
1970). A peu près à la même époque, la capacité de transformation de l'EBV a été
confortée par la potentialité du virus de transformer efficacement, in vitro, des
lymphocytes B au repos et d'induire des tumeurs chez les primates non humains (Henle et
al, 1967; Miller, 1974; Pope et al, 1968).
Les études faites dans les années 80 ont prouvé que l'EBV est associé à de
nombreuses variétés de tumeurs humaines comprenant des maladies des lymphocytes B
telles que la maladie de Hodgkin (HD) et les maladies lymphoprolifératives surgissant
après immunosuppression des patients sidéens, des lymphomes T, et des tumeurs
épithéliales (NPC et cancers gastriques). Toutes ces tumeurs sont caractérisées, d’une part,
par la présence de copies multiples extra-chromosomiques du génome viral circulaire
épisomal dans les cellules tumorales, et d’autre part, par l’expression des gènes de latence
codés par l’EBV, qui semblent contribuer au phénotype malin (Rickinson et Kieff, 2001).

Chapitre IV : Virus et cancer du sein
37
1.3.1.3. Structure du virusLes virus de la famille des herpesviridae sont enveloppés. L’enveloppe qui dérive
de la membrane cellulaire, contient un nombre de glycoprotéines virales variable d’un
virus à l’autre. L’EBV présente à sa surface plusieurs glycoprotéines d’enveloppe dont la
gp 350/220, les gp 85/25 et la gp 42 nécessaires à l’infection. En plus de l’enveloppe, le
virus est constitué : du tégument qui correspond à une structure protéique fibreuse, d’une
capside protéique icosaédrique d’environ 100 nm de diamètre qui comprend 162
capsomères, et d’un core qui correspond à la structure contenant le génome viral.
1.3.1.4. Génome viral de l’EBVEn 1984, le génome de l’EBV a été entièrement séquencé à partir de la souche B95-
8, lymphocytes B de marmouset infectés par l’EBV en primo-infection (Baer et al., 1984).
Dans le virus, la molécule d’ADN double brin est sous forme linéaire (fig. 12) et composée
d’environ 172 kpb. Dans la cellule infectée, le génome viral est dans la majorité des cas,
sous forme épisomale (fig. 13) et rarement intégré (Rickinson et Kieff, 1996). Des
répétitions internes en tandem, « internal repeat » (IR1) s’intercalent entre deux régions
uniques : « unique short » (US) et « unique long » (UL). D’autres séquences répétées
courtes (IR2, 3 et 4) sont présentes dans la séquence unique UL. La digestion par l’enzyme
de restriction BamHI a permis de cartographier le génome viral de la souche de référence
B95-8 en fragments de restriction qui ont été classés de A à Z et de a à h en fonction de
leur taille. Plus d’une centaine de phases ouvertes de lecture (ORF pour « Open Reading
Frame ») ont pu être mises en évidence à partir de l’analyse de séquences. Elles sont
répertoriées selon : (i) le fragment de restriction BamHI auquel elles appartiennent, (ii) le
sens de transcription (L pour « Leftward » vers la gauche et R pour « Rightward » vers la
droite), (iii) leur ordre dans le sens de transcription de ce fragment. Par exemple, BNLF1
correspond à une ORF présente dans le fragment de restriction BamHI N (BN), elle est
transcrite vers la gauche (L) et il s’agit de la première ORF situé dans BamHI N (F1). Sous
sa forme linéaire, le génome viral est encadré par deux régions répétées directes, les
répétitions terminales (TR) constituées d’un nombre variable de motifs de 500 pb suivant
les souches virales. Les TR permettent au génome viral de se circulariser. Grâce au nombre
de motifs dans les TR, il est possible de déterminer si l’infection virale est monoclonale ou
polyclonale dans les différentes pathologies EBV positives (Raab-Traub et Flynn, 1986).
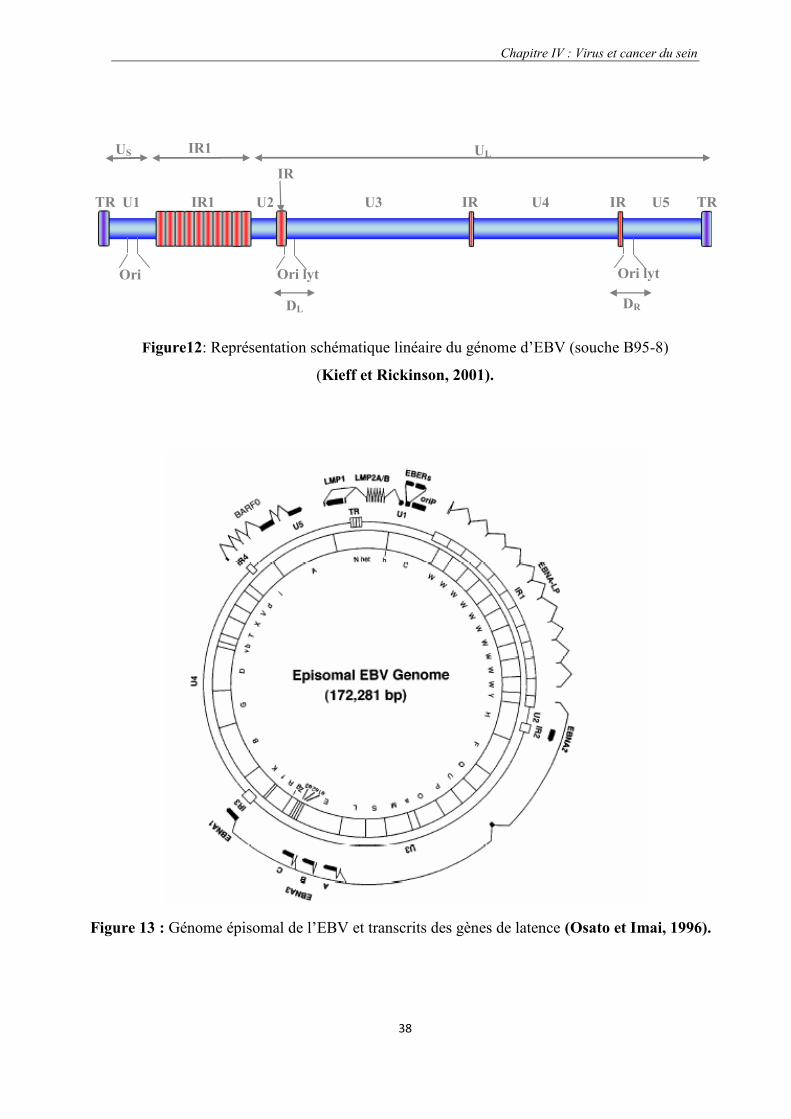
Chapitre IV : Virus et cancer du sein
38
US IR1 UL
DRDL
Ori lytOri lytOrip
U1TR TRIR1
IR2
IR3
IR4
U2 U3 U4 U5
Figure12: Représentation schématique linéaire du génome d’EBV (souche B95-8)
(Kieff et Rickinson, 2001).
Figure 13 : Génome épisomal de l’EBV et transcrits des gènes de latence (Osato et Imai, 1996).

Chapitre IV : Virus et cancer du sein
39
1.3.1.5. Polymorphismes de l’EBVL’EBV possède lui-même, un polymorphisme. En effet, il a été décrit différentes
souches virales de l’EBV, qui peuvent être classées en deux types (EBV-1 et EBV-2 ou
EBVAµ et EBV-B).Ces deux souches virales se distinguent par des polymorphismes
génomique, au niveau des gènes de latence comme BYRF1 codant EBNA2 (EBV Nuclear
Antigens 2), EBNA3A, 3B, 3C, permettant de les distinguer et de les identifier. Des études
de séroprévalence ont montré que le type 1 est majoritairement prévalent en Occident,
alors qu’en Afrique et en nouvelle Guinée, les types 1 et 2 présentent la même fréquence
d’apparition. De plus, il existe entre les différents isolats d’EBV un polymorphisme au
niveau de certaines séquences du génome viral. En effet, il peut y avoir une variation du
nombre des séquences en tandem dans les régions TRs de son génome. Enfin, pour une
même souche virale de type A ou B, un autre polymorphisme s'observe également au
niveau des régions IRs, ainsi qu’au niveau de plusieurs gènes dont BYRF1, BZLF1 (codant
Zta) et BNLF1 (Grunewald et al, 1998).
1.3.1.6. Les voies de transmissionLa transmission de l’EBV se fait essentiellement par contact direct avec la salive
(d’où l’appellation « maladie du baiser »). Ce mode de transmission peut expliquer le
caractère ubiquitaire et précoce de l’infection par l’EBV : 100% des enfants sont infectés
avant l’âge de deux ans en Afrique intertropicale alors que dans les pays développés,
l’infection peut survenir dans l’adolescence ou encore à l’âge adulte (Henle et al, 1968).
Les contaminations par transfusion sanguine (Turner et al, 1972), voies sexuelles
(Portnoy el al, 1984) (S Fafi-Kremer et al, 2007) ou lors de transplantations d’organes
(Gratama et al, 1988) ont été rapportées. La présence de l’EBV dans le lait maternel a
également était constaté (Junker et al, 1991).
1.4. Tropisme cellulaire
1.4.1. In vivoL’EBV infecte principalement les lymphocytes B matures naïfs quiescents en
induisant dans ces cellules une latence de type III. Les lymphoblastes infectés gagnent les
follicules lymphoïdes et subissent via l’expression des gènes latents du virus une
différentiation en lymphocytes B (LB) mémoires circulants. Il a été montré que chez les
porteurs sains, 1 à 10 lymphocytes circulants par million restent infectés par l’EBV
(Miyashita et al., 1997).

Chapitre IV : Virus et cancer du sein
40
Dans ces cellules, l’EBV trouve l’environnement idéal pour persister dans
l’organisme (d’où la notion de cellules réservoirs) car les lymphocytes B ont une durée de
vie très longue et n’expriment aucun antigènes cibles de lymphocytes T cytotoxiques. De
plus, l’EBV exprime la protéine nucléaire EBNA1, assurant la partition du génome viral
dans les cellules divisées (Thorley-Lawson and Gross, 2004). Les cellules T et les
cellules épithéliales peuvent aussi constituer des cibles pour l’EBV. En effet, le génome
viral a été détecté dans les cellules tumorales de certains lymphomes T (d'Amore et al,
1996), dans les carcinomes du nasopharynx (NPC) (Martel-Renoir et al, 1995), dans
certains cas de carcinomes gastriques (Tokunaga et al, 1993), d’hépatocarcinomes (Li W
et al, 2004) et de carcinomes mammaires (Arbach et al, 2006).
1.4.2. In vitroDu fait de son pouvoir d’immortalisation sur les lymphocytes B en culture, la
biologie de l’EBV repose essentiellement sur l’étude des lignées B. Les lignées dérivées de
lymphome de Burkitt EBV positifs (BLCL), les lymphocytes B humains infectés in vitro
par l’EBV (LCL), ainsi qu’une lignée de lymphocytes B de singe de marmouset infectés in
vitro (lignée de référence B95-8) sont couramment utilisés en laboratoire. L’inoculation
des LCL dans la souris SCID conduit au développement de tumeurs dont les
caractéristiques morphologiques se rapprochent des lymphoproliférations polyclonales B
des immunodéprimés (Young et al, 1989). Dans ces modèles cellulaires, le virus est
majoritairement à l’état latent. En revanche, dans 2 à 5% des cellules, le virus peut entrer
spontanément en cycle réplicatif.
Des lignées de cellules épithéliales ont également été établies à partir de biopsies de NPC
mais le virus est perdu au cours des passages (Yao et al, 1990). En revanche, des lignées
EBV postives ont été obtenues à partir de tumeurs NPC transplantées chez la souris nude
(Busson et al, 1988), et une culture infectée de façon stable a été établie à partir de
xénogreffes de NPC (Cheung et al, 1999). Par ailleurs, des lignées infectées ont été établie
à partir de tissu gastrique (Tajima et al, 1998). L’infection des cellules épithéliales par un
EBV recombinant porteur d’un gène de sélection (résistance à la néomycine), a permis
d’obtenir des clones EBV positifs. Cette infection est plus efficace quand les cellules
donneuses de virus et les cellules épithéliales sont en contact direct (Imai et al, 1998).
Grâce à ce système, des lignées épithéliales NPC (Chang et al, 1999), de carcinomes
gastriques (Nishikawa et al, 1999), d’hépatocarcinomes (Imai et al, 1998),des

Chapitre IV : Virus et cancer du sein
41
kératinocytes (Chang et al, 1999) ainsi que des cellules épithéliales mammaires (Bonnet-
Duquennoy et al, 2003) ont été infectées par l’EBV
1.5. Cycle biologique du virus d’Epstein-Barr
1.5.1. Entrée du virus dans les cellules ciblesL’infection par l’EBV des lymphocytes B est initiée par l’attachement et la fixation
du virus sur les lymphocytes B possédant à leur surface le CD21 (également connu sous le
nom de CR2). Ce récepteur est exprimé de façon ubiquitaire à la surface cellulaire de tous
les lymphocytes B et appartient à la super famille des immunoglobulines. L’ancrage de
l’EBV sur les lymphocytes B se fait par liaison de la glycoprotéine gp350/220 qui domine
l’enveloppe virale externe, au CD21 (Tanner et al, 1987). Après fusion de l’enveloppe
virale avec la membrane plasmique de la vésicule d’endocytose, la nucléocapside rentre
dans le cytosol. Cette pénétration du virus requiert un complexe de trois glycoprotéines
virales : gp25, gp85, et gp42. Les deux premières glycoprotéines sont nécessaires pour la
fusion de l’enveloppe virale avec la membrane cellulaire des lymphocytes B alors que la
gp42, considérée comme un co-récepteur, se lie aux molécules du complexe majeur
d’histocompatibilité de type II (CMH-II) et intervient dans la pénétration du virus dans la
cellule. Après décapsidation, le génome viral migre sous forme linéaire vers le noyau
(Wang et al, 1998; Haan et al, 2000), où il se circularise pour établir un état de latence.
Plusieurs études ont montré que le mécanisme d’infection des cellules épithéliales
est différent des lymphocytes B. En effet, certaines cellules épithéliales n’exprimant pas le
CD21, les glycoprotéines gp350/220 et gp42 ne sont pas nécessaire à l’infection par l’EBV
(Li Q et al, 1997). L’attachement de l’EBV aux cellules épithéliales se fait alors par
interaction du complexe gp25/gp85 avec une molécule de surface qui a une fonction
comparable au CMH-II. L’infection par l’EBV des cellules épithéliales est également
possible par phagocytose de corps apoptotiques contenant plusieurs copies du génome
d’EBV (Holmgren et al, 1999). La pénétration du virus dans les cellules épithéliales peut,
par ailleurs, se produire par contact direct entre les cellules donneuses de virus et les
cellules épithéliales (Imai et al, 1998), par transfert direct entre différentes lignées de
cellules épithéliales (Imai et al, 1998) ou par voie de transcytose des Ig A in vitro (Sixbey
et al, 1992).

Chapitre IV : Virus et cancer du sein
42
1.5.2. Latence et maintien du génomeIn vitro, il est possible d’immortaliser des lymphocytes B par l’EBV et d’obtenir
des LCLs. Les lymphocytes B infectés sont très peu permissifs à la réplication virale (cycle
lytique) et le virus peut s'y maintenir dans un état dit « de latence ». Ce statut de latence
virale repose sur deux phénomènes interdépendants. Le premier est la répression du cycle
lytique, ainsi que le maintien du génome viral en situation intranucléaire sous forme
épisomale circularisée. Le deuxième, essentiel au maintien de la latence virale, est
l'expression des gènes dits de latence. Ces gènes codent pour des protéines qui ont des
fonctions diverses dont celle d'immortaliser les lymphocytes B infectés. On compte
plusieurs gènes de latence, regroupés en 3 principales familles :
la famille des gènes codant pour les protéines nucléaires EBNAs.
la famille des gènes codant pour les protéines membranaires LMPs
la famille des gènes codant pour les ARNs EBERs, BARTs et les microARN
(miARN).
L’expression des protéines de latence contribue aux propriétés d'immortalisation
et/ou de transformation de l’EBV en influençant la «machinerie» cellulaire afin de
contrôler la croissance et/ou la survie des cellules infectées. Trois promoteurs de latence
virale sont décrits : Wp, Cp et Qp. Le profil d'expression des gènes de latence n'est pas
identique dans tous les types cellulaires infectés par l’EBV. On distingue ainsi 4 types de
latence en fonction des différents gènes exprimés. Dans chacune d’elle, les ARNs EBER1,
EBER2 et BARTs sont présents.
1.5.2.1. Les différents types de latences virales :
A- La latence de type I
Retrouvée dans des lignées de lymphomes de Burkitt et se caractérise par la seule
expression d’EBNA1. Les cellules en latence I possèdent un phénotype cellulaire
particulier avec l’expression à leur surface des marqueurs CD10 et CD77 et presque pas ou
peu d'antigènes d'activation cellulaire et de molécules d'adhésion (Cirone et al, 1995;
Gregory et al, 1990). Toutefois, in vitro, la latence I dans certaines lignées de lymphomes
de Burkitt maintenues en culture n’est pas stable. En effet, on peut constater un
changement du profil d'expression des gènes de latence dans ces cellules, ainsi que du
phénotype cellulaire aboutissant à une latence de type III (Cirone et al, 1995; Gregory et
al, 1990).

Chapitre IV : Virus et cancer du sein
43
B- La latence de type II
Caractéristique du carcinome indifférencié du nasopharynx, de la maladie de Hodgkin et
des lymphomes T. Elle se manifeste par l'expression des protéines EBNA1, LMP1,
LMP2A et LMP2B.
C- La latence de type III
Est décrite dans les lymphocytes B infectés in vitro par l’EBV (LCL) et dans
les syndromes lymphoprolifératifs des sujets immunodéprimés après transplantation
d’organes (PTLD, Post-Transplant Lymphoprolifératives Disease). Elle est caractérisée
par l’expression de la totalité des gènes de latence (EBNA-1, EBNA-2, EBNA-3A,
EBNA-3B, EBNA-3C, EBNA-LP, LMP-1, LMP-2A et LMP-2B) à partir des
promoteurs Wp puis Cp. Au cours de cette latence, le promoteur Qp n’est cependant
pas actif. Les cellules expriment différents marqueurs de d’activation (CD21, CD23,
CD30, CD39, CD70) et de marqueurs d’adhésion (CD11a, CD54, CD58). Ces trois
types de latence sont typiquement retrouvés dans certaines pathologies malignes
associées à l’EBV. (Thèse Tan-Southéa Ouk, 2008)
1.5.2.2. Les protéines de latence : description, régulation et fonction des protéines de
latence :
Les protéines de latence dérégulent un certain nombre de fonctions cellulaires en
détournant la régulation transcriptionnelle et différentes voies de signalisation cellulaire,
aboutissant à l’immortalisation de la cellule-hôte.
A- Les promoteurs des protéines de latence
Trois promoteurs sont particulièrement décrits: Cp, Wp et Qp, respectivement situé dans
les fragments BamH1 C, W et Q. Les promoteurs Cp et Wp contrôlent l’expression des
protéines EBNAs, alors que le promoteur Qp contrôle l’expression de la protéine EBNA-1
en absence d’EBNA-2. EBNA-2 est une protéine essentielle à l’immortalisation in vitro
des lymphocytes et n’est exprimée qu’au cours de la latence virale de type III.
La transcription des gènes codant les EBNAs par le promoteur Cp semble restreinte aux
lymphocytes B alors que celle par le promoteur Wp est ubiquitaire quel que soit le type
cellulaire infecté (Contreras-Brodin C et al.,1996). Les promoteurs Cp et Wp sont
exclusifs l’un l’autre (Woisetschlaeger M et al., 1989). Aux stades initiaux de l’infection
virale, Wp permet l’expression d’EBNA-LP, EBNA2 et EBNA1, alors que Cp est réprimé
par méthylation. Ensuite, après la répression de Wp par méthylation des motifs CpG, Cp

Chapitre IV : Virus et cancer du sein
44
prend le relais de la transcription des gènes de latence (Woisetschlaeger M et al., 1990 ;
Schlager S et al., 1996).
La répression du promoteur Wp dépendrait d’EBNA-2 et d’EBNA-LP puisque lors de
l’infection par des virus déficients pour le gène codant EBNA-2 et/ou ayant une mutation
sur le gène codant EBNA-LP, il n’y avait plus de switch entre Wp et Cp (Jin XW et
al.,1992).
Le promoteur Qp est dépourvu d’une boîte minimale de type TATA. Il a été également
montré que Qp est hypométhylé et constitutivement activé quelles que soient les tumeurs et
les types cellulaires étudiés. (Tao Q et al,1998)
1.5.2.3. La famille des Antigènes Nucléaires de l’EBV, les protéines EBNAs :
A- EBNA-LP :
La protéine EBNA-LP (ou EBNA-5) est la première des protéines EBNAs à
être exprimée à partir du promoteur Wp et joue un rôle important dans l’immortalisation in
vitro des lymphocytes B par l’EBV (MannickJB et al, 1991). Elle est composée d’une
partie amino-terminale (Nterm) constituée de tandems répétés codés par deux exons W1
(22 aas) et W2 (44 aas) dérivés de la région IR1 et d’une partie carboxy-terminale (Cterm)
de 45 aas codés deux exons uniques Y1 et Y2 (Sample J et al, 1986 ; Ling PD et al,
2005).
EBNA-LP est essentiellement nucléaire et est retrouvé colocalisé à des
structures particulières du noyau, les corps nucléaires PML (ProMyelocytic et al, 1996).
EBNA-LP possède, au sein des exons W2 et des régions conservées CR1 et CR2
(Conserved Region 1 et 2),
Des séquences basiques de localisation nucléaire NLS (Nuclear Localization
Signal, (Peng R et al, 2000 ; McCann EM et al, 2001). Cependant certaines études ont
retrouvé EBNA-LP colocalisé à des protéines cytoplasmiques (Kawaguchi Y et al, 2000 ;
Matsuda G et al, MicrobiolImmunol 2003 ; Forsman A et al, JProteomeRes 2008).
Une des principales fonctions d’EBNA-LP est de participer à l’activation de
l’expression des protéines membranaires de latence, dont LMP-1, par EBNA-2 et du
promoteur Cp (Nitsche F et al, 1997 ; Harada S et al, 1997 ; Peng R et al, 2000).
L’interaction entre EBNA-LP et EBNA-2 grâce aux domaines W1 et W2 est régulée par

Chapitre IV : Virus et cancer du sein
45
phosphorylation au niveau de résidus sérine S35 et S63 (Yokoyama A et al, 2001 ;
McCann EM et al, 2001).
EBNA-LP se fixe aux protéines pRB, p53, p14ARF, HA95, Sp100 ou CBP
suggérant que cet EBNA est impliqué dans la régulation de la transcription, du cycle
cellulaire et de la mort cellulaire (Szekely L et al, 1993 ; Kashuba E et al,
IntJCancer2003 ; Han I et al, 2001 ; Ling PG et al, 2005 ; Bandobashi K et al, 2001).
B- EBNA2 :
EBNA2 est l’une des premières protéines virales exprimées après infection. Il
est requis à l’immortalisation cellulaire in vitro par l’EBV (Hammerschmidt et al, 1989).
Il s’agit d’un activateur transcriptionnel qui régule l’activité des promoteurs
viraux (EBNA Cp, LMP1, LMP2A et LMP2B) mais également de nombreux promoteurs
cellulaires codant des facteurs impliqués dans la signalisation (récepteurs tels que CD21),
la prolifération (facteurs de transcriptions tels que STAT6), et la régulation du cycle
cellulaire (cycline D2). EBNA2 ne se fixe pas à l’ADN mais est recruté au niveau de ses
cibles via une interaction avec des facteurs transcriptionnels cellulaires comme RBP-Jκ
(Henkel et al, 1994). RBP-Jκ fait partie de la voie de signalisation cellulaire impliquée
dans des voies de développement cellulaire : la voie du récepteur Notch. Brièvement,
après fixation de son ligand, le domaine intracellulaire de Notch est clivé et transloqué
dans le noyau où, comme EBNA2, il s’associe à RBP-Jκ pour activer la transcription de
ses gènes cibles. De ce fait, EBNA2 est considéré comme un analogue fonctionnel de
Notch activé (ligand-indépendant). (Hayward et al, 2006)
C- EBNA-3 (-3A, -3B et -3C) :
Les protéines EBNA-3A, EBNA-3B et EBNA-3C (ou EBNA-3, EBNA-4 et
EBNA-6) sont codées par trois exons issus du fragment BamHI E et transcrits à partir du
promoteur Cp. Avec EBNA-2, les protéines EBNA-3 sont à l’origine du polymorphisme
entre les variants de l’EBV (EBV de type I et de type II. Bien qu’elles soient issues du
même fragment BamHI E, elles ne possèdent pas le même rôle. En effet, il a été démontré
qu’EBNA-3A et EBNA-3C sont essentielles pour l’immortalisation des LCLs.
Contrairement à EBNA-3B qui ne semble pas nécessaire (thèse Tan Southéa OUK 2008).

Chapitre IV : Virus et cancer du sein
46
La délétion complète de l’exon codant EBNA-3B ne perturbe ni l’immortalisation ni la
croissance des LCLs in vitro (Chen A et al, 2005).
Les EBNA-3 sont des protéines nucléaires grâce à leurs séquences NLS
(Burgess A et al, 2006 ; Buck M et al, 2006) et, comme EBNA-2, se fixent indirectement
à l’ADN sur CBF1/RBPJk.
Cependant, les fixations d’EBNA-2 et des EBNA-3 à CBF1/RBPJk,
notamment EBNA-3A et EBNA-3C, semblent mutuellement exclusives (Johannsen E et
al, 1996). En se fixant à CBF1/RBPJk, EBNA-3A et EBNA-3C déplacent la liaison de ce
facteur à l’ADN et régulent négativement ainsi la transactivation des gènes par EBNA-2
(Waltzer L et al, 1996), dont la protéine LMP-1 (LeRoux A et al, 1994). Par ce même
mécanisme, EBNA-3C est capable de réguler l’expression de tous les EBNAs en réprimant
le promoteur viral Cp (RadkovSA et al, 1997) et exercerait un rétro-contrôle afin de
maintenir les EBNAs et les LMPs à un certain niveau d’expression (Robertson ES et al,
1995 ; Lin J et al, 2002).
De plus, EBNA-3C a un rôle de répresseur transcriptionnel intrinsèque en
recrutant des HDACs (HDAC1 et HDAC2) et des co-répresseurs tels que CtBP, mSin3A et
NCoR grâce à une interaction avec la ProThymosin alpha (Radkov SA et al. 1999 ;
Touitou R et al, 2001 ; Knight JS et al, 2003). L’action répressive d’EBNA-3C ne
requiert cependant pas l’interaction avec CBF1/RBPJk (Bain M et al, 1996 ; Bourillot PY
et al, 1998). Le traitement à la TSA (TrichoStatineA), inhibiteur des HDAC n’inhibe que
partiellement l’activité déacétylase de ce complexe suggérant que d’autres protéines à
activité déacétylase insensibles à la TSA seraient impliquées (Knight JS et al, 2003).
Néanmoins, certaines études ont montré qu’EBNA-3C pourrait agir comme un
activateur transcriptionnel en induisant l’expression du CD23, de LMP-1 (Wang F et al,
1990) et de LMP-2B (Jimenez-Ramirez C et al., 2006) grâce à l’association avec les
facteurs Sp1 et PU.1 (Marshall D and Sample C,1995 ; Lin J et al., 2002). Dans ces
circonstances, EBNA-3C aurait un rôle similaire à EBNA-LP. EBNA-3C est capable
d’interagir, par l’intermédiaire de la ProThymosin alpha, avec des molécules co-
activatrices telles que l’acétyltransférase p300 (Cotter MA et al, 2000 ; Subramanian C
et al, 2002).

Chapitre IV : Virus et cancer du sein
47
EBNA-3C est essentiel à la pathogenèse associée à l’EBV en régulant la
prolifération, le cycle cellulaire et les capacités métastasiques des cellules infectées par ce
virus. EBNA-3C est capable d’interagir et de stabiliser c-Myc grâce au recrutement de son
co-activateur Skp2, augmentant ainsi la transcription des gènes dépendants de c-Myc
(Bajaj BG et al.2008).
EBNA-3C fonctionne comme une oncoprotéine en ciblant le rétinoblastome Rb
(Parker G et al, Oncogene 1996) par différents mécanismes. EBNA-3C peut, d’une part,
augmenter la phosphorylation de Rb (Knight et al, 2004 Knight JS et al, 2004) et d’autre
part recruter SCFSkp2 qui dégrade Rb (Knight JS et al, 2005) permettant la progression
du cycle cellulaire.
EBNA-3C peut également agir sur un autre inhibiteur du cycle cellulaire,
p27Kip soit en empêchant son accumulation (Parker GA et al, Oncogene 2000), soit en
augmentant sa dégradation (Knight JS et al, 2005).
EBNA-3C peut supprimer l’arrêt du cycle en G2/M en réponse à des drogues génotoxiques
(Krauer KG et al, 2004) et pourrait contribuer à l’accumulation de dommages à l’ADN
(Wade M et al ., 2000).
Enfin, EBNA-3C augmente les capacités métastasiques des cellules infectées
par l’EBV en interagissant avec le suppresseur de métastases, Nm23-H1 et inhibe la
capacité de Nm23-H1 à supprimer la migration des cellules de lymphome de Burkitt et des
cellules du cancer du sein (Subramanian C et al, 2001). Grâce à l’interaction avec Nm23-
H1, EBNA-3C induit l’expression de la métalloprotéase MMP-9 (connue pour son
implication dans les lymphomes agressifs, (Kuppers DA et al, 2005) et de la cyclo-
oxygénase COX-2, impliquée dans les phénomènes inflammatoires via les facteurs CRE,
AP1 et NFkB (Kaul R et al, 2006).
D- EBNA1 :
En latence III, la protéine EBNA-1 est transcrite à partir du promoteur Wp puis
à partir du promoteur Cp après circularisation du génome (Kieff E, Fields 1996). Lors des
latences virales de type I et II où EBNA-1 est la seule des protéines EBNAs à être produite,
les promoteurs Wp et Cp sont réprimés par méthylation et c’est le promoteur Qp qui
reprend le relais pour la transcription d’EBNA-1 (Schaefer BC et al, 1995).

Chapitre IV : Virus et cancer du sein
48
Hormis dans la latence de type 0, EBNA-1 est la seule des protéines de latence
à être exprimée dans les cellules infectées par l’EBV (Rowe DT et al, 1986). La protéine
EBNA-1 est la protéine de latence requise pour la réplication et la maintenance de
l’épisome viral. EBNA-1 se fixe au niveau de l’origine de réplication latente OriP (Lupton
S et al., 1985).
Bien qu’EBNA-1 soit essentiel à la maintenance du génome sous forme
épisomique, il semblerait qu’EBNA-1 ne soit pas nécessaire à l’immortalisation des
lymphocytes B in vitro (Humme S et al, 2003).
En effet, l’équipe d’Aloys Schepers a réussi à établir des LCLs après infection
des lymphocytes B par des virions déficients en EBNA-1.
EBNA-1 est décrit comme un activateur transcriptionnel et augmente
l’expression de gènes viraux à partir des promoteurs viraux Cp et de LMP-1 (Gahn TA
and Sugden B, 1995 ; Kieff E, 1996) grâce à son domaine UR1 (Altmann M et al, 2006).
1.5.2.4. Les protéines membranaires de latence :
A- LPM-1 (protéine membranaire de latence-1) :
LMP-1 est transcrit à partir de la région BamHI N du génome viral et du gène
BNLF1 (BamHI N Leftward Frame1). LMP-1 est indispensable à l’immortalisation par
l’EBV et est considéré comme l’oncogène majeur du virus (Eliopoulos AG et al., 2001).
LMP-1 se comporte comme un homologue fonctionnel de la famille des récepteurs du
TNFα, notamment du récepteur CD40. In vivo, LMP1 peut se substituer à CD40 en
induisant l’expression d’un grand nombre de gènes cellulaires impliqués dans les processus
d’activation et de prolifération cellulaire dans les lymphocytes B (Uchida J et al, 1999).
Contrairement aux récepteurs cellulaires dont l’activation dépend d’un ligand, LMP-1 est
constitutivement actif et son expression se traduit par un détournement et une dérégulation
chronique de voies de transduction et de leurs gènes cibles impliqués notamment dans des
processus prolifératifs (Gires O et al, 1997).
B- LMP-2 (protéine membranaire de latence-2) :
Les ARNm de LMP2A et LMP2B sont transcrits à partir du même gène
mais de promoteurs différents. Le promoteur de LMP2B est en fait pLMP1 puisque ce
promoteur est bidirectionnel. Ainsi, la protéine LMP2B est une version tronquée de
LMP2A à laquelle il manque tout le domaine amino-terminal cytosolique.

Chapitre IV : Virus et cancer du sein
49
LMP2A joue un rôle important dans le maintien à l’état latent du virus in
vivo (Merchant, et al. 2001). En effet, LMP2A possède des motifs ITAM
(Immunoreceptor Tyrosine-based Activation Motifs) qui permettent de lier et de séquestrer
les tyrosine kinases Lyn et Syk nécessaires à la signalisation du récepteur des lymphocytes
B (BCR).
La protéine LMP2A recrute également les ligases (E3) qui vont
ubiquitinyler les protéines Lyn et Syk et donc activer leur dégradation par le protéasome
(Winberg, et al. 2000).
En bloquant la voie de signalisation du BCR, la protéine LMP2A inhibe le
passage de la latence au cycle lytique, normalement induit par la signalisation du BCR
(Merchant, Swart et al. 2001). Cependant, LMP2A stimule à un certain degré les
tyrosines kinases et fournit des signaux de survie mimant l’activation du BCR, empêchant
ainsi les lymphocytes B de suivre leur développement normal dans les centres germinatifs -
c’est-à-dire mourir par apoptose en l’absence de sélection via le BCR- (Merchant, Swart
et al. 2001).
Les transcrits de LMP2A ou LMP2B sont fréquemment détectés dans les
lymphomes de Hodgkin, le carcinome du nasopharynx et dans le Lymphome de Burkitt
(Bell, Groves et al. 2006) suggérant que LMP2A est un facteur clé dans la capacité d’EBV
à contribuer aux cancers humains. LMP2A est en effet capable de participer à la
transformation des cellules épithéliales notamment grâce à l'activation des voies de
signalisation cellulaire PI3K-Akt et β-caténine (Morrison and Raab-Traub 2005), mais
également à leur pouvoir métastatique (Pegtel, Subramanian et al. 2005). Cette donnée a
été récemment confirmée par les travaux de Longnecker, dans des lignées de cellules
épithéliales et de cellules de carcinome gastrique (Fukuda et al., 2007). Idée également
supportée par de récents travaux rapportant l’effet protecteur de LMP2A sur l’apoptose
induite par une translocation c-myc dans un modèle de Lymphome de Burkitt, ainsi que sa
participation à la prolifération cellulaire et à l’accélération de la mise en place du
lymphome (Bultema, et al. 2009). Ceci suggère que LMP2A peut avoir un rôle clé non
seulement dans le maintien de la latence chez les porteurs sains, mais également en tant
qu’activateur de la carcinogénèse EBV-dépendante (Pang, Lin et al. 2009).

Chapitre IV : Virus et cancer du sein
50
Les ARN EBERs :
Les EBERs (Epstein-Barr Encoded RNA), composés d’EBER-1 et EBER-2,
sont les transcrits viraux les plus abondants dans les cellules infectées par l’EBV (Rymo L,
1979). EBER-1 et EBER-2 sont de petits ARN non-polyadénylés de 167 et 172 nucléotides
respectivement, (Rosa MD et al, 1981). Contrairement à la majorité des gènes de l’EBV
qui sont transcrits par l’ARN polymérase II, les EBERs sont transcrits par l’ARN
polymérase III (Rosa MD et al, 1981 ; Arrand JR et al, 1982 ; Jat P et al., 1982),
(l’ARN polymérase II est responsable de la synthèse des ARNm (ARN messager),
Alors que l’ARN polymérase III permet la synthèse des ARNt et ARNr 5S (ARN de
transfert et ARN ribosomal 5S). Les EBERs sont exprimés aussi bien lors de l’infection
latente que lors de l’infection lytique (Weigel R et al, 1985).
Cependant, il a été montré que la transcription des EBERs est diminuée lors de
l’induction de l’infection lytique. (Greifenegger N et al, 1998)
Les EBERs sont essentiellement localisés dans le noyau au niveau de
complexes ribonucléoprotéiques mais ils ont également été détectés dans le cytoplasme au
niveau périnucléaire dans le réticulum endoplasmique rugueux (REG) et dans le golgi
(Howe JG et al., 1986 ; Schwemmle M et al, 1992).
De nombreuses données indiquent que les EBERs jouent un rôle essentiel dans
la persistance de l’EBV chez son hôte. En effet, les EBERs sont capables d’induire
l’expression de la molécule antiapoptotique Bcl-2 (Komano J et al, 1999 ; Ruf IK et al,
2000 ; Yamamoto N et al, 2000) et contribuant à la malignicité du lymphome de Burkitt.
Ensuite, les EBERs semblent induire l’expression de l’IL-10.
Cette cytokine immuno-suppressive maintient la croissance des cellules de BL
(Kitawaga N et al, 2000), grâce à une boucle autocrine et contribue à l’inhibition de la
réponse cytotoxique Th1. De plus, l’expression de LMP-1 peut être induite par l’IL10 (Kis
LL et al, 2006).

Chapitre IV : Virus et cancer du sein
51
1.5.2.5. Les transcrits de la région BamHI A (BART, BamHI A Rightward
Transcripts)
Les transcrits de la région BART ont été découverts dans des coupes de tissus de
carcinomes du naso-pharynx (NPC) et représentent les transcrits viraux les plus abondants
(Hitt MM et al, 1989 ; Gilligan K et al, 1990). Les transcrits BARTs sont exprimés dans
les LCLs et dans tous les types de latence virale mais à des niveaux moindres (Brooks LA
et al, 1993). La délétion de la région BART n’altère pas l’immortalisation des lymphocytes
B primaires, ce qui semble indiquer qu’ils ne sont pas essentiels à l’établissement des
LCLs (Robertson ES et al., 1994).
Plusieurs cadres de lecture ouverts (ORF) ont été identifiés dans la région BART : BARF0
; RK-BARF0 (extension de BARF0), RPMS et BARF1 (Smith PR et al, 1993 ; Sadler
RH et al, 1995, Wei MX et al 1989).
RK-BARF0, une des protéines BARF0, est exprimée dans les cellules infectées par l’EBV
in vitro (Fries KL et al, 1997) mais semblent difficilement détectables in vivo (vanBeek J
et al, 2003). RK-BARF0 est associée à des structures nucléaires (Kienzle N et al, 1999).
RK-BARF0 interagit avec le domaine de fixation du ligand du récepteur Notch4 et induit
la translocation nucléaire de celui-ci (Kusano S et al., 2001). L’accumulation nucléaire de
Notch4 diminuerait la quantité totale de Notch fixé à CBF1/RBPJk. Cette diminution
s’expliquerait par l’induction par RK-BARF0 de leur dégradation par le protéasome
(Thornburg NJ et al, JVirol 2004) favorisant ainsi la fixation d’EBNA-2 à CBF1/RBPJk.
En absence d’EBNA-2, RK-BARF0 peut induire l’expression de LMP-1 grâce à son
interaction avec Notch et CBF1/RBPJk (Kusano S et al.,2001).
Un autre transcrit de la région BART, RPMS a un rôle de régulateur négatif de la voie
Notch et EBNA-2. BARF1, transcrit à partir de la région BamHI A, est particulièrement
intéressant. En effet, la surexpression de BARF1 conduit à la transformation de cellules
épithéliales (Wei MX, et al, 1989 ; Wei MX et al, 1997).BARF1 est considéré comme un
oncogène en induisant l’expression de Bcl2, molécule mitochondriale antiapoptotique
(Sheng W et al, Oncogene 2001 ; Wang Q et al, CancerLett 2006) et en stimulant le
cycle cellulaire via l’induction de la cyclineD1 (Wiech T et al, 2008). BARF1 est sécrété
par les cellules infectées par
L’EBV (Fiorini S et al.,2008) et se comporte comme un homologue soluble du récepteur
de la cytokine.

Chapitre IV : Virus et cancer du sein
52
1.5.3. L’infection lytique:
La fréquence avec laquelle les lymphocytes B infectés de façon latente
deviennent permissifs à la réplication virale peut être influencée par des conditions de
culture spécifiques. Spontanément, seules 2 à 10 % des cellules présentes dans les LCL
entrent en cycle lytique.
On peut augmenter ce taux en traitant les cultures cellulaires avec divers agents
chimiques ester de phorbol, butyrate de sodium ou 5-azacytidine ou biologiques (anti-
immunoglobulines de surface, TGF-B ou vecteur exprimant la protéine
EB1 /BZLF1 /Zta /ZEBRA-Z Epstein-Barr Replication Activator).
Cette faible propension à activer le cycle lytique vient du fait que les protéines
exprimées lors de la latence s’opposent à l’entrée dans le cycle lytique. La protéine LMP1
est particulièrement impliquée dans cette inhibition du cycle lytique. En effet, la protéine
LMP1 active la voie de signalisation NFB. Or, le niveau d’expression du facteur de
transcription NFB joue un rôle majeur pour le passage de la latence au cycle lytique en
bloquant à la fois l'expression et l'activité de la protéine Zta (Brown, Song et al. 2003).
Cependant, une étude semble aller à l’encontre d’un tel rôle inhibiteur de LMP1 (Ahsan,
Kanda et al. 2005). Les auteurs montrent en effet que LMP1, dont l’expression est
également induite lors du cycle lytique, est essentielle au relargage des particules virales en
fin de cycle lytique.
Néanmoins, il semble que cela soit indépendant d’une signalisation classique
induite par LMP1 car c’est surtout la forme lytique de LMP1, incapable d’activer cette
signalisation, qui permet de palier l’absence de LMP1 pour le relargage des virions.
La protéine Zta, que code le gène BZLF1, est le commutateur induisant le
passage de la latence au cycle lytique. Quand Zta peut s’exprimer, elle induit sa propre
expression mais également celle des facteurs de transcription Rta et Mta, que codent les
gènes BRLF1 et BMLF1, respectivement. De plus, elle inhibe ses propres inhibiteurs, les
facteurs de la famille NFκB (Dreyfus, Nagasawa et al. 1999).

Chapitre IV : Virus et cancer du sein
53
L’expression des protéines Zta, Rta et Mta, qui sont les antigènes précoces
immédiats (IEA: Immediate Early Antigen) du cycle lytique, va permettre l’expression
d’autres antigènes précoces (EA: Early Antigen) qui vont constituer le pool de protéines
nécessaires à la formation du complexe de réplication virale et à la biosynthèse des
antigènes tardifs que sont les antigènes de capside VCA (Viral Capsid Antigen) et
d’enveloppe MA (Membrane Antigen).
Pendant le cycle lytique, les cellules subissent un changement cytologique qui
se traduit par la formation d’une inclusion intranucléaire, une margination de la chromatine
nucléaire, l’assemblage des capsides à l’intérieur du noyau près de la membrane nucléaire,
le bourgeonnement du virus au travers de la membrane nucléaire et la formation de
vésicules cytoplasmiques. En culture, le cycle lytique se déroule sur une période de 16 à 24
heures. (Marechal et al., 2000)
1.5.4. Réactivation
La réactivation virale spontanée peut survenir dans 2 à 5% des cellules infectées. Ce
cycle lytique peut être abortif ou complet avec assemblage et libération des particules
virales induisant généralement la destruction de la cellule hôte .La protéine ZEBRA (Z
Epstein-Barr Replication Activator, aussi nommée Zta ou EB1), codée par le gène BZLF1,
est le commutateur induisant le passage de la latence au cycle lytique (Lieberman et al,
1990; Yin et al, 2004). ZEBRA induit sa propre expression mais également celle des
facteurs de transcription Rta et Mta, codés respectivement par les gènes BRLF1 et BMLF1
(Ragoczy et al, 1998; Liang et al, 2002).
L’expression des protéines ZEBRA et Rta qui sont les antigènes immédiat précoces
(Immediat Early : IE) du cycle lytique, va permettre l’expression des antigènes précoces
dont Mta (Early Antigen : EA).
Ces derniers vont constituer le pool de protéines nécessaires à la formation du
complexe de réplication viral (TK, DNA polymérase) et à la biosynthèse des antigènes
tardifs que sont les antigènes de capside (Viral Capsid Antigen : VCA) et d'enveloppe MA
(Membrane Antigen) (Kieff et al., 1996).

Chapitre IV : Virus et cancer du sein
54
In vitro, le cycle lytique de l’EBV peut être induit en traitant les cellules infectés par
les esters de phorbol tel que le TPA (12-O-Tétradecanoyl Phorbol-13-Acétate) activateur
de la protéine kinase C (PKC) (Zur Hausen et al., 1978), les inhibiteurs des histones
déacétylases comme le n-butyrate (Luka et al., 1986), les glucocorticoïdes (Bauer, 1983),
les anti-immunoglobulines de surface (Takada, 1984) ainsi que le facteur physiologique le
TGF- (Transforming Growth Factor). (Fahmi et al, 2000)
1.6 Pathologies associées au virus d’Epstein-Barr :
L’EBV est associé à un très large spectre de pathologies bénignes et malignes.
L’expression des protéines virales dans les tissus infectés conduisent à des altérations
cellulaires qui contribuent à ces pathologies :
6.6.1. Infection primaire et pathologies
La mononucléose infectieuse
La mononucléose infectieuse (MNI) est une infection bénigne généralement
spontanément résolutive. C'est la forme clinique symptomatique de la primo-infection à
EBV. Elle se traduit par une fièvre dans 90% des cas, d’adénopathies généralisées, d’une
asthénie sévère. Cette affection se caractérise par une hyperleucocytose (augmentation
anormale du taux de leucocytes) et la présence de lymphocytes atypiques de grande taille
hyperbasophiles constituant jusqu’à 30% des lymphocytes. Ces lymphocytes
correspondent majoritairement à des lymphocytes CD8+ activés. Des complications
peuvent survenir de façon peu fréquente : rupture de rate, méningite ou encéphalite. Dans
certains cas, la MNI peut évoluer vers une infection chronique active (CAEBV ou Chronic
Active Epstein-Barr Virus infection) qui persiste plus d’un an. Dans sa forme sévère, elle
est associée à une mortalité élevée et peut évoluer vers un lymphome (Kimura H et al,
2001).
1.6.2. Infection persistante et pathologies tumorales
Au-delà de la réponse à l’infection primaire, c’est finalement une réponse des
lymphocytes TCD8+ principalement dirigés contre les antigènes de la famille EBNA3
(Rickinson et al.,1997) qui contrôle l’infection à l’EBV et surtout le nombre de cellules B
infectées. Cependant, le système immunitaire ne parvient pas à totalement éliminer
l’infection à l’EBV puisqu’on retrouvera une infection latente dans les cellules du sang

Chapitre IV : Virus et cancer du sein
55
périphérique et une infection lytique dans la cavité oropharyngée (source de sécrétion de
nouveaux virus infectieux). Les patients souffrant d’une agamaglobulinémie liée à l’X ne
disposent pas de lymphocytes B et ne présentent pas de trace d’infection latente, ce qui
détermine le rôle essentiel des cellules B dans l’établissement de la latence (Faulkner,
Burrows et al. 1999).
1.6.2.1. Pathologies de l’hôte immunodéprimé
L’importance du rôle du contrôle immunitaire sur l’infection à l’EBV se comprend surtout
au regard de l’émergence de lymphomes ou de lymphoproliférations B conduites par
l’EBV chez les patients présentant une immunosuppression génétique (immunodéfiscience
combinée sévère, syndrome de Wilskott-Aldrich, Ataxie-Télangiectasie) ou acquise
(immunosuppression iatrogénique post-transplantation, SIDA).
1. Désordres Lymphoprolifératifs Post-Transplantation (PTLD)
Les PTLD comptent une large variété de proliférations lymphocytaires B (quasi
exclusivement) qui s’étalent de la pathologie à régression spontanée, à la prolifération
clonale B létale indifférenciable d’un lymphome non-Hodgkinien (LNH), en passant par
les expansions lymphoplasmacytoïdes typiques de type inflammatoire ; et dont la majeure
partie est associée à l’EBV. L’organisation mondiale de santé (OMS) a classifié ces
pathologies en quatre grands groupes : (i) réactif, hyperplasie plasmacytique, hyperplasie
lymphocytique de type MI, (ii) PTLD polymorphiques, (iii) PTLD monomorphique
correspondant à un LNH à cellules B et T (plus rare), et (iv) Lymphome de Hodgkin (LH)
et PTLD de type LH (également rare). Sur un plan histologique, il s’agit en général de
lymphomes malins non hodgkiniens de haut grade de phénotype B, à grandes cellules de
type immunoblastique ou indifférencié (Orazi, Hromas et al. 1997). Ces affections
n’apparaissent pas dans les tissus ganglionnaires, et se présentent généralement comme des
lésions multifocales dans des sites extra-ganglionnaires, impliquant fréquemment l’organe
transplanté et le tractus gastro-intestinal. Les PTLD peuvent survenir suite à une
transplantation d’organe solide ou de moelle osseuse dont l’incidence est en lien directe
avec l’immunosuppression. L’incidence rapportée des lymphoproliférations survenant
après transplantation d’organe varie ainsi de 1 à 20% selon le type de greffe d’organe.

Chapitre IV : Virus et cancer du sein
56
Le risque est augmenté d’un facteur 15 chez les patients séronégatifs pour
l’EBV (Aris, Maia et al. 1996) et est proportionnel à l’intensité du traitement
immunosuppresseur (la variation d’incidence selon le type d’organe transplanté reflète
l’intensité du traitement immunosuppresseur nécessaire) (Savoldo, Goss et al. 2001). Ces
lymphomes surviennent entre 3 mois et 5 ans (voire davantage) après la greffe, la majorité
étant toutefois observée dans les 12 premiers mois. Les lymphomes survenant précocement
sont presque toujours associés à l’EBV alors que ceux de survenue tardive peuvent être
EBV-positifs ou négatifs (Swinnen 1999). Enfin, les cellules tumorales dérivent en général
des cellules B du receveur.
Après greffe de CSH, les lymphoproliférations liées à l’EBV surviennent
principalement en cas de donneur familial HLA-mismatch, donneur non apparenté, greffe
T-déplétée, traitement immunosuppresseur intense ou déficit immunitaire associé
(Gerritsen, Stam et al. 1996; Heslop and Rooney 1997; O'Reilly, Small et al. 1997;
O'Reilly, Small et al. 1998; Curtis, Travis et al. 1999). Ainsi, si l’incidence des
lymphoproliférations n’est que de 0,25% après greffe géno-identique sans manipulation du
greffon (Deeg and Socie 1998), elle s’élève jusque 5 à 30 % en cas de greffe familiale
HLA-mismatch ou phéno-identique (Savoldo, Goss et al. 2001). Dans la plupart des cas,
ces lymphoproliférations sont de survenue précoce (1,5 - 6 mois) (Deeg and Socie 1998).
Différents travaux ont montré que, durant les premiers mois suivant la greffe, la fréquence
des lymphocytes T EBV-spécifiques était très faible voire nulle, et qu’il fallait au moins 6
mois pour retourner à des valeurs normales (Lucas, Small et al. 1996). A la différence des
lymphoproliférations survenant après greffe d’organe, les cellules tumorales proviennent
presque toujours des lymphocytes du donneur (Orazi, Hromas et al. 1997).
L’apparition d’un syndrome lymphoprolifératif EBV+ semble précédée par une nette
augmentation de la charge virale associée à la prolifération des lymphocytes B infectés par
l’EBV. Plusieurs études ont ainsi montré qu’une mesure régulière de la charge virale EBV
représentait un moyen de dépistage précoce des lymphoproliférations EBV+ à la fois après
transplantation d’organe et greffe de CSH (Rowe, Qu et al. 1997; Baldanti, Grossi et al.
2000; Green, Bueno et al. 2000; Stevens, Verschuuren et al. 2001). Une augmentation
de 2 ou 3 log de la charge virale semble prédire le développement d’une
lymphoprolifération EBV+ après greffe de CSH (Rooney, Smith et al. 1995).

Chapitre IV : Virus et cancer du sein
57
Il convient néanmoins de souligner que tous les patients présentant une charge virale
élevée (en particulier après transplantation d’organe) ne développeront pas une
lymphoprolifération (Gottschalk, Heslop et al. 2002). Des travaux récents indiquent
également que les cellules B mémoires des sujets possédant une charge virale EBV élevée
peuvent présenter des profils de latence variés, et qu’une latence de type III conférerait un
risque plus élevé de développer une lymphoprolifération (Rose, Green et al. 2001).
effet qu’elles présentent une latence de type III (programme par défaut), incluant de
fait l’expression de LMP1 et EBNA2, protéines critiques de la transformation cellulaire
EBV induites. Sans contrôle immunitaire, une croissance non-inhibée des cellules B
infectées par l’EBV va se développer. Cette lésion précoce est généralement de nature
polyclonale comme l’attestent des expériences de Southern blotting des séquences des TR
(Terminal Repeats) d’EBV (Raab-Traub and Flynn 1986), et de contrôle de
réarrangement des gènes des immunoglobulines. Une baisse dans le régime
immunosupressif ou encore l’infusion de cellules T cytotoxiques EBV-spécifiques peut
mener à la régression de la lésion ; preuve empirique supplémentaire de l’importance du
rôle de l’immunité cellulaire T dans le contrôle de la pathologie. Cependant, certains
défauts génétiques peuvent survenir dans les cellules B infectées par l’EBV, incluant les
gènes codant pour p53, c-myc ou encore bcl-6 (Knowles, Cesarman et al. 1995; Polack,
Hortnagel et al. 1996; Cesarman, Chadburn et al. 1998). Ces lésions alors souvent plus
agressives, présentent un profil d’expression des gènes de latence différent et ne répondent
pas à la baisse du régime immunosuppressif.
6.2.1.2. Lymphomes associés à l’EBV chez le malade du SIDA
Ces lymphomes sont un groupe hétérogène qui regroupent les lymphomes du
système nerveux central (SNC), lymphomes diffus à larges cellules, LH, lymphome de
Burkitt (LB) ou lymphome de type Burkitt, lymphomes primaires d’effusion (LPE) ; leur
fréquence est variable et dépend du type de tumeur impliquée. Les lymphomes du SNC
sont ainsi tous associés à l’EBV, probablement du fait que l’immunosuppression due au
VIH augmente le trafic des cellules B infectées par l’EBV dans le cerveau, ou encore que
la combinaison de l’immunosuppression virale à la zone de privilège immunitaire que
représente le cerveau en fait un lieu privilégié de développement des lymphoproliférations
associées à l’EBV. La plupart des LH et des lymphomes diffus à larges cellules de
morphologie immunoblastique chez les patients sidéens sont liés à l’EBV.

Chapitre IV : Virus et cancer du sein
58
Ces pathologies n’arrivent généralement qu’en fin de SIDA quand le patient est
sévèrement immunodéprimé. Au contraire, les LB et LB-like, ne sont associés à l’EBV que
dans 30% à 50% des cas (Diebold, Raphael et al. 1997). Dans ces cas, l’infection à l’EBV
et l’immunosuppression peuvent augmenter le pool de cellules B à risque de translocation
c-myc et ne seraient alors pas la cause primaire du développement de la prolifération
maligne.
A. Léiomyosarcomes liés à l’Immunodéficience
Dans de rares cas, les patients immunodéprimés développent des néoplasmes
des cellules musculaires lisses qui présentent un génome EBV clonal (McClain, Leach et
al. 1995). Ces tumeurs se développent typiquement chez les enfants au décours d’une
transplantation d’organe ou du SIDA, et sont plus communément localisées dans le tractus
gastro-intestinal ou l’arbre bronchique. Le programme de latence est alors de type III
(Rogatsch, Bonatti et al. 2000). Les cellules musculaires lisses ne sont pas des cibles de
l’EBV, cependant une expression variable du CD21, le récepteur de l’EBV, a été révélée
dans ces cellules tumorales (Lee, Locker et al. 1995; Rogatsch, Bonatti et al. 2000).
Aucune infection des cellules musculaires lisses normales adjacentes à la tumeur EBV+ n’a
été détectée, et l’EBV n’a pas été trouvée comme infectant les léiomyosarcomes des
patients immunocompétents. L’immunosuppression étant dans ce cas sûrement responsable
de cette entrée anormale de l’EBV dans les cellules musculaires lisses, apportant alors un
risque plus grand de développer une transformation maligne.
B. Leucoplasie orale chevelue
Il s’agit d’une pathologie proliférative des cellules épithéliales de la muqueuse
squameuse, que l’on rencontre de manière spécifique chez les immunodéprimés. Cette
pathologie est remarquable dans le sens ou il s’agit de la seule qui soit associée à
l’infection primaire lytique à l’EBV(Greenspan and Greenspan 1989). L’expression des
gènes viraux lytique est facilement observable, les stades plus avancés dans la réplication
virale se rencontrant dans les couches épithéliales les plus différenciées (Wolf, Bogedain
et al. 1993; Walling, Flaitz et al. 2001).

Chapitre IV : Virus et cancer du sein
59
Les lésions se développeraient à partir de cellules épithéliales en phase
d’infection latente qui deviendraient plus permissives à la réplication virale lytique suite à
l’immunosuppression. L’immunosuppression pourrait également permettre aux cellules
épithéliales de la langue d’être anormalement infectées à partir de quoi elles deviendraient
plus susceptibles à la réplication lytique excessive et à la prolifération. Un traitement par
acyclovir permet une régression des lésions, mais celles-ci réapparaissent fréquemment à
l’arrêt du traitement (Walling, Flaitz et al. 2003).
1.6.2.2. Pathologies de l’hôte immunocompétent
Le Lymphome de Burkitt, le Lymphome de Hodgkin, et le Carcinome du
Nasopharynx sont trois des plus importantes pathologies malignes associées à l’EBV chez
l’hôte immunocompétent. Elles s’établissent généralement tardivement après l’infection
primaire, l’infection à l’EBV paraît en ce sens l’un des facteurs du long et complexe
processus de transformation maligne.
1. Lymphome de Burkitt (LB)
Le lymphome de Burkitt est un lymphome malin non hodgkinien de haut
grade. Il en existe trois formes, endémiques, sporadiques, et associées à
l’immunosuppression qui diffèrent respectivement selon leur présentation, biologie et
association à l’EBV.
La forme endémique est celle initialement décrite par Denis Burkitt, et qui se
rencontre fréquemment en Afrique équatoriale et en Papouasie Nouvelle-Guinée. La
fréquence relative de l’EBV dans les cellules tumorales de LB endémique est de 100%. On
ne rencontre principalement la forme sporadique de LB que chez les jeunes adultes et les
enfants sans distribution géographique particulière ; et son incidence relative est plus faible
avec 2-3 cas par million d’individus. De même, en Europe et aux Etats-Unis d’Amérique,
où l’on rencontre cette forme de LB, l’association à l’EBV est faible (15 à 30% des cas)
(Gutierrez, Bhatia et al. 1992; Hecht et al.,2000).
L’aspect histologique le plus commun est un infiltrat diffus et monomorphique
de cellules lymphoïdes atypiques de taille intermédiaire présentant un noyau rond, une
chromatine condensée, 1-3 nucléoles centraux, un cytoplasme modérément basophile
souvent vacuolisé.

Chapitre IV : Virus et cancer du sein
60
Ce sont des cellules hautement prolifératives présentant fréquemment des
figures mitotiques et des corps apoptotiques. En accord avec le taux élevé de mort
cellulaire par apoptose, on retrouve la présence de nombreux histiocytes phagocytaires
(macrophages du tissu conjonctif) d’où l’aspect typique en « ciel étoilé » des tissus lésés
(Pelc, De Maertelaere et al. 1983).
La classification de l’OMS définit le LB par la présence invariable d’une
translocation c-myc qui résulte en une dérégulation de l’expression de la protéine c-MYC.
Le réarrangement de l’oncogène c-myc est considéré comme l’événement transformant
initial (Shapira and Peylan-Ramu 1998). Cette translocation implique un échange de
matériel génétique entre l’oncogène c-myc sur le chromosome 8 (8q24) et soit la chaîne
lourde des immunoglobulines sur le chromosome 14 bande q32 [t(8 ;14)] (80% des cas),
soit la chaîne légère kappa sur le chromosome 2 (2p11) [t(8 ;2)] (15% des cas), ou enfin
avec la chaîne légère lambda sur le chromosome 22 (22q11) [t(8 ;22)] (5% des cas). Dans
la forme endémique, les points de cassure du chromosome 8 se trouvent dans la région 5’
non-codante de l’exon 1 de c-myc, et les points de cassure sur le chromosome 14 se
trouvent dans les régions de jonction sur des sites possibles d’hypermutation somatique qui
à lieu durant le développement des centres germinatifs-. Dans les formes sporadiques et
associées au SIDA, les points de cassures du chromosome 8 se situent entre l’exon 1 et
l’exon 2 de c-myc et dans la zone du switch des chaînes lourdes des immunoglobulines sur
le chromosome 14 (Hecht et al.,2000)- zone impliquée dans la commutation de classe
isotypique des immunoglobulines durant le développement des centres germinatifs B-. La
présence des translocations dans des régions qui subissent des cassures chromosomiques
durant la maturation des centres germinatifs renforce l’idée que ces translocations sont des
erreurs qui ont lieu durant le développement normal des cellules B. Elles dérégulent
l’expression de c-MYC, menant les cellules vers le cycle cellulaire et activant les voies
anti-apoptotiques.
Des travaux suggèrent que l’infection à EBV ne serait qu’un événement tardif
dans la genèse des lymphomes de Burkitt (Gutierrez, Bhatia et al. 1997). Un modèle de
développement de LB décrit ainsi l’EBV comme un potentialisateur dans la formation des
tumeurs mais avec un rôle plus restreint dans le maintien du phénotype des cellules
tumorales.

Chapitre IV : Virus et cancer du sein
61
Ce modèle implique une action précoce de l’EBV dans le processus
lymphomagénique via une augmentation du nombre d’instabilités géniques probables dans
les cellules B infectées, telle qu’une translocation du gène c-myc, résultant alors en un LB.
Un modèle alternatif décrit l’EBV comme augmentant le potentiel de cellules B présentant
déjà une translocation c-myc. Ce modèle pourrait expliquer la présence permanente de
l’EBV dans les cellules tumorales, puisqu’elles en retirent un avantage biologique.
Certaines études suggèrent.
En effet, qu’EBNA1 (via une régulation contrôlée lors du cycle cellulaire) ou
les EBERs (qui induisent clonalité, résistance à l’apoptose, tumorigénicité et induction
d’IL-10) pourraient avoir un effet sur la croissance cellulaire dans des modèles
expérimentaux de LB (Ruf and Sample 1999; Nanbo et al.,2002).
2. Le lymphome de Hodgkin
Environ 2/3 des lymphomes de Hodgkin (LH ou HL, Hodgkin Lymphoma) sont
associés à l’EBV. Le LH est caractérisé par la présence de cellules néoplasiques
particulières appelées cellules de Reed-Sternberg et la désorganisation de l’architecture
ganglionnaire (Kapatai G and Murray P, 2007). Le LH représente près de 30% des
lymphomes. Les cellules de Reed-Sternberg, qui dérivent du centre germinatif, constituent
1 à 3% de la masse tumorale. Ces cellules sont souvent de grandes cellules binucléées avec
des nucléoles proéminents et un immunophénotype peu commun : CD15+ et CD30+. De
plus, ces cellules présentent une latence virale de type II avec l’expression d’EBNA-1,
LMP-1 et LMP-2 ainsi que l’expression des ARNs EBER et les transcrits de la région
BamHI (BARTs).
3. Le carcinome du rhinopharynx
Ces tumeurs sont relativement rares, mais certaines régions du monde
présentent une incidence très élevée de carcinomes indifférenciés du nasopharynx: le
Maghreb, le sud-est asiatique (provinces du sud de la Chine, Nord-Vietnam, Malaisie,
Indonésie, Nord-Thaïlande), l’Afrique de l’est (Tanzanie, Kenya, Ouganda), le Groenland
et l’Alaska (certaines peuplades d’Esquimaux) (Nicholls, Agathanggelou et al. 1997).
Une explication potentielle de l’augmentation du risque dans ces régions,
pourrait reposer sur des traits culturels, alimentaires ou encore une prédisposition

Chapitre IV : Virus et cancer du sein
62
génétique au développement des CNP associés à l’EBV. Cette pathologie représente dans
ces contrées plus de 99% des tumeurs malignes du nasopharynx et sont souvent le premier
cancer de l’homme (20% des cancers chez l’homme en Asie du sud-est) avec un sex-ratio
compris entre 2 et 3.
Les données épidémiologiques suggèrent donc l’existence de facteurs
génétiques et/ou environnementaux en plus du rôle de l’EBV (Hsu et al., 2000). L’âge de
découverte des carcinomes indifférenciés du nasopharynx est compris entre 20 et 40 ans.
Des anticorps spécifiques de l’EBV sont détectés à un taux élevé au diagnostic. Leur titre
est corrélé à l’extension de la maladie et à la réponse au traitement, qui repose
principalement sur la radiothérapie.
Le rôle de l’EBV dans le développement du CNP dépend de la compréhension
de quand et comment l’EBV infecte les cellules épithéliales. On sait clairement que la
cavité orale est la porte d’entrée du virus, et que le virus se répand via les sécrétions
oropharyngées. L’idée de cellules épithéliales en infection persistante est soumise à
controverse dans la mesure où l’on n’a pas encore identifié de manière univoque de
cellules épithéliales oropharyngées en infection persistante chez le porteur sain. Les CNP
peuvent survenir à partir de cellules épithéliales en infection persistante qui subiraient
d’autres événements génétiques comme ceux présentés pour le modèle du développement
des LB. Alternativement, l’infection à l’EBV dans les cellules épithéliales de personnes à
risque peut avoir lieu plus tardivement dans leur vie, certaines études suggérant qu’une
exposition précoce du nasopharynx au carcinogènes environnementaux rendrait
l’épithélium plus sensible à l’infection par l’EBV (Knox, Li et al. 1996). Ainsi, le CNP
n’évoluerait pas à partir de cellules épithéliales déjà infectées par l’EBV, mais
représenterait une infection rare de cellules épithéliales rendues plus susceptibles due à une
exposition aux carcinogènes, ou à d’autres changement prénéoplasiques. Bien que le stade
précis auquel a lieu l’infection à l’EBV ne soit pas connu, il est clair que toutes les cellules
tumorales dans tous les CNP associés à l’EBV portent un génome viral monoclonal
(Niedobitek, Young et al. 1992; Pathmanathan, Prasad et al. 1995).
Le profil d’expression des gènes de latence est intermédiaire à celui de LB
(latence I) et des LH (latence II). EBNA1 et les EBERs sont toujours présents quelque
soient les cas de CNP EBV+, les transcrits de LMP2A et -2B sont amplifiables dans la

Chapitre IV : Virus et cancer du sein
63
plupart des tumeurs, et LMP2A peut jouer un rôle sur la croissance des cellules épithéliales
(Morrison et al., Raab-Traub 2005) ; cependant cette protéine n’est détectée que dans
50% des cas (Heussinger, Buttner et al. 2004). L’ARNm de LMP1 est plus difficilement
détectable et la protéine n’est actuellement identifiée que dans 35% des cas (Young,
Dawson et al. 1988; Brooks, Yao et al. 1992).
La présence de LMP1 dans les lésions pré-invasives suggère cependant que
l’expression de LMP1 est nécessaire dans les lésions précoces, mais pas essentielle à
l’établissement du carcinome. La prolifération des cellules épithéliales induite par l’EBV
augmente le risque que d’autres événements génétiques surviennent contribuant ainsi à la
tumorigènese.
L’expression occasionnelle de la protéine lytique BZLF1 a été détectée dans
des échantillons de CNP (Cochet, Martel-Renoir et al. 1993) ; cependant aucune autre
protéine du cycle lytique n’a été retrouvée ni même la présence d’ADN linéaire ce qui
implique que les cellules tumorales ne sont pas permissives à la réplication virale (Gulley,
Amin et al. 1995). Cependant, on enregistre fréquemment la présence d’IgA anti-VCA,
EA, ou encore anti-MA chez les patients CNP, qui peuvent être utilisés comme tests pour
le screening des patients des zones endémiques de CNP (Low, Leong et al. 2000). De
même, les taux d’ADN libres dans le sérum, peuvent servir au diagnostique, au pronostic
et comme information thérapeutique pour les patients CNP (Chan et al., Lo 2002). Nous
ne savons pas très bien si l’ADN retrouvé est de l’ADN nu ou encapsidé, mais sa
résistance à la DNase et les masses tumorales relativement faibles comparativement au
taux d’ADN sérique suggèrent que le virus pourrait survenir d’une réplication active plutôt
que des cellules tumorales mourantes.
4. Maladies associées à l’EBV dans les cibles cellulaires non-conventionnelles
Parmi les pathologies touchant des cibles cellulaires non conventionnelles de
l’EBV, on trouve : des carcinomes de type lymphoépithéliome, des sous type de
carcinomes gastriques, des désordres lymphoprolifératifs des cellules T, des tumeurs
associées aux cellules dendritiques folliculaires, et quelques carcinomes hépatiques et
mammaires.

Chapitre IV : Virus et cancer du sein
64
Ces pathologies ont en commun le fait que la présence de l’EBV n’y représente
qu’un événement rare, mais si l’infection à lieu, l’expression des gènes viraux conduit à la
tumorigenèse via de nombreuses voies que l’on rencontre dans les lymphomes B.
A. Carcinome gastrique et carcinome de type lymphoépithéliome.
Des carcinomes se présentant comme des CNP indifférenciés (de type
lymphoépithéliome) ont été identifiés dans d’autres sites que l’oropharynx dont le thymus,
le larynx, les amygdales, les glandes salivaires, les poumons, la peau, le col de l’utérus, la
vessie et l’estomac. L’incidence de l’infection à l’EBV dans les tumeurs de type
lymphoépithéliome de l’intestin antérieur est endémique à certaines populations, comme
Asie du Sud-est, chez les Esquimaux du Groenland, et en Amérique du Sud (Burke, Yen
et al. 1990; Serraino, Piselli et al. 2005). L’histologie évoque celle du CNP, suggérant un
mécanisme pathogénique similaire. Les résultats d’une étude rétrospective faite au Texas
ont plus récemment été publiés.
Sur 235 cas de cancers gastriques primaires, seuls 12 étaient associés à l’EBV.
Cette association n’a été révélée que dans les cellules tumorales et non dans les cellules des
tissus avoisinants. 8 de ces 12 patients présentaient des métastases ganglionnaires associées
à l’EBV. Ce qui suggère une réplication du génome virale de l’EBV dans les cellules
tumorales (Truong, Feng et al. 2009). Il a été convenu qu’une fraction des carcinomes
gastriques conventionnels sans morphologie de type lymphoépithéliome était également
associée à l’EBV (Imai, Koizumi et al. 1994). L’étude d’une grande cohorte de cas de
carcinomes gastriques (n=970) à montré la présence d’EBER1 (par hybridation in-situ)
dans 6,9% des tumeurs faiblement à modérément différenciées (Tokunaga, Land et al.
1993). Cependant, l’épithélium gastrique comme cible normale d’infection à l’EBV et le
rôle exact de celui-ci dans ces pathologies malignes reste incertain.
B. Maladies lymphoprolifératives à cellules T
Les thymocytes et les cellules T expriment le récepteur à l’EBV (CD21/CR2) à
bas bruit ; cependant même durant la phase aiguë de la MI, l’infection par l’EBV des
cellules T reste un événement rare.

Chapitre IV : Virus et cancer du sein
65
L’association la plus constante de l’EBV à des désordres cellulaires T se
rencontre dans les cas de lymphome des cellules T du syndrome hémophagocytaire associé
au virus (SHAV), un lymphome nasal NK/T, où quasiment 100% des tumeurs sont
positives à l’EBV. Les lymphomes à cellules T SHAV sont des lymphoproliférations rares,
de nature monoclonale des cellules T matures qui se produisent chez les patients en
infection chronique active à l’EBV ou occasionnellement après une infection primaire
aiguë (Jones, Shurin et al. 1988; Kanegane, Bhatia et al. 1998; Kanegane, Miyawaki et
al. 1999). Comme son nom l’indique, elle est associée à une hémophagocytose (syndrome
d’activation macrophagique) caractéristique souvent de nature fulminante (Su, Hsu et al.
1993). Les cellules présentent une infection clonale à l’EBV et un profil de latence I/II
(Kanegane, Bhatia et al. 1998). Cette hémophagocytose qui accompagne les
proliférations des cellules T néoplasiques semble être médiée par l’expression de TNF-
par les cellules T, qui en combinaison à l’IFN- et probablement d’autres cytokines, active
les macrophages (Lay, Tsao et al. 1997). Le profil d’expression des gènes viraux est
quasiment le même dans le cas des lymphomes NK/T nasaux, avec une expression clonale
à l’EBV dans tous les cas. Ces lymphomes sont plutôt agressifs et les tumeurs destructives
qui impliquent la ligne médiane des tissus nasopharyngés sont plus communément
rencontrées dans les populations d’Asie du Sud-est.
L’absence de CD3 et de TCR dans la majorité des cas, associé à l’absence de
réarrangements des gènes du TCR et à la présence du CD56 (marqueur des cellules NK),
indiquent que ces tumeurs sont dérivées des cellules NK (Jaffe, Chan et al. 1996;
Cheung, Chan et al. 2003). Peu de choses sont actuellement connues sur le rôle de l’EBV
dans ces pathologies notamment dû au manque de modèles de cellules T immortalisées par
l’EBV.
C- Pseudotumeurs inflammatoires de type Tumeurs des cellules dendritiquesfolliculaires (CDF)
Ces tumeurs ont récemment été identifiées et reconnues en tant que variant des
tumeurs CDF (Cheuk, Walford et al. 2001). Ces tumeurs sont fortement prédominantes
chez les femmes, fréquemment localisées dans des sites intra-abdominaux, spécialement la
rate et le foie et associées à une présence fréquente de symptômes systémiques
(Rosenbaum, Fekrazad et al. 2009).

Chapitre IV : Virus et cancer du sein
66
D’un point de vue histologique, les pseudotumeurs inflammatoires de type
CDF, présentent des cellules ovoïdes ou fuselées avec des noyaux tordus ou pliés, et
occasionnellement vésiculaires, avec un nucléole distinct, sur un fond
lymphoplasmacytique inflammatoire. Les cellules fuselées dérivent de cellules
dendritiques folliculaires, elles expriment CD21 (CR2), CD35, CD23, R4/23, Ki-M4 et
CNA.42 (Selves, Meggetto et al. 1996; Shek, Ho et al. 1996; Cheuk, Walford et al.
2001; Horiguchi, Matsui-Horiguchi et al. 2004). Enfin, des analyses en Southern Blot
impliquent la présence de clones identiques d’épisomes d’EBV, et expriment LMP1,
EBNA1, et les EBERs, mais sont EBNA2 négatives.
D. Le cancer du sein
L’association entre l’EBV et ce cancer a été suggérée par plusieurs équipes.
Des études réalisées sur l’identification de gènes de l’EBV ou des produits de ces gènes
dans la tumeur. Ces études ont utilisé plusieurs méthodes. La PCR est une méthode
potentiellement très sensible et spécifique pour détecter l’ADN de l’EBV. Parmi les études
utilisant la PCR pour détecter l’EBV dans les tumeurs du sein, le virus a été retrouvé dans
0 à 66 % des échantillons (Bonnet et al, 1999; Grinstein et al, 2002; Labrecque et al,
1995; Preciado et al, 2005; Xue et al, 2003). Les travaux de Bonnet et al. Ont permis de
mettre en évidence la présence du génome viral de l’EBV dans 51% des cas étudiés. De
plus, l’EBV n’a été retrouvé spécifiquement que dans les cellules tumorales et sa présence
a été corrélée à l’existence de facteurs de mauvais pronostic.
Plus récemment, la même équipe a montré que l’EBV augmentait la résistance
des cellules à l’agent chimiothérapique paclitaxel (Arbach et al, 2006). L’association entre
l’EBV et le cancer du sein reste néanmoins l’objet de beaucoup de discussions (Chu et al,
2001; Murray et al, 2003; Perrigoue et al, 2005).

Chapitre IV : Virus et cancer du sein
67
- Premières études
1. Etudes sérologiques
Aucune étude publiée n’a regardé les marqueurs sérologiques classiques de
l’EBV chez les patientes atteintes d’un cancer du sein. Récemment, les anticorps anti-
BFRF1, une protéine de l’EBV associée à la réplication virale, n’ont pas été détectés chez
les 71 patientes atteintes d’un cancer du sein mais ont été retrouvés chez des patients
atteints d’autres cancers associés à l’EBV (Angeloni et al, 2001).
2. Etudes de la tumeur
La plupart des résultats en faveur d’une association EBV / cancer du sein
proviennent des études réalisées sur l’identification de gènes de l’EBV ou des produits de
ces gènes dans la tumeur. Ces études ont utilisé plusieurs méthodes.
La PCR est une méthode potentiellement très sensible et spécifique pour
détecter l’ADN de l’EBV. Cependant, il est impossible de différencier l’EBV des cellules
tumorales de l’EBV des lymphocytes infiltrants, une limitation dans l’étude de tumeurs
comme le cancer du sein qui possède des lymphocytes infiltrants.
Des études utilisant la PCR pour détecter l’EBV dans les tumeurs du sein, le
virus a été retrouvé dans quelques des échantillons. La prévalence est plus importante
lorsque la PCR est utilisée pour détecter les EBERs et la séquence BamH1 W (Labrecque
et al, 1995) (Luqmani et al, 1995) ; (Bonnet et al, 1999) ; (Brink et al, 2000) ; (Fina et
al, 2001) ;(Grinstein et al, 2002). La prévalence de l’EBV dans les échantillons est plus
modérée lorsque la PCR est utilisée pour détecter le gène LMP1 ou EBNA-4 (Brink et al,
2000) ; (Chu et al, 2001) et plus faible lorsque la PCR est utilisée pour détecter EBNA-1
(Gaffey et al, 1993) ; (McCall et al, 2001).

II : Matériel e méthodes
68
L’objectif de notre travail vise la recherche des marqueurs sérologiques du virus
Epstein-Barr (EBV) dans les sérums de patientes ayant un cancer du sein et issues de
l’Ouest Algérien.
1. Population de l’étudeNotre étude a porté sur vingt quatre patientes admises au service de gynécologie, aile
oncologie médicale du centre hospitalier universitaire d’Oran, toutes les patientes sont
suivies pour un cancer du sein.
Ces patientes ont été triées de façon aléatoire parmi une population générale, elles
ont fait l’objet d’une enquête réalisée par le biais d’un questionnaire, soulevant plusieurs
paramètres : cliniques, socioéconomiques, etc… (Annexe 1). Il est important de noter que
toutes ces patientes étaient consentantes pour le prélèvement des échantillons biologiques
en signant un consentement éclairé (Annexe 2).
2. Prélèvements
2.1. Prélèvement de biopsiesDès l’ablation du sein au niveau du bloc opératoire, nous avons procédé à la
récupération des tumeurs. Une partie de la tumeur a été immédiatement congelée à -80°C,
pour servir à une étude moléculaire ultérieure, quant à l’autre partie, elle a été fixée pour la
réalisation de l’étude histologique afin de déterminer la classification TNM et le SBR (le
grading de Scarff-Bloom et Richardson).
2.2. Prélèvement de SérumsDans des tubes secs nous avons procédé au prélèvement de 10ml de sang total
sur toutes nos patientes avant ablation de la tumeur, le sang a été centrifugé à 1800 rpm, et
les sérums ont été récupérés et conservés à -20°C pour permettre la réalisation de l’étude
sérologique.
3. Etude sérologiqueCette étude nous a permis de rechercher les différents marqueurs sérologiques
(anticorps anti-EBV) dirigés contre les antigènes viraux exprimés par l’EBV, il s’agit de
l’antigène EA, IgG, IgA/VCA, IgG, IgM, IgA, EBNA1. Pour cela nous avons utilisé trois
différentes techniques : ELISA (enzyme linked immunoassays), IFI (immunofluorescent
assays) et western blot.

II : Matériel e méthodes
69
Il existe plusieurs anticorps anti-EBV, dirigés contre les différents constituants du
virus : anticorps anti-VCA (Viral Capsid Antigen), anticorps anti-EA (Early Antigen) et
anticorps anti EBNA1.
3.1. Technique Elisa (enzyme-lynked immunoessay)
3.1.1. Principe de la techniqueCette technique permet la réalisation d’un dosage semi-quantitatif in vitro. Elle
permet aussi de mesurer le titre en anticorps spécifiques humains de classe IgA, IgG, IgM
dirigés contre les antigènes EBV-CA, EBV-EA dans le sérum ou le plasma. On adsorbe
l’antigène sur la plaque qui contient des barrettes de microtitration de 8 puits sécables,
chacun contenait les antigènes EBV-CA, EBV-EA.
Dans la première étape de la réaction, les échantillons dilués de sérum ou plasma des
patientes sont incubés dans les puits. Dans le cas d’échantillon positif, les anticorps
spécifiques de classe IgA, IgM, IgG, se fixent aux antigènes. Pour détecter les anticorps
fixés, une seconde incubation est réalisée en utilisant un anticorps anti-IgA (IgM, IgG)
humain couplé à une enzyme (conjugué enzymatique), ce conjugué est capable de générer
une réaction colorée.
3.1.2. Protocole expérimental Incubation des échantillons : transférer 100 ml de calibreur, de contrôle positif et
négatif ou des échantillons des patientes, Incuber pendant 60 minutes à température
ambiante (+18°C à 25°C) ;
Lavage : vider et laver les puits 3 fois ;
Incubation du conjugué pendant 60 minutes, pipeter 100 ml du conjugué
enzymatique dans chaque puits de la microplaque à température ambiante ;
Lavage : vider et laver les puits ;
Incubation du substrat pendant 20 minutes, pipeter 100 ml de la solution substrat
dans chaque puits de la microplaque ;
Arrêt de la réaction ;
Pipeter 100 ml de la solution d’arrêt dans chaque puits ;
Lecture : mesure photométrique de l’intensité de la coloration.

II : Matériel e méthodes
70
3.2. Technique IFI (immunofluorescence indirecte)
3.2.1. Principe de la techniqueCette technique est exclusivement destinée au dosage quantitatif et ou qualitatif in
vitro des anticorps humains dans le sérum.
Les sérums des patientes censés contenir les AG viraux : EBV/VCA EBV/EBNA
ont été dilué et incubé en présence des anticorps de classe IgA, IgG, IgM, dirigés contre
ces antigènes.
3.2.2. Protocole expérimental Déposer 20 ml de sérum dilué dans chaque puits de réaction ;
Commencer la réaction en mettant chaque lame à BIOCHIPs dans la position
correspondante à température ambiante ;
Rincer les lames avec un flux de tampon PBS-Tween ;
Déposer 20 ml de complément fraîchement reconstitué dans chaque puits de
réaction ;
Incuber : sortir une lame du PBS-Tween, dans les 5 secondes, essuyer le dos et le
coté bas 30 mn dans une température ambiante. (+18 à 25°C) ;
Rincer les lames avec un flux de tampon PBS-Tween ;
Déposer 20 ml de globuline anti-Ig humaine marquée à fluorescéine ;
Incubation : sortir une lame du tampon PBS-Tween, dans les 5 secondes, essuyer le
dos et le coté bas 30 mn dans une température ambiante. (+18 à 25°C) ;
Lavage : rincer les lames avec un flux de tampon PBS-Tween ;
Inclusion : Placer des lamelles couvre-objet en verre sur le support d’inclusion en
polystyrène. Déposer sur ces lamelles des gouttes de 10 ml de Glycérol/PBS par
puits de réaction ;
Evaluation : interpréter la fluorescence au microscope.
3.3. Technique de Western Blot
3.3.1. Principe de la techniqueLe test Western blot est un test qualitatif in vitro pour la détection des anticorps
humain dirigé contre le virus Epstein-Barr (EBV). Ce kit Contient des bandelettes avec les
extraits d’antigènes de l’EBV séparés par électrophorèse. Les bandelettes de blot doivent
être dans la première étape, incubées dans le sérum patient dilué.

II : Matériel e méthodes
71
3.3.2. Protocole expérimental Saturation : remplir un nombre de rigoles du bac d’incubation avec le nombre
d’échantillon à tester avec 1,5 ml de tampon universel plus dilué fourni 10x
concentré. Avant la préparation du tampon universel agiter soigneusement le flacon
de 50 ml. La quantité requise doit être prélevé du flacon avec une pipette propre et
ensuite diluée au 1 :10 avec de l’eau distillée ;
Incuber pendant 15 minutes ;
Incubation des sérums : remplir chaque rigole avec 1,5 ml de sérum patient dilué et
incuber 30 minutes à température ambiante ;
Lavage : aspirer et éliminer le liquide contenu dans les rigoles et laver ensuite sur
un agitateur à bascule 3 fois pendant 5 minutes ave 1,5 ml de tampon universel ;
Incubation du conjugué : déposer 1,5 ml de conjugué enzymatique dilué et laisser
incuber 30 minutes.
Lavage : aspirer et éliminer le liquide contenu dans les rigoles et laver ensuite sur
un agitateur à bascule 3 fois pendant 5 minutes ave 1,5 ml de tampon universel ;
Incubation : déposer 1,5 ml de solution substrat dans chaque rigole 10 minutes à
température ambiante ;
Arrêt de la réaction : Aspirer et éliminer le liquide de chaque rigole et laver ensuite
3 fois 1mn avec de l’eau distillée.
4. Techniques histologique :
La technique histologique à pour but l'obtention de coupes minces, transparentes de tissus
et d'organes observables au microscope, le plus souvent après coloration par des colorants
spécifiques qui donnent aux diverses parties des teintes différentes
II : Matériel e méthodes
72
Tableau : 1
Réactions Produits Durée
POST FIXATION Formol 1/10eme 45 mn
CIRCULATION Acétone I 45 mn
Acétone II 45 mn
Acétone III 45 mn
Toluène I 45 mn
Toluène II 45 mn
Toluène III 45 mn
Paraffine I 24 H
Paraffine II 1 H
INCLUSION Paraffine à l’aide de moules métalliques.
Après la coupe au microtome les lames ont été réalisées selon le tableau 2 :
II : Matériel e méthodes
73
Réactions Produits Durée
Déparaffinage Toluène I 10 minutes
Toluène II 10 minutes
Réhydratation Ethanol 96 ° 30 secondes
Ethanol 70 ° 30 secondes
Ethanol 50 ° 30 secondes
Eau courante 1 minute
Coloration Hémalun de HARIS 5 minutes
Eau courante 1 minute
Eau acide 1 minute
Bicarbonate de Lithium 1 minute
Ethanol 50 ° 30 secondes
Eosine alcoolique 1 minute
Déshydratation Ethanol 96 ° 30 secondes
Ethanol 96 ° 30 secondes
Ethanol 96 ° 30 secondes
Acétone 5 minutes
Eclaircissement Toluène III 10minutes
Toluène IV 10 minutes
Montage Lamelle montée sur lame avec de l’Eukitt
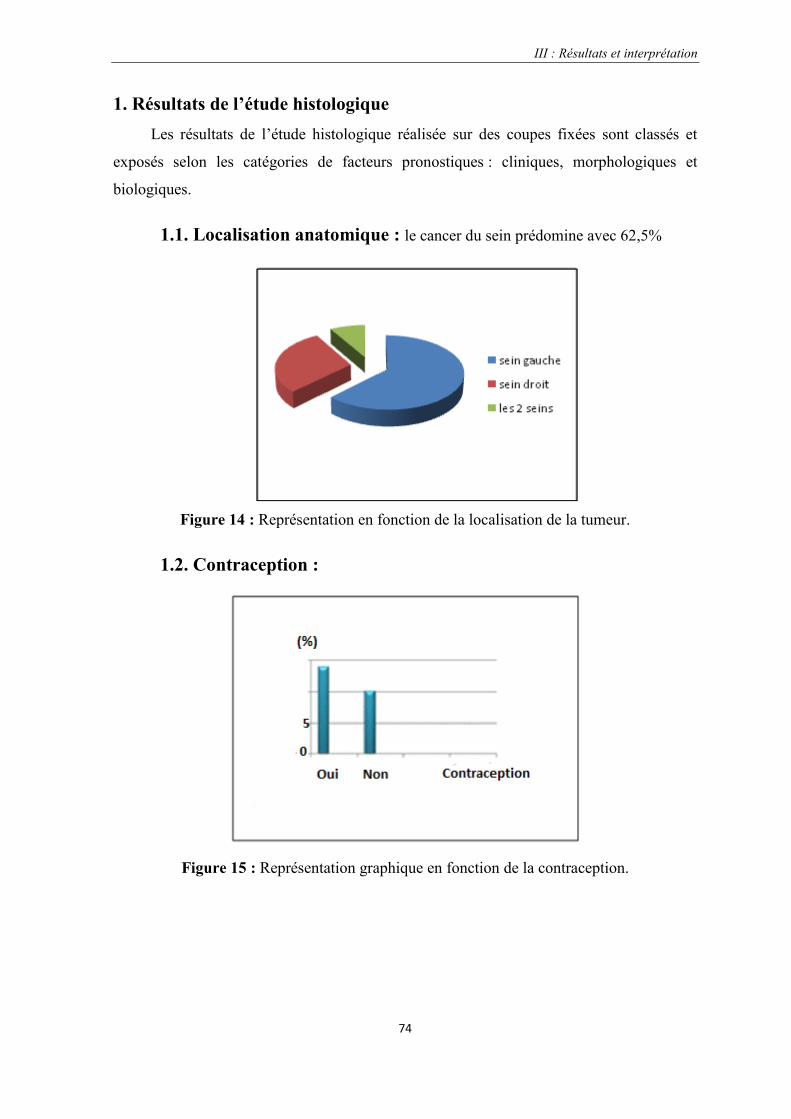
III : Résultats et interprétation
74
1. Résultats de l’étude histologiqueLes résultats de l’étude histologique réalisée sur des coupes fixées sont classés et
exposés selon les catégories de facteurs pronostiques : cliniques, morphologiques et
biologiques.
1.1. Localisation anatomique : le cancer du sein prédomine avec 62,5%
Figure 14 : Représentation en fonction de la localisation de la tumeur.
1.2. Contraception :
Figure 15 : Représentation graphique en fonction de la contraception.
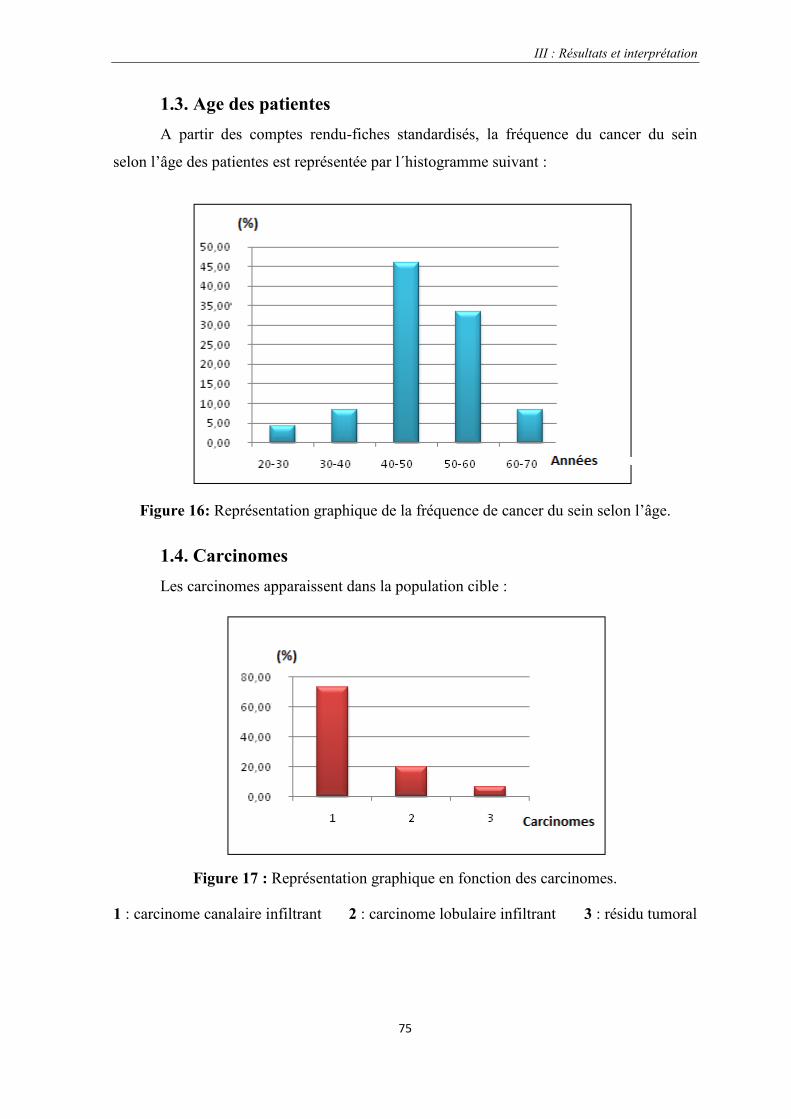
III : Résultats et interprétation
75
1.3. Age des patientesA partir des comptes rendu-fiches standardisés, la fréquence du cancer du sein
selon l’âge des patientes est représentée par l´histogramme suivant :
Figure 16: Représentation graphique de la fréquence de cancer du sein selon l’âge.
1.4. CarcinomesLes carcinomes apparaissent dans la population cible :
Figure 17 : Représentation graphique en fonction des carcinomes.
1 : carcinome canalaire infiltrant 2 : carcinome lobulaire infiltrant 3 : résidu tumoral

III : Résultats et interprétation
76
1.5. Grade
Figure 18 : Représentation des grades apparus dans la population cible.
2. Résultats de l’étude sérologiqueDans cette partie du travail, nous avons exploré les différents marqueurs
sérologiques dirigés contre les différentes protéines virales : les VCA, les anticorps anti
EBNA et les anticorps anti EA (antigène précoce) par trois techniques différentes :
technique Elisa, Immunofluorescence et western blot
Ces résultats ne montrent pas de différence significative entre elles ; la majorité des
sérums explorés, ont tous affichés des profils sérologiques positifs cependant, les titrages
des différents anticorps recherchés n’ont pu être réalisé par manque de réactifs pour
estimer et quantifier significativement le taux de positivité, cependant la densité optique
nous a permis de faire une comparaison qualitative des différents sérums explorés.
Sur les 24 sérums testés nous avons obtenue les résultats suivant :
Pour les anticorps IgG VCA, 20 sur 24 patientes ont affiché un profil sérologique
positif ce qui représente 83,33% de positivité, cette dernière a été estimée à partir d’une
densité optique supérieure à 0,483, ainsi 5 sérums ont montré une densité optique située
entre 2 et 2,8 ; 9 sérums avaient une densité optique située entre 1 et 1,90 ; 6 sérums entre
0,4 et 1. Parmi les 15 témoins (12 femmes saines et 3 patientes atteintes de cancer du
nasopharynx), étudiés pour ces paramètres, 5 d’entre elles, y compris les 3 NPC
présentaient une positivité (fig. 19).
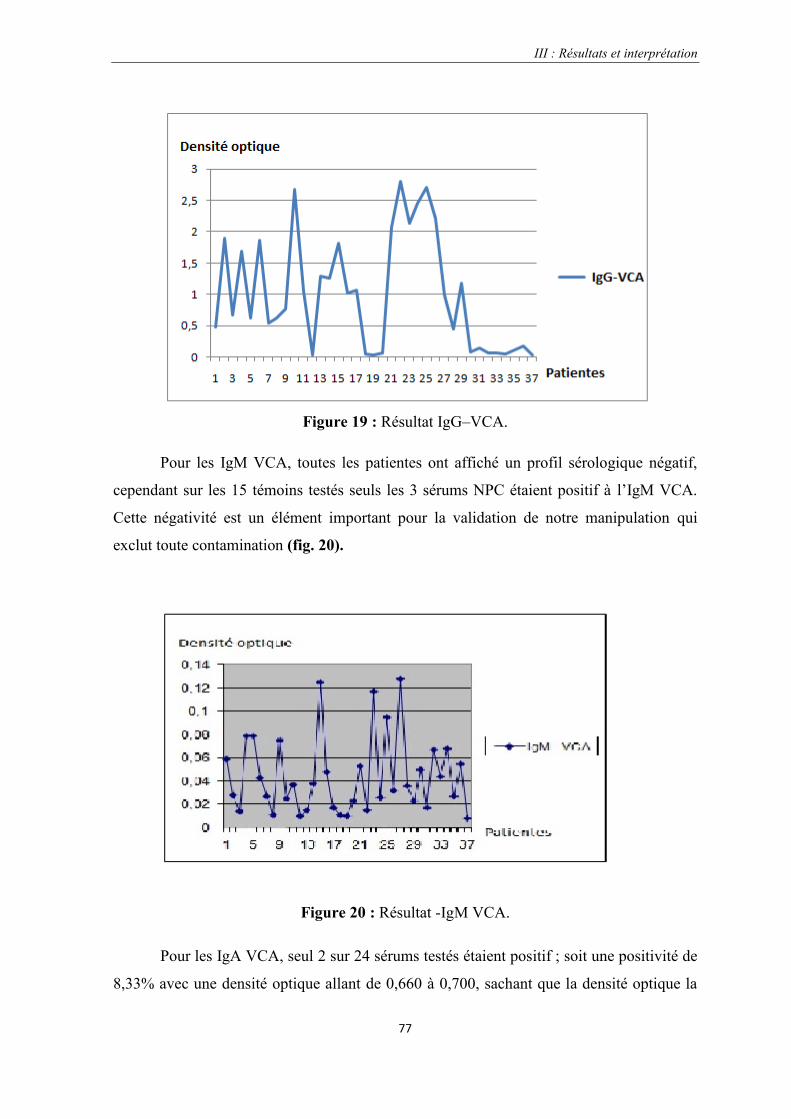
III : Résultats et interprétation
77
Figure 19 : Résultat IgG–VCA.
Pour les IgM VCA, toutes les patientes ont affiché un profil sérologique négatif,
cependant sur les 15 témoins testés seuls les 3 sérums NPC étaient positif à l’IgM VCA.
Cette négativité est un élément important pour la validation de notre manipulation qui
exclut toute contamination (fig. 20).
Figure 20 : Résultat -IgM VCA.
Pour les IgA VCA, seul 2 sur 24 sérums testés étaient positif ; soit une positivité de
8,33% avec une densité optique allant de 0,660 à 0,700, sachant que la densité optique la
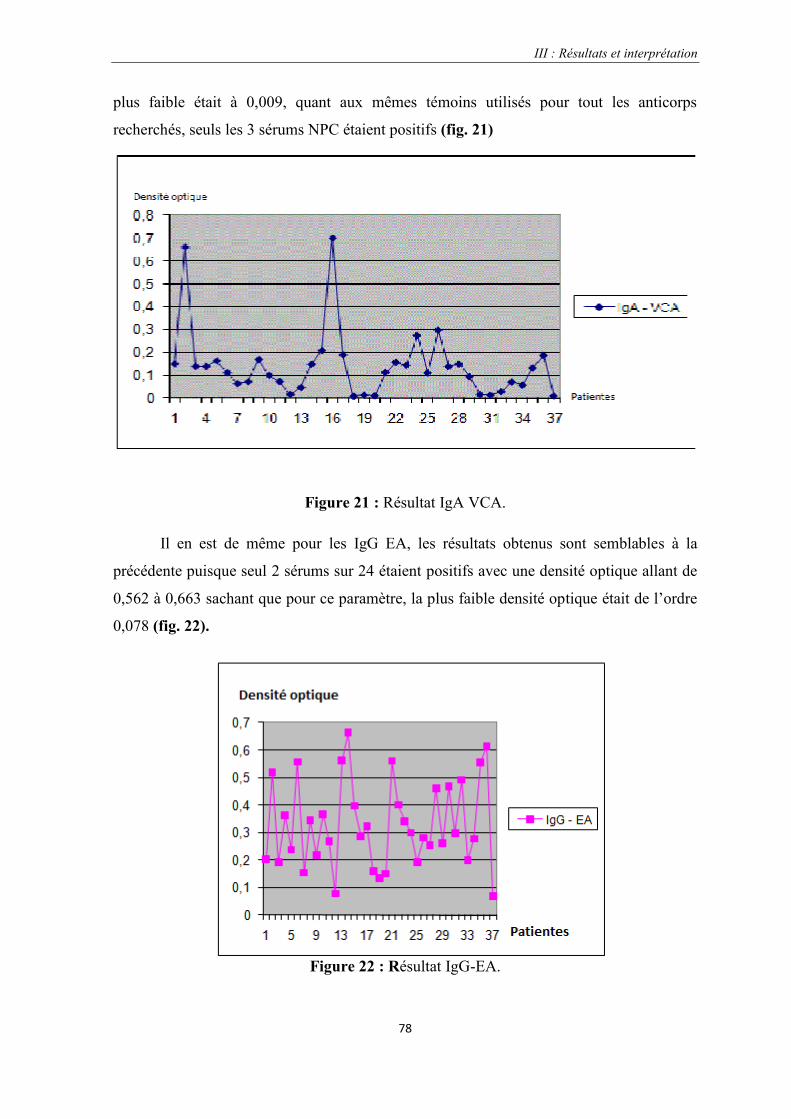
III : Résultats et interprétation
78
plus faible était à 0,009, quant aux mêmes témoins utilisés pour tout les anticorps
recherchés, seuls les 3 sérums NPC étaient positifs (fig. 21)
Figure 21 : Résultat IgA VCA.
Il en est de même pour les IgG EA, les résultats obtenus sont semblables à la
précédente puisque seul 2 sérums sur 24 étaient positifs avec une densité optique allant de
0,562 à 0,663 sachant que pour ce paramètre, la plus faible densité optique était de l’ordre
0,078 (fig. 22).
Figure 22 : Résultat IgG-EA.
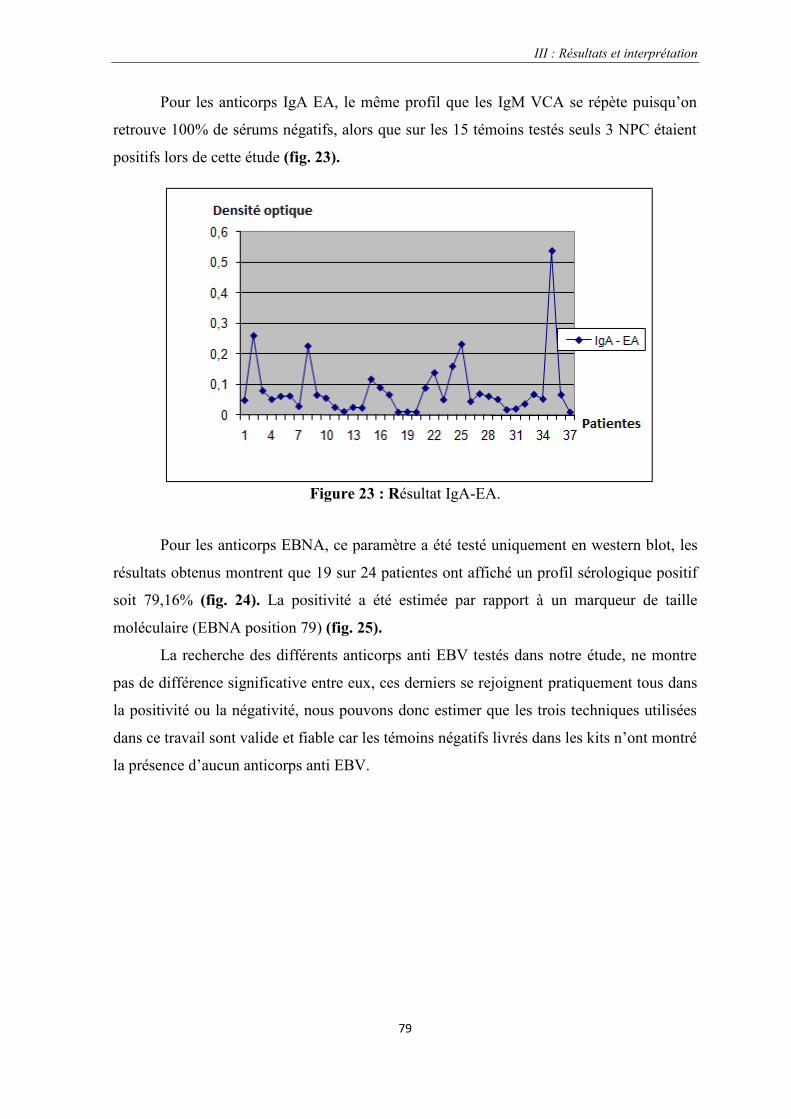
III : Résultats et interprétation
79
Pour les anticorps IgA EA, le même profil que les IgM VCA se répète puisqu’on
retrouve 100% de sérums négatifs, alors que sur les 15 témoins testés seuls 3 NPC étaient
positifs lors de cette étude (fig. 23).
Figure 23 : Résultat IgA-EA.
Pour les anticorps EBNA, ce paramètre a été testé uniquement en western blot, les
résultats obtenus montrent que 19 sur 24 patientes ont affiché un profil sérologique positif
soit 79,16% (fig. 24). La positivité a été estimée par rapport à un marqueur de taille
moléculaire (EBNA position 79) (fig. 25).
La recherche des différents anticorps anti EBV testés dans notre étude, ne montre
pas de différence significative entre eux, ces derniers se rejoignent pratiquement tous dans
la positivité ou la négativité, nous pouvons donc estimer que les trois techniques utilisées
dans ce travail sont valide et fiable car les témoins négatifs livrés dans les kits n’ont montré
la présence d’aucun anticorps anti EBV.
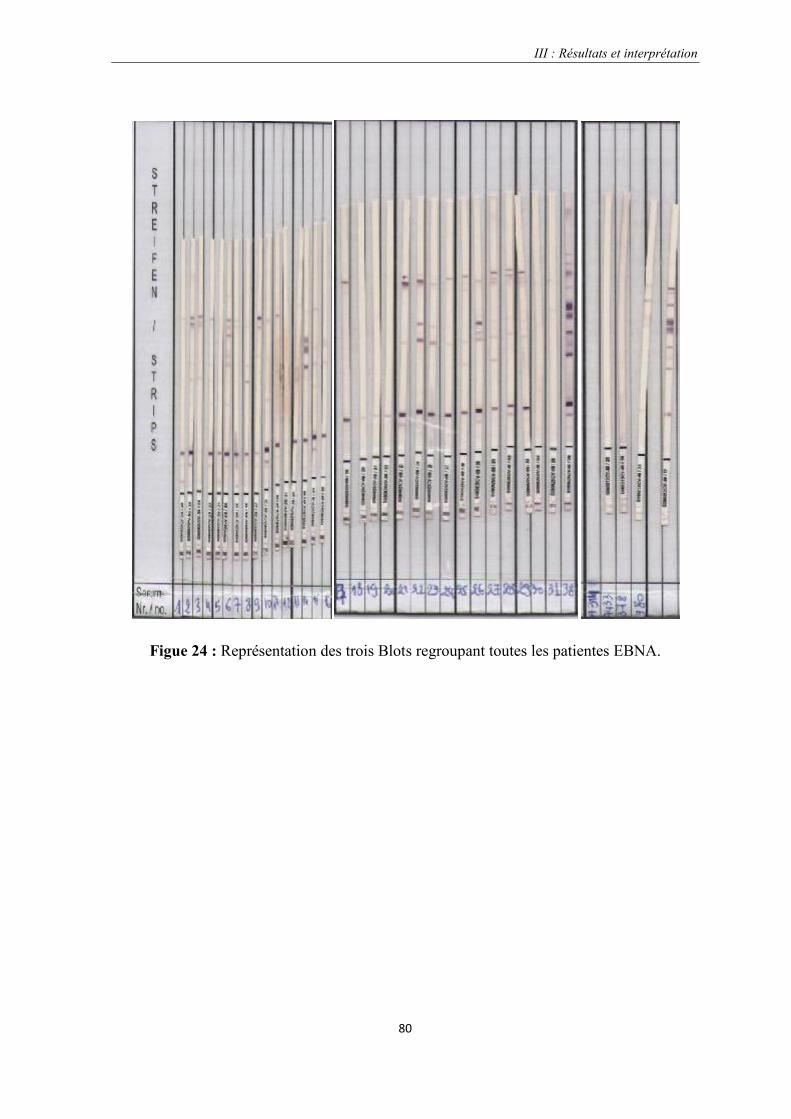
III : Résultats et interprétation
80
Figue 24 : Représentation des trois Blots regroupant toutes les patientes EBNA.
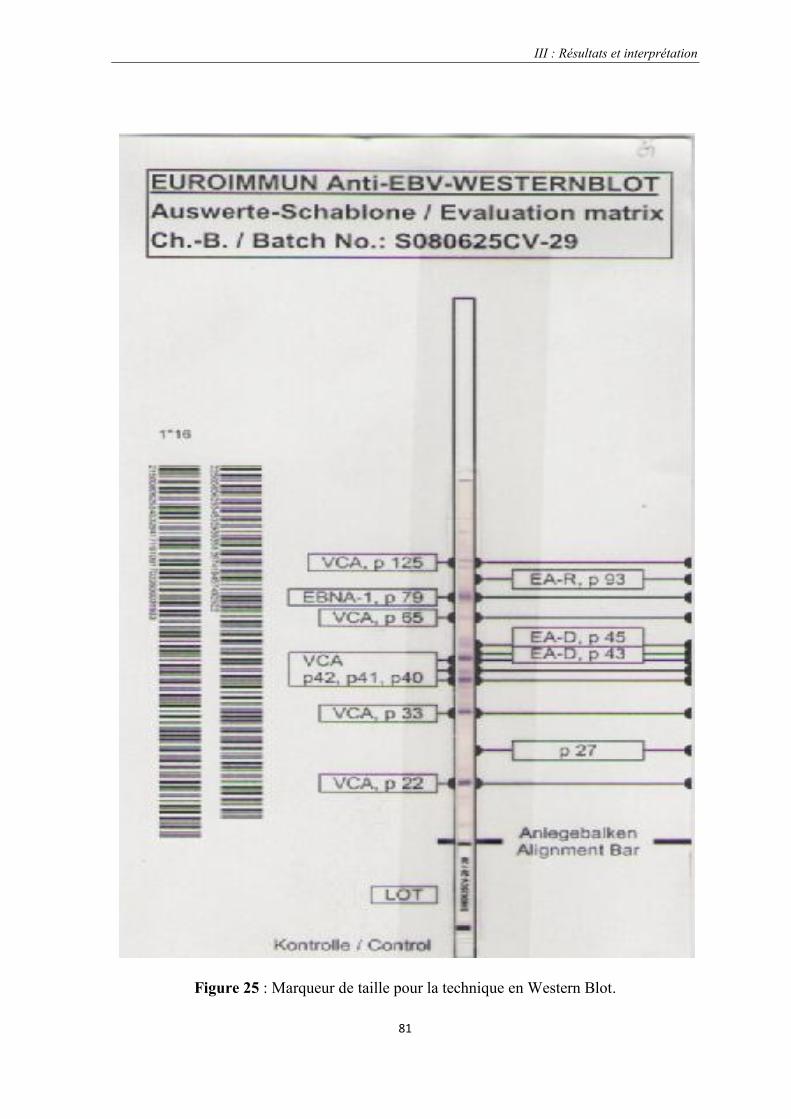
III : Résultats et interprétation
81
Figure 25 : Marqueur de taille pour la technique en Western Blot.

CHAPITR IV DISCUSSION, CONCLUSION ET PERSPECTIVE
82
Discussion et conclusion
Nous avons abordé dans ce mémoire un des cancers les plus meurtriers chez la femme,
il s’agit du cancer du sein, en effet ce cancer est le plus répandu dans le monde avec 1,15
millions de nouveaux cas par an et une mortalité de 410.000 (Kamangar F et al 2006).
En Algérie ce cancer occupe la première place parmi tous les cancers féminins et
constitue la première cause de mortalité par cancer. Un grand nombre de cancers du sein à un
stade avancé avec métastases ne sont pas diagnostiqués dans beaucoup de pays, en raison de
conditions socioéconomiques défavorables.
Il s’agit d’une maladie dont la compréhension est sombre, son étiologie est encore très
mal élucidée avec une multitude de facteur de risque, cependant de nombreux cas de cancer
du sein ne sont liés à aucun facteur de risque et la cause de leur survenue est inconnue.
(Welcsh P.L et al 2001)
Depuis quelques années les facteurs viraux, comme l’infection précoce par l’EBV mais
aussi par d’autres virus ont été rapportés dans la genèse des carcinomes mammaires. Des
études menées par plusieurs équipes de recherche considèrent l’EBV comme cofacteur de
cancer du sein (Arbach et Joab 2005 ; Arbach et al 2006, Derkaoui et al, 2008), il s’agit d’un
virus oncogène lié aux lymphomes comme les NPC et au carcinome, comme le carcinome
gastrique dans plus de 30% et carcinome mammaires dans 20 à 50% des cas.
In vivo l’infection par ce virus permet d’établir une infection latente persistante dans
l’organisme toute la vie, les cellules infectées vont exprimer peu de protéines virales ce qui
leurs permettra d’échapper à la surveillance du système immunitaire. Cependant cette phase
est interrompue par la réactivation du virus ce qui va aboutir à la production de nouvelles
particules virales, destruction de la cellule hôte et survenue de cancer, on peut donc considérer
que les cancers induits par l’EBV sont le reflet d’un déficit sévère du système immunitaire.
(Pagano et al… 2004).
Le travail présenté dans ce mémoire est une continuation au travail déjà mené par
Madame Derkaoui et qui a fait l’objet de sa thèse de doctorat.

CHAPITR IV DISCUSSION, CONCLUSION ET PERSPECTIVE
83
Nous avons essayé d’apporter un éclaircissement quant à la présence des marqueurs
sérologiques tumoraux impliqués dans le cancer du sein. Notre étude a porté sur 24 patientes
toutes atteintes de cancer du sein et issues de l’Ouest Algérien.
Les résultats obtenus a l’issu de l’étude histologique font ressortir une atteinte
prédominante du sein gauche avec 62.5% par rapport au sein droit. (Figure14)
Les travaux de Diallot et al (1996) ont retrouvé une atteinte de 54,32% de seins gauches,
contre 43,68% de seins droits.
L’âge d’apparition du cancer du sein varie entre 28 et 73 ans. Parmi les 24 patientes, la plus
part avait un âge comprit entre 40 et 50 ans avec une moyenne de 45.83%.( Figure16)
Bekkouche et al (2000) rapportent l’âge des patientes compris entre 25 et 78 ans, l’âge moyen
étant de 47 ans. Dans l’étude de 2007, ces mêmes auteurs retrouvent un âge moyen de 48 ans.
D’après Delozier (2002) la majorité des patientes étudiées (68%) est âgée de plus de 50 ans
(âge médian 57 ans).Dans l’étude de l’équipe tunisienne Ben Ahmed et al (2002), l’âge
moyen au diagnostic était de 50 ans avec un pic de fréquence entre 41et 50 ans (46,88%).
Senhadji (2004) rapporte une moyenne d’âge de 43 ans.
Zaoui et al (2009) ont trouvé que la tranche d’âge la plus touchée est comprise entre 40 et 50
ans (46%) avec une moyenne de 44 ±11.93 ans.
Le type carcinome canalaire infiltrant a été retrouvé chez 11 patientes avec un pourcentage de
73.33% contre 20% de carcinome lobulaire infiltrant qui a été retrouvé chez 3 patientes, le
type carcinome canalaire infiltrant de type comédocarcinome de grade III a été retrouvé chez
10 patientes soit 46.66%, Il ressort que le type histologique le plus répandu dans notre étude
est le CCI. (Figure17) . Selon Spielmann et al. (2006) pour le groupe SBR I et II, la survie à
5 ans est respectivement de 94% et 91%. Pour le groupe SBR III, la survie est de 84% pour
les patientes traitées par chimiothérapie et de 79% pour les patientes non traitées (Figure18).
Bekkouche et al. (2000) ont retrouvé 3% de grade SBR I, 74% de grade SBR II et 23% de
grade SBR III. Delozier (2002) remarque l’incidence élevée des grades SBR I (39%) pour les
cancers non métastatiques.
Notre travail a été axé principalement sur l’étude sérologique. Trois techniques
différentes ont été nécessaires à la réalisation de cette partie la technique
ELISA,IFI ,Westernblot ; les différents anticorps anti EBV recherchés ont montré peu ou pas

CHAPITR IV DISCUSSION, CONCLUSION ET PERSPECTIVE
84
de différences significatives entre eux, ces derniers se rejoignent pratiquement tous dans la
positivité ou la négativité.
Cependant un des marqueurs sérologique exploré ressort plus positif que les autres, il
s’agit de l’anticorps IgG VCA, positif chez 20 patientes sur 24, ce qui nous poussent à penser
soit à la conséquence d’une phase de réactivation virale et donc cancérisation ou tout
simplement une infiltration lymphocytaire.
Les travaux de Derkaoui et ses collaborateurs (2008) sur la sérologie EBV, ont retrouvé
que sur 61 sérums testés, 26 (42.62%) ont montré un profil sérologique normale, 24 sérums
étaient positifs en IgG VCA et EA IgG, 6 sérums (9.83%) ont montré un titre élevé en IgG
VCA et 18 (29.5%) était positif en EA IgG avec 3 patientes ayant un titre très élevé en IgG-
EA.
Nous pouvons conclure que nos résultats sont en concordance avec ceux retrouvé dans
la littérature puisqu’il a été rapporté que l’infection par l’EBV se produit dans la petite
enfance et la séropositivité est presque invariable avant l’âge adulte.
Dans les pays endémiques en EBV, la séroconversion se produit généralement tard dans
la vie et seulement 60 à 70% des adultes sont séropositifs. Une étude cas-témoins a souligné
un risque élevé chez des femmes ayant eu tardivement la MNI de développer un cancer du
sein (Yasui et, 2001). Par conséquent, pour tester cette hypothèse, il est important de
déterminer le statut immunitaire de ce virus chez des femmes saines.
L’étude sérologique menée par Richardson et al, 2004 sur 208 patientes atteintes de
cancer du sein par rapport à une population témoin saine de 169 femmes à la recherche de
l’anticorps VCA-IgG par technique Elisa, montre qu’il n’y a aucune différence significative
entre les taux de positivité chez les deux groupes , il suggère alors qu’il n’y a aucune preuve
palpable quant à l’association entre des taux élevés d’IgG EBV et le cancer du sein .
Dans le même contexte, Rickinson et al, 2007 affirment que des antigènes exprimés lors
d’une infection par l’EBV en phase proliférative tels que les EA et les VCA, mais aussi ceux
exprimés en phase d'infection latente comme EBNA, LMP peuvent déclencher des réponses
immunitaires et produire ainsi la production d'anticorps spécifiques, cette idée est confortée

CHAPITR IV DISCUSSION, CONCLUSION ET PERSPECTIVE
85
par le fait que le taux des anticorps anti EBV comme VCA IgG, EBNA sont
systématiquement élevés dans les infections dues à l’EBV.
En 2009 une étude menée par JIAN-RONG HE et al. sur la relation EBV cancer du sein
infirme toute relation entre ces deux paramètres , il en est de même que l’étude de Angeloni et
al, sur 71 patientes atteintes de cancer du sein où la protéine BFRF1 (protéine de réplication) a
été retrouvée négative dans tout les sérums testés mais des résultats positifs concernant la
même protéine ont été observés chez les patients NPC et des Burkitt, ce qui rejoint l’idée que
l’infection par l’EBV dans les carcinomes mammaires reste encore inconnu et suscite de plus
en plus de travaux sur une cohorte importante , avec en premier lieu l’identification du virus
lui-même comme agent inducteur de cancer mammaire par des techniques fines et fiables
telles que la PCR Quantitative en Temps Réel, mais aussi la technique des micro-array
(Derkaoui et al 2008).
L’EBV peut avoir un rôle dans le cancer du sein humain mais son rôle possible comme
facteur étiologique reste à déterminer.
La sérologie virale est considérée comme un excellent marqueur tumoral qui peut être
utilisé comme moyen diagnostique dans les cancers viro induit ce qui pourrait contribuer à la
prévention de ces cancers à un stade précoce.

CHAPITR IV DISCUSSION, CONCLUSION ET PERSPECTIVE
86
Perspectives
Malgré toutes les avancées scientifiques concernant la compréhension et la prise
en charge du cancer du sein, hélas cette pathologie continue a frapper de plus en plus
des femmes à tout âge et surtout des sujets jeunes représentant ainsi la première cause de
mortalité par cancer .
Le cancer du sein reste donc une pathologie énigmatique avec une multitude de
facteurs de risque connue tel que : la génétique, l’environnement, les hormones, les
rayonnements ultraviolets, les virus, etc. mais aussi d’autres facteurs encore inconnus
jusqu'à maintenant.
Tous ces paramètres, représentent un obstacle majeur dans la prise en charge du cancer
du sein d’autant plus difficile dans l’ouest Algérien , à cause des conditions socio-
économiques des patientes qui sont le plus souvent défavorables, ce qui ne les poussent
pas à consulter à temps, ceci conduit inévitablement à la propagation de la maladie et quand
ces patientes arrivent au centre de soin; la maladie est déjà a un stade très avancé ne
laissant aucune chance à la survie.
Le cancer du sein en Algérie est un véritable problème de santé publique d’où la
nécessité d’alerter les pouvoirs publiques pour cerner cette épidémie.
Il est donc impératif de créer une équipe multidisciplinaire composé de
médecins, chirurgiens, chercheurs, biologistes et surtout psychologues pour
une prise en charge optimale des patientes.
Création de registre de cancer regroupant toutes les patientes issues de l’ouest
algérien, ceci peut être réalisé grâce au concours et aux efforts de toutes les
équipes médicale et paramédicale mené par un réseau d’épidémiologistes et
statisticiens
Impliquer de plus en plus de virologues au diagnostic des cancers viro induits,
banaliser les techniques sérologiques telles que les techniques Elisa et immuno
assays.

CHAPITR IV DISCUSSION, CONCLUSION ET PERSPECTIVE
87
Accéder aux nouveaux moyens diagnostiques tel que la PCR Quantitative en
Temps Reel qui nous fournit une amplitude de renseignements quant à
l’amplification et la détection des gènes viraux mais surtout la quantification la
charge virale.
Sur le plan recherche, il est nécessaire de regarder les profils moléculaires tels
que l’établissement de carte d’identité moléculaire des tumeurs du sein pour
déboucher sur une classification plus fine de la maladie.
A la fin il serait nécessaire de faciliter l’accès au diagnostic du cancer du sein en offrant
les moyens de dépistage en créant une unité mobile se déplaçant dans les zones rurale et
isolées des centres de soins afin d’offrir la chance à ces patientes de consulter à temps et
accéder aux différents moyens d’imagerie tels que la mammographie, l’échographie et à titre
gratuit pour les patientes les plus démunies , ceci permettra peut être de minimiser cette
épidémie et surtout permettre d’établir un diagnostic précoce

Annexe
Annexe 1

Annexe
Annexe 2
Consentement éclairé
Je soussigné (e)………………………………………………………………………….
Avoir pris connaissance du document m’informant que des praticiens de la santé ainsi
qu’un groupe de chercheur ont pour mission de prélever des échantillons biologique
(sérum, cellules, tissus et tumeurs), sur ma personne, et que ces prélèvement seront
utilisés à des fin purement scientifique, ce qui permettra de faire avancer la recherche
médicale et la compréhension des mécanismes sur le processus cancéreux, les
nouvelles connaissance sur le cancer sont le plus souvent produite grâce au
rapprochement des résultats d’analyses biologique avec des observations clinique de
la maladie et de son évolution.
Les résultats des études sont confidentielles et de ce fait ne seront transmis qu’a moi-
même et en aucun cas à un autre membre de ma famille, et que les informations
utilises dans cette recherche ne permettrons en aucun cas de m’identifier.
De ce fait l’anonymat des patients faisant objet de cette étude est strictement préservé
et les informations sont confidentielle et ne seront partagés qu’au sein de la
communauté scientifique.
Je considère donc disposer de toutes les informations qui me permette de décider de :
□ Donner mon accord pour que des prélèvements biologiques effectuer sur ma
personne soit utilisés à des fin s de recherche
□ Je m’oppose à ce que mes prélèvements biologiques soient utilisés à des fins de
recherche.
PATIENT MEDECIN Responsable de Recherche
Nom, prénom Nom, prénom Nom, prénom
Signature Signature Signature
Version : 2009-2010
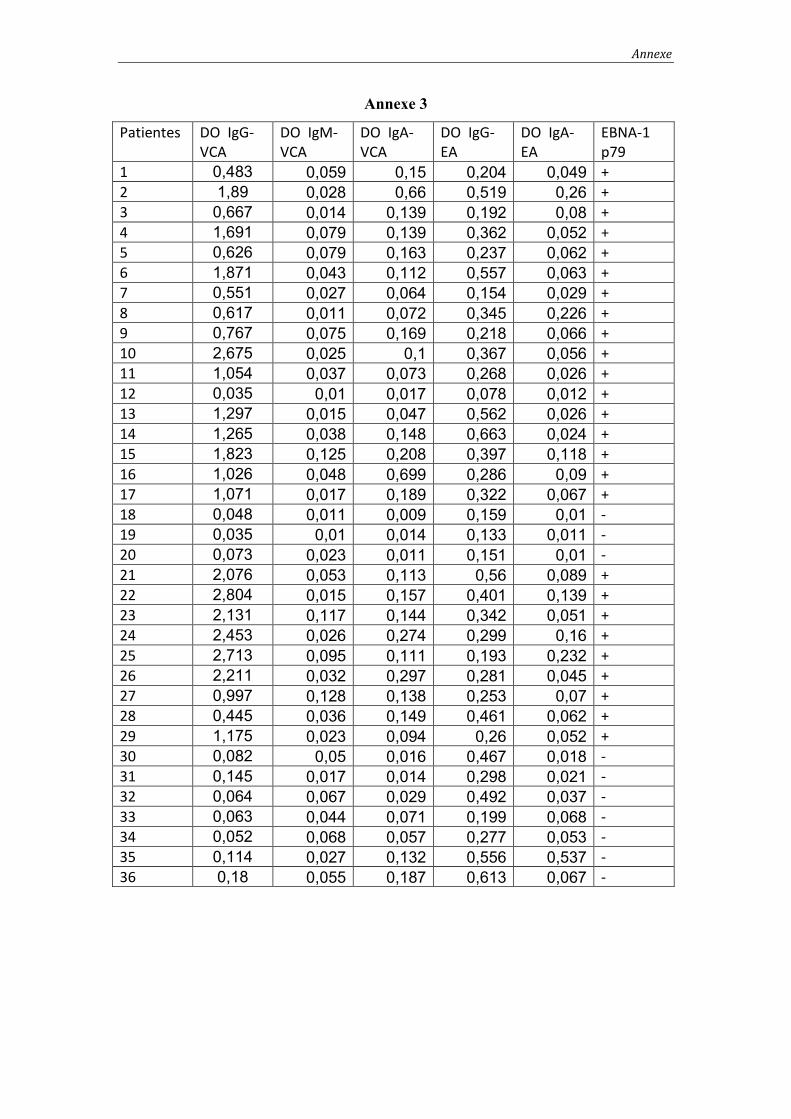
Annexe
Annexe 3
Patientes DO IgG-VCA
DO IgM-VCA
DO IgA-VCA
DO IgG-EA
DO IgA-EA
EBNA-1p79
1 0,483 0,059 0,15 0,204 0,049 +2 1,89 0,028 0,66 0,519 0,26 +3 0,667 0,014 0,139 0,192 0,08 +4 1,691 0,079 0,139 0,362 0,052 +5 0,626 0,079 0,163 0,237 0,062 +6 1,871 0,043 0,112 0,557 0,063 +7 0,551 0,027 0,064 0,154 0,029 +8 0,617 0,011 0,072 0,345 0,226 +9 0,767 0,075 0,169 0,218 0,066 +10 2,675 0,025 0,1 0,367 0,056 +11 1,054 0,037 0,073 0,268 0,026 +12 0,035 0,01 0,017 0,078 0,012 +13 1,297 0,015 0,047 0,562 0,026 +14 1,265 0,038 0,148 0,663 0,024 +15 1,823 0,125 0,208 0,397 0,118 +16 1,026 0,048 0,699 0,286 0,09 +17 1,071 0,017 0,189 0,322 0,067 +18 0,048 0,011 0,009 0,159 0,01 -19 0,035 0,01 0,014 0,133 0,011 -20 0,073 0,023 0,011 0,151 0,01 -21 2,076 0,053 0,113 0,56 0,089 +22 2,804 0,015 0,157 0,401 0,139 +23 2,131 0,117 0,144 0,342 0,051 +24 2,453 0,026 0,274 0,299 0,16 +25 2,713 0,095 0,111 0,193 0,232 +26 2,211 0,032 0,297 0,281 0,045 +27 0,997 0,128 0,138 0,253 0,07 +28 0,445 0,036 0,149 0,461 0,062 +29 1,175 0,023 0,094 0,26 0,052 +30 0,082 0,05 0,016 0,467 0,018 -31 0,145 0,017 0,014 0,298 0,021 -32 0,064 0,067 0,029 0,492 0,037 -33 0,063 0,044 0,071 0,199 0,068 -34 0,052 0,068 0,057 0,277 0,053 -35 0,114 0,027 0,132 0,556 0,537 -36 0,18 0,055 0,187 0,613 0,067 -

ReferencesAAdler, B., Schaadt, E., Kempkes, B., Zimber-Strobl, U., Baier, B. & Bornkamm,G.W. (2002). Control of Epstein-Barr virus reactivation by activated CD40 and virallatent membrane protein 1. Proceedings of the National Academy of Sciences of theUnited States of America, 99, pp. 437-442.
Anderson, L. J. & Longnecker, R. (2008). An auto-regulatory loop for EBVLMP2A involves activation of Notch. Virology, 371, pp. 257-266.
Angeloni A, Farina A, Gentile G, et al. (2001). Epstein-Barr virus and breastcancer: search for antibodies to the novel BFRF1 protein in sera of breast cancerpatients. J Natl Cancer Inst 93:560-1.
d'Amore F, Johansen P, Houmand A, Weisenburger DD, Mortensen LS.(1996).Epstein-Barr virus genome in non-Hodgkin's lymphomas occurring inimmunocompetent patients: highest prevalence in nonlymphoblastic T-cell lymphomaand correlation with a poor prognosis. Danish Lymphoma Study Group, LYFO.Blood. 87:1045-55.
Arbach H, Viglasky V, Lefeu F, Guinebretiere JM, Ramirez V, Bride N,Boualaga N, Bauchet T, Peyrat JP, Mathieu MC, Mourah S, Podgorniak MP,Seignerin JM, Takada K, Joab I. (2006). Epstein-Barr virus (EBV) genome andexpression in breast cancer tissue: effect of EBV infection of breast cancer cells onresistance to paclitaxel (Taxol). J Virol. 80:845-53.
Altmann, M., Pich, D., Ruiss, R., Wang, J., Sugden, B. & Hammerschmidt, W.(2006). Transcriptional activation by EBV nuclear antigen 1 is essential for theexpression of EBV's transforming genes. Proceedings of the National Academy ofSciences of the United States of America, 103, pp. 14188-14193.
Ahsan, N., T. Kanda, et al. (2005). "Epstein-Barr virus transforming protein LMP1plays a critical role in virus production." J Virol 79(7): 4415-24.
Aris, R. M., D. M. Maia, et al. (1996). "Post-transplantation lymphoproliferativedisorder in the Epstein-Barr virus-naive lung transplant recipient." Am J Respir CritCare Med 154(6 Pt 1): 1712-7.

BBaer R., Bankier A. T., Biggin M. D., Deininger P. L., Farrell P. J., Gibson T. J.,Hatfull G., Hudson G. S., Satchwell S. C., Seguin C., and et al. (1984). DNAsequence and expression of the B95-8 Epstein-Barr virus genome. Nature 310: 207-11.
Band V., Zajchowski D., Kulesa V., and Sager R. (1990). Human papilloma virusDNAs immortalize normal human mammary epithelial cells and reduce their growthfactor requirements. Proc Natl Acad Sci U S A 87: 463-7.
Blaschke S., Schwarz G., Moneke D., Binder L., Muller G., and Reuss-Borst M.(2000). Epstein-Barr virus infection in peripheral blood mononuclear cells, synovialfluid cells, and synovial membranes of patients with rheumatoid arthritis. J Rheumatol27: 866-73.
Bikfalvi A (2003). Tumor angiogenesis. Bull Cancer. 90: 449-58.
Bonnet M, Guinebretiere JM, Kremmer E, Grunewald V, Benhamou E,Contesso G, Joab I. Detection of Epstein-Barr virus in invasive breast cancers. J NatlCancer Inst. 91(16): 1376-81.
Bonnet-Duquennoy M, Arbach H, Vandest B, Lefeu F, Joab I. (2003). Abscenceof Estein-Barr virus genome in breast cancer-derived cell lines. J Natl Caner Inst.95:1254-1255.
Bratthauer G. L., Tavassoli F. A., and O'Leary T. J. (1992). Etiology of breastcarcinoma: no apparent role for papillomavirus types 6/11/16/18. Pathol Res Pract188: 384-6.
Brink A. A., ten Berge R. L., van den Brule A. J., Willemze R., Chott A., andMeijer C. J. (2000). Epstein-Barr virus is present in neoplastic cytotoxic T cells inextranodal, and predominantly in B cells in nodal T non-Hodgkin lymphomas. JPathol 191: 400-6.
Brousset P., Schlaifer D., Meggetto F., Bachmann E., Rothenberger S., Pris J.,Delsol G., and Knecht H. (1994). Persistence of the same viral strain in early andlate relapses of Epstein-Barr virus-associated Hodgkin's disease. Blood 84: 2447-51.
Brooks J. S. (1999). Differential diagnosis of thymic lesions. Acta Cytol 43: 527-9.
Busson P, Ganem G, Flores P, Mugneret F, Clausse B, Caillou B, Braham K,Wakasugi H, Lipinski M, Tursz T. (1988). Establishment and characterization ofthree transplantable EBV-containing nasopharyngeal carcinomas. Int J Cancer.42:599-606.
Bajaj, B. G., Murakami, M., Cai, Q., Verma, S. C., Lan, K. & Robertson, E.S.(2008). Epstein-Barr virus nuclear antigen 3C interacts with and enhances the stabilityof the c-Myc oncoprotein. Journal of virology, 82, pp. 4082-4090.

Burkitt, D. (1958). A sarcoma involving the jaws in African children. The Britishjournal of surgery, 46, pp. 218-223.
Babcock, G. J., Hochberg, D. & Thorley-Lawson, A.D. (2000). The expressionpattern of Epstein-Barr virus latent genes in vivo is dependent upon the differentiationstage of the infected B cell. Immunity, 13, pp. 497-506..Balandraud, N., Roudier, J. & Roudier, C. (2004). Epstein-Barr virus andrheumatoid arthritis. Autoimmunity reviews, 3, pp. 362-367.
Bandobashi, K., Maeda, A., Teramoto, N., Nagy, N., Székely, L., Taguchi, H.,Miyoshi, I., Klein, G. & Klein, E. (2001). Intranuclear localization of thetranscription coadaptor CBP/p300 and the transcription factor RBP-Jk in relation toEBNA-2 and -5 in B lymphocytes. Virology, 288, pp. 275-282.
Bhende, P. M., Seaman, W. T., Delecluse, H. & Kenney, S.C. (2005). BZLF1activation of the methylated form of the BRLF1 immediate-early promoter isregulated by BZLF1 residue 186. Journal of virology, 79, pp. 7338-7348.
Brielmeier, M., Mautner, J., Laux, G. & Hammerschmidt, W. (1996). The latentmembrane protein 2 gene of Epstein-Barr virus is important for efficient B cellimmortalization. The Journal of general virology, 77 ( Pt 11), pp. 2807-2818.
Brooks, L. A., Lear, A. L., Young, L. S. & Rickinson, A.B. (1993). Transcriptsfrom the Epstein-Barr virus BamHI A fragment are detectable in all three forms ofvirus latency. Journal of virology, 67, pp. 3182-3190.
Boisserie-Lacroix M, Lafitte JJ, Sirben C, Latrabe V, Grelet P, Zeinoun R, et al.Les lésions inflammatoires et infectieuses du sein : contribution de l'échographie. JChir (Paris) 1993;130(10):408-15
Buchberger W, Strasser K, Heim K, Muller E, Schrocksnadel H. Phylloidestumor: findings on mammography, sonography, and aspiration cytology in 10 cases.AJR Am J Roentgenol 1991; 157: 715-9
Bazzocchi F, Santini D, Martinelli G, et al. Juvénile papillomatosis (épitheliosis) ofthe breast. Am J Clin Pathol 1986;86:745-8.
Bell, A. I., K. Groves, et al. (2006). "Analysis of Epstein-Barr virus latent geneexpression in endemic Burkitt's lymphoma and nasopharyngeal carcinoma tumourcells by using quantitative real-time PCR assays." J Gen Virol 87(Pt 10): 2885-90
Bultema, R., R. Longnecker, et al. (2009). "Epstein-Barr virus LMP2A acceleratesMYC-induced lymphomagenesis." Oncogene 28(11): 1471-6.
Brown, H. J., M. J. Song, et al. (2003). "NF-kappaB inhibits gammaherpesvirus lyticreplication." J Virol 77(15): 8532-40.

Baldanti, F., P. Grossi, et al. (2000). "High levels of Epstein-Barr virus DNA inblood of solid-organ transplant recipients and their value in predicting posttransplantlymphoproliferative disorders." J Clin Microbiol 38(2): 613-9
Brooks, L., Q. Y. Yao, et al. (1992). "Epstein-Barr virus latent gene transcription innasopharyngeal carcinoma cells: coexpression of EBNA1, LMP1, and LMP2transcripts." J Virol 66(5): 2689-97.
Burke, A. P., T. S. Yen, et al. (1990). "Lymphoepithelial carcinoma of the stomachwith Epstein-Barr virus demonstrated by polymerase chain reaction." Mod Pathol3(3): 377-80.
CChang K. L., and Weiss L. M. (1996). The association of the Epstein-Barr viruswith malignant lymphoma. Biomed Pharmacother 50: 459-67.
Chu PG, Chang KL, Chen YY, Chen WG, Weiss LM. (2001) No significantassociation of Epstein-Barr virus infection with invasive breast carcinoma. Am JPathol 159: 571-8.
Cohen J. I., and Lekstrom K. (1999). Epstein-Barr virus BARF1 proteinidispensable for B-cell transformation and inhibits alpha interferon secretion frommononuclear cells. J Virol 73: 7627-32.
Carmeliet P, Jain RK (2000). Angiogenesis in cancer and other diseases. Nature.407: 249-57.
Carmeliet P. (2000). Mechanisms of angiogenesis and arteriogenesis. Nat Med.6:389-95.
Chang Y, Tung CH, Huang YT, Lu J, Chen JY, and Tsai CH. (1999).Requirement for cell-to-cell contact in Epstein-Barr virus infection of nasopharyngealcarcinoma cells and keratinocytes. J Virol. 73: 8857-66.
Cheung ST, Huang DP, Hui AB, Lo KW, Ko CW, Tsang YS, Wong N, WhitneyBM, Lee JC. (1999). Nasopharyngeal carcinoma cell line (C666-1) consistentlyharbouring Epstein-Barr virus. Int J Cancer. 83:121-6.
Cirone M, De Maria R, D'Alessandro A, Frati L, Faggioni A, Ragona G. (1995).Epstein-Barr virus DNA is present both in CD10/CD77 positive and negative subsetsof human tonsillar lymphocytes. Cancer Lett. 89:125-8.
Conway EM, Collen D, Carmeliet P (2001). Molecular mechanisms of bloodvessel growth. Cardiovasc Res 49: 507-21.

Chevallier-Greco, A., Gruffat, H., Manet, E., Calender, A. & Sergeant, A. (1989).The Epstein-Barr virus (EBV) DR enhancer contains two functionally differentdomains: domain A is constitutive and cell specific, domain B is transactivated by theEBV early protein R. Journal of virology, 63, pp. 615-623.
Cohen, J. I., Wang, F., Mannick, J. & Kieff, E. (1989). Epstein-Barr virus nuclearprotein 2 is a key determinant of lymphocyte transformation. Proceedings of theNational Academy of Sciences of the United States of America, 86, pp. 9558-9562.
Contreras-Brodin, B., Karlsson, A., Nilsson, T., Rymo, L. & Klein, G. (1996). Bcell-specific activation of the Epstein-Barr virus-encoded C promoter compared withthe wide-range activation of the W promoter. The Journal of general virology, 77 ( Pt6), pp. 1159-1162.
Carter,C.L., Corle,D.K., Micozzi,M.S., Schatzkin,A., and Taylor,P.R. (1988). Aprospective study of the development of breast cancer in 16,692 women with benignbreast disease. Am. J. Epidemiol. 128, 467-477.
Claus,E.B., Risch,N., and Thompson,W.D. (1991). Genetic analysis of breastcancer in the cancer and steroid hormone study. Am. J. Hum. Genet. 48, 232-242
Cotter, M. A. 2. & Robertson, E.S. (2000). Modulation of histone acetyltransferaseactivity through interaction of epstein-barr nuclear antigen 3C with prothymosin alpha.Molecular and cellular biology, 20, pp. 5722-5735.
Cesarman, E., A. Chadburn, et al. (1998). "BCL-6 gene mutations inposttransplantation lymphoproliferative disorders predict response to therapy andclinical outcome." Blood 92(7): 2294-302.
Cochet, C., D. Martel-Renoir, et al. (1993). "Expression of the Epstein-Barr virusimmediate early gene, BZLF1, in nasopharyngeal carcinoma tumor cells." Virology197(1): 358-65.
Chan, K. C. and Y. M. Lo (2002). "Circulating EBV DNA as a tumor marker fornasopharyngeal carcinoma." Semin Cancer Biol 12(6): 489-96.
Cheung, M. M., J. K. Chan, et al. (2003). "Natural killer cell neoplasms: adistinctive group of highly aggressive lymphomas/leukemias." Semin Hematol 40(3):221-32.
Cheuk, W., N. Walford, et al. (2001). "Primary histiocytic lymphoma of the centralnervous system: a neoplasm frequently overshadowed by a prominent inflammatorycomponent." Am J Surg Pathol 25(11): 1372-9.

Dd'Amore F., Johansen P., Houmand A., Weisenburger D. D., and Mortensen L.S. (1996). Epstein-Barr virus genome in non-Hodgkin's lymphomas occurring inimmunocompetent patients: highest prevalence in nonlymphoblastic T-cell lymphomaand correlation with a poor prognosis. Danish Lymphoma Study Group, LYFO. Blood87: 1045-55.
Dawson C. W., Dawson J., Jones R., Ward K., and Young L. S. (1998). Functionaldifferences between BHRF1, the Epstein-Barr virus-encoded Bcl-2 homologue, andBcl-2 in human epithelial cells. J Virol 72: 9016-24.
Delecluse H. J., Marafioti T., Hummel M., Dallenbach F., Anagnostopoulos I.,and Stein H. (1997). Disappearance of the Epstein-Barr virus in a relapse ofHodgkin's disease. J Pathol 182: 475-9.
Di Lonardo A, Venuti A, Marcante ML. (1992). Human papillomavirus in breastcancer. Breast Cancer Res Treat. 21(2): 95-100.
Dolcetti R., and Menezes J. (2003). Epstein-Barr virus and undifferentiatednasopharyngeal carcinoma: new immunobiological and molecular insights on a long-standing etiopathogenic association. Adv Cancer Res 87: 127-57.
Dumitrescu,R.G., and Cotarla,I. 2005. Understanding breast cancer risk -- where dowe stand in 2005? J. Cell Mol.Med. 9:208-221.
Dreyfus, D. H., M. Nagasawa, et al. (1999). "Inactivation of NF-kappaB by EBVBZLF-1-encoded ZEBRA protein in human T cells." J Immunol 163(11): 6261-8.
Deeg, H. J. and G. Socie (1998). "Malignancies after hematopoietic stem celltransplantation: many questions, some answers." Blood 91(6): 1833-44.
Diebold, J., M. Raphael, et al. (1997). "Lymphomas associated with HIV infection."Cancer Surv 30: 263-93.
EEliceiri BP, Paul R, Schwartzberg PL, Hood JD, Leng J, Cheresh DA. (1999).Selective requirement for Src kinases during VEGF-induced angiogenesis andvascular permeability. Mol Cell. 1999. 4:915-24.
Epstein, M. A., Achong, B. G. & Barr, Y.M. (1964). Virus particles in culturedlymphoblasts from Burkitt's lymphoma. Lancet, 1, pp. 702-703.
Eliopoulos, A. G. & Young, L.S. (2001). LMP1 structure and signal transduction.Seminars in cancer biology, 11, pp. 435-444.

Ewertz,M., Duffy,S.W., Adami,H.O., Kvale,G., Lund,E., Meirik,O.,Mellemgaard,A., Soini,I., and Tulinius,H. (1990). Age at first birth, parity and riskof breast cancer: a meta-analysis of 8 studies from the Nordic countries. Int. J. Cancer.46, 597-603
Easton,D.F., Bishop,D.T., Ford,D., and Crockford,G.P. (1993). Genetic linkageanalysis in familial breast and ovarian cancer: results from 214 families. The BreastCancer Linkage Consortium. Am. J. Hum. Genet. 52, 678-701
FFahmi H., Cochet C., Hmama Z., Opolon P. and Joab I. (2000). TransformingGrowth Factor Beta 1 stimulates expression of Epstein-Barr virus BZLF1 immediate-early gene product ZEBRA by an indirect mechanism which requires the MAPKkinase pathway. J Virol 74(13): 5810-18.
Fina F., Romain S., Ouafik L., Palmari J., Ben Ayed F., Benharkat S., BonnierP.,Spyratos F., Foekens J. A., Rose C., Buisson M., Gerard H., Reymond M. O.,Seigneurin J. M., and Martin P. M. (2001). Frequency and genome load of Epstein-Barr virus in 509 breast cancers from different geographical areas. Br J Cancer 84:783-90.
Flavell K. J., and Murray P. G. (2000). Hodgkin's disease and the Epstein-Barrvirus. Mol Pathol 53: 262-9.
Feederle, R., Kost, M., Baumann, M., Janz, A., Drouet, E., Hammerschmidt, W.& Delecluse, H.J. (2000). The Epstein-Barr virus lytic program is controlled by theco-operative functions of two transactivators. The EMBO journal, 19, pp. 3080-3089.
Fiorini, S. & Ooka, T. (2008). Secretion of Epstein-Barr virus-encoded BARF1oncoprotein from latently infected B cells. Virology journal, 5, p. 70.
Fixman, E. D., Hayward, G. S. & Hayward, S.D. (1992). trans-acting requirementsfor replication of Epstein-Barr virus ori-Lyt. Journal of virology, 66, pp. 5030-5039.
Flemington, E. & Speck, S.H. (1990). Autoregulation of Epstein-Barr virus putativelytic switch gene BZLF1. Journal of virology, 64, pp. 1227-1232.
Forsman, A., Rüetschi, U., Ekholm, J. & Rymo, L. (2008). Identification ofintracellular proteins associated with the EBV-encoded nuclear antigen 5 using anefficient TAP procedure and FT-ICR mass spectrometry. Journal of proteomeresearch, 7, pp. 2309-2319.
Fries, K. L., Sculley, T. B., Webster-Cyriaque, J., Rajadurai, P., Sadler, R. H. &Raab-Traub, N. (1997). Identification of a novel protein encoded by the BamHI Aregion of the Epstein-Barr virus. Journal of virology, 71, pp. 2765-2771.

Fu, Z. & Cannon, M.J. (2000). Functional analysis of the CD4(+) T-cell response toEpstein-Barr virus: T-cell-mediated activation of resting B cells and induction of viralBZLF1 expression. Journal of virology, 74, pp. 6675-6679.
Fukuda, M. & Longnecker, R. (2005). Epstein-Barr virus (EBV) latent membraneprotein 2A regulates B-cell receptor-induced apoptosis and EBV reactivation throughtyrosine phosphorylation. Journal of virology, 79, pp. 8655-8660.
FERRIS C. F. , KULKARNI P. , SULLIVAN J. M., Jr. , HARDER J. A. ,MESSENGER T. L. , FEBO M. Pup suckling is more rewarding than cocaine:evidence from functional magnetic resonance imaging and three-dimensionalcomputational analysis, The Journal of Neuroscience, 25(1):149-156, 2005
Fornage BD, Lorigan JG, Andry E. Fibroadenoma of the breast: sonographicappearance. Radiology 1989;172(3):671-5.
Fukuda, M. and R. Longnecker (2007). "Epstein-Barr virus latent membraneprotein 2A mediates transformation through constitutive activation of the Ras/PI3-K/Akt Pathway." J Virol 81(17): 9299-306.
Faulkner, G. C., S. R. Burrows, et al. (1999). "X-Linked agammaglobulinemiapatients are not infected with Epstein-Barr virus: implications for the biology of thevirus." J Virol 73(2): 1555-64.
GGaffey M. J., Frierson H. F., Jr., Mills S. E., Boyd J. C., Zarbo R. J., Simpson J.F., Gross L. K., and Weiss L. M. (1993). Medullary carcinoma of the breast.Identification of lymphocyte subpopulations and their significance. Mod Pathol 6:721-8.
Gan Y. J., Razzouk B. I., Su T., and Sixbey J. W. (2002). A defective, rearrangedEpstein-Barr virus genome in EBER-negative and EBER-positive Hodgkin's disease.Am J Pathol 160: 781-6.
Gahn, T. A. & Sugden, B. (1995). An EBNA-1-dependent enhancer acts from a distanceof 10 kilobase pairs to increase expression of the Epstein-Barr virus LMP gene. Journalof virology, 69, pp. 2633-2636.
Gregory C. D., Murray R. J., Edwards C. F., and Rickinson A. B. (1988).Downregulation of cell adhesion molecules LFA-3 and ICAM-1 in Epstein-Barrvirus-positive Burkitt's lymphoma underlies tumor cell escape from virus-specific Tcell surveillance. J Exp Med 167: 1811-24.
Grinstein S., Preciado M. V., Gattuso P., Chabay P. A., Warren W. H., DeMatteo E., and Gould V. E. (2002). Demonstration of Epstein-Barr virus incarcinomas of various sites. Cancer Res 62: 4876-8.

Groux H., Cottrez F., Montpellier C., Quatannens B., Coll J., Stehelin D., andAuriault C. (1997). Isolation and characterization of transformed human T-cell linesinfected by Epstein-Barr virus. Blood 89: 4521-30.
Gutensohn N., and Cole P. (1981). Childhood social environment and Hodgkin'sdisease. N Engl J Med 304: 135-40.
Gratama JW, Oosterveer MA, Zwaan FE, Lepoutre J, Klein G, Ernberg I.(1988). Eradication of Epstein-Barr virus by allogeneic bone marrow transplantation:implications for sites of viral latency. Proc Natl Acad Sci U S A. 85: 8693-6.
Grunewald V, Bonnet M, Boutin S, Yip T, Louzir H, Levrero M, SeigneurinJM, Raphael M, Touitou R, Martel-Renoir D, Cochet C, Durandy A, Andre P,Lau W, Zeng Y, Joab I. (1998). Amino-acid change in the Epstein-Barr-virusZEBRA protein in undifferentiated nasopharyngeal carcinomas from Europe andNorth Africa. Int J Cancer. 75:497-503.
Gilligan, K., Sato, H., Rajadurai, P., Busson, P., Young, L., Rickinson, A., Tursz,T. & Raab-Traub, N. (1990). Novel transcription from the Epstein-Barr virusterminal EcoRI fragment, DIJhet, in a nasopharyngeal carcinoma. Journal of virology,64, pp. 4948-4956.
Gires, O., Zimber-Strobl, U., Gonnella, R., Ueffing, M., Marschall, G., Zeidler,R., Pich, D. & Hammerschmidt, W. (1997). Latent membrane protein 1 of Epstein-Barr virus mimics a constitutively active receptor molecule. The EMBO journal, 16,pp. 6131-6140.
Greifenegger, N., Jäger, M., Kunz-Schughart, L. A., Wolf, H. & Schwarzmann,F. (1998). Epstein-Barr virus small RNA (EBER) genes: differential regulation duringlytic viral replication. Journal of virology, 72, pp. 9323-9328.
Grogan, E., Jenson, H., Countryman, J., Heston, L., Gradoville, L. & Miller, G.(1987). Transfection of a rearranged viral DNA fragment, WZhet, stably convertslatent Epstein-Barr viral infection to productive infection in lymphoid cells.Proceedings of the National Academy of Sciences of the United States of America, 84,pp. 1332-1336.
Gruffat, H., Batisse, J., Pich, D., Neuhierl, B., Manet, E., Hammerschmidt, W. &Sergeant, A. (2002). Epstein-Barr virus mRNA export factor EB2 is essential forproduction of infectious virus. Journal of virology, 76, pp. 9635-9644.
Goodwin,P.J. and Boyd,N.F. (1987). Critical appraisal of the evidence that dietaryfat intake is related to breast cancer risk in humans. J. Natl. Cancer Inst. 79, 473-485.
Gail,M.H., Brinton,L.A., Byar,D.P., Corle,D.K., Green,S.B., Schairer,C., andMulvihill,J.J. (1989). Projecting individualized probabilities of developing breastcancer for white females who are being examined annually. J. Natl. Cancer Inst. 81,1879-1886.

Gerritsen, E. J., E. D. Stam, et al. (1996). "Risk factors for developing EBV-relatedB cell lymphoproliferative disorders (BLPD) after non-HLA-identical BMT inchildren." Bone Marrow Transplant 18(2): 377-82.
Gottschalk, S., H. E. Heslop, et al. (2002). "Treatment of Epstein-Barr virus-associated malignancies with specific T cells." Adv Cancer Res 84: 175-201.
Greenspan, J. S. and D. Greenspan (1989). "Oral hairy leukoplakia: diagnosis andmanagement." Oral Surg Oral Med Oral Pathol 67(4): 396-403.
Gutierrez, M. I., K. Bhatia, et al. (1992). "Molecular epidemiology of Burkitt'slymphoma from South America: differences in breakpoint location and Epstein-Barrvirus association from tumors in other world regions." Blood 79(12): 3261-6.
Gutierrez, M. I., K. Bhatia, et al. (1997). "Intraclonal molecular heterogeneitysuggests a hierarchy of pathogenetic events in Burkitt's lymphoma." Ann Oncol 8(10):987-94.
Gulley, M. L., M. B. Amin, et al. (1995). "Epstein-Barr virus is detected inundifferentiated nasopharyngeal carcinoma but not in lymphoepithelioma-likecarcinoma of the urinary bladder." Hum Pathol 26(11): 1207-14.
HHamilton-Dutoit S. J., Rea D., Raphael M., Sandvej K., Delecluse H. J.,Gisselbrecht C., Marelle L., van Krieken H. J., and Pallesen G. (1993). Epstein-Barr virus-latent gene expression and tumor cell phenotype in acquiredimmunodeficiency syndrome-related non-Hodgkin's lymphoma. Correlation oflymphoma phenotype with three distinct patterns of viral latency. Am J Pathol 143:1072-85.
Hennig E. M., Di Lonardo A., Venuti A., Holm R., Marcante M. L., and NeslandJ. M. (1999). HPV 16 in multiple neoplastic lesions in women with CIN III. J ExpClin Cancer Res 18: 369-77.
Holmes R. D., and Sokol R. J. (2002). Epstein-Barr virus and post-transplantlymphoproliferative disease. Pediatr Transplant 6: 456-64.
Huang H., Pan X. H., Zhou J. H., Yu L., Kong X. T., Zhou S. M., Li Z. J., Fu Q.,and Sun X. Y. (1998). The effect of Epstein-Barr virus gene BHRF1 expression onradioresistance of nasopharyngeal carcinoma cells. ORL J Otorhinolaryngol RelatSpec 60: 329-33.
Haan KM, Kwok WW, Longnecker R, Speck P. (2000). Epstein-Barr virus entryutilizing HLA-DP or HLA-DQ as a coreceptor. J Virol. 74:2451-4.

Hochberg D, Middeldorp JM, Catalina M, Sullivan JL, Luzuriaga K, Thorley-Lawson DA. (2004). Demonstration of the Burkitt's lymphoma Epstein-Barr virusphenotype in dividing latently infected memory cells in vivo. Proc Natl Acad Sci US A. 101:239-44.
Holmgren L, Szeles A, Rajnavolgyi E, Folkman J, Klein G, Ernberg I, Falk KI.(1999). Horizontal transfer of DNA by the uptake of apoptotic bodies. Blood. 93:3956-63.
Han, I., Harada, S., Weaver, D., Xue, Y., Lane, W., Orstavik, S., Skalhegg, B. &Kieff, E. (2001). EBNA-LP associates with cellular proteins including DNA-PK andHA95. Journal of virology, 75, pp. 2475-2481.
Harada, S. & Kieff, E. (1997). Epstein-Barr virus nuclear protein LP stimulatesEBNA-2 acidic domain-mediated transcriptional activation. Journal of virology, 71,pp. 6611-6618.
Henderson, S., Huen, D., Rowe, M., Dawson, C., Johnson, G. & Rickinson, A.(1993). Epstein-Barr virus-coded BHRF1 protein, a viral homologue of Bcl-2,protects human B cells from programmed cell death. Proceedings of the NationalAcademy of Sciences of the United States of America, 90, pp. 8479-8483.
Henle, G., Henle, W. & Diehl, V. (1968). Relation of Burkitt's tumor-associatedherpes-ytpe virus to infectious mononucleosis. Proceedings of the National Academyof Sciences of the United States of America, 59, pp. 94-101.
Henle, W. & Henle, G. (1970). Evidence for a relation of Epstein-Barr virus toBurkitt's lymphoma and nasopharyngeal carcinoma. Bibliotheca haematologica, , pp.706-713.
Hitt, M. M., Allday, M. J., Hara, T., Karran, L., Jones, M. D., Busson, P., Tursz,T., Ernberg, I. & Griffin, B.E. (1989). EBV gene expression in an NPC-relatedtumour. The EMBO journal, 8, pp. 2639-2651.
Holley-Guthrie, E. A., Quinlivan, E. B., Mar, E. C. & Kenney, S. (1990). TheEpstein-Barr virus (EBV) BMRF1 promoter for early antigen (EA-D) is regulated bythe EBV transactivators, BRLF1 and BZLF1, in a cell-specific manner. Journal ofvirology, 64, pp. 3753-3759.
Howe, J. G. & Steitz, J.A. (1986). Localization of Epstein-Barr virus-encoded smallRNAs by in situ hybridization. Proceedings of the National Academy of Sciences ofthe United States of America, 83, pp. 9006-9010.
Hummel, M. & Kieff, E. (1982). Mapping of polypeptides encoded by the Epstein-Barr virus genome in productive infection. Proceedings of the National Academy ofSciences of the United States of America, 79, pp. 5698-5702.
Huneeus A, Schilling A, Horvath E, Pinochet M, Carrasco O. Retroareolar cysts inthe adolescent. J Pediatr Adolesc Gynecol 2003;16(1):45-9.

Henderson,B.E., Powell,D., Rosario,I., Keys,C., Hanisch,R., Young,M.,Casagrande,J., Gerkins,V., and Pike,M.C. (1974). An epidemiologic study ofbreast cancer. J. Natl. Cancer Inst. 53, 609-614
Humme, S., Reisbach, G., Feederle, R., Delecluse, H., Bousset, K., Hammerschmidt,W. & Schepers, A. (2003). The EBV nuclear antigen 1 (EBNA1) enhances B cellimmortalization several thousandfold. Proceedings of the National Academy of Sciencesof the United States of America, 100, pp. 10989-10994.
Heslop, H. E. and C. M. Rooney (1997). "Adoptive cellular immunotherapy forEBV lymphoproliferative disease." Immunol Rev 157: 217-22.
Hecht, J. L. and J. C. Aster (2000). "Molecular biology of Burkitt's lymphoma." JClin Oncol 18(21): 3707-21.
Hecht, J. L. and J. C. Aster (2000). "Molecular biology of Burkitt's lymphoma." JClin Oncol 18(21): 3707-21.
Hsu, J. L. and S. L. Glaser (2000). "Epstein-barr virus-associated malignancies:epidemiologic patterns and etiologic implications." Crit Rev Oncol Hematol 34(1):27-53.
Heussinger, N., M. Buttner, et al. (2004). "Expression of the Epstein-Barr virus(EBV)-encoded latent membrane protein 2A (LMP2A) in EBV-associatednasopharyngeal carcinoma." J Pathol 203(2): 696-9.
IImai S, Nishikawa J, Takada K. (1998). Cell-to-cell contact as an efficient mode ofEpstein-Barr virus infection of diverse human epithelial cells. J Virol. 72: 4371-8.
JJohannessen I., Asghar M., and Crawford D. H. (2000). Essential role for T cellsin human B-cell lymphoproliferative disease development in severe combinedimmunodeficient mice. Br J Haematol 109: 600-10.
Junker A. K., Thomas E. E., Radcliffe A., Forsyth R. B., Davidson A. G., andRymo L. (1991). Epstein-Barr virus shedding in breast milk. Am J Med Sci 302: 220-3.
Jin, X. W. & Speck, S.H. (1992). Identification of critical cis elements involved inmediating Epstein-Barr virus nuclear antigen 2-dependent activity of an enhancerlocated upstream of the viral BamHI C promoter. Journal of virology, 66, pp. 2846-2852.

J. Amice - Faculté de Médecine de Brest LA GLANDE MAMMAIREGENERALITES Sa présence.
Jones, J. F., S. Shurin, et al. (1988). "T-cell lymphomas containing Epstein-Barrviral DNA in patients with chronic Epstein-Barr virus infections." N Engl J Med318(12): 733-41.
Jaffe, E. S., J. K. Chan, et al. (1996). "Report of the Workshop on Nasal and RelatedExtranodal Angiocentric T/Natural Killer Cell Lymphomas. Definitions, differentialdiagnosis, and epidemiology." Am J Surg Pathol 20(1): 103-11.
Jian-Rang he, Er Wei Song, Ze fang Ren(2009) .Reasearch advencement onrelation ship between Epstein-Barr virus and breast cancer.
KKieff E. (1996). Epstein-Barr virus. In: Fileds, B.N., K.N.I.P.E., D.M. & Howley Et,A.L. Editors. Virology. Philadelphia: Lippincott-Raven Press. 2343 - 2396.
Koide J., Takada K., Sugiura M., Sekine H., Ito T., Saito K., Mori S., TakeuchiT., Uchida S., and Abe T. (1997). Spontaneous establishment of an Epstein-Barrvirus-infected fibroblast line from the synovial tissue of a rheumatoid arthritis
Kalluri R (2003). Basement membranes: structure, assembly and role in tumourangiogenesis. Nat Rev Cancer 3: 422-33.
Kerbel R, Folkman J. (2002). Clinical translation of angiogenesis inhibitors. NatRev Cancer. 2: 727-39.
Krauer KG, Buck M., Sculley, T. (1999). Characterization of the transcriptionalrepressor RBP in Epstein-Barr virus-transformed B cells. J Gen Virol. 80 :3217-3226.
Kapatai, G. & Murray, P. (2007). Contribution of the Epstein Barr virus to themolecular pathogenesis of Hodgkin lymphoma. Journal of clinical pathology, 60, pp.1342-1349.
Kashuba, E., Mattsson, K., Pokrovskaja, K., Kiss, C., Protopopova, M., Ehlin-Henriksson, B., Klein, G. & Szekely, L. (2003). EBV-encoded EBNA-5 associateswith P14ARF in extranucleolar inclusions and prolongs the survival of P14ARF-expressing cells. International journal of cancer. Journal international du cancer,105, pp. 644-653.
Kawaguchi, Y., Nakajima, K., Igarashi, M., Morita, T., Tanaka, M., Suzuki, M.,Yokoyama, A., Matsuda, G., Kato, K., Kanamori, M. & Hirai, K. (2000).Interaction of Epstein-Barr virus nuclear antigen leader protein (EBNA-LP) withHS1-associated protein X-1: implication of cytoplasmic function of EBNA-LP.Journal of virology, 74, pp. 10104-10111.

Kienzle, N., Buck, M., Greco, S., Krauer, K. & Sculley, T.B. (1999). Epstein-Barrvirus-encoded RK-BARF0 protein expression. Journal of virology, 73, pp. 8902-8906.
Kimura, H., Hoshino, Y., Kanegane, H., Tsuge, I., Okamura, T., Kawa, K. &Morishima, T. (2001). Clinical and virologic characteristics of chronic activeEpstein-Barr virus infection. Blood, 98, pp. 280-286.
Kis, L. L., Takahara, M., Nagy, N., Klein, G. & Klein, E. (2006). IL-10 can inducethe expression of EBV-encoded latent membrane protein-1 (LMP-1) in the absence ofEBNA-2 in B lymphocytes and in Burkitt lymphoma- and NK lymphoma-derived celllines. Blood, 107, pp. 2928-2935.
Komano, J., Maruo, S., Kurozumi, K., Oda, T. & Takada, K. (1999). Oncogenicrole of Epstein-Barr virus-encoded RNAs in Burkitt's lymphoma cell line Akata.Journal of virology, 73, pp. 9827-9831.
Kusano, S. & Raab-Traub, N. (2001). An Epstein-Barr virus protein interacts withNotch. Journal of virology, 75, pp. 384-395.
Kaul, R., Verma, S. C., Murakami, M., Lan, K., Choudhuri, T. & Robertson, E.S.(2006). Epstein-Barr virus protein can upregulate cyclo-oxygenase-2 expression throughassociation with the suppressor of metastasis Nm23-H1. Journal of virology, 80, pp.1321-1331.
Kuppers, D. A., Lan, K., Knight, J. S. & Robertson, E.S. (2005). Regulation of matrixmetalloproteinase 9 expression by Epstein-Barr virus nuclear antigen 3C and thesuppressor of metastasis Nm23-H1. Journal of virology, 79, pp. 9714-9724.
Krauer, K. G., Burgess, A., Buck, M., Flanagan, J., Sculley, T. B. & Gabrielli, B.(2004). The EBNA-3 gene family proteins disrupt the G2/M checkpoint. Oncogene, 23,pp. 1342-1353.
Knight, J. S., Sharma, N. & Robertson, E.S. (2005). SCFSkp2 complex targeted byEpstein-Barr virus essential nuclear antigen. Molecular and cellular biology, 25, pp.1749-1763.
Knight, J. S. & Robertson, E.S. (2004). Epstein-Barr virus nuclear antigen 3C regulatescyclin A/p27 complexes and enhances cyclin A-dependent kinase activity. Journal ofvirology, 78, pp. 1981-1991.
Knowles, D. M., E. Cesarman, et al. (1995). "Correlative morphologic andmolecular genetic analysis demonstrates three distinct categories ofposttransplantation lymphoproliferative disorders." Blood 85(2): 552-65.
Knox, P. G., Q. X. Li, et al. (1996). "In vitro production of stable Epstein-Barr virus-positive epithelial cell clones which resemble the virus:cell interaction observed innasopharyngeal carcinoma." Virology 215(1): 40-50.

Kanegane, H., K. Bhatia, et al. (1998). "A syndrome of peripheral blood T-cellinfection with Epstein-Barr virus (EBV) followed by EBV-positive T-celllymphoma." Blood 91(6): 2085-91.
Kanegane, H., T. Miyawaki, et al. (1999). "Development of EBV-positive T-celllymphoma following infection of peripheral blood T cells with EBV." LeukLymphoma 34(5-6): 603-7.
LLabrecque L. G., Barnes D. M., Fentiman I. S., and Griffin B. E. (1995). Epstein-Barr virus in epithelial cell tumors: a breast cancer study. Cancer Res 55: 39-45.
Levin L.I. and Levine P.H. (1998). The epidemiology of Epstein-Barr virusassociated human cancers. Gann Monograph on cancer research. 45: 51-74.
Lopategui J. R., Gaffey M. J., Frierson H. F., Jr., Chan J. K., Mills S. E., ChangK. L., Chen Y. Y., and Weiss L. M. (1994). Detection of Epstein-Barr viral RNA insinonasal undifferentiated carcinoma from Western and Asian patients. Am J SurgPathol 18: 391-8.
Luka J., Miller G., Jornvall H., and Pearson G. R. (1986). Characterization of therestricted component of Epstein-Barr virus early antigens as a cytoplasmicfilamentous protein. J Virol 58: 748-56.
Luqmani Y. A., Mortimer C., Yiangou C., Johnston C. L., Bansal G. S., SinnettD., Law M., and Coombes R. C. (1995). Expression of 2 variant forms of fibroblastgrowth factor receptor 1 in human breast. Int J Cancer 64: 274-9.
Li Q, Spriggs M.K, Kovats S, Turk S.M, Comeau M.R, Nepom B, Hutt-FletcherL.M. (1997). Epstein-Barr virus uses HLA class II as a cofactor for infection of Blymphocytes. J Virol. 71:4657-62.
Li W, Wu BA, Zeng YM, Chen GC, Li XX, Chen JT, Guo YW, Li MH, Zeng Y.(2004). Epstein-Barr virus in hepatocellular carcinogenesis. World JGastroenterol.10:3409-13.
Lieberman PM, Hardwick JM, Sample J, Hayward GS, Hayward SD. (1990).The zta transactivator involved in induction of lytic cycle gene expression in Epstein-Barr virus-infected lymphocytes binds to both AP-1 and ZRE sites in target promoterand enhancer regions. J Virol. 64:1143-55.
Lin J, Johannsen E, Robertson E, Kieff E. (2002). Epstein-Barr virus nuclearantigen 3C putative repression domain mediates coactivation of the LMP1promoter with EBNA-2. J Virol. 76:232-42.
Laux, G., Perricaudet, M. & Farrell, P.J. (1988). A spliced Epstein-Barr virus geneexpressed in immortalized lymphocytes is created by circularization of the linear viralgenome. The EMBO journal, 7, pp. 769-774.

Levitskaya, J., Coram, M., Levitsky, V., Imreh, S., Steigerwald-Mullen, P. M.,Klein, G., Kurilla, M. G. & Masucci, M.G. (1995). Inhibition of antigen processingby the internal repeat region of the Epstein-Barr virus nuclear antigen-1. Nature, 375,pp. 685-688.
Ling, P. D., Peng, R. S., Nakajima, A., Yu, J. H., Tan, J., Moses, S. M., Yang, W.,Zhao, B., Kieff, E., Bloch, K. D. & Bloch, D.B. (2005). Mediation of Epstein-Barrvirus EBNA-LP transcriptional coactivation by Sp100. The EMBO journal, 24, pp.3565-3575.
Longnecker, R., Miller, C. L., Miao, X. Q., Marchini, A. & Kieff, E. (1992). Theonly domain which distinguishes Epstein-Barr virus latent membrane protein 2A(LMP2A) from LMP2B is dispensable for lymphocyte infection and growthtransformation in vitro; LMP2A is therefore nonessential. Journal of virology, 66, pp.6461-6469.
Longnecker, R., Miller, C. L., Miao, X. Q., Tomkinson, B. & Kieff, E. (1993). Thelast seven transmembrane and carboxy-terminal cytoplasmic domains of Epstein-Barrvirus latent membrane protein 2 (LMP2) are dispensable for lymphocyte infection andgrowth transformation in vitro. Journal of virology, 67, pp. 2006-2013.
Longnecker, R., Miller, C. L., Tomkinson, B., Miao, X. Q. & Kieff, E. (1993).Deletion of DNA encoding the first five transmembrane domains of Epstein-Barrvirus latent membrane proteins 2A and 2B. Journal of virology, 67, pp. 5068-5074.
Lupton, S. & Levine, A.J. (1985). Mapping genetic elements of Epstein-Barr virus thatfacilitate extrachromosomal persistence of Epstein-Barr virus-derived plasmids in humancells. Molecular and cellular biology, 5, pp. 2533-2542.
Lee, E. S., J. Locker, et al. (1995). "The association of Epstein-Barr virus withsmooth-muscle tumors occurring after organ transplantation." N Engl J Med 332(1):19-25.
Low, W. K., J. L. Leong, et al. (2000). "Diagnostic value of Epstein-Barr viralserology in nasopharyngeal carcinoma." Otolaryngol Head Neck Surg 123(4): 505-7.
Lay, J. D., C. J. Tsao, et al. (1997). "Upregulation of tumor necrosis factor-alphagene by Epstein-Barr virus and activation of macrophages in Epstein-Barr virus-infected T cells in the pathogenesis of hemophagocytic syndrome." J Clin Invest100(8): 1969-79.

MM Lucas, K. G., T. N. Small, et al. (1996). "The development of cellular immunity
to Epstein-Barr virus after allogeneic bone marrow transplantation." Blood87(6): 2594-603.
Martel-Renoir D., Grunewald V., Touitou R., Schwaab G., and Joab I. (1995).Qualitative analysis of the expression of Epstein-Barr virus lytic genes innasopharyngeal carcinoma biopsies. J Gen Virol 76 ( Pt 6): 1401-8.
McCall SA, Lichy JH, Bijwaard KE, Aguilera NS, Chu WS, Taubenberger JK.(2001) Epstein-Barr virus detection in ductal carcinoma of the breast. J Natl CancerInst 93: 148-50
Moriuchi M., and Moriuchi H. (2003). Increased susceptibility to HIV-1 ofperipheral blood lymphocytes in acute infection with Epstein-Barr virus. J Med Virol71: 343-6.
Mueller N., Evans A., Harris N. L., Comstock G. W., Jellum E., Magnus K.,Orentreich N., Polk B. F., and Vogelman J. (1989). Hodgkin's disease and Epstein-Barr virus. Altered antibody pattern before diagnosis. N Engl J Med 320: 689-95.
Murray P. G., Lissauer D., Junying J., Davies G., Moore S., Bell A., Timms J.,Rowlands D., McConkey C., Reynolds G. M., Ghataura S., England D., CarollR., and Young L. S. (2003). Reactivity with A monoclonal antibody to Epstein-Barr virus (EBV) nuclear antigen 1 defines a subset of aggressive breast cancers inthe absence of the EBV genome. Cancer Res 63: 2338-43.
Mannick, J. B., Cohen, J. I., Birkenbach, M., Marchini, A. & Kieff, E. (1991).The Epstein-Barr virus nuclear protein encoded by the leader of the EBNA RNAs isimportant in B-lymphocyte transformation. Journal of virology, 65, pp. 6826-6837.
Matsuda, G., Nakajima, K., Kawaguchi, Y., Yamanashi, Y. & Hirai, K. (2003).Epstein-Barr virus (EBV) nuclear antigen leader protein (EBNA-LP) formscomplexes with a cellular anti-apoptosis protein Bcl-2 or its EBV counterpart BHRF1through HS1-associated protein X-1. Microbiology and immunology, 47, pp. 91-99.
Miller, C. L., Lee, J. H., Kieff, E. & Longnecker, R. (1994). An integral membraneprotein (LMP2) blocks reactivation of Epstein-Barr virus from latency followingsurface immunoglobulin crosslinking. Proceedings of the National Academy ofSciences of the United States of America, 91, pp. 772-776.
Miyashita, T. & Reed, J.C. (1995). Tumor suppressor p53 is a direct transcriptionalactivator of the human bax gene. Cell, 80, pp. 293-299.

McDivitt RW, Stevens JA, Lee NC, Wingo PA, Rubin GL, Gersell D and the Cancerand Steroid Hormone Study Group, Histologic types of benign breast disease and therisk for breast cancer, Cancer, 1992, 69 : 1408-1414
Morrow M, Wong S, Venta L. The evaluation of breast masses in women youngerthan forty years of age. Surgery 1998;124(4):634-40; discussion 640-1
MacMahon,B., Trichopoulos,D., Brown,J., Andersen,A.P., Cole,P., deWaard,F.,Kauraniemi,T., Polychronopoulou,A., Ravnihar,B., Stormby,N., andWestlund,K. (1982). Age at menarche, urine estrogens and breast cancer risk. Int. J.Cancer. 30, 427-431
Malkin,D., Li,F.P., Strong,L.C., Fraumeni,J.F., Jr., Nelson,C.E., Kim,D.H.,Kassel,J., Gryka,M.A., Bischoff,F.Z., and Tainsky,M.A. (1990). Germ line p53mutations in a familial syndrome of breast cancer, sarcomas, and other neoplasms.Science 250, 1233-1238
Marshall, D. & Sample, C. (1995). Epstein-Barr virus nuclear antigen 3C is atranscriptional regulator. Journal of virology, 69, pp. 3624-3630.
Morrison, J. A. and N. Raab-Traub (2005). "Roles of the ITAM and PY motifs ofEpstein-Barr virus latent membrane protein 2A in the inhibition of epithelial celldifferentiation and activation of {beta}-catenin signaling." J Virol 79(4): 2375-82.
Marechal, V. and T. Piolot (2000). "Lytic infection by double-strand DNA virusesand cell cycle alterations." Pathol Biol (Paris) 48(3): 289-300.
McClain, K. L., C. T. Leach, et al. (1995). "Association of Epstein-Barr virus withleiomyosarcomas in children with AIDS." N Engl J Med 332(1): 12-8.
Morrison, J. A. and N. Raab-Traub (2005). "Roles of the ITAM and PY motifs ofEpstein-Barr virus latent membrane protein 2A in the inhibition of epithelial celldifferentiation and activation of {beta}-catenin signaling." J Virol 79(4): 2375-82.
NNaresh K. N., Johnson J., Srinivas V., Soman C. S., Saikia T., Advani S. H.,Badwe R. A., Dinshaw K. A., Muckaden M., Magrath I., and Bhatia K. (2000).Epstein-Barr virus association in classical Hodgkin's disease provides survivaladvantage to patients and correlates with higher expression of proliferation markers inReed-Sternberg cells. Ann Oncol 11: 91-6.
Neri A., Barriga F., Inghirami G., Knowles D. M., Neequaye J., Magrath I. T.,and Dalla-Favera R. (1991). Epstein-Barr virus infection precedes clonal expansionin Burkitt's and acquired immunodeficiency syndrome-associated lymphoma. Blood77: 1092-5.
Niedobitek G., Lisner R., Swoboda B., Rooney N., Fassbender H. G., KirchnerT., Aigner T., and Herbst H. (2000). Lack of evidence for an involvement of

Epstein-Barr virus infection of synovial membranes in the pathogenesis of rheumatoidarthritis. Arthritis Rheum 43: 151-4.
Nishikawa J., Imai S., Oda T., Kojima T., Okita K., and Takada K. (1999).Epstein-Barr virus promotes epithelial cell growth in the absence of EBNA2 andLMP1 expression. J Virol 73: 1286-92.
Nitsche, F., Bell, A. & Rickinson, A. (1997). Epstein-Barr virus leader proteinenhances EBNA-2-mediated transactivation of latent membrane protein 1 expression:a role for the W1W2 repeat domain. Journal of virology, 71, pp. 6619-6628.
Nanbo, A. and K. Takada (2002). "The role of Epstein-Barr virus-encoded smallRNAs (EBERs) in oncogenesis." Rev Med Virol 12(5): 321-6.
Nicholls, J. M., A. Agathanggelou, et al. (1997). "The association of squamous cellcarcinomas of the nasopharynx with Epstein-Barr virus shows geographical variationreminiscent of Burkitt's lymphoma." J Pathol 183(2): 164-8.
Niedobitek, G., L. S. Young, et al. (1992). "Expression of Epstein-Barr virus genesand of lymphocyte activation molecules in undifferentiated nasopharyngealcarcinomas." Am J Pathol 140(4): 879-87.
OOsato T, Imai S. (1996). Epstein-Barr virus and gastric carcinoma. Semin CancerBiol. 7:175-82.
Ottman,R., Pike,M.C., King,M.C., and Henderson,B.E. (1983). Practical guidefor estimating risk for familial breast cancer. Lancet 2, 556-558.
Orazi, A., R. A. Hromas, et al. (1997). "Posttransplantation lymphoproliferativedisorders in bone marrow transplant recipients are aggressive diseases with a highincidence of adverse histologic and immunobiologic features." Am J Clin Pathol107(4): 419-29.
O'Reilly, R. J., T. N. Small, et al. (1997). "Biology and adoptive cell therapy ofEpstein-Barr virus-associated lymphoproliferative disorders in recipients of marrowallografts." Immunol Rev 157: 195-216.
O'Reilly, R. J., T. N. Small, et al. (1998). "Adoptive immunotherapy for Epstein-Barr virus-associated lymphoproliferative disorders complicating marrow allografts."Springer Semin Immunopathol 20(3-4): 455-91.
PPallesen G., Hamilton-Dutoit S. J., Rowe M., Lisse I., Ralfkiaer E., Sandvej K.,and Young L. S. (1991a). Expression of Epstein-Barr virus replicative proteins inAIDS-related non-Hodgkin's lymphoma cells. J Pathol 165: 289-99.

Pallesen G., Hamilton-Dutoit S. J., Rowe M., and Young L. S. (1991b).Expression of Epstein-Barr virus latent gene products in tumour cells of Hodgkin'sdisease. Lancet 337: 320-2.
Pathmanathan R., Prasad U., Chandrika G., Sadler R., Flynn K., and Raab-Traub N. (1995). Undifferentiated, nonkeratinizing, and squamous cell carcinoma ofthe nasopharynx. Variants of Epstein-Barr virus-infected neoplasia. Am J Pathol 146:1355-67.
Purtilo D. T., Strobach R. S., Okano M., and Davis J. R. (1992). Epstein-Barrvirus-associated lymphoproliferative disorders. Lab Invest 67: 5-23.
Polyak K. Biochim Biophys Acta. (2001). On the birth of breast cancer. 1552:1-13.Review.
Portnoy J, Ahronheim GA, Ghibu F, Clecner B, Joncas JH. (1984). Recovery ofEpstein-Barr virus from genital ulcers. N Engl J Med. 311:966-8.
Pugh CW, Ratcliffe PJ. (2003). Regulation of angiogenesis by hypoxia: role of theHIF system.. Nat Med. 9:677-84. Review.
Peng, R., Tan, J. & Ling, P.D. (2000). Conserved regions in the Epstein-Barr virusleader protein define distinct domains required for nuclear localization andtranscriptional cooperation with EBNA2. Journal of virology, 74, pp. 9953-9963.
Prince, S., Keating, S., Fielding, C., Brennan, P., Floettmann, E. & Rowe, M.(2003). Latent membrane protein 1 inhibits Epstein-Barr virus lytic cycle inductionand progress via different mechanisms. Journal of virology, 77, pp. 5000-5007.
PONS JY. Hormones et sein in << les mastopathies bénignes >> Eds ArnetteBlackwell. 17es Journées Nationales de la Société Française de sénologie et depathologie Mammaire - Bordeaux, 11-13 octobre 1995
PAGE DL, DUPONT WD, ROGERS LW et al.Atypical hyperplastic lesions of thefemale breastCancer, 1985 ; 55 : 2698-2708
Parker, G. A., Touitou, R. & Allday, M.J. (2000). Epstein-Barr virus EBNA3C candisrupt multiple cell cycle checkpoints and induce nuclear division divorced fromcytokinesis. Oncogene, 19, pp. 700-709.
Parker, G. A., Crook, T., Bain, M., Sara, E. A., Farrell, P. J. & Allday, M.J. (1996).Epstein-Barr virus nuclear antigen (EBNA)3C is an immortalizing oncoprotein withsimilar properties to adenovirus E1A and papillomavirus E7. Oncogene, 13, pp. 2541-2549.
Pang, M. F., K. W. Lin, et al. (2009). "The signaling pathways of Epstein-Barrvirus-encoded latent membrane protein 2A (LMP2A) in latency and cancer." Cell MolBiol Lett 14(2): 222-47.

Polack, A., K. Hortnagel, et al. (1996). "c-myc activation renders proliferation ofEpstein-Barr virus (EBV)-transformed cells independent of EBV nuclear antigen 2and latent membrane protein 1." Proc Natl Acad Sci U S A 93(19): 10411-6.
Pelc, S., E. De Maertelaere, et al. (1983). "Burkitt's lymphoma--a correlated lightand electron microscopic study on the malignancy in the CSF." Clin Neuropathol2(2): 69-74.
Pathmanathan, R., U. Prasad, et al. (1995). "Clonal proliferations of cells infectedwith Epstein-Barr virus in preinvasive lesions related to nasopharyngeal carcinoma."N Engl J Med 333(11): 693-8.
RRaab-Traub N., and Flynn K. (1986). The structure of the termini of the Epstein-Barr virus as a marker of clonal cellular proliferation. Cell 47: 883-9.
Richardson A. (1997). Is breast cancer caused by late exposure to a common virus?Med Hypotheses 48: 491-7.
Rickinson AB. and Kieff E. (2001). Epstein-Barr virus. In: Knipe USA, ed. FieldsVirology 4th Edition.Lippincott Williams & Wilkins: 2575-627.
Rouesse J., Martin P.M. and Contesso G. (1997). Le praticien face au cancer dusein. Ed Arnette.
Ragoczy T, Heston L, Miller G. (1998). The Epstein-Barr virus Rta protein activateslytic cycle genes and can disrupt latency in B lymphocytes. J Virol. 72:7978-84.
Radkov, S. A., Bain, M., Farrell, P. J., West, M., Rowe, M. & Allday, M.J.(1997). Epstein-Barr virus EBNA3C represses Cp, the major promoter for EBNAexpression, but has no effect on the promoter of the cell gene CD21. Journal ofvirology, 71, pp. 8552-8562.
Radkov, S. A., Touitou, R., Brehm, A., Rowe, M., West, M., Kouzarides, T. &Allday, M.J. (1999). Epstein-Barr virus nuclear antigen 3C interacts with histonedeacetylase to repress transcription. Journal of virology, 73, pp. 5688-5697.
Ragoczy, T. & Miller, G. (1999). Role of the epstein-barr virus RTA protein inactivation of distinct classes of viral lytic cycle genes. Journal of virology, 73, pp.9858-9866.
Ragoczy, T. & Miller, G. (2001). Autostimulation of the Epstein-Barr virus BRLF1promoter is mediated through consensus Sp1 and Sp3 binding sites. Journal ofvirology, 75, pp. 5240-5251.
Rechsteiner, M. P., Berger, C., Weber, M., Sigrist, J. A., Nadal, D. & Bernasconi,M. (2007). Silencing of latent membrane protein 2B reduces susceptibility to

activation of lytic Epstein-Barr virus in Burkitt's lymphoma Akata cells. The Journalof general virology, 88, pp. 1454-1459.
Rechsteiner, M. P., Bernasconi, M., Berger, C. & Nadal, D. (2008). Role of latentmembrane protein 2 isoforms in Epstein-Barr virus latency. Trends in microbiology, ,
Robertson, E. S., Lin, J. & Kieff, E. (1996). The amino-terminal domains ofEpstein-Barr virus nuclear proteins 3A, 3B, and 3C interact with RBPJ(kappa).Journal of virology, 70, pp. 3068-3074.
Robertson, E. S., Tomkinson, B. & Kieff, E. (1994). An Epstein-Barr virus with a58-kilobase-pair deletion that includes BARF0 transforms B lymphocytes in vitro.Journal of virology, 68, pp. 1449-1458.
Rosa, M. D., Gottlieb, E., Lerner, M. R. & Steitz, J.A. (1981). Striking similaritiesare exhibited by two small Epstein-Barr virus-encoded ribonucleic acids and theadenovirus-associated ribonucleic acids VAI and VAII. Molecular and cellularbiology, 1, pp. 785-796.
Ruf, I. K., Rhyne, P. W., Yang, C., Cleveland, J. L. & Sample, J.T. (2000).Epstein-Barr virus small RNAs potentiate tumorigenicity of Burkitt lymphoma cellsindependently of an effect on apoptosis. Journal of virology, 74, pp. 10223-10228.
REID SE, MURTHY MS, KAUFMAN M, SCANLON, EF. Endocrine andparacrine hormones in the promotion, progression and recurrence of breast cancer.British Journal of Surgery. 1996 ; 83, 1037-1046
Rowe, D. T., Rowe, M., Evan, G. I., Wallace, L. E., Farrell, P. J. & Rickinson, A.B.(1986). Restricted expression of EBV latent genes and T-lymphocyte-detected membraneantigen in Burkitt's lymphoma cells. The EMBO journal, 5, pp. 2599-2607.
Rickinson, A. B. and D. J. Moss (1997). "Human cytotoxic T lymphocyte responsesto Epstein-Barr virus infection." Annu Rev Immunol 15: 405-31.
Rooney, C. M., C. A. Smith, et al. (1995). "Use of gene-modified virus-specific Tlymphocytes to control Epstein- Barr-virus-related lymphoproliferation." Lancet345(8941): 9-13.
Rose, C., M. Green, et al. (2001). "Pediatric solid-organ transplant recipients carrychronic loads of Epstein-Barr virus exclusively in the immunoglobulin D-negative B-cell compartment." J Clin Microbiol 39(4): 1407-15.
Rogatsch, H., H. Bonatti, et al. (2000). "Epstein-Barr virus-associated multicentricleiomyosarcoma in an adult patient after heart transplantation: case report and reviewof the literature." Am J Surg Pathol 24(4): 614-21.
Ruf, I. K. and J. Sample (1999). "Repression of Epstein-Barr virus EBNA-1 genetranscription by pRb during restricted latency." J Virol 73(10): 7943-51.

Rosenbaum, L., M. H. Fekrazad, et al. (2009). "Epstein-Barr virus-associatedinflammatory pseudotumor of the spleen: report of two cases and review of theliterature." J Hematop.
SSemmes O. J., Chen L., Sarisky R. T., Gao Z., Zhong L., and Hayward S. D.(1998). Mta has properties of an RNA export protein and increases cytoplasmicaccumulation of Epstein-Barr virus replication gene mRNA. J Virol 72: 9526-34.
Sheng W., Decaussin G., Ligout A., Takada K., and Ooka T. (2003). Malignanttransformation of Epstein-Barr virus-negative Akata cells by introduction of theBARF1 gene carried by Epstein-Barr virus. J Virol 77: 3859-65.
Shimakage M., Horii K., Tempaku A., Kakudo K., Shirasaka T., and SasagawaT. (2002). Association of Epstein-Barr virus with oral cancers. Hum Pathol 33: 608-14.
Shimizu N., Yoshiyama H., and Takada K. (1996). Clonal propagation of Epstein-Barr virus (EBV) recombinants in EBV-negative Akata cells. J Virol 70: 7260-3.
Shinkura R., Yamamoto N., Koriyama C., Shinmura Y., Eizuru Y., andTokunaga M. (2000). Epstein-Barr virus-specific antibodies in Epstein-Barr virus-positive and -negative gastric carcinoma cases in Japan. J Med Virol 60: 411-6.
Sugawara Y., Mizugaki Y., Uchida T., Torii T., Imai S., Makuuchi M., andTakada K. (1999). Detection of Epstein-Barr virus (EBV) in hepatocellularcarcinoma tissue: a novel EBV latency characterized by the absence of EBV-encodedsmall RNA expression. Virology 256: 196-202.
Silvestre JS, Lévy BI.(2002). Role of matrix proteolysis in angiogenesis. JournAnnu Diabetol Hotel Dieu. 2002:23-32.
Sixbey JW, Yao QY. (1992). Immunoglobulin A-induced shift of Epstein-Barrvirus tissue tropism. Science.255:1578-80.
Sadler, R. H. & Raab-Traub, N. (1995). Structural analyses of the Epstein-Barrvirus BamHI A transcripts. Journal of virology, 69, pp. 1132-1141.
Sample, J., Hummel, M., Braun, D., Birkenbach, M. & Kieff, E. (1986).Nucleotide sequences of mRNAs encoding Epstein-Barr virus nuclear proteins: aprobable transcriptional initiation site. Proceedings of the National Academy ofSciences of the United States of America, 83, pp. 5096-5100.
Sample, J., Liebowitz, D. & Kieff, E. (1989). Two related Epstein-Barr virusmembrane proteins are encoded by separate genes. Journal of virology, 63, pp. 933-937.

Schlager, S., Speck, S. H. & Woisetschläger, M. (1996). Transcription of theEpstein-Barr virus nuclear antigen 1 (EBNA1) gene occurs before induction of theBCR2 (Cp) EBNA gene promoter during the initial stages of infection in B cells.Journal of virology, 70, pp. 3561-3570.
Schwemmle, M., Clemens, M. J., Hilse, K., Pfeifer, K., Tröster, H., Müller, W. E.& Bachmann, M. (1992). Localization of Epstein-Barr virus-encoded RNAs EBER-1and EBER-2 in interphase and mitotic Burkitt lymphoma cells. Proceedings of theNational Academy of Sciences of the United States of America, 89, pp. 10292-10296.
Semmes, O. J., Chen, L., Sarisky, R. T., Gao, Z., Zhong, L. & Hayward, S.D.(1998). Mta has properties of an RNA export protein and increases cytoplasmicaccumulation of Epstein-Barr virus replication gene mRNA. Journal of virology, 72,pp. 9526-9534.
Sharp, T. V., Schwemmle, M., Jeffrey, I., Laing, K., Mellor, H., Proud, C. G.,Hilse, K. & Clemens, M.J. (1993). Comparative analysis of the regulation of theinterferon-inducible protein kinase PKR by Epstein-Barr virus RNAs EBER-1 andEBER-2 and adenovirus VAI RNA. Nucleic acids research, 21, pp. 4483-4490.
Sheng, W., Decaussin, G., Sumner, S. & Ooka, T. (2001). N-terminal domain ofBARF1 gene encoded by Epstein-Barr virus is essential for malignant transformationof rodent fibroblasts and activation of BCL-2. Oncogene, 20, pp. 1176-1185.
Swift,M., Morrell,D., Massey,R.B., and Chase,C.L. (1991). Incidence of cancer in161 families affected by ataxia-telangiectasia. N. Engl. J. Med. 325, 1831-1836.
Sinclair, A. J. & Farrell, P.J. (1995). Host cell requirements for efficient infection ofquiescent primary B lymphocytes by Epstein-Barr virus. Journal of virology, 69, pp.5461-5468.
Smith, P. R., Gao, Y., Karran, L., Jones, M. D., Snudden, D. & Griffin, B.E.(1993). Complex nature of the major viral polyadenylated transcripts in Epstein-Barrvirus-associated tumors. Journal of virology, 67, pp. 3217-3225.
Szekely, L., Pokrovskaja, K., Jiang, W. Q., de The, H., Ringertz, N. & Klein, G.(1996). The Epstein-Barr virus-encoded nuclear antigen EBNA-5 accumulates inPML-containing bodies. Journal of virology, 70, pp. 2562-2568.
Szekely, L., Selivanova, G., Magnusson, K. P., Klein, G. & Wiman, K.G. (1993).EBNA-5, an Epstein-Barr virus-encoded nuclear antigen, binds to the retinoblastomaand p53 proteins. Proceedings of the National Academy of Sciences of the UnitedStates of America, 90, pp. 5455-5459.
Stehr KG, Lebeau A, Stehr M, Grantzow R. Fibroadenoma of the breast in an 11-year-old girl. Eur J Pediatr Surg 2004;14(1):56-9
Stringel G, Perelman A, Jimenez C. Infantile mammary duct ectasia: a cause ofbloody nipple discharge. J Pediatr Surg 1986;21(8):671-4

Stavraky,K. and Emmons,S. (1974). Breast cancer in premenopausal andpostmenopausal women. J. Natl. Cancer Inst. 53, 647-654
Staszewski,J. (1971). Age at menarche and breast cancer. J. Natl. Cancer Inst. 47,935-940.
Sattin,R.W., Rubin,G.L., Webster,L.A., Huezo,C.M., Wingo,P.A., Ory,H.W.,and Layde,P.M. (1985). Family history and the risk of breast cancer. JAMA 253,1908-1913
Schaefer, B. C., Strominger, J. L. & Speck, S.H. (1995). Redefining the Epstein-Barrvirus-encoded nuclear antigen EBNA-1 gene promoter and transcription initiation site ingroup I Burkitt lymphoma cell lines. Proceedings of the National Academy of Sciences ofthe United States of America, 92, pp. 10565-10569.
Subramanian, C., Cotter, M. A. 2. & Robertson, E.S. (2001). Epstein-Barr virusnuclear protein EBNA-3C interacts with the human metastatic suppressor Nm23-H1: amolecular link to cancer metastasis. Nature medicine, 7, pp. 350-355.
Sakorafas,G.H., Krespis,E., and Pavlakis,G. 2002. Risk estimation for breast cancerdevelopment; a clinical perspective. Surg. Oncol. 10:183-192.
Shapira, J. and N. Peylan-Ramu (1998). "Burkitt's lymphoma." Oral Oncol 34(1):15-23.
Savoldo, B., J. Goss, et al. (2001). "Generation of autologous Epstein-Barr virus-specific cytotoxic T cells for adoptive immunotherapy in solid organ transplantrecipients." Transplantation 72(6): 1078-86.
Swinnen, L. J. (1999). "Overview of posttransplant B-cell lymphoproliferativedisorders." Semin Oncol 26(5 Suppl 14): 21-5.
Stevens, S. J., E. A. Verschuuren, et al. (2001). "Frequent monitoring of Epstein-Barr virus DNA load in unfractionated whole blood is essential for early detection ofposttransplant lymphoproliferative disease in high-risk patients." Blood 97(5): 1165-
Su, I. J., Y. H. Hsu, et al. (1993). "Epstein-Barr virus-containing T-cell lymphomapresents with hemophagocytic syndrome mimicking malignant histiocytosis." Cancer72(6): 2019-27.
Selves, J., F. Meggetto, et al. (1996). "Inflammatory pseudotumor of the liver.Evidence for follicular dendritic reticulum cell proliferation associated with clonalEpstein-Barr virus." Am J Surg Pathol 20(6): 747-53.
Shek, T. W., F. C. Ho, et al. (1996). "Follicular dendritic cell tumor of the liver.Evidence for an Epstein-Barr virus-related clonal proliferation of follicular dendriticcells." Am J Surg Pathol 20(3): 313-24.
T

Takada K. (1999). [Virus-associated malignancies]. Ryoikibetsu Shokogun Shirizu:421-4.
Takada K., and Ono Y. (1989). Synchronous and sequential activation of latentlyinfected Epstein-Barr virus genomes. J Virol 63: 445-9.
The Eurogast Study Group (1993). An international association betweenHelicobacter pylori infection and gastric cancer. Lancet 341: 1359-62.
Timms J. M., Bell A., Flavell J. R., Murray P. G., Rickinson A. B., Traverse-Glehen A., Berger F., and Delecluse H. J. (2003). Target cells of Epstein-Barr-virus(EBV)-positive post-transplant lymphoproliferative disease: similarities to EBV-positive Hodgkin's lymphoma. Lancet 361: 217-23.
Tokunaga M.,UemuraY.,TokudomeT., Ishidate T.,Masuda H.,OkazakiE.,Kaneko K.,Naoe S., Ito M., Okamura A., and et al. (1993). Epstein-Barr virusrelated gastric cancer in Japan: a molecular patho-epidemiological study. Acta PatholJpn 43: 574-81.
Tajima M, Komuro M, Okinaga K. (1998). Establishment of Epstein-Barr virus-positive human gastric epithelial cell lines. Jpn J Cancer Res. 89:262-8.
Takada K. (1984). Cross-linking of cell surface immunoglobulins induces Epstein-Barr virus in Burkitt lymphoma lines. Int J Cancer. 33:27-32.
Tanner J, Weis J, Fearon D, Whang Y, Kieff E. (1987). Epstein-Barr virusgp350/220 binding to the B lymphocyte C3d receptor mediates adsorption, capping,and endocytosis. Cell. 50: 203-13.
Tokunaga M, Land CE, Uemura Y, Tokudome T, Tanaka S, Sato E. (1993).Epstein-Barr virus in gastric carcinoma. Am J Pathol. 143:1250-4.
Turner AR, MacDonald RN, Cooper BA. (1972). Transmission of infectiousmononucleosis by transfusion of pre-illness plasma. Ann Intern Med.77:751-3.
Tao, Q., Robertson, K. D., Manns, A., Hildesheim, A. & Ambinder, R.F. (1998).The Epstein-Barr virus major latent promoter Qp is constitutively active,hypomethylated, and methylation sensitive. Journal of virology, 72, pp. 7075-7083.
Thorley-Lawson, D. A. & Gross, A. (2004). Persistence of the Epstein-Barr virusand the origins of associated lymphomas. The New England journal of medicine, 350,pp. 1328-1337.
Thornburg, N. J., Kusano, S. & Raab-Traub, N. (2004). Identification of Epstein-Barr virus RK-BARF0-interacting proteins and characterization of expression pattern.Journal of virology, 78, pp. 12848-12856.
Tsurumi, T., Daikoku, T., Kurachi, R. & Nishiyama, Y. (1993). Functionalinteraction between Epstein-Barr virus DNA polymerase catalytic subunit and itsaccessory subunit in vitro. Journal of virology, 67, pp. 7648-7653.

Truong, C. D., W. Feng, et al. (2009). "Characteristics of Epstein-Barr virus-associated gastric cancer: a study of 235 cases at a comprehensive cancer center inU.S.A." J Exp Clin Cancer Res 28: 14.
Tokunaga, M., C. E. Land, et al. (1993). "Epstein-Barr virus in gastric carcinoma."Am J Pathol 143(5): 1250-4.
UUchida, J., Yasui, T., Takaoka-Shichijo, Y., Muraoka, M., Kulwichit, W., Raab-Traub, N. & Kikutani, H. (1999). Mimicry of CD40 signals by Epstein-Barr virusLMP1 in B lymphocyte responses. Science, 286, pp. 300-303.
VVan Baarle D., Hovenkamp E., Callan M. F., Wolthers K. C., Kostense S., Tan L.C., Niesters H. G., Osterhaus A. D., McMichael A. J., van Oers M. H., andMiedema F. (2001). Dysfunctional Epstein-Barr virus (EBV)-specific CD8(+) Tlymphocytes and increased EBV load in HIV-1 infected individuals progressing toAIDS-related non-Hodgkin lymphoma. Blood 98: 146-55.
Vo QN, Geradts J, Gulley ML, Boudreau DA, Bravo JC, Schneider BG. (2002).Epstein-Barr virus in gastric adenocarcinomas: association with ethnicity andCDKN2A promoter methylation. J Clin Pathol. 55(9): 669-75.
WWang X, Mori I, Tang W, Nakamura M, Nakamura Y, Sato M, Sakurai T,Kakudo K. (2002). P63 expression in normal, hyperplastic and malignant breasttissues. Breast Cancer. 9(3): 216-9.
Wang Y, Holland JF, Bleiweiss IJ, Melana S, Liu X, Pelisson I, Cantarella A,Stellrecht K, Mani S, Pogo BG. (1995). Detection of mammary tumor virus envgene-like sequences in human breast cancer. Cancer Res. 55(22): 5173-9.
Wang Y, Go V, Holland JF, Melana SM, Pogo BG. (1998). Expression of mousemammary tumor virus-like env gene sequences in human breast cancer. Clin CancerRes. 4(10): 2565-8.
Webb PM. and Forman D. (1995). Helicobacter pylori as a risk factor for cancer.Baillères Clin Gastroenterol 9: 563-82.
Webster-Cyriaque J., Middeldorp J., and Raab-Traub N. (2000). Hairyleukoplakia: an unusual combination of transforming and permissive Epstein-Barrvirus infections. J Virol 74: 7610-8.
Wrede D., Luqmani Y. A., Coombes R. C., and Vousden K. H. (1992). Absence ofHPV 16 and 18 DNA in breast cancer. Br J Cancer 65: 891-4.

Wang F, Gregory C, Sample C, Rowe M, Liebowitz D, Murray R, Rickinson A,Kieff E. (1990b). Epstein-Barr virus latent membrane protein (LMP1) and nuclearproteins 2 and 3C are effectors of phenotypic changes in B lymphocytes: EBNA-2and LMP1 cooperatively induce CD23. J Virol. 64:2309-18.
Wang X and Hutt-Fletcher LM. (1998). Epstein-Barr virus lacking glycoproteingp42 can bind to B cells but is not able to infect. J Virol. 72:158-63.
Wan, J., Zhang, W., Wu, L., Bai, T., Zhang, M., Lo, K., Chui, Y., Cui, Y., Tao,Q., Yamamoto, M., Akira, S. & Wu, Z. (2006). BS69, a specific adaptor in thelatent membrane protein 1-mediated c-Jun N-terminal kinase pathway. Molecular andcellular biology, 26, pp. 448-456.
Wei, M. X. & Ooka, T. (1989). A transforming function of the BARF1 gene encodedby Epstein-Barr virus. The EMBO journal, 8, pp. 2897-2903.
Wei, M. X., de Turenne-Tessier, M., Decaussin, G., Benet, G. & Ooka, T. (1997).Establishment of a monkey kidney epithelial cell line with the BARF1 open readingframe from Epstein-Barr virus. Oncogene, 14, pp. 3073-3081.
Weigel, R., Fischer, D. K., Heston, L. & Miller, G. (1985). Constitutive expressionof Epstein-Barr virus-encoded RNAs and nuclear antigen during latency and afterinduction of Epstein-Barr virus replication. Journal of virology, 53, pp. 254-259.
Wiech, T., Nikolopoulos, E., Lassman, S., Heidt, T., Schöpflin, A., Sarbia, M.,Werner, M., Shimizu, Y., Sakka, E., Ooka, T. & zur Hausen, A. (2008). CyclinD1 expression is induced by viral BARF1 and is overexpressed in EBV-associatedgastric cancer. Virchows Archiv : an international journal of pathology, 452, pp. 621-627.
Wilson, A. D., Redchenko, I., Williams, N. A. & Morgan, A.J. (1998). CD4+ Tcells inhibit growth of Epstein-Barr virus-transformed B cells through CD95-CD95ligand-mediated apoptosis. International immunology, 10, pp. 1149-1157.
Woisetschlaeger, M., Yandava, C. N., Furmanski, L. A., Strominger, J. L. &Speck, S.H. (1990). Promoter switching in Epstein-Barr virus during the initial stagesof infection of B lymphocytes. Proceedings of the National Academy of Sciences ofthe United States of America, 87, pp. 1725-1729.
Willett,W.C., Stampfer,M.J., Colditz,G.A., Rosner,B.A., Hennekens,C.H., andSpeizer,F.E. (1987). Moderate alcohol consumption and the risk of breast cancer. N.Engl. J. Med. 316, 1174-1180
Wooster,R., Neuhausen,S.L., Mangion,J., Quirk,Y., Ford,D., Collins,N.,Nguyen,K., Seal,S., Tran,T., and Averill,D. (1994). Localization of a breast cancersusceptibility gene, BRCA2, to chromosome 13q12-13. Science 265, 2088-2090.
Wade, M. & Allday, M.J. (2000). Epstein-Barr virus suppresses a G(2)/M checkpointactivated by genotoxins. Molecular and cellular biology, 20, pp. 1344-1360.

Winberg, G., L. Matskova, et al. (2000). "Latent membrane protein 2A ofEpsteinBarr virus binds WW domain E3 protein-ubiquitin ligases that ubiquitinate B-cell tyrosine kinases." Mol Cell Biol 20(22): 8526-35.
Wolf, H., C. Bogedain, et al. (1993). "Epstein-Barr virus and its interaction with thehost." Intervirology 35(1-4): 26-39.
Walling, D. M., C. M. Flaitz, et al. (2001). "Persistent productive Epstein-Barr virusreplication in normal epithelial cells in vivo." J Infect Dis 184(12): 1499-507.
Walling, D. M., C. M. Flaitz, et al. (2003). "Epstein-Barr virus replication in oralhairy leukoplakia: response, persistence, and resistance to treatment withvalacyclovir." J Infect Dis 188(6): 883-90.
XXue SA, Lampert IA, Haldane JS, Bridger JE, Griffin BE. (2003). Epstein-Barrvirus gene expression in human breast cancer: protagonist or passenger? Br J Cancer89: 113-9.
YYanai H., Takada K., Shimizu N., Mizugaki Y., Tada M., and Okita K. (1997).Epstein-Barr virus infection in non-carcinomatous gastric epithelium. J Pathol 183:293-8.
Yip T. T., Ngan R. K., Lau W. H., Poon Y. F., Joab I., Cochet C., and Cheng A.K. (1994). A possible prognostic role of immunoglobulin-G antibody againstrecombinant Epstein-Barr virus BZLF-1 transactivator protein ZEBRA in patientswith nasopharyngeal carcinoma. Cancer 74: 2414-24.
Young L.S., Dawson C.W., Clark D., Rupani H., Busson P., Tursz T., Jonhson A.and Rickinson A.B. (1984). Epstein-Barr virus gene expression in nasopharyngealcarcinoma. J. Gen. Virol. 69: 1080-1085.
Yao KT, Zhang HY, Zhu HC, Wang FX, Li GY, Wen DS, Li YP, Tsai CH,Glaser R. (1990). Establishment and characterization of two epithelial tumor celllines (HNE-1 and HONE-1) latently infected with Epstein-Barr virus and derivedfrom nasopharyngeal carcinomas. Int J Cancer. 45:83-9.
Young L, Alfieri C, Hennessy K, Evans H, O'Hara C, Anderson KC, Ritz J,Shapiro R S, Rickinson A, Kieff E, and et al. (1989). Expression of Epstein-Barrvirus transformation-associated genes in tissues of patients with EBVlymphoproliferative disease. N Engl J Med. 321:1080-5.
Yamamoto, N., Takizawa, T., Iwanaga, Y., Shimizu, N. & Yamamoto, N. (2000).Malignant transformation of B lymphoma cell line BJAB by Epstein-Barr virus-encoded small RNAs. FEBS letters, 484, pp. 153-158.

Yokoyama, A., Tanaka, M., Matsuda, G., Kato, K., Kanamori, M., Kawasaki, H.,Hirano, H., Kitabayashi, I., Ohki, M., Hirai, K. & Kawaguchi, Y. (2001).Identification of major phosphorylation sites of Epstein-Barr virus nuclear antigenleader protein (EBNA-LP): ability of EBNA-LP to induce latent membrane protein 1cooperatively with EBNA-2 is regulated by phosphorylation. Journal of virology, 75,pp. 5119-5128.
Young, L. S., C. W. Dawson, et al. (1988). "Epstein-Barr virus gene expression innasopharyngeal carcinoma." J Gen Virol 69 ( Pt 5): 1051-65.
ZZur Hausen H, O'Neill FJ, Freese UK, Hecker E. (1978). Persisting oncogenicherpesvirus induced by the tumour promotor TPA. Natur.
Zhang, Q., Gutsch, D. & Kenney, S. (1994). Functional and physical interactionbetween p53 and BZLF1: implications for Epstein-Barr virus latency. Molecular andcellular biology, 14, pp. 1929-1938.
Zimber-Strobl, U., Kremmer, E., Grässer, F., Marschall, G., Laux, G. &Bornkamm, G.W. (1993). The Epstein-Barr virus nuclear antigen 2 interacts with an