Embed Size (px)
Citation preview

ENZYMOLOGIE I) Propriétés générales des enzymes
1.1. Application
- En médecine pour le dépistage des maladies (dosage des prises de sang), par exemple une augmentation de concentration en amylase dans le sang cause un disfonctionnement du pancréas
- Outils industriels : lessive, fabrication de papier, de textile, agroalimentaire…
1.2. Propriétés de catalyseur
1.2.1. Efficacité Un catalyseur accélère une réaction et n’est pas consommé au cours de cette réaction, il est actif en faible concentration et ne modifie pas le cours thermodynamique de la réaction. 1.2.2. Spécificité Au niveau des liaisons coupées ou transformées :
- Chimiosélectivité ; ex : protéases qui coupent les liaisons peptidiques NH-CO et séparent les acides aminés
- Régiosélectivité ; ex : trypsine coupe après un acide aminé basique, chymotrypsine après un acide aminé aromatique
- Stéréosélectivité ; ex : L-amino-oxydase ou D-amino-oxydase, α ou β galactosidase, furamase qui réagit sur le furamate en configuration E pour donner du malate…
1.2.3. Régulation
La concentration en calcium intracellulaire, la phosphorylation ou l’allostérie (modification de conformation) peut moduler l’activité des enzymes.
1.3. Classification des enzymes
- Les oxydoréductases catalysent le transfert d’électrons et vont avoir comme substrat l’oxydant et le réducteur. (1)
- Les transférases catalysent le transfert d’un groupe d’atomes R de A vers B ou inversement. (2)
- Les hydrolases coupent la liaison souvent entre deux carbones ou entre un carbone et un soufre/oxygène/azote : C—X + H2O → C—OH + HX. Les réactions sont déplacées vers la rupture de la liaison. (3)
- Les lyases lient deux groupement d’atomes : A=B + X—Y → X—A—B—Y (4) - Les isomérases permettent la formation d’un isomère d’une molécule donnée :
X—A—B ←→ A—B—X (5) - Les ligases ou synthétases : produisent l’action inverse des hydrolases. (6)

II) Cinétique des réactions enzymatiques 2.1. Rappel sur la cinétique chimique La thermodynamique nous renseigne sur l’énergie de l’état initial et de l’état final, ce qui permet de déterminer si une réaction est faisable, mais ne renseigne pas sur sa vitesse. La cinétique renseigne sur la vitesse (variation de concentration en fonction du temps). La vitesse est fonction de la concentration et de la nature du substrat et du produit, de la température, de la pression, du pH…
!
V =dP
dt= "
dS
dt
2.1.1. Cinétique d’ordre 1
Substrat → Produit
!
V = k1S[ ] = "
dS
dt
k1 est la constante de vitesse d’ordre 1 (s-1).
!
lnS[ ]S0[ ]
= k1t
La courbe V = f (S) est une droite de pente k1. T1/2 est indépendant de la concentration initiale en substrat.
!
T1/ 2
=ln2
k1
Exemple : saccharose + H2O → glucose + fructose
!
V = k saccharose[ ] H2O[ ] = k
1saccharose[ ]
2.1.2. Cinétique d’ordre 2 A + B → P A + B → P + Q
!
V =dP
dt= "
dA
dt
!
V = k2A[ ] B[ ] (k2 en M-1.s-1)
!
1
A0" B
0
lnB0[ ] A[ ]A0[ ] B[ ]
= k2t si
!
A[ ] = B[ ],
!
V = k2A[ ]
2
!
1
A[ ]= k
2t +
1
A0[ ]
2.1.3. Cinétique d’ordre 0 V = k0, la vitesse est indépendante de la concentration.
!
V =dP
dt= k
0 et
!
P[ ] = k0t

2.2. Notion de vitesse initiale 2.2.1. État stationnaire k1 kcat E+ S ES E + P
!
V = kcat
ES[ ] k-1
- Phase pré-stationnaire (ms) - Phase stationnaire d’ordre 0 (Vi, en s, min) ≠ d’état d’équilibre - Phase post-stationnaire
Pour une unité d’enzyme : Vi est en µmol/min Pour un katal (énergie nécessaire pour transformer une mole de substrat, en SI) : Vi est en mol/s. 2.2.2. Vi = f(E) Si on fait varier la concentration en enzyme pour une concentration de substrat fixe, la formation de produit en fonction du temps reste linéaire, plus il y a d’enzyme, plus la réaction est rapide. Si la concentration en enzyme est exprimée en mg/L, on parle d’activité spécifique de l’enzyme. La cinétique est d’ordre 1 par rapport à l’enzyme.
!
Vi= k E[ ]
2.2.3. Vi = f(S) On fait varier la concentration en substrat pour une concentration en enzyme fixe. La vitesse initiale augmente de façon parabolique quand la concentration initiale en substrat augmente. 2.3. Équation de Michaelis Mentes Elle nécessite plusieurs conditions :
1) la concentration en produit doit être très inférieure à celle en substrat, la réversibilité de la réaction est ainsi proche de 0
2) la concentration en ES doit être constante au cours du temps k1 kcat E+ S ES E + P k-1
!
Vformation = k1E[ ] S[ ]
!
Vdisparition = k"1 ES[ ] + kcat ES[ ] = (k"1 + kcat ) ES[ ] À l’équilibre :
!
Vf =Vd donc
!
k1E[ ] S[ ] = (k"1 + k
cat) ES[ ]
!
Et[ ] = E[ ] + ES[ ] donc
!
E[ ] = Et[ ] " ES[ ]
!
k1Et[ ] " ES[ ][ ] # S[ ] = (k"1 + k
cat) ES[ ]

!
V = kcat
ES[ ] donc
!
V =kcatk1Et[ ] S[ ]
k1S[ ] + (k"1 + k
cat)
!
Vm = kcat
Et[ ] et
!
Km =k"1 + k
cat
k1
Donc :
!
V =Vm S[ ]S[ ] + Km
Cette équation n’est valable que dans les premiers temps de la réaction. Signification des constantes :
- Km : concentration initiale en substrat pour laquelle
!
Vi=1
2Vm
- Quand la concentration initiale en substrat est très supérieure à Km,
!
Vi=Vm = k
catEt[ ] (kcat est l’efficacité catalytique de l’enzyme, en s-1)
- Quand la concentration initiale en substrat est très inférieure à Km,
!
Vi=kcat
KmEt[ ] S[ ] (
!
kcat
Km est la constante de spécificité, de l’ordre de 108-109)
La comparaison de différentes constantes de spécificité permet de déterminer le meilleur substrat pour une enzyme donnée.
!
kcat
Km=
k1kcat
k"1 + kcat
Si kcat est très supérieure à k-1,
!
kcat
Km tend vers k1.
Représentations linéaires :
- Linewearer-Buck :
!
1
Vi
= f1
S[ ]0
"
# $ $
%
& ' '
!
1
V=
Km
Vm S[ ]0
+1
Vm (voir cours chimie du vivant)
- Eadie-Hofstee :
!
V = fV
S[ ]0
"
# $ $
%
& ' '
!
Vm =V S[ ] + Km[ ]
S[ ]0
=V +V
S[ ]Km
!
V =Vm "KmV
S[ ]0
2.4. Des effecteurs de l’activité enzymatique 2.4.1. Le pH
- pH optimum entre pK1 et pK2 : forme zwitterion - dénaturation aux pH extrêmes (réversible ou non), changement de conformation et
perte d’activité - substrat ionisable : l’activité va dépendre de sa forme ionique (et donc du pH)
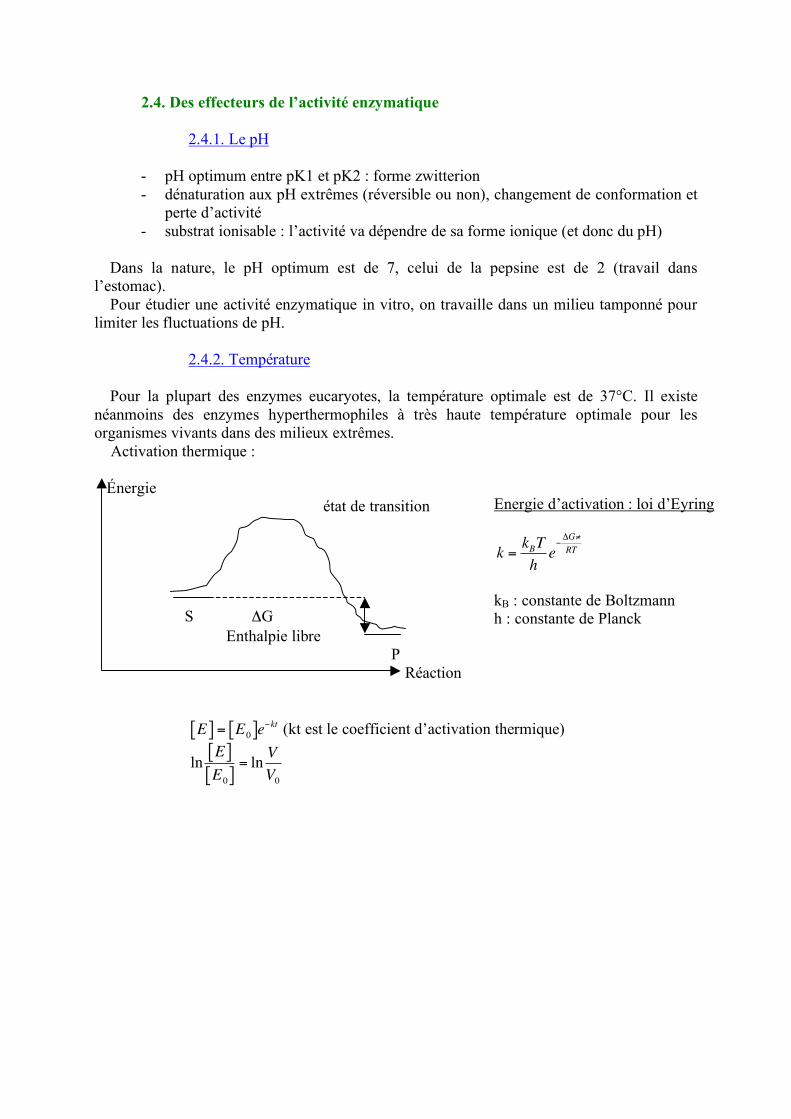
Dans la nature, le pH optimum est de 7, celui de la pepsine est de 2 (travail dans l’estomac). Pour étudier une activité enzymatique in vitro, on travaille dans un milieu tamponné pour limiter les fluctuations de pH. 2.4.2. Température Pour la plupart des enzymes eucaryotes, la température optimale est de 37°C. Il existe néanmoins des enzymes hyperthermophiles à très haute température optimale pour les organismes vivants dans des milieux extrêmes. Activation thermique : Énergie état de transition S ΔG Enthalpie libre
P Réaction
!
E[ ] = E0[ ]e"kt (kt est le coefficient d’activation thermique)
!
lnE[ ]E0[ ]
= lnV
V0
Energie d’activation : loi d’Eyring
!
k =kBT
he"#G$
RT
kB : constante de Boltzmann h : constante de Planck

2.4.3. Effets des inhibiteurs 2.4.3.1. Inhibiteurs irréversibles 2.4.3.2. Inhibiteurs réversibles
• Compétitifs : exemple du malonate Le malonate est l’inhibiteur du succinodeshydrogénase. Si l’inhibiteur est compétitif :
!
V =VMS[ ]
KM1+
I[ ]K
I
"
# $
%
& ' + S[ ]
!
KMapp = KM 1+I[ ]KI
"
# $
%
& ' , si I augmente, KMapp augmente
La vitesse maximale est invariable.
• Non compétitifs L’inhibiteur peut agir au niveau de E + S (KI) ou de ES (KI’). Si KI = KI’, on parle d’inhibition non compétitive mixte Si KI ≠ KI’, on parle d’inhibition non compétitive stricte
!
V =VMS[ ]
1+I[ ]K
I
"
# $
%
& ' KM
+ S[ ]( )
!
VMapp =VM
1+I[ ]KI
"
# $
%
& '
, si I augmente, VMapp diminue
Le KM reste invariable.
• Anti-compétitifs (ou incompétitifs) Inhibition au niveau de ES, formation d’ESI.
!
V =VMS[ ]
KM
+ S[ ] 1+I[ ]K
I
"
# $
%
& '
!
VMapp =VM
1+I[ ]KI
"
# $
%
& '
et
!
KMapp =KM
1+I[ ]KI

III) Régulation de l’activité enzymatique 3.1. Introduction La cellule doit réguler cette activité par souci d’économie d’énergie chimique, sans régulation enzymatique, les réactions enzymatiques du vivant atteindraient leurs états d’équilibre. Il existe différents points de contrôle de la cellule : 3.1.1 Contrôle à long terme
- Régulation de la quantité de protéines synthétisées : la cellule peut intervenir sur la transcription, traduction et dégradation des protéines
- Compartimentation cellulaire : isoenzymes (formes différentes d’une même activité enzymatique mais dont les propriétés catalytiques sont différentes)
- Compartimentation des réactions : E1 A ←→ B En régulant E1 ou E’1, on contrôle la quantité de A ou de B E’1
Exemple : Fruc-6P + ATP → Fruc 1-6 diP + ADP (phosphofructokinase) Fruc-6P + Pi ← Fruc 1-6 diP + H2O (fructose biphosphatase) 3.1.2. Contrôle immédiat
- Production d’inhibiteurs - Disponibilité du substrat : on fait évoluer le Km pour le faire tendre vers la
concentration cellulaire - Inhibition par le produit, exemple le Glc-6P inhibe l’hexokinase qui catalyse sa
formation
3.1.3. Contrôle intrinsèque
- Allostérie - Modification covalente (phosphorylation) - Clivage protéolytique
3.2. Allostérie
Il s’agit d’une régulation par changement conformationnel 3.2.1. Concept général Les enzymes sont soumises à des phénomènes de rétrocontrôle, il y a changement de conformation de ces enzymes allostériques qui possèdent une structure quaternaire. Leur comportement est non michaelien (effet coopératif). Ex : Aspartate transcarbamylase.

3.2.2. Effet coopératif
Courbe sigmoïde. Équation de Hill :
!
lnV
VM"V
#
$ %
&
' ( = n ln S[ ] " lnK
(n : indice de coopérativité) 3.2.3. Monod-Wyman-Changeux (modèle)
- Enzyme allostérique : oligomères (plusieurs sous-unités) - Chaque protomère (sous-unité) ne contient qu’un seul site pour un ligand - L’enzyme peut exister sous deux états conformationnels : relâché (forte affinité
pour le substrat) ou tendu (faible affinité pour le substrat) - Deux sous-unités de différentes conformations ne peuvent pas s’associer - Les propriétés cinétiques de l’enzyme sont différentes d’un état à l’autre
Effets des effecteurs allostériques :
- activateurs - inhibiteurs La courbe sigmoïde est déplacée vers la gauche ou la droite
3.3. Modifications covalentes (phosphorylation)
Phénomène de tout ou rien, l’enzyme régulée peut exister sous deux formes : E native ou E-X modifiée E1L1 ⇔ E1 + L1 X E E-X Actif Inactif E2L2 ⇔ E2 + L2 Exemple du pyruvate déshydrogénase NAD + CoA + pyruvate → NADH + acetyl CoA (cycle de Krebs) Glycolyse ATP NADH ADP acetyl CoA PDH PDH-P inactif Phosphatase ← Ca2+

Il y a activation de la voie de signalisation cellulaire (200 gènes pour les kinases, 2000 pour les phosphatases). 3.4. Clivage protéolytique La cellule synthétise des protéines sous forme inactive : ce sont les zymogènes, précurseurs d’enzymes. Un zymogène va être transporté sur son lieu d’action et va subir un clivage protéolytique pour donner une enzyme active. Exemple de la trypsine, synthétisée sous forme de trypsinogène qu’une entérokinase va cliver entre Lys et Ile (acides aminés 6 et 7) en une trypsine. On peut assister à une cascade enzymatique, par exemple la thrombine obtenue à partir du fibrinogène va catalyser la transformation du fibrinogène en fibrine insoluble. Gène Transcription ARNm Traduction
dégradation (protéosome) Protéines Clivage protéolytique phosphorylation Enzyme active enzyme inactive Allostérie substrat, produit, inhibiteur S P
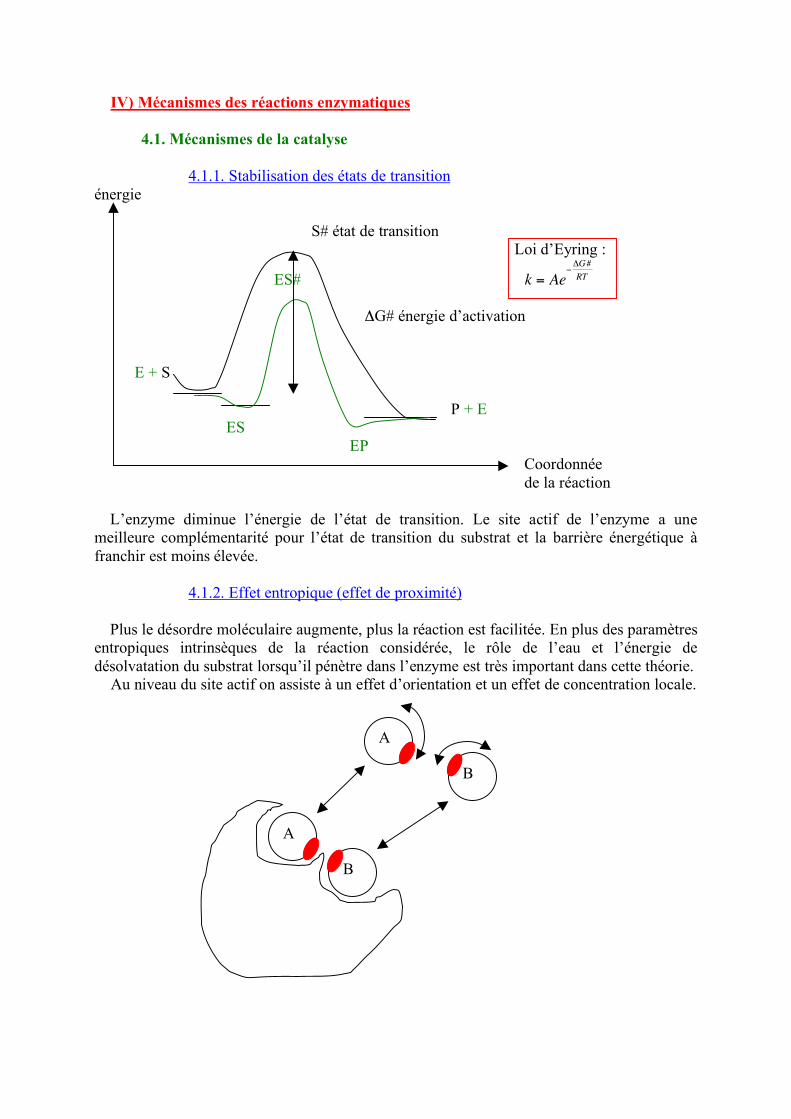
IV) Mécanismes des réactions enzymatiques 4.1. Mécanismes de la catalyse 4.1.1. Stabilisation des états de transition énergie S# état de transition Loi d’Eyring :
ES#
!
k = Ae"#G#
RT ΔG# énergie d’activation E + S P + E ES EP Coordonnée de la réaction L’enzyme diminue l’énergie de l’état de transition. Le site actif de l’enzyme a une meilleure complémentarité pour l’état de transition du substrat et la barrière énergétique à franchir est moins élevée. 4.1.2. Effet entropique (effet de proximité) Plus le désordre moléculaire augmente, plus la réaction est facilitée. En plus des paramètres entropiques intrinsèques de la réaction considérée, le rôle de l’eau et l’énergie de désolvatation du substrat lorsqu’il pénètre dans l’enzyme est très important dans cette théorie. Au niveau du site actif on assiste à un effet d’orientation et un effet de concentration locale.
A
B
A
B
4.1.3. Catalyse acide-base concertée Elle est induite par les groupes réactifs apportés par les chaînes latérales de l’enzyme. Milieu basique OH- O—R R— C O HA milieu acide 4.1.4. Formation d’un intermédiaire covalent énergie S# état de transition ES#1 ES#2 ΔG# énergie d’activation E + S EX P + E ES EP Coordonnée de la réaction Exemple de la protéase à sérine qui forme un ester intermédiaire. 4.2. Cofacteurs des réactions enzymatiques Il s’agit de substances qui aident l’enzyme en apportant des groupements réactifs non présents dans les protéines. On trouve deux types de cofacteurs :
Les ions métalliques : - activateurs (Na+, K+, Ca2+, Mg2+) - métalloenzymes (Mn2+, Co3+, Fe2+, Cu2+, Zn2+)
Les coenzymes : - cosubstrat (fixation réversible à l’enzyme) - groupement prosthétique (fixation permanente à l’enzyme)
4.2.1. Les ions métalliques
Ils assurent la fixation du substrat dans le site actif, son orientation et le masquage des charges. Les ions Mg2+ sont employés par exemple dans la réaction kinase → ATP. Ils assurent également des réactions d’oxydoréduction (apparition de hème) et des catalyses nucléophiles (anydrase carbonique).
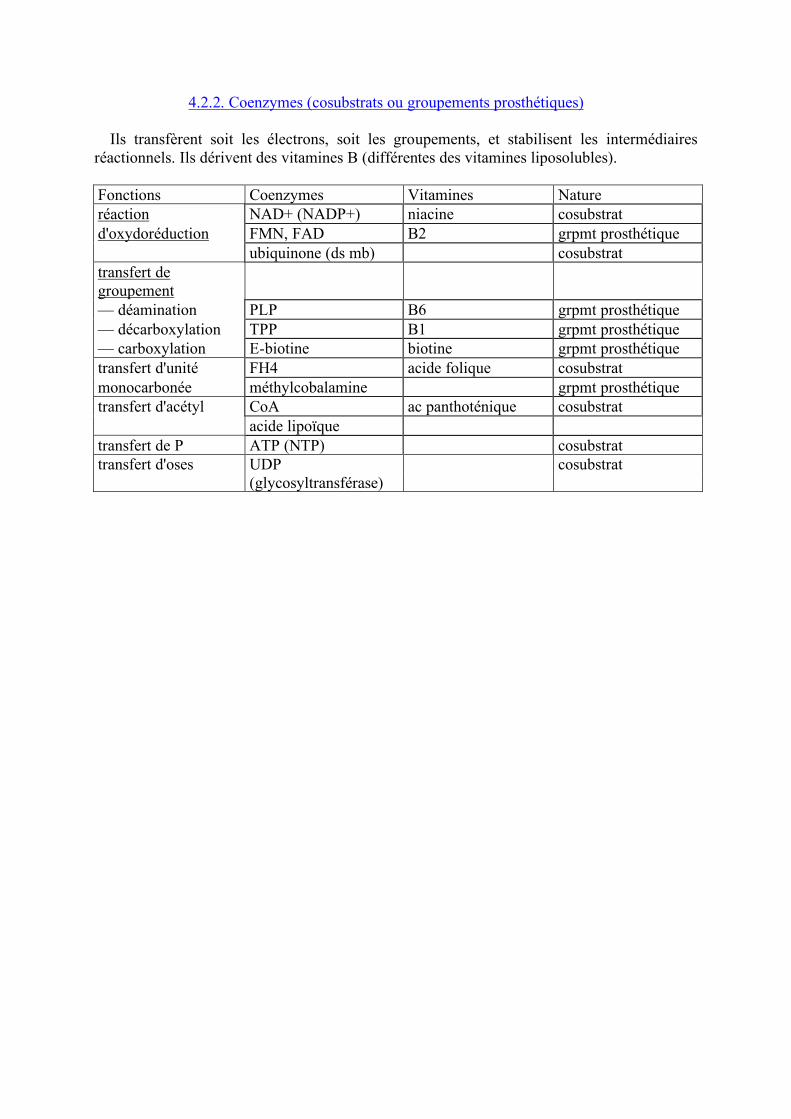
4.2.2. Coenzymes (cosubstrats ou groupements prosthétiques) Ils transfèrent soit les électrons, soit les groupements, et stabilisent les intermédiaires réactionnels. Ils dérivent des vitamines B (différentes des vitamines liposolubles). Fonctions Coenzymes Vitamines Nature réaction NAD+ (NADP+) niacine cosubstrat d'oxydoréduction FMN, FAD B2 grpmt prosthétique ubiquinone (ds mb) cosubstrat transfert de groupement — déamination PLP B6 grpmt prosthétique — décarboxylation TPP B1 grpmt prosthétique — carboxylation E-biotine biotine grpmt prosthétique transfert d'unité FH4 acide folique cosubstrat monocarbonée méthylcobalamine grpmt prosthétique transfert d'acétyl CoA ac panthoténique cosubstrat acide lipoïque transfert de P ATP (NTP) cosubstrat transfert d'oses UDP cosubstrat (glycosyltransférase)





























