Embed Size (px)
Citation preview

République Algérienne Démocratique et Populaire
Ministère de l’Enseignement Supérieur et de la Recherche Scientifique
Université d’Oran
Faculté des Sciences
Département de Biologie
Laboratoire Réseau de Surveillance Environnementale
Mémoire présenté pour l’obtention du diplôme de
Magister
en
Sciences de l’Environnement
Option : Biologie et pollution marine
Etude bactériologique du sable de quatre plages
(Beau Séjour, Eden, les Andalouses et Madagh)
du littoral ouest algérien.
Présenté par : Naima MESSAOUI
Soutenu le : / / 2011 devant la Commission du Jury composée de :
PRESIDENT : T. SAHRAOUI Professeur, Université d’Oran
EXAMINATEURS : M. HADADJI Maitre de Conférences, Université d’Oran
M. BOUDERBALA Maitre de Conférences, Université d’Oran
ENCADREUR Z. BOUTIBA Professeur, Université d’Oran
Co-ENCADREUR A. MATALLAH-BOUTIBA Maitre de Conférences, Université d’Oran

Remerciements
Je tiens avant tout à remercier Monsieur le Professeur Z. BOUTIBA Directeur
du Laboratoire Réseau de Surveillance Environnementale (LRSE) du
Département de Biologie, Faculté des Sciences de l’Université d’Oran qui m’a
permis de mener ce travail dans son laboratoire avec la plus grande liberté
d’action et de pensée.
J’exprime ma gratitude à Mme A. BOUTIBA-MATALLAH, Maître de
Conférences au Département de Biologie de l’Université d’Oran, qui m’a
encadré tout au long de ces trois années, m’a formé au métier de chercheur et
m’a permis de travailler dans d’excellentes conditions. Merci pour votre écoute,
votre gentillesse, pour toutes les réponses et explications à mes questions
incessantes, pour votre soutien moral et le grand intérêt que vous avez
manifesté à l’égard de ce mémoire. Merci pour votre bienveillance sur le bon
déroulement de ce travail, pour les nombreuses discussions scientifiques.
Je remercie vivement les membres du jury qui ont accepté d’évaluer ce travail,
remplissant encore un peu plus des agendas déjà chargés.
Monsieur le Professeur T. SAHRAOUI m’a fait l'honneur de participer au
Jury de soutenance et de le présider, je l’en remercie profondément.
Messieurs, les Maitres de Conférences au Département de Biologie de
l’Université d’Oran; M. BOUDERBALA et M. HADADJI, ont accepté
d'examiner ce travail, je les remercie vivement pour leur participation au Jury.
Ils ont également contribué par leurs nombreuses remarques et suggestions à
améliorer la qualité de ce mémoire et je leur en suis très reconnaissante.
Mes remerciements s’adressent aussi à tout le personnel du laboratoire Réseau
de Surveillance Environnementale (Département de biologie Université d’Oran);
et toute ma promotion de Magister.
Je remercie ma collègue et amie Asma de m’avoir soutenu, de m’être très
compréhensive et pour son soutien moral aussi.

Je n’oublierai pas ma famille, mes parents, frères et sœurs, qui m’ont beaucoup
donné et soutenu afin de réussir.
Toutes les personnes dont les noms apparaissent clairsemés dans ce manuscrit
sont ici remerciées. Acceptez ici l’expression de toute ma gratitude.

Résumé
Les plages présentent un vecteur d’attraction majeur, des sites hautement
fréquentés ; bon nombre de visiteurs de l’été, excursionnistes ou plaisanciers s’y
rendent uniquement dans la perspective de profiter des longues plages sableuses.
Suite à la grande fréquentation, une possible contamination du sable par des
communautés bactériennes pourrait constituer une source de transmission de
certaines bactéries pathogènes.
Un dénombrement des différentes flores bactériennes qui existent dans ce milieu
pourrait donner une estimation de son état de contamination.
Dans le cadre de ce travail, on souhaite mettre en évidence la présence de
certaines bactéries qui peuvent être source de maladie pour les baigneurs et
comparer les différents niveaux de contamination sur les quatre plages.
Les sables des plages Beau Séjour, Eden, Andalouses et Madagh ont été
analysés au cours de la saison pluvieuse et la saison sèche, de décembre 2010 à
juin 2011 en utilisant les biodindicateurs de contamination fécale.
Un total de trois points d’échantillonnage sur quatre différents sites était
sélectionné en fonction du nombre de baigneurs important et du montant des
eaux usées. Les échantillons ont été prélevés bimensuellement dans les quatre
plages de la côte oranaise. Dans ces sites de collectes un échantillon de sable
sec, sable humide et eau de mer ont été recueillis pendant six mois (saison sèche
et saison de pluie).
Des tests statistiques basés sur des modèles linéaires généraux ont été réalisés et
ont indiqué un niveau de signification pour les paramètres physico-chimiques et
climatiques pour les échantillons de sable.
Mots clés : Pollution, Coliformes fécaux, E. coli, plages, Beau Séjour, Eden, les
Andalouses, Madagh, sable.

Abstract
The beaches are a major vector of attraction, highly frequented sites, many
visitors in the summer, hikers and boaters go there only with a view to enjoy the
long sandy beaches.
Following the large attendance, possible contamination of the sand by bacterial
communities could be a source of transmission of certain pathogenic bacteria.
A count of the different Bacterial flora that exists in this environment could give
an estimate of its state of contamination.
As part of this work we want to show the presence of certain bacteria that can
cause illness to swimmers and compare the different levels of contamination on
the four beaches.
Sand beach of Beau Séjour, Eden, Andalouses and Madagh was analyzed during
the dry season and rainy season from December 2010 to June 2011 using
biodindicators of fecal contamination.
A total of three points on four different sampling sites were selected based on
the large number of bathers and the amount of wastewater. Samples were
collected fortnightly in the four beaches on the coast of Oran, in these sites to
collect a sample of dry sand, wet sand and seawater were collected during six
months (dry season and rainy season.
Statistical tests based on general linear models were made and indicated a level
of significance for the physical and chemical parameters for climate and sand
samples.
Keywords:
Pollution, Fecal coliformes, E. coli, beaches, Beau Séjour, Eden, les Andalouses,
Madagh, sand.

ملخص
تمثل الشواطئ عامل جذاب هام، أماكن مرتادة للغاية، عدد هام من الزوار في الصيف و المتجولين
.يذهبون اليها بغية التمتع بالشواطئ الرملية الطويلة
و يكون مصدرا النتقال البكتيريا تبعا للحضور الكبير في هذه األماكن، يحتمل أن يكون تلوث بكتيري
.المسببة لألمراض
.ب عدد مختلف أنواع البكتيريا في هذه البيئة، قد يمكننا من تقدير حالة التلوثحسا
، نريد اظهار وجود بكتيريا يحتمل أن تتسبب في عدة أمراض للمصطافين كما نود أن ي اطار هذا العملف
.نقارن مختلف مستويات التلوث في الشواطئ المدروسة
رمال Beau Séjour, Andalouses، Eden ،Madagh من حللت خالل الموسم الجاف و الممطر
.باستعمال المؤشرات الحيوية للتلوث البرازي 1022 حتى جوان 1020من ديسمبر
على مستوى أربعة مواقع اختيرت و ذلك استنادا للعدد الكبير من محبي أخذ للعينات مجموع ثالث نقاط
.السباحة ومستوى مياه الصرف الصحي المتدفق
جمع العينات مرتين شهريا في الشواطئ األربعة على مستوى ساحل وهران وقد تم جمع عينات وقد تم
.ف الرطب و المياه خالل ستة أشهر خالل الموسم الممطر و الموسم الجافامن الرمل الج
وكانت االختبارات اإلحصائية معتمدة على النماذج الخطية العامة و أشارت الى مستوى الترابط بين
.ات الفيزيو كيميائية و المناخية للرمالالمعلم
بقاء جميل ،عدن، األندلسيات، مداغ، ،القولونية الشاطئ، ،القولونيات البرازية التلوث:كلمات البحث
.الرمل

LISTE DES ABREVIATIONS
AIEA: Agence Internationale de l'Energie Atomique.
AM: Ampicilline.
AMC: Amoxicilline.
ATB: Antibiotique.
BCPL: Bouillon lactose au pourpre de bromocrésol.
D/C: Double concentration.
EMB: Milieu Eosine Bleu de Méthylène.
EPA: U.S. Environmental Protection Agency.
FAO: Food and agriculture organization.
GFCM General Fisheries Commission for the Mediterranean.
GM: Gentamycine.
GN: Gélose nutritive.
IOC: Inversion of Control.
MH : Milieu Muller Hinton.
OMS : Organisation mondiale de la Santé.
PNUE/UNEP : Programme des Nations Unies pour l'environnement.
S/C: Simple concentration.
UFC: unite formant colonie.
WHO: World Health Organization.

LISTE DES FIGURES
Figure 1 : Site d’Ain Turk.
Figure 2 : Site d’Eden.
Figure 3 : Site des Andalouses.
Figure 4 : Site de Madagh.
Figure 5 : Echantillons récoltés des quatre sites.
Figure 6 : Sites d’échantillonnage.
Figure 7 : Etapes suivies pour une identification bactérienne.
Figure 8 : Galerie biochimique API E20.
Figure 9 : Dépôt de disque d’ATB sur milieu MH.
Figure 10 : Ensemencement sur milieu gélosé.
Figure 11 : Principe de la recherche des coliformes.
Figure 12 : Lecture de la galerie API 20E.
Figure 13 : Variations du pH selon les saisons.
Figure 14 : Variation de la température selon les saisons.
Figure 15 : Concentration des flores dans les quatre sites durant toute la période
de l’étude.
Figure 16 : Variations en charges bactériennes mensuelles (ufc/100ml) dans le
sable sec, sable humide et l’eau dans les quatre sites.
Figure 17 : Corrélation entre les Coliformes fécaux dans l’eau et les Coliformes
fécaux dans le sable pour les quatre sites.
Figure 18 : Corrélation entre les Coliformes fécaux et les Cliformes totaux dans
le sable sec pour les quatre sites.
Figure 19 : Abondance des germes selon les différentes saisons.
Figure 20 : Abondance des flores bactériennes selon les sites.
Figure 21 : Dominance entre les flores bactériennes.

LISTE DES TABLEAUX
Tableau 1 : Temps de liaison de plusieurs particules selon le diamètre.
Tableau 2 : Principaux groupes de virus pathogènes excrétés dans la matière
fécale et les maladies transmises.
Tableau 3 : Principaux agents bactériens pathogènes présents dans les selles et
les maladies transmises.
Tableau 4 : Stations d’échantillonnage.
Tableau 5 : Classification caractéristique des antibiotiques utilisés dans cette
étude.
Tableau 6 : Seuils de sensibilité - résistance des différents antibiotiques testés.
Le diamètre est exprimé en mm.
Tableau 7 : Tableau des corrélations.

LISTE DES ANNEXES
Annexe 1: Les variations du pH pendant la période de l’étude.
Annexe 2: Les variations de la température pendant la période de l’étude.
Annexe 3: La composition des milieux de culture.
Annexe 4: Test de Levène pour les germes totaux.
Annexe 5: Test de Levène pour les Staphylocoques.
Annexe 6: Test de Levène pour les Pseudomonas.
Annexe 7: Test de Levène pour les Coliformes totaux.
Annexe 8: Test de Levène pour les Coliformes fécaux.
Annexe 9: Test de Levène pour les E. coli.

SOMMAIRE
Introduction …………………………………………………………………..….1
PREMIERE PARTIE : Revues bibliographiques sur les peuplements
bactériologiques du sable.
1. Les plages……………………………………………………………………..3
2. La diversité bactérienne dans différentes conditions
environnementales……………………………………………………………….7
3. Evaluation de la pollution microbienne en méditerranée ……………....17
3.1. Sort de la pollution microbienne en Méditerranée………………………...17
4. Les polluants hydriques…………..………………………………………….20
5. La flore des sables de plage……………………………...……………….....21
5.1. Microorganismes d’indication fécale……………………………………...21
5.2. Staphylocoques ……………………….………………………………23
5.3. Pseudomonas aeruginosa ………………….……………….………….…24
5.4. Vibrio. ……………………………………………………………….........24
5.5. Bactéries entériques …………………………………………………….....24
5.6. Mycètes…………...……..……………………...…….……………...….…25
5.6. 1. Mycètes saprophytiques ……………………...……………………........25
5.7. Virus et parasites ………...……………………………………..............27
5.8. Composition du sable en agents pathogènes ……….……………….......27
6. Dispersion et destin des microorganismes en sable de plage ….…………...28
7. Le transport bactérien dans le sable ………………………………………....32
7.1. Adhésion bactérienne …………………………………..…….............35
7.1.1. Les étapes de l’adhésion bactérienne …………………………………..36
a) Accès à la surface …………………….…………………………….…..36
b) Adhésion initiale ……………………….………………………………37
c) Attachement ………………………………...…………..................37
d) Colonisation de la surface ………………………………………....37
7.1.2. L’approche physicochimique ………………………………………....38
7.1.2 .1.Sensibilité des interactions électrostatiques à l’environnement…….…38
a) La force ionique…..………………………….....………………………...38
b) L’effet du pH de la solution sur les interactions électrostatiques ………..39
7.1.3. Les facteurs biologiques ………………………………...………………40
7.1.3.1. Polymères de la surface cellulaire ……………………………….….40
a) Les polymères extracellulaires (ou EPS)………...……………………….40
b) Les Lipopolysaccharides (LPS).……………….…………………….…..41

c) Les pili …………………………………………………………………...41
7.1.3.2. Motilité cellulaire et transport bactérien dans le sable………………...42
7.1.3.3. Etat et condition physiologique…………………………………….….43
7.1.3.4. Densité cellulaire…….……………………………………………...…44
7.2. La filtration………………………………………………….………..........45
7.2.1. Le milieu poreux ……………………………………………………….45
7.2.2. La morphologie des bactéries …………………….…………………...47
7.2.3. L’obstruction des pores ……….……………………………………….48
7.2.4. Saturation en eau et charge hydraulique en sable ……………………..48
7.3. Forces Hydrodynamiques ………………………………………………..49
8. Le transport de bactéries pathogènes dans le sable………………………….49
8.1. Le sort des bactéries pathogènes entérique dans un sol………………......51
8.1.1. La survie ………………………………….…………………………..51
8.1.2. Le déplacement ……………………………………………….............51
8.1.3. Détection de pollutions fécales ………….………………………….......52
DEUXIEME PARTIE: Matériels et méthodes.
1. Sites d’étude……….…………………...……………………………............57
2. Echantillonnage………..………………………………………………….....59
3. Mesure du pH …………………………………………………………...…60
4. Mesure de la température ……………………………………...................60
5. Préparation des échantillons aux analyses microbiologiques ……….….......60
5.1. Les dilutions ……….…………………………………………..………....60
6. Dénombrement des germes aérobies totaux …………………………....61
7. Dénombrement des Coliformes ……………………..………………......62
7.1. Coliformes fécaux ……………………………………………..……...…63
7.1.1. Colimétrie en milieu liquide…..………………...…………….................64
7.1.2. Numération par la méthode NPP (Nombre le Plus Probable)……..........64
7.2. Dénombrement d’Escherichia coli …………………………………..65
8. Dénombrement des Staphylocoques totaux ……………………………66
9. Dénombrement des Pseudomonas totaux ……..……………………………66
10. Tests de confirmation et d’identification …………………………………..67
10.1 .Caractérisation d’enzymes …………………………………………67
10.1.1. Mise en évidence d’enzymes respiratoires : Test
cytochrome oxydase…………………………………………………...….…....67
10.1.2. Test catalase ………………………………….………...……….....67
10.2. Coloration de Gram…………..…………………………………....……..68

10.3. Antibiogramme………………………………………………....…..…….69
10.3.1.Galerie Api 20E [Biomérieux]….…...……………………………..…...69
10.3.1.1. Préparation de l’inoculum………………..……………………..........70
TROISIEME PARTIE: Résultats et discussion
1. Les résultats……………………………………………..…………………..75
1.1. Les paramètres abiotiques …………………………………………..……75
a) Le pH …………………...………………………………..........................75
b) La température ………………………………..………………….….76
1.2. Suivi des différentes populations bactériennes …………………………....77
1.2.1. Estimation des bactéries mésophiles cultivables
(germes totaux) ……………………………………………….......................81
1.2.2. Estimation des polluants fécaux………….……………………………...81
1.2.3. Estimation des Staphylocoques totaux……….…………………..……..82
1.2.4. Estimation des Pseudomonas totaux…………………………………..83
1.3. Etude comparative des différents germes dans le sable des
quatre plages……………………..…………….……………………………….83
2. Discussion….………………………………………………………………...91
Conclusion……………………………………………………………………...97
Références bibliographiques………….…………………………………..…....99
Annexes..………………………..……………………………..………….......131

Introduction

Introduction
1
La Méditerranée dans son ensemble compte environ 45 000 km de côtes.
La population totale des pays côtiers méditerranéens est de 450 millions
d’habitants environ (7% de la population mondiale) à laquelle il faut ajouter
environ 200 millions touristes internationaux. Avec près de la moitié de la
population méditerranéenne vivant près des côtes, le littoral méditerranéen
figure parmi les secteurs les plus densément peuplés et les plus fortement
urbanisés de la planète. D’ici 2025, ce pourcentage d’occupation des côtes
devrait passer de 50% à 88%, et l’artificialisation de son littoral de 40 à 50%
(Ifremer, 2007). Le littoral oranais est de plus en plus agressé de par toutes les
nuisances du monde civilisé: activités industrielles, tourisme intensif et
urbanisation massive avec comme corollaire une ampleur sans cesse croissante
d’une pollution d’origine domestique (Boutiba et al, 2003).
La pression humaine le long du littoral est très importante. On estime à 1.5
millions le nombre d’habitants qui résident en permanence sur la côte ouest et
près de dix fois plus durant la période estivale (Boutiba et al, 2003). La frange
côtière subit un accroissement des rejets d’eaux usées sans aucun traitement, à
l’origine d’une pollution microbienne et d’une contamination des eaux de
baignade par des matières fécales (Boutiba et al, 2003).
Sur toutes les plages, ce sont en général les détritus qui gâchent l’agrément du
lieu. On trouve toutes sortes de déchets dans l’eau ou échoués sur le sable :
déchets alimentaires et emballages, bouteilles et boîtes de conserves, mégots,
poissons morts, préservatifs et serviettes hygiéniques usagées, seringues,
aiguilles et autres déchets médicaux(OMS, 2004).
La plupart des activités domestiques pour ne pas dire toutes génèrent des déchets
(solides, liquides et gazeux) qui sont responsables d’intrants dans le sol, donc
une modification de ce milieu surviendra. Mimouni et al, (2002) soulignent
l’influence du rejet des eaux usées sur la qualité microbiologique des plages et
confirmaient que le sable concentrait les polluants.

Introduction
2
Des bactéries, des champignons, des parasites et des virus ont été isolés dans le
sable des plages. Certains peuvent être pathogènes. Les facteurs qui influent sur
la survie et la dispersion des agents pathogènes sont la nature de la plage, les
marées, la pollution par les eaux usées, la saison, le passage d’animaux et la
fréquentation par les nageurs. La transmission peut se faire par contact direct
entre personnes ou par d’autres voies, mais aucun mode de transmission n’a été
mis en évidence avec certitude (OMS, 2004).
On a émis l’hypothèse que le sable des plages et d’autres matériaux similaires
pourraient être des réservoirs ou des vecteurs d’infection, mais le pouvoir
infectant des microorganismes mis en évidence dans le sable n’a toujours pas été
démontré, de sorte qu’on ignore quels risques ils présentent au juste pour la
santé publique. Par conséquent, aucun élément ne justifie le calcul d’un seuil
pour les bactéries servant d’indicateurs ou pour les microorganismes pathogènes
qui se trouvent dans le sable (OMS, 2004).
Les excréments d’animaux, en particulier les excréments de chien, sont la
principale source d’infection pour l’homme sur les plages et les lieux semblables
(OMS, 2004).

Revues
bibliographiques sur
les peuplements
bactériologiques du
sable

Revues bibliographiques sur les peuplements bactériologiques du sable
3
1. Les plages
Les plages représentent le sédiment non consolidé qui se trouve à la jonction
entre l'eau (océans, lacs et fleuves) et la terre et se composent habituellement de
sable, boue ou cailloux. (Nestor et al, 1984 ; Roses Codinachs et al, 1988 ;
Mendes et al, 1997). Une plage est une accumulation de sédiment, parfois
grossiers (plage de galets et de blocs, sur la côte extérieure) ou plus fins- c’est
le sable- qui pour le géologue est constitué de particules dont la taille est
comprise entre 0,2 et 2 mm. Cette définition pose d’emblée le problème de
l’origine de ces particules (Boughaba, 1994)
Les sables et les galets des plages proviennent de diverses sources. Ils sont transportés le
long de la côte par la dérive littorale, courant né de l'obliquité de la houle par rapport au
rivage et ils sont déposés là où la houle s'affaiblit (Piriou et al, 1991).
Cependant, l'énergie libérée par les vagues qui déferlent, comme celle liée aux courants
de marée, est capable de les remettre en mouvement. Lorsque, sur une plage, il y a plus
de matériaux qui arrivent que de matériaux qui s'en vont, son «budget sédimentaire»
est positif et elle s'élargit. On dit qu'elle prograde. Dans le cas contraire, elle démaigrit
et perd du terrain au profit de la mer. Si le budget est équilibré, elle est stable (Bird,
1985).
D’abord, comment sont les petites particules amenées par la mer et pourquoi se
déposent-elles ? Une loi assez curieuse, résultant de frottements dans les fluides
et d’une force liée à la rotation de la terre, la force de Coriolis, veut que lorsque
le vent souffle sur l’eau, l’eau se mette en mouvement… non dans le sens du
vent mais dans une direction faisant un angle avec celui-ci de 20 à 40° vers la
droite ; on parle de dérive littorale (Boughaba, 1994).
Le domaine marin participe à l'alimentation des plages, surtout grâce aux produits du
recul des falaises. Les vagues de tempête sont aussi capables de remonter des sédiments
depuis la proche avant-côte, en particulier ceux de nature biodétritique, comme le
sont les sables coquilliers. Cependant, l'essentiel des matériaux des plages a une

Revues bibliographiques sur les peuplements bactériologiques du sable
4
origine terrigène et, à cet égard, l'apport des cours d'eau est aujourd'hui primordial
(McBride, 1991).
Les mouvements de la mer entraînant donc les particules ; mais pourquoi se
déposent-elles parfois pour former des plages ? Toutes les particules ont
tendance à se déposer dans l’eau, mais l’agitation les remet en suspension. Plus
l’agitation est forte, plus les grosses particules sont entraînées, plus l’agitation
est faible, plus les petites particules peuvent se déposer. Dans des conditions
moins agitées, ce sont des galets mais aussi des sables qui se déposent. (Piriou
et al, 1991).
L’origine des matériaux de la plage est multiple. Beaucoup sont des constituants
peu ou lentement altérables des roches soumises à l’érosion et à l’altération,
arrachée aux continents par des fleuves, mais aussi aux falaises par la mer. Il
existe donc deux types de côtes :
*Les côtes d’érosion où l’attaque par la mer
*Les côtes de dépôts, où domine la sédimentation (Paskoff, 1998).
Il existe un autre constituant, d’origine biologique celui-là ; le calcaire. Cette
substance, altérable provient des coquilles d’animaux marins.
Les plages sont marquées par les organismes vivants. D’abord, il s’échoue une
grande quantité de matière organique, les « laisses de mer ». En effet, poussés
par les vagues, les objets flottants finissent le plus souvent à la côte (Gillet et al,
2000).
Divers animaux prolifèrent sur les laisses de mer ; c’est le cas de la Puce de mer
(Talitres saltator), par exemple, mais aussi d’autres Crustacés et Insectes, ainsi
que de nombreuses bactéries et organismes unicellulaires moins visibles. Les
grains de sable ménagent entre eux de larges trous, qui laissent bien passer
l’oxygène et permettent à ces organismes d’aller chercher ces débris, même
quand ils sont enfouis (Gillet et al, 2000).

Revues bibliographiques sur les peuplements bactériologiques du sable
5
Il n y a pas d’accumulation de matière organique sous les plages, à l’inverse de
ce qui se passe lorsque les sédiments sont plus fins et donc moins aérés, la
richesse en matière organique reflète la couleur grise des sables de plage
(Paskoff, 1998).
Les laisses de mer sont donc remobilisées et disparaissent de la plage, tant que
l’apport n’est pas trop important. Par exemple, les excréments des petits
animaux détritivores, finement broyés, vont être remportés à marée haute par la
mer, où ils constituent ensuite un engrais pour les organismes microscopiques du
plancton. On sait aussi que de nombreux oiseaux fréquentent les plages pour se
nourrir : beaucoup, comme tournepierre à collier, consomment les animaux
détritivores. Les fientes qu’ils produisent à terre jouent un rôle fertilisant pour
les sols des régions côtières. En ce sens, la plage, comme tous les autres points
du littoral, est une porte d’échange de matière entre écosystèmes continentaux et
marins. (Gillet et al, 2000).
Mais la vie de la plage encore variée à l’échelle microscopique, entre les grains
de sables, bien aérés mais aussi tenus à humides par les eaux, soit salées laissées
par la marée haute, soit plus douces, issues du ruissellement des eaux de pluie
(Pasqualini ,1997).
Dans les parties superficielles, des algues, comme les Diatomées ou de petites
algues vertes filamenteuses, et des Cyanobactéries, équivalentes des plantes
dans nos écosystèmes terrestres, effectuent la photosynthèse (Pasqualini ,1997).
Des organismes unicellulaires, comme des Ciliés et de petits animaux
consomment ces algues ; ils sont à leur tour consommés par des animaux
carnivores, tout aussi microscopiques. Les animaux de ces sables appartiennent
aux crustacés et aux Vers, mais aussi à des groupes moins connus comme les
Tardigrades et les Nématodes (Paskoff, 1993).
Sous les serviettes de plage, un petit monde nombreux et invisible mène donc
une vie active, dont un rôle majeur est l’épuration des plages. C’est ainsi que

Revues bibliographiques sur les peuplements bactériologiques du sable
6
disparaissent lentement les traces d’hydrocarbures amenés par la mer, au moins
lorsqu’elles arrivent en petite quantité (Miossec, 1998).
A l’échelle de l’année, on observe souvent que le sable est emporté par les
tempêtes d’hiver ; ce processus dit de « démaigrissement », annuel, s’explique
par les grandes tempêtes d’hiver et l’importance du ruissellement des eaux de
pluie (Miossec, 1998).
L’entraînement concerne sélectivement les minéraux les moins denses, ce qui
explique la reconcentration des minéraux lourds, qui colore certaines plages en
hiver (Begin et al, 1989).
Les plages connaissent des variations saisonnières qui font alterner des phases de
démaigrissement et des phases d'engraissement. Aux latitudes tempérées, pendant
l'hiver, les vagues de tempête leur enlèvent des sédiments qui s'accumulent sur
l'avant-côte sous la forme de bancs immergés. Ils seront ensuite restitués pendant
les périodes de beau temps qui voient ces bancs s'effacer. Ce processus naturel de
restauration peut être plus ou moins long. C'est seulement lorsqu'il est incomplet que
l'on doit conclure à un état d'érosion sur le long terme. La méconnaissance de cette
évolution naturelle fait que l'on décide parfois à la hâte, après de grandes tempêtes,
des travaux de défense contre la mer qui ne sont pas justifiés si la situation de crise est
seulement passagère (Begin et al, 1989).
Le vent peut aussi prendre en charge le sédiment de la plage : c’est l’origine des
dunes littorales. Les dunes qui constituent des espaces complémentaires des
plages en arrière desquelles elles se situent. Ces dunes sont caractérisées par la
présence d'une couverture végétale et peuvent également abriter des
microorganismes symbiotiques tels les Rhizobia et les champignons
endomycorhiziens (Salerno, 2006).
Les Rhizobia sont des bactéries fixatrices d'azote atmosphérique une fois
associées à leur plante hôte. Malheureusement, la littérature traitant l'activité de
ces bactéries dans les dunes est très fragmentaire (Pourcher et al, 2007).

Revues bibliographiques sur les peuplements bactériologiques du sable
7
2. La diversité bactérienne dans différentes conditions environnementales
La recherche de vie dans les habitats terrestres extrêmes occupe aussi une place
centrale dans les interrogations sur les origines de la vie et sur la possibilité de
vie extra-terrestre.
Au cours des dernières décennies, la gamme connue d’environnements habités
s’est très largement étendue, y compris à des environnements dépourvus d’eau
liquide.
Une des plus surprenantes découvertes a été celle, en 1977, des écosystèmes
océaniques profonds associés aux sources hydrothermales, premier écosystème
connu totalement basé sur la production primaire de bactéries
chémiosynthétiques (Prieur et al, 1995).
L’endroit le plus aride de la planète, le désert d’Atacama au Chili, est le seul
environnement naturel probablement stérile à la surface de la Terre (Navarro-
Gonzalez et al, 2003).
Marion et al, (2003) donnent les caractéristiques extrêmes des environnements
dans lesquels la vie existe de façon certaine :
1. la gamme de température s’étend de 20°C à 121°C) ;
2. la gamme de salinité, mesurée en termes d’activité de l’eau, va de 0,6 à 1 ;
3. le pH va de 0 à 13 ;
4. la pression (hydrostatique) va de 0 (vide poussé) à 1100 bar.
Les extrêmophiles, « organismes dans des habitats où la température, le pH, la
salinité ou la pression sont extrêmes » (Ciaramella et al, 2002) sont regroupés
en catégories selon le stress auquel ils sont adaptés :
– les thermophiles et psychrophiles sont respectivement adaptés aux hautes et
basses températures, des plus chaudes sources hydrothermales (Deming, 1993)
jusqu’au permafrost sibérien (Bakermans et al, 2003).
-Les barophiles sont adaptés aux fortes pressions1, comme celles des grands
fonds océaniques (Margesin, 2004) ;

Revues bibliographiques sur les peuplements bactériologiques du sable
8
– Les acidophiles, alcalinophiles et halophiles sont adaptés à l’acidité, la basicité
ou la forte salinité du milieu (comme les bassins hypersalins et anoxiques au
fond de la mer méditerranée, qui résultent de la dissolution de dépôts de sel du
Miocène, (van der Wielen et al, 2005).
– Les aérophiles résistent à un transport atmosphérique (Lighthart et Shaffer,
1994).
Le développement des activités humaines élargit encore la gamme de conditions
dites « Extrêmes ».
L’industrialisation et les pollutions associées, notamment chimiques,
organiques et radiologiques, créent des environnements artificiels qui sont aussi
en quasi-totalité colonisés par des microorganismes adaptés (Backman et
Jansson, 2004).
Même l’exploration spatiale a permis d’isoler des bactéries, contaminant les
systèmes d’eau de la station spatiale internationale, tolérant la microgravité
(Baker et Leff, 2005) ou capables, sur une courte durée, de survivre au vide (et
donc à la dessiccation) et aux radiations UV intenses du milieu spatial (Saffary
et al, 2002).
Des tests balistiques sont menés, qui visent à reproduire le choc de la chute
d’une météorite et à prouver que des spores portée par la météorite pourraient y
survivre (Benardini et al, 2003).
Les organismes capables de vivre dans les environnements pollués, en dégradant
les composés polluants, ont un intérêt évident même si les connaissances
nécessaires à l’élaboration de procédés de dépollution efficaces sont très
importantes (Salerno, 2006).
Les utilisations technologiques des organismes extrêmophiles dépassent très
largement la bioremédiation. De nombreuses protéines thermostables,
maintenant indispensables en recherche et en industrie, ont été isolées à partir
de bactéries du genre Thermus, les chercheurs exploitent maintenant les données

Revues bibliographiques sur les peuplements bactériologiques du sable
9
issus du séquençage intégral du génome d’organismes extrêmophiles comme
Thermus thermophilus (Benardini et al, 2003).
Par rapport aux environnements cités plus haut, il apparait nécessaire de
distinguer la faculté des organismes de survivre à l’application d’un stress
(ponctuelle ou plus fréquente ou prolongée), de la faculté d’avoir une activité
métabolique normale ou quasi-normale en présence de ce stress. Les déserts
chauds se caractérisent par le caractère intermittent des extrêmes, de
température et d’humidité. La condition extrême, la chaleur d’après-midi
associée à une très forte dessiccation, ne permet aucune activité microbienne.
Par contre, les organismes qui y survivent, à l’´etat dormant, profitent des
périodes de conditions plus clémentes ; ces organismes sont qualifiés
d’« anhydrobiotiques ». (Benardini et al, 2003).
Ciaramella et al, (2002) décrit il y a plus de trois cents ans la reviviscence des
animalcules (Rotifères), initialement séchés, lors de leur réhydratation. Le cas le
plus étudié actuellement est celui des croûtes microbiennes des déserts.
Garcia-Pichel et Pringault, (2001), ont étudié les bad-lands d’Espagne et ont
montré que les cyanobactéries se déplacent activement verticalement en réponse
à l’humidification ou à l’assèchement de la surface du sol.
Ceci permet d’assurer au mieux la production primaire, photosynthétique, quand
les conditions sont favorables. Toutefois, les perturbations de l’écosystème,
comme le piétinement par le bétail, augmentent la mortalité des bactéries,
qu’elles soient dormantes ou non ( Garcia-Pichel et Pringault, 2001),
Cable et Huxman, (2004), ont mesuré la part des croûtes microbiennes dans la
respiration du sol du désert de Sonora en fonction de l’intensité des pulses
expérimentaux de précipitations. Lors des évènements les plus intenses, les
plantes et bactéries du sol contribuent à la quasi-totalité du flux de CO2.
Lors des évènements de faible intensité, qui constituent l’essentiel des
précipitations en Arizona, le « réveil » des organismes des croûtes participe à

Revues bibliographiques sur les peuplements bactériologiques du sable
10
hauteur de 80% à la production de CO2 par le sol. Les croûtes contribuent
fréquemment à la production de l’écosystème.
Billi et Potts (2002), posent les questions fondamentales de l’étude des
mécanismes de tolérance à la dessiccation. Comment certaines espèces
bactériennes font-elles face au déficit en eau alors que d’autres ne le peuvent
pas ? Combien de temps les cellules desséchées restent-elles viables ? Deux
processus entrainent un efflux de l’eau des cellules, le dessèchement par l’air et
le stress hypertonique.
Mais même chez les plus extrêmes des bactéries halophiles, la perte d’eau reste
très inférieure à celle des cellules anhydrobiotiques. Chez ces dernières, le
contenu résiduel est souvent moins de 0,1 g/g poids sec (et cette eau ne suffit
même pas à maintenir une monocouche d’eau autour des macromolécules,
rendant notamment impossible toute réaction enzymatique). On considère que
les bactéries sensibles à la dessiccation sont celles qui meurent quand leur
contenu en eau est réduit à 0,3 g/g poids sec. Les premiers stades du
desséchement entrainent un stress osmotique, dont les micro-organismes
se prémunissent principalement par l’accumulation intracellulaire de solutés.
Les solutés organiques accumulés, comme le tréhalose (un disaccharide), la
proline (un acide aminé) et la glycine-bétaine (carboxyméthyl-
triméthylammonium) sont dits « compatibles » car ils ne perturbent pas le
fonctionnement des macromolécules et le métabolisme cellulaire (Kempf et
Bremer, 1998). Une telle contrainte de « compatibilité » explique sans doute le
fait que ces solutés sont les mêmes chez les bactéries, les archées et les
eucaryotes selon les mêmes auteurs.
La cellule maintient une activité plus ou moins normale tant que l’accumulation
des solutés évitent des pertes d’eau trop importantes. Surviennent ensuite les
dommages liés à la dessiccation. Les changements de conformation des
protéines entrainent un mauvais fonctionnement des enzymes, des chaînes de
transport d’électrons. Le stress oxydant peut alors entraîner des dommages

Revues bibliographiques sur les peuplements bactériologiques du sable
11
chimiques aux molécules biologiques, notamment à l’ADN. Le manque d’eau
perturbe aussi la structure des membranes, car les interactions électrostatiques et
’’hydrophile-hydrophobes ’’qui maintiennent normalement en place les lipides
membranaires sont perturbées (Kempf et Bremer, 1998).
Inversement, certains organismes possèdent des systèmes qui minimisent es
pertes d’eau ; à la différence des spores, qui sont presque totalement
déshydratées, les kystes ont une teneur en eau comparable à celle des cellules
végétatives. Le processus d’enkystement inclut la formation d’une paroi
cellulaire plus ou moins épaisse qui limite les échanges avec l’extérieur
(Singleton et Sainsbury, 1994). Les kystes, dont Azotobacter est l’exemple le
plus connu, sont généralement métaboliquement dormants et incapables de se
diviser. Cette distinction renvoie à la différence entre les notions d’évitement et
de tolérance. L’évitement regroupe tous les processus qui maintiennent la cellule
ou l’organisme en déséquilibre thermodynamique avec son milieu, par
l’établissement de barrières physiques qui isolent l’individu ou par une
exclusion constante du stress par une barrière chimique ou métabolique. La
tolérance est la capacité pour l’organisme, à l’équilibre thermodynamique avec
le stress, de ne pas en souffrir en prévenant, diminuant ou réparant les
dommages causés par le stress. La résistance de certains organismes implique à
la fois des mécanismes de tolérance et d’évitement, dont les contributions
respectives sont délicates à estimer. La situation du sable est différente de celle
d’un sol tempéré. Les bactéries sont en relation directe avec les particules
minérales, ou avec les rares particules riches en matière organique (Sugiyama et
Nikara, 2004).
Dans les systèmes naturels (les poussières atmosphériques étudiées par (Tong et
Lighthart, 1998) ou artificiels (les réseaux de distribution d’eau étudiées par
(Wu et al, 2005), les particules minérales protègent les bactéries associées des
effets délétères des radiations solaires ou des UV utilisés pour la désinfection.

Revues bibliographiques sur les peuplements bactériologiques du sable
12
De même, les associations avec les particules décrites par Lunsdorf et al, (2000)
créent des micro-habitats aux conditions physico-chimiques différentes
(plus favorables) de celles du sol « moyen ».
L’étude des interactions entre les bactéries et les minéraux dans les
environnements arides possède donc un double intérêt. Outre les quantifications
des activités métaboliques géochimiquement pertinentes, l’étude de la
participation des minéraux aux mécanismes de résistance des bactéries aux
stress de l’environnement prend dans les déserts tout son sens.
L’étude de la diversité d’une communauté bactérienne est nécessaire avant les
études fonctionnelles ou physiologiques. La diversité d’une communauté repose
sur trois points (Dunbar et al, 1999). La composition est l’inventaire des types
bactériens présents (au niveau taxonomique choisi), la richesse est le nombre de
types et la structure et l’abondance relative des différents types.
Les méthodes moléculaires permettent de mieux appréhender chacun de ces trois
points et de s’affranchir des lacunes des méthodes basées uniquement sur les
cultures pures (Amann et al, 1995).
Quelle que soit la complexité du système étudié, il est possible d’obtenir les
séquences des bactéries majoritaires. Les méthodes d’hybridation in situ par des
sondes oligonucléotides, basées sur les séquences des bactéries supposées
majoritaires, permettent d’estimer leurs effectifs et donc de réaliser une
estimation de la structure de la communauté. La structure peut être appréhendée
de plus en plus précisément par l’emploi successif de sondes de spécificité
croissante. Dans certains cas, ces méthodes d’hybridation permettent même
d’obtenir des informations morphologiques ou physiologiques (comme le
contenu cellulaire en ARN, qui renseigne sur le taux de croissance et l’activité
des bactéries). Les approches basées sur la mise en culture des micro-
organismes, malgré les limites évoquées plus haut, conservent un fort intérêt
écologique (Garland et al, 2001).

Revues bibliographiques sur les peuplements bactériologiques du sable
13
Garland et al, (2001), considèrent que les bactéries « cultivables » sont celles
qui sont capables de coloniser rapidement un milieu peu contraignant. La
proportion de ces bactéries « opportunistes » dans la flore totale serait donc un
bon indicateur de l’état, par rapport à la succession écologique, des
communautés.
Malgré les avancées récentes permises par ces méthodes, la diversité absolue
des bactéries est inconnue et souvent considérée comme hors de portée
(Curtis et al, 2002, Cases et Lorenzo, 2002), et ce quelle que soit l’échelle
ou l’environnement. Il est généralement impossible de réaliser un inventaire
exhaustif des types bactériens présents, mais les méthodes statistiques utilisant
des modèles de structure de communauté permettent d’extrapoler les
abondances des types majoritaires pour estimer le nombre de taxa présents.
Les études synthétisées par Torsvik et al (2002), ont été menées dans des
environnements très divers, autant « naturels » (tempérés comme les sols
forestiers, sédiments marins ou extrêmes comme des bassins saumâtres saturés
en sel) que perturbés par les activités humaines (sols agricoles, ou zones
marines à forte pollution organique). En analysant l’hybridation des ADN
extraits.
Curtis et al, (2002), étudiant les relations entre la diversité locale et la diversité
globale, émettent aussi l’hypothèse que la diversité bactérienne globale est
constituée d’un nombre relativement faible de taxa ubiquitaires.
Une estimation quelque peu précise de l’abondance des deux ou trois taxa
majoritaires améliorerait significativement notre connaissance de la diversité
globale. Elle fournirait aussi un test très intéressant de l’hypothèse log-normale
de distribution des espèces bactériennes, qui constitue la base de la plupart des
modélisations mathématiques en écologie microbienne (Curtis et al, 2002).
Zhou et al, (2002), ont étudié différents sols et leurs horizons contenant
différentes teneurs en eau et en matière organique. Un sol de surface, non saturé
en eau et pauvre en carbone, a montré une communauté microbienne très

Revues bibliographiques sur les peuplements bactériologiques du sable
14
uniforme, au sein de laquelle toutes les espèces étaient également abondantes.
L’horizon inférieur du même sol, lui aussi pauvre en carbone mais saturé en eau,
montrait une distribution spécifique plus classique, dominée par quelques taxa.
On considère très généralement que la dominance est le résultat d’interactions
compétitives (Zhou et al, 2002).
Les conséquences liées aux déplacements bactériens dans un profil sol
concernent de nombreuses thématiques environnementales, agronomiques et de
santé. Aussi l’intérêt pour cette problématique est grandissant car les
perspectives d’application sont nombreuses. Ainsi certaines souches sont
spécialisées dans la dégradation de polluants (biodégradation), d’autres
capables de rééquilibrer un écosystème ou encore d’apporter des sources d’azote
à des plantes de culture. L’apport et le transfert de ces microorganismes dans un
sol pourraient donc servir à la protection de l’environnement. (Walker et al,
2004).
Cependant les déplacements de microbes dans un sol peuvent aussi être
nuisibles. Le transfert de bactéries pathogènes, exogènes ou génétiquement
modifiées dans la zone non saturée peut aboutir à la contamination de nappes
phréatiques ou à la perturbation d’un écosystème. De plus les bactéries peuvent
être vecteurs pour le transport de polluants tel que des métaux lourds, des
éléments radioactifs ou des pesticides. De nombreux déchets pouvant contenir
des concentrations élevées en bactéries entériques pathogènes sont
régulièrement répandu à la surface de sols. Des études récentes ont montré que
le sol pouvait jouer un rôle important dans la transmission de maladies
entériques. Dans les pays en voie de développement. (Grasso et al, 1996)
Zhou et al, (2002). Proposent quatre facteurs susceptibles d’entraîner la
répartition uniforme qu’ils ont observée. Une grande abondance de ressources
réduit la compétition, permettant l’établissement d’une grande diversité.
L’existence de la ressource sous différentes formes peut entraîner une
spécialisation des espèces, réduisant la compétition. La séparation spatiale des

Revues bibliographiques sur les peuplements bactériologiques du sable
15
ressources peut résulter en une séparation physique des populations, évitant
encore la compétition. Des conditions environnementales fluctuantes peuvent
enfin entretenir une situation hors-équilibre avec des populations plus diverses.
Les études de la diversité bactérienne dans les zones arides sont plus
fréquemment menées dans des zones où un couvert végétal, même très clairs
existe, que dans des zones véritablement dépourvues de végétation. Elles
comparent souvent les caractéristiques de la communauté microbienne du sol
près ou loin des plantes (Zhou et al, 2002).
Kuske et al, (2002), ont comparé les rhizosphères de trois plantes du plateau
aride du Colorado et les espaces sans végétation mais portant une croûte
microbienne (à trois profondeurs différentes, de la surface du sol à 30 cm). Deux
plantes étaient autochtones typiques et la troisième était une espèce invasive. La
quantité d’ADN extraite des sols rhizosphériques, et donc la biomasse
bactérienne, était supérieure à celle des espaces inter-plantes. Le concept d’«
îlots de fertilité » correspond dans les environnements arides, où les nutriments
sont peu abondants et répartis inégalement, à la plus nombreuse population
microbienne dans la rhizosphère des plantes. Les microorganismes favorisent la
croissance des plantes en fixant l’azote, limitant des pathogènes et favorisant la
dissolution des minéraux. Mais la notion souvent admise d’« ˆilots de fertilité »
était infirmée dans l’étude de Kuske et al, (2002) par des quantités de carbone
organique du sol et un nombre de bactéries hétérotrophes cultivables partout
similaires. Toutefois, les structures des communautés bactériennes étaient
notablement différentes.
L’étude plus précise du groupe des Acidobacteria, très divers et connus dans de
nombreux types de sols et environnements dans le monde entier, révélait des
différences plus marquées encore. Le rôle fonctionnel de ces bactéries dans les
L’´etude plus précise du groupe des Acidobacteria, très divers et connus dans de
nombreux types de sols et environnements dans le monde entier, révélait des
différences plus marquées encore. Le rôle fonctionnel de ces bactéries dans les

Revues bibliographiques sur les peuplements bactériologiques du sable
16
sols n’est pas connu, mais cette étude a démontré que différents membres de
cette division occupent différentes niches écologiques et seraient donc très
intéressants à suivre dans la perspective de changements de type de couvert
végétal liés aux modifications de l’environnement (Kuske et al, 1997).
Bird et al, (2002) ont étudié des sols arides du Nouveau-Mexique, constitués de
patches de végétation avec des espaces qui les séparent, mais n’ont pas observé
de teneurs uniformes (Kuske et al, 1997) en matière organique dans le sol
rhizosphérique et entre les plantes. Ils en ont déduit que la teneur en carbone, en
est très variable, tant à l’échelle du patch de végétation qu’à l’échelle du
paysage. Cette conclusion est en accord avec l’hypothèse des « ilots de fertilité »
Dunbar et al, (1999), ont étudié la rhizosphère de pins pignons et les espaces
entre les arbres sur deux sites dans une région aride boisée du nord de l’Arizona,
soit quatre échantillons. Le champ de cendres vieux de 900 ans d’un volcan
éteint, constitue un des sites étudiés. L’autre est un sol sableux typique de la
région. Les deux sites connaissent le même régime de précipitations, mais le sol
cendreux est plus grossier et mieux drainé. De plus, sa couleur très sombre crée
un environnement plus chaud que le sol sableux (sécheresse édaphique, (Kuske
et al, 1997). Contrairement au sol sableux qui porte quelques herbes, les espaces
entre les arbres ne portent pas de végétation sur le sol cendreux. Les auteurs ont
comparé la diversité bactérienne par séquençage des gènes, tant sur des isolats
bactériens cultivées qu’après extraction de l’ADN du sol et clonage (Kuske et
al, 1997).
L’étude de l’altération des minéraux en milieux arides est motivée par le fait que
de très nombreuses découvertes de météorites ont lieu dans les déserts, chauds
ou froids ( Aguilera et al, 1999).
Ces découvertes ayant généralement lieu longtemps après la chute, l’altération
terrestre est susceptible de brouiller le message chimique porté par les
météorites ; De nombreux travaux sont en cours, qui visent à déterminer les

Revues bibliographiques sur les peuplements bactériologiques du sable
17
modes de résistance des bactéries aux stress de l’environnement (notamment la
dessiccation et les rayonnements solaires) (Aguilera et al, 1999).
La connaissance des conditions de vie des micro-organismes vise aussi à estimer
la participation des déserts aux cycles géochimiques à la surface de la Terre.
3. Evaluation de la pollution microbienne en méditerranée
IL existe deux sortes principales d'exposition humaine aux polluants microbiens en
Méditerranée ; la consommation de poissons et crustacés contaminés et le contact
direct avec les agents contaminateurs au cours de la baignade, dans ce dernier cas par
ingestion d'eau de mer. On peut illustrer l'ampleur du danger potentiel crée par cette
exposition en mentionnant qu'environ 100 millions de personnes vivent de façon
permanente sur les côtes de la Méditerranée et à peu près autant s'y rendent tous les
ans dans un but de plaisance (GFCM, 1983).
La pollution microbienne est un des résultats directs du déversement d'eaux usées non
traitées dans le milieu marin. Dans la région méditerranéenne, plus de 90% des
déchets municipaux sont déversés à l'état brut (PNUE/FAO/OMS/AIEA, 1990).
L'importance d'assurer une qualité convenable d'eau de mer est rendue plus aigue
par les faits que l'un des charmes les plus populaires des loisirs le long de la côte
méditerranéenne est la plage destinée à la baignade et que, puisque les températures
sont relativement élevées, les baigneurs demeurent plus longtemps dans l'eau qu'ils ne
le feraient normalement dans les climats tempérés, donnant donc une période
d'exposition plus longue à une contamination possible (UNEP/IOC/IAEA/FAO,
1990).
3.1. Sort de la pollution microbienne en Méditerranée
Les sources de pollution bactérienne sont nombreuses, et les germes proviennent
généralement de :
Rejets urbains : ces germes sont issus de l’épuration domestique et industrielle ;
lorsque certaines stations négligent de les traiter ; lorsqu’il n y a pas
d’assainissement, que les réseaux sont défaillants ou lorsque la capacité

Revues bibliographiques sur les peuplements bactériologiques du sable
18
d’assainissement est dépassée, en période estivale notamment par exemple, la
capacité totale des installations d’épuration déjà réalisées en Algérie représente
environ 18,3% du besoin national (Bentir, 1996).
Trois groupes d'indicateurs de pollution fécale ont servi essentiellement de base à
l'évaluation de la pollution microbienne en mer Méditerranée ; les Coliformes
totaux, les Coliformes fécaux et les Streptocoques fécaux. rien qu'ils ne
répondent pas à toutes les exigences d'un "indicateur idéal", ils sont en général
considérés et utilisés comme des indicateurs acceptables pour déterminer la qualité
sanitaire des eaux à usage récréatif (OMS/PNUE, 1990 )
L'eau de mer n'est pas le milieu naturel de la plupart des microorganismes
déversés dans les effluents d'eaux usées, en particulier ceux provenant des voies
intestinales de l'homme ou d'autres animaux à sang chaud. Donc, on peut s'attendre à
ce que les trois indicateurs microbiens cités au paragraphe ci-dessus ne demeurent
pas inchangés dans les eaux de mer réceptrices mais plutôt disparaissent au fur et à
mesure (OMS/PNUE, 1979).
La salinité, la lumière naturelle, la température, les substances dissoutes et les
prédateurs naturels comptent au nombre des facteurs connus affectant la survie de ces
microorganismes dans l'eau de mer. Plus particulièrement, on a démontré que le
rayonnement solaire est un des seuls facteurs importants responsable de l'inactivation
microbienne (Prado et al, 1994).
Les résultats disponibles de Natsch et al ( 1996) montrent que les lésions sublétales
provoquées par le rayonnement solaire dans le système enzymatique de catalase
d’Escherichia coli rendent les cellules sensibles à des concentrations de peroxyde,
inoffensives dans le cas contraire. Bien que les techniques de culture normales ne
permettent pas de retrouver toutes les cellules microbiennes agressées, l'addition de
cellules errantes de peroxyde et particulièrement l'enzyme de catalase lui-même,
permet de retrouver une partie considérable des cellules d’Escherichia coli
affectées. Cependant, si l'on considère le haut niveau d'enzyme supplémentaire
nécessaire dans les cultures de laboratoire pour retrouver les cellules ayant subi des

Revues bibliographiques sur les peuplements bactériologiques du sable
19
lésions, on peut s'attendre à ce que ce processus d'enrichissement ne se produise pas
dans des conditions naturelles et par conséquent que la plupart des organismes
ayant subi des lésions par la lumière du soleil ne puissent pas survivre, étant
ainsi perdus de façon permanente après un certain temps.
Les résultats d'études in situ, menées au cours de (PNUE / OMS, 1979) et ailleurs,
ont démontré les différents modèles de survie des trois indicateurs microbiens.
Alors que les Coliformes totaux et les Coliformes fécaux semblent être inactivés
dans l'eau de mer assez rapidement et progressivement dans des conditions
naturelles, les Streptocoques montrent une vitesse d'inactivation plus faible.
La floculation des cellules microbiennes et leur sédimentation consécutive au
fond de la mer sont considérées comme le mécanisme responsable de
l'enrichissement microbien des sédiments dans les zones situées autour des
déversements d'eaux usées (Mitchell et Chamberlain, 1975).
Les remous naturels et les courants de mer peuvent devenir un mécanisme plausible
par lequel les sédiments contaminés peuvent être resuspendus, avec consécutivement
un endommagement de la qualité microbienne de l'eau de mer sus-jacente
(Volterra et Aulicino, 1980).
Cependant, le fond de la mer n'est pas le milieu naturel de la plupart des
microorganismes apportés par les effluents d'eaux usées; on peut donc s'attendre à
ce qu'en mettant fin aux déversements et en les améliorant, avec la déplétion des
substrats organiques qui s'ensuit, la survie de ces micro-organismes soit fortement
compromises (Volterra et Aulicino, 1980).
De façon semblable à ce qui se produit dans l'eau de mer, des résultats d'études
pratiques ont démontré que les Streptocoques fécaux peuvent survivre plus longtemps
que les Coliformes fécaux jusqu'à les dépasser en nombre, contrairement à ce que
l'on observe normalement dans les effluents d'eaux usées municipales non
t ra i t ées (Volterra et Aulicino, 1980).
Tous les résultats précédents permettent de soutenir fortement l'inclusion des
Streptocoques fécaux comme indicateur de la pollution fécale en plus des

Revues bibliographiques sur les peuplements bactériologiques du sable
20
Coliformes fécaux et totaux afin de les utiliser dans les programmes de
surveillance continue courants. Les connaissances actuelles soutiennent l'utilisation
tant des Coliformes fécaux que des Streptocoques fécaux comme paramètres de
routine pour la surveillance de la qualité des eaux côtières parce qu'ils sont
importants comme indicateur individuel et aussi parce qu'ils fournissent des
renseignements supplémentaires de valeur lorsqu'on les compare quant à
l'origine et au temps de séjour dans les eaux de mer des effluents d'eaux usées
(Geldreich, 1976).
4. Les polluants hydriques
Les eaux usées domestiques non traitées sont utilisées à des fins d’irrigation en
agriculture. Cette méthode est une source de prolifération de souches pathogènes
dans les sols et de contamination des nappes phréatiques, des puits ou des eaux
de récréations. Mais cette pratique commence également à se répandre dans les
pays développés notamment dans les régions souffrant de déficits hydriques
importants. Par exemple dans l’Arizona de nombreux parcs municipaux sont
irrigués avec des eaux usées. Les données sur le déplacement de souches
pathogènes dans un profil sol sont encore rares dans la littérature. La possibilité
que le sol puisse être un réservoir et un lieu de transport de bactéries pathogènes
n’a été que peu étudiée. Il semble logique de penser que des bactéries apportées
à un sol peuvent atteindre les nappes phréatiques grâce à la circulation de l’eau
dans les pores du sol. Cependant le lien entre l’origine des maladies entériques
et le sol n’a pas encore été parfaitement établi.
(Choi et al, 2007)
En 1980 on estimait que 2 milliards d’hommes dans le monde n’avaient pas
accès à un point d’eau potable, pendant cette même période estimait que 80 des
maladies sur la planète sont transmises par les eaux contaminées par les
polluants chimiques et organiques (O.M.S, 1987).

Revues bibliographiques sur les peuplements bactériologiques du sable
21
Certaines espèces bactériennes normalement absentes dans l’intestin d’une
personne en bonne santé, peuvent être sécrétées de façon intermittente et en
quantités variables selon le lieu et l’état de santé de la personne, ces bactéries
pathogènes, ou potentiellement pathogènes, sont responsables de la plupart des
maladies infectieuses qui sévissent en Afrique subtropicale : choléra, fièvre
typhoïde, dysenterie, gastro-entérite, maladies diarrhéiques (Redman et al,
2002).
Généralement transmises à l’homme par voie digestive liée à la consommation
d’eau ou d’aliments contaminés, les bactéries pathogènes jouent un rôle
déterminant dans la pollution biologique de la nappe phréatique à partir d’une
latrine (Li et al, 2007).
Les bactéries pathogènes ne sont pas toujours omniprésentes dans les matières
fécales contrairement aux bactéries indicatrices de la pollution fécale. (Redman
et al, 2002)
5. La flore des sables de plage
5.1 Micro-organismes d’indication fécale
Ils sont des microorganismes non pathogènes employés pour indiquer le degré
de la contamination fécale. Ils sont généralement présents dans des nombres
bien plus grands que les microorganismes pathogènes et il est facile de les isoler,
identifier et énumérer (Reasoner et Geldreich, 1985).
Les microorganismes d’indication fécale incluent des Coliformes (Coliformes
totales, Coliformes thermotolérants et Escherichia coli), des entérocoques
intestinaux, des bactériophages et Clostridium (Collins et al, 1994).
La présence des Coliformes totales, des Coliformes thermotolérants, de
l'Escherichia coli et d'entérocoques en sable de plage et le rapport entre leurs
nombres en sable de plage et leurs nombres dans les eaux adjacentes ont
comporté un domaine de recherche significatif, avec des résultats apparent
contradictoires (Koneman et al, 1997).

Revues bibliographiques sur les peuplements bactériologiques du sable
22
Des Coliformes totaux, des Coliformes thermotolérants et des entérocoques
intestinaux ont été isolés dans les échantillons extérieurs de sable à Marseille et
Agde en France. Le nombre d'entérocoques intestinaux, provenant
probablement des animaux, étaient plus haut que le nombre d'autres indicateurs
(Conseil Supérieur d' Hygiène Publique de France, 1990).
Des nombres très élevés des Coliformes thermotolérants et d’entérocoques
intestinaux étaient Isolés en sable de plage le long des eaux côtières de Tarente
en Italie (Signorile et al, 1992).
Des nombres plus peu élevés des microorganismes d’indication fécale ont été
enregistrés dans des secteurs de natation à Avive, Israël, et à Barcelone,
l'Espagne (Figueras et autres, 1992 ; Ghinsberg et al, 1994).
Des nombres peu élevés de ces indicateurs ont été récupérés en sable sec d'une
plage le long de la côte de Thyrrenian (Italie). Escherichia coli a été récupérée
dans 61% des échantillons et les entérocoques ont dépassé des Coliformes en
nombre (Bonadonna et al, 2002).
Dans une étude italienne, une corrélation significative a été trouvée entre la
contamination de plages et la contamination des eaux de mer adjacentes, bien
que le sable ait généralement eu des recensements de bactéries plus élevés que
l'eau (Aulicino et al, 1985).
Une tendance semblable a été trouvée aux plages de Barcelone ; contrairement à
l'étude italienne, cependant, le niveau de la contamination n'était pas
sensiblement différent entre le sable et l'eau de mer (Roses Codinachs et al,
1988).
Papadakis et al, (1997) n'ont trouvé aucune corrélation entre les indicateurs de la
pollution fécale comptés sur la partie humide de la plage et les comptes de
Staphylococcus aureus ou présence de mycètes. Une corrélation statistiquement
significative a été détectée entre les levures et les moules, l'Escherichia coli, les
entérocoques et les spores de Clostridium sulfitoréducteur et entre les spores et

Revues bibliographiques sur les peuplements bactériologiques du sable
23
des Staphylocoques dans une recherche sur les sables secs et humides en Italie
(Bonadonna et al, 2002).
Dans une étude épidémiologique effectuée sur deux plages à Malaga, l'Espagne,
des microorganismes d’indication fécale, particulièrement, les coliphages, ont
été fortement et sensiblement corrélés avec les champignons dermatophytes
(mycètes microscopiques qui se développent sur la peau et les muqueuses) sur
une des deux plages. Seulement E. coli a montré une corrélation significative
avec des Candida albicans (une mycète pathogène).
À l'autre plage, les entérocoques intestinaux ont montré la meilleure corrélation
avec des champignons dermatophytes. Encore, les coliphages présentaient une
meilleure corrélation avec des Candida albicans. (Borrego et al, 1991).
5.2. Staphylocoques
Selon quelques études, les espèces de Staphylocoque prédominent au-dessus
des autres flores dans le sable (Dowidart et Abdel-Monem, 1990). D'un total de
85 contraintes de coques gram positif isolés dans l'eau et le sable de plage
situés à deux plages populaires dans le Chili, dont 31% ont été classifiés comme
des Staphylocoques épidermite, 9% comme S.haemolyticus, 24% comme
Staphylocoques doré et 36% comme autres espèces de Staphylocoque (Prado et
al, 1994). L'origine des Staphylocoques en sable de plage est attribuée à
l'activité humaine. Son nombre est corrélé avec le nombre de nageurs sur la
plage, et les nombres de Staphylocoques dorés montrent une corrélation avec la
présence des levures d'origine humaine dans des échantillons de sable
(Papadakis et al, 1997).
De plus, de grandes quantités de Staphylocoques doré ont été récupérées du
sable et de l'eau en été, quand il y avait une densité plus élevée de nageurs sur la
plage, qu'en hiver (Ghebremedhin et al, 2008).

Revues bibliographiques sur les peuplements bactériologiques du sable
24
En outre, de plus grandes quantités de Staphylocoques doré ont été récupérées
du sable en le comparant avec les échantillons d'eau (Ghinsberg et al, 1994 ;
Papadakis et al ; 1997).
Les investigations effectuées le long de la côte tyrrhénienne ont montré des
densités plus élevées des espèces de Staphylocoque en sable des secteurs
caractérisés par des brise-lames qu'en sables trouvés dans des terrains
découverts. Les Staphylocoques épidermites, était les prédominantes
(Bonadonna et al, 1993).
5.3. Pseudomonas aeruginosa
Dans une étude en Israël, l'eau de mer et le sable sur un certain nombre de
plages, a contenu de divers niveaux de Pseudomonas aeruginosa. L'isolement
de Pseudomonas aeruginosa. et d'autres espèces de Pseudomonas était
proportionnellement plus haut en sable que dans des échantillons d'eau de mer
(Ghinsberg et al, 1994). Les Pseudomonas aeruginosa ont été isolées dans les
plages sablonneuses au Portugal dans des conditions diverses de marée,
(Mendes et al, 1993).
5.4. Vibrio
Des isolats de Vibrio parahaemolyticus ont été trouvés dans l'eau marine ou
saumâtre et les spécimens rassemblés des échantillons de sable en Afrique
(Aldova, 1989). Vibrio harvey a été isolé dans l'eau de bord de la mer et dans
les échantillons de sable rassemblés sur la plage (Aldova, 1989).
5.5. Bactéries entériques
Ce sont des espèces de bactéries qui peuvent causer la gastroentérite, elles ont
été isolées dans des échantillons de sable. Cependant, leur présence ne constitue
aucune menace apparente de santé. Les sables des plages au Portugal ont
contenu des valeurs semblables de Clostridium perfringens dans de diverses
conditions de marée (Mendes et al, 1993).

Revues bibliographiques sur les peuplements bactériologiques du sable
25
Bonadonna et al (1993) ont suggéré que C. perfringens pourrait être un bon
indicateur de contamination fécale en sable. Des niveaux bas de
Campylobacter jejuni ont été enregistrés dans les eaux côtières et le sable sur
des plages israéliennes, le sable de plage a contenu une plus grande quantité de
bactéries entériques que le rivage adjacent (Ghinsberg et al, 1994).
Au Royaume-Uni, sur la zone intertidale, les sédiments ont semblé servir de
réservoir substantiel aux campylobactéries thermophiles, qui pourraient
contribuer de manière significative aux nombres bactériens en eaux de surface
(Obiri-Danso et Jones, 1997).
Dabrowski (1982) a isolé des espèces de Shigella à partir des échantillons de
sable et d'eau sur une plage dans le compartiment de Danzig (Pologne).
5.6. Mycètes
Mycètes qui sont souvent trouvés dans l'environnement pendant que les
saprophytes peuvent agir en tant que microbes pathogènes opportunistes (Hoog
et al, 2000).
Les études par Soussa (1990) dans les régions côtières centrales portugaises ont
montré la présence des dermatophytes dans 42% des plages de sable analysées.
Les plus communs étaient les Trichophyton mentagrophytes, le T. rubrum et le
Microsporum Nanum.
Toutes ces espèces sont associées aux infections de la peau, avec des T.
mentagrophytes étant l'agent le plus commun du dermatomycosis en Europe et
T. rubrum L'agent le plus commun dans le monde entier (Hoog et autres, 2000).
5.6.1. Mycètes saprophytiques
Les Candida et Aspergille (A. ochraceus et A. fumigatus) ont été isolés dans
les secteurs inondés et intermédiaires en conditions de marée élevées (Izquierdo
et al, 1986).
Des Candida albicans et d'autres espèces de Candida ont été isolés dans des
plages de sable dans les sud de la France (Bernard et al, 1988). Dans la même

Revues bibliographiques sur les peuplements bactériologiques du sable
26
étude, 8 mycètes keratinophiliques (c'est-à-dire, ceux capables se développer
sur la kératine, un terrain communal caractéristique aux dermatophytes) et 11
espèces non-keratinophiliques, qui sont des microorganismes potentiellement
pathogènes, ont été isolés.
Izquierdo et al (1986) ont isolé 16 espèces de mycètes dans le sable de plage le
long du nord-est de la côte méditerranéenne de l'Espagne, dont quelques
espèces sont potentiellement pathogènes. La plupart des espèces ont appartenu
aux genres Penicillium, Aspergille et Cladosporium.
En Israël, Ghinsberg et al (1994) ont isolé des mycètes dans tous les
échantillons de sable de plage, mais pas dans des échantillons d'eau de mer.
Dans une étude en Guadeloupe, Boiron et al (1983) ont étudié des espèces
fongiques en sable d'eau de mer et de bord de la mer, concluant que la similitude
des espèces bactériennes en sable et eau de mer, en même temps que le fait
qu'aucune Candida albicans n'a été isolée, a corroboré leur hypothèse que les
levures isolées étaient d'origine marine. Les mycètes isolées ont appartenu aux
espèces C. tropicalis, C. parapsilosis, C. langeronii, C. guilliermondii,
Trichosporon cutaneum et espèces de Thorulopsis. Plus souvent les genres
isolés dans les échantillons de sable de plage dans une étude espagnole étaient
Penicillium, Aspergillus, Cladosporium, Altenaria, Mucor, Monilia,
Cephalosporium, Verticillium et Chrysosporium (Roses Codinachs et al, 1988).
Absence ou présence limitée de C. albicans a été également enregistrée par
d'autres chercheurs (Roses Codinachs etl, 1988 ; Figueras et al, 1992).
La densité fongique de 180 échantillons de sable rassemblés de 42 plages
espagnoles méditerranéennes a atteint plusieurs centaines de mille de colonies
formants unités par gramme d'échantillon. Les genres les plus isolés
généralement étaient Penicillium, Cladosporium, Aspergillus, Acremonium,
Altenaria et Fusarium (Larrondo et Calvo, 1989). Dans une étude effectuée dans
la région d'Attica de la Grèce, les isolats fongiques ont inclus des Candida

Revues bibliographiques sur les peuplements bactériologiques du sable
27
albicans, C. krusei, C. tropicalis, C. puilliermondi, C. rugosa, Pitirosporum.
orbiculare, Fusarium, Penicillium, Mucor, Helminthosporium et Aspergillus
(Papadakis et al, 1997), dont un certain nombre d’espèces est pathogènes
(Hoog et al, 2000).
5.7. Virus et parasites
Très peu d’informations existent au sujet de la présence de virus et de parasites
en sable de plage. Dans une étude de trois ans en Roumanie par Nestor et al
(1984), la présence des entérovirus dépendait de la saison, étant présents en
sable de l'eau et de plage pendant des saisons de non-vacances.
Dans une étude de deux plages de sable à Marseille, France, Toxocara canis
s'est avéré le parasite le plus commun, étant présent en moyenne dans 150 g du
sable (Conseil Supérieur d' Hygiène Publique de France, 1990). Cependant, dans
une étude effectuée sur le sable à Perth, Australie, un total de 266 échantillons
n'a montré aucune trace d’œufs de Toxocara canis ou d'autres œufs ou larves de
parasites nématodes (Dunsmore et al, 1984).
5.8. Composition du sable en agents pathogènes
L’un des risques majeurs sur la santé humaine liés aux déchets est sans doute
leur contamination microbiologique par divers agents pathogènes tels que les
bactéries, les protozoaires, les virus et autres. Le suivi de certains paramètres
microbiologiques dans le compost, comme l’Aspergillus fumugatus par exemple,
permet de déterminer rapidement son état sanitaire ; et il est démontré que la
présence d’une grande quantité de moisissures implique automatiquement la
présence d’autres agents pathogènes (EPA, 2002).
D’autre part, il est important de mettre en relief cette caractéristique pour qu’elle
puisse être prise en compte dans d’éventuelles mises en place de programme de
valorisation et de recyclage des rejets atténuant ainsi leur impact sur la
santé(EPA, 2002) .

Revues bibliographiques sur les peuplements bactériologiques du sable
28
Elle peut servir aussi à la sensibilisation des personnes en contact direct avec les
déchets et qui sont le plus souvent non protégées aussi bien dans les pays
industrialisés que dans les autres pays. Hassen et al, (2001) ont identifié
plusieurs microorganismes présents en nombre important dans les déchets au
cours du compostage (spores bactériens, Coliformes fécaux, Escherichia coli,
Streptocoques fécaux, Staphylocoques, Salmonelles et Shiguelles). D’après
Hoornweg et al. (2000), ces différents agents pathogènes trouvés dans les
déchets sont d’origine humaine ou animale et peuvent provenir des boues de
vidange, des couches-culottes ou des déchets des animaux domestiques.
6. Dispersion et destin des microorganismes en sable de plage
La croissance des micro-organismes en sable de plage est limitée par les
éléments nutritifs. Les études de laboratoire ont prouvé que les aliments
traversent la communauté bactérienne (Khiyama et Makemson, 1973). Autres
études ont prouvé que la contamination microbienne est plus haute en sable que
dans les eaux ; le sable se comporte comme port passif pour la pollution
cumulative (Oliveira et Mendes, 1993; Oshiro et Fujioka, 1995).
On a trouvé des niveaux plus élevés de Coliformes, d’Escherichia coli et
d’entérocoques en sable que dans l’eau dans le compartiment de Hanauma
(Hawaï) provenus de falaises entourant le compartiment (Oshiro et Fujioka,
1995). La matière fécale des pigeons et de mangoustes étaient également et
vraisemblablement une source de contamination de sable de plage.
Cette étude a conclu que le sable souillé pourrait être la source principale des
niveaux élevés de bactéries dans l'eau. La contamination de sable est fortement
variable, rendant l'interprétation des résultats difficile (Aubert et al, 1987 ;
Figueras et al, 1992 ; Oshiro et Fujioka, 1995).
La survie des bactéries entériques sur la surface du sable sec peut être de courte
durée, ces bactéries sont détruites la plupart du temps, par la pression
environnementale. Le sable humide, le secteur où les enfants passent

Revues bibliographiques sur les peuplements bactériologiques du sable
29
typiquement la majeure partie de leur temps sur la plage, est le plus approprié à
la survie des bactéries. Le sable humide, enrichi avec les substances organiques,
fournit un environnement favorable pour les bactéries entériques et qui leur
permet de survivre plus longtemps qu'en eau de mer (Papadakis et al, 1997).
Les divers facteurs qui favorisent la survie et la dispersion de microorganismes
pathogènes et les indicateurs de contamination fécale sur le sable de plage sont
multiples, notamment la nature de la plage, les phénomènes de marée, les
sorties d'eaux d'égout, la saison, la présence des animaux et le nombre de
baigneurs.
Obiri-Danso et Jones (1997) ont analysé des échantillons de sédiments dans le
Royaume uni, pour une étude sur les Campylobactéries thermophiles et les
microorganismes, indicateurs de contamination fécale dans différentes
conditions de marrée sur une période de douze mois. 53% des échantillons
étaient positifs pour les Campylobactéries avant couverture de marée ; ce
chiffre était sensiblement plus bas que les 64% récupérés après disposition de
marée. Cependant, il n'y avait aucune différence significative dans les nombres
d'indicateurs de contamination fécale en ce qui concerne les échantillons
prélevés avant ou après la couverture de marée. Dans la même étude, on a
observé une variation saisonnière dans les Campylobactéries, avec le taux
d'isolement le plus élevé en hiver (100%), suivi des crêtes secondaires au
printemps (33-67%) et en automne (67-78%). Les plus basses valeurs ont été
trouvées en été, qui sont corrélées avec la présence des Campylobactéries en
eaux de surface.
En revanche Mendes et al (1993) ont étudié l'influence des marées sur le
nombre de microorganismes pathogènes et les indicateurs de contamination
fécale en sable sans trouver aucune différence claire.
Nestor et al (1984) ont constaté que la présence de quelques germes
pathogènes dépendait de la saison, les virus ne sont pas présentés en eau de
mer et le sable des plages en dehors de la saison des vacances.

Revues bibliographiques sur les peuplements bactériologiques du sable
30
Borrego et al (1991) ont rapporté des recensements des bactéries plus élevés et
de longue survie en plages près des sorties d'eaux d'égout.
Conformément à la section précédente, des mycètes sont souvent produites en
sable, et leur survie est plus longue que celle des bactéries entériques, cela dues
à leur capacité de former les spores résistantes. On a suggéré que la présence
des mycètes indique une contamination indirecte provenant des résidus ou
détritus des utilisateurs de plage et/ou de l'influence de marée (Mendes et al,
1998).
Dans une étude in vitro, Anderson (1979) a constaté que quatre mycètes
pathogènes (Trichosporon cutaneum , les Candida albicans, Microsporum
gypseum et Trichophyton mentagrophytus ) ont survécu pour au moins 1 mois
en sable non stérile inoculé avec des propagules de tels mycètes. Dans une étude
semblable, les espèces des dermatophytes (Epidermophyton floccosum
,Microsporum canis , M. gypseum, Trichophyton mentagrophytes et T. rubrum)
Et des Scopulariopsis brevicaulis ont survécu pendant une période qui varie
entre 25 et 360 jours (Carillo-Muñoz et al, 1990).
Les secteurs d’eau intensivement utilisés présentent des moyens de la
transmission microbienne avec préavis des germes pathogènes (par exemple,
dermatophytes). La transmission peut se produire parce que les individus ont
jeté des microbes pathogènes sur le sable, par le contact direct ou par des autres
moyens, bien que, excepté la transmission par l'intermédiaire de l'eau polluée,
aucun de ces derniers n’a été franchement démontré (Aubert et al, 1987).
Papadakis et al (1997) ont rassemblé des échantillons d’eau et de sable de deux
plages dont l’un est plus populaires que l’autre en l'été et en hiver, et les
nombres de nageurs présents sur les plages ont été comptés. Des Coliformes
totaux, des Coliformes thermotolérants, des entérocoques, des Staphylocoques
doré, des levures et des moules ont été également étudiés. Les échantillons de
l'eau présentaient des valeurs basses que celles notées sur sable étaient pour les
indicateurs de contamination fécale. Des espèces de levures étaient présentes en

Revues bibliographiques sur les peuplements bactériologiques du sable
31
eau et en sable des deux échantillons provenant des deux emplacements.
Staphylocoques doré a été isolée dans des échantillons de l'eau et de sable
seulement deux fois en hiver. Une corrélation significative apparue entre les
nombres de nageur sur les plages et le nombre de Staphylocoques doré dans les
échantillons d’eau, la corrélation étant plus prononcée sur la plage la plus
populaire.
Dans des échantillons de sable, les nombres de S.doré Se sont corrélés avec le
nombre de nageurs sur la plage, donc une moyenne plus élevée dans la plage la
plus populaire, les mêmes observations ont été notées pour les levures.
L'évidence épidémiologique pour des risques sanitaires liés à l’exposition aux
plages sablonneuses n'a pas été trouvée. Les études épidémiologiques ont visé à
étudier les causes-effet ou à examiner un rapport possible de réponse à des doses
données liant la qualité microbienne de la plage de sable avec la peau, l'œil,
l'oreille et les symptômes gastro-intestinaux. (Chabasse et al, 1986 ; Conseil
Supérieur d’Hygiène Publique de France, 1990).
La surveillance systématique de plage en tant qu'élément de la lutte contre la
pollution est une surveillance relativement limitée, et généralement non
justifiée. Cependant, on l’a souvent recommandée pour la recherche.
WHO/UNEP (1992, 1994) a indiqué que le sable et les sédiments humides de
plage devraient être une partie d'études épidémiologiques et microbiologiques
corrélant la qualité de l'eau avec les effets sur la santé, mais l'évidence indique
jusqu'ici que le sable de plage ne semble pas constituer un risque infectieux
(Chabasse et al, 1986 ; Conseil Supérieur d' Hygiène Publique de France, 1990).
Dans quelques pays, le nettoyage mécanique de sable est à la pratique commune
qui peut éliminer les déchets évidents qui se mélangent au sable, et qui réduit
une quantité de matière organique et donc de réduit le développement ultérieur
de microorganismes (Bartram et Rees, 2000).
Cependant, le nettoyage mécanique peut toucher à l'écologie de sable (Llewellyn
et Shackley, 1996). Les recherches qui ont étudié la qualité microbiologique du

Revues bibliographiques sur les peuplements bactériologiques du sable
32
sable ont montré à cela une amélioration claire qui a été réalisé en raison
d'élever les niveaux généraux de l'hygiène et de la propreté (Fernandez et Ferrer,
1982).
Des produits chimiques tels que les désinfectants sont parfois appliqués au sable
sans prendre en considération leurs effets écotoxologiques possibles. Le
traitement du sable par des méthodes nécessaires et simples, telles que le
balayage et l'aération, pourrait être appliqué (Figueras et al, 1992), ainsi que la
surveillance constante de plage afin d'empêcher l'accès par des animaux.
L'utilisation des serviettes propres pour l'usage sur la plage, bon personnel
l'hygiène, la prohibition des animaux et le nettoyage mécanique régulier sont
considérés comme étant, par quelques autorités, importants (Conseil Supérieur
d'Hygiène Publique de France, 1990).
7. Le transport bactérien dans le sable
Wood (1953) a fait un examen approfondi de ce phénomène et chiffre les
relations existantes entre la dimension des particules, leur densité et le nombre
de germes. Des recherches récentes de Meadows et Anderson (1966) relatives à
la fixation des micro-organismes sur les grains de sable, montrent l'adsorption
des entérovirus par les floculats, les boues activées et les précipites de sulfate
d'alumine peut atteindre 99 %.
Les bactéries du milieu marin, et celles qui y sont charriées, se fixent
pratiquement toujours sur les particules en suspension dans l'eau. Pour s'en
convaincre, il suffit de plonger une lame de verre dans 1'eau de mer, de la
reprendre au bout de 24 heures et de faire sur cette lame une coloration de Gram.
(Jacobs et al, 2007)
L’étude du transport bactérien, dans un sable et dans un sol, d’une communauté
bactérienne issue d’une boue de station d’épuration a montré les points suivants
-une forte réduction de la diversité microbienne et de la concentration
bactérienne ;
-parmi les espèces transportées se trouvaient des Coliformes fécaux ;

Revues bibliographiques sur les peuplements bactériologiques du sable
33
-les bactéries ayant traversé les colonnes sont chargées négativement ;
- Malgré ces conditions défavorables à l’adhésion pendant le transport de la
communauté bactérienne des boues, la rétention des bactéries dans les milieux
poreux était importante (Tian et al, 2002).
Cette forte réduction peut être due à la difficulté de disperser la boue, ce qui a
probablement eu pour conséquence de laisser des amas bactériens qui ont pu être
filtrés ou à la distribution des propriétés d’adhésion au sein du mélange. (Tian et
al, 2002).
Ces résultats confirment le rôle important des interactions électrostatiques sur la
migration des bactéries en milieux poreux (Tian et al, 2002).
Le transport bactérien n’est pas identique pour toutes les souches. Le nombre de
cellules constituant la communauté bactérienne est également fortement réduit
par la traversé des milieux poreux : 60% de rétention sur le sable et 90% sur les
colonnes de sol (Joy et al, 1998).
Le transport bactérien est fortement dépendant des propriétés physicochimiques
du sol et les écoulements qui y ont lieu. Pour des milieux homogènes ce sont les
interactions électrostatiques qui dominent le transport. En revanche pour les
milieux plus complexes les caractéristiques hydrodynamiques et porales sont
plus déterminantes. (Jacobs et al, 2007).
Aujourd’hui il est admis que deux mécanismes principaux empêchent le
déplacement de cellules microbiennes dans un sol : l’adhésion bactérienne et la
filtration. Les phénomènes d’adhésion sont contrôlés par des interactions
physicochimiques entre la bactérie et la phase solide. Néanmoins cette approche
nécessite une caractérisation de la surface des cellules et des grains du milieu
poreux qui est complexe (Jacobs et al, 2007).
De plus l’adhésion bactérienne est couplée à des processus biologiques propre à
la nature des bactéries. En effet de nombreuses espèces bactériennes produisent
des composées extracellulaires dont le rôle est soit de promouvoir soit
d’empêcher l’adhésion. De ce fait, le caractère vivant des cellules microbiennes

Revues bibliographiques sur les peuplements bactériologiques du sable
34
complique beaucoup l’étude de l’adhésion bactérienne en sable. Le second
mécanisme qui s’oppose au transport microbien est la filtration. Ce facteur est
très dépendant de la composition et de la porosité des milieux poreux. Aussi le
choix des milieux poreux utilisés pour ce type d’études est-il primordial (Jacobs
et al, 2007).
Les premières études sur le transport microbien en milieux poreux,
particulièrement le sable, remontent aux années 70 (Gerba, 1975; Rajagopalan,
1976). Pendant les années 90 le nombre d’articles augmente considérablement
impliquant des scientifiques de disciplines très diverses. En effet les processus
mis en jeu dans les déplacements de bactéries sont très nombreux et complexes.
La modélisation et la compréhension du déplacement microbien en milieux
poreux est typiquement un couplage entre des mécanismes physiques (filtration,
forces hydrodynamiques…), chimiques (hydrophobicité, liaisons faibles…) et
biologiques (caractéristiques membranaire, mobilité…). Cette dernière
composante complique beaucoup l’étude du transport microbien dans un profil
sol à cause du caractère vivant des bactéries. (Webb et al, 1999).
Les premiers travaux ont principalement porté sur deux axes : les interactions
des bactéries avec la surface solide (Marshall et al, 1971 ; Marshall, 1976;
Marshall, 1980) et la modélisation du transport (Yao, 1971 ; Rajagopalan,
1976).
En effet les interactions des cellules avec le sable peuvent aboutir à l’adhésion
des bactéries et empêcher leur transport. Ces interactions ont également étudiées
dans le cadre de la problématique des biofilms (Marshall et al, 1971 ; Costerton
et al, 1978; Costerton 1985 ; Lappin-Scott, 1989 ; Costerton et al, 1995;
Costerton et al, 1999).
Dans l’étude pionnière par Marshall et al (1971) les auteurs suggèrent que la
sorption de bactéries à la surface implique une étape initiale de sorption
réversible suivi par une sorption plus lente dépendant de la surface conduisant à
une adsorption irréversible (Marshall, 1971).

Revues bibliographiques sur les peuplements bactériologiques du sable
35
En effet un nombre croissant d’études suggère que le transport microbien est
fortement influencé par la physicochimie des surfaces cellulaires (Gannon,
1991 ; DeFlaun et al, 1999 ; Hermansson, 1999; Chen et Strevett, 2001; Chen et
Strevett, 2002; Chen et Strevett, 2003 ; Redman et al, 2004).
Cependant les seules interactions physicochimiques ne suffisent pas à décrire le
comportement des cellules bactériennes dans un sable et de nombreux articles
ont décrit l’importance des forces hydrodynamiques (Smith 1985) ou encore les
caractéristiques du milieu poreux traversé (Pekdeger, 1983; Corapcioglu, 1984).
A cela il faut ajouter que le comportement des bactéries dans un sable peut
varier d’une espèce à l’autre en fonction de ses caractéristiques morphologiques
(Corapcioglu, 1984), de surface cellulaire ou sa capacité de survie (Ward, 1980).
Cette synthèse bibliographique propose un état actuel des connaissances sur le
déplacement de bactéries dans le sable, une attention particulière sera portée sur
les 3 mécanismes principaux qui régissent le transport bactérien dans le sable:
l’adhésion, la filtration et les forces hydrodynamiques (Wan et al, 1994).
7.1. Adhésion bactérienne
L’étude de l’adhésion bactérienne est devenu un enjeu crucial depuis que de
nombreuses observations ont montré que 99,9% des bactéries vivent attachées à
une surface (Costerton et al, 1995). Les communautés bactériennes attachées à
une surface sont appelées biofilms. En 1995, Costerton et al ont soutenu que les
biofilms pouvait adhérer à des surfaces et les uns aux autres, incluant dans cette
définition les populations adhérentes dans les milieux poreux (Costerton et al,
1995).
Tout comme pour les biofilms il est essentiel de comprendre ces phénomènes
d’adhésion dans le sable, qui peuvent ralentir les cellules dans leur progression à
travers ce sol. Etudier l’adhérence de cellules bactériennes en sable revient à
étudier les étapes initiales du développement d’un biofilm (busscher, 1984).

Revues bibliographiques sur les peuplements bactériologiques du sable
36
7.1.1. Les étapes de l’adhésion bactérienne
L’adhésion bactérienne à lieu en 4 étapes (van Loosdrecht et al, 1990):
a. Accès à la surface
Dépend du mode de transport qui permet à une cellule bactérienne d’atteindre la
surface. Ce mode de transport peut être par diffusion, par convection (flux du
liquide) ou actif si la cellule dispose de moyens de propulsion. Dans ce dernier
cas le chimiotactisme joue également un rôle. Avant d’adhérer la cellule doit
être transportée vers la surface et plusieurs mécanismes permettent le
déplacement bactérien dans un sol pour augmenter les opportunités de
contact (Marshall, 1986)
- Les particules colloïdales sont animées de mouvements aléatoires. Les
bactéries, qui peuvent être assimilées à des particules colloïdales, présentent un
mouvement non négligeable (40μm.h-1) en moyenne (Marshall, 1976) visible
sous microscope. Ce phénomène contribue à augmenter les opportunités de
contact entre particules solides du sol et cellules bactériennes (Marshall, 1976;
Marshall, 1986).
- Les mouvements de diffusion/sédimentation sont lents par rapport aux vitesses
d’écoulement de l’eau du sol. Les forces de cisaillements créées par
l’écoulement peuvent faciliter le rapprochement de la cellule vers une surface du
sol (pour l’adsorption) (Rijnaarts, 1993) ou au contraire arracher la cellule
adsorbée (McClaine et Ford, 2002).
- Enfin certaines bactéries ont leur propre mobilité grâce à la présence
d’appendices spécifiques (flagelles...) (McCaulou, 1995 ; Camesano et Logan,
1998 ; McClaine et Ford, 2002 ;Becker et al, 2004) et peuvent répondre à des
stimuli chimiotactique (Olson et al, 2004). L’ensemble de ces phénomènes
contribue à la mobilité des bactéries dans un milieu poreux et influent en
conséquence sur leur déplacement dans un sol.

Revues bibliographiques sur les peuplements bactériologiques du sable
37
b. Adhésion initiale
Procédé essentiellement physico-chimique qui peut être réversible. L’adhésion
résulte d’interactions physicochimiques entre la cellule et la phase solide.
(Absolom et al,1983). Un nombre croissant de publications suggère que la
rétention des cellules microbiennes est fortement influencée par des
caractéristiques de la paroi des cellules (Lovly, 2003).
Dans les années 80 plusieurs études établissent un rapport entre les propriétés
physico-chimiques de la surface cellulaire et le phénomène d’adhésion cellulaire
(Busscher 1984; van Loosdrecht, 1989). On parle aussi d’interactions à « longue
distance » car la distance qui sépare la cellule de la surface est alors supérieure à
1nm (van Loosdrecht, 1989).
c. Attachement
grâce à des appendices cellulaires (fibrilles,polymères…) qui forment des
liaisons fortes entre la cellule et la surface, liaisons qui peuvent être
irréversibles. La phase d’attachement est d’ordre biologique avec l’implication
d’appendices, cellulaires (Vidal et al. 1998) et/ou la production de biopolymères
exopolysaccharides…) faisant office de colle. Lorsque les macromolécules de
surface ont une affinité pour la surface un pontage peut se former ce qui a pour
effet d’ancrer la bactérie à la surface (Vidal et al, 1998).
A noter que certains composants extracellulaires au contraire peuvent
empêchent l’adhésion à cause d’interactions stériques répulsives (van
Loosdrecht et al, 1990 ; Rijnaarts et al, 1994 ; Liu et al, 2007).
La phase d’attachement concerne donc seulement les interactions stériques
attractives (ou interactions de « pontage ») (Vidal et al, 1998).
d. Colonisation de la surface
Il s’agit de la formation du biofilm proprement dit, produit par l’ensemble de
bactéries sur la surface du grain de sable (van Loosdrecht et al, 1990).

Revues bibliographiques sur les peuplements bactériologiques du sable
38
7.1.2 L’approche physicochimique
L’importance des interactions électrostatiques est très dépendante de la présence
d’ions dans le milieu (van Loosdrecht, 1989) (force ionique et espèce). Dans le
cas où le colloïde et la surface ont des charges de même signe et présentent aussi
des interactions négatives Le colloïde est attiré vers la. Parce que les bactéries et
les grains du sable ont généralement une charge électrique globale de même
signe (négatifs) ce qui empêche l’adhésion. En revanche ces interactions
contribuent à la rétention des bactéries près de la surface. Il se forme un puits à
quelques dizaines de nanomètres de la surface dont la profondeur diminue au fur
et à mesure que la distance de séparation colloïde-surface augmente (van
Loosdrecht, 1989).
7.1.2 .1.Sensibilité des interactions électrostatiques à l’environnement
a)La force ionique
Plusieurs articles montrent la relation entre force ionique, forces électrostatiques
et adhésion cellulaire sur différentes surfaces (Loosdrecht, 1989 ;Rijnaarts et al ,
1999; van Li et Logan 2004).
Selon la conclusion de ces études l’augmentation de la force ionique augmente
l’adhésion et inversement (Rijnaarts, 1995).
La relation entre force ionique, interactions électrostatiques et transport
microbien dans le sable a également été étudiée (Johnson et al, 1996; Li et
Logan, 1999; Rijnaarts et al, 1999 ; Redman et al, 2004 ; Choi et al, 2007;).
Ces publications mettent en évidence l’importance du rôle des électrolytes sur
l’adhésion et par conséquent le déplacement de cellules bactériennes à travers
une matrice poreuse Lorsque la molarité de la solution est nulle la double
couche électronique est très épaisse ce qui se traduit par de fortes répulsions
électrostatiques entre les bactéries et le sable créant ainsi des conditions
défavorable pour l’adhésion. Dans de telles conditions le transport bactérien
dans le sable est favorisé (Redman et al, 2004).

Revues bibliographiques sur les peuplements bactériologiques du sable
39
Redman et al (2004); constatent le transport seulement partiel d’E. coli, à travers
une colonne de sable malgré la présence de la barrière électrostatique (Redman
et al, 2004). Ils suggèrent que les cellules qui n’ont pas été transportées sont
retenues dans le minimum secondaire. Ces interactions n’étant pas influencées
par la composition et la concentration de la solution, les interactions
électrostatiques peuvent devenir prépondérantes lorsque la force ionique est
faible (Redman et al, 2004).
Redman et al (2004) constatent alors qu’en diminuant la force ionique,
l’ensemble des bactéries retenues dans le sable sortent de la colonne. Ils en
déduisent que ces bactéries étaient retenues dans le minimum secondaire éliminé
par la diminution de la force ionique. Tong et Johnson(2006) ont obtenu des
résultats similaires avec des microsphères (Tong et Johnson, 2006)
b) L’effet du pH de la solution sur les interactions électrostatiques
L’adhésion bactérienne est sensible au changement de pH du milieu (Hamadi et
al. 2005).
La plupart des études montrent une meilleure adhésion lorsque le pH de la
suspension bactérienne est proche du point isoélectrique (PIE) (Webb et al.
1999). Le PIE est la valeur du pH pour laquelle la charge électrique nette d’une
surface est nulle.
Rijnaarts et al (1999), ont utilisé le PIE des bactéries comme un indicateur de
présence de polymères à la surface cellulaire qui inhibent l’adhésion (Rijnaarts
et al, 1995).
Van der et Kooij (1991) ont montré que les groupements anioniques à la surface
des cellules bactériennes dominent les groupements cationiques (van der et
Kooij, 1997).
le PIE des bactéries se situe généralement en pH très acide. Or dans bien des
cas, le transport bactérien concerne des environnements avec des pH proches de
la neutralité (entre 6 et 8) valeurs bien supérieures au PIE des bactéries. Si on

Revues bibliographiques sur les peuplements bactériologiques du sable
40
fait exception des environnements extrêmes, la charge électrique nette d’une
cellule bactérienne dans un sol est donc généralement négative. (Webb et al,
1999)
7.1.3 Les facteurs biologiques
La composition chimique d’une surface cellulaire est complexe et évolue au
cours du temps. Les membranes peuvent être modifiées en fonction de
l’environnement, du stress ou encore de l’apport nutritionnel (Sanin et al, 2003).
Or les propriétés d’adhésion sont en partie déterminées par les caractéristiques
physicochimiques (hydrophobicité par exemple) de la membrane cellulaire (van
Loosdrecht et al, 1987; van Loosdrecht, 1990).
De plus de nombreuses espèces bactériennes sont capables de produire des
composés et/ou des appendices extracellulaires qui peuvent à leur tour modifier
les propriétés d’adhésion des cellules (Zita et Hermansson, 1997 ; Iwabuchi et
al, 2003).
A cause de l’implication de ces caractéristiques biologiques des cellules dans la
formation de biofilms la littérature est très riche et propose de nombreuses
études. En revanche l’influence des facteurs biologiques (polymères de surface,
motilité cellulaire et conditions physiologique) sur le transport microbien dans le
sable a reçu moins d’attention (van Loosdrecht et al. 1987; van Loosdrecht
1990).
7.1.3.1. Polymères de la surface cellulaire
a. Les polymères extracellulaires (ou EPS)
Les EPS sont composés d’une mixture de macromolécules très diverses telles
que des polysaccharides, des protéines ou des composés lipidiques (Nielsen et
al. 1997). Les EPS sont impliqués dans la formation et le développement de
biofilms (Costerton et al, 1995).
Cependant le rôle exact des EPS dans les phénomènes d’adhésion reste encore
malconnu. En effet la composition chimique des EPS est très hétérogène et leur

Revues bibliographiques sur les peuplements bactériologiques du sable
41
production varie en fonction des souches modifiant ainsi les propriétés
d’adhésion (Tsuneda et al. 2003).
Certaines études mentionnent l’effet positif des EPS sur l’adhésion bactérienne
Costerton, 1985; Dufrene et al, 1996) tandis que d’autres mentionnent le
contraire (Gomez-Suarez et al, 2002).
b. les Lipopolysaccharides (LPS)
Les lipopolysaccharides sont des macromolécules constituées d’une chaîne
polysaccharidique attachée à la membrane cellulaire par une partie lipidique
(lipide A) (DeFlaun et al, 1999).
Les chaînes latérales (ou antigène O) sont de nature polysaccharidique et sont
spécifiques à chaque souche. Le « core » est également de nature saccharidique.
Il y a plus de 3,5 millions de molécules LPS sur la face externe de la membrane
des cellules Gram négatif. L’impact des LPS sur le transport microbien dans le
sable a été abondamment abordé (DeFlaun et al, 1999;Abu-Lail et Camesano,
2003; Walker et Redman , 2004; Walker et al, 2004 ; Liu et al, 2007).
c. Les Pili
Le pilus (ou fimbriae, pili au pluriel) est un appendice se situant à la surface de
la paroi de nombreuses bactéries à Gram négatif (et exceptionnellement des
bactéries à Gram positif) et qui peut mesurer jusqu’à 2μm. On en distingue deux
types : les pili communs et les pili sexuels. Les pili communs peuvent intervenir
dans les phénomènes d’adhésion spécifique notamment avec des récepteurs
situés à la surface des cellules eucaryote (Bullitt et Makowski, 1998).
Le caractère hydrophobe des pili pourrait être à l’origine des propriétés
adhésives qu’on leur attribue généralement (Ward, 1980).
Cependant la littérature ne mentionne aucune étude de l’impact des pili sur le
transport microbien dans le sable (Ward, 1980).

Revues bibliographiques sur les peuplements bactériologiques du sable
42
7.1.3.2. Motilité cellulaire et transport bactérien dans le sable
Certaines cellules bactériennes possèdent leurs propres moyens de propulsion
grâce des appendices extracellulaires appelés flagelles. Ces structures
extracellulaires agissent comme des hélices ou des nageoires. Plusieurs études
ont été menées pour connaître l’influence de la motilité bactérienne sur leur
transport dans le sable (Camper et al, 1993 ; Camesano et Logan, 1998;).
Certaines comparaisons entre bactéries motiles et non motiles en chambre à
écoulement (flow cell) (McClaine et Ford, 2002) ou sur colonnes.
(Becker et al, 2004) montre que le taux d’attachement est plus élevé pour les
cellules mobiles alors que d’autres études obtiennent des résultats contraires
(Camesano et Logan, 1998).
Gannon et al, (1991) n’ont pas constaté d’influence de la motilité des cellules
sur le transport bactérien en milieu poreux (Gannon et al, 1991).
Le mécanisme par lequel la motilité bactérienne influe sur le déplacement des
cellules dans le sable demeure inconnu mais de nombreuses hypothèses ont été
faites. La mobilité bactérienne pourrait par exemple augmenter le taux de
collision avec la surface du milieu poreux et donc augmenter les chances
d’adhésion de la bactérie (van Loosdrecht, 1989).
Mc Caulou et al, (1995) affirment que la motilité bactérienne augmenterait le
taux de désorption grâce à la force de locomotion qui permet à la cellule de se
détacher de la surface (McCaulou et al, 1995).
Ces auteurs en déduisent que le transfert de bactéries motiles vers des aquifères
profond serait significativement accru grâce à leur motilité propre. Enfin,
McClaine et Ford (2002) suggèrent que le comportement de détachement de
bactéries motile est très influencé par la vitesse d’écoulement du fluide
(McClaine et Ford, 2002). Les flagelles pourraient aider à augmenter
l’attachement ou faciliter le détachement des bactéries de la surface selon les
forces hydrodynamiques du système (McClaine et Ford, 2002).

Revues bibliographiques sur les peuplements bactériologiques du sable
43
Dans certaines conditions le chimiotactisme peut influer sur le transport
microbien dans le sable en modifiant la direction de déplacement des cellules
motiles (Sen et al, 2005).
En effet certaines bactéries motiles peuvent se déplacer vers un environnement
plus favorable en réponse à un stimulus. Par exemple ce type de bactéries est
capable de se diriger vers une zone où la concentration en nutriments bénéfiques
est élevée ou de s’éloigner de substances nuisibles (McClaine et Ford, 2002).
Olson et al (2004) ont montré que le chimiotactisme peut jouer un rôle
important dans le déplacement microbien dans les aquifères (Olson et al, 2004).
Plusieurs auteurs ont intégré le facteur chimiotactisme dans leurs modèles pour
prédire le transport microbien dans un profil sol (Barton et Ford, 1997; Nelson et
Ginn, 2001; Sen et al, 2005). Barton et Ford n'ont observé aucune différence
significative du transport microbien (en colonne de sables) en présence ou
absence d'un gradient chimique (Barton et Ford, 1995).
Cependant les auteurs attribuent cette absence d’effet du chimiotactisme au
gradient chimique trop faible utilisé pendant leurs expériences (Barton et Ford,
1997).
7.1.3.3. Etat et condition physiologique
L’état physiologique d’une bactérie peut influencer les caractéristiques
physicochimiques de la surface des cellules et par conséquent les propriétés
d’adhésion. Par exemple l’hydrophobicité de la cellule peut être modifiée par la
phase de croissance et le taux de multiplication cellulaire (McEldowney, 1986).
Van Loosdrecht et al (1987) ont observé que les bactéries deviennent plus
hydrophobes pendant la phase de croissance exponentielle (van Loosdrecht et al,
1987).
La nutrition des cellules peut également modifier les propriétés d’adhésion des
cellules (Sanin et al, 2003).

Revues bibliographiques sur les peuplements bactériologiques du sable
44
Chen et Strevet (2003) ont mis en évidence l’impact des sources nutritives (ratio
de carbone sur azote) sur les propriétés de surface d’une souche d’E. coli (Chen
et Strevett 2003).
Les mêmes auteurs observent par ailleurs que la phase de croissance, la phase
stationnaire et la phase de dégénérescence n’ont pas d’effet significatif sur les
propriétés de surface (en particulier γLW) des souches d’ E. coli, Pseudomonas
fluorescence et Bacillus subtilis (Chen et Strevett, 2001). Grasso et al (1996) ont
fait le même constat (Grasso et al, 1996).
Smet et al(1999) ont étudié l’impact de l’état physiologique de P. fluorescence
sur l’adhésion et le transport à travers des colonnes remplies de billes de verre
(Smets et al, 1999). Les trois états physiologiques testés ont été définis par
phase de croissance, la phase stationnaire et la phase de dégénérescence.
Les cellules en phase exponentielle adhérent mieux sur le verre que les cellules
en phase stationnaire ou en dégénérescence. Les auteurs constatent également
que les cellules en phase exponentielle sont le plus hydrophiles, en contradiction
avec les travaux cités ci-dessus (Smets et al, 1999).
Ceci est d’autant plus surprenant que cette hydrophilicité augmente les
répulsions entre le verre et les cellules bactériennes et devrait donc
théoriquement diminuer l’adhésion. L’explication pourrait venir du potentiel
zêta plus faible des cellules en phase de croissance. En effet les auteurs
suggèrent que les cellules en phase exponentielle, du fait de leur faible potentiel
zêta, sont vraisemblablement retenues dans un minimum secondaire qui est pour
cette phase de croissance plus important que pour les autres états
physiologiques. (Smets et al, 1999).
7.1.3.4. Densité cellulaire
Il a été postulé que plus la concentration en cellules est élevée, plus le taux
d’adsorption bactérienne sera élevé. Bengtsson et Lindqvist (1995) ont constaté
que lorsque la concentration cellulaire dans la phase liquide augmente la

Revues bibliographiques sur les peuplements bactériologiques du sable
45
quantité de cellules attachées aux particules de sol augmente également jusqu’à
ce que la surface disponible à l’adhésion arrive à saturation (Bengtsson et
Lindqvist, 1995).
Il est a noté que Bradford et Battahar(2006), utilisant des colloïdes artificiels,
ont observé une diminution de la rétention quand la concentration d’injection
augmente (Bradford et Bettahar 2006). Les auteurs postulent des interactions
répulsives entre colloïdes en solutions et adsorbés pour expliquer leurs
observations.
7.2. La filtration
Le mécanisme de filtration correspond à un blocage physique des cellules par
des pores dont la taille est inférieure à celles des cellules bactériennes
(Corapcioglu, 1984).
Les facteurs qui influent sur la filtration dans un milieu poreux sont : la forme et
la taille des cellules, le niveau de saturation en eau du milieu poreux et
l’obstruction éventuelle des pores (Stevik et al, 2004).
7.2.1. Le milieu poreux
La taille des grains constituant le milieu poreux est un facteur important dans le
transport microbien comme l’attestent plusieurs travaux (Fontes et al, 1991;
Stevik et al, 1999 ;Ausland et a, 2002; Bradford et al, 2006)
Plus la taille des grains est petite plus la taille des pores est réduite limitant ainsi
le passage des bactéries. L'argile, le limon et le sable fin induisent des tailles de
pores de l’ordre de grandeur de la plupart des cellules bactériennes. La filtration
peut ainsi être un mécanisme limitant le mouvement bactérien à travers ce type
de matériaux (Matthess, 1985).
La présence de macro pores et de fractures dans une matrice poreuse au
contraire facilite le transport microbien ( Natsch et al, 1996 ; Artz et al, 2005).
La présence de galeries formées par des vers de terre par exemple facilite le
transport d’E. coli O157:H7 tandis que le compactage du sol limite son transport

Revues bibliographiques sur les peuplements bactériologiques du sable
46
(Artz et al, 2005). Ibaraki et Sudicky (1995) ont réalisé des simulations
numériques de transport colloïdal par un réseau de fractures dans un milieu
poreux (grès et argile) ( Ibaraki et Sudicky, 1995).
Leurs travaux montrent que le manque de données sur les réseaux de fractures et
les coefficients de filtration augmentent l’incertitude de prédiction du transport
colloïdal pour des applications à grande échelle. En effet lorsque l’eau coule
dans les pores de grande taille les vitesses d’écoulement sont plus rapides
augmentant la vitesse et la distance de transport (Ibaraki et Sudicky, 1995).
Bradford et al (2005) ont publié récemment une série d’articles insistant sur
l’importance de l’effet filtration sur le transport des colloïdes ou de bactéries en
milieu poreux (Bradford et al, 2003 ; Bradford et al, 2005; Bradford et Bettahar,
2006)
En testant le transport de la souche E. coli O157:H7 dans des colonnes de sables
de porosité variables Bradford et al (2005), ont clairement mis évidence
l’influence de la taille des grains de sable.
Dans ce type d’étude les sédiments utilisés ont généralement des tailles de grain
très uniforme. Cependant, la plupart des sédiments normaux ont une distribution
de la taille des grains fortement non uniforme : dans les sédiments naturels 10%
des pores sont assez petits (>1μm) pour gêner transport bactérien (Jordan et al,
2004). Brown et al (2002) ont également constaté l’influence de la forme des
grains de sables sur le transport microbien, les grains ovales étant moins
propices au transport que les grains ronds (Brown et al, 2002).
Plusieurs travaux ont tenté d’établir un ratio entre la taille des bactéries et la
taille des pores (ou encore tailles des grains) à partir duquel la filtration opère
(Matthess 1985). Mais ce ratio varie beaucoup en fonction des auteurs. Brouwer
estime que la filtration débute lorsque le diamètre des grains constituant les
milieux poreux est inférieur ou égal à 5 fois le diamètre des colloïdes en
suspension (Brouwer 1984). Selon Bradford et al(2005). La filtration se produit

Revues bibliographiques sur les peuplements bactériologiques du sable
47
quand la ration entre le diamètre des particules en suspension (de) et le diamètre
moyen des grains (d50) est supérieur à 0.0017 (Bradford et al, 2005).
Bradford et al (2005) utilisent le même calcul (de/d50) pour déterminer que la
filtration d’E. coli O157:H7 par des sables fins se produit lorsque de/d50 est
supérieur à 0.005 (Bradford et al, 2005).
Un autre phénomène qui peut être assimilé à la filtration par un milieu poreux
est le blocage des colloïdes au niveau des zones de contact entre grains
constituant la matrice poreuse (Bradford et al, 2003; Bradford et al, 2005) .Plus
récemment le blocage de colloïdes par des zones de contact entre grains d’un
milieu poreux a pu être visualisé et est considéré comme un mécanisme
important de déposition (Li et al, 2006). Il a aussi été montré de façon théorique
que le phénomène de blocage dans une matrice porale peut se produire même en
présence d’une barrière d’énergie (Johnson et al, 2007).
7.2.2. La morphologie des bactéries
Les études sur l’influence de la forme et de la taille des bactéries sur la filtration
sont peu nombreuses. Gannon et al(1991) ont trouvé une bonne corrélation entre
la filtration et la taille des cellules (Gannon et al, 1991).
Weiss et al (1995) ont étudié l’influence de la morphologie cellulaire (en
calculant le ratio largeur sur longueur cellulaire) de 14 souches sur le transport
microbien dans le sable (Weiss et al, 1995). Le résultat de leurs études suggère
que les bacilles allongés sont les plus filtrés par le sable. De même ils montrent
que les plus petites cellules sont les mieux transportés (Weiss et al, 1995). Plus
récemment Salerno et al (2006), ont utilisé des microsphères de latex
artificiellement déformées pour étudier l’effet de la morphologie des colloïdes
sur la filtration en milieux poreux (Salerno et al, 2006).
Les résultats montrent que la filtration dans les colonnes remplies de billes de
verre (diamètre 40μm) est d’autant plus élevée que le ratio longueur sur largeur
est grand (Salerno et al, 2006). Le contraste est surtout remarquable avec les
colloïdes témoins qui sont parfaitement sphériques. A noter que la chimie de

Revues bibliographiques sur les peuplements bactériologiques du sable
48
surface et le potentiel zêta ne jouent pas un rôle discriminant dans cette
expérience puisque ces caractéristiques sont identiques pour tous les colloïdes
utilisés. Les auteurs concluent que la forme allongée de certaines particules,
telles que les bactéries de type bacille, est un facteur fortement impliquée dans
les phénomènes de rétention en milieu filtrant (Salerno et al, 2006).
7.2.3. L’obstruction des pores
La diminution du volume poral par l’obstruction des pores peut être une cause
de filtration dans un environnement poreux tel que le sable. Une des causes
majeures de l’obstruction peut être la formation de biofilms (Thullner et al,
2002) ou des cellules agrégées (Bradford et al. 2006) qui remplissent les pores.
D’après Bradford et al (2006), des cellules d’E. coli non agglomérées peuvent
se déposer au niveau d’étranglement de pores et s’agglomérer pour former un
bouchon. Les cellules agglomérées peuvent ensuite être relâchées dans la phase
aqueuse sous l’effet de forces hydrodynamiques (Bradford et al, 2006).
Iliuta et Larachi (2006) ont simulé l’agrégation de cellules de Pseudomonas
putida dans un bioréacteur (lit poreux alimenté goutte à goutte avec des eaux
usées) (Iliuta et Larachi, 2006).
Rijnaarts et al (1996) ont constaté que l’obstruction des pores par des cellules
bactériennes dépend de la taille cellulaire et de la possibilité d’agrégation des
cellules (Rijnaarts et al, 1996).
7.2.4. Saturation en eau et charge hydraulique en sable
Lorsque le flux hydrique appliqué à la surface d’un milieu poreux insaturé est
faible (c'est-à-dire, inférieur à la conductivité hydraulique à saturation) l’eau
circule préférentiellement dans les petits pores favorisant ainsi les phénomènes
de filtration. Quand le flux appliqué est suffisamment élevé pour augmenter le
niveau de saturation en eau dans le sable, l’eau circule préférentiellement à
travers les pores les plus grands (loi de Poiseuille). Dans ces conditions les
phénomènes de filtrations sont réduits (Smith, 1985).

Revues bibliographiques sur les peuplements bactériologiques du sable
49
Une augmentation du flux hydrique contribue donc au transport microbien dans
le sable. De plus en conditions non saturées, la présence d’air dans le sable crée
des interfaces air-eau. Depuis les années 70 plusieurs études montrent que les
bactéries ou les colloïdes ont tendance à s’accumuler à l’interface eau-air
(Blanchard, 1972; Corapcioglu et Choi, 1996; Powelson et Mills, 1996; Schafer
et al, 1998).
Wan et al(1994) ont même visualisé l’accumulation de cellules bactérienne au
niveau d’interfaces eau-air après le transport de bactéries à travers des colonnes
de sables insaturées (Wan et al, 1994). Ces études suggèrent fortement que la
présence d’air dans un sol joue un rôle de rétention lors du transport bactérien.
7.3. Forces Hydrodynamiques
De nombreuses études on montré que le transport des bactéries à travers le sable
est amélioré lorsque les vitesses d’écoulement augmentent (Camper et al, 1993;
Sarkar et al, 1994; Tan et al, 1994).
Ainsi Huysman et Verstraete (1993) ont constaté que le transport bactérien à
travers des filtres était beaucoup plus élevé. Comme expliqué précédemment
l’effet de la filtration est amoindri lorsque le flux d’eau à la surface du milieu
filtrant augmente (Huysman et Verstraete, 1993).
A l’intérieur des pores les forces hydrodynamiques peuvent être responsables
du détachement de bactéries (phénomènes de cisaillement) mais également de la
rétention des cellules dans des zones mortes (Tan et al, 1994).
8. Le transport de bactéries pathogènes dans le sable
Il existe plusieurs sources de bactéries pathogènes susceptibles de contaminer
des nappes phréatiques ou des eaux de baignades après transport à travers un sol.
Parmi les sources les plus souvent citées on trouve les eaux usées (Stevik et al,
2004), les boues des stations d’épuration ou le fumier des élevages intensifs
(Cools et al, 2001).

Revues bibliographiques sur les peuplements bactériologiques du sable
50
Plusieurs auteurs ont fait le point sur les études faites en matière de transport de
souches pathogènes dans des environnements aquifères (Ferguson et al, 2003;
Santamaria et Toranzos, 2003; Stevik et al, 2004).
Cependant la littérature comporte un nombre limité de travaux sur les
conséquences liées au déplacement microbien dans un sol. Unc et Goss (2004)
relatent l’événement tragique de mai 2000 à Walkerton (Ontario, Canada) où
l’eau du robinet fut contaminée par les souches pathogènes Escherichia coli
0157:H7 et Campylobacter (Unc et Goss, 2004).
Cette contamination a entraîné l’hospitalisation de 2300 personnes dont 7 sont
décédées sur un total de 5000 habitants que compte la ville deWalkerton.
L’enquête sur les origines du drame a montré que la contamination était
vraisemblablement due à des bactéries issues de lisiers qui ont réussi à atteindre
l’aquifère (Unc et Goss, 2004).
Dans les zones rurales l’utilisation de systèmes d’assainissement des eaux usées
autonome se développement rapidement. Aux Etats-Unis 25% de la population
bénéficie de ce type de système (US Environmental Agency, 1997).
Les eaux usées sont d’abord stockées dans une fosse septique pour permettre la
décantation de la phase solide. Le surnageant est envoyé dans le sol via un
réseau de drains. Ce type d’assainissement a été conçu en considérant que le sol
agit comme un filtre. En effet des expériences ont démontré que les
microorganismes issus de ce type de système d’assainissement sont filtrés sur
des distances relativement courtes en milieu poreux insaturé (Hagedorn, 1981).
Cependant Gerba et al (1975) ont constaté que des bactéries Coliformes pouvait
être transportées jusqu’à une distance de 450 mètres à partir du point de
contamination (Gerba et al, 1975).
La migration de souches pathogènes peut ainsi aboutir à la contamination des
réserves en eaux potables et porter atteinte à la santé publique. Selon Scandura
et Sobsey (1997) les effluents des fosses septiques sont une source importante
responsable de la contamination des nappes phréatiques (Scandura et Sobsey,

Revues bibliographiques sur les peuplements bactériologiques du sable
51
1997). Les bactéries pathogènes importante dans ce type de contamination sont :
Salmonella sp., Shigella sp., Vibrio cholerea, Yersinia enterocolitica, Y.
pseudotuberculosis, Leptospira sp., Francisella tularensis, Dyspepsia coli, et
des souches productrices d’entérotoxines comme E. coli et Pseudomonas
(Matthess, 1985).
8.1. Le sort des bactéries pathogènes entérique dans un sol
8.1.1. La survie
Les bactéries entériques vivent normalement exclusivement dans le tractus
digestif des mammifères. De ce fait ces microorganismes sont peu adaptés à la
survie dans d’autres environnements et en particulier le sol. Néanmoins
plusieurs auteurs ont constaté que des Coliformes fécaux issues de stations
d’épuration pouvaient survivre plusieurs mois dans un sol (Ngole, 2006;
Pourcher, 2007). Une augmentation de l’humidité du sol, la diminution de la
température ou encore la quantité de matière organique disponible favoriserait la
survie des souches entériques (Ngole, 2006).
8.1.2. Le déplacement
L’argile favoriserait la réduction du transport de bactéries entériques dans un sol
par filtration et adsorption. En effet l’argile est constitué de particules très fines
(<2μm) bloquant ainsi le passage des cellules bactériennes (Huysman et
Verstraete, 1993). De la présence de groupements chargés positivement sur les
particules d’argile favorise l’adhésion électrostatique des bactéries généralement
de charge opposée (Fletcher, 1979).
Scandura et Sobsey (1997) soutiennent également que des sols avec au moins
15% d’argile sont capable de retenir des bactéries et des virus et réduisent
fortement les risques de contamination (Scandura et Sobsey, 1997). Cependant
Conboy et Goss en réalisant une surveillance de plusieurs puits ont remarqué
que les sols riches en argile sont les plus susceptibles à permettre une
contamination bactérienne d’origine fécale (Conboy, 2000).

Revues bibliographiques sur les peuplements bactériologiques du sable
52
Les événements pluvieux favoriseraient le déplacement de bactéries entériques
dans un sol. Plusieurs auteurs ont observé le déplacement de Coliformes sur plus
de 150 mètres lors d’épisodes pluvieux importants (Gerba, 1984 ; Celico et al,
2004).
8.1.3. Détection de pollutions fécales
Traditionnellement les Coliformes fécaux (aussi appelés Coliformes thermo
tolérants) ont été utilisés comme indicateur de contamination fécale. Plus
récemment l’Agence de Protection de l’Environnement des Etats-Unis (EPA) et
l’Organisation Mondiale de la Santé (OMS) proposent d’utiliser E. coli et les
entérocoques de l’intestin comme indicateurs de pollution fécale. De plus, des
techniques de biologie moléculaire ont été développées pour la détection d’E.
coli dans des environnements comme le sol (Sabat, 2000) ou les eaux usées
(Bertrand et Roig, 2007). Bertrand et Roig (2007) ont développé une technique
PCR pour détecter spécifiquement la souche pathogène E. coli 0157 H7 avec
une sensibilité de 200CFU/litre (Bertrand et Roig, 2007). La conférence
annuelle de la Société Américaine de Microbiologie (ASM) de 2007 (Toronto,
Ontario, Canada) a montré l’essor de méthodes de détection de souches
pathogènes dans l’environnement (Esseilli, 2007; Haffar, 2007; Hallier-Soulier,
2007).
Plusieurs études ont donc permis de détecter des pollutions fécales dans
l’environnement au moyen de ces techniques. Cependant la littérature ne
mentionne pas de lien clair entre une pollution fécale et le transport de bactéries
fécales en milieu poreux. (Conboy, 2000)
Le déplacement bactérien en milieux poreux résulte d’un couplage complexe
entre plusieurs mécanismes :
- Rétention : immobilisation des cellules par adhésion, blocage ou filtration dans
la matrice poreuse.
- Détachement : remobilisation des cellules faiblement retenues sur la matrice et/

Revues bibliographiques sur les peuplements bactériologiques du sable
53
ou décrochage par les forces de cisaillements exercées par l’écoulement.
- Transport : déplacement de cellules bactériennes dans un milieu poreux lié aux
lois de la convection dispersion. (Olson et al, 2004).
Une des lacunes majeure dans la littérature concerne le faible nombre d’études
du transport bactérien dans les milieux poreux hétérogènes tel qu’un sol. On n’a
trouvé que très peu de travaux rapportant des expériences de déplacement dans
des milieux autres qu’une colonne de sable ou de billes de verre. Dans une
matrice porale homogène l’hydrodynamique et la physicochimie de surface sont
simplifiées. (Sanin et al, 2003).
Dans ce sens les expériences en milieux homogènes (billes de verre, grains de
sables calibrés) sont généralement privilégiées parce que dans de telles
conditions l’étude de l’impact des mécanismes de déposition et de décrochage
est plus simple et peut se focaliser sur des aspects physicochimiques. Cependant,
on peut imaginer que, par analogie avec les phénomènes de transport de soluté,
le mécanisme de rétention étant un processus cinétique, il va être en compétition
avec le transport par l’écoulement ; Or, l’écoulement peut présenter des
hétérogénéités bien plus importantes (distribution des vitesses par exemple) dans
un milieu naturel que dans une colonne de matériaux fin et homogène. Par
conséquent, il nous semble nécessaire de conduire des expériences dans des
milieux simples mais aussi dans des milieux un peu plus compliqués pour
essayer de se rapprocher du milieu naturel du point de vue de la complexité des
écoulements ( Olson et al, 2004)
Moyennant cette gamme de complexité, il nous semble que l’on pourra en partie
évaluer dans quelle mesure les mécanismes de déposition entrent en compétition
avec le transport( Simoni et al,1998).
Le diagramme suivant illustre la durée qui est exigée pour que des particules de
différentes tailles se lient ensemble.
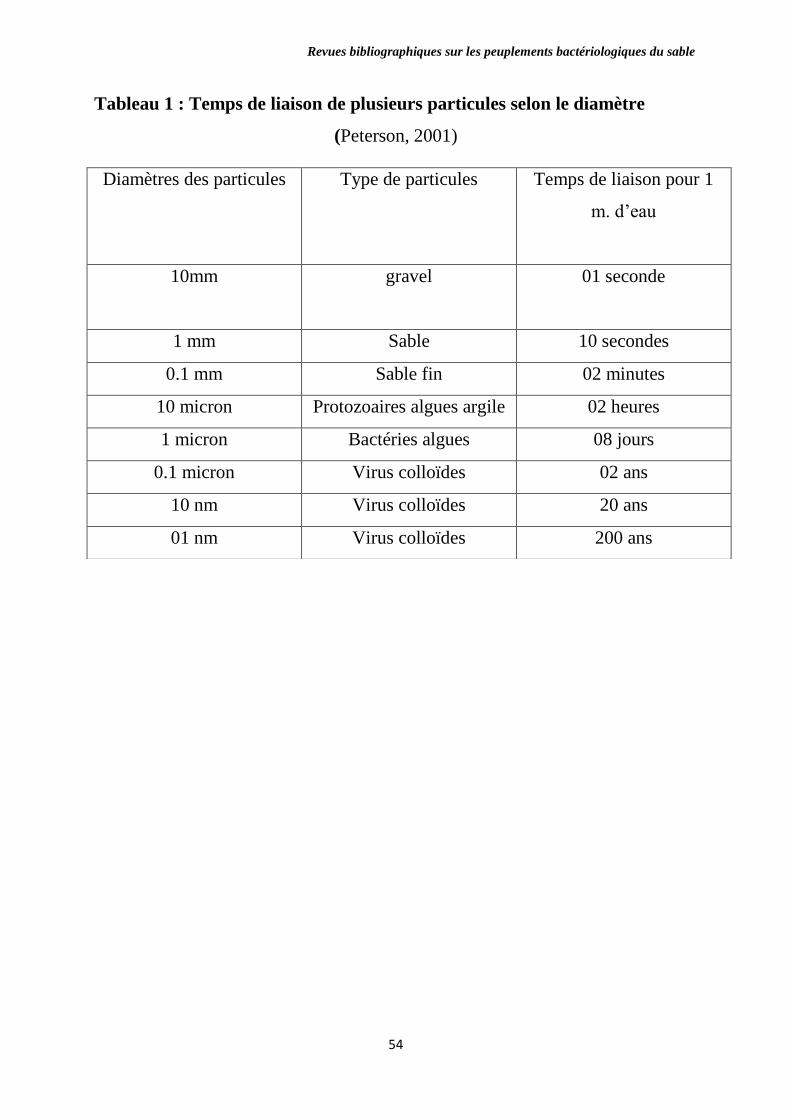
Revues bibliographiques sur les peuplements bactériologiques du sable
54
Tableau 1 : Temps de liaison de plusieurs particules selon le diamètre
(Peterson, 2001)
Diamètres des particules Type de particules Temps de liaison pour 1
m. d’eau
10mm
gravel 01 seconde
1 mm Sable 10 secondes
0.1 mm Sable fin 02 minutes
10 micron Protozoaires algues argile 02 heures
1 micron Bactéries algues 08 jours
0.1 micron Virus colloïdes 02 ans
10 nm Virus colloïdes 20 ans
01 nm Virus colloïdes 200 ans
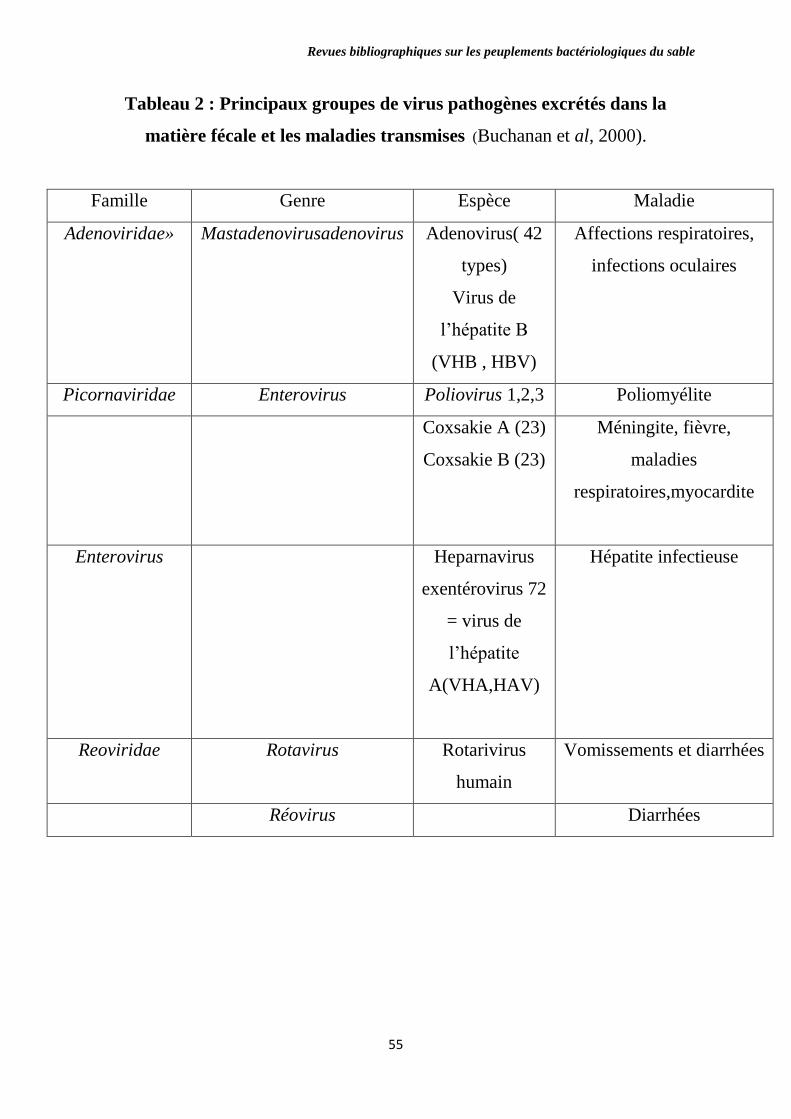
Revues bibliographiques sur les peuplements bactériologiques du sable
55
Tableau 2 : Principaux groupes de virus pathogènes excrétés dans la
matière fécale et les maladies transmises (Buchanan et al, 2000).
Famille Genre Espèce Maladie
Adenoviridae» Mastadenovirusadenovirus Adenovirus( 42
types)
Virus de
l’hépatite B
(VHB , HBV)
Affections respiratoires,
infections oculaires
Picornaviridae Enterovirus Poliovirus 1,2,3 Poliomyélite
Coxsakie A (23)
Coxsakie B (23)
Méningite, fièvre,
maladies
respiratoires,myocardite
Enterovirus
Heparnavirus
exentérovirus 72
= virus de
l’hépatite
A(VHA,HAV)
Hépatite infectieuse
Reoviridae Rotavirus
Rotarivirus
humain
Vomissements et diarrhées
Réovirus Diarrhées
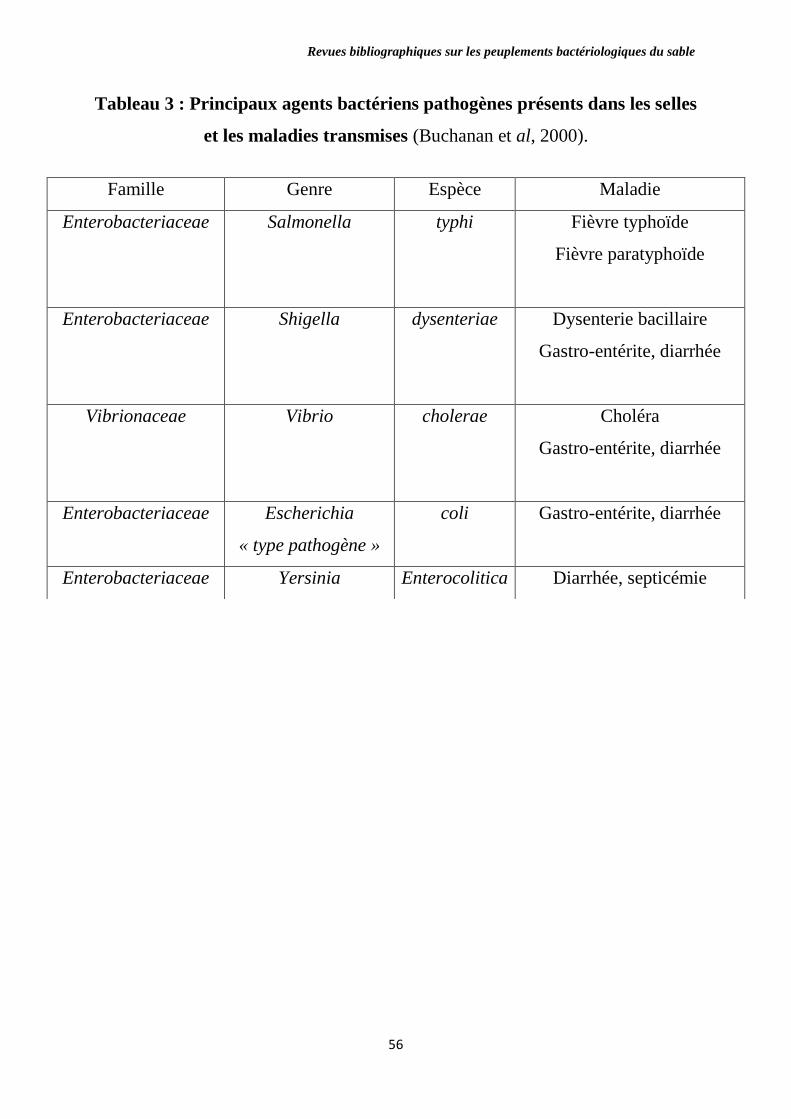
Revues bibliographiques sur les peuplements bactériologiques du sable
56
Tableau 3 : Principaux agents bactériens pathogènes présents dans les selles
et les maladies transmises (Buchanan et al, 2000).
Famille Genre Espèce Maladie
Enterobacteriaceae Salmonella typhi Fièvre typhoïde
Fièvre paratyphoïde
Enterobacteriaceae Shigella dysenteriae Dysenterie bacillaire
Gastro-entérite, diarrhée
Vibrionaceae Vibrio cholerae Choléra
Gastro-entérite, diarrhée
Enterobacteriaceae
Escherichia
« type pathogène »
coli Gastro-entérite, diarrhée
Enterobacteriaceae Yersinia Enterocolitica Diarrhée, septicémie

Matériels et méthodes

Matériels et méthodes
57
1. Sites d’étude
Les quatre sites retenus pour cette étude sont situés dans différents points de la
côte oranaise (Tableau 4), Ain El-Turk( Figure 1) est située à 15 km en nord-
ouest de la wilaya d’Oran, une zone marquée par l’urbanisation incontrôlée de
son rivage, les Andalouses à 25 km, une plage caractérisée par un espace vaste
de sable fin et doré, hautement fréquentée par les touristes.
Beau séjour, Andalouses (Figure 3) sont deux plages sous l’influence de
l’émissaire de la communauté urbaine de Ain El- Turk et El Ançor.
Madagh (Figue 4) est une zone non impactée puisqu’elle est située loin de la
métropole oranaise d’environ 40 km vers l’Ouest et où l’action anthropique est
très peu marquée (Sahnouni, 2003 ; Kherraz, 2004 ; Moffouk, 2005).
Tableau 4 : Caractérisation des quatre sites d’échantillonnage.
Sites Géoréférencement Commune Caractéristiques
Beau Séjour N 35° 44' 54.81"
W 0° 46' 09.18"
Ain El-Turck Rejet des effluents
urbains et industriels
Eden N 35° 45' 13.46"
W 0° 46' 50.81"
Ain El-Turck Zone touristique
Les Andalouses N 35° 42' 23.18"
W 0° 53' 13.05"
El Ançor Zone touristique
Madagh N 35° 37' 952"
W 000° 104' 243"
Ain El Karma Aire marine protégée

Matériels et méthodes
58
Figure 1 : Site d’Ain El-Turk. Figure 2 : Site d’Eden.
Figure 3 : Site des Andalouses. Figure 4 : Site de Madagh.

Matériels et méthodes
59
2. Echantillonnage
Un total de 144 échantillons a été analysé durant la saison hivernale et
estivale. Après prélèvement, les échantillons sont mis dans des flacons marqués
(Figure 5) et transportés après leur conservation dans une glacière à 4°C.
Figure 5 : Echantillons récoltés des quatre sites.
Figure 06 : Sites d’échantillonnage (Google 2011)
L’eau est recueillie dans des bouteilles en plastique stériles de 250 ml. Le sable
sec a été prélevé et mis dans des grands tubes à vis en plastique de 250 ml,
dans les zones non inondées par l’eau de mer (au niveau de l’étage supralittoral)

Matériels et méthodes
60
tandis que le sable humide a été prélevé dans la zone intermédiaire entre le
sable sec et l'eau de mer (étage médiolittoral).
Les échantillons ont été mis dans une glacière (4°C) et transportés dans des
conditions stériles au laboratoire de recherche Réseau de Surveillance
Environnementale (LRSE) et analysés de suite.
3. Mesure du pH
Le pH de l’eau est mesuré en introduisant l’électrode du pH mètre dans un
bécher contenant la solution à analyser.
Le pH du sable a été mesuré ainsi :
On introduit 10 g de sable dans 90 ml d’eau déminéralisé et après décantation,
on plonge l’électrode du pH mètre dans le liquide décanté.
4. Mesure de la température
On introduit la sonde de température dans l’échantillon à analyser (eau ou sable)
et on passe à la lecture affichée sur le thermomètre.
5. Préparation des échantillons aux analyses microbiologiques
5.1. Les dilutions
Entre la préparation de la suspension, ses dilutions et la mise en culture, il ne
doit pas s’écouler plus de 45 mn. Les dilutions suivent des séries logarithmiques
dont les termes sont en progression géométriques ; par exemple les dilutions
décimales : 0,1 (10-1) ; 0,01 (10-2).
Prélever une capacité de 1 g de sable à l’aide d’une spatule stérile et le
transférer dans un tube stérile contenant 9 ml d’eau distillée stérile ou eau
physiologique (recherche de Staphylocoques) = Obtention de la suspension
mère.
De même pour l’analyse d’eau en prenant en considération que la solution mère
est l’eau de mer. Ensuite on prélève 1 ml de cette eau et l’introduire dans 9 ml
d’eau distillée stérile.

Matériels et méthodes
61
Prélever aseptiquement 1 ml de la suspension mère à l’aide d’une pipette
graduée stérile de 1 ml munie d’une poire à aspiration ; l’homogénéisation du
prélèvement se fait après aspiration est refoulement 3 fois.
Transférer aseptiquement le 1 ml prélevé dans le 1er tube (10-1), la pipette ne
devrait pas pénétrer dans les 9 ml du diluant.
Jeter la pipette utilisée dans un conteneur approprié. A l’aide d’une 2ème pipette
stérile de 1 ml, procéder du même du tube 10-1 au tube 10-2.
Faire de même pour les deux derniers tubes en utilisant à chaque prélèvement
une pipette nouvelle.
On peut naturellement utiliser des volumes différents en respectant le facteur de
dilution souhaité.
6. Dénombrement des germes aérobies totaux
Ce dénombrement reflète la qualité microbiologique générale d’un produit et
permet d’en suivre l’évolution. Le nombre de germes « totaux » pour donner une
indication de l’état de décomposition du produit : Il peut aussi dans certains cas
constituer un indice de la qualité sanitaire. Au cours des traitements
technologiques, le dénombrement de la flore « globale » permettra de juger de
l’incidence des diverses opérations (Guiraud, 1998).
La culture en surface d’un milieu gélosé, dite surface count, est recommandée
lorsque la description des colonies peut être utile à l’identification des germes
dénombrés.
La gélose nutritive est un milieu riche permettant le développement de la
plupart des microorganismes. Sa formule comprend une peptone riche en acide
aminés libres et de l’extrait de levure. L’association de ces deux constituants
fournie au milieu de nombreux facteurs de croissance (Multon, 1994).
- On fait fondre le milieu de base et le refroidir jusqu’à 45°c.
- On coule en boites de pétri stériles.

Matériels et méthodes
62
-Après solidification, sécher la surface du milieu à l’étuve à 45°c, couvercle
entrouvert, et laisser refroidir couvercle fermé.
-Marquer les boites (10-1, 10-2,…).
-A partir de la prise d’essai bien homogénéisée, porter aseptiquement 0,1 ml (2
gouttes) dans la boite de pétri contenant la gélose nutritive
- Etaler l’inoculum à l’aide d’un étaleur stérile « pipette râteau » que l’on passe à
la surface de la gélose pendant que l’on imprime à la boite un mouvement
circulaire horizontal.
- Laisser sécher 15 mn à la température du laboratoire.
- Incuber les boites à 37 °c pendant 48 h. La lecture s’effectue par comptage
visuel. Dans tous les cas, seules les boites contenant entre 20 à 300 colonies sont
utilisées.
Lorsque le nombre de colonies est compris entre 20 et 300, on recherche les
germes incriminés en effectuant une identification microbienne basée sur l’étude
microscopique et l’ensemencement d’une galerie biochimique selon la Figure 7.
7. Dénombrement des Coliformes
Les Coliformes totaux regroupent plusieurs espèces bactériennes de la famille
des entérobactéries qui se présentent sous formes de bâtonnet, Gram négatives,
non sporulant, possédant l'enzyme â-galactosidase permettant l'hydrolyse du
lactose à 37 °C afin de produire des colonies rouges avec des reflets métalliques
sur un milieu bien approprie. Le groupe des Coliformes est utilisé depuis la fin
du 19ème siècle comme indicateur de pollution fécale (Lee et al, 2006). La
plupart des espèces de ce groupe se retrouveront naturellement dans le sol ou la
végétation (Edberg et al, 2000).
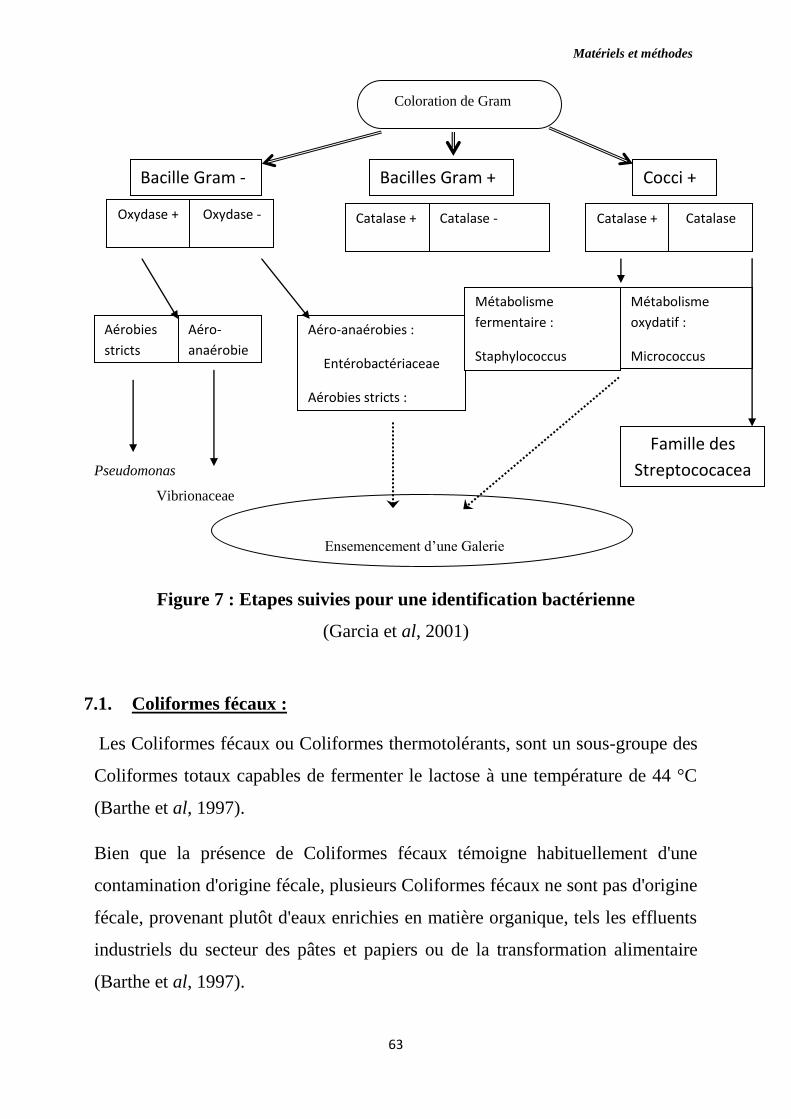
Matériels et méthodes
63
Coloration de Gram
Pseudomonas
Vibrionaceae
Ensemencement d’une Galerie
Figure 7 : Etapes suivies pour une identification bactérienne
(Garcia et al, 2001)
7.1. Coliformes fécaux :
Les Coliformes fécaux ou Coliformes thermotolérants, sont un sous-groupe des
Coliformes totaux capables de fermenter le lactose à une température de 44 °C
(Barthe et al, 1997).
Bien que la présence de Coliformes fécaux témoigne habituellement d'une
contamination d'origine fécale, plusieurs Coliformes fécaux ne sont pas d'origine
fécale, provenant plutôt d'eaux enrichies en matière organique, tels les effluents
industriels du secteur des pâtes et papiers ou de la transformation alimentaire
(Barthe et al, 1997).
Bacille Gram -
Oxydase + Oxydase -
Aérobies
stricts
Aéro-
anaérobie
s
Aéro-anaérobies :
Entérobactériaceae
Aérobies stricts :
Acinetobacter
Bacilles Gram +
Catalase + Catalase -
Bacillus Closridium
Cocci +
Catalase + Catalase
-
Métabolisme
fermentaire :
Staphylococcus
Métabolisme
oxydatif :
Micrococcus
Famille des
Streptococacea
e

Matériels et méthodes
64
L'intérêt de la détection de ces Coliformes, à titre d'organismes indicateurs,
réside dans le fait que leur survie dans l'environnement est généralement
équivalente à celle des bactéries pathogènes et que leur densité est généralement
proportionnelle au degré de pollution produite par les matières fécales (Barthe et
al, 1997).
7.1.1. Colimétrie en milieu liquide
La colimétrie en milieu liquide permet la caractérisation et dénombrement des
Coliformes. Il ne s’agit bien souvent que d’un test présomptif qui demande
confirmation et qui nécessite l’emploi d’un milieu solide de différenciation et
d’isolement.
La colimétrie en milieu liquide consiste en une numération par la méthode NPP.
Le milieu utilisé est le milieu BCPL (bouillon lactosé au pourpre de
bromocrésol) avec cloche (Milieu pour le dénombrement des Coliformes)
[OXOID].
La culture se fait dans des tubes avec cloche de Durham. Les milieux sont
incubés 24 à 48 heures à une température de 37˚C. Le milieu devient trouble et
jaunâtre lors de la pousse bactérienne. Si elle s’accompagne d’une importante
production de gaz, ceci est une présomption de la présence de Coliformes. Le
caractère positif se traduit par un dégagement de gaz de 1/10e au moins du
volume de la cloche.
7.1.2. Numération par la méthode NPP (Nombre le Plus Probable)
Cette méthode est basée sur le fait qu’après ensemencement d’un milieu liquide,
toute croissance microbienne indique la présence d’au moins un germe (UFT :
Unité Formant Trouble).
C’est une estimation statistique du nombre de microorganismes supposés
distribués dans l’échantillon, les bactéries se multiplient librement dans le milieu

Matériels et méthodes
65
liquide. En cas de présence, l’ensemble du milieu liquide inoculé vire à la «
positivité » (trouble ou virage de l’indicateur).
Un jugement quantitatif est possible en jouant sur les volumes de la prise
d’essai.
Le développement peut être apprécié visuellement par dégagement gazeux dans
une cloche de Durham.
Une série de dilutions est effectuée à partir de la suspension à. Une petite
quantité (1 ml) de ces dilutions est placée dans des tubes contenant du bouillon
BCPL (9ml).
Après culture, on procède à la lecture des résultats. Sont comptés positifs les
tubes qui présentent une croissance (dégagement de gaz) et négatifs les autres.
Un test de confirmation peut être réalisé pour vérifier la présence des Coliformes
en incubant une petite quantité de la culture des tubes positifs sur un milieu
sélectif tel que le milieu à l’éosine et au bleu de méthylène de Teague-Levine
(EMB) à 37˚C (Booysen et al, 2002)
Les Coliformes thermotolérants, parfois appelés " Coliformes fécaux " ou
" Escherichia coli présomptifs ", peuvent être recherchés et dénombrés dans les
mêmes conditions mais l’incubation se fait à 44˚C. S’il y a dégagement de gaz,
une confirmation par culture en milieu solide est nécessaire (milieu EMB).
D’autres Entérobactéries (Citrobacter, Klebsiella,...) peuvent donner une
réaction positive. Il conviendra donc de réaliser une identification plus précise
via un isolement sur milieu EMB puis pour confirmation plus précise de réaliser
une galerie API 20 E.
7.2. Dénombrement d’Escherichia Coli
Escherichia coli est un coliforme thermotolérant qui entre autre :
-Produit de l’indole à partir du tryptophane à 44°C,
-Donne un résultat positif à l’essai au rouge de méthyle,
-Ne produit pas de l’acéthyl méthyl carbinol,

Matériels et méthodes
66
- N’utilise pas le citrate comme source unique de carbone.
Pour la recherche d’E. coli, un isolement sur milieu EMB réalisé à partir des
derniers tubes positifs de la colimétrie permet de confirmer la présence de
E. coli dans notre échantillon. L’ensemencement de la gélose EMB se fait par
transfert à l’anse et étalé directement sous formes de stries d’épuisement à la
surface de la gélose. L’incubation dure 24 heures à 44˚C. Le milieu EMB
contient deux colorants, l’éosine et le bleu de méthylène qui inhibent la majeure
partie de la flore Gram+ (sauf Streptocoques D). Bien que les Entérobactéries
lactose - puissent s’y développer, la culture des Entérobactéries lactose + y est
favorisée (Sinton et al, 1999).
Le lactose est un critère de différenciation du milieu : l’utilisation du lactose se
traduit par un centre foncé d’aspect métallique des colonies (lactose +), dans le
cas contraire les colonies sont incolores (lactose -). Les Escherichia Coli
apparaissent comme des colonies isolées, de 2 à 3 mm de diamètre, ayant
tendance à confluer et présentant un reflet métallique verdâtre en lumière
réfléchie et un centre noir pourpre en lumière transmise(Sinton et al, 1999) .
8. Dénombrement des Staphylocoques totaux
L’isolement direct est pratiqué sur le milieu sélectif (Chapman). A l’aide d’une
anse de platine ou d’une pipette Pasteur on prélève 1 g de sable ou 1 ml d’eau du
prélèvement que nous ensemençons par épuisement sur gélose en boîte de Pétri,
de façon à obtenir des colonies bien isolées après une incubation à 37°C pendant
24 heures. L’incubation pour le milieu d’enrichissement peut être faite pendant
24 heures et plus lorsque cela est nécessaire (Sinton et al, 2002).
9. Dénombrement des Pseudomonas totaux
On ensemence 1 g de sable (1 ml pour l’eau) sur des boites de Pétri contenant le
milieu King A et King B on coule une deuxième couche de ces milieux après
ensemencement et on incube à 30°C pendant 72H (Biely et al, 1996).

Matériels et méthodes
67
10. Tests de confirmation et d’identification
Des tests de confirmation d’identification peuvent être réalisés pour toutes les
souches obtenues en vue de confirmer notre présomption d’identification.
10.1 .Caractérisation d’enzymes :
10.1.1. Mise en évidence d’enzymes respiratoires : Test cytochrome
oxydase
Les oxydases interviennent à la fin des étapes de déshydrogénation et des
chaînes de cytochromes. Il en existe plusieurs types. Elles sont mises en
évidence par leur propriété de catalyser la réaction d’oxydation d’un substrat
organique par l’oxygène de l’air. Le substrat utilisé est le N-diméthyl
paraphénylène diamine, qui est incolore sous forme réduite et rouge-violet sous
forme oxydée ou le chlorohydrate de tétraméthyl paraphénylène diamine qui
donne une coloration pourpre. Le caractère oxydase + signifie que la souche
possède une enzyme capable d’oxyder le substrat employé et ne signifie pas la
présence d’une oxydase particulière. Le test est réalisé sur papier filtre" OX ",
imprégnés de réactif. La colonie est prélevée et déposée sur le papier filtre. Une
coloration bleue est considérée comme positive. Toutes les entérobactéries sont
cytochrome oxydase négative.
10.1.2. Test catalase
La catalase est une enzyme catalysant la dismutation de l'eau oxygénée
(peroxyde d'hydrogène).Cette enzyme est utilisée en bactériologie systématique
pour l'identification des bactéries. Il s'agit de mettre en contact une colonie de la
bactérie à étudier en présence d'eau oxygénée.
Ce test est important pour la première orientation dans l'identification d'une
souche pure bactérienne.

Matériels et méthodes
68
Pour effectuer l'identification complète d'une bactérie il faut connaître le type
respiratoire.Il faudra donc poursuivre l'identification de la souche par
l'ensemencement de milieux: sélectifs, hostiles, ou faisant partie de la batterie de
test prévue pour la différenciation des genres et l'identification de la bactérie.
La recherche de la catalase permet de différencier:
-Les bactéries des genres Staphylococcus et Micrococcus ( catalase +) ;
-Des bactéries des genres Enterococcus, Streptococcus, Pediococcus,
Lactococcus et Leuconostoc (catalase -).
Sur une lame de verre propre, déposer une goutte d’eau oxygénée, puis la mettre
en contact avec une colonie isolée, prélevée directement avec une pipette
Pasteur boutonnée ou une anse plastique à usage unique. Il ne faut pas utiliser
une anse en métal car elle serait alors oxydante.
Une effervescence (dû à un dégagement de dioxygène) signe la présence d'une
catalase.
S’il y a une formation de bulles, la bactérie possède la catalase.
Si rien n'est observable, la bactérie ne possède pas l'enzyme.
10.2. Coloration de Gram
La coloration différentielle la plus connue est celle de Gram, qui permet de
diviser les bactéries en deux grands groupes : Gram + et Gram -. Cette méthode
de coloration est basée sur la différence de structure de paroi chez les 2
groupes : les bactéries Gram + contiennent une forte proportion de lipides (20%)
et retiennent le violet de gentiane (premier colorant) après lavage à l’alcool ; les
bactéries Gram - contiennent une faible proportion de lipides, sont décolorées à
l’alcool et prennent ensuite la couleur du second colorant.
Le traitement par l’alcool extrait les lipides chez les Gram – entrainant une
décoloration des organismes au premier colorant qui vont alors prendre la
coloration du second colorant (rouge). Les parois Gram + au contraire sont
déshydratées par l’alcool : leur perméabilité diminue, le premier colorant ne peut

Matériels et méthodes
69
être extrait et elles gardent donc la coloration du premier colorant (bleu).
(Genebio, 2006)
Les souches vont subir une identification plus précise via des galeries Api 20 E.
10.3. Antibiogramme
Un antibiogramme est une technique de laboratoire visant à tester la sensibilité
d’une souche bactérienne vis-à-vis d’un ou plusieurs antibiotiques. Le principe
consiste à placer la culture de bactéries en présence du ou des antibiotiques à
tester et à observer les conséquences sur le développement et la survie de celle-
ci.
On place alors plusieurs pastilles d’antibiotiques sur une souche bactérienne
étalée dans une boite de Mueller-Hinton (Figure 9) (Ce milieu est une gélose
riche pour tester l’action des différents antibiotiques). Il existe 3 types
d’interprétation selon le diamètre du cercle qui entoure le disque d’antibiotique :
Souche sensible, intermédiaire ou résistante. Ce diamètre représente la zone
d’inhibition de l’antibiotique sur la souche donnée. Il représente la concentration
minimale inhibitrice (CMI) de l’antibiotique sur cette souche.
Le diamètre de la zone d’inhibition de l’antibiotique correspond au CMI ou
concentration minimale inhibitrice.
Nous recherchons via l’antibiogramme les résistances à ces antibiotiques ainsi
que la présence de β-lactamase à spectre étendu (BLSE). Ce terme désigne les
enzymes β-lactamases produites surtout par Klebsiella sp. et E. coli, codant pour
la résistance aux β-lactamines à large spectre, qui sont habituellement actives
contre les bacilles à Gram négatif.
10.3.1. Galerie Api 20E [Biomérieux]
Galerie de 20 microtubes prêts à l’emploi permettant de réaliser 23 tests
biochimiques afin d’identifier des bacilles Gram - appartenant à la famille des
Enterobateriaceae (Figure 8).

Matériels et méthodes
70
10.3.1.1. Préparation de l’inoculum
Nous isolons une colonie bien isolée ou nous prélevons une souche pure et nous
l’introduisons dans 5 ml d’eau distillée stérile.
Ensemencement de la galerie : nous introduisons la suspension bactérienne dans
chaque tube à l’aide d’une pipette Pasteur stérile, pointe appuyée à l’intérieur et
sur le côté pour éviter la formation de bulles.
Figure 8 : Galerie biochimique API E20
L’incubation dure 24 heures et la lecture permet d’identifier la souche.
Pour certains caractères, il faut remplir le tube et la cupule (CIT, VP, GEL).
Pour d’autres caractères, il faut remplir le tube et recouvrir d’huile de paraffine
(ADH,LDC, ODC, H2S, URE)(conditions anaérobies).
Figure 9 : Dépôt de disque d’ATB sur milieu MH.
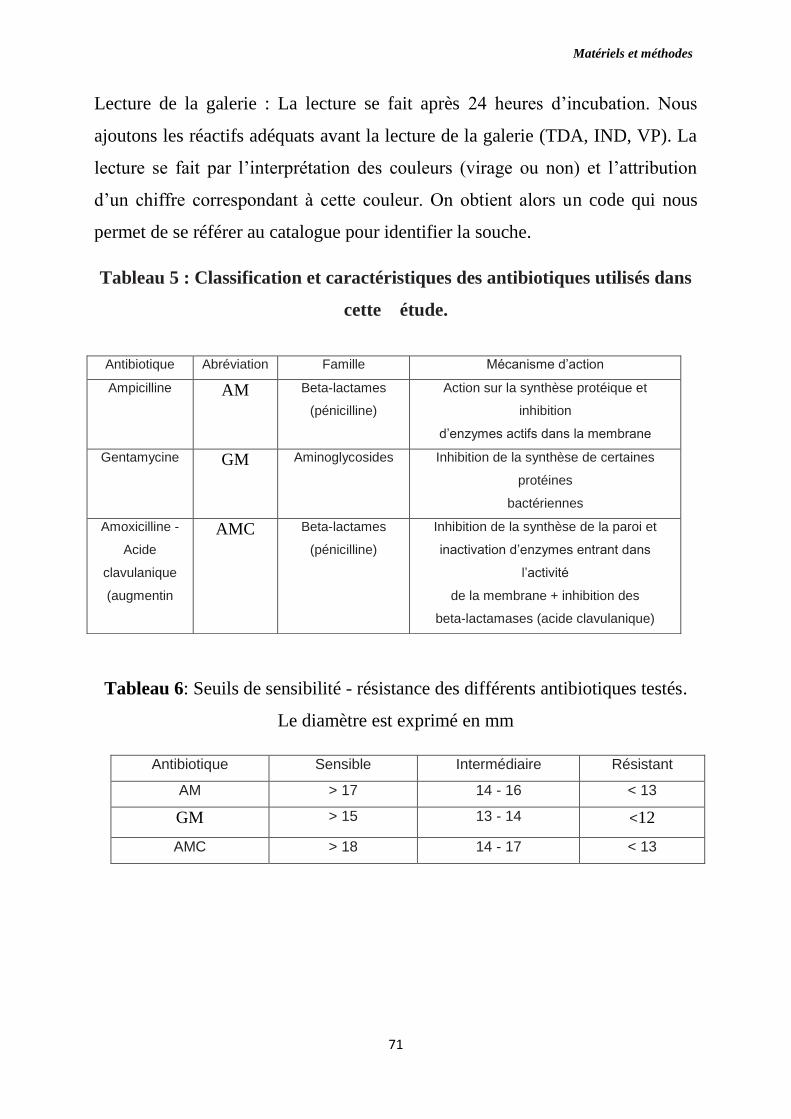
Matériels et méthodes
71
Lecture de la galerie : La lecture se fait après 24 heures d’incubation. Nous
ajoutons les réactifs adéquats avant la lecture de la galerie (TDA, IND, VP). La
lecture se fait par l’interprétation des couleurs (virage ou non) et l’attribution
d’un chiffre correspondant à cette couleur. On obtient alors un code qui nous
permet de se référer au catalogue pour identifier la souche.
Tableau 5 : Classification et caractéristiques des antibiotiques utilisés dans
cette étude.
Tableau 6: Seuils de sensibilité - résistance des différents antibiotiques testés.
Le diamètre est exprimé en mm
Antibiotique Sensible Intermédiaire Résistant
AM > 17 14 - 16 < 13
GM > 15 13 - 14 <12
AMC > 18 14 - 17 < 13
Antibiotique Abréviation Famille Mécanisme d’action
Ampicilline AM Beta-lactames
(pénicilline)
Action sur la synthèse protéique et
inhibition
d’enzymes actifs dans la membrane
Gentamycine GM Aminoglycosides Inhibition de la synthèse de certaines
protéines
bactériennes
Amoxicilline -
Acide
clavulanique
(augmentin
AMC Beta-lactames
(pénicilline)
Inhibition de la synthèse de la paroi et
inactivation d’enzymes entrant dans
l’activité
de la membrane + inhibition des
beta-lactamases (acide clavulanique)
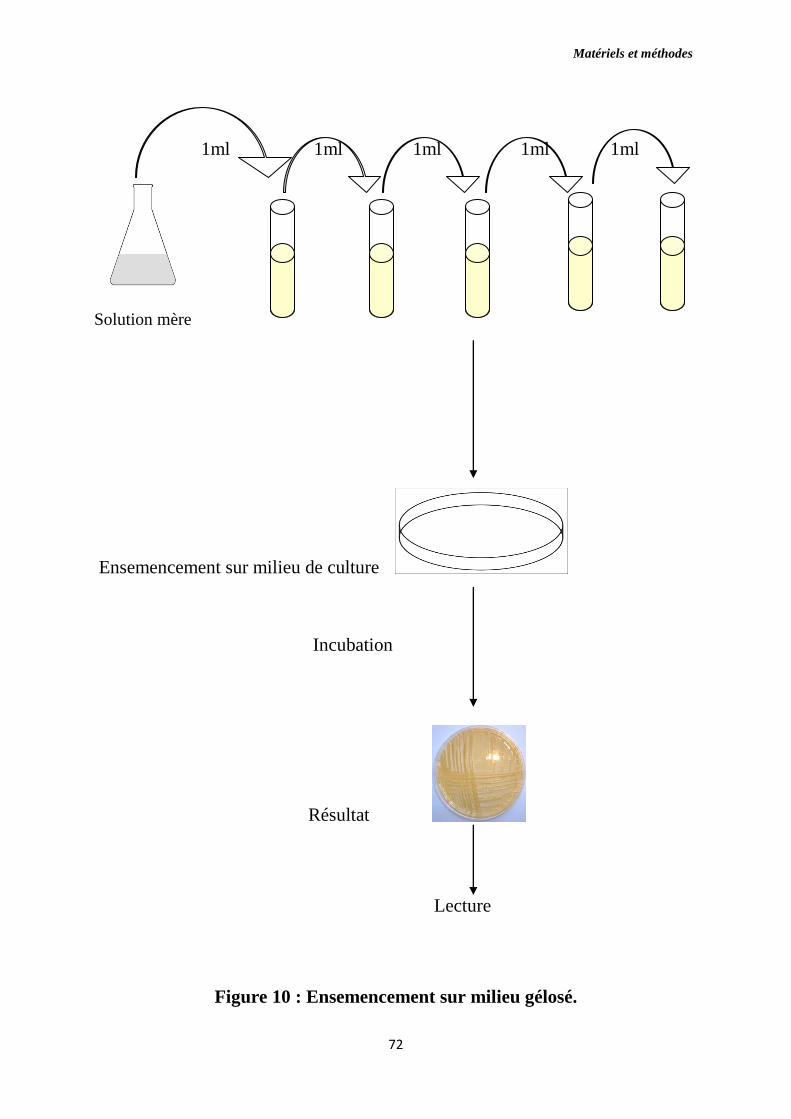
Matériels et méthodes
72
1ml 1ml 1ml 1ml 1ml
Solution mère
Ensemencement sur milieu de culture
Incubation
Résultat
Lecture
Figure 10 : Ensemencement sur milieu gélosé.
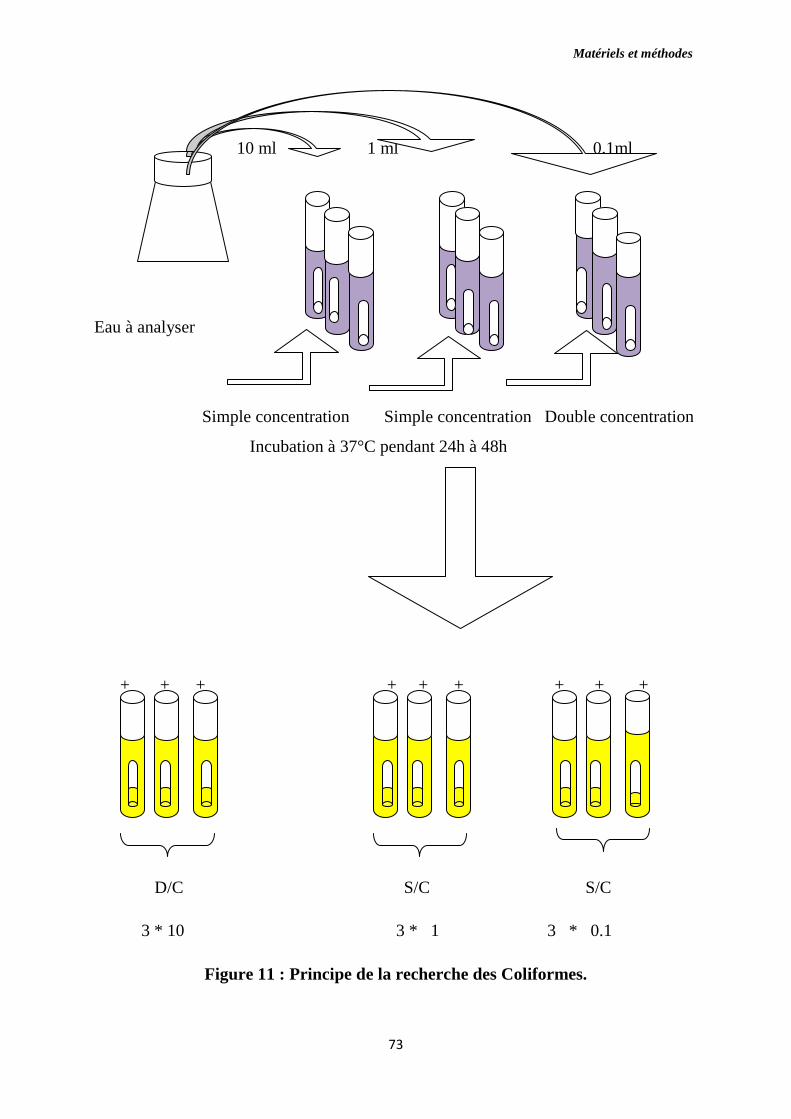
Matériels et méthodes
73
10 ml 1 ml 0.1ml
Eau à analyser
Simple concentration Simple concentration Double concentration
Incubation à 37°C pendant 24h à 48h
+ + + + + + + + +
D/C S/C S/C
3 * 10 3 * 1 3 * 0.1
Figure 11 : Principe de la recherche des Coliformes.
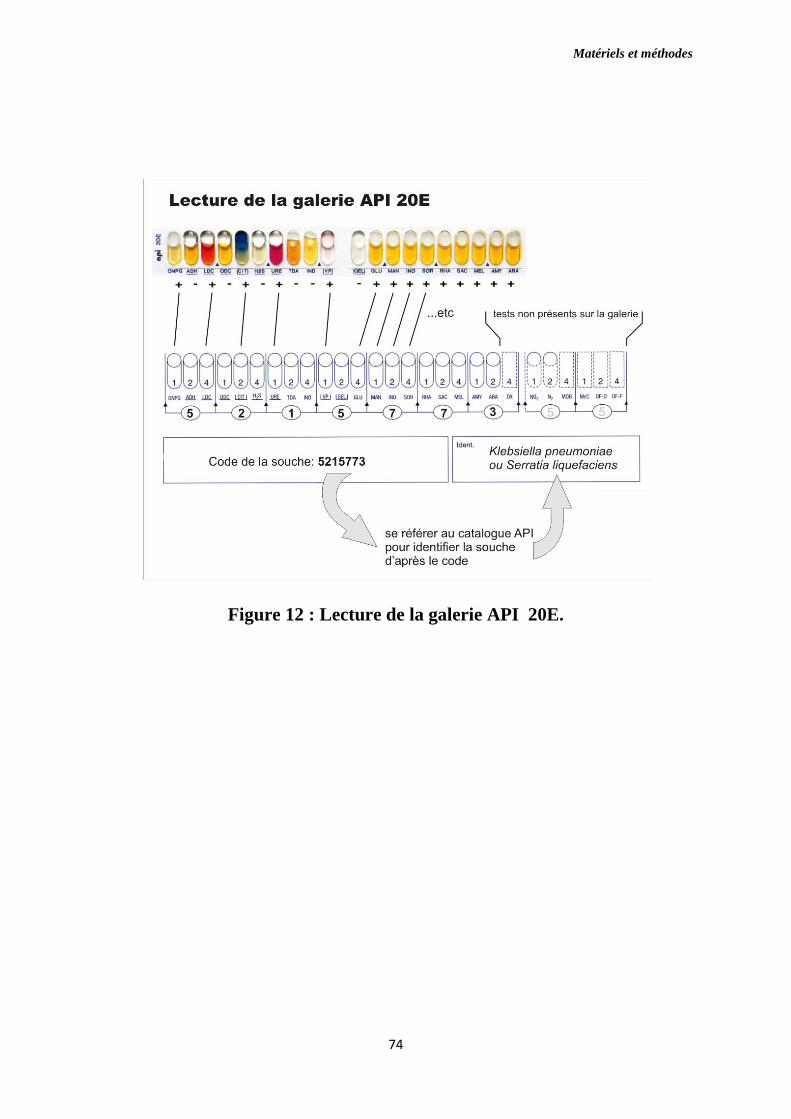
Matériels et méthodes
74
Figure 12 : Lecture de la galerie API 20E.

Résultats et discussion
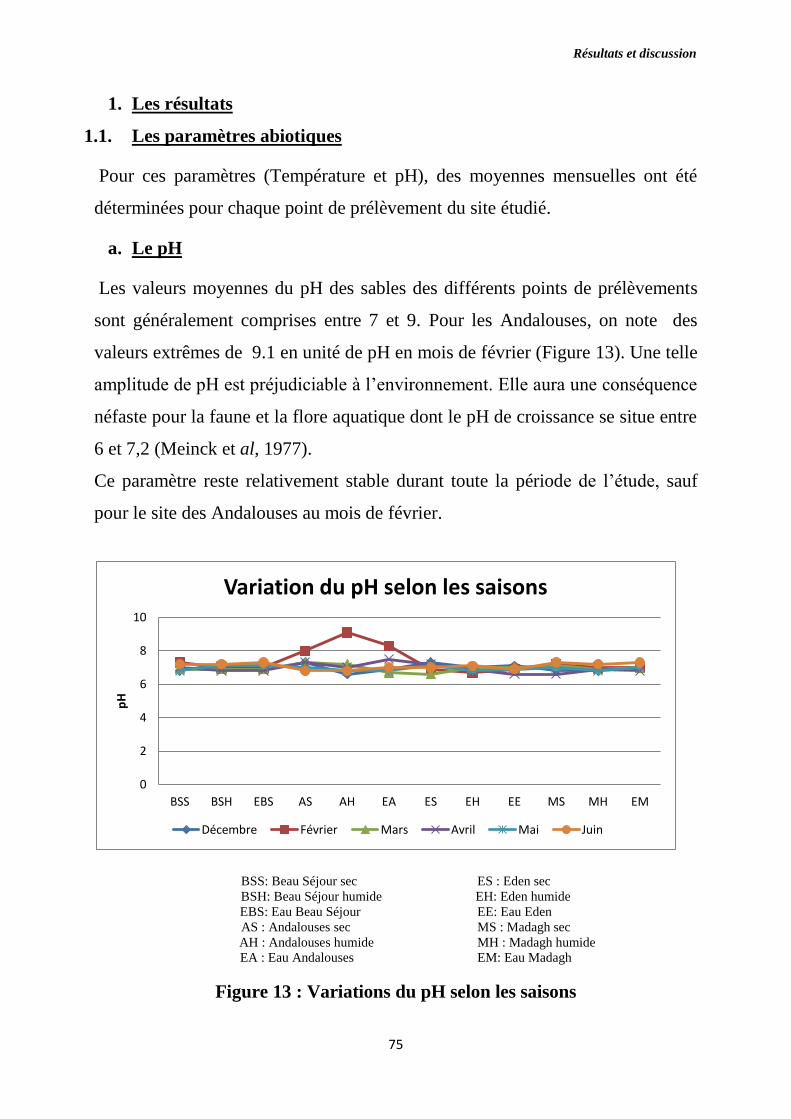
Résultats et discussion
75
1. Les résultats
1.1. Les paramètres abiotiques
Pour ces paramètres (Température et pH), des moyennes mensuelles ont été
déterminées pour chaque point de prélèvement du site étudié.
a. Le pH
Les valeurs moyennes du pH des sables des différents points de prélèvements
sont généralement comprises entre 7 et 9. Pour les Andalouses, on note des
valeurs extrêmes de 9.1 en unité de pH en mois de février (Figure 13). Une telle
amplitude de pH est préjudiciable à l’environnement. Elle aura une conséquence
néfaste pour la faune et la flore aquatique dont le pH de croissance se situe entre
6 et 7,2 (Meinck et al, 1977).
Ce paramètre reste relativement stable durant toute la période de l’étude, sauf
pour le site des Andalouses au mois de février.
BSS: Beau Séjour sec ES : Eden sec
BSH: Beau Séjour humide EH: Eden humide
EBS: Eau Beau Séjour EE: Eau Eden
AS : Andalouses sec MS : Madagh sec
AH : Andalouses humide MH : Madagh humide
EA : Eau Andalouses EM: Eau Madagh
Figure 13 : Variations du pH selon les saisons
0
2
4
6
8
10
BSS BSH EBS AS AH EA ES EH EE MS MH EM
pH
Variation du pH selon les saisons
Décembre Février Mars Avril Mai Juin
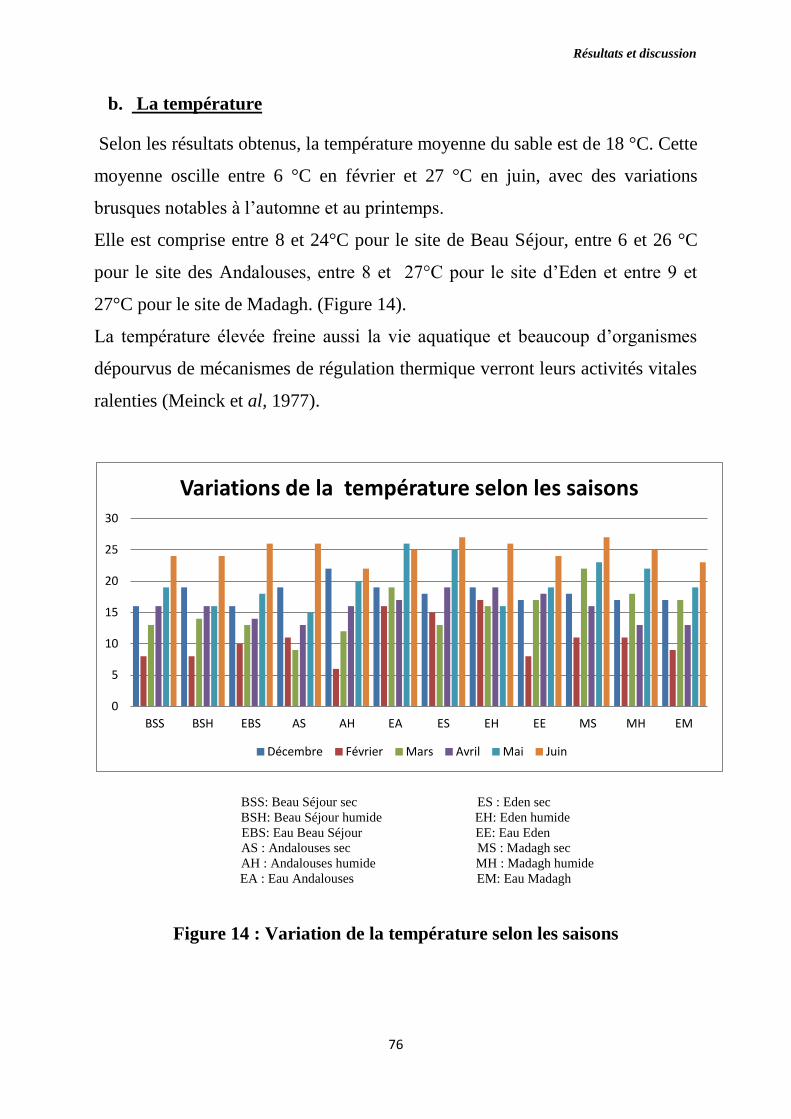
Résultats et discussion
76
b. La température
Selon les résultats obtenus, la température moyenne du sable est de 18 °C. Cette
moyenne oscille entre 6 °C en février et 27 °C en juin, avec des variations
brusques notables à l’automne et au printemps.
Elle est comprise entre 8 et 24°C pour le site de Beau Séjour, entre 6 et 26 °C
pour le site des Andalouses, entre 8 et 27°C pour le site d’Eden et entre 9 et
27°C pour le site de Madagh. (Figure 14).
La température élevée freine aussi la vie aquatique et beaucoup d’organismes
dépourvus de mécanismes de régulation thermique verront leurs activités vitales
ralenties (Meinck et al, 1977).
BSS: Beau Séjour sec ES : Eden sec
BSH: Beau Séjour humide EH: Eden humide
EBS: Eau Beau Séjour EE: Eau Eden
AS : Andalouses sec MS : Madagh sec
AH : Andalouses humide MH : Madagh humide
EA : Eau Andalouses EM: Eau Madagh
Figure 14 : Variation de la température selon les saisons
0
5
10
15
20
25
30
BSS BSH EBS AS AH EA ES EH EE MS MH EM
Variations de la température selon les saisons
Décembre Février Mars Avril Mai Juin
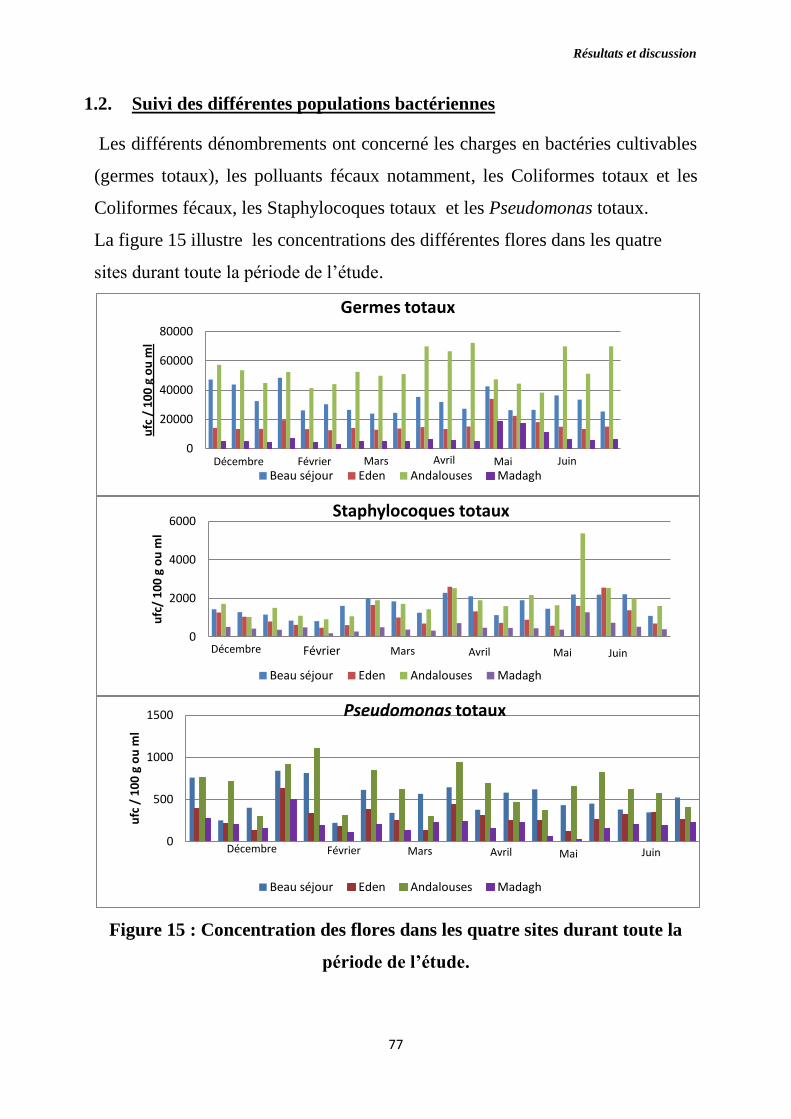
Résultats et discussion
77
1.2. Suivi des différentes populations bactériennes
Les différents dénombrements ont concerné les charges en bactéries cultivables
(germes totaux), les polluants fécaux notamment, les Coliformes totaux et les
Coliformes fécaux, les Staphylocoques totaux et les Pseudomonas totaux.
La figure 15 illustre les concentrations des différentes flores dans les quatre
sites durant toute la période de l’étude.
Figure 15 : Concentration des flores dans les quatre sites durant toute la
période de l’étude.
0
20000
40000
60000
80000
ufc
/ 1
00
g o
u m
l
Germes totaux
Beau séjour Eden Andalouses Madagh Décembre Février Mars
Avril
Mai Juin
0
2000
4000
6000
ufc
/ 1
00
g o
u m
l
Staphylocoques totaux
Beau séjour Eden Andalouses Madagh
Décembre Février Mars Avril Mai Juin
0
500
1000
1500
ufc
/ 1
00
g o
u m
l
Pseudomonas totaux
Beau séjour Eden Andalouses Madagh
Décembre Février Mars Avril Mai Juin
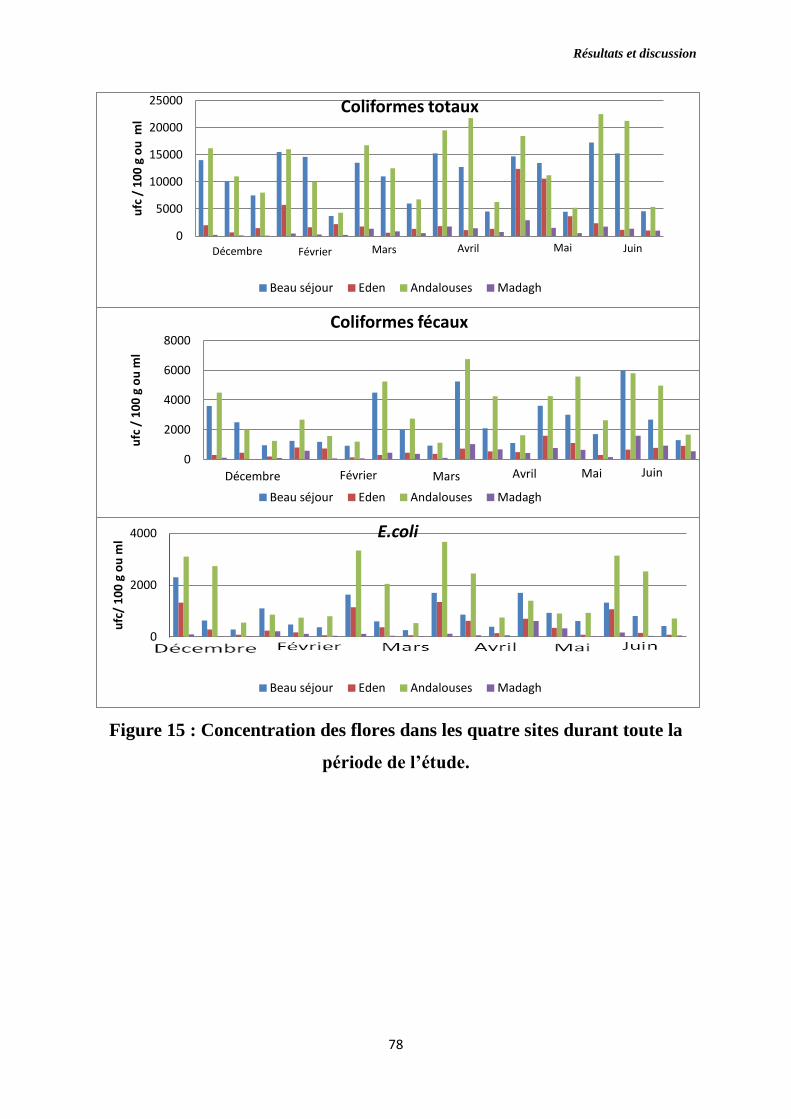
Résultats et discussion
78
Figure 15 : Concentration des flores dans les quatre sites durant toute la
période de l’étude.
0
5000
10000
15000
20000
25000
ufc
/ 1
00
g o
u m
l
Coliformes totaux
Beau séjour Eden Andalouses Madagh
Décembre Février Mars Avril Mai Juin
0
2000
4000
6000
8000
ufc
/ 1
00
g o
u m
l
Coliformes fécaux
Beau séjour Eden Andalouses Madagh
Décembre Février Mars Avril Mai Juin
0
2000
4000
ufc
/ 1
00
g o
u m
l
E.coli
Beau séjour Eden Andalouses Madagh
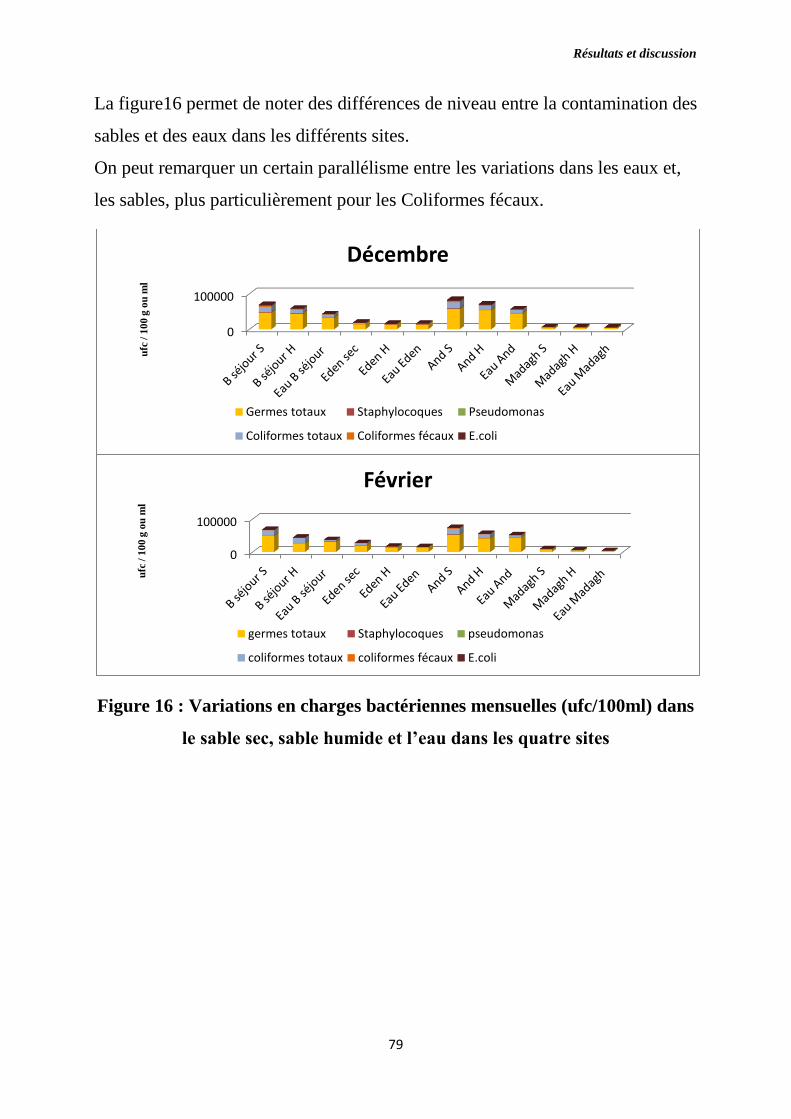
Résultats et discussion
79
La figure16 permet de noter des différences de niveau entre la contamination des
sables et des eaux dans les différents sites.
On peut remarquer un certain parallélisme entre les variations dans les eaux et,
les sables, plus particulièrement pour les Coliformes fécaux.
Figure 16 : Variations en charges bactériennes mensuelles (ufc/100ml) dans
le sable sec, sable humide et l’eau dans les quatre sites
0
100000
ufc
/ 1
00
g o
u m
l
Décembre
Germes totaux Staphylocoques Pseudomonas
Coliformes totaux Coliformes fécaux E.coli
0
100000
ufc
/ 1
00
g o
u m
l
Février
germes totaux Staphylocoques pseudomonas
coliformes totaux coliformes fécaux E.coli

Résultats et discussion
80
Figure 16 : Variations en charges bactériennes mensuelles (ufc/100ml) dans le sable sec,
sable humide et l’eau dans les quatre sites.
0
100000
ufc
/ 1
00
g o
u m
l
Mars
germes totaux Staphylocoques pseudomonas
coliformes totaux coliformes fécaux E.coli
0
200000
Ta
ux d
e g
erm
es
/10
0 g
ou
ml
Avril
germes totaux Staphylocoques pseudomonas
coliformes totaux coliformes fécaux E.coli
0
100000
Ta
ux d
e g
erm
es
/10
0g
ou
ml
Mai
germes totaux Staphylocoques Pseudomonas
coliformes totaux coliformes fécaux E.coli
0
200000
Ta
ux d
e g
erm
es
/ 1
00
g o
u m
l
Juin
germes totaux Staphylocoques pseudomonas
coliformes totaux coliformes fécaux E.coli

Résultats et discussion
81
1.2.1 Estimation des bactéries mésophiles cultivables (germes totaux)
Les résultats relatifs aux germes totaux ont révélé des charges assez importantes
pour les différents types de prélèvements. Cependant, les sables secs sont
plusieurs fois plus chargés en ces bactéries que l’eau (Figure15).
Le traitement statistique de ces résultats a révélé une corrélation positive entre
ces charges bactériennes dans le sable sec et la température, pour les quatre
sites ; Beau Séjour, Eden, Andalouses et Madagh (respectivement r = 0,71 ; r=
0,26 ; r= 0,60 et r= 0,72), d’une part, et avec les charges en polluants fécaux,
pour les Coliformes fécaux (r = 0,69 ;r= 0,46 ; r= 0,31 et r= 0,26) et pour les E.
coli (r = 0,48 ; r=0,42 ; r= 0,61 et r= 0,24 ).
Cette corrélation expliquerait l’apport important en polluants fécaux par le
déversement en provenance du rejet des eaux usées en période pluvieuse (mois
d’avril et juin) (Figure 16).
Les densités maximales des germes totaux sont enregistrées au niveau du sable
sec au niveau du site des Andalouses (7,2.107
ufc /100ml) en mois de mai, alors
que les abondances minimales sont notées au niveau de l’eau de mer au niveau
du site de Madagh (Figure 16), légèrement polluée (3,10 4)
ufc 100/ml.
Par ailleurs, une corrélation positive a été révélée entre la température et la
charge en germes totaux, dans les eaux de mer dans les quatre sites
(respectivement, r = 0,31 ; r = 0,61 ; r=0,77 et r=0,48).
1.2.2. Estimation des polluants fécaux
Les résultats relatifs à ces indicateurs fécaux ont révélé que les sables secs
(moyennes mensuelles plus importantes que dans l’eau) notamment en périodes
automnale et hivernale (Figure15). Le taux maximal des Coliformes fécaux a été
enregistré sur le site des Andalouses dans le sable sec d’une valeur de 6,7 .10
4
ufc / 100 g aussi l’abondance minimale en Coliformes fécaux a été noté au
niveau du sable humide dans le site de Madagh en décembre 30 ufc /100 g
(Figure 16).

Résultats et discussion
82
Cependant, les variations saisonnières, notamment pour les E. coli, sont peu
importantes pour les différents compartiments étudiés.
Les Coliformes fécaux dans le sable sec sont significativement corrélés avec la
température pour les sites de Beau Séjour, Andalouses et Madagh (r= 0,79 ;
r=0,27 et r= 0,48), cette corrélation est négative et non significative pour le site
Eden (r= -0,012).
Les E. coli présentent une corrélation positive avec la température pour les
sites de Beau Séjour, Eden et Madagh (respectivement r = 0,17 ; 0,27 ; 0,22).
Pour le site des Andalouses la corrélation est négative et faiblement significative
(r= -0,22).
Une corrélation significativement positive est établie entre les E. coli et les
Coliformes fécaux dans le sable sec sauf pour le site d’Eden où la corrélation
est significativement négative (r= -0,51).
E. coli représente plus de 90 % des Coliformes présents dans les excréments
humains, le reste étant constitué de Klebsiella sp, d’Enterobacter sp. et de
Cirobacter sp. (Dufour, 1977).
1.2.3. Estimation des Staphylocoques totaux
La figure 16 illustre les résultats relatifs aux variations en Staphylocoques totaux
au niveau des différents sites ; On a estimé une densité maximale en
Staphylocoques en mois de mai enregistrée dans l’eau de mer du site des
Andalouses (5,104
ufc/ml) en mois de mai, une valeur beaucoup moins
importante est notée dans le sable humide du site de Madagh en mois de
février (186 ufc / 100 g).
Les taux estimés ont montré que les sables secs et les sables humides sont plus
concentrés que l’eau notamment en saison estivale (Figure 15).
Par ailleurs, une corrélation positive a été révélée entre la température et la
charge en Staphylocoques, dans les sable sec de la plage du Beau Séjour
(r=0,71) pour les plages des Andalouses, Eden et Madagh une corrélation

Résultats et discussion
83
significativement négative a été révélée entre le taux des Staphylocoques et la
température (respectivement, r = -0,45 ; r =- 0,76 ; r = -0,79).
Les taux en Staphylocoques dans l’eau sont également associés avec la
température (r = 0,48 ; r = 0 ,25 ; r= 0,81 ; r= 0,67).
Les concentrations de Staphylocoques totaux sont corrélées (r=0,47; r = 0,19 ;
r = 0,74 ; r = 0,37) avec les concentrations de Coliformes fécaux (Tableau 7).
Certaines études épidémiologiques ont identifié les Staphylocoques totaux
comme un indicateur potentiel de salubrité dans un contexte de baignade
(Seyfried et al, 1985 ; Cheung et al, 1991). En eau douce, les Staphylocoques
totaux semblent être les indicateurs les plus appropriés pour prédire le taux
d’infection et de maladie parmi les baigneurs (Seyfried et al, 1985). En eau
salée, les Staphylocoques totaux pourraient servir d’indicateurs de la densité des
baigneurs et du risque d’infection croisée parmi ceux-ci plutôt que d’indicateur
de contamination fécale (Cheung et al, 1991).
1.2.4. Estimation des Pseudomonas totaux
L'analyse des résultats obtenus montre une corrélation négative entre les
concentrations des Pseudomonas et la température pour le Beau séjour, Eden,
Andalouses et Madagh (respectivement, r = -0,86 ; -0,45 ; -0,67 ; -0,79)
Une corrélation significativement négative avec les Coliformes fécaux sauf
pour le site des Andalouses où la corrélation est positive (r=0,85). Avec les
bactéries indicatrices de la pollution fécale, la corrélation n'est significativement
positive qu'en milieux très pollués.
1.3. Etude comparative des différents germes dans le sable des quatre
plages
Les concentrations saisonnières des germes totaux, Coliformes fécaux,
Staphylocoques et Pseudomonas sont illustrés dans la figure 15, ainsi, une
comparaison des taux de chaque flore entre les quatre sites dans la figure 16.

Résultats et discussion
84
Les deux endroits les plus contaminés en Coliformes fécaux sont les sites qui
sont sous l’influence des rejets des eaux usées à savoir Beau Séjour et
Andalouses avec des taux qui varient entre 1.104
ufc / 100 g et 6,7 .10 4
ufc
/100g pour les Andalouses et entre 9. 102 ufc /100 g
et 5,8. 10
4 ufc /100g pour
le site de Beau Séjour. Les deux autres sites présentent une pollution moins
importante avec des taux de Coliformes qui varient entre 30 ufc/g et 1,6.103
ufc /g pour le site de Madagh et entre 110 ufc/100g et 1,6.10
2 ufc/g
pour le site
d’Eden.
Les concentrations des Staphylocoques totaux sont beaucoup plus élevées dans
l’eau de mer du site des Andalouses (Figure 16) et les deux endroits les plus
contaminés en Staphylocoques totaux sont les sites qui sont sous l’influence
directe des rejets des eaux usées, c’est-à-dire celui des Andalouses et du Beau
Séjour. D’après les résultats de la présente étude, les eaux usées seraient une
source importante de Staphylocoques, ainsi le nombre des estivants; en effet les
eaux usées traitées mais non désinfectées sont rejetées dans la mer des
Andalouses et du Beau Séjour. Les Staphylocoques colonisent la peau et les
muqueuses en abondance (SBESC, 1992) et se retrouvent également dans les
matières fécales humaines et animales (Seyfried et Harris, 1990), ainsi que dans
l’urine (Cheung et al, 1991). Le milieu aqueux ne constitue pas un milieu
naturel pour les Staphylocoques qui sont en général incapables de s’y multiplier.
Ils ont besoin de beaucoup d’éléments nutritifs et ne peuvent se développer que
dans un milieu dont la température est d’environ 20 °C (SBESC, 1992). Il est
donc probable que les Staphylocoques se retrouvent en concentration importante
dans les sables des plages et possible qu’ils s y multiplient à cause de
l’abondance des éléments nutritifs présents. Malheureusement, on connaît peu
l’importance de certaines sources potentielles, tels les animaux et les eaux de
ruissellement (SBESC, 1992).
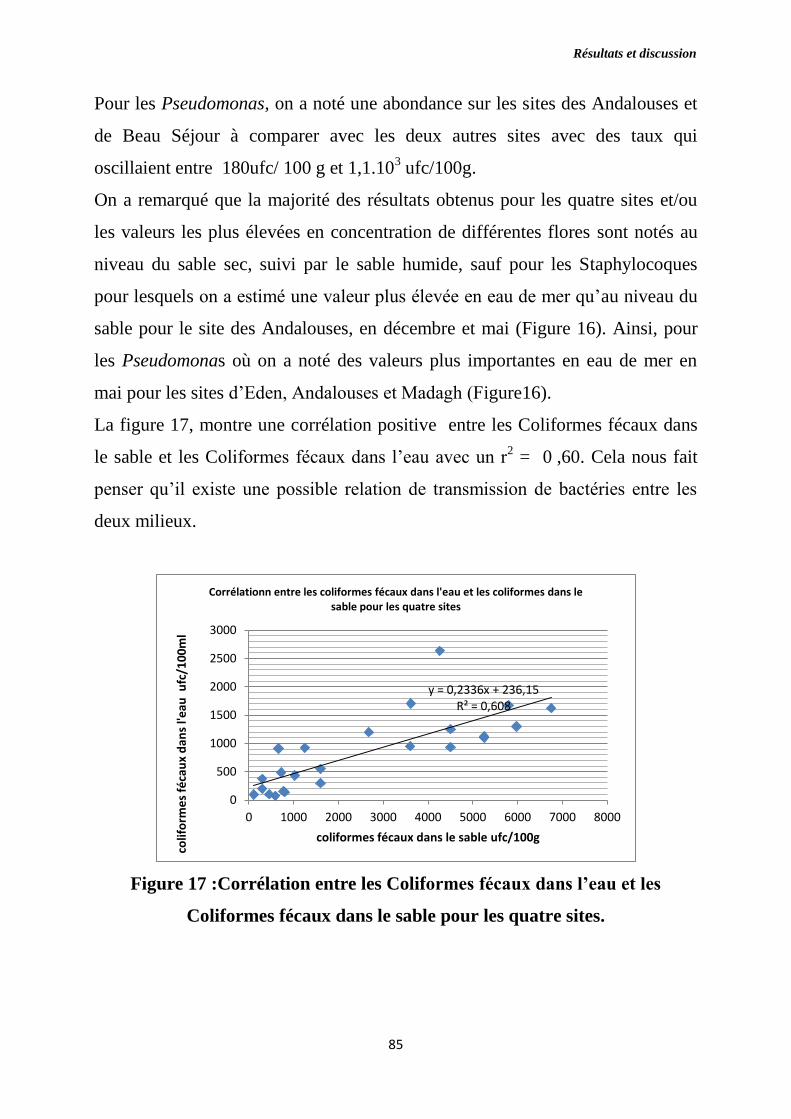
Résultats et discussion
85
Pour les Pseudomonas, on a noté une abondance sur les sites des Andalouses et
de Beau Séjour à comparer avec les deux autres sites avec des taux qui
oscillaient entre 180ufc/ 100 g et 1,1.103 ufc/100g.
On a remarqué que la majorité des résultats obtenus pour les quatre sites et/ou
les valeurs les plus élevées en concentration de différentes flores sont notés au
niveau du sable sec, suivi par le sable humide, sauf pour les Staphylocoques
pour lesquels on a estimé une valeur plus élevée en eau de mer qu’au niveau du
sable pour le site des Andalouses, en décembre et mai (Figure 16). Ainsi, pour
les Pseudomonas où on a noté des valeurs plus importantes en eau de mer en
mai pour les sites d’Eden, Andalouses et Madagh (Figure16).
La figure 17, montre une corrélation positive entre les Coliformes fécaux dans
le sable et les Coliformes fécaux dans l’eau avec un r2 = 0 ,60. Cela nous fait
penser qu’il existe une possible relation de transmission de bactéries entre les
deux milieux.
Figure 17 :Corrélation entre les Coliformes fécaux dans l’eau et les
Coliformes fécaux dans le sable pour les quatre sites.
y = 0,2336x + 236,15 R² = 0,608
0
500
1000
1500
2000
2500
3000
0 1000 2000 3000 4000 5000 6000 7000 8000
colif
orm
es
féca
ux
dan
s l'e
au u
fc/1
00
ml
coliformes fécaux dans le sable ufc/100g
Corrélationn entre les coliformes fécaux dans l'eau et les coliformes dans le sable pour les quatre sites
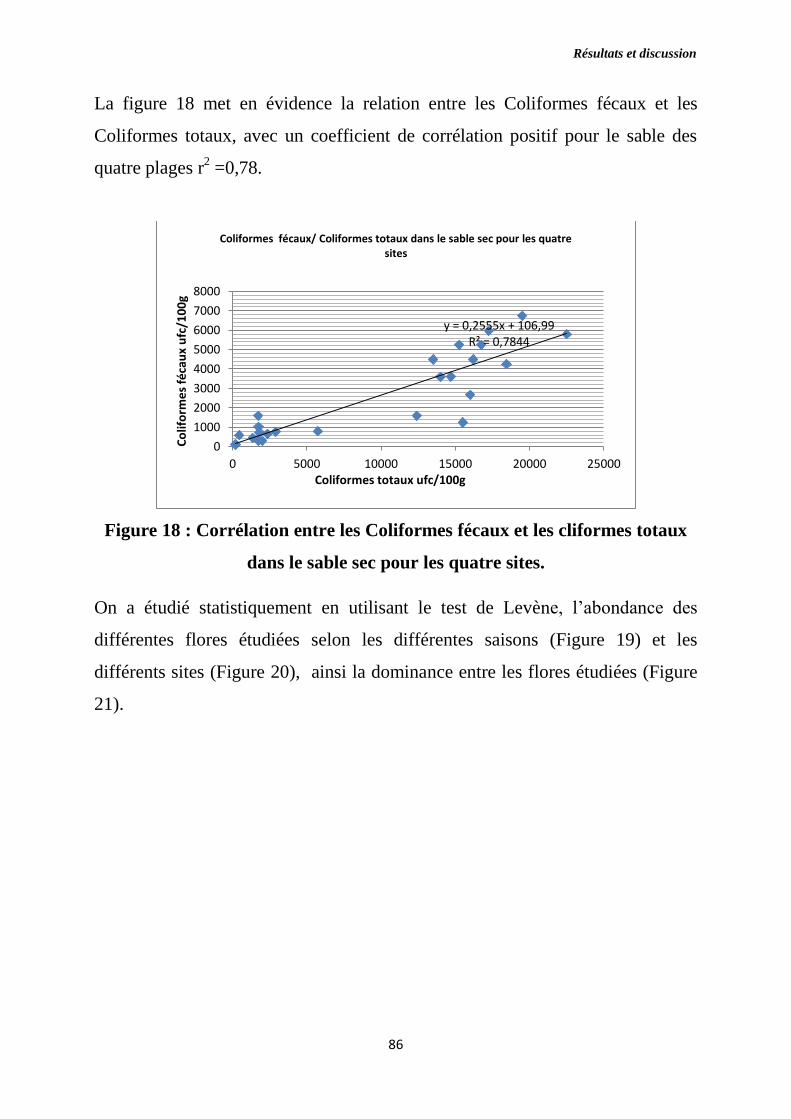
Résultats et discussion
86
La figure 18 met en évidence la relation entre les Coliformes fécaux et les
Coliformes totaux, avec un coefficient de corrélation positif pour le sable des
quatre plages r2 =0,78.
Figure 18 : Corrélation entre les Coliformes fécaux et les cliformes totaux
dans le sable sec pour les quatre sites.
On a étudié statistiquement en utilisant le test de Levène, l’abondance des
différentes flores étudiées selon les différentes saisons (Figure 19) et les
différents sites (Figure 20), ainsi la dominance entre les flores étudiées (Figure
21).
y = 0,2555x + 106,99 R² = 0,7844
0
1000
2000
3000
4000
5000
6000
7000
8000
0 5000 10000 15000 20000 25000
Co
lifo
rme
s fé
cau
x u
fc/1
00
g
Coliformes totaux ufc/100g
Coliformes fécaux/ Coliformes totaux dans le sable sec pour les quatre sites
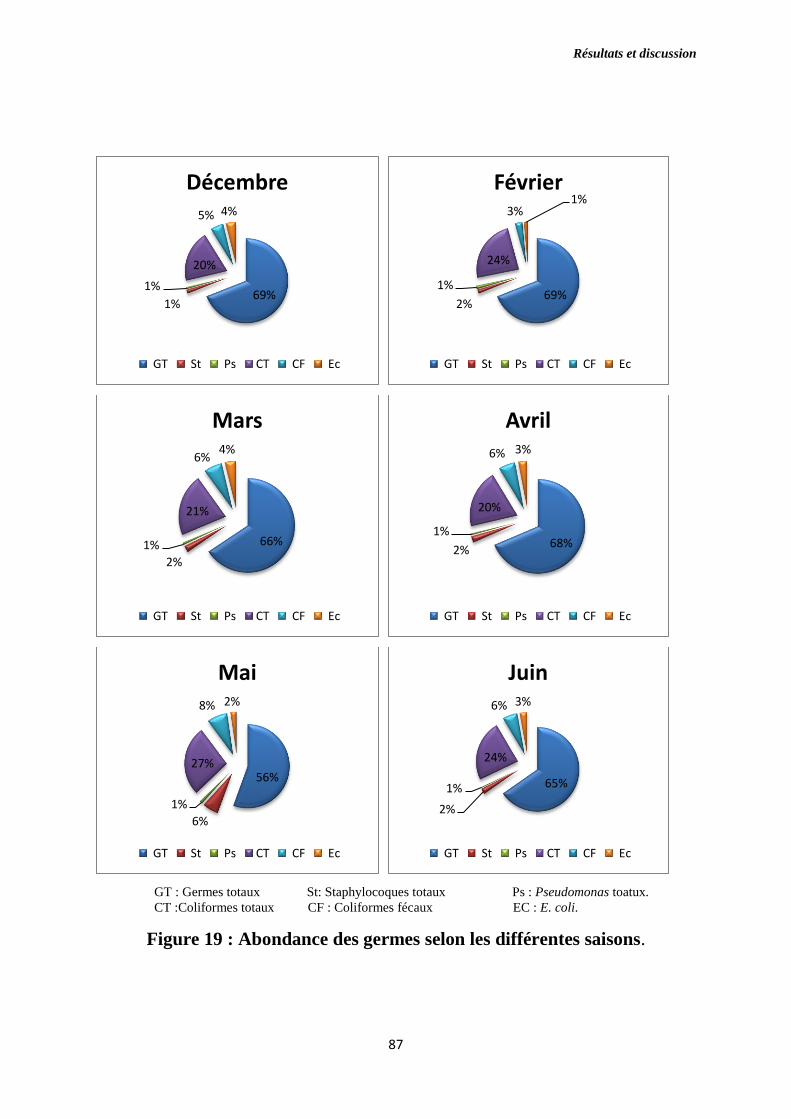
Résultats et discussion
87
GT : Germes totaux St: Staphylocoques totaux Ps : Pseudomonas toatux.
CT :Coliformes totaux CF : Coliformes fécaux EC : E. coli.
Figure 19 : Abondance des germes selon les différentes saisons.
69% 1%
1%
20%
5% 4%
Décembre
GT St Ps CT CF Ec
69% 2%
1%
24%
3% 1%
Février
GT St Ps CT CF Ec
66%
2%
1%
21%
6% 4%
Mars
GT St Ps CT CF Ec
68% 2%
1%
20%
6% 3%
Avril
GT St Ps CT CF Ec
56%
6%
1%
27%
8% 2%
Mai
GT St Ps CT CF Ec
65%
2%
1%
24%
6% 3%
Juin
GT St Ps CT CF Ec

Résultats et discussion
88
Les résultats ont révélé une présence très abondante en germes totaux au mois
de février avec un pourcentage de 69% à comparer aux autres mois où le
pourcentage de cette flore bactérienne présentait un pourcentage moins
important, en mois de mai, les Coliformes totaux (27%), les Coliformes fécaux
(8% ) et les Staphylocoques (6%) ont été plus abondants que dans les autres
mois de prélèvement, E. coli, a dominé au mois de décembre et en mois de
mars avec un pourcentage de 4 %.
Les sites de Beau Séjour et des Andalouses, dominaient les sites d’Eden et de
Madagh en matière d’abondance des flores étudiées (Figure20), le site des
Andalouses comprenait jusqu’à 29% de germes de l’ensemble des flores
bactérienne étudiées, 20% pour le site du Beau Séjour. Les sites de Madagh et
Eden ont présenté des pourcentages moins importants.
La figure 19 illustre la flore la plus dominante à comparer avec les autres flores,
pendant la période de l’étude, ce sont les germes totaux qui ont dominé toutes
les autres flores avec une apparition de 76%, suivi par les Coliformes totaux
14 %.
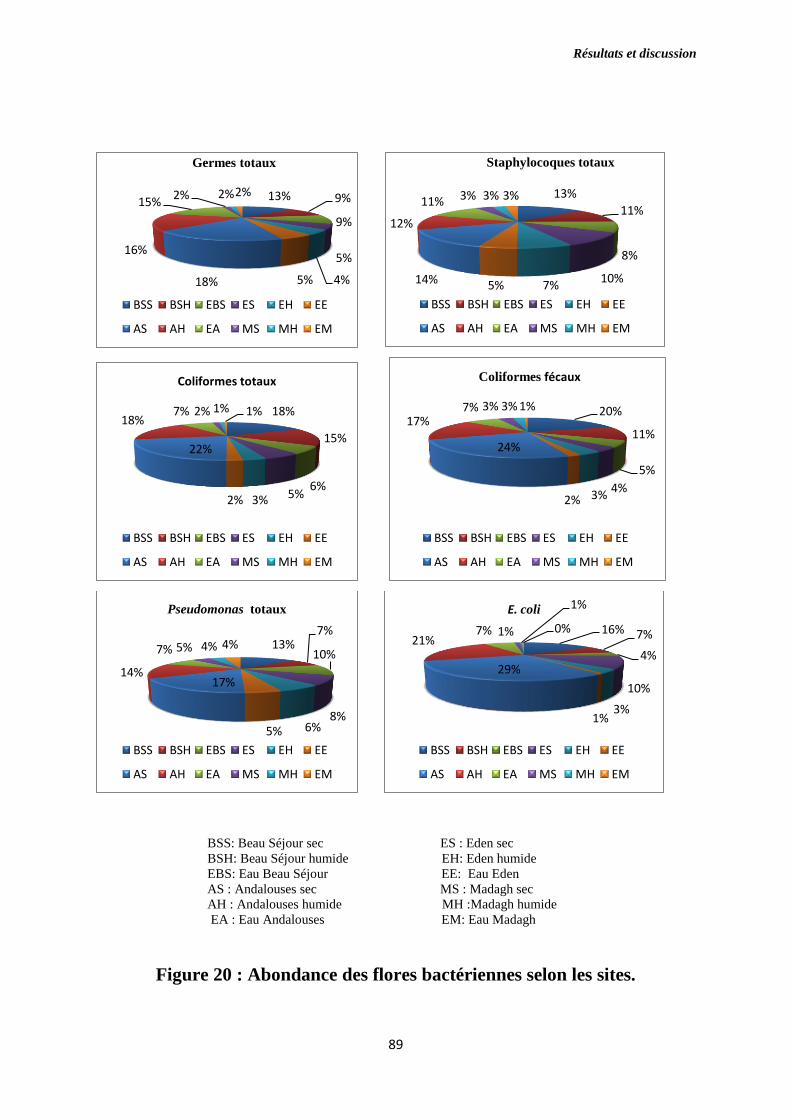
Résultats et discussion
89
BSS: Beau Séjour sec ES : Eden sec
BSH: Beau Séjour humide EH: Eden humide
EBS: Eau Beau Séjour EE: Eau Eden
AS : Andalouses sec MS : Madagh sec
AH : Andalouses humide MH :Madagh humide
EA : Eau Andalouses EM: Eau Madagh
Figure 20 : Abondance des flores bactériennes selon les sites.
13% 9%
9%
5%
4% 5% 18%
16%
15% 2% 2% 2%
Germes totaux
BSS BSH EBS ES EH EE
AS AH EA MS MH EM
18%
15%
6% 5% 3% 2%
22%
18% 7% 2% 1% 1%
Coliformes totaux
BSS BSH EBS ES EH EE
AS AH EA MS MH EM
20%
11%
5%
4% 3% 2%
24%
17% 7% 3% 3% 1%
Coliformes fécaux
BSS BSH EBS ES EH EE
AS AH EA MS MH EM
13% 7%
10%
8% 6% 5%
17% 14%
7% 5% 4% 4%
Pseudomonas totaux
BSS BSH EBS ES EH EE
AS AH EA MS MH EM
16% 7%
4%
10%
3% 1%
29%
21% 7% 1%
1%
0%
E. coli
BSS BSH EBS ES EH EE
AS AH EA MS MH EM
13%
11%
8%
10% 7% 5% 14%
12%
11% 3% 3% 3%
Staphylocoques totaux
BSS BSH EBS ES EH EE
AS AH EA MS MH EM
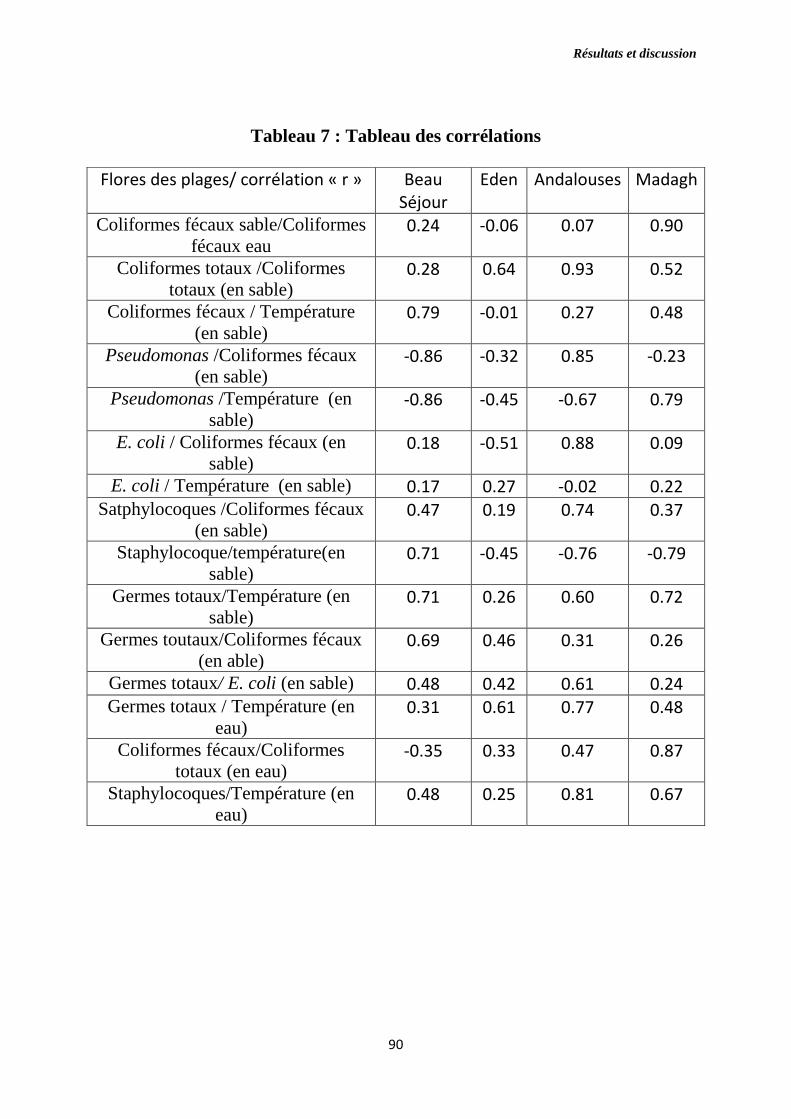
Résultats et discussion
90
Tableau 7 : Tableau des corrélations
Flores des plages/ corrélation « r » Beau Séjour
Eden Andalouses Madagh
Coliformes fécaux sable/Coliformes
fécaux eau 0.24 -0.06 0.07 0.90
Coliformes totaux /Coliformes
totaux (en sable) 0.28 0.64 0.93 0.52
Coliformes fécaux / Température
(en sable) 0.79 -0.01 0.27 0.48
Pseudomonas /Coliformes fécaux
(en sable) -0.86 -0.32 0.85 -0.23
Pseudomonas /Température (en
sable) -0.86 -0.45 -0.67 0.79
E. coli / Coliformes fécaux (en
sable) 0.18 -0.51 0.88 0.09
E. coli / Température (en sable) 0.17 0.27 -0.02 0.22 Satphylocoques /Coliformes fécaux
(en sable) 0.47 0.19 0.74 0.37
Staphylocoque/température(en
sable) 0.71 -0.45 -0.76 -0.79
Germes totaux/Température (en
sable) 0.71 0.26 0.60 0.72
Germes toutaux/Coliformes fécaux
(en able) 0.69 0.46 0.31 0.26
Germes totaux/ E. coli (en sable) 0.48 0.42 0.61 0.24 Germes totaux / Température (en
eau) 0.31 0.61 0.77 0.48
Coliformes fécaux/Coliformes
totaux (en eau) -0.35 0.33 0.47 0.87
Staphylocoques/Température (en
eau) 0.48 0.25 0.81 0.67
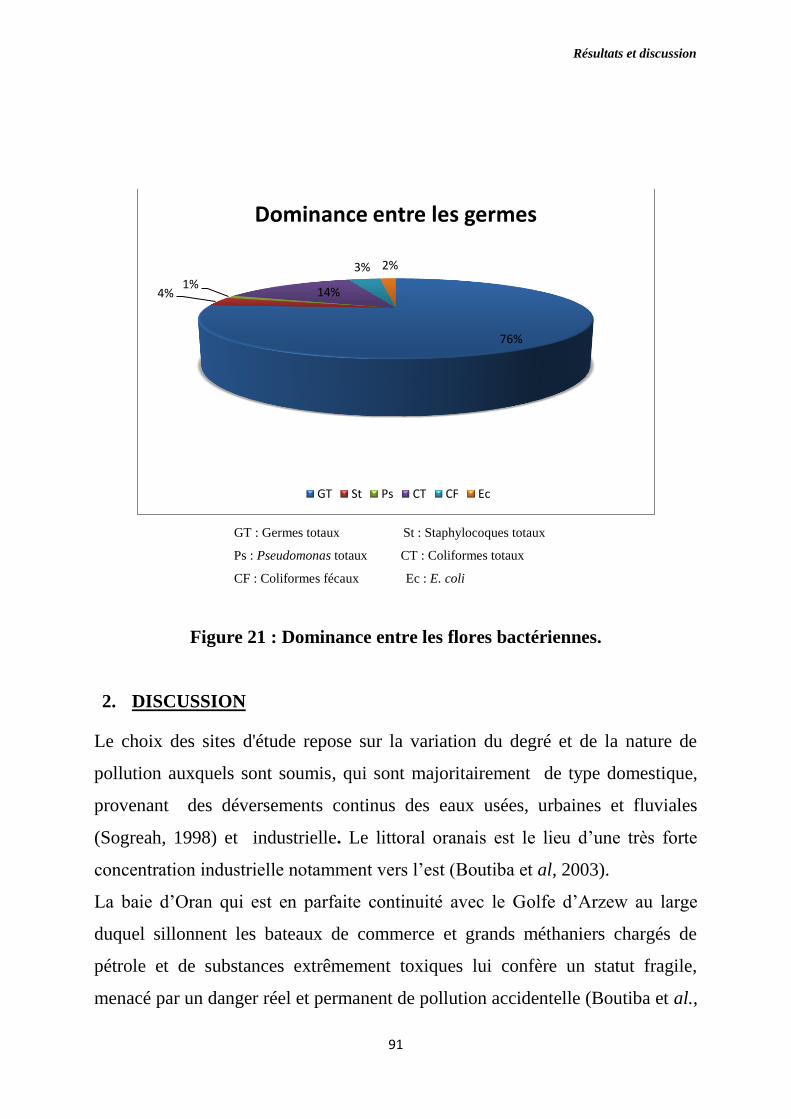
Résultats et discussion
91
GT : Germes totaux St : Staphylocoques totaux
Ps : Pseudomonas totaux CT : Coliformes totaux
CF : Coliformes fécaux Ec : E. coli
Figure 21 : Dominance entre les flores bactériennes.
2. DISCUSSION
Le choix des sites d'étude repose sur la variation du degré et de la nature de
pollution auxquels sont soumis, qui sont majoritairement de type domestique,
provenant des déversements continus des eaux usées, urbaines et fluviales
(Sogreah, 1998) et industrielle. Le littoral oranais est le lieu d’une très forte
concentration industrielle notamment vers l’est (Boutiba et al, 2003).
La baie d’Oran qui est en parfaite continuité avec le Golfe d’Arzew au large
duquel sillonnent les bateaux de commerce et grands méthaniers chargés de
pétrole et de substances extrêmement toxiques lui confère un statut fragile,
menacé par un danger réel et permanent de pollution accidentelle (Boutiba et al.,
76%
4% 1%
14%
3% 2%
Dominance entre les germes
GT St Ps CT CF Ec

Résultats et discussion
92
1996). Une très grande pression et agression par les activités humaines liées aux
industriels des villes côtières, Oran, Arzew, Ghazaouet, …; et des grandes
agglomérations urbaines génèrent une pollution intense caractérisée par les
rejets d’eaux usées. Très rares sont les stations d’épurations fonctionnelles dans
les villes côtières (Boutiba et al, 1996). Tous ces déchets se déversent
directement dans le milieu marin entraînant des effets nuisibles en provoquant
de grands dommages aux ressources biologiques qui induisent un réel danger
pour la santé humaine (Terbeche, 2007).
Ainsi, en se basant sur le rapport CF/CT qui est supérieur à 40% ce qui peut être
expliqué par une pollution bactérienne au niveau des sables de plages d'origine
humaine ou animale.
Les activités, touristique et pastorale peuvent être considérées parmi les
principales sources de contamination de ces sables de plages.
D'après les résultats obtenus, les bactéries mésophiles sont présentes dans
l’ensemble des sites prospectés, mais à des concentrations variables (Figure 15
et Figure 20).Cette flore a dominé les autres flores, ce qui peut être dû à la
facilité de la prolifération de cette flore et son adaptation aux différents milieux
dans les différentes conditions. Avec les germes indicateurs de la pollution
fécale, la corrélation positive statistiquement et significative est enregistrée au
niveau des zones moyennement et très polluées (tableau 7).
Au niveau des sites légèrement pollués, en revanche, les abondances des
Pseudomonas sont comparables à celles des bactéries indicatrices de la
pollution fécale en période hivernale (Figure 16 et Figure 19). Ces résultats
corroborent ceux déjà obtenus pour les analyses de l’eau de mer par certains
auteurs (Araujo et al, 1989; Schubert, 1991; Rhodes & Kator, 1994).ces germes
sont présents dans les eaux de surface, leur nombre peut indiquer la présence de
substances organiques (Zmyslowska et al, 2000). La corrélation entre les
densités des germes indicateurs de la pollution fécale et celles des Pseudomonas
au niveau des sites très pollués suggère que ces deux groupes bactériens sont

Résultats et discussion
93
issus probablement d'une même origine (Figure 20 et tableau 7). Dans ce sens,
Schubert (1991) a considéré l'eau usée comme l'un des principaux réservoirs de
ces bactéries. L'absence de bactéries témoins de contamination fécale ne
présume pas de celle de microorganismes non fécaux à pouvoir pathogène tels
Pseudomonas aeruginosa et Legionella qui devront être recherchées en compte-
tenu du risque spécifique qu'ils représentent par eux mêmes (Schubert ; 1975).
L'analyse de l'évolution de la température en comparaison avec les densités des
différentes flores montre que la majorité des flores étudiées évoluent dans le
même sens, sauf pour les Pseudomonas qui sont considérés comme flore
psychrophile (Figure 15).
Les fortes abondances des germes totaux, Staphylocoques et les indicateurs de
pollution fécale coïncident avec les températures moyennes qui varient entre 19
et 25 °C (Figure 15 et Figure 19). En effet, Le climat de la région d'Oran est de
type méditerranéen, chaud en été (35°C maximum) et doux en hiver (9°C
minimum), avec une saison sèche très marquée entre la mi-juin et la mi-
septembre. Ces conditions sont dues à l'alternance de brise de mer fraîche et
humide et de brise de terre chaude et sèche (Sahnouni, 2003). Ainsi, le climat
peut réagir en réduisant ou en augmentant le nombre de bactéries (Curtis et al,
1992). Nous pensons donc que ce phénomène peut jouer un rôle très important
dans l’énumération des bactéries lors de notre étude.
D'après l'étude de la composition bactériologique des sables, les Coliformes
fécaux sont présent en abondance dans les sables des quatre plages. Cette flore
est très dominante dans les eaux usées et dans les milieux recevant ce type
d'eau. Cependant, au niveau des sables légèrement pollués, le pourcentage de
cette flore diminue remarquablement en les comparants avec les sables subissant
une forte pollution (Figure 16).
En période estivale, la pollution fécale augmente au niveau des sables des
plages étudiées. En période froide, les abondances des Pseudomonas sont plus

Résultats et discussion
94
fortes ; ce qui laisse supposer que la dynamique de ces flores est très
probablement liée à la température du sable (Figure 15).
Jusqu'à présent, la recherche des Pseudomonas n'est jamais prise comme critère
d'intérêt sanitaire pour la détermination de la qualité d’un environnement
(Schubert, 1991). Malgré que ces bactéries soient reconnues responsables de
plusieurs types d'infections et d’altération (Dufour, 1986; Janda et Duffey,
1988).
Vu que la plupart des bactéries soient autochtones des milieux aquatiques, la
contamination de ces milieux par la pollution fécale ne peut qu'accentuer le
risque sanitaire lié à ces germes. En effet, plusieurs auteurs ont démontré la
capacité de multiplication de ces germes en présence de la matière organique
(Monfort et Baleux, 1991; Van der et Kooij, 1991). Suite à cela, le contact avec
des milieux contenant de la matière organique, doit être effectué avec beaucoup
de précaution.
Les résultats relatifs aux dénombrements des populations bactériennes ont
montré que les sables sont relativement plus chargés que l’eau. Ceci refléterait
la survie de ces bactéries dans les sables, ce qui serait conséquent à de fortes
concentrations en matière organique (Dellali et al, 2000; Rozen et Belkin, 2001).
Les auteurs ont expliqué l’augmentation des charges bactériennes sédimentaires
par la richesse en matières organiques issues des eaux usées (Essid et al, 2007).
Selon Laliberte et Grimes (1982), les sédiments représentent un réservoir
d’accumulation de différentes formes bactériennes. Leur effet protecteur pour
les espèces de bactéries pathogènes, a été souligne par Wiklund (1995).
D’autre part, les présents résultats ont révélé que les sables humides présentent
des charges bactériennes plus concentrées de celles des eaux (Figure 20).
Les présents résultats ont révélé que les charges des bactéries fécales sont très
importantes en saison pluvieuse (Figure 15), ce qui serait dû aux apports de
rejets urbains drainés par les fortes pluies. Ainsi, nous avons constaté que les

Résultats et discussion
95
taux élevés en ces charges bactériennes pendant les mois d’avril et juin seraient
associés avec l’augmentation pluviométrique (Figure 16 et Figure 19).
Cependant, la survie de ces types de bactéries serait sous l’influence de la
température (Rozen et Belkin, 2001).
Dans le cas de la présente étude nous avons révélé une corrélation négative entre
la température et les Coliformes fécaux (Tableau 7) dans les site du Beau Séjour
et Eden( Figure 14 et Figure 19), des résultats similaires ont été obtenus par
Sinton et al (2002) dans les eaux de la nouvelle Zélande ; Aslanyilmaz et al
(2004) dans les eaux de la Turquie ; Chandrana et Hatha (2005) dans les sables
de l’Inde. Ces auteurs ont montré que l’augmentation en température, pendant la
saison estivale pourrait inactiver les polluants fécaux.
Les résultats relatifs aux Staphylocoques ont révélé que leurs charges sont plus
importantes que celles des bactéries fécales pour les prélèvements analysés dans
les sites les moins pollués (Madagh, Eden) (Figure 15). Ces Staphylocoques
semblent être une composante de la microflore des sables des plages.
Selon la présente étude, les charges bactériennes dans les eaux de mer ainsi que
dans les sables sont importantes en période pluvieuse, ce qui serait dû à l’effet
d’une concentration dans le sable par le drainage des eaux usées. Aussi,
l’analyse statistique a montré l’influence de la température sur les
Staphylocoques (Figure 19 et tableau 7). Castaneda et al (2005) ont montré que
l’abondance des Staphylocoques dans les eaux de mer et le sable des plages au
Mexique, est corrélée significativement avec l’augmentation de la température.
Par ailleurs, Stabili et al (2005) en Italie, soulignent une corrélation positive
entre les charges en Staphylocoques dans les sédiments et la température de
l’eau .
Dans ce sens on pense à une relation établie entre la température de l’eau et
l’abondance des Staphylocoques dans le sable.
En outre, nous avons obtenu une corrélation positive entre les charges en
Staphylocoques et celles des Coliformes fécaux dans les sables.

Résultats et discussion
96
Les polluants fécaux ont montré une corrélation significativement positive entre
leur charge en eau et en sable dans tous les sites étudiés excepte à Eden (Figure
17) et pour ce nous pensons à une relation qui peut lier l’eau de mer et le sable
des plages.
La concentration observée dans le sable paraît liée à la charge bactérienne dans
l'eau et qui augmente au niveau des sites très pollués (Figure 15).
Une grande différence est apparue dans la concentration des indicateurs de
contamination fécale entre les eaux de mer et les sables de plage (Figure 16).
Cette différence dans les facteurs de concentration pourrait s'expliquer par une
dilution plus importante des contaminants fécaux à l'intérieur du milieu marin.
La contamination des E. coli dans les sables est corrélée avec l’abondance en
germes totaux ; donc la présence de cette flore indicatrice de contamination
fécale est liée à la présence de matières organiques dans l’environnement marin.
Selon les résultats obtenus pour les différentes flores bactériennes, notamment,
les indicateurs de la contamination fécale, nous pensons être en présence d’une
contamination dont l’origine est commune que ce soit pour l’eau de mer ou le
sable de plage.

Conclusion

Conclusion
97
Le climat exceptionnel, la beauté et la diversité du littoral oranais expliquent
aisément sa croissance démographique accélérée, avec les maxima estivaux bien
connus. Cette vocation touristique, alliée au développement des populations
riveraines s'est naturellement accompagnée d'une augmentation considérable du
volume des déchets domestiques, bactériologiquement très pollués, qui sont
systématiquement rejetés dans cet environnement aquatique.
Dans notre étude, nous pouvons résumer que le sable présente un habitat où les
bactéries peuvent vivre, croître et proliférer en s’adaptant à différentes
conditions environnementales.
Les résultats ont démontré que la distribution des différentes flores bactériennes,
n’était pas régulière sur les différents sites étudiés.
Cette étude démontre que le sable sec contient un taux de germes beaucoup plus
important que le sable humide et l’eau de mer; cela peut être expliqué par le
pouvoir auto-épurateur de l’eau de mer où les germes sont déversées et sont
soumis aux phénomènes de dilutions et de dispersion (hydrodynamisme actif).
Bien que l’effusion de la matière organique peut stimuler la croissance des
bactéries dans le sable, il est également probable que leur croissance est stimulée
par la perturbation physique du sable (marche, jeux…etc).
Le sable peut être considéré comme un filtre efficace pour éliminer les bactéries
de la colonne d’eau.
Les prédateurs qui se trouvent dans l’eau de mer (benthos, necton, plancton)
peuvent être aussi une raison du nombre réduit des germes par rapport au sable
et contribuent ainsi à leur disparition.

Conclusion
98
Les indicateurs de contamination fécale présentaient une concentration élevée en
sable de la plage des Andalouses et Beau Séjour, cela peut être expliqué par la
proximité de ces deux plages aux rejets des eaux usées domestiques.
D'une manière générale, ce milieu marin particulier va promouvoir des
sélections qui entrainent une éradication ou tout au moins une mise en minorité
rapide des espèces pathogènes par rapport au sable. En outre, les bactéries sont
soumises à des facteurs physiques, chimiques, biologiques telles que la
température, la lumière, la salinité (facteur de sélection) et les carences
alimentaires. Tous ces facteurs abiotiques et biotiques s’associent pour
augmenter ou réduire leur nombre.
Si l'effort consenti par les collectivités riveraines se poursuit et qu’elles
conjuguent leurs bonnes volontés, les pollutions bactériennes en mer ou au
niveau des plages devraient, dans l'ensemble, accuser une nette régression au
cours des années à venir.
Mais nous ne voudrions pas en terminer sans avoir fait remarquer que ce n'est en
aucune façon un argument permettant de minimiser le danger des nuisances
apportées par les eaux domestiques.
Ces effluents, dont le volume déversé en mer va sans cesse en augmentant,
transportent en particulier de nombreux éléments polluants chimiques :
hydrocarbures, pesticides, métaux lourds, détergents surtout, dont l'activité
délétère peut être très grave sur les écosystèmes marins et peut y entraîner des
modifications irréversibles (Boutiba et al, 2003 ; Boutiba, 2006 ; Kherraz,
2004 ). Les conséquences de ces phénomènes nuisibles associés se feront
inévitablement sentir dans le domaine des pollutions microbiennes puisque les
capacités de lutte du milieu marin contre les agressions par ces microorganismes
terrestres dépendent étroitement du maintien de son intégrité.

Références
bibliographiques

Références bibliographiques
99
Abu-Lail, N. I., Camesano, T. A., (2003). Role of lipopolysaccharides in
the adhesion retention, and transport of Escherichia coli JM109.
Environmental Science and Technology 37(10):2173-2183.
Aguilera, L. E. ., Gutierrez, J. R. ., and Meserve, P. L., (1999)
Variation in soil micro-organisms and nutrients underneath and outside
the canopy of adesmia bedwellii (papilionaceae) shrubs in arid coastal
chile following drought and above average rainfall. Journal of Arid
Environments, 42(1) :61–70.
Aldova, E. (1989) Serovars of Vibrio parahaemolyticus. Journal of
Hygiene, Epidemiology and Immunology, 33(2): 219–228.
Amann, R. I., Ludwig, W., and Schleifer, K. H., (1995). Phylogenetic
identification and in-situ detection of individual microbial-cells without
cultivation. Microbiological Reviews, 59(1) :143–169..
Anderson, JN. (1979) In-vitro survival of human pathogenic fungi in
Hawaii USA beach sand. Sabouraudia, 17(1): 13–22.
Araujo,R., Arribas. R., Lucena. F., Pares.R. (1989) Relation between
Aeromonas and faecal coliforms in fresh waters. J. Appl. Bacteriol. 67,
213-217.
Artz, R. R. E., Townend,J., Brown,K., Towers, W., Killham,
K.(2005). Soil macropores and compaction control the leaching potential
of Escherichia coli O157 : H7. Environmental Microbiology 7(2):241-
248.
Aslanyilmaz, A., Okus, E.,Ovez, S. (2004). Bacteriological indicators of
anthropogenic impact prior to and during the recovery of water quality in
an extremely polluted estuary, Golden Horn, Turkey, Mar. Pollut. Bull.,
49, 951-958

Références bibliographiques
100
Aubert, M., Mathonnet, S., Gillemaut,C. (1987) Dépollution des plages
par l’eau de mer éléctrolysée. Effects sur les micro-organismes
pathogènes. [Cleaning of beaches with electrolyzed seawater. Effects on
pathogenic microorganisms.] Revue Internationale d’Océanographie
Médicale, 85–86: 222–227.
Aulicino, FA., Volterra, L., Donati, G. (1985) Faecal contamination of
shore-line sands. Bollettino della Società Italiana di Biologia
Sperimentale, 61(10): 1469–1476. 124.
Ausland, G., Stevik, T. K., Hanssen, J. F., Kohler, J. C., Jenssen, P.
D., (2002). Intermittent filtration of wastewater - removal of fecal
coliforms and fecal streptococci. Water Research 36(14):3507-3516.
Backman, A and Jansson, J. K., (2004) Degradation of 4-chlorophenol
at low temperature and during extreme temperature fluctuations by
Arthrobacter chlorophenolicus A6. Microb Ecol, 48(2):246–53.
Baker, P. W. , and Leff, L. G., (2005). Intraspecific differences in
bacterial responses to modelled reduced gravity. J Appl Microbiol, 98(5)
:1239–46.
Bakermans,C., Tsapin, A., I, Souza-Egipsy,V., Gilichinsky, D. A and
Nealson ,K.H., (2003). Reproduction and metabolism at -10 degrees C of
bacteria isolated from Siberian permafrost. Environmental Microbiology,
5(4):321–326.
Bannefond,G,. Fabre,H., Mandrou,B., and Simeon,M., Buochberg,
D.,Pharm,J., (1990) HH Tennesen amd AL Grislingaas, Int. J. Pharm.,
60, 157. 222.
Bar,M., von Hardenberg,J., Meron,E., and Provenzale,A.,(2002)
Modelling the survival of bacteria in drylands : the advantage of being
dormant. Proc Biol Sci, 269(1494) :937–42, J. M.

Références bibliographiques
101
Barth, G., Gaillardin, C., (1997). Physiology and genetics of the
dimorphic fungus Yarrowia lipolytica. FEMS Microbiol. Lett. 19, 219-
237.
Barton, J. W., Ford, R. M., (1995). Determination of Effective
Transport-Coefficients for Bacterial Migration in Sand Columns. Applied
and Environmental Microbiology 61(9):3329-3335.
Bartram, J., Rees, G., (2000) Monitoring bathing waters: a practical
guide to the design and mplementation of assessments and monitoring
programmes. London, E & FN Spon. Published on behalf of the World
Health Organization, Commission of the European Communities and US
Environmental Protection Agency.
Begin, Y., Aresenault, S., Lavoie,J., (1989) : Dynamique d’une bordure
forestière par suite de la hausse récente du niveau marin, rive sud-ouest du
golfe du Saint-Laurent, Nouveau-Brunswick. Géographie Phys. Quat.,
vol. 43, n°3, p. 355-366.
Benardini,J.N., Sawyer,J., Venkateswaran,K., and
Nicholson,W.L.,(2003) Spore uv and acceleration resistance of endolithic
bacillus pumilus and bacillus subtilis isolates obtained from sonoran
desert basalt : implications for lithopanspermia. Astrobiology, 3(4) :709–
17.
Bengtsson, G., Lindqvist, R., (1995). Transport of Soil Bacteria
Controlled by Density Dependent Sorption Kinetics. water Resources
research 31(5):1247-1256.
Bentir, M., (1996) - L'épuration des eaux usées urbaines en Algérie : Etat
actuel et perspectif. Edil. Inf- eau, trimestre n° 11, juin 1996 : 4-6.
Bernard, P., Gueho, E., Pesando, D., (1988) Recherche de
dermatophytes et de moisissures pathogènes dans les sables des plages,
1986–87. [Research on dermatophytes and pathogenic moulds in sea sand,

Références bibliographiques
102
1986–87.] Athens, United Nations Environment Programme,
Mediterranean Action Plan, Mediterranean Marine Pollution Monitoring
and Research Programme (MED POL Research Project Final Report).
Bertrand, R., Roig, B., (2007). Evaluation of enrichment-free PCR-based
detection on the rfbE gene of Escherichia coli O157 - Application to
municipal wastewater.Water Research 41:1280-1286.Biol (Noisy-le-
grand), 50(4) :429–36.
Biely, P., Heinrichova, K., Kuzikova, M.,(1996). Induction and inducers
of the pectolytic system in Pseudomonas.sp. Current Microbiol. 33, 6-10.
Billi, D,. M, Potts,. (2002) Life and death of dried prokaryotes. Res
Microbiol, 153(1) :7–12.
Bird, E.C.F., (1985) ,coastline changes, a global review, chichester : John
Wiley.
Bird, S. B., Herrick, J. E., Wander, M. M., and Wright, S. F. , (2002).
Spatial heterogeneity of aggregate stability and soil carbon in semi-arid
rangeland. Environmental Pollution, 116(3) :445–455.
Blanchard, D. C., Syzdek, L.D., (1972). Concentration of bacteria in jet
drops from bursting bubbles. Journal of Geophysic Research 77:5087-
5099.
Boiron, P., Agis, F., Nguyen, VH,. (1983) Study of yeast flora of
medical interest on the beach of Saint Anne in Guadeloupe. Bulletin de la
Société de Pathologie Exotique et de ses Filiales, 76(4): 351–356.
Bonadonna, L., Dal Cero, C., Liberti, R., Pirrera, A., Santamaria, C.,
Volterra L (1993) Clostridium perfringens come indicatore in sedimenti
marini. [Clostridium perfringens as an indicator in marine sediments.]
Ingegneria Sanitaria Ambientale, 1: 28–30.

Références bibliographiques
103
Bonadonna, L., Briancesco, R., Cataldo C, Di Girolamo, I., (2002)
[Microbiological quality of seaside sands: a beach in Latium]. Igiene e
Sanità Pubblica, LVIII: 43–54. (In Italian).
Booysen,C., Dicks, L.M.T., Meijering, I., Ackermann. A.(2002).
Isolation, identification and changes in the composition of coliforms
bacteria during the malting of two different barley cultivars. Int. J. Food
Microbiol. 76, 63-73.
Borrego, JJ., Romero, P., Marino, F., (1991) Epidemiological study on
bathers from selected beaches in Malaga. Athens, United Nations
Environment Programme, Mediterranean Action Plan, pp.1–27 (MAP
Technical Reports Series No. 53).
Boughaba, A., (1994) : Morphodynamique littorale entre Cap Negro et
Cap Mazari : l’exemple de la plage de Martil. Revue de la Fac. des Lettres
pp 59-74.
Boutiba, Z., Taleb, Z., Abdelghani, F., (1996). Teneur en organochlorés
chez la Bogue (Boops boops) péchée dans la baie d’Oran. Edit.inf. EAU.
Boutiba ,Z., Taleb, M.Z., Abi-Ayed, S.M., (2003). Etat de la pollution
marine de la côte oranaise. Ed: Dar El Gharb. Oran (Algérie) : 69p.
Boutiba, Z., (2006). Pollution: la mer en danger. Le Quotidien d’Oran.
SPA Edition – ORAN.
Bradford, S. A., Simunek, J., Bettahar, M., Van Genuchten, M. T.,
Yates, S. R., (2003). Modeling colloid attachment, straining, and
exclusion in saturated porous media. Environmental Science &
Technology 37(10):2242-2250.
Bradford, S. A., Bettahar, M., (2005). Straining, attachment, and
detachment of Cryptosporidium oocysts in saturated porous media.
Journal of Environmental Quality 34(2):469-478.

Références bibliographiques
104
Bradford, S. A., Simunek, J., Bettahar, M., Tadassa, Y. F., van
Genuchten, M. T., Yates, S. R., (2005). Straining of colloids at textural
interfaces. water Resources research 41(10).
Bradford, S. A., Bettahar, M., (2006). Concentration dependent
transport of colloids in saturated porous media. Journal of Contaminant
Hydrology 82(1-2):99-117.
Brouwer, H., (1984). Ground water pollution microbiology. In: C.P., B.
G. a. G., editor. Element of soil science and ground water hydrology. New
York: Wiley. p 9-38.
Brown, D. G., Stencel, J. R., Jaffe, P. R., (2002). Effects of porous
media preparation on bacteria transport through laboratory columns.
Water Research 36(1):105-114.
Buchanan , R.L., Smith, J.L., Long, W., (2000) Microbial risk
assessment : dose-response relations and risk characterization,
International journal of food microbiology, 58, 159-172.
Bullitt, E., Makowski, L., (1998). Bacterial adhesion pili are
heterologous assemblies of similar subunits. Biophysical Journal
74(1):623-632.
Busscher, H. J., Weerkamp, A. H., van der Mei, H. C., van Pelt, A.
W., de Jong, H. P., Arends, J., (1984). Measurement of the surface free
energy of bacterial cell surfaces and its relevance for adhesion. Appl
Environ Microbiol 48(5):980-983.
Cable,J.M., and Huxman,T.E., (2004) Precipitation pulse size effects
on sonoran desert soil microbial crusts. Oecologia, 141(2) :317–24.
Camesano, T. A., Logan, B. E., (1998). Influence of fluid velocity and
cell concentration on the transport of motile and nonmotile bacteria in
porous media. Environmental Science & Technology 32(11):1699-1708.

Références bibliographiques
105
Camper, A. K., Hayes, J. T., Sturman, P. J., Jones, W. L.,
Cunningham, A. B., (1993). Effects of Motility and Adsorption Rate
Coefficient on Transport of Bacteria through Saturated Porous-Media.
Applied and Environmental Microbiology 59(10):3455-3462.
Carillo-Muñoz, AJ., Torres-Rodriguez, JM., Madrenys-Brunet, N.,
Dronda-Ayza, A., (1990) Comparative study on the survival of 5 species
of dermatophytes and Scopulariopsis brevicaulis in beach sand under
laboratory conditions. Revista Iberoamericana de Microbiologia, 7(2):
36–38.
Cases,I., and de Lorenzo,V., (2002) The grammar of (micro)biological
diversity. Environmental Microbiology, 4 :623–627.
Castaneda chavez, M., Sedas,V.P., Borunda,E.O., Reynoso,F.L.,
(2005). Influence of water temperature and salinity on seasonal
occurrences of Vibrio cholerae and enteric bacteria in oyster-producing
areas of Veracruz, Mexico. Mar. Pollut. Bull., 50, 1641-1648.
Celico, F., Varcamonti, M., Guida, M., Naclerio, G., (2004). Influence
of precipitation and soil on transport of fecal enterococci in fractured
limestone aquifers. Applied and Environmental Microbiology 70(5):2843-
2847.
Chabasse D, Laine P, Simitzis-Le-Flohic AM, Martineau B, el Hourch
M, Becaud JP (1986) Relationship between microbial water quality and
environnemental conditions in coastal recreational waters: the Fylde
coast, UK. Water Res., 35, 4029-4038.
Chandrana, A., et Hatha,M., (2005). Relative survival of Escherichia
coli and Salmonella typhimurium in tropical estuary. Water Res., 39,
1397-1403.
Chen, G., Strevett, K. A., (2001). Impact of surface thermodynamics on
bacterial transport. Environmental Microbiology 3(4):237-245.

Références bibliographiques
106
Chen, G., Strevett, K. A., (2002). Surface free energy relationships used
to evaluate microbial transport. Journal of Environmental Engineering-
Asce 128(5):408-415.
Chen, G., Strevett, K. A., (2003). Microbial surface thermodynamics and
interactions in aqueous media. Journal of Colloid and Interface Science
261(2):283-290 Chile. Revista Iberoamericana de Microbiologia, 36(2):
71–77.
Chen, G., Rockhold, M., Strevett, K. A., (2003). Equilibrium and
kinetic adsorption of bacteria on alluvial sand and surface thermodynamic
interpretation. Research in Microbiology 154(3):175-181.
Cheung, W.H., Chang,K.C., Hung, R.P., (1991). « Variations in
microbial indicator densities in beachwater and health-related assessment
of bathing water qualities », Epidemiology and infection, 106(2) : 329-
344.
Choi, N. C., Kim, D. J., Kim, S. B., (2007). Quantification of bacterial
mass recovery as a function of pore-water velocity and ionic strength.
Research in Microbiology 158(1):70-78.
Ciaramella, M., Pisani, F. M ., and Rossi M., (2002). Molecular biology
of extremophiles : recent progress on the hyperthermophilic archaeon
Sulfolobus. Antonie Van Leeuwenhoek, 81(1-4):85–97.
Collins, M.D., Lawson, P.A, Willems, A., Cordoba, J.J., Fernandez-
Garayzabal, J, Garcia, P., (1994) Thephylogeny of the genus
Clostridium: proposal of five new genera and eleven new species
combinations. Int J Syst Bacteriol ;44:812-26.
Conboy, M. J., and Goss, M.J., (2000). Natural protection of
groundwater against bacteria of fecal origin. Journal of Contaminant
Hydrology 43:1-24.

Références bibliographiques
107
Conseil Supérieur d’Hygiène Publique de France (1990) Qualité
microbiologique des sables: essai d’un nouveau précédé de désinfection.
[Microbiological quality of sea sand: a trial of a new disinfection
protocol.] Paris.
Cools, D., Merckx, R., Vlassak, K., Verhaegen, J., (2001). Survival of
E. coli and Enterococcus spp. derived from pig slurry in soils of different
texture. Applied Soil Ecology 17(1):53-62.
Corapcioglu, M. Y., and Haridas, A., (1984). Transport and fate of
microorganisms in porous media: a theoretical investigation. journal of
hydrology 72:149-169.
Corapcioglu, M. Y., Choi, H., (1996). Modeling colloid transport in
unsaturated porous media and validation with laboratory column data.
water Resources research 32(12):3437-3449.
Costerton, J. W., Geesey, G. G., Cheng, K. J., (1978). How bacteria
stick. Scientific American 238(1):86-95.
Costerton, J. W., Lewandowski, Z., Caldwell, D. E., Korber, D. R.,
Lappin-Scott, H. M., (1995). Microbial biofilms. Annual Review of
Microbiology 49:711-7457
Costerton, J. W., Marie, C. B., and Cheng K.J., (1985). Phenomena of
bacterial adhesion. In: Fletcher, D. C. S. a. M., editor. Bacterial Adhesion.
New York: Plenum. p 3-43.
Costerton, J. W., Stewart, P. S., Greenberg, E. P., (1999). Bacterial
biofilms: A common cause of persistent infections. Science
284(5418):1318-1322.
Curtis, T. P., Mara, D. D. et Silva, S. A., (1992) Influence of pH,
oxygen and humic substances on the ability of sunlight to damage fecal
coliforms in waste stabilization pond water. Appl. Environ. Microbiol. 58,
1335-1343.

Références bibliographiques
108
Curtis, T. P., Sloan, W. T. ,and Scannell J. W., (2002) Estimating
prokaryotic diversity and its limits. Proc. Nat. Acad. Sci. USA, 99:10494–
10499. 5(4):321–326, 2003. 65(3–4): 305–328.
Dabrowski, J., (1982) Isolation of the Shigella genus bacteria from beach
sand and water of the bay of Gdansk. Bulletin of the Institute of Maritime
and Tropical Medicine in Gdyinia, 33(1–2):49–53.
DeFlaun, M. F., Oppenheimer, S. R., Streger, S., Condee, C. W.,
Fletcher, M., (1999). Alterations in adhesion, transport, and membrane
characteristics in an adhesion-deficient pseudomonad. Applied and
Environmental Microbiology(2):759-765.
Dellali, M., M, El bour ., P, AISSA., (2000). Evaluation de la pollution
bacterienne dans la lagune de Bizerte : resultats preliminaires. J. Rech.
Oceanogr., 26, 18-28.
Deming, W., and Baross, J. A.,(1993). Deep-sea smokers : windows to
a subsurface biosphere ? Geochim Cosmochim Acta, 57(14) :3219–30,
1993.
Dowidart, A. , Abdel-Monem, M.H., (1990) Effect of chemical
pollutants on bacterial counts in El-Temsah Lake area, Ismailia, Egypt.
Journal of the Egyptian Public Health Association, 65(3–4): 305–328.
Dufour, A. P., (1977). « Escherichia coli: The fecal coliform », Am. Soc.
Test. Mater. Spec. Tech. Publ., 635 : 48-58.
Dufour, A. P., (1986) Diseases caused by water contact. In: Waterborne
Diseases in the United States (ed. by G. F. Craun), 23-41. CRC, Boca
Raton, Florida.
Dufrene, Y. F., Boonaert, C. J. P., Rouxhet, P. G., (1996). Adhesion of
Azospirillum brasilense: Role of proteins at the cell-support interface.
Colloids and Surfaces BBiointerfaces 7(3-4):113-128.

Références bibliographiques
109
Dunbar,J., Takala,S., Barns,S.M., Davis,J.A., and Kuske,C.R. (1999)
,Levels of bacterial community diversity in four arid soils compared by
cultivation and 16S rRNA gene cloning. Applied and Environmental
Microbiology, 65(4) :1662–1669.
Dunsmore, J.D., Thomson, R.C., Bates, I.A., (1984) Prevalence and
survival of Toxocara canis eggs in the urban environment of Perth,
Australia. Veterinary Parasitology, 16(3–4): 303–311.
Engen, S., and Lande, R., (1996) Population dynamic models generating
the lognormal species abundance distribution. Math Biosci, 132(2):169–
83.
EPA (2002). RCRA Waste Sampling Draft Technical Guidance,
Planning, Implementation and Assessment, EPA530-D-02-002, Office of
Solid Waste and Emergency Response, Office of Solid Waste.
Washington, DC, 353 pages.
Esseilli, M., and Kassem, I.I., (2007). A new method to identify sources
of fecal pollution: DGGE fingerprints of E. coli communities. ASM
General Meeting. Toronto (Canada).
Essid, N., Mahmoudi, E., Boufahja ;F ., Dellali,M., Beyrem,H,. et
Aissa,P., (2007). Impact des pseudo-feces de moules sur les densites des
bacteries heterotrophes dans le secteur mytilicole de la lagune de
Bizerte (Tunisie). Rev.Sci. Eau, 20, 383-392.
FAO GFCM-PC version (1997) et FAO Fishstat-PC, version (1998).
Ferguson, C., Husman, A. M. D., Altavilla, N., Deere, D., Ashbolt, N.,
(2003). Fate and transport of surface water pathogens in watersheds.
Critical Reviews in Environmental Science and Technology 33(3):299-
361.

Références bibliographiques
110
Fernandez, F., Ferrer, M.D., (1982) Estudio microbiologico de la arena
de las playas de Barcelona.[Microbiological study of Barcelona beaches.]
Gazeta Sanitaria, 3: 13–17.
Figueras, M.J., Guarro, J., Soler, L., Inza, I., Polo F,. (1992) Estudio
piloto sobre la contamination de las playas del litoral Catalan. [A pilot
study on the contamination of Catalounian beaches.] Proceedings of the
1° Congreso Nacional del Medio Ambiente, Madrid.
Fletcher, M., and, Loeb, G.I., (1979). Influence of substratum
characteristics on the attachment of marine pseudomonas to soil surfaces
Applied and Environmental Microbiology 37:67-72.
Fontes, D. E., Mills, A. L., Hornberger, G. M., Herman, J. S., (1991).
Physical and chemical factors influencing transport of microorganisms
through porous media. Applied and Environmental Microbiology
57(9):2473-2481.
Gannon, J. T., Manilal, V.B., and Alexander, M., (1991). Relationship
between cell surface properties and transport of bacteria through soil.
Applied and Environmental Microbiology 57(1):190-193
Garcia-Pichel. F., and Pringault, O., (2001) Microbiology -
Cyanobacteria track water in desert soils. Nature, 413(6854) :380–381.
Garland,J.L., Cook, K.l., Adams,J.L., and Kerkhof,L., (2001)
Culturability as an indicator of succession in microbial communities.
Microb Ecol, 42(2) :150–8.
Geldreich ,E ., (1967). -The coliform group, II, Réactions in the EC
Medium at 45° C. -Appl, Microbiol., 6 347-8.
Gerba, C. P. W. C., and Melnick J.L., (1975). Fate of wastewater
bacteria and viruses in soil. Journal of Irrigation and Drain Division
101:157-174.

Références bibliographiques
111
Gerba, C.P., and Bitton, G., (1984) Groundwater pollution
microbiology. John Wiley & Sons, USA.
Ghebremedhin, B., Layer, F., König, W., König, B., (2008). "Genetic
classification and distinguishing of Staphylococcus species based on
different partial gap, 16S rRNA, hsp60, rpoB, sodA, and tuf gene
sequences". J. Clin. Microbiol. 46 (3): 1019–1025.
Ghinsberg, R.C., Leibowitz, P., Witkin, H., Mates, A., Seinberg, Y.,
Bar, D.L., Nitzan, Y., Rogol, M., (1994) Monitoring of selected bacteria
and fungi in sand and seawater along the Tel-Aviv coast.
Gillet,P., Barrat, J. A. , Heulin, T. , Achouak, W. , Lesourd, M. ,
Guyot, F. , and Benzerara, K., (2000). Bacteria in the Tatahouine
meteorite : nanometricscale life in rocks. Earth Planet Sci Lett, 175(3-4)
:161–7.
Gomez-Suarez, C., Pasma, J., van der Borden, A. J., Wingender, J.,
Flemming, H. C., Busscher, H. J., van der Mei, H. C., (2002).
Influence of extracellular polymeric substances on deposition and
redeposition of Pseudomonas aeruginosa to surfaces. Microbiology 148(Pt
4):1161-1169.
Grasso, D., Smets, B. F., Strevett, K. A., Machinist, B. D., VanOss, C.
J., Giese, R. F., Wu, W., (1996). Impact of physiological state on surface
thermodynamics and adhesion of Pseudomonas aeruginosa.
Environmental Science and Technology 30(12):3604-3608.
Guiraud, J.P., (2003) Microbiologie alimentaire, Dunod, Paris, RIA (Le
mensuel de l’innovation alimentaire), 651 p.
Haffar, M., Handoko Y., Gilbride K., (2007). The detection and
enumeration of bacterial pathogens using FISH and quantitative Real-
Time PCR. ASM General Meeting. Toronto (Canada).

Références bibliographiques
112
Hagedorn, C., McCoy E.L., Rahe T.M., (1981). The potential for
groundwater contamination from septic effluents. Journal of
Environmental Quality(10):1-8.
Hallier-Soulier, S., Racaud, and Chablain P., (2007). Development of a
duplex quantitative PCR test for the monitoring of fecal pollution in
environmental waters. ASM General Meeting. Toronto (Canada).
Hamadi, F., Latrache, H., Mabrrouki, M., Elghmari, A., Outzourhit,
A., Ellouali, M., Chtaini, A., (2005). Effect of pH on distribution and
adhesion of Staphylococcus aureus to glass. Journal of Adhesion Science
and Technology 19(1):73-85.
Hassen A., Jedidi N., K. Belguith, M. Cherif M. and Boudabous A.
(2001) Microbial characterization during composting of municipal solid
waste. Bioresources Technology, 80, 231 - 239.
Henne, A., Bruggemann,H., Raasch,C., Wiezer,A., Hartsch,T.,
Liesegang,H., Johann,A., Lienard,T., Gohl,O., Martinez-Arias,R.,
Jacobi,C., Starkuviene,V., Schlenczeck,D., Dencker,S., Huber,R.,
Klenk,H.P., Kramer,A., Merkl,R., Gottschalk,G., and Fritz,H.G.,
(2004) The genome sequence of the extreme thermophile Thermus
thermophilus. Nat Biotechnol, 22(5) :547–53.
Hermansson, M., (1999). The DLVO theory in microbial adhesion.
Colloids and Surfaces B-Biointerfaces 14(1-4):105-119.
Hoog, G.S,. Guarro, J., Gené, J., Figueras, M.J., (2000) Atlas of
clinical fungi. Second edition.Utrecht, Centrallbureau voor
Schimmelculture.
Hoornweg ,D., Thomas, L., et Otten, L., (2000). Composting and Its
Applicability in Developing Countries, Urban Waste management,
Published for the Urban Development Division, The World Bank,
Washington DC, 52 pages.

Références bibliographiques
113
Huysman, F., Verstraete, W., (1993). Water-Facilitated Transport of
Bacteria in Unsaturated Soil Columns - Influence of Cell-Surface
Hydrophobicity and Soil Properties. Soil Biology and Biochemistry
25(1):83-90.
Ibaraki, M., Sudicky, E. A., (1995). Colloid-facilitated contaminant
transport in discretely fractured porous media .2. Fracture network
examples. water Resources research 31(12):2961-2969.
Ifremer, (2007) P06 Annexes - Le REBENT Méditerranée – APS février
, Ingegneria Sanitaria Ambientale, 1: 28–30.
Iliuta, I., Larachi, F., (2006). Dynamics of cells attachment, aggregation,
growth and detachment in trickle-bed bioreactors. Chemical Engineering
Science 61(15):4893-4908.
Iwabuchi, N., Sunairi, M., Anzai, H., Morisaki, H., Nakajima, M.,
(2003). Relationships among colony morphotypes, cell-surface properties
and bacterial adhesion to substrata in Rhodococcus. Colloids and Surfaces
B-Biointerfaces 30(1-2):51-60.
Izquierdo, J., Piera, G., Aledany, M.C., Lucena, F., (1986) Estudio de
la flora fungica de la arena de la playa de Barcelona. [A study of the
fungal flora of the beaches in Barcelona.] Athens, United Nations
Environment Programme, Mediterranean Action Plan, Mediterranean
Marine Pollution Monitoring and Research Programme (MED POL
Research Project Final Report.
Jacobs,A., Lafolie,F., Herry,S., Debroux,J.M., (2007).Colloids and
Surfaces B : Biointerfaces, volume 59 , p: 35-45.
Janda, J. M., Duffey, P. S., (1988) Mesophilic aeromonads in human
disease : current taxonomy, laboratory identification and infectious
disease spectrum. Rev. Infect. Dis. 10, 980-997.

Références bibliographiques
114
Johnson, W. P., Logan, B. E., (1996). Enhanced transport of bacteria in
porous media by sediment-phase and aqueous-phase natural organic
matter. Water Research 30(4):923-931.
Johnson, W. P., Li, X. Q., Yal, G., (2007). Colloid retention in porous
media: Mechanistic confirmation of wedging and retention in zones of
flow stagnation. Environmental Science and Technology 41(4):1279-
1287.
Jordan, F. L., Sandrin, S. K., Frye, R. J., Brusseau, M. L., Maier, R.
M., (2004). The influence of system complexity on bacterial transport in
saturated porous media.Journal of Contaminant Hydrology 74(1-4):19-38.
Joy, D.M., Lee, H., Reaume, C.M., Whiteley, H.R. and Zelin, S. (1998)
Microbial contamination of subsurface tile drainage water from field
applications of liquid manure, Canadian Agricultural Engineering, 40 (3):
153-160.
Kempf.B and Bremer.E.,(1998) Uptake and synthesis of compatible
solutes as microbial stress responses to high-osmolality environments.
Arch Microbiol, 170(5) :319–30,.
Kherraz, D., (2004). Etat de la pollution bactériologique au niveau de la
côte oranaise – cas des plages d’Aïn El Turk, de Coralès et de Maddagh.
Mém. Magister , Univ. Oran, Algérie : 120p.
Khiyama, H.M., Makemson. J.C., (1973) Sand beach bacteria:
enumeration and characterization. Applied Microbiology, 26(3): 293–297.
Koneman, E.W., Allen. S. D., Janda,W. M., Schreckenberger. P .C.,
Winn, W.C. J., (1997) Color Atlas and Textbok of Diagnostic
Microbiology. 5th Philadelphia: Lippincott Williams & Wilkins; p. 709-
84.
Kuske,C.R., Barns, S.M.,Busch,J.D.,(1997) Diverse uncultivated
bacterial groups from soils of the arid southwestern united states that are

Références bibliographiques
115
present in many geographic regions. Appl Environ Microbiol, 63(9)
:3614–21.
Kuske, C.R., Ticknor, L.O., Miller, M.E., Dunbar, J.M., Davis, J.A.,
Barns, S.M., and Belnap, J. (2002) Comparison of soil bacterial
communities in rhizospheres of three plant species and the interspaces in
an arid grassland. Appl Environ Microbiol 68: 854-63.
Laliberte, P., et Grimes,D., (1982). Survival of Escherichia coli in lake
bottom sediment. Appl. Environ. Microbial., 43,623-628.
Lappin-Scott, H. M., and Costerton, J.W., (1989). Bacterial biofilms
and surface fouling. Biofouling(1):332-342.
Larrondo, J.V,, Calvo, M.,A., (1989) Fungal density in the sands of the
Mediterranean coast beaches. Mycopathologia, 108(3): 185–194.
Dunsmore JD, Thomson RC, Bates IA (1984) Prevalence and survival of
Toxocara canis eggs in the urban environment of Perth, Australia.
Veterinary Parasitology, 16(3–4): 303–311.
Lee, C.M., T.Y. Yin, C.C. Lin, G.A. Kohbodi, A. Bhatt, R. Lee, and
J.A. Jay. (2006). Persistence of fecal indicator bacteria in Santa Monica
Bay beach sediments. Water Research 40:2,593–2,602.
Li, B. K., Logan, B. E., (2004). Bacterial adhesion to glass and metal-
oxide surfaces. Colloids and Surfaces B-Biointerfaces 36(2):81-90.
Li, M., Y. Lai, A. E. Villaruz, D. J. Cha, D. E. (2007) Sturdevant, and
M. Otto.. Gram-positive three-component antimicrobial peptide-sensi
system. Proc Natl Acad Sci U S A 104:9469-9474.
Li, X. Q., Lin, C. L., Miller, J. D., Johnson, W. P., (2006). Role of
grain-to-grain contacts on profiles of retained colloids in porous media in
the presence of an energy barrier to deposition. Environmental Science &
Technology 40(12):3769-3774.

Références bibliographiques
116
Lighthart,B., and Shaffer,B.T., (1994) Bacterial flux from chaparral
into theatmosphere in midsummer at a high desert location. Atmospheric
Environment, 28(7) :1267–1274.
Liu, J. L., Xu, W. P., Zhao, Q. J., Qiao, M. Q., Zhang, H. Y., Chen,
G., (2007). Bacterial retention in lipopolysaccharide coated silica sand.
Separation Science and Technology 42(5):1031-1047.
Llewellyn, P.J., Shackley, S.E., (1996) The effect of mechanical beach-
cleaning on invertebrate populations. British Wildlife, 7(3): 147–155
“recul du trait de côte le long du littoral aquitain » l’ Hydraulique et la
maitrise du littoral, société hydrotechnique de France, Paris, p. II.6.1.-
II.6.9.
Lorin, J., Mignot, C., (1984) “recul du trait de côte le long du littoral
aquitain » l’Hydraulique et la maitrise du littoral, société hydrotechnique
de France, Paris, p. II.6.1.-II.6.9.
Lovley, R., (2003) Cleaning up with genomics : applying molecular
biology to bioremediation. Nat Rev Microbiol, 1(1):35–44.
Lunsdorf,H., Erb, R.W., AbrahamW.R., and Timmis,N.,(2000)
Clay hutches’: a novel interaction between bacteria and clay minerals.
Environ Microbiol, 2(2) :161–8, 2000.
Margesin, R., Nogi, Y., (2004). Psychropiezophilic microorganisms. Cell
Mol Biol (Noisy-le-grand), 50(4) :429–36.
Marion, G.M. , Fritsen,C.H., Eicken,H., and Payne,M.C., (2003) The
search for life on Europa : limiting environmental factors, potential
habitats, and Earth analogues. Astrobiology, 3(4) :785–811.
Marshall, K. C., (1976). Interfaces in microbial ecology. Cambridge,
Mass: Harvard University Press.
Marshall, K. C., (1980). Asorption of Microorganims to Soils and
sediments. New- York: Wiley.

Références bibliographiques
117
Marshall, K. C., (1986). Adsorption and adhesion processes in microbial
growth at interfaces. Advances in Colloid and Interface Science(25):59-
86.
Marshall, K. C., Stout, R., Mitchell, R., (1971). Selective sorption of
bacteria from seawater. Canadian Journal of Microbiology 17(11):1413-
1416.
Matthess, G., and Pekdeger A., (1985). Survival and transport of
pathogenic bacteria and viruses in groundwater. In: Ward, C. H., Giger,
W., and McCarty, P.L.,editor. Ground water quality. New York: Wiley. p
472-482.
McBride, R. A.,(1991). Shorline changes of the Islanand arcs, Louisiana,
from 1853 to 1989, Denver: U.S. geological survey.
McCaulou, D. R., R.C., Bales, and, R.G., Arnold, (1995). Effect of
temperaturecontrolled motility on transport of bacteria and microspheres
through saturated sediment. water Resources research 31(2):271-281.
McClaine, J. W., Ford, R. M., (2002). Characterizing the adhesion of
motile and nonmotile Escherichia coli to a glass surface using a parallel-
plate flow chamber. Biotechnology and Bioengineering 78(2):179-189.
McEldowney, S., and, Fletcher M., (1986). The effect of growth
conditions and surface characteristics of aquatic bacteria on their
attachment on solid surfaces. Journal of General Microbiology(132):513-
523.
Meadows, P. S., and J. G. Anderson. (1968). Microorganisms attached
to marine sand grains. J. Mar. Biol. Ass. U.K. 48:161-175
Meinck, F., Stooff, H., Kohlschutter., (1977). Les eaux résiduaires
industrielles, 2ème Ed. Masson, paris, 863p.

Références bibliographiques
118
Mendes B, Nascimento MJ, Oliveira JS (1993) Preliminary
characterisation and proposal of microbiological quality standard of sand
beaches. Water Science and Technology, 27(3–4): 453–456
Mendes B, Urbano P, Alves C, Lapa N, Norais J, Nascimento J,
Oliveira JFS (1997) Sanitary quality of sands from beaches of Azores
islands. Water Science and Technology, 35(11–12): 147–150.
Mendes B, Urbano P, Alves C, Morais J, Lapa N, Oliveira JS (1998)
Fungi as environmental microbiological indicators. Water Science and
Technology, 38(12); 155–162.
Mimouni, R., Aitalla, A., Anajjarel, M., Finance, C., et Moukrim, A.,
(2002)- Impact du rejet des eaux usées sur la qualite microbilogique des
plages de la baie d’Agadir (Maroc)- Journal Europeen d’hydrologie, 33, 1,
pp. 115-123.
Ministère des mines de l’énergie et de l’hydraulique. (1996). .Etude
des eaux souterraines pour l’alimentation en eau potable de
l’agglomération de Bamako, Mai.
Ministère de l’aménagement du territoire et de l’environnement,
(1997). La défense des cotes contre l’érosion marine, Paris, 142 p
Miossec, A., (1998), les littoraux entre nature et aménagement , Paris :
Sedes, 192 p.
Monfort, P. et Baleux, B., (1990) Dynamics of Aeromonas hydrophila,
Aeromonassobria and Aewmonascaviae in a sewage treatment pond. Appl.
Environ. Microbiol. 56, 1999-2006.
Mouffok, N., (2005). Etat de la contamination bactériologique de la côte
oranaise (plages : Ain El Turk, Bousfer et Madagh). Mém. Magister,
Univ. Oran, Algérie.
Natsch, A., Keel, C., Troxler, J., Zala, M., VonAlbertini, N., Defago,
G., (1996). Importance of preferential flow and soil management in

Références bibliographiques
119
vertical transport of a biocontrol strain of Pseudomonas fluorescens in
structured field soil. Applied and Environmental Microbiology 62(1):33-
40.
Navarro-Gonzalez, R. , Rainey, F. A. , Molina, P. , Bagaley, D. R. ,
Hollen, B., de la Rosa, J., Small, A. M. , Quinn, R. C. , Grunthaner, F.
J. , Caceres, L., Gomez-Silva, B., and McKay,. C. P., (2003) Mars-like
soils in the Atacama Desert, Chile, and the dry limit of microbial life.
Science,302(5647) :1018–1021.
Nelson, K. E., Ginn, T. R., (2001). Theoretical investigation of bacterial
chemotaxis in porous media. Langmuir 17(18):5636-5645.
Nestor I, Costin-Lazar L, Sovrea D, Ionescu N (1984) Detection of
enteroviruses in sea water and beach sand. Zentralblatt für Bakteriologie,
Mikrobiologie und Hygiene Abteilung 1, 178(5–6):527–534.
Ngole, V., Mpuchane, S., Totolo, O., (2006). Survival of faecal
coliforms in four different types of sludge-amended soils in Botswana
EUROPEAN JOURNAL OF SOIL BIOLOGY 42.
Nielsen, P. H., Jahn, A., Palmgren, R., (1997). Conceptual model for
production and composition of exopolymers in biofilms. Water Science
and Technology 36(1):11-19.
O.M.S., (1977)Surveillance de la qualité de l’eau de boisson. Genève .
O.M.S., (1986). Directives de qualité pour l’eau de boisson. Vol. 3 :
contrôle de la qualité de l’eau de boisson destinée à l’approvisionnement
des petites collectivités. Genève 1986.
O.M.S., (1987).Technologie de l’approvisionnement en eau et de
l’assainissement dans les pays en développement. Série de rapports
techniques 742. Genève 1987.
O.M.S/PNUE., (1990). L'utilisation des pesticides en agriculture et ses
conséquences pour la santé publique. Genève, OMS, 145 p.

Références bibliographiques
120
O.M.S., (2004). Directives pour la sécurité des eaux de baignades.
Volume 1, eaux côtières et eaux douces, Genève (Suisse).
O.M.S / P.N.U.E., (1995)- Recommandations pour la surveillance
sanitaire deszones côtières à usage récréatif et des zones conchylicoles:
quelques agents bactériens pathogènes, III (3) : 1-14.
Obiri-Danso, K., Jones, K., (1997) Seasonality of thermophilic
Campylobacter and faecal indicators in sediments from the intertidal zone
of Morecambe Bay. In: Proceedings of the 9th International Workshop on
Campylobacter, Helicobacter and Related Organisms. Cape Town, 15–19
September. Abstracts.
Oliveira JS, Mendes BS (1992) Qualidade da Agua do Litoral Portugues.
[Water quality in Portugal.] 1° Congresso da Agua, Vol. 2. Lisbon,
Portuguese Association of Water Resources (APRH), pp. 155–179.
Olson, M. S., Ford, R. M., Smith, J. A., Fernandev, E. J., (2004).
Ouantification of bacterial chemotaxis in porous media using magnetic
resonance imaging. Environmental Science and Technology 38(14):3864-
3870.
Oshiro, R., Fujioka, R., (1995) Sand, soil and pigeon droppings: sources
of indicator bacteria in the waters of Hanauma Bay, Oahu, Hawaii. Water
Science and Technology, 31(5–6): 251–254.
Papadakis, J.A., Mavridou, A., Richardson, S.C., Lambiri, M.,
Marcelou, U., (1997) Bather related microbial and yeast populations in
sand and seawater. Water Research, 31(4): 799–804
Paskoff ,R.,(1993). Côtes en danger, Paris : Masson, 250 p.
Paskoff, R., (1998). Les littoraux, impact des aménagements sur leur
évolution, Paris : Armand Colin.

Références bibliographiques
121
Pasqualini, C., (1997 ) Caractérisation des peuplements et types de
fonds le long du littoral corse (Méditerranée, France). Thèse de doctorat
Université de Corse 1-190.
Pekdeger, A., and Matthess, G., (1983). Factor of bacteria and virus
transport in groundwater. Environmental Geology 5(49-52).
Peterson, H. G. (2001). Rural Drinking Water and Waterborne Illness.
In: Maintaining Drinking Water Quality, Lessons from the Prairies and
Beyond, Proceedings of the Ninth National Conference on Drinking
Water. Regina, Saskatchewan, Canada. May 16-18. Canadian Water and
Wastewater Association. W. Robertson (Editor).
Piriou, J.Y., Menesguen ,A. et Salomon , J.C., (1991). Les marées
vertes à ulves: conditions nécessaires, évolution et comparaison de sites.
In: ELLIOTT M. et DUCROTOY J.P., eds., Estuaries and Coasts:
Spatial and Temporal Intercomparisons, Olsen et Olsen:117-122.
PNUE / OMS, (1979). Recommandation pour la surveillance sanitaire
des zones côtières à usage récréatif et des zones conchylicoles. Bureau
régional de l'OMS pour l'Europe, Copenhague, 168p.
PNUE/FAO/OMS/AIEA, (1990). Evaluation de l'état de la pollution de
la mer Méditerranée par les composés organohalogénés. MAP Tech. Rep.
Ser., (39):105–216.
Pourcher, A. M., Picard-Bonnau, F., Ferre V., Agnieszka, G.,
Vasilica, S., Moguedet G., (2007). Survival of faecal indicators and
enteroviruses in soil after landspreading of municipal sewage sludge.
Applied Soil Ecology 35:473-479.
Powelson, D. K., Mills, A. L., (1996). Bacterial enrichment at the gas-
water interface of a laboratory apparatus. Applied and Environmental
Microbiology 62(7):2593-2597.

Références bibliographiques
122
Prado B, Bernal P, Contreras M, Savedra M, del Moral A, Joyas M
(1994) Numerical taxonomy of staphylococci isolated from water and
beach sand from Valparaiso and Vina del Mar,Chile. Revista
Iberoamericana de Microbiologia, 36(2): 71–77.
Prieur,D., Erauso,G. , Jeanthon,C. (1995). Hyperthermophilic life at
deepsea hydrothermal vents. Planet Space Sci, 43(1-2) :115–22.
Rajagopalan, R., and C. Tien, (1976). Trajectory analysis of deep bed
filtration using the sphere in cell porous media model. AIChe J(22):523 à
533.
Readman J.W., Fillmann G., Tolosa I., Bartocci J., Villeneuve J.-P.,
Catinni C., Mee L.D., (2002) - Petroleum and PAH contamination of the
Black Sea, Marine Pollution Bulletin, 44, 48-62.
Reasoner, D.J. et Geldreich, E.E. (1985). A new medium for the
enumeration and subculture of bacteria from potable water. Appl.
Environ. Microbiol., 49 : 1-7.
Redman, J. A., Walker, S. L., Elimelech, M., (2004). Bacterial adhesion
and transport in porous media: role of the secondary energy minimum.
Environmental Science and Technology 38(6):1777-1785.
Rhodes, M. W. & Kator, H. (1994) Seasonal occurrence of mesophilic
Aeromonas spp. as a function of biotype and water quality in temperate
freshwater lakes. Wat. Res. 28, 2241-2251.
Rijnaarts,H.H.M., Norde,W., Bouwer,J., (1993). Bacterial adhesion
under static and dynamic conditions. Appl Environ Microbiol
59(10):3255-3265.
Rijnaarts, H. H. M., W. Norde, E.J. Bouwer, J. Lyklema, and A.J.B.
Zehnder, (1995). Reversibility and mechanism of bacterial adhesion.
Colloids and surfaces B: Biointerfaces(4):5-22.

Références bibliographiques
123
Rijnaarts, H. H. M., Norde, W., Bouwer, E. J., Lyklema, J., Zehnder,
A. J. B., (1996). Bacterial deposition in porous media related to the clean
bed collision efficiency and to substratum blocking by attached cells.
Environmental Science &Technology 30(10):2869-2876.
Rijnaarts, H. H. M., Norde, W., Lyklema, J., Zehnder, A. J. B.,
(1999). DLVO and steric contributions to bacterial deposition in media of
different ionic strengths. Colloids and Surfaces B-Biointerfaces 14(1-
4):179-195.
Roses Codinachs, M., Isern Vins, A.M., Ferrer Escobar, M.D.,
Fernandez Perez ,F., (1988) Microbiological contamination of the sand
from the Barcelona city beaches. Revista de Sanidad e Higiene Publica,
62(5–8): 1537–1544.
Rozen,Z. et Belkin.,K (2001). Survival of enteric bacteria. FEMS
Microbiol. Rev., 25, 513-529.
Sabat, G., Rose P., Hickey W.J., Harkin J.M., (2000). Selective and
sensitive method for PCR amplification of Escherichia coli 16S rRNA
genes in soil. Applied and Environmental Microbiology 66:844-849.
Saffary, R., Nandakumar, R., Spencer, D., Robb, F. T., Davila, J. M.,
Swartz, M. L., Ofman, R. J. ,Thomas,H., and DiRuggiero, J., (2002)
Microbial survival of space vacuum and extreme ultraviolet irradiation :
strain isolation and analysis during a rocket flight. FEMS Microbiology
Letters, 215:163–168.
Sahnouni, F., (2003). Surveillance de la qualité des eaux marines du
littoral oranais. Cas des baies de Cap Falcon, des Andalouses et de
Madagh. Mém. Magister, Univ. Oran, Algérie: 145p.
Salerno, M. B., Flamm, M., Logan, B. E., Velegol, D., (2006).
Transport of rodlike colloids through packed beds. Environmental Science
& Technology 40(20):6336-6340.

Références bibliographiques
124
Sanin, S. L., Sanin, F. D., Bryers, J. D., (2003). Effect of starvation on
the adhesive properties of xenobiotic degrading bacteria. Process
Biochemistry 38(6):909-914.
Santamaria, J., Toranzos, G. A., (2003). Enteric pathogens and soil: a
short review. International Microbiology 6(1):5-9.
Santé et Bien Etre Social Canada (SBESC)., (1992). Recommandations
au sujet de la qualité des eaux utilisées à des fins récréatives au Canada,
ministère des Approvisionnements et Services Canada, Ottawa, 110 p.
Sarkar, A. K., Georgiou, G., Sharma, M. M., (1994). Transport of
Bacteria in Porous- Media .1. An Experimental Investigation.
Biotechnology and Bioengineering 44(4):489-497.
Scandura, J. E., Sobsey, M. D., (1997). Viral and bacterial
contamination of groundwater from on-site sewage treatment systems.
Water Science and Technology 35(11-12):141-146.
Schafer, A., Ustohal, P., Harms, H., Stauffer, F., Dracos, T., Zehnder,
A. J. B., (1998). Transport of bacteria in unsaturated porous media.
Journal of Contaminant Hydrology 33(1-2):149-169.
Schubert, R. H. W. (1975). The relation of aerogenic to anaerogenic
aeromonads of the "Aeromonas punctata group" in river water depending
on the load of waste. Zbt. Bakt. Hyg. B160, 237-245.
Schubert, R. H. W. (1991). Aeromonas and their significance as potential
pathogens in water. J. Appl. Bacteriol.- Symposium Sup. 70, 131S-135S.
Sen, T. K., Das, D., Khilar, K. C., Suraishkumar, G. K., (2005).
Bacterial transport in porous media: New aspects of the mathematical
model. Colloids and Surfaces a- Physicochemical and Engineering
Aspects 260(1-3):53-62.

Références bibliographiques
125
Seyfried,P.L., Tobin, R.S., Brown, N.E. et P.F., Ness. (1985). « A
prospective study of swimming-related illness I. Swimming-Associated
Health Risk », Am. J. Public Health, 75 (9) :1068-1070.
Seyefried , P.L., et Harris,E., (1990). Bacteriological characterization
of feces and source differentiation, Ontario Ministry of the Environment,
Water Ressources Branch, Toronto, 149 p.
Signorile G, Montagna MT, Sena G, Cavallo RA (1992)
Bacteriological surveys in waters and sands of Taranto coastal areas.
L’Igiene Moderna, 98(3): 475–483.
Simoni, S. F., Harms, H., Bosma, T. N. P., Zehnder, A. J. B., (1998).
Population heterogeneity affects transport of bacteria through sand
columns at low flow rates. Environmental Science and Technology
32(14):2100-2105.
Singleton,P., and Sainsbury,D., (1994). Dictionary of microbiology and
molecular biology. Wiley.
Sinton L.W., Finlay R.K et Lynch P.A. (1999). Sunlight Inactivation of
fecal bacteriophages and bacteria in sewage polluted seawater. Appl.
Environ. Microbiol., 65, 8, 3605- 3613.
Sinton, L.W., Hall, C. ., Lynch, P.A. , et Daviescolley, R.J. ,(2002).
Sunlight inactivation of fecal indicator bacteria and bacteriophages from
waste stabilization pond effluent in fresh and saline water. Appl. Environ.
Microbiol,.68, 1122-1131.
Smets, B. F., Grasso, D., Engwall, M. A., Machinist, B. J., (1999).
Surface physicochemical properties of Pseudomonas fluorescens and
impact on adhesion and transport through porous media. Colloids and
Surfaces B-Biointerfaces 14(1-4):121-139.

Références bibliographiques
126
Smith, G. W. T., R.E. White D. and Ritonga, (1985). Transport of E.
coli through intact and disturbed columns. Journal of Environmental
Quality 14:87-91.
SOREGAH (1998). Etude d’assainissement du groupement urbain
d’Oran, Juin 1984.
Soussa, M.L.R., (1990) Micoses. [Fungi.] Lisbon, National Institute of
Health (INSA), Centre of Epidemiologic Surveillance of Transmissible
Diseases (Epidemiology Bulletin No. 5).
Stabili,L.,Immacolata,M.,Cavallo,R.A.,(2005).Mytilus galloprovincialis
filter feeding on the bacterial community in a Mediterranean coastal area
(Northern Ionian Sea, Italy). Water Res., 39, 469-477
Stevik, T. K., Ausland, G., Hanssen, J. F., Jenssen, P. D., (1999). The
influence of physical and chemical factors on the transport of E-coli
through biological filters for wastewater purification. Water Research
33(18):3701-3706.
Stevik, T. K., Aa, K., Ausland, G., Hanssen, J. F., (2004). Retention
and removal of pathogenic bacteria in wastewater percolating through
porous media: a review. Water Research 38(6):1355-1367.
Sugiyama. S. and Nikara. C. ( 2004) Differential contribution of
avoidance and tolerance to dehydration resistance in populations of
perennial rye grass, Lolium perenne L. Australian Journal of Agricultural
Research, 55 :33–37.
Tan, Y., Gannon, J. T., Baveye, P., Alexander, M., (1994). Transport of
Bacteria in an Aquifer Sand - Experiments and Model Simulations. water
Resources research 30(12):3243-3252.
Terbeche,M.,(2007). Tendance de la contamination bactériologique et
métallique chez la crevette rouge (Aristeus antennatus). Mém. Magister,
Univ. Oran, Algérie.

Références bibliographiques
127
Thullner, M., Zeyer, J., Kinzelbach, W., (2002). Influence of microbial
growth on hydraulic properties of pore networks. Transport in Porous
Media 49(1):99-122.
Tian, Y. Q., Gong, P., Radke, J. D., Scarborough, J., (2002). Spatial
and temporal modeling of microbial contaminants on grazing farmlands.
Journal of Environmental Quality 31(3):860-869.
Tong, M.P., and Lighthart. B., (1998). Effect of simulated solar
radiation on mixed outdoor atmospheric bacterial populations. FEMS
Microbiology Ecology, 26:311–316.
Tong, M. P., Johnson, W. P., (2006). Excess colloid retention in porous
media as a function of colloid size, fluid velocity, and grain angularity.
Environmental Science and Technology 40(24):7725-7731.
Torsvik, V., and Thingstad, T. F., (2002). Prokaryotic diversity–
magnitude, dynamics, and controlling factors. Science, 296(5570) :1064–
6.
Tsuneda, S., Aikawa, H., Hayashi, H., Yuasa, A., Hirata, A., (2003).
Extracellular polymeric substances responsible for bacterial adhesion onto
solid surface. Fems Microbiology Letters 223(2):287-292.
Unc, A., Goss, M. J., (2004). Transport of bacteria from manure and
protection of water resources. Applied Soil Ecology 25(1):1-18.
UNEP ., (1992) Health risks from bathing in marine waters.
Mediterranean Marine Pollution Monitoring and Research Programme
(MED POL) Phase II. Annex 5. Copenhagen, WHO Regional Office for
Europe, p. 63.
UNEP/FAO/IOC/IAEA., (1993) Guidelines for monitoring chemical
contaminants in the sea using marine organisms. Reference Methods for
Marine Pollution Studies No. 6. UNEP.

Références bibliographiques
128
US Environmental Protection Agency ., (1997). Response to congress
on use of decentralized wastewater treatment systems. Washington,
DC.
van der Wielen, P., Bolhuis, H., Borin, S., Daffonchio, D. , Corselli,
C. , Giuliano, L. , D’Auria, G. , de Lange, G. , Huebner, A., Varnavas,
S. P. , Thomson, J. , Tamburini, C., Marty, D., McGenity, T. J. ,
Timmis, K. N., and Party, B. S., ( 2005). The enigma of prokaryotic life
in deep hypersaline anoxic basins. Science, 307(5706) :121–3.
Van Der,L., Kooij, D. (1991) Nutritional requirements of Aeromonas
and their multiplication in drinking water. Experientia 47,444-469.
Van Loosdrecht, M. C., Lyklema, J., Norde, W., Schraa, G., Zehnder,
A. J., (1987). Electrophoretic mobility and hydrophobicity as a measured
to predict the initial steps of bacterial adhesion. Applied and
Environmental Microbiology 53(8):1898-1901.
Van Loosdrecht, M. C. M., J. Lyklema, W. Norde, and A.J.B.
Zehnder, (1989). Bacterial adhesion : A physochemical approach.
Microbial Ecology(17):1-15.
Van Loosdrecht, M. C., Norde, W., Lyklema, L. and Zehnder, J.,
(1990). Hydrophobic and electrostatic parameters in bacterial adhesion.
Aquatic Science 51(103-114).
Van Oss, C. J., (1994). Interfacial forces in aqueous media. Interfacial
forces in aqueous media. New York: Dekker.
Vidal, O., Longin, R., Prigent-Combaret, C., Dorel, C., Hooreman,
M., Lejeune, P., (1998). Isolation of an Escherichia coli K-12 mutant
strain able to form biofilms on inert surfaces: Involvement of a new ompR
allele that increases curli expression. Journal of Bacteriology
180(9):2442-2449.

Références bibliographiques
129
Walker, S.E. , Mostaghimi, S., Dillaha, T.A. and Woeste, F.E., (2004)
Modeling animal waste management practice: impacts on bacteria levels
in runoff from agricultural lands, Transations of the ASAE, 33 (3): 807-
817.
Wan, J. M., Wilson, J. L., Kieft, T. L., (1994). Influence of the Gas-
Water Interface on Transport of Microorganisms through Unsaturated
Porous-Media. Applied and Environmental Microbiology 60(2):509-516.
Ward, J. B., and Berkeley R.C.W., (1980). The microbial cell surface
and adhesion. In: Berkeley, R. C. W., Lynch J.M., Melling J., Rutter P.R.
and Vincent D., editor. Microbial adhesion to surfaces. Chichester: Ellis
Horwood. p 47-66.
Webb, J. S., Van der Mei, H. C., Nixon, M., Eastwood, I. M.,
Greenhalgh, M., Read, S. J., Robson, G. D., Handley, P. S., (1999).
Plasticizers increase adhesion of the deteriogenic fungus Aureobasidium
pullulans to polyvinyl chloride. Applied and Environmental Microbiology
65(8):3575-3581.
Weiss, T. H., Mills, A. L., Hornberger, G. M., Herman, J. S., (1995).
Effect of Bacterial-Cell Shape on Transport of Bacteria in Porous-Media.
Environmental Science and Technology 29(7):1737-1740.
WHO/UNEP., (1992) Health risks from bathing in marine waters.
Mediterranean Marine Pollution Monitoring and Research Programme
(MED POL) Phase II. Annex 5. Copenhagen, WHO Regional Office for
Europe, p. 63.
WHO/UNEP., (1994) Microbiological quality of coastal recreational
waters. Mediterranean Marine Pollution Monitoring and Research
Programme (MED POL) Phase II. Annex 1. Copenhagen, WHO Regional
Office for Europe, p. 5.

Références bibliographiques
130
Wiklund, T., (1995). Survival of atypical Aeromonas salmonicida in
water and sediment microcosms of different salinities and temperature.
Dis Aquat. Org., 21, 137-143.
Wood, E. J. F.,(1959) Some aspects of marine microbiology. J. Mar.
Biol. Assoc. India, , 1, 26–32.
Wu, Y., Clevenger, T. , and Deng, B.,(2005). Impacts of goethite
particles on UV disinfection of drinking water. Applied and
Environmental Microbiology, 71 :4140–4143..
Yao, K. M., Habibian, M.T., O'Melia, C.R., (1971). Water and waste-
water filtration: concepts and applications. Environmental Science &
Technology(5):1105-1112.
Zhou,J., Xia,B., Treves,D.s., Wu,L.Y., Marsh,T.L., O’Neill,
R.V.,Palumbo,V., and Tiedje,J.M., (2002) Spatial and resource factors
influencing high microbial diversity in soil. Appl Environ Microbiol,
68(1) :326–34.
Zita, A., Hermansson, M., (1997). Effects of bacterial cell surface
structures and hydrophobicity on attachment to activated sludge flocs.
Applied and Environmental Microbiology 63(3):1168-1170.
Zmyslowska,l; Gola, I., Filipkowska,Z., Lewandowska,D., (2002).
Potentially pathogenic bacteria from the family Enterobacteriaceae,
Pseudomonas sp. and Aeromonas sp. in waters designated for drinking
and household purposes. Pol. J. Environ. Stud. 11 (4): 325-330.

Annexes
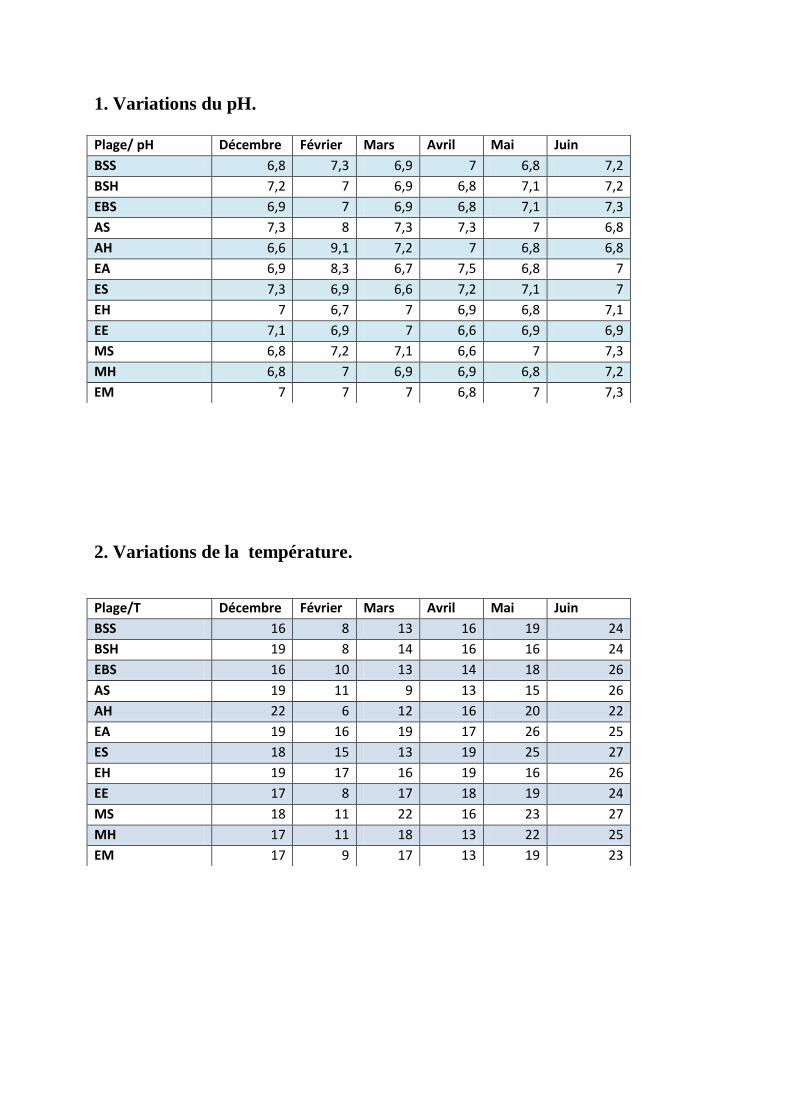
1. Variations du pH.
2. Variations de la température.
Plage/ pH Décembre Février Mars Avril Mai Juin
BSS 6,8 7,3 6,9 7 6,8 7,2
BSH 7,2 7 6,9 6,8 7,1 7,2
EBS 6,9 7 6,9 6,8 7,1 7,3
AS 7,3 8 7,3 7,3 7 6,8
AH 6,6 9,1 7,2 7 6,8 6,8
EA 6,9 8,3 6,7 7,5 6,8 7
ES 7,3 6,9 6,6 7,2 7,1 7
EH 7 6,7 7 6,9 6,8 7,1
EE 7,1 6,9 7 6,6 6,9 6,9
MS 6,8 7,2 7,1 6,6 7 7,3
MH 6,8 7 6,9 6,9 6,8 7,2
EM 7 7 7 6,8 7 7,3
Plage/T Décembre Février Mars Avril Mai Juin
BSS 16 8 13 16 19 24
BSH 19 8 14 16 16 24
EBS 16 10 13 14 18 26
AS 19 11 9 13 15 26
AH 22 6 12 16 20 22
EA 19 16 19 17 26 25
ES 18 15 13 19 25 27
EH 19 17 16 19 16 26
EE 17 8 17 18 19 24
MS 18 11 22 16 23 27
MH 17 11 18 13 22 25
EM 17 9 17 13 19 23
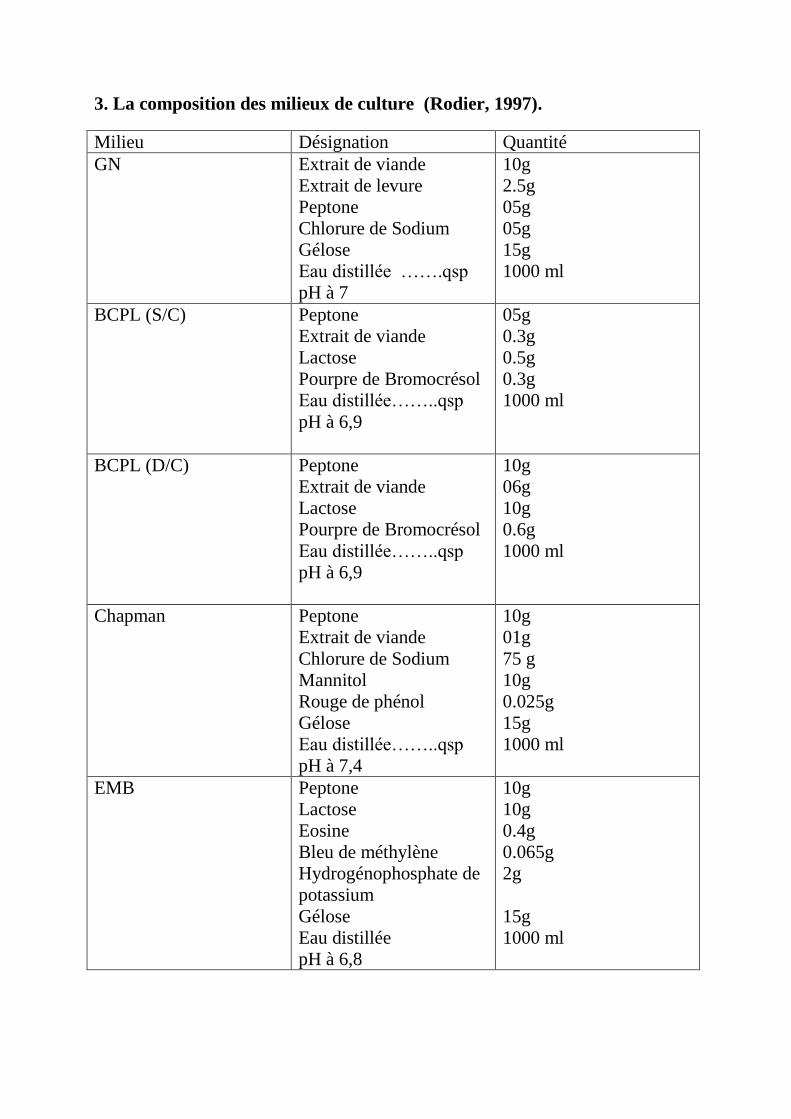
3. La composition des milieux de culture (Rodier, 1997).
Milieu Désignation Quantité
GN Extrait de viande
Extrait de levure
Peptone
Chlorure de Sodium
Gélose
Eau distillée …….qsp
pH à 7
10g
2.5g
05g
05g
15g
1000 ml
BCPL (S/C) Peptone
Extrait de viande
Lactose
Pourpre de Bromocrésol
Eau distillée……..qsp
pH à 6,9
05g
0.3g
0.5g
0.3g
1000 ml
BCPL (D/C) Peptone
Extrait de viande
Lactose
Pourpre de Bromocrésol
Eau distillée……..qsp
pH à 6,9
10g
06g
10g
0.6g
1000 ml
Chapman Peptone
Extrait de viande
Chlorure de Sodium
Mannitol
Rouge de phénol
Gélose
Eau distillée……..qsp
pH à 7,4
10g
01g
75 g
10g
0.025g
15g
1000 ml
EMB Peptone
Lactose
Eosine
Bleu de méthylène
Hydrogénophosphate de
potassium
Gélose
Eau distillée
pH à 6,8
10g
10g
0.4g
0.065g
2g
15g
1000 ml
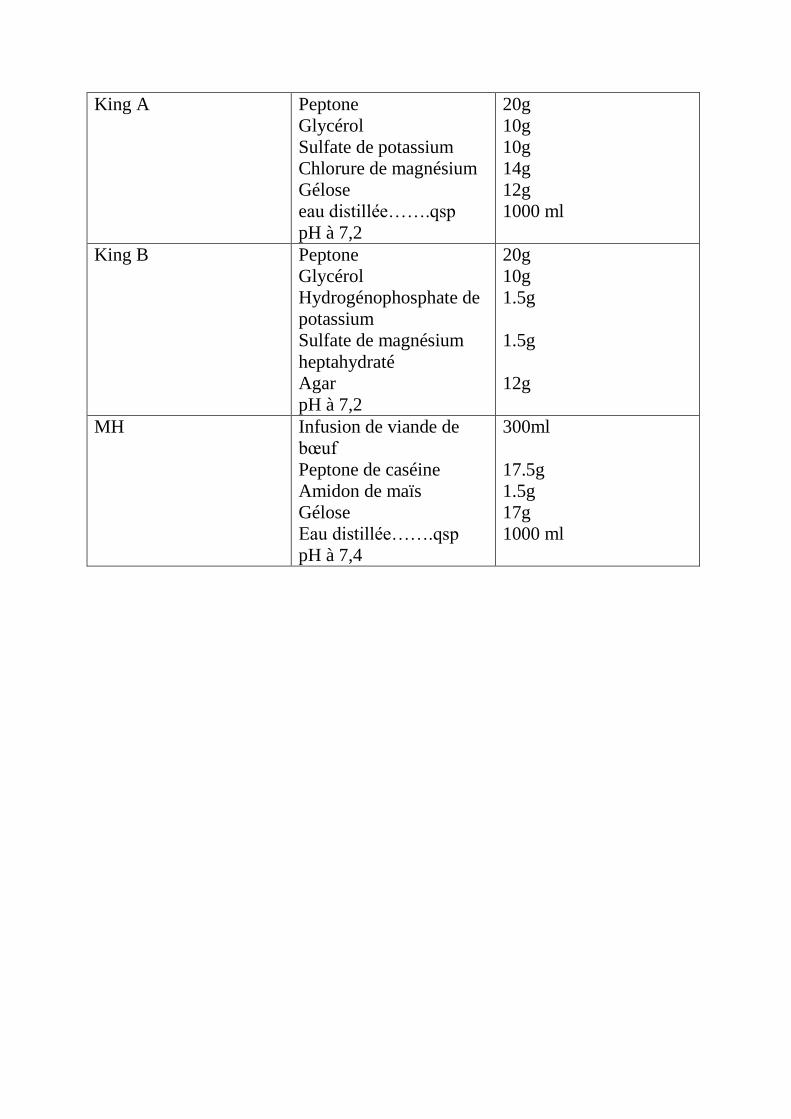
King A Peptone
Glycérol
Sulfate de potassium
Chlorure de magnésium
Gélose
eau distillée…….qsp
pH à 7,2
20g
10g
10g
14g
12g
1000 ml
King B Peptone
Glycérol
Hydrogénophosphate de
potassium
Sulfate de magnésium
heptahydraté
Agar
pH à 7,2
20g
10g
1.5g
1.5g
12g
MH Infusion de viande de
bœuf
Peptone de caséine
Amidon de maïs
Gélose
Eau distillée…….qsp
pH à 7,4
300ml
17.5g
1.5g
17g
1000 ml
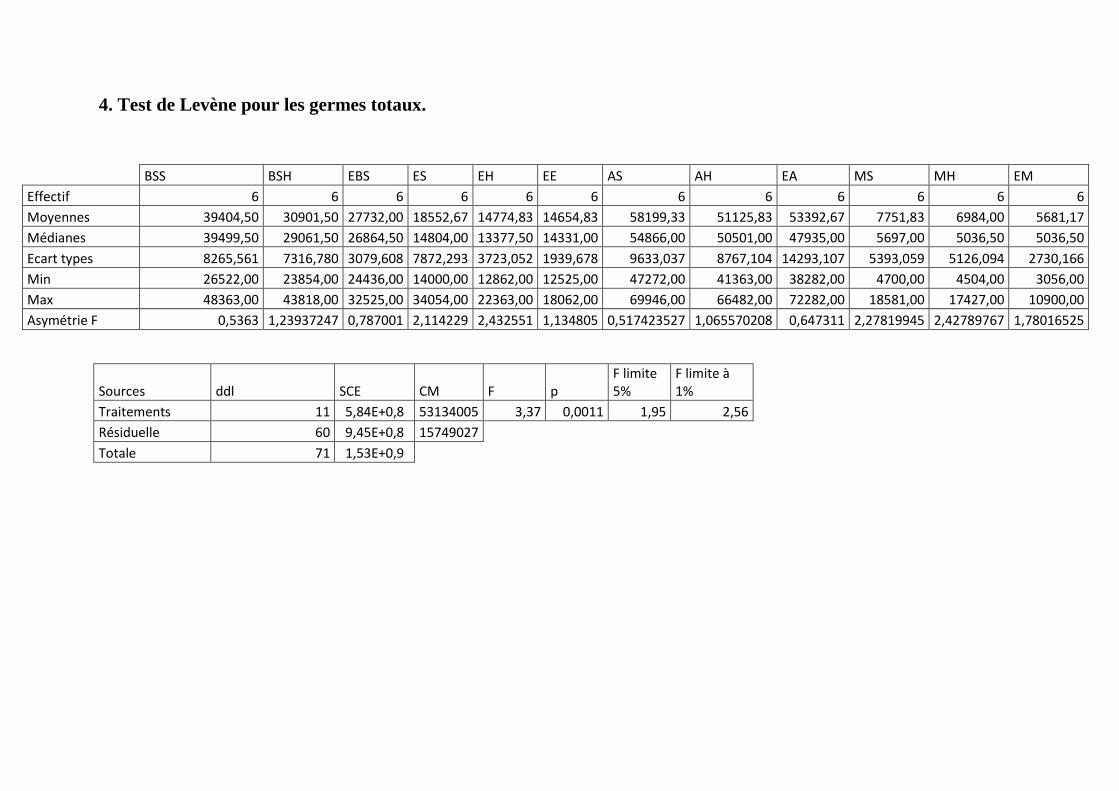
4. Test de Levène pour les germes totaux.
BSS BSH EBS ES EH EE AS AH EA MS MH EM
Effectif 6 6 6 6 6 6 6 6 6 6 6 6
Moyennes 39404,50 30901,50 27732,00 18552,67 14774,83 14654,83 58199,33 51125,83 53392,67 7751,83 6984,00 5681,17
Médianes 39499,50 29061,50 26864,50 14804,00 13377,50 14331,00 54866,00 50501,00 47935,00 5697,00 5036,50 5036,50
Ecart types 8265,561 7316,780 3079,608 7872,293 3723,052 1939,678 9633,037 8767,104 14293,107 5393,059 5126,094 2730,166
Min 26522,00 23854,00 24436,00 14000,00 12862,00 12525,00 47272,00 41363,00 38282,00 4700,00 4504,00 3056,00
Max 48363,00 43818,00 32525,00 34054,00 22363,00 18062,00 69946,00 66482,00 72282,00 18581,00 17427,00 10900,00
Asymétrie F 0,5363 1,23937247 0,787001 2,114229 2,432551 1,134805 0,517423527 1,065570208 0,647311 2,27819945 2,42789767 1,78016525
Sources ddl SCE CM F p F limite 5%
F limite à 1%
Traitements 11 5,84E+0,8 53134005 3,37 0,0011 1,95 2,56
Résiduelle 60 9,45E+0,8 15749027 Totale 71 1,53E+0,9
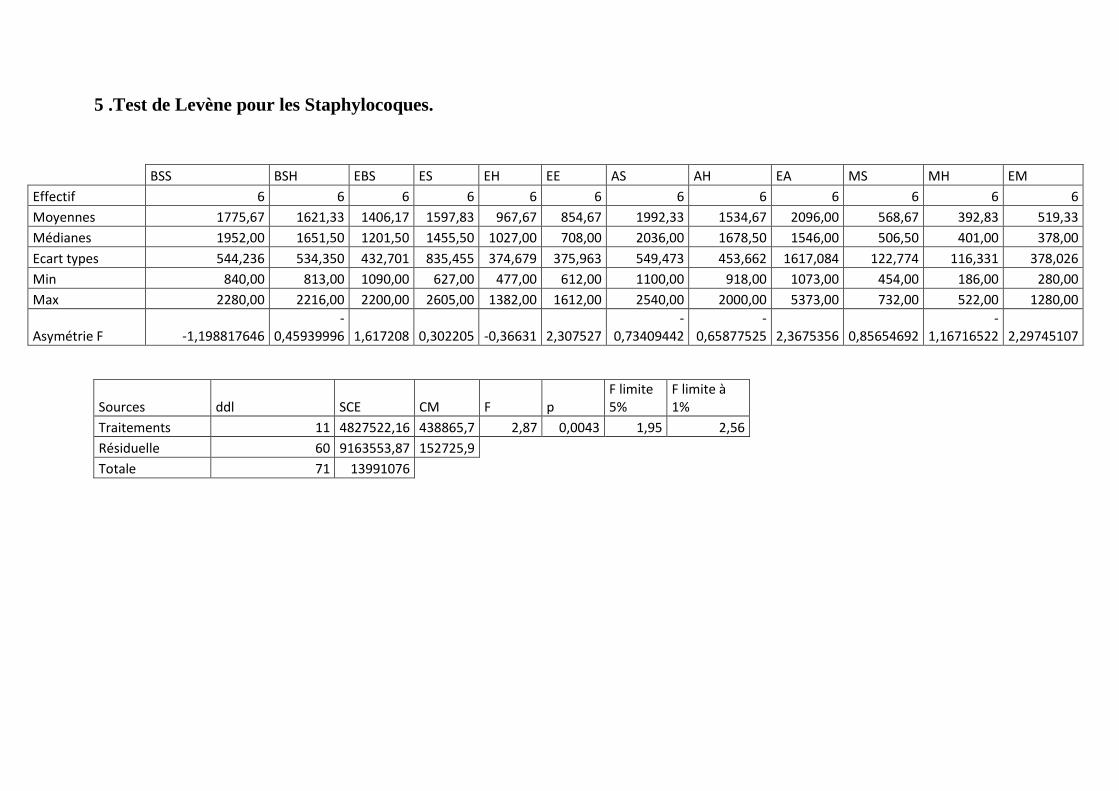
5 .Test de Levène pour les Staphylocoques.
BSS BSH EBS ES EH EE AS AH EA MS MH EM
Effectif 6 6 6 6 6 6 6 6 6 6 6 6
Moyennes 1775,67 1621,33 1406,17 1597,83 967,67 854,67 1992,33 1534,67 2096,00 568,67 392,83 519,33
Médianes 1952,00 1651,50 1201,50 1455,50 1027,00 708,00 2036,00 1678,50 1546,00 506,50 401,00 378,00
Ecart types 544,236 534,350 432,701 835,455 374,679 375,963 549,473 453,662 1617,084 122,774 116,331 378,026
Min 840,00 813,00 1090,00 627,00 477,00 612,00 1100,00 918,00 1073,00 454,00 186,00 280,00
Max 2280,00 2216,00 2200,00 2605,00 1382,00 1612,00 2540,00 2000,00 5373,00 732,00 522,00 1280,00
Asymétrie F -1,198817646 -
0,45939996 1,617208 0,302205 -0,36631 2,307527 -
0,73409442 -
0,65877525 2,3675356 0,85654692 -
1,16716522 2,29745107
Sources ddl SCE CM F p F limite 5%
F limite à 1%
Traitements 11 4827522,16 438865,7 2,87 0,0043 1,95 2,56
Résiduelle 60 9163553,87 152725,9 Totale 71 13991076
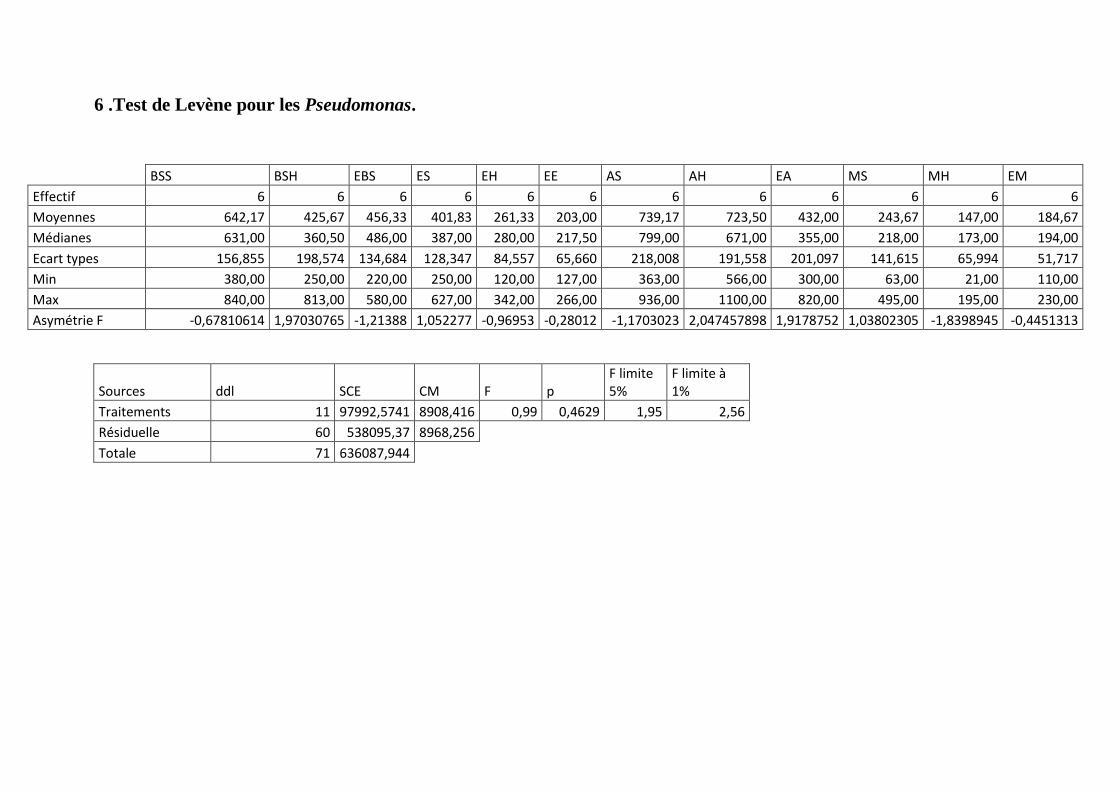
6 .Test de Levène pour les Pseudomonas.
BSS BSH EBS ES EH EE AS AH EA MS MH EM
Effectif 6 6 6 6 6 6 6 6 6 6 6 6
Moyennes 642,17 425,67 456,33 401,83 261,33 203,00 739,17 723,50 432,00 243,67 147,00 184,67
Médianes 631,00 360,50 486,00 387,00 280,00 217,50 799,00 671,00 355,00 218,00 173,00 194,00
Ecart types 156,855 198,574 134,684 128,347 84,557 65,660 218,008 191,558 201,097 141,615 65,994 51,717
Min 380,00 250,00 220,00 250,00 120,00 127,00 363,00 566,00 300,00 63,00 21,00 110,00
Max 840,00 813,00 580,00 627,00 342,00 266,00 936,00 1100,00 820,00 495,00 195,00 230,00
Asymétrie F -0,67810614 1,97030765 -1,21388 1,052277 -0,96953 -0,28012 -1,1703023 2,047457898 1,9178752 1,03802305 -1,8398945 -0,4451313
Sources ddl SCE CM F p F limite 5%
F limite à 1%
Traitements 11 97992,5741 8908,416 0,99 0,4629 1,95 2,56
Résiduelle 60 538095,37 8968,256 Totale 71 636087,944
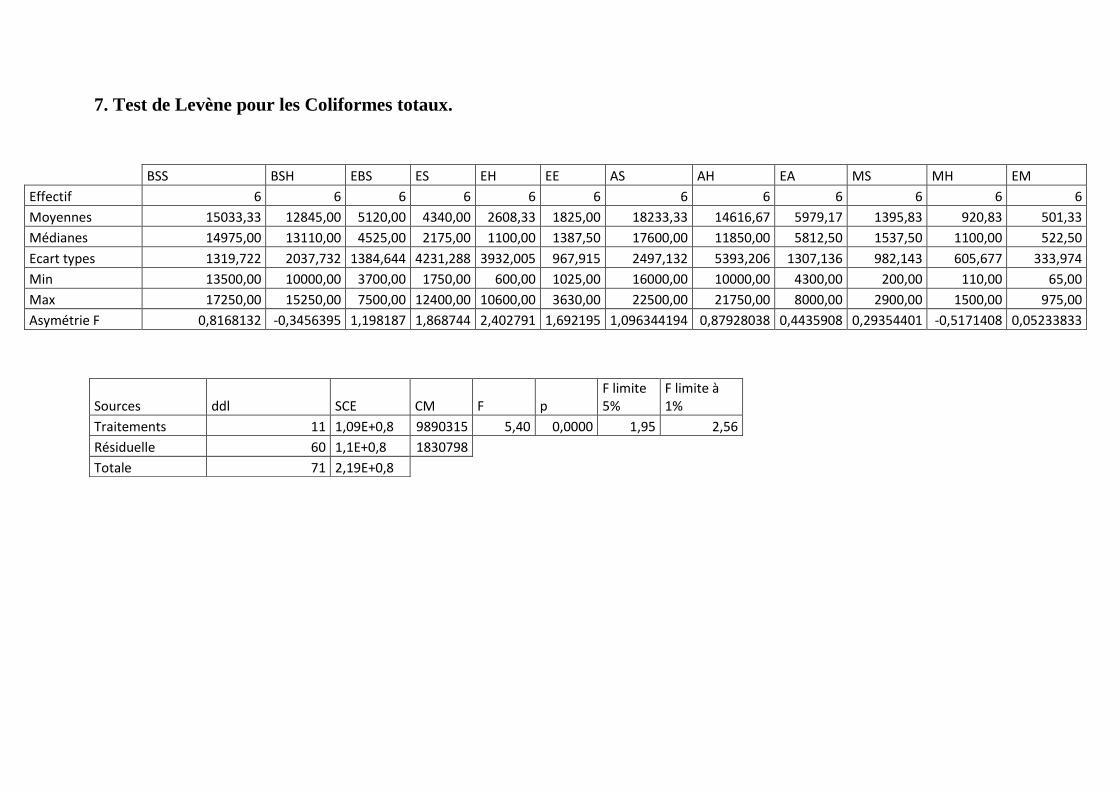
7. Test de Levène pour les Coliformes totaux.
BSS BSH EBS ES EH EE AS AH EA MS MH EM
Effectif 6 6 6 6 6 6 6 6 6 6 6 6
Moyennes 15033,33 12845,00 5120,00 4340,00 2608,33 1825,00 18233,33 14616,67 5979,17 1395,83 920,83 501,33
Médianes 14975,00 13110,00 4525,00 2175,00 1100,00 1387,50 17600,00 11850,00 5812,50 1537,50 1100,00 522,50
Ecart types 1319,722 2037,732 1384,644 4231,288 3932,005 967,915 2497,132 5393,206 1307,136 982,143 605,677 333,974
Min 13500,00 10000,00 3700,00 1750,00 600,00 1025,00 16000,00 10000,00 4300,00 200,00 110,00 65,00
Max 17250,00 15250,00 7500,00 12400,00 10600,00 3630,00 22500,00 21750,00 8000,00 2900,00 1500,00 975,00
Asymétrie F 0,8168132 -0,3456395 1,198187 1,868744 2,402791 1,692195 1,096344194 0,87928038 0,4435908 0,29354401 -0,5171408 0,05233833
Sources ddl SCE CM F p F limite 5%
F limite à 1%
Traitements 11 1,09E+0,8 9890315 5,40 0,0000 1,95 2,56
Résiduelle 60 1,1E+0,8 1830798 Totale 71 2,19E+0,8
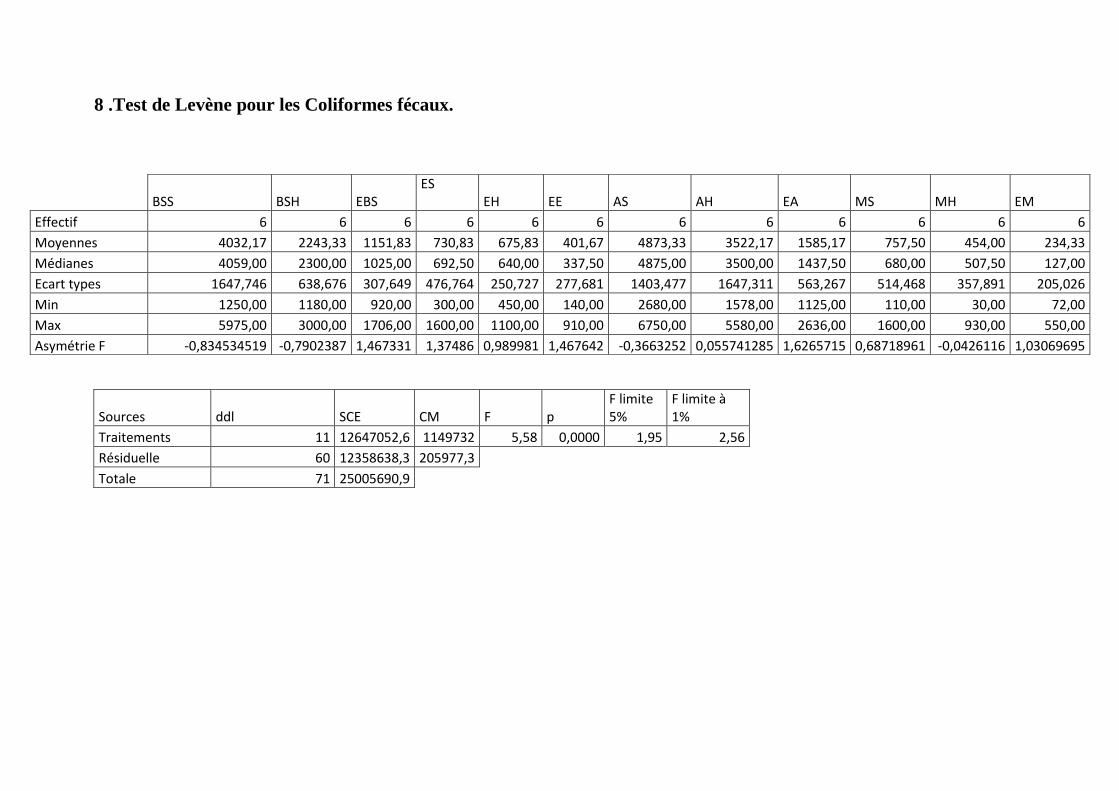
8 .Test de Levène pour les Coliformes fécaux.
Sources ddl SCE CM F p F limite 5%
F limite à 1%
Traitements 11 12647052,6 1149732 5,58 0,0000 1,95 2,56
Résiduelle 60 12358638,3 205977,3 Totale 71 25005690,9
BSS BSH EBS
ES EH EE AS AH EA MS MH EM
Effectif 6 6 6 6 6 6 6 6 6 6 6 6
Moyennes 4032,17 2243,33 1151,83 730,83 675,83 401,67 4873,33 3522,17 1585,17 757,50 454,00 234,33
Médianes 4059,00 2300,00 1025,00 692,50 640,00 337,50 4875,00 3500,00 1437,50 680,00 507,50 127,00
Ecart types 1647,746 638,676 307,649 476,764 250,727 277,681 1403,477 1647,311 563,267 514,468 357,891 205,026
Min 1250,00 1180,00 920,00 300,00 450,00 140,00 2680,00 1578,00 1125,00 110,00 30,00 72,00
Max 5975,00 3000,00 1706,00 1600,00 1100,00 910,00 6750,00 5580,00 2636,00 1600,00 930,00 550,00
Asymétrie F -0,834534519 -0,7902387 1,467331 1,37486 0,989981 1,467642 -0,3663252 0,055741285 1,6265715 0,68718961 -0,0426116 1,03069695
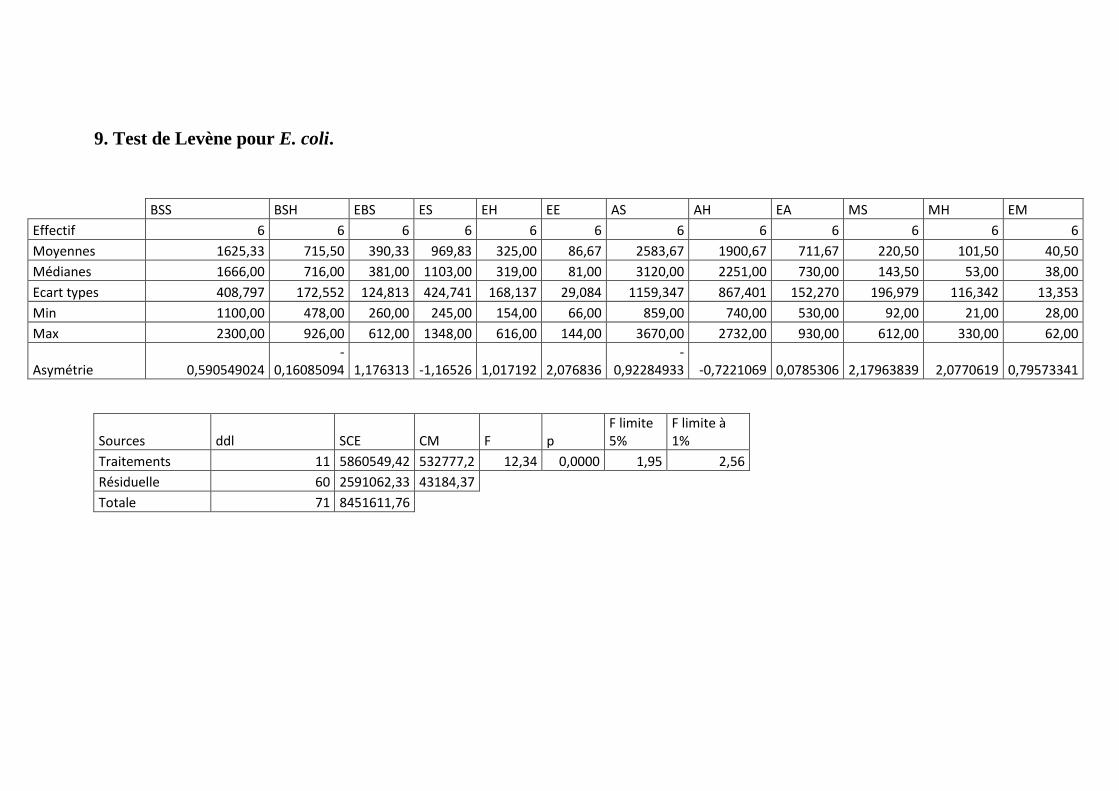
9. Test de Levène pour E. coli.
BSS BSH EBS ES EH EE AS AH EA MS MH EM
Effectif 6 6 6 6 6 6 6 6 6 6 6 6
Moyennes 1625,33 715,50 390,33 969,83 325,00 86,67 2583,67 1900,67 711,67 220,50 101,50 40,50
Médianes 1666,00 716,00 381,00 1103,00 319,00 81,00 3120,00 2251,00 730,00 143,50 53,00 38,00
Ecart types 408,797 172,552 124,813 424,741 168,137 29,084 1159,347 867,401 152,270 196,979 116,342 13,353
Min 1100,00 478,00 260,00 245,00 154,00 66,00 859,00 740,00 530,00 92,00 21,00 28,00
Max 2300,00 926,00 612,00 1348,00 616,00 144,00 3670,00 2732,00 930,00 612,00 330,00 62,00
Asymétrie 0,590549024 -
0,16085094 1,176313 -1,16526 1,017192 2,076836 -
0,92284933 -0,7221069 0,0785306 2,17963839 2,0770619 0,79573341
Sources ddl SCE CM F p F limite 5%
F limite à 1%
Traitements 11 5860549,42 532777,2 12,34 0,0000 1,95 2,56
Résiduelle 60 2591062,33 43184,37 Totale 71 8451611,76