Embed Size (px)
Citation preview

ECOLE NATIONALE VETERINAIRE DE NANTES ANNEE 2008
Etude de la pathogénicité d’Anaplasma phagocytophilum chez les populations de
chevreuils (Capreolus capreolus) de Franche Comté
THESE Pour le diplôme d’Etat
De DOCTEUR VETERINAIRE
Présentée et soutenue publiquement Le
Devant la faculté de médecine de Nantes Par
Julien PORTIER Né le 09 février 1985 à Clamart
JURY
Président : Monsieur Michel Marjolet Professeur à la faculté de médecine de Nantes
Membres : Madame Suzanne Bastian-Orange
Maître de Conférences à l’Ecole Nationale Vétérinaire de Nantes Monsieur Albert Agoulon Maître de Conférences à l’Ecole Nationale Vétérinaire de Nantes

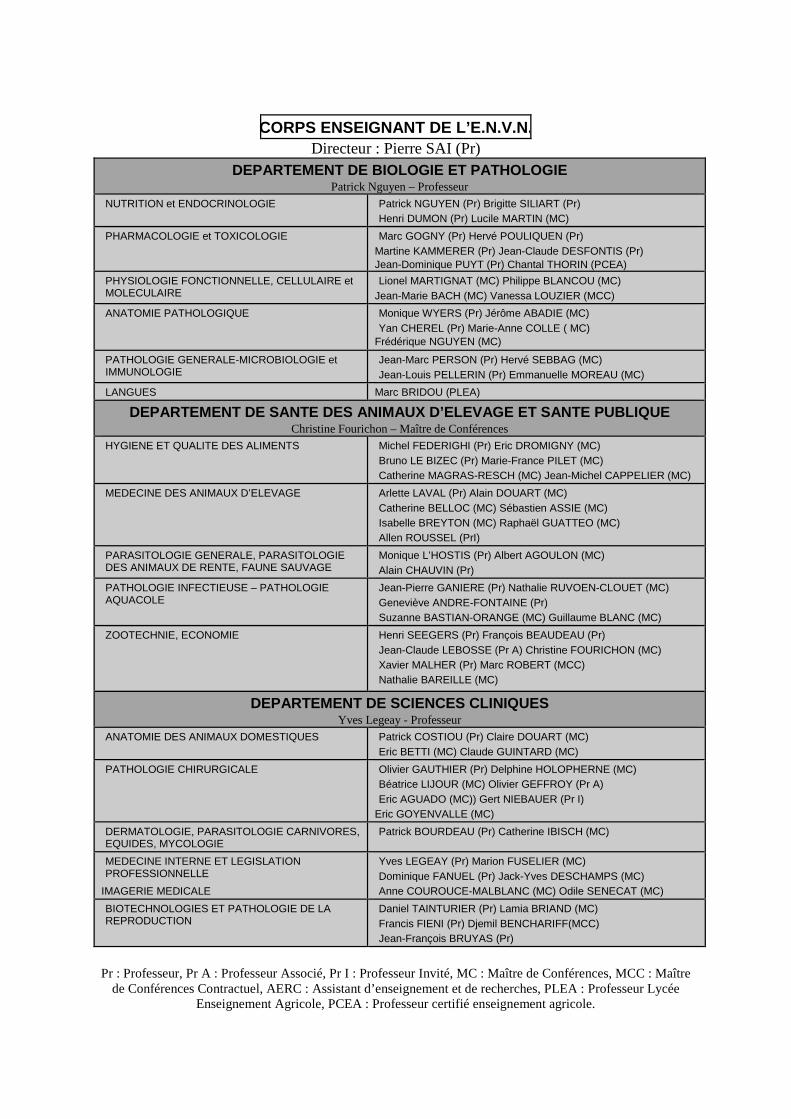
CORPS ENSEIGNANT DE L’E.N.V.N. Directeur : Pierre SAI (Pr)
DEPARTEMENT DE BIOLOGIE ET PATHOLOGIE Patrick Nguyen – Professeur
NUTRITION et ENDOCRINOLOGIE Patrick NGUYEN (Pr) Brigitte SILIART (Pr) Henri DUMON (Pr) Lucile MARTIN (MC)
PHARMACOLOGIE et TOXICOLOGIE Marc GOGNY (Pr) Hervé POULIQUEN (Pr) Martine KAMMERER (Pr) Jean-Claude DESFONTIS (Pr) Jean-Dominique PUYT (Pr) Chantal THORIN (PCEA)
PHYSIOLOGIE FONCTIONNELLE, CELLULAIRE et MOLECULAIRE
Lionel MARTIGNAT (MC) Philippe BLANCOU (MC) Jean-Marie BACH (MC) Vanessa LOUZIER (MCC)
ANATOMIE PATHOLOGIQUE Monique WYERS (Pr) Jérôme ABADIE (MC) Yan CHEREL (Pr) Marie-Anne COLLE ( MC)
Frédérique NGUYEN (MC)
PATHOLOGIE GENERALE-MICROBIOLOGIE et IMMUNOLOGIE
Jean-Marc PERSON (Pr) Hervé SEBBAG (MC) Jean-Louis PELLERIN (Pr) Emmanuelle MOREAU (MC)
LANGUES Marc BRIDOU (PLEA)
DEPARTEMENT DE SANTE DES ANIMAUX D’ELEVAGE ET SANTE PUBLIQUE Christine Fourichon – Maître de Conférences
HYGIENE ET QUALITE DES ALIMENTS Michel FEDERIGHI (Pr) Eric DROMIGNY (MC) Bruno LE BIZEC (Pr) Marie-France PILET (MC) Catherine MAGRAS-RESCH (MC) Jean-Michel CAPPELIER (MC)
MEDECINE DES ANIMAUX D’ELEVAGE Arlette LAVAL (Pr) Alain DOUART (MC) Catherine BELLOC (MC) Sébastien ASSIE (MC) Isabelle BREYTON (MC) Raphaël GUATTEO (MC) Allen ROUSSEL (PrI)
PARASITOLOGIE GENERALE, PARASITOLOGIE DES ANIMAUX DE RENTE, FAUNE SAUVAGE
Monique L’HOSTIS (Pr) Albert AGOULON (MC) Alain CHAUVIN (Pr)
PATHOLOGIE INFECTIEUSE – PATHOLOGIE AQUACOLE
Jean-Pierre GANIERE (Pr) Nathalie RUVOEN-CLOUET (MC) Geneviève ANDRE-FONTAINE (Pr) Suzanne BASTIAN-ORANGE (MC) Guillaume BLANC (MC)
ZOOTECHNIE, ECONOMIE Henri SEEGERS (Pr) François BEAUDEAU (Pr) Jean-Claude LEBOSSE (Pr A) Christine FOURICHON (MC) Xavier MALHER (Pr) Marc ROBERT (MCC) Nathalie BAREILLE (MC)
DEPARTEMENT DE SCIENCES CLINIQUES Yves Legeay - Professeur
ANATOMIE DES ANIMAUX DOMESTIQUES Patrick COSTIOU (Pr) Claire DOUART (MC) Eric BETTI (MC) Claude GUINTARD (MC)
PATHOLOGIE CHIRURGICALE Olivier GAUTHIER (Pr) Delphine HOLOPHERNE (MC) Béatrice LIJOUR (MC) Olivier GEFFROY (Pr A) Eric AGUADO (MC)) Gert NIEBAUER (Pr I)
Eric GOYENVALLE (MC)
DERMATOLOGIE, PARASITOLOGIE CARNIVORES, EQUIDES, MYCOLOGIE
Patrick BOURDEAU (Pr) Catherine IBISCH (MC)
MEDECINE INTERNE ET LEGISLATION PROFESSIONNELLE
IMAGERIE MEDICALE
Yves LEGEAY (Pr) Marion FUSELIER (MC) Dominique FANUEL (Pr) Jack-Yves DESCHAMPS (MC) Anne COUROUCE-MALBLANC (MC) Odile SENECAT (MC)
BIOTECHNOLOGIES ET PATHOLOGIE DE LA REPRODUCTION
Daniel TAINTURIER (Pr) Lamia BRIAND (MC) Francis FIENI (Pr) Djemil BENCHARIFF(MCC) Jean-François BRUYAS (Pr)
Pr : Professeur, Pr A : Professeur Associé, Pr I : Professeur Invité, MC : Maître de Conférences, MCC : Maître
de Conférences Contractuel, AERC : Assistant d’enseignement et de recherches, PLEA : Professeur Lycée Enseignement Agricole, PCEA : Professeur certifié enseignement agricole.


Remerciements A Monsieur le professeur Michel Marjolet,
pour nous avoir fait l’honneur de présider cette thèse,
hommage respectueux.
Au Docteur Suzanne Bastian-Orange,
qui a accepté d’assurer la direction de cette thèse,
pour nos discussion passionnantes dans et hors sujet,
Hommages respectueux.
Au, Docteur Raphaël Guattéo,
pour son œil acéré et assuré d’assesseur,
Hommage respectueux.
Nous remercions également
Professeur Monique L’Hostis
pour la confiance que vous m’avez témoignée et pour la réciproque,
toutes mes amitiés.
Docteur Charlotte Dunoyer
pour avoir été le maître de stage que tout étudiant voudrait avoir,
pour votre calme, votre confiance et votre humour,
profonde gratitude.
Docteurs Guy Joncour et Philippe Gibert,
pour vos conseils et remarques avisées,
merci.
Mademoiselle Charlette Chandosné,
Pour votre disponibilité et pour le plaisir que j’ai eu à ramasser des productions de lynx avec vous,
Merci et bonne chasse.
L’ensemble des personnes ayant collaboré de près de loin à ce travail,
M. Viry, M. Hesseman, M. leHong, Mme Vassalo et M.Nicollet, pour leurs qualités d’analyse quel que soit le domaine…
M. Marillier, M. Martin, M. Clerc et M. deMeulemester pour leur disponibilité et pour m’avoir donné envie de revenir en Franche-Comté
A tous ces amis de la FNC,
à bientôt !
A tous les gens qui ont permis par leur travail sur le terrain l’accomplissement de ce travail, Bravo !


Remerciements personnels A…
Ma famille, que je ne citerai pas intégralement, l’objet de cette thèse n’étant pas d’écrire une encyclopédie des prénoms, mais quand même :
Papa, Maman, Benjy, Robin, Ghislain, Duncan, Tristan et Marin.
N’oublions pas Papy, Mamie,
and don’t forget GrandMa, Jenny and Steve
Je vous remercie pour votre soutien
A ces amis que je connais depuis longtemps et, j’espère, pour longtemps
A Vincent, qui parmi tous m’est le plus cher,
Et pour tous les plaisirs que nous avons partagé
A Gabriel, un ami qui vous veut du bien
A Cloé, mon amie la plus ancienne
« The Bat kindly salutes you »
A Marion, colloc’ des grandes heures et des veillées pré-examinatoires
Viel Glück !
Aux Minimoys, à Nono et à Blanche Neige qui donnent une teinte banlieue à ma vie
Aux amis retrouvés récemment…
A cette belle-famille que je supporte, finalement….
Monsieur et Madame Le Gal, pour leur soutien moral et parfois pour leur mécénat bienveillant.
A Chris, pour sa fraîcheur sauf au petit matin
A mon chat primordial
A ce chien qui parfois à l’air heureux, parfois seulement, pauvre victime
Je terminerai par une dernière catégorie, la femme qu’on aime parce qu’on la connaît jusqu’au bout des doigts…
A tout ces amis qui savent que s’ils ne sont pas cités, c’est uniquement par manque de place et qu’il y aura toujours une place pour eux…. A la votre


« On aurait pu dire…Oh !Dieu !...Bien des choses en somme… » Cyrano de Bergerac, Edmond Rostand


1
Sommaire
Introduction
Première partie: étude bibliographique
.I. BIOLOGIE DU CHEVREUIL
.II. ETUDE BIBLIOGRAPHIQUE DE A. PHAGOCYTOPHILUM
Deuxième partie: étude expérimentale
.I. CADRE ET OBJECTIFS DE CETTE ETUDE
.II. ETUDE EXPERIMENTALE : ETUDE DU ROLE PATHOGENE D’A. PHAGOCYTOPHILUM DANS LA
POPULATION DE CHEVREUILS DE FRANCHE-COMTE SUR LA SAISON DE CHASSE 2007/2008
Discussion
.I. ETUDE CAS-TEMOIN : UN PORTAGE PLUS IMPORTANT CHEZ LES MALADES
.II. DES PREVALENCES ET SEROPREVALENCE ELEVEES ET UNE CIRCULATION HOMOGENE
.III. UNE SAISON 2007/2008 MARQUEE PAR UNE FAIBLE MORTALITE
.IV. UNE BACTERIE RETROUVEE TOUT AU LONG DE L’HIVER DANS LE SANG DES CHEVREUILS
.V. LE ROLE POTENTIEL DU CHEVREUIL DANS LE CYCLE EPIDEMIOLOGIQUE D’A. PHAGOCYTOPHILUM
.VI. INTEGRATION DES DONNEES DE SUIVI DES POPULATIONS
.VII. LE ROLE PATHOGENE D’A. PHAGOCYTOPHILUM DANS LES POPULATIONS DE CHEVREUILS DE
FRANCHE-COMTE EN 2007/2008 ET AVANT
Conclusion
Références bibliographiques
Annexes

2

3
Liste des figures
Figure 1: Répartition du Chevreuil en France (Réseau ONCFS/FNC/FDC,"ongulés sauvages") .......... 11
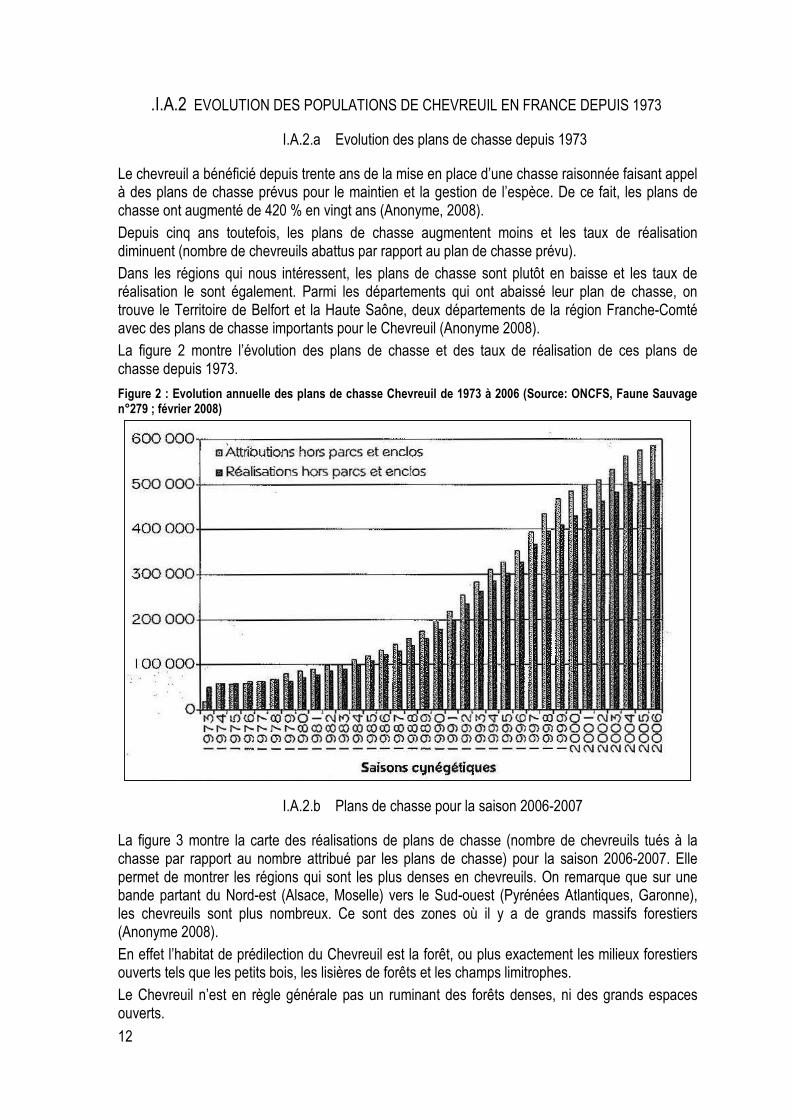
Figure 2 : Evolution annuelle des plans de chasse Chevreuil de 1973 à 2006 (Source: ONCFS, Faune Sauvage n°279 ; février 2008) ............................................................................................................... 12
Figure 3 : Réalisations de plans de chasse en France en 2006/2007 (Source: ONCFS, Faune Sauvage n°279 ; février 2008) .............................................................................................................................. 13
Figure 4: Schémas détaillés de la tête du Chevreuil et traits caractéristiques, vue de profil gauche (g) et vue de face (d) (Boisaubert & Boutin, 1988) .......................................................................................... 14
Figure 5: Le cycle annuel des bois du Chevreuil (d’après Boisaubert & Boutin, 1988) .......................... 15
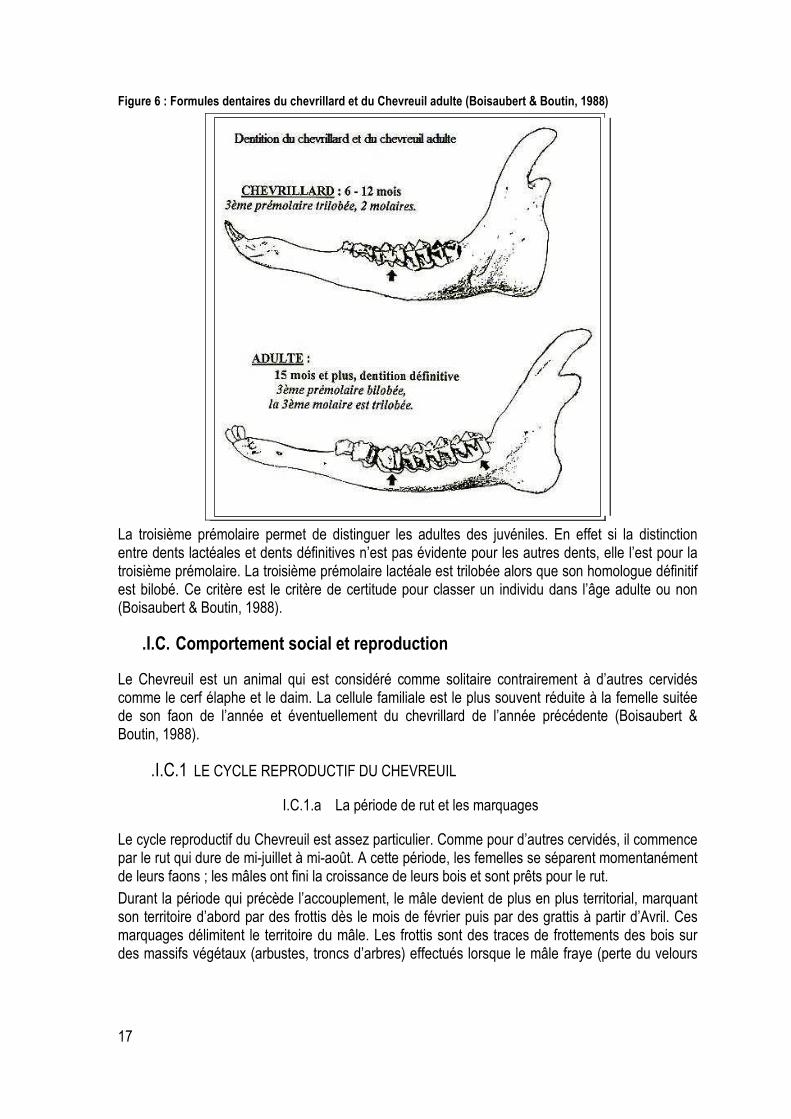
Figure 6 : Formules dentaires du chevrillard et du Chevreuil adulte (Boisaubert & Boutin, 1988) .......... 17
Figure 7 : Les étapes successives de la reproduction de la chevrette (d’après Boisaubart & Boutin, 1988) ...................................................................................................................................................... 18
Figure 8: Phylogénie actuelle du genre Anaplasmatacée (L'échelle représente le nombre de différence pour 1000 paires de bases) (Dumler et al., 2005) .................................................................................. 27
Figure 9: Cycle infectieux d'A. phagocytophilum et rôle de la tique Ixodes ricinus ................................. 29
Figure 10: Le cycle biologique d'Ixodes ricinus ...................................................................................... 36
Figure 11: Répartition des sites équilibre Faune-Flore sur la région Franche-Comté ............................ 48
Figure 12:Distribution temporelle des prélèvements de chevreuils sains et moribonds ......................... 56
Figure 13: Composition en âge des échantillons malade et sain ........................................................... 57
Figure 14: Composition en sexe des échantillons malade et sain .......................................................... 57
Figure 15:Comparaison par cartographie de la distribution géographique des prélèvements de chevreuils malades et sains par rapport à l’emplacement des sites Equilibre Faune-Flore ................... 59
Figure 16: Distribution des résultats de PCR semi-quantitative sur sang pour les classes de chevreuils sain et malade ........................................................................................................................................ 61
Figure 17: Distribution des résultats de PCR semi-quantitative sur sang pour les classes de chevreuils jeune et adulte ....................................................................................................................................... 61
Figure 18: Distribution des résultats en PCR sur sang selon les dates de prélèvement ........................ 62
Figure 19: Répartition géographique des chevreuils positifs en Franche-Comté ................................... 63
Figure 20: distribution de la présence de tiques sur les chevreuils prélevés .......................................... 64

4

5
Liste des tableaux
Tableau 1 : Formule dentaire du faon (Boisaubert & Boutin, 1988) ....................................................... 16
Tableau 2 : Formule dentaire du Chevreuil adulte (Boisaubert & Boutin, 1988)..................................... 16
Tableau 3 : Pousse des molaires chez le Chevreuil (Boisaubert & Boutin, 1988) .................................. 16
Tableau 4 : Remplacement des dents lactéales chez le Chevreuil (Boisaubert & Boutin, 1988) ........... 16
Tableau 5 : Les taux de survie des faons à la limite de capacités d’un milieu dans leur première année et conséquences (Delorme, communication personnelle) ...................................................................... 20
Tableau 6: Les différents indicateurs de changements écologiques utilisés en France ......................... 24
Tableau 7: Les hôtes d'A. phagocytophilum et les liens entre hôtes ...................................................... 33
Tableau 8 : Les modifications biochimique dues à l'ehrlichiose chez la chèvre et le mouton ................. 40
Tableau 9: Nombre de chevreuils à prélever par classe et par département ......................................... 47
Tableau 10: Les prélèvements disponibles selon la classe de chevreuil ............................................... 49
Tableau 11: Nombre de chevreuils prélevés et inclus lors de la saison de prélèvement 2007/2008 ...... 52
Tableau 12: Tableau récapitulatif de l'ensemble des résultats toutess analyses confondus (PCR tous tissus, IFI) pour les classes malade et sain ............................................................................................ 53
Tableau 13 : Tableau de contingence pour la PCR sur sang et les classes de chevreuils malade et sain ............................................................................................................................................................... 53
Tableau 14: Résultats du test de Fisher de comparaison malade/sain des prévalences en PCR sur sang ............................................................................................................................................................... 54
Tableau 15: Tableau de contingence pour les PCR sur sang et rate et les classes de chevreuils malade et sain .................................................................................................................................................... 54
Tableau 16: Résultats du test de Fisher de comparaison malades/sain des prévalences en PCR tous tissus ...................................................................................................................................................... 55
Tableau 17; Tableau de contingence pour l'IFI et pour les classes malade et sain................................ 55
Tableau 18: Résultats du test de Fisher de comparaison malade/sain des séroprévalences obtenues par IFI ..................................................................................................................................................... 56
Tableau 20: Nombre de chevreuils obtenus par classe et par département par rapport aux objectifs ... 60
Tableau 21: Tableau de croisement des données des analyses IFI et PCR (sang ou rate) par individu 60
Tableau 22: Les évolutions des ICE sur les sites Equilibre-Faune-Flore suivis depuis au moins trois ans ............................................................................................................................................................... 67

6

7
Liste des abréviations
FNC : Fédération nationale des chasseurs
FRC : Fédération régionale des chasseurs
FRCFC : Fédération régionale des chasseurs de Franche-Comté
FDC : Fédération départementale des chasseurs
FDC25 : Fédération départementale des chasseurs du Doubs
FDC39 : Fédération départementale des chasseurs du Jura
FDC70 : Fédération départementale des chasseurs de Haute-Saône
FDC90 : Fédération départementale des chasseurs du Territoire de Belfort
ONCFS : Office National de la Chasse et de la Faune Sauvage
EGB : Ehrlichiose Granulocytaire Bovine
EGH : Ehrlichiose Granulocytaire Humaine
ICE : Indicateur de Changement Ecologique
A. phagocytophilum : Anaplasma phagocytophilum

8

9
Introduction
La France compte un très petit nombre de grands herbivores sauvages sur son territoire qui ont tous une image emblématique auprès du grand public. Parmi eux on trouve le Chevreuil (Capreolus capreolus), un petit ongulé d’une grande importance pour le milieu cynégétique.
En 1995, un premier cas de mortalité massive a été rapporté dans le Cher et depuis d’autres cas sont apparus associés à des baisses des observations de chevreuils dans certains massifs (Marti-Brizen, 2007). Les performances de ces populations étaient également moins bonnes que les années précédentes. Les chevreuils retrouvés morts partageaient certaines caractéristiques (animaux amaigris, diarrhéiques) au sein d’un même massif et entre des massifs différents. Ces caractéristiques communes ont amené l’ensemble des acteurs de la chasse à supposer qu’un agent pathogène unique pourrait être à l’origine de cette mortalité.
Plusieurs études ont été menées par les réseaux de surveillance (SAGIR) en 1999 puis en 2000-2001 dans le but de trouver une nouvelle maladie touchant le chevreuil mais aucune n’a donné à l’heure actuelle pleine satisfaction. Les différents acteurs de la chasse et de l’environnement (fédérations de chasses, Office Nationale de la Chasse et de la Faune Sauvage, écoles vétérinaires, vétérinaires praticiens) se sont rassemblés en 2005 à l’initiative de la Fédération Nationale des chasseurs suite à de nouveaux cas de mortalités massives (Marti-Brizen, 2007). L’absence de cause de mort commune chez les chevreuils autopsiés a conduit l’ensemble des experts à penser à l’existence d’une maladie infectieuse immunosuppressive à partir de 2006. Deux pistes ont été envisagées : les pestiviroses et Anaplasma phagocytophilum, l’agent de l’ehrlichiose bovine/ovine. Par ailleurs, on considère aujourd’hui qu’il pourrait ne pas y avoir un seul agent pathogène en cause mais plusieurs et que la dégradation des conditions de vie du Chevreuil pourrait avoir également joué un rôle.
Au cours de cette thèse nous allons chercher à mettre en évidence la corrélation entre l'infection par Anaplasma phagocytophilum chez le chevreuil et la survenue de maladie ou de mortalité, par une étude cas-témoins comparant des animaux morts ou malades à des animaux sains tués à la chasse. L'hypothèse d'une responsabilité de facteurs d'environnement sera explorée grâce à l'analyse de données de suivi des populations de chevreuils de Franche-Comté depuis 1995, sur des sites dénommés « Equilibre Faune-Flore »

10

11
Première Partie : étude bibliographique
.I. BIOLOGIE DU CHEVREUIL
.I.A. La répartition du Chevreuil en France, l’évolution des populations depuis 1973 et les habitats
Le Chevreuil (Capreolus capreolus, Linnaeus, 1758) est capable de coloniser un grand nombre de milieux et vit dans tous les types d’habitats qu’offrent les climats tempérés.
C’est le seul Capréolidé en Europe où il est présent sur un large territoire. Il est en effet présent de l’Ouest de la France à la frontière occidentale de la Russie, du Nord de la Turquie au cercle polaire en Scandinavie (Delorme & Van Laere 2007).
Il est absent uniquement au Sud de l’Espagne, en Italie, en Grèce (climat trop aride) et dans certaines régions de Grande Bretagne et d’Irlande.
Le Chevreuil n’est présent qu’en Europe. Ce sont d’autres espèces du genre Capreolus qui colonisent le reste du monde et notamment l’Asie (Delorme & Van Laere 2007).
.I.A.1 REPARTITION DU CHEVREUIL EN FRANCE : UNE PREDILECTION POUR LES GRANDS MASSIFS FORESTIERS DE L’EST ET DU SUD-OUEST
C’est un gibier qui est chassé sur l’ensemble du territoire français avec dans certaines régions de l’Est des tableaux de chasse atteignant plus de 11000 individus par an. L’intérêt cynégétique de cette espèce n’est plus à démontrer et le ralentissement de l’accroissement des populations depuis trois ans a relancé l’intérêt pour cette espèce (Anonyme, 2008).
Le Chevreuil est un ongulé qui colonise l’ensemble du territoire français à l’exception de la Corse. Il est particulièrement abondant dans les régions de l’Est de la France (Alsace, lorraine, Franche-Comté, Centre, Bourgogne). La figure 1 présente la répartition du Chevreuil en France.
Figure 1: Répartition du Chevreuil en France (Réseau ONCFS/FNC/FDC,"ongulés sauvages")
12
.I.A.2 EVOLUTION DES POPULATIONS DE CHEVREUIL EN FRANCE DEPUIS 1973
I.A.2.a Evolution des plans de chasse depuis 1973
Le chevreuil a bénéficié depuis trente ans de la mise en place d’une chasse raisonnée faisant appel à des plans de chasse prévus pour le maintien et la gestion de l’espèce. De ce fait, les plans de chasse ont augmenté de 420 % en vingt ans (Anonyme, 2008).
Depuis cinq ans toutefois, les plans de chasse augmentent moins et les taux de réalisation diminuent (nombre de chevreuils abattus par rapport au plan de chasse prévu).
Dans les régions qui nous intéressent, les plans de chasse sont plutôt en baisse et les taux de réalisation le sont également. Parmi les départements qui ont abaissé leur plan de chasse, on trouve le Territoire de Belfort et la Haute Saône, deux départements de la région Franche-Comté avec des plans de chasse importants pour le Chevreuil (Anonyme 2008).
La figure 2 montre l’évolution des plans de chasse et des taux de réalisation de ces plans de chasse depuis 1973.
Figure 2 : Evolution annuelle des plans de chasse Chevreuil de 1973 à 2006 (Source: ONCFS, Faune Sauvage n°279 ; février 2008)
I.A.2.b Plans de chasse pour la saison 2006-2007
La figure 3 montre la carte des réalisations de plans de chasse (nombre de chevreuils tués à la chasse par rapport au nombre attribué par les plans de chasse) pour la saison 2006-2007. Elle permet de montrer les régions qui sont les plus denses en chevreuils. On remarque que sur une bande partant du Nord-est (Alsace, Moselle) vers le Sud-ouest (Pyrénées Atlantiques, Garonne), les chevreuils sont plus nombreux. Ce sont des zones où il y a de grands massifs forestiers (Anonyme 2008).
En effet l’habitat de prédilection du Chevreuil est la forêt, ou plus exactement les milieux forestiers ouverts tels que les petits bois, les lisières de forêts et les champs limitrophes.
Le Chevreuil n’est en règle générale pas un ruminant des forêts denses, ni des grands espaces ouverts.

13
Il est couramment repéré dans des zones d’élevage partageant la pâture avec des bovins surtout dans les zones boisées.
Cependant, depuis quelques années le Chevreuil a adopté un nouveau comportement et on le retrouve dans des milieux très ouverts. Il n’est pas anormal de retrouver aujourd’hui des groupes de plusieurs chevreuils dans des milieux de plaines très ouverts tels que la Champagne-Ardennes, la Beauce et d’autres grandes plaines céréalières (Delorme & Van Laere 2007).
Cette évolution est probablement due à l’augmentation des populations de chevreuils, qui, de ce fait se sont orientés vers des milieux plus ouverts lorsque les milieux originels ont été surchargés (Delorme & Van Laere 2007).
Figure 3 : Réalisations de plans de chasse en France en 2006/2007 (Source: ONCFS, Faune Sauvage n°279 ; février 2008)
Le total des attributions était de 585520 chevreuils pour la saison 2006-2007 avec un taux de réalisation de 87 % (Anonyme, 2008).
Le Chevreuil occupe donc la totalité du territoire français et colonise plusieurs milieux variés. Il est nécessaire pour le gestionnaire de savoir distinguer cette espèce des autres ongulés sauvages et d’identifier l’âge et le sexe de l’individu observé ou abattu. Il est donc important de connaître les caractères morphologiques et les critères de distinctions de l’âge et du sexe.

14
.I.B. Morphologie, critères d’âges et de sexe
.I.B.1 LA TETE DU CHEVREUIL
La figure 4 présente les principales caractéristiques de la tête du Chevreuil en vue de profil gauche et en vue de face.
Figure 4: Schémas détaillés de la tête du Chevreuil et traits caractéristiques, vue de profil gauche (g) et vue de face (d) (Boisaubert & Boutin, 1988)
La tête d’un chevreuil est souvent la partie du corps la plus facilement observable car du fait de sa petite taille, le Chevreuil est souvent tête dressée pour surveiller ses alentours, le reste du corps étant masqué par la végétation.
Nous nous intéressons plus particulièrement à la tête du Chevreuil car son observation permet souvent de renseigner sur le sexe de l’animal et surtout de faire la distinction entre les juvéniles et les adultes par l’examen des dents. A distance, la vue de la tête permet aussi au chasseur de vérifier s’il s’agit d’un adulte ou d’un juvénile.
Seul le mâle possède des bois qui sont implantés à la verticale sur le sommet du crâne et mesurent jusqu’à 25 cm chez l’adulte. Les oreilles sont de grande taille et implantées sur le même plan que les bois, à leur extérieur.
.I.B.2 ETUDE DES BOIS
Les bois, ou broches, d’un chevreuil constituent le trophée. La structure des bois est spécifique à l’individu et la complexité de cette structure ainsi que la taille font la valeur du trophée.
La base des bois, constituée d’un bourrelet osseux est appelée la meule. Elle est surmontée d’un corps unique qui peut se diviser en plusieurs parties plus haut formant ainsi des andouillers voire plusieurs. Les irrégularités sur le corps du bois sont appelées des perles (Boisaubert et Boutin, 1988).
Le nombre de corps portés par le Chevreuil est déterminé par le nombre de terminaisons de chaque bois.
Le croissance de bois suit un cycle qui commence dès la deuxième année. Le mâle ne porte pas de bois entre octobre et décembre. Il peut alors être confondu avec la femelle surtout à distance.
La figure 5 illustre le cycle annuel de la pousse des bois chez le Chevreuil.

15
Figure 5: Le cycle annuel des bois du Chevreuil (d’après Boisaubert & Boutin, 1988)
Contrairement à ce qui est généralement supposé, la forme des bois n’est pas un bon indicateur de l’âge. En règle générale, les bois sont plus élaborés avec l’âge mais il y a beaucoup d’exceptions entre les chevreuils précoces, les tardifs ou les malformés (Boisaubert & Boutin, 1988).
La première année, jusqu’en novembre, les chevrillards n’ont que des petites bosses à la place des bois et par la suite, ils n’ont que des petits bois appelés broches et qui tombent à la fin de l’hiver. Ces broches sont observés uniquement chez le chevrillard alors que les chevreuils adultes ont perdus leurs bois à la même époque. Des broches de un à quatre cm sont observées unique chez le chevrillard.
.I.B.3 DETERMINATION DE L'AGE DU CHEVREUIL
Plusieurs caractéristiques distinguent les adultes des jeunes.
L’aspect général tout d’abord peut nous renseigner : l’adulte est plus grand mais il est également plus massif. Les jeunes chevreuils ont une encolure fine et une masse musculaire moins développée qui leur donne un aspect gracile alors que les adultes sont plus lourds, plus massifs ; l’encolure et le thorax sont plus larges et donne un aspect un peu plus majestueux. Plusieurs auteurs ajoutent que le comportement s’ajoute à ses observations ; les jeunes sont souvent plus alertes, plus joueurs alors que les adultes sont plus calmes.
Les bois peuvent donner une indication mais cette indication n’est jamais une certitude sauf pour les chevrillards. Le seul élément pouvant renseigner précisément sur l’âge est la dentition de l’individu. On ne peut donner l’âge d’un individu à distance, il faut examiner sa mâchoire. C’est le seul critère qui soit utilisé pour la distinction des âges des individus abattus pour les plans de chasse.
La structure dentaire du Chevreuil nous renseigne également sur sa façon de s’alimenter.
Le Chevreuil possède une dentition de ruminant : il ne possède pas d’incisives supérieures mais un bourrelet fibreux qui lui permet de prendre appui pour couper les tiges ou feuilles dont il se nourrit.
La dentition du Chevreuil évolue avec son âge de façon précise et permet de donner l’âge du chevreuil examiné (Boisaubert & Boutin, 1988).
16
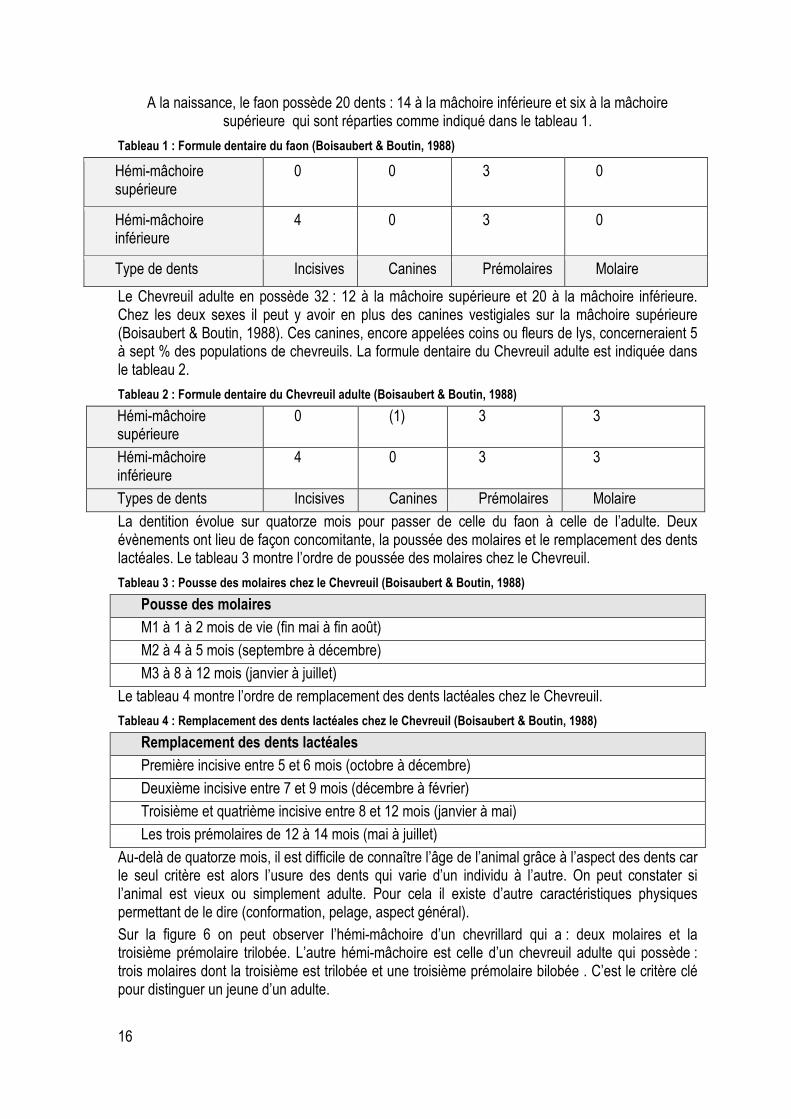
A la naissance, le faon possède 20 dents : 14 à la mâchoire inférieure et six à la mâchoire supérieure qui sont réparties comme indiqué dans le tableau 1.
Tableau 1 : Formule dentaire du faon (Boisaubert & Boutin, 1988)
Hémi-mâchoire supérieure
0 0 3 0
Hémi-mâchoire inférieure
4 0 3 0
Type de dents Incisives Canines Prémolaires Molaire
Le Chevreuil adulte en possède 32 : 12 à la mâchoire supérieure et 20 à la mâchoire inférieure. Chez les deux sexes il peut y avoir en plus des canines vestigiales sur la mâchoire supérieure (Boisaubert & Boutin, 1988). Ces canines, encore appelées coins ou fleurs de lys, concerneraient 5 à sept % des populations de chevreuils. La formule dentaire du Chevreuil adulte est indiquée dans le tableau 2.
Tableau 2 : Formule dentaire du Chevreuil adulte (Boisaubert & Boutin, 1988)
Hémi-mâchoire supérieure
0 (1) 3 3
Hémi-mâchoire inférieure
4 0 3 3
Types de dents Incisives Canines Prémolaires Molaire
La dentition évolue sur quatorze mois pour passer de celle du faon à celle de l’adulte. Deux évènements ont lieu de façon concomitante, la poussée des molaires et le remplacement des dents lactéales. Le tableau 3 montre l’ordre de poussée des molaires chez le Chevreuil.
Tableau 3 : Pousse des molaires chez le Chevreuil (Boisaubert & Boutin, 1988)
Pousse des molaires
M1 à 1 à 2 mois de vie (fin mai à fin août)
M2 à 4 à 5 mois (septembre à décembre)
M3 à 8 à 12 mois (janvier à juillet)
Le tableau 4 montre l’ordre de remplacement des dents lactéales chez le Chevreuil.
Tableau 4 : Remplacement des dents lactéales chez le Chevreuil (Boisaubert & Boutin, 1988)
Remplacement des dents lactéales
Première incisive entre 5 et 6 mois (octobre à décembre)
Deuxième incisive entre 7 et 9 mois (décembre à février)
Troisième et quatrième incisive entre 8 et 12 mois (janvier à mai)
Les trois prémolaires de 12 à 14 mois (mai à juillet)
Au-delà de quatorze mois, il est difficile de connaître l’âge de l’animal grâce à l’aspect des dents car le seul critère est alors l’usure des dents qui varie d’un individu à l’autre. On peut constater si l’animal est vieux ou simplement adulte. Pour cela il existe d’autre caractéristiques physiques permettant de le dire (conformation, pelage, aspect général).
Sur la figure 6 on peut observer l’hémi-mâchoire d’un chevrillard qui a : deux molaires et la troisième prémolaire trilobée. L’autre hémi-mâchoire est celle d’un chevreuil adulte qui possède : trois molaires dont la troisième est trilobée et une troisième prémolaire bilobée . C’est le critère clé pour distinguer un jeune d’un adulte.
17
Figure 6 : Formules dentaires du chevrillard et du Chevreuil adulte (Boisaubert & Boutin, 1988)
La troisième prémolaire permet de distinguer les adultes des juvéniles. En effet si la distinction entre dents lactéales et dents définitives n’est pas évidente pour les autres dents, elle l’est pour la troisième prémolaire. La troisième prémolaire lactéale est trilobée alors que son homologue définitif est bilobé. Ce critère est le critère de certitude pour classer un individu dans l’âge adulte ou non (Boisaubert & Boutin, 1988).
.I.C. Comportement social et reproduction
Le Chevreuil est un animal qui est considéré comme solitaire contrairement à d’autres cervidés comme le cerf élaphe et le daim. La cellule familiale est le plus souvent réduite à la femelle suitée de son faon de l’année et éventuellement du chevrillard de l’année précédente (Boisaubert & Boutin, 1988).
.I.C.1 LE CYCLE REPRODUCTIF DU CHEVREUIL
I.C.1.a La période de rut et les marquages
Le cycle reproductif du Chevreuil est assez particulier. Comme pour d’autres cervidés, il commence par le rut qui dure de mi-juillet à mi-août. A cette période, les femelles se séparent momentanément de leurs faons ; les mâles ont fini la croissance de leurs bois et sont prêts pour le rut.
Durant la période qui précède l’accouplement, le mâle devient de plus en plus territorial, marquant son territoire d’abord par des frottis dès le mois de février puis par des grattis à partir d’Avril. Ces marquages délimitent le territoire du mâle. Les frottis sont des traces de frottements des bois sur des massifs végétaux (arbustes, troncs d’arbres) effectués lorsque le mâle fraye (perte du velours

18
autour des bois) et après la fraye. La période de fraye a lieu au mois de février et le mâle continue ses frottis jusqu’à la période du rut1.
Les grattis, qui apparaissent en mars sont des marques laissées par le brocard qui frotte le sol avec ses antérieurs, laissant ainsi une marque visible et odoriférante. Souvent grattis et frottis sont associés, ils sont alors appelés régalis (Boisaubert & Boutin, 1988).
L’ensemble de ces marques -frottis, grattis et régalis- est utilisé comme indicateur de présence d’un brocard territorial et la distribution de ces marques peut être utile pour le suivi des populations.
Durant la période de rut, le Chevreuil recherche la femelle et chasse les autres mâles. Une parade précède l’accouplement qui est difficile à observer2.
I.C.1.b Du rut à la mise bas : onze longs mois
Après l’accouplement et la fécondation, la particularité de la chevrette est que l’œuf fécondé ne s’implante pas rapidement. Au contraire il entre en dormance pendant 170 jours, il s’agit alors d’une blastula libre. La blastula ne s’implante qu’à la fin décembre. La gestation dure 130 jours, la mise bas a lieu vers mai juin. Ainsi de la fécondation à la mise-bas il s’écoule 11 mois alors que la gestation effective ne dure que 4 mois et demi (Boisaubert & Boutin, 1988).
La figure 7 effectue la chronologie des étapes de la reproduction chez la chevrette.
Figure 7 : Les étapes successives de la reproduction de la chevrette (d’après Boisaubart & Boutin, 1988)
La mise bas à lieu vers mai-juin. La chevrette met bas à des faons qui pèsent environ 1,5 kg dès la naissance. La période de rut a lieu seulement un mois et demi plus tard, la chevrette va donc abandonner momentanément les faons.
.I.C.2 RAPPORTS CHEVRETTE/FAONS : LA PHASE CRITIQUE JUVENILE
C’est la caractéristique majeure de la vie sociale du Chevreuil : la femelle garde ses faons auprès d’elle de leur naissance jusqu’au mois de mai de l’année suivante lorsqu’ils ont atteint environ douze mois.
Dans les semaines qui précèdent la mise bas, la chevrette devient très territoriale et sur un domaine d’environ un hectare, elle empêche l’entrée d’autres chevreuils et de prédateurs (renard).
1 Il a été observé que l’intensité des frottis augmente avec le taux de testostérone circulant dans le sang. 2 La chevrette n’est en œstrus que deux jours ce qui rend la probabilité d’observer un accouplement assez faible. On observe d’ailleurs des mâles monogames qui accompagnent une chevrette toute l’année ce qui permettrait au mâle d’assurer sa descendance en ne ratant pas les chaleurs.

19
Ce comportement territorial va continuer durant les mois qui suivent la mise bas et s’atténuer vers la fin de l’été.
Les faons naissent autour du mois de mai et sont pris en charge par la mère uniquement. Durant les premières semaines de vie, les faons sont laissés dans un endroit que la mère repère avant de partir se nourrir à une distance qui peut aller jusqu’à 150 mètre des faons. Durant cette période, les faons s’immobilisent en cas de danger et choisissent leurs reposées (litière où ils se reposent) en fonction des conditions climatiques.
Ce comportement rend les faons très vulnérables quand ils sont découverts et le danger majeur est la fenaison agricole qui a lieu durant le mois de juin. Le passage d’engins agricoles dans les champs où des faons se cachent provoque de nombreuses morts. La survie au premier mois des faons dépend de la période de moisson qui dépend des conditions climatiques. Ainsi selon l’année, la moisson pourra avoir lieu pendant les premières phases de vie des faons ou plus tard lorsqu’ils sont plus mobiles (Marillier, communication personnelle).
Par la suite, les faons vont commencer progressivement à accompagner la mère dans ses déplacements ; au début un seul faon sur les deux puis les deux vers 80 jours.
La chevrette mobilise toute son énergie lors de la gestation pour mettre bas deux faons bien développés de 1 à 1,5 kg le plus souvent1. Après la mise-bas, la chevrette n’a plus de réserves énergétiques Par la suite, la croissance des faons est extrêmement rapide : de 150 à180 gr par jour.
Les premières étapes de vie des faons sont primordiales car comme la chevrette ne dispose pas de réserves graisseuses pour l’allaitement des jeunes, cet allaitement est directement inféodé aux ressources existant sur le moment. Si les ressources sont trop faibles, les faons et la chevrette vont dépérir jusqu’au point où la chevrette abandonne ses petits pour survivre. Les faons abandonnés ne survivent pas et sont rapidement tués par les prédateurs du Chevreuil (Renard roux, Martre, Chat sylvestre, Lynx).
La force et la faiblesse du mode de reproduction de la chevrette est qu’elle donne toute son énergie à la procréation avec un taux de natalité et un poids à la naissance importants mais que ce don peut coûter cher si les ressources nécessaires pour un bon allaitement ne sont pas suffisantes. Si une chevrette abandonne ces faons pour survivre, il n’y a pas de répercussions sur la reproduction de cette chevrette l’année suivante (Delorme et al., 2007).
Ainsi les mauvaises années (en terme de ressources) seront caractérisées par la présence de moins de chevrillards durant l’hiver et par un mauvais état des chevrillards survivants qui n’assureront pas une bonne reproduction les années suivantes. Ceci s’explique par le fait qu’il faut un poids et un âge suffisants pour entrer en reproduction et que les chevrettes nées une mauvaise année ont des retards de croissance et entrent en reproduction plus tardivement et mettent bas moins de faons car le nombre de faons est corrélé au poids à la fécondation (Delorme et al., 2007).
Ce décalage se ressent sur plusieurs années pour une cohorte née dans de mauvaises années. Ces mauvaises années sont rares dans des situations d’équilibre mais courantes dans des conditions de déséquilibre dans des massifs fortement peuplés. Une cohorte née une mauvaise année va être caractérisée par des chevrettes de plus petite taille, de longévité réduite et entrant en reproduction parfois avec un an de retard, à trois ans. Les tailles de portées sont également réduites puisque des chevrettes en bonne santé ont plus souvent des gestations gémellaires (Delorme et al., 2007).
Dans ces massifs le taux de survie à un an peut descendre à 20 % dans des conditions défavorables. La tableau 5 donne une indication des taux de survie des faons selon les conditions.
1 La gestation est souvent gémellaire chez la chevrette surtout dans des milieux riches.

20
Tableau 5 : Les taux de survie des faons à la limite de capacités d’un milieu dans leur première année et conséquences (Delorme, communication personnelle)
Conditions favorables
(rare, 1 ou 2 années par décennies)
Conditions défavorables
(les plus courantes)
Survie estivale
Survie hivernale
Total Survie estivale
Survie hivernale
Total
70 % 80 % 56 % 30 % 70 % 21 %
Répercussions sur la cohorte en années défavorables
Peu de femelles suitées
Peu de chevrillards
Retards à l’entrée en reproduction
Mauvaises performances reproductrices de cette cohorte sur toute sa vie
Répercussions sur les cohortes suivantes
Le poids des chevrillards est un bon indicateur de santé des populations, il est utilisé comme tel pour l’établissement de plans de chasse dans certaines régions et pour le suivi des populations notamment dans les réserves de Chizé (79) et Trois fontaines (51). Nous verrons comment il peut être utilisé ultérieurement.
Cette cellule familiale va persister jusqu’au mois de février. Par la suite, les chevrillards vont progressivement s’éloigner de la mère. Le groupe se séparera définitivement vers mai.
Les jeunes acquièrent un domaine vital qui leur est propre. Il semblerait que les domaines vitaux des jeunes chevauchent celui de la mère et que les jeunes chevrettes rejoignent leur mère, formant ainsi des petits groupes familiaux.
Les jeunes mâles quant à eux s’éloignent pour former leur domaine propre, séparé de celui de la mère.
Les domaines vitaux qui se chevauchent au départ vont progressivement se séparer avec l’âge du chevrillard. A terme, une chevrette a un territoire propre séparé de celui des autres femelles et chevauché à certaines périodes de l’année par celui d’un ou de plusieurs mâles.
.I.D. Alimentation du Chevreuil, impacts sur les cultures et les forêts françaises
.I.D.1 REGIME ALIMENTAIRE DU CHEVREUIL
Le régime alimentaire d’un Chevreuil est très varié. En somme tout ce qui se trouve à sa hauteur est susceptible d’être consommé.
Le Chevreuil sélectionne les végétaux qu’il consomme en fonction de ses besoins et de leur valeur énergétique. Sur des ronces il va par exemple sélectionner les bourgeons, jeunes pousses, mûres qui sont les éléments les plus nutritifs.
De plus le Chevreuil semble être capable de consommer des végétaux toxiques pour d’autres animaux. Il aurait même besoin de consommer des végétaux riches en tanins alors que ces mêmes tanins sont toxiques aux mêmes doses pour d’autres animaux (BoisAubert, 1989).
Le comportement alimentaire très éclectique et sélectif du Chevreuil lui permet de ne pas être en compétition avec d’autre espèces telle que le Cerf élaphe ou les bovins domestiques qui ont un comportement alimentaire non sélectif de brouteur.

21
Les végétaux de prédilection du Chevreuil sont les arbustes et arbres de petite taille. Citons comme exemples le Hêtre, le Roncier ainsi que le Lierre et l’Aubépine. Mais le Chevreuil consomme également des pousses d’arbres, des graminées (pousses, tiges, graines) des fleurs ou des champignons.
Nous le verrons ultérieurement, le Chevreuil consomme les végétaux d’une façon particulière : il saisi la tige entre ses incisives et prémolaires inférieures et le bourrelet cartilagineux qui compose sa mâchoire supérieure et tire sur la tige. Il en résulte une tige coupée nette et arrachée sur sa longueur par le bourrelet cartilagineux. Cette cassure caractéristique est bien visible lorsqu’elle concerne tout un massif végétal et est utilisée comme indicateur de présence.
On parle d’abroutissement et ces marques permettent de mettre en évidence la présence du Chevreuil.
Paradoxalement, le Chevreuil est une espèce qui s’implante très bien dans des milieux qui ne sont pas tout à fait ses milieux de prédilection. En effet même s’il s’agit d’une espèce sélective, son régime alimentaire très varié lui permet d’utiliser des ressources autres que celles consommées habituellement.
La sélectivité alimentaire du Chevreuil fait de lui un herbivore pouvant survivre dans des conditions défavorables où la disponibilité en végétaux est très faible. Ainsi dans des milieux pauvres, où les aliments sont plus rares le Chevreuil survit mieux que d’autres espèces comme le Cerf élaphe. Ceci explique la prédominance du Chevreuil dans les massifs forestiers.
Le Chevreuil est une espèce dont le mode d’alimentation ne favorise pas en temps ordinaires la colonisation de milieux ouverts (ex : la plaine française) car il n’est pas capable d’optimiser son alimentation sur ces terrains. C’est un herbivore paisseur qui sélectionne les éléments consommés et ne consomme que des petites quantités à chaque prise alimentaire.
Cependant, il colonise ces milieux car il n’y est pas en concurrence avec des herbivores brouteurs comme le Cerf Elaphe.
.I.D.2 DEGATS AUX CULTURES ET AUX FORETS
Le Chevreuil s’alimente sur de nombreux supports et de ce fait il a un impact non négligeable sur la forêt et sur les cultures.
Le Chevreuil cueille souvent les parties les plus énergétiques du végétal auquel il s’attaque. Souvent les parties les plus attaquées sont les jeunes pousses, les bourgeons et les baies ce qui gêne la régénération du sous-sol forestier et pour le maintien des pépinières.
D’autre part le Chevreuil a un odorat très développé qui lui permet de sélectionner les éléments les plus riches des plantes mais aussi de sélectionner les plants les plus riches. Les arbres et arbustes de pépinières utilisés pour l’entretien des forêts par les forestiers sont souvent plus riches en énergie et préférés aux arbres autochtones par le Chevreuil. Ainsi des zones entières de plantation peuvent être ravagées sévèrement par des chevreuils surtout dans des zones où les ressources alimentaires sont insuffisantes.
Enfin le Chevreuil ne coupe pas net une tige, il en arrache une partie ce qui nuit à la régénération de la plante. Souvent on observe des plants dont la totalité des tiges sont ainsi coupées et ne peuvent se régénérer correctement.
Les dégâts aux cultures peuvent être importants puisqu’une culture est un véritable silo à grain pour le Chevreuil surtout en cas de surpopulation dans les sous-bois.
L’impact sur la végétation n’est pas uniquement négatif car le Chevreuil permet la régénération du sous-sol en consommant des végétaux qui ne sont pas consommés par d’autres espèces et en limitant leur expansion.

22
Le Chevreuil est donc un animal qui provoque des dégâts modérés dans des conditions d’équilibre mais qui peut avoir un impact beaucoup plus important dès qu’il y a un déséquilibre avec le milieu ; soit parce que les densités de chevreuils sont très élevées, soit parce que les ressources du milieu sont trop faibles.
L’impact sur la végétation est un signe de présence du Chevreuil dans un massif forestier. Il est même utilisé comme indicateur biologique pour effectuer le suivi des populations. En effet, afin d’adapter les plans de chasse pour permettre un maintien raisonné du Chevreuil, les fédérations de chasse ont mis en place plusieurs indicateurs de santé des populations. L’utilisation de ces indicateurs permet également de dépister des problèmes démographiques liés au milieu, aux maladies… Nous allons maintenant décrire ces indicateurs en insistant sur ceux d’entre eux qui sont utilisés en France Comté.
.I.E. Principe du suivi des populations de chevreuils
Le principe du suivi des populations d’ongulés sauvages a beaucoup évolué depuis vingt ans. En effet le comptage des animaux était la règle il y a quinze ans et c’est sur la base de cette méthode que les plans de chasses étaient établis. Cette méthode a montré ses limites à travers des études comparatives avec les méthodes de capture-marquage-recapture dans les réserves françaises de Chizé et Trois-fontaines. Cette technique s’est avérée peu précise et peu juste. Les erreurs liées à cette technique se sont ressenties sur le terrain avec des plans de chasse mal adaptés à une situation qui n’était pas retranscrite par les comptages (Groupe Chevreuil, 1996).
Aujourd’hui les plans de chasses sont établis avec des indicateurs qui ne donnent pas directement la densité de population mais qui permettent un meilleur suivi dans la durée et surtout donnent des indications sur la santé des populations suivies.
.I.E.1 LES INDICATEURS DE PRESENCE
L’ensemble des activités du Chevreuil laisse des traces qui sont facilement repérables et identifiables par l’observateur. Grâce à l’utilisation de ces indicateurs on peut repérer les domaines vitaux de différents individus en distinguant le sexe (Albaret et al., 1991).
I.E.1.a Empreintes
Comme pour toutes les espèces, le premier indicateur de présence connu chez le Chevreuil est son empreinte. L’empreinte de ce mammifère est relativement facile à identifier car dans la plupart des régions de France, c’est le seul artiodactyle qui ait des empreintes de cette taille.
Les observateurs expérimentés peuvent distinguer l’âge et le sexe d’un individu par ses empreintes. Les empreintes sont uniquement un indicateur de présence et permette difficilement de tirer une notion d’abondance (BoisAubert, 1989).
I.E.1.b Les reposées
Le Chevreuil adulte passe environ sept heures par jour couché dans la végétation et le faon passe 90 % de son temps couché. Lors du couchage, le Chevreuil ménage une couche ou reposée en grattant le fond et en repoussant les débris végétaux. Ces couches sont bien visibles et dans des milieux végétaux variés. Leur abondance donne une idée de l’abondance de chevreuils dans un secteur donné (Delorme, 2007a).

23
I.E.1.c Grattis, frottis et régalis
Les grattis et frottis ont l’intérêt de marquer les bordures de territoire d’un brocard qui fait ces marques tout autour de son territoire pour en marquer le contour. Ces marques ne sont visibles que la moitié de l’année, de février à la fin de l’été mais peuvent être intéressantes pour repérer le territoire d’un mâle pendant la période du rut notamment.
.I.E.2 LES INDICATEURS DE CHANGEMENT ECOLOGIQUE
I.E.2.a Conditions d’utilisation des indicateurs de changement écologiques (ICE) et qualités d’un ICE
Les indicateurs que nous allons présenter doivent être utilisés par le gestionnaire sur plusieurs années et dans des conditions très standardisées. Le recueil d’informations doit se faire si possible :
• Au moins trois ans (Delorme, communication personnelle)
• avec le même observateur
• sur les mêmes périodes et les mêmes tracés
Les plans de chasse basés sur l’utilisation de ces indicateurs sont efficaces uniquement si la population est suivie depuis plusieurs années et si l’impact des plans de chasses est pris en compte d’une année sur l’autre. Contrairement aux plans de chasses anciens qui faisaient appel aux données de l’année, les plans de chasses actuels sont établis sur la base du suivi pluriannuel de la population et sur la base de l’impact des plans de chasses précédents (Anonyme, 1999).
Un plan de chasse est établi d’une part sur l’évolution en nombre des chevreuils mais aussi sur la santé des populations et en vue d’objectifs précis qui sont le plus souvent :
• une population d’ongulés en équilibre avec son milieu avec un impact réduit sur la végétation forestière et les cultures,
• un nombre précis d’ongulés au plan de chasse,
• des ongulés de grande taille pour la chasse au trophée.
On distingue des indicateurs de présence, qui ne marquent que la présence d’individus dans un massif ou dans une zone précise, et des indicateurs de changement écologique qui permettent un suivi d’une population (cf. infra).
Les indicateurs de changements écologiques donnent des indications sur la santé des population. Ils sont utiles au gestionnaire dans les mêmes conditions d’utilisation et d’interprétation que les indices kilométriques, c'est-à-dire par des utilisations standardisées sur plusieurs années. Les ICE doivent présenter un certain nombre de qualités.
Tout d’abord ils doivent être simples à mesurer et être répétables d’une session sur une autre. Ce sont des indicateurs dont l’évolution est significative d’une évolution du système « individu-population-environnement ».
Les ICE sont suffisamment sensibles pour retransmettre des variations du système et leur variation doit être attribuable à un type d’évolution précis et unique dans les interactions « individu-population-environnement », donnant une réponse précise au gestionnaire.
Mais une variation d’un indicateur peut avoir plusieurs causes dans la nature. Il est donc plus intéressant d’utiliser plusieurs ICE pour pouvoir les utiliser simultanément dans le but d’avoir une idée précise de l’évolution des populations suivies.
Il y a plusieurs ICE différents utilisés en France qui sont donnés dans le tableau 6.

24
Tableau 6: Les différents indicateurs de changements écologiques utilisés en France
Les différents Indicateurs Ecologiques de changement
• Nombre moyen de faons par femelle
• Suivi pondéral des animaux et les mesures morphologiques
• Tendance grégaire hivernale
• Taux de parasitisme, les réseaux lipidiques
• La longueur de la mandibule
• Le taux d’utilisation des végétaux
I.E.2.b L’indice kilométrique
L’indice kilométrique est un des outils les plus utilisés pour le suivi des populations de chevreuils. Il est basé sur le principe d’une probabilité de rencontre d’un individu sur un tracé précis, à une période précise par le même observateur et n’a d’intérêt que s’il est reconduit sur plusieurs années. Un indice kilométrique réalisé sur une seule année n’a aucune valeur et n’est pas suffisamment précis, ni juste pour permettre d’estimer une densité ce qui n’est pas son rôle. Le rôle de l’indice kilométrique est de mettre en évidence des variations de densités et non de donner un chiffre de densité (Anonyme, 1991).
Le protocole pour les indices kilométriques (IK) est de tracer plusieurs circuits parcourant la zone à explorer qui soient le plus nombreux possibles sans se chevaucher pour éviter les risques de double comptage. Ces circuits mesurent cinq à sept km et sont parcourus chacun par un observateur. Le réussite de ce système repose sur l’utilisation des mêmes circuits chaque année par les mêmes observateurs et sur les mêmes périodes.
Les résultats sont présentés de la façon suivante :
• I.K.Ci= Nombre de contacts sur le circuit i/nombre de kilomètres du circuit i,
• I.K.Sj=somme de I.K.Ci de la série J/ nombre de circuits (=n),
• I.K.a= somme des I.K.Sj/ nombre de séries.
Les résultats sont interprétables au bout de quelques années. On considère qu’il faut trois ans de données pour pouvoir avoir une estimation suffisamment précise de la situation.
Plus il y a de séries, plus l’intervalle de confiance est réduit donc plus les résultats sont précis.
L’indice kilométrique donne une indication sur la population de chevreuils en nombre mais ne donne pas d’indications sur la santé de la population, ni sur sa composition. Ces indications sont données par les indicateurs de changement écologique.
Sont associées à cette technique des méthodes d’évaluation de la santé des populations :
• la mesure du poids des chevrillards,
• la mesure de la longueur du postérieur,
• la longueur de la mâchoire.
Ces trois méthodes donnent des indicateurs de changements biologiques.
I.E.2.c Le nombre de faons par femelle
Cette technique consiste à surveiller le nombre de chevrettes suitées durant l’été. Cet indicateur n’est pas jugé fiable car les chevrettes ont tendance soit à abandonner tous leurs faons soit à les

25
garder tous ce qui conduit à avoir des résultats trop manichéens que l’on ne peut affiner et qui ne donnent des renseignements intéressants que dans des situations extrêmes. Cet indicateur est trop peu sensible pour être une méthode de référence. (Delorme, communication personnelle)
I.E.2.d Le suivi pondéral des animaux
Nous l’avons vu, la phase critique se situe dans les premières semaines de vie du faon. Dans ces conditions, il est très intéressant de suivre d’une année sur l’autre le poids des faons ou chevrillards (Groupe Chevreuil, 1996).
Le poids des jeunes est un indicateur facile à obtenir pour autant que le chasseur pense à peser les chevrillards qu’il abat et qu’il le fasse de façon rigoureuse (une estimation au kilogramme près n’est pas idéale pour un bon suivi). Le suivi de l’évolution de ces poids d’une année sur l’autre est facilement réalisable et peut renseigner directement sur la santé des populations.
Le chevrillard doit être pesé non vidé et la pesée doit être précise. Le poids du chevrillard est ensuite corrigé pour standardiser l’ensemble des poids recueillis. En effet un chevrillard tué en septembre n’a pas le même poids que s’il avait été tué en décembre. Il faut donc pondérer les poids pour les harmoniser à ce qu’ils auraient été à une date précise, commune pour tous les poids.
Si le poids moyen baisse au cours des années il peut y avoir un problème de densité-dépendance ou un problème de ressources. Si au contraire le poids moyen augmente, les ressources sont plus importantes ou les populations sont plus réduites. Cet indicateur n’a donc pas de valeur utilisé seul.
I.E.2.e La longueur de la mandibule et la longueur de la patte arrière
Le principe de cette méthode est le même que la mesure du poids des chevrillards : on mesure la longueur de la mandibule avec précision et on ramène cette longueur à une date de référence.
C’est encore le même principe pour la longueur de la patte arrière (Delorme, 2007b).
I.E.2.f Le taux d’utilisation de la végétation : les indices de pression sur la flore
Le mode d’alimentation du Chevreuil laisse beaucoup de traces bien identifiables sur les massifs végétaux. En effet le Chevreuil saisi une tige entre ses incisives inférieures et son bourrelet fibreux supérieur ce qui fait que la tige est partiellement sectionnée du côté des dents et arrachée du côté du bourrelet. Les traces laissées sur la végétation sont bien identifiables et souvent bien visibles car plusieurs traces sont visibles sur un même massif. L’ensemble de ces traces est appelé abroutissement. Ils revêtent une importance particulière car ce sont des indicateurs de présence et de densité. Sur les cultures, ces traces se voient bien et permettent d’attribuer ces dégâts au Chevreuil. Ces abroutissements sont très mutilants pour le végétal de plus les Chevreuils consomment en premier lieu les éléments énergétiques d’un végétal qui sont par définition des éléments vitaux (bourgeon, jeunes plants).
L’impact sur la végétation est utilisé comme indicateur d’abondance. Il est également lors de l’établissement des plans de chasse qui tiennent compte non seulement de la densité d’animaux mais également de l’impact du Chevreuil sur la forêt et les cultures en plus de la densité.
D’autres méthodes ont été utilisées pour étudier la dynamique d’une population de chevreuils. Citons par exemple l’examen des corps jaunes chez les chevrette abattues pour évaluer la fécondation (Jolibois, 2007). Par ailleurs d’autres méthodes sont toujours à l’études et pourraient être utilisées à terme : vitesse de développement du faon, ratio poids Chevreuil/ poids du faon (Delorme et al., 2007).

26
.II. Etude bibliographique de A. phagocytophilum
.II.A. Systématique et biologie
.II.A.1 HISTORIQUE DE LA NOMENCLATURE D’A. PHAGOCYTOPHILUM
L’ehrlichiose est une maladie bactérienne due à la bactérie appelée aujourd’hui A. phagocytophilum. Cette maladie a été diagnostiquée pour la première fois par Gordon et al. en 1932 sur des moutons en Ecosse sans que l’agent causal ne soit identifié. Depuis il a été identifié et a changé plusieurs fois de nomenclature jusqu’en 2001 (Chimier, 2003)
L’ensemble de ces changements suit le cheminement des découvertes faites sur cet agent depuis 1932 et à l’heure actuelle sa nomenclature est encore soumise à discussion. Voici comment la classification de cette bactérie a évolué.
En 1949 Foggie la range dans le groupe des rickettsiales car la transmission de la maladie se fait par les tiques. Il la nomme Rickettsia phagocytophila ovis.
Cette bactérie sera par la suite replacée dans le genre Ehrlichia par Philip en 1962. L’ehrlichiose, bien que connue depuis 1932 n’a été diagnostiquée sur aucune autre espèce que le mouton jusqu’en 1969.
En Californie en 1969 que Gribble découvre une bactérie de genre Ehrlichia provoquant une maladie mortelle chez des chevaux. Il nomme la maladie « Ehrlichiose granulocytaire équine » du fait de la localisation de morulas dans les granulocytes. La bactérie alors décrite est nommée Ehrlichia equi par Lewis et al. en 1975.
Il faut attendre 1994 pour qu’une ehrlichiose clinique soit diagnostiquée chez un patient humain par Chen et al . La maladie humaine est appelée l’ehrlichiose granulocytaire humaine (EGH).
Ainsi sont décrites trois bactéries différentes : Ehrlichia phagocytophila, Ehrlichia equi et l’agent de l’Ehrlichiose granulocytaire humaine (EGH). Grâce à l’apparition des outils de biologie moléculaire, le séquençage de certains composants de ces trois bactéries, nommément le gène codant pour l’ARN ribosomal 16S ainsi que les gènes groESL, gltA et ankA a permis de mettre en évidence un grand degré d’homologie entre les trois bactéries (Weisburg et al., 1991, Sumner et al., 1997). De plus les trois bactéries possèdent des caractéristiques biologiques et épidémiologiques très proches.1
Un changement de nomenclature a été proposé pour placer les trois bactéries sous la même dénomination : Anaplasma phagocytophila puis Anaplasma phagocytophilum (Dumler et al., 2001).
Sur la base de divergences sur le gène gltA et biologiques2 entre les trois ehrlichia et le genre Anaplasma, un nouveau groupe a été proposé, regroupant les trois Ehrlichia hors du genre Anaplasma (Inokuma et al., 2001). Ces travaux ont été publiés la même année mais après ceux de Dumler qui n’a pu les prendre en compte. En résumé, la bactérie qui nous intéresse se nomme Anaplasma phagocytophilum et regrouperait les espèces responsables de trois maladies historiquement décrites, respectivement chez les ovins, les chevaux et l’Homme. Euzéby nomme trois variantes dans son dictionnaire de bactériologie vétérinaire : A. phagocytophilum biovar phagocytophila, A. phagocytophilum biovar equi et A. phagocytophilum biovar EGH car il considère que les trois maladies sont dues à trois biovars distincts.
Aujourd’hui, le classification d’A. phagocytophilum se situe comme le montre la figure 8.
1 Les trois bactéries sont transmises par des tiques, provoquent des maladies proches et colonisent les granulocytes 2 Les bactéries du genre Anaplasma colonisent les cellules rouges alors que Ehrlichia colonise les granulocytes

27
Figure 8: Phylogénie actuelle du genre Anaplasmatacée (L'échelle représente le nombre de différence pour 1000 paires de bases) (Dumler et al., 2005)
.II.A.2 LA PROBLEMATIQUE DES VARIANTS D’A. PHAGOCYTOPHILUM
Ces trois biovars que donne Euzéby ne sont pas repris dans la littérature. En effet, l’étude par séquençage de plusieurs gènes a dégagé l’existence de plusieurs variants mais on peine toujours à les regrouper dans les trois biovars précités. Les gènes étudiés sont le gène de l’ARNr 16S, les gènes codant pour la protéine de surface p44 (gènes msp2 et msp4) (De la Fuente et al., 2005) , le gène ankA, l’opéron groESL (Bown et al., 2007)
En Amérique du Nord, le séquençage des gènes de protéines de surface a fait la distinction entre des variants qui avaient des propriétés biologiques et écologiques différentes. En revanche, en Europe, des variants distingués par séquençage ont été retrouvés chez les mêmes hôtes. L’apparition de la méthodes de Multilocus Variable-Number Tandem-Repeat Analysis utlisée par Bown et al. (2007) permet une distinction plus fine des souches et les travaux de 2007 ont montré que sur 20 variants de bactéries issues d’hôtes et de pays différents (Danemark, Slovénie et Grande Bretagne), seulement deux partageaient les mêmes allèles sur 4 loci distincts.
Chaque variant atteint une certaine catégorie d’espèces avec un pouvoir pathogène propre mais plusieurs variants peuvent cohabiter chez un même hôte (Stuen et al., 2002) et un seul variant peut infecter plusieurs hôtes. Ce potentiel pathogène étant en partie lié à la protéine p44 on comprend que le séquençage des gènes de cette protéine dégage des variants avec des potentiels pathogènes variés et que le nombre de variants obtenus soit plus important qu’avec d’autres gènes comme l’ARNr16s.
On ne peut donc affirmer, si l’on s’en tient aux études récentes, qu’il y ait trois biovars clairement définis par leur génome et leur spectre d’hôtes. On peut supposer que certains variants ont des caractéristiques proches, regroupées éventuellement en trois dominantes et que les trois espèces touchées historiquement (ovins, chevaux et humains) l’étaient peut-être par des souches

28
différentes. Ce n’est pas le sujet de notre étude, nous ne nous attarderons pas plus longtemps sur ce point.
Gardons à l’esprit que pour l’espèce hôte qui nous intéresse, c’est à dire le Chevreuil, d’une part A. phagocytophilum est présent et d’autre part il y a sûrement plusieurs variants qui touchent cette espèce, chaque souche ayant sa propre virulence et peut-être ses propres réservoirs.
Bien qu’il soit établi que plusieurs variants existent, l’ensemble de ces variants partage le même vecteur et provoque les mêmes signes cliniques par les mêmes mécanismes pathogéniques, en France en tout cas. Nous ne chercherons donc pas à faire la distinction entre les souches dans l’étude bibliographique de la maladie qui suit.
Ici nous nous attacherons donc à présenter tous les hôtes qui se sont été révélés porteurs d’A. phagocytophilum. Dans un second temps nous essayerons de dégager des spectres d’hôtes pour les trois souches décrites. Il faudra ici tenir compte du fait que les connaissances sur les spectres d’hôtes ne sont pas complètes en ce qui concerne A. phagocytophilum.
.II.A.3 DONNEES SUR LES CARACTERISTIQUES BIOLOGIQUES D’A. PHAGOCYTOPHILUM
II.A.3.a Généralités
L’ehrlichiose est une maladie bactérienne qui touche de nombreux mammifères. Il s’agit d’une maladie vectorielle stricte transmise par les tiques du genre Ixodes. La bactérie colonise les cellules de la lignée myéloblastique et provoque, outre un syndrome inflammatoire, une immunodéficience chez l’hôte.
Cette maladie est due une bactérie intra-cellulaire stricte GRAM négatif. Elle partage les caractéristiques biologiques typiques d’une bactérie intracellulaire (Liz 1994).
II.A.3.b Morphologie et propriétés antigéniques de la bactérie
A. phagocytophilum est une petite bactérie en forme de coque ou coccoïde de 0,4 à 0,5µm de diamètre, colorée négativement à la coloration de GRAM, observable au microscope à immersion. Au microscope électronique on distingue une paroi externe ondulée doublée d’une membrane interne. Le cytoplasme contient des amas ribosomiques sous la membrane et l’ADN (Liz, 1994).
Ces bactéries sont observables dans les granulocytes neutrophiles infectés. Les premiers stades de la multiplication ne sont observables qu’au microscope électronique car ils sont caractérisés par des corps élémentaires de 0,4 à 2µm de diamètre au sein d’une vacuole d’endocytose.
Ce stade précoce est suivi par la multiplication des bactéries par fission binaire au sein de la vacuole d’endocytose et 24 heures plus tard on peut observer au microscope optique des amas de bactéries appelés morulae du fait de leur aspect de mûres. Ces amas mesurent de six à huit micromètres de diamètre (Liz 1994).
Il s’agit d’une bactérie Gram-1 qui à l’instar d’autres bactéries intracellulaires ne possède pas de lipopolysaccharide ou de structure proche qui sont les structures antigéniques des bactéries GRAM-. Les structures porteuses d’antigènes sont des protéines membranaires qui permettent l’attachement et la pénétration de la bactérie dans les cellules cibles.
A ce stade une coloration de May-Grünwald-Giemsa permet de voir les morulae et de diagnostiquer la portage avec certitude (Liz, 1994).
1 La bactérie ne prend pas la coloration de GRAM et apparaît rosée sur la lame (couleur violette pour les GRAM+)

29
.II.B. Pouvoir pathogène d’A. phagocytophilum
.II.B.1 TRANSMISSION VECTORIELLE
A. phagocytophilum ne peut traverser les barrières cutanées et muqueuses et doit donc être injectée pour pouvoir atteindre les cellules cibles : les granulocytes neutrophiles et éosinophiles.
Ce passage à travers la barrière cutanée est permis par la piqûre des tiques du genre Ixodes. En France le vecteur est la tique Ixodes ricinus mais on trouve d’autres espèces d’Ixodes dans d’autres pays d’Europe et d’Amérique du nord.
A. phagocytophilum se maintient et se multiplie très bien dans les glandes salivaires de la tique mais la pénétration de la bactérie dans la tique nécessite un repas de sang d’au moins 30 heures sur un animal infecté. Il faut un repas de sang d’au moins 24 heures pour la transmission de tique à animal (Foley et al. 2006).
Si la tique permet effectivement un maintien de la bactérie dans son organisme, ce maintien ne se perpétue pas à travers les générations car il n’y a pas de transmission trans-ovarienne. Ceci implique qu’une tique doit effectuer au moins un repas de sang de 30 heures sur un animal infecté pour être infectée elle-même. Par contre ce repas suffit puisqu’il y a transmission transstadiale (Liz 1994).
Une tique doit donc faire un repas de sang sur un animal infecté pour être infectante mais par contre elle reste infectante sur tout son cycle après infection.
Ixodes ricinus parasite tous les mammifères de nos campagnes à des stades différents du fait de sa biologie (Guétard, 2001). La figure 9 montre le cycle épidémiologique d’A. phagocytophilum.
Figure 9: Cycle infectieux d'A. phagocytophilum et rôle de la tique Ixodes ricinus

30
.II.B.2 ENTREE DANS L’ORGANISME
La bactérie est injectée dans le sang de l’hôte de façon passive par un vecteur (tique) ou un autre moyen (transfusion).
L’entrée dans l’organisme ainsi que l’inflammation provoquée par la piqûre de la tique ont pour conséquence un afflux de leucocytes, macrophages et granulocytes, sur le lieu d’entrée de la bactérie. Ainsi les cellules cibles, les granulocytes, affluent auprès de la bactérie.
La bactérie est phagocytée par les granulocytes par attachement à leurs protéines de surface (Jason, 2003). Le délai entre l’attachement et la pénétration est de 240 minutes au cours desquelles le production d’hyperoxydes par le granulocyte est activée et entame les réserves en hyperoxydes (Carlyon et al., 2004).
.II.B.3 SURVIE ET MULTIPLICATION DANS LES GRANULOCYTES
Elle survit dans des phagosomes au sein de la cellule en inhibant la fusion entre les phagosomes et les lysosomes.
L’inhibition de la fusion phagosome-lysosome est annulée par l’administration d’oxytétracycline ce qui prouve que la bactérie doit être vivante pour que cette inhibition soit maintenue. On peut associer l’inhibition à une production continue de facteurs d’inhibition par la bactérie.
La bactérie, en plus d’inhiber la fusion phagosome-lysosome, provoque également une baisse de l’expression des protéines d’adhésion à la surface des granulocytes, diminuant de 50 % la capacité de phagocytose des granulocytes sus-décrits (Garyu et al. 2005).
Il y a plusieurs mécanismes qui permettent la survie d’A. phagocytophilum, la première est la protection que lui confère le phagosome contre le matériel enzymatique du granulocyte, la deuxième est la haute tolérance de la bactérie aux hyperoxydes et le dernier mécanisme est la dépression engendrée sur les autres cellules de l’immunité.
A. phagocytophilum ne commence à se multiplier que 48 à 72 heures après pénétration dans la cellule. Cette multiplication au sein d’un granulocyte présuppose une perturbation des mécanismes apoptotiques de la cellule car les granulocytes ont une demi-vie d’environ 48 à 72 heures.
Cette perturbation des mécanismes apoptotiques serait due au contact de la bactérie avec les protéines de surface des granulocytes. En effet l’oxytétracycline tue la bactérie mais ne raccourci pas la durée de vie de la cellule infectée ce qui mène à penser que cette vie prolongée n’est pas due à un mécanisme actif de la bactérie. Le mécanisme de l’apoptose est inhibé par l’augmentation de la transcription de gènes anti-apoptotiques (MCL1,BFL1)
.II.B.4 VIRULENCE
II.B.4.a Les protéines de surface
La virulence de la bactérie serait due à une catégorie de protéines membranaires : les protéines p44.
Ces protéines provoqueraient l’inflammation par activation de cytokines proinflammatoires. Cette présomption est confirmée par le fait que des anticorps anti-p44 neutralisent l’infection par A. phagocytophilum.
L’intérêt de cette protéine p44 est que selon l’environnement de la bactérie – un hôte différent, la tique vecteur – sa structure sera différente. A chaque environnement une structure particulière de la protéine p44 est majoritaire.

31
Plusieurs structures de la protéine p44 existent et l’une d’entre elle est exprimée en majorité selon l’environnement de la bactérie. Ce mécanisme est courant chez les bactéries intracellulaires, on le retrouve pour la protéine de surface msp2 chez Anaplasma marginale qui est apparentée à la protéine p44 d’A. phagocytophilum (Barbet et al., 2006). La protéine p44 d’A. phagocytophilum a d’abord été appelée msp2 du fait son analogie avec la protéine de surface d’Anaplasma marginale.
La caractéristique de cette protéine est que le gène codant pour la protéine p44 est hypervariable. C’est le gène msp2. En effet il y a plusieurs dizaines de loci de ce gène dans le génome de la bactérie qui sont exprimés chacun selon les pressions exercées par l’environnement.
Ainsi il y a une grande variation inter-variants et une variation à l’échelle de la vie de l’individu.
Ce mécanisme permet à la bactérie de vivre dans plusieurs organismes différents et de survivre dans la tique.
La relation entre l’immunité et la variabilité de la protéine p44 reste à établir. Il semblerait que cette variabilité permette la persistance de la bactérie dans l’organisme par perturbation de l’immunité humorale (Granquist et al., 2008).
Il y a une variabilité importante au niveau des protéines de surface et cette variabilité se traduit dans les manifestations cliniques de la maladie qui peuvent être différentes selon le variant considéré (Stuen et al., 2003).
Certaines études formulent l’hypothèse que la structure de p44 serait à l’origine des virulences variables des différentes souches d’A. phagocytophilum pour l’Homme et expliquerait notamment la différence de virulence entre les souches américaines, d’Europe septentrionale et d’Europe du nord.
II.B.4.b Les mécanismes de survie d’A. phagocytophilum
La pénétration et la multiplication de la bactérie au sein des granulocytes provoque la production d’interleukines et de la TNFα ainsi que celle du médiateur endogène leucocytaire (Woldehiwet, 2007).
Cet ensemble de facteurs sont des médiateurs de l’inflammation, leur production provoquant une inflammation et des modifications importantes de la formule sanguine.
Par ailleurs, le médiateur endogène inhibe l’absorption de fer et de zinc. Le zinc a une importance majeure dans plusieurs mécanismes notamment dans plusieurs mécanismes enzymatiques, hormonaux et pour certaines vitamines.
Il est possible d’après certaine auteurs (Gokce & Woldehiwet, 1999) que ce déficit en zinc provoque des avortements par mort fœtale, des retards de croissance, des perturbations de la spermatogénèse, une anorexie.
.II.C. L’ehrlichiose granulocytaire
.II.C.1 EPIDEMIOLOGIE
Comme pour toutes les maladies vectorielles, il y a un cycle précis suivant celui du vecteur avec des animaux réceptifs et des animaux sensibles. Il y a également des animaux réservoirs de la maladie.
Bien que le regroupement des trois ehrlichioses granulocytaires historiquement décrites soit effectif depuis 2002, on parle toujours d’ehrlichiose et non d’anaplasmose et les trois maladies sont toujours distinguées : l’ehrlichiose granulocytaire bovine, l’ehrlichiose équine et l’ehrlichiose granulocytaire humaine. A ces trois maladies sont associés directement les trois biovars dont il a déjà été question : phagocytophila, equi et EGH.

32
Cette séparation est beaucoup moins nette en réalité. En effet si l’on identifie bien trois maladies, il est difficile de distinguer trois biovars différents tant sur le plan génétique que sur le plan biologique. En effet on peine à donner trois réservoirs distincts à ces trois biovars. Pour cette raison il apparaît difficile de donner un tableau ferme et définitif des espèces sensibles et réceptives pour chaque biovar. Nous essaierons cependant de distinguer d’éventuelles spécificités de souches.
II.C.1.a Les hôtes d’A. phagocytophilum
De nombreuses espèces sont réceptives ou sensibles à A. phagocytophylum. Nous l’avons vu trois biovars sont donnés mais il existe une grande variété de variants et il est difficile de savoir quelle souche touche quel animal.
On peut cependant décrire plusieurs groupes touchés par l’ehrlichiose :
Les ongulés domestiques et sauvages : l’ehrlichiose est une maladie bien connue chez les bovins les caprins et surtout chez les ovins. Elle est appelée fièvre à tiques, de l’anglais « tick borne fever » ou maladie des gros pâturons. Nous étudierons plus en détail l’ehrlichiose granulocytaire bovine car c’est chez les bovins que la maladie a été décrite pour la première fois en France.
Les ongulés sauvages sont également touchés. Le Chevreuil l’est mais d’autres espèces le sont également : le Cerf élaphe, le Daim, le Chamois et aux Etats-Unis le Cerf de virginie (Stuen et al., 2002,)
Les chevaux : les cas sont rares en France, il y en a beaucoup aux Etats Unis
Les micromammifères : de nombreux mammifères ont été trouvés positifs par PCR à l’ehrlichiose : Apodemus sylvaticus, Apodemus flavicolis, Clethrionimys glareolus, Sorex araneus en Europe et d’autres aux Etats unis : Peromyscus leucopus, Neotoma fuscipes, Tamias striatus, Clethrionimys gapperi (Bown et al., 2003, Barandika et al., 2007). A l’heure actuelle, le rôle des micromammifères dans l’épidémiologie de l’ehrlichiose granulocytaire reste à établir. En effet si il avait semblé que ces espèces pouvaient être un réservoir de la maladie et permettre le maintien de la maladie durant l’hiver, aucun portage long n’a pu être mis en évidence. Les auteurs ayant travaillé sur le rôle des micromammifères considèrent que la persistance de la bactérie durant l’hiver serait due surtout au maintien chez la tique.
Le chien est également une espèce hôte.
L’Homme est touché par cette maladie. Aux Etats Unis il s’agit d’une zoonose reconnue à laquelle on peut rattacher des cas cliniques et même mortels. En Europe en revanche, il n’y a eu que des cas cliniques modérés en Europe centrale malgré des taux de séropositivité élevés dans des zones à tiques partout en Europe (Strle, 2004). L’ehrlichiose granulocytaire n’est pas une zoonose majeure en Europe.
Il semblerait que les oiseaux aient également un rôle de réservoir. C’est en tout cas ce qui a été observé aux Etats Unis sur le Merle d’amérique (Turdus migratorius) et la Grive fauve (Catharus fuscescens) (Daniels et al., 2002) . Ceci n’a pas été retrouvé en Europe parce qu’il n’y a pas eu de travaux sur le sujet.
Toutes les espèces citées sont touchées avec une gravité plus ou moins importante mais on ne retrouve pas les mêmes souches selon l’espèce considérée.
Aucune souche commune entre ongulés sauvages et humains n’a été trouvée. Par contre il existe des souches communes à l’Homme et aux chevaux qui sont pathogènes pour les deux.
On a pu établir que les souches de ruminants domestiques sont les mêmes que pour les ruminants sauvages.
Des souches communes ont été retrouvées chez plusieurs hôtes et il a été montré des communautés de spectres d’hôtes. Le tableau 7 montre les différents liens qui ont pu être établis dans les spectres des différents variants.
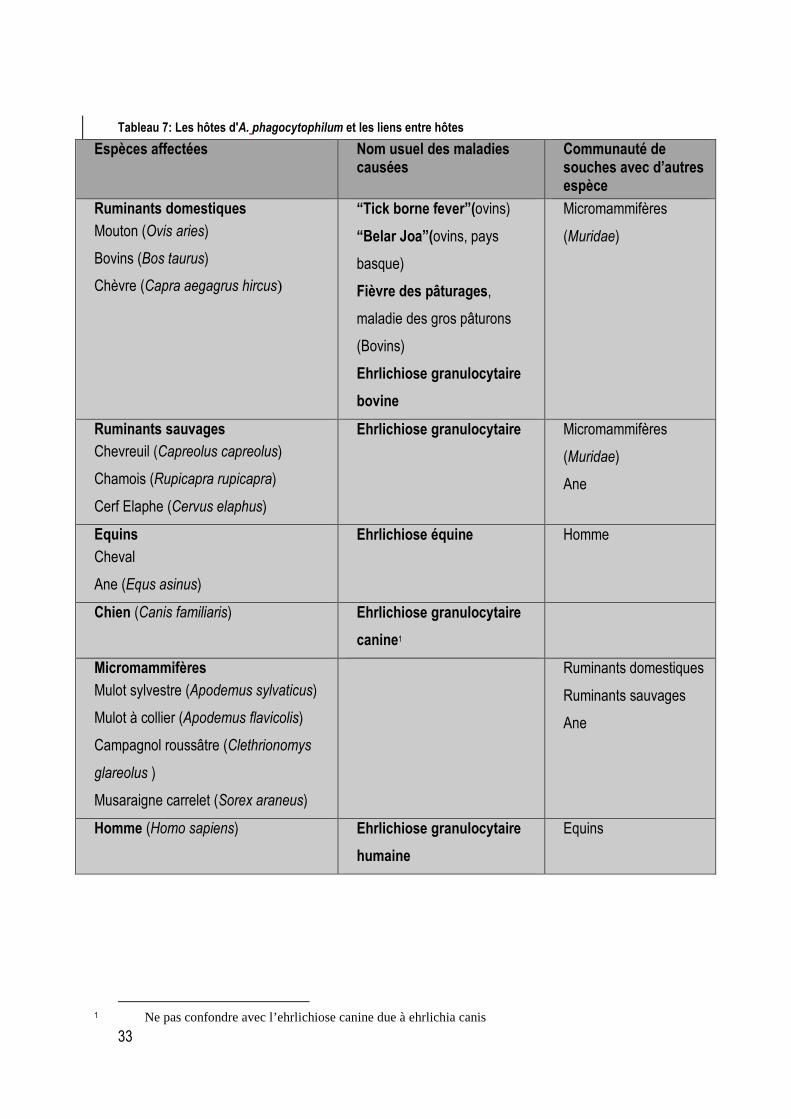
33
Tableau 7: Les hôtes d'A. phagocytophilum et les liens entre hôtes
Espèces affectées Nom usuel des maladies causées
Communauté de souches avec d’autres espèce
Ruminants domestiques
Mouton (Ovis aries)
Bovins (Bos taurus)
Chèvre (Capra aegagrus hircus)
“Tick borne fever”(ovins)
“Belar Joa”(ovins, pays
basque)
Fièvre des pâturages,
maladie des gros pâturons
(Bovins)
Ehrlichiose granulocytaire
bovine
Micromammifères
(Muridae)
Ruminants sauvages
Chevreuil (Capreolus capreolus)
Chamois (Rupicapra rupicapra)
Cerf Elaphe (Cervus elaphus)
Ehrlichiose granulocytaire Micromammifères
(Muridae)
Ane
Equins
Cheval
Ane (Equs asinus)
Ehrlichiose équine Homme
Chien (Canis familiaris) Ehrlichiose granulocytaire
canine1
Micromammifères
Mulot sylvestre (Apodemus sylvaticus)
Mulot à collier (Apodemus flavicolis)
Campagnol roussâtre (Clethrionomys
glareolus )
Musaraigne carrelet (Sorex araneus)
Ruminants domestiques
Ruminants sauvages
Ane
Homme (Homo sapiens) Ehrlichiose granulocytaire
humaine
Equins
1 Ne pas confondre avec l’ehrlichiose canine due à ehrlichia canis

34
.II.C.2 ROLE DE LA TIQUE IXODES RICINUS COMME VECTEUR D’A. PHAGOCYTOPHILUM
II.C.2.a Biologie de la tique
C’est un arthropode chélicérate de la classe des arachnidae. C’est une tique de la famille des Ixodidés (Tiques dures) qui regroupe d’autres espèces jouant également un rôle dans la transmission de l’ehrlichiose dans d’autres pays : (Guétard, 2001)
• Ixodes trianguliceps (Royaume Uni)
• Ixodes scapularis (Etats Unis)
• Ixodes ventalloi (Portugal)
• Ixodes pacificus (Etats Unis)
• Ixodes persulcatus (Pays de l’Est, Russie, Chine et Japon)
Ixodes ricinus est un ectoparasite qui effectue des repas de sang sur les mammifères, les oiseaux mais aussi les reptiles.
La tique se fixe sur son hôte et migre de son site de fixation vers des sites où la peau est fine (pavillons, encolure, plis de l’aine ; scrotum) où elle transperce la peau à l’aide de son rostre.
II.C.2.b Répartition et habitats d’Ixodes ricinus
On retrouve Ixodes ricinus sur l’ensemble du territoire français. En effet les biotopes français sont idéaux pour la tique qui colonise des milieux abrités au moins une partie de l’année par une végétation dense. Ainsi les forêts, lisières, bordures ou les haies bocagères sont des milieux très propices au maintien des populations d’Ixodes ricinus. Elle est cependant absente sur le pourtour méditerranéen dans les zones de garrigues.1 (Guétard, 2001)
La tolérance d’Ixodes ricinus pour des conditions plus difficiles -les grands froids et inversement les grandes chaleurs- lui permet de coloniser des milieux qui paraissent moins favorables. De ce fait on la retrouve en Grande Bretagne, en Scandinavie, en Europe orientale et septentrionale jusque dans le tiers Nord de l’Espagne.
Seuls le Sud de l’Espagne et la Grèce ne sont pas colonisés par Ixodes ricinus.
On retrouve même Ixodes ricinus dans des pays chauds notamment les pays du Maghreb avec un cycle légèrement différent2.
Les seuls milieux qui ne soient pas colonisés par Ixodes ricinus en Europe sont les zones de moyenne et haute montagne. En effet les densités de population diminuent à partir de 1000 mètre pour être en principe nulles vers 1500 mètres. Il semblerait que le réchauffement climatique provoque une remontée en altitude de ces tiques et qu’on les retrouve aujourd’hui dans des zones qui en étaient exemptes auparavant ( Communications personnelles)
II.C.2.c Cycle biologique d’Ixodes ricinus
Cette tique a un cycle qui dure deux ans. On distingue trois stades : larve, nymphe et adulte.
A chaque stade à lieu au moins un repas de sang qui dure 30 à 60 heures.
1 Des températures trop élevée ainsi qu’une humidité trop faible sont probablement responsables de l’absence d’Ixodes ricinus dans ces régions. 2 Les températures étant plus élevées dans le Maghreb, le pic d’activité à lieu en Hiver et non au printemps

35
Chaque stade a ces particularités.
Les larves naissent à partir de nids d’œufs situés dans des micro-climats propices à leur développement dont elles ne bougent pas jusqu’à la fixation sur le premier hôte. Les larves sont très peu mobiles car très dépendantes des températures et de l’hygrométrie qui règnent dans le nid. Elles peuvent cependant se mettre à l’affut à quelques centimètres de hauteur. Avant cet affut, les larves sont inactives pendant environ deux semaines pour finir leur mue et se débarrasser des reste d’éclosion (Guétard, 2001).
La phase d’attente d’un hôte dure quelques semaines mais peut durer plus longtemps (jusqu’à 520 jours) en cas de conditions défavorables.
Les larves peuvent se nourrir sur tous les types d’hôtes : petits et grands mammifères, oiseaux, reptiles, humains. Cependant du fait de leur méthode de chasse – la larve monte à quelques centimètres de hauteur pour s’accrocher à un hôte sur son passage – elles se fixent en grande majorité sur des petits mammifères, des oiseaux ou des reptiles. Voici quelques espèces qui sont classiquement infestées par des larves d’Ixodes ricinus :
• Rongeurs et insectivores : mulot sylvestre (Apodemus sylvaticus), mulot fauve (apodemus flavicollis), campagnol roussâtre (Clethrionomys glareolus), campagnol agreste (Microtus agrestis), campagnol des champs (Microtus arvalis) ainsi que la musaraigne carrelet (Sorex araneus) et le hérisson (Erinaceus europaeus).
• Oiseaux de la famille des turdidés : grive, merle (Turdus), rouge gorge familier (Erithacus rubecula)
• Lézards :lézard vert (Lacerta viridis)
• Le repas de sang dure trois à sept jours et à la fin de ce repas les larves se laissent tomber au sol. La phase de mue arrive environ quatre à huit semaines après le repas de sang ; c’est le délai de digestion du sang.
A la fin de cette mue, la larve devient une nymphe de poids et taille équivalent à la larve gorgée (Guétard, 2001).
Les nymphes sont, comme les larves, peu mobiles car elles sont elles aussi très sensibles à la dessication. Elle peuvent cependant se mettre à l’affût quelques centimètres en hauteur et sont plus dispersées dans la végétation contrairement au larves qui sont en nids (Guétard, 2001).
L’affut peut commencer deux à trois semaines après la mue et dure jusqu’au passage d’un hôte soit quelques semaines le plus souvent.
Les nymphes se fixent indifféremment sur des petits hôtes ou des grands. Ainsi les hôtes que nous venons de citer pour les larves sont également infestés par des nymphes et ceux que nous allons citer pour les femelles adultes le sont également (Guétard, 2001).
Le repas de sang dure trois à six jours et il est suivi d’une phase de repos menant à la mue en deux à cinq mois.
La tique adulte se met à l’affût jusqu’à 70 cm de haut dans la végétation. En effet le stade adulte est beaucoup moins inféodé aux conditions du milieu et se déplace plus librement (Guétard, 2001).
Les adultes se fixent uniquement sur des grands mammifères car elles se fixent à plus d’un mètre de hauteur dans l’attente du passage d’un hôte ce qui rend impossible l’infestation d’espèces se déplaçant au niveau du sol. Les hôtes infestés par des adultes sont :
• Les grand ongulés sauvages : Chevreuil, cerf élaphe, daim
• Le sanglier et le porc
• Les grands animaux d’élevage : bovins, ovins, caprins
• Les équidés : cheval, âne ; baudet…
• Les humains

36
Le repas dure 4 à 14 jours et il est suivi chez la femelle d’une phase de préponte liée à la digestion du sang et à la maturation des œufs.
Le femelle pond 500 à 3000 œufs sur cinq à 20 jours dans un milieu propice aux larves à quelques centimètres du point de chute après le repas de sang. A la fin de la ponte, la femelle meurt en quelques jours (Guétard, 2001).
La maturation des ovocytes que porte la femelle nécessite une fécondation par un mâle adulte. Nous allons donc voir maintenant le rôle du mâle adulte dans le cycle.
La figure 10 présente le cycle évolutif d’Ixodes ricinus.
Figure 10: Le cycle biologique d'Ixodes ricinus
Tous les stades du cycle sont inféodés à deux conditions importantes du milieu : l’hygrométrie et la température.
Dans les zones tempérées telles que la France, c’est surtout la température qui est importante. Ixodes ricinus est capable de survivre de -10 à 32°c mais n’entre en activité qu’à partir de 7°c. Des températures basses rallongent les durées de chaque phase surtout sur les stades larvaire et nymphal (Guétard, 2001).
Ce sont les températures qui déterminent chaque année les pics d’activité (ie les pics d’infestation) et des modèles prédictifs d’infestation basés sur les températures ont été testés avec une grande fidélité aux réels niveaux d’infestation.
II.C.2.d Interactions hôte-Ixodes ricinus
La tique se fixe sur son hôte puis migre vers une zone peu poilue, où la peau est plus fine. Les zones de fixation des tiques sont souvent autour de l’encolure, au niveau des pavillons auriculaires, dans les plis de l’aine, au niveau de l’arse, sur le scrotum (Guétard, 2001).
Une fois que la tique a trouvé un point de fixation idéal, elle mort son hôte. Cette morsure se passe en deux temps :
• Les chélicère et les crochets percent la peau et permettent l’enfoncement de l’hypostome dans l’organisme.

37
• Un cément est sécrété autour du rostre et solidifie la fixation à l’hôte.
Une fois la morsure effectuée, la tique commence son repas de sang à partir d’un foyer de lyse à l’extrêmité de son rostre.
L’ensemble de ces opérations provoque une réponse inflammatoire non spécifique avec un afflux de polynucléaires neutrophiles et éosinophile sur le site de morsure au bout de 48 heures (délai d’apparition des granulocytes lors d’une inflammation). On comprend ici l’intérêt pour A. phagocytophilum de l’afflux de granulocytes sur un site de morsure de tique : les cellules cibles arrivent directement sur le site d’injection de la bactérie (Guétard, 2001).
On observe, associé à la morsure des lésions de types macules ou erythèmes. Lors de contacts réitérés, la réaction inflammatoire peut devenir allergique
.II.C.3 L’EHRLICHIOSE : ETUDE CLINIQUE CHEZ LES RUMINANTS
L’ehrlichiose est une maladie qui a été diagnostiquée tardivement en France (premier cas en 1991). Les raisons de ce diagnostic tardif sont multiples. L’une d’entre elles est la méconnaissance que les vétérinaires avaient de cette maladie, pourtant bien connue par les éleveurs dans certaines régions de la France comme au pays Basque par exemple où l’agent de la Belar Joa, maladie d’estive des ovins, s’est avéré être due à A. phagocytophilum.
Il s’agit d’une maladie difficile à diagnostiquer- surtout quand on n’en connaît pas l’existence- parce que les signes cliniques qu’elle engendre sont peu spécifiques et souvent peu exprimés. Il faut donc une bonne connaissance de la maladie pour pouvoir la diagnostiquer.
Les données sur les signes cliniques de l’ehrlichiose granulocytaire chez le Chevreuil étant très rares, nous avons décidé de présenter la maladie chez l’espèce la mieux étudiée : les bovins. Nous compléterons cette description par les données existant chez le Chevreuil et chez les ovins/caprins.
La raison de l’absence de données sur le Chevreuil est due au fait qu’il est très difficile de maintenir des Chevreuils en captivité pour prélever du matériel biologique tous les jours sans modifier les résultats et sans faire face à des mortalités dues au stress.
II.C.3.a Le diagnostic clinique de l’ehrlichiose bovine
L’ehrlichiose bovine est une maladie sous-diagnostiquée en France dont les signes cliniques sont difficiles à objectiver. Le diagnostic reste faisable en prenant en compte les conditions d’apparition de la maladie, associées à la biologie du vecteur Ixodes ricinus (Joncour, 2003).
L’ehrlichiose bovine est une maladie qui apparaît dans un troupeau sur un mode annuel bi phasique calqué sur les périodes de gorgement des tiques qui atteignent leurs plus hauts niveaux au printemps et à l’automne. Ces périodes varient de façon importante en fonction de la température et du climat. Ainsi on pourra observer de l’ehrlichiose pendant un hiver doux ou un été pluvieux. Le mode biphasique est donc purement théorique et il faut tenir compte des conditions climatiques lors d’une suspicion d’ehrlichiose, quelle que soit la période de l’année (Joncour, 2003).
Cette maladie peut atteindre un seul individu dans un troupeau mais peut revêtir un aspect épizootique avec plusieurs animaux qui expriment les mêmes signes cliniques sur un court intervalle.
L’inoculation du germe se fait 30 à 36 heures après la morsure par la tique, à la fin du gorgement de la tique et est suivie d’une incubation de cinq à huit jours. Cette incubation longue empêche parfois de trouver des ectoparasites pendant l’examen clinique qui pourraient orienter vers une maladie vectorielle.
Les symptômes sont peu spécifiques de la maladie ce qui rend le diagnostic difficile, ces symptômes étant partagés par d’autres maladies.

38
Les principaux signes cliniques sont liés à l’inflammation que provoque la multiplication d’A. phagocytophilum.
II.C.3.b Signes cliniques chez les bovins
Chez les bovins, seule une infection aiguë peut être diagnostiquée. Cette infection aiguë est généralement de courte durée et observable uniquement en début d’épisode au sein d’un troupeau.
Les signes cliniques de l’ehrlichiose sont liés directement à l’inflammation provoquée par la multiplication de la bactérie (Joncour, 2003).
Le premier signe d’alerte pour l’éleveur et souvent le motif d’appel du vétérinaire est l’agalaxie qui est souvent brutale (chutes de plus de 50 % de la production habituelle).
Ce premier signe s’accompagne d’une forte hyperthermie qui oscille entre 39,5 et 42°c et plus forte en début d’infection.
Une anorexie est souvent repérée. Elle est parfois sélective et mène à un amaigrissement à terme.
Une syndrôme grippal est observé, il est caractérisé par une toux sèche inductible par palpation de la trachée, de la polypnée et un essoufflement. La toux est parfois grasse et productive.
Des troubles de la locomotion sont observés ainsi que de l’asthénie. Il peut y avoir un œdème des partie déclives des membres : pâturons surtout et parfois jarrets
On peut observer plus rarement des troubles de la reproduction (troubles de la spermatogénèse, infertilité) ainsi que des avortements.
II.C.3.c Signes cliniques chez la Chèvre
Afin de se rapprocher le plus possible du modèle Chevreuil pour les signes cliniques et les modifications biochimiques et cellulaires, plusieurs travaux de recherche ont utilisé la chèvre comme modèle de travail à la place d’ongulés sauvages (Gokce & Woldehiwet, 1999a).
La chèvre semble moins sensible que le Mouton dans des conditions expérimentales mais développe quand même des signes cliniques.
Les signes cliniques chez la chèvre sont proches de ceux des bovins :
• élévation abrupte de la température rectale,
• anorexie, affaiblissements, tremblements, tachypnée, toux d’été.
En plus de ces signes cliniques, la Chèvre peut émettrer des « grognements » et plus souvent des sécrétions lacrymales (Gokce & Woldehiwet, 1999a).
Les signes cliniques sont donc très proches chez les différentes espèces et se caractérisent par un syndrome inflammatoire avec fièvre forte et précoce suivie de signes liés à l’inflammation (abattement, œdème des pâturons, anorexie, avortements). Ce tableau clinique doit évoquer l’ehrlichiose granulocytaire bovine. Bien qu’ils soient plus rares, les troubles de la fertilité et les avortements peuvent être attribués à l’ehrlichiose. Cette maladie ne doit donc pas être oubliée parmi les différents agents provoquant des avortements (Gokce & Woldehiwet, 1999a).
Il est à noter que si les cas de mortalité sont rares en France, dans les pays scandinaves et en Ecosse il a été observé des cas de mortalité chez les ovins atteignant jusqu’à 30% dans certains troupeaux.
II.C.3.d Les ehrlichioses sub-aiguës et chroniques
Les cas d’ehrlichiose subaiguë sont problématiques car difficiles à objectiver et lourds de conséquences quand ils touchent tout un troupeau.

39
En effet on observe des signes cliniques très frustes : principalement une baisse diffuse de la production lactée, des poussées de fièvre et une sensibilité accrue aux infections.
Si un cas d’ehrlichiose clinique n’est pas diagnostiqué (et donc pas traité), la Vache survit dans la plupart des cas, mais sa récupération est très lente surtout pour la formule sanguine qui peut subir des rechutes régulières. Cet état de récupération peut durer plusieurs mois au cours desquels l’individu est sensible aux infections (Reid, 1986).
Certaines infections sont classiquement associées à l’ehrlichiose (Liz, 1994) :
• pyohémie à tiques (abcès pultiples à Staphylococcus aureus),
• pneumonies à pasteurelles, RSV, fièvre Q (Coxiella burnetii),
• listérioses oculaires (Listeria monocytogenes),
• mammites et augmentation du taux de cellules dans le lait,
• entérotoxémies (Clostridium sp),
• babésiose (Babesia divergens),
• anaplasmose (Anaplasma marginale),
Nombre de ces maladies sont des maladies vectorielles transmises par les tiques (Fièvre Q, Babésiose, anaplasmose) ; la présence de l’une d’entre elles sur une exploitation doit donc faire penser aux autre maladies vectorielles. La présence d’une babésiose doit donc appeler à rechercher A. phagocytophilum si les signes cliniques sont évocateurs.
Des portages multiples ont d’ailleurs été mis en évidence chez les bovins et dans la faune sauvage.
Les ehrlichioses chroniques n’ont pas encore été décrites chez les bovins bien qu’il soit possible qu’il y en ait. Les bovins semblent réagir de façon plus violente à la maladie. De ce fait la bactérie provoquerait une maladie dans la plupart des cas mais ne persisterait pas longtemps ; constat qui est peu compatible avec un portage à long terme. Chez les ovins et caprins en revanche un portage long et asymptomatique avec possibilité de transmission a été relevé.
.II.C.4 LES MODIFICATIONS DE LA NUMERATION CELLULAIRE ET DE LA BIOCHIMIE
II.C.4.a Les modifications de la numération cellulaire
A. phagocytophilum pénètre dans les granulocytes neutrophiles et éosinophiles et s’y multiplie provoquant leur mort. Il y a donc des modifications importantes de la formule sanguine lors d’infections cliniques. Ces modifications sont les mêmes chez les bovins, ovins et caprins bien que l’importance relative de chaque modification ne soit pas exactement la même du fait des différences de sensibilités (Gokce et al., 1999a).
Ici sont évoquées études expérimentales menées sur les ovins et caprins (Woldehiwet et al., 1999a).
On observe une leucocytopénie précoce qui commence quelques jours seulement post-infection (3 à 4 jours). Ce sont les lymphocytes qui sont touchés les premiers car ils chutent dès le début de l’infection.
Par la suite, les granulocytes neutrophiles baissent à partir de quatre à cinq jours post-infection après avoir augmenté sur les premiers jours de l’infection (Woldehiwet et al., 1999a).
La baisse des neutrophiles est inversement proportionnelle au nombre de leucocytes infectés qui augmente pour atteindre un pic vers cinq à six jours post-infection.
On observe également une éosinopénie et une monocytose.
La numération érythrocytaire diminue fortement environ cinq jours post-infection ; une étude a montré que chez des chèvres, la numération érythrocytaire passe en 12 jours d’environ

40
15,41.106Cellules/ml à 11,7.106Cellules/ml. Il y a une baisse forte du nombre d’hématies. Cette baisse est durable, la même étude ayant montré l’absence de remontée de la numération 21 jours post-infection (Woldehiwet et al., 1999a).
Il y a une baisse concomitante de la concentration en hémoglobine et du taux d’hématocrite.
Une thrombocytopénie est observée durant une semaine à partir du cinquième jour mais la quantité de plaquettes remonte et atteint même des niveaux très supérieurs aux niveaux pré-infection. Ces niveaux élevés se maintiennent longtemps ( les niveaux se maintiennent à 21 jours post-infection).
Il peut y avoir une baisse du taux d’hématocrite provoquant l’apparition sporadique de pétéchies sur les muqueuses.
Dans un cas clinique aigu il n’y a donc pas uniquement les risque liés directement à l’ehrlichiose. Il faut également prendre en compte les risques liés à l’immunodépression.
Cette immunodépression rend possibles des infections par des germes opportunistes (pasteurelles, staphylocoques) ainsi que des infestations parasitaires massives (Woldehiwet et al., 1999a).
II.C.4.b Les modifications biochimiques
Les modifications énoncées ici sont issues d’une étude menée sur des chèvres et des ovins (Gokce et al., 1999b) Des modifications significatives de plusieurs paramètres d’importance ont été observées et sont rapportées dans le tableau 8.
Tableau 8 : Les modifications biochimique dues à l'ehrlichiose chez la chèvre et le mouton
Modifications biochimiques observées Nature de la modification
La Phosphatase alkaline sérique
Baisse
Zinc et fer
Baisse
Bilirubine totale
Augmentation
Urée Augmentation
Créatinine Augmentation
L’ensemble de ces modifications est classiquement observé pour les maladies dues à des bactéries intra-cellulaires car ces infections provoquent de façon concomitante la production de facteurs pyrogènes (TNF α) par les leucocytes et celle du médiateur leucocytaire endogène. La production du médiateur leucocytaire endogène baisse les concentrations en fer et en zinc et la production de phosphatase alcaline est dépendante de la concentration en zinc (le zinc catalyse la réaction de production de la phosphatase alcaline).
Le déficit en zinc pourrait avoir un impact négatif sur l’immunité car le taux de lymphocytes T et l’activité des macrophages seraient liés aux concentrations en zinc.
Le déficit en fer pourrait être dû à la production d’interleukine 1 par les macrophages activés.
La bilirubine totale ainsi que l’urée et la créatinine sont dues à la dégradation des hématies dans la rate et le foie.
En résumé, la réduction des concentrations en zinc pourrait avoir un impact tout à fait important dans la pathogénie de l’ehrlichiose granulocytaire et expliquer la présence de signes cliniques tels que les troubles de la fertilité et les avortements. D’autre part la diminution de la concentration en zinc pourrait avoir un impact à plus long terme avec des retards de croissance chez des individus atteints d’ehrlichiose jeunes.

41
.II.C.5 ETUDE DES LESIONS D’EHRLICHIOSE
Il y a peu de documentation sur ce sujet ; la plupart des éléments présentés dans cette partie sont issues d’une seule publication (Campbell et al., 1994).
II.C.5.a Lésions macroscopiques d’ehrlichiose
Macroscopiquement, on observe une hyperplasie progressive de l’ensemble des tissus lymphoïdes et de la rate. C’est le seul élément macroscopique évocateur de l’ehrlichiose.
On a pu cependant observer sur un chevreuil euthanasié suite à une suspicion d’ehrlichiose clinique des pétéchies sur l’endorcarde et sous la capsule de la rate.
II.C.5.b Histologie
Des lésions visibles à l’histologie sont présentes sur plusieurs organes.
Les poumons : certains auteurs (Campbell et al., 1994) décrivent une alvéolite sublobulaire précoce avec des pneumocytes et des macrophages gros et pléomorphiques . Il y a une faible infiltration par les granulocytes neutrophiles et des petits lymphocytes apparaissent accompagnés de lymphoblastes. On note également un léger œdème des alvéoles et des sacs alvéolaires avec une très léger dépôt de fibrine.
Certains sacs alvéolaires atteints sont collabés
Nœuds lymphatiques : on note une réaction des tissus réticulés et lymphoblastiques, progressive et aigüe durant les deux premières semaines post-infection.
Rate : on note une hyperplasie réactionnelle des follicules lymphoïdes et une hyperplasie générale de la rate
Reins : le mésangium apparaît hypertrophié et les cellules mésangiales apparaissent pléomorphiques
.II.D. Diagnostic de laboratoire
Comme pour toutes les maladies bactériennes, il y a deux éléments qui peuvent être détectés : il s’agit de l’agent bactérien lui-même (recherche directe) et des anticorps dirigés contre cet agent bactérien (recherche indirecte).
Nous explorerons les méthodes de recherche de l’infection par Anaplasma sur cet axe en commençant par les méthodes de détection directe.
.II.D.1 DIAGNOSTIC DIRECT D’EHRLICHIOSE
II.D.1.a La cyto-hématologie : une méthode simple et rapide
La cyto-hématologie est une méthode simple qui est effectuée par le vétérinaire lui-même. Elle présente l’intérêt d’être rapide et d’être efficace en période clinique (Vassalo, 2003).
Le principe est le suivant : une goutte de sang est étalée sur une lame et colorée avec un colorant de routine tel que May-Grünwald-Giemsa. Par la suite cet étalement est observé au microscope optique à immersion. La présence de morulae dans les granulocytes neutrophiles signe une infection à A. phagocytophilum.

42
Cette méthode demande tout de même un peu de pratique pour repérer efficacement les morulae qui ne sont pas toujours faciles à observer mais lors de la phase clinique d’infection, jusqu’à 90 % des granulocytes sont infectés ce qui rend l’observation plus facile.
Il s’agit donc de la méthode de choix lorsque le vétérinaire est face à un cas clinique suspecté d’ehrlichiose.
Cette technique n’est cependant pas assez sensible pour détecter des ehrlichioses sub-aigües ou chroniques et ne permet pas de mettre en évidence un passage antérieur d’ehrlichiose au sein d’un élevage.
II.D.1.b La PCR
La PCR est une technique beaucoup plus sensible et très efficace pour la détection d’ehrlichiose au sein d’un individu ou d’un troupeau.
Il existe plusieurs méthodes de PCR publiées pour la détection d’A. phagocytophilum. Ces méthodes ciblent chacune une portion définie de l’ADN de la bactérie.
Il existe des PCR nichées (Nested PCR) ciblant l’ADN du gène codant pour l’ARNr16s. Il existe également une PCR visant opéron Gro-ESL (Chevallier et al, 2003).
Il existe enfin une technique de PCR temps-réel visant le gène codant pour l’ARNr16S.
Il semblerait que les techniques de PCR temps-réel et nested proposées pour l’ARNR16S ne soient pas suffisamment sensibles pour détecter certaines souches d’A. phagocytophilum et que la spécificité de ces techniques ne soit pas de 100 % à cause de la détection de Ehrlichia ruminantum, Anaplasma platys, Anaplasma marginale (Drazenovich et al., 2006).
Il semblerait qu’une technique PCR visant le gène de la protéine msp2 soit plus sensible et permette la détection de souches régionales et garde une excellent spécificité. En effet, le grand nombre de copies du gène msp2 a pour effet que l’amplification est beaucoup plus facile que pour un gène avec un seul locus. C’est devenu une technique de référence pour la détection d’A. phagocytophilum.
.II.D.2 DIAGNOSTIC INDIRECT DE L’EHRLICHIOSE
L’immunofluorescence indirecte permet de détecter les anticorps anti-A. phagocytophilum. Il s’agit donc d’une technique qui ne permet pas au vétérinaire de confirmer un cas d’ehrlichiose quand il est face à un individu malade.
En revanche c’est un technique très efficace pour savoir si il y a eu passage d’ehrlichiose au sein d’un troupeau. Les IgM et IgG apparaissent dans les deux à trois semaines qui suivent l’infection et persistent longtemps (6 semaines) ce qui permet de détecter facilement un passage d’ehrlichiose (Vassalo, 2003).
Cette méthode permet surtout de situer le statut d’un troupeau vis-à-vis de l’ehrlichiose. Cela peut être intéressant de connaître le statut d’un troupeau surtout si il se situe dans une zone à risque, par exemple à la lisière d’un bois ou dans un champs entouré de haies.

43
.II.E. Données sur l’ehrlichiose chez le Chevreuil
Bien qu’il y ait peu de données sur les signes cliniques de l’ehrlichiose chez le Chevreuil, nous avons certaines données sur le comportement de la bactérie chez le Chevreuil.
A. phagocytophilum chez le Chevreuil a été dépistée dans plusieurs pays : la France (URGTV Bretgne 2003, Chimier 2005), les pays scandinaves, la slovénie, l’Ecosse et d’autres pays européens comme l’Espagne (Barandika 2007, De la Fuente 2007). Il s’agit donc d’une infection largement répandue sur l’Europe qui semble présente sur l’ensemble de l’aire de répartition du Chevreuil.
La bactérie a été retrouvée par PCR et par immunofluorescence indirecte dans l’ensemble de ces pays. La séroprévalence a souvent été très élevée chez cette espèce, autour de 70 % dans le plupart des cas (Joncour 2003, Stuen 2003).
Les premières investigations concernant A. phagocytophilum chez le Chevreuil en France ont été menées par Joncour en 2003. Cette étude a mené sur les chevreuils des côtes d’Armor et a donné pour le grand Ouest de la France une prévalence globale de 38 % d’A. phagcocytophilum lors d’une campagne de prélèvement des chevreuils à la chasse.
Chimier (2006) a passé en revue l’ensemble des études menées en Europe et montre que ce taux français est le plus élevé.
Lors d’études comparatives avec d’autres espèce sauvages (Chamois, Elan, Cerf élaphe), la séroprévalence a toujours été la plus élevée chez le Chevreuil (Stuen, 2003). Il semble donc que les rapports entre A. phagocytophilum et Chevreuil ne soient pas communs à toute la faune sauvage.
Plusieurs variants d’A. phagocytophilum ont été retrouvées chez le Chevreuil, notamment des variants qui touchent les moutons, chèvres et les chevaux. Aucune communauté de variants n’a été trouvée entre bovins et Chevreuil.
Il a été démontré que A. phagocytophilum pouvait être transmise du Chevreuil à la tique mais la persistance de la bactérie chez le Chevreuil n’a pas été mise en évidence au-delà de trois semaines. A priori, le portage à long terme de la bactérie par le Chevreuil n’a pas été démontré contrairement au cas du mouton par exemple.
En ce qui concerne l’ehrlichiose bovine, il a été montré que le Chevreuil est surtout infesté par des tiques adultes qui ne peuvent par la suite infecter un autre hôte (cf bibliographie, pas de passage trans-ovarien).
Le Chevreuil pourrait être un indicateur de présence d’A. phagocytophilum sur un territoire (Chimier, 2006).
Les cas de mortalité attribuables directement à une ehrlichiose sont rarement décrits chez le Chevreuil. On rapporte un cas en Suède (Stuen et al, 2006), et un cas en France (Chimier, 2005).
En résumé, l'expression clinique des infections à A. phagocytophilum chez le Chevreuil est mal connue. Les cas de mortalité liés directement à l'infection sont certainement sous-diagnostiqués. Compte-tenu du fait que l'effet pathogène est essentiellement indirect, via des mécanismes peu spécifiques, l'incrimination de la bactérie dans les mortalités groupées observées sur le terrain ne peut se raisonner que sur la base d'une étude épidémiologique cas-témoins, où l'exposition à A. phagocytophilum est un facteur de risque. Les résultats seront ensuite discutés à la lumière de l'analyse de facteurs environnementaux, tels qu'ils ressortent de données de suivi des populations et de leurs habitats, collectées depuis 1995 dans cette région.

44

45
Deuxième partie : études expérimentale et rétrospective
.I. Cadre et objectifs de cette étude
.I.A. Cadre de l’étude
Cette étude a été proposée par la Fédération Régionale des Chasseurs de Franche-Comté (FRCFC) et par les Fédérations Départementales des Chasseurs du Doubs (FDC25) du Jura (FDC39), de Haute Saône (FDC70) et du territoire de Belfort (FDC90) à la Fédération Nationale des Chasseurs (FNC). Guy Joncour (vétérinaire praticien en Bretagne) avait lancé l’idée que A. phagocytophilum pouvait provoquer une maladie chez le Chevreuil et durant l’été 2007, trois chevreuils retrouvés morts en Franche-Comté étaient porteurs de la bactérie (positifs en PCR sang ou rate). Ce sont ces trois découvertes qui ont déclenché la demande des Fédérations Régionale et Départementales des Chasseurs auprès de la Fédération Nationale des Chasseurs. Cette proposition a été retenue parce que le milieu de la chasse est très actif en Franche-Comté et que cela permettait de mobiliser de nombreux chasseurs pour cette étude. La région France Comté effectue par ailleurs un suivi efficace de la dynamique des populations de Chevreuils depuis plusieurs années qui pouvait être intégré à cette étude.
.I.B. Objectifs
L’objectif principal de cette étude était d’incriminer l'infection par A. phagocytophilum dans la survenue de morbidité/mortalité au sein d’une population de Chevreuils sur une saison donnée.
Est-ce que la bactérie A. phagocytophilum peut être tenue responsable de maladies chez le Chevreuil ?
.II. Etude expérimentale : Etude du rôle pathogène d’A. phagocytophilum dans la population de Chevreuils de Franche-Comté sur la saison de chasse 2007/2008
.II.A. Rappel des objectifs
L’objectif principal de cette étude était de savoir si l'infection par A. phagocytophilum peut être incriminée dans la survenue de maladies chez le Chevreuil.
Est-ce que la bactérie A. phagocytophilum provoque une maladie chez le Chevreuil ?
Quelles sont les manifestations cliniques d’une ehrlichiose granulocytaire chez le Chevreuil ?
Quelles sont les lésions consécutives à une ehrlichiose granulocytaire chez le Chevreuil ?

46
.II.B. Matériel et méthodes de l’étude expérimentale
Ain de déterminer si A. phagocytophilum est pathogène ou non pour le Chevreuil, il y a deux systèmes d’étude possibles.
Le premier système d’études est l’inoculation de la bactérie à des chevreuils naïfs de différentes classes en enclos. Ce système permet une observations rapprochées après inoculation. Il permet également de suivre des paramètre biologiques éventuellement intéressants pour l’étude.
Le Chevreuil supporte mal des conditions de captivité et c’est une espèce qui se manipule très difficilement. Outre d’éventuels problèmes de mortalité liés au stress, la mesure de paramètres biologiques pourrait être modifiée par le stress. Enfin, l'inoculation d'agents pathogènes à des animaux en semi-liberté (en parc) pose des problèmes de biosécurité pour le reste de la faune.
Cette méthode a été exclue d’emblée.
La deuxième méthode consiste à effectuer une étude comparative entre des individus malades et des individus sains au sein d’une population sauvage. La l'association statistique entre maladie et infection permet de suspecter un rôle pathogène de l’agent . Elle s'applique à une une population in situ ,en veillant à ce que les lots d'animaux comparés ne diffèrent pas par d'autres facteurs qui pourraient influer sur la maladie.
.II.B.1 LA FRANCHE-COMTE COMME SITE D’ETUDE
La Franche-Comté s’est avérée un bon site d’étude puisque tout d’abord l’ensemble des acteurs potentiels en Franche-Comté était demandeur et motivé pour le projet, que ce soit les techniciens de fédérations départementales des chasseurs, de la fédération régionale des chasseurs ou les laboratoires.
De plus, la Franche-Comté est une région où les plans de chasses sont élevés ce qui devait rendre plus facile le prélèvement des chevreuils sains, mais aussi des chevreuils moribonds (en première analyse, plus il y a de chevreuils, plus on voit de malades).
C’est également une région pionnière en matière de suivi des populations puisque sur l’ensemble des quatre départements ont été mis en place des sites dits équilibre faune-flore ou le suivi des populations de chevreuils est effectué depuis plusieurs années avec des indicateurs de changement écologiques validés scientifiquement et appliqués dans le cadre prescrit.
Le travail effectué en Master par Anne-Sophie Hesler en 2006 a permis de définir des unités biogéographiques de populations de chevreuils sur la région Franche-Comté. Chaque unité biogéographique présente l’avantage de représenter une population de chevreuils qui évolue sur un milieu homogène, dans des conditions climatiques homogènes (les conditions climatiques sont variables en Franche-Comté parce que les altitudes sont très variées) et qui est séparé des autres populations par des barrières géographiques (anthropiques ou naturelles).
.II.B.2 ECHANTILLONNAGE
II.B.2.a Les chevreuils prélevés
Les prélèvements ont eu lieu sur l’ensemble de la région Franche-Comté de la fin Septembre 2007 au 20 mai 2008. L’objectif de cette saison de prélèvement était de récupérer des chevreuils sains durant la saison de chasse 2007/2008 et des chevreuils trouvés malades ou morts depuis peu durant cette saison de chasse et jusqu’au 20 mai 2008.
Devant la difficulté de procéder à la récupération de matériel biologique de chevreuils sains avec un prélèvement homogène sur l’ensemble de la région, il a été décidé (comme pour la plupart des études sur la faune sauvage chassable) que les chevreuils sains seraient des chevreuils tués à la

47
chasse et prélevés juste après la mort. Ceci permettait de contrôler les prélèvements et d’avoir des prélèvements géographiquement homogènes mais également de pouvoir contrôler le nombre d’individus prélevés par classe prévue.
Nous avons décidé d’apparier les cas et les témoins sur la localisation géographique et sur l’âge car des observations de terrain semblaient refléter une différence entre les statuts des chevreuils adultes et les jeunes chevreuils (moins de un an) face au portage d’A. phagocytophilum (Joncour, communication personnelle).
Les chevreuils malades ou morts récemment étaient tous ramenés aux fédérations départementales par les personnes qui les trouvaient. Les chevreuils malades étaient abattus avant d’être rapatriés ce qui impliquait soit une autorisation préfectorale de tir exceptionnelle (Doubs) soit une autorisation de tir individuelle délivrée par les agents de l’Office National des Forêts.
II.B.2.b Nombre de chevreuils à prélever
Ces deux classes d’individus devaient être séparées appariées sur la localisation géographique et sur l’âge (jeune de moins de 1 an , adulte de plus d’un an).
Les objectifs de prélèvement étaient de 50 individus par classe soit
• 50 chevreuils sains tirés à la chasse et jeunes (moins de un an)
• 50 chevreuils sains tirés à la chasse et adultes (plus de un an)
• 50 chevreuils moribonds ou morts et jeunes (moins de un an)
• 50 chevreuils moribonds ou morts et adultes (plus de un an)
Lors de l’établissement du protocole, il a été décidé de chercher à avoir 105 individus sains et 105 malades afin d’être sûr d’en avoir au moins 100 par classe en cas de problèmes liés aux prélèvements ou à leur acheminement.
La décision du nombre d’individus à prélever par classe a été faite sur la base d’une marge d’erreur minimale pour la prévalence et la séroprévalence et pour pouvoir comparer les résultats entre les quatre classe sans problèmes statistiques. Ces chiffres ont été choisis sur la base de la prévalence observée durant l’étude de l’URGTV Bretagne (Joncour, 2003) qui était d’environ 70% chez les chevreuils.
L’autre impératif était la faisabilité : pour les chevreuils tirés à la chasse il n’y avait pas de problème pour augmenter le nombre de prélèvements mais le nombre de chevreuils malades était difficilement augmentable. Le nombre de chevreuils à prélever pour chaque département et chaque classe est indiqué dans le Tableau 9.
Tableau 9: Nombre de chevreuils à prélever par classe et par département
FDC 25 FDC 39 FDC 70 FDC 90
Sains
(Tués à la chasse)
48
(8 sites x 6)
36
(6 sites x 6)
24
(4 sites x 6)
12
(2 sites x 6)
Moribonds 30 30 30 15
La distinction des âges a été faite avec les indicateurs donnés dans l’étude bibliographique soit la conformations globale de l’individu et la dentition.
Ces prélèvements se faisaient sous la surveillance des techniciens des Fédérations Départementales des Chasseurs des quatre départements de la région Franche-Comté : Le Doubs, Le Jura, La Haute Saône et le Territoire de Belfort.
Plusieurs éléments étaient récupérés pour chaque chevreuil.

48
Les chevreuils sains étaient prélevés de préférence sur, ou à proximité des sites Equilibre-Faune-Flore afin de coupler les données d’analyse des chevreuils et celles du suivi des populations.
La figure 11 montre la localisation des différents sites Equilibre-Faune-Flore. Ceux-ci sont disposés de façon homogène sur l’ensemble de la Franche-Comté offrant ainsi un maillage qui couvre l’ensemble des différents écotypes de la région.
Figure 11: Répartition des sites équilibre Faune-Flore sur la région Franche-Comté
.II.C. Nature des prélèvements
Tout d’abord, une fiche commémorative était remplie renseignant la date et le lieu de prélèvement, le préleveur ainsi que l’âge (renseigné par les dents), le sexe, la conformation du chevreuil (bon état, amaigri). Une fiche type est inscrite en annexe 1.
Deux tubes de sang étaient remplis, un tube sec pour l’immunofluorescence et un tube EDTA pour la PCR (le choix des analyses est discuté ultérieurement). Le sang était prélevé le plus vite possible soit à la veine jugulaire soit dans le caillot cardiaque si le prélèvement à la veine jugulaire était impossible. Ces prélèvements étaient effectués par les chasseurs pour les individus tués à la chasse et par les techniciens de fédérations pour les individus malades.
La rate était prélevée et congelée. Le choix de prélever la rate était motivé par le rôle de cet organe qui filtre le sang et de ce fait à un rôle potentiellement important dans la protection contre l’ehrlichiose. De plus, il a été trouvé des cas de chevreuils positifs sur rate alors qu’ils avaient été négatifs sur sang (G. Joncour, communication personnelle). Ainsi la rate pouvait permettre de détecter des chevreuils apparemment négatifs sur sang mais porteurs d’A. phagocytophilum dans
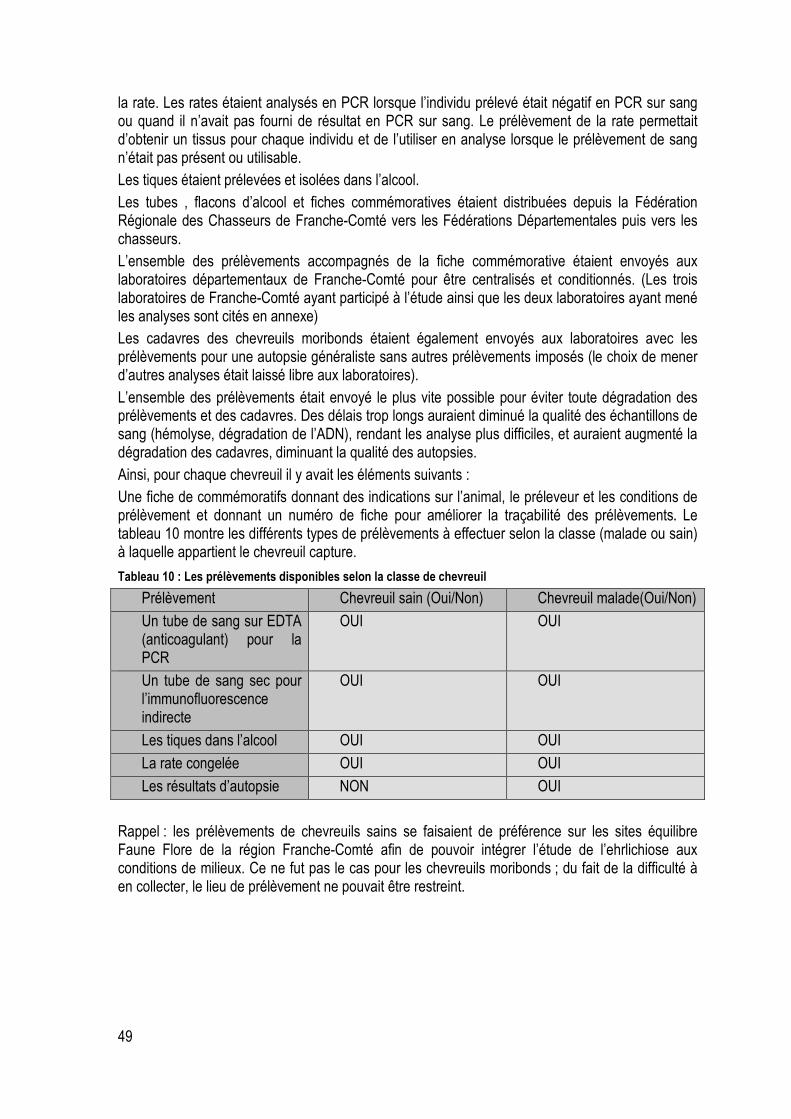
49
la rate. Les rates étaient analysés en PCR lorsque l’individu prélevé était négatif en PCR sur sang ou quand il n’avait pas fourni de résultat en PCR sur sang. Le prélèvement de la rate permettait d’obtenir un tissus pour chaque individu et de l’utiliser en analyse lorsque le prélèvement de sang n’était pas présent ou utilisable.
Les tiques étaient prélevées et isolées dans l’alcool.
Les tubes , flacons d’alcool et fiches commémoratives étaient distribuées depuis la Fédération Régionale des Chasseurs de Franche-Comté vers les Fédérations Départementales puis vers les chasseurs.
L’ensemble des prélèvements accompagnés de la fiche commémorative étaient envoyés aux laboratoires départementaux de Franche-Comté pour être centralisés et conditionnés. (Les trois laboratoires de Franche-Comté ayant participé à l’étude ainsi que les deux laboratoires ayant mené les analyses sont cités en annexe)
Les cadavres des chevreuils moribonds étaient également envoyés aux laboratoires avec les prélèvements pour une autopsie généraliste sans autres prélèvements imposés (le choix de mener d’autres analyses était laissé libre aux laboratoires).
L’ensemble des prélèvements était envoyé le plus vite possible pour éviter toute dégradation des prélèvements et des cadavres. Des délais trop longs auraient diminué la qualité des échantillons de sang (hémolyse, dégradation de l’ADN), rendant les analyse plus difficiles, et auraient augmenté la dégradation des cadavres, diminuant la qualité des autopsies.
Ainsi, pour chaque chevreuil il y avait les éléments suivants :
Une fiche de commémoratifs donnant des indications sur l’animal, le préleveur et les conditions de prélèvement et donnant un numéro de fiche pour améliorer la traçabilité des prélèvements. Le tableau 10 montre les différents types de prélèvements à effectuer selon la classe (malade ou sain) à laquelle appartient le chevreuil capture.
Tableau 10 : Les prélèvements disponibles selon la classe de chevreuil
Prélèvement Chevreuil sain (Oui/Non) Chevreuil malade(Oui/Non)
Un tube de sang sur EDTA (anticoagulant) pour la PCR
OUI OUI
Un tube de sang sec pour l’immunofluorescence indirecte
OUI OUI
Les tiques dans l’alcool OUI OUI
La rate congelée OUI OUI
Les résultats d’autopsie NON OUI
Rappel : les prélèvements de chevreuils sains se faisaient de préférence sur les sites équilibre Faune Flore de la région Franche-Comté afin de pouvoir intégrer l’étude de l’ehrlichiose aux conditions de milieux. Ce ne fut pas le cas pour les chevreuils moribonds ; du fait de la difficulté à en collecter, le lieu de prélèvement ne pouvait être restreint.

50
.II.C.1 LA PCR COMME METHODE DE DETECTION DIRECTE D’A. PHAGOCYTOPHILUM
Cette étude visait à rechercher à la fois la présence d’une réponse immunitaire chez chaque individu et la présence de la bactérie dans le sang des chevreuils. Il fallait donc une méthode pour détecter les anticorps anti-A. phagocytophilum et une méthode pour détecter A. phagocytophilum.
Les PCR sur sang ont été effectuées au laboratoire de Sèvres-Atlantiques (79)
II.C.1.a Utilisation du kit ADIAVET® ANA PHA REALTIME comme technique de PCR temps-réel
C’est une technique de PCR temps-réel qui a été utilisée pour la recherche directe d’A. phagocytophilum car c’était le test qui présentait les meilleures qualités pour ce travail.
Un seul kit est actuellement commercialisé en France
ADIAVET® ANA PHA REALTIME
La PCR (Polymérase Chain Reaction) est une technique qui permet de détecter des concentrations très faibles d’ADN au sein d’un mélange en amplifiant des portions de cet ADN. Le principe de la PCR temps-réel est d’amplifier une portion d’ADN grâce à des amorces spécifiques et de suivre l'accroissement de la quantité amplifiée au cours des cycles d’amplification grâce à des fluorochromes qui deviennent fluorescents lorsqu’ils sont intégrés à des portions amplifiées. Le franchissement d’un seuil d’intensité de fluorescence indique la présence de la séquence recherchée et donc la positivité de l’échantillon pour la bactérie recherchée. La positivité du test est signée par l’allure sigmoïde de la courbe de fluorescence donnée et par l’inflexion de la courbe.
Ici il s’agissait d’une PCR nichée qui consiste à amplifier une première séquence large puis à amplifier une plus petite séquence au sein de cette séquence large. Cette technique permet une meilleure spécificité de la PCR et une meilleure sensibilité puisqu’il y a une double amplification.
II.C.1.b Qualités du test ADIAVET® ANA PHA REALTIME
Pour ce test, ce sont des amorces provenant du laboratoire ADIAGENE qui ont été utilisées, permettant une nested-PCR du gène 1655rRNA codant pour un fragment de 913 paires de bases.
Il n'y a pas de réactions croisées pour Borrelia burgdorferi, Borrelia afzelli, Borrelia garinii, Coxiella burnetii, Ehrlichia canis et Rickettsia conorii ainsi que pour Anaplasma marginale.(communication personnelle Adiagène)
Un témoin interne d’amplification permet d’utiliser ce test en routine et de mettre en évidence d’éventuelles erreurs dans le protocole.
La sensibilité de cette technique est excellente par rapport aux autre techniques existantes. Hors des autres techniques de PCR, la seule technique de mise en évidence d’A. phagocytophilum est la cyto-hématologie qui ne permet de mettre en évidence la bactérie que lors d’épisodes cliniques.
II.C.1.c Utilisation du kit ADIAVET ANA PHA REALTIME
Les échantillons ont été décongelés dans un premier temps. L’ADN des échantillons a ensuite été extrait grâce à l’utilisation d’un kit d’extraction. Le kit utilisé était un kit QIAGEN, QIAamp® DNA mini kit.
L’ADN était donc extrait puis purifié en utilisant ce kit puis la PCR était lancée en utilisant le kit ADIAVET® ANA PHA REALTIME avec les recommandations l’accompagnant.

51
II.C.1.d Utilisation du kit ADIAVET ANA PHA REALTIME en PCR semi-quantitative
L’entreprise ADIAGENE a mis à disposition du laboratoire départemental des Deux-Sèvres un plasmide contenant la séquence ARNr16S détectée par le kit ANA PHA REALTIME.
Le plasmide est distribué en échantillon de concentrations connues et est utilisé comme témoin quantitatif lors des étapes de thermocyclage. En effet, le nombre de cycles qu’il faut à un échantillon pour franchir le seuil de positivité est corrélé à la quantité d’ADN dans le mélange.
Utilisé en plusieurs dilutions, le plasmide permet de donner une valeur en quantité d’ADN pour la durée pour atteindre le seuil de positivité.
On a donc une indication sur la quantité d’ADN d’A. phagocytophilum dans les échantillons mais pas la quantité exacte car les conditions d’accession à un plasmide ne sont pas les mêmes que celles d’un ADN bactérien pour les enzymes utilisés lors d’une PCR. On n’a donc pas les mêmes quantités d’ADN bactérien et de plasmide pour un même cycle seuil.
Les résultats obtenus ont donc été interprétés de façon relative les uns par rapport aux autres. Ainsi les expressions retenues sont les suivantes : par rapport aux autre prélèvements, un prélèvement est négatif, faiblement positif, moyennement positif ou fortement positif. Le choix du nombre de cycles seuils est discuté plus loin.
Un problème supplémentaire se pose pour l’interprétation de ces résultats, au-delà de trente cycles d’amplification, la répétabilité du test diminue très vite ce qui n’influe pas sur le résultat qualitatif (positif ou négatif) mais qui peut gêner l’interprétation semi-quantitative.
La cyto-hématologie aurait pu être utilisée pour détecter la présence de la bactérie dans le sang. Cela n’a pas été effectué parce que c’est une technique peu sensible et qu’il n’était pas certain que les prélèvements obtenus sur les chevreuils seraient de suffisamment bonne qualité pour permettre une lecture de qualité des lames lors de cet examen.
Cette technique aurait également pu être utilisée pour effectuer une numération formule sur chaque prélèvement. Cela n’a pas été effectué non plus car il n’y a pas de données de références pour la numération formule du chevreuil ce qui n’aurait pas permis de commenter les résultats obtenus. Ici aussi la qualité des prélèvements aurait pu poser problème (la lyse des cellules dans l’échantillon n’aurait pas permis d’obtenir une numération exacte).
.II.C.2 DIAGNOSTIC INDIRECT DE L’EHRLICHIOSE : L’IMMUNOFLUORESCENCE INDIRECTE
Les analyses d’Immunofluorescence indirecte ont été menées au laboratoire départemental des Côtes d’Armor (22)
Les immunofluorescences étaient effectuées sur des lames contenant des granulocytes de cheval infectés.
La sensibilité de ce test est difficile à établir car il n’existe pas de test de référence auquel comparer cette technique. Ce test est néanmoins positif jusqu’à une dilution au 1/32è.
La spécificité de ce test souffre également de l’absence de test de référence. On sait qu’il peut y avoir des réactions croisées avec Anaplasma marginale mais cette bactérie n’est pas présente en France Métropolitaine (anonyme, 2005).
.II.C.3 ANALYSE DES DONNEES DE SUIVI SUR LES POPULATIONS DE CHEVREUILS
Aux données collectées sur les chevreuils, nous avons pu ajouter des données sur les populations de chevreuils. La fédération régionale des chasseurs de Franche-Comté a gracieusement mis à notre disposition les données sur la santé des populations des chevreuils : les indicateurs de santé

52
des population présentés dans l’étude bibliographique de la situation du Chevreuil en Franche-Comté.
Ces études ont été menées sur les sites Equilibre-Faune-flore et ont commencé entre 2002 et 2006 selon le site concerné. Nous avons pu intégrer ces données aux résultats d’analyse obtenus sur les chevreuils.
Les autres données portent sur les entités biogéographiques de populations de chevreuils. Un travail mené en 2005 sur l’ensemble de la Franche-Comté a permis de définir des populations de chevreuils précises, séparées entre elles par des obstacles naturels ou humains et qui partagent les mêmes caractéristiques intrinsèques et les mêmes milieux (Hesler, 2006).
Par ailleurs, cette analyse des données a été accompagnée d’une entrevue avec au moins un technicien de chaque fédération départementale des chasseurs de Franche-Comté et de la visite de sites Equilibre Faune-Flore représentatifs des différents milieux existant en Franche-Comté.
.II.C.4 TRAITEMENT DES DONNEES
L’ensemble des données sur les individus, prélèvements, commémoratifs, autopsies, ont été rentrés dans un tableau de données sous le logiciel EXCEL (Microsoft Office 2007).
Par la suite l’exploitation statistique des données a été effectuée avec le logiciel R 2.6.1.
Les test utilisés sont les suivants : Test de Fischer, calcul de fréquence et d’écart-type.
Nous avons utilisé le logiciel ARCVIEW (ARCGis 9.2 ) qui est un logiciel de Système d’Information Géographique pour saisir manuellement la localisation (lieu-dit, commune) des cas et des témoins et analyser visuellement leur distribution spatiale, notamment la proximité avec les sites équilibre faune-flore.
.III. Résultats
.III.A.1 REALISATIONS, NOMBRE DE CHEVREUILS PRELEVES
Au cours de l’étude, 168 chevreuils ont été prélevés au total dont 114 chevreuils sains (témoins) et 36 chevreuils malades (cas).
Sur ces 168 chevreuils, 18 chevreuils ont été exclus de l’étude parce que les délais entre prélèvement et arrivée au laboratoire. Ces 18 chevreuils étaient des chevreuils sains prélevés en début de saison de chasse ( septembre 2008).
Le tableau 11 montre les résultats de la campagne de prélèvement 2007/2008.
Tableau 11: Nombre de chevreuils prélevés et inclus lors de la saison de prélèvement 2007/2008
Campagne de prélèvements 2007/2008 Total prélevés
Nombre total de chevreuils prélevés 168
Nombre de chevreuils inclus à l’étude 150 (18 chevreuils exclus, délais d’acheminement au laboratoire trop longs)
Chevreuils témoins(abattus à la chasse) 114
Chevreuils cas (retrouvés malades ou morts) 36
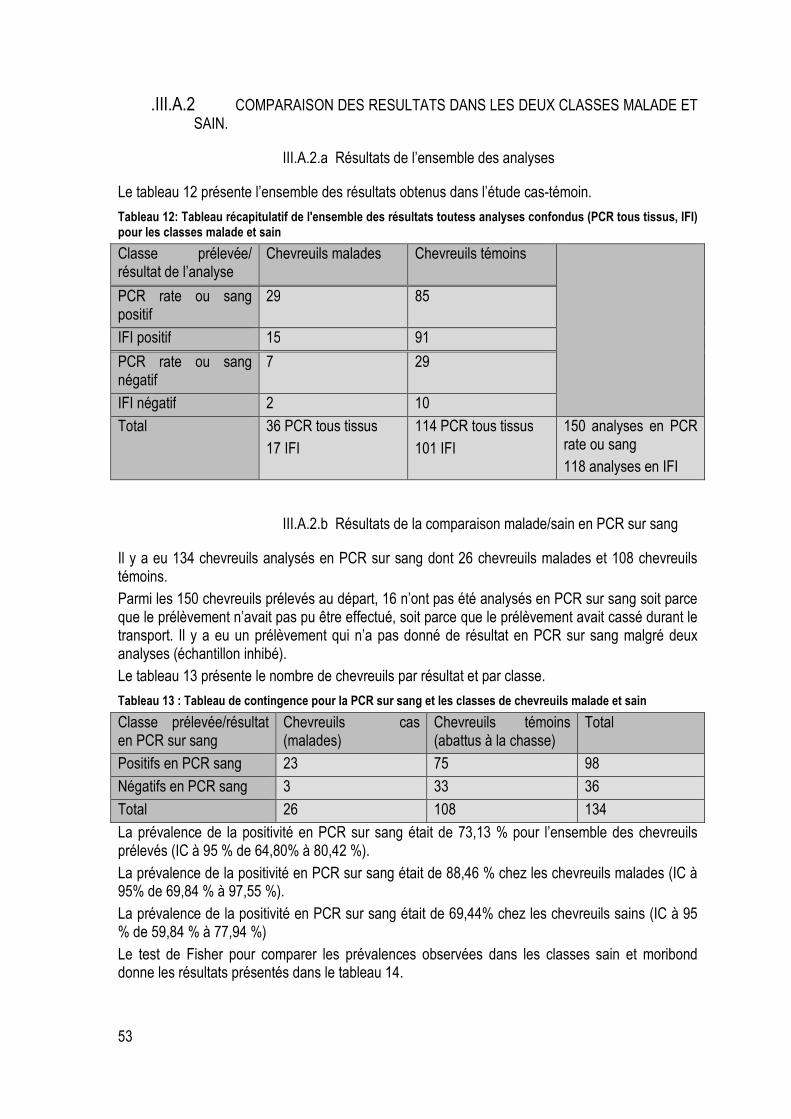
53
.III.A.2 COMPARAISON DES RESULTATS DANS LES DEUX CLASSES MALADE ET SAIN.
III.A.2.a Résultats de l’ensemble des analyses
Le tableau 12 présente l’ensemble des résultats obtenus dans l’étude cas-témoin.
Tableau 12: Tableau récapitulatif de l'ensemble des résultats toutess analyses confondus (PCR tous tissus, IFI) pour les classes malade et sain
Classe prélevée/ résultat de l’analyse
Chevreuils malades Chevreuils témoins
PCR rate ou sang positif
29 85
IFI positif 15 91
PCR rate ou sang négatif
7 29
IFI négatif 2 10
Total 36 PCR tous tissus
17 IFI
114 PCR tous tissus
101 IFI
150 analyses en PCR rate ou sang
118 analyses en IFI
III.A.2.b Résultats de la comparaison malade/sain en PCR sur sang
Il y a eu 134 chevreuils analysés en PCR sur sang dont 26 chevreuils malades et 108 chevreuils témoins.
Parmi les 150 chevreuils prélevés au départ, 16 n’ont pas été analysés en PCR sur sang soit parce que le prélèvement n’avait pas pu être effectué, soit parce que le prélèvement avait cassé durant le transport. Il y a eu un prélèvement qui n’a pas donné de résultat en PCR sur sang malgré deux analyses (échantillon inhibé).
Le tableau 13 présente le nombre de chevreuils par résultat et par classe.
Tableau 13 : Tableau de contingence pour la PCR sur sang et les classes de chevreuils malade et sain
Classe prélevée/résultat en PCR sur sang
Chevreuils cas (malades)
Chevreuils témoins (abattus à la chasse)
Total
Positifs en PCR sang 23 75 98
Négatifs en PCR sang 3 33 36
Total 26 108 134
La prévalence de la positivité en PCR sur sang était de 73,13 % pour l’ensemble des chevreuils prélevés (IC à 95 % de 64,80% à 80,42 %).
La prévalence de la positivité en PCR sur sang était de 88,46 % chez les chevreuils malades (IC à 95% de 69,84 % à 97,55 %).
La prévalence de la positivité en PCR sur sang était de 69,44% chez les chevreuils sains (IC à 95 % de 59,84 % à 77,94 %)
Le test de Fisher pour comparer les prévalences observées dans les classes sain et moribond donne les résultats présentés dans le tableau 14.

54
Tableau 14: Résultats du test de Fisher de comparaison malade/sain des prévalences en PCR sur sang
Fisher's Exact Test for Count Data
p-value = 0.0526
alternative hypothesis: true odds ratio is not equal to 1
Intervalle de confiance à 95 % :
0.9139453 18.6182849
sample estimates:
Odds ratio : 3.348334
La différence observée entre les deux classes n’est pas significative au seuil de 5 % puisque la p-value est supérieure à ce seuil.
On observe une différence statistiquement significative pour le caractère positif en PCR sur sang entre chevreuils malades et chevreuils sains au seuil de 5 %.
L’Odds ratio est de 3,35 avec un Intervalle de confiance à 95% de 0,91 à 18,61.
III.A.2.c Résultats de la comparaison malade/sain en PCR tous tissus confondus
Le tableau 15 montre le nombre de chevreuils obtenu pour chaque classe et chaque résultat
Tableau 15: Tableau de contingence pour les PCR sur sang et rate et les classes de chevreuils malade et sain
Classe prélevée/résultat en PCR sur sang ou rate
Chevreuils cas (malades)
Chevreuils témoins (abattus à la chasse)
Total
Positifs en PCR sang ou rate
29 85 114
Négatifs en PCR sang ou rate
7 29 36
Total 36 114 150
La prévalence totale pour la positivité en PCR sang ou rate est de 76,51 % pour l’ensemble des chevreuils prélevés (IC à 95% de 68,88 % à 83,06 %).
La prévalence totale pour la positivité en PCR sang ou rate est de 80,56 % pour les chevreuils malades (IC à 95 % de 63,98 % à 91,81 %).
La prévalence totale pour la positivité en PCR sang ou rate est de 74,56 % pour les chevreuils sains (IC à 95 % de 65,55 % à 82,25 %).
Le test de Fisher permettant de comparer les prévalences observées dans les deux classes donne le résultat présenté dans le tableau 16.

55
Tableau 16: Résultats du test de Fisher de comparaison malades/sain des prévalences en PCR tous tissus
Fisher's Exact Test for Count Data
p-value = 0.5115
alternative hypothesis: true odds ratio is not equal to 1
Intervalle de confiance à 95 %
0.5287736 4.2291520
sample estimates:
odds ratio
1.410341
Le test n’a pas permis de mettre en évidence une différence pour le caractère positif en PCR sang ou rate entre chevreuils malades et chevreuils sains au seuil de 5%.
Il y a eu huit chevreuils sains qui se sont avérés positifs sur rate alors qu’ils étaient négatifs sur sang, deux chevreuils sains qui se sont avérés positifs sur rate sans avoir été testé sur sang et six chevreuils malades positifs sur rate sans avoir été testé sur sang.
Il y a eu 4 chevreuils sains négatifs et 4 chevreuils malades négatifs parmi les chevreuils qui n’avaient pas été testé en PCR sur sang,
Parmi les chevreuils négatifs en PCR, 19 chevreuils sains étaient négatifs en PCR sur rate et3 chevreuils malades étaient négatifs également.
III.A.2.d Résultats de la comparaison malade/sain en immuno-fluorescence indirecte
Le tableau 17 montre le nombre de chevreuils obtenu pour chaque classe et chaque résultat.
Tableau 17; Tableau de contingence pour l'IFI et pour les classes malade et sain
Classe prélevée/résultat en IFI
Chevreuils cas (malades)
Chevreuils témoins (abattus à la chasse)
Total
Positifs en IFI 15 91 106
Négatifs en IFI 2 10 12
Total 17 101 118
La prévalence pour la positivité en IFI est de 89,83 % pour l’ensemble des chevreuils prélevés (IC à 95 % de 82,91 % à 94,63 %).
La prévalence pour la positivité en IFI est de 88,24 % pour les chevreuils malades (IC à 95 % de 63,56 % à 98,54 %).
La prévalence pour la positivité en IFI est de 90,10 % pour les chevreuils sains (IC à 95 % de 82,55 % à 95,15 %).
Le test de Fisher permettant de comparer les prévalences observées dans les deux classes sain et malades donne le résultat présenté dans le tableau 18.
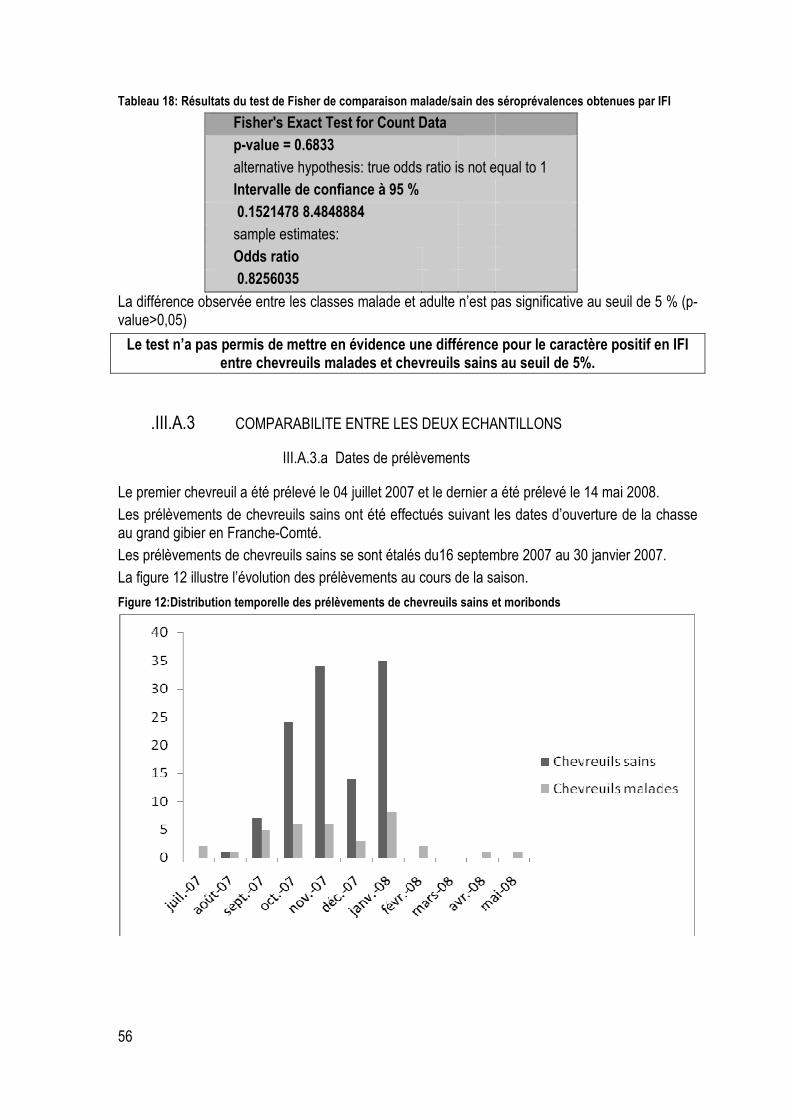
56
Tableau 18: Résultats du test de Fisher de comparaison malade/sain des séroprévalences obtenues par IFI
Fisher's Exact Test for Count Data
p-value = 0.6833
alternative hypothesis: true odds ratio is not equal to 1
Intervalle de confiance à 95 %
0.1521478 8.4848884
sample estimates:
Odds ratio
0.8256035
La différence observée entre les classes malade et adulte n’est pas significative au seuil de 5 % (p-value>0,05)
Le test n’a pas permis de mettre en évidence une différence pour le caractère positif en IFI entre chevreuils malades et chevreuils sains au seuil de 5%.
.III.A.3 COMPARABILITE ENTRE LES DEUX ECHANTILLONS
III.A.3.a Dates de prélèvements
Le premier chevreuil a été prélevé le 04 juillet 2007 et le dernier a été prélevé le 14 mai 2008.
Les prélèvements de chevreuils sains ont été effectués suivant les dates d’ouverture de la chasse au grand gibier en Franche-Comté.
Les prélèvements de chevreuils sains se sont étalés du16 septembre 2007 au 30 janvier 2007.
La figure 12 illustre l’évolution des prélèvements au cours de la saison.
Figure 12:Distribution temporelle des prélèvements de chevreuils sains et moribonds
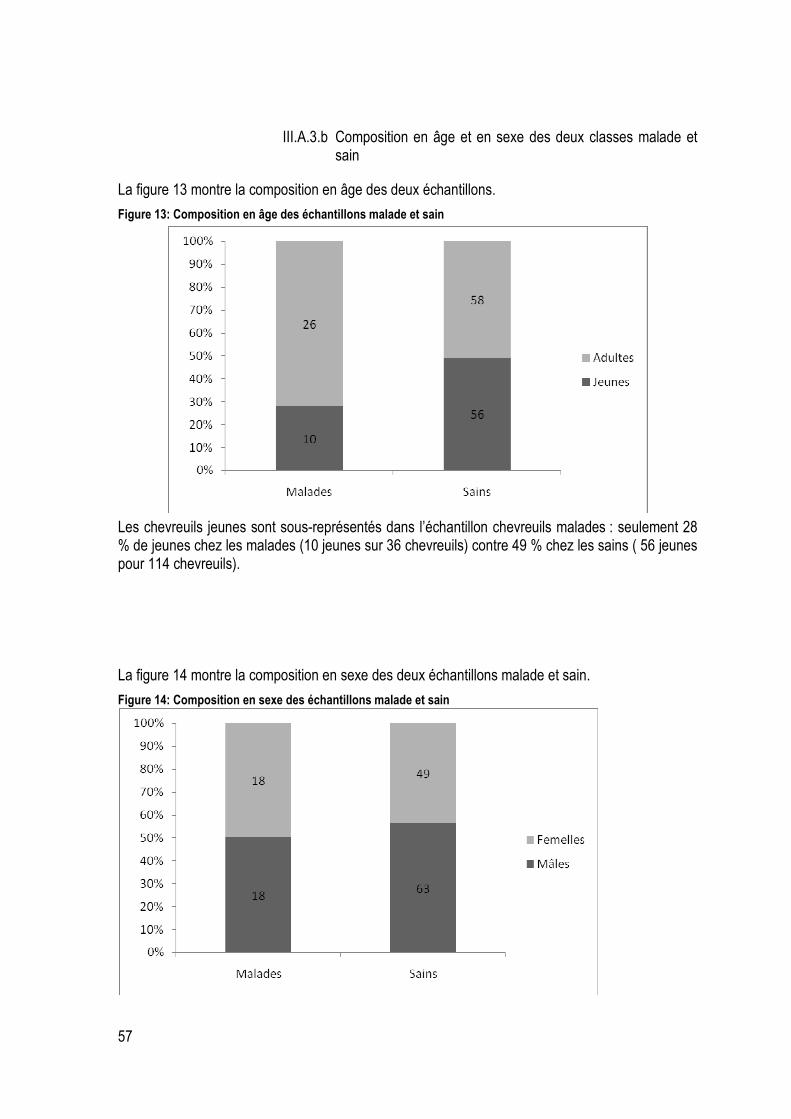
57
III.A.3.b Composition en âge et en sexe des deux classes malade et sain
La figure 13 montre la composition en âge des deux échantillons.
Figure 13: Composition en âge des échantillons malade et sain
Les chevreuils jeunes sont sous-représentés dans l’échantillon chevreuils malades : seulement 28 % de jeunes chez les malades (10 jeunes sur 36 chevreuils) contre 49 % chez les sains ( 56 jeunes pour 114 chevreuils).
La figure 14 montre la composition en sexe des deux échantillons malade et sain.
Figure 14: Composition en sexe des échantillons malade et sain

58
La répartition des sexes est homogène entre les deux échantillons. Il y a 50% de chaque sexe chez les malades (18 chevreuils pour chaque sexe) et 43, 75 % de femelles dans l’échantillon sains (49 femelles pour 112 chevreuils).
Il y a deux chevreuils sains (un jeune et un adulte) pour lesquels le sexe n’a pas été renseigné.
III.A.3.c Lieux de prélèvement
Le traitement des données de prélèvements par système d’information géographique (ArcView) a donné les figure 15 qui montre la répartition des prélèvements des chevreuils sains et malades entre eux et par rapport aux sites Equilibre Faune Flore.
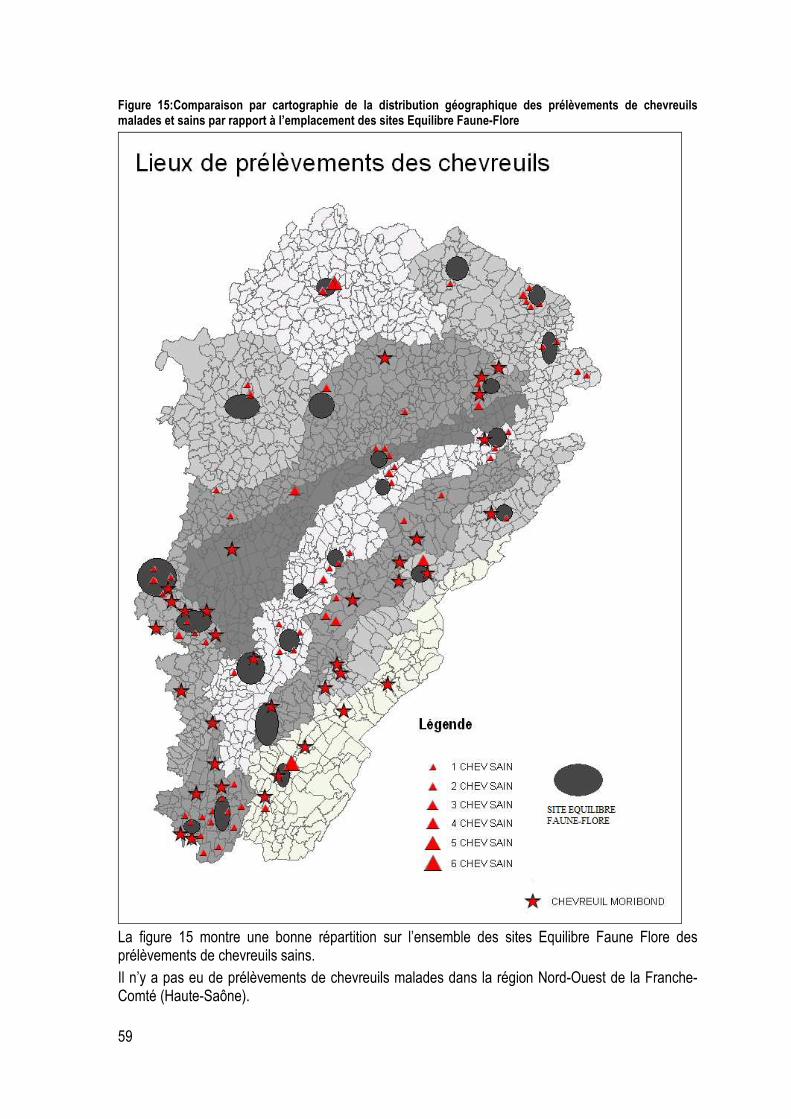
59
Figure 15:Comparaison par cartographie de la distribution géographique des prélèvements de chevreuils malades et sains par rapport à l’emplacement des sites Equilibre Faune-Flore
La figure 15 montre une bonne répartition sur l’ensemble des sites Equilibre Faune Flore des prélèvements de chevreuils sains.
Il n’y a pas eu de prélèvements de chevreuils malades dans la région Nord-Ouest de la Franche-Comté (Haute-Saône).
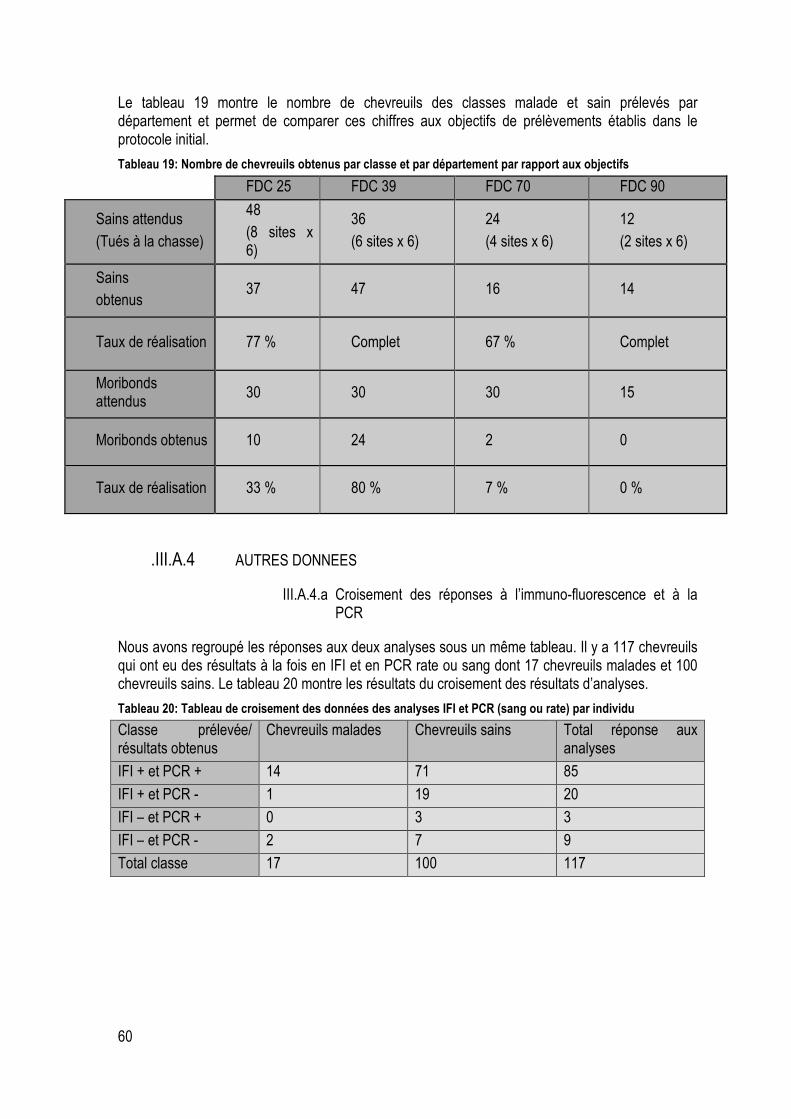
60
Le tableau 19 montre le nombre de chevreuils des classes malade et sain prélevés par département et permet de comparer ces chiffres aux objectifs de prélèvements établis dans le protocole initial.
Tableau 19: Nombre de chevreuils obtenus par classe et par département par rapport aux objectifs
FDC 25 FDC 39 FDC 70 FDC 90
Sains attendus
(Tués à la chasse)
48
(8 sites x 6)
36
(6 sites x 6)
24
(4 sites x 6)
12
(2 sites x 6)
Sains
obtenus 37 47 16 14
Taux de réalisation 77 % Complet 67 % Complet
Moribonds attendus
30 30 30 15
Moribonds obtenus 10 24 2 0
Taux de réalisation 33 % 80 % 7 % 0 %
.III.A.4 AUTRES DONNEES
III.A.4.a Croisement des réponses à l’immuno-fluorescence et à la PCR
Nous avons regroupé les réponses aux deux analyses sous un même tableau. Il y a 117 chevreuils qui ont eu des résultats à la fois en IFI et en PCR rate ou sang dont 17 chevreuils malades et 100 chevreuils sains. Le tableau 20 montre les résultats du croisement des résultats d’analyses.
Tableau 20: Tableau de croisement des données des analyses IFI et PCR (sang ou rate) par individu
Classe prélevée/ résultats obtenus
Chevreuils malades Chevreuils sains Total réponse aux analyses
IFI + et PCR + 14 71 85
IFI + et PCR - 1 19 20
IFI – et PCR + 0 3 3
IFI – et PCR - 2 7 9
Total classe 17 100 117
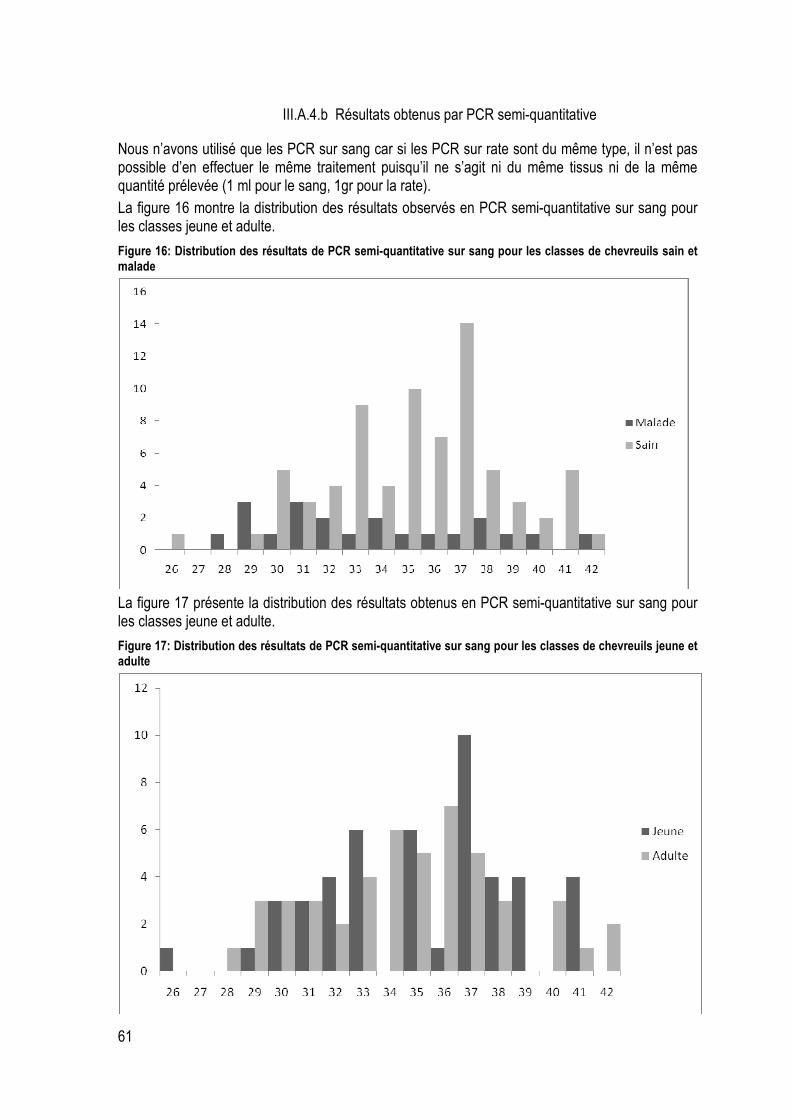
61
III.A.4.b Résultats obtenus par PCR semi-quantitative
Nous n’avons utilisé que les PCR sur sang car si les PCR sur rate sont du même type, il n’est pas possible d’en effectuer le même traitement puisqu’il ne s’agit ni du même tissus ni de la même quantité prélevée (1 ml pour le sang, 1gr pour la rate).
La figure 16 montre la distribution des résultats observés en PCR semi-quantitative sur sang pour les classes jeune et adulte.
Figure 16: Distribution des résultats de PCR semi-quantitative sur sang pour les classes de chevreuils sain et malade
La figure 17 présente la distribution des résultats obtenus en PCR semi-quantitative sur sang pour les classes jeune et adulte.
Figure 17: Distribution des résultats de PCR semi-quantitative sur sang pour les classes de chevreuils jeune et adulte

62
Pour indication,
• 30 ct équivalent à 105 copies d’ADN par ml
• 34 ct équivalent à 104 copies d’ADN par ml
III.A.4.c Répartition temporelle des chevreuils positifs
La figure 18 montre la distribution dans le temps des résultats obtenus en PCR sur sang.
Figure 18: Distribution des résultats en PCR sur sang selon les dates de prélèvement
III.A.4.d Répartition géographique des chevreuils positifs
Nous avons choisi ici de représenter tous les chevreuils pour lesquels au moins une analyse s’est révélée positive (IFI, PCR sur sang, PCR sur rate). La figure 19 présente la répartition géographique des chevreuils positifs pour au moins une analyse sur les trois. Le fond de carte de la figure 19 représente le découpage des différentes unités biogéographiques de populations de Chevreuils en région Franche-Comté.
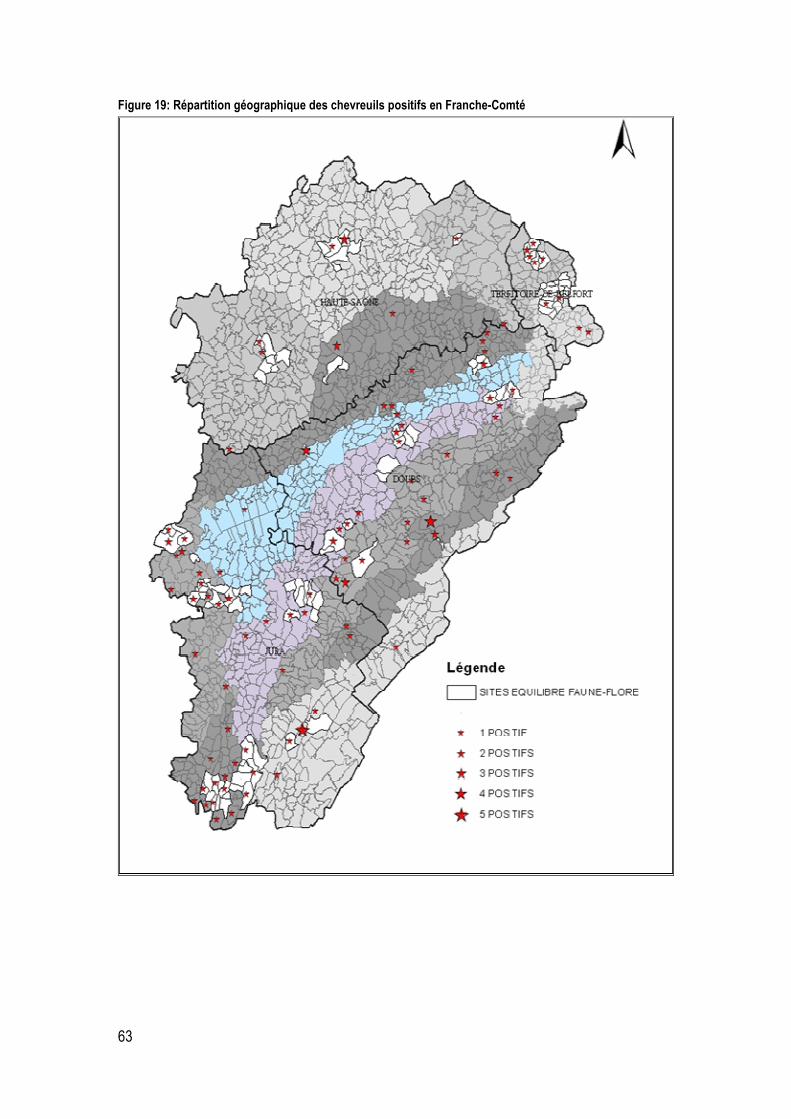
63
Figure 19: Répartition géographique des chevreuils positifs en Franche-Comté

64
.III.A.5 RESULTATS OBTENUS POUR LES AUTOPSIES ET LES TIQUES
III.A.5.a Les autopsies
Il y a eu 30 autopsies réalisées dont une ne pouvait pas être exploitée, l’animal étant dans un état de lyse avancé.
Sur ces trente chevreuils, 6 sont sont morts suite à un traumatisme (collision avec une voiture ou autre). Sur les 24 autopsies restantes, 19 autopsies ont permis de dégager des causes potentielles de mort et 5 n’ont pas dégagé de lésion significative.
Pour les 19 autopsies ayant donné un résultat, voici le type d’atteinte majeure rapporté pour chaque chevreuil:
• 8 chevreuils avec un poly-parasitisme important
• 4 méningo-encéphalites dues probablement à des combats
• 2 pasteurelloses respiratoires
• 1 hépatite à Fasciola hepatica (cause certaine de la mort)
• 1 pleuro-pneumonie abcédative chronique à Corynebacterium
• 1 suspicion de septicémie
Un certain nombre de facteurs communs sont apparus entre les autopsies. Les chevreuils étaient souvent porteurs de grandes quantités de parasites intestinaux et de bactéries dans le foie, la rate et les nœuds lymphatiques mésentériques. Ces bactéries étaient le plus souvent : Echerichia coli, Streptococcus bovis, staphylocoques, micrococcus et pseudomonas.
III.A.5.b Les tiques
Il n’y a pas pu y avoir de comptage précis et d’indication des stades de tiques infestant. La figure 20 montre le nombre chevreuils porteurs ou exempts de tiques en fonction du temps.
Il y a beaucoup de chevreuils porteurs de tiques jusqu’au 30 novembre.
Une seule tique a été prélevée entre le premier décembre et le 15 janvier sur 27 chevreuils.
Il y avait six chevreuils porteurs de tiques entre le 15 janvier et le 01 février.
Figure 20: distribution de la présence de tiques sur les chevreuils prélevés

65
.III.B. Etude rétrospective de la dynamique des populations de chevreuils de Franche-Comté, des conditions climatiques et des milieux
.III.B.1 OBJECTIFS
Les objectifs de cette deuxième étude étaient d’identifier et de caractériser les évolutions des populations de chevreuils ainsi que les conditions (climat et milieu) dans lesquelles elle ont évolué sur la saison de prélèvement 2007-2008.
Nous avons vu précédemment qu’il est judicieux de suivre une population de chevreuils sur plusieurs années pour bien comprendre les évolutions qu’elle suit. Nous nous sommes donc intéressés à la période 2003-2008.
.III.B.2 RESSOURCES UTILISEES POUR LE SUIVI
Nous avons fait la synthèse des résultats de suivi des populations de chevreuils sur les sites équilibre Faune-Flore sur la saison 2003-2008. Ces résultats nous étaient fournis sous format informatique par les fédérations départementales des chasseurs. Nous n’avons examiné que les indices de suivi qui avaient été mis en place depuis 3 ans ou plus pour chaque site. Ces comptes rendus de suivi des populations contenaient une brève description des milieux ce qui nous a permis de connaître les milieux qui forment le paysage franc-comtois.
Un compte rendu type de suivi d’une population de chevreuils sur un site EqFF est présenté en annexe 5.
Afin de connaître les conditions climatiques et environnementales dans lesquelles les chevreuils ont évolué entre 2003 et 2008, nous avons interrogé les techniciens de FDC. Il s’agissait de la source la plus fiable, ce personnel étant sur le terrain quotidiennement. Ces informations devaient complémentaires de celles recueillies par le suivi sur sites EqFF.
Ainsi une entrevue avec au moins un technicien de chaque FDC a eu lieu ainsi qu’une sortie sur une partie des sites EqFF pour identifier et caractériser de visu les différents milieux.
.III.B.3 RESULTATS DE L’ETUDE RETROSPECTIVE
III.B.3.a Historique de l’évolution des populations et de leurs conditions de vie
Les premiers cas de surmortalité interviennent en 1995/1996 dans le Doubs et le Territoire de Belfort (près de 600 chevreuils morts sur deux ans) (Martin, communication personnelle, de Meulemester, communication personnelle). Par la suite, le problème a disparu jusqu’en 2003.
L’année 2003 a été marquée par la canicule estivale qui a fait d’énormes dégâts dans les populations de chevreuils : le taux de survie estivale a été très faible chez les faons et la conformation insuffisante des faons à l’entrée de l’hiver a diminué le taux de survie hivernale. Le taux de survie à l’année des faons de l’année 2003 a été exceptionnellement faible. Les dégâts de l’année 2003 ont continué en 2004 car les chevrettes nées en 2003 ont eu des retards à l’entrée en reproduction et donc des difficultés à mener à terme l’élevage des faons de 2004.
Ce problème a été amplifié par une mauvaise prise en compte de l’impact de 2003 pour l’établissement des plans de chasse de 2004.(Techniciens de fédérations départementales de Franche-Comté, communication personnelle). Depuis, les plans de chasse sont plus faibles qu’auparavant et les taux de réalisation sont également plus faibles. A titre d’exemple ; le site de Levier dans le Doubs présentait un plan de chasse de 542 chevreuils en 2003 avec un taux de

66
réalisation de 87 % (474 chevreuils abattus). En 2006, le plan de chasse comptait 274 chevreuil avec un taux de réalisation de 89 % (244chevreuils abattus).
Depuis, la population a repris grâce à un printemps 2006 très humide, idéal pour la reproduction du chevreuil et la survie des faons. Aujourd’hui, les cas de mortalité sont plus faibles, les techniciens sont tous d’avis que la Franche-Comté est en queue de problème avec moins de cas de mortalités. Il n’y a pas eu de problèmes de surmortalité observés pour la saison 2007-2008.
La Franche Comté est une région particulière par le nombre de biotopes différents rencontrés ce qui a pour conséquences que les populations de chevreuils n’ont pas les mêmes relations avec leurs milieux d’une zone à l’autre.
La Haute Saône est une région où les plaines céréalières prédominent avec des massifs forestiers parsemés, de taille moyenne (une dizaine d’hectares en général). Ces massifs forestiers sont généralement assez clairs avec des essences feuillues et un sous-sol très riche. Ce sont des forêts très vertes et très humides. Les techniciens de la fédération départementale de Haute-Saône ont observé peu de problèmes de mortalités et de baisse des populations sur leurs sites malgré des densités importantes (les densités des plans de chasses sur certaines zones sont parmi les plus importante de la région).
Progressivement, du Nord vers le Sud le paysage s’inverse : on passe de milieux très ouverts de plaines avec des inclusions plus ou moins grandes de bois et de forêts a des milieux de plus en plus fermés avec des massifs forestiers de plus en plus couvrant et des inclusions de champs.
Au Sud, les milieux sont moins riches, il y a de plus en plus de résineux dans les forêt jusqu’à ce que ce soient les essences quasi-exclusives sur le haut plateau du Jura avec le Buis (Buxus sp.), une essence qui pousse typiquement sur les sols pauvres.
Le Doubs et le Jura présentent des milieux forestiers plus pauvres, plus secs, plus rocailleux et plus accidentés. De plus ces milieux très fermés offrent peu de champs céréaliers où les chevreuils et autres grands herbivores puissent se replier en cas de baisse des ressources dans les sous-bois.
Ces deux départements sont des départements où le lynx est maintenant bien installé et pose problème surtout dans les élevages ovins de la région. Ce prédateur peut avoir une part de responsabilité dans la baisse des observations de chevreuils sur ces départements. Il semble que le chamois aie déjà changé de comportement dans le Jura suite à l’arrivé du Lynx (Marillier, communication personnelle). C’est surtout sur ces départements qu’il y a eu une baisse des indicateurs et des cas de mortalités « massives ».
III.B.3.b Evolution de la santé des populations de chevreuils sur les sites Equilibre Faune Flore
Sur 16 sites équilibre Faune-Flore, il n’y a pas de récolte de données depuis au moins trois ans ce qui ne permet pas d’interpréter les résultats obtenus. Ces 16 sites ont été exclus de l’analyse.
Pour les quatre départements il y a des sites où des données sont récoltées depuis trois ans ou plus. Le tableau 21 donne les orientations des différents indicateurs de suivi depuis 2004 sur les 6 sites où les suivi est suffisamment ancien. Globalement, on constate que sur l’ensemble de la région Franche-Comté, les populations de chevreuils sont légèrement en baisse ou stable depuis 2004 alors qu’ils avaient été généralement en hausse avant 2003.
On constate le plus souvent une baisse de l’indice de consommation ce qui tendrait à soutenir l’impression de baisse du nombre de chevreuil. Il faut se méfier ici car sur la plupart des sites, les indices de consommation n’ont remplacé les indices de pression sur la flore que depuis un à deux ans.
Le poids des chevrillards évolue différemment selon le site étudié.
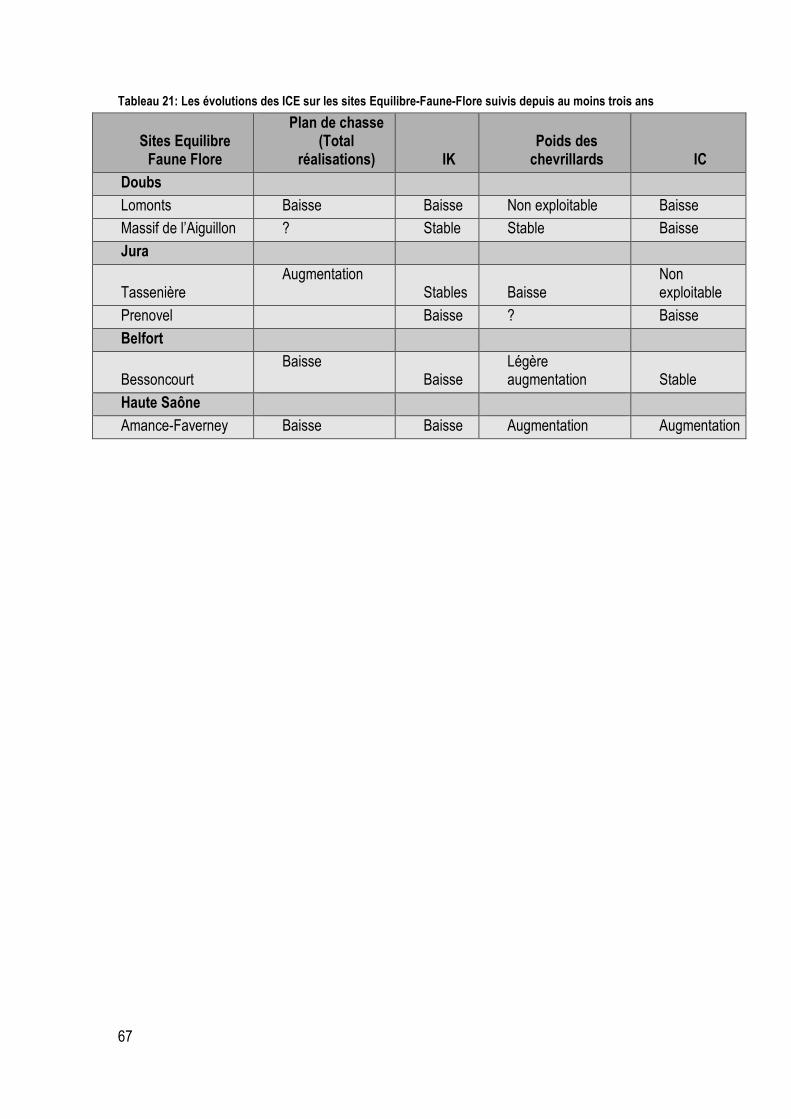
67
Tableau 21: Les évolutions des ICE sur les sites Equilibre-Faune-Flore suivis depuis au moins trois ans
Sites Equilibre Faune Flore
Plan de chasse (Total
réalisations) IK Poids des chevrillards IC
Doubs
Lomonts Baisse Baisse Non exploitable Baisse
Massif de l’Aiguillon ? Stable Stable Baisse
Jura
Tassenière Augmentation
Stables Baisse Non exploitable
Prenovel Baisse ? Baisse
Belfort
Bessoncourt Baisse
Baisse Légère augmentation Stable
Haute Saône
Amance-Faverney Baisse Baisse Augmentation Augmentation

68

69
Discussion
.I. Etude Cas-témoin : un portage plus important chez les malades
.I.A. Les chevreuils malades sont plus porteurs de la bactérie que les chevreuils sains
.I.A.1 INTERPRETATION DES RESULTATS DE L’ETUDE CAS TEMOINS
La comparaison des statuts entre chevreuils malades et sains n’a pas permis de dégager des différences significatives entre les deux classes. Le test exact de Fischer n’a en effet pas permis de dégager de différences significatives entre les deux échantillons pour les analyses IFI, PCR sur sang et PCR sur sang ou rate.
Cependant, ce test dégage un Odds-ratio de 3,35 pour la PCR sur sang qui est presque significatif au seuil de 5 % (p-value pour le test de 0,0526) ce qui signe une réelle différence dans le portage dans le sang entre chevreuils malades et sains. Nous pouvons considérer ici que les chevreuils malades sont réellement plus souvent porteurs que les chevreuils sains. Cependant, il faut d’autres éléments pour pouvoir considérer que A. phagocytophilum soit pathogène pour le chevreuil.
Le test exact de Fischer donne pour la PCR sur rate ou sur sang, un Odds-ratio de 1,41 non significatif (p-value de 0,51). L’ajout des données obtenues en PCR sur rate à celle de la PCR sur sang a permis d’augmenter le nombre de chevreuils cas qui est passé de 26 à 36. Ce test n’a pas mis en évidence la même différence que pour la PCR sur sang. Ce test ne montre pas que les chevreuils malades sont plus souvent porteurs d’A. phagocytophilum que les chevreuils sains. Ceci amène a conclure que le portage de la bactérie dans la rate n’a pas la même signification qu’une bactériémie.
L’étude de la séroprévalence a montré que les statuts entre malades et sains étaient très proches avec un Odds-ratio de 0,8 non significatif (p-value de 0,68). Ceci s’explique par les très forts taux de séroprévalence observés (entre 74 et 80 % selon l’échantillon considéré). Nous avons présenté et expliqué les principaux résultats de cette étude. Il nous faut maintenant vérifier que les méthodes d’analyse employées sont adéquates et ne posent pas de problèmes pour l’interprétation des résultats.
.I.A.2 QUALITES DES ANALYSES UTILISEES
Le faible nombre de chevreuils malades analysés (26 en PCR sur sang,36 en PCR tous tissus, 17 en IFI) a eu pour conséquence que la puissance des tests utilisés à été plus faible que prévue. A cause de ce faible nombre de chevreuils malades, la précision des tests à été moins bonne et on observe des intervalles de confiance importants (le test exact de Fischer donne un intervalle de confiance de presque 18 pour l’Odds-ratio de la PCR sur sang). Il est possible de ce fait que le test exact de Fischer pour la PCR sur sang aurait donné un résultat significatif et un Odds-ratio plus précis. Par ailleurs, la technique d’IFI requiert des prélèvements en bon état (pas de lyse cellulaire) ce qui a eu pour conséquences que 32 échantillons n’ont pas été analysés.
Nous disposons de peu d’informations sur la sensibilité de la méthode d’IFI utilisée car il n’existe pas de méthode de référence à laquelle nous pouvons la comparer. Cependant les résultats

70
obtenus sont élevés ce qui signifie qu’il est peu probable que la technique ne soit pas suffisamment sensible. Quant à la spécificité de cette technique, il existe des réaction croisées avec Anaplasma marginale (Vassalo, communication personnelle) mais cette bactérie n’est pas présente en Franche-Comté, c’est une bactérie qui est surtout présente dans les régions tropicales et subtropicales (Anonyme, 2005). Ce test ne pose donc en principe pas de problème de spécificité.
La méthode de PCR utilisée est sensible et spécifique d’A. phagocytophilum ce qui permet de considérer que les résultats obtenus avec cette technique sont relativement certains. Il est en effet très peu probable qu’il y ait eu des chevreuils porteurs d’A. phagocytophilum qui n’aient pas été détectées ou que des chevreuils non porteurs aient eu un résultat positif.
Pour répondre aux objectifs de cette étude, il avait été choisi d’utiliser une méthode de PCR semi-quantitative. Le but était d’avoir des résultats donnés en quantité d’ADN dans chaque échantillon pour effectuer des comparaisons d’intensités de portage, notamment entre sains et malades. Ceci nous aurait permis de mettre en évidence d’éventuelles différences d’intensités de portage entre malades et sains et aurait pu confirmer les orientations retenues à l’examen des fréquences de positivité.
Les résultats en quantité d’ADN présentés dans ce travail n’ont pas pu être utilisés pour plusieurs raisons.
Si la PCR ne nécessite pas un échantillon d’excellente qualité pour détecter A. phagocytophilum dans un échantillon, ce n’est pas le cas si on cherche à obtenir une notion de quantité. Il est difficile d’interpréter des différences de portage sur des échantillons qui ont été prélevés dans des conditions différentes et qui ont voyagé plus ou moins longtemps entre le lieu de prélèvement et le laboratoire.
Le deuxième problème observé est la répétabilité du test qui chute très vite au-delà d’un seuil d’amplification de 30 ct (C.Maingourd, communication personnelle) : un échantillon qui sort positif au seuil de 35 ct pourrait sortir positif au seuil de 36 ou 37 ct. Or durant cette étude, seulement six échantillons sont ressortis positifs en deçà de 30 ct. Ainsi les résultats obtenus en PCR semi-quantitative ne sont pas interprétable. Deux questions sont soulevées par ces résultats : le portage d’A. phagocytophilum est-il faible chez le chevreuil ou s’agit-il plutôt d’une baisse de sensibilité de la PCR semi-quantitative ? Nous ne pouvons répondre à cette question en l’état actuel des connaissance sur cette technique. La raison pour laquelle nous ne pouvons avoir de quantités absolues en PCR est que la gamme d’étalonnage est réalisée avec un plasmide sur lequel se trouve la portion de gène à amplifier.
La PCR quantitative pourra être un outil très intéressant pour l’étude approfondie du pouvoir pathogène d’A. phagocytophilum car elle permettra de travailler sur les intensités de portage entre malades et sains. Ceci permettra d’identifier des différences de portage entre jeunes et adultes et éventuellement d’en identifier la cause.
La corrélation entre présence de la bactérie et maladie n’est pas suffisante pour considérer qu’A. phagocytophilum puisse provoquer une maladie. Voici les arguments qui nous permettent de penser que cette bactérie peut provoquer un maladie chez le Chevreuil.
.I.A.3 ARGUMENTS BIOLOGIQUES EN FAVUER DE LA PATHOGENICITE D’A. PHAGOCYTOPHILUM
I.A.3.a L’ensemble de espèces proches du Chevreuil est sensible à la bactérie
L’ehrlichiose granulocytaire est une maladie qui est largement répandue chez les espèces d’élevage. On retrouve cette maladie chez les équidés, les bovins, les ovins et les caprins.

71
La pathogénie de cet agent est la même chez toutes les espèces d’élevage : A. phagocytophilum colonise les granulocytes neutrophiles et sa multiplication dans ces cellules provoque une inflammation et une immunodépression transitoire.
De ce fait , les signes cliniques sont proches d’une espèce à l’autre. Il est probable que les symptômes d’une ehrlichiose granulocytaire chez le Chevreuil soient proches de ceux observés chez les animaux d’élevages. Rappelons qu’il y a deux types de manifestations : celles liées à l’inflammation et celles liées à l’immunodépression.
Le syndrome inflammatoire se manifeste surtout par des symptômes généraux: chute de lait, infertilité, avortement, fièvre, anorexie et affaiblissement (Gokce & Woldehiwet, 1999a, Joncour, 2003).
L’immunodépression est exprimée par des chutes des taux de granulocytes neutrophiles, de granulocytes éosinophiles et des monocytes(Woldehiwet et al., 1999a).
Au vu de l’homogénéité de la pathogénie d’A. phagocytophilum et des manifestations cliniques engendrée par la maladie quelle provoque, il est normal de considérer que le Chevreuil puisse être sensible à cette bactérie. Si nous pouvons admettre que le Chevreuil soit sensible à la bactérie, nous ne pouvons savoir si il est plus ou moins sensible que ses homologues d’élevage.
Il y a de forte possibilités pour que le Chevreuil soit sensible à A. phagocytophilum et qu’il soit atteint d’ehrlichiose granulocytaire lorsque la bactériémie est suffisamment importante. Cette étude montre que la bactériémie est fréquente chez les Chevreuils. Par ailleurs, les mêmes variants de la bactérie ont été retrouvés chez la Chèvre, les ovins, les chevaux et le Chevreuil. Le Chevreuil héberge donc des variants qui sont pathogènes pour les autres espèces. Il y a une communauté de variant entre la Chèvre et le Chevreuil.
Si nous pouvons admettre qu’il n’y a pas d’argument biologique pour contredire l’idée qu’A. phagocytophilum soit pathogène pour le Chevreuil, il y a eu en revanche peu de cas documentés sur l’ehrlichiose granulocytaire du Chevreuil. Nous avons vu dans l’étude bibliographique qu’il y avait très peu de données sur cette maladie. Si A. phagocytophilum est pathogène chez le chevreuil, il semble que cette bactérie soit très discrète.
I.A.3.b L’ehrlichiose granulocytaire est difficile diagnostiquer même en élevage
Même chez les espèces domestiques, la maladie n’est pas évidente à détecter car son expression est courte et les signes cliniques sont non spécifiques (Joncour 2003). Chez les espèces d’élevage, la suspicion d’ehrlichiose clinique est basée sur la saison, sur le contexte écologique (presence de tiques, conformation du champs) et éventuellement sur la présence d’autres maladies à tiques (Joncour, 2003). Par ailleurs, cette maladie n’a été découverte qu’en 1991 en France alors qu’il y a une forte suspicion qu’elle existe depuis bien plus longtemps sur le territoire français (Joncour, communication personnelle).
Le diagnostic de l’ehrlichiose granulocytaire en élevage est donc plus souvent basé sur le contexte écoépidémiologique que sur ces manifestations cliniques. S’il est difficile de mettre en évidence la maladie chez les bovins, il est d’autant plus difficile de mettre en évidence cette même maladie chez des animaux sauvages.
Les 30 autopsies menées sur les chevreuils malades n’ont pas mis en évidence de lésions attribuables à une ehrlichiose granulocytaire clinique. Les lésions supposées de l’ehrlichiose granulocytaires sont principalement une splénomégalie et une hypertrophie de certains nœuds lymphatiques (Campbell et al., 1994). Or ces lésions sont très peu spécifiques : il y a beaucoup de causes potentielles pour une splénomégalie (bactériémie, lésion agoniques) ou une hypertrophie de nœuds lymphatiques (bactériémie, parasitisme intestinal). Ces lésions étant peu spécifiques, elles ont pu être attribuées à d’autres causes.

72
Par ailleurs, Les autopsies de chevreuils sont rares en laboratoire et une splénomégalie est difficile à objectiver. Il est fort possible que des splénomégalies soient passées inaperçues.
La connaissance apportée par l’étude bibliographique de cette thèse doit permettre aux laboratoires de connaître les lésions éventuellement associées à une ehrlichiose granulocytaire chez le Chevreuil et de rechercher plus spécifiquement ces lésions lors d’une suspicion.
Il n’y a pas eu de lésions évocatrices d’une ehrlichiose clinique mais nous l’avons vu les lésions sont difficile à objectiver. C’est le cas également pour les manifestations cliniques. On peut supposer que si le chevreuil manifeste les mêmes signes cliniques d’une ehrlichiose granulocytaire que des bovins ou des caprins, ces signes passent inaperçus. L’absence de lésions ne signifie pas qu’il n’y a pas eu d’atteinte clinique des chevreuils positifs.
L’inflammation a été exclue pour les 30 chevreuils autopsiés mais rien ne permet d’affirmer qu’il n’y ait pas eu un effet immunodépresseur chez certains chevreuils autopsiés.
I.A.3.c Effet immunodépresseur potentiel
Les autopsies ont permis d’établir avec certitude que 10 chevreuils étaient morts des suites de traumatismes. Il y a 5 chevreuils pour lesquels aucune cause de mort n’a été trouvée. Pour les 15 autres chevreuils, il y a plusieurs causes évoquées. Pour huit chevreuils, la cause suspectée de la mort est un fort parasitisme (strongles digestifs et respiratoires, Ostartagia, trichures…) intestinal et respiratoire. Il y a un chevreuil qui est mort d’une fasciolose hépatique. Les six autres chevreuils sont morts d’une infection bactérienne. Nous pouvons supposer ici qu’une immunodépression pourrait avoir favorisé l’expression de ces maladies.
Il est possible de se demander si le fait de classer 10 chevreuils traumatisés dans les malades n’a pas eu pour effet de ranger des chevreuils sains hormis le traumatisme dans l’échantillon de malades. Ceci étant dit, un chevreuil qui a été percuté par une voiture pouvait souffrir d’une maladie qui l’avait affaibli au préalable. Des hyperthermies ou une atteinte de la vision auraient pu favoriser des collisions ou des accidents. Sur ces 10 individus, 4 sont morts suite à des combats (encéphalites) ce qui semblerait les exclure complètement du cadre des malades.
Le fait qu’il y ait significativement plus de chevreuils malades porteurs de la bactérie montre que cette bactérie a eu un rôle, quel qu’il soit, dans l’apparition d’au moins une partie de ces maladies. Il est probable que pour certains des chevreuils malades, il y ait eu une immunodépression due à A. phagocytophilum qui aurait favorisé l’apparition de maladies parasitaires ou bactérienne.
Cette étude ne nous permet pas d’affirmer avec certitude qu’A. phagocytophilum provoque une immunodépression chez le Chevreuil bien qu’elle apporte des éléments pour le suspecter.
Cette bactérie n’est peut-être pas directement cause de mortalité surtout pour des chevreuils en bonne santé. En revanche, chez des individus affaiblis, A. phagocytophilum pourrait être cause indirecte de mortalité. Dans des populations où les chevreuils sont en mauvais état corporel et sanitaire, A. phagocytophilum pourrait intervenir en tant que facteur de surmortalité.
.I.A.4 VALIDITE DU MODELE
I.A.4.a Une sous estimation de l’âge
Il y a un déséquilibre dans la composition en âge entre les échantillons sain et malade. En effet, les jeunes sont sous représentés dans l’échantillon malade (moins de 30 % chez les malades contre 50 % chez les sains). Ce déséquilibre n’a pas pu être corrigé par le choix du chevreuil par sa classe de sexe contrairement à ce qui a été fait pour l’échantillon de chevreuils sains. En effet, tous les chevreuils malades ont été inclus à l’étude alors que les chevreuils sains étaient choisis au sein de l’ensemble des chevreuils tirés à la chasse. Le fait de n’avoir trouvé que 30 % de jeunes chevreuils

73
est attribuable au fait qu’il est plus difficile de trouver un jeune chevreuil malade parce qu’il est plus petit et donc plus difficile a trouver, meurt plus rapidement et disparaît plus rapidement.
Il y a donc une sous représentation des jeunes chevreuils chez les malades. Il semblerait qu’il y ait une différence entre les statuts des jeunes et des adultes vis-à-vis d’A. phagocytophilum. De ce fait, il est très possible que l’âge soit un facteur de confusion potentiel.
Il est possible que les jeunes chevreuils soient moins bien immunisés que les adultes surtout après la levée de l’immunité colostrale et qu’ils soient de ce fait plus souvent porteurs d’A. phagocytophilum que les adultes. Nous ne pouvons conclure sur ce point qu’il faudra approfondir dans des études à venir. Les premières phases de vie des chevrillards étant cruciales pour la santé des populations, il est important de savoir quel est l’impact d’A. phagocytophilum sur cette étape de la vie.
I.A.4.b Répartitions géographique et temporelle
Les chevreuils sains ont été prélevés sur une période de 5 mois entre septembre 2007 et janvier 2008. Les chevreuils malades ont été prélevés sur une période plus longue : le premier chevreuil a été prélevé le 4 juillet 2007 et le dernier le 14 mai 2008.
Les périodes de prélèvements ont donc été différentes, huit chevreuils malades ont été prélevés hors période de prélèvement des chevreuils sains. L’idéal aurait été que les deux périodes soient les mêmes car la transmission d’A. phagocytophilum dépend de l’activité des tiques qui est saisonnière.
Le faible nombre de chevreuils malades n’a pas permis de sélectionner les chevreuils malades sur la période de prélèvement ce qui aurait été le cas si il y avait eu suffisamment de chevreuils malades prélevés.
Les prélèvements de chevreuils devaient être effectués de préférence autour des sites Equilibre Faune-Flore. Cette consigne a été bien suivi pour les chevreuils sains puisqu’il y a eu des prélèvements de chevreuils sains sur tous les sites équilibre Faune-Flore de Franche-Comté ce qui permet d’obtenir un maillage complet de la Franche-Comté. En revanche il y a eu certains sites Equilibre Faune-Flore où il y a eu beaucoup plus de prélèvements notamment dans le Jura et le territoire de Belfort. Ceci s’explique par le fait que le Jura a dépassé les objectifs de chevreuils sains alors que le Doubs où la Haute-Saône étaient en deçà des objectifs. Il y a donc un effet de regroupement sur les cartes de prélèvement pour certains sites, notamment Tassenière (39), Recépage (39) ou Faye (90).
Si les chevreuils sains sont répartis de façon homogène malgré des effets de regroupement sur certains sites, les chevreuils malades sont moins bien répartis. En effet il y en a eu 24 prélevés dans le Jura, 10 dans le Doubs et deux en Haute Saône. Il n’y en a aucun dans le territoire de Belfort. Les prélèvements de chevreuils malades ont donc été regroupés sur le Sud de la région et forment une flèche dont la pointe est orienté vers le Sud Est de la région et dont le corps suit le deuxième plateau du Jura. Au vu du faible nombre de chevreuils prélevés au total, on ne peut dire s’il s’agit d’une réelle répartition des cas de maladies ou si cette différence est liée à d’autres paramètres (pression d’observation, type de milieu). Toujours est-il que si la répartition des chevreuils sains est relativement homogène, celle des chevreuils malades ne l’est pas. Ce n’est pas surprenant au vu du faible nombre de chevreuils malades. Les lieux de prélèvement des chevreuils malades sont rarement liés à un site Equilibre Faune-Flore. Comme pour les dates de prélèvements, les lieux de prélèvements sont différents entre malades et sains et l’explication est la même : appliquer les mêmes contraintes aux deux classes n’aurait pas permis d’obtenir suffisamment de chevreuils malades. Ceci étant, il y a eu plusieurs prélèvements de chevreuils malades sur les sites Equilibre Faune-Flore. La cause de cet effet est que la pression d’observation a été plus forte sur les sites Equilibre Faune-Flore.

74
.II. Des prévalences et séroprévalence élevées et une circulation homogène
.II.A. De nombreux chevreuils positifs
L’étude menée est une étude cas témoins ce qui veut dire que les chevreuils ont été échantillonnés pour représenter d’une part les malades et d’autre part les sains et non pour représenter la population totale. Les échantillons n’étant pas représentatifs de la population, nous ne pouvons considérer que les résultats obtenus sont applicables à la population totale des chevreuils.
Ceci étant, les résultats obtenus donnent une orientation et montrent que le portage et la séroprévalence sont très élevés. En PCR rate ou tissus, le portage dans l’échantillon sain est de 74,56 % (IC à 95 % de 65,55 82,25 %) et en IFI la séroprévalence dans l’échantillon sain est de 90,10 % (IC à 95 % de 82,55 % à 95,15 %).
Ceci montre qu’A. phagocytophilum circule beaucoup dans les populations de chevreuils. Ces chiffres sont plus élevés que ceux obtenus dans d’autres publications qui donnent des séroprévalences aux alentours de 60 à 70 %.
L’IFI dose des immunoglobulines de type M qui ont une persistance estimée à six semaines (Vassalo, communication personnelle). La séroprévalence globale observée est donc élevée soit parce qu’il y a un contact régulier avec A. phagocytophilum (ré-inoculation régulière par les tiques) soit parce qu’A. phagocytophilum persiste plus longuement chez le chevreuil que chez les bovins chez lesquels la persistance de la bactérie est d’une dizaine de jours. La séroprévalence élevée n’est pas due à une persistance longue des anticorps détectés par l’IFI. Il y a donc eu une circulation importante de la bactérie au sein des populations de chevreuils de Franche-Comté pour la saison de chasse 2007/2008.
.II.B. Une répartition homogène sans foyers de positivité
L’observation par cartographie des cas de chevreuils positifs soit par IFI soit par PCR montre qu’il y a un maillage complet de la région. Il y a cependant des zones où il semble y avoir eu plus de chevreuils positifs. Ces zones coïncident avec celles où il y a eu plus de prélèvements. Il ne s’agit donc pas de foyers de cas positifs mais d’un biais lié à la répartition initiale des prélèvements. La répartition des chevreuils positifs peut donc être considérée comme homogène sur l’ensemble de la région Franche-Comté. La positivité des analyse ne se cantonne pas à un ou quelques massifs qui pourraient être considérés comme des foyers de positivité.
Il y a donc une circulation importante et homogène d’A. phagocytophilum dans les populations de chevreuils de Franche-Comté.
Les données sur l’ehrlichiose granulocytaire dans les cheptels ovins et bovins de Franche-Comté rapportent l’existence de cette maladie sur le territoire Franc-comtois depuis 15 ans (TRIMAILLE, Communication personnelle). Il est possible que la présence de la bactérie en Franche-Comté soit encore antérieure à 15 ans car c’est l’acquisition de connaissances sur la maladie qu’elle engendre chez les bovins qui a permis de la mettre en évidence.
Cela fait donc 15 ans qu’une bactérie pathogène circule chez les populations de chevreuils. Or les premiers cas de surmortalité remontent à 1995 soit il y a 13 ans.
La question qui suit est est-ce que les premiers cas de mortalité des chevreuils en Franche-Comté coïncide avec l’arrivée de la bactérie sur ce territoire ? Nous pensons que la bactérie est présente depuis plus longtemps et que les deux évènements (arrivée de la bactérie et mortalité des chevreuils) sont éloignés.

75
Il est possible cependant que A. phagocytophilum ait eu un rôle dans ces mortalités mais il n’est probablement pas du à son arrivée sur le territoire franc-comtois.
Nous avons vu qu’il y avait une distribution importante et homogène d’A. phagocytophilum. Les analyse en PCR ont montré également que le nombre de chevreuils sains porteurs d’A. phagocytophilum était important.
.III. Une saison 2007/2008 marquée par une faible mortalité
Il y a eu 36 chevreuils malades prélevés sur une période de huit mois (de septembre 2007 à mai 2008). La pression d’observation a été maintenue pendant toute la durée du protocole par une information par le biais d’articles dans les journaux de chasse de Franche-Comté et par une information sur le terrain assurée par les techniciens des FDC. Le faible nombre de chevreuils malades prélevés n’est donc pas attribuable à un manque d’observation sur le terrain comme nous l’avons déjà indiqué. Il a pu y avoir des cas de chevreuils malades qui n’ont pas été signalés. La principale cause de ce faible nombre de malades observés est avant tout le faible nombre de malades existant sur la région Franche-Comté. La saison de prélèvements a couvert l’automne 2007 et le printemps 2008, or c’est à l’automne et au printemps que les mortalités de chevreuils étaient le plus souvent observés (techniciens de FDC, communication personnelle). Le faible de nombre de morts retrouvés corrobore les impressions des techniciens de FDC qui ont constaté une baisse de la mortalité des chevreuils depuis début 2007.
L’objectif de cette étude était de savoir si A. phagocytophilum pouvait être tenue responsable de maladies chez le Chevreuil.
La saison de prélèvement 2007/2008 a duré 8 mois pour les chevreuils malades et seulement 36 chevreuils malades ont été recrutés. Cette situation n’est pas comparable à celles observées les années précédentes.
Nous avons vu lors de l’étude rétrospective des populations de chevreuils que les saison précédentes avaient été marquées par des mortalités observés importantes et cela alors que la pression d’observation était moindre. Par ailleurs, les indicateurs de suivi semblent se stabiliser depuis 2006.
Le faible nombre de chevreuils malades retrouvés n’est pas du à un manque d’observation car l’ensemble des chasseurs était mobilisé pour cette étude et cette mobilisation a été maintenue pendant toute la durée de l’étude.
Si une campagne de prélèvement qui dure huit mois et qui réunit l’ensemble des chasseurs sur une région entière ne permet de détecter que 36 chevreuils malades, c’est que sur la période de prélèvement, il n’y a pas eu de surmortalité. Sur la saison de chasse 2007/2008, il n’y a pas eu de problèmes de mortalités de chevreuils. Les problèmes de surmortalités des chevreuils observés depuis 1995 se caractérisaient par des mortalités groupées dans le temps et dans l’espace, or ici les 36 prélèvements se sont fait de façon homogène dans le temps (jamais plus de trois chevreuils par semaine de prélèvement) et ne sont pas regroupés autour d’un massif ou de plusieurs massifs (cf carte de répartition des prélèvements).
Si A. phagocytophilum est pathogène pour le Chevreuil, cette bactérie n’a pas causé beaucoup de mortalité sur la saison de chasse 2007/2008. La présence importante de cette bactérie n’a pas eu d’impact important en mortalité pour la saison 2007/2008.
La pathogénicité d’une bactérie s’exprime par la maladie et les lésions qu’elle engendre avant tout. Il est donc intéressant de savoir si les chevreuils malades étaient porteurs de lésions pouvant être attribuées à une ehrlichiose clinique.

76
.IV. Une bactérie retrouvée tout au long de l’hiver dans le sang des chevreuils
Le suivi de la prévalence en PCR sur sang a permis de montrer qu’on retrouvait des chevreuils porteurs tout au long de l’hiver. Parallèlement, la récolte des tiques diminue avec l’arrivée de cette saison.
Il est difficile cependant d’en tirer des conclusions puisque la recherche de tiques sur un chevreuil nécessite un examen très approfondi de tout le corps de l’animal. Certaines zones telles que l’intérieur des conduits auditifs ou l’intérieur des narines sont difficiles d’accès mais sont des lieux très souvent parasités par les tiques en hiver. De nombreux parasitologues (L’Hostis, communication personnelle) estiment que l’activité des tiques se maintient durant l’hiver malgré un ralentissement. Le fait de n’avoir pas trouvé de tiques sur certaines périodes de la saison de prélèvement ne veut donc pas dire que l’activité des tiques était nulle à ce moment.
Même si l’activité des tiques n’est pas nulle en hiver, la présence de chevreuils positifs en PCR tout au long de l’hiver permet de poser l’hypothèse d’une portage à long terme. Ceci expliquerait qu’on ait retrouvé des chevreuils positifs même dans des conditions de températures extrêmes pour la survie de la tique. Si cette étude ne permet pas de répondre à cette hypothèse, elle permet de l’avancer.
Aucune recherche par PCR d’A. phagocytophilum n’a été entreprise sur les tiques. La raison de cette absence de recherche est avant tout qu’il y avait trop peu de tiques pour faire une recherche efficace et que cette recherche n’apporterait rien de neuf sur les connaissances existant déjà sur les rapports entre tique et bactérie. Nous savons déjà qu’il y a un pourcentage faible de tiques porteuses de la bactérie qu’il y a passage trans-stadial mais pas trans-ovarien (Liz, 1994). Le faible nombre de tiques prélevés ne permettait pas d’approfondir les connaissances déjà existantes sur le rôle d’Ixodes ricinus dans le cycle d’A. phagocytophilum.
.V. Le rôle potentiel du Chevreuil dans le cycle épidémiologique d’A. phagocytophilum
Les rares études qui ont porté sur la comparaison des statuts immunitaires entre chevreuil et autre ongulés sauvages ont montré qu’il y avait une séroprévalence significativement plus importante chez le chevreuil par rapport aux autres espèces (Stuen, 2003). Ici nous observons également une séroprévalence élevée ce qui montre que le chevreuil n’entretient pas les mêmes rapports avec A. phagocytophilum que les autres ongulés sauvages. Une séroprévalence plus importante peut s’expliquer de deux façons.
La première est que le chevreuil est plus en contact avec la bactérie que les autres espèces ce qui ne peut s’expliquer alors que par le fait que le chevreuil soit plus parasité par les tiques que les autres. Le chevreuil est en effet considéré comme un très bon hôte pour les tiques (Joncour, 2003). La deuxième explication possible est que le chevreuil héberge plus longtemps la bactérie que les autres ongulés. En augmentant le temps de contact, il augmente également la séroprévalence. La présente étude ne permet pas de conclure sur ce point.
On peut alors se demander si le chevreuil n’a pas un rôle de réservoir de l’ehrlichiose en tant qu’animal réceptif mais peu sensible. Cette question est importante puisque nous savons que l’ehrlichiose granulocytaire est une maladie qui touche de nombreux mammifères, entre autres les bovins et les ovins ainsi que les équins. Rappelons également qu’il s’agit d’une zoonose importante

77
en Amérique du Nord. On sait que le chevreuil est capable de transmettre A. phagocytophilum à la tique, ce n’est donc pas un cul de sac épidémiologique.
Les stades de tiques qui infestent le chevreuil sont surtout les nymphes et les adultes ce qui fait que dans la plupart des cas, une tique qui s’est gorgée de sang infecté sur un chevreuil ne peut transmettre qu’une seule fois la bactérie (pas de transmission trans-ovarienne). Ceci mène à penser que si le chevreuil est effectivement un bon porteur pour la bactérie, le rayonnement infectieux d’un individu reste faible sauf durant les périodes d’activité importante des tiques.
Les pics d’activité des tiques du genre Ixodes se situent dans des conditions climatiques normales au printemps et à l’automne car les températures et l’hygrométrie sont alors idéales. Ces pics d’activité coïncident avec les cas d’ehrlichiose clinique chez les animaux de rente.
Lors des analyses, huit chevreuils sains se sont révélés positifs en PCR sur rate alors qu’ils avaient été négatifs en PCR sur sang. Ceci serait du au rôle d’organe filtre que joue la rate, qui aurait tendance à filtrer les pathogènes du sang et à les concentrer en son sein. Nous supposons ici que les chevreuils positifs en PCR sur rate et négatifs en PCR sur sang étaient soit en fin d’infection soit en début d’infection avec une absence de bactériémie ou une bactériémie non détectable et une concentration du pathogène dans la rate qui serait précoce ou tardive. La phase de bactériémie étant considérée comme très précoce (cinq à six jours post-infection), il semble plus probable qu’il s’agisse d’une persistance tardive de la bactérie. Rappelons que pour beaucoup de bactéries et parasites intracellulaires du sang, les individus splénectomisés sont souvent de bons réservoirs. Ainsi on peut supposer que la rate a un rôle extrêmement important dans l’élimination d’A. phagocytophilum.
.VI. Intégration des données de suivi des populations
.VI.A. Impacts des évènements climatiques sur les populations de chevreuils
L’épisode caniculaire de l’été 2003 a eu des répercussions sur la quasi-totalité du territoire avec pour conséquences une baisse des taux de survie juvénile sur l’année, une augmentation de la mortalité, une baisse globale des populations et une baisse des poids des jeunes.
Cet épisode a eu un impact moindre voire absent sur les zones forestières les plus riches de Haute-Saône. Pour rappel, la Haute-Saône est une région plus riche et plus humide que les trois autres départements de Franche-Comté. Sur le site Equilibre Faune-Flore d’Amance-Faverney il n’y a pas eu d’impact majeur observé de la canicule alors que sur les sites du Doubs et du Jura, les impacts ont été très importants.
Les répercussions de la canicule de l’été 2003 ont été étudiées sur les deux sites de références nationaux : Chizé et Trois-Fontaines. Les répercussions décrites sont les mêmes que pour la Franche-Comté (Van Laere et al. 2006).
Précisons ici qu’il y a eu une augmentation de la mortalité observée sur ces deux réserves.
Par la suite, les printemps de 2004 et 2005 ont été trop secs pour permettre une bonne reproduction et une réussite de la survie des juvéniles. La bonne qualité du printemps 2007 a permis une reprise de la reproduction.
Ces différents évènements climatiques ont eu des effets plus ou moins marqués sur les populations de chevreuils.

78
.VI.B. Mécanismes explicatifs des répercussions des mauvaises conditions de milieux
La répercussion importante de ces évènements climatiques s’explique principalement par le mode de reproduction du Chevreuil. Ce n’est par sur les chevreuils adultes hors reproduction qu’il y a des répercussion car le Chevreuil est capable de vivre sur des milieux pauvres. Nous avons vu que la chevrette sacrifie beaucoup d’énergie à la gestation, donnant naissance à un faon déjà très développé (Delorme et al., 2007). Par ailleurs, l’augmentation rapide du poids du faon est inféodée à la richesse du milieu. Les chevrettes doivent atteindre un poids et une conformation suffisamment importants pour pouvoir soutenir la demande en énergie de la gestation et de la croissance du faon par la suite (Delorme et al., 2007). Si les ressources sont insuffisantes au moment de la mise bas, les chevrettes ne vont pas pouvoir mener à bien la croissance de leurs faons et vont même jusqu’à les abandonner. Le manque de ressources sur une saison a donc un impact direct sur la survie des faons. Par la suite, les chevrettes issues de cette génération vont avoir une conformation trop faible l’année suivante pour entrer en reproduction. Il y aura donc une baisse de la natalité l’année suivante et des retards d’entrée en reproduction (Delorme et al., 2007). Une année à problème a donc un impact au moins deux années voire plus.
Nous l’avons dit, dans les zones les plus refermées et les moins riches, les effets ont été plus marqués. Ceci nous conduit à penser que les populations de chevreuils existant dans ces milieux sont plus exposés lors de perturbations climatiques. La raison de ce constat est très probablement due au fait que les milieux les plus pauvres ont des capacités d’accueil en densités de chevreuils plus faibles. Il n’est pas exclu que des problèmes de populations puissent être observés à l’avenir dans les autres massifs de la région Franche-Comté.
Chaque milieu a une limite d’accueil liée à la quantité d’aliments potentiels pour les chevreuils. Cette limite est liée aux ressources disponibles, aux capacités de régénération du milieu et à la qualité de ces ressources. Il est important de garder à l’esprit que la densité acceptable en chevreuils n’est pas la même quelque soit le milieu concerné : un milieu pauvre pourra supporter moins de chevreuils.
Hors conditions exceptionnelles, si un milieu est à sa limite, la dynamique de la population de chevreuils n’est pas excellente mais les effets ne peuvent être observés que par un suivi de bonne qualité et de longue durée (Delorme et al., 2007). Nous avons vu dans l’étude bibliographique que d’après Delorme (2007), les taux de survie sont faibles mais pas catastrophiques (plus de 50 %) sur ces milieux durant les bonnes années. En revanche en cas de perturbation majeure, on observe les effets dont nous avons déjà discuté. Ces effets là sont beaucoup plus observables : baisse du taux de survie juvénile très important (moins de 25 %), baisse des populations, mortalité de l’ensemble des classes d’âge.
Si ces conditions de déséquilibre ne sont pas visibles durant des saisons normales, il est possible qu’elles durent sur plusieurs années. Alors les conséquences d’une saison anormale paraissent d’autant plus brutales et importantes. De plus, comme elles atteignent généralement des massifs où les populations sont très denses, les mortalités observées, les baisses de densités les mortalités juvéniles sont encore plus observables. Ceci pourrait mener l’observateur à croire à un évènement du à une maladie plus qu’à un déséquilibre population-milieu.
Il est possible que les répercussions exceptionnelles de la canicule de 2003 soient aussi dus à la baisse de la disponibilité en eau, il semble en effet que le Chevreuil soit plus dépendant de la qualité des ressources en eau que des ressources alimentaires (Wallach et al., 2007)

79
.VI.C. Relations entre maladies et conditions du milieu
Un déséquilibre population-milieu est souvent caractérisé par une forte densité. Les fortes densités sont plus propices à l’expression de maladies infectieuses car la promiscuité entre individus est plus importante. Par ailleurs un milieu en déséquilibre est un milieu où les ressources viennent à manquer ce qui fait que les chevreuils peuvent se regrouper autours de zones plus riches voire autour de zones d’agrainage et d’élevage dans le milieu. Qui plus est, un milieu en déséquilibre produit des chevreuils plus faibles et avec un système immunitaire débilité ce qui les rend plus vulnérables. Il y a donc une accentuation des manifestations de ce déséquilibre qui se manifeste par une augmentation de la mortalité par des maladies infectieuses et parasitaires.
Il est probable que les problèmes observés chez les populations de chevreuils en Franche-Comté - baisses de densités, baisses des poids des jeunes, baisse des taux de réalisation - soient dus avant tout aux conditions du milieu et qu’ils se soient manifestés par l’apparition de mortalités dues à des maladies diverses.
L’étude bibliographique a montré que le Chevreuil était présent sur tout le territoire français et que sur les massifs de l’Est et du Sud-ouest, les plans de chasse étaient au moins aussi élevés qu’en Franche-Comté (Delorme & Van Laere, 2007). Si la topographie et le climat de la Franche-Comté sont propres à cette région, les problèmes de densité-dépendance qui y ont probablement eu lieu sont eux applicables aux autres régions. On a rapporté des cas de surmortalités importantes dans d’autres régions de France qui partageaient les mêmes caractéristiques que ceux observés en Franche-Comté (Marti-Brizen, 2007). Par ailleurs, la stagnation des plans de chasse et la régression des taux de réalisation est observée sur l’ensemble du territoire français (Anonyme, 2008) Dans la plupart des cas de surmortalité français, le mécanisme explicatif doit être proche de celui que nous avons présenté. Il s’ensuit que les conclusions et les recommandations de cette étude sont applicables à la plupart des régions où de tels phénomènes ont été observés. Nous excluons de ceci le Chevreuil dit de plaine puisqu’il évolue sur des milieux radicalement différents et qu’il semble ne pas rencontrer de problèmes (Dunoyer, communication personnelle).
Nous constatons que seulement six des 22 sites EqFF ont donné pour 2007 des orientations interprétables. Pour les 16 autres sites EqFF, la durée du suivi a été insuffisantes pour permettre d’interpréter les résultats. Ceci est du avant tout au délai entre mise en place du protocole et validité des résultats qui est de trois ans.par ailleurs, pour de nombreux sites les indicateurs ont été changé, notamment les indice de pression sur la flore qui ont été remplacé par les indice de consommation. Notons que pour la plupart des sites, les résultats seront interprétable dès l’année 2008 ou l’année 2009. Le problème du suivi des populations de chevreuils est qu’il est extrêmement lourd a mettre en place, que ce soit en temps, en personnel ou en logistique. Il est difficile de mettre en place un dispositif qui fait appel à plusieurs centaines de personnes sur plusieurs années à des dates fixées et sur des transects également fixes. Cependant les données, bien que lacunaires, que nous avons recueillies sont très utiles et montrent l’intérêt de continuer à effectuer un suivi de qualité des populations de chevreuils comme il est fait en Franche-Comté.
.VII. Le rôle pathogène d’A. phagocytophilum dans les populations de chevreuils de Franche-Comté en 2007/2008 et avant
Nous avons démontré que A. phagocytophilum circulait de façon très importante au sein des populations de chevreuils de la région Franche-Comté et nous avons mis en évidence que le portage de la bactérie dans le sang était plus important chez les chevreuils malades par rapport aux

80
chevreuils sains. Nous avons également montré qu’il y avait suffisamment d’argument biologique pour suspecter un pouvoir pathogène à cette bactérie et une manifestation clinique par une inflammation et une immunosuppression.
Enfin le faible nombre de chevreuil retrouvés malades montre qu’A. phagocytophilum n’a pas eu un impact important sur les populations de chevreuils de Franche-Comté sur la saison de chasse 2007/2008.
Nous avons montré que les problèmes de surmortalité observés dans les populations de chevreuils de Franche-Comté sont apparus suite à des perturbations climatiques. Les problèmes de surmortalité sont donc apparus parce que les populations de chevreuils ont été mises dans des conditions défavorables. Les populations de chevreuils, très denses sur certains massifs, étaient plus faibles et en plus mauvais état que des populations évoluant dans de bonnes conditions.
Il y a sûrement eu des mortalités directes (morts de faims, faons abandonnés…) mais la majorité de la mortalité observée était due à plusieurs agents pathogènes.
.VII.A. Impact potentiel de la présence d’A. phagocytophilum
Les chevreuils évoluant sur les milieux les plus pauvres étaient en moins bonne santé et étaient donc plus réceptifs à des infections opportunistes. L’impact d’A. phagocytophilum sur ces chevreuils a pu être important. Il y a deux questions qui se posent devant cette supposition. Tout d’abord, y a-t-il eu des cas de mortalités dus à des ehrlichioses granulocytaires aiguës (maladie inflammatoire) ? Ensuite, y a-t-il eu un impact important de l’effet immunosuppresseur d’A. phagocytophilum durant ces années problématiques, favorisant d’autant plus l’apparition de maladies opportunistes ? nous ne pouvons pas donner une réponse exacte sur le rôle qu’a joué A. phagocytophilum lors des années où une surmortalité a été observée car la saison 2007/2008 n’a pas été marquée par ces problèmes. Nous ne pouvons donc appliquer directement ce qui a été observé durant l’étude à ce qui s’est passé avant.
Il est probable qu’il y ait eu un impact de la présence importante de cette bactérie au sein des populations de chevreuils lors des années à problèmes. Mais cette bactérie n’est pas le seul agent pathogène à avoir joué un rôle dans ces problèmes. La cause de ces problèmes est avant tout un problème de densité-dépendance dans certains massifs. La mortalité par des maladies infectieuses et parasitaires n’en a été qu’une expression et A. phagocytophilum n’est pas le seul agent pathogène en cause. Il faudra de nouvelles investigations pour mieux connaître la pathogénie de la bactérie chez le chevreuil afin de savoir quel rôle cette bactérie peut jouer dans la mortalité des chevreuils sauvages. Des études par inoculation en stations expérimentales pourraient répondre aux questions qui se posent à la suite de cette étude.
.VII.B. Mortalité naturelle
Pour finir cette discussion, il est intéressant de noter que la Franche-Comté est une région à fortes densités en chevreuils. Il y a toujours de la mortalité naturelle au sein d’une population quelle qu’elle soit et la mortalité observable n’est qu’une fraction de celle-ci. Philippe Gibert (communication personnelle) précise qu’une mortalité de 20 % par an n’a rien d’anormal dans une population sauvage. Sur des populations de faible densité, ces 20 % ne représentent pas un grand nombre de chevreuils et la mortalité observable sera encore plus faible. Mais sur une plus grande population, 20 % représentent un grand nombre d’individus et la mortalité observable peut devenir très élevée. Il est donc difficile, ne connaissant pas les densités exactes de chevreuils évoluant sur un massif, de connaître précisément le taux de mortalité dans cette population. Ceci ne remet pas en cause les observations de surmortalité de Franche-Comté puisqu’elles sont basées sur une comparaison d’année en année ; il y a bien eu une augmentation de la mortalité à partir de 2003. En revanche cela remet en cause l’impact que cela a eu sur les populations. Il est très difficile

81
d’estimer l’impact qu’a eu cette mortalité. Seul le suivi nous permettra de voir si les populations de chevreuils sont toujours en situation de densité-dépendance ou si la surmortalité a eu un effet bénéfique, faisant revenir les chevreuils à des densités inférieures et évitant ainsi un problème de densité-dépendance.

82

83
Conclusion
L’étude expérimentale que nous avons mené nous a permis de montrer que la bactérie A. phagocytophilum était susceptible de provoquer une maladie chez le Chevreuil : l’ehrlichiose granulocytaire. Au vu des résultats, il y a une forte suspicion pour que l’ehrlichiose granulocytaire s’exprime par immunodépression chez le chevreuil et que son impact sur une population soit plus important quand cette populations évolue dans de mauvaises conditions de vie.
En Franche-Comté de la canicule de 2003 a été suivi d’une surmortalité importante chez les chevreuils qui a continué jusqu’en 2007. Cette surmortalité a été plus importante sur les populations de chevreuils évoluant en grandes densités sur des milieux déjà pauvres. Ces populations étaient probablement en situation de densité-dépendance. Les populations de chevreuils en situation de densité-dépendance sont des populations qui sont en mauvais état sanitaire et démographique. La survenue d’une évènement climatique exceptionnel sur ces populations peut provoquer une mortalité importante due à des maladies infectieuses ou parasitaires. Il est probable que la présence d’A. phagocytophilum au sein de ces population ait eu un impact au moins par un effet immunodépresseur et peut être par des syndromes inflammatoires connus chez cette bactérie pour les animaux d’élevage.
Si A. phagocytophilum a eu un impact sur ces populations, ce n’est certainement pas le seul agent pathogène en cause. Cette bactérie fait partie d’une mosaïque de maladies classiques du Chevreuil qui sont favorisées par les conditions de vie.
Ce sont les conditions de vie qui sont la cause de surmortalité. La surmortalité par infections n’est qu’un symptôme d’un mauvais état de la population. Il est donc très important de suivre les populations de chevreuil de près pour prévoir et anticiper le type de problème observé en Franche-Comté. Par ailleurs, les problèmes observés en Franche-Comté sont comparables à d’autres problèmes observés dans plusieurs grands massifs français. Les conclusions de ce travail sont donc probablement applicables à d’autres régions de France. La nécessité du suivi de populations est elle un devoir pour l’ensemble des régions où le Chevreuil est une espèce d’intérêt.
Les conclusions de ce travail restent lacunaires du fait des limites de l’étude d’une population sauvage. Les manifestations cliniques de l’ehrlichiose granulocytaires du Chevreuil, l’intensité et la durée du portage sont des élément qui ne pourront être étudiés que par l’inoculation en station expérimentale malgré les problèmes d’éthiques que cela suscite.
Par ailleurs, si le chevreuil est atteint d’une ehrlichiose granulocytaire, la question se pose de sa capacité à transmettre cet agent pathogène aux espèces d’élevage. Il serait intéressant d’établir le rôle que joue le Chevreuil dans le cycle épidémiologique de l’ehrlichiose.Si l’étude en station peut apporter des réponses, une étude préalable sur la communauté de variants est nécessaire. C’est cette étude qui sera mené en Franche-Comté afin de savoir si le Chevreuil et les Bovins partagent les mêmes variants ou non. Pour l’instant, rien ne permet d’affirmer que le Chevreuil puisse jouer un rôle important.

84

85
Bibliographie
.I. Références bibliographiques
Albaret M, Péroux R, Gaillard JM, Lartigues A (1991), Les relevés d’indices de présence comme indicateurs des variations d’effectif chez le chevreuil (Capreolus capreolus) Gibier Faune Sauvage, 8, 231-250
Anonyme (1991), Méthodes de suivi des populations de chevreuils en forêt de plaine. Exemple : l’indice kilométrique (I. K), Sup Bull Mens ONCFS, 157, Fiche n°70
Anonyme (1996), Les bio-indicateurs : futurs outils de gestion des populations de chevreuils ?, Sup Bull Mens ONCFS, 209, Fiche n°90
Anonyme (1996), Un indicateur biologique fiable : La masse corporelle des jeunes chevreuils, Sup Bull Mens ONCFS, 209, Fiche n°91
Anonyme (1999), La gestion des populations de chevreuils par l’utilisation d’indicateurs population-environnement, Sup Bull Mens ONCFS, 244, Fiche Technique n°95
Anonyme (2008), supplément plans de chasse, Faune Sauvage, 279
Argenté G, Collin E, Morvan H (1992), Ehrlichiose bovine (fièvre des pâtures) : une observation en France, Le Point Vétérinaire, 24, 89–90.
Barandika JF et al. (2007), Tick-Borne Zoonotic Bacteria in Wild and Domestic Small Mammals in Northern Spain, Appl and Environ Microbiol, 73, 6166-6171
Barbet AF et al. (2002), Expression of multiple outer membrane protein sequence variants from a single genomic locus of Anaplasma phagocytophilum, Infect Immun, 1706-1718.
Barbet AF et al. (2006), Structure of the Expression Site Reveals Global Diversity in MSP2 (P44) Variants in Anaplasma phagocytophilum, Infect Immun, 74, 6429–6437.
Boisaubert B, Boutin JM (1988), Le Chevreuil, Hatier : Faune Sauvage, 236p
Bown KJ et al. (2007), High-resolution genetic fingerprinting of European strains of Anaplasma phagocytophilum by use of multilocus variable-number tandem-repeat analysis, J Clin Microbiol,. 45, 1771-1776
Boyard C, Gasqui P, Barnouin J, Vourc’h G (2007), Comment diminuer le risque de maladies transmises par les tiques chez les bovins au pâturage, Bull GTV, 41, 67-72
Brouqui P, Ehrlichiose à Anaplasma phagocytophilum en France et en Europe In : Recueil du Colloque européen francophone Rickettsioses-Zoonoses et autres Arbo-bactérioses-zoonoses, ISPAIA Zoopôle Saint-Brieuc-Ploufragan, 11 et 12 septembre 2003, 49
Campbell et al. (1994), Sequential pathology of tick-borne fever, J Comp Pathol, 111, 303-313
Carlyon JA, Latif DA, Pypaert M, Lacy P, Fikring E (2004), Anaplasma phagocytophilum utilizes multiple host evasion mechanisms to thwart NADPH oxidase-mediated killing during neutrophil infection, Infect Immun, 72, 4772-4783

86
Casey ANJ et al. (2004), Groupings of highly similar major surface protein (p44)-encoding paralogues: a potential index of genetic diversity amongst isolates of Anaplasma phagocytophilum, Microbiology, 150, 727-734
Chevallier B, Civelli B, Blanchard B (2003), Développement d’un test PCR permettant la détection d’Anaplasma phagocytophilum In : Recueil du Colloque européen francophone Rickettsioses-Zoonoses et autres Arbo-bactérioses-zoonoses, ISPAIA Zoopôle Saint-Brieuc-Ploufragan, 11 et 12 septembre 2003, 56-57
Chimier AME (2005). Contribution à l’étude de la prévalence de l’infection à Anaplasma phagocytophilum dans la faune sauvage en France, Thèse Med. Vet, Ecole Nationale Vétérinaire de Maisons-Alfort, Créteil
Davoust B (2003), Anaplasma phagocytophilum chez le chien et le chat en France et en Europe In : Recueil du Colloque européen francophone Rickettsioses-Zoonoses et autres Arbo-bactérioses-zoonoses, ISPAIA Zoopôle Saint-Brieuc-Ploufragan, 11 et 12 septembre 2003, 48
De la Fuente J et al. (2007), Evidence of Anaplasma infections in European roe deer (Capreolus capreolus) from southern Spain, Res Vet Science, doi:10.1016/j.rvsc.2007.05.018
De la Fuente J et al. (2005), Sequence analysis of the msp4 gene of Anaplasma phagocytophilum strains, J Clin Microbiol, 43, 1309-1317
Delorme D (2007), La longueur de la patte arrière : un indicateur fiable du suivi des populations de chevreuils en forêts, Sup Faune Sauvage, 275, Fiche Technique n°97
Delorme D, Bédarida G, Van Laere G (2008), De la MAC à la MEC : de la mortalité anormale du chevreuil à la mortalité expliquée du chevreuil, Grande Faune, 117,23-30
Delorme D, Van Laere (2007), Le Chevreuil, Belin : Eveil Nature, 87p
Delorme D, Van Laere G, Gaillard JM, Klein F (2007), La phase juvénile chez le chevreuil, Faune sauvage, 275, 15-22
Drazenovich N, Foley J, Brown RJ (2006), Use of Real-time Quantitative PCR targeting the msp2 Protein Gene to identify cryptic Anaplasma phagocytophilum infections in wildlife and domestic animals, Vector-borne and zoonotic diseases, 6, p83-90
Dumler JS et al (2005), Human Granulocytic Anaplasmosis and Anaplasma phagocytophilum, Emerg Infect Dis, 11, 1828-34.
Dumler JS et al. (2001), Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and 'HGE agent' as subjective synonyms of Ehrlichia phagocytophila, Int J Syst Evol Microbiol., 51, 2145-2165
Foley J, Nieto N (2006), Anaplasma phagocytophilum subverts tick salivary gland proteins, Trends parasitol, 23, p3-5
Garyu JW, Dumler J S (2005), Anaplasma phagocytophilum infection reduces expression of phagocytosis-related receptors on neutrophils, Ann N Y Acad Sci, 1063, 416-9
Gokce HI, Woldehiwet Z, (1999), Differential haematological effects of tick-borne fever in sheep and goats, Zentralbl Veterinarmed B, 46, p 105-115
Gokce HI, Woldehiwet Z, (1999), The effects of ehrlichia (Cytoecetes) phagocytophila on the clinical chemistry of sheep and goats, Zentralbl Veterinarmed B, 46, p 93-103

87
Granquist EG, Stuen S, Lundgren AM, Bråten M, Barbet AF (2008), Outer Membrane Protein Sequence Variation in Lambs Experimentally Infected with Anaplasma phagocytophilum, Infect Immun, 76, 120–126.
Guetard M (2001), Ixodes ricinus : morphologie, biologie, élevages, données bibliographiques, Thèse Med.Vet, Ecole Nationale Vétérinaire de Toulouse, Toulouse
Hesler AS (2006), Etude des populations d’ongulés sauvages et relation avec la présence du Lynx (Lynx lynx) en Franche-Comté, Mémoire de stage de master 2, 56p
Inokuma H, Brouqui P, Drancourt M, Raoult D (2001), Citrate Synthase Sequence: a new tool for phylogenetic analysis and identification of Ehrlichia, J Clin Microbiol, 39, 3031-3039
Inokuma H, Terada Y, Kamio T, Raoult D, Brouqui P (2001), Analysis of the 16S rRNA Gene Sequence of Anaplasma centrale and Its Phylogenetic Relatedness to Other Ehrlichiae, Clin Diagn Lab Immunol, 8, 241–244
Jason A et al. (2003), Murine neutrophils require 1,3-fucosylation but not PSGL-1 for productive infection with Anaplasma phagocytophilum, Blood, 102, 3387-95.
Jolibois C (2007), Chercher les corps jaunes, Plaisirs de la chasse, 665, 30-31
Joncour G (2003), Rickettsioses-zoonoses et autres arbo-bactérioses-zoonoses Restitution des données (1999-2003) In : Recueil du Colloque européen francophone Rickettsioses-Zoonoses et autres Arbo-bactérioses-zoonoses, ISPAIA Zoopôle Saint-Brieuc-Ploufragan, 11 et 12 septembre 2003, 58-77
Joncour G (2007), L’ehrlichiose granulocytaire bovine/ovine à Anaplasma phagocytophilum et la faune sauvage, Bulletin des GTV, 40, p33_36
Joncour G, Collin E (2003), La diagnostic clinique de l’ehrlichiose bovine In : Recueil du Colloque européen francophone Rickettsioses-Zoonoses et autres Arbo-bactérioses-zoonoses, ISPAIA Zoopôle Saint-Brieuc-Ploufragan, 11 et 12 septembre 2003, 50-53
Joncour G, Pouliquen G, Kaufmann P, Mayaux P (2006), Anaplasma phagocytophilum, agent de l’ehrlichiose granulocytaire bovine (EGB) et d’avortements chez les bovins, proposition de protocole et d’aide au diagnostic, Bull GTV, 35, 95-104
Lin Q et al. (2002), Analysis of sequence and loci of p44 homologs expressed by Anaplasma phagocytophila in acutely infected patients, J Clin Microbiol, 40, 2981-2988
Lin Q et al. (2004), Anaplasma phagocytophilum has a functionnal msp2 gene that is distinct from p44, Infect Immun, 72, 3883-3889
Liz JS (1994), Ehrlichia phagocytophila : Aspects épidémiologiques, hématologiques et sérologiques de l’infection chez les bovins en Suisse, Thèse Sciences, Université Neuchâtel, Neuchâtel
Liz JS, Sumner JW, Pfister K, Brossard M (2002), PCR detection and serological evidence of granulocytic ehrlichial infection in Roe deer (Capreolus capreolus) and Chamois (Rupicapra rupicapra), J Clin Microbiol, 40, 892-897
Marti-Bizen E (2007), La mortalité anormale (ou normale ?) du chevreuil en France : état des lieux, avancée des travaux et propositions de recherches, Rapport de stage de titularisation de l’Ecole Nationale des Services Vétérinaires, 52p
Massung RF, Mather TN, Levin ML (2006), Reservoir competency of goats for the Ap-Variant 1 strain of Anaplasma phagocytophilum, Infect Immun, 74, 1373-1375
Reid HW et al. (1986), Response of sheep to experimental concurrent infection with tick-borne fever (Cytoecetes phagocytophila) and louping-ill virus, Res Vet Sci, 41, 56-62

88
Strle F (2004), Human granulocytic ehrlichiosis in Europe, Int J Med Microbiol, 293, p 27-35.
Stuen S et al. (2006), A paretic condition in an Anaplasma phagocytophilum infected Roe Deer Calf, Journal of wildlife diseases, 42, p 170-174
Stuen S, Akerstedt J, Bergström K, Handeland K (2002), Antibodies to granulocytic ehrlichia in moose, red deer, and roe deer in Norway, Journal of Wildlife Diseases, 38, pp. 1–6
Stuen S, Bergström K, Petrovec M, Van de Pol I, Schouls LM (2003), Differences in clinical manifestations and hematological and serological responses after experimental infection with genetic variants of Anaplasma phagocytophilum in sheep, , Clin Diagn Lab Immunol, 10, 692-695
Sumner JW, Nicholson WL, Massung RF (1997), PCR amplification and comparison of nucleotide sequences from the groESL heat shock operon of Ehrlichia species, J Clin Microbiol, 35, 2087-92.
Van Laere G, Toïgo C, Gaillard JM, Renaud G (2006), Les effets de la canicule de l’été 2003 sur le chevreuil, Faune Sauvage, 273, 15-19
Vassalo N, Lamanda P, Ehrlichiose bovine à A. phagocytophilum : Diagnostic de laboratoire In : Recueil du Colloque européen francophone Rickettsioses-Zoonoses et autres Arbo-bactérioses-zoonoses, ISPAIA Zoopôle Saint-Brieuc-Ploufragan, 11 et 12 septembre 2003, 54-55
Wallach A, Inbar M, Scantlebury M, Speakman J, Shanas U, Water requirements as a bottleneck in the reintroduction of European roe deer to the southern edge of its range, Canadian Journal of zoology, 85, 1182-1192
Weisburg WG, Barns SM, Pelletier DA, Lane DJ (1991), 16S ribosomal DNA amplification for phylogenetic study, J Bacteriol, 173, 697-703
Woldehiwet Z (1987), Depression of lymphocyte response to mitogens in sheep infected with tick-borne fever, J Comp Pathol, 97, 637-643
Woldehiwet Z (1991), Lymphocyte subpopulations in peripheral blood of sheep experimentally infected with tick-borne fever, Res Vet Sci, 51, 40-43
Woldehiwet Z (2007), Immune evasion and immunosuppression by Anaplasma phagocytophilum, the causative agent of tick-borne fever of ruminants and human granulocytic anaplasmosis, the veterinary journal, 175, 37-44
.II. Autres documents consultés
ADIAGENE, ADIAVET ANA PHA REALTIME, Test pour la détection des trois biovars d’Anaplasma phagocytophilum par amplification enzymatique de gène, Manuel d’instruction
ADIAGENE, ADIAVET ANA.PHA REALTIME (2008), Données de validation
.II.A. Sites internet consultés
Anonyme (2005), Manuel des tests de diagnostic et des vaccins pour les animaux terrestres de l'OIE 2005 URL: http://www.oie.int/fr/normes/mmanual/pdf_fr/Chapitre%20final05%202.3.7_Anaplasmose.pdf,
consulté le 13/12/2008
Bown KJ, Begon M, Bennett M, Woldehiwet Z, Ogden NH (2003), Seasonal dynamics of Anaplasma phagocytophila in a rodent-tick (Ixodes trianguliceps) system, United Kingdom, Emerg Infect Dis, URL: http://www.cdc.gov/ncidod/EID/vol9no1/02-0169.htm, consulté le 05/10/2008

89
Daniels T J, Battaly G R, Liveris D, Falco R C (2002) Avian reservoirs of the agent of human granulocytic ehrlichiosis? Emerg Infect Dis, URL: http://www.cdc.gov/ncidod/EID/vol8no12/01-0527.htm, consulté le 05/10/2008
Euzéby J (2008), Dictionnaire de bactériologie vétérinaire, http://www.bacterio.cict.fr/bacdico/aa/phagocytophila.html, consulté le 05/10/2008
.II.B. Logiciels utilisés
Arcview 9.2, ESRI
R version 2.6.1 (2007), The R Foundation for Statistical Computing

90

91
Table des matières
SOMMAIRE ...................................................................................................................................................... 1
LISTE DES FIGURES ........................................................................................................................................... 3
LISTE DES TABLEAUX ........................................................................................................................................ 5
LISTE DES ABBREVIATIONS ............................................................................................................................... 7
INTRODUCTION ................................................................................................................................................ 9
PREMIER PARTIE: ETUDE BIBLIOGRAPHIQUE .................................................................................................. 10
.I. BIOLOGIE DU CHEVREUIL ...................................................................................................................... 11
.I.A. LA REPARTITION DU CHEVREUIL EN FRANCE, L’EVOLUTION DES POPULATIONS DEPUIS 1973 ET LES HABITATS ............ 11 .I.A.1 REPARTITION DU CHEVREUIL EN FRANCE : UNE PREDILECTION POUR LES GRANDS MASSIFS
FORESTIERS DE L’EST ET DU SUD-OUEST ..................................................................................................... 11 .I.A.2 Evolution des populations de Chevreuil en France depuis 1973 .................................................... 12
I.A.2.a Evolution des plans de chasse depuis 1973 .............................................................................................. 12 I.A.2.b Plans de chasse pour la saison 2006-2007 ................................................................................................ 12
.I.B. MORPHOLOGIE, CRITERES D’AGES ET DE SEXE ............................................................................................... 14 .I.B.1 La tête du Chevreuil ....................................................................................................................... 14 .I.B.2 Etude des bois ................................................................................................................................ 14 .I.B.3 Détermination de l'âge du Chevreuil ............................................................................................. 15
.I.C. COMPORTEMENT SOCIAL ET REPRODUCTION ................................................................................................. 17 .I.C.1 Le cycle reproductif du Chevreuil ................................................................................................... 17
I.C.1.a La période de rut et les marquages .......................................................................................................... 17 I.C.1.b Du rut à la mise bas : onze longs mois ...................................................................................................... 18
.I.C.2 Rapports chevrette/faons : la phase critique juvénile ................................................................... 18 .I.D. ALIMENTATION DU CHEVREUIL, IMPACTS SUR LES CULTURES ET LES FORETS FRANÇAISES ........................................ 20
.I.D.1 Régime alimentaire du Chevreuil .................................................................................................. 20
.I.D.2 Dégâts aux cultures et aux forêts .................................................................................................. 21 .I.E. PRINCIPE DU SUIVI DES POPULATIONS DE CHEVREUILS ..................................................................................... 22
.I.E.1 Les indicateurs de présence ........................................................................................................... 22 I.E.1.a Empreintes ................................................................................................................................................ 22 I.E.1.b Les reposées ............................................................................................................................................. 22 I.E.1.c Grattis, frottis et régalis ............................................................................................................................ 23
.I.E.2 Les indicateurs de changement écologique ................................................................................... 23 I.E.2.a Conditions d’utilisation des indicateurs de changement écologiques (ICE) et qualités d’un ICE .............. 23 I.E.2.b L’indice kilométrique ................................................................................................................................ 24 I.E.2.c Le nombre de faons par femelle ............................................................................................................... 24 I.E.2.d Le suivi pondéral des animaux .................................................................................................................. 25 I.E.2.e La longueur de la mandibule et la longueur de la patte arrière ............................................................... 25 I.E.2.f Le taux d’utilisation de la végétation : les indices de pression sur la flore ............................................... 25
.II. ETUDE BIBLIOGRAPHIQUE DE A. PHAGOCYTOPHILUM .......................................................................... 26
.II.A. SYSTEMATIQUE ET BIOLOGIE ...................................................................................................................... 26 .II.A.1 Historique de la nomenclature d’A. phagocytophilum .................................................................. 26 .II.A.2 La problématique des variants d’A. phagocytophilum .................................................................. 27 .II.A.3 Données sur les caractéristiques biologiques d’A. phagocytophilum ............................................ 28
II.A.3.a Généralités .......................................................................................................................................... 28 II.A.3.b Morphologie et propriétés antigéniques de la bactérie ...................................................................... 28
.II.B. POUVOIR PATHOGENE D’A. PHAGOCYTOPHILUM ........................................................................................... 29 .II.B.1 Transmission vectorielle ................................................................................................................ 29 .II.B.2 Entrée dans l’organisme ................................................................................................................ 30 .II.B.3 Survie et multiplication dans les granulocytes .............................................................................. 30 .II.B.4 Virulence ........................................................................................................................................ 30

92
II.B.4.a Les protéines de surface ...................................................................................................................... 30 II.B.4.b Les mécanismes de survie d’A. phagocytophilum ............................................................................... 31
.II.C. L’EHRLICHIOSE GRANULOCYTAIRE ............................................................................................................... 31 .II.C.1 Epidémiologie ................................................................................................................................ 31
II.C.1.a Les hôtes d’A. phagocytophilum.......................................................................................................... 32 .II.C.2 Rôle de la tique Ixodes ricinus comme vecteur d’A. phagocytophilum .......................................... 34
II.C.2.a Biologie de la tique .............................................................................................................................. 34 II.C.2.b Répartition et habitats d’Ixodes ricinus ............................................................................................... 34 II.C.2.c Cycle biologique d’Ixodes ricinus .............................................................................................................. 34 II.C.2.d Interactions hôte-Ixodes ricinus .......................................................................................................... 36
.II.C.3 L’ehrlichiose : étude clinique chez les ruminants ........................................................................... 37 II.C.3.a Le diagnostic clinique de l’ehrlichiose bovine ..................................................................................... 37 II.C.3.b Signes cliniques chez les bovins ........................................................................................................... 38 II.C.3.c Signes cliniques chez la Chèvre ................................................................................................................. 38 II.C.3.d Les ehrlichioses sub-aiguës et chroniques ........................................................................................... 38
.II.C.4 Les modifications de la numération cellulaire et de la biochimie .................................................. 39 II.C.4.a Les modifications de la numération cellulaire ..................................................................................... 39 II.C.4.b Les modifications biochimiques .......................................................................................................... 40
.II.C.5 Etude des lésions d’ehrlichiose ..................................................................................................... 41 II.C.5.a Lésions macroscopiques d’ehrlichiose ................................................................................................ 41 II.C.5.b Histologie ............................................................................................................................................. 41
.II.D. DIAGNOSTIC DE LABORATOIRE ................................................................................................................... 41 .II.D.1 Diagnostic direct d’ehrlichiose ..................................................................................................... 41
II.D.1.a La cyto-hématologie : une méthode simple et rapide ......................................................................... 41 II.D.1.b La PCR .................................................................................................................................................. 42
.II.D.2 Diagnostic indirect de l’ehrlichiose ................................................................................................ 42 .II.E. DONNEES SUR L’EHRLICHIOSE CHEZ LE CHEVREUIL .......................................................................................... 43
DEUXIEME PARTIE: ETUDES EXPERIMENTALE ET RETROSPECTIVE .................................................................. 43
.I. CADRE ET OBJECTIFS DE CETTE ETUDE ................................................................................................... 45
.I.A. CADRE DE L’ETUDE .................................................................................................................................. 45
.I.B. OBJECTIFS.............................................................................................................................................. 45
.II. ETUDE EXPERIMENTALE : ETUDE DU ROLE PATHOGENE D’A. PHAGOCYTOPHILUM DANS LA
POPULATION DE CHEVREUILS DE FRANCHE-COMTE SUR LA SAISON DE CHASSE 2007/2008 .......................... 45
.II.A. RAPPEL DES OBJECTIFS ............................................................................................................................. 45
.II.B. MATERIEL ET METHODES DE L’ETUDE EXPERIMENTALE .................................................................................... 46 .II.B.1 LA FRANCHE-COMTE COMME SITE D’ETUDE ................................................................................ 46 .II.B.2 Echantillonnage ............................................................................................................................. 46
II.B.2.a Les chevreuils prélevés ........................................................................................................................ 46 II.B.2.b Nombre de chevreuils à prélever ........................................................................................................ 47
.II.C. NATURE DES PRELEVEMENTS ..................................................................................................................... 48 .II.C.1 La Pcr comme méthode de detection directe d’A. phagocytophilum ............................................ 50
II.C.1.a Utilisation du kit ADIAVET® ANA PHA REALTIME comme technique de PCR temps-réel .................... 50 II.C.1.b Qualités du test ADIAVET® ANA PHA REALTIME ................................................................................. 50 II.C.1.c Utilisation du kit ADIAVET ANA PHA REALTIME ........................................................................................ 50 II.C.1.d Utilisation du kit ADIAVET ANA PHA REALTIME en PCR semi-quantitative ......................................... 51
.II.C.2 Diagnostic indirect de l’ehrlichiose : l’immunofluorescence indirecte ........................................... 51
.II.C.3 Analyse des données de suivi sur les populations de chevreuils .................................................... 51
.II.C.4 Traitement des données ................................................................................................................ 52
.III. RESULTATS ....................................................................................................................................... 52
.III.A.1 Réalisations, nombre de chevreuils prélevés ............................................................................ 52
.III.A.2 Comparaison des résultats dans les deux classes malade et sain. ........................................... 53 III.A.2.a Résultats de l’ensemble des analyses .................................................................................................. 53 III.A.2.b Résultats de la comparaison malade/sain en PCR sur sang ................................................................. 53 III.A.2.c Résultats de la comparaison malade/sain en PCR tous tissus confondus ........................................... 54 III.A.2.d Résultats de la comparaison malade/sain en immuno-fluorescence indirecte ................................... 55
.III.A.3 Comparabilité entre les deux échantillons ................................................................................ 56

93
III.A.3.a Dates de prélèvements ........................................................................................................................ 56 III.A.3.b Composition en âge et en sexe des deux classes malade et sain ........................................................ 57 III.A.3.c Lieux de prélèvement .......................................................................................................................... 58
.III.A.4 Autres données ......................................................................................................................... 60 III.A.4.a Croisement des réponses à l’immuno-fluorescence et à la PCR .......................................................... 60 III.A.4.b Résultats obtenus par PCR semi-quantitative ..................................................................................... 61 III.A.4.c Répartition temporelle des chevreuils positifs .................................................................................... 62 III.A.4.d Répartition géographique des chevreuils positifs ............................................................................... 62
.III.A.5 Résultats obtenus pour les autopsies et les tiques ................................................................... 64 III.A.5.a Les autopsies ....................................................................................................................................... 64 III.A.5.b Les tiques ............................................................................................................................................. 64
.III.B. ETUDE RETROSPECTIVE DE LA DYNAMIQUE DES POPULATIONS DE CHEVREUILS DE FRANCHE-COMTE, DES CONDITIONS
CLIMATIQUES ET DES MILIEUX ................................................................................................................................... 65 .III.B.1 Objectifs .................................................................................................................................... 65 .III.B.2 Ressources utilisées pour le suivi .............................................................................................. 65 .III.B.3 Résultats de l’étude rétrospective ............................................................................................ 65
III.B.3.a Historique de l’évolution des populations et de leurs conditions de vie ............................................. 65 III.B.3.b Evolution de la santé des populations de chevreuils sur les sites Equilibre Faune Flore .................... 66
DISCUSSION ................................................................................................................................................... 66
.I. ETUDE CAS-TEMOIN : UN PORTAGE PLUS IMPORTANT CHEZ LES MALADES .......................................... 69
.I.A. LES CHEVREUILS MALADES SONT PLUS PORTEURS DE LA BACTERIE QUE LES CHEVREUILS SAINS ................................. 69 .I.A.1 Interprétation des résultats de l’étude Cas témoins ...................................................................... 69 .I.A.2 Qualités des analyses utilisées ...................................................................................................... 69 .I.A.3 Arguments biologiques en favuer de la pathogenicite d’A. phagocytophilum .............................. 70
I.A.3.a L’ensemble de espèces proches du Chevreuil est sensible à la bactérie .................................................. 70 I.A.3.b L’ehrlichiose granulocytaire est difficile diagnostiquer même en élevage ............................................... 71 I.A.3.c Effet immunodépresseur potentiel .......................................................................................................... 72
.I.A.4 Validité du modèle......................................................................................................................... 72 I.A.4.a Une sous estimation de l’âge .................................................................................................................... 72 I.A.4.b Répartitions géographique et temporelle ................................................................................................ 73
.II. DES PREVALENCES ET SEROPREVALENCE ELEVEES ET UNE CIRCULATION HOMOGENE .......................... 74
.II.A. DE NOMBREUX CHEVREUILS POSITIFS .......................................................................................................... 74
.II.B. UNE REPARTITION HOMOGENE SANS FOYERS DE POSITIVITE ............................................................................. 74
.III. UNE SAISON 2007/2008 MARQUEE PAR UNE FAIBLE MORTALITE .................................................... 75
.IV. UNE BACTERIE RETROUVEE TOUT AU LONG DE L’HIVER DANS LE SANG DES CHEVREUILS ................ 76
.V. LE ROLE POTENTIEL DU CHEVREUIL DANS LE CYCLE EPIDEMIOLOGIQUE D’A. PHAGOCYTOPHILUM ...... 76
.VI. INTEGRATION DES DONNEES DE SUIVI DES POPULATIONS ............................................................... 77
.VI.A. IMPACTS DES EVENEMENTS CLIMATIQUES SUR LES POPULATIONS DE CHEVREUILS ................................................. 77
.VI.B. MECANISMES EXPLICATIFS DES REPERCUSSIONS DES MAUVAISES CONDITIONS DE MILIEUX ...................................... 78
.VI.C. RELATIONS ENTRE MALADIES ET CONDITIONS DU MILIEU ................................................................................. 79
.VII. LE ROLE PATHOGENE D’A. PHAGOCYTOPHILUM DANS LES POPULATIONS DE CHEVREUILS DE
FRANCHE-COMTE EN 2007/2008 ET AVANT ................................................................................................... 79
.VII.A. IMPACT POTENTIEL DE LA PRESENCE D’A. PHAGOCYTOPHILUM.......................................................................... 80
.VII.B. MORTALITE NATURELLE ............................................................................................................................ 80
CONCLUSION .................................................................................................................................................. 79
BIBLIOGRAPHIE .............................................................................................................................................. 80
.I. REFERENCES BIBLIOGRAPHIQUES .......................................................................................................... 85
.II. AUTRES DOCUMENTS CONSULTES ........................................................................................................ 88
.II.A. SITES INTERNET CONSULTES ....................................................................................................................... 88
.II.B. LOGICIELS UTILISÉS .................................................................................................................................. 89

94
TABLE DES MATIERES ..................................................................................................................................... 85
ANNEXES ........................................................................................................................................................ 89
.I. ANNEXE 1: QUESTIONNAIRE PROPOSE LORS DES PRELEVEMENTS DES CHEVREUILS ............................. 96
.II. ANNEXE 2: LES COURBES DE PCR TEMPS-REEL ...................................................................................... 97
.III. ANNEXE 3 : LISTE DES LABORATOIRES ET ORGANISMES AYANT PARTICIPE A L’ETUDE ..................... 98
.IV. ANNEXE 4 : FEDERATIONS DE CHASSEURS AYANT PARTICIPE A L’ETUDE .......................................... 99
.V. ANNEXE 5 : EXEMPLE D’UN COMPTE RENDU DE SUIVI DES POPULATIONS DE CHEVREUILS (EXEMPLE
SITE EQFF DE L’AIGUILLON) .......................................................................................................................... 100

95
ANNEXES

96
.I. Annexe 1: Questionnaire proposé lors des prélèvements des chevreuils
97
.II. Annexe 2: Les courbes de PCR temps-réel
COURBES IPC (CONTROLE INTERNE GAPDH)
COURBES ANA PHA GAMME QUANTIFICATION
DROITE REGRESSION GAMME QUANTIFICATION ANA PHA
COURBES ANA PHA ECHANTILLONS + GAMME ANA PHA QUANTIFICATION

98
.III. Annexe 3 : Liste des laboratoires et organismes ayant participé à l’étude
Laboratoire départemental des côtes d’Armor
Contact : Natalie Vassalo
Zoopôle, 3, rue du Sabot, BP 54 22 440 PLOUFRAGAN,
Tél. : 02 96 01 37 22, Fax : 02 96 01 37 50
Laboratoire départemental de Sèvre-Atlantique LASAT
Contact : Philippe Nicollet
210 avenue de la Venise Verte - 79022 NIORT Cedex
Tél. : 05 49 17 10 57 Fax : 05 49 09 08 70
Laboratoire départemental du Doubs
Contact : Marc Hesseman
13, rue Gay-Lussac, BP 1981, 25 020 BESANÇON CEDEX,
Tél. : 03 81 25 88 50, Fax : 03 81 25 88 51
Laboratoire Départemental d'Analyses du Jura
Contact : Alain Viry
59 RUE DU VIEIL HOPITAL
39800 Poligny
Téléphone : 03.84.73.73.40
Fax : 03.84.37.12.14
Laboratoire départemental de Haute-Saône
Contact : Pierre Lehong
29, Rue Lafayette
70000 Vesoul
03 84 76 70 03

99
.IV. Annexe 4 : Fédérations de chasseurs ayant participé à l’étude
Fédération nationale des chasseurs
Contact : DMV Charlotte Dunoyer
13 rue du Général Leclerc
92136 ISSY LES MOULINEAUX CEDEX
Tél. : 01 41 09 65 10
Fax. : 01 41 09 65 20
Fédération régionale des chasseurs de Franche Comté
Contact : Charlette Chandosné
26 A route de Lyon25720 BEURE
Tél : 03 81 51 06 10 –
Fédération départementale des chasseurs du Doubs
Contact : David Clerc
Rue du Châtelard
25360 GONSANS
Tél. : 03.81.61.23.87 –
Fax : 03.81.61.21.13
Fédération départementale des chasseurs du Jura
Contact : Mickaël Marillier
Maison de la Nature et de la Faune Sauvage
Rue de la Fontaine Salée
39140 ARLAY
Tél. : 03 84.85.19.19
Fax : 03.84.85.19.10
Fédération départementale des chasseurs de Haute Saône
Contact : Romain Martin
Rue de Verdun
70000 NOIDANS LES VESOUL
Tél. : 03.84.97.13.53
Fax : 03.84.75.08.92
Fédération départementale des chasseurs du Territoire de Belfort
Contact : Jérôme Demeulemester
6, rue Denfert Rochereau
90000 BELFORT
Tél. : 03.84.22.28.71
Fax : 03.84.54.02.54

100
.V. Annexe 5 : Exemple d’un compte rendu de suivi des populations de chevreuils (exemple site EqFF de l’Aiguillon)
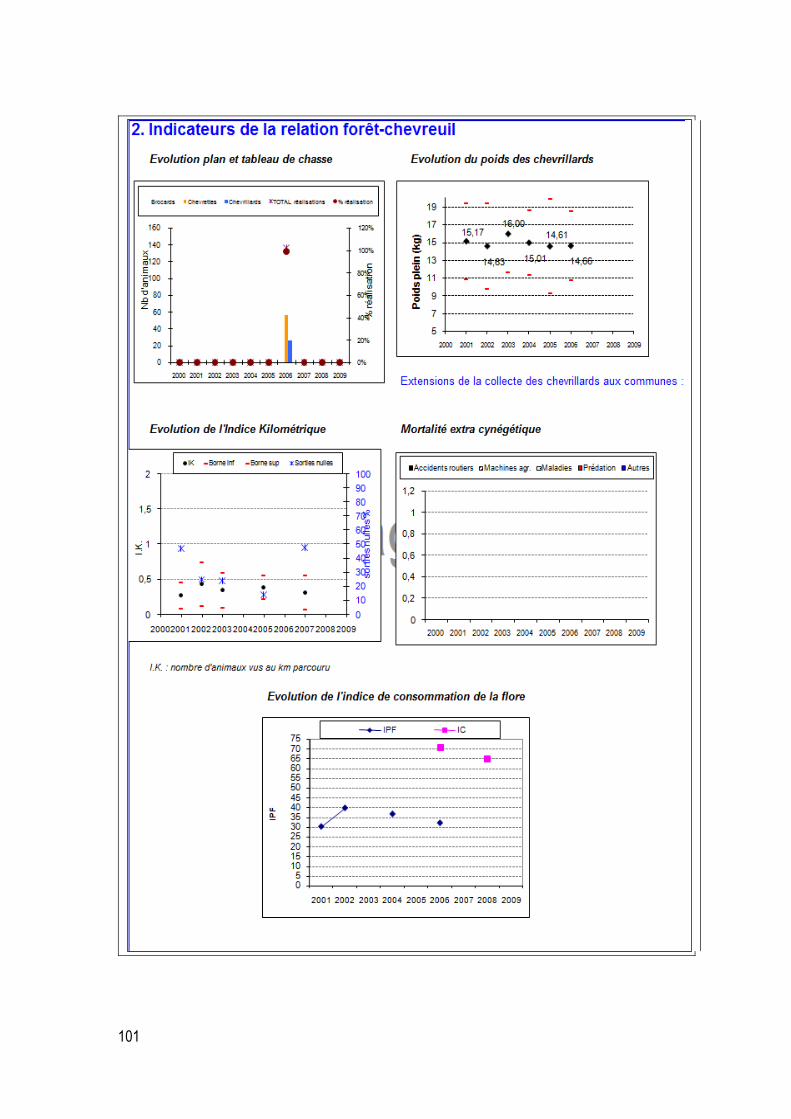
101
102

Etude de la pathogénicité d’Anaplasma phagocytophilum chez les populations de
chevreuils (Capreolus capreolus) de Franche Comté
Julien PORTIER
Résumé
Cette étude cherche à mettre en évidence le rôle pathogène joué par Anaplasma phagocytophilum, dans des problèmes de mortalité et d’altération de la dynamique des populations chez des populations de chevreuils en Franche-Comté. Il s’agit d’une étude cas-témoin comparant le portage de la bactérie et l’immunité envers cette bactérie entre des chevreuils malades et des chevreuils sains. Cette étude met en évidence un portage plus important chez les chevreuils malades. Il est probable que Anaplasma phagocytophilum provoque une maladie chez le chevreuil qui pourrait se manifester par un effet immunosupresseur dont l’impact serait non négligeable chez des chevreuils en mauvais état. La conclusion de ce travail est qu’il est possible qu’Anaplasma phagocytophilum ait eu un impact sur les populations de chevreuils mais que ce n’est pas le seul agent responsable et que ce sont des mauvaises conditions du milieu qui déclenchent le genre de problèmes observés.
Mots clés : Chevreuil, Franche-Comté, Anaplasma phagocytophilum, Etude expérimentale, Etude Cas-témoin, Dynamique des populations, Agent pathogène
Jury :
Président : Pr. Michel Marjolet
Directeur : Dr. Suzanne Bastian-Orange
Assesseur : Dr. Raphaël Guattéo
Adresse de l’auteur :
M Julien Portier
106 avenue Jules Vallès
38100 Grenoble


Study of the pathogenic potential of Anaplasma phagocytophilum in Roe deer (Capreolus
capreolus) populations of the Franche-Comté region
Julien PORTIER
Summary
The Roe Deer (Capreolus capreolus) is a wild ungulate which has a great importance in the French hunting tradition. During the last fifteen years, unusually high death rates were observed on different hunting grounds in France that shared the same characteristics. Besides these deaths, roe deer population dynamics have been altered in these regions. It is now assumed that these deaths are not due to one pathogen but that they might be due to a series of pathogens. Many experts suppose that troubled relations between roe deer populations and their environment might trigger the problems observed.
This case-control study explores the possibility that the bacteria Anaplasma phagocytophilum plays a part in these deaths through a comparative serological and PCR study between healthy and ill Roe deer in the Franche-Comté region. Data on the survey of Roe deer populations and meteorological data were also collected.
The PCR results show that the prevalence of Anaplasma phagocytophilum is significantly higher in ill Roe deer. This could indicate that Anaplasma phagocytophilum can provoke illness in roe deer. A high prevalence shows that the bacteria is widely spread over the region and that it can have an impact on roe deer populations.
There is a strong suspicion that the 2003 drought triggered the disorders observed in roe deer population and that the visible manifestation of these disorders was a high death rate due to a variety of pathogens including Anaplasma phagocytophilum.
Key-words : Roe deer, Franche-Comté, Anaplasma phagocytophilu, Exerimental study, Case-control study, Population dynamics, pathogen
Jury :
Président : Pr. Michel Marjolet
Directeur : Dr. Suzanne Bastian-Orange
Assesseur : Dr. Raphaël Guattéo
Adresse de l’auteur :
M Julien Portier
106 avenue Jules Vallès
38100 Grenoble





























