Embed Size (px)
Citation preview

Étude descriptive et expérimentale du comportement de capture de Ranatra linearis au coursde son ontogenèseAuthor(s): Ann CloarecSource: Behaviour, Vol. 35, No. 1/2 (1969), pp. 84-113Published by: BRILLStable URL: http://www.jstor.org/stable/4533297 .
Accessed: 25/06/2014 00:43
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
BRILL is collaborating with JSTOR to digitize, preserve and extend access to Behaviour.
http://www.jstor.org
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

]TUDE DESCRIPTIVE ET EXPERIMENTALE DU COMPORTEMENT DE CAPTURE DE RANATRA LINEARIS
AU COURS DE SON ONTOGEN~SE
par
ANN CLOAREC
(Laboratoire d'Ethologie, Faculte des Sciences de Rennes, France)
(Avec 7 Figures) (Regu 6 - III - 1969)
SOMMAIRE
I - Introduction . . . . . . . . . . . . . . . . 85 II - Materiel et methodes . . . . . . . . . . . . . .. 85
III - Description du comportement de capture des proies et d'alimentation . . 86
A) Comportement complet . . . . . . . . . . . 86 a) Posture de preparation a la capture . .. 86
Position du corps par rapport au support . .. 86 Position des pattes par rapport au corps . .. 87
b) Prise d'intert . . . . . . . . . .. 87 c) Orientation . . . .. 88 d) Capture proprement-dite . . . . .. 89 e) Consolidation de la prise . . . .. go f) Exploration rostrale .. go g) Piqfire et succion . . . . .. 9I h) Fin du repas . . . . . . .. 9
B) Variations dans le comportement de capture. . 91 C) Ontogenese du comportement de capture et d'alimentation . . . . 94
a) Posture de preparation a la capture . . . .94 b) Capture . . . . . .95 c) Alimentation . . . . . . .. 96 d) Ontogenese. 96
IV - Etude experimentale des facteurs de causalite de la capture des proies. . 103 A) Caracteristiques optiques et mecaniques d'une proie . I03
a) Forme et taille de la proie ... 103 b) Couleur . . . . ... 04 c) Mouvement . . ... 104 d) Poids et nature . . . ... I09
e) Action repere du leurre . . . . . . Io9 B) Espace efficace . . . . . . . . . . . . 09
V - Conclusion .. IIo Resume . . . . . . . . . . . Ouvrages cites . . . . . . . . . .. Summary. 112
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA 85
I. INTRODUCTION
Dans le present travail, nous nous proposons d'etudier quelques aspects du comportement alimentaire de la Ranatre (Ranatra linearis Fab.). Cet
heteroptere aquatique frequente les nombreuses pieces d'eau de la region, ou les conditions ecologiques sont relativement stables et bien definies. Ceci nous a permis d'entretenir au laboratoire un elevage dans lequel ces individus
pouvaient etre constamment compares aux Insectes des biotopes naturels. La Ranatre a servi d'objet a de nombreux travaux. Citons parmi eux ceux
de la TORRE-BUENO (I906), HOLMES (I906), POISSON (I924), HOFFMAN
(I930), LARSEN (I936) et RAGHUNATHO RAO (I962) pour la biologie et le
cycle de developpement; ceux de DUFOUR (I820), LOCY (I884), MARSHALL et SEVERIN (1904), BAUNACKE (I912), NEISWANDER (I926) et l'importante ceuvre de LARSEN (I949) pour la morphologie et l'anatomie; ceux de HOLMES
(1905) sur les reactions a la lumiere et de ABBOTT (I940) sur les reactions a des stimuli de contact apres diverses operations effectuees sur la chaine nerveuse. Aucun de ces auteurs n'a etudie de maniere precise le comporte- ment alimentaire. Certains, cependant, signalent que la Ranatre chasse a l'afftut et saisit sa proie a l'aide de la premiere paire de pattes transformee en organe prehensile.
Dans ces conditions, et compte tenu du fait d'etudes de comportement alimentaire effectuees sur des Insectes predateurs appartenant au meme milieu que la Ranatre (Odonates) ou possedant des organes de predation homologues (Mantides), il nous a paru interessant de poursuivre une analyse methodique de ce comportement chez la Ranatre. Nous nous sommes place dans l'optique de notre laboratoire et nous tentons une etude neurophysiolo- gique de la regulation du comportement en comparant les animaux a eux- memes au cours de leur ontogenese.
La presente note regroupe toutes nos observations de base sur l'adulte et les jeunes dans les conditions naturelles et dans les conditions experimentales d'utilisation de leurres.
II. MATERIEL ET MRTHODES
Les imagos recoltes dans la nature sont maintenus au laboratoire soit en isolement dans de petits bacs parallelipipediques (IO X 20 X 15 cm), soit
par couples dans de grands aquariums (80 X 30 X 40 cm). Ils sont nourris une fois par semaine en general avec des larves de chironomes, mais ils peuvent manger toutes sortes de petits animaux aquatiques passant a leur portee. L'accouplement a lieu en avril - mai; les larves eclosent environ 3 semaines apres la ponte si les oeufs sont maintenus a une temperature de
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

ANN CLOAREC
20-22°C. (le taux d'eclosion est actuellement plus faible malheureusement
pour les oeufs pondus au laboratoire que pour les ceufs recoltes dans la
nature). Des leur eclosion, les larves sont placees par petits groupes dans des
recipients de Io X Io X I6 cm dont la hauteur de remplissage en eau n'excede pas 2 a 3 cm. Nous leur fournissons une grande quantite d'organis- mes planctoniques. Dans ces conditions le cannibalisme est rare. Le develop- pement se fait en 5 stades larvaires; pour une observation plus facile, a
partir du 4e stade les larves sont isolees dans des bocaux de 7 X 7 X I I cm; elles sont alors nourries de larves de chironomes. Pour une temperature de 2I°, un rythme d'eclairage I2h/I2h, et une nourriture abondante les durees des stades ont ete:
TABLEAU I
Duree de developpement
Stade Duree moyenne minimale maximale (en jours)
I 6 5 8 2 7 5 9 3 9 5 II
4 9 6 12 5 12 8 15
III. DESCRIPTION DU COMPORTEMENT DE CAPTURE DES PROIES ET D'ALIMENTATION
L'etude de ce comportement a ete faite soit par observation directe soit
par analyse cinematographique (64 imageslseconde). Dans ce dernier cas les films etaient pris a l'aide de deux cameras perpendiculaires qui assuraient la
possibilite de comparer deux aspects d'une meme sequence de capture (en general de profil et par dessus). Cette methode nous a permis de preciser les positions relatives des articles des pattes ravisseuses aux divers instants
caracteristiques de la capture.
A. COMPORTEMENT COMPLET
a. Posture de preparation a la capture.
Position du corps par rapport au support. La Ranatre se tient en general sur une plante aquatique, les conditions
nec,ssaires d'un support etant la quasi immobilite et une rigidite suffisante
pour resister au poids de l'animal. La Ranatre est subparallele au support; elle est en general au-dessus de lui avec la tete plus basse que l'abdomen; la tete se situe toujours dans le prolongement du thorax. Cette position est
86
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA
independante de la hauteur d'eau recouvrant l'insecte (dans certaines con- ditions, en captivite au moins, la Ranatre peut se tenir juste sous la surface de l'eau sans l'aide d'un support solide).
Position des pattes par rapport au corps. Le Figure I precise les divers elements angulaires caracteristiques de
cette position dans la periode preparatoire a la capture. Les tarses meso et
metathoraciques servent de points d'appui, tandis que les pattes prothoraci- ques ne touchent nulle part.
V H
H S
A \; H k -/S
Figure I. Position d'attente en vue verticale (V.) et en vue horizontale (H.) indiquant les differentes valeurs angulaires en degres. S = Support.
V H
Patte SA = angle tibia-femur. prothoracique B = angle femur-coxa.
Patte mesothoracique
Patte metathoracique
C = angle corps-femur. D = angle femur-tibia.
/E = angle tibia-tarse.
F = angle corps-femur. G = angle femur-tibia. H = angle tibia-tarse.
95° (90o-0oo°) 75° (70°-90°)
500 (40°-70°) 65° (40°-8o°) 120° (8o°-i70°)
90° (8o°-i200) 45° (30°-70°) 1700 (80o°-700)
90° (90°-ioo00) 90° (80°-io0°)
60° (2o° -6°) 60° (40-900°) 1700 (80o°-700)
20° (00-30°)
I35° (90°-I70°) I45° (8o°-I7o0)
b. Prise d'intere,t.
Des qu'une proie potentielle est per;ue par la Ranatre, celle-ci redresse
legerement les femurs prothoraciques qu'elle rapproche de son corps: dimi- nution de l'angle forme par l'articulation femurlcoxa.
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

88 ANN CLOAREC
c. Orientation. (v. Figures 2 et 3).
Apres un delai plus ou moins long, s'observent:
- un changement de position de tout le corps de la Ranatre par un mouvement lent (exemple: remontee de la tete de 5 mm en 2 seconde) qui amene l'axe sagittal dans la direction de la proie. Ce sont les pattes meso et metathoraciques qui interviennent dans ce mouvement en modifiant les
A A
::::::. :::
Figure 2. Schemas montrant differentes postures observees au cours d'une capture. Les grandes fleches marquent l'enchainement. Les petites montrent le mouvement effectue.
Les lettres majuscules sont les memes que celles employees sur la Figure 3. A = position de repos. images I a 42. B = prise d'interet. duree 31i6e" images 42 a 61. C = orientation du corps. duree = I sec. puis attente. images 61 a 151. D = orientation des extremites des femurs et des tibio-tarses. duree = o,I sec. images 151 a 157. E = capture proprement dit = saisie de la proie par la patte gauche. duree = 0,03". images 157 a I59. F = amortissement du mouvement = abaissement des femurs et du corps. duree - 0,I sec. images I59-160. G = preparation de la saisie par la 2e patte: orientation du femur de la patte droite puis ouverture du tibio-tarse droit. duree = 0,I sec. images 160 a i66. H = orientation du tibio-tarse droit. duree = o,o6 sec. mages 166 a I70. I = saisie de la proie par la patte droite. duree = 47 millisec. images 170 a 173. J = mouvement d'approche de la proie du rostre. duree = 20" en moyenne. images: a partir de I73. K = exploration rostrale. duree tres variable: de quelques secondes a plusieurs minutes; puis, toujours dans la meme position, piqure et succion.
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions
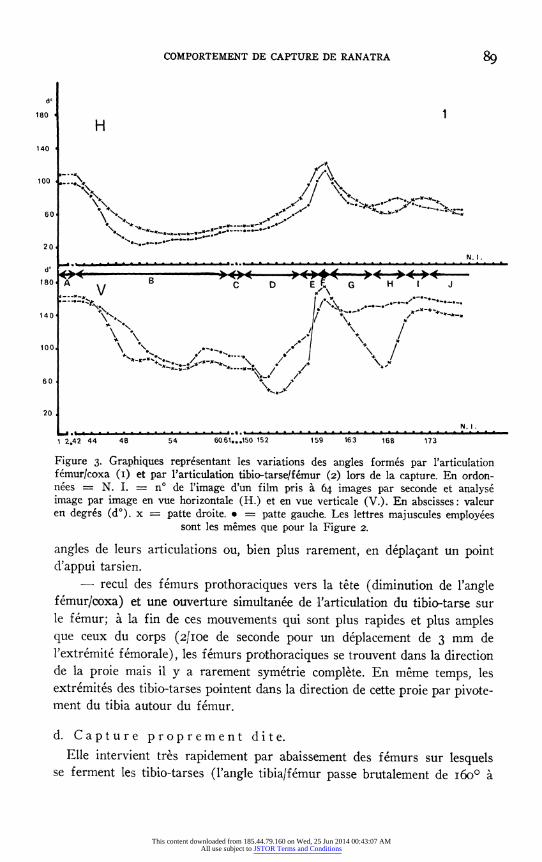
COMPORTEMENT DE CAPTURE DE RANATRA
1 H
1 2.42 44 48 54 60.61...150 152 159 163 168 173
Figure 3. Graphiques representant les variations des angles formes par l'articulation femur/coxa (I) et par l'articulation tibio-tarse/femur (2) lors de la capture. En ordon- nees = N. I. = n° de l'image d'un film pris a 64 images par seconde et analyse image par image en vue horizontale (H.) et en vue verticale (V.). En abscisses: valeur en degres (d°). x = patte droite. · = patte gauche. Les lettres majuscules employees
sont les memes que pour la Figure 2.
angles de leurs articulations ou, bien plus rarement, en depla;ant un point d'appui tarsien.
- recul des femurs prothoraciques vers la tete (diminution de l'angle femur/coxa) et une ouverture simultanee de l'articulation du tibio-tarse sur le femur; a la fin de ces mouvements qui sont plus rapides et plus amples que ceux du corps (2/Ioe de seconde pour un deplacement de 3 mm de l'extremite femorale), les femurs prothoraciques se trouvent dans la direction de la proie mais il y a rarement symetrie complete. En meme temps, les extremites des tibio-tarses pointent dans la direction de cette proie par pivote- ment du tibia autour du femur.
d. Capture proprement dite. Elle intervient tres rapidement par abaissement des femurs sur lesquels
se ferment les tibio-tarses (l'angle tibia/femur passe brutalement de 6o0° a
180
140
100
89
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions
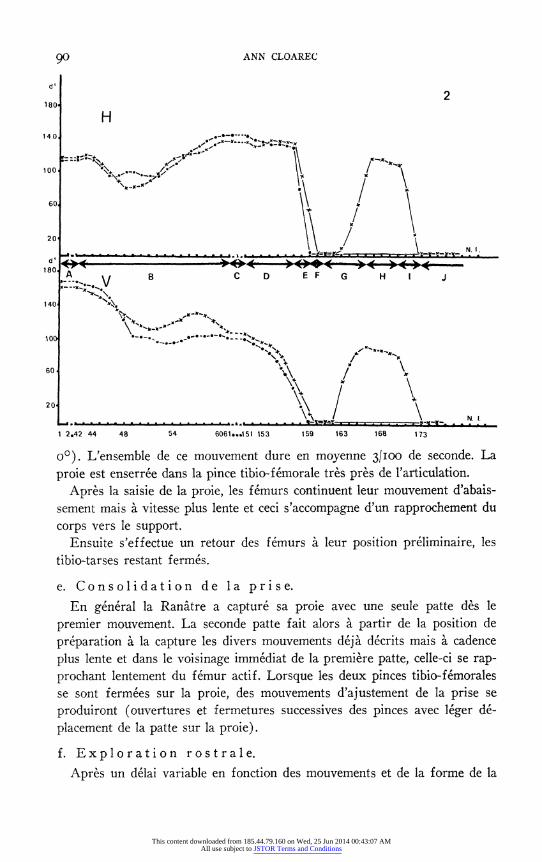
ANN CLOAREC
1 2.42 44 48 54 6061...151 153 159 163 168 173
o°). L'ensemble de ce mouvement dure en moyenne 31Ioo de seconde. La
proie est enserree dans la pince tibio-femorale tres pres de l'articulation. Apres la saisie de la proie, les femurs continuent leur mouvement d'abais-
sement mais a vitesse plus lente et ceci s'accompagne d'un rapprochement du
corps vers le support. Ensuite s'effectue un retour des femurs a leur position preliminaire, les
tibio-tarses restant fermes.
e. Consolidation de la prise. En general la Ranatre a capture sa proie avec une seule patte des le
premier mouvement. La seconde patte fait alors a partir de la position de
preparation a la capture les divers mouvements deja decrits mais a cadence
plus lente et dans le voisinage immediat de la premiere patte, celle-ci se rap- prochant lentement du femur actif. Lorsque les deux pinces tibio-femorales se sont fermees sur la proie, des mouvements d'ajustement de la prise se
produiront (ouvertures et fermetures successives des pinces avec leger de-
placement de la patte sur la proie).
f. Exploration rostrale.
Apres un delai variable en fonction des mouvements et de la forme de la
90
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA
proie, les pattes ravisseuses rapprochent lentement la proie de l'extremite du rostre qui se souleve et decrit des mouvements en tous sens. Lorsque le contact s'etablit, les deplacements du 3e article rostra1 (le plus mobile) per- sistent encore un certain temps sur la cuticule de la proie bien tendue entre les 2 pattes. L'exploration s'arrete quand ie rostre trouve une rupture de
surface; si cette irregularite n'est pas decouverte, la Ranatre modifie sa prise par deplacement de ses pattes et recommence.
g. Piqfire et succion.
Les stylets maxillaires sont enfonces perpendiculairement dans la proie suivis des stylets mandibulaires. Leurs mouvements de va-et-vient persistent pendant tout le repas, la prise des stylets etant suffisamment solide pour per- mettre a l'une puis a l'autre patte prothoracique de se relacher totalement.
h. Fin du repas. La Ranatre reprend la depouille de sa proie avec ses pattes ravisseuses et
apres retraction des stylets la laisse tomber en ouvrant les tibio-tarses. I1 n'y a pas de toilette apres la fin du repas.
B. VARIATIONS DANS LE COMPORTEMENT DE CAPTURE La description ci-dessus correspond a un acte complet; certains mouve-
ments peuvent disparaitre, mais surtout on observe une variabilite assez
grande dans les angles caracteristiques des diverses postures de repos. Le Tableau 2 precisera les limites de cette variabilite:
TABLEAU 2
Variabilite des postures des pattes
angle pattes meso et pattes en degrees metathoraciques prothoraciques
Tarse/tibia 70 a 200° toujours I70°
Tibia/femur o a I85° 0° a I90°
Femur/coxa o a 180° 5° a I80°
Coxa/axe I5° a 2400 (de profil)
60o a 2000 (de par-dessus)
Pour la commodite de la suite de l'expose nous avons designe par des combinaisons de lettres et de chiffres certaines positions particulieres. Ces
sigles sont explicites dans le Figure 4. Dans certains cas, la capture peut etre menee a bien a partir d'une position
de preparation du type 5a, 6a, 7a, 8a, 9a, ioa. Cependant il n'y a jamais de
capture lorsque la Ranatre nage.
91
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

ANN CLOAREC
H V Y://.
X: 180° Z:.
x:145°.
y:20:25° 3. z
x: 145°.
\/y:30°40°
'< Vz:15:20' x:135°. \ 2
5. \/Y: 50°.
\^ \a z:25: x: 120'.
(ii)
H. ,7NA/ a .
6
7
H V \ /y.60°
2 :30? x: 120°
\/:40o
:
9/0 Vz :15°-20
x: 90°
8. /Y 50'
8 \z:3 25' x: 90o
x:\ /602 z 30 3O'70:
x: 90 .
o.v :25V x: 60°
). C.
Figure 4. Schemas montrant certaines positions des pattes ravisseuses, observees lorsque la Ranatre est au repos.
(i) - Postures definies par les differentes positions des 2 femurs et 2 coxas les uns par rapport aux autres. H.: vue horizontale. V.: vue verticale. X.: angle forme par l'articu- lation femur/coxa, en vue horizontale. Y.: angle forme par les 2 femurs entre eux, en vue verticale. Z.: angle forme par les 2 coxas entre eux, en vue verticale. Toutes les valeurs angulaires sont exprimees en degres. Chaque chiffre designe une position des pattes ravisseuses definie par une valeur de l'angle (X) forme par l'articulation femur/ coxa vu horizontalement, et par les valeurs des angles (Y), forme par les 2 femurs
entre eux, et (Z), forme par les 2 coxas entre eux, tous vus verticalement.
(ii) - Postures definies par les differentes positions des tibio-tarses par rapport aux femurs. a: Les 2 pinces sont ouvertes; (les 2 angles formes par les femurs et les tibio- tarses = 9o0). b: I pince est ouverte, I pince est fermee; (I angle forme par le tibio- tarse par rapport au femur = go90, l'autre angle = o°). c: les 2 pinces sont fermees; (les 2 angles formes par les femurs et les tibio-tarses = o°). Chaque lettre (a, b ou c) designe une position du femur par rapport au tibio-tarse considere pour les 2 pattes ravisseuses a la fois, et chaque lettre peut se combiner a n'importe quel chiffre. Ainsi
8a a ete appele posture de capture.
92
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA 93
La phase de prise d'interet peut manquer, mais la proie est toujours decelee a distance. Lorsque la proie est trop grosse ou trop vive, la Ranatre se bloque en flexion totale des articulations de ses pattes prothoraciques (reaction ,,d'evitement").
Si la phase d'orientation du corps peut souvent manquer il y a presque toujours orientation (plus ou moins rapide, plus ou moins ample) des pattes ravisseuses, sur la proie et le mouvement de capture s'effectue toujours ensuite; cependant ces derniers mouvements peuvent etre plus ou moins dis-
symetriques, plus ou moins decales dans le temps. Lorsque la Ranatre tient
deja une proie dans une patte ravisseuse l'autre peut capturer seule; en dehors de ce cas, les deux pattes reagissent. La capture proprement dite
peut se limiter a la fermeture des tibio-tarses sur les femurs (moins de 2% des captures reussies par les imagos).
Sur 550 ensembles de mouvements analyses, nous avons observe 82% de captures reussies et i8% de captures manquees. Parmi les premieres, 67% etaient effectuees par une seule patte et 33% par les deux pattes ensemble. Dans les cas de capture avec une patte, les animaux observes avaient souvent une tendance preferentielle: 40% des Ranatres etaient gau- cheres, 40% droitieres alors que 20% paraissaient ambidextres (ces derniers chiffres sont donnes a partir d'observation de capture d'une proie tombant exactement dans le plan sagittal de la Ranatre). Lorsque la capture a ete effectuee par une patte, l'autre continue son abaissee environ I/5eme de seconde puis sa pince s'ouvre et elle capture a son tour la proie deja tenue
par la premiere patte. Dans les cas de captures manquees a la premiere tentative, la Ranatre
repete l'ensemble de ces mouvements, ou l'enchainement s'arrete et l'insecte
reprend la position d'attente. La phase d'exploration rostrale existe toujours a moins que le rejet de la
proie ait eu lieu avant; la phase de piqfire et de succion n'existe que si l'ex-
ploration rostrale a confirme la qualite de la proie. Inversement, piqfire et succion ne peuvent intervenir que si la capture a eu lieu. Nous avons souvent tente et toujours vainement de faire piquer une proie convenable par une Ranatre au repos. II semble donc que l'enchainement precedemment decrit des mouvements des pattes soit necessaire pour le declenchement des mouve- ments des stylets buccaux. La fin du repas peut intervenir meme avant que la proie soit completement videe, soit parce que la Ranatre est rassasiee, soit
pour une toute autre raison. Elle peut aussi lacher la proie sans la toucher avec les pattes ravisseuses, uniquement par retraction des stylets.
Le schema suivant recapitulera l'ensemble des enchainements observables.
Behaviour XXXV 7
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions
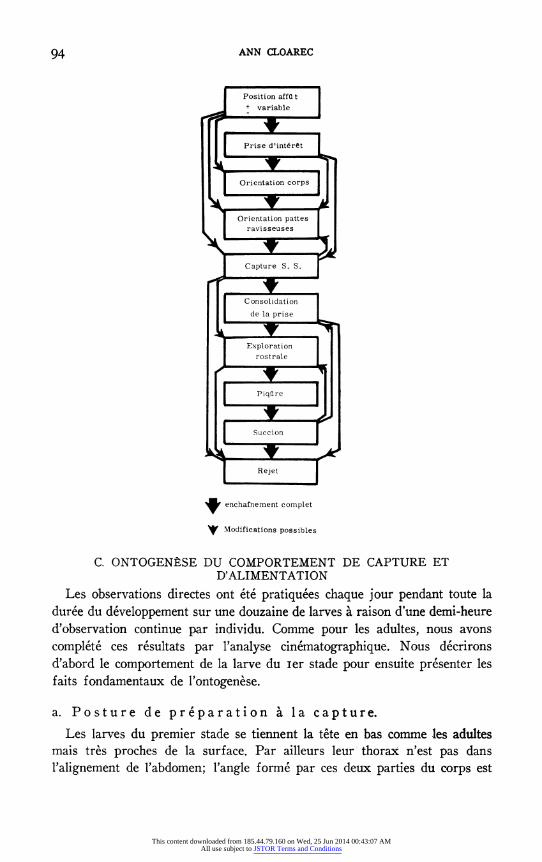
94 ANN CLOAREC
Position affQ t + variable
Prise d'int ret
Orientation corps
Orientation pattes ravisseuses
Capture S. S.
C onsolidation
de la prise
Exploration rostrale
Piqare
Succion
Rejet
t enchafnement complet
i Modifications possibles
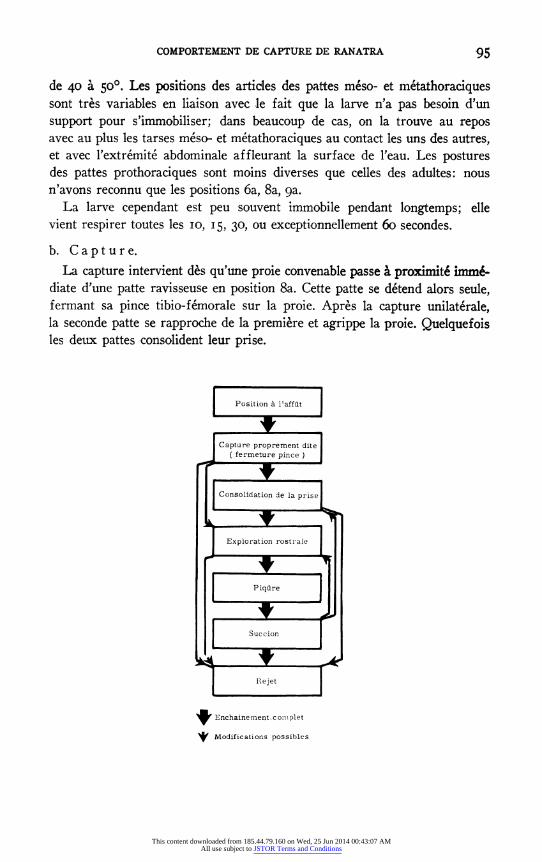
C. ONTOGENRSE DU COMPORTEMENT DE CAPTURE ET D'ALIMENTATION
Les observations directes ont ete pratiquees chaque jour pendant toute la duree du developpement sur une douzaine de larves a raison d'une demi-heure d'observation continue par individu. Comme pour les adultes, nous avons
complete ces resultats par l'analyse cinematographique. Nous decrirons d'abord le comportement de la larve du Ier stade pour ensuite presenter les faits fondamentaux de l'ontogenese.
a. Posture de preparation a la capture. Les larves du premier stade se tiennent la tete en bas comme les adultes
mais tres proches de la surface. Par ailleurs leur thorax n'est pas dans
l'alignement de l'abdomen; l'angle forme par ces deux parties du corps est
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions
COMPORTEMENT DE CAPTURE DE RANATRA
de 40 a 50°. Les positions des articles des pattes meso- et metathoraciques sont tres variables en liaison avec le fait que la larve n'a pas besoin d'un
support pour s'immobiliser; dans beaucoup de cas, on la trouve au repos avec au plus les tarses meso- et metathoraciques au contact les uns des autres, et avec l'extremite abdominale affleurant la surface de l'eau. Les postures des pattes prothoraciques sont moins diverses que celles des adultes: nous n'avons reconnu que les positions 6a, 8a, 9a.
La larve cependant est peu souvent immobile pendant longtemps; elle vient respirer toutes les 10, 15, 30, ou exceptionnellement 60 secondes.
b. Capture. La capture intervient des qu'une proie convenable passe a proximit6 imm&
diate d'une patte ravisseuse en position 8a. Cette patte se detend alors seule, fermant sa pince tibio-femorale sur la proie. Apres la capture unilaterale, la seconde patte se rapproche de la premiere et agrippe la proie. Quelquefois les deux pattes consolident leur prise.
9 Enchainement conm plet
1 Modifications possibles
95
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

ANN CLOAREC
c. Alimentation.
Apres un temps plus ou moins long, les pattes amenent la proie au contact du rostre qui effectue les memes mouvements d'exploration, de piquire, de succion et de rejet que nous avons decrits chez l'adulte.
Ainsi nous avons dresse le schema ci-contre pour resumer les phases du
comportement de capture de la larve du premier stade pour laquelle nous
soulignons trois differences fondamentales par rapport a l'adulte: - peu de diversite des postures preliminaires; - capture seulement a tres faible distance sans orientation prealable; - quasi independance des deux pattes ravisseuses.
d.: Ontogene se.
C'est surtout dans les postures generales et les mouvements du corps ou des pattes que se marquent les changements au cours de l'ontogenese.
L'angle forme par le thorax sur l'abdomen diminue de la maniere suivante: Ier stade: 40-50°; 2e stade: 35-45°; 3e stade: 25-40°; 4e stade: 20-30°; 5e stade: Io-I5°; adulte: o-5°.
Les postures des pattes prothoraciques se diversifient progressivement: Ier stade = 6a, 8a, 9a. 2e stade = 5a, 6a, 8a, 9a. 3e stade = 4a, 5a, 6a, 8a, 9a. 4e stade = 4a, 5a, 6a, 7a, 8a, 9a, Ioa.
5e stade = 3a, 4a, 5a, 6a, 7a, 8a, 9a, Ioa.
Les mouvements des pattes ravisseuses aboutissant a la capture se com-
pliquent; nous avons pu, au cours de nos seances d'observation (40 a 80 mouvements de capture pour une larve observee pendant I/2 heure), recon- naitre les divers types suivants que nous designerons ulterieurement par leurs
sigles (v. Figure 5). - la proie vient au voisinage de l'articulation tibio-femorale; la pince se
referme sans que l'autre patte intervienne (CrI). - meme chose apres leger contact de la proie et de la face interne de la
pince (CrI.t.). - la proie vient entre les 2 pattes ravisseuses, la Ranatre la saisit apres
fermeture d'une seule pince, mais accompagnee d'un rapprochement symetri- que des femurs des 2 pattes (CrI.F).
- meme mouvement de rapprochement des femurs, saisie avec les deux pinces (Cr2.F).
- meme chose, mais une des pinces se referme dans le vide (Cr2G.F ou Cr2D.F selon que c'est la pince gauche ou la droite qui a saisi la proie).
- des mouvements identiques de prise bilaterale (Cr2) ou unilaterale
96
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA 97
CrN \4r.t k/IV
Crl. F. C r2.F. C r 2 G. F.
Cr.2. R2F.Cr2.
A2F.Cr2G.
Figure 5. Schemas representant quelques mouvements observes au cours de differentes captures. Aires en pointilles = region ou se trouve la proie future. Pour l'explication
des sigles employes voir texte.
(Cr2G, Cr2D) peuvent s'observer apres fermeture simultanee des 2 pinces sans deplacement lateral des femurs. Dans d'autres cas, et comme c'est la
regle chez l'adulte, les mouvements de prises tibio-femorales se font apres un recul des extremites femorales vers la tete R2F et saisie soit par I, soit
par les 2 pinces se fermant simultanement, (R2F.Cr2, R2F.Cr2G, R2F.
Cr2D); comme precedemment et meme si les deux femurs se sont rapproches de la tete, chaque patte peut capturer independamment de l'autre (R2 (ou Ri) F.CrI). Enfin la capture peut intervenir non pas apres recul des femurs vers la tete mais apres abaissement plus ou moins lent des extremites femo- rales: ces cas seront traduits par la substitution de la lettre ,,A" a la lettre ,,R" dans nos sigles (A2F).
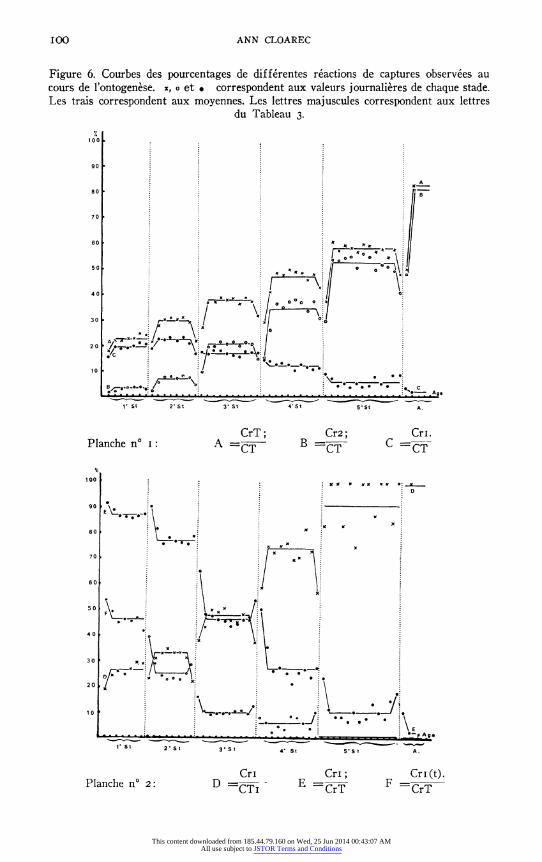
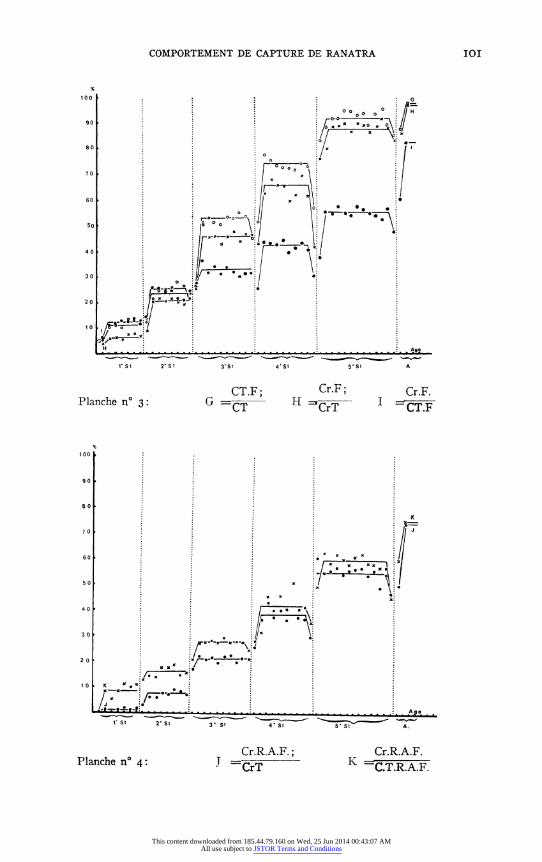
Tous ces mouvements peuvent s'observer sans reussite de la capture (rem- placement de Cr. par Cm.); mais le nombre des captures reussies parmi toutes les tentatives effectuees augmente au cours du vieillissement: Cr.T./ C.T.: courbe A: 22,5% au ler stade, 29,6% au 2e, 37,6% au 3e, 46,7% au 4 e, 57.2% au 5e, 82,0% chez l'adulte. (voir Figure 6 et Tableau 3).
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

98 ANN CLOAREC
TABLEAU 3
Rapports et pourcentages de differentes reactions de capture observees au cours de l'ontogenese; (les moyennes ont ete faites pour chaque stade sans tenir compte des observations faites le premier et le dernier jour de ce stade)
Ier St 2e St 3e St 4e St 5e St Adulte
Cr. T. 469 8o0 I I43 / 393 / 566 / 459
ACT. 2080/ 2702/ 3037/ 841 988 559- /
/ 22,5 / 29,6 / 37,6 / 46,7 / 57,2 / 82,0
Cr. I. 408 615 525 / 106 57 II D C T. . 1555/ 1852/ IIo8/ I42/ 62
/ 26,2 / 33,2 / 47,3 / 74,6 / 9I,9 / I00
Cr. i. 408 615 525 /io6 57 Ii / E Cr. T. 469/ 801I/ 1 43/ 393/ 566 45 9
86,9 / 76,7 / 45,9 / 26,9 1 , I / 2,5
Cr. i. (t) 218 199 116 - 22 2 O
Cr. T. 469 / 801 o 1143/ 2 393 566 6 459/
/ 46,4 / 24,8 / 10, 1 / 5,5 / 0,4 / o
Cr. i. 408 6I15 525 I 0o6 57 1l C T. I 208o 2702 303 / 841 988 559 /
/ IC3,6 /^ 22,7 / I7,2 1 I2,6 / 5.7 / I,9
Cr. 2. 61 / 186 / 6i8 287 509 /448 B C T. 2080/ 2702/ 841 I 988 559
/ 2,9 / 6,8 / 20,3 / 34,I I 51,5 / 80,0
CT. F. 242 / 682 1608 /624 / 901 /542 / C T. 2080/ 2702/ 3037/ 84I/ 988 559
/ I11,6 25,2 5 . 52,9 / 74,I / 9I,9 I 96,9
Cr. F. 30 162 / 523 265 494 / 445 / H Cr. T. 469/ 801 o / 43/ 393 566/ 459
// 6,3 / 20,2 / 45,7 2 67,4 87,3 / 96,9
Cr. F. 30 I62 523 265 494 445 ICT. F. 242 682 i6o8/ 624 9 901 542
12,3 / 23,7 / 32,5 / 42,4 / 54^ , / 82,I
C r. R.A.E. F 6 63 246 / i6o 315 / 345)
J Cr. T. 469 8/ o 43/ 393/ 566 45( 1,2 7,8 6 2I,5 2 40,7 7 55,6 / 75,I
K CT RAF. 71 \ - 879 6 389 535 / 452 / 8,4 2/9 96,93
ingeres 140 431 725 357 553 452 L Cr. T. 46) 8oi/ I43/ 393 566 459
3429,8 / 53,8 / 63,4 / 90.8 9747 / 98,4
ingeres 140 431I 725 357 553 452 M C T. 2080/ 2702 3037 841 98 559
/ 6,7 / I5,9 / 23,8 / 42,4 .55,9 /4 80,8
Cr.T. = Toutes les captures reussies. C.T. = Tous les mouvements de captures obser-
ves, reussis ou non. Cr.i. = captures reussies apres mouvement d'une seule patte ravis- seuse: seulement fermeture de la pince; (generalement). C.T.I. = Tous les mouvements de capture faits par une seule patte. Cr.i.(t). = Captures reussies apres que la proie ait touche la patte ravisseuse. Cr.2. = Toutes les captures reussies par reaction des 2 pattes ravisseuses ensemble. Cr.F. et C.T.F. = captures reussies, et toutes les mouvements de
captures comportant un deplacement des femurs (verticalement ou horizontalement). Cr.R.A.F. et C.T.R.A.F. = captures reussies et toutes les mouvements de capture avec deplacement vertical des femurs (recul vers tete ou abaissement). Les lettres majuscules
correspondent aux courbes de la Figure 6. Pour plus d'explications voir texte.
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA 99
Un fait interessant au cours de l'ontogenese est la diminution progressive du nombre de captures reussies apres mouvement d'une seule patte ravis- seuse; par rapport au nombre de captures reussies [CrI/CrT]-courbe E; ou par rapport au nombre total de mouvements de capture (CrI/CT)-courbe C- apres le 2e stade; bien que le pourcentage de reussites pour les mouve- ments d'une seule patte augmente (CrI/CTI) - les rares fois que les larves
agees et les imagos capturent a l'aide d'une seule patte, s'expliquent souvent
par le fait que l'autre patte enserre deja une proie. D'autre part, on s'aperqoit que le pourcentage des captures reussies du
type 2 (Cr2, Cr2.F, R2F.Cr2, etc. ...) parmi l'ensemble des tentatives de
capture (C.T.) voir courbe B, ou par rapport au captures reussies (CrT) voir courbes H et J, augmente; il augmente meme plus rapidement que le
pourcentage de captures reussies. Ceci indique la perte de l'independance des deux pattes ravisseuses l'une par rapport a l'autre au cours du vieillissement de l'animal.
Un autre fait doit etre souligne: si les captures apres contact de la proie avec le tibia ou le femur representent 45% de toutes les reussites de la larve du Ier stade, elles ne sont plus que I ou 2% au 4e stade et elles deviennent
exceptionnelles au 5e stade ou chez l'adulte (voir courbe F.). Ceci signifie que de plus en plus s'installe une detection de la proie a distance. D'ailleurs, cette variation s'accompagne d'une augmentation de la quantite de mouve- ments de capture dans lesquels s'observent des deplacements importants des extremites femorales. L'augmentation progressive de la proportion de mouve- ments de capture et de capture reussie, apres mouvement des femurs (d'abord rapprochement symetrique des 2 femurs: Cr.F, puis recul ou abaissement des femurs R2F ou A2F comptes ensemble: R.A.F.), montre que la visee des larves se precise en vieillissant. (voir courbes G.H. et I; et J. et K.).
Si l'orientation des femurs prealable a la capture peut s'observer dans
2,5 a 3% des tentatives chez les larves du Ier stade, cette orientation devient
progressivement la regle; l'orientation du corps est plus tardive: tres rare au 2e stade, elle ne s'observe bien qu'a partir du 3' stade.
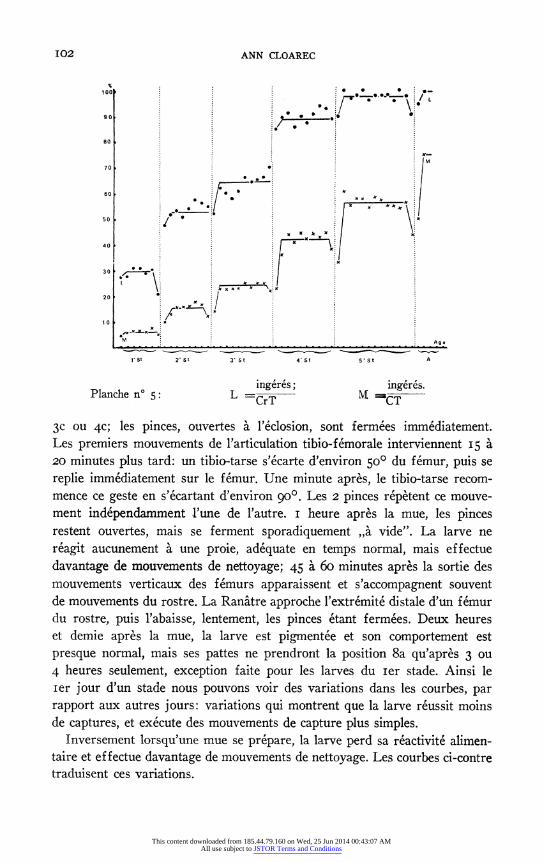
Precisons que le nombre de proies ingerees par rapport au nombre de
proies capturees augmente egalement et que, des le be stade larvaire, ce rapport est d'environ 50%. Ceci implique sans doute egalement une meilleure dis- crimination a distance (voir courbes L. et M., Figure 6 et Tableau 3).
Enfin, nous soulignerons l'influence de la mue sur le comportement de
capture. Nous avons suivi la modification des postures des pattes apres une mue au cours du durcissement de la cuticule larvaire et nous avons note les
temps des premieres tentatives de capture. Pendant 5 a Io minutes apres son
degagement, la larve reste sur l'exuvie, les pattes prothoraciques en position
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions
IOO ANN CLOAREC
Figure 6. Courbes des pourcentages de differentes reactions de captures observees au cours de l'ontogenese. x, o et * correspondent aux valeurs journalieres de chaque stade. Les trais correspondent aux moyennes. Les lettres majuscules correspondent aux lettres
du Tableau 3.
100 »
80 *
50. .
40. :
A.. .j^ ir-t---^ i
30 f
1: : ~
° : * o e ·.o . . C
.° b :, ,,,,, ,:............. , ,,,, , .,,,,,,,,:,*-
1' St 2' St 3' St 4'St 5'St
Planche n° I:
100
90 \ . s
j
80
.
70,
60 i
CrT; Cr2; A CT B CT
CrI. C =CT
O
i`
50
40,
30
20 ,
10
1 St 2' St 3- St 4' St 5-St A.
CrI CrI; Cr (t). DCTi E CrT F CrT
A.
. . - - .6 - - - - - - - - w
i X x x r. r : -A - D
)( rr~~~~~~
Planche n° 2:
·i
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions
C)
II
471
It Q-3
5d >
i IJ
It
CD
C~)
TT nIll
U nfl
C-3 ;Tj
II
H11 r-q .,
Yt.7
o 5C
tI
0
d
c3
z
I
H
0
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions
ANN CLOAREC
190 i i i ... *
90 . ' i/. *
i .
80
*0 i f _/
30, .. 6
20 /
310 - i \|
10:
M . Aga
1'St 2' St 3' St 4 St 5-St A
ingeres; ingeres. Planche n° 5: L =CrT CT
3c ou 4c; les pinces, ouvertes a l'eclosion, sont fermees immediatement. Les premiers mouvements de l'articulation tibio-femorale interviennent 15 a 20 minutes plus tard: un tibio-tarse s'ecarte d'environ 50° du femur, puis se replie immediatement sur le femur. Une minute apres, le tibio-tarse recom- mence ce geste en s'ecartant d'environ go°. Les 2 pinces repetent ce mouve- ment independamment l'une de l'autre. I heure apres la mue, les pinces restent ouvertes, mais se ferment sporadiquement ,,a vide". La larve ne
reagit aucunement a une proie, adequate en temps normal, mais effectue
davantage de mouvements de nettoyage; 45 a 60 minutes apres la sortie des mouvements verticaux des femurs apparaissent et s'accompagnent souvent de mouvements du rostre. La Ranatre approche l'extremite distale d'un femur du rostre, puis l'abaisse, lentement, les pinces etant fermees. Deux heures et demie apres la mue, la larve est pigmentee et son comportement est
presque normal, mais ses pattes ne prendront la position 8a qu'apres 3 ou 4 heures seulement, exception faite pour les larves du Ier stade. Ainsi le Ier jour d'un stade nous pouvons voir des variations dans les courbes, par rapport aux autres jours: variations qui montrent que la larve reussit moins de captures, et execute des mouvements de capture plus simples.
Inversement lorsqu'une mue se prepare, la larve perd sa reactivite alimen- taire et effectue davantage de mouvements de nettoyage. Les courbes ci-contre traduisent ces variations.
I02
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA I03
IV. ETUDE EXPiRRIMENTALE DES FACTEURS DE CAUSALIT: DE LA CAPTURE DES PROIES
Nous avons effectue cette etude a l'aide de leurres simulant une larve de chironome presentes a des larves des 4e et 5e stades ou a des adultes; pour les larves plus jeunes, jamais les leurres utilises n'ont declenche des reactions semblables aux proies vivantes et nous avons fait quelques experiences avec celles-ci.
A. CARACTiRISTIQUES OPTIQUES ET MICANIQUES D'UNE PROIE
Des experiences semblables a celles de KOEHLER (I924) sur les Libellules
(presentation d'une proie ou d'un leurre derriere une paroi de verre) nous
permettent d'affirmer que les stimuli visuels jouent un role fondamental dans le declenchement de la capture. Les mouvements sont les memes que ceux d'une capture normale quand on introduit le leurre dans un tube de verre dans l'aquarium ou se trouve la Ranatre, quand on deplace devant elle un
spot lumineux ou meme quand on agite un leurre dans l'air au-dessus d'une Ranatre qui se trouve pres de la surface de l'eau.
Cependant, si les excitations visuelles sont tres importantes dans le cas
normal, elles ne sont pas seules en cause. En effet, la Ranatre adulte parvient a capturer en lumiere rouge tres faible ou a l'obscurite ou encore en etant
aveuglee soit que la proie touche la patte ravisseuse, soit qu'elle descende a vitesse convenable entre les deux pattes. Ceci prouve que les excitations
mecaniques (vibration ou contact) jouent egalement un role dans le declen- chement de la capture.
a. Forme et taille de la proie. Nous avons recherche les limites de diametre pour nos leurres cylindriques
et de surface pour des leurres plats qui permettent d'observer soit une
capture totale, soit des elements de capture, soit des reactions d'evitement. Les resultats des experiences sont consignes dans le Tableau 4, et exprimes en mm.
TABLEAU 4 Tailles limites de leurres
Reaction Capture reussie Reaction de capture Evitement cylindre forme plate cylindre forme cylindre forme plate Taille
Stade plate tibiotarse I <0,5 <o,75 X 0,75 0,6-0,7 I X I >0,7 >2 X I 1,3 mm 2 <o,75 <I,5 X I 0,8-I 2 X I >1,2 >3 X 1,2 1,85 mm 3 <I <2 X 1,5 I,I,5 4 X 4 > 1,8 >6 X 4 2,4 mm 4 0,2<c<I,75 <2,5 X 2,5 1,8-2,2 9 X 6 >2,5 >9 X 9 3,5 mm 5 o,2<c<2,5 <6 X 4 2,6-3,5 9 X 12 >3,75 > 2 X 12 4,4 mm
A o,2<c<3,5 2I2 X 15 3,8-4,5 I5 X 15 >5 >15 X I8 5,4 mm
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

ANN CLOAREC
TABLEAU 5
Reactions des imagos a un leurre cylindrique Diametre Reaction
3,5 mm tous lachent immediatement 30% lachent immediatement
3,2 mm- 50% lachent apres une duree moyenne de 13" 20% procedent a une exploration rostrale apres une moyenne de 20"
Pour chaque stade, la prise d'une proie depend de la longueur du tibio- tarse mais les reactions de capture sont tentees pour des tailles tres differentes de celle-la. Dans tous les cas les insectes capturent des leurres plats plus grands que des leurres cylindriques. Ceci s'observe bien lorsque l'on tente de nourrir des Ranatres avec des larves de Criquets: la prise est impossible si l'abdomen depasse une certaine taille; cependant en presentant le Criquet de telle sorte que la Ranatre le capture par une patte, l'alimentation devient realisable.
b. Couleur.
Nous n'avons observe aucune influence de la couleur propre du leurre ni de son contraste avec le fond de l'aquarium dans la frequence des captures. Afin de nous rapprocher cependant des conditions naturelles, toutes les
experiences autres que celles correspondant au present chapitre ont ete reali- sees avec des leurres rouges.
c. Mouvement.
L'introduction dans l'aquarium au voisinage d'une Ranatre, d'un leurre de taille et de forme convenable declenche aussitot le relevement femoral
calacteristique de la prise d'interet. Si le leurre est alors maintenu immobile, le deroulement des sequences de capture s'interrompt. Des que le leurre est
agite a nouveau, la capture peut intervenir.
Lorsque la vitesse de deplacement vertical et les mouvements du leurre sont irreguliers, le pourcentage de captures reussies est eleve, mais la reussite des captures dans un secteur determine par rapport a la larve depend de la
posture de depart des pattes.
TABLEAU 6
Postures permettant le succes de la capture Presentation a Postures permettant le succes
o° 4a, 5a, 6, 7, 8, 9, io 45° 5a, 6, 7, 8, 9, Io o90 6a, 8, ga
I35° 9a
I04
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA
Les angles sont mesures en degres dans un plan horizontal, a partir du
plan de symetrie de la Ranatre, de l'avant vers l'arriere et ayant toujours pour sommet un point situe entre les yeux.
Lorsque les mouvements sont reguliers, le leurre est sans doute moins efficace mais la regularite permet une meilleure definition des conditions
experimentales. Nous avons tente de faire capturer le leurre soit pendant sa descente, soit pendant sa montee, soit en mouvement horizontal, en con- dition symetrique ou dissymetrique.
Pour les mouvements de descente, nous avons presente le leurre aux posi- tions utilisees pour les mouvements irreguliers, tandis que pour les mouve- ments de montee, nous n'avons employe que la position o°.
Les vitesses du leurre employees sont, en mm/sec: o,8; I,I5; 1,45; 5; 6,9; 8,8; 12,5; I7; 22; 25; 34; 44; 50; 69; 88.
Un leurre qui se deplace a une vitesse inferieure a 0,8 mm/sec. ne declenche aucune reaction. A une vitesse superieure a 88 mm/sec., il est trop rapide pour que les captures aient le temps d'etre declenchees. Nous sommes donc restee entre ces limites et nos resultats codes sont fournis dans le Tableau
7, A, B, C, D, E et F.
TABLEAU 7 A a F Tableaux montrant les differents degres de reaction des pattes ravisseuses en fonction de leur position au depart (voir fig. n° 4), de la vitesse du leurre, de sa position et du mouvement (descente, montee ou mouvement horizontal). Chaque position des pattes ravisseuses a ete testee, pour chaque categorie de
mouvemients, 3 ou 4 fois sur une vingtaine d'insectes A
Leurre descente a o° Vitesses en mm/sec. 1,45 6,9 17 et 34 88 Positions
IC o o o o Ia o 6o%: o 5ao%:o o
o.%: pt F2T. 5o%: pt E2F. 3C o 5o%: o 0 0
50%o: E2F. 3a o 6o%: F2T. Sso%:R2F. 5o%:o
4o%: R2F.F2T. 50%: R2F.F2T. (so%: F2T. 5c 0 ptR2F. ptR2F. o 5a R2F. R2F. R2F.F2T. S6o%: F2T.
40o%: F2T.R X 2F. 7c 0 R2F. R2F. pt R2F. 7a 50% :o 5o%: R2F. 166%: R2F. R2F.F2T.
So%: R2F. o5%: Cr. ?33 : Cr. 8c S66%:o R2F. 50%:R2F. R2F.
p33%: pt R2F. o50%: R2F.02T. 8a Cr. Cr. Cr. 20%: Cm.
8o%: Cr. ga Cr. Cr. Cr. Cr.
I05
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

ANN CLOAREC
B
Leurre dcscente a 450
Vitesses en mm/sec. Positions
IC Ia 3C 3a
6,9.
0 0 0 0
5C E2F.
17
o 0 o 0 o
50%: E2F. FI+ 5o 50%: F2T. MC + 5O.
pt R2F
5a pt R2F. F -- IO+ 5o%: R2F.F2T. 50%: R2F.F1 +- io
7C 0 o2T. 7a $5o%: F2T. R2F.
i50%: R2F.F2T.
8c 150%: OT1. R2F.02T. i50%: O
8a Cr. (MC. + 50) Cr. (MC.+ ±io)). ga Cr. (MC. + Ioo) Cr. (MC. ±- io).
34
0 0
88
0 0
0 0 F2T.ptR2F. MC+ IO' o
50%': R2F. 50%: E2F. 50%: R2F. FI+ io. 50%: R2F.F2T.
0 R2F.
E2F.F1 +- io1.
Cr. (MC. + I5°). Cr. (MC. +- Io).
0
R2F.FI -+- io.
0
50% : F2T.ptR2F. 50%: F2T.ptR2F.
+MC. +50 0
Cm. (MC. +- Io). Cr. (MC. + IO0).
. 1,45 mm/sec.: aucune reaction sauf pour la position 8a (ptRFi.FI (Cr.MC +- 50).
Leurre descente Leurre descente a 900
Vitesses en mm/sec. Positions
17 34
IC 0 ia 3c 3a
0 0 0
5C 0 5¢ o 5a 25%' o
175%: FT1. 7c o 7a RF1. 8¢ o 8c 0 8a S25%: R2F.FT1,
75%: Cm.sansMC. 9a Cr. (R2F. MC. +- 300
puis C.)
0 0 0 0 0 R2F.F1> F2. FT1.
0
ptR2F. Fi > F2. 0
30%: R2F.F2T. 70%: Cm.sans MC.
Cr. (R2F.MC. - 20W. puis C.)
0 0 0 0 0
o ptR2F. MC. ± i00
0
ptR2F. Fi> F2. 0 MC. +- Io.FT1.R2F.
Fl+ Io°. Cr. (MC. - 200)
0 0 0 0 0
R2F.FT1.
0 R2F. Fi. + 50 0
MC. + ioo.R2F.J
Cm. sans MC.
o o o o
R2F. FT1.
o R2F. Ft -]- 5°. o MC. q-t I o.ReF.]
Cm. sans MC.
a 1,45 mm/sec.: aucune reaction sauf pour la position 8a (RF1) et 9a (Cr.MC. + 300).
io6
+- 50) et 9a
88
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA17
D
Leurre descente 'a I35-
17
IC 0 la 0
3C 3a 5C
0 0 0 0
7C 0 7a 0
8c 0 8a 25o
~75%1: RFi ou R2F.Fi>E2.
9a Cm. sans MC.
0 0 0 0 0 0 0
pt R2F. 0
50%1: R2F.MC. + io0
50%: R2F.FT1. MC. + Io .
CM. (MC. +200).
0 0 0 0 0 0 0 R2F. Ft + io00 0
S50o%: R2F.MC± + o ~50%: R2F.F2T
sans MC. 5o%: Cm. (MC. + 50)
50%: R2F.F1 + io'
a I145 mmn/sec. aucune reaction sauf pour les positions 8a (50%b: Ot. et 50%: pelil mouvement des fe'murs: Y2EF1 et ¼/ pt RF-i.) et ga (Cm. sans MC.)
E
Leurre monte'e 'a 00
Vitesses en mm/sec. Positions
3c 3a
6,9
0 0 0 pt E2F.
5c pt R2F. 5a 5o%: R2F.
0o% : R2F. -I-
7a R2F. F2T. 8c 55o%: pt R2F.
So%: pt R2F.o2 8a C. 9a C.
17
0 0 0
50: pt E2F. 5o% : F2T.
R2F. 5o:R2F. ±
cp. 50%: R2F.F2T. 5o%: R2F.
~50%: R2F. o2T. R2F. + cp. R2F. o2T.
'T. C. + cp. C.
34
0 0 0
0 0 0
$so% : o 0 ~ o:E2F.
pt R2F. 0 cp. 5o%: RZ2F. + cp. R2F.
5o%: R2F. F2T. 0
R2F. ± cp. R2F. o2T.
C.' C.
0
JR2-?F . pt R2F.
Cm. C. + cp.
I ,45 mm/sec. aucune reaction sauf pour les positions -a (pt R2F.) et 8a et Qa (C.).
Vitesses en mim/sec. Positions
6,9 34 88
0 0 0 0 0 0 0
pt R2F.F2T. 0 R2F.F2T. MC.+ +
Cm. sans MC.
88
W7
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

ANN CLOAREC
F
Leurre horizontal Vitesses en mm/sec. 6,4 23 43 85 Positions
IC o o o o ia o o o o 3c o o0 0 3a 50%:0o 50%: R2F. !50%:R2F. S50%: o
5o%: pt E2F. i50%: R2F. 15o%: R2F. F2T. 50% : F2T. 5c o R2F. ptE2F. o 5a R2F. 550%: Cm. R2F.FT1. pt R2F. F2T
50%o: C. 7c o50%:o pt R2F. o o
i5o%: pt R2F. 7a 50%: Cm. s5o%:R2F. Cm. (MC.+ IO°). 5o%:Cm.
o50%:R2F. i50%:Cm. 50%: R2F. 8c R2F. R2F. R2F.OTi. 0 8a Cr. Cr. Cr. Cm. 9a Cr. (quand le leurre Cr. (pas d'ant- Cr. (petite Cm. -- MC. en
se trouve dans le icipation.) anticipation suivant le plan de symetrie.) par les pattes.) leurre.)
Lcgende: 0: aucune reaction. E2F: ecartement des 2 femurs du plan de symetrie. 02T: ecartement des 2 tibiotarses des femurs: ouverture de la pince. F2T: fermeture des 2 tibiotarses sur les femurs. FTI: fermeture du tibiotarse du c6te du leurre. R2F: recul des 2 femurs vers la tete. Cr: capture reussie. Cm: capture manquee. MC: mouve- ment du corps lateralement. cp: mouvement du corps vers le haut. pt: mouvement peu
ample. FI: femur du c6te du leurre. F2: femur eloign6 du leurre.
Nous pouvons conclure de ces experiences que le mouvement vertical de descente d'un leurre est plus efficace que le mouvement vertical de montee ou que le mouvement horizontal; par ailleurs, le nombre de captures reussies decroit lorsqu'on s'eloigne du plan sagittal anterieur de l'Insecte, les reactions etant tres rares a la suite d'une presentation &a 35°.
Les leurres animes d'un mouvement horizontal, sont cap'ures pres du plan de symetrie par mouvements symetriques des 2 pattes ravisseuses; mais pour un leurre mu par vitesse plus elevee (>45 mm/sec.), la capture a lieu
lorsque le leurre a depasse le plan de symetrie, et dans ce cas, on observe une orientation du corps.
Les mouvements des 2 pattes ravisseuses sont symetriques pour un leurre descendant dans le plan de symetrie. Quand le leurre est a 45°, il y a souvent deplacement des 2 femurs sur le cote; le femur le plus loin du leurre se deplacant le plus, tandis que l'autre patte seule saisit la proie lorsqu'il y a capture.
Quand le leurre est a 9o0, les mouvements sont moins amples quand il n'y a pas capture; autrement en plus du deplacement des femurs lateralement, on observe une orientation de tout le corps, un pivotement de jusqu'a 40°.
Io8
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA 109
d. Poids et nature.
En utilisant des cylindres de diametre identique (2,5 mm) de divers mate- riaux, il a ete possible d'explorer les qualites mecaniques du leurre. Les Ranatres adultes etudiees capturent puis effectuent l'exploration rostrale a. condition que le poids du leurre presente soit inferieur a 9 cg. Elles capturent encore des leurres de idg., mais ne l'explorent pas: des apres la capture, une reaction speciale se manifeste: le corps se rapproche de la verticale, les
pattes ravisseuses enserrant le leurre tombent et parfois presentent un trem- blement qui rappelle un tetanos imparfait. Tout leurre d'un poids superieur a idg. est lache immediatement apres capture.
La nature irreguliere du leurre est une condition fondamentale de l'explo- ration rostrale, les substrats lisses etant rapidement rejetes.
e. Action repete du leurre.
En repetant les stimulations, on aboutit a une extinction de la reaction de capture. Le Tableau 8 en fournit 2 exemples.
TABLEAU 8
Position des pattes Reaction Temps de Position des pattes ravisseuses au depart latence ravisseuses apres
8a capture immediate 8a 8a capture apres 1,5 sec. 8a
(I) 8a capture apres 4 sec. 8a 8a capture apres 8 sec. 6a 6a capture apres 50 sec. 5a puis se met a nager.
8a capture immediate 8a 8a capture apres I sec. 8a
(2) 8a capture apres 7 sec. 5a 6a capture apres 12 sec. 4a 4a rien F2T allongement 3c
pattes ravisseuses
Ceci nous a conduite a laisser au moins I/4 d'heure de repos a un Insecte entre deux tests successifs de la meme serie d'experiences.
B. ESPACE EFFICACE
Ayant determine les qualites physiques d'un leurre optimal, nous l'intro- duisons dans une position mesuree en l'approchant progressivement de la
Ranatre. Nous reperons la distance a laquelle il declenche la reaction d'orien-
tation seule puis la reaction d'orientation suivie de capture (reaction dans au moins 8o% des cas). Chaque essai est repris 3 fois sur chaque individu dans chaque direction pour les distances efficaces. Le groupe experimental
7*
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions
I I ANN CLOAREC
V H
B
E F
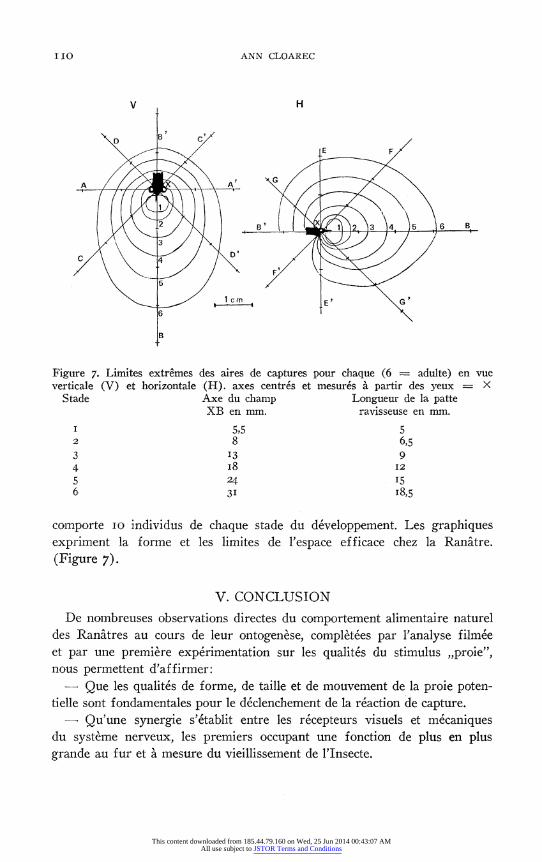
Figure 7. Limites extremes des aires de captures pour chaque (6 = adulte) en vue verticale (V) et horizontale (H). axes centres et mesures a partir des yeux = X
Stade Axe du champ Longueur de la patte XB en mm. ravisseuse en mm.
I 5,5 5 2 8 6,5 3 I3 9 4 I8 12 5 24 15 6 3I I8,5
comporte 10 individus de chaque stade du developpement. Les graphiques expriment la forme et les limites de l'espace efficace chez la Ranatre.
(Figure 7).
V. CONCLUSION
De nombreuses observations directes du comportement alimentaire naturel des Ranatres au cours de leur ontogenese, completees par l'analyse filmee et par une premiere experimentation sur les qualites du stimulus ,,proie", nous permettent d'affirmer:
- Que les qualites de forme, de taille et de mouvement de la proie poten- tielle sont fondamentales pour le declenchement de la reaction de capture.
Qu'une synergie s'etablit entre les recepteurs visuels et mecaniques du systeme nerveux, les premiers occupant une fonction de plus en plus grande au fur et a mesure du vieillissement de l'Insecte.
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA
- Que l'espace efficace vis-a-vis des reactions de capture est essentiellement un volume antero-dorsal par rapport a l'animal.
- Que la reaction de capture est une reaction sequentielle enchainee mais non obligatoire dans toutes ses phases, le milieu exterieur agissant toujours sur le deroulement de l'acte.
- ue s'etablissent au cours de l'ontogenese divers types de coordination
posturale entre les regions thoracique et abdominale du corps, entre les differentes pattes ipsi- et contra-laterales. L'independance des pattes ravis- seuses, grande au debut de la vie, fait place progressivement a une liaison
temporelle tres nette. - Que l'expression du comportement alimentaire se trouve integree dans
l'ensemble de l'expression des comportements de la Ranatre: un adulte qui nage ou qui respire en surface ne capture pas, ou en tout cas, ne suce jamais une proie saisie fortuitement dans la patte ravisseuse. Le fait que le degre de preparation a la capture puisse s'exprimer dans la position des pattes prothoraciques; le fait aussi que tout ceci varie au cours de l'ontogenese, permet de penser que les facteurs neuroendocrines et les facteurs de matura- tion jouent un grand role dans le comportement alimentaire de cet Insecte.
L'analyse de ces facteurs est commencee dans la voie tracee par nos precedes- seurs (MINNICK, I93I; DETHIER, I958; MITTELSTAEDT, I957; BUCHHOLTZ, 196I, entre autres) et fera l'objet d'un prochain memoire.
RigSUMi
Diverses phases ont ete distinguees lors d'une sequence complete de capture: (I) prise d'interet; (II) orientation du corps, puis des pattes prothoraciques; (III) capture pro- prement dite; (IV) consolidation de la prise; (V) exploration rostrale; (VI) piqire et succion; (VII) fin du repas.
Une etude ontogenetique montre les differences essentielles de comportement de la larve du I° stade par rapport a celui de l'adulte. Les postures des pattes prothoraciques sont moins nombreuses; la capture se fait a tres faible distance sans orientation prealable. Les 2 pattes ravisseuses sont quasi independantes. Divers mouvements amenant a la capture sont decrits ainsi que leur evolution au cours de l'ontogenese.
Une partie experimentale etudie quelques facteurs de causalite de la capture: forme, taille, couleur, lumiere, poids, nature, type de mouvement, vitesse et position par rapport a la Ranatre.
OUVRAGES CITES
ABBOT, C. (I940). Some reflex responses of Ranatra fusca to contact stimuli. - Bull. Brooklyn Entom. Soc. 35, p. I33-I34.
BAUNACKE, W. (1912). Statische sinnesorgane bei den Nepiden. - Zool. Jahrb. 34, p. I79-346.
BUCHHOLTZ, C. (I96I). Eine verhaltensphysiologische Analyse der Beutefanghandlung von Calopteryx splendens Harr. (Odonata) unter besonderer Beriicksichtigung des opti- schen AAM nach partiellen R6ntgenbestrahlungen des Protocerebrums. - Verh. d. Dtsch. Zool. Ges. 25, p. 402-412.
III
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

II2 ANN CLOAREC
DETHIER, V. G. (I957). Chemoreception and the behavior of insects. - In: Survey of Biological Progress 3, p. 49-83.
- & BOrENSTEIN, D. (1958). Hunger in the blowfly. - Zsch. f. Tierpsychol. 15, p. I29-I40.
DUFOUR, L. (1820). Recherches anatomiques sur la Ranatre lineaire et la Nepe cendree. - Ann. Gen. Sc. Phy. Brux. 7, p. 194-213.
HOFFMANN, W. E. (1930). Notes on the life history of Ranatra chinensis Magr. - Proc. Nat. Hist. Fukien Christian Univ. China 3, p. 31-37.
HOILMES, S. J. (1905). The reaction of Ranatra to light. - Journ. comp. neurol. Psychol. I-5.
- (I9o6). Observation on the young of Ranatra quadridentata (Stal). - Biol. Bull. Marine Biol. La. 12, p. 158-164.
KOEHLER, 0. (I924). Sinnesphysiologische Untersuchungen an Libellenlarven. - Verb. Dt. Zool. Ges. 29, p. 83-9I.
LARSEN, 0. (1936). Zur Biologie von Ranatra linearis. - Opuscula entomologica I.I, p. 112-119.
- (I949). Die Ortsbewegungen von Ranatra linearis L. - Lunds Univ. Arsskrift N.F. 2, p. 45-6.
LOCY, W. A. (1883). The anatomy and physiology of the family Nepidae.- American Nat. I8, p. 250.
MARSHALL, W. S. & SEVERIN, H. (I904). Some points in the anatomy of Ranatra furca. - Trans. Wisc. Acad. 14, p. 487-508.
MINNICK, D. E. (I931). The sensitivity of the oral lobes of the proboscis of the blowfly Calliphora vomitoria Linn. to various sugars. - J. exp. Zool. 60, p. 121-139.
MITTELSTAEDT, H. (I957). Prey capture in Mantids. - In: Recent Advances in Inverte- brate Physiology, Univ. Oregon Pub. p. 51-71.
NEISWANDER (1926). On the anatomy of the head and thorax in Ranatra (Heteroptera). - Trans. Ann. Ent. Soc. 51, p. 311-320.
POISSON, R. (1924). Contribution a l'etude des Hemipteres aquatiques. - Bull. Biol. Fr. Belg. 58, p. 49-305.
RAGHUNATHA RAO, T. K. (I962). On the biology of Ranatra elongata Fabr. (Heteroptera Nepidae) and Sphaeroderma annulatum Fabr. (Heteroptera Belostomatidae). - Proc. Roy. Entom. Soc. London, A. Gen. Entom. 37, 4-6, p. 61-64.
TORRE-BUENO, J. R. de la (Ioo6). Life histories of North American Waterbugs. 1. Ranatra quadridentata. - Canad. Ent. 38, p. 189-197.
SUMMARY The catching and feeding behaviour of imagos of Ranatra lincaris preying upon
Chironomus larvae is described. This behaviour starts from a particular posture: the precapture posture. A typical prey-capture sequence consists of the following phases: (I) Arousal of attention; (II) orientation of the body on the resting place, followed by orientation of the fore-legs while the body remains still; (III) seizure of the prey; (IV) consolidation of the grasp on the prey; (V) exploration of the surface of the prey with the rostrum; (VI) insertion of the stylets and sucking; (VII) discarding of the prey. This pattern may vary in this respect that some phases may drop out. A number of different postures of the prothoracic legs have been distinguished. When swimming Ranatra never capture a prey.
This same behaviour was studied during the ontogenesis of Ranatra. In the first instar the prothoracic legs show a smaller number of postures than in the imago; more possible postures are added after each moult. The main differences observed between the behaviour of first instar larvae and of imagos are the following: capture is not preceded by an orientation phase; a prey can be seized only if it is very near - the two prothoracic legs may move completely independantly during a capture. Different
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions

COMPORTEMENT DE CAPTURE DE RANATRA I13
movements of the leg in grasping a prey have been described: (I) only one tibia presses down on the femur, sometimes after a prey has actually touched the inside of a claw; (2) a horizontal movement towards the insect's plane of symmetry by one or by both femurs, followed by the closing of one or both claws; (3) a more complex movement by the fore-legs in which they are either moved back towards the head or lowered. The percentage of catching attempts which are successful increases with the age of the insect. The fore-leg movements become more and more complex; the percentage of successful catching attempts made after a movement of only one prothoracic leg progressively decreases although the percentage of successful captures after a movement of only one fore-leg increases. At the same time the percentage of successful attempts made by both legs increases. This shows that there is a certain gradual loss of independance of the fore-legs. The number of captures after a prey has touched the inside of a claw decreases rapidly; such catches are exceptional in 5th instar larvae and in imagos. Prey detection at a distance becomes the rule. The proportion of capture movements including preliminary femur movements gradually increases. Preliminary femur orientation is quite rare during the Ist instar, but progressively becomes the rule in later stages. Proper orientation of the body can only be observed from the 3rd instar on; capture behaviour of Ranatra just before or just after a moult shows the influence of morphological and physiological changes associated with moulting.
This work was completed by an experimental study of some factors relating to prey catching, such as the visual and mechanical characteristics of the optimal prey, and the form and size of a rectangular or cylindrical dummy. Weight, movement, speed and position with regard to predator prove to be important for Ranatra. Colour differences have no effect. A position of the prey in the plane of symmetry of Ranatra and a speed of the prey of about 6 to 34 mm/sec. is optimal for catching. A vertical downward movement is more efficacious than a vertical upward movement, or a horizontal one; but an irregular movement is even more efficacious. The efficacious space for successful catches lies above and in front of Ranatra.
This content downloaded from 185.44.79.160 on Wed, 25 Jun 2014 00:43:07 AMAll use subject to JSTOR Terms and Conditions