Embed Size (px)
Citation preview

L’Encéphale, 2006 ;
32 :
253-62, cahier 1
253
NEUROPSYCHOLOGIE
Existe-t-il un déficit d’inhibition lors du vieillissement ? Confrontation de l’hypothèse dorso-ventrale et de l’hypothèse frontale dans l’effet d’amorçage négatif
S. MARTIN
(1)
, D. BROUILLET
(2, 3)
, E. GUERDOUX, R. TARRAGO
(1) [email protected].(2) Laboratoire de Psychologie Expérimentale de la Mémoire et de la Cognition (LaMeCo), EA 3021, Université Paul-Valéry, Route de Mende,34199 Montpellier cedex 05.(3) [email protected]
Travail reçu le 21 juin 2004 et accepté le 17 décembre 2004.
Tirés à part :
S. Martin (à l’adresse ci-dessus).
Inhibition and resource capacity during normal aging : a confrontation of the dorsal-ventral and frontal models in a modified version of negative priming
Background.
Over the past decades, cognitive psychology contribution to our understanding of aging relies on two majorperspectives, focusing on the selective impact of age on either cognitive multiple-systems or global factors of cognition :slowing, working memory and inhibition. In the latter, reduction in inhibitory control during aging (in its access, deletionor restraint functions) is associated with poorer performance on a variety of tasks referring to memory, comprehensionor language [Hasher, Zacks and May
(16)
]. The attractiveness of inhibition as an explanatory factor results in part in theabsence of negative priming during aging. Negative priming refers to the slow down of latencies when individuals haveto respond to recently ignored informations, compared to unrelated informations. The dissociation, between a preservedlocation negative priming and an absence of identity negative priming during aging, supports the dorsal-ventral model ofinhibition which suggests that spatial and identity inhibition are supported by different and independent visual pathways.An alternative model, directly at odds, is that inhibitory mechanisms are supported by the frontal lobe. In this perspective,inhibition is not a central process responsible for the control of working memory contents, but an automatic and localmechanism whose triggering depends on controlled attention. Therefore, working memory drives efficient inhibition bysustaining task instructions and appropriate responses throughout task execution. This hypothesis is consistent with Hough-ton and Tipper’s
(17)
architecture of selective attention. According to the authors, the presence or absence of automaticinhibition is very closely linked to a Match/Mismatch field whose function is to compare the present stimulus to an internalself-generated internal template. When an information fails to match the subject’s current goals, the match/mismatchfield causes an automatic inhibitory imbalance which reduces the to-be-ignored properties’ responsiveness. In contrast,information matching subjects’ goal is enhanced through an automatic excitatory imbalance. The accurate functioning ofthe Match/Mismatch field requires efficient executive functioning responsible for the uphold of goals and correct responses.In the case of negative priming, manipulating the efficiency of working memory is of interest as it should affect the triggeringof slowing,
ie
, an indirect inhibitory deficit, when the task is resource demanding [Conway
et al. (6)
]. Moreover, if inhibition,as reflected by negative priming, is mediated by individual resource capacity, then NP should disappear during agingonly when individuals are engaged in a resource-demanding task.
Objectives.
To address this issue, we examine whethercognitive control load in a gender decision task contributed to the presence or absence of NP during aging. Accordingto the dorsal-ventral model, task complexity should not have any impact on performance, since gender decision taskrelies on a conceptual analysis of information. In turn, the frontal model predicts that age differences in performanceprofile will only differ when individual resource capacity is overloaded.
Design of the study.
Sixty-four participants (32young and 32 older adults) performed a gender categorisation task through two experiments. Trials involved two stimulipresented successively at the same location. A word served as a prime and a word as a target. Both prime and target

S. Martin
et al.
L’Encéphale, 2006 ;
32 :
253-62, cahier 1
254
Résumé.
L’évaluation des processus d’inhibition a une placecentrale en neuropsychologie comme en psychopathologie.De nombreux praticiens et chercheurs tiennent pour acquisqu’un déficit d’inhibition peut être évoqué pour rendre comptedes performances lors du vieillissement normal et pathologi-que. Si les fonctions essentielles remplies par les mécanis-mes inhibiteurs sont largement détaillées dans la littérature,la nature précise des processus sous-jacents mis en jeudemeure peu abordée. À ce propos, dans un article de 1997,Burke
(4)
met en exergue l’urgence de se concentrer sur unespécification de la théorie de l’inhibition qui demeure à ce jourvague et intuitive. Autrement dit, les phénomènes inhibiteursobservés sur les performances (
ie
ralentissement des tempsde réponse) ne sont pas interprétables de façon univoque parl’existence de processus cognitifs inhibiteurs centraux. Ceciest d’autant plus vrai dans le contexte de l’amorçage négatif,paradigme couramment employé lors de protocoles hospita-liers de recherche clinique (notamment en neuropsychologieet psychopathologie). Selon l’interprétation dorso-ventrale del’amorçage négatif, il existe 2 processus d’inhibition indépen-dants, l’un basé sur la localisation, l’autre sur l’identité desinformations. Néanmoins, la question de la multiplicité desfonctions inhibitrices et de la validité psychologique de l’inhi-bition comme processus organisateur de la pensée demeureà ce jour source d’oppositions. Pour notre part, nous propo-sons qu’une approche de l’inhibition en termes de niveauxde traitement autorise une compréhension unitaire et auto-matique de cette dernière. Nous défendons ainsi l’hypothèsefrontale de l’amorçage négatif selon laquelle la capacité àinhiber dépend des capacités de contrôle et de maintien desobjectifs des individus. Dans ce but, nous avons réalisé 2expériences à partir d’une version modifiée du paradigmed’amorçage négatif associé à une tâche de catégorisation surle genre. Nous montrons que la compatibilité des couplesamorce/cible donne lieu à un ralentissement des temps dedécision comparativement à une condition incompatible.Nous appelons ce ralentissement Effet de compatibilité néga-tive (NCE) que nous interprétons comme étant un indicateurde la mise en œuvre d’un mécanisme d’inhibition automati-
que. Le profil des performances entre jeunes et âgés ne dif-fère pas dès lors que la tâche nécessite peu d’attention con-trôlée (expérience 1). En revanche, les personnes âgées neprésentent pas de NCE dès lors que la tâche est complexe(expérience 2). Notre travail suggère qu’il est essentiel deprendre en compte la capacité générale des sujets, notam-ment leur capacité à maintenir les buts en cours, lorsque nouspostulons qu’il existe un déficit d’inhibition lors du vieillisse-ment. Nous discutons de ces résultats au regard de la théoriedes niveaux de traitement et des données récentes issuesde la neuro-imagerie.
Mots clés :
Amorçage négatif ; Contrôle ; Hypothèse dorso-ven-trale et frontale ; Inhibition ; Vieillissement.
INTRODUCTION
Considérations théoriques
D’un point de vue neurobiologique, les déficits liés àl’âge sont attribués à un dysfonctionnement du cortex pré-frontal (CPF), notamment des modulations dopaminergi-ques permettant la régulation de l’attention en directiondes informations pertinentes et des réponses appro-priées. Une diminution en récepteurs dopaminergiquesaurait pour conséquence une augmentation du rapportsignal/bruit au sein du CPF, réduisant ainsi sa sensibilitéaux signaux afférents et subséquemment une moindre dif-férenciation de ces derniers (23). D’un point de vue cogni-tif, la conséquence directe de ce phénomène serait unebaisse de l’élaboration des connaissances au cours duvieillissement du fait d’une régulation attentionnelle moinsefficiente. Un certain nombre de théories cognitives rendcompte de cette implication des lobes frontaux dans larégulation des comportements. Le modèle proposé parNorman et Shallice (28, 29), sur lequel repose l’évaluationneuropsychologique, représente la première systématisa-tion des fonctions cognitives dites exécutives sous-ten-
could be male or female. When prime and target matched on gender, we talked about VALID pairs (or compatible). Whenprime and target mismatched on the manipulated features, we talked about INVALID pairs (or incompatible). Participants’task was to identify the gender of the target. They were explicitly instructed not to respond to primes but to read themsilently. Our interest was in response latencies for valid
versus
invalid pairs. We manipulated task complexity by theabsence (experiment 1) or presence (experiment 2) of a distractor during probe trials.
Results.
For younger adults, primespresented before targets gave rise to behavioural costs when pairs were mapped to the same response compared topairs that were mapped to opposite ones. This slowing, called Negative Compatibility Effect (NCE), was independent ofthe presence or absence of a distractor. NCE was reliable for the elderly patients only under condition of no interferinginformation during the probe trial : pattern of performance of older participants was identical to that of young adults (expe-riment 1). This effect disappeared as task complexity increased (experiment 2).
Discussion.
This result suggests thatNCE triggering is dependant on the amount of cognitive control engaged by the task, and therefore that the ability toinhibit irrelevant information is secondary to a general capacity of the working memory.
Conclusion.
The implications ofour data are consistent with the level of processing account, as well as the recent neuroimaging contributions whichsuggest, for example, the involvement of the dorso-lateral prefrontal cortex (sensitive to aging) when task demands arehigh, and a ventro-lateral prefrontal implication when demands are low [see Eenshuistra
et al.
for a review
(10)
].
Key words :
Aging ; Control ; Dorsal-ventral and frontal model ; Inhibition ; Negative priming.

L’Encéphale, 2006 ;
32 :
253-62, cahier 1 Existe-t-il un déficit d’inhibition lors du vieillissement ?
255
dues par le cortex frontal. À mi-chemin entre la mémoireet l’attention, les fonctions exécutives regroupent touteactivité mentale impliquée dans la planification, l’initiationet la régulation du comportement. Selon Shallice (32), levieillissement des lobes frontaux aurait pour conséquenceune moindre efficience du système de supervision (SAS),
a fortiori
du contrôle exercé par l’administrateur central dela mémoire de travail. Ce contrôle s’effectue à partir d’inhi-bition et d’activation locale de schémas d’action en fonc-tion de leur pertinence. La pratique clinique fait une grandeplace à l’évaluation de l’efficience du fonctionnement exé-cutif chez tout patient présentant une plainte cognitive.Cette évaluation aboutit souvent à la conclusion d’un trou-ble de l’inhibition. Néanmoins, elle ne repose sur aucunmodèle théorique de l’inhibition. Si celui de Norman etShallice (28, 29) fait référence en la matière, le modèleconnexionniste de l’inhibition développé par Houghton etTipper (17, voir aussi 18, 38) nous semble tout à fait enmesure de rendre compte de la théorie frontale du vieillis-sement
(figure 1)
. Pour ces auteurs, l’attention aurait pourrôle principal de moduler les inputs perceptifs
(bottom-up)
en fonction des objectifs du sujet
(top-down)
, et ce dansle but de diriger ses actions et pensées de façon pertinenteet adaptée. En l’absence d’une telle modulation sélective,l’organisme serait éminemment distractible et sensible àl’interférence.
L’inhibition est conçue dans ce modèle comme unmécanisme sélectif et local, c’est-à-dire qui ne touche queles dimensions des informations distractrices entrant encompétition avec les buts que le sujet s’est fixé. Tipper
et al.
(38) argumentent largement leur hypothèse desélectivité à partir de données neurophysiologiques.
Le modèle de l’attention sélective de Houghton
et al.
(18) rend compte de façon heuristique du paradigmed’amorçage négatif (8, 24, 36). Ce paradigme est souventmis en place dans les protocoles hospitaliers de rechercheclinique car il est considéré comme l’indicateur de l’effi-cience avec laquelle un sujet est capable d’inhiber l’infor-mation distractrice issue de son environnement. Dans sondéveloppement classique, le paradigme d’amorçagenégatif implique la présentation de 2 essais se succédantrapidement. Le premier est appelé « amorce » et lesecond « sonde ». Lors de chaque essai, les participantsont pour tâche de ne traiter qu’une cible présentéeconjointement avec un ou des distracteurs. L’amorçagenégatif renvoie au fait qu’il faut plus de temps pour traiterune cible en sonde lorsque cette dernière est identiqueou entretient un lien avec le distracteur de la phase amorce
(figure 2)
.
Au cours du vieillissement, nous observons uneabsence de ralentissement en situation d’amorçage néga-tif dès lors que l’inhibition s’applique à l’identité des infor-mations distractrices. En revanche, les personnes âgéesralentissent de la même façon que les sujets plus jeunesdès lors que la dimension inhibée concerne la localisationdes distracteurs (5, 14). La dissociation obtenue avecl’âge entre inhibition de localisation (présence d’un AN,donc préservée) et inhibition d’identité (absence d’un AN,donc altérée) a donné lieu au postulat qu’il existe2 processus d’inhibition indépendants et différentielle-ment affectés par le vieillissement, le premier automatiqueet le second contrôlé. Selon l’hypothèse dorso-ventrale del’inhibition cognitive (21, 39), ces 2 processus seraientsous-tendus par des voies corticales distinctes en lienavec le cortex préfrontal, à savoir un chemin dorsal occi-pito-pariétal pour l’inhibition de localisation, et un cheminventral occipito-temporal pour l’inhibition de l’identité desstimuli. L’obtention d’une telle dissociation permet à cer-
FIG. 1. —
Architecture générale de l’attention sélective.
(Schéma modifié de Houghton, Tipper, Weaver & Shore, 1996, avecl’autorisation des auteurs.)
Output
Inputs externes
Système de réponse(liage des paramètres
des schémasde réponse activés)
Champ de l’objet(représentationd’ensemblesde percepts)
dirigé par les données
Détecteur descorrespondances(Match/Mismatch)
Champ de la cible(description interne
des stimuli attendus)
Système de supervision
Activation et maintiendes schémas de réponse
Activation et maintiende la description des cibles
FIG. 2. —
Situation d’amorçage négatif.
Les participants ont pour tâche de dénommer les cibles (en gras) etd’ignorer les distracteurs. Les temps de réponses sont plus lents pourla situation AN que pour la situation contrôle. La phase de sondage enAN permet donc d’avoir une mesure réactive des mécanismes inhibi-teurs mis en place sur le distracteur en phase d’amorçage (dans notreexemple sur CHAT).
Phased’amorçage (n)
Phasede sondage (n+1)
Situation AN TABLE
TABLE
AVION
AVION
CHAT CHAT
CHAT
Situation contrôle
CADRE

S. Martin
et al.
L’Encéphale, 2006 ;
32 :
253-62, cahier 1
256
tains auteurs de militer en faveur d’une famille de fonctionsinhibitrices plutôt qu’en faveur d’un mécanisme unique.Néanmoins, les résultats de la littérature sont inconstants,certains démontrant une préservation de l’inhibition baséesur l’identité au cours du vieillissement (20, 22, 31, 35).Cette absence de convergence forte des dissociations(22) ainsi que l’absence de corrélation entre différentesmesures d’inhibition censées reposer sur une même fonc-tion (11, 33) affaiblissent la position multifactorielle desprocessus inhibiteurs (11). Ce point de désaccord a par-ticulièrement retenu notre attention.
Le modèle de Houghton et Tipper (17) permet de pré-dire qu’une absence d’amorçage négatif n’indique pas defaçon univoque un déficit d’inhibition. Une absence d’effetpeut aussi bien être consécutive à un déficit d’inhibitionqu’à un échec du maintien du patron attentionnel concer-nant à la fois les buts du sujet et les moyens d’y parvenir.Ce point est essentiel dans le champ du vieillissement etde la psychopathologie où l’absence d’amorçage négatifconduit systématiquement à la conclusion d’un déficit desprocessus d’inhibition lors de la sénescence normale. Or,l’alternative frontale (9, 27, 41) à l’hypothèse dorso-ven-trale de l’inhibition nous paraît tout à fait appropriée pourenvisager les dissociations observées au cours du vieillis-sement, non plus en termes de processus indépendants,mais en termes de niveau de contrôle présidant à la miseen œuvre d’un mécanisme unitaire. Si, pour Hasher,Zacks et May (16), la conséquence d’un trouble de l’inhi-bition entraîne une moindre efficience du contrôle desinformations qui entrent et sortent de la mémoire de travail,nous proposons que c’est une moindre efficience du con-trôle, dépendant de l’intégrité du cortex préfrontal, quientraîne une absence de déclenchement du mécanismed’inhibition (15). Il s’agit dans ce travail de pointer la néces-sité de distinguer la mise en œuvre du mécanisme d’inhi-bition du mécanisme lui-même : existe-t-il réellement undéficit d’inhibition chez la personne âgée ? S’il s’avère queles personnes âgées ne sont pas déficitaires en inhibitionmais présentent une moindre efficience lors de la mise enœuvre des traitements coûteux, il sera alors indispensablede considérer l’inhibition non plus comme un ensemble desous-processus mais comme un mécanisme unitaire,local et automatique faisant partie tantôt de routines, tantôtd’opérations contrôlées. Nous pensons que la démons-tration d’une dissociation en fonction du niveau de traite-ment engagé par une même tâche au sein d’un mêmegroupe d’âge (âgés) et non entre les groupes d’âge (jeu-nes
vs
âgés) permettra de répondre à cette question.
Dans ce but, nous avons réalisé 2 expériences avecdes participants jeunes et âgés, nous basant sur un para-digme d’amorçage négatif modifié associé à une tâche decatégorisation sur le genre. À notre connaissance, uneseule étude s’est appuyée sur la même procédure quenous (40). Les deux expériences nous ont permis de fairevarier le niveau de contrôle engagé par la tâche qui néces-site le traitement d’une dimension conceptuelle de l’infor-mation non fournie par l’environnement, à savoir le genre.
Dans la première expérience
, les participants voient2 mots se succéder à l’écran, une amorce (phase d’amor-
çage) et une cible (phase de sondage) sans lien apparentet présentées isolément. Ils ont pour consigne de donnerle genre grammatical des cibles. Toute réponse associéeaux amorces est explicitement interdite, le respect de cetteconsigne devant donner lieu à une inhibition des informa-tions amorces considérées comme distractrices. La moitiédes couples amorce/cible partage le même genre, l’autremoitié présente des genres différents. De plus, sans queles participants en soient informés, amorces et cibles ren-voient ou non à la même catégorie (naturelle
vs
artificielle).Nous manipulons ainsi la compatibilité des amorces et descibles sur la base de leur genre et de la catégorie à laquelleelles renvoient. Nous prédisons des temps de réponseplus longs lorsque amorce et cible sont compatibles parrapport à la situation où amorce et cible sont incompati-bles. Nous appellerons ce ralentissement « effet de com-patibilité négative » (ECN). Nous pensons que l’ECN estun phénomène sélectif permettant aux sujets de s’adapteraux demandes liées à la tâche en cours. De fait, seulesles propriétés d’une information entretenant un lien directavec les buts du participant seront sujettes à inhibition.Autrement dit, seule la compatibilité sur le genre engen-drera un ralentissement des temps de réponse alors quela compatibilité sur la catégorie favorisera la performance.
Dans la seconde expérience
, la tâche des participantsest identique. La seule différence repose sur le fait queles cibles sont présentées simultanément avec un distrac-teur. Nous augmentons ainsi le contrôle cognitif néces-saire à l’atteinte des buts : les participants doivent à la foisrésister au traitement des amorces en phase d’amorçage,mais aussi à celui des distracteurs en phase de sondage.Selon l’hypothèse frontale de l’inhibition cognitive, nousprédisons que les personnes âgées presenteront le mêmeprofil de performances que les participants jeunes dès lorsque la tâche est routinisable, c’est-à-dire dès lors qu’ellerepose sur un faible contrôle cognitif (expérience 1). Enrevanche, les participants âgés ne présenteront pasd’effet de compatibilité négative (ECN) dès lors que latâche nécessite la mise en œuvre contrôlée des mécanis-mes d’inhibition (expérience 2). Selon l’hypothèse dorso-ventrale, le groupe âgé ne devrait pas présenter d’ECN,et ce quel que soit le niveau de traitement engagé par latâche, cette dernière reposant sur une analyse concep-tuelle des informations.
EXPÉRIENCE 1
Participants
Au total, 16 adultes jeunes (moyenne d’âge = 22,6 ans+/– 1,52 ans) et 16 âgés (moyenne d’âge = 71,71 +/– 4,9 ans) ont participé à cette première expérience lorsd’une session unique de 20 minutes. Le groupe de parti-cipants âgés était constitué de sujets recrutés par voied’affichage ou par recommandation, vivant de façon auto-nome à domicile. Tous présentaient un score de 26 et plusau MMSE ainsi qu’un score inférieur à 15 à l’échelle dedépression MADRS. Aucun ne rapportait de plainte cogni-tive spontanée significative. Les participants âgés étant

L’Encéphale, 2006 ;
32 :
253-62, cahier 1 Existe-t-il un déficit d’inhibition lors du vieillissement ?
257
évalués uniquement l’après-midi (entre 14 h et 18 h), àsavoir au moment de la journée le moins optimal du pointde vue de leur capacité cognitive (25, 26, 42, 43), nousavons aussi testé les participants jeunes au moment lemoins optimal au regard leur propre performance, à savoirle matin de 8 h à 12 h et ce afin de ne pas maximiser lesdifférences liées à l’âge. Tous les participants avaient unevue normale ou corrigée et ont tous été testés individuel-lement sur un ordinateur HP Pavillon ze4300 Athlon àl’aide du logiciel E-prime (Psychology SoftwareTools, Inc.,2001). Les participants jeunes ont été évalués à l’Univer-sité des Lettres et des Sciences Humaines de Montpellier,et l’ensemble des participants âgés à leur domicile. Aucundes participants n’était rémunéré.
Matériel
Au total, 128 mots ont été utilisés afin de construire 64paires amorce/cible. Amorces et cibles ne se différencientni sur leur fréquence (entre 4 et 27 occurrences par mil-lion*), ni sur leur nombre de lettres (entre 3 et 9), ni surleur nombre de syllabes (entre 1 et 3). Les paires nerimaient pas et n’entretenaient aucun lien sémantique évi-dent. La moitié des couples était constituée d’amorces etde cibles partageant le même genre (conditioncompatible : G+), l’autre moitié, d’amorces et de cibles degenres différents (condition incompatible : G–). De plus,50 % des couples étaient constitués de mots appartenantà la même catégorie (C+), et de 50 % de mots appartenantà des catégories différentes (C–). La compatibilité sur legenre était contrebalancée entre les items : un même motassocié à une amorce compatible (
ie
peigne/briquet) était,pour un autre participant, associé à une amorce incom-patible (
ie
poubelle/briquet). Le
tableau I
présente unexemple de chaque type de couples utilisés dans cettepremière expérience.
Procédure
L’expérience débute par un bloc d’entraînement de32 essais, suivi d’un bloc expérimental de 64 essais. Ledéroulement de chaque essai est présenté sur la
figure 3
.Nous avons systématiquement pondéré le ralentissementcognitif lié au vieillissement en appliquant un coefficientmoyen de ralentissement de 1,6 sur les temps d’expositiondes amorces et des cibles chez les âgés (2, 30).
Les participants avaient pour consigne de placer leurregard sur la croix de fixation au début de chaque nouvelessai. Leur tâche était de donner, le plus rapidement(maximum 1 200 ms) et correctement possible, le genredes mots cibles. Les touches réponses étaient contreba-lancées entre les sujets. Un ordre aléatoire a été construitpour chaque sujet. Les sujets n’avaient pas consciencede la manipulation de la compatibilité sur la catégorie. Illeur était demandé de lire silencieusement les amorces,
mais de ne pas y répondre. Les essais pour lesquels lesparticipants n’ont pas respecté cette interdiction de répon-dre aux amorces ont été éliminés.
Résultats
Les temps de réaction moyens
(tableau II)
ont été sou-mis à une analyse de variance avec l’âge comme facteurinter-sujet et les facteurs genre et catégorie comme facteursintra-sujets. Nous éliminons des analyses les temps liés auxréponses erronées ainsi que ceux s’écartant de 2,5 dévia-tions standard de la moyenne du groupe, ce qui représentemoins de 3 % des données. L’analyse statistique ne révèleaucune interaction significative d’aucun ordre entre les dif-férents facteurs manipulés. De façon non surprenante,nous obtenons un effet de l’âge [F(1, 30) = 10,45,p < 0,001)] avec un temps moyen de réponse plus lent pourles personnes âgées. Les effets globaux des facteurs genre[F(1, 30) = 23,96, p < 0,001)] et catégorie [F(1, 30) = 29,91,p < 0,001)] sont tout deux significatifs et opposés : lestemps de réponses sont plus lents lorsque amorce et ciblespartagent le même genre, en revanche, les participantsrépondent plus rapidement lorsque les couples renvoientà la même catégorie. Nous obtenons ainsi un effet signifi-catif de la compatibilité négative liée au genre (ECNG) etde la compatibilité positive liée à la catégorie (ECPC).
Les analyses partielles réalisées pour chaque grouped’âge sont d’un plus grand intérêt en ce qui concerne notreproblématique. L’effet de compatibilité négative liée augenre (ECNG) demeure significatif chez les adultes jeu-nes [coût de 19,58 ms, F(1, 15) = 11,24, p < 0,005)]comme chez les âgés [coût de 25,69 ms, F(1, 30) = 12,84,p < 0,005)]. Ces résultats sont consistants dans le sensoù 87,5 % des participants jeunes (z = 2,53, p < 0,01, Wil-coxon rank) et 81,25 % des participants plus âgés
* Lexique.org, News et Ferrand, 2003, pour la version la plusrécente.
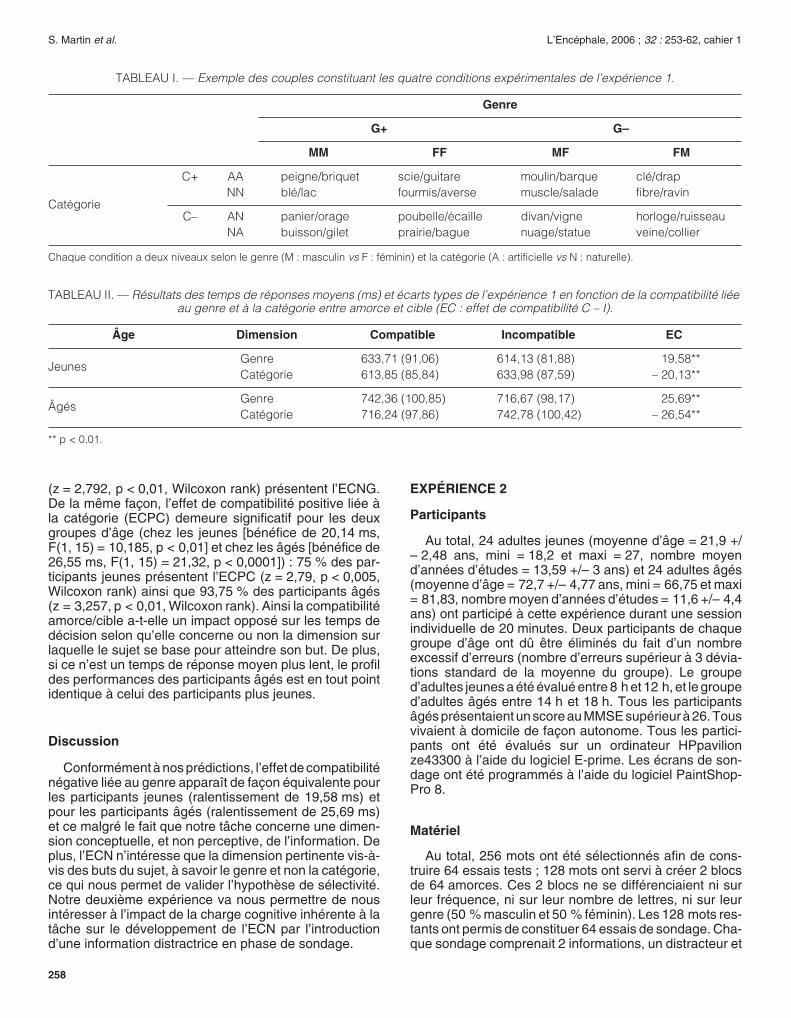
FIG. 3. —
Exemple d’essai de l’expérience 1 pour le groupe de participants jeunes.
Les informations sont toutes écrites en blanc sur écran noir en policeTimes 20. La durée d’exposition des amorces et des cibles variait doncen fonction de l’âge des sujets, à savoir 350 ms pour les sujets jeuneset 560 ms pour les plus âgés. Les participants doivent décider si lescibles renvoient à un mot masculin ou féminin. Ils n’étaient pas avertisde la manipulation sur la catégorie. Il leur était explicitement interdit derépondre aux amorces.
AMORCE
+
CIBLE
1 000 ms
500 ms
350 ms
1 000 ms
350 ms
Temps
S. Martin
et al.
L’Encéphale, 2006 ;
32 :
253-62, cahier 1
258
(z = 2,792, p < 0,01, Wilcoxon rank) présentent l’ECNG.De la même façon, l’effet de compatibilité positive liée àla catégorie (ECPC) demeure significatif pour les deuxgroupes d’âge (chez les jeunes [bénéfice de 20,14 ms,F(1, 15) = 10,185, p < 0,01] et chez les âgés [bénéfice de26,55 ms, F(1, 15) = 21,32, p < 0,0001]) : 75 % des par-ticipants jeunes présentent l’ECPC (z = 2,79, p < 0,005,Wilcoxon rank) ainsi que 93,75 % des participants âgés(z = 3,257, p < 0,01, Wilcoxon rank). Ainsi la compatibilitéamorce/cible a-t-elle un impact opposé sur les temps dedécision selon qu’elle concerne ou non la dimension surlaquelle le sujet se base pour atteindre son but. De plus,si ce n’est un temps de réponse moyen plus lent, le profildes performances des participants âgés est en tout pointidentique à celui des participants plus jeunes.
Discussion
Conformément à nos prédictions, l’effet de compatibiliténégative liée au genre apparaît de façon équivalente pourles participants jeunes (ralentissement de 19,58 ms) etpour les participants âgés (ralentissement de 25,69 ms)et ce malgré le fait que notre tâche concerne une dimen-sion conceptuelle, et non perceptive, de l’information. Deplus, l’ECN n’intéresse que la dimension pertinente vis-à-vis des buts du sujet, à savoir le genre et non la catégorie,ce qui nous permet de valider l’hypothèse de sélectivité.Notre deuxième expérience va nous permettre de nousintéresser à l’impact de la charge cognitive inhérente à latâche sur le développement de l’ECN par l’introductiond’une information distractrice en phase de sondage.
EXPÉRIENCE 2
Participants
Au total, 24 adultes jeunes (moyenne d’âge = 21,9 +/– 2,48 ans, mini = 18,2 et maxi = 27, nombre moyend’années d’études = 13,59 +/– 3 ans) et 24 adultes âgés(moyenne d’âge = 72,7 +/– 4,77 ans, mini = 66,75 et maxi= 81,83, nombre moyen d’années d’études = 11,6 +/– 4,4ans) ont participé à cette expérience durant une sessionindividuelle de 20 minutes. Deux participants de chaquegroupe d’âge ont dû être éliminés du fait d’un nombreexcessif d’erreurs (nombre d’erreurs supérieur à 3 dévia-tions standard de la moyenne du groupe). Le grouped’adultes jeunes a été évalué entre 8 h et 12 h, et le grouped’adultes âgés entre 14 h et 18 h. Tous les participantsâgés présentaient un score au MMSE supérieur à 26. Tousvivaient à domicile de façon autonome. Tous les partici-pants ont été évalués sur un ordinateur HPpavilionze43300 à l’aide du logiciel E-prime. Les écrans de son-dage ont été programmés à l’aide du logiciel PaintShop-Pro 8.
Matériel
Au total, 256 mots ont été sélectionnés afin de cons-truire 64 essais tests ; 128 mots ont servi à créer 2 blocsde 64 amorces. Ces 2 blocs ne se différenciaient ni surleur fréquence, ni sur leur nombre de lettres, ni sur leurgenre (50 % masculin et 50 % féminin). Les 128 mots res-tants ont permis de constituer 64 essais de sondage. Cha-que sondage comprenait 2 informations, un distracteur et
TABLEAU I. —
Exemple des couples constituant les quatre conditions expérimentales de l’expérience 1.
Genre
G+ G–
MM FF MF FM
Catégorie
C+ AA peigne/briquet scie/guitare moulin/barque clé/drapNN blé/lac fourmis/averse muscle/salade fibre/ravin
C– AN panier/orage poubelle/écaille divan/vigne horloge/ruisseauNA buisson/gilet prairie/bague nuage/statue veine/collier
Chaque condition a deux niveaux selon le genre (M : masculin
vs
F : féminin) et la catégorie (A : artificielle
vs
N : naturelle).
TABLEAU II. —
Résultats des temps de réponses moyens (ms) et écarts types de l’expérience 1 en fonction de la compatibilité liée au genre et à la catégorie entre amorce et cible (EC : effet de compatibilité C – I).
Âge Dimension Compatible Incompatible EC
JeunesGenre 633,71 (91,06) 614,13 (81,88) 19,58**Catégorie 613,85 (85,84) 633,98 (87,59) – 20,13**
ÂgésGenre 742,36 (100,85) 716,67 (98,17) 25,69**Catégorie 716,24 (97,86) 742,78 (100,42) – 26,54**
** p < 0,01.
L’Encéphale, 2006 ;
32 :
253-62, cahier 1 Existe-t-il un déficit d’inhibition lors du vieillissement ?
259
une cible, présentés au-dessus et au-dessous d’une croixde fixation blanche sur fond d’écran noir. Dans la mesuredu possible, cible et distracteur étaient appariés sur leurnombre de lettres et leur fréquence. Nous avons manipuléla relation sur la dimension du genre entre amorce et ciblemais aussi entre amorce et distracteur. Nous obtenons4 conditions comprenant chacune 25 % des essais
(tableau III)
:1) Condition CI : les paires amorce/cible étaient com-
patibles et amorce/distracteur étaient incompatibles.2) Condition CC : les paires amorce/cible et amorce/
distracteur étaient compatibles.3) Condition II : les paires amorce/cible et amorce/dis-
tracteur étaient incompatibles.4) Conditions IC : les paires amorce/cible étaient
incompatibles et amorce/distracteur étaient compatibles.Notre objectif n’étant pas d’étudier l’influence des
dimensions ne participant pas à la sélection, nous avonsfait le choix de bloquer la variable catégorie préalablementmanipulée. Dans ce but, nous avons exclusivement cons-truit des essais compatibles sur leur catégorie (50 % natu-relle et 50 % artificielle).
Les conditions expérimentales CC et II n’ont pas faitl’objet d’un traitement statistique car elles impliquent quela cible et le distracteur soient du même genre. Il nous estdonc impossible de savoir si le participant a répondu à la
cible ou au distracteur, autrement dit si les réponses desparticipants sont correctes ou erronées. Nous avonsnéanmoins inclus ces 2 situations afin de rendre la sélec-tion plus difficile par l’augmentation de la confusionqu’elles entraînent. Nous espérons de la sorte réduiretoute stratégie individuelle basée sur la détection d’uneopposition entre le genre de la cible et celui du distracteursi ces deux informations étaient systématiquement incom-patibles. Nous analyserons ainsi l’effet de la compatibilitéliée au genre uniquement sur les essais pour lesquels cibleet distracteur sont de genre différent.
Procédure
L’expérience débute avec 20 essais d’entraînement sui-vis de 64 essais tests. La session test commence unique-ment si les participants ont répondu correctement à aumoins 90 % des essais d’entraînement. Chaque essai com-mence par un écran sur lequel est inscrit « Prêt » quidemeure affiché jusqu’à ce que le participant appuie sur labarre espace. Cet écran permet à l’expérimentateur ou auparticipant d’interrompre l’expérience lorsqu’il le souhaite.Tout comme l’expérience 1, nous avons pondéré le ralen-tissement cognitif. La suite du déroulement de chaque essaiest présentée sur la
figure 4
. Une expérience pilote a mon-
TABLEAU III. —
Exemple de chaque condition de l’expérience 2. En gras, la cible (M = masculin et F = féminin).
LienAmorce/Cible
LienAmorce/
DistracteurConditions
Amorce Masque ISI Cible
350 ms 100 ms 1 000 ms Max 2 000 ms
Incompatible CI (Test)
M §#§#§#§
M
+ F
F §#§#§#§
F
+ M
Compatible
Compatible CC
M §#§#§#§
M
+ M
F §#§#§#§
F
+ F
Incompatible IC (Test)
F §#§#§#§
M
+ F
M §#§#§#§
F
+ M
Incompatible
Compatible II
F §#§#§#§
M
+ M
M §#§#§#§
F
+ F
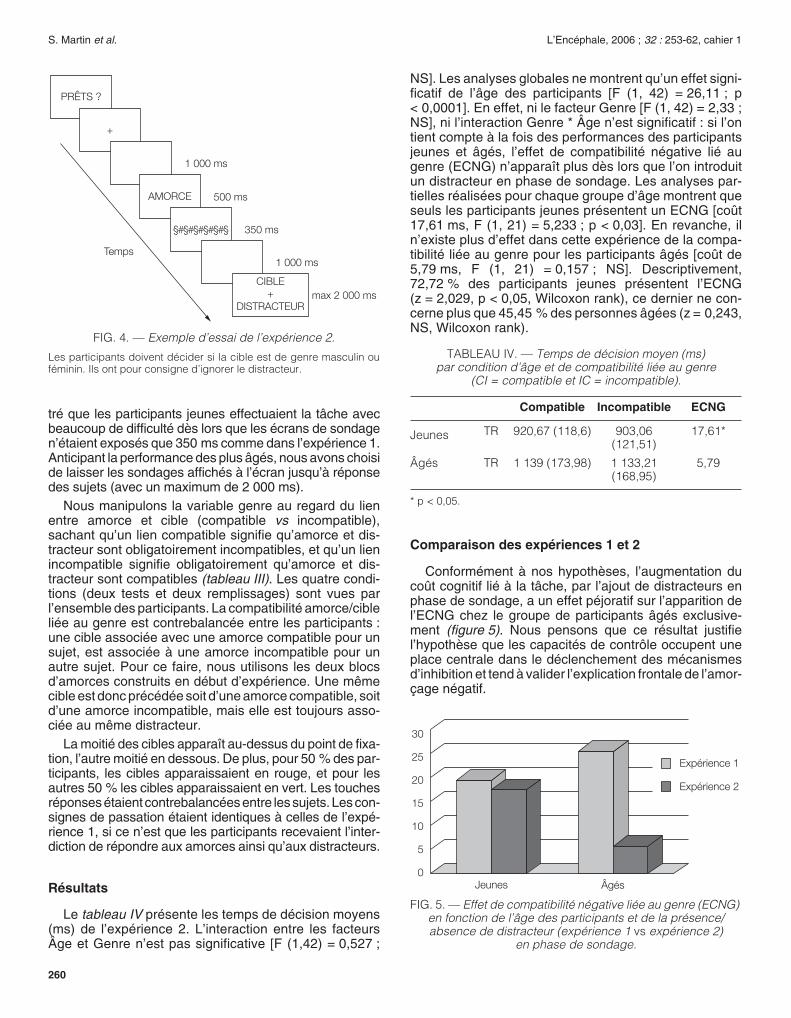
S. Martin
et al.
L’Encéphale, 2006 ;
32 :
253-62, cahier 1
260
tré que les participants jeunes effectuaient la tâche avecbeaucoup de difficulté dès lors que les écrans de sondagen’étaient exposés que 350 ms comme dans l’expérience 1.Anticipant la performance des plus âgés, nous avons choiside laisser les sondages affichés à l’écran jusqu’à réponsedes sujets (avec un maximum de 2 000 ms).
Nous manipulons la variable genre au regard du lienentre amorce et cible (compatible
vs
incompatible),sachant qu’un lien compatible signifie qu’amorce et dis-tracteur sont obligatoirement incompatibles, et qu’un lienincompatible signifie obligatoirement qu’amorce et dis-tracteur sont compatibles
(tableau III)
. Les quatre condi-tions (deux tests et deux remplissages) sont vues parl’ensemble des participants. La compatibilité amorce/cibleliée au genre est contrebalancée entre les participants :une cible associée avec une amorce compatible pour unsujet, est associée à une amorce incompatible pour unautre sujet. Pour ce faire, nous utilisons les deux blocsd’amorces construits en début d’expérience. Une mêmecible est donc précédée soit d’une amorce compatible, soitd’une amorce incompatible, mais elle est toujours asso-ciée au même distracteur.
La moitié des cibles apparaît au-dessus du point de fixa-tion, l’autre moitié en dessous. De plus, pour 50 % des par-ticipants, les cibles apparaissaient en rouge, et pour lesautres 50 % les cibles apparaissaient en vert. Les touchesréponses étaient contrebalancées entre les sujets. Les con-signes de passation étaient identiques à celles de l’expé-rience 1, si ce n’est que les participants recevaient l’inter-diction de répondre aux amorces ainsi qu’aux distracteurs.
Résultats
Le
tableau IV
présente les temps de décision moyens(ms) de l’expérience 2. L’interaction entre les facteursÂge et Genre n’est pas significative [F (1,42) = 0,527 ;
NS]. Les analyses globales ne montrent qu’un effet signi-ficatif de l’âge des participants [F (1, 42) = 26,11 ; p< 0,0001]. En effet, ni le facteur Genre [F (1, 42) = 2,33 ;NS], ni l’interaction Genre * Âge n’est significatif : si l’ontient compte à la fois des performances des participantsjeunes et âgés, l’effet de compatibilité négative lié augenre (ECNG) n’apparaît plus dès lors que l’on introduitun distracteur en phase de sondage. Les analyses par-tielles réalisées pour chaque groupe d’âge montrent queseuls les participants jeunes présentent un ECNG [coût17,61 ms, F (1, 21) = 5,233 ; p < 0,03]. En revanche, iln’existe plus d’effet dans cette expérience de la compa-tibilité liée au genre pour les participants âgés [coût de5,79 ms, F (1, 21) = 0,157 ; NS]. Descriptivement,72,72 % des participants jeunes présentent l’ECNG(z = 2,029, p < 0,05, Wilcoxon rank), ce dernier ne con-cerne plus que 45,45 % des personnes âgées (z = 0,243,NS, Wilcoxon rank).
Comparaison des expériences 1 et 2
Conformément à nos hypothèses, l’augmentation ducoût cognitif lié à la tâche, par l’ajout de distracteurs enphase de sondage, a un effet péjoratif sur l’apparition del’ECNG chez le groupe de participants âgés exclusive-ment
(figure 5)
. Nous pensons que ce résultat justifiel’hypothèse que les capacités de contrôle occupent uneplace centrale dans le déclenchement des mécanismesd’inhibition et tend à valider l’explication frontale de l’amor-çage négatif.
FIG. 4. —
Exemple d’essai de l’expérience 2.
Les participants doivent décider si la cible est de genre masculin ouféminin. Ils ont pour consigne d’ignorer le distracteur.
§#§#§#§#§#§
AMORCE
PRÊTS ?
+
CIBLE+
DISTRACTEUR
1 000 ms
500 ms
350 ms
1 000 ms
max 2 000 ms
Temps
TABLEAU IV. —
Temps de décision moyen (ms) par condition d’âge et de compatibilité liée au genre
(CI = compatible et IC = incompatible).
Compatible Incompatible ECNG
Jeunes TR 920,67 (118,6) 903,06 (121,51)
17,61*
Âgés TR 1 139 (173,98) 1 133,21 (168,95)
5,79
* p < 0,05.
FIG. 5. —
Effet de compatibilité négative liée au genre (ECNG) en fonction de l’âge des participants et de la présence/absence de distracteur (expérience 1
vs
expérience 2) en phase de sondage.
0Jeunes Âgés
5
10
15
20
25
30
Expérience 1
Expérience 2

L’Encéphale, 2006 ;
32 :
253-62, cahier 1 Existe-t-il un déficit d’inhibition lors du vieillissement ?
261
DISCUSSION
L’hypothèse actuellement dominante est qu’il existe2 processus d’inhibition indépendants, un basé sur lalocalisation des informations, l’autre sur leur identité(hypothèse dorso-ventrale). La validité de cette hypothèses’appuie largement sur les dissociations obtenues aucours du vieillissement dont nous proposons une alterna-tive interprétative : il n’existe pas 2 processus indépen-dants mais un seul mécanisme d’inhibition, dont la miseen jeu, tantôt automatique tantôt contrôlée, dépend desdemandes liées à la tâche (hypothèse frontale). Nos résul-tats (expérience 1) montrent que le profil des performan-ces entre jeunes et âgés ne diffère pas dès lors que latâche permet la mise en œuvre d’une activité à faible con-trôle attentionnel, c’est-à-dire réalisable sans l’interven-tion du système de supervision attentionnel (32). Enrevanche, les participants âgés ne présentent plus deralentissement dès lors que la tâche implique un contrôlecomportemental important (expérience 2). Nous pensonsque ces données vont dans le sens d’une implication fortedu cortex frontal dans la mise en œuvre de l’inhibition. Plusprécisément, elles indiquent que le déclenchement del’inhibition dépend d’un facteur intermédiaire, la capacitéde contrôle de la mémoire de travail (7), autrement dit dela capacité de travail de la mémoire, plus ou moins solli-citée selon le type de traitement en cours (6, 34). Dansce sens, nos résultats permettent de considérer certainesdonnées de la littérature avec un regard différent consis-tant à envisager les tâches en termes de niveau de trai-tement plutôt qu’en termes de paradigmes qui, du statutd’outils acquièrent souvent celui d’objet d’étude.
Ainsi, à l’hypothèse dorso-ventrale de l’amorçagenégatif qui repose sur une dissociation explicable par desaspects structurels de la cognition, nous préférons unehypothèse frontale qui implique de s’intéresser auxaspects fonctionnels de cette dernière. Dans ce contexte,l’amorçage négatif d’identité (19, 37) serait une tâcheimpliquant une analyse conceptuelle des informationsreposant sur un contrôle cognitif élevé, alors que l’amor-çage négatif de localisation serait une tâche réalisable àpartir d’une analyse perceptive de l’environnement, doncpeu coûteuse en ressource (5). Les données issues de laneuro-imagerie montrent qu’il existe des corrélats neuro-anatomiques cohérents avec l’hypothèse des niveaux detraitements (1) : la réalisation de tests conceptuels est cor-rélée avec des changements métaboliques du cortex tem-poral et frontal gauche, alors que la réalisation d’une tâchede complétion de fragments, reposant sur une analyseperceptive des informations, engage les régions corticalespostérieures, notamment le cortex occipital. De même, lestravaux de Gabrieli
et al.
(12)
et de Gardiner
et al.
(13)révèlent une activation prépondérante du cortex antérieurlors d’un encodage conceptuel de l’environnement, et ducortex postérieur lors de son traitement perceptif. Finale-ment, Eenshuistra
et al.
(10) rappellent l’existence d’unedissociation au sein même du cortex préfrontal, avec uneimplication majeure du cortex dorso-latéral (sensible auxeffets de l’âge) lorsque la tâche en cours nécessite un con-trôle cognitif élevé et un engagement ventro-latéral lors-
que le maintien des buts repose sur une implication mini-male de la mémoire de travail.
CONCLUSION
Nous pensons que l’ensemble des résultats issus dela littérature ainsi que les observations cliniques gagne-raient à être réinterprétés en termes de niveau de traite-ment dont l’avantage est de prendre en compte le fonc-tionnement du sujet en interaction avec sonenvironnement (3). Ce point est essentiel dans le domainede la prise en charge neuropsychologique mais aussi psy-chopathologique car il pourrait permettre d’éviter la con-fusion entre une manifestation observable (inhibition com-portementale) et le processus interne censé lui donnernaissance (inhibition cognitive) : une inhibition comporte-mentale peut être consécutive à l’incapacité de maintenirà moyen terme les buts en cours ou les moyens d’y par-venir, et non à un déficit des mécanismes responsablesde l’élimination des informations ou actions non perti-nentes.
Références
1. BLAXTON TA. Investigating dissociations among memorymeasures : support for a transfert-appropriate processing fra-mework. J Exp Psychol Learn Mem Cogn 1989 ; 15 : 657-68.
2. BRINLEY JF. Cognitive sets, speed and accuracy of performancein the elderly.
In
: Welford AT, Birren JE, eds. Behavior, Aging, andthe Nervous System. Springfield, IL : Thomas, 1965 : 114-49.
3. BROUILLET D, MARTIN S. La psychologie cognitive à l’écoute duvieillissement.
In
: Talepin JM, ed. À définir (Ouvrage collectif sur levieillissement). Paris : Dunod, 2005.
4. BURKE DM. Language, aging and inhibitory deficits : evaluation ofa theory. J Gerontol Psychol Sci 1997 ; 52B : 254-64.
5. CONNELLY S L, HASHER L. Aging and the inhibition of spatial loca-tion. J Exper Psychol Hum Percept Perform 1993 ; 19 : 238-50.
6. CONWAY ARA, TUHOLSKI S W, SHISLER RJ
et al
. The effect ofmemory load on negative priming : an individual differences inves-tigation. Mem Cogn 1999 ; 27 : 1042-50.
7. COWAN N. Evolving conceptions of memory storage, selectiveattention and their mutual constraints within the human information-processing system. Psychol Bull 1988 ; 104 : 163-91.
8. DALRYMPHLE-ALFORD EC, BUDAYR B. Examination of someaspects of the Stroop color-word test. Percept Mot Skills 1966 ; 23 :1211-4.
9. DEMPSTER FN. The rise and fall of the inhibitory mechanism :toward a unified theory of cognitive development and aging. DevelopRev 1992 ; 12 : 45-75.
10. EENSHUISTRA RM, RIDDERINKHOF KR, VAN DER MOLEN MW.Age-related changes in antisaccade task performance : inhibitorycontrol or working-memory engagement ? Brain Cogn 2004 ; 56 :177-88.
11. FRIEDMAN NP, MIYAKE A. The relations among inhibition and inter-ference control functions : a latent-variable analysis. J Exper PsycholGen 2004 ; 133 (1) : 101-35.
12. GABRIELI JDE, DESMOND JE, DEMB JB et al. Functional magneticresonance imaging of semantic memory processes in the frontallobes. Psychol Sci 1996 ; 7 : 278-83.
13. GARDINER JM, RICHARDSON-KLAVEHN A, RAMPONI C et al.Involuntary levels of processing effects in perceptual and conceptualpriming. In : Naveh-Benjamin M, Moscovitch M, Roediger HL, eds.

S. Martin et al. L’Encéphale, 2006 ; 32 : 253-62, cahier 1
262
Perspectives of human memory and cognitive aging : essays inhonour of Fergus Craig. New-York : Psychology Press NY, 2002.
14. GUIDI A. Inhibition et vieillissement : l’amorçage négatif dans unetâche d’identification ou de localisation. Unpublished Experimentaland Cognitive Psychology Thesis, Université Paul Valery, 2003.
15. HAMBRICK DZ, ENGLE RW. Effects of domain knowledge, workingmemory capacity, and age on cognitive performance : an investiga-tion of the knowledge-is-power hypothesis. Cogn Psychol 2002 ; 44 :339-87.
16. HASHER L, ZACKS RT, MAY CP. Inhibitory control, circadian arou-sal, and age. In : Gopher D, Koriat A, eds. Attention and PerformanceXVII, Cognitive Regulation of Performance : Interaction of Theoryand Application. Cambridge : MA : MIT Press, 1999 : 653-75.
17. HOUGHTON G, TIPPER SP. A model of inhibitory mechanisms inselective attention. In : Dagenbach D, Carr TH, eds. Inhibitory pro-cesses in attention, memory, and language. San Diego, CA : Aca-demic Press, 1994 : 53-112.
18. HOUGHTON G, TIPPER SP, WEAVER B et al. Inhibition and inter-ference in selective attention : some tests of a neural network model.Vis Cogn 1996 ; 3 : 119-64.
19. KANE MJ, HASHER L, STOLTZFUS ER et al. Inhibitory attentionalmechanisms and aging. Psychol Aging 1994 ; 9 : 103-12.
20. KIELY JM, HARTLEY AA. Age-related equivalence of identity sup-pression in the Stroop color-word task. Psychol Aging 1997 ; 12 :22-9.
21. KOSHINO H, BOESE GA, FERRARO FR. The relationship betweencognitive ability and positive and negative priming in identity and spa-tial priming tasks (statistical data included). J Gen Psychol 2000 ;127 (4) : 372-82.
22. KRAMER AF, HUMPHREY DG, LARISH JF et al. Aging and inhibi-tion : beyond a unitary view of inhibitory processing in attention. Psy-chol Aging 1994 ; 9 : 491-512.
23. LI SC, LINDENBERGER U, SIKSTRÖM S. Aging cognition : fromneuromodulation to representation. Trends Cogn Sci 2001 ; 5 (11) :479-85.
24. MARTIN S. De l’inhibition contrôlée au contrôle de l’inhibition : uneapproche fonctionnelle du déficit lors du vieillissement. UnpublishedExperimental and Cognitive Psychology Thesis, Paul Valéry, Mont-pellier, 2004.
25. MAY CP, HASHER L. Synchrony effects in inhibitory control overthought and action. J Exper Psychol Hum Percept Perform 1998 ;24 : 363-79.
26. MAY CP, HASHER L, STOLTZFUS ER. Optimal time of day andthe magnitude of age differences in memory. Psychol Sci 1993 ; 4 :326-30.
27. MILLER EK, COHEN JD. An integrative theory of prefrontal cortexfunction. Annu Rev Neurosci 2001 ; 24 : 167-202.
28. NORMAN D, SHALLICE T. Attention to action : willed and automaticcontrol of behavior (CHIP Report n° 99). San Diego : University ofCalifornia, 1980.
29. NORMAN DA, SHALLICE T. Attention to action : willed and auto-matic control of behaviour. In : Davidson RJ, Schartz GE, ShapiroD, eds. Consciousness and self-regulation (vol 4). New York : Ple-num Press, 1986 : 1-18.
30. SALTHOUSE TA. Speed of behaviour and its implications for cogni-tion. In : Schaie KW, Birren JE, eds. Handbook of the Psychologyaging. New York : Van Nostrand Reinhold, 1985 : 400-26.
31. SCHOOLER C, NEUMANN E, CAPLAN LJ et al. Continued inhibi-tory capacity throughout adulthood : conceptual negative priming inyounger and older adults. Psychol Aging 1997 ; 12 (4) : 667-74.
32. SHALLICE T. Multiple levels of controls processes. In : Umita C,Moscovitch M, eds. Attention and performance, XV. Conscious andnonconscious information processing. Cambridge, MA : Bradford/MIT Press, 1994 : 395-420.
33. SHILLING VM, CHETWYND A, RABBITT PMA. Individual inconsis-tency across measures of inhibition : an investigation of the constructvalidity of inhibition in older adults. Neuropsychologia 2002 ; 40 :605-19.
34. STUSS DT, TOTH JP, FRANCHI D et al. Dissociation of attentionalprocesses in patients with focal frontal and posterior lesions. Neu-ropsychologia 1999 ; 37 : 1005-27.
35. SULLIVAN MP, FAUST ME. Evidence for identity inhibition duringselective attention in old adults. Psychol Aging 1993 ; 8 : 589-98.
36. TIPPER SP. The negative priming effect : inhibitory priming by igno-red objects. Quart J Exper Psychol Hum Exper Pychol 1985 ; 37A :571-90.
37. TIPPER SP. Less attentional selectivity as a result of declining inhi-bition in older adults. Bull Psychonom Soc 1991 ; 29 : 45-7.
38. TIPPER SP, WEAVER B, HOUGHTON G. Behavioural goals deter-mine inhibitory mechanisms of selective attention. Quart J ExperPsychol 1994 ; 47A : 809-40.
39. UNGERLEIDER LG, MISHKIN M. Two cortical visual systems. In :Ingle MAG, Mansfield RJW, eds. Analysis of visual behavior. Cam-bridge, MA : MIT Press, 1982 : 549-86.
40. VERSACE R, ALLAIN G. Negative priming in a gender decision taskand in a semantic categorization task. Acta Psychol 2001 ; 108 : 73-90.
41. WEST RL. An application of prefrontal cortex function theory tocognitive aging. Psychol Bull 1996 ; 120 : 272-92.
42. WINOCUR G HASHER L. Circadian rhythms and memory in agedhumans and animals. In : Squire L, Schacter D, eds. Neuropsycho-logy of Memory (3rd ed). New York NY : Guilford Publishers, 2002 :273-85.
43. YOON C, MAY CP, HASHER L. Aging, circadian arousal patterns,and cognition. In : Park DC, Schwarz D, eds. Cognitive aging : a pri-mer. Philidelphia, PA : Psychology Press, 2000 : 151-71.



























![L’hypothèse en grec ancien [docx TNR]](https://img.pdfslide.fr/doc/110x75/62b4a6c3234c1048f81c9800/lhypothse-en-grec-ancien-docx-tnr.jpg)

